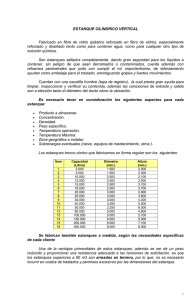
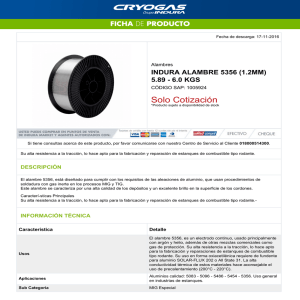
4 Fauna Asociada en Relación con la Operación de Estanques
Anuncio

Fauna Asociada en Relación con la Operación de Estanques Camaronícolas en Sistemas Lagunares 4 Fauna Asociada en Relación con la Operación de Estanques Camaronícolas en Sistemas Lagunares HENDRICKX ANTONIO MICHEL E. y MEDA MARTÍNEZ MARCO Unidad Académica Mazatlán, Instituto de Ciencias del Mar y Limnología, UNAM, Apdo. postal 811, Mazatlán 82000, Sinaloa. E-mail: [email protected] 4.1. 4.2. 4.3. 4.4. Introducción ......................................................................................................................................... 51 Asentamiento de comunidades biológicas en las compuertas de entrada de estanques de cultivo ................................................................................................................................................... 54 Macrofauna asociada a los estanques de cultivo de camarón......................................................... 62 Discusión y conclusiones.................................................................................................................... 68 Agradecimientos................................ ................................ ................................ ................................ .. 69 Bibliografía ........................................................................................................................................... 70 4.1. Introducción El desarrollo actual de la camaronicultura en el sur del estado de Sinaloa ha provocado el interés, por parte, de varios grupos de investigación, para determinar en que medida estos sistemas artificiales se comportan como unidades ecológicas y en que medida pueden afectar las condiciones ambientales de los sistemas costeros naturales adyacentes. En el área de Mazatlán, por ejemplo, existe ya una producción relativamente elevada de camarones de cultivo y todo 51 Camaronicultura y Medio Ambiente parece indicar que la superficie total ocupada por las granjas aumentará comparativamente con la superficie de cuerpos de aguas naturales (Páez-Osuna, 1992). El análisis de los procesos biológicos no relacionados con el crecimiento del camarón y que tienen lugar en los estanques de cultivo son importantes por diversos motivos: primero permiten obtener una imagen del estado de salud del sistema; en efecto, la aparición de especies asociadas pueden servir de indicadores para el seguimiento o monitoreo de la calidad del agua, por ejemplo mediante la medición de la concentración de ciertos metabolitos o contaminantes en representantes de estas especies sin tener que recurrir directamente al monitoreo del recurso cultivado. Segundo, la presencia de especies depredadoras puede influir negativamente sobre el resultado final de la cosecha, ya que pueden alimentarse de postlarvas o de juveniles de camarones. Tercero, ciertas especies colonizadoras pueden, si son exitosas, representar una competencia significativa a nivel del consumo del alimento proporcionado al camarón de cultivo; consecuentemente, la producción por unidad de área puede sufrir bajas sensibles. Cuarto, la presencia de poblaciones abundantes de organismos filtradores puede ayudar en el control de la carga de partículas en suspensión que suele provocar problemas de hipoxia o anoxia, o bien eliminar algas tóxicas que pueden afectar la calidad terminal del producto. Quinto, algunas de las especies colonizadoras de los estanques de cultivo podrían representar un producto con potencial de explotación, sin que forzosamente actúen en deterioro de la producción principal (i.e., el camarón) que se pretende obtener. En Sinaloa, todas las actividades de cultivo de camarón son de tipo "monocultivo", es decir que interviene una sola especie. El abastecimiento de agua es, por lo general, a partir de cuerpos de agua naturales y consiste en agua de salinidad variable obtenida de sistemas lagunares o bien o de sistemas basados en el uso de agua marina (salinidad del orden de 34 /oo) que se diluye con aportes de agua dulce. Las granjas más productivas son, lógicamente, aquellas que pueden tener un abastecimiento continuo de agua de calidad adecuada. En Sinaloa, eso es un problema fundamental debido a la desecación progresiva de los cuerpos de agua lagunares durante la época de sequía con un consecuente incremento de la salinidad que, en última instancia, rinde esta agua impropia para el uso en estanquería. El bombeo de agua desde los cuerpos naturales se realiza en general de manera directa, es decir sin un sistema de prefiltración selectivo. El abastecimiento hacia los estanques puede ser directo o vía un canal que permite un abastecimiento por gravedad continuo y controlable mediante estructuras con compuertas ajustables. Pocos estudios han sido orientados al análisis de la fauna colonizadora y de sus posibles efectos en los estanques de cultivo de camarón de tipo semi-intensivo, como aquellos que se encuentran en el sur de Sinaloa. Con el doble propósito de determinar cuales