Anexo VI: Informe Universidad de Aberdeen
Anuncio

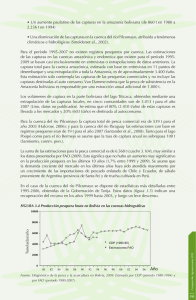
Índice: 1. Introducción 2. Material y Métodos 2.1 Esfuerzo de muestreo y área de estudio 2.2 Métodos de muestreo 2.3 Análisis de datos 3. Resultados: 3.1 Esfuerzo de pesca, capturas, captura accidental y pérdidas debidas a la depredación 3.2 Capturas y depredación en campañas sin observadores científicos a bordo de palangreros 3.3 Capturas, captura accidental y depredación en campañas con observadores científicos a bordo de palangreros 3.4 Recogida de datos acústicos durante las campañas con observadores científicos a bordo de los palangreros 3.5 Avistamientos de cetáceos registrados por observadores científicos en palangreros 3.6 Depredación 4. Discusión: 5. Conclusiones: 6. Agradecimientos: 7. Bibliografía: 1 RESULTADOS DEL ESTUDIO A PEQUEÑA ESCALA DE LAS INTERACCIONES DE CETÁCEOS Y PESQUERÍAS DE PALANGRE EN LAS AGUAS DEL ATLÁNTICO: RESULTADO DE LAS CORRELACIONES AMBIENTALES CON LAS CAPTURAS Y LAS INCIDENCIAS DE DEPREDACIÓN G. Hernández Milián(1); Goetz, S.(2); Varela Dopico, C.(3,4); Rodríguez Gutiérrez, J.(4); Romón, J.(4); Fuertes Gamundi, J. R.(4); Ulloa, E.(4); Tregenza, N.J.C.(5); Smerdon, A.(6); Otero, M.G.(7); Tato, V.(7); Wang, J.(1); Santos, M.B.(2); López, A.(8); Lago, R.(8); Portela, J. (2); Pierce, G. J.(1,2) 1) University of Aberdeen; Tillidrone Avenue, AB24 2TZ, Aberdeen, UK. E-mail: [email protected] 2) Instituto Español de Oceanografía, Centro Oceanográfico de Vigo; Cabo Estai, Canido, 36200 Vigo, Spain 3) Estación de Biología Mariña da Graña, University of Santiago de Compostela, Casa do Hórreo e Casa da Estrela, Rúa da Ribeira 1-4, A Graña 15590, Ferrol, Spain 4) Cooperativa de Armadores de Pesca del Puerto de Vigo s/n, Sociedad Coop. Ltda.; Puerto Pesquero, Edif. Cooperativa de Armadores, Apdo. 1078 36202 Vigo, Spain 5) Chelonia Limited, Beach Cottage; 5 Beach Terrace, Long Rock, Cornwall, TR20 8JE, 6) Aquatec Group Ltd.; High Street, Hartley Wintney, Hampshire, RG27 8NY, UK 7) MG Otero Consultores SL, Porto Pesqueiro, Nvo Tinglado Xeral Empaque Of 8, PO UK Box 1132. 36202 Vigo, Spain 8) CEMMA Apdo. 15, 36380 Gondomar, Pontevedra, Spain Resumen Se presentan los resultados de las campañas realizadas en las aguas del Atlántico central tropical y del oeste y noroeste del archipiélago de las Azores, dentro del proyecto financiado por la Secretaría General de Pesca Marítima Española “Desarrollo de un prototipo acústico para la reducción de la depredación ocasionada por mamíferos marinos en las pesquerías de pez espada”. En el Atlántico, las pérdidas económicas han sido registradas para pesquerías de palangre de tiburones, pez espada y túnidos debido a la depredación por cetáceos. Hemos examinado las interacciones de los cetáceos odontocetos con las pesquerías de palangre que faenaron en aguas cercanas a Brasil y el archipiélago de Azores 2 durante 2006-07, analizando las relaciones entre las capturas, la depredación sobre las capturas, los avistamientos de cetáceos, los registros acústicos de la presencia de cetáceos y las variables ambientales. Los datos fueron obtenidos a través de la información facilitada por los patrones de seis de los barcos y por observadores científicos en dos de los barcos. El porcentaje de lances depredados por cetáceos fue bajo (variando entre 1 y 9 % del total de lances por barco) pero la proporción de captura dañada fue elevada (hasta un 100%) cuando la depredación ocurrió. Las capturas estaban relacionadas con la fase lunar, la cobertura de nubosidad, temperatura superficial del mar y profundidad del área de pesca mientras que los avistamientos de cetáceos estuvieron principalmente relacionados con las capturas. En particular, se encontró una asociación positive entre los avistamientos de Delphinus delphis y las capturas de pez espada y los avistamientos de Stenella frontalis y las capturas de marrajo. La detección acústica fue muy baja cuando se registró depredación por falsa orca aunque elevada proporción de chasquidos fue detectada cuando los delfínidos fueron avistados y las falsas orcas capturadas accidentalmente. Esto podría indicar que las falsas orcas no están ecolocalizando mientras se alimentan del pescado capturado en los anzuelos del palangre. Palabras clave: cetáceos, falsa orca, pesquería de palangre, depredación, hidrófonos, comportamiento, modelos de hábitat Abstract: RESULTS OF A SHORT STUDY OF INTERACTIONS OF CETACEANS AND LONGLINE FISHERIES IN ATLANTIC WATERS: ENVIRONMENTAL CORRELATES OF CATCHES AND DEPREDATION EVENTS Results of comercial longline fisheries operating in the Central Tropical Atlantic waters and west-north west of Archipiélago of Azores, within the proyect financed by the Secretaría General de Pesca Marítima Española “Depredation by killer whales ( Orcinus orca ) and false killer whales ( Pseudorca crassidens ) on European longline fisheries: evaluation and mitigation of economic impact”. In the Atlantic, economic losses have been reported from shark, swordfish and tuna longline fisheries due to depredation by cetaceans. We examined interactions of odontocete cetaceans with commercial longliners operating in waters off Brazil and 3 the Azores archipelago during 2006-07, analysing relationships between catches, depredation on hooked fish, cetacean sightings, acoustic records of cetacean presence and environmental variables. Data were provided by skippers of six vessels and by on-board observers for two vessels. The percentage of longline sets depredated by cetaceans was low (ranging from 1 to 9 % of total sets per ship) but the proportion of fish damaged was high (up to 100%) when depredation occurred. Catches were related to the phase of the moon, cloud cover, sea surface temperature and water depth whereas cetacean sightings were primarily related to catches. In particular there was a positive association between Delphinus delphis sightings and catches of swordfish and Stenella frontalis sightings and mako catches. Acoustic detection was low when depredation by false killer whales occurred although high rates of clicks were detected when delphinids were sighted and false killer whales were by-caught. This may indicate that false killer whales are not echolocating when feeding on fish hooked on a longline. Keywords: cetaceans, false killer whale, longline fishery, depredation, hydrophones, behaviour, habitat modelling 4 1. Introducción: La flota comercial palangrera de altura que opera en las aguas pelágicas del Atlántico tiene como principales especies objetivo los túnidos, el pez espada, distintas especies de la familia Istiophoridae (marlines, agujas, pez vela) y diferentes especies de tiburones (quenlla o tiburón azul, marrajo) (Brothers et al. 1999a). Las capturas de estas especies en el océano Atlántico no se han visto incrementadas en la última década, al contrario de lo ocurrido con las capturas que se realizaron en el océano Pacífico (ICCAT 2007). Las áreas de pesca palangrera de altura más importantes en el océano Atlántico se concentran en la zona central tropical del Atlántico sur y en el oeste-noroeste del archipiélago de Azores (Lewison et al. 2004). Las pesquerías palangreras de altura españolas cuya especie objetivo son los túnidos y el pez espada constituye una de las más importantes en el Atlántico (ICCAT 2007), y supone un importante recurso económico para el sector pesquero español (Garza Gil et al. 2003). Los palangreros comerciales utilizan artes cuya longitud aproximada es de 50 millas de largo y donde cuelgan entre 1200 a 1250 anzuelos de media. Las actividades de estos barcos palangreros favorecen la atracción de otras especies acompañantes como los mamíferos marinos, en particular odontocetos, ya que proveen de un recurso alimenticio de gran tamaño que es de fácil acceso. La eliminación y destrozo de la captura, así como la eliminación de la carnada, por parte de los cetáceos es una de las pérdidas más importantes que se da en las pesquerías palangreras en todo el mundo (Northridge 1984; Dahlheim 1988; Ashford et al. 1996; Capdeville 1997; Dalla Rosa y Secchi 2002, 2007; Donoghue et al. 2002; Gilman et al. 2006; Zollett y Read 2006; Ramos-Cartelle y Mejuto 2007). Algunos autores han sugerido que estas especies habrían podido desarrollar cierta familiaridad con el sonido producido por estas pesquerías (como el sonido del motor, el sonido producido por el palangre en la virada y los equipos electrónicos) y así, como se ha observado frecuentemente, seguir durante días a estos barcos y obtener un beneficio de las capturas (Gilman et al. 2006; Ramos-Cartelle y Mejuto 2007). Además, Gilman et al. (2007) han sugerido que las tasas de depredación tienden a ser mayores cuanto más tiempo permanece el arte en el agua, favoreciendo la disponibilidad de alimento para estos animales. 