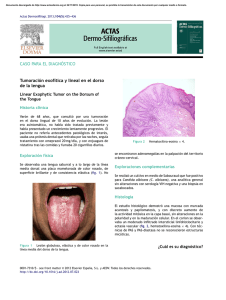
Factores de virulencia en Candida sp
Anuncio

Dermatología Rev Mex 2005;49:12-27 Artículo de revisión Factores de virulencia en Candida sp Laura Estela Castrillón Rivera,* Alejandro Palma Ramos,* Carmen Padilla Desgarennes** RESUMEN La incidencia cada vez mayor de infecciones intrahospitalarias por Candida y su asociación individuos susceptibles que cursan con pdf elaborado por en medigraphic ciertas inmunodeficiencias hacen posible que estos microorganismos, considerados comensales, se vuelvan patógenos. Conocer los factores de virulencia del microorganismo hace posible determinar, de manera más precisa, cómo se modifica la relación que establece con su huésped una vez que los mecanismos de resistencia se han deteriorado. Los principales factores de virulencia asociados con Candida se relacionan con su dimorfismo, secreción enzimática, cambio de fenotipo, expresión diferencial de genes en respuesta al ambiente, síntesis de adhesinas y su capacidad para formar biopelículas. En este trabajo se presenta una revisión de la descripción de estos factores y cómo éstos se relacionan con el proceso infeccioso. Palabras clave:Candida, factores de virulencia, morfogénesis, adhesinas, enzimas, biopelículas. ABSTRACT The higher incidence of intrahospitalary infections by Candida and its association in susceptible individuals that course with certain immunodeficiency states make possible that these microorganisms considered commensals become pathogens. To know the virulence factors of this microorganism allows to determine in a more precisely way how the relationship that it establishes with the host is modified once the resistance mechanisms have been deteriorated. The main virulence factors associated with Candida are related to dimorphism, enzymatic secretion, phenotype change, differential gene expression to environmental response, adhesions synthesis and their ability to form biofilms. In this work we present a review of the description of these factors and how they are related to the infection process. Key words: Candida, virulence factors, morphogenesis, adhesins, enzymes, biofilms. C andida sp son organismos comensales en el intestino de individuos sanos; se ha visto que están presentes universalmente. La colonización por Candida albicans puede llevar a la infección sistémica cuando el huésped tiene varios factores de riesgo, como el uso de antibióticos de amplio espectro, esteroides u otros agentes inmunosupresores, diabetes mellitus, SIDA, depresión de las funciones fagocíticas o alteraciones locales del sistema gastrointestinal. Estas situaciones pueden * Departamento de sistemas biológicos, Universidad Autónoma Metropolitana Xochimilco. ** Centro Dermatológico Pascua. Correspondencia: Dra. Laura Estela Castrillón Rivera. Departamento de sistemas biológicos, Universidad Autónoma Metropolitana Xochimilco. Calzada del Hueso 1100, colonia Villa Quietud, CP 04960, México, DF. E-mail: [email protected] Recibido: noviembre, 2004. Aceptado: diciembre, 2004. 12 ocasionar candidiasis gastrointestinal o diseminación hematógena.1 Candida albicans es un microorganismo muy versátil, por su capacidad para sobrevivir como comensal en varios sitios anatómicamente distintos (intestino, cavidad oral y vagina), y puede causar enfermedad cuando se le presenta la oportunidad. La limitación por nutrientes y la competencia entre bacterias y hongos (microbiota) en las superficies mucosas proporcionan una presión selectiva que ocasiona la eliminación de los microorganismos menos adaptados. Candida albicans tiene varios atributos de virulencia para colonizar el huésped y ocasiona daño de forma directa, al activar, resistir o desviar los mecanismos de defensa del mismo. Los factores de virulencia expresados o requeridos por el microorganismo para causar infección pueden variar según el tipo, el sitio y la naturaleza de las defensas del huésped.2 El delicado equilibrio entre el huésped y el hongo patógeno puede convertirse en una relación parásita Dermatología Rev Mex Volumen 49, Núm. 1, enero-febrero, 2005 Factores de virulencia en Candida sp y resultar en enfermedad grave. Los hongos no son participantes pasivos en el proceso infeccioso. La interacción entre el hongo y el medio ambiente está afectada por su variabilidad antigénica, el cambio fenotípico y la transición dimórfica. Existen diversos factores potenciales de virulencia, como la morfología celular, la actividad enzimática extracelular, el cambio fenotípico y los factores de adhesión, que favorecen la formación de biopelículas. A continuación se describen, de manera independiente, cada uno de estos factores. MORFOGÉNESIS Candida albicans es polimórfica, ya que existe en forma de levadura (blastosporas) o como filamentos (pseudohifa o hifa). La morfogénesis se refiere a la transición entre las levaduras (unicelulares) y la forma de crecimiento filamentosa del microorganismo, que puede convertirse de forma reversible a células de levadura, con crecimiento de hifa o pseudohifa. conversión la forma unicelular de levadura pdfLa elaborado porde medigraphic al crecimiento filamentoso es esencial para la virulencia de Candida albicans. La morfogénesis, por sí misma, está bajo múltiples controles y rutas de transducción de señales. La transición de levadura a hifa es uno de los atributos de virulencia que capacitan a Candida albicans para invadir los tejidos. Se ha comprobado que el crecimiento de forma filamentosa tiene ventajas sobre la levadura en la penetración de la célula o tejido, y aunque la hifa puede ser idónea para abrir la brecha entre las barreras tisulares, gracias a que su punta es el sitio de secreción de enzimas capaces de degradar proteínas, lípidos y otros componentes celulares, ésta facilita su infiltración en sustratos sólidos y tejidos.3 En general, las levaduras predominan durante la colonización de la mucosa en el huésped sano, pero la hifa emerge cuando las defensas de éste declinan. Por lo tanto, ambas formas de crecimiento podrían desempeñar un papel importante en la patogénesis y encontrarse en muchos microambientes diferentes en el huésped.4 La hifa se produce en el estado temprano de la colonización, mientras que las levaduras se observan comúnmente durante la enfermedad o en el tejido necrótico, justo cuando el crecimiento de la hifa se revierte por el suero a la forma de levadura y las proteínas se degradan por proteinasas in vivo e infiltran tejidos haciéndolos necróticos. En otras palabras, la morfogénesis de levadura a hifa se revierte conforme avanza la infección y quizá sea el resultado de cambios temporales en señales que el hongo recibe de su medio ambiente (figura 1). Episodios tempranos Episodios tardíos Activación de genes de polimorfismo y señalización celular Levadura Tubo germinativo Reconocimiento por receptores de manosa de células dendríticas. Activación Respuesta TH 1 = Protección Levaduras gemantes Hifa Reconocimiento por receptores Penetración a tejidos C3 o FcR de células dendríticas. Activación Respuesta TH 2 = Producción de anticuerpos Figura 1. Morfogénesis en Candida sp durante la infección. Dermatología Rev Mex Volumen 49, Núm. 1, enero-febrero, 2005 13 Castrillón Rivera LE y col. También se ha propuesto que existe asociación entre la morfogénesis y la virulencia en los hongos dimórficos, ya que un morfotipo existe en el medio ambiente o durante el comensalismo y el otro, en el huésped durante el proceso de la enfermedad.5 Sin embargo, se ha sugerido que es el cambio de la forma de levadura a hifa (morfogénesis) y no la forma del hongo lo que es importante para la virulencia.6 Otro tipo de pruebas que apoyan a la morfogénesis como factor de virulencia es que las lesiones clínicas a menudo se marcan por la existencia de células de varias morfologías, por lo que es legítimo asumir que todas las formas del hongo son necesarias para mantener la infección por Candida albicans. Existen tres hallazgos que sustentan la hipótesis de que la filamentación se requiere para la virulencia por este hongo:7 1. La formación de filamentos se estimula a 37 ºC en presencia de suero, con pH neutro. 2. Los filamentos recién formados (llamados tubos germinativos) son más adherentes a las células mamíferas que las levaduras y la adherencia es el requisito para la penetración tisular. 