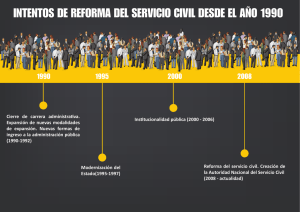
experiencia recepcional
Anuncio

lí/f UNIVERSIDAD VERACRUZANA FACULTAD DE BIOANALISIS LICENCIATURA EN QUIMICA CLINICA "EXPERIENCIA RECEPCIONAL" “Efecto del número de entrenamientos sobre la curva de aprendizaje espacial en crias de ratas Wistar” TESIS Presenta Sandra Aneli Hernández Landa Directora M. en C. Isela Santiago Roque Catedrática de Experiencia Recepcional E.D. Graciela Guadalupe Nava Kuri Xalapa Veracruz Agosto 2013 Agradecimientos A mis padres: Emigdio e Hilda, gracias a su apoyo constante y cariño, ahora puedo culminar uno de ios anhelos mas importantes en mi vida. A mi directora de tesis la M. en C. Isela Santiago Roque por su amistad, confianza, paciencia y apoyo brindado para la realización de este trabajo. A la M. en C. Cynthia Fernández Pomares, por el apoyo para la realización del análisis estadístico. A mis amigos: Lizeth Hernández Acosta,Ana Gabriela Gordillo Aguilar y Mauricio Villanueva Olivo quienes ayudaron para la captura del video de la prueba de laberinto acuático de Morris. Juan David Olivares Y Montserrat Correa por brindarme su apoyo, comprensión, preocupación y cariño. A todas las personas que han estado conmigo a lo largo de esta etapa y que me han brindado su confianza... mil gracias !! Contenido 1 Resumen.....................................................................................................................4 2 Introducción................................................................................................................. 5 3 Antecedentes...............................................................................................................7 3.1 Consideraciones metodológicas........................................................................ 12 3.1.1 Diferencias por Sexo.................................................................................12 3.1.2 Diferenciasentre Especies y cepas.......................................................... 13 3.1.3 Edad.......................................................................................................... 13 3.1.4 Nutrición.................................................................................................... 14 3.1.5 Estrés........................................................................................................ 14 3.1.6 Otros factores.................................................................................................15 3.2 Protocolos de entrenamiento, estrategias de nado y parámetros analizados 16 3.3 Neurobiologia del aprendizaje y memoria.......................................................... 20 3.3.1 Aprendizaje...................................................................................................20 3.3.2 Memoria........................................................................................................23 3.4 Navegación espacial...........................................................................................29 3.5 Áreas del cerebro relacionadas con el aprendizaje y memoria......................... 33 3.6 Modelos experimentales para el estudio de la memoria en animales............... 38 4 Planteamiento del problema..................................................................................... 41 5 Justificación............................................................................................................... 41 6 Hipótesis.................................................................................................................... 41 7 Objetivos.................................................................................................................... 42 8 9 7.1 Objetivo general..................................................................................................42 7.2 Objetivos particulares......................................................................................... 42 Material y métodos....................................................................................................43 8.1 Sujetos experimentales...................................................................................... 43 8.2 Evaluación de los grupos................................................................................... 43 8.3 Técnica para uso de Laberinto Acuático de Morris............................................45 Análisis estadístico....................................................................................................47 10 Resultados.............................................................................................................47 11 Discusión................................................................................................................ 52 12 Conclusión.............................................................................................................. 54 13 Bibliografía.............................................................................................................. 55 1 Resumen El aprendizaje y la memoria se relacionan con la capacidad de adquirir y retener asociaciones de las características del ambiente, esto permite que los humanos y mamíferos puedan ubicarse en un espacio. Para poder evaluar este tipo de aprendizaje y la memoria denominada espacial en los roedores se utiliza frecuentemente el Laberinto Acuático de Morris (MWM, por sus siglas en ingles). En esta prueba, los roedores nadan en una piscina hasta localizar una plataforma oculta en un tiempo determinado (tiempo de latencia), utilizando señales extra-laberinto (señales de referencia). Este procedimiento se hace en forma repetida a través de varios intentos y días consecutivos de entrenamiento y se observa como consecuencia del aprendizaje una disminución del tiempo en alcanzar la plataforma, produciendo una curva de aprendizaje progresiva. A pesar de que se conoce que el número de intentos genera la adquisición del aprendizaje y que existe una diversidad de protocolos, en el Laboratorio de Neurotoxicología se utiliza un protocolo que consiste en entrenar al roedor por tres días y en cada uno de ellos se realiza seis intentos, así como una prueba de memoria. Sin embargo no conocemos si la curva de aprendizaje continua siendo progresiva al incrementar el número de intentos o los días de entrenamiento, por ello se propone en este trabajo determinar si un mayor número de intentos y días de entrenamiento utilizados en el protocolo implementado en el laboratorio de Neurotoxicología produce una curva de aprendizaje progresiva. Para ello se utilizó 24 ratas Wistar machos de 21 días de edad, divididas en cuatro grupos, las cuales fueron sometidas a la prueba de laberinto acuático de Morris, realizando 6 a 8 intentos durante 3 ó 5 días de entrenamiento de acuerdo al grupo al que integraron. Los resultados indicaron que el incrementar el número de intentos y días de entrenamiento no produce una curva de aprendizaje progresiva y que es suficiente 18 intentos distribuidos en tres días de entrenamiento para obtener una curva progresiva tal y como se ha utilizado en el Laboratorio de Neurotoxicología. 2 Introducción El ser humano posee la capacidad de aprender y recordar (memoria), característica que comparte con otros mamíferos entre ellos los roedores. A través de estas capacidades, los humano puede reconocer estímulos que ha visto antes, realizar asociaciones entre ellos e incluso orientarse de acuerdo con las relaciones que establece; organizar acontecimientos pasados a través de la memoria, es decir, todo lo que aprende se agrupa y almacena en el cerebro, que posteriormente utiliza para adaptarse a situaciones futuras o resolver tareas que se le encomienda. De la misma manera, los roedores también se orientan en el espacio para organizar sus conductas parental, reproductiva, alimentarias, de sobrevivencia para regresar al nido como mecanismo de defensa ante un agente agresor. Esta capacidad de aprendizaje y memoria puede ser evaluada. La prueba más empleada es el Laberinto Acuático de Morris (MWM, por sus siglas en ingles), en donde los roedores tienen que nadar en una piscina circular hasta localizar una plataforma oculta en un tiempo determinado (tiempo de latencia), guiándose por señales que se localizan alrededor del laberinto (señales de referencia). Procedimiento que se hace en forma repetida a través de varios intentos y días consecutivos (Vicens, Redoiat & Carrasco, 2003) y que se denomina entrenamiento. En esta fase el tiempo promedio empleado por el roedor para alcanzar la plataforma nos proporciona evidencia de que ha adquirido una estrategia, lo que nos permite graficar el tiempo promedio versus día de entrenamiento y obtener una curva de aprendizaje progresiva. Para la evaluación de la memoria, se realiza una prueba de ensayo que consiste en un intento con tiempo limitado, el cual se lleva a cabo en el último día del experimento, para ello se quita la plataforma y se valora el tiempo de nado del roedor en el lugar donde se encontraba la plataforma que debe ser mayor que en el resto de la piscina (Vicens, Redoiat & Carrasco, 2003; Morris, 1984). La utilidad que ofrece el MWM es evaluar efectos que ejercen fármacos, tóxicos, patologías crónicas degenerativas sobre los procesos de aprendizaje y memoria. Por ello, en el laboratorio de Neurotoxicología, se ha implementado este test como herramienta en la evaluación de los efectos ocasionados por tóxicos, enfermedades crónicas y neurodegenerativas sobre el aprendizaje y la memoria espacial en ratas Wistar de 21 días de edad, adultas y ratones BALB/c. No obstante el protocolo implementado en el laboratorio para el MWM, consiste en un total de 18 intentos distribuidos en tres días de entrenamiento y un día para evaluar la memoria. Del cual se han obtenido curvas de aprendizaje progresivas que estadísticamente muestran una disminución significativa en el promedio del tiempo de latencia, que se asocia con la capacidad de aprendizaje del roedor, sin embargo se desconoce si al incrementar el número de intentos y los días de entrenamiento esta curva de aprendizaje mejore, es decir el tiempo promedio de latencia sea menor y genere a un más progresión de la curva comparada con la obtenida por los roedores que realizan 18 intentos. 3 Antecedentes El laberinto Acuático de Morris (MWM) fue desarrollado en 1981 por Richard G. Morris con el objetivo de estudiar y evaluar el aprendizaje y la memoria espacial en ratas de laboratorio, desde entonces es uno de los más utilizado en las neuroclencias conductuales para el estudio de los procesos psicológicos y mecanismos de aprendizaje y memoria (Soriano et al., 2007). Sin duda, una de las razones de su éxito es la simplicidad y el hecho de que evita los problemas generados por las claves olfativas que se acumulan en los laberintos tradicionales A lo largo de los años se le ha dado varios nombres como: piscina de Morris, laberinto de Morris, laberinto acuático, laberinto de nado, tarea de navegación espacial; el nombre más común y utilizado actualmente “laberinto acuático de Morris” (D’Hooge &De Deyn, 2001). El MWM está constituido por una piscina circular grande generalmente de 1,5 a 2 metros de diámetro llena de agua, la cual se mantiene a una temperatura que oscila entre los 18 y los 27°C, según se utilicen ratas o ratones, en la que se sumerge una plataforma que debe ser localizada por el roedor. Anteriormente se opacaba el agua con leche o alguna sustancia no tóxica como el látex para que el roedor no viera la plataforma. Sin embargo, se ha demostrado que esto no es necesario porque el roedor nada con la cabeza por encima del agua lo que le impide que vea la plataforma (Vicens, Redoiat & Carrasco, 2003). Aunque originalmente Morris (1984) describió dos piscinas para ratas: uno de 132 cm. y otro de 214 cm. de diámetro; el laberinto también se adapta para ratones donde las medidas de la piscina son de 73-180 cm. de diámetro y 28-57 cm. de altura, así como la plataforma de escape de 6 X 6 cm. En la versión tradicional del MWM, las ratas nadan desde diferentes puntos de salida, situados en el perímetro de la piscina, hasta encontrar la plataforma escondida bajo el agua. Para localizar la plataforma, el