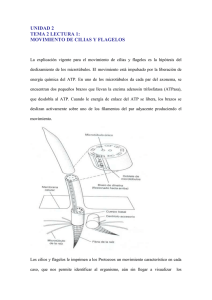
DIVISIÓN HETEROKONTOPHYTA (CHRYSOPHYTA, HETEROCONTÓFITOS O CRISÓFITOS) CARACTERÍSTICAS GENERALES •Comprende una gran diversidad de tipos morfológicos de organización, aunque constituyen un grupo uniforme en cuanto a sus estructuras ultramiscroscópicas. •Los plastos (cromatóforos) son generalmente amarilos, pardo-amarillentos o pardos, debido a la presencia de clorofilas a y c, ß-caroteno (α-caroteno en diatomeas) y diversas xantofilas (diadinoxantina, diatoxantina, fucoxantina, heteroxantina, vauqueriaxantina y zeaxantina). •Los plastos están rodeados exteriormente por un repliegue del retículo endoplasmático que envuelve la típica doble membrana de los mismos; inmediatamente por debajo de ésta se sitúa la llamada laminilla zonal que está constituida por un grupo de tres tilacoides periféricos. •Como sustancias de reserva almacenan principalmente crisolanarina, laminarina y manita, aunque también aceites. •La pared celular presenta en ciertos grupos una matriz de celulosa con alginatos (polisacáridos) y en otros está reforzada con capas suplementarias (escamas o valvas silíceas) •Las células flageladas llevan dos flagelos y son heterocontas; de los dos flagelos, uno es largo, dirigido hacia delante (tractor) y provisto de dos filas de bárbulas o mastigonemas (disposición pennada); el otro, dirigido hacia atrás (impulsor) es liso y está ensanchado en su base. •La posición de los flagelos puede ser apical (cel. Acrocontas) o lateral (cel. Pleurocontas). •En algunas ocasiones las células móviles presentan un único flagelo, y en casos especiales, los organismos unicelulares o las células reproductoras no poseen flagelos. •La reproducción sexual responde a los siguientes tipos: • conjugación isógama o anisógama, • planogamia isógama o anisógama y • oogamia externa o interna. •En cuanto a ciclos biológicos, los heterocontofitos presentan una gran diversidad: • monogenético haplo y diplofásico, • digenéticos isomórficos diplohaplofásicos, • digenéticos heteromóficos con gametófito dominante diplohaplofásicos y • digenéticos heteromórficos con esporófito dominante diplohaplofásicos. CLASIFICACIÓN (ANDERSEN, 2004) 1. Chrysophyceae (golden-brown algae) 2. Synurophyceae 3. Eustigmatophyceae 4. Pinguiophyceae 5. Dictyochophyceae (silicoflagellates) 6. Pelagophyceae 7. Bolidophyceae 8. Bacillariophyceae (diatoms) 9. Raphidophyceae (chloromonads) 10. Xanthophyceae (yellow-green algae) 11. Phaeothamniophyceae 12. Phaeophyceae (brown algae) CLASE: CRYSOPHYCEAE (ALGAS DORADAS) •Son predominantemente de agua dulce, reconocido primero por Stein (1878) y Klebs (1893). •Ha sido separado en diversas clases por Andersen (1999 y 2007). •Las incongruencias entre la clasificación basada en morfología microscópica y la filogenética moderna. •Basados en estudios molecular, ultraestructural y pigmentos han apoyado la eliminación de Haptophyta, Dictyochophyceae, Pelagophyceae y Phaeothamniophyceae de Chrysophyceae. •Poseen clorofilas a, c1 y c2 (Andersen y Mulkey, 1983) •Dos flagelos insertados en la célula perpendicular entre sí, un fotorreceptor en el flagelo corto que generalmente está sombreado por una mancha ocular en el parte anterior del cloroplasto, vacuolas contráctiles en la parte anterior de la célula, retículo endoplásmico del cloroplasto y escamas de sílice simétricas radial o biradialmente (si están presentes). •El producto de almacenamiento es crisolaminarina. •Muchos miembros de la clase producen estatosporas encerradas en una pared silicificada con un poro terminal. •La mayoría de las especies en las Chrysophyceae son de agua dulce y se encuentran en aguas blandas (bajas en calcio). •Muchas de las especies de agua dulce se encuentran en el plancton de los lagos donde están presentes en abundancia. •Los géneros cocoides y filamentosos se encuentran principalmente en manantiales y arroyos fríos, donde ocurren como crecimientos gelatinosos en piedras y madera. •La mayoría de las crisófilas son sensibles