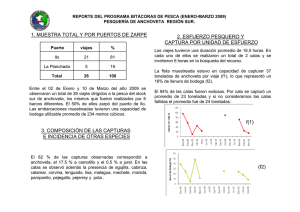
Informe vol. 35 número 2, Cruceros de evaluación de la
Anuncio

CONSEJO DIRECTIVO DEL IMARPE 2008 Presidente Contralmirante (r) Héctor Soldi Soldi Vicepresidente Contralmirante Mauro Cacho de Armero Directores Doctor Luis Icochea Salas Ingeniero José A. Sarmiento MadueÑo Ingeniero Jorge Vértiz CalderÓn Director Ejecutivo Economista Godofredo CaÑote Santamarina INSTITUTO DEL MAR DEL PERÚ INFORME ISSN 0378 - 7702 Volumen 35 Número 2 Cruceros de evaluación de la biomasa desovante de la anchoveta peruana. Método de la producción de huevos (MPH). Invierno 2003, 2004, 2005 Abril - Junio 2008 Callao, Perú CONTENIDO Cruceros de evaluación de la biomasa desovante de la anchoveta peruana. Método de la producción de huevos (MPH). Invierno 2003, 2004, 2005 1. Biomasa desovante de la anchoveta peruana. Método de producción de huevos (MPH). Invierno 2003 The Peruvian anchoveta spawning biomass. Eggs production Method (EPM). Winter 2003 Patricia AyÓn Dejo, Betsy BuitrÓn Díaz ………………………………………………...........................… 77 2. Producción diaria de huevos de la anchoveta peruana en agosto - setiembre 2003 The Peruvian anchoveta eggs daily production in August – September 2003 Patricia AyÓn Dejo ……………………………………………………………………….......................…… 81 3. Evaluación hidroacústica de anchoveta y otros recursos pelágicos. Agosto y setiembre 2003 Peruvian anchovy and other pelagic resources hydroacoustic evaluation. August and September 2003 Ramiro Castillo, Salvador Peraltilla, Marceliano Segura …………………….........................…… 87 4. Características biológicas de la anchoveta peruana y otros recursos pelágicos durante agosto- setiembre 2003 Biological characteristics of Peruvian anchoveta and other pelagic resources during August – September 2003. Julio Mori, Cecilia PeÑa, Arturo Ventosilla ……………………………………….......................…… 97 5. Fitoplancton en el invierno 2003. Phytoplankton in winter 2003. Flor Chang, Elcira Delgado ………………………………………………………….........................…… 103 6. Condiciones oceanográficas físicas en invierno 2003 Physical oceanographic conditions in winter 2003 Luis Vásquez Espinoza …………………………………………………………………….........................… 109 7. Biomasa desovante de la anchoveta del stock norte centro en el invierno 2004 Anchoveta spawning biomass of the north and central stock during winter 2004 Patricia AyÓn, Angel Perea …………………………………………………………............................… 115 8. El fitoplancton durante el invierno 2004 Phytoplankton during winter 2004 Sonia Sánchez, Nelly Jacobo ……………………………………………………............................……… 119 9. Características oceanográficas físicas en invierno 2004 Physic oceanographic characteristics in Winter 2004 Luis Vásquez Espinoza ………………………………………………………………….............................… 123 10. Evaluación acústica de la distribución y abundancia de especies pelágicas y mesopelágicas. Invierno 2004 Acoustic assessment of distribution and abundance of pelagic and mesopelagic species during winter 2004 Mariano Gutiérrez , Salvador Peraltilla ………………………………………................................… 129 11. Biomasa desovante de anchoveta en la zona norte centro del mar peruano (4o36,5’S– 14o40’S) a finales del invierno austral 2005 The anchovy spawning biomass in the north central zone of Peruvian sea (4o36,5’ – 14o40’S) at the late austral winter 2005 Patricia AyÓn, M. Ángel Perea ………………………………………….…...........................................…. 141 12. Biomasa de anchoveta y otros recursos pelágicos estimados por el método hidroacústico a finales del invierno 2005 Peruvian anchovy and other pelagic resources estimated by the hydroacoustic method at late winter 2005. Ramiro Castillo, Salvador Peraltilla, Mariano Gutiérrez ………..........................................……. 145 13. Fitoplancton del mar peruano a fines del invierno 2005 Peruvian sea phytoplankton during late winter 2005 Elcira Delgado, Flor Chang …………………………………………………..............................… ............153 14. Aspectos biológicos de los principales recursos pelágicos peruanos a finales del invierno 2005 Biological aspects of the major Peruvian pelagic resources at late winter 2005 Cecilia PeÑa , Carlos Goicochea, Julio Mori ……………………………….........................................… 159 15. Parámetros reproductivos de adultos de anchoveta peruana a finales del invierno 2005 Reproductive parameters of adult Peruvian anchovy at the end of winter 2005 Ángel Perea De La Matta, Cecilia Roque García …………………………............................................. 167 16. Condiciones ambientales físicas del mar peruano a fines del invierno 2005 Peruvian marine physical environmental conditions during late winter 2005 Luis Vásquez Espinoza……………………..................................................................................................... 173 Instrucciones para los autores ……………………………………………………………...................................…… 181 Biomasa desovante de la anchoveta peruana (MPH). Invierno 2003 Ayón y Buitrón BIOMASA DESOVANTE DE LA ANCHOVETA PERUANA. MÉTODO DE PRODUCCIÓN DE HUEVOS (MPH). INVIERNO 2003 THE PERUVIAN ANCHOVETA SPAWNING BIOMASS. EGGS PRODUCTION METHOD (EPM). WINTER 2003 Patricia Ayón Dejo Betsy Buitrón Díaz RESUMEN AyÓn P, BuitrÓn B. 2008.