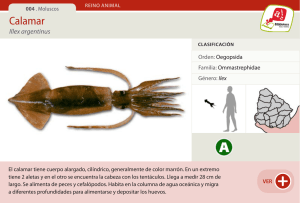
Calamar (Illex argentinus): Stock Sud-patago nico al
Anuncio
: Stock Sud-patago nico al")
Calamar (Illex argentinus): Stock Sud-patago nico al sur de 45 LS Prenski, Leszek Bruno & Morales-Yokobori, Marcelo L. Terra Curanda www.terracuranda.org Buenos Aires, Febrero 2011 Summary Argentina recovered the leadership as a squid (Illex argentinus) production country in 1993 although with declining yields since 1997. In 1997, the captures of squid were higher than 900,000 tons considering different fleets, while in 2007 were estimated in more than 800,000 tons. This work analyses the possible causes for the declining of the squid stock, arising consequently declines on captures. Reproductive capacity and prey-predator relationship aspects are analysed in this work in order to provide an insight on the population dynamics of this species. Relationships with hake (Merluccius hubbsi) which is considered a predator of squid adults and juveniles are also analysed. It is observed that good years for the squid fishery alternate with good ones for the hake fishery. As well, feed habits of the squid, including cannibalism, are analysed considering ranges of depth. It is observed that the squid maintains the proportion of its diet across years. Also, environmental aspects like the Malvinas Current, with determining impacts on the dynamics of the species, are analysed. Finally, biological and environmental variables are analysed in order to predict stock status in order to provide basis for management decisions. Contenido Summary .................................................................................................................................. 1 Contenido ................................................................................................................................. 1 Breve reseña histórica............................................................................................................... 2 Distribución estacional del calamar ........................................................................................... 5 Relación merluza-calamar. ........................................................................................................ 7 Efecto indirecto de la pesca sobre la merluza. ....................................................................... 8 Análisis de la Relación Merluza – Calamar (Figura 9)............................................................ 10 Cambios generales percibidos ............................................................................................. 12 Alimentación del calamar........................................................................................................ 16 Zona Patagónica.................................................................................................................. 17 Composición de la dieta con respecto a la profundidad ....................................................... 17 Consumo del alimento del calamar en el área austral en relación al zooplancton ................ 18 Simulación del consumo de calamar.................................................................................... 20 Densidad de Zooplancton y el reclutamiento del calamar y otras especies y la intensidad pesquera. ............................................................................................................................ 24 Canibalismo como mecanismo compensador e índice de deficiencia en la alimentación ..... 27 Condiciones del manejo del calamar y su repercusión en el ecosistema pesquero................... 30 Medioambiente y reclutamiento de calamar ....................................................................... 30 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori Transporte de la Corriente de Malvinas y reclutamiento ..................................................... 31 Manejo del calamar y su repercusión en el ecosistema pesquero ........................................ 36 Seguimiento y alerta temprano de cierre de la pesquería de Illex argentinus. Tasa de escape fija o dinámica..................................................................................................................... 38 Procedimiento del criterio de alerta temprano y recomendación de cierre de la pesquería (Dinámico) .......................................................................................................................... 40 Relación entre el reclutamiento de calamar y reclutamiento de merluza y su relación con la pesca .................................................................................................................................. 41 Breve reseña histórica La pesca del calamar ha existido en Argentina desde 1946. Hasta 1977, la especie fue capturada como acompañante en la pesca de merluza y los desembarcos totales no superaron las 5.000 t anuales. A partir de 1978 se intensifica la demanda externa y la magnitud de las capturas. Los buenos rendimientos obtenidos en esos años alentaron el arribo de flotas extranjeras de ultramar, especialmente de Polonia y Japón, que comenzaron a operar por fuera de la ZEE, sobre las concentraciones pre-reproductivas de calamar entre febrero y junio. Al mismo tiempo, arrastreros argentinos iniciaron la pesca dirigida a la especie en la región bonaerense-norpatagónica, lo cual condujo a una notable expansión de la pesquería. Se observa en el siguiente gráfico la captura de distintas Flotas. Los desembarcos alcanzaron un máximo cercano a las 500.000 t en las temporadas 1987/88, pero sólo un 10% de ellos correspondieron a la Argentina. Argentina recuperó su posición de liderazgo como país productor de calamar a partir de 1993, condición que ostentaba hasta el 2007, aunque con rendimientos decrecientes desde 1997, año para el que se consignan declaraciones máximas de captura, cercanas a las 400.000 toneladas. Debe destacarse la progresiva declinación de las capturas acumuladas de todas las flotas que participaron de la pesquería en los últimos años, las que básicamente se reducen en un orden de magnitud, pasando de casi un millón de toneladas en 1997 a menos de cien mil toneladas en 2004 y 2005 al Sur de 45° LS. En el año 2008 las capturas en alta mar fueron estimadas por fuentes privadas (CAPeCA ) en 450.000t. Terra Curanda www.terracuranda.org/fisheries 2 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori CAPTURA DE CALAMAR POR DISTINTO TIPO DE FLOTAS 1000000 900000 800000 TONELADAS 700000 600000 500000 400000 300000 200000 100000 0 1991 1992 1993 1994 1995 1996 1997 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 ANIOS SUR 45 Argentina Malvinas Alta Mar Figura 1: Fuente: elaboración propia en base a datos extraídos de Brunetti et al., Informes Técnicos INIDEP: 77108/1996, 143/1997, 29-147/1998, 54-65/99, 93/2000, 86/2001, 87/2002, 103/2003, 70/2004, 50/2005, 53/2006, 61/2007, 52/2008; Revista Redes; Información CAPeCA. Se recupera en el 2006 y en el 2007 más plenamente alcanzando al sur de 45° LS una captura de más de 800000 toneladas. En el 2008 superan las 700000 toneladas. Las causas de la caída de la biomasa de calamar y por lo tanto de la captura sobre la especie se analizarán en el curso del presente trabajo, en el cual se perfila el problema como un tema de sobrepesca y de años de cambios ambientales probablemente no favorables para el desarrollo del calamar. En el gráfico inferior se observan las capturas por flotas de poteros y arrastreros tanto argentinos como de Malvinas. Se puede observar que las capturas son generadas en su mayoría por los poteros y el año de mayor presencia de unidades pesqueras fue 1997. Figura 2: Fuente: extraído de Brunetti et al., Informe Técnico INIDEP 61/2007. En el siguiente gráfico se observan las capturas totales de calamar asociados al reclutamiento del stock Sur Patagónico SSP. Terra Curanda www.terracuranda.org/fisheries 3 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori CAPTURA DE CALAMAR SUR DE 45 LS E ÍNDICES DE RECLUTAMIENTO 1200 5,0 CAPTURA EN TON EN MILES 4,0 3,5 800 3,0 600 2,5 2,0 400 1,5 1,0 200 RECLUTAMIENTO DEL SSP EN N 10/9 4,5 1000 0,5 0 0,0 2009 2008 2007 2006 2005 2004 2003 2002 2001 2000 1999 1998 1997 1996 1995 1994 1993 1992 1991 1990 1989 1988 1987 1986 1985 1984 1983 1982 1981 1980 1979 1978 AÑOS Captura Sur 45 en miles t N Ind. Recl 10/9 Figura 3: Fuente: elaboración propia en base a datos extraídos de Brunetti et al., Informes Técnicos INIDEP: 77108/1996, 143/1997, 29-147/1998, 54-65/99, 93/2000, 86/2001, 87/2002, 103/2003, 70/2004, 50/2005, 53/2006, 61/2007, 52/2008: Csirke, J., FAO Doc. Téc. de Pesca 286; Revista Redes; Información CAPeCA. Si bien los datos de reclutamiento tienen dos orígenes diferentes se observa un grado de relación elevada entre la captura y los índices de reclutamiento del SSP. En las especies de ciclo vital corto, tal el caso del calamar, debe recordarse que la aplicación del concepto de Rendimiento Máximo Sostenible no es siempre la más adecuada. El cálculo de dicha magnitud ha sido concebido para especies longeva o relativamente longevas, como una forma de asegurar niveles de extracción que puedan ser mantenidos a largo plazo sin afectar negativamente la capacidad de producción de las poblaciones naturales. En cambio, la disponibilidad de recursos pesqueros con ciclos de vida anuales, o casi anuales, depende en gran medida del éxito de una temporada de reproducción y de los siguientes eventos que conducirán al reclutamiento. Terra Curanda www.terracuranda.org/fisheries 4 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori Variaciones del reclutamiento y de los desovantes que le dieron origen desovante en Numero del SSP corridos un año, para una relación directa 5000 1400 4500 Reclutamiento en Numero 1000 3500 3000 800 2500 600 2000 1500 400 1000 Biomasa desovante en Numero 1200 4000 200 500 0 19 78 19 79 19 80 19 81 19 82 19 83 19 84 19 85 19 86 19 87 19 88 19 89 19 90 19 91 19 92 19 93 19 94 19 95 19 96 19 97 19 98 19 99 20 00 20 01 20 02 20 03 20 04 20 05 20 06 20 07 20 08 20 09 0 Años SSP Numero de Reclutas 10/6 SSP Biomasa Desovante N Figura 4: Fuente: elaboración propia en base a datos de Brunetti et al., Informes Técnicos INIDEP: 77-108/1996, 143/1997, 29-147/1998, 54-65/99, 93/2000, 86/2001, 87/2002, 103/2003, 70/2004, 50/2005, 53/2006, 61/2007, 52/2008. En términos generales, el manejo y evaluación del calamar se realiza a partir del conocimiento del tamaño de la población reclutada anualmente y de su evolución a lo largo de la temporada de pesca. El objetivo es permitir un escape suficiente de desovantes, que asegure un adecuado reclutamiento el siguiente año. Esto se logra regulando la captura total, o bien el esfuerzo por temporada de pesca. Distribución estacional del calamar “En otoño se observan importantes concentraciones pre-reproductivas a lo largo de la plataforma externa y talud continental, que no son coincidentes en el tiempo. Al sur de los 44° LS dichas concentraciones ocurren entre marzo y mayo, mientras que al norte se reconocen entre abril y julio, correspondiendo las mismas a diferentes unidades demográficas: Subpoblación Sur-patagónica (SSP) y Bonaerense–Nor-patagónica (SBNP), respectivamente. En invierno, desaparecen totalmente los grandes centros de concentración de adultos al sur de los 44° LS, indicando que la SSP ha completado su emigración de desove. Núcleos de agregación permanecen al norte de la latitud antes señalada y corresponden a la SBNP, que concreta su emigración hacia fines de agosto-setiembre. En primavera, las mayores concentraciones se localizan en la plataforma bonaerense-norpatagónica, entre los 50 y 100 m de profundidad, y comprenden juveniles (SSP, SBNP) de hábitos pelágicos, que han completado o completarán hacia el final de la temporada su migración al área de crianza y su reclutamiento al fondo, pre-adultos de la Subpoblación Desovante de Verano (SDV) y adultos de la Subpoblación Desovante de Primavera (SDP). Terra Curanda www.terracuranda.org/fisheries 5 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori En verano se detectan dos áreas de altas concentraciones, una entre los 43°S y 45°S, correspondiente a individuos en reproducción de la SDV, y la otra entre los 46°S y 48°S, conformada por pre-adultos de la SSP (Brunetti, N. “Diagnostico de las pesquerías argentinas”, INIDEP, 2000). 