a los cambios en el medio ambiente y sobrevive a los períodos por medio de estatosporas. ESTRUCTURA CELULAR : FLAGELO Y MANCHA OCULAR •Muchos de los Chrysophyceae tienen un flagelo de oropel que se inserta en el extremo anterior de la célula paralelo al eje de la célula y un flagelo de latigazo que se inserta perpendicular al flagelo de oropel. •El flagelo del latigazo a menudo es corto. •Los pelos en el flagelo en oropel son usualmente pelos microtubulares tripartitos (Hill y Outka, 1974), aunque se han reportado pelos tripartitos y fibrilares en Ochromonas (Bouck, 1971). • El flagelo posterior en latigazo tiene una hinchazón en su base, la misma que contiene un área densa de electrones denominada fotorreceptor. •La hinchazón flagelar contiene retina, el cromóforo de las proteínas similares a la rodopsina, lo que sugiere que una proteína similar a la rodopsina es el fotorreceptor (Walne et al., 1995). •La hinchazón flagelar encaja en una depresión de la célula inmediatamente debajo de la cual, dentro del cloroplasto, está la mancha ocular. •La mancha ocular está formada por lípidos. DEPÓSITOS EXTRACELULARES •Las paredes celulares compuestas de celulosa, loricas y escamas y paredes silicificadas, aparecen en algunas de las crisófilas. •Las escamas de sílice, como las de Paraphysomona, son radial o biradialmente simétricas. •Las escamas están dispuestas sin apretar fuera de la membrana plasmática sin ningún patrón claramente definido. •Al igual que las Bacillariophyceae y Synurophyceae, las escamas de las Chrysophyceae se forman dentro de una vesícula de deposición de sílice que se deriva del retículo endoplásmico. •Esta disposición difiere de la de las escamas calcificadas de las Prymnesiophyceae y la Chlorophyta, que se forman por el aparato de Golgi. •La formación de escamas también es similar a la formación de frústulas en las diatomeas. •La lórica de O. malhamensis está compuesta por la polisacárida quitina. •La lórica se puede mineralizar como es el caso de C. rufescens. Esta alga tiene una lorica básicamente mucilaginosa que rodea la célula. En las células jóvenes, la lorica es muy transparente, probablemente consiste solo en mucílago. •Anthophysa vegetans, un alga colonial de agua dulce produce un tallo mineralizado que puede contener sales de calcio, hierro o manganeso. Los minerales en el ambiente determinan el color y la composición mineral del tallo. ESTATOSPORAS • La formación de un quiste o estatospora o espora en reposo es una de las características por las cuales un miembro de Chrysophyceae o Synurophyceae puede ser reconocido inequívocamente. • Son generalmente esféricas, elipsoidales u ovadas, y la superficie exterior puede ser lisa o adornada de diversas maneras con verrugas, espinas o brazos. • La ornamentación de la pared es específica de la especie. • En la formación de una estatospora, la célula es inmóvil y existe una considerable actividad de vacuolas contráctiles • Una vesícula casi esférica llamada vesícula de deposición de sílice se forma en el citoplasma. • Se forma una esfera completa de sílice, interrumpida únicamente por el poro y el collar en desarrollo. El núcleo, el cloroplasto, los cuerpos basales flagelares, las mitocondrias, el cuerpo de Golgi, la vesícula de crisolaminarina y los ribosomas se segregan en el interior de la vesícula de deposición de sílice, mientras que en el exterior hay mitocondrias, ribosomas, vacuolas contráctiles y pequeñas vesículas. • Después de que se hayan formado las espinas, el poro y el collar en la vesícula de deposición de sílice, el citoplasma forma un tapón en el área de los poros. • Con la formación del tapón no siliciado, el contacto se pierde entre el protoplasma dentro de la estatospora y el exterior. • Cuando una estatospora germina, se disuelve el tapón o se separa de la pared de la espora. Luego, el protoplasto sale de la estatospora mediante un movimiento ameboide, formando flagelos a medida que se mueve. NUTRICIÓN •Puede ser fototrófica, fagotrófica