- Biomasa desovante de la anchoveta peruana. Método de producción de huevos (MPH). Invierno 2003. Inf Inst Mar Perú 35(2): 77-80.- La biomasa desovante de la anchoveta peruana del stock norte centro en agosto-setiembre 2003 fue calculada en 1.265.581 t. El valor de los parámetros estimados fueron: proporción sexual 0,52; fecundidad 11.407 ovocitos/hembra, frecuencia de desove 9,6%, peso 18,2 g. Se discute y compara los pesos y fecundidad de los últimos años, con relación al estimado final de la biomasa desovante. Palabras clave: anchoveta peruana, biomasa desovante, método de producción de huevos, invierno 2003 ABSTRACT AyÓn P, Buitron B. 2008 .- The Peruvian anchoveta spawning biomass. The eggs production method (HPM). Winter 2003. Inf Inst Mar Peru 35 (2): 77-80.- The spawning biomass of north-central anchovy stock for August-September was assessed at 1,265,581 t. The values of the estimated parameters were: sex ratio 0.52, fecundity 11,407 eggs / female, spawning frequency 9.6%, weight 18.2 g. We discuss and compare the weights and fertility of recent years in relation to the final estimate of the spawning biomass. Keywords: Peruvian anchovy, spawning biomass, eggs production method, winter 2003. 83° W INTRODUCCIÓN El estimado de la biomasa de la anchoveta por el método acústico en el verano 2003 fue de 7,7 millones de toneladas con un límite de confianza (LC) de ± 13,27%. Se distribuyó principalmente en el norte entre Paita y Chimbote con las mayores concentraciones dentro de las 20 mn. En el sur se halló desde Atico hasta Morro Sama dentro de las 30 mn de la costa. En otoño, la distribución se amplió hasta las 120 mn de la costa, principalmente en la región norte-centro, con las mayores concentraciones frente a Pimentel, Chicama, Chimbote, y de Huacho a Pisco, siempre dentro de las 60 mn de la costa y asociadas al enfriamiento de las aguas en la zona (IMARPE 2003). Los estimados de la biomasa desovante utilizando el MPH, en los últimos años han sido congruentes con los estimados de la biomasa por el método acústico así como el de población virtual, habiéndose obtenido para el invierno 2002 una biomasa mayor a los 5,5 millones de toneladas métricas (AyÓn y 82° W 81° W 80° W 79° W 78° W 77° W 76° W 75° W 74° W Pto. Pizarro Pta. Sal 4° S 4° S Trayectos y lances realizados Talara 5° S 5° S Paita Pta. Gobernador BIC SNP.2 Pta. La Negra 6° S BIC J.OLAYA B. 6° S BIC J. Olaya B. Mórrope Pim entel 7° S BIC SNP.2 7° S Chérrepe Chicama 8° S 8° S Salaverry Punta Chao 9° S 9° S Chimbote Casma Punta Lobos 10° S 10° S Huarmey Punta Bermejo Supe 11° S 11° S Huach o Chancay 12° S 12° S Callao Pucusana 13° S 13° S Cerro Azul Tambo de Mora Pisco 14° S 14° S B. Independencia Punta Infiernillos Pta. Caballas 15° S 83° W 82° W 81° W 80° W 79° W 78° W 77° W 76° W 75° W 15° S 74° W Figura 1.- Carta de trayecto. Crucero BIC Olaya y SNP2 0308-09 BuitrÓn 2002). Para el invierno 2003 se programó la ejecución de un crucero para utilizar el MPH, bajo un escenario donde, en las capturas de los últimos meses, la incidencia de juveniles causó la 77 disminución de los desembarques, constituyendo un factor importante a considerar tanto para el desarrollo del crucero como para el estimado final de la biomasa. Informe IMARPE Vol. 35(2): Abril-Junio 2008 el caso de la fecundidad se utilizó 101 hembras de un rango de tamaño de 12,0 a 16,5 cm, se halló la ecuación lineal que fue aplicada a las primeras 25 hembras de las calas que entraron en los cálculos. MATERIAL Y MÉTODOS El crucero BIC Olaya y SNP2, se llevó a cabo del 13 agosto al 16 de setiembre 2003, frente al área comprendida entre caleta La Cruz y Bahía Independencia. El BIC SNP2 trabajó dentro de las 30 mn de la costa desde caleta La Cruz hasta Pucusana, y el BIC Olaya, entre las 30 y 90 mn de la costa, pero hacia el sur, entre Pucusana y Bahía Independencia cubrió hasta 50 mn. Algunos perfiles alcanzaron distancias >120 mn y en otras, fueron <50 mn de la costa. Se ejecutaron 64 perfiles, la distancia entre ellos fue de 12,5 mn, al norte de Punta La Negra y al sur de Casma y en los otros fue de 10 mn (Figura 1). La producción diaria de huevos se calculó de acuerdo al trabajo de AyÓn (2009, en este informe), aplicándose el método iterativo programado en un software escrito en Gwbasic, modificado de Santander et al. (1984), para una PC. RESULTADOS Parámetros adultos.- El peso promedio de las hembras obtenido fue de 18,2 g ± 4%; rango 12 - 17 cm LT. La proporción sexual tuvo un promedio de 0,52 con un coeficiente de variación de 5%. La ecuación para la fecundidad fue: E= -92,598 + 631,4 (W). Al aplicar esta ecuación se obtuvo un promedio en la fecundidad parcial de 11.407 ovocitos/hembra ± 16%. La frecuencia de hembras con desove diario fue el 9,6%, con un coeficiente de variación del 18%. Producción diaria de huevos.