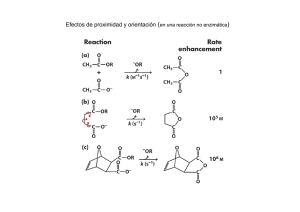
Evaluació celul ón de la lolíticos degrada aislado ación de os de
Anuncio

1 ón de la degradaación deel tamo de arro oz por hongos Evvaluació celullolíticos aislado os de sueelos de cultivo de arro oz An ndrea Meelissa Sán nchez Gaaribello Universid dad Nacionaal de Colom mbia Instittuto de Biottecnología B Bogotá, Colombia 2013 2 Evaluación de la degradación del tamo de arroz por hongos celulolíticos aislados de suelos de cultivo de arroz Andrea Melissa Sánchez Garibello Tesis o trabajo de investigación presentada(o) como requisito parcial para optar al título de: Magister en Ciencias ‐ Microbiología Directora: M. Sc., PhDc. Ivonne del Socorro Gutiérrez Rojas Codirector: PhD . Daniel Uribe Vélez Universidad Nacional de Colombia Instituto de Biotecnología Bogotá, Colombia 2013 3 A mi mamá por el amor, y los sacrificios para hacer de mí la persona que soy. A mis abuelos por la enseñanza de vida, el amor incondicional, y sus cuidados ahora desde el cielo. A mi hermana por estar para mí siempre y por todo su amor. 4 Agradecimientos Agradezco enormemente a Dios por cada día de aprendizaje y por poner en mi camino a tantas personas maravillosas, porque sin todas ellas nada de esto sería posible. A la Universidad Nacional de Colombia por darme la oportunidad de crecer como profesional, a la doctora Ivonne Gutiérrez Rojas por acogerme y creer en mi trabajo, al doctor Daniel Uribe por el acompañamiento constante y los aportes a este proyecto, a la Pontifica Universidad Javeriana, especialmente al Grupo de biotecnología Ambiental e Industrial, Laboratorio de Biotecnología, por brindarme la oportunidad de desarrollar el proyecto en sus instalaciones ,y por todo el apoyo, enseñanzas y compromiso a la doctora Balkys Quevedo Hidalgo. A mi familia, especialmente a mi mamá quien me ha apoyado desde el principio en este difícil pero hermoso camino, a mi papá, a mi hermana que siempre tiene palabras de aliento, a mi cuñado por sus desvelos y cuidados y a mis sobrinos porque le imprimen el toque de alegría a todo lo que hago. A mis amigos del alma y compañeros de trabajo , por apoyarme siempre, creer en mí, y por todo su cariño, Marcela Ramos, Shirley Rojas, Carel Carvajal, Iván Ávila, María Alejandra Jaramillo, Laura Muñoz, Marco Rincón, Jessica Díaz, Nicolás Cárdenas, Sergio Ordúz, Ángela Alvarado , Yara Cruz, Viviana Ávila, Deisy Toloza, Mauricio Aguirre, Mónica López , Camila Cardozo, Jorge Moreno. A la doctora María Ximena Rodríguez, Aura Marina Pedroza, Martha Fontanilla y Socorro Prieto por su colaboración. Y finalmente a quienes ya no están en mi vida pero de una forma u otra influyeron para que mi camino tomara este rumbo, el cual no cambiaría por nada del mundo. 5 6 V Resumen El cultivo del arroz es de importancia económica para el país ya que anualmente registra una producción de alrededor de 20x105 toneladas de arroz paddy seco, lo cual se traduce en ganancias importantes en el sector agrícola. Sin embargo después del proceso de cosecha se genera una cantidad de residuo igual o mayor al producto obtenido. El objetivo de esta investigación fue evaluar la capacidad de los microorganismos aislados de suelos del cultivo de arroz para degradar el residuo conocido como tamo o paja para ser reincorporado al suelo. Se estableció que de los 16 aislamientos evaluados Penicillium sp MF 3‐64 y Paecilomyces sp. presentaron actividad degradativa promisoria debido a la producción de celulasas totales a partir de caldo tamo, halos de hidrolisis en agar celulosa y producción de CO2 a partir de la degradación del sustrato. Se caracterizó la actividad enzimática de los dos aislamientos seleccionados por medio de un diseño factorial irregular, empleando como factores la fuente de carbono y nitrógeno con 2 y 3 niveles respectivamente las variables de respuesta fueron las actividades enzimáticas endoglucanasa, celobiohidrolasa, β‐ glucosidasa, encontrándose que la carboximetilcelulosa y el sulfato de amonio tienen una influencia positiva en la producción enzimática (p˂0,005). Adicionalmente, se determinó la influencia de la fuente de nitrógeno en la producción de xilanasas obteniéndose mayor actividad enzimática en presencia de la misma. Se encontró que Penicillium sp. produce 4000 UE/L de β‐glucosidasa a diferencia de Paecilomyces sp. que produce 225 UE/L; esta tendencia se encontró en las demás actividades enzimáticas evaluadas. En la evaluación de Penicillium sp. en fermentación solida empleando el residuo como sustrato, se obtuvieron actividades enzimáticas de 747 UE β‐glucosidasa/g, 138 UE Celobiohidrolasa/g, y 10 UE Endoglucanasa/g, así como una reducción en el porcentaje de celulosa (4,6%) y hemicelulosa (4,4%), Adicionalmente se observó el cambio en la estructura del residuo y la colonización de los microorganismos sobre el mismo por medio de MEB. Palabras clave: Cultivo de arroz, tamo, celulasas, xilanasas, Penicillium sp. VII 7 Ab bstract The rice crop has an econo omic importtance for Co olombia duee to the hugge annual p production whicch is about 20x105 d dry tonnes of paddy rice. Howeever after harvest thee process geneerates an amount of w waste greateer than the product ob btained. In tthe presentt study we evaluated the ability of m microorganisms isolateed from soils to degraade the ricee straw in ordeer to reinstaatement thee waste to tthe culture soil. It was established d through qualitative, quantitative and indirect eevaluations tthat Penicilllium sp MF 3‐64 and Pa aecilomycess sp MF 4‐ 71 h have a prom mising cellulo olytic activitty unlike the 14 remain ning isolates tested. Ad dditionally by aan irregular factorial deesign using as a factorr the sourcee of carbon and nitroggen with 2 and 3 levels respectively, werre measurred enzym me activitties endogglucanase, cello obiohydrolaase and β‐‐glucosidasee. It was found thaat carboxymethylcellu ulose and amm monium sullfate have a positive influence in enzyme production. In the case of the xylanase was fo ound equally influenced by the nittrogen sourrce. It was fo ound that P Penicillium produces 40 000 UE/L of β‐glucosid dase in conttrast with P Paecilomycees sp. with 225 UE/L; sp. p this trend was ffound on the other enzzymatic activvities. In th he evaluatio on of Penicilllium sp. in solid fermeentation em mploying thee residue as substrate the activity was w 747 EU β‐glucossidase/g, 1 138 EU cellobiohydro olase/g, and 10 EU Endo oglucanase//g, it was found a reeduction in n the perceentage of cellulose c (4 4,6%) and hem micellulose ((4.4%), additionally by SEM it was observed changes in n the straw structure and the colonizzation of thee microorganism. words: Ricee crop, rice sstraw, cellulases, xylanases, Peniciillium sp. Keyw VII 8 Contenido Resumen ……………………………………………………………………………………………………. V Lista de figuras …………………………………………………………………………………………… X Lista de tablas …………………………………………………………………………………………….. XI Lista de abreviaturas ………………………………………………………………………………….. XII Introducción ………………………………………………………………………………………………. 13 1.Capítulo 1 Revisión teórica ……………………………………………………………………………. 15 1.1 El Cultivo del arroz ………………………………………………………………………………….. 15 1.2 Composición del residuo agrícola ………………………………………………………… 16 1.2.1 La celulosa ………………………………………………………………………………………….. 18 1.2.2 La Hemicelulosa …………………………………………………………………………………. 18 1.3 Enzimas celulolíticas ……………………………………………………………………………….. 20 1.4 Enzimas hemicelulolíticas ………………………………………………………………………. 23 1.5 Microorganismos productores de enzimas celuloliticas ………………………….. 24 1.6 Microorganismos productores de xilanasas …………………………………………. 24 1.7 Tratamiento de los residuos agrícolas y alternativas de uso de los 26 productos …………………………………………………………………………………………………… 1.7.1 Metodologías térmicas ………………………………………………………………………. 27 1.7.2 Metodologías químicas ……………………………………………………………………… 28 1.7.3 Tratamiento Biológico: Biotransformación de la biomasa 29 lignocelulósica ……………………………………………………………………………………………………… 1.8 El tamo de arroz como fuente renovable de carbono …………………………….. 32 2 .Capítulo 2. Planteamiento del problema ……………………………………………………. 36 2.1 Justificación ……………………………………………………………………………………… 36 2.2. Pregunta de investigación …………………………………………………………………. 38 2.3. Objetivos …………………………………………………………………………………………. 38 2.3.1 Objetivo General ………………………………………………………………………………… 38 2.3.2 Objetivos Específicos …………………………………………………………………………. 38 2.4 Metodología …………………………………………………………………………………….. 38 2.4.1 Selección de las cepas celulolíticas …………………………………………………… 39 2.4.2 Evaluación de la producción enzimática bajo diferentes fuentes de 38 carbono y nitrógeno …………………………………………………………………………………………….. 2.4.3 Evaluación de la degradación del sustrato y la producción enzimática 38 en fermentación solida ………………………………………………………………………………………… 3. Capítulo 3. Selección de los aislamientos con actividad celulolítica ……………… 41 3.1 Introducción ……………………………………………………………………………………… 41 3.2 Materiales y métodos ……………………………………………………………………….. 42 3.2.1 Microorganismos ………………………………………………………………………………. 42 3.2.2 Evaluación de la actividad cualitativa en agar celulosa …………………….. 42 3.2.3. Análisis de los resultados ………………………………………………………………….. 42 3.2.4 Evaluación de la producción de celulasas en el sustrato tamo de arroz 43 3.2.4.1 Cultivo del microorganismo……………………………………………………….. 43 VIII 9 3.2.4.2 Obtención del extracto ……………………………………………………………… 3.2.4.3 Actividad enzimática FPasas …………………………………………………….. 3.2.5 Evaluación de la producción de CO2 como índice de la actividad fúngica sobre el tamo de arroz …………………………………………………………………………….. 3.2.5.1 Obtención del inoculo ……………………………………………………………….. 3.2.5.2 Montaje del microcosmos ………………………………………………………….. 3.2.5.3 Determinación de CO2 ……………………………………………………………………………………………… 3.2.5.4 Análisis de los resultados …………………………………………………………… 3.3 Resultados y Discusión ……………………………………………………………………………. 3.3.1 Actividad celulolítica cualitativa …………………………………………………………. 3.3.2 Evaluación de la actividad celulolítica cuantitativa ………………………….. 3.3.4 Evaluación indirecta de la degradación del sustrato vegetal (tamo) 4.Capítulo 4. Evaluación de la producción enzimática bajo diferentes fuentes de carbono y nitrógeno ……………………………………………………………………………………. 4.1 Introducción ……………………………………………………………………………………… 4.2 Materiales y Métodos ……………………………………………………………………….. 4.2.1 Evaluación de la expresión enzimática en diferentes fuentes de carbono ……………………………………………………………………………………………………………… 4.2.2 Cultivo de los microorganismos para la evaluación de celulasas………. 4.2.3 Cultivo de los microorganismos para la evaluación de xilanasas………. 4.2.4 Obtención de los extractos ………………………………………………………………… 4.2.5 Actividades Enzimáticas …………………………………………………………………….. 4.2.5.1 Actividad endo‐1,4‐β‐glucanasa (EG)……………………………………….. 4.2.5.2 Actividad exo‐1,4‐β‐glucanasa (BCH)…………………………………………. 4.2.5.3 Actividad de 1‐4‐β‐glucosidasa (BGL) ………………………………………. 4.2.5.4 Actividad xilanasa ………………………………………………………………………. 4.2.6 Análisis de los resultados …………………………………………………………………… 4.3 Resultados y Discusión ……………………………………………………………………… 4.3.1 Perfil de la producción enzimática celulolítica ………………………………….. 4.3.2 Actividad Enzimática Endoglucanasa …………………………………………………. 4.3.3 Actividad enzimática Exoglucanasa (CBH) ………………………………………… 4.3.4 Actividad enzimática β‐glucosidasa …………………………………………………… 4.3.5 Perfil de la actividad enzimática xilanolítica ……………………………………… 4.3.6 Actividad Xilanasa ………………………………………………………………………………. 4.3.7 Influencia de la fuente de carbono y nitrógeno en la producción enzimática de Penicillium sp. y Paecilomyces sp …………………………………………………… 5. Capítulo 5. Evaluación de la degradación del tamo por el aislamiento seleccionado ……………………………………………………………………………………………… 5.1 Introducción ……………………………………………………………………………………… 5.2 Materiales y Métodos ……………………………………………………………………….. 5.2.1 Evaluación de la actividad enzimática “in vitro” de los microorganismos sobre el sustrato ………………………………………………………………………. 5.2.1.1 Obtención de los extractos enzimáticos …………………………………… 5.2.2 Determinación de lignina celulosa y hemicelulosa del residuo ………… 43 43 44 44 44 45 46 46 46 48 52 56 56 57 58 58 58 58 59 59 59 59 60 60 60 61 66 67 68 70 71 72 75 75 76 76 77 78 10 IX 5.2.3 Análisis cualitativo de la degradación ……………………………………………….. 5.3 Resultados y discusión ……………………………………………………………………….. 5.3.1 Actividad Enzimática ………………………………………………………………………….. 5.3.2 Determinación de Celulosa y Hemicelulosa ………………………………………. 5.3.3 Análisis cualitativo de la degradación del sustrato por MEB ……………. 6. Conclusiones y Recomendaciones ………………………………………………………………… 6.1 Conclusiones ……………………………………………………………………………………………… 6.2 Recomendaciones ……………………………………………………………………………………… 7. BIBLIOGRAFIA ………………………………………………………………………………………… A. Anexo: Composición de medios de cultivo …………………………………………. B. Anexo: Curvas de calibración para los métodos analíticos………………. C. Anexo: Análisis estadísticos ……………………………………………………………… D. Anexos : Determinación de Celulosa y Hemicelulosa ………………………….. 78 78 79 79 85 89 89 91 92 102 103 106 117 X 11 Lista de figuras Figura1.1 Composicion de la pared celular vegetal ……………………………………………………………………………………………………………………….. Figura 1. 2 Esquema propuesto de la acción de las enzimas celulolíticas sobre la celulosa ……………………………………………………………………………………………………………… Figura 1.3 Mecanismos de acción de las celulasas sobre el sustrato …………………. Figura 1.4 Procesos de conversión de biomasa más relevantes ………………………….. Figura 1.5 Efectos de la adición de materia orgánica obtenida a partir de residuos de cosecha en el suelo …………………………………………………………………………………………. Figura 2.1 Esquema de la metodología propuesta para la selección, caracterización y evaluación de degradación del residuo de los microorganismos celulíticos …………………………………………………………………………………………………………… Figura 3.1 Halos de hidrólisis en milímetros de los aislamientos evaluados ……… Figura 3.2 Determinación de la actividad enzimática FPasa ……………………………….. Figura 3.3 Determinación de CO2 en microcosmos en función del tiempo de las cepas MF 3‐64 Penicillium sp. MF 4‐71 Paecilomyces sp MF 9‐61Aspergillus sp Figura 3.4 Ensayo de microcosmos empleando el residuo como sustrato…………….. Figura 4.1 Cinéticas de actividad enzimática bajo diferentes fuentes de carbono y nitrógeno de Penicillium sp. …………………………………………………………………………………. Figura 4.2 Cinéticas de actividad enzimática bajo diferentes fuentes de carbono y nitrógeno de Paecilomyces sp. ……………………………………………………………………………. Figura 4.3 Área bajo la curva (ABC) de las unidades enzimáticas celulolíticas de Penicillium sp. y Paecilomyces sp. ………………………………………………………………………. Figura 4.4 Evaluación de la producción de xilanasas bajo diferentes fuentes de nitrógeno de Penicillium sp. y Paecilomyces sp. …………………………………………………. Figura 4.5 Área bajo la curva (ABC) de las unidades enzimáticas Xilanasas de Penicillium sp. (A) y Paecilomyces sp. (B) Figura 5.1 Actividad enzimática en la fermentación sólida. Endoglucanasa (EG), Xilanasa (XS), β –glucosidasa (BGS), Celobiohidrolasa (CBH Figura 5.2 Actividad enzimática en la fermentación sólida. Endoglucanasa (EG), Xilanasa (XS), y Azucares reductores producidos Figura 5.3 Microscopia electrónica de barrido (MEB) de las muestras al finalizar la fermentación solida 17 21 22 27 34 40 47 50 53 54 63 64 69 70 71 80 83 84 12 XI Lista de tablas Tabla 1.1 Composición de lignocelulosa de las clases de plantas ………………………. Tabla 1.2 Pretratamientos físicos empleados para la conversión de la biomasa .. Tabla 1.3 Pretratamientos químicos empleados para la conversión de la biomasa Tabla 1.4 Clases de Tratamientos biológicos ………………………………………………………. Tabla 1.5 Microorganismos empleados en la bioconversión de biomasa lignocelulósica ……………………………………………………………………………………………………… Tabla 1.6 Desechos lignocelulósicos y la producción anual mundial de los mismos Tabla 4.1 Diseño factorial irregular para evaluar la producción de celulasas ……… Tabla 5.1 Microorganismos empleados en la degradación de biomasa lignocelulósica …………………………………………………………………………………………………….. Tabla 5.2 Porcentaje de Celulosa y Hemicelulosa en el residuo antes y después de la fermentación sólida …………………………………………………………………………………………. 18 28 29 30 32 35 57 81 84 XII 13 Lista de abreviaturas Abreviatura Termino BGL Β –glucosidasa CBD Cellulose Binding Domain CBH Celobiohidrolasa CMC Carboximetilcelulosa EG Endoglucanasa Ha Hectáreas K Potasio MEB Microscopia Electrónica de Barrido Mg Magnesio Msnm Metros sobre el nivel del mar N Nitrógeno S Azufre Tm Toneladas métricas XS Xilanasas 14 Introducción El arroz se cultiva en más de cien países, en un rango de ambientes ecológicos, que van desde arroz secano a inundado, desde los 0 a 3000 msnm, en climas templados y tropicales en suelos de condiciones químicas y físicas muy variadas. Para obtener rendimientos satisfactorios es necesario un suministro adecuado de nutrientes. Las condiciones más apropiadas para el arroz se presentan en suelos con pH entre 5 y 7, con texturas medianas a pesadas y con contenido moderado de materia orgánica (FEDEARROZ 2000). En Colombia algunos suelos difícilmente proporcionan adecuadamente todos los nutrientes que requiere un cultivo como el del arroz, debido a esto se requiere recurrentemente realizar una fertilización química con elementos como N, P,K,S, Mg. En especial el cultivo del arroz requiere de grandes cantidades de fertilización ya que el sistema de riego, las actividades de labranza y las características de los suelos donde se cultiva no son propicios para el mantenimiento de los nutrientes en el suelo. (Castilla et al. 2012). Adicionalmente la demanda del cereal es alta por lo cual se requiere altos rendimientos por lo que la fertilización juega un papel importante en el proceso de siembra. Por otro lado la rápida expansión de los cultivos, la creciente demanda de alimentos, el agotamiento de combustibles fósiles y el rápido deterioro de los suelos ha llevado a la aparición de problemas ambientales, los cuales requieren de la aplicación de tecnologías que permitan utilizar de forma más eficaz los recursos. Sumado a esto dentro del proceso de cosecha del arroz se generan grandes cantidades de desechos, ya que por cada tonelada de producto se genera la misma cantidad de tamo o paja (Espinal 2005). Este residuo representa un problema para el agricultor ya que puede servir como propagador de plagas y así mismo puede aumentar los problemas fitosanitarios dentro del cultivo. Estos residuos representan un bajo valor económico y muchas veces son apilados, quemados o reincorporados al suelo sin una degradación previa, lo que puede 15 causar diferentes problemas en el suelo como por ejemplo la iniciación de procesos anaeróbicos y la propagación de plagas. Una alternativa para estos residuos podría ser la reincorporación al suelo después de haber sido degradados por microorganismos capaces de producir enzimas celulolíticas y hemicelulolíticas las cuales llevan a cabo la conversión de estos residuos a fuentes más fácilmente asimilables. El residuo del cultivo de arroz, es una fuente importante de nutrientes por su composición en variadas fuentes de carbono, sin embargo su reutilización se ve disminuida por el costo y trabajo que demandan los procesos tanto físicos como químicos que se deben realizar para poner a disposición sustratos más simples producto de su degradación Los microorganismos productores de enzimas celulolíticas ligninoliticas y hemiceluloliticas pueden ofrecer una alternativa a la degradación de estos residuos. Se han estudiado géneros como Trichoderma y Aspergillus tratando de entender el sistema enzimático celulolítico y en algunos casos hemicelulolítico (Sohail et al. 2009). Sin embargo estos microorganismos presentan dificultades en alcanzar una degradación eficiente ya que presentan bajas producciones de β – glucosidasa, enzima necesaria para completar la hidrólisis del polímero. En Colombia se ha estudiado el uso de Trichoderma ressei para la degradación de estos residuos, sin embargo es de interés la búsqueda de microorganismos que presenten la producción del sistema enzimático y que además presenten una eficiente degradación. En el presente estudio se plantea la biodegradación del residuo para su posterior incorporación al suelo como una alternativa sustentable que podría brindar una solución a los problemas que se tiene actualmente con el tamo de arroz debido a su excesiva producción y a al poco aprovechamiento que se le da a este tipo de residuos. Para esto se planteó la selección de los aislamientos con la mejor actividad celulolítica, y la evaluación de la producción de las enzimas que componen el complejo enzimático. Así mismo se evaluó el potencial de degradación del residuo para establecer los mejores aislamientos en producción enzimática, colonización y degradación del residuo. 16 Capítulo 1. Revisión teórica 1.1 El Cultivo del arroz El arroz (Oryza sativa L.) es un alimento básico para más de la mitad de la población mundial. Muchos lo consideran el cultivo más importante del mundo, principalmente si se toma en cuenta la extensión cultivada y la cantidad de gente que depende de su cosecha. Más del 40% de la población mundial depende del arroz para el 80% de su dieta y proporciona el 20% del consumo de calorías per cápita en todo el mundo (Benavides & Segura 2005). El cultivo del arroz tiene gran importancia económica. En América Latina y el Caribe se siembran cerca de 6.7 millones de hectáreas de arroz, siendo los principales productores: Brasil, Colombia, Ecuador, Perú y Venezuela (Pantoja 1997). En Colombia, el arroz ocupa el primer lugar en términos de valor económico entre los cultivos de ciclo corto y es el tercer producto agrícola en extensión, después del café y el maíz. Representa el 13% del área cosechada y el 30% de los cultivos transitorios (Espinal 2005) (Benavides & Segura 2005). En el proceso de producción del arroz, además del producto comercial conocido como arroz blanco, proveniente del arroz paddy (o con cascara), se generan varios tipos de desechos según la etapa de producción. Una vez efectuado el proceso de trilla (que consiste en retirar la cáscara al paddy), se obtienen los primeros dos subproductos: el arroz integral (o brown) y la cascarilla del arroz. Esta última se considera como desecho, aunque en ocasiones es usada como combustible para el proceso de secamiento, o en viveros y cultivos. Enseguida, el arroz integral (que también puede destinarse directamente al consumo humano), se pasa por un proceso de pulimento, a partir del cual se obtienen el arroz blanco y la harina de pulimento (o salvado de arroz). Esta última, se utiliza generalmente 17 como materia prima en la industria de alimentos balanceados para animales. Por otro lado durante el proceso de molinería por ejemplo, se obtienen granos que no cumplen con el estándar de calidad los cuales pueden ser vendidos como subproductos ( Ministerio de Agricultura y Desarrollo Rural Observatorio Agrocadenas Colombia 1996). En la etapa de cosecha se generan grandes cantidades de tamo o paja debido a que alrededor del 50% del peso seco de la planta corresponde a este residuo, por lo que por cada tonelada de grano cosechado se producen aproximadamente entre 1 y 1.35 toneladas de tamo que no posee un valor comercial (Ministerio de Agricultura y Desarrollo Rural 1996) (Singh et al. 2010). 