o mixotrófica (organismo fotosintético capaz de absorber partículas y moléculas del medio) (Zhang y Watanabe, 2001). • Ejm Dinobryon, tiene la capacidad de competir con crustáceos, rotíferos y ciliados en la captura de microorganismos. En condiciones de poca luz, Dinobryon puede ingerir un promedio de tres células bacterianas cada cinco minutos (Bird y Kalff, 1986). •Los crisófitos tienen la capacidad de ingerir presas hasta 30 veces más grandes que ellos mismos. •La capacidad para captar microorganismos parece ser aproximadamente la misma para los crisofitos pigmentados y no pigmentados. •Al igual que otros crisofitos mixotróficos, Epipyxis pulchra fagocita partículas de alimentos que son vivas (bacterias, algas pequeñas o incluso células de su tipo) y no vivas (detritus, materia fecal. • El isofloridosido, un producto de la fotosíntesis en Ochromonas malhamensis, se utiliza para adaptar las células a los cambios de presión osmótica, evitando la pérdida de agua al medio. ORDEN CHROMULINALES • Tienen una sola célula con dos flagellas de inserción apical en algún momento de su ciclo. • Por lo general, hay dos cloroplastos parietales, un núcleo central y una vesícula de crisolaminarin posterior grande. • Dentro del orden hay una progresión de la forma unicelular a la colonial. Ejm Uroglena y Anthophysa. • Algunos como Dinobryon, tienen células rodeadas por una lorica. En Chrysococcus, la célula está rodeada por una lorica que tiene poros en ella. • Hay una tendencia hacia la pérdida de la actividad fotosintética y adaptación a diversas formas de fagotrofia y quimio-organotrofia.Ejm Anthophysa. • En Dinobryon cylindricum, la reproducción sexual es heterotálica y dioica, y morfológica y fisiológicamente anisógama. • En D. divergens, la mayoría de las estatosporas se asientan en el fondo del lago y germinan a principios del próximo año. • Las estatosporas de Dinobryon cylindricum producidas de forma asexual pueden ocurrir en poblaciones exponenciales o en fase estacionaria, dependiendo de los clones involucrados. • La producción continua de un bajo número de estatosporas resistentes por parte de una población de Dinobryon actúa como una cobertura contra un rápido cambio desfavorable en las condiciones ambientales, lo cual mataría a las células vegetativas. • La estatospora madura tiene dos núcleos, dos plastos y un rico suministro de reservas de energía en forma de aceite y criosolaminarina. ORDEN PARMALES • Células pequeñas, generalmente de 2 a 5 um de diámetro, cada una con un cloroplasto y una pared celular silicificada compuesta de cinco a ocho placas. • En Pentalaminaceae hay cinco placas de pared. • En Octolaminaceae hay ocho placas de pared. • Puede haber 105 cél/L en aguas marinas polares y subpolares, lo que los convierte en uno de los grupos más abundantes en estas aguas. • La ornamentación de varios tipos (papilas, alas, espinas, quillas) se produce en las placas. ORDEN CHRYSOMERIDALES • Estas algas tienen zoosporas con los flagelos más o menos lateralmente insertados en el cuerpo celular. • Hay una mancha ocular en el cloroplasto y el pigmento accesorio violaxantina está presente. • Las similitudes en las zoosporas de las algas en este orden han llevado a la especulación de que las Phaeophyceae (algas pardas) probablemente evolucionaron de un alga similar a Giraudyopsis en este orden. • Las algas en los Chrysomeridales, sin embargo, carecen de los esporangios uniloculares y pluriloculares, así como de los plasmodesmas y alginatos característicos de las Phaeophyceae.. CLASE: EUSTIGMATOPHYCEAE (EUSTIGMATÓFITOS) •Organismos unicelulares de color verde amarillo que se producen en agua dulce, agua salobre y agua de mar, así como en el suelo. •Son similares a las de las Xantófilas, difieren en tener una mancha ocular fuera del cloroplasto. •Otras características de la clase incluyen una inflamación basal del flagelo del oropel adyacente a la mancha ocular. •Solo clorofila a •Retículo endoplásmico de cloroplasto no conectado