3. Las levaduras capturadas por macrófagos producen filamentos y son capaces de lisar a éstos, por lo tanto, la formación de filamentos es una forma de evadir los mecanismos de defensa del huésped.8,9 Se han realizado varios intentos para demostrar la relación que existe entre la forma filamentosa de Candida albicans con su virulencia, pero hasta ahora no hay una demostración inequívoca acerca de este punto, aunque hay datos de que la virulencia se atenúa cuando se obtienen mutantes que se confinan a la forma de levadura o filamentosa. Por ejemplo, cepas mutantes de Candida albicans incapaces de formar hifas son generalmente avirulentas en modelos de ratón con candidiasis diseminada. Por lo tanto, se sugiere que los defectos de la cepa, para formar hifas, podrían reducir la virulencia del hongo.7-10 La correlación de la morfogénesis del hongo con la virulencia se basa en los estudios realizados en animales; no obstante, el análisis de la virulencia fúngica se limita por los modelos inadecuados comparados con la enfermedad en humanos. Una limitante importante para poder llegar a estas conclusiones es que aún no existen los modelos de 14 infección crónica que semejen la enfermedad en humanos y por ello es importante reconocer que un modelo experimental inapropiado puede llevar a conclusiones erróneas, en relación con los factores de virulencia de los hongos.11 La morfogénesis de levadura a hifa es reversible in vivo y se controla por redes reguladoras que modifican otros aspectos de la biología de Candida albicans. Durante la infección evolucionan distintas fases morfogenéticas quemedigraphic pueden ser influidas por microampdf elaborado por bientes locales en el huésped, éstas son:4 1) Episodios tempranos en la colonización por células de levadura que pueden afectar su decisión de generar un tubo germinativo hasta la formación de la hifa. 2) Episodios subsecuentes que promueven el crecimiento de la hifa extendida. 3) En las fases tardías de la infección, las células necróticas a menudo se acompañan de la transición de hifa a levadura y generan células en gemación. Estas fases implican cambios sutiles en el ciclo celular, cambios importantes en la polaridad de crecimiento, alteraciones en la composición de la pared celular y modulación de factores de virulencia. De ahí que las rutas morfogenéticas se integran con rutas de señalización que controlan otros procesos. Por lo que se refiere a la respuesta del huésped ante el dimorfismo de Candida, se ha observado que la capacidad de Candida albicans para cambiar entre la forma de levadura saprofita y la forma filamentosa patogénica del hongo permite a las células dendríticas (responsables de la inmunidad innata) detectar una forma específica, al ocasionar respuestas dependientes de células T cooperadoras, protectoras o no protectoras. Lo anterior, en virtud de que dichas células poseen distintos tipos de receptores y a que contribuyen a disparar patrones diferentes de reactividad, observados de forma local en respuesta al desafío por Candida albicans.12 Estos hallazgos ofrecen nuevas claves para interpretar los mecanismos de virulencia de los hongos: más que el dimorfismo, la inclusión de diferentes tipos de receptores en las células dendríticas puede seleccionar el modo de internalización del hongo y de la presentación antigénica, lo que condiciona las respuestas de las células T cooperadoras y favorece el saprofitismo o la infección 2 (figura 1). Dermatología Rev Mex Volumen 49, Núm. 1, enero-febrero, 2005 Factores de virulencia en Candida sp Genética molecular del polimorfismo de Candida Candida albicans crece como levaduras o como hifas. La hifa es la forma de adaptación para la adherencia y penetración de los epitelios y células endoteliales. Se identificó que el gen EFG1 en Candida albicans codifica para un regulador transcripcional que tiene homologación con las proteínas PHD1 y StuA, que son las proteínas que controlan la morfogénesis en Saccharomyces cerevisiae y Aspergillus nidulans, respectivamente. Los estudios del EFG1 sugieren que es un activador transcripcional y un represor esencial para la morfogénesis de la levadura, hifa y pseudohifa.1 Los estudios bioquímicos que establecen las rutas de transducción de señales, que activan la filamentación de Candida albicans, concluyen que existen dos rutas principales, una es la cascada de cinasas MAPK y la segunda está representada por el factor transcripcional EFG1p. Las mutantes que les faltan a ambas rutas no son filamentosas y son avirulentas.13 ENZIMAS Las pueden proponerse como determinantes pdf enzimas elaborado por medigraphic de virulencia en Candida, ya que tienen la capacidad de romper polímeros que proporcionan nutrientes accesibles para el crecimiento de los hongos, así como de inactivar las moléculas útiles en la defensa del organismo. Las principales enzimas extracelulares relacionadas con la patogénesis de Candida son las proteasas, fosfolipasas y lipasas. Proteasas En Candida albicans se han descrito varios miembros de una gran familia de enzimas de secreción aspártico proteinasas (SAP), que han sido bastante estudiadas en estos hongos. En particular, las aspartil proteinasas secretadas (Saps) son codificadas por los genes de la familia SAP, que cuenta con diez miembros y que está regulada diferencialmente; además, sus distintos miembros se expresan bajo una variedad de condiciones de crecimiento de laboratorio y durante las infecciones experimentales in vivo e in vitro. La contribución de las aspartil proteinasas secretadas a la patogénesis de Candida albicans se ha demostrado con el uso de mutantes deficientes en SAP e inhibidores de proteasas. La presencia de los genes de la familia Dermatología Rev Mex Volumen 49, Núm. 1, enero-febrero, 2005 SAP en Candida albicans proporciona al hongo un sistema proteolítico eficiente y flexible que puede garantizar su éxito como patógeno oportunista.3 Se ha sugerido que las aspartil proteasas secretadas desempeñan un papel importante en la patogénesis de C. albicans, ya que se han obtenido mutantes con varios genes SAP alterados y se ha demostrado que SAP1-3 y SAP6 son importantes en la infección oral, mientras que SAP1 y SAP2 lo son en la candidiasis vaginal. El papel de dichas enzimas es esencial en las infecciones de mucosas en las fases iniciales, pero no cuando el hongo se ha infiltrado en los vasos sanguíneos. Asimismo, hay pruebas clínicas que correlacionan la secreción de estas enzimas con la candidiasis vaginal.1 Caracterización de las aspartil proteinasas de Candida albicans Los diez genes SAP codifican pre y proenzimas de aproximadamente 60 a 200 aminoácidos, mayor que la proteína madura. El segmento señal N-terminal es fragmentado por una señal peptidasa en el retículo endoplásmico. El propéptido se remueve para activar la enzima por una proteinasa semejante a subtilisina Kex-2 en el aparato de Golgi, antes de que sea transportada, vía vesículas, hacia la superficie de la célula para la secreción o glucosilación. Se sabe que existen otros procesos alternativos para la activación de las SAP de C. albicans, como la autoactivación que ocurre extracelularmente para SAP1, SAP2, SAP3 y SAP6 a ciertos valores de pH.14 Las proteínas maduras contienen una secuencia de motivos típicos de todas las aspartil proteasas, que incluyen los dos residuos aspartato conservados en el sitio activo. Es posible que los residuos cisteína conservados mantengan la estructura tridimensional de las enzimas.15 Las proteínas que comprenden la familia SAP no se limitan sólo a Candida albicans, ya que se ha demostrado su presencia en C. dubliniensis, C. tropicalis y C. parapsilosis.16,17,18 La presencia de los genes de la familia SAP es única en las especies patógenas de Candida y están ausentes en la levadura no patógena S. cerevisiae, lo cual apoya que estas proteinasas están implicadas en su virulencia. Aunque las consecuencias de la secreción de estas enzimas durante las infecciones en humanos aún no 15 Castrillón Rivera LE y col. se conocen con precisión, los estudios in vitro, en animales y en humanos, han implicado a las proteinasas de C. albicans en una de las siguientes formas:1 9 1. Correlación entre la producción de SAP in vitro y la virulencia. 