- La producción diaria de huevos en el área en estudio fue de 4,00 E+13 huevos/día, con un coeficiente de variación de 13,9%. Para el peso promedio se hizo una corrección de 4% y se obtuvo una ecuación adecuada a este fin. En ovocitos/hembra peso (g) 40 30 20 10 0 12,5 13,5 La biomasa desovante de la anchoveta calculada mediante el MPH, en este crucero 0308-09, fue baja en comparación con la estimada por el método acústico, durante el mismo periodo y área de muestreo. Es importante tomar en consideración algunos factores que podrían haber incidido drásticamente en estos resultados. Por un lado, el estimado acústico corresponde a toda la biomasa, que incluye individuos adultos (activos como inactivos) y juveniles. La incidencia de juveniles de acuerdo a lo registrado en el seguimiento de las pesquerías habría sido importante, sobrepasando la tolerancia máxima del 10% en las capturas (informe interno del seguimiento de pesquerías), hecho corroborado cuando en los 7°S, la captura de juveniles llegó a casi 90%. Se debe enfatizar que el porcentaje de juveniles durante el crucero fue 74,71%, que en peso representó un 50,42% de la biomasa acústica total (IMARPE 2003). B = Biomasa desovante Po = Producción diaria de huevos W = Peso promedio F = Frecuencia de desove E = Fecundidad R = Proporción sexual En el cálculo de peso (W) y fecundidad (E) se utilizaron las primeras 25 hembras; para la proporción sexual (R) se usaron los primeros 800 g. Para la frecuencia de desove (F) se emplearon otras 25 hembras, colectadas de manera especial para poder determinar los folículos post-ovulatorios. 11,5 DISCUSIÓN El estimado final de la biomasa desovante de la anchoveta se obtuvo de acuerdo a la ecuación general B=PoW/FER, en donde: Durante el crucero se ejecutaron 103 lances, 48 el SNP2 y 55 el Olaya. Para calcular la biomasa de la anchoveta se usaron 44 calas positivas. Para colectar adultos y huevos se siguió la metodología empleada por Santander et al. (1984). Adicionalmente a la red CalVET, para colección de huevos de anchoveta, se utilizó el equipo CUFES a fin de obtener información de la distribución de huevos, temperatura, salinidad y clorofila-a. 10,5 Biomasa desovante de la anchoveta.- De acuerdo con los parámetros estimados por el método de producción de huevos (MPH), para el periodo agostosetiembre del presente año, en el área comprendida entre caleta La Cruz y Bahía Independencia se estimó una biomasa desovante de anchoveta equivalente a 1.265.581 toneladas métricas (t). 14,5 15,5 16,5 17,5 El peso promedio ha sido ligeramente más alto que en el 2002; pero fue 30% menor que los registros de otros años. Los pesos 30000 20000 10000 0 10,5 longitud (cm) 12 13,5 15 16,5 18 longitud (cm) 2000 2001 2002 2003 2000 Figura 2.- Variación del peso promedio de hembras de anchoveta en los periodos de desove de invierno-primavera, entre el 2000 y el 2003. 2001 2002 2003 Figura 3.- Variación de la fecundidad de hembras de anchoveta en los periodos de desove de invierno-primavera, entre el 2000 y el 2003. 78 Biomasa desovante de la anchoveta peruana (MPH). Invierno 2003 Ayón y Buitrón que en el verano anterior mostraron influencia de las Aguas Subtropicales Superficiales (ASS) en la zona norte-centro, con fuertes proyecciones hacia la costa, que fueron más significativas frente a Chimbote (25 mn), Pisco (10 mn) y Morro Sama (20 mn) (IMARPE 2003), con grandes procesos de mezcla entre ACF y ASS principalmente al norte de punta Infiernillos hasta punta La Negra. En los años anteriores (Figura 5) la influencia de las ASS tuvo un menor impacto, con mayor dominio de las ACF. I.G. Tallas de anchoveta Regiòn Norte-Centro 8 7 6 IG 5 4 3 2 Veda 1 0 E02 1F 1M 1A 5A 4M 3J 3J 2A 2S 2O 1N 1D E03 5E 4F 4M 4A 3M 3J 12 - 14 SEMANAS cm 3J 2A 2S 2O 1N 1D > 14 cm Figura 4.- Promedio del índice gonadosomático (IGS) mensual por rango de tallas. Periodo 2002 a 2003 (J. Mori, com. pers.) 84 W 82 W 80 W 78 W 76 W 74 W 72 W 70 W Pto. Pizarro 4S Pta. Sal SALINIDAD SUPERFICIAL DEL MAR ( ups) Talara 5S Paita 6S Pimentel 7S Cr Chicama 8S Huarmey .2 Supe 1/ 11 S Chancay 12 S Callao Pucusana 3 -03 2-0 02 0 2/ 00 00 .2 12 S 10 S - 03 02 Cr Cr 11 S 9S 3/ 00 Punta Chao Chimbote 10 S 8S .2 Salaverry 9S 13 S 13 S Cer ro Azul Pisco 14 S 14 S Punta Infiernillos 15 S 15 S San Juan 16 S 16 S Atico 17 S 17 S Mollendo Ilo 18 S M. Sama 76 W ASS ACF 5S 6S Pta. La Negra 7S 4S Es evidente que la producción diaria de huevos fue muy baja, similar a 1990 (3,009E+13) pero con un CV alto (Carrasco y AyÓn informe interno), tal vez, debido a que hubo un submuestreo en la parte costera. Para este año, 2003, los huevos tuvieron distribución diferente, en donde el CV sólo fue 13%. La frecuencia de estaciones positivas para huevos de anchoveta, en comparación con otros años, fue muy baja, pues normalmente se obtiene más del 50%. En este crucero el porcentaje de estaciones con huevos de anchoveta fue 36,3%. 74 W 72 W 18 S 70 W AES Figura 5.- Distribución de las masas de agua de acuerdo con las características de la salinidad (L. Vásquez, com. pers.) por rango de talla, en el 2003 fueron algo mayores que en 2002, excepto en 17,0 cm, pero mucho menores a los del 2000 en todo rango de tallas (Figura 2). 