especies colonizan las estructuras por medio de las cuales se abastecen de agua los estanques y cuales son las especies de la macrofauna que logran invadir y desarrollarse en los mismos estanques, se 52 Fauna Asociada en Relación con la Operación de Estanques Camaronícolas en Sistemas Lagunares realizó un estudio piloto en la granja "Clementina" S.C.L., ubicada a unos 25 km. de Mazatlán, Sin., ésta es una granja camaronícola de tipo semi-intensivo ubicada al final del sistema lagunar conocido como “Estero Urías”. Se encuentra rodeada por los esteros “La Sirena”, “Barrón” y “El Confite”, comunicados entre sí y formando parte del “Estero Urías”. Los sistemas de abastecimiento de agua de la granja se basan en el bombeo a partir de uno de los esteros naturales del sistema lagunar adyacente y en la repartición del agua a los estanques mediante un canal de distribución (canal de llamada). En esta granja, este sistema permite un recambio del 5 al 8 % del volumen del estanque al día mediante compuertas de entrada ("monjes") cerradas con una malla rígida de prefiltración de 5 mm de abertura y otra, interna, de 1 mm de abertura. Antes de iniciar el proceso de siembra de las postlarvas de camarones, el estanque permanece vacío por un período bastante largo que impide la sobrevivencia de especies acuáticas. Eso permite tomar como hipótesis de partida que ninguna especie colonizadora se encuentra presente previamente al inicio del estudio. Debido al tamaño de la malla, un alto porcentaje de organismos planctónicos pueden introducirse en los estanques desde el momento que éstos se inundan al inicio de un nuevo ciclo de cultivo. De una u otra manera, la introducción de diversas especies en el estanque desencadena un proceso de competición por alimento, espacio y, hasta cierto punto, por el oxígeno disuelto disponible. Curiosamente, en el Pacífico mexicano todavía no se ha desarrollado un interés para estudios integrales, ya sea cualitativos o cuantitativos que permitan determinar la dinámica de colonización o los patrones de crecimiento de las especies que logren introducirse en los estanques. La información incluida en el presente trabajo se refiere esencialmente a dos tipos de fauna: la macrofauna (tamaño > 0.5 mm) que participa en los procesos de sucesión en las comunidades que se establecen en las estructuras de entrada de agua (compuertas o “monjes”); la macrofauna asociada con los estanques que se introduce desde el principio o a lo largo del cultivo, cuando se llenan los estanques, y que permanece hasta el momento de la cosecha, alcanzando tallas considerables. Para analizar la primera, se requiere aplicar un método de seguimiento a través del ciclo de cultivo, método que conlleva a determinar los patrones de colonización (i.e., determinar cuales especies participan y cuales dominan) y de una eventual sucesión de especies (i.e., determinar cuales son las especies pioneras y especies oportunistas) en el sustrato. Para el estudio de la fauna que coloniza los estanques, es posible realizar muestreos durante el ciclo de cultivo, pero afectarían el propio cultivo por los métodos de captura que se requiere implementar. Además, la representatividad de las muestras obtenidas podría ser baja. Por estos motivos, cuando se trata de realizar un estudio preliminar de la composición y biomasa de la macrofauna asociada al cultivo, es preferible realizar las capturas al final del cultivo, cuando se realiza la cosecha del producto vaciando progresivamente el estanque. De esta forma, la captura es más representativa con ca. 100% de la fauna y no se basa en una 53 Camaronicultura y Medio Ambiente evaluación a partir de muestreos aleatorios. 4.2. Asentamiento de comunidades biológicas en las compuertas de entrada de estanques de cultivo El mecanismo de asentamiento implica, en general, la presencia inicial de especies sésiles que operan a manera de especies pioneras, permitiendo el asentamiento de una mayor diversidad de especies, incluso algunas especies móviles que encuentran un lugar adecuado (un nicho ecológico) para refugiarse y alimentarse entre las comunidades de especies sésiles pioneras. En muchos casos es necesario que el sustrato sea modificado por el tránsito del agua (erosión leve) o por depósitos de tipo inorgánico (arenas, lodos) u orgánicos (partículas inertes, película de microorganismos) antes de que se realice el asentamiento de especies pioneras. Un esquema sencillo del patrón de asentamiento (Fig. 4.1) refleja el constante papel que tiene el flujo de agua para mantener la comunidad en un proceso activo de colonización mediante aportes elevados de oxígeno (agua en cascada) y de partículas orgánicas nutritivas. El número de especies involucradas en los asentamientos es variable y depende de dos factores principales: la riqueza faunística de especies sésiles (i.e., que viven fijas a un sustrato) en el ambiente natural, lo cual definirá el número potencial de especies capaces de realizar un proceso de colonización; y la estabilidad de las características ambientales que definirán si las condiciones son propicias para el asentamiento