5 Las interacciones entre las orcas (Orcinus orca, Linnaeus 1758) y las falsa orcas (Pseudorca crassidens, Owen 1846) y las flotas palangreras, que tienen como especies objetivos los túnidos y el pez espada, son comunes en las aguas tropicales y subtropicales del océano Atlántico (Dalla Rosa y Secchi, 2002, 2007; Dalla Rosa et al. 2006; Ramos-Cartelle y Mejuto 2007). Ambas especies de cetáceos tienen una distribución mundial, aunque las orcas presentan cierta preferencia por aguas más frías y cercanas a costa (normalmente en los primeros 800 km costeros, concentrándose en la plataforma continental); en cambio, las falsas orcas suelen localizarse en aguas templado-cálidas, subtropicales y tropicales, más alejadas de costa y donde las aguas son más profundas. Ambas especies se alimentan preferentemente de pescado, cefalópodos y otros mamíferos marinos (Jefferson et al. 1993; Stacey et al. 1994; Carwadine 1995). Las características ambientales y oceanográficas, tales como la temperatura del agua, la batimetría, los frentes oceánicos, los ciclos lunares, así como factores espacio-temporales, parecen jugar un importante papel en la distribución tanto de los cetáceos como de sus presas (Damalas et al. 2007; Romo et al. 2007; De Stephanis et al. 2008). El estudio de los mamíferos marinos lejos de costa suele realizarse mediante el registro de avistamientos por observadores científicos a bordo de barcos. Sin embrago, la utilización de métodos acústicos pasivos, como por ejemplo la disposición de T-PODs (www.chelonia.com), puede aumentar el rango de detección de estos animales; esto es especialmente importante cuando las condiciones de visibilidad son bajas o los animales sólo salen a la superficie durante cortos períodos de tiempo, ya que el rango de detección será mayor que si sólo se recogieran los datos visuales o de avistamientos (Cartensen et al. 2006; Leeney y Tregenza 2006; Philpott et al. 2007). El presente estudio pretende describir las interacciones de las pesquerías de palangre de túnidos y pez espada en dos áreas del Atlántico (Brasil y Azores) con los cetáceos, en particular las interacciones producidas con la orca y la falsa orca, determinando las relaciones entre las capturas, la presencia de cetáceos, el índice de depredación y las variables ambientales disponibles. Específicamente se pretende responder a las siguientes cuestiones: (1) ¿existe alguna relación entre las capturas y 6 las condiciones ambientales en las distintas zonas analizadas?, (2) ¿hay alguna relación entre la presencia de cetáceos y/o la incidencia de depredación con determinadas condiciones ambientales?, (3) ¿hay alguna relación entre la depredación y la presencia de determinadas especies de cetáceos? ¿Está esto relacionado con la captura total obtenida? Por ultimo, debido a que la captura accidental de tortugas es bastante frecuente, se analizó la posible relación entre la captura accidental (by-catch), el total de capturas y las condiciones ambientales. 2. Material y Metodos: 2.1 Esfuerzo de muestreo y área de estudio Durante el período de un año (junio de 2006-junio de 2007) se registraron los datos de ocho barcos de palangre de superficie españoles de pesca comercial que estuvieron operando en dos áreas del océano Atlántico: siete de los barcos palangreros centraron sus actividades pesqueras en aguas cercanas a la costa de Brasil extendiéndose por las aguas centro Atlánticas (V1-V7), mientras que el octavo (V8) se centró en la zona oeste del archipiélago de las Azores. Sólo los barcos V7 y V8 llevaron observadores científicos a bordo. Ver figura 1. El área de pesca en el Atlántico central y sur se encuentra dominada por la corriente ecuatorial del sur que, al llegar a la costa de Brasil, se divide en dos formando, hacia el norte, la corriente norte de Brasil y, hacia el sur, la corriente de Brasil. La corriente de Brasil se caracteriza por ser una corriente débil en su extremo más occidental, donde lleva aguas subtropicales con temperaturas entre 18ºC y 28ºC; discurre por toda la costa oriental de Brasil desde los 9º de latitud sur hasta los 38º de latitud sur, quedando confinada a los primeros 600 metros de la columna de agua (Memery et al. 2000; Zavilov et al. 1999). La pesquería más importante en las aguas afectadas por la corriente de Brasil está constituida por los túnidos, donde el Thunnus albacares adquiere mayor importancia en la región sur y el T. alalunga en la región norte (Zavala-Camin &yAntero da Silva 1991). Estas especies son capturadas preferentemente en áreas donde los montes submarinos y los bancos de arena son dominantes. 7 La otra área de pesca estudiada se ubica en la zona oeste de las islas Azores. El archipiélago de las Azores está compuesto por un grupo de 9 islas volcánicas situadas en la cordillera centro Atlántica entre las latitudes 37º y 41º norte y las longitudes 25º y 31º oeste, cuyas aguas están dominadas por la corriente del Golfo. La riqueza de los recursos pesqueros en las aguas cercanas a las Azores se debe a una compleja relación caracterizada por una alta temperatura y salinidad del agua, un bajo contenido en nutrientes (Johnson y Stevens 2000) y una serie de ecosistemas influenciados por montes submarinos. Las especies pesqueras objetivo son, fundamentalmente, túnidos y pez espada, aunque algunas especies de tiburones – principalmente quenlla o tiburón azul (Prionace glauca) y marrajo (Isurus oxyrinchus)– pueden llegar a superar en número a las capturas de pez espada realizadas en palangre en una proporción de 10:1 (Morato et al. 2001). 2.2 Métodos de muestreo Los datos de las capturas realizadas por seis de los barcos palangreros (V1-V6) estudiados que operaron en el Atlántico central y sur fueron facilitadas por los propios patrones. Además, se registraron los datos de la actividad pesquera, avistamientos de cetáceos, registros acústicos de odontocetos, depredación detectada sobre la captura y variables ambientales de dos palangreros con observadores científicos a bordo centrados en cada una de las áreas estudiadas (V7 en aguas cercanas a Brasil y V8 al oeste de Azores) (tabla 1 y 2). Los datos tomados durante la actividad pesquera incluyen la hora y la posición de los lances, el número de anzuelos por línea, el total de la captura (tanto en número como en biomasa de pesca por especie) y la captura accidental de mamíferos marinos y/o tortugas. Las especies de peces capturadas fueron clasificadas para el análisis posterior como: pez espada (Xiphias gladius), marrajo (Isurus oxyrinchus), quenlla o tiburón azul (Prionace glauca), atún (Thunnus spp., principalmente T. alalunga, T. albacore y T. Obesus), peces de la familia Istiophoridae (marlines, agujas, picudos y peces vela), dorado (Coryphaena hippurus) y barracuda (Sphyraena spp.). 8 La depredación causada por cetáceos, tiburones o tortugas se clasificó según el tipo de marcas encontradas en el pescado (ver figura 2). Los tiburones suelen morder el animal a la mitad del cuerpo, dejando las marcas de una mordedura limpia, o realizan numerosos mordiscos relativamente pequeños. En cambio la orca y la falsa orca arrancan el cuerpo del animal, dejando mordeduras con los bordes rugosos o dejando, a menudo, solamente los labios y la mandíbula superior del pescado en el anzuelo (Secchi & Vaske 1998; Donoghue et al. 2002; Gilman et al. 2006; VarelaDopico, obs. pers.). Los patrones han denunciado en muchas ocasiones que los cetáceos pueden llegar a eliminar totalmente el pescado del anzuelo. Por otro lado, las tortugas dejan numerosos mordiscos de pequeño tamaño en el pescado, comiéndose principalmente las partes comerciales del mismo (Varela-Dopico, obs. pers.). Mediante una cámara digital compacta, se tomaron fotos de los peces depredados, así como de otros peces capturados. Los datos ambientales recogidos por los observadores fueron los siguientes: el estado del mar según la escala de Douglas (Sea), la cobertura nubosa en una escala del 0 al 8 (Cd), la fase lunar (M1: luna nueva, M2: luna creciente, M3: luna llena, M4: luna menguante), la temperatura superficial del mar (SST) y la profundidad de la zona (Depth). Además, la batimetría y la línea de costa fue obtenida a través del programa GEBCO Atlas 2003, y el mapa fue realizado mediante el software Arc/View 3.3 (figura 1). Los avistamientos de cetáceos se realizaron mediante localización visual primero, para posteriormente ser identificados a través de prismáticos con una óptica de 10x42. Estos datos fueron recogidos durante toda la actividad pesquera por los observadores científicos en los barcos V7 y V8 anotando las coordenadas del momento del avistamiento, el número de animales y la especie avistada; mientras que en los barcos sin observadores científicos a bordo (V1-V6) los datos fueron recogidos por los patrones de los barcos y considerados como oportunistas. Las especies y categorías identificadas fueron las siguientes: cachalote (Physeter macrocephalus), orca (Orcinus orca), falsa orca (Pseudorca crassidens), delfín común (Delphinus delphis), delfín moteado del Atlántico (Stenella frontalis), Odontocetos no identificados y Misticetos. 9 Para el registro de sonidos acústicos producidos por los cetáceos se utilizaron hidrófonos de detección acústica pasiva (T-PODs –figura 3– diseñados por el Dr. Nick J.C. Tregenza de Chelonia Ltd., www.chelonia.co.uk). Se dispuso de 4 T-PODs de la versión 5 en cada campaña, de los cuales se colocaron entre uno y tres por lance (figura 4, tabla 3), uno a cada extremo de la línea y otro en la zona media, a una profundidad aproximada de 5 m a lo largo de la línea de palangre. Para evitar posibles pérdidas del aparato en caso de corte de la línea madre, se colocaron unidos a un gallo o boya (figura 5). Los T-PODs registran los chasquidos (clicks) y los trenes de chasquidos (train clicks) producidos por los cetáceos para los que se ha programado el hidrófono y que se encuentran en la zona. Estos aparatos están constituidos por un hidrófono, un procesador analógico, un sistema de registro de chasquidos ecolocalizados y un software capaz de filtrar los chasquidos producidos por los cetáceos dentro de una banda de frecuencia determinada. Se programaron para detectar los chasquidos emitidos por falsa orca cuando se realizaran los lances, de manera que se ajustaron los valores del aparato a los calculados con anterioridad por Madsen et al. (2004) para falsa orca en libertad (filtro A = 41 kHz, filtro B = 16 kHz, banda de frecuencia= 4-6, sensibilidad = varias, duración mínima de chasquido = 40μs). Los hidrófonos son capaces de registrar el número de chasquidos producidos por los odontocetos y clasificarlos en función de que la probabilidad de que procedan de la especie para la que se han ajustado los parámetros (en el caso de este estudio para falsa orca) sea alta, baja, dudosa o muy dudosa (tabla 4). Los T-PODs se programaron con los parámetros preestablecidos en Pseudorca 41 selectivity trial.pds, Pseudorca 41 guess.pds y, en algún caso, Pseudorca frequency trial.pds. Estos tres archivos establecen parámetros para el hidrófono de forma que recoja los trenes de chasquidos de ecolocalización