1.2 Composición del residuo agrícola La pared celular de las células vegetales consiste en una mezcla compleja de polisacáridos y otros polímeros secretados por la célula y ensamblados en una organización entramada y equilibrada por medio de un conjunto de enlaces covalentes y no covalentes, está compuesta por una matriz amorfa que contiene pectina, proteínas, lignina y hemicelulosa dentro de la cual se encuentra embebida la fase microfibrilar cristalina compuesta por celulosa; a este polímero junto con la lignina y hemicelulosa se les conoce como lignocelulosa (Fig 1.1) (Linda 1997). 18 Figura 1 1.1 Composición de la pared celulaar vegetal. M Modificado (Park & Kim m 2012) La ligno ocelulosa esstá compuessta entoncees por tres ttipos de po olímeros, celulosa (C6H10O5)n, hemicellulosa (C5H8O4)m y lign nina [C9H100O3(OCH3)0.99‐1.7]n, fuerteemente unidos por enlaces covalentes y no covvalentes (Singh et al. 2 2010). Entree estos com mpuestos la celulosa occupa el primer lugar en concentració ón, seguidaa por la heemicelulosa y finalmen nte la lignin na. La porción de llas mismas varia de una especie de planta a o otra depend diendo compossición y prop sus caraacterísticas, es decir si son plantass productorras de grand des cantidades de polímeros de alta rrigidez o no o. (Tabla 1.1 1), (Sánchez 2009). En n el caso dell residuo deel cultivo de arroz, se han encontrad do valores de compo osición de Lignina (1 12.1%), cellulosa (39.1 1%) y hemicellulosa (23.6 6%) (Park & & Kim 2012 2), lo cual ubica al residuo en laa clasificación de pastos, que tiene ccomo caraccterística tener bajo co ontenido dee lignina y alto a conteniido de hemicellulosa y de ccelulosa. 19 Tablla 1.1 Comp posición de lignocelulossa de las trees clases de plantas. Tomado de (M Malherbe & Clloete 2002) 1.2..1 La celu ulosa ntes de carb bono en el p planeta, la La celulosa es cconsideradaa una de las más imporrtantes fuen ual por parrte de las plantas y aalgas marin nas se encu uentra alreededor de biossíntesis anu 8.5xx1010 tonelaadas, por este e motivo, la celulo osa es empleada en diferentes procesos indu ustriales paara la obteención de d diferentes tipos de productos p aaprovechab bles como azuccares y así m mismo para la producción de energgía (Niranjane et al. 200 07). Estee polisacárid do está locaalizado en la pared celular vegetaal en dondee se encuen ntra como unid dades micro oscópicas dee forma alargada conoccidas como micelas. A su vez, estaas micelas se arreglan en estructurass más grand des, las micrrofibrillas, las cuales esstán suficientemente emp paquetadas para prevenir la penettración no so olo de enzim mas sino dee pequeñas moléculas sem mejantes al aagua (Kamid de 2005), (A Andersson ett al. 2003). Las fibras celuló ósicas están n compuesttas por un p polímero lin neal, cuya unidad básicca es la D‐ gluccosa que se enlaza suceesivamente a través de un enlace gglucosídico en la configguración β (1‐4 4) para form mar moléculas de celobiosa. Cadaa unidad esstructural ((β‐ D‐ gluco opiranosa) oxilos libress (uno prim mario y dos secundario os). Las molléculas de conttiene tres ggrupos hidro 20 celulosa tienen una fuerte tendencia a formar enlaces por puentes de hidrogeno intra e intermoleculares. La existencia de estos enlaces tiene un efecto importante en la reactividad que presentan las cadenas celulósicas. Los enlaces de hidrógeno intermoleculares permiten una estructura fibrilar terciaria de alta cristalinidad, las zonas que presentan elevada cristalinidad son difíciles de penetrar, por el contrario las zonas relativamente más desordenadas (amorfas) son más accesibles y más susceptibles a todas las reacciones químicas (Singhania et al. 2013), (Kamide 2005) . Se ha estimado que la proporción de región cristalina es del 50 al 90% (Gan et al. 2003). Existen otras características de la molécula a parte de la cristalinidad que influyen en el proceso de degradación, como el área superficial accesible a las enzimas y las dimensiones de los poros de las fibras. Sin embargo dependiendo el origen, la celulosa puede variar en cuanto a la longitud del polímero y el grado de interacción entre las cadenas que lo conforman, así como el grado de polimerización alterando así las características anteriores (Gan et al. 2003). 1.2.2 La Hemicelulosa La hemicelulosa es la estructura que sirve de conexion entre la lignina y las fibras de celulosa proporcionando estabilidad al entramado lignocelulosico, tiene un peso molecular mas bajo que la celulosa y está formada por ramificaciones con cadenas laterales cortas que consistenen diferentes azucares, por lo que se denomina como un heteropolimero. Algunos polisacáridos representativos de la hemicelulosa encontrados en la pared celular son: xilanos, mananos, glucomananos, galactoglucomananos, xiloglucanos y calosa.(Hendriks & Zeeman 2009). El compuesto hemicelulolítico más abundante en las maderas duras (angiospermas) es el xilano, mientras que en las maderas blandas (gimnospermas) es el galactoglucomanano, aunque esto puede variar dependiendo de la especie de planta. En general el xilano es el componente mas abundante de la hemicelulosa. Es un complejo polisacarido compuesto 21 por unidades de xilosa unidas por enlaces β‐1,4, modificados por una gran variedad de sustituyentes en cadenas laterales como acetil, arabinosil, uronil, unidos principalmediante en el C2 y el C3 de las xilosas (Mo et al. 2010). Adicionalmente los componentes de la hemicelulosa forman complejos entramados con la lignina por medio de enlaces tipo ester con los acidos ferúlicos y cumarílicos lo que proporciona la estabilidad a la lignocelulosa. 1.3 Enzimas celulolíticas Para el aprovechamiento de los sustratos vegetales, algunos microorganismos producen enzimas capaces de degradar los componentes de la pared celular de las plantas, y a su vez esta habilidad es aprovechada en la industria para realizar varios tratamientos a fibras lignocelulósicas. Estas enzimas se conocen con el nombre de celulasas y son secretadas como moléculas libres por hongos filamentosos, actinomicetos y bacterias aeróbicas (Singhania et al. 2010). Las celulasas son enzimas que hidrolizan el enlace β‐1,4‐ glicosídico de la celulosa, y generan como productos primarios glucosa, celobiosa y celo‐oligosacáridos. Este complejo enzimático, comprende las Endoglucanasa (EG) (EC3.2.1.9), Celobiohidrolasa (CBH) (EC3.2.1.4), y las β‐glucosidasas (BGL) (EC3.2.1.21). En la Figura 1.2 se muestra el esquema general de acción de complejo, en donde la EG produce cortes en el polímero de celulosa principalmente en las regiones amorfas, exponiendo terminaciones reductoras y no reductoras, las CBH actúan por medio de estas terminaciones reductoras y no reductoras para liberar celo‐oligosacáridos y celobiosa, y las BGL hidrolizan la celobiosa para liberar glucosa completando la hidrolisis. El sistema de celulasas comprende a las tres enzimas, las cuales actúan sinérgicamente para convertir la celulosa en glucosa (Singhania et al. 2013). 22 Figura 1 1. 2 Esquem ma propuesto o de acción de las enzim mas celulolííticas, tomado de (Nideetzky B. et al 1993). Las enzzimas EG y CBG consttan usualmeente de do o y el os dominioss, el domin nio catalítico dominio o de unión a la celulossa (CBD), esste último se encuentra clasificad do en famillias en base a ssimilitudes een las secueencias de am minoácidos y proporcio ona las diferrencias en ccuanto al modo o de anclaje al polímero o. En el caso o del dominio catalítico o, la hidrólissis enzimáticca que se prod duce se lleva a cabo po or medio de un mecanismo de ccatálisis acid da. El mecanismo envuelvve la donacción de un protón al enlace β 1‐4‐ glicosíídico, por m medio del grupo carboxíllico del ácid do glutámico residual p presente en n el dominio o de la enziima, producciendo la rotura del enlacee y la estab bilización deel carbocatió ón intermed dio. La secu uencia finaliza por el ataqu ue nucleofíílico de unaa molécula de agua, obteniéndo o ose los prod ductos finales de hidrolisiis. En función del carb bocatión inttermedio fo ormado, la configuración del átom mo de carbono o en el prod ducto puedee ser retenid da, es decir que conserrva la misma estereoqu uímica o inverttida, dando el nombre a la clase dee mecanism mo empleado por la enzzima (retención o inversió ón) (Fig 1.3) (Carrillo Naavarrete 200 02). 23 Figu ura 1.3 Mecaanismos dee acción de las celulasas sobre el ssustrato a. M Mecanismo de Reteención b. Mecanismo d de Inversión (Ee 1990). Por otro lado sse sabe quee la síntesis de celulasaas está influ uenciada po or diferentes factores entrre los que sse encuentrra el microo organismo eempleado, las condicio ones del cultivo y las clasees de susttrato. La reelación enttre estas vvariables tieene un efeecto marcaado en la prod ducción del complejo enzimático o celulolíticco, el cual se encuentra complejamente regu ulado (Niran njane et al. 2 2007). Los catabolitoss solubles de bajo p peso moleccular que se liberan de los co ompuesto polim méricos porr la acción de enzima b basales pueeden entrar a la célula yy señalar la presencia del ssustrato en el medio, h haciendo qu ue se produ uzca la induccción de la síntesis aceelerada de las eenzimas celulolíticas. LLa síntesis d de estas enzzimas está ssujeta a ind ducción y/o represión cataabólica, com mpuestos co omo celulosa, y sus derrivados, celo obiosa, xilano, pectina, soforosa y lacctosa puedeen actuar co omo inducto ores. Así mismo la sínttesis de las enzimas puede verse reprrimida por la presencia de glucosaa y otros azu ucares fácilm mente asimilables. El p papel de la 24 celobiosa, soforosa, glucosa y otros azucares como inductores o represores varia con cada microorganismo, ha sido difícil encontrar mecanismos universales de inducción o represión pero en general es evidente que los mejores sustratos para la producción de estas enzimas son los celulósicos insolubles (Lockington et al. 2002), (Kaur et al. 2006), (Saibi et al. 2007). 1.4 Enzimas hemicelulolíticas La hemicelulosa es la segunda fracción más abundante en la naturaleza después de la celulosa y comparada con esta, posee una heterogeneidad en su composición, haciendo que su proceso de degradación sea más complejo. Las enzimas hemicelulolíticas necesarias para tal degradación se clasifican de acuerdo a su acción sobre diferentes sustratos produciendo azúcares manoméricos y ácido acético como principales productos (Sunna & Antranikian 1997) El xilano es el principal carbohidrato encontrado en la hemicelulosa. La biodegradación de xilano requiere de complejos enzimáticos del tipo esterasas y glucanasas, estas enzimas son producidas por bacterias y hongos y son en su mayoría extracelulares (Sunna & Antranikian 1997). Dentro de estos complejos enzimáticos se encuentran las enzimas endoxilanasas (E.C. 3.2.1.8); las cuales actúa al azar sobre la cadena principal del xilano, produciendo xilooligosacaridos y la xilosidasa (E.C. 3.2.1.37); que libera xilosa de oligosacáridos cortos. Adicionalmente es necesaria la acción de enzimas accesorias como la α‐4‐O‐metil glucoronidasa, la arabinofuranosidasa (E.C. 3.2.1.55); que remueve 1‐arabinofuranosa, la D‐ Glucuronidasa (E.C. 3.2.1.139); que hidroliza residuos de ácido glucorónico metilado, la acetil xilano esterasa (E.C. 3.1.1.72); que hidroliza grupos acetato de la cadena principal, y la feruloil (E.C. 3.1.1.73) y cumaril esterasa (E.C. 3.1.1.‐) las cuales hidrolizan los ácidos aromáticos unidos a los residuos de arabinofuronidasa (Chávez et al. 2006). La mayor parte de la caracterización de las enzimas hemiceluloliticas se lleva a cabo sobre las endoxilanasas, debido a la dificultad que existe para purificar las otras enzimas. Se ha encontrado que los complejos enzimáticos se producen en una amplia variedad con 25 diferente especificidad. Estudios demuestran que algunos tipos de endoxilanasas producen principalmente xilosa y xilobiosa, mientras que otras producen oligosacáridos con altos grados de polimerización. (Elegir G. et al 1994), (Chávez et al. 2006). 1.5 Microorganismos productores de enzimas celuloliticas Existe una gran variedad de microorganismos envueltos en la producción de celulasas incluyendo bacterias aeróbicas y anaeróbicas, hongos de podredumbre blanca y microorganismos anaerobios (Singhania et al. 2010). Las celulasas de muchas bacterias celulolíticas se organizan en complejos multienzimáticos discretos, llamados celulosomas, las múltiples subunidades de celulosomas se componen de numerosos dominios funcionales, que interactúan entre sí y con el sustrato celulósico; estas estructuras han sido identificadas y caracterizadas en bacterias anaerobias como Clostridium thermocellum (Bayera et al. 1994). En hongos aún no se ha reportado una estructura similar a la de las bacterias anaerobias, sin embargo se han informado de la existencia de otras proteínas accesorias, que son parte de los complejos enzimáticos y pueden afectar la degradación de materiales celulósicos, pero no muestran comportamiento hidrolítico. Estas proteínas son producidas por hongos y plantas y se conocen como expansinas. Estas enzimas actúan de forma tal que pueden generar una hinchazón en la red de celulosa, perturbando la estructura del sustrato, y así aumentando la accesibilidad de las celulasas (Luciano et al. 2012). Actualmente se sabe que los más eficientes degradadores de celulosa son los hongos filamentosos, los cuales presentan un sistema enzimático completo de celulasas capaces de degradar parcial o totalmente la celulosa en celobiosa y glucosa. La capacidad de degradación eficiente de los hongos radica en el crecimiento micelial que permite que el hongo transporte nutrientes que pueden resultar limitantes como nitrógeno y hierro a lugares distantes (Niranjane et al. 2007), (Singh et al. 2010). Dentro de los hongos que presentan más eficiencia en la degradación de compuestos celulósicos se encuentran: 26 Trichoderma viride, Trichoderma reesei, Penicillium pinophilum, Sporotrichum pulverulentum, Fusarium solani, Talaromyces emersonii Trichoderma koningii, y Phlebia gigantea (Latifian et al. 2007), (Niranjane et al. 2007)(Jørgensen et al. 2004)(Juhász et al. 2005). Otros microorganismos productores de celulasas incluyen hongos aeróbicos termofílicos (Sporotrichum thermophile, Thermoascus aurantiacus y Humicola insolens), hongos anaeróbicos mesofílicos (Neocallimastix frontales, Piromonas communis, Sphaeromonas communis), bacterias aeróbicas mesofílicas y termofílicas (Bacillus sp., Cellulomonas sp., Cellvibrio sp., Microbispora bispora y Thermomonospora sp.), bacterias anaeróbicas mesofílicas y termofílicas (Acetivibrio cellulolyticus, Bacteroides succinogenes, Ruminococcus albus, Ruminococcus flavefaciens y Clostridium thermocellum) y actinomicetos (Streptomyces sp., Thermomonospora sp. y Thermobifida sp.) (Singh et al. 2010) (Niranjane et al. 2007), (López & Freitas 2012a). 1.6 Microorganismos productores de xilanasas El xilano es el mayor componente de la hemicelulosa, en la naturaleza la degradación del xilano es llevada a cabo por diferentes microorganismos como bacterias, levaduras y hongos (Abdel‐Sater & El‐Said 2001). Esta capacidad está ampliamente distribuida entre los microorganismos saprófitos, así como entre la microbiota del rumen. Estos microorganismos poseen sistemas enzimáticos complejos para llevar a cabo la degradación del xilano. Dentro de los microorganismos productores de xilanasas se encuentran bacterias como Streptomyces sp y Bacillus subtilis. y hongos como Gloeophyllum trabeum Penicillium oxalicum y Aspergillus sp (Li et al. 2010) (Sa et al. 2002), (Li et al. 2007), (Chapla et al. 2010). Similar a lo encontrado con bacterias celulolíticas, se han reportado estructuras llamadas xilanosomas, similares a los celulosomas las cuales pueden jugar un importante papel en la degradación de la hemicelulosa. Esta clase de estructuras se han encontrado en bacterias como Butyvibrio fibrisolvens y Clostridium papyrosolvens (Jiang et al. 2005). 27 Los hongos filamentosos son particularmente importantes para la industria por la producción de enzimas xilanolíticas ya que son capaces de secretar a medio grandes cantidades a diferencia de bacterias y levaduras (Li et al. 2006). Así mismo han demostrado producir un amplio rango de enzimas xilanolíticas, siendo Aspergillus sp y Trcichoderma sp. los géneros más ampliamente estudiados (Abdel‐Sater & El‐Said 2001). Al realizar aislamientos de microorganismos degradadores de xilano provenientes de residuos agrícolas se han encontrado especies como: Aspergillus avus A. niger A. ochraceus A. terreus A. ustus, Penicillium chrysogenum P. corylophilum P. duclauxii P. funiculosum P. oxalicum y Trichoderma harzianum y viride, asi mismo se encontraron microorganismos como Cladosporium herbaru, Emericella nidulans Epicoccum nigrum Fusarium oxysporum Paecilomyces variotii (Abdel‐Sater & El‐Said 2001). 1.7 Tratamiento de los residuos agrícolas y alternativas de uso de los productos Dependiendo del origen y sus características la biomasa lignocelulósica puede emplearse para diversos fines, entre ellos se encuentra la alimentación animal, generación de energía, la generación de otros productos como por ejemplo papel y la utilización como abono orgánico dentro del mismo cultivo (Biomass Users Network (BUN‐CA) 2002). Dentro de este campo, la mayoría de desarrollo se ha centrado en la generación de energía, para este fin los residuos deben ser transformados en diversos subproductos que darán lugar a diversas formas de energía como la electricidad, el vapor, y la producción de combustibles líquidos empleados para tal fin. Otro tipo de alternativas contemplan el uso de estos residuos para la producción de compuestos de interés como azucares y polímeros utilizados ampliamente en diversas industrias (Fig. 1.4) (Sánchez 2009). Sin embargo es necesario el empleo de diferentes metodologías para el tratamiento de estos compuestos, estas se clasifican en térmicas o dependientes de altas temperaturas, químicas y biológicas, centrándose la atención en esta última por la amplia variedad de 28 sistemas enzimáticos produccidos por diferentes clases de microorgan nismos com mo se mencion nó anteriorm mente. Figura 1 1.4 Procesoss de conversión de biom masa más reelevantes. TTomado de (Sánchez 20 009) 1.7.1 M Metodologgías térmicas Durantee este pretrratamiento la biomasa lignocelulósica es caleentada, en temperaturaas que oscilan entre 90 aa 180 °C d dependiend do el proceeso (Tabla 1.2), en eeste processo los nentes se so olubilizan co omenzando o por la hem micelulosa yy continuando con la lignina. compon Durantee el proceso o térmico u una parte dee la hemiceelulosa es h hidrolizada formando f á ácidos, los cualees se asumee que particcipan en la d degradación n total del compuesto. Con respecto a la solubilizzación de la lignina sse pueden producir co ompuestos en su mayyoría fenólicos y heteroccíclicos, esto os compuesstos reaccio onan fácilm mente y son n de difícil remoción, puede 29 prod ducirse así mismo furffural entre otros comp puestos. Po or otro lado o altas tem mperaturas den incluso causar un iincremento en el índicee de cristalin pued nidad. Es im mportante mencionar que con essta clase dee metodologgías, es neccesaria iguaalmente la inveersión de en nergía en su u conversión a producttos utilizables, lo cual conlleva a aumentar los ccostos de lo os procesos (Hendriks & & Zeeman 20 009). Tablla 1.2 Algun nos Pretrataamientos físicos empleaados para laa conversión n de la biom masa. Tom mado de (Taherzadeh & & Karimi 200 08) 1.7..2 Metodo ologías qu uímicas El p pretratamien nto químico o se lleva acabo emp pleando differentes claases de com mpuestos, entrre estos los más usados son los áccidos y las b bases. El preetratamiento químico ccon ácidos es rrealizado p para increm mentar la d digestibilidaad anaerob bia del resiiduo, el ob bjetivo es solu ubilizar la hemicelulosaa, y por meedio de estto hacer la celulosa m más accesible para la degrradación. LLa principaal reacción que ocurrre en estee caso es la hidrolisis de la hem micelulosa, especialmente el xilan no debido a que el glucomanano es relattivamente estaable a pH accido. Para ttal fin se em mplean com mpuestos co omo el ácido o nítrico en ntre otros. 30 Los oliggómeros solubilizados pueden esstar sujetos a reaccion nes de hidrrolisis que puede igualmeente produccir monómeros, furfural, y otros co ompuestos volátiles. Durantee el pretrattamiento alcalino la prrimera reaccción que tiiene lugar es e la solvattación. Esto pro ovoca un esstado hinchado de la b biomasa y laa hace más accesible p para las enziimas y bacteriaas (Hendrikss & Zeeman n 2009). Estee tipo de prre tratamien nto incluye, la utilizació ón de , hidróxid do, de sodio o, hidróxido de calcio en ntre otros (TTabla 1.3) ((Park & Kim m 2012). Tabla 1..3 Algunos p pretratamieentos químiccos empleados en la biomasa. Tom mado de (Taherzaadeh & Kariimi 2008) 1.7.3 TTratamiento Biológiico: Biotraansformacción de la biomasa llignoceluló ósica d enzimáticaa de los miccroorganism mos es En el caaso de la deegradación biológica, laa capacidad clave en e la degrradación yy bioconversión, perm mitiendo obtener o differente tip po de subprod ductos, y así mismo se s evita el uso de compuestos q químicos o energía paara su degradaación. 