36° Illex argentinus SBNP SDP 40° SDV 44° SSP Masas de huevos 48° 52° AREAS DE DESOVE 70° 66° 62° 58° 54° 50° 46° 56° 42° Figura 5: Fuente: extraído de Brunetti, N. Diagnóstico de las pesquerías argentinas, INIDEP, 2000. •VERANO •En verano las mayores concentraciones ocurren 34° 36° Verano •Concentraciones reproductivas del SDV (14(14-25 cm 38° 40° 42° •SDV •SSP 44° 48° 52° 54° 68° 66° 64° 62° 60° 58° 56° ML) en el norte (43(43-46° 46°S). •Concentraciones tró tróficas del SSP (17(17-29 cm ML) en el sur (46° (46°S - 51° 51°S). % 25 20 15 10 5 0 46° 50° 70° sobre la plataforma patagó patagónica (43(43-55° 55°S), donde dos stocks está están presentes: 54° 10 15 15 20 % 20 15 10 5 0 20 ML (cm) 25 ML (cm) 25 30 30 35 52° Figura 6: Fuente: extraido de Brunetti, N.. Diagnóstico de las pesquerías argentinas, INIDEP, 2000 Terra Curanda www.terracuranda.org/fisheries 6 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori Tal como se describió sumariamente en el párrafo precedente el ciclo anual del calamar para el área austral es solo relevante, desde el punto de vista de la pesquería, en las estaciones de verano y otoño. De Brunetti N. (2000) se exponen los gráficos correspondientes a esas estaciones para comprender la dinámica del ecosistema pesquero del área austral. 34° •OTO ÑO Otoño •En otoñ otoño las mayores concentraciones se encuentran a lo largo de la plataforma externa y talud continental. 36° •Al sur de los 43° 43°S las mayores concentraciones ocurren en marzo - mayo y corresponden al SSP. •SBNP 38° 40° • •Al norte de los 43° 43°S las concentraciones se dan en mayo - julio y corresponden al SBNP. 42° 44° •SSP 46° 15 48° 50° 52° 54° 70° 68° 66° 64° 62° 60° 58° 56° 54° 20 15 % 10 5 0 20 20 15 % 10 5 0 15 52° 20 25 ML (cm) 25 30 30 35 35 ML (cm) Figura 7: Fuente: extraído de Brunetti, N., Diagnóstico de las pesquerías argentinas, INIDEP, 2000. Relación merluza-calamar. Esta relación es usada para comprender la dinámica más general que abarcan todas las especies australes que interesa observar el área. Desde un punto de vista general se trata en realidad de una relación de predador-presa en la cual están involucradas las especies Moroteuthis ingens, Illex argentinus, Loligo sanpaulensis y L. gahi. En este análisis del ecosistema pesquero austral se describirán calamares como Illex argentinus y Moroteuthis ingens y, bajo la denominación de Loligo sp., a las dos especies de loligo anteriormente mencionadas. Existe significativa información acerca de las estimaciones de los efectivos de calamares por distintas áreas de pesca del Mar Argentino (Brunetti, N.; Informe Técnico INIDEP: 77-108/1996, 143/1997, 29-147/1998, 54-65/99, 65/2000, 86/2001, 87/2002, 103/2003, 70/2004, 50/2005, 53/2006). La información del área no regulada más allá de la milla 201 se obtuvo de CAPECA, a través de la relación intercámaras e información proveniente de otras flotas del “Falkland Fisheries Statistics Bulletin”. Las capturas comerciales nacionales e internacionales, particularmente para la especie Illex argentinus, experimentaron un aumento continuo en la década del ‘80 desde 150.000 t. a 600.000 t/año para luego estabilizarse en aproximadamente 700.000 t., aunque en algunos años registraron picos de mas de 900.000 t y en 2005 y 2006 se produjo una abrupta caída a 100.000 t. debido a una falla de reclutamiento. Terra Curanda www.terracuranda.org/fisheries 7 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori Efecto indirecto de la pesca sobre la merluza. Para la merluza, la presa de mayor importancia trófica dentro del ítem calamares es la especie Illex argentinus (calamar común), cuyos efectivos se hallan diseminados en toda el área de distribución ecológica de la merluza común. En el siguiente gráfico, y en la medida de poder explicar desde el punto de vista trófico el aumento de la biomasa de calamar, se trataron de integrar los datos obtenibles de las campañas de evaluación de la biomasa de la merluza común, en este caso total, que incluye prácticamente toda la distribución de la especie. Este dato fue así integrado en función de que el ciclo de vida del calamar abarca la distribución de la merluza, y por lo tanto ésta preda sobre juveniles y adultos. Más adelante se verá esta relación de estructura. Lo que puede observarse es una disminución importante de la biomasa y por lo tanto de la predación. Se anexaron los datos de la totalidad de la captura de la especie calamar del área austral obtenidos de diferentes fuentes: Subsecretaría de Pesca y Acuicultura, CAPeCA (del área no regulada) y del Falkland Fishery Bulletin. Debido a la significancia de la captura, se anexaron los datos de Csirke (1987), que a través de datos de CPUE, calculó el reclutamiento del calamar de la zona austral, como reflejo de las etapas iniciales de la pesca sobre el calamar. Para el período posterior se utilizaron los datos de Brunetti N., Inf. Técnico INIDEP: 77-108/1996, 143/1997, 29-147/1998, 54-65/99, 65/2000, 86/2001, 87/2002, 103/2003, 70/2004, 50/2005, 53/2006; Cordo D y Renzi M. & Irusta G. INIDEP 36-39/2006. CAPTURA DE CALAMAR, ÍNDICES DE RECLUTAMIENTO DE CALAMAR Y RELACIÓN CON LA BIOMASA DE MERLUZA POR CAMPAÑAS, ÁREA TOTAL 1200 5,0 4,5 CAPTURA EN TON EN MILES 4,0 3,5 800 3,0 600 2,5 2,0 400 1,5 1,0 RECLUTAMIENTO DEL SSP EN N 10/9 1000 200 0,5 0,0 19 78 19 79 19 80 19 81 19 82 19 83 19 84 19 85 19 86 19 87 19 88 19 89 19 90 19 91 19 92 19 93 19 94 19 95 19 96 19 97 19 98 19 99 20 00 20 01 20 02 20 03 20 04 20 05 20 06 20 07 20 08 20 09 0 AÑOS Captura Sur 45 en miles t N Ind. Recl 10/9 Campañas área total Bio Merl 10/6 Figura 8: Fuente: elaboración propia en base a datos obtenidos de Brunetti, N., Inf. Tecnico INIDEP: 77-108/1996, 143/1997, 29-147/1998, 54-65/99, 65/2000, 86/2001, 87/2002, 103/2003, 70/2004, 50/2005, 53/2006; 61/2007, 52/2008, 15/2009; Cordo, D. y Renzi, M. & Irusta, G.; INIDEP 36-39/2006; Irusta, G., Renzi, M. ; D’ Átri, l.; Informe Terra Curanda www.terracuranda.org/fisheries 8 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori INIDEP 43 / 2007; Irusta, G., Renzi, M.; Informe INIDEP 47 /2007; información de DINPyA, CAPeCA, Fishery Statistical Bulletin Falkland Island. De esto se puede deducir a través de los análisis anteriores que la baja de la biomasa de merluza provocó un neto aumento de las biomasas de calamar, con un efecto secundario que lo favoreció, ya que al ser la merluza un pez que preda sobre el zooplancton, también dejó libre zooplancton que fue aprovechado como alimento por el calamar. Con ello el efecto fue doblemente importante. El gráfico y descripción subsiguiente comprende la dinámica de predación en toda el área de distribución de la merluza. El interés de esta descripción detallada es su posible utilización como base interpretativa de la relación predador presa para las demás especies que se asocian sobre el mencionado ítem, no solo como especie sino sobre los stocks del cefalópodo. El sector denominado austral abarca la sección comprendida entre 45° LS y el área más austral de Argentina. En la presentación de Angelescu, V. & Prenski, L. B. (1987) (en la Tabla 15 del trabajo original) se dan los resultados del muestreo agrupados por clases de largo del predador con los valores promedio y la amplitud de variación de la longitud del manto (LM) y del peso total individual de las presas. Así mismo, el estudio de la dinámica de la predación efectuado por zonas latitudinales y batimétricas a lo largo del año, permite establecer los ciclos estacionales de los desplazamientos en el plano horizontal de los cardúmenes de la merluza, en seguimiento de los efectivos de la presa según la aparición y desaparición de sus cohortes en el área del hábitat trófico común. Resulta de este cotejo que la predación ocurre sobre distintos grupos de tamaño de calamar con amplia variación de la longitud del manto, al tratarse de incidencias sucesivas espacio-temporales desde el grupo de los juveniles al de los adultos, o a la vez sobre los cuatro stocks de calamar. En el gráfico siguiente se puede observar la coexistencia de tamaños totalmente diferentes de la misma especie, que se corresponde con la evolución de los stocks de calamar en especial con el SSP y el SDV que se pueden observar en los gráficos precedentes de distribución de calamar obtenidos de Brunetti, N. (2000), en referencia al área austral (D). Si bien el trabajo de Angelescu, V. & Prenski, L. B. trata de la predación sobre calamar de la merluza en su total dispersión, la parte asociada al área austral es la que corresponde al área D. Terra Curanda www.terracuranda.org/fisheries 9 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori Figura 9: Relación Merluza – Calamar con la variación del largo de manto (LM cm) de las presas, según zonas latitudinales y batimétricas y meses del ciclo anual. (I) = de 1-10 individuos (III) de 10-30 individuos. Fuente: extraido de Angelescu, V.; Prenski, L. B.; 1987, INIDEP. Análisis de la Relación Merluza – Calamar (Figura 9) a) Ciclo de predación en las zonas latitudinales A y B (36°-40° LS; isobatas de 70 -400 m). Se trata de un ciclo anual completo de predación que comienza en el lapso de juniojulio con la aparición de una cohorte de juveniles de la presa con el largo del manto comprendido entre 1-2 cm, originada de los desoves invernales del efectivo norteño; los mismos juveniles alcanzan en el mes de noviembre un tamaño de 9 a 14 cm LM y en el lapso de diciembre-enero sobrepasan los 18 cm del LM para convertirse entre los meses de mayo y junio en adultos maduros (20 - 30 cm LM). Estos ya en estado de desove se mezclan en el mes siguiente (julio o agosto) con la nueva generación y después desaparecen del área del hábitat trófico común. Por lo tanto, le amplitud de predación se extiende sobre todas las clases de tamaño de la presa (1 - 30 cm LM) y las incidencias aumentan a medida que los juveniles se adentran durante los meses de invierno y primavera en la región interna de la plataforma bonaerense, en la cual se encuentran también los juveniles y los adultos del predador con una variación de talla Terra Curanda www.terracuranda.org/fisheries 10 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori de 15 a 60 cm Lt. Se puede destacar que aquí las incidencias son totales en la relación predador-presa, mientras que en la región de aguas profundas del talud continental (200-400 m) se produce un corrimiento hacia tamaños mayores de los individuos de ambos grupos integrantes de dicha relación. b) Ciclo de predación en la zona latitudinal C (40° - 42°S; isobatas de 100 - 250 m). Igualmente que en las zonas anteriores, el ciclo anual se inicia en el mes de julio con una cohorte proveniente del mismo efectivo norteño del calamar que está integrada por juveniles de tamaño algo mayor (7 cm LM), por provenir de los desoves más tempranos de los meses de mayo y junio; éstos llegan a la talla máxima de adulto (28 30 cm LM) en el lapso junio-julio del año siguiente cuando el ciclo queda concluido. El impacto mayor de predación se comprueba entre los meses del otoño y principio del invierno sobre las mismas clases de tamaño dé la presa mencionadas para las cohortes de las zonas A y B. c) Ciclo de predación en la zona latitudinal D (42° - 48°S; isobatas de 100 - 250 m). En el área que comprende esta zona hay un cambio de situación en la relación predadorpresa que se debe a la presencia alternante de individuos de la presa de dos efectivos diferentes, según la época y sus áreas de desoves (Brunetti, N.; 1981). El ciclo anual se inicia en el lapso junio-julio con la aparición de una cohorte de juveniles (2 - 7 cm LM) en la región comprendida entre el talud continental y la plataforma externa, originada de los desoves otoñales (abril-mayo) del efectivo de aguas profundas. Los adultos que integran este efectivo son de tamaño grande (28 - 30 cm LM) y llegan en su dispersión hasta el sector norteño de la zona latitudinal E (48° - 52°S). El segundo efectivo es propio de la región de aguas de profundidad menor de la plataforma (< 150 m) y se halla distribuido en toda la extensión latitudinal de la zona D. El ciclo correspondiente a este efectivo comienza en el lapso de enero-febrero con la aparición de la cohorte de la nueva generación. Los juveniles se mantienen en la misma región de su origen y alcanzan el tamaño de adulto (20 - 22 cm LM) en el mes de enero del año siguiente. En ese momento ocurren la puesta y seguidamente la muerte post-desove. Resulta de este proceso cíclico entre las cohortes intra-anuales de dos efectivos diferentes una permanente mezcla de las clases de tamaño del calamar en el área del hábitat trófico del predador que a su vez está representado por individuos secundarios, pre-adultos y adultos hasta un límite máximo de talla de 35 cm LM. En consecuencia, debido a la mezcla permanente de tamaños de la presa, existe una variación continua de amplitud dimensional-somática en el espectro trófico de los individuos de merluza que se hace más evidente en los meses de