roedor depende de unos puntos de referencia (señales extra-laberinto), por ejemplo figuras localizadas alrededor de la circunferencia de la piscina (Ripoll, et al, 2010) (figura 1). Figura 1 Laberinto Acuatico de Morns Tomada de http://www.mcg edu/Core/Labs/sabc/Morriswatermaze htm Respecto al numero de objetos distales al laberinto suficientes para garantizar el aprendizaje, se ha demostrado que una cortina alrededor de la piscina retrasa la adquisicion de la tarea (Morris, 1981) y que cuando mas simple es el entorno exterior al laberinto mas facil es el aprendizaje de lugar (Lamberty & Gower, 1991). No obstante, las ratas de los experimentos de Prados y Trobalon (1998) y Rodrigo y cols. (1997) tuvieron dificultades para encontrar la plataforma escondida cuando solo se utilizaba un unico punto de referenda. Por otro lado, la precision con la que las ratas pueden estimar la distancia entre las diferentes claves y la plataforma varia inversamente con la distancia (Chamizo et al., 2006). Chamizo y Rodrigo (2004) corroboro que a menor distancia es mas probable que una unica clave sea suficiente para localizar la plataforma. Ellas entrenaron a ratas con una unica clave presente y registraron la latencia que tardaron en localizar la plataforma en los ensayos de entrenamiento y la proporcion de tiempo que las ratas pasaron en el cuadrante target en los ensayos de prueba. En ambas medidas, la ejecucion de las ratas fue mejor cuando la unica clave se encontraba suspendida inmediatamente sobre la plataforma que cuando se encontraba sobre la pared de la piscina y mejor cuando la clave se colocaba sobre la pared de la piscina en el cuadrante opuesto al de la plataforma. La proximidad de la clave a la plataforma es un factor importante que debe tomarse en cuenta a la hora de diseñar el procedimiento experimental. Otros experimentos han comprobado que los roedores normales (sin lesión, ni déficit cognitivo asociado a la edad) aprenden a localizar rápidamente la plataforma sumergida usando vías directas que indican que la navegación a un punto fijo y oculto en el espacio puede ser guiada por información de la memoria sobre su relación espacial y también la memoria de trabajo, cuando esta relación cambia en cada ensayo. La memoria de referencia es independiente de los ensayos y permite aprender el procedimiento general de la ejecución de la tarea y la memoria de trabajo en cambio es una memoria temporal que depende de cada ensayo. El MWM puede utilizarse también como tarea no espacial. En este procedimiento la plataforma es visible y puede llevar acoplada alguna señal, por lo que los animales resuelven la tarea sin hacer uso, necesariamente de la información que el espacio proporciona (esencial para la versión de la plataforma oculta). Ambas versiones (oculta y visible) no son igual de sensibles a los déficits conductuales. Un deterioro en la tarea visible sugeriría que el tratamiento experimental está afectando a otras capacidades más que al aprendizaje “per se” (Morris, 1984). Sin embargo, incluso con la plataforma visible, los animales podrían hacer uso de información espacial para localizar la plataforma. Únicamente una ejecución correcta de la versión oculta junto con un deterioro de la visible indicaría que los déficits motivacionales o sensoriomotores no contribuyen significativamente al aprendizaje espacial (D’Hooge & De Dyen, 2001) En la mayoría de los estudios conductuales se distinguen tres fases; habituación de la piscina, adquisición o aprendizaje y retención del aprendizaje también llamado prueba de ensayo. Fase de habituación de la piscina: Se introduce al roedor con el hocico apuntando hacia las paredes de la piscina y se deja para que busque la plataforma durante un tiempo de 60-120 segundos; cuando el roedor (por si solo) encuentra la plataforma se debe dejar de 20-30 segundos sobre ella; en caso de que no la encuentre, el experimentador lo guiara hacia la plataforma y lo dejara la misma cantidad de tiempo que en el caso anterior. En estos ensayos de habituación las señales extra-laberinto no están presentes, lo que se pretende es familiarizar al roedor con el ambiente y así disminuir los efectos interferentes sobre el mismo causados por las respuestas emocionales a los estímulos nuevos (Dieter, 1977). Fase de adquisición o aprendizaje; se introduce el roedor al agua de igual manera que en la fase anterior y se deja nadar durante 60-120 segundos para que busque la plataforma. En estos ensayos las claves espaciales están presentes. Se considera que el roedor ha encontrado la plataforma cuando permanece en ella 5-10 segundos; en caso de no encontrarla, es guiado hacia ella por el experimentador y se deja 30 segundos. Una vez transcurrido este tiempo, se retira el roedor de la plataforma y se deja descansar brevemente en la caja de ambientación antes de que inicie el siguiente ensayo. Este procedimiento se repite en los distintos ensayos a lo largo del entrenamiento; la capacidad del roedor para localizar la plataforma depende de la utilización de las señales extra-laberinto. Esta fase puede durar varios días. Prueba de retención del aprendizaje o prueba de ensayo: se realiza al terminar la fase de adquisición o aprendizaje. En esta prueba se retira la plataforma de la piscina y se permite nadar al roedor entre 60-120 segundos y se registra el tiempo que este pasa en el resto del cuadrante target, el tiempo que pasa en el resto de los cuadrantes y la trayectoria de nado. Esta es una prueba espacial en la que, sí el animal ha aprendido a localizar la plataforma con base a las claves distales al laberinto, nadara más tiempo en el cuadrante target, es decir en el que debería estar situado la plataforma (figura 2) (Vicens, Redoiat y Carrasco, 2003). 10 LABERINTO DE A G UA (A) (B) A D Q U ISIC IÓ N Primer (O día RETENCIÓN Figura 2. (A) Laberinto de agua y lugar donde se sitúa la platafonna su­ mergida (repivsentada por un pequeño cuadrado). (B) Trayectoría que re­ aliza e l animal para alcanzar la platafom ta sumeigida durante la fase de adquisición. Fn el prim er día de entrenamiento muestra un patrón de bús­ queda desorientado, mientras que e l últim o día su trayectoria es directa hacia la platafonna sumergida. (C) Trayectoria del animal en lapnieba fi­ nal sin plataforma. En esta pnieba de retención nada más tiempo en el cuadrante donde estaba situada la platafonna Figura tomada de: Vicens, Redoiat y Carrasco, 2003 11 3.1 Consideraciones metodológicas Los investigadores se han vuelto cada vez más consciente de que el rendimiento en el MWM está influenciado por factores tales como: las características de los animales de experimentación, las diferencias en el aparato de prueba o laberinto acuático y el procedimiento de entrenamiento. Respecto a las características de los animales de experimentación deben ser controladas o tomadas en cuenta en la planificación de los experimentos o en el análisis de resultados. Factores como el peso corporal, el desarrollo físico y la edad pueden influir en la velocidad de nado (Wenk, 1998) así como la cepa, la especie y el género también pueden afectar el rendimiento en el MWM. En el caso de los machos tienden a obtener mejores resultados que las hembras, y esto puede explicarse en sus diferencias en la fuerza muscular o la resistencia, así como la diferencia entre especies. Otros factores que afectan al aprendizaje espacial pueden ser el estrés, enfermedades como la desnutrición y también la vejez. 3.1.1 D ife re n cia s p o r Sexo Se encontraron diferencias en el rendimiento entre machos y hembras en el MWM, los machos muestran una ventaja en el aprendizaje espacial, asociado a su mayor tasa de maduración comparado a las hembras, sin embargo esta diferencia se puede mantener solo hasta los seis meses en donde ambos sexos alcanzan su maduración y la diferencia en el rendimiento en el MWM desaparece. ( Bucci et al; 1995) Las hormonas sexuales podrían desempeñar un papel importante en las diferencias observadas en el rendimiento cognitivo entre machos y hembras. En el caso de las hembras se plantea que la progesterona puede ser un factor que disminuya el desempeño en el MWM, debido a que al ser tratadas con testosterona durante su primera semana de vida no difirieron de los machos en el entrenamiento (Roof; 1993) Además, otros estudios demostraron que las ratas hembras no ovarioctemizadas 12 presentaron niveles de adquisición inferiores a las ovarioctemizadas (Daniel, et al„ 1999). También Se ha observado que el rendimiento es mejor en ratas hembra durante la fase de estro que durante el proestro (Warren & Jurasca, 1997). 3.1.2 D ife re n cia s en tre E species y cepas La tarea del MWM fue diseñada originalmente para estudiar los mecanismos de localización espacial en ratas. Los ratones, a pesar de que muestran una respuesta de comportamiento muy diferente al de las ratas cuando son expuestas al medio ambiente del MWM, se han utilizado con éxito por varios autores. Con la introducción de las técnicas de manipulación genética, los ratones se han convertido en los más importantes animales de laboratorio para la construcción de modelos transgénicos, y muchos de ellos fueron evaluados en el MWM (Lipp & Wolfer, 1998). Son capaces de aprender una estrategia para encontrar la plataforma en el MWM, aunque lo hacen más lentamente que las ratas (whishaw & Tomie, 1996) una posible explicación es, por un lado, que la .flotación y tigmotaxis^ tienden a ser más pronunciadas en los ratones que en ratas, lo que puede complicar las pruebas. Por otro lado, el hecho de que los ratones en determinados paradigmas de evitación activa tienden a paralizarse (respuesta de freezing) y a aplazar la respuesta de huida más que a buscar una posible ruta de escape (D’Hooge & De Deyn, 2001). Lipp y Wolfer (1998) atribuyen el desempeño superior de las ratas a mejores habilidades de natación (el desempeño de ensayo a ensayo es más estable), y no a los diferentes procesos cognitivos. Sin embargo, también han señalado que entre especies pueden existir diferencias entre las ratas y ratones en los procesos celulares y moleculares básicos de la memoria 3.1.3 Edad Estudios realizados han establecido que el rendimiento en el MWM disminuye con la edad de los animales (Brandéis, Brandys & Yehuda, 1989; Deupree, et al., 1991; Gower & Lamberty, 1993) y puede deberse a habilidades de natación, locomoción y exploración. Además, durante el envejecimiento los animales también muestran ' Es la tendencia a nadar a lo largo de la pared de la piscina 13 alteraciones en el funcionamiento cognitivo (tal como sucede en los humanos) y esto se ha encontrado que coincide con los cambios estructurales y fisiológicos en las regiones del cerebro involucradas con estas funciones (D’Hooge &De Deyn, 2001) como el hipocampo que presenta cambios en la función y en su morfología (Gallagher & Nicolle, 1993; Geinisman, et al.. 1995). Sin embargo, ha sido difícil de encontrar marcadores del hipocampo consistentes de la disminución cognitiva relacionada con la edad. 3.1.4 N u tric ió n El estado nutricional de los animales puede influir en el rendimiento del MWM de diversas maneras. En particular, una dieta hipocalórica de por vida evita déficits asociados con la edad en ratas de 24 meses de edad (Pitsikas, et al 1990; Stewart, Mitchell & Kalant, 1989) y la restricción de comida incrementa la resistencia de las ratas a los déficits de aprendizaje inducidos por la inyección intrahipocampal de Kainato (Bruce-Keller, et al., 1999). Bedi (1992) encontró que ratas desnutridas durante los primeros 30 días de vida postnatal tienen peores resultados en el MWM comparado con ratas control, pero incluso en un corto período de rehabilitación nutricional estos resultados se invierten. 