31 Paraa llevar a cabo la biodeegradación d de la biomasa lignocelu ulósica es im mportante m mencionar que esta se lleva a cabo p por un prrocesos con n múltiples pasos los ccuales incluyyen: 1. La adso orción de la celulasa en n el sustrato o por interm medio del do ominio de u unión de la eenzima; 2. Locaalización del enlace sussceptible dee hidrolisis een la superfiicie del susttrato; 3. La fformación del complejo enzima‐sustrrato; 4. Hidrólisis de lo os enlaces β β‐glicosídico o mientras la enzima reco orre la cadena del polím mero; 5. La desorción d de las celulasas del susstrato (Yeom man et al. 2010 0). Sin em mbargo es importantee mencionaar que po or lo geneeral es neccesaria la dism minución deel tamaño de partícula del sustrrato para así a hacerlo más accesible a los micrroorganismo os a empleear en el proceso, p en n la Tabla 1.4 se men ncionan loss tipos de trataamientos biiológicos em mpleados. En ssi el objetivo o de los trattamiento co onvencionalles es reduccir la cristalinidad de laa celulosa, increementando o la rupturaa de las moléculas m d de hemicelu ulosa, aumentando el área de conttacto del miicroorganism mo con el sustrato y dee esta maneera poder accceder a la eestructura recaalcitrante dee la celulossa produciendo la máxxima obtención de azu ucares y co ompuestos fácillmente mettabolizabless (Park & Kim m 2012). En el caso deel tratamien nto biológicco además de lograr la degradación d del residuo,, se está dissponiendo aadecuadam mente de loss residuos oblemas de contaminacción adicion nales por el uso de com mpuestos qu uímicos, o sin ggenerar pro el gaasto de energía por la d degradación n como suceede con los procesos téérmicos. mientos bioló ógicos. Tom mado de (Taaherzadeh & & Karimi 200 08). Tablla 1.4 Clases de Tratam 32 Se han propuesto variados usos para los productos de la biodegradación de estos residuos, dentro de los cuales podemos encontrar la utilización como materia prima para la producción de etanol, para la manufactura de papel, como compost para el cultivo y directamente para alimentación animal (Singh & Ashok 2009). En la Tabla 1.5 se encuentran diferentes ejemplos de microorganismos empleados para la degradación de diferentes clases de materiales lignocelulósicos y los productos que se han evaluado de estas degradaciones. Muchas de las investigaciones se han llevado acabo con el propósito de encontrar un combustible alternativo, empleando métodos biológicos, debido al efecto positivo de los biocombustibles en el ambiente. El etanol por ejemplo es usado como materia prima o como aditivo para la gasolina, en Brasil y EEUU por ejemplo se produce etanol de la fermentación de azucares obtenidos de material lignocelulósico degradado biológicamente como la caña de azúcar. Así mismo desde el 2008 se han incrementado los estudios con respecto a la utilización de residuo del cultivo de arroz para la producción de etanol, países como Japón, China, India y EEUU lideran las investigaciones en este campo. Por otro lado es posible igualmente obtener diferentes tipos de compuestos como xantanos, ácidos orgánicos, aminoácidos y vitaminas empleando los azucares obtenidos de esta transformación como sustrato para la fermentación (Li et al. 2007). Sin embargo es importante aclarar que el uso del material lignocelulósico biodegradado no siempre tiene que estar condicionado a la obtención de energía o de biocombustibles, existen otros campos en donde estos residuos degradados pueden emplearse de manera eficiente, como es el caso de las reincorporaciones en los suelos de los cultivos como abono orgánico. 33 Tablla 1.5 Algun nos microorganismos empleados een la degrad dación de bio omasa ligno ocelulósica. 1.8 8 El tamo o de arrozz como fu uente renovable de carbo ono Deb bido a que el residuo o del cultivvo de arroz está comp puesto por lignina ccelulosa y hem micelulosa, eentre otros nutrientes,, el tamo seería una fueente renovaable de carrbono que podría emplearse en el aaprovecham miento denttro del missmo cultivo o. Es de im mportancia enco ontrar alterrnativas de aporte de n nutrientes d debido a qu ue no siemp pre que se ffertiliza se estáá nutriendo la planta ya y que exissten diverso os factores que afectaan la eficien ncia de la fertiilización. Deentro de esttos factoress que afectaan la dispon nibilidad dee los nutrien ntes están 34 los de orden físico, como la compactación lo cual limita el movimiento del agua en el suelo, la erosión, la cual disminuye la materia orgánica afectando los procesos bioquímicos de disponibilidad de nutrientes debido a una disminución de microorganismos en el suelo, y del orden químico (Castilla 2013). Dos factores determinan en gran parte la productividad de la tierra en que se siembra el arroz: la condiciones del suelo y el agua disponible. Con respecto a la primera, el arroz se cultiva en suelos cuyas propiedades químicas y físicas difieren mucho; por consiguiente, tanto las variedades como el manejo de los suelos han debido adaptarse a esas condiciones edáficas (Degiovanni et al. 2010), (Marentes et al. 2012). En estos suelos fértiles o de baja disponibilidad de nutrientes, se pueden obtener rendimientos aplicando los elementos nutricionales deficientes o corrigiendo las condiciones que limitan la disponibilidad de los nutrientes (Degiovanni et al. 2010). Dentro de este contexto la incorporación de residuo degradado biológicamente al suelo en el cultivo del arroz puede contribuir favorablemente en las propiedades químicas, físicas y biológicas del suelo, ayudando en la regulación del pH , a la aireación y absorción de la humedad gracias a la porosidad del residuo (Figura 1.5) (Degiovanni et al. 2010)Se ha encontrado que la incorporación puede incrementar incluso en 1 tonelada la producción, haciendo de esta metodología una alternativa viable tanto para la disposición adecuada de los residuos como para el mantenimiento de la calidad del suelo donde se lleva a cabo el cultivo (Espinal 2005). Así mismo el residuo puede servir de medio para el desarrollo de microorganismos que pueden crecer sobre los productos de transformación de los sustratos. Estos microorganismos pueden terminar la función de degradar el sustrato y continuar con otra función biológica de importancia dentro del ciclaje de otros nutrientes (Degiovanni et al. 2010). 35 Figu ura 1.5 Efecctos de la adición de materia orgánica o obtenida a partir de residuos de coseecha en el suelo. Modifficado (Degiovanni et al. 2010), (Esspinal 2005)). En eel caso de otros residuos del cultivo de arrroz como laa cascarilla, la utilizacción como abono presenta un bajo ccosto, calcu ulándose qu ue se requiere en pro omedio 8 K Kg/m para com mplementar las necesid dades nutriccionales del cultivo (Qu uiceno & Mosquera 20 010). En el caso o del tamo d de arroz, el lograr la in ncorporación n del residu uo sería de ggran utilidad para así poder disponer de la graan cantidad d de residuo generado o, ya que m mientras po or cada 8 n de arroz see generan 1 de cascarillla, en el caso de tamo lla relación toneeladas que sse obtienen es 1 1:1.5 respectivamente. A nivel mundial igualm mente se ob bserva que el residuo d del cultivo se p produce en n grandes cantidades,, anualmen nte se regiistran hastaa 731 millones de toneeladas a niivel mundiaal, valor ostensiblemeente alto ccomparado con otross residuos agríccolas (Tablaa 1.6). 36 Tabla 1..6 Algunos d de los desecchos lignoceelulósicos y la producción anual mundial de lo os mismoss tomado dee (López & FFreitas 2012 2b) 37 Capítulo 2. Planteamiento del problema 2.1 Justificación El arroz es el cereal más importante que se produce en el país, es el soporte agrícola de nueve departamentos y al año se cultivan cerca de 490.000 hectáreas, lo cual permite registrar ganancias importantes que sitúan a Colombia como el segundo productor más importante en Latinoamérica (Ministerio de Agricultura y Desarrollo Rural Observatorio Agrocadenas Colombia 1996) En el año 2003, el arroz en Colombia se cultivó en 498.486 hectáreas (Ha.) que rindieron 2.543.161 toneladas métricas (Tm.) de paddy y aproximadamente 1.729.349 Tm. de arroz blanco (Ministerio de Agricultura y Desarrollo Rural 1996). Sin embargo el cultivo del arroz como actividad agroindustrial ha llevado a la acumulación de una gran cantidad de residuos lignocelulósicos, los cuales se encuentran por el orden de 157 a 188 x 106 toneladas/año (Castilla et al. 2012). Existen varias formas de disposición de estos residuos, como las quemas agrícolas a cielo abierto con algunas restricciones para proteger la calidad del aire y el suelo (Espinal 2005), así como procesos físicos y químicos de degradación para ser utilizados en diferentes industrias, pero en ambos casos los costos y el transporte para ser degradarlos dificultan la utilización de estos métodos. Por otro lado el costo de los fertilizantes utilizados en el cultivo del arroz ha aumentado por el incremento de los precios internacionales (especialmente de los nitrogenados) y también por el efecto de la devaluación. Adicionalmente, su consumo también se ha incrementado, en especial en las zonas donde se emplea el sistema de riego, con el fin de lograr el máximo de productividad de las variedades que se están sembrando (Ministerio de Agricultura y Desarrollo Rural 1996). 38 Dentro de este contexto el uso de los residuos como fertilizantes orgánicos sería una mejor alternativa, ya que la incorporación al suelo mejora las condiciones físico químicas del mismo, incrementando la infiltración y retención del agua del suelo, protegiéndolo de la erosión y así mismo incrementando la materia orgánica disponible que puede servir como nutriente para la planta, así como para los microorganismos de la rizósfera que proporcionan sustento al crecimiento del cultivo y paralelamente representaría una alternativa sustentable del manejo de los residuos agrícolas (Pantoja Alberto 1997) (Fan et al. 2005). Sin embargo, para este fin es necesaria la acción de microorganismos que lleven a cabo la degradación del residuo, se ha centrado el interés en las enzimas celulasas y xilanasas las cuales son capaces de degradar celulosa y hemicelulosa, componentes principales del residuo del cultivo de arroz. Existen pocos reportes sobre el uso de esta técnica para la degradación de residuos lignocelulósicos “in situ”. En Colombia, FEDEARROZ ha evaluado el uso del hongo Trichoderma viride, (Marentes et al. 2012), sin embargo es necesario una investigación más a fondo que permita la búsqueda de otros microorganismos que puedan aportar eficiencia y productividad en la degradación. De este modo se torna de gran importancia no solo aislar microorganismos promisorios en la degradación de estos sustratos, sino también realizar una descripción del proceso que llevan a cabo los mismos con el fin de establecer el tipo de enzimas que se producen y las características de los complejos enzimáticos. 39 2.2. Pregunta de investigación Los microorganismos aislados del suelo de cultivo de arroz son capaces de producir enzimas celulolíticas y xilanolíticas capaces de llevar a cabo la degradación eficiente del residuo agrícola conocido como tamo o paja. 2.3. Objetivos 2.3.1 Objetivo General: Evaluar la degradación del tamo de arroz empleando microorganismos celulolíticos aislados de suelos de cultivos de arroz 2.3.2 Objetivos Específicos: 2.3.2.1 Seleccionar las cepas con mayor actividad enzimática celulolítica mediante evaluaciones cualitativas y cuantitativas. 2.3.2.2 Evaluar la producción de glucanasas, celobiohidrolasas, 1‐4‐β‐glucosidasas y xilanasas de los microorganismos seleccionados sobre diferentes fuentes de carbono y nitrógeno 2.3.2.3 Evaluar la degradación ejercida por los aislamientos sobre el residuo natural 2.4 Metodología General De acuerdo al problema planteado y para dar respuesta a los objetivos propuestos se llevó a cabo la metodología que se muestra en la Figura 2.1 y que se describe brevemente a continuación. 40 2.4.1 Selección de las cepas celulolíticas La metodología planteada en el capítulo 3 describe la selección de las cepas celulolíticas a partir de un banco de trabajo aislado de suelos de cultivo de arroz. Para ello se llevaron a cabo tres metodologías sucesivas. La evaluación cualitativa en la cual se midieron los halos de hidrólisis de las 16 cepas en agar Carboximetilcelulosa, la evaluación cuantitativa, determinando las unidades celulolíticas totales en fermentación sumergida con el residuo como sustrato y finalmente una evaluación indirecta de degradación del sustrato por medio de la técnica de respirometría obteniendo los miligramos de CO2 como producto de la degradación. 2.4.2 Evaluación de la producción enzimática bajo diferentes fuentes de carbono y nitrógeno En el capítulo 4 se plantea la evaluación de la influencia de la fuente de carbono y nitrógeno en la producción del complejo enzimático celulolítico. Para ello se realizó un diseño factorial irregular evaluándose dos fuentes de carbono y dos fuentes de nitrógeno y la ausencia de la misma. Las variables de respuesta fueron las unidades enzimáticas: exo‐1,4 β‐d‐glucanasa, endo‐1,4‐ β‐d‐glucanasa y β‐glucosidasa. Para evaluar la influencia de la fuente de nitrógeno en la producción de las xilanasas se evaluó una fuente de nitrógeno orgánica, una inorgánica y la combinación de las dos fuentes. La variable de respuesta fue la unidad enzimática xilanolítica. 2.4.3 Evaluación de la degradación del sustrato y la producción enzimática en fermentación solida Finalmente en el capítulo 5 se describe la metodología para evaluar la degradación del sustrato por el microorganismo. Se realizó una fermentación solida por 30 días evaluando la producción enzimática, igualmente se evaluó el porcentaje de lignina, celulosa y hemicelulosa presente en el sustrato antes y después del tratamiento biológico y se llevó a 41 cabo o un análisiis MEB paraa determinar cualitativvamente la colonizació ón y degrad dación del susttrato por el microorgan nismo. Figu ura 2.1 Esqu uema de la m metodologíaa propuestaa para la seleección, caraacterización y evaluación de d degradación n del residuo o de los miccroorganism mos celulítico os 42 Capítulo 3. Selección de los aislamientos con actividad celulolítica 3.1 Introducción En un cultivo de alto interés económico como lo es el de arroz es de importancia diferentes aspectos como la calidad de la semilla, el aporte de nutrientes a la planta, el sistema de riego entre otras. Dentro de estas características la calidad del suelo es de interés para los agricultores debido a que la capacidad funcional del mismo permite sostener la productividad del cultivo. En el correcto funcionamiento del ecosistema del suelo es de importancia la presencia de los diferentes grupos funcionales debido a esto la diversidad microbiana es de interés por la amplia variedad de sistemas enzimáticos que producen. Por tal motivo es relevante determinar las características de los suelos rizosféricos del cultivo para así conocer el estado de las comunidades en el suelo. El aislamiento y caracterización de los microorganismos es de interés no solo para conocer la diversidad, sino también para establecer la participación que tienen estos en los procesos de descomposición y biotransformación de los compuestos orgánicos Por lo anterior se realizó la selección de los aislamientos que podrían ser promisorias en la producción de enzimas celulolíticas, ya que de esta forma se podrían llevar a cabo procesos de degradación de los residuos del cultivo con microorganismos que presentan una mejor adaptación a las condiciones in situ. En el presente estudio se evaluaron 16 cepas fúngicas para determinar cuáles presentan la capacidad de hidrolizar la celulosa, para tal fin se llevaron a cabo tres metodologías sucesivas dando como resultado la selección de los aislamientos con actividad celulolítica promisoria. 43 3.2 Materiales y métodos 3.2.1 Microorganismos Se evaluaron aislamientos fúngicos de suelo de cultivo de arroz de fincas de los departamentos de Tolima y Meta (Gutierrez et al. 2012). Estos se encuentran conservados a 4°C en el banco de microorganismos del Laboratorio de Biotecnología Aplicada de la Pontificia Universidad Javeriana. 3.2.2 Evaluación de la actividad cualitativa en agar celulosa Como primera evaluación de las cepas se realizó una metodología cualitativa en agar celulosa. Para inducir la producción de celulasas se empleó el agar Celulosa 1% (En gL‐1 extracto de levadura 2,5, celulosa alimentaria 10.0; triptosa 2,5; sulfato de amonio 0,5; cloruro de calcio, 0,5; fosfato monobásico de potasio, 0,1; fosfato di básico de potasio, 0,1). Por medio de punción se sembró el microorganismo, las placas se incubaron por 8 días a 28°C y después de este tiempo se adicionó rojo congo al 1% (p/v), se llevaron a refrigeración por 12 hrs y posteriormente se adicionó NaCl al 0,85% evidenciándose la producción de halos de hidrólisis. (Liu, Zhang, Yang, Xu, et al. 2011). Se realizó la medición de los halos empleando la siguiente formula (ver ecuación 1.1) : ó – 10 (1.1) 3.2.3. Análisis de los resultados Los resultados de los halos de hidrólisis de las 16 cepas se analizaron con el software de estadística IBM SPSS versión 19 (Armonk, NY, USA) y Statistix. Las pruebas de normalidad se realizaron con el test de Shapiro‐Wilk y la homogeneidad de varianzas con el test de Leven. Se aplicó un análisis Kruskal Wallis . El nivel de confianza fue del 95%. 44 3.2.4 Evaluación de la producción de celulasas en el sustrato tamo de arroz 3.2.4.1 Cultivo del microorganismo Para cuantificar la producción de enzimas celulolíticas se llevó a cabo una fermentación sumergida en matraces de 250 ml con un volumen efectivo de 120 ml utilizando caldo tamo de arroz (En gL‐1 Extracto de levadura, 2,5; triptosa 2,5; sulfato de amonio 0,5; cloruro de calcio, 0,5; fosfato monobásico de potasio, 0,1; fosfato di básico de potasio, 0,1; cloranfenicol, 0,1; tamo de arroz 10.0) El inoculo consistió en 12 ml de una suspensión de esporas con una concentración de 106 esporas.mL‐1 de cada aislamiento. Después de inoculados los matraces se incubaron a 28°C, en una incubadora orbital a 120 rpm. El ensayo se llevó a cabo por triplicado y con un control abiótico que consistió en la utilización del mismo medio de cultivo sin inoculación del microorganismo. 3.2.4.2 Obtención del extracto Se realizaron muestreos a los 4 y 6 días debido a que en trabajos previos llevados a cabo con algunos aislamientos del presente estudio se encontró la máxima producción enzimática en este lapso de tiempo (Perez & Hernandez 2011). Se tomaron 10mL de medio de cada uno de los matraces inoculados y del control abiótico. La muestra se centrifugó a 10000 rpm durante 20 minutos a 4ºC y posteriormente a 5000rpm durante 20 minutos, esto para separar la biomasa y los componentes del medio de cultivo del extracto a partir del sobrenadante obtenido se realizaron las pruebas de actividad enzimática. 3.2.4.3 Actividad enzimática celulasas totales (FPasas) Para la evaluación cuantitativa de la actividad celulolítica se tomaron 2mL de cada extracto obtenido y 2mL de buffer citrato de sodio pH 5.0 y 100 mg de papel de filtro BOECO. El blanco de la reacción consistió en la mezcla de 2mL de cada extracto y 2mL de buffer citrato sin sustrato. Las mezclas se incubaron a 37ºC durante 60 minutos (Ghose 1987). Transcurrido este tiempo se llevó a cabo la determinación de azucares reductores en la muestra por medio de la técnica de DNS (Miller 1959). 45 Una unidad celulolítica se definió como la cantidad de enzima que libera 1μmol de azúcares reductores por minuto por litro (μmol/min.L). 3.2.5 Evaluación de la producción de CO2 como índice de la actividad fúngica sobre el tamo de arroz Posteriormente a la evaluación enzimática se llevó a cabo un ensayo in vitro por el método de respirometría con las cepas que presentaron actividad enzimática sobresaliente, lo anterior para obtener información de la actividad degradativa fúngica sobre el sustrato por medio de la liberación de CO2. 3.2.5.1 Obtención del inoculo Para la obtención del inoculo se realizó un cultivo en matraces de 250 ml con 120 ml de caldo Extracto de Malta (En gL‐1 extracto de Malta 20,0, peptona 1,0, glucosa 10,0) en el cual se inocularon discos de agar del microorganismo, el cultivo se incubó a 28°C a 120 rpm y al cabo de 8 días se recuperó la biomasa por centrifugaciones sucesivas a 8.000 rpm a 4°C con agua destilada estéril. La biomasa recuperada se secó en papel filtro. 3.2.5.2 Montaje del microcosmos Se emplearan frascos de vidrio de 500 ml, donde se introdujeron 10 g de tamo de arroz como sustrato, adicionado de 40 ml de solución de nutrientes definida por (Mandels & Weber 1969) (en gL‐1 fosfato de potasio 0,3, Sulfato de Magnesio 0,83, cloruro de calcio 0,3, sulfato de hierro 0,005, sulfato de manganeso 1,56 mg, Sulfato de zinc 1,4 mg tween 80 0,2 ml pH 6.0.) y un inóculo de 1,0 g de biomasa húmeda de los microorganismos seleccionados. Dentro del frasco se introdujo a su vez un frasco estéril con 20 ml de NaOH 0,3N, al cual se le confirmó la concentración por medio de titulación empleando biftalato de potasio y fenolftaleína como indicador. Los frascos se sellaron con tapones de caucho los cuales constaban de una jeringa de 5 cm para liberar la tensión del gas producido dentro del microcosmo y se incubaron por 28 días a 28°C. Cada 4 días se realizaron muestreos destructivos del hidróxido y se remplazó por una nueva trampa. El ensayo se llevó a cabo 46 por cuaadriplicado con un co ontrol abióttico que co onsistió en el mismo montaje, ccon la solución n de nutrien ntes y el sustrato pero ssin inoculacción del micrroorganismo. 