primavera (octubre y noviembre), cuando los preadultos del efectivo con desoves en verano SDV y los juveniles descendientes del efectivo de aguas profundas SSP se encuentran en un área de distribución colindante o superpuesta en la plataforma patagónica correspondiente a la zona D. Por lo general, el consumo de calamares incide en la mayoría de los casos sobre individuos de tamaños mediano a grande (> 15 cm LM); el número de presas ingeridas por un individuo de merluza es reducido, con una variación de 1 a 2 presas, correspondiendo estos números a casi 96% de la totalidad de merluzas con calamares en el contenido estomacal. La relación de tamaño entre el predador (Lt) y las presas examinada mediante la aplicación de una regresión de ajuste a una función potencial, demuestra que los calamares aparecen como un "alimento generalizado" en la dieta de la merluza; sin embargo, alcanzan una preferencia a partir de la talla de 40 cm Lt en adelante.” La pesca intensiva del SSP en el área no regulada, especialmente por no respetar los porcentajes de escape que deben mantenerse para permitir un buen reclutamiento en el año Terra Curanda www.terracuranda.org/fisheries 11 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori siguiente, tiene efectos diferenciales en las relaciones de las distintas especies. La acción de la pesca sobre el SSP provoca modificaciones en el medio, beneficiando a la merluza de cola por un aumento del alimento disponible de crustáceos pelágicos y perjudicando a la merluza común si los reclutamientos del calamar no son los adecuados para mantener la captura trófica y a la pesca en un nivel suficiente. Esta relación debe ser tomada en cuenta en el manejo de las especies del área austral. La pesca no regulada plantea serios problemas, tanto en el tema del futuro reclutamiento por la sobrepesca de stock desovante, como sobre la interrelación inter-específica. Cambios generales percibidos 1- El aumento continuo de las capturas comerciales de merluza común en la "Zona Económica Exclusiva" (ZEE) del Mar Argentino, especialmente en el período 1970 1983, tuvo como efecto la disminución de la predación sobre los calamares, que a su vez causó un probable aumento de la biomasa de sus efectivos en dicha zona de pesca provocando el aumento de la presión pesquera sobre el calamar. 2- La pesca intensiva de calamares dentro y fuera del área circundante a las Islas Malvinas a partir del año 1984, habría afectado probablemente a los individuos de merluza común de las clases superiores a los 50 cm Lt, causando una mayor mortalidad natural por la merma del alimento principal en esta área. 3- Un mayor aumento del esfuerzo de pesca sobre los efectivos de calamares en la región austral del Mar Argentino comprendida entre las latitudes de 45° y 50° LS, habría provocado una disminución de la disponibilidad del alimento principal para las especies de peces predadores como por ejemplo las merluzas común y austral, la merluza de cola y otras (ver tabla siguiente sobre las distintas especies que predan sobre el calamar). Las investigaciones tróficas realizadas en el año 1987 en la región mencionada respaldaron lo observado por Prenski & Angelescu (1993) mostrando que el alimento principal estaba constituido por un 75% de especies de calamares. En cambio, para los años 1992 y 1993, esa cantidad había disminuido de 6 a 5 veces en el consumo instantáneo de todas las especies del área austral. Ver ampliación del tema en Consumo instantáneo. En cuanto al concepto de ciclicidad entre especies en el siguiente gráfico se expusieron valores de reclutamiento de calamar (N+10/9) del SSP obtenidos de los informes de la Dra. Norma Brunetti y a los cuales se le adjuntó los valores de reclutamiento de la merluza de la edad 1, efectivo Norte y Sur de 41° LS obtenidos por modelo, y del efectivo sur de la campaña de juveniles, información extraída de los informes de INIDEP. Terra Curanda www.terracuranda.org/fisheries 12 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori RELACIÓN ENTRE EL RECLUTAMIENTO DE CALAMAR SSP Y MERLUZA 5000 4500 NÚMERO DE IND 10/6 4000 3500 3000 2500 2000 1500 1000 500 0 1986 1987 1988 1989 1990 1991 1992 1993 1994 1995 1996 1997 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 AÑOS SSP Numero 10/6 Reclutamiento de la merluza Norte edad 1 Modelo Reclutamiento de la merluza Sur edad 1 Modelo Reclutamiento edad 1 Camp. De juveniles Figura 10: Fuente: elaboración propia en base a datos obtenidos de Brunetti, N.; Inf. Tecnico INIDEP: 77-108/1996, 143/1997, 29-147/1998, 54-65/99, 65/2000, 86/2001, 87/2002, 103/2003, 70/2004, 50/2005, 53/2006, 61/2007, 52/2008, 15/2009; Cordo, D. y Renzi, M. & Irusta, G.; INIDEP 36-39/2006; Irusta, G.; Renzi, M.; D’ Átri, l.; Informe INIDEP 43 / 2007; Irusta, G.; Renzi, M.; Informe INIDEP 47 /2007; Renzi M et al Informe Tec 46/2008, 32/2008 Louge, E.; Santos, B.; Castrucci, R., 10-2002; Louge, E.; Reta, R.; Santos, B.; 103-2005; Cordo, H.; 39-2006; Dato, C.; Pajaro M.; 8-2007. Observando el gráfico se puede decir que parecería que años buenos de calamar se contraponen con años malos de merluza. Se incluyeron los dos efectivos debido a que las larvas y pre-adultos de calamar migran por la plataforma al sur siendo objeto de captura trófica por ambos efectivos de merluza. Las posibles explicaciones pueden ser dos; la primera apunta a cambios oceanográficos que actúan de manera diferencial en una relación trófica o causalidad. De este modo, como se observa en el gráfico, ésto podría indicar al año 2007 como un año malo para merluza en el reclutamiento de la edad 1 y bueno para el calamar. Como conclusión podemos decir que por ser una pesquería basada en pocas clases de edad 2 y 3, cualquier disminución en el reclutamiento inmediatamente se reflejará en la potencialidad de la pesquería. También se puede interpretar de una manera diferente; es decir que años con buenos reclutamientos de merluza común actúan de tal manera que el importante número de juveniles preden intensamente sobre las larvas y juveniles de calamar disminuyendo así su reclutamiento. Terra Curanda www.terracuranda.org/fisheries 13 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori Cotoperca gobio Merluccius hubbsi Macruronus magellanicus Acanthistius brasilianus Schroederichties bivius Dipturus chilensis Squalus acanthias Patagonothotem ramsayii Sebastes ocullatus Merluccius australis Dissostichus eleginoides Micromesistius australis Bathyraja albomaculata Psamobatis scobina Bathyraja scaphiops Bathyraja griseocauda Bathyraja brachiurops Terra Curanda 10,96 3,23 9,62 19,61 11,19 --23,66 11,11 24,88 0,85 0,00 0,00 1,92 0,00 8,11 0,00 7,69 6,90 7,69 1,09 0,00 0,83 3,57 0,23 --9,71 15,18 0,00 0,00 0,00 23,08 0,00 0,00 0,00 ------- 0,00 0,00 0,00 0,37 1,43 --0,91 0,00 1,26 0,00 0,00 0,00 0,00 0,57 --0,00 5,88 0,00 --- www.terracuranda.org/fisheries 1,61 0,00 0,45 1,64 5,08 --0,99 0,00 3,10 0,65 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 8,70 4,4 --20,5 28,86 ----21,8 1,0 10,4 --40,7 11,5 31,13 ----0,50 --0,00 0,00 ----6,44 0,00 --3,1 0,00 20,48 ----3,90 --0,00 ----------- N tot muestr. VER95 0,00 0,00 1,64 0,00 0,31 --2,04 0,00 14,81 0,69 --1,43 0,00 0,00 5,00 0,00 ----0,00 VER 94 8,35 0,22 3,70 21,49 2,64 6,25 13,46 0,00 24,27 0,27 0,36 0,00 0,00 0,00 2,56 1,52 0,00 0,00 --- INV93 OTO 93 Salilota australis VER93 Genypterus blacodes PRIM 92 PECES, PREDADORES SOBRE CALAMAR OTO92 ESTACIONES ANUALES VER 92 Tabla 1: Relación de predación de las especies de peces sobre calamar. Predación sobre calamar, expresadas en frecuencias de ocurrencia (estómagos observados con el ítem calamar relacionado con el total observado, en porcentaje) de todas las especies presentes en los lances de pesca australes. Fuente: elaboración propia datos obtenidos de información no publicada del Proyecto Especies Australes, Prenski, L.B. y Sanchez, Felisa et al, Informe Técnico INIDEP, 88/2002. 3271 1663 788 9053 22906 16 1229 56 2311 3320 486 1159 162 2341 214 295 50 69 159 14 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori Figura 11: Áreas de predación ictícola sobre calamar. Fuente: elaboración propia datos obtenidos de información no publicada del Proyecto Especies Australes Terra Curanda www.terracuranda.org/fisheries 15 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori Figura 12: Distribución, abundancia y composición de tallas y estadios de madurez sexual de lIIex argentinus durante las cinco temporadas de verano analizadas en la región patagónica (1992 – 1995). Fuente: extraído de Marcela Ivanovic, Alimentación y relaciones tróficas del calamar Illex argentinus en el ecosistema pesquero, Tesis de Doctorado Universidad de Mar del Plata, 1996. Alimentación del calamar Illex argentinus es un predador oportunista, cuya dieta esta compuesta exclusivamente por organismos pelágicos, es poco diversificada y su composición esta condicionada por la edad del individuo y la disponibilidad de sus presas. Los calamares juveniles habitan profundidades comprendidas entre 50 y 100 m, donde se alimentan principalmente del macro-zooplancton disponible, aunque también consumen calamares y peces pequeños debido a sus pequeñas tallas. Los pre-adultos y adultos se ubican en zonas más profundas, donde son capaces de capturar presas grandes (peces y calamares). Cuando la disponibilidad del macro-zooplancton es suficiente para los requerimientos poblacionales, los ejemplares de todas las tallas se alimentan de dicho macro-zooplancton, resultando la diversidad de la dieta muy baja, especialmente en la zona patagónica. Terra Curanda www.terracuranda.org/fisheries 16 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori Zona Patagónica Las presas pequeñas siempre fueron preponderantes en la dieta durante las cinco temporadas de verano analizadas, tanto por sus frecuencias de aparición como por su contribución al peso total del alimento consumido. Los anfípodos se constituyeron en la presa más importante con frecuencias de ocurrencia que oscilaron entre el 49% en 1994 y el 79% en 1996, seguidos por eufáusidos (6% en 1996 – 39% en 1993). La frecuencia de ocurrencia máxima de calamares y peces en conjunto fue del 15% en 1996. Si bien la contribución de las presas grandes respecto al peso proporcional fue más importante que su frecuencia de ocurrencia, en general osciló entre el 11 y el 14%, y solo alcanzó el 26% en 1994. Merece destacarse que la mayor proporción de anfípodos se observó en 1996 (frecuencia de ocurrencia 79%, peso proporcional 83%) cuando los calamares analizados habían sido capturados en horas de la noche. Durante el otoño las presas pequeñas, principalmente anfípodos, continúan siendo preponderantes tanto en su frecuencia de aparición como en su contribución al peso total del alimento. Los peces solo fueron consumidos, y en muy baja proporción, en 1992, y los calamares alcanzaron una alta frecuencia de ocurrencia en 1994 (24%) debido al consumo de ejemplares muy pequeños por parte de los juveniles analizados ese año. Durante la primavera, la dieta de los juveniles muestreados se compuso casi exclusivamente de anfípodos en 1992 y de eufáusidos en 1997. Peces y calamares muy pequeños solo fueron consumidos en este ultimo año y en conjunto su frecuencia de ocurrencia no superó al 3%. Composición de la dieta con respecto a la profundidad Se analizaron las variaciones del porcentaje de ocurrencia (PO) y del índice de dominancia (ID) de los cuatro ítems presa considerando tres rangos de profundidad: de 70 – 100, de 101 – 200 y de 201 – 400 m. En la zona patagónica durante el verano el mayor el mayor PO siempre correspondió a los anfípodos, con la única excepción del rango de mayor profundidad en 1994. El PO de eufáusidos siempre superó el 25% para profundidades inferiores a los 200 m, pero no fue consumido en todos los casos, así como no se registró el consumo de ellos a profundidades superiores a los 200 m. Aunque el consumo de peces en general fue escaso, los mayores PO se registraron en los individuos capturados a profundidades superiores a los 200 m (1993 y 1994). El consumo de calamar fue bajo y similar entre los distintos rangos de profundidad. Con la única excepción del rango 201 – 400 en 1994, la presa dominante siempre la constituyeron los anfípodos. Para analizar el otoño se dispusieron de ejemplares adultos durante 1992 (sin muestras del rango 201 – 400 m) y juveniles durante 1994. El mayor PO siempre correspondió a los anfípodos, quienes también mostraron los mayores ID. El consumo de eufáusidos fue considerable durante 1992, en particular entre los 101 – 200 m, donde su ID fue ligeramente inferior al de los anfípodos, y fue nulo en 1994. El consumo de peces y calamares