y persistencia de algunas de estas especies. Debido a la limpieza que efectúa el personal de las granjas en “los monjes” al término de cada ciclo de cultivo y a la exposición al sol por varias semanas (i.e., sin escurrimiento de agua), la superficie de concreto de “los monjes” de entrada se encuentra desprovista de cualquier tipo de sustrato (inerte o vivo) cuando se inicia el llenado con agua del canal de llamada. Al igual que en otros ambientes sólidos, “los monjes” de los estanques de cultivo reciben depósitos (arena y lodo) que se acumulan progresivamente desde el inicio cuando se llenan los estanques. La formación de esta capa de sustrato modifica la capacidad receptora para el asentamiento de especies. Así, durante los muestreos en ambos ciclos de cultivo, el piso de los monjes se cubre de una película de depósitos de sedimento y de material orgánico que es rápidamente colonizada por algas clorofíceas filamentosas (Enteromorpha clathrata) (Fig. 4.2). Conforme progresa el asentamiento de las especies animales colonizadoras, los depósitos orgánicos e inorgánicos se modifican o desaparecen, simultáneamente con la disminución o desaparición de la capa de algas filamentosas en los lugares de asentamiento de la fauna. Esta alga solamente se mantiene a las orillas de los pisos de “los monjes”, al límite de la zona de flujo de agua. Las especies móviles aparecen rápidamente, buscando refugio y alimento aprovechando la colonización inicial por parte de las algas. 54 Fauna Asociada en Relación con la Operación de Estanques Camaronícolas en Sistemas Lagunares ´ DEPOSITOS Orgánicos Inorgánicos ALGAS VERDES FILAMENTOSAS SUSTRATO LIM PIO Flujo de agua Flujo de agua Flujo de agua Sedimentos suspendidos Nutrientes Materia orgánica particular Oxígeno disuelto Estadíos larvarios ESPECIES COLONIZADORAS I ´ Sesiles pioneras ESPECIE S COLONIZADORA S II ´ Sesiles asociadas Flujo de agua ESPECIES DOMINA NTES ESPECIES ASOCIADA S Móviles pioneras Competencia por es pacio y alimento Crecimiento en tallas y en tamaño de población Fig. 4.1. Diagrama de asentamiento de las comunidades de macro-invertebrados en las compuertas de entrada de agua en una granja de cultivo del sur de Sinaloa. detalle de las ramificaciones su variación Enteromorpha clathrata Fig. 4.2. Alga verde filamentosa colonizadora en los estanques de cultivo de camarones, Sinaloa (retomado de Zertuche-González et al., 1995). 55 Camaronicultura y Medio Ambiente Las especies sésiles encontradas en “los monjes” comprenden, además de la especie de alga verde citada, especies pertenecientes a tres grupos faunísticos: los moluscos, los poliquetos y los crustáceos. Las especies móviles corresponden a tres especies de crustáceos y a dos especies de poliquetos de la familia Nereidae (Cuadro 4.1). La presencia de Enteromorpha clathrata representa un factor determinante en el patrón de colonización; en efecto, la presencia de algas puede facilitar el mecanismo de fijación de larvas en sustratos duros. Cuadro 4.1. Especies de invertebrados encontradas en las comunidades establecidas en las compuertas de entrada de estanques de cultivo y tipo de alimentación Esta clasificación se realizó en base a la literatura especializada (Ogle et al., 1982; Barnes, 1974; Ewing, 1984; Salazar-Vallejo et al , 1988; Brusca y Brusca, 1990; Hernández-Alcantara, 1992). Especie Grupo taxonómico Poliquetos Poliquetos Grado de locomoción Sésil Sésil Estrategia de alimentación Suspensívoro, detritívoro Detritívoro Polydora cornuta Capitella capitata Laeonereis culveri Poliquetos Móvil Detritívoro Neanthes succinea Poliquetos Móvil Detritívoro Sabella melanostigma Poliquetos Sésil Suspensívoro Ficopomatus sp. Poliquetos Sésil Suspensívoro Corophium cf. baconi Anfípodos Móvil Detritívoro Corophium cf. panamense Anfípodos Móvil Suspensívoro Caprella sp. Anfípodos Móvil Suspensívoro Uromunna sp. Isopodos Móvil Detritívoro Sinelobus stanfordi Tanaidaceos Móvil Detritívoro Balanus inexpectatus Cirripedos Sésil Suspensívoro Callinectes arcuatus Braquiuros Móvil Detritívoro/oportunista (juv.) Mytella strigata Pelecípodos Sésil Suspensívoro Protothaca asperrima Pelecípodos Sésil Suspensívoro Tagelus affinis Pelecípodos Sésil Suspensívoro Acteocina inculta Gasterópodos Móvil Detritívoro En la granja estudiada, se reconocieron 14 especies de invertebrados que participan en los 56 Fauna Asociada en Relación con la Operación de Estanques Camaronícolas en Sistemas Lagunares procesos de colonización. Estas 14 especies comprenden 5 especies de anélidos poliquetos (Fig. 4.3), 4 especies de moluscos (Fig. 4.4) y 5 especies de crustáceos (Fig. 4.5). Además de pertenecer a grupos faunísticos distintos y, como consecuencia, tener una morfología característica, las especies animales encontradas presentan un tipo de alimentación distinto. Sin entrar en detalles, se pueden clasificar en dos grupos principales: (a) especies con una alimentación basada en partículas orgánicas vivas o inertes en suspensión (suspensívoras); (b) especies con una alimentación a base de depósitos o detritus (detritívoras) (Cuadro 4.1). Los crustáceos