3.1.5 E strés La función cognitiva se sabe que está influenciada por el estrés, tanto en animales como en seres humanos. En el MWM el aprendizaje está motivado por comportamientos aversivos y aun cuando se hace lo posible por reducir los factores de aversión, la adquisición de esta tarea será un evento estresante para los animales involucrados (Hölscher, 1999). En estas condiciones, el estrés adicional podría tener profundos efectos en el rendimiento cognitivo de tal forma que no se adquiere el aprendizaje y la memoria (Hölscher, 1999) y este se asocia a un aumento de la corticosterona en el plasma (Quervain, Roozendaal & McGauhg, 1998). La evidencia indica que las respuestas autónomas y endocrinas al estrés y la activación del eje hipotálamo-pituitario-adrenal subyacen a los efectos del estrés sobre el aprendizaje y la 14 memoria. Sandí (1998) sugiere que la liberación de las hormonas esteroide-adrenales como consecuencia del estrés, influye directamente en la formación de la memoria. 3.1.6 Otros factores Por otro lado, algunos agentes infecciosos han demostrado ser perjudiciales directa o indirectamente en el rendimiento cognitivo en la tarea de MWM. Las infecciones virales y bacterianas alteran el rendimiento en las ratas o ratones infectados (McLean, et al., 1993; Gibertini, et al., 1995). Existen otros factores que también logran deteriorar el aprendizaje y la memoria espacial tales como el alcohol, sustancias toxicas y privación de sueño De forma sorprendente el aislamiento social, aunque altera otras conductas, como la habituación en el campo abierto, no parece inducir un deterioro espacial, sino que incluso puede mejorar la adquisición del laberinto acuático (Wongwitdecha y Marsden, 1996). 15 3.2 P ro to c o lo s de e n tre n a m ie n to , e s tra te g ia s de nado y parám etros a n a liza d o s Los protocolos de entrenamiento básico incluyen: el entrenamiento de adquisición o aprendizaje, el test de prueba y el test de working memory (reversal test) (Wenk, 1998). No obstante, otros autores han descrito variaciones de estos protocolos incluyendo diferentes procedimientos de preentrenamiento, plataformas flotantes y protocolos mixtos de adquisición y retención (Brandéis 1989; Stewart & Morris 1993). El entrenamiento estándar en la versión plataforma escondida, usualmente consiste en bloques (entrenamientos) de 4 ensayos (intentos) cada uno, en los que se va variando aleatoriamente el punto de salida del animal. Como ya se mencionó anteriormente, cada ensayo tiene una duración de entre 60 y 120 segundos, tras los cuales el animal es guiado por el experimentador hacia la plataforma donde permanecerá un corto periodo de tiempo (por ejemplo 30 segundos). Los datos obtenidos en cada sesión de adquisición se utilizan para obtener un promedio por bloque de ensayos. Normalmente el parámetro más utilizado para establecer este promedio es la latencia de escape de cada roedor. Tras una serie de bloques de ensayos de adquisición, se realiza uno o varios ensayos de prueba o ensayos de transferencia, durante los cuales se retira la plataforma de la piscina y se permite al animal nadar durante un periodo de tiempo determinado (por ejemplo 60 o 120 segundos). Durante el ensayo de prueba, la precisión espacial del roedor está determinada y representada por el tiempo que el roedor pasa en el cuadrante donde debiera encontrarse la plataforma (cuadrante target) o por el número de veces que el roedor pasa por el área virtual que forma la plataforma. Los roedores que están bien entrenados muestran mayor preferencia por el cuadrante target y pasa una media de tiempo igual o superior al 50 % de su tiempo libre de nado explorando ese cuadrante. Alternativamente, estos bloques de ensayos de adquisición pueden estar seguidos por bloques de ensayos en los que se ha cambiado la posición de la plataforma y en los que el roedor debe de establecer una nueva relación espacial entre la plataforma y las claves visuales. Por otro lado, el entrenamiento de adquisición de la 16 tarea plataforma visible se ha usado a veces como una tarea de control no espacial. No obstante, se ha visto que incluso durante el entrenamiento en esta tarea los ratones parecen usar información espacial para localizar la plataforma (Hauben, et al., 1999). Básicamente, un animal puede usar tres estrategias diferentes para llegar a la plataforma de escape durante un ensayo de nado (Brandéis, Brandys & Yehuda, 1989): 1. - Estrategia praxis, se refiere a utilizar una secuencia aprendida de los movimientos que lo llevan a la plataforma. 2. - Estrategia taxis, puede acercarse a la plataforma por medio de señales proximales. 3. -estrategia espacial o mapeo. Puede navegar hasta la plataforma con información de señales distales sobre la ubicación de la plataforma, dentro de la configuración espacial. Dalm, et al. (2000), observaron que un animal podría utilizar tanto estrategias praxícas como espaciales para localizar la plataforma escondida incluso cuando las posiciones de salida del animal se aleatorizan. La piscina, así como los procedimientos de capacitación han variado a través de varios estudios, se han utilizado líquidos para opacar el agua como leche pura o en polvo, pintura blanca no tóxica, o sintéticos como el látex. Su uso en ocasiones se vuelve compleja debido a que en el caso de la leche puede descomponerse fácilmente si se utiliza una temperatura por encima de 25°C (Lamberty & Gower, 1993). Sin embargo cada vez está en desuso, porque se ha comprobado que no afecta el dejar de usarlos. El tamaño de la piscina utilizada para esta prueba puede variar de acuerdo a los sujetos experimentales que se requiere evaluar. Morris (1984) originalmente describe dos piscinas para el uso en ratas: uno de 132 cm y otro de 214 cm de diámetro, además de 60 cm y 40 cm de profundidad respectivamente. Jett, et al.( 1997) utilizaron para ratas adultas una piscina de 180 cm de diámetro y 70 cm de profundidad; para ratas de 21 días observaron una considerable diferencia de tamaño corporal en comparación con las adultas, por lo tanto, utilizaron una piscina de 90 cm de diámetro y de 50 cm de profundidad, con la finalidad de eludir el problema potencial de agotamiento y una 17 mayor dificultad de la tarea; las dimensiones de la piscina son similares a la utilizada por Morris(1981) en sus experimentos de ratas pre y post-destete. Varios autores han reducido las dimensiones de la piscina para su uso en ratones. Por ejemplo: Ikonen, Schmidt & Riekkinen (1999) utiliza una piscina de 120 cm de diámetro, Lamberty & Gower(1993) utilizaron una piscina de 90 cm de diámetro; Means, Higgins & Fernández (1993) utilizaron una piscina de 61 cm de diámetro para ratones. Se han empleado piscinas de 150 cm de diámetro en trabajos de modelos de ratones transgénicos (D’Hooge, et al., 1999). Las medidas conductuales más utilizadas son latencia de escap>e en los ensayos de adquisición y el porcentaje de tiempo que el sujeto pasa en el cuadrante target durante la prueba del test. La adquisición se refleja en menores latencias de ensayo a lo largo de los días y la retención en el mayor porcentaje de tiempo en el cuadrante target. Estas medidas se registran mediante cronómetros o métodos computarizados (Gerlai, 2001 ). Lindner (1997) argumenta que la longitud del trayecto podría ser el índice más apropiado de rendimiento cognitivo en el MWM. Varios autores han recomendado el uso de medidas alternativas, como la ruta de direccionalidad (Stewart & Morris, 1993) o la distancia acumulada a la plataforma (Dalm, et al., 2000) para poder permitir una distinción más precisa entre los patrones de natación o las estrategias de búsqueda. Las medidas de desempeño, a menudo agrupadas por cada bloque de prueba, pueden ser graficadas para cada grupo experimental de animales para obtener una curva de aprendizaje progresiva. El análisis estadístico de las diferencias entre los grupos en la medición repetida de estos parámetros se puede realizar mediante el análisis de varianza (ANOVA) de dos vías para medidas repetidas, con el grupo experimental y los ensayos como fuentes de variación. Las pruebas de comparación múltiple subsiguiente permitirá la identificación de diferencias estadísticamente significativas entre los pares de medios. Rendimiento de exploración puede ser representado como el tiempo en 18 diferentes zonas de la piscina o el número de cruces de la zona de destino, y los procedimientos de ANOVA, será usualmente suficiente, para evaluar el significado de las diferencias entre las medias (Dalm, et al., 2000). 19 3.3 Neurobiologia del aprendizaje y memoria El aprendizaje y la memoria son dos procesos estrechamente ligados y están presentes en procesos cerebrales como la percepción sensorial, las emociones o el lenguaje y también participa en funciones conductuales. Se relacionan con la capacidad de adquirir y retener asociaciones de las características del ambiente, lo que permite al organismo desenvolverse en el espacio (Vicens, Redoiat y Carrasco, 2003;). En el “lenguaje” del sistema nervioso, aprender significa producir una referencia básica a través de los patrones neuronales construidos e incorporados a lo largo de las diversas etapas comprendidas entre el nacimiento y la madurez (Martel, et al., 2006). El aprendizaje es el proceso gracias al cual los animales y seres humanos adquirimos conocimientos sobre el ambiente que nos rodea. La memoria es la capacidad del individuo de retener y utilizar informaciones de diferentes maneras y en diferentes periodos (Levitt, Fugelsang & Crossiey 2006). En consecuencia, el aprendizaje y la memoria son procesos interrelacionados, ya que sin aprendizaje no hay memoria (Morgado, 2005) 3.3.1 A p re n d iza je El aprendizaje es un proceso continuo por el cual los organismos modifican su conducta para adaptarse a las condiciones cambiantes e impredecibles del medio que los rodea. Junto a las fuerzas selectivas de la evolución, el aprendizaje constituye el modo principal de adaptación de los seres vivos. Cuanto más cambiante es el entorno más plástica debe ser la conducta, por lo que los organismos que viven en medios diferentes presentan también grados diferentes de plasticidad conductual. Cuanto más plasticidad tiene su sistema nervioso más posibilidades de aprendizaje tiene un animal. Por tanto, el aprendizaje puede considerarse como un cambio en el sistema nervioso que resulta de la experiencia y que origina cambios duraderos en la conducta de los organismos. Lo que aprendemos es retenido o almacenado en nuestro cerebro y constituye lo que denominamos memoria. 20 Existen varias formas de aprendizaje: aprendizaje no asociativo, aprendizaje asociativo y aprendizaje cognitivo (figura 3) Figura 3. Formas de aprendizaje Aprendizaje no asociativo En el aprendizaje no asociativo, el organismo modifica su respuesta cuando se ve expuesto varias veces al mismo tipo de estímulo y se observa a través del comportamiento. Las formas más comunes de aprendizaje no asociativo se encuentran representadas por la habituación y la sensibilización. La habituación se entiende como la reducción de la eficiencia al estímulo que, por no ser significativo, conduce a ignorarlo. La sensibilización es la intensificación de la respuesta en función de un fuerte estímulo capaz de producir incluso la percepción de estímulos que anteriormente al hecho, se consideraban débiles (Simons-Weidenmaier, et al., 2006). Aprendizaje asociativo. El aprendizaje asociativo se caracteriza por asociaciones entre hechos, se distinguen dos tipos de procesos: el condicionamiento clásico y el operante. En el condicionamiento clásico se produce la relación estímulo-respuesta, donde el primer estímulo evoca una respuesta que se puede medir de forma diferente a aquella que se produce como resultado de la aplicación del segundo estímulo. El primero es el 21 estímulo incondicionado, que no requiere entrenamiento o preparación anticipada a la respuesta; el segundo es la estimulación condicionada, que necesita del entrenamiento específico sobre la acción motora pretendida (Hoover, et al., 2006). El condicionamiento operante es la asociación de un comportamiento particular a un estímulo significativo, una relación estímulo-comportamiento orgánico donde el acto motor es consecuencia de una respuesta neurofisiológica a un suceso (Eamshaw & Bressioff, 2006). Con base a este proceso, el organismo se vuelve capaz de prever, a partir de un comportamiento suyo, la posterior verificación del acontecimiento deseado, y esto explica porque un animal tiende a repetir un comportamiento espontaneo (innato o casual) si éste determina una variación favorable en el ambiente, como una recompensa o el alojamiento de un estímulo perjudicial. Aprendizaje cognitivo Implica una serie de operaciones mentales, como el análisis del estímulo, su confrontación con información ya presente en la memoria o su asignación a una determinada categoría. Ha sido sobre todo el estudio de aprendizaje cognitivo el que ha demostrado que la memoria es un fenómeno dinámico en virtud del cual el modo en que la información adquirida se conserva varía con el paso del tiempo 22 3.3.2 Memoria La memoria es la capacidad de retener y de evocar eventos del pasado, mediante procesos neurobiológicos de almacenamiento y de recuperación de la información, básica en el aprendizaje y en el pensamiento (Etchepareborda y AbadMas, 2005) En los primeros años de la vida, la memoria es de carácter sensitivo, guarda sensaciones o emociones. Más tarde aparece la memoria de las conductas: se ensayan movimientos, se repiten y poco a poco se van grabando. Finalmente, se desarrolla la memoria del conocimiento, o capacidad de introducir datos, almacenarlos correctamente y evocarlos cuando sea oportuno (Etchepareborda & Abad-Mas, 2005). El sistema de la memoria está integrado por tres procesos básicos (Etchepareborda & Abad-Mas, 2005): 1) Codificación de la información. La codificación o adquisición es el proceso en donde se prepara la información para que se pueda guardar. La información puede codificarse como una imagen, sonidos, experiencias, acontecimientos o Ideas significativas; las circunstancias que rodean este momento resultan fundamentales para el éxito o fracaso de la memoria. Es importante en este proceso inicial, la atención, la concentración y el estado emocional del sujeto. 