2. Degradación de proteínas humanas y análisis estructural al determinar la especificidad del sustrato de SAP. 3. Asociación de la producción de SAP con otros procesos de virulencia de C. albicans. 4. Producción de la proteína SAP y reacción inmunitaria en infecciones animales y humanas. 5. Expresión de genes SAP durante las infecciones por Candida. 6. Modulación de la virulencia de C. albicans por inhibidores de aspartil proteinasas. 7. Uso de mutantes de SAP para analizar la virulencia de C. albicans. Las proteínas SAP tienen funciones especializadas durante el proceso infeccioso e incluyen la digestión de moléculas proteínicas para adquirir nutrientes, digerir o distorsionar las membranas del huésped y facilitar la adhesión, la invasión a tejidos y la digestión de moléculas del sistema inmunitario del huésped para evitar o resistir el ataque antimicrobiano de éste. Aunque la actividad proteolítica extracelular se descubrió a mediados de la década de 1960,20 no fue sino hasta el inicio de 1990 cuando los métodos moleculares se introdujeron al campo de Candida y los científicos comenzaron a comprender la complejidad genética de este hongo, demostrando diez genes SAP que codifican para estas enzimas. Mientras se hacían esfuerzos por recolectar datos de los genes SAP se despertó el interés hacia su papel y función durante el proceso infeccioso. Es claro que hay diferencias temporales y espaciales en la expresión de los genes SAP, cada uno de los cuales desempeña diferente papel en el establecimiento de la enfermedad y en la invasión de los tejidos.21 Se investigó el papel de las aspartil proteinasas secretadas durante la invasión de tejidos y su asociación con las diferentes morfologías de C. albicans. Se demostró que dentro de las primeras 72 horas postinfección, las proteinasas SAP1, SAP2, SAP4, SAP5 y SAP9 fueron los principales genes expresados. 16 Los antígenos SAP1 a SAP3 se encontraron en las células de levadura e hifa, mientras que los antígenos SAP4 a SAP6 se apreciaron sobre todo en las células de hifa que están en contacto muy estrecho con las células del huésped, en particular con leucocitos y eosinófilos.22 La presencia de los genes SAP indica que diferentes proteinasas pueden tener distintas acciones en las células del huésped y en los tejidos durante el proceso infeccioso, lo cual que su expresión puede ser pdf elaborado por sugiere medigraphic regulada de manera diferencial bajo diversas condiciones de laboratorio y durante el proceso infeccioso. En este sentido, se ha demostrado que bajo condiciones de laboratorio la principal enzima proteolítica expresada por C. albicans es SAP2, que está regulada por un mecanismo de retroalimentación positiva donde los péptidos resultantes de su acción inducen su síntesis.23 En cambio, los genes de SAP1 y SAP3 se expresan diferencialmente durante el cambio de fenotipo en ciertas cepas, 24 mientras que la expresión de SAP8 está regulada por la temperatura.25 Se ha demostrado que las diferentes Saps tienen un valor de pH óptimo para su actividad. La SAP2 actúa sobre todo en valores de pH ácidos (cercanos a 4.0), las Saps 4 a 6 son activas al pH fisiológico y la SAP3 al pH de 2.0. Esta característica proporciona a C. albicans un rango muy amplio de actividad proteolítica, que va de 2.0 a 7.0, lo que es esencial para la adaptación específica hacia diferentes condiciones ambientales del huésped.26 La contribución de los genes de la familia SAP en la manifestación de la enfermedad se ha estudiado por RT-PCR, cepas mutantes y por métodos inmunoquímicos. Con estos estudios se ha demostrado que las Saps 1 a 6 se requieren para la enfermedad invasiva, mientras que en modelos experimentales de vaginitis la SAP2 se necesita para el desencadenamiento de la enfermedad. En modelos in vitro de epidermis humana se midió la expresión temporal de Saps por la técnica de RTPCR. El orden de expresión fue SAP1, SAP2, seguido, de forma secuencial, por SAP8, SAP6 y SAP3. La expresión tuvo correlación con la invasión del tejido, es decir, invasión temprana (SAP1 y 2), penetración extensiva (SAP8) y crecimiento extensivo de la hifa (SAP6). Dermatología Rev Mex Volumen 49, Núm. 1, enero-febrero, 2005 Factores de virulencia en Candida sp Correlación entre morfogénesis y secreción enzimática La pared celular es el punto de contacto entre Candida albicans y su huésped. Durante la morfogénesis de levadura a hifa ocurren cambios distintos en la arquitectura de la pared, como resultado de la regulación de las enzimas biosintéticas de la pared celular. En resumen, los genes que codifican a las enzimas de secreción aspartil proteinasas se regulan diferencialmente durante el desarrollo de la hifa: los genes SAP4 a 6 se activan cuando la formación de la hifa se induce en un medio que contiene polipéptidos como única fuente de nitrógeno. También la hifa de C. albicans es capaz de producir rompimientos en las superficies, lo que quizás promueva la penetración del hongo en células y membranas endoteliales del huésped. Esta propiedad (conocida como tigmotropismo) depende del crecimiento de la hifa. La morfogénesis de levadura a hifa está muy relacionada con otros factores de virulencia, que incluyen la biosíntesis de la pared celular, la adhesión, la producción de enzimas hidrolíticas extracelulares y el tigmotropismo. demostración de los diferentes patrones de pdfLa elaborado por medigraphic expresión de las distintas Saps en la levadura e hifa, así como en células que han cambiado su fenotipo indican que dicha expresión es un proceso altamente regulado; sin embargo, no aseguran si este proceso contribuye a la patogenicidad de C. albicans in vivo. Por tal motivo, es decisivo saber si la expresión diferencial también ocurre durante el proceso infeccioso, para lo cual se han utilizado modelos de infección in vitro e infecciones experimentales en animales. Con estos estudios se demostró que las SAP1 a SAP3 son las proteinasas principales expresadas durante las infecciones superficiales,27 en contraste con las SAP1 a SAP4, que se expresan sobre todo en la enfermedad sistémica.28 Estos hallazgos se apoyan en el uso de cepas mutantes de enzimas específicas en diferentes modelos de infección. Las mutantes de EFG1 (gen del factor transcripcional Efg1) tienen capacidad reducida para formar hifas. También se asocian con la capacidad limitada de sintetizar las enzimas SAP4 a SAP6, y se ha demostrado que las mutantes que carecen de SAP1 a SAP3 tienen propiedades invasivas comparables con las cepas silvestres. En cambio, las mutantes de SAP4 Dermatología Rev Mex Volumen 49, Núm. 1, enero-febrero, 2005 a SAP6 reducen su capacidad invasiva, aunque puedan lograr la transición a hifa. Por lo tanto, las cepas que producen hifas pero que carecen de las proteinasas asociadas con este morfotipo son menos invasivas. Esto se demostró con mutantes deficientes de uno o de ambos genes y de su interacción con el epitelio oral y con neutrófilos polimorfonucleares, que en comparación con la cepa silvestre, las mutantes cph, efg1 y las cph/efg1 fueron incapaces de producir hifas, de dañar a las células, de expresar los genes SAP1 y SAP3 y de disminuir la expresión de SAP4, que es un gen asociado con la morfología de hifa. Con estos resultados se concluye que la virulencia reducida de mutantes deficientes en la formación de hifa no sólo se debe a esta propiedad, sino a la expresión modificada de los genes SAP, que por lo general se asocia con esta morfología. Por lo tanto, estos estudios ofrecen una prueba de la regulación combinada de factores de virulencia, como el polimorfismo y la expresión de genes SAP.29 Las principales características de estas enzimas se presentan en el cuadro 1. Posibles blancos de las proteinasas de Candida A nivel más básico, un papel de las Saps de Candida es la digestión de proteínas para proporcionar nitrógeno a las células; sin embargo, las Saps también pueden haberse adaptado y participar en funciones de virulencia. Por ejemplo, podrían contribuir a la adhesión en tejidos del huésped y a la invasión por degradación o distorsión de las estructuras en la superficie de las células del huésped o por destrucción de moléculas del sistema inmunitario del huésped para evitar o resistir el ataque antimicrobiano. La matriz extracelular y las proteínas de superficie del huésped, como queratina, colágena, laminina, fibronectina y mucina, son