de la misma banda de frecuencia en la que la falsa orca (Pseudorca crassidens) emite para detectar a sus presas. P. crassidens produce sonidos de ecolocalización dentro de la banda de frecuencia de 30-70 kHz (Madsen et al. 2004). Sin embargo, se superpone a la banda de frecuencia de otras especies de delfínidos, como Delphinus delphis cuyo pulso de frecuencia se encuentra entre 20–100 kHz (Wood & Evans 1980) y Stenella spp. que emiten chasquidos entre 30-85 kHz (Lammers et al. 2003) o muestran un espectro de chasquidos bimodal con picos en las bandas de frecuencia 40-60 kHz y 120-140 10 kHz (Schotten et al. 2003). Por otro lado, el grado de especificidad de especies en las detecciones depende también de las especies presentes en el área de estudio. Al finalizar la virada de cada lance, se descargaron los datos recogidos por los TPODs y se almacenaron en un ordenador portátil para su posterior análisis. Debido a problemas técnicos, no todos los T-PODs colocados en la campaña realizada en las aguas de Brasil funcionaron; sin embrago, el la campaña de Azores todos los T-PODs funcionaron correctamente y debido a las malas condiciones ambientales no se largó en uno de los lances. Los registros acústicos recogidos por los T-PODs en cada lance, son clasificados por el programa del hidrófono como chasquidos probablemente emitidos por falsa orca (“positivos”, corresponden a la suma de las categorías “CetHi” y “CetLo”, o también conocidas como “CetAll”, ver www.chelonia.co.uk), y chasquidos considerados como dudosamente procedentes de falsa orca (“?” y “??”, cuya suma correspondió a los denominados “negativos”) (figura 6). También, se calcularon otros dos índices de actividad de cetáceos (Tougaard et al. 2004; Skov et al. 2002; Leeney & Tregenza 2006): la media proporcional de chasquidos (número de chasquidos positivos dividido por el total de minutos que el hidrófono registró sonidos) e intensidad de chasquidos (media del número de chasquidos positivos durante los minutos en que se registraron chasquidos). Ambos índices fueron calculados para cada hidrófono y cada lance. 2.3 Análisis de datos Para el análisis de los datos, se utilizó la composición de la captura, la incidencia y cantidad de animales depredados por captura, los avistamientos de cetáceos, los sonidos emitidos por los cetáceos, y los datos ambientales (ver tabla 5). Primeramente, se realizó una exploración de las variables utilizando el análisis de correlación de Spearkman, tratando los datos de cada lance como una muestra única y analizando los datos de cada barco independientemente. Posteriormente, para los dos barcos con observadores científicos a bordo (V7 y V8), se realizó un análisis más profundo para detectar las relaciones entre las variables respuesta y las variables explicativas. Para ello se utilizaron los análisis de 11 redundancias (RDA) y los análisis de regresión de Modelos Aditivos Generalizados (GAMs) y Modelos Lineales Generalizados (GLMs). El análisis de RDA es un análisis multivariante que indica la proporción de la variación de la variable respuesta explicada por la variable explicativa. La significación estadística de los efectos de las variables explicativas se obtuvieron mediante la prueba de permutación Monte Carlo para n=4999 permutaciones. La relación entre las variables respuesta y explicativa se disponen mediante un gráfico de dos entradas de puntos y vectores (ver Zuur et al, 2007). Los datos recogidos para ambas áreas (Brasil y Azores) fueron analizadas de manera independiente y, para ambos, se utilizaron como variables respuesta en el análisis de RDA las (a) la cantidad de captura (en número) de pez espada, atún, marrajo y quenlla o tiburón azul y (b) los datos de acústica recogidos por los T-PODs (proporción de chasquidos e intensidad de chasquidos). En el caso de la campaña de Azores, se dispusieron de datos suficientes como para poder tratar también como variables respuesta en el análisis de redundancias (c) los avistamientos de cetáceos (número de animales avistados de Delphinus delphis, Stenella frontalis, Orcinus orca, Pseudorca crassidens, Physeter macrocephalus, Mysticetos y Odontocetos no identificados) y (d) la incidencia de depredación (número de pez espada, marrajo y escolares depredados). En total se llevaron a cabo seis RDA (tabla 6). El resto de variables disponibles fueron tratadas como variables explicativas. Cuando las variables respuesta utilizadas fueron el registro acústico o la incidencia de depredación, los avistamientos de cetáceos (variables explicativa) fueron utilizados en función de la presencia/ausencia de los mismos, ya que se consideró que la presencia/ausencia del avistamiento de los distintos grupos de cetáceos era más importante que el número de animales de cada especie. Cuando los resultados de RDA indicaron una relación significativa entre las variables respuesta y explicativa, se realizó un estudio más profundo mediante el uso de los Modelos Aditivos Generalizados (GAMs) y los Modelos Lineales Generalizados (GLMs). Un modelo aditivo generalizado (GAM) es un modelo de regresión univariante de la forma: 12 yi= α + ƒ1(xi1)+ … + ƒm(xim) + βnxin + … + βpxip+ εi εi ~N (0,σ2) donde yi es la variable respuesta, ƒj() son los suavizadores, βn son los coeficientes para los términos paramétricos (por ejemplo, las falsas variables fueron generadas a partir de las variables categóricas), y ε es el error aleatorio (Zuur et al. 2007). Los modelos se ajustan utilizando un algoritmo de selección de variables (fowards, backwards), eliminando paso a paso las variables que resultan no significativas hasta obtener un modelo con todos los términos significativos. Cuando ninguna de las variables que quedan en los modelos finales es continua o no pueden ser tratadas como continuas, el modelo se ajusta utilizando los modelos lineales generalizados (GLM). Para confirmar la bondad de ajuste del modelo se realizó la gráfica de los residuos. utilizando el paquete Los modelos RDA, GAMs y GLMs fueron realizados estadístico Brodgar 2.5.2 (www.brodgar.com). Más información sobre estás técnicas analíticas puede encontrarse en Zuur et al. (2007). Se analizó cada variable respuesta utilizada en el análisis de RDA con cada uno de los cuatro grupos de variables (capturas, detección acústica, avistamientos y depredación), de manera que se tuvieron en cuenta las relaciones no lineales. En el análisis, las distribuciones de las variables repuesta fueron consideradas como binomiales, cuando las variables eran nominales (presencia/ausencia), o Poisson, cuando las variables eran continuas (número de peces, chasquidos, etc). Además, se incluyó un parámetro de dispersión en aquellos modelos en los que se detectó sobre dispersión. Sólo fue necesaria la transformación en una de las variables (intensidad de chasquidos) para conseguir que la distribución de la misma se aproximara a una distribución Gaussiana, aplicando la transformación logarítmica antes de realizar el análisis. La proporción media de chasquidos tenía una distribución aproximadamente Gaussiana sin transformación previa. En la campaña realizada en aguas cercanas a Brasil, ambos índices acústicos dieron resultados muy bajos y, por tanto, se utilizó el número de chasquidos (“positivos” y “negativos”) como variable respuesta cuando se realizó el análisis de regresión. En los análisis de regresión (GAMs y GLMS) las variables de avistamientos de cetáceos y depredación fueron utilizados en la forma presencia/ausencia; en cambio, la cobertura de nubes (en una escale de 0 a 8) fue tratada como una variable continua. Para evitar un sobre-ajuste 13 en los modelos, se impuso que los grados de libertad de las variables explicativas continuas fueran menores de 5. La captura accidental de tortugas fue elevada durante todas las campañas, incluidas aquellas que no disponían de observadores científicos a bordo. Por ello, se analizaron los posibles factores que podrían estar relacionados con dicha interacción (condiciones ambientales y los tipos de captura). Se utilizó la prueba de KruskalWallis para comparar la captura obtenida por cada barco y las condiciones ambientales, cuando fue posible, con los lances con y sin captura accidental de tortugas. 3. Resultados: 3.1 Esfuerzo de pesca, capturas, captura accidental y pérdidas debidas a la depredación El esfuerzo de pesca realizado se centró en el área afectada por la Corriente Ecuatorial del Sur (V1-V7) y la Corriente del Golfo (V8) (figura 1). Se realizaron un total de 635 operaciones de pesca en el período de junio de 2006 a julio de 2007 (793 días de campaña en total), con una duración media de 15.68 h por lance, una media de 1275 anzuelos por línea y con una captura total de 1185.5 toneladas (1.4 toneladas de media por barco y lance) de pescado comerciable. Las capturas estuvieron dominadas por pez espada, tiburones (principalmente quenlla y marrajo), atún y especies de la familia Istiophoridae (marlines, agujas, picudos y peces vela), en orden decreciente (Figura 7, tabla 2). Las campañas con observadores a bordo difirieron en el tipo de capturas; mientras que la composición mayoritaria de la pesca del palangrero que operó en aguas cercanas a Brasil era de túnidos y pez espada (figura 8), las capturas del palangrero en aguas occidentales de Azores estuvieron compuestas, fundamentalmente, por tiburones (figura 9). La depredación producida por los cetáceos varió entre el 1% y el 9% de los lances (por barco), con una media por barco de 0.2% a 8.6% de pérdidas del total de 14 las capturas. Para las campañas realizadas por los palangreros V1, V2, V3, V4, V5 y V6, se consideró que había incidencia de depredación tanto cuando el pescado se encontró dañado y no comerciable como aquel que fue eliminado del los anzuelos; para las campañas realizadas por los palangreros V7 y V8, sólo se consideró el pescado que se encontró dañado (tabla 1). Las capturas accidentales de mamíferos marinos solo se dieron en un lance de la campaña realizada en Azores, donde dos falsas orcas fueron capturadas en el palangre. Por el contrario, un 11.2% de los lances (por barco) registraron capturas accidentales de tortugas. El número de tortugas capturadas varió entre 1 y 5 por lance en todas las campañas, excepto en la realizada por el barco V3 que no registró ninguna captura accidental. Las especies que sufrieron un mayor número de capturas accidentales fueron la tortuga laúd Dermochelys coriacea, constituyendo el 39% de las tortugas capturadas, y la tortuga verde Chelonia mydas, constituyendo el 31% de las capturas accidentales; las otras especies capturadas fueron la tortuga boba Caretta caretta (19%) y la tortuga lora Lepidochelys olivacea (11 %). La mayoría de las tortugas fueron capturadas vivas y, posteriormente, liberadas (tabla 1). 