3.2.5.3 Determinac D ción de CO22 Para deeterminar laa cantidad de CO2 producida en el microccosmo se lllevó a cabo una titulació ón acido‐base. Para ello se preparró HCl al 0,4 4 N y se reaalizó una co onfirmación n de la normalidad del ácido por meedio de la ttitulación de d Carbonatto de calcio o con naran nja de metilo ccomo indicaador. Al rettirar la tram mpa de NaO OH del micrrocosmo see adiciono B BaCl2 y fenolftaaleína y se realizó una titulación con el HCl 0,4N confirrmado anteeriormente..(Page 1982) .P Para la deteerminación de los miliggramos de CO2 produ ucidos duran nte el proceeso se realizó eel siguiente calculo (ver ecuacion1 1.2). (1.2) Dónde: aOH inicial q que se obtuvieron de laa concentración inicial empleada een las tramp pas de g de Na hidróxid do del micro ocosmo. g de NA AOH R se obtuvieron d de la relació ón entre las moles del ácido que sse empleó p para la titulació ón y las moles de NaOH H que reaccionaron. PM CO22 es el peso molecular d del compuesto PM NaO OH es el pesso molecular del compu uesto 47 3.2.5.4 Análisis de los resultados Los resultados se analizaron con el software de estadística IBM SPSS versión 19 (Armonk, NY, USA). Las pruebas de normalidad se realizaron con el test de Shapiro‐Wilk y la homogeneidad de varianzas con el test de Leven. Se aplicó un ANOVA de un factor y prueba de Tukey. El nivel de confianza fue del 95%. 3.3 Resultados y Discusión 3.3.1 Actividad celulolítica cualitativa Del banco de microorganismos empleado en la investigación, se cuenta con 16 aislamientos de los cuales 9 pertenecen al género Penicillium, 5 al género Aspergillus y 2 al género Paecilomyces. Reportes indican que los primeros dos géneros han sido ampliamente reconocidos como microorganismos capaces de producir complejos enzimáticos celulolíticos de manera eficiente, sin embargo para el género Paecilomyces son menos los estudios en este ámbito (Singhania et al. 2013). De acuerdo a un estudio anterior realizado por (Reyes & Aguirre 2009) estos aislamientos presentaron actividades celulolíticas en agar Celulosa, sin embargo para el presente estudio fue necesario confirmar dicha actividad e identificar los aislamientos celulolíticos promisorios. Para este fin se llevó a cabo una evaluación cualitativa por medio de la medición de halos de hidrolisis. La determinación cualitativa de producción de celulasas se lleva a cabo por medio de la utilización de carboximetil celulosa como sustrato, esta es una clase de celulosa amorfa que permite identificar la hidrolisis de la celulosa al adicionar el rojo Congo ya que este colorante es capaz de formar fuertes interacciones con polisacáridos unidos por enlaces β 1‐4 y al producirse la hidrólisis del compuesto la interacción entre el colorante y el polisacárido se pierde, observándose un área de aclaramiento cuando el microorganismo secreta las enzimas celulolíticas, así que el halo sería equivalente al potencial celulolítico del microorganismo (López & Freitas 2012a). 48 De acuerdo a los resultados y por medio de un análisis de diferencia de medias se encontró que la cepa MF 3‐69 (Aspergillus sp) presentó el menor halo de hidrolisis, las cepas MF 3‐65 Y 471 del genero Paecilomyces sp. MF 6‐68, MF 10‐63, MF 10‐60, MF 14‐57, MF 12,59, MF 10‐62, MF 14‐56, y MF 3‐70 pertenecientes al género Penicillium, al igual que las cepas MF 7‐67, MF 8‐66, MF 9‐58 del genero Aspergillus presentaron un halo de hidrolisis intermedio y las cepas MF 9‐61 (Aspergillus sp) y MF 3‐64 (Penicillium sp.) generaron el mayor valor con halos de 29 y 33 mm respectivamente presentándose diferencia estadísticamente significativa entre los tratamientos (p<0,05) (Fig 3.1) Existen diferentes especies del genero Penicillium las cuales han sido reportadas como productoras de enzimas celulolíticas, algunos géneros reportados corresponden a P. achinulatum, P. decumbens, P. occitans, P. funiculosum, P. phinophilum y P. brasilianum.(Adsul et al. 2007), (Maeda et al. 2013), (Camassola & Dillon 2009). Así mismo dentro del género Aspergillus se reportan las especies A. niger, A. nidulans, A. fumigatus entre otras.(Bansal et al. 2012), (Liu, Zhang, Yang, Wu, et al. 2011), (Lockington et al. 2002) . Por otro lado Trichoderma sp. ha sido especialmente estudiado para dilucidar el sistema de regulación enzimática celulolítica en hongos filamentosos (Ma et al. 2013). Debido a esto se evaluaron dos aislamientos de Trichoderma sp. de diferente origen, empleados como bioinoculantes microbianos, ya que existen reportes sobre la producción de celulasas de este género en la literatura, (Liming & Xueliang 2004), (Latifian et al. 2007), (Gutierrez‐ correa et al. 1999),asi mismo uno de los aislamientos es empleado para la degradación artesanal del residuo en campo sin embargo no se encontró producción de halo de hidrolisis en las condiciones del ensayo (Fig 3.1). Figura 3.1 Halos de hidrólisis en mm de las cepas 3‐70 (Penicillium sp.), 3‐65 (Paecilomyces lilacinum), 9‐61 (Aspergillus sp), 3‐64 (Penicillium oxalicum), 4‐71 (Paecilomyces sp.), 9‐58 (Aspergillus sp), 14‐56 (Penicillium sp) 8‐66 (Aspergillus sp.), 10‐62 (Penicillium sp), 12‐56 (Penicillium palitans), 14‐57, Trichoderma sp (A), Trichoderma sp. (B). 49 Para realizar la selección de las cepas fue necesario plantear otra metodología que permitiera evaluar efectivamente la producción de celulasas por parte de los aislamientos debido a que la evaluación cualitativa no fue concluyente al contrastar los resultados con las cepas empleadas para degradar el tamo en campo, en este caso Trichoderma sp. el cual no presento un halo de hidrolisis medible. Y por otro lado solo 6 aislamientos presentan halos por encima de la media con respecto al MF 3‐64 que fue el aislamiento que genero mayor halo de hidrolisis. 3.3.2 Evaluación de la actividad celulolítica cuantitativa Posterior a la evaluación cualitativa se llevó a cabo la cuantificación enzimática celulolítica de los 16 aislamientos en fermentación sumergida con tamo como fuente de carbono y con papel filtro como sustrato de la reacción. La cuantificación se realizó a los 4 y 6 días ya que por estudios anteriores dentro del grupo de investigación se estableció que en este tipo de medio de cultivo la máxima producción enzimática celulolítica se obtuvo en este lapso de tiempo (Perez & Hernandez 2011). 50 Como resultado de la cuantificación de las celulasas totales por la técnica con papel filtro como sustrato en la reacción, se encontró que de los 16 aislamientos solamente las cepas MF 3‐64 (Penicillium sp. y MF 3‐69 (Aspergillus sp.) presentaron actividad celulítica, obteniéndose 68 UE, y 0,6 UE respectivamente a los cuatro días (Fig 3.2). Estos resultados contrastan con los obtenidos en la evaluación cualitativa ya que el aislamiento 3‐69 que presento menor halo de hidrolisis, en este caso fue una de las dos cepas de las que se obtuvieron unidades celulolíticas totales. Por otro lado el aislamiento MF 3‐64 fue el único que presento correspondencia entre las dos técnicas siendo el microorganismo que presento mayor actividad cualitativa y cuantitativa. Como se expuso anteriormente son varios los reportes que se encuentran sobre la expresión de enzimas celulolíticas por parte de diferentes cepas del genero Penicillium al que corresponde el aislamiento MF 3‐64, Sin embargo existen estudios donde las unidades enzimáticas expresadas como FPasas debido a la utilización de papel filtro como sustrato son mucho mayores que las obtenidas por el aislamiento de estudio. Debido a que se sabe qué factores como el pH, la temperatura y el tipo de sustrato influye en la reacción enzimática es posible que el resultado anterior se deba a la utilización de una clase de papel filtro diferente a la reportada por (Ghose 1987) ya que se sabe que al emplear sustratos diferentes en la reacción se pueden obtener actividades variables. En estudios realizados por (Camassola & J.P. Dillon 2012) se encontró que al emplear papel filtro Whatman N°1, Avicel, Celulosa SigmaCell‐20 y discos de papel filtro en la reacción con la misma preparación de celulasa comercial se obtuvieron valores de 60.4 FPU/ mL, 14.1 FPU/mL 34.7 FPU/mL y 35.2 FPU/ mL respectivamente, demostrándose que aun al emplear la misma concentración de enzima los resultados pueden ser variables dependiendo el sustrato empleado en la reacción. Existen otros factores que pudieron influir en la poca actividad celulolítica registrada. En la técnica propuesta por (Ghose 1987) empleando papel filtro como sustrato de la reacción enzimática es necesario tener en cuenta que si bien es un método rápido y ampliamente empleado para la determinación de celulasas totales, existen ciertos factores a tener en 51 cuenta. Por ejemplo es necesario que el extracto enzimático en la reacción tenga una concentración adecuada capaz de convertir o degradar el 4% del sustrato en 60 minutos para ser cuantificable. Por otro lado se sabe que la técnica es adecuada para la determinación de extractos concentrados, siendo inadecuado para preparaciones con títulos bajos enzimáticos, los cuales se pueden encontrar en sobrenadantes de cultivos de microorganismos que no ha sido purificados o concentrados. (Luciano et al. 2012), siendo este el caso, ya que los extractos se obtuvieron por centrifugación sin realizar metodologías que permitieran concentrar o purificar las enzimas celulolíticas. Por otro lado es probable que el tiempo de fermentación no haya sido el suficiente para obtener la máxima producción enzimática, o que se haya encontrado en días que no se evaluaron dentro del diseño experimental. Es importante aclarar que la determinación de azucares reductores en el trabajo de (Perez & Hernandez 2011) de donde se obtuvieron los tiempos máximos de actividad sobre esta misma fuente de carbono se llevó a cabo por la técnica de Somogy‐Nelson, a diferencia del presente estudio donde se empleó la técnica de DNS. En el estudio realizado por (Marjamaa et al. 2013) se encontró que la mayor producción enzimática celulolítica se obtuvo entre los 4 y 6 días de cultivo cuando se empleó celulosa microcristalina como fuente de carbono. Esto sugeriría que al emplear una fuente de carbono compleja como el tamo el tiempo de máxima producción podría incluso superar los seis días de cultivo. Al igual que en el ensayo cualitativo se evaluaron los aislamientos de Trichoderma y en esta ocasión tampoco se encontraron valores de actividad enzimática. Aunque se reporta que este microorganismo es capaz de producir grandes cantidades de enzimas celulolíticas, en la investigación de (Marjamaa et al. 2013) se encontró que una cepa de Penicillium produjo incluso dos veces más actividad enzimática celulolítica comparado con Trichoderma ressei, microorganismo ampliamente empleado para procesos de producción enzimática por los altos títulos de obtención. 52 Figura 3.2 Determinación de actividad celulolítica (FPasa) de 3‐64 (Penicillium oxalicum), 3‐ 70 (Penicillium sp.), 3‐69 (Aspergillus sp), Trichoderma sp. (A), Trichoderma sp. (B) con Papel filtro como sustrato de la reacción enzimática. Actividad Celulasa (U/L) Es importante mencionar que la diferencia entre los resultados cualitativos y cuantitativos puede estar relacionada con el medio de cultivo y el sustrato empleado en las dos metodologías. Mientras en la evaluación cualitativa se presenta en medio sólido el cual puede dificultar la difusión de la enzima, en la evaluación cualitativa se encuentra en medio líquido y con un sustrato que es de mayor complejidad ya que se empleó el tamo como fuente de celulosa, el cual está compuesto por otros polímeros que pueden dificultar o demorar la degradación. En el estudio realizado por (López & Freitas 2012a) se encontró un comportamiento similar, ya que los microorganismos que presentaron actividad celulolítica en agar celulosa no presentaron actividad enzimática FPasa ni EG. Debido a que los resultados en ambas metodologías al parecer no tienen correspondencia se decide llevar a cabo una metodología indirecta de cuantificación de degradación del sustrato para lo cual se pre seleccionaron los aislamientos Penicillium sp. MF 3‐64, que presento mejor actividad tanto cualitativa como cuantitativa, el aislamiento MF 3‐69 que presento actividad cuantitativa celulolítica, y adicionalmente se seleccionó el aislamiento Paecilomyces sp. MF 4‐71, que si bien no presentó el mayor halo de hidrolisis, presento un 53 halo de 20 mm el cual se encontraba entre los más altos y por otro lado este aislamiento representa uno de los tres géneros que componen el banco de aislamientos con potencial celulolítico. 3.3.4 Evaluación indirecta de la degradación del sustrato vegetal (tamo) Algunos de los parámetros evaluados en una fermentación solida son el consumo de oxígeno y/o la producción de CO2. La metodología de respirometría puede dar información de la actividad microbiana en el sustrato, siguiendo la producción de CO2 se puede tener una visión clara de la actividad metabólica que se está llevando a cabo en el microcosmo. Este parámetro permite asegurar la biodegradabilidad del sustrato en el medio sólido (Willcock & Magan 2000), (Bellon‐Maurel et al. 2003). Además de esto los procesos en fase solida permiten que le microorganismos crezca en condiciones similares a las que se encuentran en la naturaleza, produciendo conidios aéreos que son más resistentes (Sofía et al. 2001). Debido a lo anterior la evaluación en microcosmos permitió simular las condiciones de la degradación del sustrato en estado natural por medio del seguimiento de la producción de CO2. De acuerdo a los resultados del análisis de varianza, existen diferencias entre los tratamientos evaluados (p<0.005). Como se observa en la Figura 3.3, el aislamiento MF 3‐64 presento la mayor producción de CO2 obteniéndose al cabo de 28 días alrededor de 635 mg, seguido de Paecilomyces sp. con 299 mg y Aspergillus sp. con 180 mg. Lo anterior puede estar relacionado entre otros factores con la producción de enzimas capaces de degradar eficientemente el residuo, lo cual corresponde con los resultados obtenidos en las evaluaciones anteriores en donde el aislamiento MF 3‐64 (Penicillium sp. ) presento la mayor actividad celulolítica, así mismo se tienen reportes sobre la amplia diversidad enzimática que producen diferentes especies de Penicillium, encontrándose producción de enzimas tanto celulolíticas como hemicelulolíticas (Camassola & Dillon 2009), (Terrasan et al. 2010), (Chen et al. 2010). 54 Figura 3.3 Determinación de CO2 en microcosmos en función del tiempo de las cepas MF 3‐ 64 Penicillium sp. MF 4‐71 Paecilomyces sp. MF 9‐61Aspergillus sp. 600 500 mg CO2 /g de sustrato MF 3-64 MF 4-71 MF 9-61 400 300 b 200 c 100 0 0 a 5 10 15 20 25 30 Tiempo (dias) El aislamiento 4‐71 (Paecilomyces sp.) presento visualmente mayor colonización del sustrato comparado con Penicillium sp. (Fig 3.4), sin embargo produjo menor cantidad de CO2. Este comportamiento puede estar sujeto a diversos factores como la naturaleza del residuo, la producción enzimática del microorganismo y la colonización superficial del sustrato. Se ha encontrado que algunas especies de Paecilomyces son capaces de crecer en diversos hábitats como por ejemplo suelo, madera en descomposición entre otros materiales lignocelulósicos. Para obtener nutrientes de este tipo de materiales Paecilomyces produce diversas enzimas como EG, hemicelulasas y en algunos casos ligninasas, (Kluczek‐Turpeinen et al. 2007), de acuerdo a lo anterior, aunque el microorganismo no produjo la misma cantidad de CO2 que Penicillium sp. no se puede descartar que sea un aislamiento promisorio en la degradación del residuo, ya que la actividad metabólica puede estar sujeta a factores como el porcentaje de humedad, la temperatura de incubación, y la adición de nutrientes que pueden inducir la actividad enzimática (Yang et al. 2006), condiciones que no se evaluaron en el ensayo. 55 Adiccionalmentee es posible que Paeecilomyces requiera dee una etap pa de adap ptación al susttrato, ya que al observaar la produccción de CO O2 entre el o octavo y el vigésimo díía, esta se man ntiene consstante y ess a los 28 días de eevaluación que se prresenta un aumento significativo en la degradacción. ura 3.4 Ensaayo de micro ocosmos em mpleando ell residuo como sustrato o Figu A Penicilliu A. um sp MF 3 3‐64. B. Paeccilomyces sp p.MF 4‐71 A B En el caso de Aspergilllus sp., au unque exissten reporttes de la utilización de este micrroorganismo o para la degradación de com mpuestos lignocelulóssicos y la eficiente prod ducción de enzimas caapaces de d degradar la celulosa y la hemicelu ulosa (Gomathi et al. 2012 2)(Gottschaalk et al. 2010) (Ban nsal et al. 2012), la producció ón de CO2 no fue sobrresaliente. EEs posible q que exista iggualmente u una influenccia del sustrrato en la producción enzimática y d degradación n del mismo o, ya que según el estudio de (Bansal ( et al. 2012), pleando Asp pergillus sp. se encontraaron mejorees actividad des enzimáticas CMCasa, FPAsa y emp β‐ glucosidasaa en salvado de trigo como sustrrato a diferrencia de o otro tipo dee residuos com mo, cascarillaa de arroz, rresiduo de ccultivo de m maíz, bagazo o de caño p paja de trigo o. En otros estu udio se encontraron mayores aactividades con rastro ojo de maííz como su ustrato, a diferencia de taamo de arro oz (Liu, Zhan ng, Yang, Wu u, et al. 201 11) 56 Merece mencionar que la tendencia en la producción de CO2 para el caso de los tres microorganismos es continuar aumentando, ya que no se observó una fase de disminución de la producción de CO2 en este tiempo, lo cual podría sugerir que la degradación completa del sustrato requiere llevarse a cabo en un tiempo más prolongado de cultivo. En algunos estudios en fermentación solida empleando sustratos vegetales como el tamo el tiempo de evaluación supera los 40 días (Henriksen & Breland 1999). Igualmente se encontró que el control abiótico presento una producción constante de CO2 la cual puede ser consecuencia de la composición normal de este compuesto en el aire, ya que en promedio se obtuvieron 730 ppm valor que corresponde al encontrado en zonas urbanas para este compuesto. De acuerdo a los resultados de la actividad celulolítica cualitativa, cuantitativa y a la producción de CO2 se decide que Penicillium sp. y Paecilomyces sp. son los aislamientos seleccionados para continuar con el siguiente objetivo del proyecto, ya que presentaron valores significativos en una o en las tres metodologías de selección. 57 Capítulo 4. Evaluación de la producción enzimática bajo diferentes fuentes de carbono y nitrógeno 4.1 Introducción La función de cada una de las enzimas que componen el complejo celulolítico es de interés para alcanzar la transformación del residuo en compuestos aprovechables tanto por la microbiota del suelo como por la planta debido a su acción sinérgica y cooperativa. Por este motivo es de importancia establecer que los microorganismos empleados en la biotransformación de los residuos agrícolas sean capaces de sintetizarlas eficientemente. Así mismo el estudio de la influencia de diferentes fuentes de carbono y nitrógeno en la inducción de la producción enzimática puede aportar información para identificar las características de los complejos enzimáticos. Las celulasas y hemicelulasas son sistemas enzimáticos inducibles por diferentes fuentes de carbono, sin embargo existen otro tipo de fuentes nutricionales que podrían producir una respuesta en la actividad enzimática. Se ha reportado que la expresión de celulasas está regulada en mayor medida por la fuente de carbono y en una menor proporción por la fuente de nitrógeno, es así que la mayoría de investigaciones se han enfocado en la evaluación de diferentes fuentes de carbono y se ha subestimado la influencia que podría tener la fuente de nitrógeno en el cultivo, y más aún cuando los microorganismos que se emplean para la producción enzimática están habituados a ciertas condiciones nutricionales propias del ambiente donde se desarrollan que podrían influir en la síntesis de estas enzimas. Por otro lado muchos microorganismos han sido clasificados como celulolíticos, pero solo unos pocos poseen un completo sistema celulolítico capaz de llevar a cabo una eficiente degradación de la celulosa (Ahamed & Vermette 2008). Para el presente objetivo se planteó la evaluación del complejo enzimático celulolítico comprendido por dos clases de enzimas denominadas: exo‐1,4 β‐d‐glucanasa (EC 3.2.1.91), 58 endo‐1,4‐ β‐d‐glucaanasa (EC 3 3.2.1.4) las cuales actúan cooperaativamente y sinérgicam mente para depolimerizarr la celulosa a celobiosaa y oligosacáridos corto os los cuales son conveertidos a glucossa por la en nzima β‐glu ucosidasa (EEC 3.2.1.21)) la cual se evaluó iguaalmente. Aunque esta enzima no acctúa directamente sobre la celulo osa, su activvidad es im mportante p porque puede reducir la acción inh hibitoria que ejerce la celobiossa sobre eendoglucanaasas y exoglucanasas. Assí mismo see realizó la evaluación n de la enzima xilanasaa como parrte del complejjo enzimáticco necesario o para degraadar la hem micelulosa. 