pertenecen en su mayoría al muy diversificado grupo de los peracarídeos, unos pequeños organismos que, en general, no rebasan los 10 mm de longitud total y que se encuentran en todos los ambientes marinos y de agua salobre en el Pacífico mexicano. El conocimiento es limitado de la biología y ecología de estas especies en los ambientes lagunares y estuarinos, algunas de estas especies logran establecerse éxitosamente debido a las condiciones ambientales. La identificación a nivel específico de estos organismos es problemática por la falta de estudios de esta fauna en las regiones tropicales del Pacífico este. El único cirrípedo encontrado, B. inexpectatus, forma parte de un complejo de especies cuya taxonomía es difícil y requiere de un examen detallado de las placas y de los apéndices del organismo. Común en ambientes intermareales, los cirrípedos asociados a lagunas costeras muestran una variabilidad limitada; B. inexpectatus es considerada una de las especies dominantes en el sistema estudiado. El tanaidáceo Sinelobus standfordi ha sido reportado en aguas salobres de Colombia, a pesar de ser una especie intermareal previamente citada para ambientes marinos. La autecología de los anfípodos encontrados (géneros Corophium y Caprella) es igualmente mal conocida en el área. La identificación de las especies de Corophium resulta incierta por no existir estudios a fondo en los ambientes estuarino-lagunares del área. La fauna de poliquetos asociada a los sistemas lagunares es, por los mismos motivos que los crustáceos, reducida en comparación con los ambientes típicamente marinos. La especie encontrada del genero Ficopomatus pertenece a un complejo de especies distribuidas a nivel mundial; sin embargo, se desconoce la existencia de variaciones geográficas suficientes, entre poblaciones establecidas en diversos lugares, para justificar el reconocimiento de varias especies. Las especies adicionalmente reconocidas en los estanques de cultivo son típicamente intermareales y han sido identificadas previamente en ambientes arenosos o lodosos. En cuanto a los moluscos, se incluyen dos especies de pelecípodos típicas de los sistemas estuarinos (Mytella strigata y Tagelus affinis) y una especie de gasterópodo (Acteocina inculta) citada esencialmente como una especie asociada a bahías poco profundas. Desde el punto de vista cuantitativo, se pueden observar variaciones estacionales importantes tanto en la abundancia de organismos asentados como en la composición de la fauna a largo del año. En un ciclo de estudio realizado de marzo a junio de 1993, los valores de densidad media 2 obtenidos para “los monjes” de los cuatro estanques fueron del orden de 700 orgs./m en marzo, 57 Camaronicultura y Medio Ambiente 2 2 2 12,000 orgs./m en abril, 3,000 orgs./m en mayo y 11,500 orgs./m en junio (Fig. 4.6). Parápodo posterior Parte an terior Parápodo anterior Neuroseta Nereidae típico Parte anterior contraida Parte anterior alargada Capitella capitata Setígera Setígera Sabella melanostigma Fig. 4.3. Especies de poliquetos colonizadoras de las compuertas de entrada de agua de los estanques de camarones, Sinaloa (retomado de Hartman, 1968, 1969; Méndez y Cardell, 1994; Salazar-Vallejo y Carrera-Parra, 1997). 58 Fauna Asociada en Relación con la Operación de Estanques Camaronícolas en Sistemas Lagunares Tagelus affinis Protothaca asperrima valva izquierda (interior) valva derecha (exterior) Acteocina inculta Mytella strigata valva izquierda (interior) valva derecha (exterior) Fig. 4.4. Especies de moluscos colonizadoras de las compuertas de entrada de agua de los estanques de camarones, Sinaloa (retomado de Keen, 1971; Poutiers, 1995). 59 Camaronicultura y Medio Ambiente Balanus inexpectatus Caprellidae Corophium. Sinelobus standfordi Uromunna Fig. 4.5. Especies de crustáceos colonizadoras de las compuertas de entrada de agua de los estanques de camarones, Sinaloa (retomado de Gardiner, 1975; Henry y McLaughlin, 1975; Zullo, 1979; Brusca, 1980). 60 Fauna Asociada en Relación con la Operación de Estanques Camaronícolas en Sistemas Lagunares 20000 2 Densidad (orgs./m ) 25000 15000 CICLO I CICLO II 10000 5000 0 MES 1 MES 2 MES 3 MES 4 Fig. 4.6. Densidad media observada en las cuatro compuertas de entrada de agua (suma de 4 estanques) durante los dos ciclos (Ciclo I, marzo a junio; Ciclo II, agosto a noviembre). 2 La mayor densidad fue de 29,700 orgs./m en abril, pero resultó un dato algo excepcional. Las especies que contribuyen en su mayor parte a este valor elevado de densidad en los asentamientos en este período son esencialmente crustáceos peracáridos: Caprella sp. y Sinelobus aff. stanfordi. En cambio, el mejillón de estero, Mytella strigata, no fue abundante pero contribuyó notablemente a la biomasa al final del ciclo por ser un organismo grande y pesado, con un ciclo de crecimiento más largo. En el segundo ciclo, realizado de agosto a noviembre, los valores de abundancia crecieron más rápido debido principalmente a la contribución de los anélidos en la fase inicial (hasta 96% de las muestras), seguido por una dominancia del mejillón Mytella strigata y de los crustáceos peracáridos. Los valores de densidad media estimados para 2 los cuatro “monjes” en este segundo ciclo fueron los siguientes: 150 orgs./m en agosto, 6,800 2 2 2 orgs./m en septiembre; 13,000 orgs./m en octubre y 22,000 orgs./m en noviembre (Fig. 4.6). 