3.2 Capturas y depredación en campañas sin observadores científicos a bordo de palangreros En las campañas realizadas sin observadores científicos a bordo (V1-V6), los patrones fueron los encargados de registrar el total de captura dañada debido a la depredación por mamíferos marinos. El porcentaje medio de los lances que registraron depredación fue relativamente bajo (4.6%), pero la pérdida de las capturas en estos lances varió desde el 2% hasta el 100% del total de pescado capturado. Además, dos tercios de las ocasiones se registraron pérdidas de las capturas de más del 25%. En el análisis de correlación, en dos de los seis barcos, la incidencia de depredación resultó significativamente negativa respecto a la captura de pesca (V2: r= -0.47, p= 0.001; V3: r= -0.24, p= 0.022) (tabla 1), sugiriendo que la depredación podría estar produciendo una reducción significativa de la captura total de pescado en los lances. 15 3.3 Capturas, captura accidental y depredación en campañas con observadores científicos a bordo de palangreros El total de las capturas de las dos campañas con observadores científicos a bordo (V7 y V8) fue de 136.7 toneladas (3645 individuos) de pescado, donde el 87% (en número de pescados) fue comerciable. Las principales especies capturadas (en número) durante la campaña de Brasil (V7) fueron el atún (54.5%), el pez espada (29.4%) y los marlines y especies relacionadas (7.5%). En cambio, en la campaña realizadas en las aguas cercanas a las Azores (V8), las principales especies capturadas (en número) fueron tiburones (73.6%, quenlla y marrajo fundamentalmente) y pez espada (24.6%). Durante estas dos campañas, de los 86 lances realizados se registraron 19 lances con animales depredados: nueve ocasiones en Brasil y diez en Azores. Sin embargo, al inspeccionar el tipo de depredación producida en el pescado, solo tres de las 19 veces estuvo relacionada con la depredación producida por cetáceos, en particular producida por falsa orca: una en la campaña de Brasil (3.3% de los lances) y dos en la de Azores (3.6% de los lances) (tabla 1). Los otros 16 lances con depredación correspondieron a pescados depredados por tiburones (en doce de las ocasiones) y la atribuida a tortugas (en cuatro ocasiones). En ningún caso se registró depredación por más de un grupo de depredadores. La captura dañada por los cetáceos se dio sólo en un 0.2% en la campaña del Atlántico sur y de un 0.9% en la campaña de Azores. Los cetáceos depredaron principalmente sobre pez espada (85.7% en número), seguido del marrajo (14.3% en número). Los análisis de redundancia indicaron que en la campaña de Brasil, las capturas de las principales especies estaban significativamente relacionadas con la captura accidental de tortugas y la fase lunar (luna menguante) (tabla 6; figura 10). Además, el análisis de modelos lineales generalizados indicó la existencia de una débil relación significativa entre la captura de pez espada y la cobertura nubosa, así como cuando se utilizó como variable la interacción entre la fase lunar y la cobertura nubosa. La gráfica de datos condicionados de co-plot (figura 11) muestra la interacción entre los efectos de la cobertura nubosa y la fase lunar en relación con la captura de pez espada. Por el contrario, cuando se analizaron las capturas de túnidos, el único efecto que resultó significativo (de forma débil también) fue el 16 término de la interacción, con ambos efectos principales (cobertura nubosa y fase lunar) independientes no significativos (tabla 7). Debido a que el tamaño muestral es bastante bajo, no se pudieron hacer investigaciones más profundas en relación a estas interacciones. El resultado del análisis de redundancias (RDA) obtenido para el barco V8, indicó que las capturas de las principales especies comerciales estaban influenciadas por la fase lunar, la profundidad de la zona y la temperatura superficial del mar (tabla 6; figura12). Además, los análisis de regresión GAMs indicaron que las capturas de pez espada estaban relacionadas con la profundidad de la zona, la fase lunar, la cobertura nubosa y los avistamientos de Stenella frontalis (tabla 8). Cabe destacar que las capturas de esta especie comercial presentaron un pico máximo cuando la cobertura nubosa era relativamente baja (valor de 2) (figura 13a) y un pico mínimo cuando la profundidad del área de pesca era de alrededor de 3000m, produciéndose un aumento de las capturas en aguas de mayor y menor profundidad (figura 13b). Las capturas de esta especie también resultaron disminuidas por la presencia de los delfínidos del género Stenella; sin embargo, los delfines moteados no suelen alimentarse de especies de gran tamaño capturadas por el palangre, por lo que la interpretación de dicha relación no ha podido ser aclarada en este estudio. En la campaña de Azores, las variables de fase lunar, la temperatura superficial del mar, la proporción de chasquidos obtenida por los hidrófonos y los avistamientos de Delphinus delphis y Stenella frontalis mostraron una influencia sobre las capturas de marrajo en cada lance (tabla 8); de manera que en aguas de mayor temperatura y con registros acústicos más elevados, las capturas de esta especie de tiburón fueron más bajas (figura 13c). La presencia de delfines influyó de manera contraria en las capturas en función de la especie presente: cuando los delfines comunes estaban presentes, la captura de marrajo tendía a disminuir (relación negativa), mientras que cuando los delfines que se registraban eran los delfines moteados, las capturas de estos tiburones tendían a aumentar (relación positiva). En esta misma campaña, las capturas de quenlla resultaron ser mayores cuando la luminosidad nocturna era mayor (luna llena o cercana a luna llena) (tabla 8). 17 Las capturas accidentales (by-catch) de tortugas se dieron en un tercio de los lances realizados en la campaña V7 (Brasil), donde en 10 de los 30 lances se registró by-catch. En ningún caso se encontró ninguna relación entre esta interacción y las variables ambientales (profundidad del área de pesca, temperatura superficial del mar, fase lunar y estado del mar) o con las capturas de túnidos o pez espada, pero si se observó una incidencia mayor con elevadas capturas de tiburones (prueba de Kruskal-Wallis, P=0.002) y con el grupo de especies clasificado como “otras especies” (prueba de Kruskal-Wallis, P=0.001). Sin embargo, en la campaña realizada en Azores (V8), la captura accidental de tortugas se dio sólo en siete de los 56 lances y se observó una incidencia ligeramente mayor cuando la captura total de marrajos aumentaba (prueba de Kruskal-Wallis, P=0.035). 3.4 Recogida de datos acústicos durante las campañas con observadores científicos a bordo de los palangreros Los T-PODs fueron colocados en el 96.5% (85 lances) de las líneas. Debido a problemas técnicos y de condiciones ambientales adversas, no fue posible colocar tres T-PODs en cada línea, pero, al menos, en el 92% de los lances se pudieron colocar dos o tres hidrófonos para la recogida de datos acústicos (tabla 3). Los TPODs funcionaron durante 3496.4 horas, donde 814.7 horas fueron analizadas en la campaña de Brasil y 2681.7 horas fueron analizadas en la campaña de Azores. En total los T-PODs funcionaron durante el 96% del tiempo que estuvieron en el agua, correspondiendo ese 4% restante a problemas técnicos en los aparatos de acústica. El número de trenes de chasquidos recogidos durante la campaña de Brasil fue muy bajo. El 84.7% fueron clasificados como “positivos”, es decir, que posiblemente fueron emitidos por falsa orca. En la campaña desarrollada en aguas cercanas a Azores, el número de chasquidos registrados fue considerablemente más alto, pero sólo el 28.7% fueron clasificados como “positivos” (tabla 4). En ambas campañas, se detectó una intensidad de chasquidos baja cuando se observó depredación sobre las capturas debido a falsa orca (figuras 14a,b). Sin embargo, en el lance donde aparecieron cinco pez espada depredados por falsa orca y, además, dos falsas orcas fueron accidentalmente capturadas (lance 20), los T-PODs situados más cerca de 18 donde las falsas orcas fueron capturadas registraron una mayor intensidad de chasquidos. El análisis de redundancias realizado para la campaña de Brasil, indicó que el número de chasquidos positivos estaba influenciado por la fase lunar, más concretamente cuando la luna se encontraba en fase creciente (tabla 6; figura 15). El análisis de regresión realizado confirmó el efecto de la fase lunar, pero indicó que los datos acústicos positivos se veían aumentados cuanto mayor era la profundidad de la zona (tabla 7; figura 13d). En esta campaña sólo se detecto depredación por falsa orca en un único lance. La tabla 5 muestra los resultados del análisis de redundancias realizado en la campaña de Azores, donde se muestra una relación significativa entre la detección de chasquidos y los avistamientos de pequeños delfínidos (Delphinus delphis y Stenella frontalis) (figura 13). Sin embargo, los resultados del análisis de regresión GLM indicaron un débil aumento de la intensidad de chasquidos cuando se registraron avistamientos de delfines moteados y la fase lunar se encontraba en fase menguante. No se encontró relación alguna con otros factores ambientales o especies capturadas. También se detectó un relativo aumento de la proporción de chasquidos cuando aumentaba la profundidad de la zona de pesca (tabla 8). 