2003 no se evidenció claramente el reposo reproductivo en otoño (Figura 4), encontrándose, por lo tanto, un número menor de ovocitos. En la fecundidad parcial por rango de tallas, se apreció diferencias claras entre los cuatro años (Figura 3). A pesar que el peso promedio en 2003 fue mayor que en 2002, la fecundidad para la misma talla fue mucho menor. Esto podría relacionarse con los resultados del seguimiento de las pesquerías, donde la clasificación macroscópica y el análisis microscópico de los ovarios, coinciden en que en el Otra evidencia del desove en el otoño fue la presencia de larvas de anchoveta con edades entre 15 a 40 días, indicando que antes del invierno habría ocurrido un desove importante. Este prolongado desove correspondió solamente a la fracción adulta de mayor talla (>14 cm). Estas observaciones podrían atribuirse a las condiciones oceanográficas observadas antes y durante del desarrollo del crucero, 79 CONCLUSIONES Teniendo en cuenta todas estas apreciaciones con relación a los parámetros que intervienen en la estimación de la biomasa desovante de la anchoveta, vemos: • Por un lado, hay un desove continuado desde el verano; y • Por otro lado, un alto porcentaje de juveniles en toda la zona explorada, que constituyen más del 50% del peso total obtenido por el método acústico. Todos estos cambios observados en estas variables podrían estar explicados, en parte, por las condiciones oceanográficas. Se puede mencionar que, en la zona de distribución del stock norte centro, previo a este crucero existió una fuerte influencia de las Aguas Subtropicales Superficiales (ASS), y de las Aguas Ecuatoriales Superficiales (AES) (IMARPE 2003). De acuerdo con el análisis micros- Informe IMARPE Vol. 35(2): Abril-Junio 2008 cópico, se observó diferencias en la fracción desovante por distancia a la costa. Se apreció mayor actividad desovante en la zona muy costera (dentro de las 15 mn) y por fuera de las 70 mn de la costa; y entre las 30 y 70 mn de la costa la fracción desovante se estimó en 6,7% debido en parte a que los cardúmenes en esta zona estaban constituidos básicamente por juveniles. Sin embargo, la baja frecuencia de atrésicos estaría indicando que el crucero se desarrolló en pleno proceso reproductivo. Esto concuerda con las catalogaciones macroscópicas, donde se observó un mayor porcentaje en estado de desove (estadio V) que alcanzó el 64,5% con un IGS de 6,73 y un FC de 0,6480 (IMARPE 2003). REFERENCIAS AyÓn P. 2009. Producción diaria de huevos de la anchoveta peruana en agosto-setiembre 2003. Inf Inst Mar Perú 35(2):81-84. AyÓn P, BuitrÓn B. 2002. Informe sobre la biomasa desovante de la anchoveta en agosto del 2002 entre caleta La Cruz y Punta Infiernillos (BIC Olaya y SNP-2 0208). Informe interno. Carrasco S, AyÓn P. 1990. Informe sobre la Biomasa desovante de la anchoveta en invierno 1990. informe interno. IMARPE. 2003a. Informe Ejecutivo del crucero 0302-04 d Evaluación Hidroacústica de Recursos Pelágicos Tacna a Tumbes. BIC José Olaya Balandra –BIC SNP-2 L/P Imarpe 26 de febrero al 06 de 80 febrero al 06 de abril 2003. IMARPE. 2003b. Informe sobre el desarrollo de la pesquería pelágica enero-julio 2003. Informe interno. Agosto 2003. IMARPE. 2003c. Informe Ejecutivo. Crucero de estimación de la biomasa desovante de anchoveta por el método de producción de huevos (MPH), entre caleta La Cruz y Bahía Independencia (13 agosto-16 setiembre 2003). Santander H, Alheit J, Smith P. 1984. Estimación de la biomasa de la población desovante de anchoveta peruana Engraulis ringens en 1981 por aplicación del Método de Producción de Huevos. Bol. Inst. Mar Perú. 8(6):209-250. Producción diaria de huevos de anchoveta en invierno 2003 Ayón PRODUCCIÓN DIARIA DE HUEVOS DE LA ANCHOVETA PERUANA EN AGOSTO - SETIEMBRE 2003 THE PERUVIAN ANCHOVETA EGGS DAILY PRODUCTION IN AUGUST – SEPTEMBER 2003 Patricia Ayón Dejo1 RESUMEN AyÓn P. 2008. Producción diaria de huevos de la anchoveta peruana en agosto - setiembre 2003. Inf Inst Mar Perú 35(2): 81-85.- Se presenta un análisis de los resultados del estimado de la producción diaria de huevos de Engraulis ringens, utilizando dos diseños de muestreo diferentes y dos métodos de agrupación. Los resultados muestran que el cálculo de la producción diaria de huevos con el nuevo diseño de muestreo y las regiones móviles da un buen estimado, con una reducción importante en el coeficiente de variación. Con el nuevo diseño de muestreo es posible estimar la biomasa del recurso anchoveta aplicando dos métodos directos de evaluación de manera simultánea como el Método de Producción de Huevos y el Método Hidroacústico. Palabras clave: MPH, producción diaria de huevos, anchoveta peruana. ABSTRACT AyÓn P. 2008. The Peruvian anchoveta eggs daily production during August – September 2003. Inf Inst Mar Perú 35(2): 81-85.