2 Las densidades más altas al final del ciclo fueron del orden de 26,500 orgs./m . Una comparación simple entre los resultados observados entre los dos ciclos, muestra una fuerte variación en la dinámica de asentamiento. En el ciclo de agosto a noviembre, la abundancia total fue mayor con 2 valores indicativos (suma de las densidades medias de cada mes) de 42,448 org/m contra 2 solamente 6,739 orgs./m en el ciclo anterior. 61 Camaronicultura y Medio Ambiente 4.3. Macrofauna asociada a los estanques de cultivo de camarón Un análisis de la megafauna asociada a los cultivos de camarón al término de dos ciclos de cultivo realizados en la granja citada anteriormente, permitió detectar la presencia de un número importante de especies de peces. Estas especies, cuyas larvas deben penetrar a través de la malla de filtración y colonizan el sistema, encuentran en los estanques un ambiente poco agresivo por la falta de depredadores adultos. En efecto, considerando que antes de cada ciclo de cultivo los estanques se vacían por completo, el crecimiento de los organismos colonizadores se inicia al momento de la inundación del sistema, es decir poco tiempo antes de la introducción de las postlarvas de camarones (organismos de cultivo). Según los registros obtenidos (Hendrickx et al., 1996), 22 especies de peces y dos de crustáceos (jaibas) tienen la capacidad de invadir los estanques y de proliferar en ellos, alcanzando en ocasiones una talla final considerable. La riqueza específica encontrada rebasa en ocasiones la riqueza citada en la literatura para algunos sistemas lagunares naturales en la región. Sin embargo, la biomasa que representan estos organismos dada su talla es muy reducida (entre aproximadamente 30 y 85 kg. al término de un ciclo de 4 meses) y no rebasa el 0.34-0.87 % de la biomasa del camarón producida al final del ciclo de cultivo (Cuadro 4.2). Eso podría indicar que la competencia por alimento o por espacio es alta, que el reclutamiento de larvas de peces es bajo o que las condiciones de sobrevivencia de la ictiofauna no son ideales. Esta última hipótesis, sin embargo, no parece ser válida para todas las especies ya que en algunos casos los especímenes recolectados al finalizar un ciclo (en particular el ciclo II) alcanzaron una talla representando hasta un 66-91% de la talla máxima conocida (Cuadro 4.3; véase infra). Las dos especies de cangrejo nadadores ("jaibas") del género Callinectes encontradas (Fig. 4.7) son comunes en los sistemas estuarino-lagunares de la región. Callinectes arcuatus, en particular, es ciertamente la especie dominante en el “estero de Urías”, adyacente a la granja estudiada (Alvarez León, 1980; Paul, 1981). La segunda especie, C. toxotes, ha sido registrada en el mismo sistema pero con una frecuencia menor (Alvarez León, 1980). Las 22 especies de peces registradas en los estanques pertenecen a grupos faunísticos muy distintos, tales como las anchoas, los gobios y los robalos (Figs. 4.8-4.10). Solamente tres especies mostraron cierta constancia en su abundancia en los ciclos de estudio (Pomadasys macracanthus, Lile stolifera y Dormitator latifrons mexicanus) y el análisis comparativo de la fauna encontrada en sistemas naturales de la región indica que la riqueza muestreada es similar a aquella que ha sido citada en sistemas estuarino-lagunares naturales adyacentes (Hendrickx et al., 1996), pero con una participación de especies significativamente distinta. Por ejemplo, es de subrayar que ningún espécimen de los géneros Mugil, Arius y Galeichthys, especies comunes en los sistemas lagunares naturales de la región, fue encontrado en los estanques de cultivo durante el estudio. 62 Fauna Asociada en Relación con la Operación de Estanques Camaronícolas en Sistemas Lagunares dientes centrales bien marcados diente lateral triangular base más ancha bordes divergentes pedúnculo ocular corto fisura supraorbital bien visible, pero angosta telson formando un triángulo equilatero 6 5 diente lateral largo caparazón en vista dorsal abdomen (macho) en vista ventral abdomen (hembra) en vista ventral Callinectes arcuatus frente con dientes centrales bien marcados dientes laterales en forma de lóbulo telson formando un triángulo alargado pedúnculo ocular corto 6 base angosta bordes casi paralelos 5 diente lateral largo caparazón en vista dorsal abdomen (macho) en vista ventral abdomen (hembra) en vista ventral Callinectes toxotes Fig. 4.7. Especies de Callinectes (Portunidae) encontradas en los estanques de cultivo de camarón, Sinaloa (según Hendrickx et al., 1996) (retomado de Hendrickx,1995). 63 Camaronicultura y Medio Ambiente Anchovia macrolepidota Anchoa mundeola Centropomus nigrescens Cynoscion xanthulus Achirus mazatlanus Chanos chanos Centropomus robalito Diapterus peruvianus Fig. 4.8. Especies de peces encontradas en los estanques de cultivo de camarones, Sinaloa (según Hendrickx et al., 1996) (retomado de Fisher et al., 1995). 