fue muy bajo. Durante las dos temporadas de primavera, el alimento excluyente fue el zooplancton: anfípodos en 1992 y eufáusidos en 1997 (Ivanovic, M., 1996). Terra Curanda www.terracuranda.org/fisheries 17 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori Tabla 2: Zona patagónica. Lista de las presas consumidas por lIIex argentinus y número de calamares donde se identificó cada una durante las cinco campañas analizadas para las temporadas de verano. Fuente: extraído de Marcela Ivanovic, Alimentación y relaciones tróficas del calamar Illex argentinus en el ecosistema pesquero, Tesis de Doctorado, Universidad de Mar del Plata, 1996. Consumo del alimento del calamar en el área austral en relación al zooplancton En la tabla siguiente se muestran las relaciones entre el peso del estomago y el peso total del cuerpo de los calamares obtenidas a partir de las campañas de evaluación de pre-reclutas al sur de los 45° LS desarrolladas entre 1993 y 1996. Los pesos totales de los calamares capturados en el área patagónica oscilaron entre 50 y 500 g, con la aparición de algunos ejemplares mas grandes en 1994; en tanto que los pesos de los estómagos en general lo hicieron entre 3 y 20 gr en el caso de la zona bonaerense-nor-patagónica. El rango de pesos totales osciló entre 12 y 730 gr y el de los pesos estomacales entre 0,3 y 30 gr (Ivanovic M, 2000, Tesis de Doctorado U. M. del P). Terra Curanda www.terracuranda.org/fisheries 18 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori Tabla 3: En el 2006 La fauna zoo-planctónica representó el 74% de las presas consumidas, representada en número por un 52% de anfípodos y 22% de eufáusidos. Los calamares representaron el 14% del alimento total en tanto que los peces fueron la categoría menos consumida, alcanzando un 6% del total. Ivanovic M. (2007) Informe de Asesor. Y Trans. INIDEP 31-2006. Fuente: Ivanovic M, 2000 Tesis de Doctorado UNMdP. Informes INIDEP de Asesor. Y Transf. 31/2006, 48/2007; Brunetti N., et al Informes Tecnicos INIDEP: 77-108/1996, 143/1997, 29-147/1998, 5465/99, 93/2000, 86/2001, 87/2002, 103/2003, 70/2004, 50/2005, 53/2006, 61/2007. AÑOS 1993 1994 1995 1996 1997 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007 N RECLUTAS 2,187E+09 0,784E+09 0,629E+09 1,130E+09 1,836E+09 1,352E+09 3,815E+09 2,628E+09 1,299E+09 0,654E+09 0,931E+09 0,047E+09 0,170E+09 2,019E+09 4,215E+09 PROPORCIÓN EN PESO ANFÍPODOS EUFAUSIDOS 0,5136 0,3565 0,4504 0,2961 0,6431 0,2379 0,8121 0,0337 0,52 0,45 FUNCIÓN PARA ESTIMAR LA RACIÓN DIARIA PECES CALAMARES 0,0688 0,0611 0,1567 0,0968 0,0529 0,0661 0,1109 0,0332 PROPORCIÓN EN NÚMERO 0,22 0,06 0,42 0,4 R=-0,6622+0,0482PT R=-1,5571+0,0428PT R=-0,3830+0,0382PT R=-1,4210+0,0327PT 0,14 0,9 Para el año 2007, el 87% del alimento total consumido por el calamar estuvo conformado en número por componentes del zooplancton. Los anfípodos, representados por una única especie, el hipérido Themisto gaudichaudii, contribuyeron en un 45%; en tanto que el 42% restante lo aportaron los eufáusidos. Calamares y peces representaron el 9% y el 4% de las presas, respectivamente (Ivanovic M. (2007) Informe de Asesor. y Trans. INIDEP 48-2007). Si bien los valores de la proporcionalidad son en número, sirven como documentación junto a las opiniones vertidas por el grupo calamar en las reuniones de la Comisión del Calamar, celebradas en el Consejo Federal Pesquero. En ellas expresó que la proporcionalidad de la alimentación por ítem se mantuvo en el curso de los años. Los parámetros para la aplicación del modelo de estimación del consumo utilizados para cada año y área se presentan en la tabla anterior. Las mayores abundancias, al sur de los 45° LS en el período considerado se produjeron en 1993, mientras que durante 1994 y 1995 disminuyeron cuanto menos un 66%, y volvieron a ascender en 1996 un 35% aproximadamente. Las presas mas consumidas fueron los anfípodos, cuyos porcentajes en peso oscilaron entre 45,04% y 81,21%, seguidos por los eufáusidos (con excepción del año 1996), en tanto que peces y calamares fueron los ítems menos frecuentes. La abundancia al norte de los 44º LS en el año 1996 fue muy inferior a la registrada en el sur ese mismo año y comparable a aquellas de 1994 y 1995. La presa mas consumida fue el calamar con un 36,26% del cual el 67% correspondió a I. argentinus y el 33% restante no pudo ser identificado. Anfípodos, peces y eufáusidos fueron consumidos en proporciones similares (19,44% – 24,07%) (Ivanovic M., 2000) Las estimaciones del consumo total por año y el área de los cuatro ítems alimentarios se muestran en las tablas siguientes. En la región patagónica, el consumo total de anfípodos fue de 1.701.871 t en 1993, producto del elevado número de reclutas, mientras que rondó las 260.000 t en 1994 y 1995, alcanzando las 502.900 t en 1996 como resultado de la elevada proporción de este ítem en la dieta durante el año señalado. El consumo de eufáusidos también mostró valores muy dispares en los cuatro años considerados, que oscilaron entre Terra Curanda www.terracuranda.org/fisheries 19 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori 20.869 t (1996) y 1.181.303 t (1993). El consumo de peces fluctuó entre 21.529 y 88.884 t en el periodo 1994-96 y había alcanzado las 227.976 t en 1993. Los calamares también presentaron variaciones menores de su consumo en los tres últimos años (20.559 – 54.907 t) y un valor notablemente superior en 1993 (202.461 t). Tabla 4: Estimaciones del consumo anual de los cuatro principales ítems de la dieta alimentaria del calamar. Fuente: Ivanovic M, 2000 Tesis de Doctorado UNMdP. CONSUMO EN TONELADAS DE LOS CUATRO ITEMS IMPORTANTES ANFÍPODOS EUFAUSIDOS PECES CALAMARES TOTAL 1993 1701871 1181303 227976 202461 3313613 1994 255478 167955 88884 54907 567225 1995 261731 96821 21529 26902 406984 1996 502900 20869 68676 20559 613004 Merece destacarse que de los cuatro ítems alimentarios considerados, solo en el caso de los anfípodos el consumo total puede ser adjudicado a una única especie: Themisto gaudichaudii. Las estimaciones presentadas aquí del consumo de los cuatro tipos de ítems alimentarios que conforman la dieta del calamar correspondieron a la segunda mitad del ciclo de vida anual de la especie, lapso en el cual los individuos se encuentran en sus áreas tróficas y reclutadas a la pesquería. Estos ejemplares se encuentran, aproximadamente, entre los 7 y 12 meses de vida. Lógicamente, por tratarse de una especie anual, el consumo está directamente influenciado por la abundancia anual de las cohortes y por sus tasas de crecimiento, las cuales a su vez estarán condicionadas por la disponibilidad de alimento. Además, la mortalidad por pesca, factor externo a la cohorte, también tiene una gran incidencia en el consumo total producido por ella (Ivanovic M., 2000). Debido a la ausencia de estimaciones de abundancia de las subpoblaciones desde el comienzo del ciclo vital, o al menos desde el estadio juvenil, es imposible estimar el consumo total producido por cada cohorte (Ivanovic M., 2000). Simulación del consumo de calamar A los efectos de poder observar el consumo y su relación con otras especies se decidió simularlo con los datos que figuran en el texto. Esta parte del trabajo es compartida con Aníbal Aubone a los efectos tanto de la consistencia matemática de lo propuesto como de una visión independiente. Tal como se expresara en el texto al existir sobrada documentación (Ivanovic M., Tesis de Doctorado UNMdP, 2000; Informes INIDEP de Asesor. y Transf. 31/2006, 48/2007; Brunetti N. et al, Informes Técnicos INIDEP 77-108/1996, 143/1997, 29-147/1998, 54-65/99, 93/2000, 86/2001, 87/2002, 103/2003, 70/2004, 50/2005, 53/2006, 61/2007; Brunetti, 1988; y Brunetti N, Ivanovic M., 1992) el calamar mantuvo la proporcionalidad de su dieta. A los efectos comparativos se utilizaron todos los datos previamente expuestos para realizar una simulación del consumo para los años que no se contaba con datos de proporcionalidad y pesos de los estómagos. Sí en cambio se contó con datos de peso medio de los individuos, además de datos de las evaluaciones de pre-reclutas obtenidos de Brunetti N et al., Informes Tecnicos INIDEP: 77-108/1996, 143/1997, 29-147/1998, 54-65/99, 93/2000, 86/2001, 87/2002, 103/2003, 70/2004, 50/2005, 53/2006, 61/2007. Terra Curanda www.terracuranda.org/fisheries 20 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori Los valores de consumo totales están expuestos en la tabla siguiente donde no figura el año 1998 debido a una diferencia entre los valores obtenidos por área barrida de ese año y aquellos de las capturas. En cuanto al año 2001, se hicieron dos evaluaciones dada la separación del SSP en dos grupos dados en llamar el del Este y el del Sud oeste, donde también se obtuvieron los datos totales. Los mayores valores se obtuvieron en el 2007 con un consumo total de aproximadamente de más de 4 millones de toneladas, de las cuales 3,7 corresponden a crustáceos pelágicos. Las cifras de calamares se elevan a valores superiores a 200 mil toneladas; una parte importante son de Illex argentinus y le sigue Loligo sp. (Ivanovic M., 2007). El canibalismo se discutirá más adelante, debido a que es un fenómeno de compensación importante, especialmente en el caso que no existan otros alimentos disponibles. Tabla 5: Consumo total del calamar por ítem. Fuente: (1997-2007) Elaboración Aubone Anibal & Prenski L. B. con datos de Ivanovic M, 2000 Tesis de Doctorado UNMdP. Informes INIDEP de Asesor. y Transf. 31/2006, 48/2007; Brunetti N., et al Informes Técnicos INIDEP: 77-108/1996, 143/1997, 29-147/1998, 54-65/99, 93/2000, 86/2001, 87/2002, 103/2003, 70/2004, 50/2005, 53/2006, 61/2007. (1993-1996) Ivanovic M, (2000) 1993 1994 1995 1996 1997 1998 1999 2000 sl 2001 este 2001 tot 2001 2002 2003 2004 2005 2006 2007 Consumo tot del Calamar por item Anf. Euf. Pec. 1701871 1181303 227976 255478 167955 88884 261731 96821 21529 502900 20869 68676 736040 281187 118444 2075212 1551426 415854 287595 703443 303701 288175 117935 161253 1197745 2702452 792787 592687 158868 109869 268734 116022 110091 45054 61603 457571 1032410 333945 249657 66920 46280 113199 48872 46373 18978 25949 192742 434881 Cal. 202461 54907 26902 20559 78253 Total 3313613 567225 406984 613004 1213924 220629 164942 44212 30576 74787 32288 30638 12538 17144 127340 287314 3422573 2558711 685854 425392 1160163 500883 475276 194507 246927 1975398 4390732 En cuanto al ítem peces, en el 2007 aparece como importante la nototenia y luego la merluza. De todas maneras, en el caso de la sobrepesca de la merluza, es importante tener en cuenta cualquier posible mortalidad por predación. Terra Curanda www.terracuranda.org/fisheries 21 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori CONSUMO TOTAL DEL CALAMAR SEMANAL 350000 CONSUMO SEMANAL DEL CALAMAR EN t 2007 2006 300000 2005 2004 250000 2003 2002 200000 2001 2000 150000 1999 1997 100000 1996 1995 50000 1994 1993 0 0 5 10 15 20 25 30 semana Figura 13: Fuente: Elaboración de Aubone Anibal & Prenski L. B. con datos de Ivanovic M, Tesis de Doctorado UNMdP, 2000; Informes INIDEP de Asesor. y Transf. 31/2006, 48/2007; Brunetti N., et al Informes Técnicos INIDEP: 77-108/1996, 143/1997, 29-147/1998, 54-65/99, 93/2000, 86/2001, 87/2002, 103/2003, 70/2004, 50/2005, 53/2006, 61/2007. El gráfico precedente muestra la disminución del consumo total semanal a medida que la cohorte se extingue y al momento de su desove. Tal como se expresó, el consumo que aquí se muestra es parcial ya que se cubre la cohorte en la mayoría de los casos en la semana 8, cuando el calamar ya tiene 5-7 meses de vida; salvo en algunos años en donde se dispuso de una serie mas larga de datos. La simulación trae otras posibles formas de ver la alimentación del calamar especialmente el zooplancton y su relación directa con lo fenómenos ambientales. Si se toma el consumo individual promedio semanal, el calamar, al ser un animal de ciclo anual, tiene un crecimiento rápido debido a una transformación eficiente de su alimento. Se puede observar que el consumo en toneladas de una semana es en promedio un 30% de la biomasa en toneladas existente al comienzo de dicha semana, y esto para todos los años. Terra Curanda www.terracuranda.org/fisheries 22 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori CONSUMO/EJEMPLAR DE CALAMAR POR SEMANA 1997 0,0003 1999 2000 Consumo individual semanal en t 0,00025 2001este 2001sudoeste 0,0002 2002 2003 0,00015 2004 2005 0,0001 2006 2007 0,00005 1993 1994 0 0 5 10 15 20 Semanas 25 30 1995 1996 Figura 14: Consumos semanales que aumentan en forma directa a medida que el calamar aumenta de peso Fuente: Elaboración Aubone Anibal & Prenski L. B. con datos de Ivanovic M, 2000 Tesis de Doctorado UNMdP. Informes INIDEP de Asesor. y Transf. 