- This report shows the results about the egg daily production of Engraulis ringens. Two different sampler designs and two kind data sort were used. The egg daily production obtained with the new sampler design and moving regions was good. Using this new sampler design is possible to evaluate the anchovy biomass with egg daily production method and Hydroacoustic Method at the same time. Keywords: EDPM, egg daily production, Peruvian anchoveta. INTRODUCCIÓN Uno de los parámetros importantes para aplicar el Método de Producción de Huevos (MPH) en especies de peces, es la producción diaria de huevos (PDH), que consiste en conocer la cantidad de huevos que se depositan en el mar en un momento dado. Esta producción dependerá directamente de las características biológicas de los reproductores, y del área en que se colecten las muestras. Hacer un diseño de muestreo adecuado es crucial para el estimado final de la producción diaria de huevos, el mismo que requiere ser analizado y probado, a fin de tratar de reducir la variabilidad del estimado. El diseño de muestreo que se ha estado utilizando hasta ahora en la anchoveta, estaba basado en función al patrón de distribución de los huevos observado entre los años 1964 y 1980, cuando el desove se concentraba principalmente dentro de las 30 mn de la costa. Este 1 método exigía un mayor esfuerzo en la parte costera (dentro de las 30 mn de la costa), y una intensidad de muestreo menor a mayores distancias. Los transectos se fijaron perpendiculares a la costa, con 18 perfiles de 90 mn y 35 de 30 mn; con 10 mn de separación entre sí (Santander et al. 1982, 1984). En los últimos años ha cambiado mucho este patrón de distribución y concentración, y se han observado núcleos importantes de huevos hasta 100 mn de la costa, frente a Punta Chao (AyÓn 2001), lo cual nos lleva a un replanteamiento del diseño así como los cálculos finales para estimar la PDH. Durante el crucero 2003, para estimar la biomasa desovante de la anchoveta del stock nortecentro se utilizó el nuevo trayecto; el estimado de la PDH se hizo utilizando nuevas regiones, permitiendo una mejor estimación de esta producción con un coeficiente de variación aceptable. Dirección de Investigaciones en Oceanografía Biológica, [email protected] 81 MATERIAL Y MÉTODOS En el crucero 0308-09, del 13 agosto al 16 setiembre 2003, se aplicó el método PDH para estimar la biomasa desovante de la anchoveta. Se colectaron 1.166 muestras de ictioplancton, mediante una red CalVET de 27 cm de diámetro de boca, operada en jales verticales desde 70 m de profundidad hacia la superficie, provista de una malla nytal de 300 micras de abertura. Las muestras fueron fijadas en formaldehído al 2%, en frascos de plástico de 300 mL; en el laboratorio se separaron, contaron y estudiaron los huevos de anchoveta. Diseño de muestreo.- El nuevo diseño de muestreo varió sustancialmente en el trayecto. Los perfiles cortos planteados por Santander et al. (1984) fueron reemplazados por perfiles largos, que podrían extenderse o cortarse de acuerdo a la presencia de los huevos de anchoveta (Figura 1). Informe IMARPE Vol. 35(2): Abril-Junio 2008 Pto. Pizarro Pto. Pizarro Pta. Sal Pta. Sal Talara Talara 5°S 5°S Paita Paita Pta. La Negra Pta. La Negra Mórrope Mórrope Pimentel 7°S Pimentel 7°S Chérrepe Chérrepe Chicama Chicama Salaverry Salaverry Pta. Chao Pta. Chao 9°S 9°S Chimbote Chimbote Casma Casma Pta. Lobos Pta. Lobos Huarmey Huarmey Pta. Bermejo Pta. Bermejo Supe 11°S Supe 11°S Huacho Huacho Chancay Chancay Callao Callao Pucusana Pucusana 13°S Cerro Azul 13°S Cerro Azul Tambo de Mora Tambo de Mora Pisco 84°W Pta. Infiernillos 82°W 80°W 78°W 76°W 15°S B. Independencia b) B. Independencia a) 15°S Pisco 84°W Pta. Infiernillos 82°W 80°W 78°W 76°W Figura 1.- Estaciones de muestreo: a) Diseño tradicional, b) Diseño modificado. Pto. Pizarro Pto. Pizarro Pta. Sal 3 5˚S Pta. Sal Talara 2 1 Talara 1 5˚S Paita Pta. La Negra Paita Pta. La Negra M—rrope M—rrope Pimentel 7˚S 13 12 11 2 7˚S ChŽrrepe Pimentel ChŽrrepe Chicama Chicama 6 Salaverry Salaverry 3 Pta. Chao 9˚S 9˚S Chimbote 23 22 21 Chimbote Casma Casma Pta. Lobos Pta. Lobos Huarmey Huarmey 4 Pta. Bermejo Pta. Bermejo Supe 11˚S Pta. Chao Supe 11˚S Huacho Huacho Chancay Chancay Callao Callao 31 13˚S 33 32 a) 15˚S 84˚W 7 Pucusana 13˚S Cerro Azul Cerro Azul Tambo de Mora Pisco Pisco b) Pta. Infiernillos 80˚W 78˚ W Pucusana Tambo de Mora B. Independencia 82˚ W 5 76˚W 15˚S B. Independencia Pta. Infiernillos Pta. Caballas 84˚ W 82˚ W 80˚ W Figura 2.- Distribución de las regiones: a) Rígidas, b) Móviles. 82 78˚ W 76˚ W Producción diaria de huevos de anchoveta en invierno 2003 Ayón Pto. Pizarro Pto. Pizarro Pta. Sal Pta. Sal Talara 5°S Talara 5°S Paita Paita Pta. La Negra Pta. La Negra Mórrope Mórrope Pimentel 7°S Pimentel 7°S Chérrepe Chérrepe Chicama Chicama Salaverry Salaverry Pta. Chao 9°S Pta. Chao 9°S Chimbote Chimbote Casma Casma Pta. Lobos Pta. Lobos Huarmey Huarmey Pta. Bermejo Pta. Bermejo Supe 11°S Supe 11°S Huacho Huacho Chancay Chancay Callao Callao Pucusana Pucusana 13°S Cerro Azul 13°S Cerro Azul Tambo de Mora Tambo de Mora Pisco Pisco B. Independencia B. Independencia Pta. Infiernillos Pta. Infiernillos 15°S Pta. Caballas 84°W 82°W 80°W 78°W 15°S 76°W Pta. Caballas 84°W 82°W 80°W 78°W 76°W Figura 3.