3.5 Avistamientos de cetáceos registrados por observadores científicos en palangreros Los resultados de avistamientos obtenidos por los observadores fueron diferentes en las distintas áreas. En la campaña realizada en las aguas cercanas a Brasil sólo se registraron doce falsas orcas y un cachalote. Debido a este bajo número de avistamientos concentrados en tres lances (lances 13, 16 y 17), no se pudieron realizar análisis en mayor profundidad. 613 cetáceos fueron registrados en la campaña realizada al oeste de las islas Azores, donde el 94% correspondieron a especies de Odontocetos (Stenella frontalis, Delphinus delphis, Pseudorca crassidens, Physeter macrocephalus y Orcinus orca en orden descendiente en número de animales avistados). Durante esta campaña, 19 los hidrófonos registraron picos elevados de datos acústicos en aquellos lances donde se registraron avistamientos de delfínidos, falsa orca y orca. El análisis multivariante mostró que los avistamientos de cetáceos estaban relacionados con las capturas totales de pez espada y marrajo, con la fase lunar, con la proporción de chasquidos obtenida y con la incidencia de depredación (tabla 6; figura 17). Debido al número de animales avistados de cada especie, sólo fue posible realizar modelos de regresión para las dos especies mayormente avistadas (Delphinus delphis and Stenella frontalis). Los resultados de los GLM indicaron que las capturas de pez espada eran mayores y las de marrajo menores cuando se detectaba la presencia de D. delphis. Por el contrario, la presencia de Stenella frontalis estaba asociada con elevadas capturas de marrajo y bajas de pez espada. El registro acústico clasificado como chasquidos “positivos”, también resultó ser más elevado cuando ambas especies estaban presentes en el área. De todas las variables ambientales tomadas, sólo la fase lunar resulto estar relacionada con los avistamientos de una de las especies de delfínidos; el número de Stenella frontalis en la zona eran menos numerosos cuando la fase lunar se encontraba en luna creciente o luna llena (tabla 8). 3.6 Depredación Un total de 24 peces fueron encontrados con signos de depredación, nueve en la campaña realizada por el palangrero V7 y quince en la campaña realizada por el palangrero V8. Sólo la depredación sobre siete de los pescados fue clasificada como posiblemente debida a la falsa orca. Un único pez espada apareció depredado en la campaña de Brasil, por lo que no fue posible realizar análisis más profundos. En la campaña realizada en Azores, la depredación se produjo sobre quince peces y afectó a tres especies: pez espada, marrajo y escolar; pero la depredación por parte de las falsas orcas se dio en seis ocasiones y, fundamentalmente, sobre pez espada. El análisis multivariante de las distintas especies de peces depredados sugirió que la depredación podría estar relacionada con los avistamientos de falsa orca registrados (tabla 6; figura 18). Sin embargo, el análisis de regresión GLM reveló que la incidencia de depredación aumentaba cuando la proporción de 20 chasquidos recogidos por los hidrófonos eran elevados y los avistamientos de delfínidos (D. delphis y S. frontalis) estaban ausentes (tabla 8). 4. Discusión: Las capturas de los palangreros de superficie de aguas lejanas parecen tener una composición similar, aunque en zonas menos tropicales (V1 y V8) los tiburones se convierten en el grupo dominante del total del pescado capturado. Se observó que las capturas de las principales especies de pescados, en las campañas en que observadores científicos recogieron los datos, estaban afectadas por las variables ambientales tales como las condiciones de luminosidad (cobertura de nubes y fase lunar), temperatura del agua y profundidad de la zona donde se realizaba la pesca. Los túnidos, el pez espada y los tiburones (especies objetivo de las pesquerías estudiadas) son especies oceánicas migratorias, que suelen encontrarse fundamentalmente en aguas superficiales templadas que están influenciadas por frentes oceánicos y zonas de afloramiento (Collette & Nauen 1983; Bigelow et al. 1999; Brill et al. 1999; Dagorn et al. 2000; De Stephanis et al. 2008). Los palangreros, que tienen estas especies como especies objetivo, largan las líneas con la puesta de sol, dejando el palangre toda la noche y recogiéndolo al amanecer, beneficiándose del comportamiento migratorio nictimeral de estas especies, que ascienden a las capas más superficiales por la noche para alimentarse mientras que durante el día permanecen en capas más profundas (Nakamura 1985; Bigelow et al. 1999; Domokos et al. 2007). La fase lunar ejerce un efecto importante sobre las capturas de pez espada y tiburones, al igual que se ha demostrado en otros estudios en otras áreas del mundo como el Pacífico (Pallares y Garcia-Mamolar 1985; Bigelow et al. 1991) y el Mediterráneo (Damalas et al. 2007). Estudios realizados en el Atlántico sur demostraron que cuando la luminosidad (dependiendo de la fase lunar y la cobertura nubosa) la captura accidental de aves marinas en el palangre aumentaba (Cherel et al. 1996; Brothers et al. 1999a, 1999b); es posible que, con una mayor luminosidad (luna llena y fases cercanas) y una cobertura nubosa baja la carnada de las líneas se 21 haga más visible y, por tanto, atraiga más peces y otras especies acompañantes (aves, tortugas y/o mamíferos marinos) al arte de pesca (Hutchinson, 1996). Las capturas de pez espada fueron menores cuando la profundidad de la zona de pesca era de 3000 metros pero, en cambio, aumentaban cuando el área de pesca tenía menor o mayor profundidad. Bigelow et al. (1999) encontraron que la CPUE de pez espada en el océano Pacífico disminuía cuando la pesca se realizaba en zonas donde la profundidad oscilaba entre 2000-3000 m, y aumentaba con profundidades mayores de 3000m. Los resultados obtenidos sugieren que en el Atlántico podría darse una relación similar. El número de avistamientos registrados de falsa orca fue muy bajo. Las actividades de alimentación suelen realizarlas debajo del agua (si se alimentan de los peces capturados en el palangre de superficie lo hacen a 15-100 metros de profundidad) y, por tanto, fuera del alcance de la vista de los observadores. Es posible que la baja frecuencia de avistamientos de esta especie pueda deberse a este factor. Sin embargo, otros delfines fueron avistados con mayor frecuencia; los delfines comunes y los delfines moteados son especies que suelen nadar cerca de los barcos saltando, en muchas ocasiones, fuera del agua, lo que les hace más visibles. Los avistamientos de Delphinus delphis registrados en la campaña de Azores fueron mayores cuando las capturas de pez espada fueron elevadas y las de marrajo fueron bajas; por el contrario, las capturas de marrajo fueron mayores y las de pez espada menores cuando la especie avistada fue Stenella frontalis. Esto sugiere que ambas especies de delfines comparten el mismo hábitat, aunque se alimentarían de distintas presas. Este resultado es contradictorio con otros estudios que han sugerido que la ecología trófica de estas dos especies de delfines es bastante similar (Aguiar dos Santos y Haimovici 2001) Es poco probable que depreden directamente sobre pez espada y marrajo de gran tamaño, pero, en cambio, que depreden sobre el mismo tipo de presa (peces y cefalópodos). En otras pesquerías se ha observado un fuerte asociación entre túnidos y delfines (D. delphis y Stenella) (por ejemplo: Hall & Donovan 2002; Reeves & Reijnders 2002); esta asociación ha sido explotada, por ejemplo, por los pescadores en el Pacífico Tropical Oriental donde el atún de aleta amarilla nada por debajo de 22 los delfines y éstos ayudan a la localización de los mismos (Archer et al., 2001). Au y Pitman (1986) sugirieron que los atunes podrían empujar a las capas superficiales del agua a las presas de éstos y los mamíferos marinos, actuando más como competidores. Esta misma asociación es posible que también se de en otras especies, como la sugerida en este estudio entre el pez espada y los delfines. Los T-PODs son instrumentos útiles para poder investigar la actividad de los cetáceos odontocetos debajo del agua. Los hidrófonos fueron programados para registrar los chasquidos emitidos en una banda de frecuencia determinada, en la que mayormente emite la falsa orca. Sin embargo, el análisis de acústica en relación con los avistamientos sugiere que la mayoría de los chasquidos recogidos por los hidrófonos procedían de pequeños delfínidos, los cuales emiten sonidos de ecolocalización en un rango de frecuencias similar al que emite la falsa orca. La falsa orca, al igual que otros odontocetos, utiliza el sistema de biosonar para la ecolocalización de sus presas, realizando un gasto energético en su uso. Los peces capturados en el palangre pelágico suponen una presa de acceso fácil y el uso de la ecolocalización puede ser no necesario para alimentarse y, por tanto, reducir el gasto energético de las actividades de alimentación. Los datos relacionados con la depredación en el palangre, recogidos tanto por los observadores científicos como por los patrones de los barcos, indican que la incidencia de depredación en los palangreros que operan en el Atlántico, cuyas especies objetivo son los túnidos, el pez espada y los tiburones, es baja. Los datos variaron en función de la disponibilidad de observadores científicos a bordo; los observadores registraron menos de un 1% de depredación sobre el total de captura por campaña, en cambio los datos registrados por los patrones indican una pérdida general ligeramente mayor. En este último caso, la captura perdida registrada por lance superaba el 25% en la mayoría de las ocasiones, aumentando hasta el 100% de la perdida