2) Almacenamiento de la información. Esta etapa se caracteriza por el ordenamiento, categorización o simple titulación de la información mientras se desarrolla el proceso en curso. Esto requiere tanto como de una metodología como de estructuras intelectuales que ayuden a la persona a clasificar los datos. Una vez que es codificada la experiencia y almacenada por cierto tiempo, esta se presenta de manera automática. El almacenamiento es un sistema complejo y dinámico que cambia con las experiencias a las que el sujeto es expuesto. 3) Evocación o recuperación de la información. Es el proceso por el cual se recupera la información. Si ésta ha sido bien almacenada y clasificada será más fácil localizarla y utilizarla en el momento en que se solicite. 23 La memoria se desarrolla a través de una variable temporal. Esta situación ha permitido dividirla en etapas o niveles temporales (figura 4), de acuerdo al momento en que se encuentre. Así, se reconocen tres tipos de memoria: inmediata, de corto plazo (mediata) y de largo plazo (diferida) Memoria Corto plazo (mediata) Memoria de trabajo Implicita o Procedimental (no declarativa) Episódica (Autobiográfica) Largo plazo (diferida) Explicita (declarativa) Figura 4. Etapas de la memoria Memoria inmediata Este tipo de memoria está relacionado con lo que se denomina registro sensorial. Está vinculada con la información que no ha sido procesada y que viene de los sentidos. Esta información entra, permanece un lapso de tiempo y luego se procesa o se pierde (Etchepareborda y Abad-Mas, 2005). La memoria sensorial puede retener representaciones efímeras de prácticamente todo lo que vemos, oímos, olemos o sentimos (Etchepareborda y Abad-Mas, 2005). Memoria a corto plazo Es un sistema en el que se retiene la información reciente y en donde se almacena una cantidad limitada de información que está disponible sólo durante unos segundos hasta un minuto luego de transcurrido el aprendizaje. Es una memoria inmediata y transitoria de los estímulos que acaban de ser percibidos. Se considera 24 frágil e inestable, debido a que puede ser vulnerable a cualquier tipo de interferencia (Ruetti et al-, 2009). La memoria a corto plazo se basa en actividad o cambios efímeros, eléctricos o moleculares, en las redes de neuronas que procesan la información (Morgado 2005) Memoria de trabajo Baddeley (1983) describe este tipo de memoria como un mecanismo de almacenamiento temporal que permite retener a la vez algunos datos de información en la mente, compararlos, contrastarlos o en su lugar, relacionarlos entre si. Se responsabiliza del almacenamiento a corto plazo, a la vez que manipula la información necesaria para los procesos cognitivos de alta complejidad. Esta clase de memoria suele utilizarse durante el aprendizaje de tareas complejas, que requieren que dos clases de información permanezcan accesibles de manera simultánea (Ruetti et al., 2009). Según Baddeley, el estímulo, al ser atendido y percibido, se transfiere a la memoria de trabajo. Esta memoria nos capacita para recordar la información pero, es limitada y susceptible de interferencias. Esta vulnerabilidad del proceso le imprime un carácter de enorme flexibilidad, que nos permite estar siempre “abiertos” a la recepción de nueva información. La memoria de trabajo participa en por lo menos dos tipos de procesos (Etchepareborda y Abad-Mas, 2005): 1. -Control ejecutivo: hace referencia al mecanismo de procesamiento de la información. 2. -Sostenimiento activo: constituye el concepto de almacenamiento temporal. Y está formada por tres componentes (Etchepareborda y Abad-Mas, 2005): 1.-Bucle articulatorio: encargado de mantener activa y manipular la información presentada por medio del lenguaje. Por tanto, está implicado en tareas puramente lingüísticas, como la comprensión, la lectoescritura o la conversación, así como en el manejo de palabras, números, descripciones, etc. 25 2. -Agenda visuo-espacial: encargada de elaborar y manipular información visual y espacial. Se ha comprobado que está implicada en la aptitud espacial, como por ejemplo el aprendizaje de mapas geográficos. 3. -Ejecutivo central: realiza dos funciones: a) Distribuir la atención que se asigna a cada una de las tareas a realizar y b) Vigilar la atención de la tarea y su ajuste a las demandas del contexto; a medida que una tarea se domina, necesita menos atención y permite la ejecución otras tareas compatibles. Memoria a largo plazo Es un sistema que permite almacenar gran cantidad de información durante períodos prolongados de tiempo, que pueden variar en su extensión desde meses hasta años, y en algunos casos, toda la vida. Por lo tanto, la característica fundamental de este tipo de memoria es que constituye un almacén duradero de la información adquirida y que es menos susceptible de ser alterada por interferencias (Morgado, 2005). Se considera que para que la información forme parte de la memoria a largo plazo, debe pasar previamente por la fase anterior, es decir, en primer lugar se procesa en la memoria a corto plazo y finalmente pasa a formar parte de una memoria más persistente. El proceso gradual por el que la reiteración de la información retenida a corto plazo produce cambios neurales que originan la memoria de largo plazo declarativa, se denomina consolidación de la memoria (Ruetti et al., 2009). El procesamiento a largo plazo de la información se lleva a cabo en una secuencia determinada: existe una primera fase de adquisición que ocurre durante el aprendizaje; luego tiene lugar la consolidación de la memoria, un proceso que dura pocas horas y en el que los recuerdos pasan de un estado lábil hacia otro más estable. Datos más recientes sugieren la existencia de una forma de consolidación que dura varios días e incluso meses (Izquierdo et al., 2006). 26 Una de las principales distinciones de los sistemas de memoria es la que se realiza entre la memoria declarativa y una forma no consiente de memoria, la memoria no declarativa (Milner et al., 1998) Memoria declarativa La memoria declarativa o explícita es aquella capaz de ‘retener” números, hechos, acontecimientos y otras situaciones conscientes; por este motivo, también se denomina memoria explícita (Fell, et al., 2006); es preposicional, es decir, puede ser correcta o incorrecta, pues está involucrada en la manipulación de la información y de las experiencias del mundo externo (Ruetti et al., 2009). Dado que puede adquirirse rápidamente, también se puede olvidar con la misma velocidad (Manns & Einchenbaum 2006). Por esta razón, esta memoria provoca cambios poco significativos en las conexiones sinópticas que se distribuyen por toda la corteza (Kohier, et al., 2005). En la región del lóbulo temporal se localizan las principales estructuras nerviosas implicadas en este tipo de memoria, como el hipocampo y la corteza entorrinal; además de la asociación que mantienen entre sí, también se comunican con otras regiones corticales. Existen regiones, denominadas ‘moduladoras’, que actúan en la formación de la memoria declarativa. Entre otras estructuras se encuentran la amígdala localizada en la fase inicial del lóbulo temporal-, la sustancia negra, los núcleos del rafe y el núcleo basal de Meynert (Paz, et al., 2006). La amígdala es la estructura que recibe el primer impacto de las hormonas periféricas (corticoides, adrenalina) liberadas en la sangre como resultado del estrés o fuertes emociones en el momento de formación de la memoria (Hammond, et al., 2006). Las otras regiones reguladoras de los estados de ánimo, la ansiedad, la alerta y las emociones son responsables de la liberación de los neurotransmisores dopamina, noradrenalina, serotonina y acetilcolina. Este proceso se lleva a cabo mediante el contacto de sus axones con el hipocampo, la amígdala y la corteza entorrinal, la corteza cingulada y la corteza parietal (Paz, et al., 2006). 27 Dentro de esta clasificación se encuentran a su vez la memoria semántica, que contiene el conocimiento sobre hechos generales, y la memoria episódica que abarca los datos autobiográficos (Ruetti et al.2009) Memoria no declarativa La memoria no declarativa se encarga de informaciones más subjetivas, como la sensación de miedo que afecta a la amígdala y el aprendizaje de las habilidades motoras, los hábitos y comportamientos. Ese tipo de almacenamiento se denomina ‘memoria de procedimientos’ (Hotermans, et al., 2006). Aunque la memoria de procedimientos también se almacena en la corteza, su recuperación es diferente a la de la memoria declarativa; la memoria de procedimientos se recupera de manera inconsciente (De Carvalho-Machado, et al., 2005). La memoria no declarativa, frecuentemente denominada ‘memoria implícita’, es el resultado de la experiencia adquirida a lo largo del tiempo por el individuo. La formación de este tipo de memoria requiere cambios duraderos en las conexiones sinópticas y se adquiere mediante la práctica y la repetición de los hechos que deben aprenderse, dando lugar a un aprendizaje de larga duración (Press, et al., 2005). Las regiones responsables son, básicamente, el núcleo caudado (inervado por la sustancia negra) y el cerebelo. Sin embargo, algunas veces la memoria de procedimientos hace uso también de los lóbulos temporales. Ese tipo de memoria difícilmente falla y sufre modulación por variables emocionales. Su mayor influencia proviene de la vía sustancia negra-núcleo caudado. Algunas formas de memoria no declarativa son fenómenos de aprendizaje tales como habituación, sensibilización y condicionamiento clásico e instrumental (Ruetti et al,. 2009). 