31/2006, 48/2007; Brunetti N., et al Informes Tecnicos INIDEP: 77-108/1996, 143/1997, 29-147/1998, 54-65/99, 93/2000, 86/2001, 87/2002, 103/2003, 70/2004, 50/2005, 53/2006, 61/2007,15/2009. En el gráfico se observan dichos consumos semanales que aumentan en forma directa a medida que el calamar aumenta de peso. La conexión es clara ya que el calamar aumenta de peso en la medida que hay alimento disponible y se puede ver en el gráfico que casi todos los años presentan relaciones casi lineales que se acercan o alejan del eje “y” pero en cuatro años 1996, 2003, 2004 y 2005 el comportamiento es diferente. Lo descriptivo del modelo es que el aumento de peso es un dato real obtenido de los informes mencionados. Esto, como se expresó con anterioridad, está en directa relación con el alimento, y en este caso con el zooplancton que es su alimento más importante; por ello el aumento de peso está relacionado con su concentración. El año 1996 presenta dos situaciones diferentes, la primera a. un nivel bajo del consumo individual promedio que solo se asocia con los años expuestos en el gráfico siguiente b. un corte que parece pasar de un estado bajo a otro menor como si se hubiera extinguido una determinada cantidad de alimento de un lugar provocando una disminución del consumo individual promedio, para luego reiniciar el consumo desde otro nivel, por el probable nuevo encuentro con otra concentración del mismo. Comparando las cohortes del año 1996 de ambas áreas Austral y nor-patagónica, se observa que el crecimiento en peso fue similar, la abundancia y el consumo de la SSP superaron en aproximadamente un 40% a los de la SBNP pero la mortalidad por pesca fue muy superior para la subpoblación del sur, lo cual indicaría que finalmente el consumo total no debería haber arrojado diferencias tan notables. Ellas se debieron a las diferencias en la ración diaria media de ambas subpoblaciones, ya que la estimada para la SSP supero en aproximadamente un 30% a la de la SBNP. El hecho de que los calamares de la SBNP, con una dieta compuesta Terra Curanda www.terracuranda.org/fisheries 23 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori principalmente por calamares y peces, incorporaron un 30% menos de alimento en peso que aquellos de la SSP (dieta basada en anfípodos) y alcanzaron tasas de crecimiento en peso similares, podría considerarse bajo la hipótesis de Dawe (1988), plantea que la eficiencia de la dieta basada en presas grandes es mayor porque la demanda de energía asociada a la captura es menor que aquella asociada a la captura de presas pequeñas (crustáceos). En este caso el 36% de la dieta estuvo conformado por calamares y el 66% de ellos fueron juveniles de I. argentinus, lo cual también apoya la hipótesis de Caddy (1983) sobre que el canibalismo produce una ganancia metabólica mayor en los ejemplares mas grandes al predar sobre los mas pequeños y aumenta la resistencia general de la población total frente a condiciones de escasez de alimento. Ivanovic M, 2000 Tesis de Doctorado UNMdP. Para el año 2003 el corte es más evidente y parecería que no encuentra otra nueva concentración. Para los años 2004 y 2005 parece que en todo el desarrollo de esas cohortes el zooplancton es escaso. Es importante mencionar que los años mencionados fueron de cohortes de un reclutamiento muy bajo del SSP (Stock Sud patagónico). CONSUMO/EJEMPLAR DE CALAMAR POR SEMANA Años con aparente deficiencia alimentaria Consumo individual semanal en t 0,00025 0,0002 0,00015 0,0001 0,00005 0 0 5 10 15 20 25 30 Semanas 2003 2004 2005 1996 Figura 15: Consumo individual semanal en toneladas del calamar. Fuente: Elaboración Aubone Anibal & Prenski L. B. con datos de Ivanovic M, 2000 Tesis de Doctorado UNMdP. Informes INIDEP de Asesor. y Transf. 31/2006, 48/2007; Brunetti N., et al Informes Técnicos INIDEP: 77-108/1996, 143/1997, 29-147/1998, 54-65/99, 93/2000, 86/2001, 87/2002, 103/2003, 70/2004, 50/2005, 53/2006, 61/2007,15/2009. Densidad de Zooplancton y el reclutamiento del calamar y otras especies y la intensidad pesquera. Comparativamente para poder tener alguna relación con la concentración de Zooplancton como una medida de poder predecir en algo de la relacion expuesta, existen los datos de Sabatini M.(2003) Inf Tec. INIDEP 124/2003; Sabatini, Reta R., Matano/ Continental Shelf Research 24 (2004) 1359–1373; Sabatini M., Shimabokuro V. (2005), Inf Tec INIDEP 5/2005; Sabatini M. Alvarez Colombo (2006) Inf Ases. y Trans INIDEP 36/2006; Sabatini M. Alvarez Colombo (2007) Inf Ases. y Trans INIDEP 51/2007. Ana Fazio, Paola Scarcia (2003) UNBA Terra Curanda www.terracuranda.org/fisheries 24 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori (seminario Oceano Biol.) contrib INIDEP. Esto permite construir una serie de densidades de biomasa cronológicas para la Plataforma Austral. Esta serie es un aporte importante, que no permite tener a futuro una mirada mas específica sobre determinadas situaciones que ofenden convertise en hipótesis de trabajo. (ver, en zooplancton, relaciones tróficas, y relación con la circulación de las masas de agua pg 49). Lo primero que se visualiza en el gráfico es la variación de la densidad media de la biomasa de zooplancton a través de los distintos años con una tendencia decreciente, ajustándole una función exponencial. De los valores de reclutamiento de calamar obtenidos de los mencionados autores, también pueden observarse tres épocas, una inicial de equilibrio en las etapas híncales de la pesquería austral, una intermedia de desarrollo muy intensivo de la pesquería del calamar a través del desarrollo de una biomasa explosiva y otra de variaciones. Podemos interpretar este grafico, al cual luego le anexaremos otros adjuntando otras especies, se puede conjeturar que la abundancia de zooplancton inicial causada por la disminución de las biomasas por pesca de otros predadores sobre el mencionado zooplancton sumada a una disminución de la predación sobre el calamar, han permitido el desarrollo de una gran biomasa y por lo tanto una gran pesquería. 5000,0 4500 4500,0 4000 Reclut de Calamar 4000,0 3500 3500,0 3000 3000,0 2500 2500,0 2000 2000,0 1500 1500,0 1000 1000,0 Biomasa media de Zooplancto PH mg por m3 Relación entre la Densidad media de la Biomasa del Zooplancton y el Reclutamiento del Calamar 500 500,0 0 19 78 19 79 19 80 19 81 19 82 19 83 19 84 19 85 19 86 19 87 19 88 19 89 19 90 19 91 19 92 19 93 19 94 19 95 19 96 19 97 19 98 19 99 20 00 20 01 20 02 20 03 20 04 20 05 20 06 20 07 20 08 20 09 0,0 Anios Recl Cal SSP Den media de Biom Zoop Exponencial (Den media de Biom Zoop) Figura 16: Fuente: Elaboración propia con datos de, Brunetti N., et al Informes Tecnicos INIDEP: 77-108/1996, 143/1997, 29-147/1998, 54-65/99, 93/2000, 86/2001, 87/2002, 103/2003, 70/2004, 50/2005, 53/2006, 61/2007, 52/2008, 15/2009 Sabatini M.(2003) Inf Tec. INIDEP 124/2003; Sabatini, Reta R., Matano/ Continental Shelf Research 24 (2004) 1359–1373; Sabatini M., Shimabokuro V. (2005), Inf Tec INIDEP 5/2005; Sabatini M. Alvarez Colombo (2006) Inf Ases. y Trans INIDEP 36/2006; Sabatini M. Alvarez Colombo (2007) Inf Ases. y Trans INIDEP 51/2007. Ana Fazio, Paola Scarcia (2003) UNBA (seminario Oceano Biol.) contrib INIDEP. Las mencionada etapa intermedia de crecimiento, se convierte en una situación más permanente pero mas proclive a variaciones importantes de su biomasa producto de la desmedida intensidad pesquera del área adyacente y los cambios ambientales que controlan la bio-productividad. Si bien por el momento parecería que el calamar tiene suficiente alimento para el desarrollo explosivo que viene demostrando, si existe un quiebre en el reclutamiento por la sobrepesca como parece haber ocurrido en los años 2004 y 2005, puede ocurrir que su Terra Curanda www.terracuranda.org/fisheries 25 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori lugar puede ser ocupado por otro como los gelatinosos. También puede ocurrir que la gran presión trófica del calamar no permita la recuperación de otras especies. Para tener una dimención por lo menos de captura de las tres especies australes de mayor desarrollo de su biomasa, Polaca, Merluza de cola y Calamar podemos decir que en algunos años se llega a valores de casi 1,2 millones de toneladas, si a esto le sumamos la merluza común, el bacalao austral, abadejo, savorín, la cabrilla etc capturadas al sur de 45° dicha cantidad se incrementa. En el siguiente gráfico se ubican las cantidades de las tres especies importantes, la caída del 2003 al 2005 esta esencialmente basada en la caída del calamar. Captura de Polaca, Calamar y Merluza de cola área austral 1400000 1200000 Captura en t 1000000 800000 600000 400000 200000 19 78 19 79 19 80 19 81 19 82 19 83 19 84 19 85 19 86 19 87 19 88 19 89 19 90 19 91 19 92 19 93 19 94 19 95 19 96 19 97 19 98 19 99 20 00 20 01 20 02 20 03 20 04 20 05 20 06 20 07 20 08 0 Años CAP CALAMAR ZONA AUSTRAL Cap Polaca Cap Merl. Cola. Captura Total Figura 17: Fuente: elaboración propia con datos de la Subsecretaría de Pesca y Acuicultura y la de otras flotas del Fishery Statistical Bulletin Falkland Island, datos corregidos Giussi A. et al, Informe técnico INIDEP 28 y 49/2007, la captura de la zona no regulada también para el área Sur de CAPeCA relación intercámara. En el gráfico siguiente se amplió la observación sobre otras dos especies, presentándose los reclutamientos de la polaca y la merluza de cola, en cuanto a la merluza de cola se debe aclarar que la presencia en nuestra plataforma de individuas mas pequeños tiene dos años de vida. No existe ningún trabajo sobre el reclutamiento de la clase de edad 2 obtenido de buques de investigación y en los informes técnicos del INIDEP del Programa de especies australes sobre la evaluación de merluza de cola y polaca no figuran los datos de salida del programa en biomasa en número por edad, estos no solo son importante como dato, sino que el trabajo debe presentarse de tal manera que se pueda realiza un trabajo de auditoría independiente. Terra Curanda www.terracuranda.org/fisheries 26 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori Reclutamiento de Calamar y Merluza de cola edad 1 Biomasa de Merluza de cola y Densidad de Zooplancton 5000 900 4500 800 4000 Reclutamiento 10/6 600 3000 500 2500 400 2000 300 1500 Densidad de Zooplancton 700 3500 200 1000 100 500 0 19 78 19 79 19 80 19 81 19 82 19 83 19 84 19 85 19 86 19 87 19 88 19 89 19 90 19 91 19 92 19 93 19 94 19 95 19 96 19 97 19 98 19 99 20 00 20 01 20 02 20 03 20 04 20 05 20 06 20 07 20 08 20 09 0 Anios MC Recl edad 1 Recl Cal SSP Bio Merl Cola en miles t Zoop Den Media Figura 18: Fuente: elaboración propia con datos obtenidos de, Giussi A. et al, Informe técnico INIDEP 28 y 49/2007, Brunetti N., et al Informes Tecnicos INIDEP: 77-108/1996, 143/1997, 29-147/1998, 54-65/99, 93/2000, 86/2001, 87/2002, 103/2003, 70/2004, 50/2005, 53/2006, 61/2007, 52/2008, 15/2009 Sabatini M.