4.2 Materialess y Métodos 4.2.1 Evaluación de la expresión n de celu ulasas en diferentees fuentees de no y nitróggeno carbon Una vez determin nadas las cepas quee presentaron mayor producción de celulaasas y eficientee degradaciión del sustrato se llevó ó a cabo la evaluación de la expreesión de celulasas y hemiccelulasas en n diferentes fuentes d de carbono o y nitrógen no. Para ello se plantteó un diseño ffactorial irrregular en d donde el Faactor A fuee la fuente de carbono o y el Facto or B la fuente d de nitrógeno, las variab bles de resp puesta fuero on las actividades enzim máticas EG, CBH y BGL (Tabla 4.1) n de la exprresión de celulasas Tabla 4..1 Diseño faactorial irreggular para laa evaluación 59 4.2.2 Cultivo de los microorganismos para la evaluación de celulasas Los cultivos se realizaron en matraces de 250mL, con un volumen efectivo de 120 mL. Se empleó la solución de macronutrientes propuesta por (Mandels & Weber 1969) (En gL‐1: KH2PO4, 0,3; MgSO4.7H2O, 0,83; CaCl2, 0,3; FeSO4.7H2O, 0,005; CaCl2, 0,02; MnSO4.H2O, 1,56 mg; ZnSO4.7H2O, 1,4 mg y Tween 80, 0,2 mL, pH 6,0) y se adiciono la respectiva fuente a evaluar de acuerdo al diseño experimental en una concentración de 10 gL‐1 para las fuentes de carbono y 5 gL‐1 para las fuentes de nitrógeno. Las fuentes de carbono evaluadas fueron, Carboximetilcelulosa (SIGMA) y Avicel (SIGMA) y las fuentes de nitrógeno, Extracto de levadura (Scharlau) y Sulfato de amonio (Merck). El inóculo consistió en 12 mL de una suspensión de esporas con una concentración de 106 esporas.mL‐1, de cada microorganismo. Después de inoculados, los matraces se incubaron a 28°C , en una incubadora orbital a 120 rpm por 10 días. 4.2.3 Cultivo de los microorganismos para la evaluación de xilanasas Los cultivos se realizaron en matraces de 250mL, con un volumen efectivo de 120 mL . Se empleó la solución de macronutrientes propuesta por (Mandels & Weber 1969) adicionado de 10 gL‐1 de Xilano SIGMA (X4252) y 5 gL‐1 la fuente de nitrógeno a evaluar. El inóculo consistió en 12 mL de una suspensión de esporas con una concentración de 106 esporas.mL‐1, de cada microorganismo. Después de inoculados, los matraces se incubaron a 28°C en una incubadora orbital a 120 rpm por 10 días. 4.2.4 Obtención de los extractos Se tomaron 8 ml de muestra a los 0, 4, 6, 8, y 10 días de cultivo, las muestras fueron centrifugados a 10000 rpm durante 20 minutos a 4ºC y posteriormente a 5000 rpm durante 20 minutos, para retirar la biomasa y los residuos de cultivo, a partir del sobrenadante obtenido se realizaron las pruebas de actividad enzimática. 60 4.2.5 Actividades Enzimáticas Todas las actividades enzimáticas fueron determinadas espectrofotométricamente usando un equipo Evolution 60 UV‐VIS Thermo Scientific Co (Waltham, MA, USA). 4.2.5.1 Actividad endo‐1,4‐β‐glucanasa (EG) Se realizó mezclando 500 µL de muestra con 500 µL de carboximetil celulosa de baja viscosidad (C 5678)) (2 % p/v) en buffer citrato 50 mM (pH 5.0) a 40°C por 60 minutos. (Quevedo 2011). La cantidad de azúcares reductores en términos de equivalentes de glucosa, se determinaron por la técnica de DNS (Miller 1959). Una unidad de actividad glucanasa (UG) es la cantidad de enzima que libera 1 µmol de azúcares reductores por minuto a las condiciones del ensayo. 4.2.5.2 Actividad exo‐1,4‐β‐glucanasa (CBH) Se evaluó usando como sustrato p‐nitrofenil‐β‐D‐celobiosido (PNPC, Sigma). La reacción contenía 320 µL de PNPC 5 mM en buffer acetato de sodio 50 mM (pH 5.0) y 100 µL de muestra. Esta mezcla se incubó a 40ºC por 60 minutos. La reacción se detuvo adicionando 250 µL de carbonato de sodio 0.5 M. La absorbancia se leyó a 400 nm. (Quevedo 2011). La actividad enzimática se calculó usando la curva patrón de p‐nitrofenol. Una unidad de actividad exoglucanasa o celobiohidrolasa (UC) se define como la cantidad de enzima que libera 1 µmol de p‐nitrofenol por minuto a las condiciones del ensayo 4.2.5.3 Actividad de 1‐4‐β‐glucosidasa (BGL) Se cuantificó usando como sustrato p‐nitrofenil‐β‐D‐glucosido (PNPG, SIGMA), siguiendo el mismo método que para la actividad celobiohidrolasa.. 61 4.2.5.4 Actividad xilanasa. La actividad xilanasa se determinó mezclando 500 µL de muestra con 500 µL de xilano de árbol de haya (X 4252)) (2%p/v) en buffer citrato 50 mM (pH 5.0) a 40°C por 60 minutos. (Driss et al. 2011). La cantidad de azúcares reductores liberados se determinó por la técnica de DNS con xilosa como estándar. Una unidad de actividad xilanasa (UX) se define como la cantidad de enzima que libera 1 µmol de azúcares reductores por minuto. 4.2.6 Análisis de los resultados Para establecer la influencia de los tratamientos sobre la expresión de cada una de las enzimas, se realizó un análisis de área bajo la curva con el programa Graphpad Prisma 6.0 (GraphPad Software Inc.) en donde la variable de respuesta fue el ABC de las unidades enzimáticas endo‐1,4‐β‐glucanasa, exo‐1,4‐β‐glucanasa, 1‐4‐β‐glucosidasa y xilanasa. Posteriormente se realizó un análisis de varianza de una vía (ANOVA), y una prueba post‐ hoc de Tukey (α=0.05), en el programa estadístico SPSS (IBM 19). 4.3 Resultados y Discusión Para determinar el efecto que tienen diferentes fuentes de carbono y nitrógeno en la producción enzimática de las dos cepas de hongos celulolítico se evaluó la presencia de avicel, carboximetilcelulosa y xilano en los cultivos, así como dos fuentes de nitrógeno correspondientes a una orgánica (extracto de levadura) y una inorgánica (sulfato de amonio), la mezcla de estas dos y la ausencia de fuente de nitrógeno. Adicionalmente por medio del diseño experimental se logró establecer la habilidad que tiene cada cepa (Penicillium sp MF 3‐64 y Paecilomyces sp MF 4‐71.) para producir las enzimas que componen el complejo celulolítico y una de las enzimas del complejo hemicelulolítico. 62 4.3.1 Perfil de la producción enzimática celulolítica Dentro de la búsqueda de microorganismos productores de enzimas celulolíticas, es importante determinar que enzimas del complejo enzimático son producidas y que factores pueden tener una relación directa con la síntesis. En el ensayo se evaluaron diferentes fuentes de carbono y nitrógeno en fermentación liquida para establecer su influencia en la producción enzimática. De acuerdo a los resultados se evidencio que los dos microorganismos son capaces de producir las enzimas que componen el complejo celulolítico comprendido por la EG y la CBH (Figura 4.1, Figura 4.2), así mismo producen la enzima β glucosidasa, confirmando los resultados de las evaluaciones cualitativas y cuantitativas (Cap.3) en donde se evidencio la presencia de enzimas celulolíticas. Existen reportes donde se menciona que diferentes especies de Penicillium son capaces de producir las diferentes enzimas del complejo entre las que se encuentra endoxilanasas, celulasas, arabinofuronidasas, acetilesterasas, xilosidasas (Jørgensen et al. 2005), así mismo se reporta en la mayoría de estudios a Paecilomyces thermophila como productor de enzimas celulolíticas y xilanoliticas (Yang et al. 2006) (Li et al. 2006) (Jia et al. 2012)(Yang et al. 2009) . Sin embargo es importante mencionar que los microorganismos muestran diferencias en la magnitud de la respuesta, presentando Penicillium sp. los valores enzimáticos más altos con respecto a las 3 enzimas. Los resultados permiten establecer que existen diferencias en el comportamiento de la producción enzimática en las tres enzimas evaluadas. En el caso de Penicillium sp MF 3‐64 con respecto a la EG se encontró que indistintamente del tratamiento con nitrógeno, cuando se empleó carboximetilcelulosa como fuente de carbono, se alcanzó la mayor actividad enzimática en el día 10 de cultivo. Se obtuvieron valores enzimáticos en un rango de 20 a 70 U/L. Cuando la fuente de carbono fue avicel, la máxima producción enzimática fue diferente según los tratamientos, presentándose en algunos casos en el día 6, 8 y 10. Los valores enzimáticos se encontraron entre 10 y 50 U/L. Lo anterior sugiere que la 63 producción enzimática presenta un comportamiento irregular en el tiempo (Figura 4.1 A1, A2).Para Paecilomyces sp. la producción enzimática se da desde el día 6 en todos los tratamientos, a diferencia de Penicillium donde desde el día 4 de fermentación se obtuvieron altas actividades de esta enzima (Figura 4.2 A1,A2), por otro lado el comportamiento de la producción es diferente ya que se observó un punto máximo de producción a los 8 días y posteriormente un descenso en la actividad, que en el caso de algunos tratamientos llego a cero. Para la CBH se encontraron valores máximos de actividad entre el día 6 y 8, encontrándose en la mayoría de tratamientos un descenso en la producción hacia el final del cultivo (Figura 4.1 B1, B2). Se encontraron diferencias en los valores máximos alcanzados de acuerdo a cada tratamiento En el caso de la β‐ glucosidasa se obtuvieron valores máximos enzimáticos en la mayoría de tratamientos en los días 4 y 6 de fermentación, se identificó igualmente que después de un descenso en algunos casos drástico de la producción enzimática, esta volvió a aumentar hacia el final del cultivo. Se obtuvieron valores enzimáticos de alrededor de 4000 U/L. (Figura 4.1. C1,C2). Estos altos valores de actividad sugieren que Penicillium podría estar contrarrestando eficientemente la represión catabólica causada por la celobiosa, sintetizando grandes cantidades de enzima β glucosidasa desde el inicio de la fermentación lo cual en algunos casos según se reporta puede ser benéfico para la inducción de la producción de las otras enzimas del complejo celulolítico (Juhász et al. 2005). Por otro lado se ha sugerido que en algunas especies de Penicillium la expresión de esta enzima es constitutiva, lo cual coincide con mecanismos regulatorios de otros hongos filamentosos (Sun et al. 2008a). 64 Figura 4.1 Cinéticas de actividad enzimática bajo diferentes fuentes de carbono y nitrógeno de Penicillium sp. Endoglucanasa (A1, A2), Celobiohidrolasa (B1,B2), β –glucosidasa (C1,C2) A1 70 60 50 40 Actividad Endo 1,4--glucanasa (U/L) B1 Actividad Celobiohidrolasa (U/L) 55 50 45 40 35 30 25 30 20 20 15 10 10 5 0 0 0 2 4 6 8 0 10 350 400 250 250 200 200 150 150 100 100 50 0 0 2 5000 Actividad 1,4 -glucosidasa (U/L) 4 6 8 10 0 10 0 CMC CMC + Ext.Lev CMC + (NH 4)2SO 4 CMC + Ext.Lev + (NH 4)2SO 4 4000 2 4 4000 3000 3000 2000 2000 1000 1000 6 8 10 Avicel Avicel + Ext. Lev Avice + (NH 4)2SO 4 Avicel + Ext. Lev + (NH 4)2SO 4 5000 8 50 6 Avicel Avicel + Ext.Lev. Avicel + (NH 4)2SO 4 Avicel + Ext.Lev. + (NH 4)2SO 4 350 300 4 B2 CMC CMC + Ext. Lev CMC + (NH 4)2SO 4 CMC + Ext. Lev + (NH 4)2SO 4 300 2 450 C1 Avicel Avicel Ext.Lev. Avicel (NH 4)2SO 4 Avicel + Ext.Lev. + (NH 4)2SO 4 60 CMC CMC + Ext.Lev. CMC + (NH 4)2SO 4 CMC + Ext. Lev +(NH 4)2SO 4 400 A2 65 80 0 0 0 2 4 6 Tiempo (dias) 8 10 0 2 4 6 Tiempo (dias) 8 10 C2 65 En general las unidades enzimáticas obtenidas empleando Paecilomyces sp. fueron menores comparadas con Penicillium sp. se observó que en el caso de la enzima EG no se encuentra producción enzimática en el día 4, y este comportamiento se observa en todos los tratamientos (Figura 4.2 A1,A2), a diferencia de Penicillium donde se obtuvieron actividades en este tiempo. Se observó un punto máximo de producción a los 8 días y posteriormente un descenso en la actividad, que en el caso de algunos tratamientos llego a cero. El valor máximo alcanzado fue de 65 U/L. En el caso de la enzima CBH y β glucosidasa igualmente se observa que en el día 10 de muestreo la actividad en la mayoría de tratamientos llegar igualmente a cero. Los valores máximos de producción son de 220 U/L y 225 U/L respectivamente. De acuerdo a los resultados anteriores se observó que existen diferencias en el comportamiento de la producción enzimática para el caso de los dos microorganismos, presentando Penicillium mayor actividad enzimática en todas las enzimas evaluadas. Este aislamiento produce el doble de actividad enzimática en el caso de la CBH e inclusive supera en unidades de miles la actividad β glucosidasa con respecto a Paecilomyces. Por otro lado presenta producción enzimática más prolongada, ya que inclusive en algunos tratamientos según los resultados la tendencia de la producción es a seguir aumentando después de los 10 días de evaluación, mientras que Paecilomyces detiene la producción enzimática en algunos tratamientos desde el día 8 de cultivo. Figura 4.2 Cinéticas de actividad enzimática bajo diferentes fuentes de carbono y nitrógeno de Paecilomyces sp. Endoglucanasa (A1, A2), Celobiohidrolasa (B1,B2), β –glucosidasa (C1,C2) 66 A1 Actividad Endo 1,4--glucanasa (U/L) 80 70 60 A2 Avicel Avicel + Ext. Lev Avicel + (NH 4) 2SO 4 Avicel + Ext. Lev + (NH 4) 2SO 4 70 CMC CMC + Ext. Lev CMC + (NH4)2SO4 CMC + Ext. Lev + (NH4)2SO4 60 50 50 40 40 30 30 20 20 10 0 10 0 0 2 4 6 8 0 10 2 4 6 8 10 300 Actividad Celobiohidrolasa (U/L) B1 CMC CMC + Ext. Lev CMC + (NH4)2SO4 CMC + Ext. Lev + (NH4)2SO4 250 100 200 80 150 60 100 40 50 20 0 0 0 Actividad 1,4 -glucosidasa (U/L) C1 B2 Avicel Avicel + Ext. Lev Avicel + (NH 4) 2SO 4 Avicel + Ext. Lev + (NH 4 )2 SO 4 120 2 4 6 8 10 0 250 CMC CMC + Ext. Lev CMC + (NH4)2SO 4 CMC + Ext. Lev + (NH4)2SO 4 2 4 6 8 10 C2 Avicel Avicel + Ext. Lev Avicel + (NH 4) 2SO 4 Avicel + Ext. Lev + (NH 4)2SO 4 120 100 200 80 150 60 100 40 50 0 20 0 0 2 4 6 8 10 0 2 4 6 Tiempo (dias) Tiempo (dias) 8 10 67 4.3.2 Actividad Enzimática Endoglucanasa Después de realizar el análisis de área bajo la curva de las unidades enzimáticas, En el caso de Penicillium sp. se encontró que existe una diferencia entre los tratamientos evaluados (p<0,005), y que el tratamiento compuesto por carboximetil celulosa y sulfato de amonio presento los valores más altos de producción enzimática, en contraste con el tratamiento compuesto por esta misma fuente de carbono con una mezcla de fuente de nitrógeno orgánica e inorgánica el cual fue el tratamiento que se presentó menor respuesta de actividad enzimática. En el caso de Paecilomyces se encontró que el mejor tratamiento fue el compuesto por Avicel y Sulfato de amonio. Sin embargo es importante resaltar que los demás tratamientos con avicel presentaron muy poca respuesta. Lo anterior podría sugerir que la influencia de la producción enzimática en el caso de la EG podría estar sujeta a la interacción entre la fuente de carbono y la fuente de nitrógeno, y adicionalmente que en cada microorganismo la inducción de la producción enzimática se lleva a cabo por dos tipos de fuente de carbono diferente, sin embargo el sulfato de amonio parece ser la fuente nutricional que estaría condicionando la actividad enzimática. Aunque se sabe que las endoglucanasas son generalmente inactivas contra la celulosa cristalina, se cree que los microorganismos secretan múltiples endogluanasas con un amplio rango de especificidad por un sustrato para llevar a cabo una hidrolisis eficiente de los sustratos celulósicos (Abe & Yano 2009). Lo anterior podría explicar por qué en el caso de Paecilomyces la mejor respuesta se obtuvo con celulosa micro cristalina y no con carboximetilcelulosa como se encontró con Penicillium. Por otro lado es importante mencionar que para la determinación de la actividad enzimática de los extractos obtenidos en los diferentes tratamientos se empleó como sustrato de la reacción carboximetilcelulosa, este sustrato es considerado para la determinación de EG debido a que imposibilita el paso del polímero por la estructura en forma de túnel característica de la mayoría de exoglucanasas, haciendo que la hidrolisis solo pueda ser llevada a cabo por enzimas de tipo EG. Al determinar las unidades enzimáticas 68 por este método se está implicando que la actividad evaluada corresponde en mayor proporción a la enzima evaluada. 4.3.3 Actividad enzimática Exoglucanasa (CBH) Se estableció que para Penicillium sp. existen diferencias entre los tratamientos con respecto al area bajo la curva (ABC) de la actividad enzimática (p<0.005), y que al igual que en el caso de la EG el tratamiento compuesto por carboximetilcelulosa y sulfato de amonio (Tratamiento 3) presentó los valores más altos de actividad. Los tratamientos en los que se presentó menor respuesta corresponden a la utilización de avicel y sulfato de amonio y avicel sin fuente de nitrógeno. Para Paecilomyces sp. se encontró que nuevamente el tratamiento 3 generó la mejor respuesta. Es de interés resaltar que la enzima celobiohidrolasa es capaz de hidrolizar celulosa cristalina a diferencia de la endoglucanasa, ( Silveira et al. 2012), sin embargo las actividades enzimáticas más altas se obtuvieron con la celulosa amorfa. En estudios realizados con H. jecorina se encontró que la estructura del sitio catalítico de la exoglucanasa difiere de la endoglucanasa debido a que esta última posee un sitio activo en forma de hendidura abierta, lo que permite su unión en cualquier ubicación a lo largo de la cadena siendo más efectiva en hidrolizar celulosa amorfa, mientras que en la CBH el sitio catalítico está oculto en el interior de una estructura de túnel formado por cadenas antiparalelas que permitirían el acceso solo de las cadenas poliméricas en los extremos, por lo cual esta enzima sería más efectiva en la hidrólisis de celulosa cristalina (Luciano et al. 2012), por tal razón se esperaría que la respuesta frente al avicel fuera positiva, sin embargo en ambos casos la fuente de carbono que presento mejor respuesta fue la amorfa implicando que las enzimas celobiohidrolasas de ambos microorganismos son estimuladas por fuentes con mayor proporción de celulosa amorfa. Adicionalmente, de acuerdo a los resultados anteriores, debido a que la mayor actividad Endoglucanasa se obtuvo con Carboximetilcelulosa, es posible que la inducción de la CBH se halla dado en mayor medida en esta misma fuente debido a que se sabe que las enzimas actúan de forma sinérgica y se 69 esperaría que al haber actividad Endoglucanasa alta se estimulara la producción de CBH. Con respecto a lo anterior, en estudios llevados a cabo con T. ressei se encontró que existe una inducción cruzada de las enzimas, y que en algunos casos para llevar a cabo la hidrolisis de un sustrato en común es necesaria la expresión tanto de EG como de CBH, así cada enzima tenga cierta especificidad por un tipo de celulosa diferente. (Sun et al. 2008b) Adicionalmente se sabe que la hidrólisis enzimática en general contempla tres pasos: la adsorción de la enzima a la superficie, la hidrólisis de la celulosa y la desorción de la enzima del sustrato. Acerca de esto se ha encontrado que la hidrolisis de la celulosa amorfa se lleva a cabo más rápido que la cristalina y esto podría influir en el grado de adsorción de la enzima al sustrato (Bansal et al. 2009); este factor podría incidir en la recuperación de la enzima en el extracto y por consiguiente a las bajas actividades enzimáticas registradas cuando avicel fue la fuente de carbono. 4.3.4 Actividad enzimática β‐glucosidasa La enzima β‐ glucosidasa es altamente dependiente de las condiciones del cultivo, se ha sugerido que existen más de un tipo de β‐ glucosidasa y que estas ejercen un importante papel en la síntesis de EG y CBH por medio de la formación de inductores enzimáticos. Al igual que en las anteriores actividades enzimáticas, Penicillium sp y Paecilomyces sp presentaron nuevamente los mayores valores de actividad enzimática con el tratamiento compuesto por carboximetilcelulosa y sulfato de amonio, presentándose diferencias entre tratamientos (p<0,005). Este comportamiento ha sido igualmente reportado en otro ensayo, en donde se encontró que el sulfato de amonio presento un efecto positivo en la producción de la enzima. (Maeda et al. 2013). Por otro lado la alta actividad enzimática obtenida nuevamente en este tratamiento podría explicar el que se hayan encontrado igualmente las actividades más altas de EG y CBH en este mismo tratamiento, ya que se sabe que la β‐ glucosidasa puede contribuir a la alta actividad celulasa debido a la formación de inductores de las demás enzimas del complejo celulolítico por esta enzima.(Jun et al. 2011) 70 Figura 4 4.3 Área bajo la curva (ABC) de laas unidades enzimáticaas celulolíticcas de Peniccillium sp. (A) yy Paecilomycces sp. (B). A B 71 4.3.5 Perfil de la actividad enzimática xilanolítica Dentro del proyecto de investigación se decidió evaluar una de las enzimas que componen el complejo necesario para degradar los compuestos hemiceluloliticos, la enzima xilanasa es de importancia en los procesos degradación debido a que el xilano es el polímero que se encuentra en mayor proporción en la hemicelulosa.(Oliveira et al. 2006). Se encontró que los microorganismos producen la enzima xilanasa, sin embargo Penicillium sp. produce los valores más altos de actividad. Así mismo el perfil de producción enzimático es diferente en cada caso, ya que mientras en el caso de Penicillium sp. las mayores actividades se obtienen en el cuarto día con valores entre 400 y 600 U/L , en el caso de Paecilomyces sp. éstas se obtienen entre el octavo y décimo día de cultivo con valores entre 45 y 80 U/L. Lo anterior sugiere que Penicillium lleva a cabo una eficiente degradación del xilano a diferencia de Paecilomyces y que este a su vez requiere un tiempo de adaptación más largo al sustrato para comenzar a producir las enzimas. Figura 4.4 Evaluación de la producción de xilanasas bajo diferentes fuentes de nitrógeno de Penicillium sp. (A) y Paecilomyces sp. (B) B A 100 700 Xilano Xilano + Ext. Lev. Xilano + (NH4 )2 SO4 600 Actividad xilanasa (U/L) Xilano Xilano + Ext. Lev. 80 Xilano + Ext. Lev. + (NH4 )2 SO4 Xilano + (NH ) SO 42 4 Xilano + Ext + (NH ) SO 42 4 500 400 300 60 40 200 20 100 00 2 4 6 Tiempo (dias) 8 00 10 2 4 6 Tiempo (dias) 8 10 12 72 4.3.6 A Actividad X Xilanasa En los reesultados de actividad xilanolitica se obtuvo que en el caaso de Peniicillium sp. n no hay una influencia direecta de las ffuentes de nitrógeno eevaluadas en e la producción enzim mática, sin emb bargo los trratamientoss con fuentte de nitróggeno son d diferentes aal tratamien nto en ausencia de la fueente, lo cuaal permite inferir que aunque no hay relació ón especificca con fuente o orgánica e inorgánica, si es necesaario la adición de nitrógeno al medio. En el estudio de (Ghaanem et al. 2000) donde se evalu uaron fuenttes de nitró ógeno tanto o orgánicas como inorgánicas, se en ncontró qu ue en todo os los caso os se obtuvieron mejjores activiidades empleando fuentee de nitróggeno a difeerencia del control sin n nitrógeno o, y se enccontró onio favorecció la produ ucción enzim mática especialmente quee la utilizaciión de sulfaato de amo xilanolittica Para Pa aecilomyces sp. se encontraron diferencias eentre los cu uatro tratam mientos sien ndo el tratamieento compu uesto por xiilano y extraacto de levvadura en el que se obttuvieron maayores unidadees enzimáticcas. En geeneral la ad dición de nitrógeno estimula la mineralizació m ón del carbono o. Lo anteriior podría explicar e por qué es necesaria la adición dee nitrógeno en el medio para la p producción enzimáticca, indistintamente d de la clase de nitrógeno adicionaada.