de la pesca en algunas ocasiones. Resultados similares han sido obtenidos por Dalla Rosa y Secchi (2007), Kock et al. (2006), Poisson et al. (2007) y RamosCartelle y Mejuto (2007). Una de las posibles razones que se ha dado a esta baja incidencia de depredación, se debe a que los patrones podrían evitar las áreas de pesca donde se tiene constancia que la presencia de estos cetáceos es elevada y, así, 23 reducir las interacciones con ellos (Dahlheim 1988); los pescadores creen que estos animales han aprendido a seguir los barcos palangreros durante días para obtener beneficio de las capturas (Poisson y Taquet 2000; Donoghue et al. 2002). Esta actitud de los patrones y pescadores puede deberse a que las pérdidas económicas debidas a esta depredación puede llegar a ser de hasta un 40% del total del valor de las capturas, incluyendo los costes de las operaciones del barco y el tiempo perdido de pesca (ARVI, datos no publicados). Por otro lado, hay que tener en cuenta que la proporción de depredación obtenida en este estudio podría estar subestimada, ya que en las dos campañas con observadores científicos a bordo sólo se tuvo en cuenta el pescado en el que se registraron marcas de depredación, mientras que el pescado eliminado totalmente de los anzuelos no pudo ser contabilizado. La depredación registrada en las campañas con observadores científicos indicó que los tiburones constituyen el grupo depredador más importante con trece animales depredados (ocho en la campaña de Brasil y 5 en la de Azores), seguido de la falsa orca con siete (uno en la campaña de Brasil y 6 en la de Azores), y las tortugas con cuatro animales depredados en la campaña de Azores. La falsa orca fue el principal mamífero marino depredador que eliminó pescado del palangre, aunque solo se dio en algunas ocasiones y la depredación por parte de los tiburones fue el doble de frecuente que la atribuida a los mamíferos marinos. Los cetáceos que se avistaron con mayor frecuencia fueron D. delphis y S. frontalis, sin embargo sus registros fueron menores cuando se observó depredación sobre las capturas; por lo tanto, se sugiere que estas especies probablemente no se están alimentando del pescado capturado en el palangre. El resultado de que el registro acústico se más elevado cuando se observaron casos de depredación puede ser debido a un hecho casual o a que las falsas orcas se estuvieran comunicando con el grupo que se avistó en la zona, en el caso de las dos falsas orcas capturadas accidentalmente. En dos de las campañas sin observadores científicos a bordo de los barcos, la captura de pescado fue significativamente más baja cuando se registró depredación y avistamientos de falsa orca. También, el análisis de RDA sugiere una relación similar entre la incidencia de depredación y los avistamientos de Pseudorca en las campañas 24 donde se dispuso de observadores científicos a bordo. La captura accidental de falsa orca coincidió con la depredación de cinco peces espada en el mismo lance. Dalla Rosa y Secchi (2007) observaron que la depredación en las pesquerías de palangre del pez espada en las aguas de Brasil es principalmente causada por las orcas y, ocasionalmente, por otros cetáceos como las falsas orcas. Los resultados de estos autores pueden deberse a que realizaron el estudio en pesquerías de palangre que operan cerca de la costa del sur de Brasil, donde las orcas son más abundantes (Jefferson et al. 1993), mientras que los palangreros estudiados en este estudio operaban en aguas pelágicas y, normalmente, muy lejos de costa. Los observadores científicos en ambas campañas registraron depredación tanto por tiburones como por otras especies; sin embargo, los patrones de los otros seis barcos solo indicaron depredación en general, asumiendo que la especie responsable de la depredación fue la falsa orca, ya que en esos mismos lances se registraron avistamientos de esta especie cerca de los barcos. Donoghue et al. (2002) han sugerido que el pescado dañado por tiburones puede que haya sido registrado incorrectamente y es posible que, en muchas ocasiones, los patrones no sean capaces de distinguir entre los diferentes tipos de marcas responsabilizando, así, a los cetáceos de la incidencia de depredación. La falsa orca se alimenta principalmente de peces y cefalópodos (Koen-Alonso & Pedraza 1999; Hernandez-Garcia 2002; Ramos-Cartelle & Mejuto 2007). El pescado capturado en las líneas de palangre resulta un recurso alimenticio nuevo de fácil acceso para multitud de especies, como es el caso de los cetáceos, provocando un cambio de costumbres en la alimentación (Secchi y Vaske 1998; Gilman et al. 2006; Zollett y Read 2006). Ramos-Cartelley y Mejuto (2007) observaron que los cetáceos parecen haber aprendido que la carnada y las capturas de las pesquerías son una presa “fácil” de capturar y por tanto reduce los costes energéticos propios de la alimentación, aunque es posible que sean selectivos a la hora de elegir el pescado de las líneas (Kock et al. 2006). Al igual que en el sur de Brasil (Dalla Rosa y Secchi, 2007) y en el océano Índico (Poisson y Taquet, 2000) estas especies parecen tener una preferencia por los peces espada. En la campaña de Brasil las capturas fueron, fundamentalmente, de túnidos y la depredación por falsa orca se dio en pez espada, mientras que en la campaña de Azores los tiburones, seguidos del pez espada, 25 cubrieron la mayoría de las capturas, siendo los peces espada la especie mayormente depredada por la falsa orca. Estos resultados sugieren que el consumo de pez espada podría indicar una preferencia de esta presa por parte de los cetáceos, de igual forma que fue sugerido por Dalla Rosa y Secchi (2007) y Poisson y Taquet (2000). Las capturas accidentales de cetáceos fueron muy bajas, al igual que las indicadas por Dalla Rosa y Secchi (2002). Sólo se registraron dos falsas orcas capturadas accidentalmente, en el mismo lance en la campaña realizada en Azores. La captura accidental en las pesquerías de palangre no es considerada como uno de los factores más importantes sobre esta pesquería, ya que su incidencia parece ser ocasional; sin embargo, si es considerado un factor importante en aquellas pesquerías que utilizan redes, como las pesquerías de enmalle y arrastre (Perrin et al., 1994). La captura accidental de tortugas fue relativamente elevada, especialmente cuando las capturas de tiburones fueron elevadas. Al menos cuatro especies distintas resultaron afectadas: la tortuga laúd (Dermochelys coriacea), la tortuga verde (Chelonia mydas), la tortuga boba (Caretta caretta) y la tortuga lora (Lepidochelys olivacea). Es posible que en nuestra área de estudio los marrajos puedan depredar sobre las distintas especies de tortugas, al igual que Carranza et al. (2006) encontraron en el Atlántico ecuatorial oriental. Aunque en este estudio la mayoría de las especies fueron liberadas aparentemente vivas, la captura accidental de tortugas continúa siendo un tema importante en las pesquerías de palangre, que podría reducirse con la utilización de otro tipo de anzuelos y/o carnaza o realizando las operaciones de pesca a mayor profundidad (Gilman et al. 2007). 5. Conclusiones: Las capturas obtenidas por los palangreros que operaban en el Atlántico central tropical y el oeste del archipiélago de las Azores resultaron estar influenciadas por diversos parámetros ambientales, tales como las condiciones de luminosidad, temperatura superficial del mar y la profundidad del área de pesca; por otro lado, la presencia de determinadas especies de cetáceos sugieren una posible relación trófica con algunas de las especies pesqueras objetivo. 26 Es probable que la mayoría de los datos registrados por los hidrófonos provengan de las dos especies de delfínidos avistados, en vez de la falsa orca; además, el hecho de que los peces capturados por los palangres sean un recurso alimenticio “fácil”, puede favorecer a que estos animales no necesiten utilizar el biosonar para localizar sus presas. La incidencia de depredación registrada en este estudio fue baja, pero la proporción de la captura perdida excedió del 25%, en la mayoría de los casos. Aunque los análisis estadísticos revelan algunas relaciones potencialmente interesantes entre las capturas, la presencia de cetáceos, la depredación y las variables ambientales, es importante indicar que este es un estudio a pequeña escala, ya que sólo se analizaron en profundidad 86 lances con hidrófonos y observadores científicos a bordo. Sería necesario continuar obteniendo datos con observadores científicos a bordo para poder explorar y cuantificar estas relaciones. 6. Agradecimientos: A los patrones y marineros de los distintos barcos por su cooperación así como a los miembros de la CEMMA (Coordinadora para os estudios de mamíferos mariños) que ayudaron en el diseño de la recogida de datos de los cetáceos y en la revisión de los resultados. A Colin MacLeod y Alain Zuur por su asistencia en los análisis estadísticos y a Gema Martinez por su ayuda en la elaboración de los mapas. Este proyecto ha sido financiado por la Secretaría General de Pesca Marítima Española para “Desarrollo de un prototipo acústico para la reducción de la depredación ocasionada por mamíferos marinos en las pesquerías de pez espada (Pingers)”. 