(2003) Inf Tec. INIDEP 124/2003; Sabatini, Reta R., Matano/ Continental Shelf Research 24 (2004) 1359–1373; Sabatini M., Shimabokuro V. (2005), Inf Tec INIDEP 5/2005; Sabatini M. Alvarez Colombo (2006) Inf Ases. y Trans INIDEP 36/2006; Sabatini M. Alvarez Colombo (2007) Inf Ases. y Trans INIDEP 51/2007. Ana Fazio, Paola Scarcia (2003) UNBA (seminario Oceano Biol.) contrib INIDEP. Canibalismo como mecanismo compensador e índice de deficiencia en la alimentación El tema canibalismo será otra de las relaciones que es conveniente tener en cuenta, especialmente como mecanismo compensador ya que si los tamaños grandes no tiene acceso al zooplancton de mayor tamaño pueden alimentarse de individuos mas pequeños de calamar, que puedan tener acceso a alimento de tamaños menores y por lo tanto asegura la continuidad del reclutamiento. De Ivanovic M, 2000 Tesis de Doctorado UNMdP. “La existencia de canibalismo fue registrada en las temporadas de otoño, invierno y primavera en la región bonaerense-nor-patagonica y solo en las temporadas de verano en la región patagónica. En la zona norte el canibalismo parece ser una constante, ya que se detectó en todas las campañas de investigación analizadas en estas tres temporadas, con la excepción de dos campañas de invierno de las que solo se dispuso de juveniles para el muestreo (EH-08/94 y EH-10/97). En el sur, en cambio, cinco campañas de investigación de verano fueron analizadas, y solo se detectó en tres de ellas. El canibalismo fue practicado en la región bonaerense-nor-patagonica por los calamares desde 30 a 350 mm LM, mientras que en la zona patagónica solo se registró en adultos (151-350 mm LM). En ambas zonas los calamares consumidos fueron juveniles de hasta 140 mm LM. El Terra Curanda www.terracuranda.org/fisheries 27 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori consumo de adultos, por el contrario, parece ser ocasional, ya que fue detectado en muy pocas oportunidades (Ivanovic M, 2000 Tesis de Doctorado UNMdP). Los mayores porcentajes de canibalismo se observaron en la zona norte, donde oscilaron entre 5,6% y 15,5% en adultos (151-350 mm LM) y entre 2 y 42% en juveniles (30-150 mm LM); mientras que en la zona sur nunca superaron el 4%. La proporción representada por I. argentinus del total de los calamares consumidos oscilo entre el 39 y el 100% en el norte y 2369% en el sur. Cabe destacar que valores del 100% se registraron cuando el canibalismo fue practicado exclusivamente por calamares juveniles (3-14 cm LM). En el caso de la campada OB-04/96, se detectó consumo de calamares en los ejemplares con tallas entre 60 y 290 mm LM, pero las especies consumidas solo pudieron ser identificadas en el 66,7% de los casos, y la totalidad de ellos correspondió a canibalismo practicado por ejemplares con tallas entre 90 y 150 mm LM (Ivanovic M, 2000, Tesis de Doctorado UNMdP). Variaciones notables fueron observadas entre años para una misma temporada en la región bonaerense-nor-patagónica. Mientras que durante el otoño de 1991 se detectó canibalismo en los ejemplares con tallas superiores a los 200 mm LM y no existió entre los más pequeños; todo el canibalismo que pudo ser detectado en el año 1996 correspondió a los calamares juveniles (90-150 mm LM). Durante 1998 solo se dispuso de juveniles y se observó un porcentaje de canibalismo mucho mayor. También hubo diferencias entre los juveniles analizados durante las temporadas de invierno, ya que se detectó canibalismo entre ellos en el año 1989 pero no hubo indicios de el mismo en las campañas realizadas en 1994, 1996 y 1997. En cuanto a las temporadas de primavera, no se detectó canibalismo entre los juveniles en 1992 pero si existió en 1997, aunque su porcentaje fue muy bajo ya que apenas alcanzó el 2%. Cuando se comparan los porcentajes de canibalismo con las abundancias de calamar no se observa ningún patrón consistente. En la región bonaerense-nor-patagónica, durante el otoño, en un año de alta abundancia como 1991, se registró un porcentaje relativamente bajo (5,6%) de canibalismo entre adultos. En 1996, en condiciones de mayores abundancias que en 1998, el porcentaje de canibalismo fue menor entre los juveniles. Durante las temporadas de invierno, los porcentajes de canibalismo fueron similares en 1991 y 1996, pero las abundancias fueron considerablemente menores en el último año. Finalmente, en primavera, frente a abundancias similares en 1992 y 1997, la proporción de canibalismo fue muy superior en el año 1992. Finalmente, en la región patagónica, las abundancias de calamar y los porcentajes de canibalismo fueron similares en los tres años investigados (Ivanovic M, 2000 Tesis de Doctorado UNMdP). Los porcentajes de canibalismo (2,2%–15,5%) detectados entre adultos de calamar argentino, considerando toda el área de distribución, son muy inferiores a los determinados entre adultos de I. illecebrosus (30%-50%; O’Dor, 1983), pero comparables a los de Ommastrephes bartrami (14%; Lipinski y Likowski, 1988), Todarodes sagittatus (4,76%; Quetglas et al., 1999), Nototodarus gouldi (3,5-19,2%; O’Sullivan y Cullem 1983) y N. sloanii (20%, Uozumi, 1998). Por otra parte, en Sthenoteuthis oualaniensis, el canibalismo mostró un patrón similar al detectado en I.argentinus, ya que el porcentaje correspondiente a los calamares juveniles (19,4%; 4,3-10,5 cm LM) fue superior al doble del estimado para los ejemplares mas grandes (8,7%; 12,4-36,5 cm LM; Shchetinnikov, 1992). El canibalismo es un fenómeno que ha sido observado, al menos, en cuatro familias de cefalópodos: Sepiidae, Loliginidae, Gonatidae y Ommastrephidae (Nixon, 1987), pero su interpretación todavía esta sujeta a controversia. Hixon (1983) observó un aumento en los porcentajes de canibalismo en las áreas de desove de Loligo opalescens y lo adjudicó a una Terra Curanda www.terracuranda.org/fisheries 28 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori respuesta de los calamares frente a condiciones de alta densidad de individuos. Según Caddy (1983), el canibalismo en los calamares esta favorecido por la capacidad de estos animales de producir varias cohortes intra-anuales, originando la coexistencia de individuos de distintas tallas, donde los más grandes, en condiciones de ausencia de alimento, son capaces de predar sobre los más pequeños. Así, la fracción de población compuesta por los animales mas grandes recibe una ganancia metabólica mayor al predar sobre ejemplares pequeños, consumidores a su vez de zooplancton, lo cual aumenta la resistencia general de la población total frente a la disminución del alimento habitual. Este análisis se basa en el comportamiento alimentario de Illex illecebrosus, donde siempre se ha verificado un cambio en los componentes de la dieta de los calamares juveniles, consumidores de zooplancton, y los adultos, que predan principalmente sobre peces. Por su parte, Ehrhardt et al. (1983) atribuyeron el canibalismo observado en Dosidicus gigas a una disminución de la disponibilidad de otras presas, en tanto que Nesis (1987) puntualizó que este fenómeno no parece ser un mecanismo de control de la densidad poblacional, especialmente porque los calamares se agrupan en cardúmenes conformados por animales de las mismas tallas (Ivanovic M, 2000, Tesis de Doctorado UNMdP). En I. illecebrosus, el porcentaje de canibalismo aumenta entre los calamares mas grandes (Amaratunga, 1980) posiblemente, y al igual que en D. gigas, como reflejo de la disminución de la abundancia de otras presas, especialmente peces (Dawe y Brodziak, 1998). Dawe (1988) encontró además que, en los años de altas abundancias de calamar, el porcentaje de canibalismo aumentó considerablemente al tiempo que declinó la proporción de peces en la dieta, y lo relacionó con una abundancia insuficiente de estos últimos para sostener las demandas del calamar. Los porcentajes de canibalismo estimados para los calamares distribuidos en el área patagónica son muy bajos; en cambio, ellos son mayores en el calamar presente en la zona bonaerense-nor-patagónica. Estos resultados concuerdan con los reportados por Koronkiewcz (1986), quien también detectó bajos porcentajes de canibalismo entre lo adultos de la región patagónica, y por Santos y Haimovici (1997), quienes estimaron en 7,71% el porcentaje de canibalismo practicado por ejemplares con tallas superiores a los 100 mm LM sobre juveniles (17-131 mm LM) en la región sur de Brasil. Este porcentaje podría ser mayor, ya que el consumo de calamares no identificados alcanzo un 15,98%, en tanto que del 11,57% identificado, el 67% correspondió a I. argentinus. En la zona bonaerense-nor-patagónica, los calamares de todas las tallas practicaron canibalismo, en cambio, en la región patagónica esto solo se observo entre los adultos. De todos modos, la mayor proporción del canibalismo detectado en el calamar argentino se produjo sobre juveniles, y solo ocasionalmente se encontraron ejemplares de tallas grandes en los contenidos estomacales. Este hecho contrasta con la observación de O’Dor et al. (1980), sobre I. illecebrosus, de que el dimorfismo sexual en el crecimiento facilita la prelación sobre los machos adultos, por poseer tallas mas pequeñas que las hembras. En I. argentinus, donde el canibalismo no solo es producido por los adultos, sino también por los juveniles; existen diferencias entre las dos áreas de distribución del calamar, no se observaron relaciones entre la magnitud del consumo y la abundancia poblacional y se registraron variaciones entre años para una misma temporada, parece cumplirse el postulado de Maurer y Bowman (1985) que establece que “la disponibilidad de presas alternativas podría ser el mecanismo primario que controla el canibalismo en los calamares del genero Illex. Este sería un indicio de que el ecosistema bonaerense-nor-patagónico tendría una capacidad de soporte del calamar menor que el ecosistema patagónico. Santos y Haimovici (1997) también reportaron que las altas tasas de canibalismo observadas en el sur de Brasil Terra Curanda www.terracuranda.org/fisheries 29 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori son un reflejo de la limitada disponibilidad de alimento para el calamar en la región. En la merluza común también se observaron porcentajes de canibalismo más altos entre los ejemplares distribuidos en el ecosistema bonaerense-nor-patagónico, especialmente los juveniles que se encuentran sobre la plataforma a menos de 100 m de profundidad, como fenómeno compensador de la falta de alimento (Sánchez y García de la Rosa, 1999; Ivanovic M, 2000, Tesis de Doctorado UNMdP). Lo observado por Ivanovic M (2000, Tesis de Doctorado UNMdP) en el texto desarrollado con anterioridad, comparativamente también en diferentes especies de calamares y merluza común, son fenómenos de denso-dependencia por puro oportunismo o por falta de alimento. En general el momento de reproducción marca diferencias. Desde un punto de vista estrictamente energético y de estrategia reproductiva la existencia en el área de desove o en las cercanías de distribuciones de juveniles hace que la ecuación de fecundidad se rija por el canibalismo como efecto estabilizador de la reproducción introduciendo energía en el asegurar la nueva generación. Esto es evidentemente notorio en peces los cuales tienen un ciclo de vida de varios años y las distribuciones de tamaño son de diferencia notable entre juveniles y adultos (Ivanovic M, 2000, Tesis de Doctorado UNMdP). En el caso del Illex argentinus si bien evidencia los expresado por Caddy (1983) por la existencia de los cuatro stocks con tamaños diferentes. También es importante lo expresado por otros autores Santos y Haimovici (1997) en cuanto a diferente capacidad de soporte entre el área patagónica y la bonaerense. Pero creemos que la intensa captura de otras especies ha permitido una capacidad de soporte mayor y por lo tanto no han desarrollado un canibalismo mas intenso. Desde el punto de vista de la observación los años críticos podrían haber sido los anteriores del inicio de la pesquería y de los cuales no hay muestras tróficas. Condiciones del manejo del calamar y su repercusión en el ecosistema pesquero Medioambiente y reclutamiento de calamar Muchos autores han tratado de encontrar relaciones entre el reclutamiento y distribución del calamar y las variables ambientales. Entre ellos cabe citar: C. M. Waluda, P. G. Rodhouse, P. N. Trathan & G. J. Pierce (2001), Fish Oceanography 10:2 pg 207-216. C. M. Waluda, P. N. Trathan & P. G. Rodhouse 2004, Fish Ocenography 13:4 pg 255-266; Gastón Bazzino , Renato A. Quiñones , Walter Norbis (2005) Fisheries Research 76 401–416 ; G. Vanyushin, T. Barkanova (2005) Geophysical Research Abstracts, Vol. 7, 04442;Chih-Shin Chen, Wen-Bin Haung, and TaiSheng Chiu, 2007 Zoological Studies 46(3): 362-374. Tomando en cuenta este valioso aporte es evidente que si le anexamos lo expuesto en circulación marina y fenómenos biológicos asociados que nos muestra la complejidad de la circulación y si nos remitimos nuevamente de los crustáceos pelágicos ya que son los que siguen el ciclo de bio-producción con mayor rapidez y consideramos que los predadores siguen a las presas, una relación simple con alguna de las variables ambientales es demasiado vulnerable a una mala interpretación por lo expresado. En el texto expuesto a continuación queremos mostrar otro tipo de situaciones que puedan influir en el reclutamiento en forma directa. Terra Curanda www.terracuranda.org/fisheries 30 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori Transporte de la Corriente de Malvinas y reclutamiento En virtud de la importancia económica y biológica de poder predecir el éxito del reclutamiento del calamar con su consecuencia inmediata en los valores de abundancia. El gráfico que se presenta a continuación se expresa la relación entre trasporte de la corriente de Malvinas y éxito de reclutamiento al año siguiente. El corrimiento realizado en los valores de CPUE es debido a que la intensidad del trasporte como acción directa se percibe su efecto año siguiente. Relación entre los valores de CPUE, Taiwan retrasados un año y datos de Geostrofía de Malvinas Fedulov (1990) 25 12 CPUE Taiwan t/dia 10 1999 8 15 1984 1985 2000 1986 19851987 1997 1983 1992 1994 1996 2001 2002 1991 1986 1987 5 2003 1988 1989 1988 10 6 1998 1993 4 1995 Geostrofia de Malvinas Sv 1982 20 1990 2 0 19 78 19 79 19 80 19 81 19 82 19 83 19 84 19 