(Henriksen & Brelaand 1999). Figura 4 4.5 Área bajo la curva (A ABC) de las unidades enzimáticas X Xilanasas dee Penicillium m sp. (A) y Pa aecilomyces sp. (B) A B 73 4.3.7 Influencia de la fuente de carbono y nitrógeno en la producción enzimática de Penicillium sp. y Paecilomyces sp. Se ha determinado que la expresión de celulasas y hemicelulasas está controlada principalmente por fuentes de carbono. En presencia de D‐glucosa la transcripción se encuentra reprimida, mientras que en ausencia de ésta y en la presencia de celulosa la transcripción es fuertemente inducida. Aún más, las enzimas celulolíticas pueden co‐ inducirse por diversos mono y disacáridos, como soforosa, xylobiosa, lactosa, D‐xylosa y L‐ sorbosa. Sin embargo existen diferentes mecanismos de regulación de las enzimas celulolíticas y algunos involucran procesos más globales. En general las dos cepas evaluadas producen los mejores valores enzimáticos celulolíticos cuando la fuente de carbono es carboximetil celulosa. Este comportamiento ha sido reportado antes con cepas de Penicillium persicinum y Penicillium brasilianum, en donde se ha encontrado que la EG y la CBH son mayormente influenciadas por la presencia de esta fuente (Jørgensen et al. 2005) Con respecto a la regulación de la expresión de las celulasas se sabe que la producción de enzimas basales representa uno de los elementos más importantes. Estas bajas cantidades de enzima hidrolizan la celulosa presente en el medio produciendo oligosacáridos solubles los cuales ingresan a la célula y generan una respuesta de inducción enzimática (Suto & Tomita 2001) . Existe evidencia de que la actividad celulolítica basal es un factor limitante en la inducción de la síntesis de celulasas en presencia de celulosa. (Sun et al. 2008b). Es posible que la diferencia de la respuesta en los microorganismos este dada por los niveles de expresión que generan estas concentraciones basales y así mismo la facilidad de degradación que tenga el sustrato empleado para inducir la producción enzimática (avicel y carboximetilcelulosa), ya que en este caso la celulosa amorfa podría estar siendo degradada más rápidamente y así generar una mejor respuesta enzimática. En el caso de la influencia de la fuente de nitrógeno sobre la producción enzimática se encontró que la combinación de sulfato de amonio y CMC como fuente de carbono presento una influencia positiva en la producción enzimática de las enzimas endoglucanasa, 74 celobiohidrolasa y β glucosidasa en el caso de Penicillium sp. presentándose diferencias entre tratamientos. Este mismo comportamiento se encontró en el caso de la producción de las enzimas celobiohidrolasa y β‐ glucosidasa por Paecilomyces sp. La influencia de la fuente de nitrógeno inorgánica en la producción enzimática puede estar relacionada con los hallazgos de Lockington y colaboradores (2002), quienes mediante estudios de producción enzimática celulolítica empleando diferentes fuentes de nitrógeno encontraron una alta actividad enzimática en condiciones de represión por amonio cuando se evaluó la mutación en el gen areA de Aspergillus nidulans el cual está relacionado con la expresión de genes que codifican para enzimas involucradas en el metabolismo del nitrógeno (Lockington et al. 2002). Igualmente en el estudio de Divanery & Hobley se encontró que la adición de fuente orgánica de nitrógeno registro los mismos valores de actividad que el tratamiento sin fuente de nitrógeno, lo anterior indica que el sulfato de amonio fue suficiente para incentivar el crecimiento celular y la producción enzimática celulolítica. Aunque no hay suficiente información sobre la regulación en géneros como Penicillium y Paecilomyces, por los resultados obtenidos se podría encontrar algún grado de similitud entre el comportamiento en la regulación de la producción de celulasas entre estos dos microorganismos y Aspergillus, ya que se evidencio una marcada influencia de el sulfato de amonio en la producción enzimática y por ende en la inducción de los genes que codifican para las enzimas del complejo celulolítico. Por otro lado es importante mencionar que en los suelos de donde se obtuvieron los aislamientos la forma predominante de nitrógeno es el amonio (Castilla et al. 2012) lo anterior podría sugerir que los microorganismos se encuentran habituados a esta fuente nutricional. En el caso de las xilanasas, como se mencionó anteriormente se encontraron diferencias en la respuesta enzimática entre los tratamientos y el control, lo cual sugiere que aunque existe una estimulación de la producción enzimática cuando hay presencia de fuente de nitrógeno, esta respuesta no está directamente relacionada con una de las dos fuentes 75 estudiadas, lo cual no implica que pueda responder diferente con otro tipo de fuentes nutricionales tanto orgánicas como inorgánicas. En general se sabe que sustratos a base de celulosa son capaces de incentivar la síntesis enzimática, sin embargo en el ensayo se encontró que la fuente de nitrógeno podría estar jugando un papel determinante en la síntesis. La utilización de diferentes fuentes de carbono y nitrógeno permitieron establecer que existe influencia en la producción enzimática máxima alcanzada y en los tiempos de cultivo en los que se obtiene. Es importante identificar la máxima producción enzimática en el complejo celulolítico debido a que este es un factor que puede influir en el sinergismo que presentan las enzimas cuando llevan a cabo la hidrolisis en cooperación. La máxima producción enzimática puede llevar a la saturación de los sitios de unión a sustrato, por este motivo establecer las perfiles de producción enzimática se hace necesario para determinar en qué punto podría ser más eficiente la degradación (Gan et al. 2003). Existen otros factores que así mismo tienen una influencia en la hidrólisis, entre los que se encuentra la concentración de sustrato, el grado de polimerización, la accesibilidad a la celulosa, la capacidad de adsorción, la cristalinidad, la concentración enzimática entre otros. Debido a que las actividades enzimáticas más altas de EG, CBH, β –glucosidasa y xilanasas, se obtuvieron con la cepa Penicillium sp., y que además los tiempo a los que se alcanza la máxima producción enzimática son menores con este aislamiento, se decide seleccionar este microorganismo para continuar con la última fase de la investigación que consiste en la evaluación del microorganismo sobre el residuo determinándose distintos parámetros de degradación del mismo. 76 Capítulo 5. Evaluación de la degradación del tamo por el aislamiento seleccionado 5.1 Introducción La reutilización de los residuos de cosecha como sustrato para compostaje, biofertilizantes, y enmiendas orgánicas es de interés en cultivos donde se requiere fertilización constante y donde los suelos necesitan ser reparados constantemente por la pérdida de materia orgánica que conlleva las actividades agrícolas. Se han estudiado tanto procesos de fermentación líquida como sólida buscando los mejores rendimientos en la obtención de diferente tipo de productos y así mismo una mayor degradación de los residuos. Sin embargo la fermentación solida ofrece diferentes ventajas, como lo son el contacto directo con el sustrato y la reducción de costos por concepto de utilización de energía en el proceso. Así mismo los procesos de degradación de sustratos complejos como los residuos agrícolas involucran diferentes factores que pueden afectar el proceso enzimático que se lleva a cabo; las características del sustrato, la temperatura, la cepa empleada para la degradación y el porcentaje de humedad son algunos de los más importantes. El residuo del cultivo de arroz, se encuentra compuesto por celulosa y hemicelulosa y en menor porcentaje por lignina. Esto hace que los microorganismos que se requieren para llevar acabo la degradación deban ser capaces de producir enzimas celulolíticas y hemicelulolíticas principalmente para hidrolizar los polímeros que constituyen la pared celular vegetal y así obtener fuentes fácilmente asimilables en el suelo. Penicillium sp. se aisló de suelos del cultivo (Gutierrez et al. 2012), y se esperaría que este habituado a las características propias del mismo, así mismo a través de la investigación se logró establecer que es capaz de producir las distintas enzimas del complejo celulolítico y hemicelulolítico. Sin embargo es de interés evaluar el comportamiento del microorganismos sobre el sustrato en condiciones de poca humedad y disminución de nutrientes fácilmente asimilables. Por tal razón se 77 planteó en el presente objetivo una fermentación solida empleando el residuo como sustrato, la cual se llevó a cabo por 20 días evaluándose las distintas enzimas del complejo celulolítico, así como la disminución de ciertos componentes del residuo que pueden sugerir que se está llevando a cabo una degradación eficiente del sustrato. 5.2 Materiales y Métodos 5.2.1 Evaluación de la actividad enzimática “in vitro” de los microorganismos sobre el sustrato Los ensayos de degradación directa por el microorganismo, se realizaron en fermentación sólida utilizando como sustrato el residuo. La fermentación se realizó en frascos con 10g de tamo, se adicionó el medio base de (Mandels & Weber 1969) (En gL‐1: KH2PO4, 0,3; MgSO4.7H2O, 0,83; CaCl2, 0,3; FeSO4.7H2O, 0,005; CaCl2, 0,02; MnSO4.H2O, 1,56 mg; ZnSO4.7H2O, 1,4 mg y Tween 80, 0,2 mL, pH 6,0), el ensayo se llevó a cabo con una humedad del 63%, el inoculo consistió en una suspensión de esporas con una concentración de 106 esporas/ml. Se incubó en condiciones estáticas a 28°C durante 20 días. 5.2.1.1 Obtención de los extractos enzimáticos Se retiraron 3 frascos cada cinco días y se adicionaron 90 ml de buffer acetato pH 5.0, 50 mM, para realizar la extracción enzimática, se mantuvo en agitación por 5 horas, posteriormente se centrifugó a 10.000 g por 20 min y a 8.000 g por 15 min para separar la biomasa y el sustrato del sobrenadante. 78 5.2.1.2 Actividad Enzimática Las actividades enzimáticas fueron determinadas a partir del extracto obtenido anteriormente. Se empleó un equipo Evolution 60 UV‐VIS Thermo Scientific Co (Waltham, MA, USA). 5.2.1.3 Actividad endo‐1,4‐β‐glucanasa Se realizó mezclando 500 µL del extracto con 500 µL de CMC de baja viscosidad (C5678) (2 % p/v) en buffer citrato 50 mM (pH 5.0) a 40°C por 60 minutos. (Quevedo 2011). La cantidad de azúcares reductores en términos de equivalentes de glucosa, se determinaron por la técnica de DNS (Miller 1959). Una unidad de actividad glucanasa (UG) es la cantidad de enzima que libera 1 µmol de azúcares reductores por minuto a las condiciones del ensayo. 5.2.1.4 Actividad exo‐1,4‐β‐glucanasa. Será evaluó usando como sustrato p‐nitrofenil‐β‐D‐celobiosido (PNPC, Sigma). La reacción contenía 320 µL de PNPC 5 mM en buffer acetato de sodio 50 mM (pH 5.0) y 100 µL de muestra. Esta mezcla se incubó a 40ºC por 60 minutos. La reacción se detuvo adicionando 250 µL de carbonato de sodio 0.5 M. La absorbancia se leyó a 400 nm. (Quevedo 2011). La actividad enzimática se calculó usando la curva patrón de p‐nitrofenol. Una unidad de actividad exoglucanasa o celobiohidrolasa (UC) se define como la cantidad de enzima que libera 1 µmol de p‐nitrofenol por minuto a las condiciones del ensayo 5.2.1.5 Actividad de 1‐4‐β‐glucosidasa. Se cuantificó usando como sustrato p‐nitrofenil‐β‐D‐glucosido (PNPG, SIGMA), siguiendo el mismo método que para la actividad celobiohidrolasa.. 79 5.2.1.6 Actividad xilanasa. La actividad xilanasa se determinó mezclando 500 µL de extracto con 500 µL de xilano de árbol de haya (SIGMA) (2%p/v) en buffer citrato 50 mM (pH 5.0) a 40°C por 60 minutos. (Driss et al. 2011). La cantidad de azúcares reductores liberados se determinó por la técnica de DNS con xilosa como estándar. Una unidad de actividad xilanasa (UX) se define como la cantidad de enzima que libera 1 µmol de azúcares reductores por minuto. 5.2.2 Determinación de lignina celulosa y hemicelulosa del residuo La determinación fue realizada por el Laboratorio de Nutrición Animal de la Universidad Nacional de Colombia. Se realizó con el sustrato antes y después de la fermentación sólida, empleando en cada caso la misma cantidad de sustrato (60 g). Se determinó la fibra en detergente neutro y fibra en detergente acido, así como el porcentaje de lignina, celulosa y hemicelulosa según la metodología de (Soest et al. 1991). 5.2.3 Análisis cualitativo de la degradación Para realizar un registro cualitativo de la degradación del sustrato se observaron mediante microscopia electrónica de barrido (MEB) a 500 y 1000 X muestras del residuo al finalizar los 20 días de fermentación sólida y del control abiótico. Las determinaciones se realizaron en el Laboratorio de Microscopia Electrónica de la Universidad de los Andes, donde se cuenta con un microscopio JOEL, modelo JSM 6490‐LV, las muestras se observaron en platina de enfriamiento. 5.3 Resultados y discusión Como objetivo final del proyecto se planteó la evaluación de la degradación del residuo en fermentación sólida por el microorganismo seleccionado (Penicillium sp.) haciendo un seguimiento de la producción enzimática, la disminución del contenido de celulosa y 80 hemicelulosa y la confirmación cualitativa de la colonización y la degradación del sustrato por MEB. 5.3.1 Actividad Enzimática En la Figura 5.1 se muestran los resultados de actividad de las enzimas tanto celulolíticas como xilanasa, se observó que esta enzima requerida para degradar la hemicelulosa, tuvo valores similares en los primeros 10 días de cultivo y posteriormente presentó un aumento del doble de la actividad registrada anteriormente, así mismo las enzimas EG y β – glucosidasa presentaron un aumento en la actividad en este mismo tiempo. Lo anterior sería de esperarse teniendo en cuenta las características de composición de sustratos complejos como el tamo, en la cual la hemicelulosa se encuentra recubriendo la celulosa y por este motivo es esencial degradar primero este heteropolimero para así poder acceder a la celulosa. Los altos valores enzimáticos registrados en el cultivo estarían permitiendo la degradación de la hemicelulosa y de esta forma la celulosa amorfa y cristalina se hace más accesible para el microorganismo. Por esta razón los valores más altos de las demás enzimas se encuentran a partir de este tiempo de cultivo, ya que al estar en contacto con el sustrato se produce una inducción de la producción enzimática, además de otro tipo de azúcares que pueden producirse en la hidrólisis de la hemicelulosa y que pueden estimular la producción de celulosas. Por otro lado la CBH presenta mayor actividad al finalizar la fermentación probablemente porque la celulosa cristalina podría estar más disponible en el residuo en este tiempo y así mismo por la existencia de extremos reductores producto de la acción de la EG. Se ha demostrado que existe una correlación entre la superficie accesible para las enzimas celulolíticas y la degradación de los materiales lignocelulolíticos. Cuando el área de contacto de la enzima aumenta, se hace más factible la acción hidrolitica sobre el sustrato, este acceso a esta área puede estar relacionado con el grado de cristalinidad, la proporción de lignina, hemicelulosa o por todos los factores anteriores (Bansal et al. 2009) 81 Figura 5.1 Actividad enzimática en la fermentación sólida. Endoglucanasa (EG), Xilanasa (XS), β –glucosidasa (BGS), Celobiohidrolasa (CBH). 40 20 0 Actividad EG ( U / g ) Actividad XS ( U / g ) 60 140 800 120 600 8 400 6 4 200 2 100 80 60 40 Actividad CBH ( U / g ) 10 80 EG XS BGS CBH 12 100 ( Actividad BGS ( U / g ) 20 0 0 0 5 10 15 0 20 Tiempo (dias) A partir de los anteriores ensayos se determinó la capacidad que tiene el asilamiento Penicillium sp. MF 3‐64 para producir las distintas enzimas necesarias para la degradación de la celulosa y así mismo es posible que sintetice diferentes enzimas capaces para degradar la hemicelulosa, probándose la producción de la xilanasa en fuentes nutricionales simples. En la fermentación solida se logró establecer del mismo modo que el sustrato tiene una influencia positiva en la inducción de la actividad enzimática. Se obtuvieron 747,64 U/g de β glucosidasa, 138,38 Celobiohidrolasa U/g, 10,01 U/g Endoglucanasa y 88,47 U/g xilanasas, estos valores de actividad enzimática al ser comparados con los obtenidos frente a otros residuos empleando otros microorganismos celulolíticos, son de interés, debido a que en el caso de la enzima β ‐glucosidasa y Celobiohidrolasa se presentan actividades enzimáticas mayores que las reportadas por microorganismos empleados comúnmente para degradar este tipo de residuos (Tabla 5.1) . En el caso de la EG los valores de actividad son similares a los encontrados en residuos de arroz como paja y cascarilla. Sin embargo se presentaron diferencias importantes en la magnitud de la respuesta de la enzima xilanasa comparada con estos microorganismos. Lo anterior podría estar relacionado con diferentes 82 factoress, como el contenido d de celulosa y hemicelu ulosa que tengan los ssustratos y si los residuoss tuvieron o o no un pre tratamiento ya sea químico o físicco, entre ottros (Deswal et al. 2011). P Por otro lado es importtante mencionar que een el estudio o no se evalluaron xilossidasas ni otrass clases de hemicelulassas, las cuaales podrían n estar aportando unaa degradació ón del residuo igualmentte y que permitirían p la alta prroducción eenzimática EG, CBH y y β – dasa que se registra. glucosid Tabla 5..1 Actividad des enzimáticas obtenid das por difeerentes micrroorganismo os. EGa Unid dades enzimátticas EG /g d de sustrato,, BGSb β –gglucosidasa//g de sustratto, CBHC CBH/g de susttrato, XSd Xilan nasas/g de ssustrato . La actividad enzimáática obtenida con resp pecto a la β β‐ glucosidaasa es de im mportancia d debido al papell que desem mpeña esta eenzima en la hidrólisiss de la celo obiosa en do os monómeros de glucosa disminuyeendo la inh hibición ejeercida por este comp puesto. Iggualmente se ha debido estableccido una rellación entree la regulacción de los ggenes de celulasas y esta enzima d a la pro oducción dee inductorees de los sisstemas celu ulolíticos en n hongos co omo Trichod derma (Bedford & Partridgge 2001). 83 Otro aspecto de interés en la alta producción enzimática producida por Penicillium sp. se centra en la utilización de esta cepa para alcanzar una degradación más eficiente del residuo, ya que en el caso de Trichoderma, microorganismo ampliamente empleado para la producción de enzimas celulolíticas, la deficiencia en la producción de β glucosidasa disminuye el desempeño del microorganismo durante la hidrolisis de materiales lignocelulósicos. Aunque se encontró actividad xilanasa más baja comparada con otros microorganismos (Tabla 5.2), es importante resaltar que Penicillium sp. MF 3‐64 produce todas las enzimas del complejo y que la enzima β glucosidasa presenta igualmente valores más altos que los encontrados en la literatura utilizando este tipo de residuos pretratados. Igualmente es de importancia establecer el comportamiento de Penicillium sp. frente a sustratos complejos, ya que se han descrito especies de este género con la habilidad de producir altos valores de celulasas y hemicelulasas, pero no se conoce mucho sobre la regulación y producción de estas enzimas comparado con Trichoderma y Aspergillus (Jørgensen et al. 2005). Adicionalmente es importante la determinación de otros parámetros dentro de la fermentación producto de la acción enzimática sobre el sustrato. Los azúcares solubles en la muestra pueden ser un indicativo de la actividad degradativa del residuo (Liu & R 2000). En la (Figura 5.2) se muestran los azucares liberados en el medio y las actividades EG y Xilanasa. Se observa que a medida que hay puntos de actividad enzimática altos, existe un estancamiento en la concentración de azucares, lo cual podría sugerir que a medida que hay una actividad enzimática creciente, el microorganismo consume los azucares liberados por la degradación del sustrato, sin embargo es importante mencionar que la tendencia en la producción de azucares es a aumentar en el tiempo, lo cual podría deberse a la rápida actividad enzimática que produce el microorganismo, y es de interés porque podría sugerir que se está llevando a cabo efectivamente la degradación del residuo, lo cual daría paso a la obtención de fuentes carbonadas sencillas de fácil asimilación que pueden ser de utilidad al momento de incorporar el residuo en el suelo. Por otro lado, el aumento en los azucares del día 15 al 20 podría ser producto de la producción de la CBH y EG, las cuales presentan niveles máximos de producción en este lapso de tiempo. 