- Distribución de huevos de anchoveta: a) Diseño mdificado, b) Diseño tradicional (Cr. 0308-09) Pto. Pizarro Pto. Pizarro Pta. Sal Pta. Sal Talara 5°S Talara Paita 5°S Paita Pta. La Negra Pta. La Negra Mórrope Pimentel 7°S Mórrope 4001 Pimentel 7°S Chérrepe Chérrepe Chicama Chicama Salaverry 3001 1001 huevos/m2 2001 Salaverry Pta. Chao 9°S Pta. Chao 9°S Chimbote Chimbote Casma Pta. Lobos Casma Pta. Lobos Huarmey Huarmey Pta. Bermejo Pta. Bermejo Supe 11°S Supe 11°S Huacho Huacho Chancay Chancay Callao Callao 1 Pucusana 13°S Pucusana Cerro Azul 13°S Cerro Azul Tambo de Mora Pisco a) Tambo de Mora Pisco b) B. Independencia B. Independencia Pta. Infiernillos 15°S 84°W 82°W 80°W 78°W Pta. Infiernillos 15°S 76°W Pta. Caballas 84°W 82°W 80°W 78°W Figura 4.- Distribución y abundancia de huevos de anchoveta: a) Diseño Nuevo, b) Diseño antiguo (Cr. 0308-09). 83 76°W Informe IMARPE Vol. 35(2): Abril-Junio 2008 Tabla 2.- Valores de las categorias de edad, producción diaria de huevos (Po), tasa de mortalidad (Z) y sus coeficientes de variación (CV), de las regiones en los dos tipos de diseño de muestreo. Muestreo tradicional Categorías de edad Regiones rígidas S A1 A2 B1 B2 C1 C2 Po (huevos.dia -1 ) Muestreo nuevo Regiones móviles Regiones rígidas Regiones móviles 0.848E+12 2.862E+12 1.369E+12 0.985E+12 0.197E+12 0.372E+12 0.001E+12 0.709E+12 2.621E+12 1.510E+12 1.216E+12 0.343E+12 0.282E+12 0.0013E+12 0.709E+12 3.005E+12 1.618E+12 1.452E+12 0.328E+12 0.252E+12 0.0013E+12 0.700E+12 2.647E+12 1.580E+12 1.239E+12 0.385E+12 0.194E+12 0.0016E+9 4.820E+12 3.990E+12 4.600E+12 4.000E+12 CV de P o (%) 13.79 15.195 19.661 13.905 z (huevos.dia -1 ) CV de z (%) % 1.311 15.896 73.045 1.026 19.542 64.156 1.054 24.958 65.163 0.997 18.129 63.12 La separación entre los perfiles se mantuvo en 10 mn. Sin embargo, en el desarrollo del crucero la distancia entre algunos perfiles fue de 12,5 mn debido a factores de tiempo. Se estableció una separación de 4 mn entre las estaciones dentro de las 30 mn; y por fuera de las 30 millas esta distancia fue de 5 millas. Edad de los huevos.- Se identificaron los 11 estadios de desarrollo de huevos, de acuerdo a sus características morfológicas; y la asignación de la edad de los huevos estuvo en función de la hora y temperatura superficial del mar, en el momento de su colecta (Santander et al. 1984). Número total de huevos por categoría de edad.- Se obtuvo utilizando nuevas regiones, denominadas “regiones móviles” (Figura 2b), las cuales estuvieron definidas por el patrón de distribución de los huevos que se muestra en la Figura 3. Para fines comparativos, se obtuvo el número total de huevos por categoría de edad basado en las “regiones rígidas” planteadas por Santander et al. (1984) (Figura 2a). Estimado de la producción diaria de huevos.- Para el cálculo final de la producción de huevos se usaron las categorías A1, A2, B1 y B2; en este cálculo no se incluyeron la categoría S ni C, estas dos categorías no se consideraron por estar submuestreadas (Santander et al. 1984). El tiempo de cada categoría corresponde a 12 horas, equivalente a medio día (0,5 días) (Tabla 1). Tabla 1. Tiempo de las categorías de edad Categoría Tiempo (día) A1 A2 B1 B2 0,1667 0,6667 1,1667 1,6667 La producción diaria de huevos (Po) se obtuvo mediante un programa iterativo a partir de la ecuación de Picquelle y Stauffer (1985), donde el estimado de la producción diaria de huevos depositada en el mar (P), por la biomasa desovante de peces, es derivada por una regresión del conteo de los huevos en sus edades, usando un modelo de mortalidad exponencial, donde se asume una tasa de mortalidad constante. Pi = Po e -z ti Donde: Pi = Producción de huevos en la categoría i Po = Producción diaria de huevos 84 Zti = mortalidad en el tiempo i Para fines de comparación entre el nuevo diseño y el de Santander et al. (1984), se efectuó un análisis con las estaciones que se encontraban dentro de los perfiles, de acuerdo con el criterio de dicha autora. Con estas estaciones se elaboraron nuevas cartas de distribución de los huevos, abundancia y cálculos de los valores de las categorías de edad así como la producción diaria de huevos. RESULTADOS Distribución de los huevos de anchoveta.- Durante el invierno, frente al área comprendida entre caleta La Cruz y Bahía Independencia, los huevos se distribuyeron en dos zonas, una costera dentro de las primeras 15 mn de la costa entre Talara y Pisco; y otra por fuera de las 40 mn de la costa entre Pimentel y Callao (Figura 3). Se observaron varios núcleos de concentración de huevos ubicados tanto en la parte costera como oceánica. Entre Pimentel y Huacho, lejos de la costa se observó una distribución homogénea, y en la parte costera se apreciaron pequeños parches aislados (Figura 4). La frecuencia de estaciones positivas fue de 36,34%, con abundancias que oscilaron entre 20 y 24180 huevos/ m². Categorías de los huevos de anchoveta.El número de huevos por categoría de edad y la producción diaria de huevos hallados para este crucero se presentan en la Tabla 2. Producción diaria de huevos.