degradadas de manera eficaz por SAP2. Varias proteínas de defensa también son susceptibles de la hidrólisis por SAP, como la lactoferrina salival, el inhibidor de proteinasa αmacroglobulina, las enzimas del estallido respiratorio del macrófago y casi todas las inmunoglobulinas, que incluyen la IgA de secreción, la cual es resistente a muchas proteinasas bacterianas.30 La SAP2 de Candida también puede actuar en las cascadas proteolíticas del huésped, como los 17 Castrillón Rivera LE y col. Cuadro 1. Principales propiedades de las enzimas aspartil-proteasas de Candida (Sap) Característica Presentes en infecciones orales Presentes en infecciones vaginales Presentes a las 72 h de infección Presentes en la superficie de levadura e hifa Presentes en hifa Importantes en enfermedad invasiva Regulada por retroalimentación positiva Regulada por temperatura pH óptimo (2 a 5) pH óptimo (3 a 7) Proteína anclada en la membrana o pared celular Proteína anclada en la pared celular Expresadas durante el cambio de fenotipo Activas en infecciones sistémicas Se expresan en hifa con polipéptidos como fuente de N Las mutantes de esta enzima son invasoras Las mutantes de esta enzima reducen su invasividad Sap1 Sap2 Sap3 Sap4 Sap5 Sap6 X X X X X X X X X X X X X X pdf X elaborado X por medigraphic Sap7 Sap8 Sap9 X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X Referencias: 1,3,14-17,19, 21-30. precursores de la cascada de coagulación sanguínea 31 que hidrolizan al precursor del endotelio-1 (péptido vasoconstrictor), al alterar la homeostasia vascular32 y activar la citocina proinflamatoria IL-1β. Por lo tanto, esta enzima participa en la activación y mantenimiento de la respuesta inflamatoria en las superficies epiteliales in vivo.33 Fosfolipasas Otras enzimas hidrolíticas secretadas, en particular las fosfolipasas, se han relacionado con la patobiología de Candida albicans. 34 Las mutantes deficientes en fosfolipasa B1 han demostrado ser menos virulentas en modelos de infección en ratón.35 En la producción extracelular de fosfolipasas, demostrada por el aclaramiento de la yema de huevo en C. glabrata, C. parapsilosis, C. tropicalis, C. lusitaniae y C. krusei y por análisis de Southern blot, se ha demostrado la presencia de homólogos de fosfolipasa intracelular (CAPLC1) en C. dubliniensis, C. glabrata, C. parapsilosis y C. tropicalis.34 Se han identificado cuatro fosfolipasas (PLA, PLB, PLC y PLD), de las cuales sólo la PLB1 ha demostrado ser necesaria para la virulencia en modelo animal de candidiasis. Una cepa con la deleción de este gen (con menor producción de esta enzima) reduce su virulencia hasta en 60%, comparada con la cepa silvestre. La PLB1A es una glucoproteína de 84 kDa, que tiene actividad de hidrolasa y lisofosfolipasa- 18 transacilasa. Se secreta y detecta en la punta de las hifas durante la invasión a los tejidos.36 Lipasas Las lipasas secretadas por Candida albicans s e codifican por una familia de genes con, al menos, 10 miembros (LIP1-LIP10). El patrón de expresión se ha investigado en infecciones experimentales y en pacientes que sufren candidiasis oral. Se ha demostrado que la expresión de esos genes depende del estado de infección, más que de la localización del órgano.37 EXPRESIÓN DIFERENCIAL DE GENES EN REACCIÓN AL MEDIO AMBIENTE Candida albicans es capaz de sobrevivir y proliferar en un amplio rango de tejidos, ya sea como comensal o como patógeno. Durante los diferentes estados y tipos de infección las células del hongo necesitan amplia flexibilidad, ya que cada sitio anatómico tiene sus propias presiones ambientales. El que Candida albicans tenga genes que codifiquen para factores de virulencia puede reflejar la adaptación a un amplio rango de condiciones ambientales, en las que el hongo se encuentra durante su crecimiento in vivo. Por ejemplo, el pH sanguíneo es casi neutro, mientras que el de la vagina es ácido. Esta versatilidad puede sostenerse Dermatología Rev Mex Volumen 49, Núm. 1, enero-febrero, 2005 Factores de virulencia en Candida sp por un repertorio de genes que se expresan permitiendo su proliferación bajo diferentes condiciones.10 Lo anterior se explica porque Candida albicans muestra expresión diferencial de genes en respuesta al pH del medio, lo que puede ser importante para su patogénesis. El gen PHR1 codifica para una glucoproteína de superficie anclada en su membrana por el glucosilfosfatidilinositol y se expresa fuertemente cuando el pH del medio de crecimiento es mayor a 5.5, pero es indetectable por valores menores a este pH. El gen PHR2 (gen homólogo de PHR1) se identificó tiempo después y se expresó de forma complementaria al pH de 4.0, pero no al pH mayor de 6.0. Ambos genes están implicados en la morfogénesis in vitro de C. albicans. La deleción de cualquiera de estos genes causa crecimientos alterados en los pH restrictivos. La principal función del gen PHR1 se asocia con la síntesis de la pared celular, cuya expresión es óptima en pH cercanos a la neutralidad, como el que existe en el torrente circulatorio o en los tejidos, mientras que en el canal vaginal su expresión se apaga favoreciendo la función del PHR2, que es similar pero con ácido.38,39,40 Éste es un ejemplo pdf elaborado porpH medigraphic de cómo Candida albicans se adapta a condiciones fisiológicas extremas dentro del huésped. Se han clonado genes de respuesta al pH en C . dubliniensis, llamados CdPHR1 y CdPHR2,10 los cuales son homólogos a los genes de C. albicans y se requieren para la virulencia en las infecciones superficiales (vaginales) y sistémicas. Puesto que Candida albicans, que por lo general causa infecciones en la superficie de las mucosas en pacientes inmunocomprometidos, puede penetrar a tejidos profundos, entrar al torrente circulatorio y diseminarse en todo el huésped causando infecciones sistémicas, se evaluó el perfil de transcripción de genes de la misma especie expuesta a la sangre humana. Se demostró que la expresión de genes es diferente, incluyendo los que dependen de la respuesta al estrés, de la respuesta antioxidativa, del ciclo del glioxilato y de los atributos específicos de virulencia.41 Con esta información se demostró que C. albicans asegura su supervivencia en el medio ambiente hostil de la sangre y cómo el hongo escapa del torrente circulatorio como paso esencial en su diseminación sistémica. Dermatología Rev Mex Volumen 49, Núm. 1, enero-febrero, 2005 ADHESINAS Un atributo de Candida albicans es que correlaciona de forma positiva su patogenicidad con la capacidad adherente a las células del huésped. Las cepas adherentes de C. albicans son más patógenas que las que tienen fenotipo menos adherente. Una adhesina se define como una biomolécula que promueve la adherencia de C. albicans a las células del huésped o a sus ligandos específicos. Se han descrito proteínas de C. albicans que se unen a varias proteínas de la matriz extracelular de las células de mamífero, como fibronectina, laminina, fibrinógeno y colágeno tipo I y IV.42,43,44 Existen diferentes tipos de adhesinas en Candida, como Als, Hwp1p, Int1p y Mnt1p. Als Los genes de la familia Als de Candida albicans codifican glucoproteínas grandes de la superficie celular que están relacionadas con el proceso de adhesión a las células del huésped. Los genes Als también se encuentran en otras especies de Candida que han sido aisladas de muestras clínicas. Desde su caracterización inicial se notó que el producto traducido del gen Als1 comparte una secuencia con la glucoproteína de adhesión a superficies celulares llamada α-aglutinina, codificada por Saccharomyces cerevisiae (AGβ1).45 Esta molécula se requiere para el reconocimiento célula-célula. La familia Als tiene características típicas de proteínas de secreción y un residuo hidrofóbico carboxilo terminal que sugiere un ancla glucosilfosfatidilinositol. El ligando de unión se asocia con el residuo amino terminal de la proteína.46,47 Las proteínas Als están densamente N- y O-glucosiladas, sobre todo en la región de repetición variable rica en serina/ treonina y en los dominios carboxilo terminal.48 Los genes Als se describieron primero en C. albicans49 y esta familia incluye, al menos, nueve genes. Cada gen Als tiene una estructura similar de tres dominios, que incluyen un dominio 5’ de 1299 a 1308 pb, que es similar en 55 a 90% entre los