64 Fauna Asociada en Relación con la Operación de Estanques Camaronícolas en Sistemas Lagunares Oligolipes mundus Elops affinis Euleptorhampus viridis Eucinostomus argenteus Lyle stolifera Gerres cinereus Micropogonias altipinnis Pomadasys macracanthus Fig. 4.9. Especies de peces encontradas en los estanques de cultivo de camarones, Sinaloa (según Hendrickx et al., 1996) (retomado de Fisher et al., 1995). 65 Camaronicultura y Medio Ambiente Gobiidae Gobionellus vista lateral Gobiomorus Gobionellus microdon cabeza en vista ventral Dormitator Fig. 10. Géneros y especies de peces encontrados en los estanques de cultivo de camarón, Sinaloa (según Hendrickx et al., 1996) (retomado de Allen y Robertson, 1994; Fisher et al., 1995). 66 Fauna Asociada en Relación con la Operación de Estanques Camaronícolas en Sistemas Lagunares Cuadro 4.2. Abundancias total y relativa de la fauna de acompañamiento obtenida al final de los dos ciclos de cultivo (1993 y 1994). Año Camarones Número total y por ha. 1993 1994 650.10 (90.10 ) 3 3 750.10 (103.10 ) Peso fresco total y por ha. 1993 1994 3 3 8548 kg (1171 kg) 9721 kg (1332 kg) Total 2355 (100) 2623 (100) Peso fresco Total 29.18 (100) 84.96 (100) Macrofauna asociada Número de especímenes Peces Decápodos 2316 (98.3) 39 (1.7) 2571 (98.0) 52 (2.0) Peces 26.31 (90.2) 79.47 (93.5) Decápodos 2.87 (9.8) 5.49 (6.5) Como se mencionó anteriormente, quizá una de las observaciones más interesantes se relaciona con el hecho que una proporción importante de las especies más abundantes alcanzaron tallas finales (i.e. después de los cinco meses de duración del ciclo de cultivo) superiores al 50% de la talla máxima conocida (Cuadro 4.3). Sin embargo, por el hecho que el reclutamiento no es controlado y que se supone que es permanente, no hay posibilidad de contar con un registro exacto respecto al momento que los organismos entraron en el estanque en su estado larval (o juvenil) con el fin de evaluar el tiempo que se tardan para alcanzar estas tallas. Cuadro 4.3. Tallas máximas observadas para las siete especies de peces más abundantes recolectadas (ambos ciclos) comparativamente con las tallas máximas citadas en la literatura (según Hendrickx et al., 1996). Especies Talla máxima conocida (TL; cm) Pomadasys macracanthus Gerres cinereus Centropomus robalito Elops affinis Micropogonias altipinnis Lile stolifera Dormitator latifrons mexicana 35.0 28.0 30.0 40.0 60.0 12.0 25.0 Talla máxima (TL; cm) Ciclo I (feb. – jun. 1993) 12.0 (29%) 9.5 (34%) 18.0 (33%) 23.0 (58%) 15.0 (25%) 9.5 (79%) 17.0 (68%) Talla máxima (TL; cm) Ciclo II (ago. 1993 – ene. 1994) 17.0 (49%) 12.5 (45%) 23.5 (78%) 36.5 (91%) 13.5 (23%) 8.0 (67%) 16.5 (66%) 67 Camaronicultura y Medio Ambiente La producción obtenida al final de ambos ciclos para los camarones fue del orden de 1.2-1.3 toneladas por hectárea en la granja; la biomasa total recuperada en cuanto a peces y crustáceos fue de entre 30 y 85 kg por ciclo. El área útil del estanque estudiado era de aproximadamente 6 hectáreas, lo cual proporciona una producción final estimada de entre solamente 5 y 14 kilos por hectárea, lo cual es sumamente bajo. 4.4. Discusión y conclusiones Ciertos factores abióticos ligados a la naturaleza del sustrato (i.e. placas de concreto) o a las condiciones ambientales predominantes (e.g. corriente continua, poca profundidad, fuerte incidencia de la luz solar) pueden limitar el establecimiento de ciertas especies en los sustratos de “los monjes”. Algunos factores bióticos, tales como la disponibilidad del alimento adecuado, pueden ser otros factores controladores. Los resultados indican que algunas especies pioneras en la comunidad son desplazadas a lo largo del tiempo por otras más aptas o eficientes en utilizar los recursos (véase Dayton, 1971; Eltringham, 1971; Jackson, 1977; Dean y Hurd, 1980; Sousa, 1984; Henschel y Cook, 1990; Sousa y Conell, 1992). Estos factores obviamente pueden estar estrechamente vinculados, actuando a la par en cualquier combinación sobre las comunidades de flora y fauna, tal como ocurre en todos los ambientes marinos y dulceacuícolas (Odum, 1969 ; 1972; Margalef, 1974). El recubrimiento del piso de “los monjes” con una película de depósitos de sedimento y de material orgánico seguido de la colonización por algas verdes favorece el asentamiento de especies sésiles colonizadoras. Estas especies pueden alimentarse directamente de los depósitos orgánicos presentes en el sustrato, o de las partículas suspendidas que llegan con el agua o que se resuspenden en el mismo lugar (Eltringham, 1971; Vegas-Vélez, 1971). El seguimiento de los estadíos larvarios de las especies susceptibles de colonizar “los monjes” es una tarea difícil debido al alto grado de riqueza del sistema adyacente. En las comunidades estudiadas se presentaron tres grupos dominantes por su número o por su biomasa: dos grupos de peracáridos (los caprélidos y los tanaidáceos) y un grupo de pelecípodos, representados por el mitílido Mytella strigata. Si bien los