85 19 86 19 87 19 88 19 89 19 90 19 91 19 92 19 93 19 94 19 95 19 96 19 97 19 98 19 99 20 00 20 01 20 02 20 03 20 04 20 05 20 06 20 07 0 Años Taiwan cpue Corrido 1 año Geostrofia Sv Figura 19: Fuente elaboración propia datos de P.P. Fedúlov, A. V. Remesló, S.N. Burykin y J.A. Pulishchuk 1990 FRENTE MARITIMO Vol. 6, Seco A: 121-127 (1990) y Chih-Shin Chen, Wen-Bin Haung, and Tai-Sheng Chiu, 2007 Zoological Studies 46(3): 362-374. Siguiendo con el análisis propuesto en el gráfico anterior se presenta en el siguiente una nueva elaboración a la cual se le agregaron los valores de trasporte de la Corriente de Malvinas obtenidos del trabajo de Vivier y Provost (1999). Este tiene el mismo carácter exploratorio de variables ambientales que puedan influir en el buen desarrollo del reclutamiento de calamar. Sin embargo existen dos situaciones diferentes con el gráfico anterior y son comparativamente la medición diferente de la Geostrofía y el uso de los valores iniciales del reclutamiento, tanto por retro-cálculo como por campaña de investigación. Considerando la velocidad de la Corriente de Malvinas (trasporte) podemos afirmar que según las velocidades un huevo de calamar del SSP puede trasladarse a las áreas de desarrollo en un mínimo de 40 días aproximadamente. Los datos de trasporte de la corriente son diferenciales según la hipótesis de que años de buen trasporte aseguran el contacto con la Corriente de Brasil y lo contrario sería opuesta. Los valores obtenidos son los expuestos en el gráfico y corresponden a un promedio de seis meses comenzando en abril. Como se puede ver, éstos oscilan entre 30 y 40 Sverdrub. Terra Curanda www.terracuranda.org/fisheries 31 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori En referencia a los corrimientos expresados en el gráfico son el alineamiento de padres e hijos y los valores de trasporte los que determinan el movimiento de los huevos. Calamar SSP, Padres y Transporte de la Corriente de Malvinas correspondientes al año anterior y sus respectivos reclutas 5000 45 4500 40 4000 35 3500 25 2500 20 2000 Transporte Sv Numero 10/6 30 3000 15 1500 10 1000 5 500 0 0 1992 1993 1994 1995 1996 1997 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 Años SSP Numero de Reclutas 10/6 Biom Des en N corr 1 año Transp. Malv en Sv año ant. abril-sep Figura 20: Fuente: elaboración propia en base a datos obtenidos de Brunetti N., Inf. Tecnico INIDEP: 77-108/1996, 143/1997, 29-147/1998, 54-65/99, 65/2000, 86/2001, 87/2002, 103/2003, 70/2004, 50/2005, 53/2006, 61/2007; Vivier y Provost 1999. Si bien en ambos hay un acompañamiento de las curvas de CPUE, reclutamiento y su asociación con la geostrofía, creemos en virtud de lo aquí apreciado que es conveniente continuar con programas internacionales tendientes a poder integrar la dinámica de las corrientes de Malvinas y Brasil y su enorme efecto en el Atlántico Sur-occidental. De acuerdo a lo expresado sería muy conveniente trabajar con una serie mas completa de datos de trasporte y también de datos de desarrollo de fitoplancton para los momentos de desarrollo de las larvas. Los procesos de bio-producción se condicionan por la presencia de hierro en el medio y su dinámica se relaciona con la fuerza de la Corriente de Malvinas Carreto (2005). Los análisis previos no permiten rechazar la hipótesis de que las variaciones de año en año en el reclutamiento anual están reguladas por la operación compensatoria de la densidad parental en la forma de una relación stock-reclutamiento, mas aún de acuerdo a los gráficos se determina una relación de dependencia al stock desovante hasta una determinada biomasa en número que alcanza aproximadamente la suma de 100 millones de individuos. Sin embargo, el análisis revela que a partir de la cifra mencionada los factores ambientales como el transporte de la Corriente de Malvinas determinan la importancia del acarreo de los huevos durante o justo después de la condición crítica, así como el tiempo de llegada y el encuentro de mayor extensión de hábitats propicios para un oportuno desarrollo del ciclo de vida como factor clave en la determinación de las tasas de supervivencia. Faltan observaciones de reclutamiento a escala de su ubicación crítica, determinadas por el área de hábitat espacial idóneo para que los juveniles, durante o justo después del encuentro Terra Curanda www.terracuranda.org/fisheries 32 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori del mencionado hábitat idóneo, y por las condiciones del transporte de la corriente de Malvinas. La gran escala geográfica de este estudio enfatiza la importancia de los mismos factores ambientales como determinantes del tamaño de población a escalas espacial y temporal. La Corriente Circumpolar Antártica, como factor que actúa en el hemisferio, opera sobre las condiciones de caudal base sobre toda la cuenca de la corriente de Malvinas para determinar un transporte específico cada año. A partir de marzo el tipo de transporte es importante para el calamar en el acarreo de los huevos al norte. Este transporte determina el encuentro con la cantidad de hábitats idóneos para la llegada de huevos y el desarrollo de larvas, el cual es específico de la sección de corriente que se encuentra con la Corriente de Brasil y su grado de mezcla o de encuentro, determinando la cantidad de reclutas en cada sección. Consideramos que el efecto del transporte actúa en un grado relevante sobre la calidad del reclutamiento. Mientras se acumula evidencia de la importancia de factores climáticos para la dinámica de poblaciones marinas (Glantz 1992, Cushing 1996), determinantes ambientales en poblaciones de la corriente de Malvinas y otros sistemas marinos permanecen elusivos. Esto puede no ser sorprendente dada la falta de estudios a largo plazo de longitud suficiente como para capturar años favorables y no favorables, lo que parece un requisito esencial para detectar efectos ambientales capaces de producir cambios en el tamaño de población. Claramente, se hacen necesarios estudios a largo plazo para dilucidar el papel que juegan los factores ambientales en la determinación del tamaño de población. No es menos importante la asociación con la intensidad pesquera del calamar en el área no regulada y el efecto de la disminución de los predadores de esta especie como la merluza común. 1993 1400 1994 1995 1996 1997 1998 1999 2000 2001 2002 200,0 1200 150,0 1000 100,0 50,0 600 0,0 400 -50,0 200 -100,0 0 -150,0 15/ 06/ 1905 800 Reclutamiento de Calamar Residuales de la altura del mar Figura 21: Residuales de la altura del mar en el área de desove del calamar y su relación con el reclutamiento 48,3049 LS Y 60,30-59 LO 1993-2002 Terra Curanda www.terracuranda.org/fisheries 33 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori RELACIÓN CON EL NIÑO MODOKI Y EL NIÑO (3-4) CON SU EFECTO EN RETARDO EN EL ATSM ANOMALÍA DE TEMPERATURA DE SUPERFICIE DEL MAR EN EL ÁREA DE DESOVE DEL CALAMAR ENERO 2003 - JUNIO 2010 2,5 2 1,5 1 0,5 abr-10 ene-10 jul-09 oct-09 abr-09 ene-09 jul-08 oct-08 abr-08 ene-08 jul-07 oct-07 abr-07 ene-07 jul-06 oct-06 abr-06 ene-06 jul-05 oct-05 abr-05 ene-05 jul-04 oct-04 abr-04 ene-04 jul-03 oct-03 abr-03 -0,5 ene-03 0 -1 -1,5 -2 NIÑO MODOKI NIÑO (3-4) Figura 22: El calamar y su relación con las situaciones oceanográficas globales. Relación con el Niño Modoki y el Niño (3-4) con su efecto en retardo en el ATSM. Anomalía de temperatura de superficie del mar en el área de desove del calamar: Enero 2003-Junio 2010. ANOMALÍA DE TEMPERATURA DE SUPERFICIE DEL MAR EN EL ÁREA DE DESOVE DEL CALAMAR ENERO 1982 - DICIEMBRE 2002 2 1,5 1 0,5 jul-02 ene-02 jul-01 jul-00 ene-01 ene-00 jul-99 jul-98 ene-99 ene-98 jul-97 jul-96 ene-97 jul-95 ene-96 jul-94 ene-95 ene-94 jul-93 jul-92 ene-93 ene-92 jul-91 jul-90 ene-91 ene-90 jul-89 ene-89 jul-88 jul-87 ene-88 ene-87 jul-86 ene-86 jul-85 jul-84 ene-85 ene-84 jul-83 ene-83 jul-82 ene-82 0 -0,5 -1 -1,5 ATSM Lineal (ATSM) Figura 23: El calamar y su relación con las situaciones oceanográficas globales. Anomalía de temperatura de superficie del mar en el área de desove del calamar: Enero 1982-Diciembre 2002 Terra Curanda www.terracuranda.org/fisheries 34 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori ANOMALÍA DE TEMPERATURA DE SUPERFICIE DEL MAR EN EL ÁREA DE DESOVE DEL CALAMAR ENERO 2003 - JUNIO 2010 2,5 2 1,5 1 0,5 abr-10 ene-10 jul-09 oct-09 abr-09 ene-09 jul-08 oct-08 abr-08 oct-07 ene-08 jul-07 abr-07 ene-07 jul-06 oct-06 abr-06 ene-06 jul-05 oct-05 abr-05 ene-05 jul-04 oct-04 abr-04 ene-04 jul-03 oct-03 abr-03 -0,5 ene-03 0 -1 -1,5 -2 ATSM Lineal (ATSM) Figura 24: El calamar y su relación con las situaciones oceanográficas globales. Anomalía de temperatura de superficie del mar en el área de desove del calamar: Enero 2003-Junio 2010 ANOMALÍA DE TEMPERATURA DE SUPERFICIE DEL MAR ATSM EN EL ÁREA DE DESOVE DEL CALAMAR Y EL MES DE MAYO 1982- 2010 2,5 5000 2 4500 4000 1,5 3500 1 3000 0,5 2500 0 ene- 82 ene- 83 ene- 84 ene- 85 ene- 86 ene- 87 ene- 88 ene- 89 ene- 90 ene- 91 ene- 92 ene- 93 ene- 94 ene- 95 ene- 96 ene- 97 ene- 98 ene- 99 ene- 00 ene- 01 ene- 02 ene- 03 ene- 04 ene- 05 ene- 06 ene- 07 ene- 08 ene- 09 ene- 10 2000 -0,5 1500 -1 1000 -1,5 500 -2 0 Biomasa de desovantes corrido un año SSP Numero de Reclutas 10/6 ATSM Figura 25: El calamar y su relación con las situaciones oceanográficas globales. Anomalía de temperatura de superficie del mar ATSM en el área de desove del calamar y el mes de mayo: 1982-2010 Terra Curanda www.terracuranda.org/fisheries 35 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori ANOMALÍA DE TEMPERATURA DE SUPERFICIE DEL MAR ATSM MARZOJULIO Y PROMEDIO 1982- 2010 EN EL ÁREA Y ÉPOCA DE DESOVE DEL CALAMAR 5000 2,5 4500 2 4000 1,5 3500 1 3000 0,5 2500 0 2000 -0,5 1500 -1 1000 -1,5 500 Biomasa de desovantes corrido un año SSP Numero de Reclutas 10/6 Densidad de Anfipodos MARZO ABRIL MAYO JUNIO JULIO PROMEDIO 2010 2009 2008 2007 2006 2005 2004 2003 2002 2001 2000 1999 1998 1997 1996 1995 1994 1993 1992 1991 1990 1989 1988 1987 1986 1985 1984 1983 -2 1982 0 Figura 26: El calamar y su relación con las situaciones oceanográficas globales. Anomalía de temperatura de superficie del mar ATSM Marzo-Julio, promedio 1982-2010 en el área y época de desove del calamar Manejo del calamar y su repercusión en el ecosistema pesquero En el caso del calamar, como se sabe, existen cuatro stocks dos de los cuales son compartidos internacionalmente. El de mayor abundancia es compartido y se ve afectado por la sobrecaptura de quienes lo comparten y por la de terceros países en el área adyacente. Indudablemente en el 2005 hubo un aumento de la abundancia en los stocks propios, con lo cual se duplicó la captura en referencia al 2004. En el 2006 el SSP (Stock Sud-Patagónico) se recuperó aunque no hubo posibilidad de predecir este hecho, el cual continuó en 2007 y 2008. En el siguiente gráfico se muestran los valores de stock desovante, corridos un año, que generan el siguiente reclutamiento. Terra Curanda www.terracuranda.org/fisheries 36 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori Variaciones del reclutamiento y de los desovantes que le dieron origen desovante en Numero del SSP corridos un año, para una relación directa 5000 1400 4500 Reclutamiento en Numero 1000 3500 3000 800 2500 600 2000 1500 400 1000 Biomasa desovante en Numero 1200 4000 200 500 0 19 78 19 79 19 80 19 81 19 82 19 83 19 84 19 85 19 86 19 87 19 88 19 89 19 90 19 91 19 92 19 93 19 94 19 95 19 96 19 97 19 98 19 99 20 00 20 01 20 02 20 03 20 04 20 05 20 06 20 07 20 08 20 09 0 Años SSP Numero de Reclutas 10/6 SSP Biomasa Desovante N Figura 27: Fuente: elaboración propia en base a datos obtenidos de Brunetti N., et al. Inf. Tecnico INIDEP: 77108/1996, 143/1997, 29-147/1998, 54-65/99, 65/2000, 86/2001, 87/2002, 103/2003, 70/2004, 50/2005, 53/2006, 61/2007. De las reuniones de la Comisión de Seguimiento de Calamar se sospechaba que las expectativas para el 2006 eran no muy halagüeñas especialmente para el SSP. Al producirse un resultado muy bueno, se decidió explorar una relación stock-recluta con los datos existentes, donde se eliminaron los valores del 92-94. Esta relación expresada en número de padres correspondió a hijos también en número para el año que correspondía al reclutamiento. Los valores fueron tomados de informes técnicos del INIDEP, evaluándose el gráfico solamente como una exploración. Terra Curanda www.terracuranda.org/fisheries 37 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori Relacion Stock Desovante Recluta SSP 5000 2008 4500 2007 4000 Reclutas en Numero 1999 3500 3000 2000 2500 2006 2000 1997 1500 1998 1000 2001 2003 2002 500 2005 2004 0 0 100 200 300 400 500 600 700 800 