componentes de la familia, un dominio central con un número variable de secuencias repetidas de 108 pb y un dominio 3’ que tiene una longitud relativamente variable entre los 19 Castrillón Rivera LE y col. genes de esta familia. El dominio de repetición variable incluye una secuencia interna de cinco aminoácidos: Val-Ala-Ser-Glu-Ser (VASES). Los genes Als están regulados diferencialmente en C. albicans por condiciones fisiológicas relevantes, como cambios en el medio de cultivo (Als1), morfología (Als3/Als8) y fase de crecimiento.46,49 La expresión de los genes Als se ha realizado por detección inmunohistoquímica in vivo de las proteínas en la superficie de Candida albicans en tejidos de enfermedad diseminada en ratón. Con esta metodología se demostró que las proteínas Als se distribuyen en la superficie celular, más que en sitios específicos. Hwp1p El gen Hwp1 codifica una manoproteína de la superficie externa que se cree se orienta por su dominio amino terminal expuesto hacia fuera. El carboxilo terminal se integra de manera covalente con el β-glucano de la pared celular. La proteína codificada por este gen (o Hwp1) sirve como sustrato de las transglutaminasas y, por lo tanto, la unión de la hifa de C. albicans a las células epiteliales de la boca debería ser estable y covalente y no revertirse por el uso de agentes que interrumpen la unión no covalente. Int1p Candida albicans se une a varios ligandos de las proteínas de la matriz extracelular, que incluyen fibronectina, laminina y colágenas tipo I y IV. Las cepas que tienen deleción en los genes Int1 son menos virulentas o se adhieren con más lentitud a las líneas celulares epiteliales. También tienen deficiencias en la filamentación en su crecimiento en leche-tween y agar Spider, por lo tanto, el gen Int1 juega un papel importante en la adherencia y filamentación de C . albicans.50 Mnt1p Las cepas de C. albicans que tienen deleción en el gen α-1,2 manosil tranferasa (Mnt1) son menos capaces de adherirse in vitro y son avirulentas.51 La proteína que codifica a este gen o Mnt1p es una proteína de membrana tipo II que se requiere para la manosilación 20 tanto O- y N- en el hongo. La manana es el principal constituyente de la pared celular. FORMACIÓN DE BIOPELÍCULAS Las especies de Candida se reconocen como los principales agentes de infecciones adquiridas en el hospital. Son el tercer o cuarto patógeno aislado de sangre que sobrepasa la frecuencia de los bacilos gramnegativos. Su aparición como patógeno pdf elaborado por52medigraphic nosocomial es importante, ya que es un factor de riesgo asociado con los procedimientos médicos modernos, sobre todo con el uso de fármacos inmunosupresores o citotóxicos, de antibióticos potentes que suprimen la flora bacteriana normal y de los dispositivos implantados de varias clases. Casi siempre, los dispositivos, como catéteres intravasculares o urinarios y tubos endotraqueales, se asocian con infecciones y se detecta la formación de biopelículas en su superficie.53 Otros dispositivos, como válvulas cardiacas, marcapasos y reemplazos de articulaciones (cadera o rodilla), son susceptibles de infección por Candida, generalmente durante el tiempo de su colocación. La mayor capacidad de C. albicans para formar biopelículas en estas superficies es la razón por la que esta especie es más patogénica que las que son menos capaces de formar estas estructuras, como C. glabrata, C. tropicalis, C. parapsilosis y C. keyfr.54 La percepción de los microorganismos como formas unicelulares de vida se basa en el modo de crecimiento como cultivo puro; sin embargo, la mayor parte de los microbios persisten unidos a las superficies dentro de un ecosistema estructurado, como biopelículas y no como organismos libres en flotación (planctónico). Una biopelícula es una comunidad de microorganismos unidos irreversiblemente a una superficie que contiene matriz exopolimérica y que muestra propiedades fenotípicas distintivas.55 En las biopelículas, las células desarrollan características fenotípicas que son diferentes de sus contrapartes planctónicas, como el incremento en la resistencia a los agentes antimicrobianos y la protección de las defensas del huésped.56 Las biopelículas formadas por C. albicans están compuestas por una capa de células en forma de levadura que se adhieren a la superficie en la parte de arriba. Esta capa la forman las células filamentosas en Dermatología Rev Mex Volumen 49, Núm. 1, enero-febrero, 2005 Factores de virulencia en Candida sp forma de hifa, rodeadas por una matriz extensiva exopolimérica.57,58 El material extracelular producido por C. albicans en esta biopelícula contiene carbohidratos, proteínas y otros componentes desconocidos y difiere en la composición del material extracelular producido por las células planctónicas (de flotación libre). 59 Se han distinguido diversos sistemas para estudiar las propiedades de la formación de biopelículas en Candida, como los modelos de crecimiento en discos de catéter, tiras de acrílico, placas de microtitulación y filtros de celulosa.60-64 El crecimiento se monitorea por ensayos colorimétricos, que dependen de la reducción de la sal de tetrazolio (técnica XTTx) o de la incorporación de 3H-leucina. Todos estos sistemas se realizan en condiciones estáticas de incubación; sin embargo, el método de biopelículas perfundidas en fermentador permite el control preciso de la velocidad de crecimiento.64 La formación de biopelículas de C. albicans s e distingue por la adhesión inicial de las células de levadura, seguida por la germinación y formación de microcolonias, filamentación, desarrollo de monocapa, proliferación maduración. Con el uso de microscopía pdf elaboradoypor medigraphic electrónica de barrido y confocal se han visualizado las biopelículas de C. albicans, que consisten en una densa red de células de levadura e hifas embebidas dentro de un material extracelular que demuestra heterogeneidad espacial y la arquitectura típica de microcolonias, con ramales de canalizaciones de agua.65 La formación de biopelículas se manifiesta en tres fases: temprana (0 a 11 horas), intermedia (12 a 30 horas) y madura (38 a 72 horas) (figura 2). La estructura detallada de las biopelículas, después de 48 horas de incubación, consiste en una densa red de levaduras, hifas y pseudohifas. La mezcla de levaduras, hifas y material de matriz no se ha visto cuando el organismo crece en medio líquido o en una superficie de agar, lo que sugiere que la morfogénesis se dispara cuando un organismo hace contacto con la superficie y que las células de la capa basal pueden tener un papel importante en el anclaje de la biopelícula a la superficie.66,67 Las cepas mutantes de C. albicans, con defectos en las rutas de filamentación, son incapaces de formar biopelículas tridimensionales, lo que indica que la formación de hifa es importante para la formación, desarrollo y mantenimiento de biopelículas. Hay dos consecuencias importantes y con profundas implicaciones clínicas en la formación de biopelículas: el marcado aumento en la resistencia a Figura 2. Fases de formación de biopelículas de Candida sp in vitro.74 La morfogénesis se activa cuando la levadura entra en contacto con la superficie y la capa de las células basales. Tiene un papel importante de anclaje de la biopelícula a la superficie, siguiendo el orden: adhesión temprana, crecimiento, proliferación y maduración.75 Dermatología Rev Mex Volumen 49, Núm. 1, enero-febrero, 2005 21 Castrillón Rivera LE y col. los agentes antimicrobianos y la protección contra las defensas del huésped. La resistencia clínica se define como la persistencia o progresión de una infección, a pesar del tratamiento antimicrobiano apropiado. La resistencia a antifúngicos por biopelículas de Candida se demostró por primera vez en 1995, en ensayos de catéteres en disco.68 También se ha estudiado en biopelículas en celulosa, poliestireno, elastómero de silicón, poliuretano y acrílicos de dentadura. Aunque las biopelículas de C. albicans y C. parapsilosis son claramente resistentes a los nuevos triazoles (voriconazol y ravuconazol), hay reportes de actividad antibiopelículas de formulaciones con lípidos de anfotericina B y dos equinocandinas, la micafungina y la caspofungina (inhibe la síntesis de β1,3 glucano, principal componente estructural de las paredes celulares de Candida). Se han propuesto tres posibles mecanismos de resistencia de las biopelículas a los fármacos y son: 1) Penetración restringida a través de la matriz de la biopelícula. 