84 Figura 5.2 Actividad enzimática en la fermentación sólida. Endoglucanasa (EG), Xilanasa (XS), y Azucares reductores producidos 8 60 6 40 4 20 2 0 0 0 5 10 15 0,5 0,4 0,3 0,2 0,1 Azucares Reductores ( g / L ) 10 Actividad EG ( U / g ) 12 80 Actividad XS ( U / g ) XS EG Azucares Reductores 100 0,0 20 Tiempo (dias) 5.3.2 Determinación de Celulosa y Hemicelulosa A demás de la determinación enzimática, existen otros parámetros que pueden dar información de la degradación que está ocurriendo en el sustrato. La determinación de componentes estructurales de la pared celular vegetal aporta información sobre el proceso de hidrolisis que se lleva a cabo en la fermentación sólida. Después de los 20 días de fermentación solida se obtuvo una disminución de 4,6 % en el contenido de celulosa y 4,4% en el contenido de hemicelulosa (Tabla 5.2). Estos porcentajes de reducción son mayores que los obtenidos con otros tratamientos como por ejemplo el químico, donde se consiguen reducciones de tan solo del 3.2% en la cantidad de hemicelulosa en el residuo de arroz (Rahnama et al. 2013). Sin embargo se encontró un aumento en el porcentaje de lignina pasando de 3 a 5,9 %. Este resultado puede estar relacionado con diferentes factores como el tratamiento biológico en sí, ya que es posible que por la acción degradativa del microorganismo sobre los otros componentes del residuo, la lignina este más expuesta a ser detectada. Así mismo es probable que la diferencia se deba a la técnica empleada. En tratamientos químicos llevados a cabo sobre residuos agrícolas, en la determinación de los 85 com mponentes d de la pared celular, sse encuenttra un aum mento en ccompuestoss como la celu ulosa despuéés del tratamiento, lo ccual indica que despuéés del tratam miento el co ompuesto qued do más expuesto a la d detección. Aunque no eexiste un índice de referencia del porceentaje de disminución de los mponentes lignocelulóssicos que deetermine cu uando un rresiduo ha sido transfo ormado o com degrradado, la d determinaciión de los m mismos en eel proceso junto con lo os demás paarámetros evaluados podrría corroborrar que las enzimas prroducidas po or Penicilliu um sp. efecttivamente estáán llevando a cabo la hidrolisis d del residuo,, lo cual ess confirmad do igualmen nte por la pressencia de azzucares reductores en el extracto enzimático o . Por otro o lado sería necesario máss tiempo de fermentación para ob btener mayo ores porcen ntajes de reducción en estos dos polím meros. Tablla 5.2 Porceentaje de Ceelulosa y Heemicelulosa en el residu uo antes y d después de la ferm mentación só ólida. ón del susttrato por M MEB 5.3..3 Análisiss cualitativvo de la degradació Al evaluar la deegradación del residuo o se debe teener en cueenta diferen ntes parám metros. Las prop piedades esstructurales del sustratto incluyend do la proporrción de la composición química así ccomo las características de las fibrras tienen u un efecto siggnificativo een la hidrolisis. En un 86 estudio llevado a cabo sobre varias fracciones de residuo de arroz diferentes morfologicamente se encontraron diferencias en la hidrolisis y en la producción de azucares reductores. Generalmente la fracción con el promedio más bajo de tamaño de fibras, menor contenido de células del parénquima, y de la epidermis, menor porcentaje de lignina, es hidrolizado más rápidamente. El tamo de arroz es un residuo heterogéneo, sin embargo presenta bajo contenido de lignina comparado con otros residuos. Esto facilita su degradación por los microorganismos celulolíticos (Jin & Chen 2006)(Liu & R 2000). Al contrarrestar las barreras que tiene la planta, el microorganismo debe como primer paso colonizar el sustrato, la adhesión al mismo hace que pueda diseminarse de una partícula a otra. Algunos hongos son capaces de producir estructuras hifales que les proporcionan mayor penetración y adhesión al sustrato, así mismo son capaces de producir polisacáridos para tal fin. La accesibilidad al sustrato esta inicialmente dada por las hendiduras en la pared celular vegetal. En el caso del tamo se observa que presenta una estructura irregular y porosa que pueden facilitar la entrada del microrganismo. En la Figura 5.3 A se aprecia la efectiva colonización cuando se observó un corte longitudinal del sustrato a los 20 días de fermentación sólida. En la imagen se observan los conidios producidos por el microorganismo y así mismo en la Figura 5.3 B se detalla el entramado hifal que recubre el sustrato. Por otro lado la especificidad del microorganismo es crítica en la fermentación solida debido a la complejidad del sustrato la cual hace necesaria la producción de un complejo enzimático específico para su degradación. La composición del complejo enzimático depende tanto del microorganismos como del sustrato (Tengerdy & Szakacs 2003). En la Figura 5.3 C,D se evidencia el cambio de morfología del residuo con respecto al control abiótico. Se aprecia la perdida de la estructura de los haces vasculares de la planta y una desintegración de las fibras que componen las estructuras de las paredes celulares. Lo anterior producto de la continua degradación del residuo por el microorganismo. 87 Figu ura 5.3 Microscopia electrónica de barrido (MEB) de las m muestras al finalizar la ferm mentación so B), (C). Contrrol abiótico (D) olida (A), (B C A D B os resultado os de las evvaluaciones anteriores se evidencio la degrad dación del De aacuerdo a lo resid duo llevadaa a cabo por Penicilllium sp. LLa producciión de las diferentess enzimas celu ulolíticas y de las xilanasas perm mitió establlecer el papel del ressiduo como o sustrato indu uctor de la actividad. LLa determin nación de lo os componentes estru ucturales dee la pared celu ular del residuo eviden nciaron disminución d del porcentaje de celu ulosa y hem micelulosa 88 después de la fermentación sólida, aunque se registró un aumento en la concentración de lignina, y el análisis del residuo por MEB confirmo la colonización del sustrato y el cambio estructural del mismo. La evaluación del crecimiento y establecimiento del microorganismo sobre el sustrato es clave si se pretende llevar a cabo estudios que permitan realizar determinaciones de degradación en campo. Las condiciones de la fermentación solida fueron discretas en el sentido que no se adiciono ninguna fuente sencilla de carbono como sustrato inductor de crecimiento, lo cual haría que el microorganismos pudiera establecerse más rápidamente en el sustrato. Lo anterior permite establecer una visión del comportamiento del aislamiento frente a condiciones que se podrían presentar en una degradación in situ. Con respecto a esto, los suelos de donde fue obtenido el aislamiento presenta alta tendencia a la pérdida de materia orgánica debido a las constantes actividades agrícolas y así mismo presentan valores de pH ligeramente ácidos con una retención de agua moderada (Castilla et al. 2012)Lo anterior es de interés ya que el microorganismo demostró un establecimiento sobre el sustrato en condiciones de baja humedad y con el residuo como única fuente de carbono y nitrógeno. Por otro lado el proceso de fermentación solida ofrece diferentes ventajas como el poco requerimiento de energía, además puede solucionar los problemas de disposición de los residuos sólidos que se presentan en el cultivo del arroz. Generalmente se ha aceptado que en la fermentación solida se obtienen rendimientos más altos de actividad enzimática, sin embargo no hay una escala o método que permita comparar la productividad en fermentación sólida y liquida. Sin embargo se acepta que estos altos rendimiento se obtienen debido a que el microorganismo se encuentra directamente sobre el sustrato natural y esto probablemente aumenta la inducción de la producción enzimática y por ende la degradación del mismo (Ghanem et al. 2000). 89 90 6. Conclusiones y Recomendaciones 6.1 Conclusiones Las actividades agrícolas producen diferentes tipos de residuos, en algunos casos estos son aprovechables en diferentes procesos, sin embargo los costos de transporte, disposición y tratamiento dificultan su uso. En el caso del tamo de arroz, la producción del residuo es igual o mayor que el producto de interés, esto hace que la acumulación sea uno de los inconvenientes que se presenta en estos cultivos lo cual conlleva a diferentes tipos de problemas tanto sanitarios como de disposición. En la presente investigación se realizó la selección de un aislamiento celulolítico, el cual presenta actividades enzimáticas de interés para la degradación del residuo agrícola compuesto principalmente por celulosa y hemicelulosa. Lo anterior resulta de interés debido a la necesidad de emplear estos residuos como fuente adicional de materia orgánica biotransformada que permita aumentar el contenido de nutrientes en el suelo. De acuerdo a las metodologías de selección de los aislamientos Penicillium sp. MF 3‐64 fue el microorganismo que presento mejor desempeño en la degradación de celulosa de acuerdo a la metodología cualitativa por medición de halos de hidrolisis de celulosa en agar, en la cuantificación de celulasas totales a partir de la fermentación sumergida en caldo tamo y además presento mayor producción de CO2 en el ensayo de respirometría empleando le residuo como sustrato. Adicionalmente el aislamiento Paecilomyces sp. MF 4‐ 71 presentó igualmente actividad celulolítica y producción de CO2. Aunque las tres técnicas son diferentes entre si en cuanto a las variables de respuesta y a las condiciones del ensayo, estas permitieron la selección de los dos microorganismos para continuar con la evaluación del complejo enzimático. 91 En la evaluación de la producción enzimática celulolítica y hemicelulolítica bajo diferentes fuentes de carbono y nitrógeno se estableció que los microorganismos presentan diferencias en la producción enzimática siendo Penicillium sp. el microorganismos que presenta mayor actividad en todas las enzimas evaluadas, en el caso de la CBH se obtuvieron incluso valores del doble de actividad, y en el caso de la β –glucosidasa inclusive la diferencia es de 2000 Unidades enzimáticas. Así mismo se estableció que la fuente de celulosa amorfa presenta una influencia positiva en la inducción de la producción enzimática de todas las enzimas y adicionalmente la fuente de nitrógeno inorgánica demostró ser igualmente importante en la producción enzimática. Es así que el tratamiento compuesto por esta combinación presento en la mayoría de actividades enzimáticas evaluadas las mayores unidades. En el caso de las xilanasas se evidencia una influencia de la fuente de nitrógeno en la producción enzimática pero no se estableció una relación directa del tipo de fuente y la inducción de la producción. Los anteriores resultados son similares en los dos microorganismos. En la evaluación del aislamiento seleccionado (Penicicillium sp.) sobre el sustrato se encontró disminución de la proporción de celulosa y hemicelulosa en el residuo después de la fermentación sólida, por otro lado se encontró alta producción enzimática CBH y β – glucosidasa, con producciones que superan incluso en 600 unidades al ser comparados con otros estudios en residuos similares. Así mismo se encontraron valore de actividad EG de interés. Por otra parte se encontró producción de azucares reductores, lo cual permite establecer que se está llevando a cabo una degradación del residuo. Adicionalmente se estableció la efectiva colonización del sustrato y la desintegración de la estructura del mismo. Los anteriores hallazgos permiten establecer que el aislamiento Penicillium sp. MF 3‐64 es un microorganismo promisorio en la degradación eficiente del residuo del cultivo, degradando los compuestos que se encuentran en mayor proporción en la planta, permitiendo avanzar hacia la siguiente etapa del proyecto que busca evaluar consorcios de 92 microorganismos como alternativa biológica para la degradación y la reincorporación del tamo de arroz. 6.2 Recomendaciones En la metodología indirecta de selección de las cepas llevada a cabo por la técnica de respirometría se observó que la tendencia de producción de CO2 es a continuar aumentando, por lo tanto el llevar el ensayo por más tiempo que el evaluado permitiría establecer los tiempos de degradación máximos En la evaluación de diferentes fuentes de carbono y nitrógeno sobre la expresión de celulasas y hemicelulosa seria de interés evaluar otras fuentes que permitieran conocer mejor el comportamiento del aislamiento seleccionado frente a diferentes condiciones nutricionales. Así mismo es importante establecer la producción de otras enzimas necesarias para llevar a cabo la degradación de la hemicelulosa ya que en el estudio se decidió evaluar la xilanasa debido a la alta presencia de xilano en el heteropolimero, sin embargo este puede estar conformado por otro tipo de polímeros haciéndose necesario determinar otras actividades enzimáticas. En algunos microorganismos se ha encontrado la posible influencia de componentes de la hemicelulosa en el proceso de inducción de celulasas y viceversa, por tal razón la evaluación de enzimas celulolíticas empleando xilano como fuente de carbono es recomendable. Finalmente debido a los importantes resultados de colonización, producción enzimática y degradación del residuo, la evaluación del microorganismo en condiciones similares a las del cultivo sería recomendable. En algunos estudios se llega incluso a evaluar mezclas de suelo proveniente del cultivo y el residuo a degradar para determinar el establecimiento del aislamiento. 93 7. BIBLIOGRAFIA Abdel‐Sater, M.. & El‐Said, a. H.., 2001. Xylan‐decomposing fungi and xylanolytic activity in agricultural and industrial wastes. International Biodeterioration & Biodegradation, 47(1), pp.15–21. Available at: http://linkinghub.elsevier.com/retrieve/pii/S096483050000113X. Abe, K. & Yano, H., 2009. Comparison of the characteristics of cellulose microfibril aggregates of wood, rice straw and potato tuber. Cellulose, 16(6), pp.1017–1023. Available at: http://link.springer.com/10.1007/s10570‐009‐9334‐9. Adsul, M.G. et al., 2007. Strain improvement of Penicillium janthinellum NCIM 1171 for increased cellulase production. Bioresource technology, 98(7), pp.1467–73. Available at: http://www.ncbi.nlm.nih.gov/pubmed/17097876]. Ahamed, A. & Vermette, P., 2008. Culture‐based strategies to enhance cellulase enzyme production from Trichoderma reesei RUT‐C30 in bioreactor culture conditions. Biochemical Engineering Journal, 40(3), pp.399–407. Available at: http://linkinghub.elsevier.com/retrieve/pii/S1369703X07004834. Andersson, S., Serimaa, R. & Paakkari, T., 2003. Crystallinity of wood and the size of cellulose crystallites in Norway spruce ( Picea abies ). , pp.531–537. Bansal, N. et al., 2012. Production of cellulases from Aspergillus niger NS‐2 in solid state fermentation on agricultural and kitchen waste residues. Waste management (New York, N.Y.), 32(7), pp.1341–6. Available at: http://www.ncbi.nlm.nih.gov/pubmed/22503148. Bansal, P. et al., 2009. Modeling cellulase kinetics on lignocellulosic substrates. Biotechnology advances, 27(6), pp.833–48. Available at: http://www.ncbi.nlm.nih.gov/pubmed/19577626. Bayera, E.A., Moraga, E. & Lamed, R., 1994. The cellulosome A treasure‐trove for biotechnology. Trends in Biotechnology, 12(9), pp.379–386. Bedford, M. & Partridge, G., 2001. Enzymes in farm animal nutrition, London: CABI Publishing. Bellon‐Maurel, V., Orliac, O. & Christen, P., 2003. Sensors and measurements in solid state fermentation: a review. Process Biochemistry, 38(6), pp.881–896. Available at: http://linkinghub.elsevier.com/retrieve/pii/S0032959202000936. 94 Biomass Users Network (BUN‐CA), 2002. Manuales sobre energía renovable: Biomasa. , p.42. Available at: http://www.bun‐ca.org/publicaciones/BIOMASA.pdf. Camassola, M. & Dillon, A.J.P., 2009. Biological pretreatment of sugar cane bagasse for the production of cellulases and xylanases by Penicillium echinulatum. Industrial Crops and Products, 29(2‐3), pp.642–647. Available at: http://linkinghub.elsevier.com/retrieve/pii/S0926669008001957. Camassola, M. & J.P. Dillon, A., 2012. Cellulase Determination: Modifications to Make the Filter Paper Assay Easy, Fast, Practical and Efficient. Journal of Analytical & Bioanalytical Techniques, 01(S1), pp.1–4. Available at: http://www.omicsonline.org/scientific‐reports/srep125.digital/srep125.html. Carrillo Navarrete, F., 2002. Caracterización estructural de fibras lyocell y su comportamiento frente a procesos de degradación. Universidad Politecnica de Cataluña. Castilla, L. et al., 2012. Evaluacion de la fertilidad en suelos de las zonas arroceras de Tolima y Meta. In D. Uribe & L. M. Melgarejo, eds. Ecologia de microorganismos rizosfericos asociados a cultivos de arroz de Tolima y Meta. Bogota: Universidad Nacional de Colombia, pp. 33–53. Castilla, L., 2013. Revista Arroz, Bogota. Available at: http://www.fedearroz.com.co/revistanew/arroz502.pdf. Chapla, D. et al., 2010. Utilization of agro‐industrial waste for xylanase production by Aspergillus foetidus MTCC 4898 under solid state fermentation and its application in saccharification. Biochemical Engineering Journal, 49(3), pp.361–369. Available at: http://linkinghub.elsevier.com/retrieve/pii/S1369703X10000392. Chávez, R., Bull, P. & Eyzaguirre, J., 2006. The xylanolytic enzyme system from the genus Penicillium. Journal of biotechnology, 123(4), pp.413–33. Available at: http://www.ncbi.nlm.nih.gov/pubmed/16569456. Chen, M. et al., 2010. Isolation and characterization of a β‐glucosidase from Penicillium decumbens and improving hydrolysis of corncob residue by using it as cellulase supplementation. Enzyme and Microbial Technology, 46(6), pp.444–449. Available at: http://linkinghub.elsevier.com/retrieve/pii/S0141022910000207. Degiovanni, V., Martinez, C. & Motta, F., 2010. Produccion Eco‐Eficiente del arroz en America Latina Tomo I., CIAT. Deswal, D., Khasa, Y.P. & Kuhad, R.C., 2011. Optimization of cellulase production by a brown rot fungus Fomitopsis sp. RCK2010 under solid state fermentation. Bioresource 95 technology, 102(10), pp.6065–72. Available at: http://www.ncbi.nlm.nih.gov/pubmed/21470856 [Accessed March 21, 2012]. Driss, D. et al., 2011. Purification and properties of an extracellular acidophilic endo‐1,4‐β‐ xylanase, naturally deleted in the “thumb”, from Penicillium occitanis Pol6. Process Biochemistry, 46(6), pp.1299–1306. Available at: http://linkinghub.elsevier.com/retrieve/pii/S1359511311000924. Ee, J.H. van, 1990. Enzymes and Detergency. Surfactant Science series, Espinal, M., 2005. La agroindustria molinera de arroz en Colombia, Fan, M. et al., 2005. Crop Yields, Internal Nutrient Efficiency, and Changes in Soil Properties in Rice–Wheat Rotations Under Non‐Flooded Mulching Cultivation. Plant and Soil, 277(1‐2), pp.265–276. Available at: http://link.springer.com/10.1007/s11104‐005‐ 7459‐7 [Accessed June 11, 2013]. FEDEARROZ, 2000. Guia ambiental del arroz. , pp.1–34. Available at: http://www.minambiente.gov.co/documentos/guia_ambiental_subsector_arrocero.pd f. Gan, Q., Allen, S.. & Taylor, G., 2003. Kinetic dynamics in heterogeneous enzymatic hydrolysis of cellulose: an overview, an experimental study and mathematical modelling. Process Biochemistry, 38(7), pp.1003–1018. Available at: http://linkinghub.elsevier.com/retrieve/pii/S0032959202002200. Ghanem, N.B., Yusef, H.H. & Mahrouse, H.K., 2000. Production of Aspergillus terreus xylanase in solid‐state cultures: application of the Plackett–Burman experimental design to evaluate nutritional requirements. Bioresource Technology, 73(2), pp.113– 121. Available at: http://linkinghub.elsevier.com/retrieve/pii/S0960852499001558. GHOSE, T.K., 1987. MEASUREMENT OF CELLULASE ACTIVITIES. Chemistry, Applied, 59(2). Gomathi, D. et al., 2012. Submerged fermentation of wheat bran by Aspergillus flavus for production and characterization of carboxy methyl cellulase. Asian Pacific Journal of Tropical Biomedicine, 2(1), pp.S67–S73. Available at: http://linkinghub.elsevier.com/retrieve/pii/S2221169112601324 [Accessed November 9, 2012]. Gottschalk, L.M.F., Oliveira, R.A. & Bon, E.P.D.S., 2010. Cellulases, xylanases, β‐glucosidase and ferulic acid esterase produced by Trichoderma and Aspergillus act synergistically in the hydrolysis of sugarcane bagasse. Biochemical Engineering Journal, 51(1‐2), pp.72– 78. Available at: http://linkinghub.elsevier.com/retrieve/pii/S1369703X10001348. 96 Gutierrez, I. et al., 2012. Estimacion de poblaciones de microorganismos ligninoliticos y celuloliticos, y actividad beta‐glucosidasa en agrosistemas de arroz. In D. Uribe & L. M. Melgarejo, eds. Ecologia de microorganismos rizosfericos asociados a cultivos de arroz de Tolima y Meta. pp. 89–109. Gutierrez‐correa, M. et al., 1999. blO tSOIJ Ct II Mixed culture solid substrate fermentation of Trichoderma reesei with Aspergillus niger on sugar cane bagasse. , 68, pp.173–178. Hendriks, a T.W.M. & Zeeman, G., 2009. Pretreatments to enhance the digestibility of lignocellulosic biomass. Bioresource technology, 100(1), pp.10–8. Available at: http://www.ncbi.nlm.nih.gov/pubmed/18599291 [Accessed May 23, 2013]. Henriksen, T.M. & Breland, T.A., 1999. Nitrogen availability e ects on carbon mineralization , fungal and bacterial growth , and enzyme activities during decomposition of wheat straw in soil. , 31. Henry, B. & Oswaldo, S., 2005. El Entorno Internacional del Sector Arrocero Centroamericano, Jia, H. et al., 2012. Engineering a thermostable β‐1,3‐1,4‐glucanase from Paecilomyces thermophila to improve catalytic efficiency at acidic pH. Journal of biotechnology, 159(1‐2), pp.50–5. Available at: http://www.ncbi.nlm.nih.gov/pubmed/22370539. Jiang, Z.Q. et al., 2005. Characterization of a novel, ultra‐large xylanolytic complex (xylanosome) from Streptomyces olivaceoviridis E‐86. Enzyme and Microbial Technology, 36(7), pp.923–929. Available at: http://linkinghub.elsevier.com/retrieve/pii/S0141022905000256. Jin, S. & Chen, H., 2006. Structural properties and enzymatic hydrolysis of rice straw. Process Biochemistry, 41(6), pp.1261–1264. Available at: http://linkinghub.elsevier.com/retrieve/pii/S1359511305005325. Jørgensen, H. et al., 2004. Growth and enzyme production by three Penicillium species on monosaccharides. Journal of biotechnology, 109(3), pp.295–9. Available at: http://www.ncbi.nlm.nih.gov/pubmed/15066767. Jørgensen, H. et al., 2005. Production of cellulases and hemicellulases by three Penicillium species: effect of substrate and evaluation of cellulase adsorption by capillary electrophoresis. Enzyme and Microbial Technology, 36(1), pp.42–48. Available at: http://linkinghub.elsevier.com/retrieve/pii/S014102290400273X [Accessed June 11, 2013]. 