28 3.4 N avegación espacial La mayoria de los animales (entre ellos los roedores) necesitan disponer de un buen sistema de orientación espacial para organizar sus conductas en el entorno en que habitan. En condiciones naturales los roedores encuentran habitualmente el alimento y el agua en determinados lugares del espacio, por ello, han de emplear algún tipo de estrategia de navegación que les permita localizar y recordar el lugar donde se encuentra la comida. Estas capacidades de los roedores también pueden estudiarse experimentalmente en laboratorios, mediante el empleo de laberintos cuya solución depende de la utilización de información espacial (Santín et al., 2000). La navegación espacial se considera una conducta compleja, caracterizada por la búsqueda de un lugar en el entorno y por el desarrollo de movimientos autocontrolados en el espacio dirigidos a localizar un lugar no visible (Santín et al., 2000) En 1978 O’Keefe y Nadel retomaron el estudio del espacio y de los procesos psicobiológicos relacionados, los cuales ya habían sido abordados entre otros por Tolman y Hull (Chamizo, 1990), y realizaron una clasificación sistemática en la que estudiaron los problemas de la representación del espacio en el sistema nervioso. Esta formulación teórica establece un sistema clasificatorio que englobaba prácticamente todas las formas de navegación espacial mayoritariamente estudiadas en los laboratorios de experimentación animal. Los dos elementos básicos de la clasificación son la propuesta de dos sistemas de navegación, neurobiológica y psicológicamente disociables: el sistema táxico y el sistema cartográfico. El sistema táxico, a su vez, puede desarrollarse a través de dos mecanismos: el aprendizaje de orientación y el aprendizaje de guía. En el aprendizaje de orientación los animales basan su búsqueda en movimientos aprendidos durante la ejecución de la tarea; en el aprendizaje de guia aprenden asociaciones entre los estímulos señal y la meta. Estas dos formas de navegación se explican mediante paradigmas asociativos de condicionamiento. Sin embargo, el sistema cartográfico se 29 basa en la memoria sobre la localización de un lugar con respecto a la configuración de pistas disponibles en el entorno circundante (Chamizo, 1990). Recientemente, se ha prestado interés a otra forma de solucionar problemas de tipo espacial, basada en una propuesta formulada por Charles Darwin en 1873. Esta forma de navegación espacial consiste en un proceso de actualización de la información, que permite que un animal en movimiento pueda mantener en su memoria la huella sobre la localización de un lugar de salida en relación con su posición actual, medíante el procesamiento de la información sobre los movimientos recabados durante el camino recorrido (información basada en la ruta). Esta teoría sobre la navegación espacial recibe el nombre de integración de la ruta y se desarrolla principalmente cuando las pistas ambientales no ofrecen información suficiente o en ambientes desde el punto de vista de la estimulación muy homogéneos o ambiguos (Benhamou, 1997; Taube, et al., 1996; Whishaw, 1998; Santín et al., 2000)), aunque para algunos autores forma parte de la navegación normal (Whishaw 1998). A medida que la navegación progresa, la aceleración, la distancia y la dirección se miden y se integran para que la posición actual se actualice constantemente con respecto al punto de salida. Aún se desconoce cuáles son las pistas que los anímales emplean para aplicar esta estrategia, pero se cree que pueden obtener estimaciones sobre la velocidad, distancia y dirección mediante la utilización de pistas anestésicas y señales vestibulares; no obstante, no se descarta la posibilidad de emplear pistas procedentes de otros sistemas sensoriales (Whishaw 1998). Por lo tanto, los roedores pueden adoptar cuatro formas principales de navegación para la resolución de tareas espaciales: orientación, guía, cartográfica e integración de la ruta (Santín et al., 2000). Estas permiten encontrar lugares concretos en un ambiente, tanto natural como de laboratorio. En todas ellas, es necesario que los sujetos formen ‘memorias’ para poder recordar dónde se encuentra un lugar; no obstante, la memoria a formar en cada una de estas formas de navegación posiblemente sea diferente (determinada por la clase de información que se procesa y representa, así como por los circuitos nerviosos que sustentan la formación de las diferentes memorias). A pesar de 30 la existencia de memorias diferentes en cuanto al contenido y a los sistemas cerebrales subyacentes, éstas se construyen y emplean mediante los mismos procesos generales: codificación, almacenamiento y recuperación de la información que permite identificar un lugar. Estos procesos son necesarios para almacenar la información sobre el espacio y poder recuperarla cuando los individuos lo necesiten. Posiblemente, la diferencia en estos procesos, en cada una de las cuatro formas de navegación espacial, se halle en los mecanismos tanto psicológicos como neurobiológicos que los componen. De este modo, parece razonable suponer que lo que un animal ha de representar cuando aprende una tarea espacial empleando una estrategia de orientación es completamente diferente a lo que representa cuando guía su orientación utilizando estrategias. Así, en el primer caso, la conducta espacial estaría guiada por movimientos estereotipados con los que los anímales pueden llegar a un lugar; no obstante, este aprendizaje no implica establecer relaciones entre los objetos, lo que el animal aprende es la mera repetición de movimientos y giros corporales concatenados; en el segundo caso, el empleo de una estrategia cartográfica implica el aprendizaje de relaciones entre los objetos (Chamizo & Mackintosh, 1989; Roberts, Pearce, 1998). Se desconoce cuáles son las representaciones que se construyen y emplean en esta clase de aprendizajes. Principalmente se establecen dos teorías que proponen dos tipos de representaciones diferentes: 1. La teoría del mapa cognitivo (O’Keefe y Nadel, 1978): defiende que los animales representan relaciones espaciales, de tipo topográfico entre los estímulos, y para ello realizan cálculos métricos, como la estimación de la distancia y relaciones angulares entre los objetos. 2. La teoría relacíonal (Eichenbaum, 1999), propone que el aprendizaje espacial no se reduce a las formas sencillas de aprendizaje asociativo, aunque tampoco es necesario explicarlo en función del aprendizaje y uso de las relaciones topográficas entre los 31 estímulos ambientales; indudablemente niega la necesidad de fom ar mapas cognitivos en sentido estricto. 32 3.5 Á reas del ce re b ro re la cio n a d a s con el a p re n d iza je y m em oria El hipocampo es una estructura crítica para procesar y recordar información espacial y contextual, su lesión o la de sus vías de salida de información, como el fórnix, afecta al aprendizaje de tareas en los laberintos radiales o de agua. El hipocampo parece desempeñar un papel fundamental en la ejecución de tareas espaciales; en 1971, John O’Keefe y Jonathan Dostrovsky descubrieron que existen neuronas del hipocampo que se activan específicamente cuando el animal está en una localización determinada de un ambiente (Nadel & Eichenbaum, 1999). Por ello, se les llamó células de lugar; anatómicamente, estas neuronas son células piramidales de la región CAI y CA3 (Muller, 1996). Se ha propuesto que las células de lugar forman parte de un mapa cognitivo en el hipocampo y se ha considerado a estas células como elementos de una representación cartesiana del ambiente (Nadel & Eichenbaum, 1999). Se ha demostramos que se puede producir sinaptogénesis en las fibras mugosas (FM) del hipocampo en el stratum oriens (SO), después de someter a ratas adultas a sesiones repetidas de aprendizaje espacial en el laberinto acuático de Morris (RamírezAmaya et al., 1999). Ramírez-Amaya, et al (2001), discutieron la relevancia funcional de la sinaptogénesis observada en el área CAS en la formación de la memoria espacial. En este trabajo se encontró que los animales que se entrenaron durante 5 días en el laberinto acuático son los que recordaban mejor la tarea una semana y hasta un mes después, comparado con aquellos animales que se habían entrenado uno o dos días. Además, estos datos se correlacionan positivamente con un incremento sustancial de las proyecciones de las FM hacia el SO en CAS, sugiriendo que estas nuevas proyecciones podrían relacionarse con la formación de la memoria a largo plazo y la representación espacial. El hallazgo de las células de lugar hizo que, en 1978, O’Keefe y Nadel propusieran la teoría del mapa cognitivo. Según la cual, el hipocampo podría ser la estructura cerebral a través de la cual se formaría un mapa cognitivo que permitiría a la rata conducirse en su entorno, es decir, el animal gracias a su hipocampo podría establecer una representación cerebral de las relaciones espaciales de su entorno apreciando 33 distancias y relaciones entre estímulos. Experimentos con animales en el MWM apoyan esta teoría, pues las lesiones de hipocampo como de otras estructuras de formación hipocampal causan un grave deterioro en esta tarea, también sugieren la implicación del sistema hipocampal en la formación de asociaciones o representaciones complejas entre los estímulos (Eichenbaum, 1999; D’Hooge y De Deyn, 2001). Esta idea concuerda con la evidencia neuroanatómica y neurofisiológica que muestra como el hipocampo íntegra múltiples áreas corticales relevantes para la percepción y la memoria, interviene en la codificación de las asociaciones entre las características espaciales y temporales de los estímulos ambientales (Daselaar, et al., 2006; Kesseis, et al., 2001). El hipocampo, por tanto, podría actuar como un “instrumento puente” que permitiría integrar los diferentes rasgos conductuales de la información en sus ambientes (Eichenbaum, 1999; Daselaar, et al., 2006). Se ha visto que las ratas con lesión hipocampal presentan déficits en la adquisición y retención de la tarea con plataforma escondida pero no en la tarea con plataforma visible. No obstante, las ratas con lesión hipocampal son capaces de localizar la plataforma escondida si se encuentra siempre en la misma distancia y dirección de una clave visible (Pearce, et al., 1998), demostrándose que la habilidad para usar un vector director se encuentra preservada en estos animales. El deterioro del aprendizaje espacial de las ratas con lesión esta proporcionalmente relacionado con el volumen de tejido hipocampal dañado. Asimismo, las lesiones del hipocampo dorsal producen mayor deterioro que las del hipocampo ventral (Moser, et al., 1993, Moser et al., 1995). Los déficits en el aprendizaje espacial que presentan las ratas con lesión hipocampal con ácido iboténico presentaron graves dificultades para aprender la plataforma escondida pero no para adquirir una tarea de discriminación no espacial (Gallagher & Holland, 1992) o una tarea de discriminación basada en la aversión al gusto (Skinner et al., 1994). Redish y Touretzky (1998) han propuesto dos funciones hipocampales en la solución de la tarea en plataforma escondida del MWM: autolocalización y repetición de la ruta. La autolocalización hace referencia a la capacidad de los animales para determinar su posición en el ambiente y en relación al objetivo (plataforma escondida). La repetición 34 de ruta es un proceso de memoria que permite al animal hacer y deshacer rutas de forma rápida dentro de ese ambiente. De acuerdo con estos autores estas capacidades dependen de la combinación de capos espaciales hipocampales y de los procesos de potenciación a largo plazo (PLP). Algunos autores han sugerido asimismo que la PLP hipocampal sería la base neuronal del aprendizaje hipocampo-dependiente, pero esta visión aún no ha sido probada de forma concluyente (Bliss, 1998). De hecho, otros autores han demostrado que el aprendizaje en el MWM es posible en animales donde se han bloqueado los procesos de PLP y también que la ejecución en el MWM puede afectarse cuando se facilita la PLP (Caín, 1997) El hipocampo forma parte de tres fases diferentes pero relacionadas: una fase de adquisición, una segunda fase de formación de la memoria y una tercera, que es la fase de recuperación. Durante la adquisición, recibe información de la corteza prefrontal que está relacionada con la corteza entorhinal y con el giro dentado (Fuster 2000). Probablemente otras regiones corticales (Frontal, cingular y occipital) también participan en el almacenamiento a largo plazo. Por ellos, se considera al hipocampo como un almacén transitorio de la información adquirida, hasta que se transfiere a un deposito a largo plazo (fundamentalmente a la corteza cerebral). Es además una estructura clave en la consolidación de la memoria y en los procesos de integración de los estímulos del medio externo (Squire & Zola-Morgan, 1988) Se ha observado que las ratas con lesiones en la corteza prefrontal, a diferencia de las que tienen lesiones en la corteza parietal, presentan déficits en la adquisición del aprendizaje en el MWM (Mogensen, et al., 1995). Las ratas con lesiones prefrontales parecen capaces de utilizar claves distales para localizar la plataforma escondida pero presentan