peracáridos forman parte de la fauna móvil oportunista que encuentra en las comunidades colonizadoras un nicho adecuado para asentarse y desarrollarse, la presencia de los mejillones en los monjes de entrada es probablemente el factor más significativo observado en el segundo ciclo (agosto-enero). La capacidad filtradora de esta especie, ciertamente parecida a aquella de Mytilus edulis, una especie afín pero de regiones templadas, podría jugar un papel determinante para seleccionar esta especie como organismo indicador de la calidad de agua que abastece las compuertas. Por otra parte, la eliminación de una parte de la carga particular inerte o viva también debe ser 68 Fauna Asociada en Relación con la Operación de Estanques Camaronícolas en Sistemas Lagunares significativa para el sistema y podría ser, incluso, incrementada, favoreciendo el asentamiento de colonias más grandes de especies filtradoras con el aumento de la superficie semi-plana de las compuertas de entrada, donde el agua escurre antes de llenar el estanque. Experimentos de reciclaje de agua en circuitos cerrados utilizando una especie de gusano (Marphisa sanguinea) mostró una clara disminución de la carga de amonio (Hamanoet al., 1994). Por otra parte, la fauna de acompañamiento del camarón de cultivo apareció más diversificada de lo que se podría suponer. El estanque actúa como un ámbito favorable al crecimiento de especies de peces que encuentran alimento, poca competencia y ausencia de depredadores. Según Hendrickx et al. (1996), el examen de los contenidos estomacales de algunos especímenes de peces no resultó indicativo dada la alta incidencia de estómagos vacíos. El comportamiento depredador por parte de algunas especies de peces podría afectar la población de camarones, pero no se tienen evidencias al respecto. Según Brewer et al. (1995), quienes estudiaron el consumo de Penaeidae por 30 especies de peces marinos, la incidencia sobre las poblaciones de camarones está directamente relacionada con la talla del pez y el hábitat del camarón. Salini et al. (1990) también encontraron un fuerte impacto de algunas especies de peces sobre los juveniles de especies de Penaeidae comerciales en ambientes estuarinos. El intentar prevenir la colonización de los estanques por peces o jaibas es una tarea difícil; se requiere establecer mecanismos de filtración más eficaz (luz de malla más fina), pero eso a la vez provocaría el taponamiento del filtro. Un tratamiento previo del agua (sistema de filtración acoplado al bombeo) sería igual de problemático aunque el uso de filtros progresivamente más finos y el uso de agua bajo presión aumentaría la eficiencia del sistema, el uso de un sistema cerrado de abastecimiento, con reciclaje de agua filtrada y purificada evitaría la eventual sobrecarga de nutrientes y de material orgánico. Agradecimientos Se agradece a Mercedes Cordero Ruiz por el trabajo editorial y a Germán Ramírez Reséndiz por el apoyo con la preparación de las figuras. Los autores agradecen también a P. Young por la identificación de los cirrípedos, y a P. Hernández Alcántara por la identificación de los poliquetos. A J. Cillis y F. Fiers (IRSNB-KBIN, Bélgica) por el apoyo con la fotografía de microscopía electrónica de barrido (MEB). Parte de este estudio fue apoyado por el proyecto CONACyT 0625-N9110. 69 Camaronicultura y Medio Ambiente Bibliografía Allen, G.R. y Robertson, D.R. (1994). Fishes of tropical eastern Pacific. University of Hawaii Press, Honolulu, Hawaii. 332 p. Alvarez León, R. (1980). Necton y bentos de tres esteros adyacentes a Mazatlán (Sin.) México. Revista de Biología Tropical, 28 (2): 237-262. Barnes, R.D. (1974). Invertebrate zoology. Third Edition, W.B. Saunders Co., Philadelphia. 870 p. Brewer, D.T., Blaber, S.J.M., Salini, J.P. y Farmer, M.J. (1995). Feeding ecology of predatory fishes from Groote Eylandt in the Gulf of Carpentaria, Australia, with special reference to predation on Penaeid prawns. Estuarine, Coastal and Shelf Science, 40: 557-600. Brusca, R.C. (1980). Common intertidal invertebrates of the Gulf of California. Second Edition. The University of Arizona Press, Tucson, Arizona. 513 p. Brusca, R.C. y Brusca, G.J. (1990). Invertebrates. Sinauer Assoc. Inc. Publishers. Sunderland, Massachusetts. 922 p. Dayton, P.K. (1971). Competition, disturbance, and community organization: the provision and subsequent utilization of space in a rocky intertidal community. Ecological Monographs, 41 (4): 351-389. Dean, T.A. y Hurd, L.E. (1980). Development in an estuarine fouling community: The influence of early colonists on later arrivals. Oecologia, 46: 295-301. Eltringham, S.K. (1971). Life in mud and sand. The English Universities Press, London. 