Descargar ACTA
Anuncio

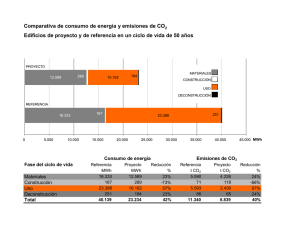
Efecto de la relación NO3-/NH4+ y enriquecimiento de CO2 sobre la actividad antioxidante de plantas de pimiento sometidas a salinidad M. Carmen Piñero1, Ginés Otálora1, Miguel Marín1, Manuel E. Porras2, Francisco M. del Amor1 1 Instituto Murciano de Investigación y Desarrollo Agrario y Alimentario (IMIDA), La Alberca, Murcia, España. 2 Instituto de Investigación y Formación Agraria y Pesquera (IFAPA), Centro La Mojonera, La Mojonera, Almería, España. [email protected] Palabras clave: nitrato, amonio, Capsicum annuum, ascorbato peroxidasa, compuestos fenólicos, peroxidación lipídica. Resumen El estrés salino es uno de los principales factores que limitan el crecimiento y la productividad de los cultivos. La salinidad provoca un estrés oxidativo el cual incrementa el contenido de compuestos fenólicos y la actividad ascorbato peroxidasa, así como genera una mayor peroxidación lipídica. Estudios previos indican que tanto la adición de NH4+ como la exposición a elevado CO2, pueden aumentar la tolerancia a la salinidad mediante la disminución del estrés oxidativo, pero no se ha descrito su efecto de manera conjunta. Por tanto, el objetivo de este experimento fue evaluar el efecto combinado de la aplicación de diferentes proporciones de NO3-/NH4+ y elevada concentración de CO2 en plantas de pimiento sometidas a estrés salino. Las proporciones de NO3-/NH4+ utilizadas fueron 100/0 y 90/10 y la concentración de sal 60 mM NaCl. Este experimento se realizó tanto a 400 y 800 µmol mol-1 CO2. Los resultados obtenidos nos permiten afirmar que con la aplicación combinada de la proporción 90/10 (NO3-/NH4+) y 800 µmol mol-1 CO2, se consigue una disminución significativa de los daños en la membrana lipídica causados por la sal, con una menor formación de compuestos fenólicos y menor actividad de la ascorbato peroxidasa, con respecto a plantas salinizadas con una nutrición sin NH4+. INTRODUCCIÓN La salinidad es un importante factor medioambiental que limita el crecimiento de las plantas y la productividad de los cultivos (Tuna et al., 2008). Debido al uso de aguas de mala calidad y al cambio climático, se están incrementando las zonas cultivables que presentan problemas de salinidad. Además, el continuo aumento de la población mundial, obliga la expansión de los cultivos a tierras salinas no utilizadas anteriormente, para mantener el suministro mundial de alimentos (Purty et al., 2008). Uno de los principales problemas que causa el estrés salino sobre las plantas se debe a un efecto osmótico por la acumulación de sales en la zona radicular (Munns, 2002). A su vez, esto provoca un desequilibrio nutricional en la planta, debido a la acumulación de Cl- y Na+, y al decrecimiento de la concentración foliar de N (Pessarakli, 1991). El desequilibrio nutricional induce un estrés oxidativo en las plantas por la generación de las llamadas especies reactivas del oxígeno (ROS), las cuales provocan daños en las células como es peroxidación lipídica (del Amor et al., 2008). La acumulación de ROS inducida por estrés es contrarrestada por los sistemas antioxidantes enzimáticos como son la ascorbato peroxidasa y los compuestos fenólicos entre otros. El rendimiento de los cultivos de pimiento (Capsicum annuum L.) y su calidad en algunas regiones mediterráneas se está viendo afectado por la salinidad. En estudios anteriores de otros investigadores, se ha comprobado que la aplicación de NO3-/NH4+ en proporciones adecuadas pueden aliviar el desequilibrio nutricional provocado por la salinidad, así como el enriquecimiento del CO2 contrarresta la limitación del crecimiento. Por todo ello, el objetivo de este trabajo fue evaluar el efecto de la aplicación combinada de NO3-/NH4+ y enriquecimiento de CO2 sobre el estrés oxidativo provocado por un estrés salino en plantas de pimiento. MATERIAL Y MÉTODOS Para la realización del estudio se utilizaron semillas de pimiento (Capsicum annuum L.), cv. Melchor (Zeta Seed S.A.) tipo Lamuyo. Las semillas fueron germinadas usando