déficits en los procesos de planificación, impidiéndose así la formación de una adecuada representación de la secuencia de movimientos necesaria para alcanzar la plataforma (Granon & Poucet, 1995; Mogense, et al., 1995). Algunos estudios han demostrad que los anímales con lesiones del fomíx pueden resolver una tarea espacial en el LAM guiándose por claves distales (Eichenbaum, et al., 1990). Lo que caracteriza la conducta de estos sujetos lesionados es la inflexibilidad 35 de su respuesta, mostrando una ejecución muy inferior a los sujetos control cuando se utilizan múltiples puntos de salida o se eliminan algunas de las claves ambientales, pero una ejecución normal cuando los ensayos reproducen exactamente las condiciones iniciales en las que se adquirió el aprendizaje (Eichenbaum, et al., 1990). McNaughton, et al., (1989) descubrieron unas células en la corteza parietal con propiedades complementarias a las células de lugar del hipocampo. Su actividad parece relacionarse con la orientación de la cabeza con respecto al entorno, independientemente de su localización, apoyando la hipótesis de que la corteza parietal integra y procesa tanto información visual como relativa al movimiento, ya que su lesión deteriora la ejecución en tareas espaciales. El hipocampo estaría más implicado en el procesamiento de señales distales y la corteza parietal con las proximales (Cressant, et al., 1997). La contribución de la corteza parietal al procesamiento de la información espacial podría depender del grado de funcionalidad del hipocampo (Thinus-Blanc, et al., 1996), aunque el papel de estas dos estructuras no puede aislarse de otras áreas cuya actividad está implicada en la integración y procesamiento que se hace del entorno. Por otro lado, se ha visto que las lesiones del núcleo accumbens, que actúa como una especie de “puente” entre el sistema límbico y motor (Ploeger, et al., 1994), deterioran la ejecución en el laberinto acuático de Morris. También se ha sugerido que el neoestriado (núcleo caudado y putamen) intervendría en la organización espacial de la conducta aunque de forma distinta al hipocampo. El putamen sería el sustrato neural de la localización espacial “alocentrica”, relacionada con la formación de los mapas cognitivos; mientras que el núcleo caudado intervendría en la localización “egocéntrica” (basada en la propia situación del cuerpo) (Devan, et al., 1996). Las lesiones cerebrales deterioran también la ejecución en el MWM, sugiriendo su implicación en los aspectos procedimentales de la tarea (D’Hooge y De Deyn, 2001). Otra región cerebral como el tálamo, participaría en el aprendizaje espacial actuando como principal estación de relevo (Conway, 1998). 36 El cerebelo es considerado el área cerebral esencial para el condicionamiento demorado de parpadeo (McCormick, et al., 1982). Este paradigma tiene la característica de ser un aprendizaje que se mantiene en animales con lesiones hipocampales. La amígdala participa fundamentalmente en algunos tipos de aprendizaje, como por ejemplo, el acercamiento a estímulos positivos o negativos, el condicionamiento del miedo y la preferencia de lugar condicionada (Davis, 1992; Fanselow & Kim, 1994; LeDoux, 2004; McDonald & White, 1993). El lóbulo temporal medial (y las estructuras díencefálicas) también se encuentran involucradas en algunos procesos de aprendizaje y memoria. Estas estructuras están implicadas en la capacidad para la recolección consiente de eventos (memoria declarativa) pero no son necesarias para algunas formas no consientes (memoria no declarativa).El sistema de memoria compuesto por el lóbulo temporal medial (hipocampo, corteza entorhinal, perirhinal y parahipocampal) funciona como un almacén temporal de la memoria, mientras que la neocorteza actuaría como un deposito más permanente y a largo plazo, y es independiente del funcionamiento del lóbulo temporal medial (P. Alvarez & Squire, 1984). El lóbulo temporal medial dirige el reconocimiento de experiencias recientes hacia los múltiples sitios corticales que constituyen una representación de la memoria; a medida que transcurre el tiempo luego del aprendizaje se produce una consolidación gradual de la memoria (Damasio, 1989; Squire, 1987). 37 3.6 Modelos experimentales para el estudio de la memoria en animales Existen distintas pruebas que fueron disenadas para estudiar procesos de memoria en animales, que consisten en exponer a los sujetos ante una situation de aprendizaje y luego evaluar la retention de la tarea realizada durante el entrenamiento. Los procedimientos experimentales mas utilizados con esta finalidad son: Laberinto radial de ocho brazos: El laberinto radial (radial-arm maze) desarrollado por Olton y Samuelson se utiliza en ratas y ratones para evaluar la memoria de trabajo (Olton & Samuelson, 1976), aunque tambien ha sido adaptado para estudiar conjuntamente este tipo de memoria y la de referencia (Beatty, 1984; Mishima, Higashitani, Teraoka & Yoshioka, 1986). El aparato utilizado habitualmente (figura 5) consiste en ocho brazos identicos, los cuales parten de un area central. Un recipiente con alimento (recompensa) se situa al final de cada brazo del laberinto. Esta tarea consiste en entrenar a los animales para que recorran el laberinto en busca del reforzador (alimento). En cuatro brazos del laberinto se coloca alimento, mientras que los restantes permanecen vacios (no reforzados). La medida dependiente que se registra es el numero de errores de la memoria de referencia (ingreso a brazos no reforzados). Tambien se tienen en cuenta otras medidas como el numero de errores de memoria de trabajo (reingreso a un brazo donde el reforzador ya fue consumido o a un brazo no reforzado), el ingreso a un brazo reforzado sin consumir la recompensa y la latencia de entrada a cada brazo (Jonasson, 2005). Figura 5 Laberinto radial de 8 brazos Tomada de http //homepage psy utexas edu/Homepage/Class/Psy308/Salinas/leaming/radialmaze g.f Laberinto de Barnes: esta prueba permite estudiar el aprendizaje y la memoria espacial, la memoria de trabajo y de referencia, y la memoria a corto y largo plazo, asi como otras tareas mas complejas en ratas y ratones. Fue disenado por Barnes en la Universidad de Arizona y consiste en un laberinto seco donde se utilizan luz y ruido como estimulos aversivos. El refuerzo que recibe el animal es escapar de estos estimulos. El laberinto esta constituido por un disco de plastico opaco elevado del suelo mediante un tripode (Figura 6). A lo largo del perimetro del disco se distribuyen 20 agujeros de 5 cm. de diametro y debajo de uno de ellos se coloca un cajon de escape de plastico opaco. Se pueden situar diversas pistas espaciales alrededor del cilindro, que permaneceran constantes durante todo el estudio, y tambien se pueden colocar debajo de algunos agujeros cajones falsos en los que parece posible introducirse, pero cuyo espacio disponible es en realidad tan pequeno que impide que el animal se pueda esconder. Un foco de luz (de unos 150W) y un generador de ruido (sobre 80 dB) constituyen los estimulos aversivos. Figuta 6 labennto de Barnes Tomada de http /www med-associates com'maies'bames l*m El procedimiento comienza con un entrenamiento que cosiste en colocar el animal en la caja de escape y dejarlo alii durante un minuto, para continuar un minuto mas tarde con las diversas sesiones de exploration. Todas las sesiones empiezan colocando el animal en el centro del laberinto en el interior de una "camara de initio", que consiste en un cilindro opaco de unos 10 cm de alto. Al cabo de 10 segundos, se retira la camara de inicio, se conectan la luz y el zumbido, y se deja que el animal explore el laberinto. El estrés producido por estos estímulos induciría al animal a buscar un espacio cerrado donde esconderse. La sesión termina cuando el animal encuentra el túnel de escape o cuando transcurren 5 minutos, momento en el que se le introduciría manualmente en dicho túnel. Cuando el animal entra en la cámara de escape, los estímulos aversivos cesan y se le deja en la oscuridad un minuto antes de devolverlo a su jaula. El túnel de escape está situado siempre debajo del mismo agujero, que se elige de forma aleatoria para cada animal. La prueba se realiza durante varios días hasta que en siete u ocho pruebas consecutivas se produzcan menos de tres errores. Se valoran el aprendizaje espacial y la memoria midiendo el tiempo que invierte el animal en encontrar el cajón refugio y el número de errores 40 4 Planteamiento del problema Como se ha descrito anteriormente las variaciones metodológicas con respecto al uso del MWM, han permitido identificar factores que pueden afectar el rendimiento del animal en dicha prueba, tales como; el sexo, especíe/cepa, edad, estado nutricional, la exposición al estrés o la infección. Así como las lesiones en las distintas áreas del cerebro: hipocampo, estriado, cerebelo, cerebro anterior basal y corteza cerebral (D’Hooge et al., 2001). A pesar de su gran utilidad, no se ha reportado cual es el número de ensayos óptimo en cada día de entrenamiento para producir una curva de aprendizaje progresiva. Lo que se sabe es que ante repetidos intentos se logra disminuir el promedio de latencia en los días de entrenamiento y que un valor de 10 segundos en promedio es indicativo para finalizar el entrenamiento (Jett et al., 1997).Sin embargo, este criterio no es utilizado en los protocolos analizados. Ante esta diversidad del número de intentos y los días utilizados para ello nos planteamos la siguiente interrogante: ¿el número de intentos y los días de entrenamiento realizados en el protocolo implementado en el laboratorio son suficientes para producir una curva de aprendizaje de las ratas Wistar macho, de 21 días post-natales? O es necesario incrementar los días de entrenamiento y los intentos por día? 5 Justificación A pesar de que existe una variedad de protocolos para evaluar el aprendizaje y la memoria espacial en roedores, no se ha encontrado algún reporte que mencione el número de intentos y dias de entrenamiento idóneos para obtener una curva de aprendizaje progresiva. Por tal motivo, en este trabajo se pretende determinar si el número de intentos y dias de entrenamiento utilizados en el protocolo implementado en el laboratorio de Neurotoxicología requiere ser modificado. 6 Hipótesis Un entrenamiento de seis intentos durante tres dias es suficiente para generar una curva de aprendizaje. 41 7 Objetivos 7.1 Objetivo general Determinar si el incrementar el número de intentos y días de entrenamiento el MWM produce una curva de aprendizaje progresiva mayor que la obtenida con el protocolo implementado en el laboratorio de Neurotoxicología. 7.2 Objetivo particular Comparar las diferentes curvas de aprendizaje progresiva obtenida con seis y ocho intentos por tres y cinco días de entrenamiento. 42 GRUPO 1 Día 21 21 22 24 23 Días posl-natales GRUPO 2 Día 22 21 21 24 23 Dias post-natales Día GRUPO 3 21 21 22 23 24 25 26 Días post-natales Dia GRUPO 4 21 21 22 23 24 25 26 Dias post-natales F ig u ra 7: Diseño experim ental para cada grupo 44 Al finalizar los días de entrenamiento cada grupo tuvo un día de “prueba de ensayo” o también conocida como “prueba final” en la cual solo se hace un intento. 8.3 Técnica para uso de Laberinto Acuático de Morris El MWM consistió en una piscina de paredes lisas con un diámetro de 85 cm y 50 cm de profundidad con una plataforma de acrílico de 4.7x 4.7 cm cuyo soporte tiene una altura de 33 cm. En las paredes del cuarto donde se colocó la piscina se pegaron 4 figuras tridimensionales (triángulo equilátero, circulo, cuadrado y pentágono) que sirven de referencia a los animales. La piscina estuvo dividida en cuatro cuadrantes. Durante la prueba la rata se colocó en un cuadrante diferente en cada intento; el cuadrante fue asignado al azar, omitiendo en el sorteo el número del cuadrante donde se encuentra la plataforma. Cada grupo se sometió a la prueba de MWM, el cual consta de las siguientes etapas: Pre-entrenamiento: se hizo únicamente el día uno de experimento, durante esta etapa la plataforma es visible, se hicieron 3 intentos y se le dan 30 segundos de nado para que encuentre la plataforma, 10 segundos de descanso sobre la plataforma y 30 segundos de descanso en la caja de ambientación. Entrenamiento: en esta etapa la plataforma fue oculta, el número de intentos y días variará de acuerdo al grupo experimental con que se esté trabajando. Se le permite a la rata 30 segundos de nado para que encuentre la plataforma, 10 segundos de descanso sobre la plataforma y 30 segundos de descanso en la caja de ambientación. 