Stock Desovante en Numero Rel. Stock Recluta Logarítmica (Rel. Stock Recluta) Figura 28: Fuente: elaboración propia en base a datos obtenidos de Brunetti N., Inf. Tecnico INIDEP: 77-108/1996, 143/1997, 29-147/1998, 54-65/99, 65/2000, 86/2001, 87/2002, 103/2003, 70/2004, 50/2005, 53/2006, 61/2007, 52/2008, 15/2009. La mencionada exploración es sobre el concepto técnico de tasa de escape del 40% sobre el total reclutado o tasa de escape fija. Seguimiento y alerta temprano de cierre de la pesquería de Illex argentinus. Tasa de escape fija o dinámica De acuerdo a Aubone (2004-2005 MS) las condiciones que se deben considerar en referencia al manejo de la población de calamar son: • Una cohorte con ciclo de vida anual • Fecundidad dependiente del tamaño de las hembras, éste a su vez dependiente de condiciones ambientales, en general desconocidas • Desarrollo de la temporada de pesca en 16 semanas promedio Según Brunetti et al (2007) Inf Tec INIDEP 61-2007, el manejo y evaluación actual del calamar se realizan a partir del conocimiento del tamaño de cada población reclutada anualmente (N0) y de su evolución a lo largo de la temporada de pesca. El objetivo es permitir un escape suficiente de desovantes, que posibilite un adecuado reclutamiento al siguiente año. Se han establecido dos áreas de manejo, una al sur de los 44°S que comprende la plataforma y el talud austral y otra al norte de esta latitud ubicada en la plataforma y el talud bonaerense norpatagónico (Res. 973/97). Además se han determinado períodos de veda que contribuyen fundamentalmente a evitar la captura de juveniles y pre-adultos, permitiendo así su crecimiento hasta alcanzar mejores rendimientos comerciales. La biomasa desovante se calcula en base a dos metodologías: Terra Curanda www.terracuranda.org/fisheries 38 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori 1) Conociéndose las capturas semanales de cada población, es posible proyectar la evolución de la población en número a través de las semanas, desde una estimación inicial del número de individuos obtenida mediante la realización de un crucero de pre-reclutas (durante el mes de febrero, al sur de los 45°S, que permite evaluar la abundancia de pre-reclutas (N0) del stock sud-patagónico (SSP)), según el modelo de decrecimiento exponencial de Malthus, donde M es la tasa instantánea de mortalidad natural semanal considerada constante, M = 0,06 y Fi es la tasa instantánea de mortalidad por pesca de la semana i. La evaluación se lleva a cabo en “tiempo real” (actualizada semanalmente) a fin de satisfacer el objetivo de manejo, consistente en un escape proporcional del 40% del efectivo. El escape está definido como la proporción de desovantes sobrevivientes a la temporada de pesca, respecto de los que hubiesen estado vivos en caso de no existir la pesquería. El porcentaje de escape para la semana i se define como: 2) Como por razones ambientales a veces existen dudas que las campañas de evaluación no cubran la distribución de los pre-reclutas a partir de 2003 también se estima el número de pre-reclutas y la evolución en número de las diferentes cohortes mediante la metodología propuesta por Aubone (Inf.Téc. Int. INIDEP N° 36/2003) que utiliza datos de captura, esfuerzo y pesos medios individuales provenientes de la flota comercial. En la temporada 2007, la relación entre la CPUE y el número medio de ejemplares de cada semana no mostró un buen ajuste y fue modificada según se detalla a continuación: En referencia a la calidad del desovante y de la cantidad que supuestamente escapó, también habría que tomar en cuenta el tamaño de los individuos ya que existe una relación entre el mismo y la fecundidad. A la dispersión adaptando un ajuste de una curva logarítmica, los valores reales de escape, pareciera ser que fuera mas apropiado un valor fijo que realmente pueda asegurar un reclutamiento. Por sobre ese valor fijo pareciera que su abundancia dependiera mas de las condiciones ambientales que de la pesca. Condicionando la captura a partir de un determinado valor. Aubone (2004-5, MS) propone un sistema de alerta temprano con escape variable, el cual va a depender de la abundancia. Esto permite que en años “buenos” escapen en proporción menos individuos y en años “malos” una proporción mayor. Aubone también propone una “ventana temporal” de biomasa desovante mínima, para asegurar de alguna forma el reclutamiento, de acuerdo al conocimiento histórico. Este nuevo sistema de alerta temprano propone establecer un criterio de escape que considere la biología reproductiva, en términos de la “calidad” de la biomasa desovante y la cantidad de la misma. Requiere de datos biológicos de cada cohorte no disponibles en el pasado en forma regular, pero posibles de ser tomados en cada temporada. En muchos años el escape proporcional en número de ejemplares fue menor de 0,40 y también el escape en biomasa fue menor de 40.000 t. Sin embargo el ciclo de vida continuó. En el año 2003 la poca biomasa desovante y tal vez condiciones ambientales adversas provocaron el colapso del ciclo de vida del Stock Sud Patagónico de Illex argentinus. En Aubone (2004-5, MS) se discute sobre si los criterios de escape tradicionales son adecuados para asegurar de algún modo la continuidad del ciclo de vida. La respuesta es, que con valores de escape suficientemente grandes, especialmente de la proporción de escape en número de Terra Curanda www.terracuranda.org/fisheries 39 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori ejemplares, la seguridad biológica es mayor. El escape en biomasa busca fijar un mínimo de biomasa desovante y cuanto mayor sea esta biomasa también será mayor la seguridad biológica. La propuesta de Aubone genera un nuevo sistema de alerta temprano de cierre y recomendación de dejar de pescar, considerando un escape proporcional en número de ejemplares variable, atendiendo al reclutamiento estimado en el crucero de investigación y a datos de captura y esfuerzo de la flota pesquera. La mayor precaución de este sistema de alerta temprano resulta por el objetivo de mantener una biomasa desovante mínima en unas semanas de alta productividad de huevos, para que el desove sea suficiente. El umbral mínimo de biomasa en las semanas de alta productividad de huevos se determina de acuerdo a estimaciones históricas. Procedimiento del criterio de alerta temprano y recomendación de cierre de la pesquería (Dinámico) 1. Se estima el número de reclutas a una semana inicial, a partir de datos del crucero de investigación. 2. Se estima el número de reclutas a una semana inicial, a partir de datos de la pesquería, y un intervalo de confianza 3. Se comparan las estimaciones del número de reclutas 4. Se estima la probabilidad de no cumplir con los objetivos biológicos de escape 5. Se realiza un seguimiento del número de individuos en la cohorte, semanalmente 6. Al superar el riesgo biológico un nivel crítico, se alerta sobre la inminente necesidad de cierre de la pesquería 7. Cuando dejan de cumplirse los criterios biológicos de conservación, se recomienda el cierre de la pesquería El sistema de alerta temprano de cierre usado históricamente (escape proporcional mínimo de 0,40 y escape en biomasa de por lo menos 40.000 t) no contempla esta biomasa desovante mínima en las semanas de mayor productividad. Es un criterio no necesariamente se garantiza un desove adecuado para la continuidad poblacional, ya que el número de ejemplares que escapan dependerá de las tasas de mortalidad por pesca, y por lo tanto los individuos que escapan pueden no ser suficientes en cantidad o no ser de la calidad reproductiva necesaria. La calidad de los desovantes (fecundidad) de esas toneladas de escape depende de cada cohorte y de la semana en que escapan. A los efectos comparativos y exploratorio, se emplearon los datos obtenidos de Brunetti N. informes técnicos INIDEP referidos al SSP y los datos de Csirke 1987, en este caso nos muestra la etapa inicial de la pesquería que se desarrolla en el área austral, si bien estos datos no discriminan stock, por su ubicación se refieren en especial al SSP. La curva logarítmica resultante es mucho más conspicua permitiendo establecer un valor básico de 100 millones de individuos desovantes. Terra Curanda www.terracuranda.org/fisheries 40 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori De todas maneras en referencia al valor mencionado debe considerar como un valor promedio que puede llegar a desarrollar un buen reclutamiento. Este valor es solo a los efectos de su definitiva valoración por un grupo de expertos que pueda analizar el tema en profundidad. RELACIÓN STOCK RECLUTA DE CALAMAR SSP DATOS DE CSIRKE Y NORMA BRUNETTI 5000 2008 4500 2007 RECLUTAS EN MILLONES 4000 1999 3500 3000 2000 2500 2006 2000 1997 1500 1998 1982 1984 1985 2003 2002 1983 1000 500 1996 2001 1979 1995 1981 2005 1980 2004 0 0 100 200 300 400 500 600 700 800 STOCK DESOVANTE EN MILLONES Figura 29: Fuente: elaboración propia en base a datos obtenidos de Csirke, Documento Tecnico N 286, FAO Brunetti N., Inf. Tecnico INIDEP: 77-108/1996, 143/1997, 29-147/1998, 54-65/99, 65/2000, 86/2001, 87/2002, 103/2003, 70/2004, 50/2005, 53/2006, 61/2007. Otro evento a tomar en cuenta en la valoración de los datos es la funcionalidad y producción del ecosistema tal como se viene desarrollando el tema. La mezcla de valores determina dos puntos en el estado de la población de calamar debido a la disminución importante de la prelación de la merluza y del excedente de crustáceos pelágicos dejados libres por la disminución de los efectivos de peces del área. Relación entre el reclutamiento de calamar y reclutamiento de merluza y su relación con la pesca Algunos de los resultados obtenidos por (Koen Alonso M. y Yodzis P., 2005) indican que la explotación pesquera del calamar común afectaría negativamente a la población de merluza, mientras que la explotación de merluza favorecería al calamar común.( ver en el texto “Impacto ambiental e indirecto de la presión pesquera sobre los distintos efectivos” y “las interacciones específicas indirectas con las pesquerías y las poblaciones de mamíferos” ). En cuanto al concepto de ciclicidad entre especies en el siguiente grafico se expusieron valores de reclutamiento de calamar (N+10/9) del SSP obtenidos de los informes de la Dra. Norma Brunetti y a los cuales se le adjunto los valores de reclutamiento de la merluza de la edad 1, efectivo Norte y Sur de 41 LS obtenidos por modelo, y del efectivo sur de la campaña de juveniles, información extraída de los informes de INIDEP. Terra Curanda www.terracuranda.org/fisheries 41 Calamar (Illex argentinus): stock sud-patagónico Prenski & Morales Yokobori Observando el grafico se puede decir que años buenos de calamar se contraponen con años malos de merluza, se incluyeron los dos efectivos debido a que las larvas y pre-adultos de calamar migran por la plataforma al sur siendo objeto de captura trófica por ambos efectivos de merluza. Las posibles explicaciones pueden ser dos, la primera apunta a cambios oceanográficos que actúan de manera diferencial en una relación trófica o causalidad. De este modo, como se observa en el grafico, esto podría indicar al año 2007 como un año malo para merluza en el reclutamiento de la edad 1 y bueno para el calamar. Como conclusión podemos decir que por ser una pesquería basada en pocas clases de edad 2 y 3, cualquier disminución en el reclutamiento inmediatamente se reflejará en la potencialidad de la pesquería. Se puede interpretar de diferente manera y seria que años de buenos reclutamientos de merluza actúan de manera que el importante número de juveniles, actúen intensamente sobre las larvas y juveniles de calamar disminuyendo así su reclutamiento. RELACIÓN ENTRE EL RECLUTAMIENTO DE CALAMAR SSP Y MERLUZA 5000 4500 NÚMERO DE IND 10/6 4000 3500 3000 2500 2000 1500 1000 500 0 1986 1987 1988 1989 1990 1991 1992 1993 1994 1995 1996 1997 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 AÑOS SSP Numero 10/6 Reclutamiento de la merluza Norte edad 1 Modelo Reclutamiento de la merluza Sur edad 1 Modelo Reclutamiento edad 1 Camp. De juveniles Figura 30: Fuente: elaboración propia en base a datos obtenidos de Brunetti N., Inf. Tecnico INIDEP: 77-108/1996, 143/1997, 29-147/1998, 54-65/99, 65/2000, 86/2001, 87/2002, 103/2003, 70/2004, 50/2005, 53/2006; 61/2007,52/2008, 15/2009; Cordo D y Renzi M. & Irusta G. INIDEP 36-39/2006; Irusta G., Renzi M, D’ Átri l., Informe INIDEP 43 / 2007; Irusta G., Renzi M., Informe INIDEP 47 /2007; Louge E, Santos B., Castrucci R.,10-2002; Louge E., Bettina Reta R., Santos B., 103-2005; Cordo H. 39-2006; Dato C., Pajaro M. Terra Curanda www.terracuranda.org/fisheries 42