2) Cambios de fenotipo que ocasionan disminución del crecimiento o limitación de los nutrientes. 3) Expresión de genes de resistencia inducidos por el contacto con la superficie. Por lo que se refiere a la penetración restringida, se ha sugerido que la matriz extracelular excluye o limita el acceso de los medicamentos a los organismos profundos de la biopelícula. Muchos estudios indican que la matriz no forma una barrera para la difusión, aunque para ciertos compuestos se retrasa la penetración.69 Se ha propuesto que el glicocalix causa la disminución de la difusión o secuestro de los antimicrobianos. Sin embargo, al disgregar las biopelículas, ajustar a una densidad celular y realizar la prueba de susceptibilidad en las biopelículas preformadas, utilizando el ensayo XTTk, los resultados demostraron que las biopelículas preformadas fueron intrínsecamente resistentes a los antifúngicos y que las células móviles mantenían su fenotipo de resistencia después de la resuspensión. Lo anterior sugiere que la integridad de la estructura de la biopelícula y el glicocalix confieren efecto mínimo en la resistencia de las células móviles. 22 Las biopelículas crecen poco a poco porque los nutrientes están limitados, lo que ocasiona disminución del metabolismo de los microorganismos. Con frecuencia, la lenta velocidad de crecimiento se asocia con la adopción de diferentes fenotipos por los microorganismos, como cambios en la cubierta celular, con lo cual se afecta la susceptibilidad del microorganismo a los agentes antimicrobianos. Para comprobar si la velocidad de crecimiento es un parámetro importante en la adquisición de resistencia pdf elaborado por medigraphic a los antifúngicos, se generaron biopelículas con diferentes velocidades de crecimiento y se comparó la susceptibilidad a la anfotericina B de las células de la biopelícula contra la de las células planctónicas que crecían a la misma velocidad. Se encontró que las células de la biopelícula fueron resistentes al fármaco a todas las velocidades de crecimiento probadas, mientras que las planctónicas fueron resistentes sólo a bajas velocidades de crecimiento.64,70 Por lo tanto, la resistencia depende de otros factores, además de la forma de crecimiento. Dado que la resistencia a los antibióticos en las biopelículas de C. albicans no puede atribuirse a la exclusión por la matriz o a la lenta velocidad de crecimiento, quizá exista un mecanismo adicional, como la expresión de genes inducidos por contacto, para adquirir otras propiedades y mediante el cual se adquiera la resistencia a los antibióticos.71 Los principales genes que contribuyen a la resistencia de los medicamentos son los que codifican a las bombas de eflujo multifármacos; su regulación resulta en un fenotipo resistente a varios agentes.72 Se determinó que cuando un microorganismo se une a la superficie y forma una biopelícula expresa un fenotipo alterado . C. albicans y C. dubliniensis poseen dos diferentes tipos de bombas de eflujo: unión adenosin trifosfato cassette (ABC), transportadores que codifican los genes CDR (CDR1 y CDR2), y los facilitadores codificados por los genes MDR. Ambos tipos de genes se regulan durante la formación y desarrollo de las biopelículas.65,73 Sin embargo, las cepas mutantes que llevan deleción simple o doble de estos genes son susceptibles al fluconazol cuando crecen de forma planctónica, pero retienen su fenotipo de resistencia cuando crecen en biopelículas.74 Por lo tanto, el mecanismo de resistencia es un proceso que Dermatología Rev Mex Volumen 49, Núm. 1, enero-febrero, 2005 Factores de virulencia en Candida sp no puede explicarse por un solo tipo de mecanismo molecular. 7. CONCLUSIÓN 9. Los factores que contribuyen a la patogénesis de C . albicans incluyen la morfogénesis (transición entre las células de levadura unicelulares y las formas de crecimiento filamentosas), las enzimas secretadas aspartil proteasas (SAP) y fosfolipasas y las biomoléculas de reconocimiento del huésped (adhesinas), que le permiten iniciar el proceso de formación de biopelículas. Asimismo, el cambio de fenotipo se acompaña por alteraciones en la expresión de antígenos, morfología colonial y afinidades de C. albicans a los tejidos. La información experimental de la contribución de estos factores de virulencia ha determinado que su participación individual no es suficiente para explicar los mecanismos de daño en el huésped, sino que existe una regulación combinada entre ellos. Esto se demostró con el polimorfismo y la expresión genética para las aspartil proteasas. estudiospor quemedigraphic se realizan hoy en día en relación pdfLos elaborado con la expresión genética de los factores de virulencia, en función de diferentes condiciones ambientales, permitirán entender con mayor precisión cómo se va modificando la actividad biológica de Candida para favorecer la adhesión o penetración y, en consecuencia, modificar su papel como comensal para transformarse en microorganismo patógeno. REFERENCIAS 1. 2. 3. 4. 5. 6. Weig M, Grob U, Muhlschlegel F. Clinical aspects and pathogenesis of Candida infection. Trends Microbiol 1998;6(12):468-70. Castrillón RL, Palma RA, Padilla DC. Reacción inmunológica en infecciones por Candida sp. Dermatología Rev Mex 2004;48(3):140-50. Hube B, Naglik J. Candida albicans proteinases: resolving the mystery of a gene family. Microbiology 2001;147:19972005. Allistair JP, Brown AR, Gow AR. Regulatory networks controlling Candida albicans morphogenesis. Trends Microbiol 1999;7(8):333-8. Kobayashi SD, Cutler JE. Candida albicans hyphal formation and virulence: is there a clearly defined role? Trends Microbiol 1998;6:92-94. Roney PJ. Linking fungal morphogenesis with virulence. Cell Microbiol 2002;4:127-37. Dermatología Rev Mex Volumen 49, Núm. 1, enero-febrero, 2005 8. 10. 11. 12. 13. 14. 15. 16. 17. 18. 19. 20. 21. 22. 23. 24. 25. 26. Mitchell AP. Dimorphism and virulence in Candida albicans . Curr Opin Microbiol 1998;1:687-92. Cutler JE. Putative virulence factors of Candida albicans. Annu Rev Microbiol 1991;45:187-218. Gow NAR, Alistair JPB, Odds FC. Fungal morphogenesis and host invasion. Curr Opin Microbiol 2002;5:366-71. Haynes K. Virulence in Candida species. Trends Microbiol 2001;9(12):591-6. Latgé JP, Calderone R. Host-microbe interactions: fungi invasive human fungal opportunistic infections. Curr Opin Microbiol 2002;5:355-8. Romani L, Bistoni F, Pucceti P. Fungi, dendritic cells and receptors: a host perspective of fungal virulence. Trends Microbiol 2002;10(11):508-14. Lo HJ, Kohler JR, DiDomenico BK, Loebenberg D, Cacciapuoti A, Fink GR. Nonfilamentous C. albicans mutants are avirulent. Cell 1997;90:939-49. Koelsh G, Rockwell N, Wang GT, Krafft GA, Fuller RS. Enzymatic characteristics of secreted aspartic proteases of Candida albicans . Biochim Biophys Acta 2000;1480:117-31. Hube B. Candida albicans secreted aspartic proteinases. Curr Top Med Mycol 1996;7:55-69. Gilfillian GD, Sullivan DJ, Haynes K, Parkinson T, Coleman DC, Gow NA. Candida dubliniensis: phylogeny and putative virulence factors. Microbiology 1998;144:829-38. Zaugg C, Borg-Von Zepelin M, Reichard U, Sanglard D, Monod M. Secreted aspartic proteinase family of Candida tropicalis. Infect Immun 2001;69:405-12. Monod M, Tongi G, Hube B, Sanglard D. Multiplicity of genes encoding secreted aspartic proteinases in Candida species. Mol Microbiol 1994;13:357-68. Naglik JR, Challacombe SJ, Hube B. Candida albicans secreted aspartyl proteinases in virulence and pathogenesis. Microbiol Mol Biol Rev 2003;67(3):400-28. Staib F. Serum-proteins as nitrogen source for yeast like fungi. Sabouraudia 1965;4:187-93. Schaller M. Invasion of Candida albicans correlates with expression of secreted aspartic proteinases during experimental infection of human epidermis. J Invest Dermatol 2000;114:712-7. Felk A, Kretschar M, Albretch A, Schaller M, Beinhauer S, Nichterlein T, et al. Candida albicans hyphal formation and the expression of the Efg-1 regulated proteinases Sap4 to Sap6 are required for the invasion of parenchymal organs. Infect Immun 2002;70(7):3689-700. Hube B, Monod M, Schofield DA, Brown AJ, Gow NA. Expression of seven members of the gene family encoding secretory aspartic proteinases in Candida albicans . Mol Microbiol 1994;14:87-89. Morrow B, Srikantha T, Soll DR. Transcription of the gene for a pepsinogen PEP1 is regulated by white-opaque switching in Candida albicans . Mol Cell Biol 1992;12:2997-3005. Monod M, Hube B, Hess D, Sanglard D. Differential regulation of SAP8 and SAP9, which encode two new members of the secreted aspartic proteinase family in Candida albicans . Microbiology 1998;144:2731-7. Borg-von Zepelin M, Beggah S, Boggian K, Sanglard D, Monod M. The expression of the secreted aspartic proteinases Sap4 to Sap6 form Candida albicans in murine macrophages. Mol Microbiol 1998;28:543-54. 