51 durante los 30 segundos que se le dan a la rata para que encuentre la plataforma no lo logra, entonces el entrenador deberá guiar a la rata hacia la plataforma y posteriormente ser tratada como a las que si encontraron la plataforma. Prueba de ensayo: se realizó el día posterior al último día entrenamiento y se hizo un intento, la plataforma se retiró de la piscina, el cuadrante del que partió la rata es elegido al azar y se dan 30 segundos de nado libre. 45 En el cuarto de conducta se colocó una cámara en la parte central del techo la cual va conectada a una computadora que se localiza en la parte exterior del cuarto, se fue grabando el recorrido que hace la rata durante la prueba y posteriormente los videos fueron analizados con el software OASPADVid para el procesamiento y adquisición de datos a través de video. 46 9 A nálisis estadístico Los datos se presentan como medias ± error estándar de la media (S.E.M.) Para la comparación de los resultados obtenido en los días de entrenamiento se utilizó ANOVA de dos vías de medidas repetidas y para comparar el tiempo de permanencia en el cuadrante target se utilizó una ANOVA de una vía, seguido de una prueba post­ hoc de Bonferroni. Utilizando el programa Sigma Stat versión 3.5. El valor de P :S0.05 se considera como significativo. 10 Resultados Respecto al test conductual tenemos los siguientes resultados: a).- Efecto del número de intentos sobre la curva de aprendizaje: GRUPO 3 GRUPO 4 — GRUPO 1 GRUPO 2 S. ra Ti 20-1 ♦- '-'í> s 2 10 ' ' i 3 O'ia Figura 8.- Efecto del núm ero de intentos sobre la curva de aprendizaje. En el grupo uno se utilizaron 6 intentos por tres días de entrenamiento. En el grupo dos se utilizaron 8 intentos por tres días de entrenamiento. Obsérvese que no existe diferencia significativa |F (2.42)=1.014; P=0.338] entre los gmpos. Los datos se representan como la m edia ± E.S.M. Figura 9 .- E fec to d e l n ú m e ro d e in ten to s s o b re la c u rv a de ap re n d iza je . En el grupo tres se utilizaron 6 intentos por dnoo días de entrenamiento. En el grupo cuatro s e utilizaron 8 intentos por cinco días de entrenamiento. O bsérvese que no existe diferencia significativa F (2 .7 0 )= 3 .9 4 ; P = 0.09 5) entre los grupos. Los datos s e representan como la m e d ia ± E.S.M. 47 b).- Al realizar el analisis del tiempo promedio de latencia obtenido en el dia tres de entrenamiento observamos lo siguiente: 20 A _ 15 v> GRUPO2 A GRUPO3 6 9 10 GRUPO 1 CS GRUPO 4 3 Oia Figura 10.- Efecto del numero de entrenamiento sobre el promedio de latencia en el tercer dia de entrenamiento. En donde osservamos que existen diferencias signrficativas (P<0.001) entre los animales de los grupos 1. 2. 3 y 4. Los datos se representan como la media ± E.S.M. c).- Distancia recorrida por los roedores durante la prueba de laberinto acuatico de Morris 40C GRUP01 GRUPO2 30C Z 20C 5 IOO P<0.001 2 Dx Figura 11.- Distancia recorrida por los roedores para localizar la plataforma en el MWM. En el grupo uno se utilizaron 6 intentos por tres dias de entrenamiento. En el grupo dos se utilizaron 8 intentos por tres dias de entrenamiento . O&servese que existe diferenaa significativa (P<0.001) entre amoos grupos. Los datos se representan como la media ± E.S.M. •• GRUPO 3 GRUPO 4 Figura 12.- Distancia recorrida por los roedores para localizar la plataform a en el MWM. En el grupo tres se utiizaron 6 intentos por cinco días de entrenamiento. En el grupo cuatro se utilizaron 8 intentos por cinco días de entrenamiento. Obsérvese que no existe diferencia significativa (P =0.096) entre los grupos. Los datos se representan como la media ± E.S.M. d).- Velocidad de nado utilizada por los roedores que conformaron los grupos de estudio. Oía Figura 13.- Velocidad de nado de los roedores en el MWM. En el grupo uno se realizaron 6 intentos por tres días de entrenamiento. En el grupo dos se realizaron 8 intentos por tres días de entrenamiento Obsérvese que existe diferenda significativa (P<0.001) entre ambos grupos Los datos se representan como la m e d ia l E.S.M. 49 •. GRUP03 •t GRUPO4 P=0.011 Figura 14.- Vélocídad de nado de los roedores en el MWM En el grupo tres se realzaron 6 intentos por cinco días de entrenamiento. En el grupo cuatro se realizaron 8 intentos por cinco días de entrenamiento. ODsétvese que existe diferencia significativa (P=0.011) entre ampos grupos. Los datos se representan como la media tE.S.M. e).-lntegrando los valores obtenidos de los parámetros evaluados al día tres de entrenamiento en el MWM podemos observar los siguientes resultados; 1 Grupo 4 Diferencia estadísticamente Grupo 1 Tiempo de latericia Velocidad de nado 12.6311.4 16.2811.16 14.6611.17 1511.39 Si P<0.001 15.0510.64 13.3910.62 9.9410.74 16.0310.74 205.21l27.83 225.81127.09 147.68l16.79 218.64124.05 Si (1vs2, 2vs4, 2vs3) P<0.001. No (1vs4) P>0.05 No P>0.05 Distancia recorrida Grupo 3 Grupo 2 Parámetros evaluados 8 intentos 6 intentos significativa Tabla 1.- Parámetros eva uados en el día tres de adquisición del aprendizaje. En la presente tabla se muestran los resultados obtenidos en los parámetros evaluados, al día tres, durante la adquisición del aprendizaje en el laberinto acuático de Morris. Los resultados se presentan como media ± E.S.M. 50 f).- Prueba de ensayo. Se grafican el promedio del tiempo de permanencia en el cuadrante en el que estuvo el roedor el mayor tiempo durante la prueba de ensayo. Coincidentemente es el cuadrante target para todos los grupos. O GRUPO 1 GS3 G R U P O 2 O GRUPO 3 Q GRUPO 4 3 Dia Figura 15. Tiempo de permanencia de los roedores en el cuadrante objetivo (target).Despues de la fase de adquisicion de aprencSzaje se evalua la memoria. Todos los roedores no mostraron diferencia significativa en el desempeno de esta prue&a (P>0.05). 11 Discusión Los resultados obtenidos en cuanto al aprendizaje, revelan que no existe diferencia significativa (P=0.338) entre realizar 18 o 24 intentos, a pesar de que gráficamente se observa una curva progresiva cuando se realizan 24 intentos (figura 8). No obstante ambas curvas reflejan la adquisición del aprendizaje espacial debido a la diferencia significativa entre los días de entrenamiento 1 y 3 (P<0.001) ó 2 y 3 (P= 0.003), criterio que ha sido observado en los trabajos de Morris, G. (1981); Jett, D., et al (1997); Prados, J. & Trobalon, F. (1998) para demostrar que el roedor adquiere un aprendizaje espacial. Lo mismo sucede al incrementar el número de intentos a 30. Sin embargo, esto se modifica cuando se entrena a los roedores con 40 intentos (figura 10), el promedio de latencia incrementa y esto se debe quizás a un cansancio del roedor al ejecutar la tarea, el cual se ve reflejado en el incremento del tiempo. Por otro lado, en la curva que se obtiene con los 40 intentos (figura 9) se observa una evidencia de aprendizaje hasta el quinto día, cuando se presenta diferencia significativa (P=0.030) al comparar el tiempo promedio de latencia del primer día contra el quinto. Dentro de las implicaciones que esto tiene se encuentra un mayor tiempo de mantenimiento de los roedores, lo que impacta en un gasto mayor en la alimentación de los mismos. En cuanto a la distancia y la velocidad de nado, que son las conductas más específicas del desempeño de los roedores(Gerlai, 2001), tenemos que el grupo de roedores que recorrió menor distancia fue el dos con 205.4 cm, lo que sugiere que ubicó la plataforma en forma más precisa y no requirió buscarla por toda el área del laberinto; no obstante su velocidad de nado fue 10.51 cm/S.( es la menor de todos los grupos) y se esperaría que fuese la mayor, debido a que ha aprendido el lugar donde se ubica la plataforma. No obstante, al comparar los parámetros evaluados entre todos los grupos el día tres de entrenamiento (tabla 1), encontramos diferencias significativas (P<0.001) en latencia entre los cuatro grupos, sin embargo, aunque existan estas diferencias, lo que para nosotros resulta relevante es el grupo que obtiene el menor tiempo en encontrar la 52 plataforma, en este caso sería el grupo uno. En cuanto a la velocidad, encontramos que el grupo dos es significativamente diferente (P<0.001) al grupo uno, tres y cuatro; en este caso lo que nos interesa es el grupo que obtiene la mayor velocidad de nado, en este caso es el grupo cuatro seguido del grupo uno, los cuales no presentan diferencias significativas (P>0.050) entre sí, por lo tanto nosotros elegimos el grupo uno, ya que sólo se realizan seis intentos por día de entrenamiento ( tres) y se obtiene un resultado similar al grupo cuatro que realiza ocho intentos por día de entrenamiento (cinco). En la distancia recorrida no encontramos diferencias significativas (P>0.050) entre los grupos, el grupo que nos interesa de este parámetro es el que recorre la menor distancia para localizar la plataforma, en este caso sería el grupo dos, pues es el que demuestra haber recorrido menor distancia para localizar la plataforma, sin embargo, también podríamos elegir el grupo uno, aunque muestra un valor más elevado comparado con el grupo dos, ya que estadísticamente no presentan diferencias significativas y en el grupo uno se realizan 3 días de entrenamiento con 6 intentos y en el grupo dos ocho intentos en igual tiempo. Por otro lado, durante la prueba de ensayo (figura 15) que valora la memoria adquirida por los roedores, no encontramos diferencias significativas (P>0.050) en el tiempo que pasaron en el cuadrante objetivo, por lo tanto, todos adquirieron la estrategia para encontrar la plataforma por lo que memorizaron la ubicación de la misma. 53 12 Conclusión Los resultados obtenidos apoyan que el mejor protocolo para obtener una curva de aprendizaje progresiva requiere 18 intentos distribuidos en tres días de entrenamiento. El incrementar los intentos o los días de entrenamiento no mejoran dicha curva. Por lo tanto, este resultado permite verificar el protocolo utilizado en el Laboratorio de Neurotoxicología para evaluar el aprendizaje y la memoria espacial de roedores de 21 días postnatal utilizando el Laberinto acuático de Morris 54 13 Bibliografía 1) Alvarez, P. & Squire, L.R. (1994). Memory consolidation and the medial temporal lobe: a simple network model. Proceedings of the National Academy of Sciences USA. 91(15). 7041-7045 2) Baddeley, A. & Hitch, G. (1979). Working Memory in G.H. Bower. The Psychology of Learning and Motivation. 8: 36-89 3) Barnes, C.A. (1973). Memory deficits associated with senescence: a neurophysiological and behavioral study in the rat. J. Comp. Phychol. 93: 74-104 4) Bliss, T.V.P. (1998). The saturation debate. Science. 281:1975-1976 5) Brandéis, R., Brandys, Y. & Yehuda, S. (1989). The use of the Morris water maze in the study of memory and learning. Int. J. Neuroscience. 48: 29-69. 6) Bruce-Keller, A.J., Umberger, G., McFall, R. & Mattson, M.P. (1999). Food restriction reduces brain damage and improves behavioral outcome following excitotoxic and metabolic insults. Annals of Neurology. 45: 8-15 7) Cain, D.P. (1997).LTP, NMDA, genes and learning. Current Opinion in Neurobiology. 7: 235-242 8) Chamizo, V.D. & Rodrigo, T. (2004). Effect of absolute spatial proximity between a landmark and a goal. Learning and Motivation 35:102-114 9) Chamizo, V.D., Manteiga, R.D., Rodrigo, T. & Mackintosh, N. J. (2006). Competition between landmarks in spatial learning. The role of proximity to the goal. Behavioural Processes. 71: 59-65 10) Conway, E. L. (1998). Brain lesions and delayed water maze learning deficits after intracerebroventricular spermine. Brain Research. 800: 10-20 11) Crabbe, J. C., .Wahlsten, D. & Dudek, B. C. (1999). Genetics of mouse behavior: interactions with laboratory environment. Science. 284: 1670-1672. 12) Cressant, A., Muller, R.U. & Poucet, B. (1997). Failure of centrally placed objects to control the firing fields of hippocampal place cells. Journal of Neuroscience. 