a la envoltura nuclear. •La mancha ocular es un gran cuerpo anaranjado en la parte anterior de la célula móvil y es completamente independiente del cloroplasto. •Los cloroplastos de las Eustigmatophyceae tienen clorofila a y B-Caroteno, siendo las dos xantofilas principales la violaxantina y Vaucheriaxantina, la única diferencia en los pigmentos en comparación con Xanthophyceae es la presencia de violaxantina y la ausencia de antheraxantina. •La vioxantina es el principal pigmento de captación de luz en la Eustigmatophyceae. •La Eustigmatophyceae es un grupo monofilético. •La mayoría de las especies producen zoosporas con un solo flagelo emergente, pero hay un segundo cuerpo basal presente, que indica que las células tenían un antepasado biflagelado. CLASE: XANTHOPHYCEAE (ALGAS AMARILLO - VERDOSAS) • Son principalmente algas de agua dulce y terrestres con unos pocos representantes marinos. • Células móviles con un flagelo dirigido hacia adelante y un flagelo de latigazo dirigido posteriormente. • Los cloroplastos contienen clorofilas a y c, carecen de fucoxantina y tienen un color verde amarillento. • La mancha ocular en las células móviles está siempre en el cloroplasto. • Los cloroplastos están rodeados por dos membranas de retículo endoplásmico cloroplasto. La membrana externa del cloroplasto E.R. generalmente es continua con la membrana externa del núcleo. •En la mayoría de las células no móviles, la pared está compuesta por dos mitades superpuestas. •Está más estrechamente relacionada con la Phaeophyceae. •Aunque la clase se llama comúnmente la Xanthophyceae, el nombre propio es Tribophyceae ya que no hay un género en la clase que pueda prestar su nombre a Xanthophyceae (Hibberd, 1981). ESTRUCTURA CELULAR : PARED CELULAR •La pared celular de Tribonema y Vaucheria están compuestas de celulosa. •En Vaucheria celulosa comprende el 90%, y 10% son polisacáridos amorfos compuestos principalmente de glucosa y ácidos urónicos. •La naturaleza de dos partes de la pared no se puede delinear con el microscopio de luz a menos que las células se hayan tratado con ciertos reactivos, como el hidróxido de potasio concentrado. •La pared se compone de dos partes, una tapa de tamaño constante ajustada sobre una porción basal tubular. A medida que la célula crece y aumenta en longitud, la porción basal tubular se alarga, pero la tapa permanece del mismo tamaño. •Una capa de material intercalar, que presumiblemente funciona como una sustancia cementante, separa la tapa de la parte tubular de la pared. CLOROPLASTO Y SUSTANCIAS DE RESERVA •Dos membranas de cloroplasto E.R. rodean a los cloroplastos, la membrana externa de cloroplasto E.R. es continua con la membrana exterior de la envoltura nuclear. •Los tilacoides se agrupan en bandas de tres y en muchos géneros hay un pirenoide en el cloroplasto. •Clorofilas a y c están presentes en los cloroplastos, siendo los carotenoides principales la diadinoxantina, la heteroxantina y el éster de vaucheriaxantina. •El manitol y la glucosa se acumulan durante la fotosíntesis en los plástidos. El principal producto de almacenamiento es, probablemente, B-1,3 glucano unido a un compuesto similar al paramilón, aunque se ha sugerido que los lípidos también son importantes REPRODUCCIÓN •Se multiplican asexualmente por fragmentación, zoosporas y aplanosporas. •Además, tienen la capacidad de formar esporas especializadas en reposo. •La fragmentación se limita a las colonias de tetrasporina y filamentosas, y se debe a la ruptura de la colonia en partes. •Las zoosporas están formadas por la mayoría de los géneros. •Las zoosporas son desnudas y generalmente piriformes. •La reproducción sexual solo se ha establecido en tres géneros: Botrydium, Tribonema y Vaucheria. CLASIFICACIÓN DE XANTÓFILAS Orden 1 Mischococcales: pequeñas células coccoides. Orden 2 Tribonematales: filamentosas, no cenocíticos. Orden 3 Botrydiales: multinucleados globosos. Talo con rizoides incoloros. Orden 4 Vaucheriales: coenocito filamentoso