27 7. Bibliografía: Aguiar dos Santos, R. y M. Haimovici, 2001. Cephalopods in the diet of marine mammals stranded or incidentally caught along southeastern and southern Brazil (21-34°S). Fisheries Research 52: 99-112. Archer, F., Gerrodette, T., Dizon, A., Abella, K. Y Southern, S. 2001. Unobserved kill of nursing dolphin calves in a tuna purse-seine fishery. Marine Mammal Science 17(3): 540-554. Ashford, J. P., P. S. Rubilar y A. R. Martin, 1996. Interactions between cetaceans and longline fishery operations around South Georgia. Marine Mammal Science 12: 452–457. Au, D.W.K. y Pitman, R.L., 1986. Seabird interactions with dolphins and tuna in the Eastern Tropical Pacific. Condor 88: 304-317 Bigelow, K. A., C. H. Boggs y X. He, 1999. Environmental effects on swordfish and blue shark catch rates in the US North Pacific longline fishery. Fisheries Oceanography 8: 178–198. Brill R. W., B. A. Block, C. H. Boggs, K. A. Bigelow, E. V. Freund y D. J. Marcinek, 1999. Horizontal movements and depth distribution of large, adult yellowfin tuna (Thunnus albacares) near the Hawaiian Islands, recorded using ultrasonic telemetry: implications for the physiological ecology of pelagic fishes. Marine Biology 133: 395-408. Brothers, N. P., J. Cooper y S. Løkkeborg, 1999a. The incidental catch of seabirds by longline fisheries: worldwide review and technical guidelines for mitigation. FAO Fisheries Circular 937. FAO, Rome. Brothers, N. P., R. Gales y T. Reid, 1999b. The influence of environmental variables and mitigation measures on seabird catch rates in the Japanese tuna 28 longline fishery within the Australian Fishing Zone, 1991-1995. Biological Conservation 88: 85-101. Capdeville, D., 1997. Interaction of marine mammals with the longline fishery around the Kerguelen Islands (Division 58.5.1) during the 1995/96 cruise. CCAMLR Science 4: 171-174. Carranza, A., A. Domingo y A. Estrades. 2006, Pelagic longlines: A threat to sea turtles in the Equatorial Eastern Atlantic. Biological Conservation 131: 52-57. Carstensen, J., O. D. Henriksen y J. Teilmann, 2006. Impacts of offshore wind farm construction on harbour porpoises: acoustic monitoring of echolocation activity using porpoise detectors (TPODs). Marine Ecology Progress Series 321: 295–308. Carwadine, M. 1995. Whales, Dolphins and Porpoises. Dorling Kindersley, London. Cherel, Y., H. Weimerskirch y G. Duhamel, 1996. Interactions between longline vessels and seabirds in Kerguelen waters and a method to reduce seabird mortality. Biological Conservation 75: 63-70. Collette, B. B. y C. E. Nauen, 1983. FAO species catalogue. Scombrids of the world. An annotated catalogue of tunas, mackerels, bonitos and related species known to date. FAO Fisheries Synopsis 125 volumen 2. FAO, Rome. Dagorn, L., P. Bach y E. Josse, 2000. Movement patterns of large bigeye tuna (Thunnus obesus) in the open ocean, determined using ultrasonic telemetry. Marine Biology 136:361-371. Dahlheim, M. E., 1988. Killer whale (Orcinus orca) depredation on longline catches of sablefish (Anoplopoma fimbria) in Alaskan waters. Seattle, Washington: Alaska Fisheries Science Centre. NWAFC processed report 88-14. Dalla Rosa, L. y E. R. Secchi, 2002. Comparative analysis of the interactions between killer whales/sharks and the tuna/swordfish fishery in southern and 29 southeastern Brazil. In Donoghue, M., R. R. Reeves y G. S. Stone (eds), Informe del seminario “Interactions Between Cetaceans and Longline Fisheries”, Apia, Samoa, 2002. Dalla Rosa, L. y E. R. Secchi, 2007. Killer whale (Orcinus orca) interactions with the tuna and swordfish longline fishery off southern and south-eastern Brazil: a comparison with shark interactions. Journal of the Marine Biological Association of the United Kingdom 87:135-140. Dalla Rosa, L., E. R. Secchi y D. Monteiro, 2006. Killer whale depredation on longline catches in Brazilian waters. Actas del seminario “Fisheries Depredation by Killer and Sperm Whales: Behavioural insights, behavioural solutions”. British Columbia, Canada, 2006. Damalas, D., P. Megalofonou y M. Apostolopoulou, 2007. Environmental, spatial, temporal and operational effects on swordfish (Xiphias gladius) catch rates of eastern Mediterranean Sea longline fisheries. Fisheries Research 84: 233-246. De Stephanis, R, T. Cornulier, P. Verborgh, J. Salazar-Sierra, N. Perez Gimeno y C. Guinet, 2008. Summer spatial distribution of cetaceans in the Strait of Gibraltar in relation to the oceanographic context. Marine Ecology Progress Series 353: 275288. Domokos, R., M. P. Seki, J. J. Polovina y D. R. Hawn, 2007. Oceanographic investigation of the American Samoa Albacore (Thunnus alalunga) habitat and longline fishing grounds. Fisheries Oceanography 16: 555-572. Donoghue, M., R. R. Reeves y G. S. Stone, 2002. Informe del seminario “interactions between cetaceans and longline fisheries”. New England Aquarium Press, Boston MA, USA. Garza Gil, M. D., M. M. Varela Lafuente y C. Iglesias Malvido, 2003. Spain’s North Atlantic swordfish fishery. Marine Policy 27: 31-37. 30 Gilman, E., N. Brothers, G. McPherson y P. Dalzell, 2006. A review of cetacean interactions with longline gear. Journal of Cetacean Research and Management 8: 215-223. Gilman, E., S. Clarke, N. Brothers, J. Alfaro-Shigueto, J. Mandelman, J. Mangel, S. Petersen, S. Piovano, N. Thomson, P. Dalzell, M. Donoso, M. Goren y T. Werner, 2007. Shark Depredation and Unwanted By-catch in Pelagic Longline Fisheries: Industry Practices and Attitudes, and Shark Avoidance Strategies. Western Pacific Regional Fishery Management Council, Honolulu, USA. Gilman, E., D. Kobayashi, T. Swenarton, N. Brothers, P. Dalzell y I. Kinan-Kelly, 2007. Reducing sea turtle interactions in the Hawaii-based longline swordfish fishery. Biological Conservation 139: 19-28. Hall, M. A. y G. P. Donovan, 2002. Environmentalists, fishermen, cetaceans and fish: Is there a balance and can science help to find it? En Evans, P. G. H. y J. A. Raga (eds), Marine mammals: biology and conservation. Kluwer Academic Press. 491-521 Hernandez-Garcia. V., 2002. Contents of the digestive tract of a false killer whale (Pseudorca crassidens) stranded in Gran Canaria (Canary Islands, central east Atlantic). Bulletin of Marine Science 71: 367-369. Hutschinson, J.D. 1996. Fisheries Interactions: The Harbour Porpoise – a Review. En: Simmonds, M.P. y Hutchinson, J.D. (eds.), The Conservation of Whales and Dolphins. John Wiley & Sons Ltd. 129-165 International Commission for the Conservation of Atlantic Tunas (ICCAT), 2007. Informe del Período Bianual, 2006-2007, Parte 1, Volumen 2 . (http://www.iccat.int) Jefferson, T., S. Leatherwood y M. Webber, 1993. FAO species identification. Marine Mammals of the World. FAO, Rome. 31 Johnson, J. y Stevens, I, 2000. A fine resolution model of the easter North Atlantic between Azores, the Canary Islands and the Gibraltar Strait. Deep Sea Research I 47: 875-899. Kock, K. H., M. Purves y G. Duhamel, 2006. Interactions between cetaceans and fisheries in the Southern Ocean. Polar Biology 29: 379-388. Koen-Alonso, M. y S. N. Pedraza, 1999. Stomach contents of false killer whales (Pseudorca crassidens) stranded on the coasts of the Strait of Magellan, Tierra del Fuego. Marine Mammal Science 15:712-724. Lammers, M. O., W. W. L. Au, R. Aubauer y P. E. Nachtigall, 2003. A comparative analysis of the pulsed emissions of free-ranging Hawaiian spinner dolphins (Stenella longirostris). En Thomas, J. E., C. F. Moss y M. Vater (eds), Echolocation in Bats and Dolphins. University of Chicago Press, Chicago. Leeney, R. H. y N. J. C. Tregenza, 2006. Actas del seminario “Static Acoustic Monitoring of Cetaceans”. Gdynia, Poland, 2006. ECS Boletín 46, Número especial. Lewison R. L., S. A. Freeman y L. B. Crowder, 2004. Quantifying the effects of fisheries on threatened species: the impact of pelagic longlines on loggerhead and leatherback sea turtles. Ecology Letters 7: 221-231. Madsen, P. T., I. Kerr y R. Payne, 2004. Echolocation clicks of two free-ranging, oceanic delphinids with different food preferences: false killer whales Pseudorca crassidens and Risso´s dolphins Grampus griseus. Journal of Experimental Biology 207: 1811-1823. Memery, L., M. Arhan, X. A. Alvarez-Salgado, M. J. Messias, H. Mercier, C. G. Castro y A. F. Rios, 2000. The water masses along the western boundary of the south and equatorial Atlantic. Progress in Oceanography 47: 69-98. Morato, T., S. Guénette y T. Pitcher, 2001. Fisheries of the Azores, 1982-1999. In D. Zeller, R. Watson, T. Pitcher y D. Pauly (eds), Fisheries impacts on North 32 Atlantic Ecosystems: Catch, effort and national/regional data sets. Fisheries Centre Research Reports, University of British Columbia 9: 214-220. Nakamura, I., 1985. FAO species catalogue. Billfishes of the World. An annotated and illustrated catalogue of marlins, sailfishes, spearfishes and swordfishes known to date. FAO Fisheries Synopsis 125 volumen 5. FAO, Rome. Northridge, S.P., 1984. World review of interactions between marine mammals and fisheries. FAO Fisheries Technical Paper 251, FAO, Rome. Pallares, P. y J. M. Garcia-Mamolar, 1985. Efectos de las fases de la luna sobre los rendimientos de la flota atunera-tropical española. Collective Volume of Scientific Papers. International Commission for the Conservation of Atlantic Tunas (ICCAT) 23: 228-236. Perrin, W. F., G. P. Donovan y J. Barlow (eds), 1994. Report of the International Whaling Commission. Special Issue 15. Gillnets and Cetaceans. International Whaling Commission, Cambridge, UK. Philpott, E., A. Englund, S. Ingram y E. Rogan, 2007. Using T-PODs to investigate the echolocation of coastal bottlenose dolphins. Journal of the Marine Biological Association of the UK 87: 11-17. Poisson F. y M. Taquet, 2000. French Swordfish longline fishery in South-West Indian Ocean: Preliminary results from the PPR Program. Actas del IOTC. WPB0006. Poisson, F., C. Marjolet, K. Mete y M. Vanpouille, 2007. Interactions of cetaceans and sharks with the Reunion Island swordfish longline fishery in the Indian Ocean between 1997 and 2000. Informe del seminario de IOTC “the depredation in the tuna longline fisheries in the Indian Ocean”. Victoria, Seychelles, 2007. 33 Ramos-Cartelle, A. y J. Mejuto, 2007. Interaction of the false killer whale (Pseudorca crassidens) and depredation on the swordfish catches of the Spanish surface longline fleet in the Atlantic, Indian and Pacific Oceans. Informe del Standing Committee on Research and Statistics SCRS de la International Commission for the Conservation of Atlantic Tunas ICCAT 25 (ICCAT/SCRS/07/25). . Reeves, R. R. y P. J. H. Reijnders, 2002. Conservation and Management. En. Hoelzel, A. R. (ed), Marine Mammal Biology: An evolutionary approach. Blackwell Science, Oxford. Romo, A., C. Casanova, J. Sanz, A. Calle y J. L. Casanova, 2007. Relation between meteorological conditions and the catching of red tuna (Thunnus thynnus) from the measurements of the TOVS and AVHRR sensors of the NOAA satellites. International Journal of Remote Sensing 28: 2671-2681. Schotten, M., W. W. L. Au, M. O. Lammers y R. Aubauer. 