- La producción diaria de huevos de acuerdo con el diseño tradicional (Santander et al. 1984) en las regiones rígidas fue equivalente a 4,82 E+12 y un CV de 13,79; en las regiones móviles fue de 3,99 E+12 y un CV de 15,19% Utilizando el nuevo diseño el estimado de la producción diaria de huevos con las regiones rígidas (Santander et al. 1984) fue de 4,6 E+12 huevos.día-1 con un Producción diaria de huevos de anchoveta en invierno 2003 Ayón coeficiente de variación de 19,66%; para las regiones móviles fue de 4,0 E+12 huevos.día-1 con un coeficiente de variación de 13,9% (Tabla 2). DISCUSIÓN El área de distribución de huevos obtenido con el nuevo diseño es similar al observado con el diseño tradicional (Figura 1). Sin embargo, se nota que con el diseño tradicional la interpolación genera áreas con mayor densidad de huevos, sobretodo en la zona oceánica (Figura 2). Esto se debe a que la resolución es menor y los puntos referenciales tienen una mayor distancia (30 mn); pero con el nuevo diseño se puede mejorar la resolución entre 10 y 12,5 mn. En los cálculos de la producción diaria de huevos, los mayores valores se han obtenido utilizando las regiones rígidas en ambos diseños de muestreo; sin embargo, es mucho mayor la producción de huevos con el muestreo tradicional. Los valores de la producción diaria de huevos con las regiones móviles son parecidos para ambos diseños, marcándose la diferencia solamente en los valores de los coeficientes de variación, siendo menor el del nuevo diseño. Comparando estos resultados, debemos analizar y ver cuál de estos valores está reflejando mejor el valor de la producción diaria de huevos en el momento del muestreo. Por un lado, se puede considerar que el valor más alto con un coeficiente de variación bajo podría ser el aceptado; sin embargo, de acuerdo a lo encontrado en la interpolación se habría dado mayor peso a zonas donde la abundancia de huevos era menor; así mismo, cuando se consideran regiones rígidas no se toma en consideración la formación de los parches, que es una característica propia de los componentes del plancton, en este caso los huevos de anchoveta. Es interesante ver como utilizando dos diseños de muestreo diferentes aplicando el mismo criterio de las regiones móviles (de acuerdo a su distribución) se obtiene un valor de la producción diaria de huevos similar, la única diferencia es que con el método nuevo el CV es menor. Esto estaría determinando que el mejor valor para la PDH es el obtenido mediante el nuevo muestreo y empleando las regiones móviles. Es importante mencionar que el tiempo de barco empleado para este nuevo muestreo es igual al que se estuvo utilizando para el muestreo tradicional. La diferencia radica en que se obtiene una mayor intensidad de muestreo en la parte oceánica, incrementando la resolución de la interpolación y ajustando mejor el estimado de la PDH. Asimismo, utilizando este nuevo trayecto, se hace compatible con el trayecto, para hacer la evaluación mediante el método acústico y estimar la biomasa de anchoveta de manera simultánea a la aplicación del MPH. CONCLUSIONES El nuevo diseño de muestreo permite obtener información homogénea por fuera de las 30 mn, lo cual mostró la importancia del desove en la zona Pimentel - 85 Huacho a esa distancia de la costa. Además con el nuevo diseño es posible realizar la evaluación de la anchoveta simultáneamente a la evaluación acústica de su biomasa Agradecimientos.- Al personal del Área de Evaluación de Producción de Huevos, quienes colectaron y analizaron las muestras para el presente trabajo. A la Blga. Soledad Guzmán por la revisión del manuscrito y sus comentarios. REFERENCIAS AyÓn P. 2001. El ictioplancton en el mar peruano durante el verano 2000. Inf. Inst. Mar Perú 159:73-84. Picquelle S, Stauffer G. 1985. Parameter estimation for an egg Production Method of Northern Anchovy Biomass assesment. Reuben Lasker (ed.) In: An egg Production method for estimating spawning biomass of pelagic fish: application to the northern anchovy, Engraulis mordax. NOAA Technical report NMFS 36. Santander H, Smith P, Alheit J. 1982. Determinación del esfuerzo de muestreo requerido para el estimado de producción diaria de huevos de anchoveta Engraulis ringens, frente al Perú. Bol. Inst. Mar Perú 7(1):1-20. Santander H, Alheit J, Smith P. 1984. Estimación de la biomasa de la población desovante de anchoveta peruana Engraulis ringens en 1981 por aplicación del Método de Producción de Huevos. Bol. Inst. Mar Perú 8(6):209-250. Smith P, Santander H, Alheit J. 1983. Detalles técnicos de un plan de crucero para estimar la biomasa desovante de anchoveta. Bol. Inst. Mar Perú 7(2):21-52. Evaluación hidroacústica de anchoveta y otros pelágicos en invierno 2003 Castillo, Peraltilla y Segura EVALUACIÓN HIDROACÚSTICA DE ANCHOVETA Y OTROS RECURSOS PELÁGICOS. AGOSTO Y SETIEMBRE 2003 PERUVIAN ANCHOVY AND OTHER PELAGIC RESOURCES HYDROACOUSTIC EVALUATION. AUGUST AND SEPTEMBER 2003 Ramiro Castillo Salvador Peraltilla Marceliano Segura RESUMEN Castillo R, Peraltilla S, Segura M. 2008. Evaluación hidroacústica de anchoveta y otros recursos pelágicos. Agosto y setiembre 2003. Inf. Inst. Mar Perú. 35(2): 87-95.