17: 2531-2542. 13) Dalm, S., Grootendorst, J., De Kloet, E.R. & Oitzl, M L. (2000) Quantification of swim patterns in the Morris water maze. Behav. Res. Methods 32: 134-139. 55 14) Damasio, A.R. (1989). Time- locked multiregional retroactivation: a systems level proposal for the neural substrates of recall and recognition. Cognition. 33(1-2). 25-62. 15) Daniel, J.M., Roberts. S.L. & Dohanich, G.P. (1999). Effects of ovarian hormones and environment on radial maze and water maze performance of female rats. Physiology and Behavior 66: 11-20 16) Daselaar, S.M., Fleck, medial temporal lobes: M.S. & Cabeza, R. (2006). Triple dissociation in the recollection, familiarity and novelty. Journal of Neurophycology. 96:1902-1911. 17) Davis, M. (1992). The role of the amygdale in conditioned fear. In J.P. Aggleton (Ed.) The amygdala: Neurobiological Aspects of Emotion Memory, and Mental Dysfunction. New York: Willey 18)De Quervain, D.J., Roozendaal, B. & McGaugh, J.L (1998). Stress and glucocorticoids impair retrieval of long-term spatial memory. Nature. 394: 787­ 790. 19) D'Hooge, R., De Deyn, P.P. (2001). Applications of the Morris water maze in the study of learning and memory. Brain Res Brain Res Rev. 36(1). 60-90. 20)D’Hooge R., Franck, F., Mucke, L. & De Deyn, P.P. (1999). Age-related behavioural deficits in transgenic mice expressing the HIV-1 coat protein gp 120, Eur. J. Neurosci. 11:4398-4402 21) Dieter, S.E: (1977) Preexposure to situational cues and shock intensity in two­ way avoindance learning. Animal Learning and Behavior. 5: 403-406 22) Eichenbaum, H., Stewart, C. & Morris, R.G. (1990). The medial temporal lobe and recognition memory. Animal Reviews of Neuroscience. 30: 123-152 23) Eichenbaum, H. (1999). The hippocampus and mechanisms of declarative memory. Behavioural Brain Research. 103:123-133 24) Etchepareborda, M.C. & Abad-Mas, L (2005).Memoria de trabajo en los procesos básicos del aprendizaje. Neurología. 40 (Sup11). S79-S83 25) Fanselow, M.S. & Kim, J.J. (1994). Adquisition of contextual paviovian fear conditioning is blocked by application of an NMDA receptor antagonist D,L-2 - 56 amino-5- phosphonovaleric acid to the basolateral amygdala. Behavioral Neuroscience. 108(1). 210-212. 26) Fraenkel, G.S., & Gunn, D.L. (1961). The orientation of animals. New York: Dover. 27) Fuster, J. M. (2000). Prefrontal neurons in networks of executive memory. Brain Research Bulletin. 52(5). 331-336. 28) Gallaher, M. & Holland, P.C. (1992). Preserved configural learning and spatial learning impairment in rats with hippocampal damage. Hippocampus. 2: 81-88 29) Gallagher, M. & Nicolle, M.M. (1993). Animal models of normal aging: relationship between cognitive decline and markers in hippocampal circuitry, Behav. Brain Res. 57: 155-162. 30) Geinisman, Y., Detoleddo-Morrell, L , Hippocampal markers of aged-related Morrell, F. & Heller, R.E. (1995) memory dysfunction: behavioral, electrophysiological and morphological perspectives. Prog. Neurobiol. 45: 223­ 252. 31) Gerlai, R. (2001) Behavioral test of hippocampal function simple paradigms complex problems. Behavioral Brain Research. 125: 269-277 32) Gibertini, M., Newton, C., Friedman, H. & Klein, T.W. (1995). Spatial learning impairment in mice infected with Legionella pneumophila administered exogenous interleukin-1-b. Brain Behav. Immun. 9: 113-128. 33) GranoN, S. & Poucet, B. (1995) Medial prefrontal lesions in the rat spatial navigation: evidence for impaired planning. Behav. Neuroscience. 109:474-484. 34) Holscher C. (1999). Stress impairs perfonnance in spatial water maze learning tasks, Behav. Brain Res. 100: 225-235. 35) lkonen, S., Schmidt, B. H. & Riekkinen Jr., P. (1999). Characterization of learning and memory behaviors and the effects of metrifonate in the C57BL strain of mice, Eur. J. Pharmacol. 372: 117-126. 36) lzquierdo, I., Bevilaquia, L.R., Rossato, J.L., Bonini, J.S., Medina, J.H. & Cammarota, M. (2006). Different molecular cascades in different sites of the brain control memory consolidation. Trends in Neurosciences. 29(9). 496-505. 57 37) Jett, A D., Kuhimann, C.A., Farmer, J S. & Guilarte, R.T. (1997). Age-dependent effects of developmental lead exposure on performance in the Morris water maze. Pharmacol Biochem Behav. 57: 271-279. 38) Kessels, R.P.C., De Haan, E.H.F., Kappele, L.J. & Postma, A. (2001). Varieties of human spatial memory: a meta-analysis on the effects of hippocampal lesions. Brain Research Reviews. 35; 295-303. 39) Lamberty, Y. & Gower, A. J. (1991). Simplifying environmental cues in a Morristype water maze improve place learning in old NMRI mice. Behavioral and Neural Biology. 56:89-100 40) Lamberty, Y. & Gower, A.J. (1993). Spatial processing and emotionality in aged NMRI mice: a multivariate analysis. Physiol. Behav. 54: 339-343. 41) LeDoux, J.E. (2004). Emotion and the amygdala. In J.P. Aggleton (ed.) The amygdala; neurobiological aspects of emotion memory and mental dysfunction. 339-351. 42) Lindner, M.D. (1997). Reliability, distribution, and validity of age-related cognitive deficits in the Morris water maze, Neurobiol. Learn Mem. 68: 203-220. 43) Lipp, H.P. & Wolfer, D.P. (1998). Genetically modified mice and cognition Curr. Opin. Neurobiol. 8: 272-280 44) Machado, S., Portella, C.E., Silva, J.G., Velasques, B., Bastos, V.H., Cunha, M., Basile, L., Cagy, M., Piedade, R.A. & Ribeiro, P. (2008). Aprendizaje y memoria implícita; mecanismos y neuropiasticídad. Neurología. 46(9): 543-549 45) McCormick, D.A., Clark, G.A., Lavond, D.G. & Thompson, R.F. (1982). Initial localization of the memory trace for a basic form of learning. Proceedings of the National Academy of Sciences USA. 79(8). 2731-2735 46) McDonald, R.J. & White, N.M. (1993). A triple-dissociation of memory systems: hippocampus, amygdala and dorsal striatum. Behavioral Neuroscience. 107(1). 3-22. 47) McLean, J.H., Shipley, M.T, Bernstein, D.l & Corbett, D. (1993). Selective lesions of neural pathways following viral inoculation of the olfactory bulb, Exp. Neurol. 122; 209-222 58 48)McNaughton, B.L., Leonard B. & Chen L.L. (1989). Cortical hippocampalinteractions and cognitive mapping. A hypotheses based on reintregation of tne parietal and inferotemporal pathways for visual processing. Psychobioloy. 17; 236-246. 49) Means, L.W., Higgins, J.L. & Fernandez, T.J. (1993). Mid-life onset of dietary restriction extends life and prolongs cognitive functioning. Physiol. Behav. 54:503-508. 50) Menzel, E. W. (1978) Cognitive mapping in chimpanzees. In S. H. Hulse, H. F. Fowler, & W. K. Honig (Eds.), Cognitive aspects of animal behaviour. Hillsdale, N.J.: Erlbaum. 51) Milner, B., Squire, L. R. & Kandel, E. R. (1998). Cognitive neuroscience and the study of memory. Neuron, 20(3): 445-468. 52) Mogensen, J., Pedersen, T. K., Holm, S. & Bang, L.E. (1995). Prefrontal cortical mediation of rats place learning in a modified water maze. Brain Research Bulletin. 38: 425-434. 53) Morgado, I. (2005). Psicobiologia del aprendizaje y la memoria. CIC (Cuadernos de información y comunicación). 10: 221-233 54) Morris, R. G. (1981). Spatial localization does not require the presence of local cues. Learning and motivation (12); 239-260. 55) Morris, R. (1984). Developments of a water-maze procedure for studying spatial learning in the rat. Journal of Neuroscience Methods. 11: 47-60 56) Morris, R. G. (2008). Morris water maze. Scholarpedia, 3(8):6315. 57) Moser, E., Moser, M.B. & Andersen, P. (1993). Spatial learning impairment parallels the magnitude of dorsal hippocampal lesions, but is hardly presents following ventral lesions. The journal of Neuroscience. 13: 3916-3925rain Research. 63: 47-60 58) 0 ’Keefe, J.A. & Dostrovsky, J. (1971). The hippocampus as spatial map preliminary evidence from unit activity in the freely-moving rat. Brain Research. 34(1): 171-175 59) 0 ’Keefe, J., Nadel, L., Keightly, S., & Kill, D. (1975). Fornix lesions selectively abolish place learning in the rat. Experimental Neurology. 48: 152-166 59 60) O’Keefe, J.A & Nadel, L. (1978). The hippocampus as a cognitive map. New York: Oxford University Press. 61) Olton, D.S., Becker, J.T., & Handleman, Cl. E. (1979). Hippocampus, space and memory. The Behavioural and Brain Sciences. 2; 313-365. 62) Pearce, J.M., Robert, A.D & Good, M. (1998) Hippocampal lesions disrupt navigation based on cognitive maps but not heading vectors. Nature. 396:75-77 63) Pitsikas, N., Carli, M., Fidecka, S. & Algeri, S. (1990). Effect of life-long hypocaloric diet on age-related changes in motor and cognitive behavior in a rat population, Neurobiol. Aging 11: 417-423. 64) Ploeger, G.E., Spruijt, B.M. & Cools, A.R. (1994). Spatial localization in the Morris water maze in rats: adquisition is affected by intr-accumbens injections of the dopaminergic antagonist haloperidol. Behavioral Neuroscience. 108: 927-934 65) Prados, J. & Trobalon, J.B. (1998). Locating an invisible goal in a water maze requires al least two landmarks. Psychobiology. 26: 42-48. 66) Redolar Ripoll, D., Moreno Alcázar, A., Robles Muñoz, N., Soriano Mas, C., Torras García, M., & Vale Martinez, A. M. (2010). Fundamentos de psicobiología. Editorial UOC. 67) Rodrigo, T., Chamizo, V.D., McLaren, I.P.L. & Mackintosh, N. J. (1997). Blocking in the spatial domain. Journal of Experimental Psychology. Animal Behavios Processes. 23: 110-118 68) Roof, R.L. (1993). Neonatal exogenous testosterone modifies sex difference in radial arm and Moms water maze performance in prepubescent and adult rats. Behavioral Brain Research. 53:1-10 69) Ruetti, E., Justel, N., & Bentosela, M. (2009) Perspectivas clásicas y contemporáneas acerca de la memoria. Suma Psicológica. 16(1): 65-83 70) Sandi, C., (1998). The role and mechanisms of action of glucocorticoid involvement in memory storage. Neural Plast. 6: 41-52 71) Santin, L.J., Rubio, S., Begega, A., Miranda, R. y Arias, J.L. (2000). Aprendizaje espacial e hipocampo. Revista de Neurología, 31:455-462. 72) Skinner, D.M., Martin, G.M., Harley, C., Kolb, B., Pridgar, A., Bechara, A. & Vander-Kooy, D. (1994). Acquisition of conditional discriminations in hippocampal 60 lesioned and decorticated rats: Evidence for learning that is separate from both simple classical conditioning and configural learning. Behavioral Neuroscience. 108:911-926. 73) Soriano Mas, C., Quillazo Blanch, G., Redolar Ripoll, D. A., Torras García, M., & Vale Martinez, A. (2007). Fundamentos de Neurociencia. Editorial UOC. 74) Squire, L.R. (1987). Memory and brain. New York. Oxford University Press. 75) Squire, L.R. & Zola-Morgan, S: (1988). Memory: brain systems and behavior. Trends is a Neurosceinces. 11(4). 170-175. 76) Squire, L. R. (2004). Memory systems of the brain: a brief history and current perspective. Neurobiology of Learning and Memory, 82(3). 171-177. 77) Stewart, J., Mitchell, J. & Kalant, N. (1989) The effects of life-long food restriction on spatial memory in young and aged Fischer 344 rats measured in the eight-arm radial and the Morris water mazes,. Neurobiol. Aging 10: 669-675. 78) Stewart, C.A. & Morris, R.G. (1993) The watermaze, in: A. Sahgal (Ed.), Behavioural Neuroscience. A Practical Approach, Vol. 1, IRL Press, Oxford. 107-122. 79) Thinus-Blanc, C., Save, E., Rossi-Amaud, C., Tozzi, A. & Ammassari-Teule, M. (1996). The differences shown by C57BL/6 and DBA/2 inbred in detecting spatial novelty are subserved by a different hippocampal and parietal cortex interplay. Behavioral Brain Research. 80: 33-40 80) Vicens, P., Redolat, R., & Carrasco, M. C. (2003) Aprendizaje espacial y laberinto de agua: metodología y aplicaciones. Psicothema 15(4), 539-544 81) Warren, S.G. & Juraska, J.M. (1997) Spatial and nonspatial learning across the ratestrous cycle, Behav. Neurosci. 111: 259-266. 82) Watson, J. B., & Lashiey, K. S. Homing and related activities & birds. Washington, D.C. Carnegie Institute. 83) Wenk, G.L. (1998). Assessment of spatial memory using radial arm and Morris water mazes. In: J. Crawley, C. Gerfen, R. McKay, M. Rogawski, D. Sibley and P. skolnick. Editors, Current Protocols in Neuroscience Wiley, New York 84) Wongwildecha, N. & Marsden, C.A. (1996). Effects of social isolation rearing on learning in the Morris water maze. Brain Research. 715:119-124 61