23 Castrillón Rivera LE y col. 27. Schaller M, Schäfer W, Korting HC, Hube B. Differential expression of secreted aspartic proteinases in a model of human oral candidosis and in patient samples from the oral cavity. Mol Microbiol 1998;29:605-15. 28. Staib P, Kretschmar M, Nichterlein T, et al. Differential activation of a Candida albicans virulence gene family during infection. Proc Natl Acad Sci USA 2000;97:6102-7. 29. Korting HC, Hube B, Oberbauer S, Januschke E, Hamm G, Albrecht A, et al. Reduced expression of the hyphalindependent Candida albicans proteinase genes SAP1 and SAP3 in the efg1 mutant is associated with attenuated virulence during infection of oral epithelium. J Med Microbiol 2003;52(Pt8):623-32. 30. Hube B. Possible role of proteinases in Candida infections. Rev Iberoam Micol 1998;15:68-71. 31. Kaminishi H, Hamatake H, Cho T, Tamaki T, Suenaga N, Jukii T, et al. Activation of blood clotting factors by microbial proteinases. FEMS Microbiol Lett 1994;121:327-32. 32. Tsushima H, Mine H. Cleavage of human big endothelin-1 by Candida albicans aspartic proteinase. FEMS Immunol Med Microbiol 1995;1:69-72. 33. Beausejour A, Grenier D, Goulet JP, Deslauriers N. Proeolytic activation of the interleukin-1β precursor by Candida albicans. Infect Immun 1998;6:676-81. 34. Ghannoum MA. Potential role of phospholipases in virulence and fungal pathogenesis. Clin Microbiol Rev 2000;3:122-43. 35. Leidich S. Cloning and disruption of CaPLB1, a phosfpholipase B gene involved in pathogenicity of Candida albicans . J Biol Chem 1998;73:26078-86. 36. Calderone AR, Fonzi AW. Virulence factors of C. albicans . Trends Microbiol 2001;9(1):327-35. 37. Sther F, Felk A, Gacser A, Kretschmar M, Mahnss B, Neuber K, et al. Expression analysis of the Candida albicans lipase gene family during experimental infections and in patient samples. FEMS Yeast Res 2004;4(4-5):401-8. 38. Saparito I. PHR1, a pH-regulated gene of C. albicans is required for morphogenesis. Mol Cell Biol 1995;15:601-13. 39. Ghannoum MA. Reduced virulence of C. albicans PHR1 mutants. Infect Immun 1995;63:4528-30. 40. Muhlschlegel FA, Fonzi WA. PHR2 of C. albicans encodes a functional homolog of the pH-regulated gene PHR1 with an inverted pattern of pH-dependent expression. Mol Cell Biol 1997;17:5960-7. 41. Fradin C, Kretschmar M, Nichterlein T, Gaillardin VC, d’Enfert C, Hube B. Stage-specific gene expression of Candida albicans in human blood. Mol Microbiol 2003;47(6):1523-43. 42. Chaffin WL. Cell wall and secreted proteins of C. albicans. Identification, function and expression. Microbiol Mol Biol Rev 1998;62:130-80. 43. Sturtevant J, Calderone R. C. albicans adhesins: biochemical aspects and virulence. Rev Iberoam Micol 1997;14:90-97. 44. Hostetter MK. Adhesins and ligands involved in the interaction of Candida spp. With epithelial and andothelial surfaces. Clin Microbiol Rev 1994;7:29-42. 45. Lipke PN. AGα1 is the structural gene for the Saccharomyces cerevisiae α-agglutinin, a cell surface glycoprotein involved in cell-cell interactions during mating. Mol Cell Biol 1989;9:3155-65. 46. Hoyer LL. Identification of Candida albicans ALS2 and ALS4 and localization of Als proteins to the fungal cell surface. J 24 Bacteriol 1998;180:5334-43. 47. Hoyer LL, Hecht JE. The ASL5 gene ofCandida albicans and analysis of the ASL5 gene of Candida albicans and analysis of the Alsp N-terminal domain. Yeast 2001;18:49-60. 48. Kapteyn JC. The cell wall architecture of Candida albicans wild-type cells and cell wall-defective mutants. Mol Microbiol 2000;35:601-11. 49. Hoyer LL. Candida albicans ALS1: domains related to a Saccharomyces cerevisiae sexual agglutinin separated by a repeating motif. Mol Microbiol 1995;15:39-54. 50. Gale CA. Linkage of adhesion, filamentous growth, and virulence in Candida albicans to a single gene. INT1. Science pdf elaborado 1998;279:1355-8. por medigraphic 51. Burman C. Molecular analysis of CaMnt1p, a mannosyl transferase important for adhesion and virulence of Candida albicans . Proc Natl Acad Sci USA 1998;95:7670-5. 52. Banerjee SN, Emori TG, Culver TH. Secular trends in nosocomial primary bloodstream infections in the United States, 1980-1989. National nosocomial infections surveillance system. Am J Med 1991;91:86S-89S. 53. Crump JA, Collignon PJ. Intravascular catheter-associated infections. Eur J Clin Microbiol Infect Dis 2000;19:1-8. 54. Nikawa H. Relationship between thigmotropism and Candida biofilm formation in vitro. Mycopathologia 1998;144:125-9. 55. Donlan RM. Biofilms: microbial life on surfaces. Emerg Infect Dis 2002;8:1-19. 56. Kumamoto AC. Candida biofilms. Curr Opin Microbiol 2002;5:608-11. 57. Baillie GS, Douglas LJ. Role of dimorphism in the development of Candida albicans biofilms. J Med Microbiol 1999;48:671-9. 58. Chandra J, Kuhun DM, Mukherjee PK, et al. Biofilm formation by the fungal pathogen Candida albicans : development, architecture and drug resistance. J Bacteriol 2001;183:5385-94. 59. Baillie GS, Douglas LJ. Matrix polymers of Candida biofilms and their possible role in biofilm resistance to antifungal agents. J Antimicrob Chemother 2000;46:397-403. 60. Baillie GS, Douglas LJ. Candida biofilms and their susceptibility to antifungal agents. Methods Enzymol 1999;310:644-56. 61. Hawser SP, Douglas LJ. Resistance of Candida albicans biofilms to antifungal agents in vitro. Antimicrob Agents Chemother 1995;39:2128-31. 62. Chandra J. Antifungal resistance of candidal biofilms formed on denture acrylic in vitro. J Dent Res 2001;80:903-8. 63. Baillie GS, Douglas LJ. Effect of growth rate on resistance of Candida albicans biofilms to antifungal agents. Antimicrob Agents Chemother 1998;42:1900-5. 64. Ramage G, Vande-Walle K, Wickles BL, Lopez-Ribot JL. Standarized method for in vitro antifungal susceptibility testing of Candida albicans biofilms. Antimicrob Agents Chemother 2001;45:2475-9. 65. Chandra J, Kuhn DM, Mulherjee PK, Hoyer LL, McCormick T, Ghannoum MA. Biofilm formation by the fungal pathogen Candida albicans: development, architecture, and drug resistance. J Bacteriol 2001;183:5385-94. 66. Douglas LJ. Medical importance of biofilms in Candida infections. Rev Iberoam Micol 2002;19:139-43. 67. Douglas LJ. Candida biofilms and their role in infection. Trends Microbiol 2003;11:30-36. 68. Hawser SP, Douglas LJ. Resistance of Candida albicans biofilms to antifungal agents in vitro. Antimicrob Agents Dermatología Rev Mex Volumen 49, Núm. 1, enero-febrero, 2005 Factores de virulencia en Candida sp Chemother 1995;39:2128-31. 69. Mah TFC, O’Toole GA. Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol 2001;9:34-39. 70. Baillie GS, Douglas LJ. Matrix polymers of Candida biofilms and their possible role in biofilm resistance to antifungal agents. J Antimicrob Chemother 2000;46:3499-505. 71. Donlan RM. Biofilms: microbial life on surfaces. Emerg Infect Dis 2002;8:1-19. 72. Ramage G, Wickes LB, Lopez-Ribot JL. Biofilms of Candida albicans and their associated resistance to antifungal agents. Am Clin Lab 2001;20(7):42-44. 73. Ramage G, Bachmann S, Patterson TF, Wickes BL, LopezRibot JL. Investigation of multidrug efflux pumps in relation to fluconazole resistance in Candida albicans biofilms. J Antimicrob Chemother 2002;49:973-80. 74. Jabra-Rizk MA, Falkler WA, Meiller TF. Fungal biofilms and drug resistance. Emerg Infect Dis 2004;10(1):14-19. 