2003, Echolocation recordings and localization of wild spinner dolphins (Stenella longirostris) and pantropical spotted dolphins (S. attenuata) using a four-hydrophone array. En Thomas, J. E., C. F. Moss y M. Vater (eds), Echolocation in Bats and Dolphins. University of Chicago Press, Chicago. Secchi, E. R. y T. Vaske Jr., 1998. Killer whale (Orcinus orca) sightings and depredation on tuna and swordfish longline catches in Southern Brazil. Aquatic Mammals 24:117-122. Skov, H., J. Teilmann, O. D. Henriksen y J. Carstensen, 2002. Investigations of harbour porpoises at the planned site for wind turbines at Horns Reef. Informe de situación: 1/1/2001 – 31/12/2001. Encargado por Tech-wise A/S. Ornis Consult A/S. http://www.hornsrev.dk/Miljoeforhold/miljoerapporter/Investigations_of_harbour _popoises.pdf Stacey, P. J., S. Leatherwood y R. W. Baird, 1994. Pseudorca crassidens. Mammalian Species 456:1-6. 34 Tougaard, J., J. Carstensen, O. D. Henriksen, J. Teilmann y J. R. Hansen, 2004. Harbour porpoises on Horns Reef - effects of the Horns Reef Wind Farm. Informe Annual de Situación 2003. Encargado por Elsam Engineering A/S. National Environmental Research Institute (NERI). Wood, F. G. y W. E. Evans, 1980. Adaptiveness and ecology of echolocation in toothed whales. In Busnel R. and J. Fish (eds), Animal sonar systems. New York Plenum. Zavala-Camin. L. A. y J. N. Antero da Silva, 1991. Histórico da pesquisa e pesca de atuns com espinhel no Brasil. Atlantica, Rio Grande 13:107-114. Zavilov, P. O., I. Wainer y J. M. Absy, 1999. Sea surface temperature variability off southern Brazil and Uruguay as revealed from historical data since 1854. Journal of Geophysical Research 105: 21-32. Zollett, E. A. y A. J. Read, 2006. Depredation of catch by bottlenose dolphins (Tursiops truncatus) in the Florida king mackerel (Scomberomorus cavalla) troll fishery. Fishery Bulletin 104:343-349. Zuur, A. F., E. N. Ieno y G. M. Smith, 2007. Analysing Ecological Data. Springer, New York. 35 FIGURAS 36 TABLAS 37 Figura 1. Mapa de zonas de pesca donde los barcos palangreros V1-V8 operaron durante el período de junio de 2006 y junio de 2007. Foto: J. Rodríguez (a) Depredación causada por tiburones Foto: C. Varela (b) Depredación causada por falsa orca Foto: C. Varela (c) Depredación causada por tortugas Figura 2. Efectos de la depredación por tiburones (a), falsa orca (b) y tortugas (c) sobre la captura Figura 3. Hidrófono (T-POD) para la recogida pasiva de sonidos emitidos por odontocetos bajo el agua. Figura 4-. Disposición de los T-PODs en el palangre 1) Foto: C.Varela 2) Foto: C.Varela 4) 3) Foto: J. Rodríguez Foto: J. Rodríguez Figura 5. Colocación del T-POD en el palangre. Figura 6. Clasificación de chasquidos recogidos por los T-PODs. Cet Hi: chasquidos con alta probabilidad de haber sido emitidos por la especie estudio; Cet Lo: chasquidos con baja probabilidad de haber sido emitidos por la especie estudio; ?: chasquidos que probablemente procedan de otra fuente distinta a los odonotocetos; ??: chasquidos que con alta probabilidad proceden de otra fuente distinta a los odontocetos; Pos: suma de Cet Hi+Cet Lo (también conocido como Cet All); Neg: suma de ?+?? Capturas de los palangreros que operaron en el Atlántico (junio 2006-junio 2007) Especies objetivo: pez espada, túnidos y tiburones Espada 120000 Atún Quenlla Marrajo 100000 Biomasa capturada (Kg) Istiophoridae Escolar Otros 80000 Otros tiburones 60000 40000 20000 0 0 1 2 3 4 5 6 7 8 9 Número de Barco Figura 7. Capturas totales de pescado durante las 8 campañas realizadas en aguas Atlánticas por palangreros españoles desde junio de 2006 hasta junio de 2007 a) b) Capturas totales por lance (V7) Capturas totales por lance (V7) 2500 Número de peces 50 Atún Barracuda Istiophoridae Otros Biomasa de pescado (kg) Pez espada Dorado Tiburones 60 40 30 20 10 Pez espada Dorado Tiburones 2000 Istiphoridae Otros 1500 1000 500 0 0 1 3 5 7 9 11 13 15 17 19 21 23 25 27 1 29 3 5 7 9 11 13 c) 15 17 19 21 23 25 27 29 Lances Lances Principales capturas de pescado (V7) Principales capturas de pescado (V7) d) 90 80 Pez espada 4500 Atún 4000 Total pescado 70 60 50 40 30 20 10 Biomasa de pescado (Kg) 100 Número de pescados Atún Barracuda Pez espada Atún 3500 Total pescado 3000 2500 2000 1500 1000 500 0 0 1 3 5 7 9 11 13 15 17 Lances 19 21 23 25 27 29 1 3 5 7 9 11 13 15 17 19 21 23 25 27 29 Lances Figura 8. Total de capturas realizadas por el palangrero V7 en número de pescados (a) y biomasa (b). Principales capturas de pescado y total de pescado en número (c) y biomasa (d). a) Pez espada Atún 2500 Quenlla Barracuda Biomasa de pescado (kg) Número de pescado 60 Marrajo Escolar Captura total de pescado (V8) b) Captura total de pescado (V8) 70 50 40 30 20 10 0 Pez espada Atún Marrajo Escolar 2000 1500 1000 500 0 1 4 7 10 13 16 19 22 25 28 31 34 37 40 43 46 49 52 55 1 4 7 10 13 16 19 22 25 Lances c) 28 31 34 37 40 43 46 49 52 55 52 55 Lances d) Principales capturas de pescado (V8) 90 Pez espada 80 Marrajo Quenlla 70 Principales capturas de pescado (V8) 3000 Biomasa de pescado (Kg) Número de pescados Quenlla Barracuda Total pescado 60 50 40 30 20 10 0 Pez espada Marrajo Quenlla Total pescado 2500 2000 1500 1000 500 0 1 4 7 10 13 16 19 22 25 28 31 Lances 34 37 40 43 46 49 52 55 1 4 7 10 13 16 19 22 25 28 31 34 37 40 43 46 49 Lances Figura 9. Total de capturas realizadas por el palangrero V8 en número de pescados (a) y biomasa (b). Principales capturas de pescado y total de pescado en número (c) y biomasa (d). Figura 10. Gráfica de RDA para las capturas realizadas por V7 (Brazil). Las variables respuesta (representadas por líneas finas) utilizadas fueron: capturas de pez espada (XGL), atún (THU), barracuda (SPH), dorado (CHI), Istiophoridae (IST), y tiburones (Shark). Las variables explicativas (representadas por líneas gruesas si son continuas o cuadrados si son nominales) utilizadas fueron: ver tabla 4 para la explicación de las abreviaturas. La gráfica se obtuvo tras eliminar aquellas variables no significativas que menos influenciaban al modelo, para una mejor visualización. Figura 11. Gráfica de datos condicionados co-plot para las capturas de pez espada en la campaña V7 (Brasil); se muestran las interacciones entre los efectos de la fase lunar y la cobertura nubosa. Figura 12. Gráfica de RDA para las capturas realizadas por V8 (Azores). Las variables respuesta (representadas por líneas finas) utilizadas fueron: capturas de pez espada (XGL), atún (THU), marrajo (IOX), y quenlla o tiburón azul (PGL). Las variables explicativas (representadas por líneas gruesas si son continuas o cuadrados si son nominales) utilizadas fueron: ver tabla 4 para la explicación de las abreviaturas. La gráfica se obtuvo tras eliminar a quellas variables no significativas que menos influenciaban al modelo, para una mejor visualización. a) b) c) d) Figura 13. Resultados de los análisis de regresión GAM para la campaña de Azores (V8) – curvas suavizadoras para los efectos parciales de (a) la cobertura nubosa y (b) la profundidad de la zona para las capturas de pez espada, y (c) la intensidad de chasquidos para las capturas de marrajo. Para la campaña de Brasil (V7) - curvas suavizadoras para los efectos parciales de (d) la profundidad de la zona para el número de chasquidos probablemente emitidos por falsa orca. Las líneas discontinuas indican el intervalo de confianza del 95%. Figura 14. Intensidad de chasquidos e incidencia de depredación para la campaña (a) V7 (Brasil) y (b) V8 (Azores), por lance. Figura 15 y 16. Gráfica de RDA para los datos acústicos obtenidos en la campaña de Brasil (V7). Las variables respuesta (representadas por líneas finas) utilizadas fueron: chasquidos probablemente emitidos por falsa orca (Pos) y chasquidos probablemente no emitidos por falsa orca (Neg). Las variables explicativas (representadas por líneas gruesas si son continuas o cuadrados si son nominales) utilizadas fueron: ver tabla 4 para la explicación de las abreviaturas. La gráfica se obtuvo tras eliminar aquellas variables no significativas que menos influenciaban al modelo, para una mejor visualización. 1 PCR dpdr 0.5 MIS axis 2 Pos Neg Depth PMA OOR IOX DDE M2 0 Hook M3 SFR Sea XGL ODO Cd -0.5 0 0.5 1 axis 1 Figura 17. Gráfica de RDA para los avistamientos de cetáceos en la campaña de Azores (V8). Las variables respuesta (representadas por líneas finas) utilizadas fueron: avistamientos de Misticetos (MIS), Delphinus delphis (DDE), Stenella frontalis (SFR), Pseudorca crassidens (PCR) and Orcinus orca (OOR), Physeter macrocephalus (PMA) y Odontocetos no identificados (ODO). Las variables explicativas (representadas por líneas gruesas si son continuas o cuadrados si son nominales) utilizadas fueron: ver tabla 4 para la explicación de las abreviaturas. La gráfica se obtuvo tras eliminar aquellas variables no significativas que menos influenciaban al modelo, para una mejor visualización. 0.5 LFLd XGLd SST hook THU axis 2 XGL Ints 0 PCR Depth Turt PGL IOXd IOX Cd -0.5 Sea -1 -0.5 0 axis 1 Figura 18. Gráfica de RDA para los avistamientos de cetáceos en la campaña de Azores (V8). Las variables respuesta (representadas por líneas finas) utilizadas fueron: depredación sobre pez espada (XGLd), marrajo (IOXd) y escolar (LFLd). Las variables explicativas (representadas por líneas gruesas si son continuas o cuadrados si son nominales) utilizadas fueron: ver tabla 4 para la explicación de las abreviaturas. La gráfica se obtuvo tras eliminar aquellas variables no significativas que menos influenciaban al modelo, para una mejor visualización.