una mezcla de turba y perlita (3:1), y una solución nutritiva de 0.5 mM CaSO4. Después de 26 días, se seleccionaron plántulas con tamaño similar y fueron trasplantadas a macetas negras de 5 l con fibra de coco (Pelemix, Alhama de Murcia, España), las cuales fueron lavadas aplicando 2 l de agua del grifo. La solución nutritiva usada en el experimento fue aplicada mediante goteros autocompensantes (2 h l-1), teniendo siempre un mínimo de 35 % de drenaje. Para evaluar la tolerancia de las plantas a la salinidad en función de la fuente de N aplicada y la concentración atmosférica de CO2, los experimentos fueron llevados a cabo en cámara climática diseñada por del Amor et al., 2010, con condiciones ambientales totalmente controladas: humedad relativa del 50 %, 16/8 h día/noche de fotoperiodo 26/18 ºC, y una radiación fotosintéticamente activa (PAR) de 250 mol m-2 s-1 proporcionada por una combinación de lámparas fluorescentes (TL-D Master réflex 830 y 840, Koninklijke Philips Electronics N.V., de Holanda) y lámparas de alta presión (Son-T Agro, Philips). Los tratamientos aplicados consistieron en dos soluciones con diferentes proporciones de NO3-/NH4+ (100/0 y 90/10) y estas mismas soluciones añadiendo sal (60 mM NaCl). Estos tratamientos fueron aplicados en dos experimentos con diferentes concentraciones de CO2, uno a concentración de CO2 ambiental (400 µmol mol-1 CO2) y otro a elevada (800 µmol mol-1 CO2), y diez plantas por tratamiento fueron usadas. Ambos experimentos finalizaron a los cuarenta y tres días después del trasplante (ddt). La actividad antioxidante fue estudiada mediante la determinación del contenido de compuestos fenólicos totales, la peroxidación lipídica y la actividad de la ascorbato peroxidasa (APOX), que fueron medidos de acuerdo a la metodología descrita por CuadraCrespo y del Amor (2010). Los datos fueron analizados estadísticamente mediante el software SPSS 13.0 por ANOVA y la prueba de rangos múltiples de Duncan (P 0.05), utilizando los tratamientos como parámetros estadísticos, para determinar diferencias significativas entre medias. RESULTADOS Y DISCUSIÓN Bajo condiciones de crecimiento normales, las ROS pueden ser eficientemente eliminadas por los sistemas antioxidantes de las plantas (Singh et al., 2010). Sin embargo, cuando las plantas son sometidas a condiciones de estrés como es la salinidad, la capacidad de los sistemas antioxidantes puede verse excedida, produciéndose daños oxidativos como la peroxidación lipídica, que es considerada como el proceso más perjudicial que se produce en todos los organismos vivos (Garg and Manchanda, 2009). Las especies vegetales tolerantes al estrés suelen incrementar su contenido en antioxidantes en respuesta al estrés, mientras que especies sensibles no lo consiguen hacer de una manera eficiente. En este estudio, investigamos el mecanismo de tolerancia del pimiento por medio de sistemas de defensa de antioxidantes y el nivel de peroxidación de la membrana lipídica. Nuestros resultados indican que en plantas sometidas a salinidad, el contenido en compuestos fenólicos fue reducido solo en aquellas plantas regadas con NH4+ (42.85 % y 70.96 %, a concentración de CO2 ambiental y elevada, respectivamente) (Fig. 1A). Sin embargo, la APOX se incrementó por la salinidad, y en este caso el incremento fue mayor en plantas regadas con NO3- como única fuente de N a ambas concentraciones de CO2 (de 0.1115 µmol g-1 FW a 0.7558 µmol g-1 FW bajo CO2 ambiental y de 0.1519 µmol g-1 FW a 0.6748 µmol g-1 FW bajo CO2 elevado) (Fig. 1B). La aplicación de CO2 por el contrario, provoca una reducción de la APOX en las plantas sometidas a estrés (12 % y 22 % en las plantas tratadas con 100/0 y 90/10, respectivamente) (Fig. 1B), lo cual fue también observado en plantas de soja por Pritchard et al. (2000). Predeciblemente, la salinidad causó daño oxidativo, como pudimos observar por el incremento en la peroxidación lipídica, pero es de destacar, este daño oxidativo desapareció a elevado CO2 (Fig. 2). Polle et al., (1997) también observaron una tendencia