75. Ramage G, Vande WK, Wickes LB, Lopez-Ribot JL. Biofilm formation by Candida dubliniensis. J Clin Microbiol 2001;39(9):3234-40. EVALUACIÓN 1. Se consideran factores de virulencia en Candida, excepto: a) Síntesis de adhesinas b) Formación de biopelículas c) Dimorfismo d) Secreción de enzimas hidrolíticas e) Formación de cápsula 2. La penetración de tejidos por la hifa: a) Ocurre durante fases tempranas de la infección b) Depende de la presencia de matriz exopolimérica c) Depende de la secreción de enzimas hidrolíticas d) Requiere señales morfogenéticas pdf elaborado por medigraphic e) Es característico de cepas mutantes de Candida 3. a) b) c) d) e) La filamentación de Candida se produce: En estados tardíos de colonización En presencia de suero a pH alcalino Sólo en infecciones sistémicas En cepas de Candida no patógenas En estados tempranos de la colonización 4. ¿Cuál de los siguientes hallazgos que apoyan que la filamentación es un factor de virulencia es incorrecto? a) La virulencia se incrementa con mutantes que no forman hifa b) Los tubos germinativos son más adherentes c) La fagocitosis de hifas induce la muerte de los macrófagos d) La presencia de suero induce la formación de filamentos e) La virulencia se atenúa con mutantes que no forman hifas 5. Las diferentes fases de la morfogénesis de Candida implican los siguientes episodios, excepto: Dermatología Rev Mex Volumen 49, Núm. 1, enero-febrero, 2005 a) b) c) d) e) Alteraciones en la pared celular Modulación de factores de virulencia Cambios en la polaridad de crecimiento Cambios en el ciclo celular Sus mecanismos de transducción de señales no se modifican 6. En relación con el reconocimiento diferencial de las células dendríticas hacia Candida ¿cuál aseveración es incorrecta? a) Las células dendríticas pueden reconocer las diferentes formas del hongo b) El reconocimiento de la forma de levadura induce respuestas TH1, por lo tanto protectoras c) El reconocimiento del hongo por estas células induce respuestas apoptóticas d) El reconocimiento de la forma de hifa induce respuestas TH2, por lo tanto no protectoras e) Las células dendríticas cuentan con diferentes receptores que le permiten reconocer a los diferentes morfotipos de Candida 7. El gen EFG1 de Candida: a) Codifica para las enzimas proteolíticas (Sap) b) Codifica para un regulador transcripcional que participa en la morfogénesis c) Es el principal componente de virulencia de Aspergillus nidulans d) Se expresa únicamente en infecciones orales e) Codifica para las adhesinas que participan en la formación de biopelículas 8. Las enzimas aspartil- proteasas (o Saps) de Candida: a) Son reguladas por cinco genes b) Se expresan de forma secuencial en infecciones gastrointestinales c) Las mutantes de estos genes han demostrado que 25 Castrillón Rivera LE y col. disminuyen su capacidad de virulencia d) Su participación como factores de virulencia se debe a su capacidad de adherirse a la superficie de las células del huésped e) Están presentes en levaduras patógenas y no patógenas 9. La asociación de las Saps como factores de virulencia en Candida se ha confirmado por los siguientes hechos, excepto: a) Hay modulación de la virulencia con el uso de inhibidores específicos de estas enzimas b) Hay expresión simultánea de estas enzimas con la de las fosfolipasas c) Las mutantes deficientes de estas enzimas son menos virulentas d) Hay correlación de su virulencia con la morfogénesis e) Se observa la expresión de estas enzimas durante el proceso infeccioso 10. Los datos que demuestran la regulación combinada entre el polimorfismo de Candida y los genes SAP se observan con las siguientes aseveraciones, excepto: a) Las mutantes del gen EFG1 tienen capacidad disminuida para formar hifas y secretar enzimas Saps b) Las cepas mutantes de SAP pueden modificar su capacidad invasiva c) El tigmotropismo está relacionado con la producción de enzimas Saps d) Las cepas que producen hifa y tienen deficiencias en su producción de Saps son menos invasivas e) Las Saps se expresan indistintamente del morfotipo de Candida 11. En relación con las características de las aspartilproteasas, ¿cuál aseveración es incorrecta? a) Saps 1, 2 y 3 tienen valor de pH óptimo de 2 a 5 b) Saps 2, 4, 6 y 7 se expresan principalmente en infecciones sistémicas c) Las Sap 1 a 6 tienen un papel decisivo en enfermedades invasivas d) Las Sap 4 a 6 son características del morfotipo de hifa e) La Sap 8 está regulada por acción de la temperatura 26 12. La Sap2: a) Tiene capacidad hidrolítica en las proteínas de la matriz extracelular b) Tiene capacidad de hidrolizar proteínas del sistema inmunitario, como las IgM c) Su valor de pH óptimo es 7.0 d) Su naturaleza química permite anclarse en la pared celular del hongo e) Está ausente durante las fases iniciales de la infección pdf elaborado por medigraphic 13. Son características de la expresión del gen PH1, excepto: a) Es un gen que se expresa diferencialmente según las condiciones del ambiente b) Codifica para una glucoproteína de superficie c) Se expresa a valores de pH de 4.0 d) Su principal función se asocia con la síntesis de pared celular e) En la vagina este gen no es activo 14. ¿Qué información de las adhesinas de Candida es incorrecta? a) Son capaces de unirse a fibrinógeno, fibronectina y colágeno b) Son biomoléculas que permiten su unión a las células del huésped c) Als, Intp y Mnt1 son ejemplos de estas moléculas d) Pueden ser reguladas diferencialmente por distintas condiciones del medio de cultivo e) Son glucoproteínas de secreción 15. La Als de Candida albicans: a) Incluye una familia de quince genes para su expresión b) Los genes ALS comparten una secuencia con la aglutinina de Aspergillus nidulans c) Son proteínas altamente glucosiladas d) Por estudios de ubicación de estas moléculas se demuestra que se encuentran en el citoplasma e) Tienen secuencias internas de repetición de tres aminoácidos 16. La adhesina Hwp de Candida albicans: a) Se codifica por un gen expresado por la familia ASP b) Esta molécula sirve como sustrato de las transglutaminasas Dermatología Rev Mex Volumen 49, Núm. 1, enero-febrero, 2005 Factores de virulencia en Candida sp c) Es una proteína de membrana tipo II que se requiere para la manosilación d) Las cepas que tienen deleción de este gen tienen deficiencias en filamentación y crecimiento en lechetween e) Codifica para una manoproteína de secreción c) Consta de cuatro fases: temprana, intermedia, madura y tardía d) La presencia de la matriz exopolimérica se aprecia hasta la fase madura e) El anclaje de la forma filamentosa a la superficie inicia el proceso 17. La descripción de una biopelícula es: a) Comunidad de microorganismos unidos irreversiblemente a una superficie formada por levaduras e hifas embebidas por una matriz exopolimérica b) Comunidad de microorganismos compuesta de células de levadura que se adhieren a la superficie y que están rodeadas por una matriz c) Comunidad de microorganismos unidos a una superficie y cuyos integrantes mantienen sus características fenotípicas idénticas d) Comunidad de microorganismos unidos reversiblemente a una superficie y que está formada por hifas y levaduras embebidas por una matriz exopolimérica e) compuesta por células filamentosas pdf Comunidad elaborado por medigraphic que se adhieren a una superficie y que se rodea por una matriz 19. La morfogénesis en las biopelículas se dispara cuando: a) Los filamentos sintetizan adhesinas que permiten la unión a la superficie b) Cuando se inicia la síntesis de matriz exopolimérica c) La levadura hace contacto con la superficie y hace el anclaje en la base d) Cuando se alcanza el máximo nivel de expresión de proteasas e) Cuando la hifa expresa el gen EFG1 18. En relación con las fases de formación de biopelículas de Candida, ¿cual aseveración es la correcta? a) El orden de aparición de las fases es: adhesión, crecimiento, maduración y proliferación b) En estudios in vitro se aprecia la estructura completa de 38 a 72 horas Dermatología Rev Mex Volumen 49, Núm. 1, enero-febrero, 2005 20. Los mecanismos de resistencia a los antimicrobianos inducidos en la formación de biopelículas pueden explicarse por los siguientes hechos, excepto: a ) Se producen cambios en el fenotipo de las células b) Hay diferencias en la velocidad de crecimiento c) Hay expresión de genes de resistencia por las células que hacen contacto en la superficie d) Hay penetración restringida a través de la matriz de la biopelícula e) Las células filamentosas inducen genes de resistencia (bombas de eflujo) 27