97 Juhász, T. et al., 2005. Characterization of cellulases and hemicellulases produced by Trichoderma reesei on various carbon sources. Process Biochemistry, 40(11), pp.3519– 3525. Available at: http://linkinghub.elsevier.com/retrieve/pii/S1359511305002035. Jun, H., Kieselbach, T. & Jönsson, L.J., 2011. Enzyme production by filamentous fungi : analysis of the secretome of Trichoderma reesei grown on unconventional carbon source. , pp.1–10. Kamide, K., 2005. Cellulose and Cellulose Derivatives E. B.V, ed., Kaur, J., Chadha, B.S. & Saini, H.S., 2006. Regulation of cellulase production in two thermophilic fungi Melanocarpus sp. MTCC 3922 and Scytalidium thermophilum MTCC 4520. Enzyme and Microbial Technology, 38(7), pp.931–936. Available at: http://linkinghub.elsevier.com/retrieve/pii/S014102290500373X [Accessed June 10, 2013]. Kluczek‐Turpeinen, B. et al., 2007. Degradation and enzymatic activities of three Paecilomyces inflatus strains grown on diverse lignocellulosic substrates. International Biodeterioration & Biodegradation, 59(4), pp.283–291. Available at: http://linkinghub.elsevier.com/retrieve/pii/S0964830506001533. Latifian, M., Hamidi‐Esfahani, Z. & Barzegar, M., 2007. Evaluation of culture conditions for cellulase production by two Trichoderma reesei mutants under solid‐state fermentation conditions. Bioresource technology, 98(18), pp.3634–7. Available at: http://www.ncbi.nlm.nih.gov/pubmed/17207619. Li, L. et al., 2006. Purification and characterization of a thermostable cellulase‐free xylanase from the newly isolated Paecilomyces themophila. Enzyme and Microbial Technology, 38(6), pp.780–787. Available at: http://linkinghub.elsevier.com/retrieve/pii/S0141022905003571. Li, X. et al., 2010. Purification and characterization of a cellulase‐free, thermostable xylanase from Streptomyces rameus L2001 and its biobleaching effect on wheat straw pulp. Biochemical Engineering Journal, 52(1), pp.71–78. Available at: http://linkinghub.elsevier.com/retrieve/pii/S1369703X10001956 [Accessed June 4, 2013]. Li, Y. et al., 2007. Improvement of xylanase production by Penicillium oxalicum ZH‐30 using response surface methodology. Enzyme and Microbial Technology, 40(5), pp.1381– 1388. Available at: http://linkinghub.elsevier.com/retrieve/pii/S0141022906005217. Liming, X. & Xueliang, S., 2004. High‐yield cellulase production by Trichoderma reesei ZU‐02 on corn cob residue. Bioresource Technology, 91(3), pp.259–262. Available at: 98 http://linkinghub.elsevier.com/retrieve/pii/S0960852403001950 [Accessed June 11, 2013]. Linda, B., 1997. Introductory Botany, Plants, People and the environment Segunda Ed., Belmont CA. USA: Thomson Higher Education. Liu, D., Zhang, R., Yang, X., Xu, Y., et al., 2011. Expression, purification and characterization of two thermostable endoglucanases cloned from a lignocellulosic decomposing fungi Aspergillus fumigatus Z5 isolated from compost. Protein expression and purification, 79(2), pp.176–86. Available at: http://www.ncbi.nlm.nih.gov/pubmed/21708265. Liu, D., Zhang, R., Yang, X., Wu, H., et al., 2011. Thermostable cellulase production of Aspergillus fumigatus Z5 under solid‐state fermentation and its application in degradation of agricultural wastes. International Biodeterioration & Biodegradation, 65(5), pp.717–725. Available at: http://linkinghub.elsevier.com/retrieve/pii/S0964830511000722 [Accessed March 14, 2012]. Liu, J.X. & R, E., 2000. Cellulase treatment of untreated and steam pre‐treated rice straw Ð effect on in vitro fermentation characteristics. , 88. Lockington, R. a et al., 2002. Regulation by carbon and nitrogen sources of a family of cellulases in Aspergillus nidulans. Fungal genetics and biology : FG & B, 37(2), pp.190– 6. Available at: http://www.ncbi.nlm.nih.gov/pubmed/12409103. López, R.M. & Freitas, S., 2012a. Selección de cepas de Bacillus sp . potenciales para la producción de hidrolasas . , pp.1–39. López, R.M. & Freitas, S., 2012b. Selección de cepas de Bacillus sp . potenciales para la producción de hidrolasas . Universidad Autónoma de Aguascalientes, Mexico. Luciano, M.H. et al., 2012. A simple and fast method for the determination of endo‐ and exo‐cellulase activity in cellulase preparations using filter paper. Enzyme and microbial technology, 51(5), pp.280–5. Available at: http://www.ncbi.nlm.nih.gov/pubmed/22975126 [Accessed June 6, 2013]. Ma, L. et al., 2013. Kinetic studies on batch cultivation of Trichoderma reesei and application to enhance cellulase production by fed‐batch fermentation. Journal of biotechnology, pp.1–6. Available at: http://www.ncbi.nlm.nih.gov/pubmed/23702163. Maeda, R.N. et al., 2013. Cellulase production by Penicillium funiculosum and its application in the hydrolysis of sugar cane bagasse for second generation ethanol production by fed batch operation. Journal of biotechnology, 163(1), pp.38–44. Available at: http://www.ncbi.nlm.nih.gov/pubmed/23123260]. 99 Malherbe, S. & Cloete, T.E., 2002. Lignocellulose biodegradation : Fundamentals and applications. , pp.105–114. Mandels, M. & Weber, J., 1969. The production of cellulases. Advances in Chemistry, 95, pp.391–414. Marentes, F. et al., 2012. Evaluacion en campo de microorganismos promotores del crecimiento y celuloliticos. In Ecologia de microorganismos rizosfericos asociados a cultivos de arroz de Tolima y Meta. Bogota, pp. 131–149. Marjamaa, K. et al., 2013. Novel Penicillium cellulases for total hydrolysis of lignocellulosics. Enzyme and microbial technology, 52(6‐7), pp.358–69. Available at: http://www.ncbi.nlm.nih.gov/pubmed/23608505 [Accessed June 11, 2013]. Miller, G., 1959. Use of dinitrosalicilic acid reagent for determination of reducing sugar. Analytical Chemistry, 31, pp.426–428. Ministerio de Agricultura y Desarrollo Rural, 1996. La cadena del arroz en Colombia, una mirada global de su estructura y dinámica 1991‐2005. Available at: http://www.agronet.gov.co/www/docs_agronet/2005112141728_caracterizacion_arr oz.pdf. Mo, X. et al., 2010. Identification and characterization of a novel xylanase derived from a rice straw degrading enrichment culture. Applied microbiology and biotechnology, 87(6), pp.2137–46. Available at: http://www.ncbi.nlm.nih.gov/pubmed/20567817. Niranjane, A.P., Madhou, P. & Stevenson, T.W., 2007. The effect of carbohydrate carbon sources on the production of cellulase by Phlebia gigantea. Enzyme and Microbial Technology, 40(6), pp.1464–1468. Available at: http://linkinghub.elsevier.com/retrieve/pii/S0141022906005400. Oliveira, L. a, Porto, A.L.F. & Tambourgi, E.B., 2006. Production of xylanase and protease by Penicillium janthinellum CRC 87M‐115 from different agricultural wastes. Bioresource technology, 97(6), pp.862–7. Available at: http://www.ncbi.nlm.nih.gov/pubmed/15953719. Page, A.L., 1982. Methods of soil analysis. Part 2. Chemical and microbiological properties, Pantoja Alberto, 1997. Manejo Integrado de Plagas en arroz Centro Internacional de Agricultura Tropical. CIAT, ed., Cali. Park, Y.C. & Kim, J.S., 2012. Comparison of various alkaline pretreatment methods of lignocellulosic biomass. Energy, 47(1), pp.31–35. Available at: 100 http://linkinghub.elsevier.com/retrieve/pii/S0360544212006226 [Accessed May 27, 2013]. Perez, E. & Hernandez, S., 2011. Evaluacion de la actividad celulolitica de aislamientos de suelo de cultivo de arroz. Pontificia Universidad Javeriana. Quevedo, B.E., 2011. Evaluación de la degradación de residuos de floricultura para la obtención de azúcares con el uso de tres hongos lignocelulolíticos. Universidad Nacional de Colombia. Quiceno, D. & Mosquera, M., 2010. Alternativas Tecnologicas para el uso de la cascarilla de arroz como combustible. Universidad Autonoma de Occidente. Rahnama, N. et al., 2013. com Effect of Alkali Pretreatment of Rice Straw on Cellulase and Xylanase Production by Local Trichoderma harzianum SNRS3 under Solid State Fermentation. , 8, pp.2881–2896. Reyes, E. & Aguirre, M., 2009. Aislamiento de microorganismos celuloliticos y ligninoliticos a partir de muestras de suelo cultivado con arroz en los departamentos de Tolima y Meta. Pontificia Universidad Javeriana. Sa, P., Costa‐ferreira, M. & Aires‐barros, M.R., 2002. Xyl II from Bacillus subtilis. , 94, pp.265–275. Saibi, W., Amouri, B. & Gargouri, A., 2007. Purification and biochemical characterization of a transglucosilating beta‐glucosidase of Stachybotrys strain. Applied microbiology and biotechnology, 77(2), pp.293–300. Available at: http://www.ncbi.nlm.nih.gov/pubmed/17938914. Sánchez, C., 2009. Lignocellulosic residues: biodegradation and bioconversion by fungi. Biotechnology advances, 27(2), pp.185–94. Available at: http://www.ncbi.nlm.nih.gov/pubmed/19100826. Singh, A. et al., 2010. Key issues in life cycle assessment of ethanol production from lignocellulosic biomass: Challenges and perspectives. Bioresource technology, 101(13), pp.5003–12. Available at: http://www.ncbi.nlm.nih.gov/pubmed/20015644. Singh, N.P. & Ashok, P., 2009. Biotechnology for Agro‐Industrial Residues Utilisation, United Kingdom: Springer. Singhania, R.R. et al., 2010. Advancement and comparative profiles in the production technologies using solid‐state and submerged fermentation for microbial cellulases. Enzyme and Microbial Technology, 46(7), pp.541–549. Available at: 101 http://linkinghub.elsevier.com/retrieve/pii/S0141022910000657 [Accessed May 21, 2013]. Singhania, R.R. et al., 2013. Role and significance of beta‐glucosidases in the hydrolysis of cellulose for bioethanol production. Bioresource technology, 127, pp.500–7. Available at: http://www.ncbi.nlm.nih.gov/pubmed/23069613 [Accessed June 11, 2013]. Soest, P.J.V.A.N., Robertson, J.B. & Lewis, B.A., 1991. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. Journal of Dairy Science, 74, pp.3583–3597. Sofía, M.C. et al., 2001. INFLUENCIA DE ALGUNOS PARÁMETROS EN LA FERMENTACIÓN EN ESTADO SÓLIDO DEL HONGO TRICHODERMA VIRIDE. , XXI(1), pp.82–87. Sohail, M. et al., 2009. Cellulase production from Aspergillus niger MS82: effect of temperature and pH. New biotechnology, 25(6), pp.437–41. Available at: http://www.ncbi.nlm.nih.gov/pubmed/19552887 [Accessed November 8, 2012]. Sun, X. et al., 2008a. The composition of basal and induced cellulase systems in Penicillium decumbens under induction or repression conditions. Enzyme and Microbial Technology, 42(7), pp.560–567. Available at: http://linkinghub.elsevier.com/retrieve/pii/S0141022908000355 [Accessed June 28, 2013]. Sun, X. et al., 2008b. The composition of basal and induced cellulase systems in Penicillium decumbens under induction or repression conditions. Enzyme and Microbial Technology, 42(7), pp.560–567. Sunna, A. & Antranikian, G., 1997. Xylanolytic enzymes from fungi and bacteria. Crit Rev Biotechnol. Suto, M. & Tomita, F., 2001. Induction and catabolite repression mechanisms of cellulase in fungi. Journal of bioscience and bioengineering, 92(4), pp.305–11. Available at: http://www.ncbi.nlm.nih.gov/pubmed/16233102. Taherzadeh, M.J. & Karimi, K., 2008. Pretreatment of lignocellulosic wastes to improve ethanol and biogas production: a review., Available at: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=2635757&tool=pmcentre z&rendertype=abstract [Accessed May 23, 2013]. Tengerdy, R.P. & Szakacs, G., 2003. Bioconversion of lignocellulose in solid substrate fermentation. Biochemical Engineering Journal, 13(2‐3), pp.169–179. Available at: http://linkinghub.elsevier.com/retrieve/pii/S1369703X02001298. 102 Terrasan, C.R.F. et al., 2010. Production of xylanolytic enzymes by Penicillium janczewskii. Bioresource technology, 101(11), pp.4139–43. Available at: http://www.ncbi.nlm.nih.gov/pubmed/20122825. Willcock, J. & Magan, N., 2000. Impact of environmental factors on fungal respiration and dry matter losses in wheat straw. Journal of stored products research, 37(1), pp.35–45. Available at: http://www.ncbi.nlm.nih.gov/pubmed/11124368. Yang, S. et al., 2009. Hydrolysis of soybean isoflavone glycosides by a thermostable β‐ glucosidase from Paecilomyces thermophila. Food Chemistry, 115(4), pp.1247–1252. Available at: http://linkinghub.elsevier.com/retrieve/pii/S0308814609000788. Yang, S.Q. et al., 2006. High‐level of xylanase production by the thermophilic Paecilomyces themophila J18 on wheat straw in solid‐state fermentation. Bioresource technology, 97(15), pp.1794–800. Available at: http://www.ncbi.nlm.nih.gov/pubmed/16230011. Yeoman, C.J. et al., 2010. Thermostable enzymes as biocatalysts in the biofuel industry. Advances in applied microbiology, 70(10), pp.1–55. Available at: http://www.ncbi.nlm.nih.gov/pubmed/20359453. 103 A. Anexo Composición de medios de cultivo Agar Celulosa 1% (p/v) Celulosa alimentaria 10g/L Extracto de levadura 2,5g/L Triptosa 2,5g/L Sulfato de amonio 0,5g/L Cloruro de calcio 0,54g/L Fosfato monobásico de potasio 0,1g/L Fosfato dibásico de potasio 0,1g/L Cloranfenicol 0,1g/L Caldo tamo de arroz al 1%(p/v) Tamo de arroz 10g/L Extracto de levadura 2,5g/L Triptosa 2,5g/L Sulfato de amonio 0,5g/L Cloruro de calcio 0,5g/L Fosfato monobásico de potasio 0,1g/L Fosfato dibásico de potasio 0,1g/L Clorafenicol 0,1g/L 104 B. Anexo Curvas de calibración para los métodos analíticos Curvas de calibración para la determinación de azucares reductores Curva de calibración empleando glucosa como patrón Para las determinación de azucares reductores en la muestra en la actividad enzimática EG se empleó glucosa como patrón en la técnica de DNS. 1,2 Absorbancia (540 nm) 1,0 0,8 0,6 y=0,562x+0,014 2 R =0,99 0,4 0,2 0,2 0,4 0,6 0,8 1,0 1,2 1,4 Concentración (g/L) 1,6 1,8 2,0 2,2 105 Curva de calibración empleando xilosa como patrón Para las determinación de azucares reductores en la muestra en la actividad enzimática Xilanasa se empleó xilosa como patrón en la técnica de DNS. 1,4 Absorbancia (540 nm) 1,2 1,0 0,8 0,6 y=0,627x-0,025 2 R =0,99 0,4 0,2 0,0 0,2 0,4 0,6 0,8 1,0 1,2 1,4 Concentracion (g/l) 1,6 1,8 2,0 2,2 106 Curva de calibración de p‐nitrofenol Determinación de p‐nitrofenol en la muestra en la actividad enzimática CBH y β ‐glucosidasa 1,2 Absorbancia (400nm) 1,0 0,8 0,6 y=0,160x+0,07 2 R =0,99 0,4 0,2 1 2 3 4 5 Concentración (µmol/mL) 6 7 107 C.Anexo Análisis estadísticos Análisis de varianza para los diámetros de los halos de hidrolisis Kruskal‐Wallis All‐Pairwise Comparisons Test of diametro by Aislamiento Aisl. Mean Homogeneous Groups 4 47.000 A 3 43.500 AB 6 41.167 AB 1 34.833 AB 5 34.667 AB 2 29.167 AB 8 28.833 AB 10 24.667 AB 13 23.500 AB 9 21.500 AB 12 20.167 AB 7 13.500 AB 11 9.8333 AB 14 9.6667 AB 15 6.6667 AB 16 3.3333 B Alpha 0.05 Critical Z Value 3,529 Critical Value for Comparison 40.343 108 Análisis de varianza para la producción de CO2 en los ensayos de microcosmos. ANOVA de un factor mg_CO2 Suma de Gl Media cuadrática F Sig. cuadrados Inter‐grupos 668425,695 Intra‐grupos Total 3 4015,581 12 672441,277 15 222808,565 334,632 665,832 ,000 mg_CO2 a HSD de Tukey Tratamientos N Subconjunto para alfa = 0.05 1 control abiotico 4 Aspergillus 4 Paecilomyces 4 4 Penicillium Sig. 2 3 4 49,12398 130,98663 249,52745 585,63245 1,000 1,000 1,000 Se muestran las medias para los grupos en los subconjuntos homogéneos. a. Usa el tamaño muestral de la media armónica = 4,000. 1,000 109 Análisis de varianza del ABC de la actividad enzimática de Penicillium sp. Análisis de varianza del ABC de la actividad EG ANOVA ABC Suma de cuadrados Media gl cuadrática Inter‐grupos 70225,983 7 10032,283 Intra‐grupos 14098,799 16 881,175 Total 84324,782 23 F Sig. 11,385 ,000 ABC Subconjunto para alfa = 0.05 Tratamientos a HSD de Tukey a 3 4 5 72,7067 AVICEL 3 115,6267 115,6267 AVICEL+EXT+SULF 3 144,1667 144,1667 CMC 3 165,7333 165,7333 CMC+EXT 3 176,3000 176,3000 176,3000 AVICEL+SULF 3 177,9000 177,9000 177,9000 AVICEL+EXT 3 229,8333 229,8333 CMC+SULF 3 ,127 CMC+EXT+SULF 3 72,7067 AVICEL 3 115,6267 AVICEL+EXT+SULF 3 CMC 3 CMC+EXT 3 AVICEL+SULF 3 3 CMC+SULF Sig. Scheffé 2 3 AVICEL+EXT a 1 CMC+EXT+SULF Sig. Duncan N 3 ,236 115,6267 251,8667 ,210 144,1667 144,1667 165,7333 165,7333 ,096 176,3000 176,3000 177,9000 177,9000 ,066 ,094 229,8333 ,218 229,8333 251,8667 ,051 ,377 CMC+EXT+SULF 3 72,7067 AVICEL 3 115,6267 115,6267 AVICEL+EXT+SULF 3 144,1667 144,1667 144,1667 CMC 3 165,7333 165,7333 165,7333 165,7333 CMC+EXT 3 176,3000 176,3000 176,3000 176,3000 110 AVICEL+SULF 3 AVICEL+EXT 3 CMC+SULF 3 Sig. 177,9000 ,053 177,9000 177,9000 229,8333 229,8333 251,8667 ,502 ,160 ,155 Se muestran las medias para los grupos en los subconjuntos homogéneos. a. Usa el tamaño muestral de la media armónica = 3,000. Análisis de varianza del ABC de la actividad CBH ANOVA ABC Suma de cuadrados Gl Media cuadrática Inter‐grupos 9890566,425 7 1412938,061 Intra‐grupos 713665,948 16 44604,122 Total 10604232,373 F 31,677 ,000 23 Sig. ABC Subconjunto para alfa = 0.05 Tratamientos a HSD de Tukey 1 2 3 4 AVICEL 3 80,1600 AVICEL+SULF 3 220,2333 CMC 3 1076,2000 CMC+EXT 3 1317,3667 CMC+EXT+SULF 3 1366,0000 AVICEL+EXT+SULF 3 1535,6667 1535,6667 AVICEL+EXT 3 1657,3333 1657,3333 CMC+SULF 3 2046,0000 ,124 Sig. Duncana N ,990 AVICEL 3 80,1600 AVICEL+SULF 3 220,2333 CMC 3 CMC+EXT 3 ,059 1076,2000 1317,3667 1317,3667 111 CMC+EXT+SULF 3 AVICEL+EXT+SULF 3 AVICEL+EXT 3 CMC+SULF 3 Sig. a Scheffé 1366,0000 1366,0000 ,429 1535,6667 1657,3333 ,130 2046,0000 ,087 3 80,1600 AVICEL+SULF 3 220,2333 CMC 3 CMC+EXT 1076,2000 3 1317,3667 1317,3667 CMC+EXT+SULF 3 1366,0000 1366,0000 AVICEL+EXT+SULF 3 1535,6667 1535,6667 AVICEL+EXT 3 1657,3333 1657,3333 CMC+SULF 3 2046,0000 ,057 ,998 ,200 Se muestran las medias para los grupos en los subconjuntos homogéneos. a. Usa el tamaño muestral de la media armónica = 3,000. Análisis de varianza del ABC de la actividad β –glucosidasa ANOVA ABC Inter‐grupos Intra‐grupos Total 1,000 AVICEL Sig. Suma de cuadrados 2,761E8 gl Media cuadrática 7 63592361,307 16 3,397E8 23 39447046,235 3974522,582 F Sig. 9,925 ,000 112 ABC Subconjunto para alfa = 0.05 Tratamientos a HSD de Tukey Duncan Scheffé CMC+EXT+SULF 3 1466,3333 CMC 3 3793,6667 3793,6667 AVICEL+SULF 3 4871,6667 4871,6667 4871,6667 AVICEL+EXT+SULF 3 7459,0000 7459,0000 7459,0000 AVICEL+EXT 3 7941,6667 7941,6667 7941,6667 CMC+EXT 3 9481,3333 9481,3333 CMC+SULF 3 ,366 AVICEL 3 1180,4667 CMC+EXT+SULF 3 1466,3333 CMC 3 3793,6667 AVICEL+SULF 3 4871,6667 AVICEL+EXT+SULF 3 AVICEL+EXT 3 CMC+EXT 3 CMC+SULF 3 ,244 4871,6667 10919,3333 ,154 7459,0000 7941,6667 7941,6667 ,052 ,440 7459,0000 9481,3333 10919,3333 ,092 ,067 AVICEL 3 1180,4667 CMC+EXT+SULF 3 1466,3333 CMC 3 3793,6667 3793,6667 AVICEL+SULF 3 4871,6667 4871,6667 4871,6667 AVICEL+EXT+SULF 3 7459,0000 7459,0000 7459,0000 AVICEL+EXT 3 7941,6667 7941,6667 7941,6667 CMC+EXT 3 9481,3333 9481,3333 CMC+SULF 3 10919,3333 ,124 a. Usa el tamaño muestral de la media armónica = 3,000. 4 ,064 Se muestran las medias para los grupos en los subconjuntos homogéneos. 3 1180,4667 Sig. 2 3 Sig. a 1 AVICEL Sig. a N ,169 113 Análisis de varianza del ABC de la actividad enzimática de Paecilomyces sp. Análisis de varianza del ABC de la actividad EG ANOVA de un factor ABC Suma de gl Media cuadrática F Sig. cuadrados Inter‐grupos Intra‐grupos Total 39749,601 2055,405 41805,007 7 5678,514 14 21 146,815 38,678 ,000 ABC Tratamientos N Subconjunto para alfa = 0.05 1 a,b HSD de Tukey Duncan 5 ,0000 AVICEL+EXT 3 1,9877 AVICEL 3 14,0600 CMC+EXT+SULF 3 23,2533 CMC 3 27,7433 CMC+EXT 3 60,3267 CMC+SULF 2 AVICEL+SULF 2 27,7433 60,3267 86,0600 ,218 ,104 AVICEL+EXT+SULF 3 ,0000 AVICEL+EXT 3 1,9877 AVICEL 3 14,0600 14,0600 CMC+EXT+SULF 3 23,2533 23,2533 CMC 3 CMC+EXT 3 CMC+SULF 2 Sig. Scheffé 4 3 AVICEL+SULF a,b 3 AVICEL+EXT+SULF Sig. a,b 2 2 27,7433 ,059 AVICEL+EXT+SULF 3 ,0000 AVICEL+EXT 3 1,9877 AVICEL 3 14,0600 ,289 1,000 60,3267 ,236 144,6000 86,0600 1,000 144,6000 1,000 1,000 114 CMC+EXT+SULF 3 23,2533 23,2533 CMC 3 27,7433 27,7433 CMC+EXT 3 60,3267 CMC+SULF 2 86,0600 AVICEL+SULF 2 60,3267 Sig. ,471 ,169 ,560 144,6000 1,000 Se muestran las medias para los grupos en los subconjuntos homogéneos. a. Usa el tamaño muestral de la media armónica = 2,667. b. Los tamaños de los grupos no son iguales. Se utilizará la media armónica de los tamaños de los grupos. Los niveles de error de tipo I no están garantizados. Análisis de varianza del ABC de la actividad CBH ANOVA de un factor ABC Suma de Gl Media cuadrática F Sig. cuadrados Inter‐grupos 449753,513 7 Intra‐grupos 202078,377 16 Total 651831,890 23 64250,502 12629,899 5,087 ,003 ABC Tratamientos N Subconjunto para alfa = 0.05 1 a HSD de Tukey a 3 AVICEL+EXT+SULF 3 57,1700 CMC 3 131,0667 CMC+EXT+SULF 3 168,5433 AVICEL 3 183,0333 CMC+EXT 3 234,5667 234,5667 AVICEL+EXT 3 270,5667 270,5667 AVICEL+SULF 3 351,9000 351,9000 CMC+SULF 3 Sig. Duncan 2 527,7333 ,079 AVICEL+EXT+SULF 3 57,1700 CMC 3 131,0667 ,082 115 CMC+EXT+SULF 3 168,5433 168,5433 AVICEL 3 183,0333 183,0333 CMC+EXT 3 234,5667 234,5667 AVICEL+EXT 3 270,5667 270,5667 AVICEL+SULF 3 CMC+SULF 3 Schefféa 351,9000 351,9000 Sig. 527,7333 ,053 ,089 ,073 AVICEL+EXT+SULF 3 57,1700 CMC 3 131,0667 CMC+EXT+SULF 3 168,5433 168,5433 AVICEL 3 183,0333 183,0333 CMC+EXT 3 234,5667 234,5667 AVICEL+EXT 3 270,5667 270,5667 AVICEL+SULF 3 351,9000 351,9000 CMC+SULF 3 527,7333 Sig. ,245 ,092 Se muestran las medias para los grupos en los subconjuntos homogéneos. a. Usa el tamaño muestral de la media armónica = 3,000. 8.3.4.3 Análisis de varianza del ABC de la actividad β –glucosidasa ANOVA de un factor ABC Suma de gl Media cuadrática F Sig. cuadrados Inter‐grupos Intra‐grupos Total 1127359,719 7 43061,285 16 1170421,004 23 161051,388 2691,330 59,841 ,000 116 ABC Tratamiento N Subconjunto para alfa = 0.05 1 a HSD de Tukey Duncan 390,0667 410,1000 441,9000 458,1000 1007,2500 3 333,3667 CMC+EXT 3 AVIEL+EXT 3 AVICEL+EXT+SULF 3 CMC+EXT+SULF 3 AVICEL+SULF 3 CMC+SULF 3 404,1000 ,370 3 AVICEL 3 CMC+EXT 3 AVIEL+EXT 3 AVICEL+EXT+SULF 3 CMC+EXT+SULF 3 AVICEL+SULF 3 Sig. a AVICEL CMC+SULF Scheffé 333,3667 237,6667 4 3 CMC 3 CMC Sig. a 2 237,6667 1,000 ,127 333,3667 390,0667 404,1000 410,1000 390,0667 404,1000 410,1000 441,9000 458,1000 3 1007,2500 1,000 ,114 ,165 1,000 CMC 3 237,6667 AVICEL 3 333,3667 333,3667 CMC+EXT 3 390,0667 390,0667 AVIEL+EXT 3 404,1000 404,1000 AVICEL+EXT+SULF 3 410,1000 410,1000 CMC+EXT+SULF 3 AVICEL+SULF 3 CMC+SULF 3 Sig. 441,9000 458,1000 ,073 Se muestran las medias para los grupos en los subconjuntos homogéneos. a. Usa el tamaño muestral de la media armónica = 3,000. 1007,2500 ,339 1,000 117 Análisis de varianza del ABC de la actividad Xilanasa ANOVA de un factor ABC Suma de gl Media cuadrática F Sig. cuadrados Inter‐grupos 19630,337 3 6543,446 Intra‐grupos 524,440 8 65,555 20154,777 11 Total 99,816 ,000 ABC Tratamientos N Subconjunto para alfa = 0.05 1 a HSD de Tukey 3 XILANO+SULF 3 XILANO+EXT+SULF 3 XILANO+EXT 3 271,4333 Duncan 3 XILANO+SULF 3 XILANO+EXT+SULF 3 XILANO+EXT 3 Sig. a Scheffé 302,7333 1,000 XILANO 271,4333 1,000 376,4000 1,000 376,4000 1,000 1,000 271,4333 XILANO+SULF 3 XILANO+EXT+SULF 3 XILANO+EXT 3 302,7333 1,000 1,000 348,1667 1,000 3 Se muestran las medias para los grupos en los subconjuntos homogéneos. 1,000 XILANO a. Usa el tamaño muestral de la media armónica = 3,000. 302,7333 4 348,1667 Sig. 3 XILANO Sig. a 2 348,1667 1,000 376,4000 1,000 1,000 118 D.Anexo Determ minación d de Celulossa y hemiccelulosa