218 p. Ewing, M.R. (1984). Capitellidae. In: Vebelarcker, J.M. y Johnson, P.G. (Eds.). Taxonomic Guide to the Polychaetes of the Northern Gulf of Mexico. Final Report to the Minerals Management Service, contract 14-12-001-29091. Barry A. Vittor and Associates, Inc., Mobile, Alabama. Fischer, W., Krupp, F., Schneider, W., Sommer, C., Carpenter, K.E. y Niem, V.H. (1995). Vols. II y III. Plantas e Invertebrados. In: Fischer, W., Krupp, F., Schneider, W., Sommer, C., Carpenter, K.E. y Niem, V.H. (Eds.). Guía FAO para la identificación de especies para los fines de la pesca. Pacífico centro-oriental. F.A.O. Roma, Italia. p. 647-1813. Hamano, T., Tsuboi, S., Imai, A., Hoshino, N., Okimoto, H., Jinnouchi, S. y Hayashi, K.I. (1994). Maintenance of water quality by rockwool and its practical application to aquaculture. Suisanzoshoku, 42 (1): 127-133. Hartman, O. (1968). Atlas of the errantiate polychaetous annelids from California. Allan Hancock Foundation, University of Southern California, Los Angeles. p. 1-828. Hartman, O. (1969). Atlas of the sedentariate polychaetous annelids from California. Allan Hancock Foundation, University of Southern California, Los Angeles. p. 1-812. Hendrickx, M.E., Salgado-Barragán, J. y Meda-Martínez, M.A. (1996). Abundance and diversity of macrofauna (fish and decapod crustaceans) in Penaeus vannamei culture ponds in Western Mexico. Aquaculture, 143: 61-73. 70 Fauna Asociada en Relación con la Operación de Estanques Camaronícolas en Sistemas Lagunares Henry, D.P. y McLaughlin, P.A. (1975). The barnacles of the Balanus amphitrite complex (Cirripedia, Thoracica). Zoologische Verhandelingen, 141: 1-254. Henschell, J.R. y Cook, P.A. (1990). The development of a marine fouling community in relation to the primary film of microorganisms. Biofouling, 2: 1-11. Hernández-Alcantara, P. (1992). Los poliquetos (Annelida: Polychaeta) de la plataforma continental del golfo de California, México. Taxonomía, abundancia numérica y distribución geográfica. Tesis de Maestría. U.A.C.P.y.P. del C.C.H. Instituto de Ciencias del Mar y Limnología. Universidad Nacional Autónoma de México. 427 p. Jackson, J.B. (1977). Competition on marine hard substrate: The adaptive significance of solitary and colonial strategies. American Naturalist, 111 (980): 743-767. Keen, A.M. (1971). Sea shells of tropical west America. Marine mollusks from Baja California to Peru. Second Edition. Stanford University Press, Stanford, California. 1064 p. Margalef, R. (1974). Ecología. Editorial Omega, Barcelona. 951 p. Méndez, M.N. y Cardell, M.J. (1994). Effects of sample fixation on body shape of Capitella capitata (Polychaeta, Capitellidae). In: Dauvin, J.C., Laubier, L. y Reish, D.J. (Eds.). Actes de la 4ème Conférence internationale des Polychètes. Mémories du Muséum national d´Histoire naturelle, Paris,162: 111-117. Odum, E.P. (1969). The strategy of ecosystem development. Science, 164 (3877): 262-270. Odum, E.P. (1972). Ecología. Tercera Edición. Nueva Editorial Interamericana, México. 639 p. Ogle, J.T., Heard, R.W. y Sieg, J. (1982). Tanaidacea (Crustacea: Peracarida) of the gulf of Mexico. I. Introduction and an annotated bibliography of tanaidacea previously reported from the gulf of Mexico. Gulf Research Reports, 7 (2): 101-104. Páez-Osuna, F. (1992). Calidad del agua: Evaluación y su importancia en la camaronicultura. Ciencias del Mar, Epoca 1 (12): 3-5. Paul, R.K.G. (1981). The development of a fishery for portunid crabs of the genus Callinectes (Decapoda, Brachyura) in Sinaloa, Mexico. Overseas Development Administration, London. 78 p. Poutiers, J.M. (1995). Bivalvos. In: Fischer, W., Krupp, F., Schneider, W., Sommer, C., Carpenter, K. E. y Niem, V.H. (Eds.). Guía FAO para la identificación de especies para los fines de la pesca. Pacífico centro-oriental. Vol. I. Plantas e Invertebrados. F.A.O. Roma, Italia. p. 99-222. Salazar-Vallejo, S.I. y Carrera-Parra, L.F. (1997). Taxonomía de Poliquetos (Annelida: Polychaeta). Ecosur, CONACyT. 64 p. Salazar-Vallejo, S.I., de León-González, J.A. y Salaices-Polanco, H. (1988). Poliquetos (Annelida: Polychaeta) de México. Universidad Autónoma de Baja California Sur, Libros Universitarios, La Paz, B.C.S. 212 p. Salini, J.P., Braber, S.J.M. y Brewer, D.T. (1990). Diets of piscivorous fishes in a tropical Australian estuary, with special references to predation on penaeid prawns. Marine Biology, 71 Camaronicultura y Medio Ambiente 105: 363-374. Sousa, W.P. (1984). Intertidal mosaics: patch size, propagule availability, and spatially variable patterns of succession. Ecology, 65 (6): 1918-1935. Sousa, W.P. y Conell, J.H. (1992). Grazing and succesion in marine algae. In: John, D.M., Hawkins, S.J. y Price, J.H. (Eds.). Plant-animal interactions in the marine benthos. Special. Vol. 46. The Systematics Association. Clarendon Press, Oxford. p. 425-442. Vegas-Vélez, M. (1971). Introducción a la ecología del bentos marino. Serie de Biología, Monografía no. 9. Secretaría General de la O.E.A., Washington, D.C. 98 p. Zertuche González, J.A., Pacheco Ruiz, I. y González González, J. (1995). Macroalgas. In: Fischer, W., Krupp, F., Schneider, W., Sommer, C., Carpenter, K.E. y Niem, V.H. (Eds.). Guía FAO para la identificación de especies para los fines de la pesca. Pacífico centro-oriental. Vol. I. Plantas e Invertebrados. F.A.O. Roma, Italia. p. 9-82. Zullo, V.A. 1979. Marine flora and fauna of the Northeastern United States. Arthropda: Cirripedia. NOAA Technical Report NMFS Circular, 425: 1-29. 72