a la reducción del contenido de malondialdehído, que es otro indicador de la peroxidación lipídica, en hojas de hayas que crecieron a elevado CO2 comparadas con las que crecieron a concentración ambiental de CO2. En resumen, basándonos en estos resultados, podemos concluir que el mejor régimen de fertilización y la mejor concentración de CO2 para mejorar la tolerancia a la salinidad del pimiento fue la proporción 90/10 y elevada concentración de CO2. Bajo estas condiciones las plantas no presentaron daños en la membrana lipídica a pesar de incrementar en menor medida la actividad de la ascorbato peroxidasa o incluso reducir el contenido de compuestos fenólicos. Por tanto, la ausencia de daño en la membrana lipídica podría deberse a que en plantas regadas con NO3-/NH4+ se produce un menor incremento de las ROS que en las plantas regadas únicamente con NO3-, a pesar de estar sometidas ambas a estrés salino. Agradecimientos Esta investigación ha sido financiada por el INIA (Proyecto: RTA2011-00026-C0201). M.C. Piñero y Manuel E. Porras recibieron el apoyo de una beca predoctoral INIA. Referencias del Amor, F.M., Serrano-Martínez, A., Fortea, Isabel and Núñez-Delicado, E. 2008. Differential effect of organic cultivation on the levels of phenolics, peroxidase and capsidiol in sweet peppers. J. Sci. Food Agric. 88:770-777. Cuadra-Crespo, P. and del Amor, F.M. 2010. Effects of postharvest treatments on fruit quality of sweet pepper at low temperature. J. Sci. Food Agric. 90:2716-2722. del Amor, F.M., Cuadra-Crespo, P., Walker, D.J., Camara, J.M. and Madrid, R. 2010. Effect of foliar application of antitranspirant on photosynthesis and water relations of pepper plants under different levels of CO2 and water stress. J. Plant. Physiol. 167:1232–1238. Garg, N. and Manchanda, G. 2009. ROS generation in plants: boon or bane? Plant. Biosys. 143:8-96. Munns, R. 2002. Comparative physiology of salt and water stress. Plant. Cell. Environ. 25:239–250. Pessarakli, M. 1991. Dry-matter yield, N-15 absorption, and water-uptake by green bean under sodium-chloride stress. Crop. Sci. 31:1633–1640. Polle, A., Eiblmeier, M., Sheppard, L. and Murray, M. 1997. Responses of antioxidative enzymes to elevate CO2 in leaves of beech (Fagus sylvatica L.) seddlings grown under a range of nutrient regimes. Plant, Cell and Environment 20:1317-1321. Pritchard, S.G., Ju, Z., van Santen, E., Qiu, J., Weaver, D.B., Prior, S.A. and Rogers, H.H. 2000. The influence of elevated CO2 on the activities of antioxidative enzimes in two soybean genotypes. Aust. J. Plant. Physiol. 27:1061-1068. Purty, R.S., Kumar, G., Singla-Pareek, S.L. and Pareek, A. 2008. Towards salinity tolerance in Brassica: an overview. Physiol. Mol. Biol. Plant. 14:39–49. Singh, S. and Tuteja, N. 2010. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant. Physiol. and Biochchemistry 48:909-930. Tuna, A.L., Kaya, C., Higgs, D., Murillo-Amador, B., Aydemir, S. and Girgin, A.R. (2008). Silicon improves salinity tolerance in wheat plants. Environ. Exp. Bot. 62:10–16. 1.000 0.008 [Acid Galic] (mg ml-1) 0.006 de de B d 0.800 c c e cd bc b bc 0.004 0.600 ab a 0.400 0.002 a a a 0.200 a 0.000 0.000 100/0 + NaCl 100/0 90/10 90/10 + NaCl 100/0 + NaCl 100/0 90/10 90/10 + NaCl Fig. 1. Efecto de la salinidad, dos formas de nitrógeno y elevada concentración de CO2 en plantas de pimiento: A) compuestos fenólicos totales y B) actividad de la ascorbato peroxidasa. Los datos son medias ±SE de diez plantas. Las letras indican diferencias significativas a p 0.05 entre tratamientos. APOX (µmol g-1 FW) 400 µmol mol-1CO2 800 µmol mol-1CO2 A 0.006 TBARS (µmol g-1 FW) 0.005 400 µmol mol-1CO2 800 µmol mol-1CO2 c bc 0.004 ab ab ab ab a a 0.003 0.002 0.001 0.000 100/0 + NaCl 100/0 90/10 90/10 +NaCl Fig. 2. Efecto de la salinidad, dos formas de nitrógeno y elevada concentración de CO2 en plantas de pimiento: peroxidación lipídica a concentración ambiental y elevada de CO2. Los datos son medias ±SE de diez plantas. Las letras indican diferencias significativas a p 0.05 entre tratamientos.