- El crucero BIC Olaya y SNP2 0308-09, para evaluación de la biomasa de anchoveta por el método de producción de huevos (MPH), del 13 agosto al 16 setiembre 2003, entre caleta La Cruz y Punta Carretas, recorrió de 0,5 a 120 mn de la costa, pero llegó hasta 150 mn de caleta La Cruz a Punta Sal. El equipo utilizado fue la ecosonda EK-500 Simrad principalmente en la frecuencia de 38 kHz. El muestreo fue el establecido por el IMARPE: de 1,0 mn en trayectos sistemáticos, perpendiculares a la línea de costa y paralelos, separados de 10 a 12,5 mn entre sí. La evaluación directa se realizó por metodología hidroacústica; la distribución se determinó por interpolación de datos; y la biomasa, por el método de estratificación por áreas isoparalitorales de 10´x30´. La anchoveta (Engraulis ringens, 8.949.966 t) fue la especie predominante con mayor abundancia entre Pimentel y Salaverry; la samasa (Anchoa nasus, 565.614 t) sobre todo entre Chérrepe a Chicama; el bagre (Galeichthys peruvianus, 480.322 t) principalmente entre Pimentel y Chimbote; la vinciguerria (Vinciguerria lucetia, 1.392.846 t) en áreas alejadas de la costa: Punta Sal, Punta La Negra, Callao, Pisco y Bahía Independencia; la pota (Dosidicus gigas, 357.243 t) dispersa de puerto Pizarro a punta Gobernador, Chérrepe a Huacho y entre Pucusana a Tambo de Mora; estas dos últimas se hallaron junto a otras especies mesopelágicas con distribución vertical hasta los 380 metros; la múnida (Pleuroncodes monodon, 597.099 t) ha disminuido en relación a los últimos cruceros. Palabras clave: anchoveta, hidroacústica, distribución, biomasa, recursos pelágicos, invierno 2003. ABSTRACT Castillo R, Peraltilla S, Segura M. 2008. Peruvian anchovy and other pelagic resources hydroacoustic evaluation. August and September 2003. Inf Inst Mar Peru. 35 (2): 87-95 .- The cruise BIC Olaya and SNP2 0308-09, to evaluate the anchoveta spawning biomass using the method of eggs production (EPM), from August 13th to September 16th 2003, between caleta La Cruz and Punta Carretas, from 0.5 to 120 mn off the coast, reached to 150 nm from caleta La Cruz to Punta Sal. The equipment used was the ecosounder SIMRAD EK-500 primarily in the frequency of 38 kHz. The sampling was that established by IMARPE: 1.0 mn in systematic trips, perpendicular to the shoreline and parallel, separated 10 to 12.5 mn between them. Direct testing was performed by hydroacoustic method; the distribution was determined by interpolation of data; and biomass, by the method of stratification on 10’x30’ isoparalitoral areas. The Peruvian anchovy (Engraulis ringens, 8.949.966 t) was the predominant species with greater abundance between Pimentel and Salaverry; the longnose anchovy (Anchoa nasus, 565,614 t), especially among Chérrepe to Chicama; the catfish (Galeichthys peruvianus, 480,322 t) mainly between Pimentel and Chimbote; the (Vinciguerria lucetia, 1,392,846 t) in areas away from the coast: Punta Sal, Punta La Negra, Callao, Pisco and Independence Bay; the giant squid (Dosidicus gigas, 357,243 t), spread from Puerto Pizarro to Punta Gobernador, Chérrepe to Huacho and between Pucusana to Tambo de Mora; these latter two were found together with other mesopelagic species with vertical distribution to the 380 meters, the munida (Pleuroncodes monodon, 597,099 t) has decreased compared to the previous cruises. Keywords: Peruvian anchovy, hydroacoustic, distribution, biomass, pelagic resources, winter 2003. INTRODUCCIÓN MATERIAL Y MÉTODOS Entre los objetivos del crucero 030809, para evaluación de biomasa desovante de la anchoveta, mediante del método de producción de huevos (MPH), se propuso realizar la evaluación de los recursos pelágicos importantes, con el método directo hidroacústico. El diseño de muestreo utilizado en este crucero ha permitido cubrir casi totalmente la distribución de anchoveta, entre caleta La Cruz y punta Carretas. En el crucero BIC Olaya y SNP2 0308-09, se realizó del 13 agosto al 16 setiembre 2003. En el recorrido entre caleta La Cruz y Pucusana, el BIC SNP-2 evaluó el área costera desde 0,5 a 32 mn de la costa y el BIC Olaya, desde 32 mn hasta 150 mn de la costa. Hacia el sur desde Pucusana a punta Carretas, el BIC Olaya evaluó desde 0,5 hasta 90 mn de la costa. El muestreo utilizado fue sistemático paralelo, 87 es decir, en trayectos paralelos y perpendiculares a línea de la costa; la separación entre transectos fue desde 10 a 12,5 mn. El intervalo del muestreo fue de 1,0 mn en todos los trayectos (Figura 1). El equipo utilizado fue la ecosonda científica Simrad EK-500 principalmente en la frecuencia de 38 kHz. El muestreo vertical fue desde 0 a 150 m (BIC SNP-2) y de 0 a 500 m (BIC Olaya). Previamente al inicio del crucero, se realizaron las cali- Informe IMARPE Vol. 35(2): Abril-Junio 2008 Figura 1.- Carta de trayectos y lances realizados en el crucero MPH 0308-09 Figura 2.- Distribución de la anchoveta Figura 3.- Distribución del jurel. braciones acústicas de las ecosondas, así como las intercalibraciones entre los equipos de ambos barcos. En total se muestrearon 6.543 mn de rastreo acústico (4.282 mn el BIC Olaya y 2.261 mn el SNP-2) en Figura 4.- Distribución de la caballa. 63 perfiles o transectos. Hubo 103 lances de pesca de comprobación. Para el tratamiento de los datos acústicos se utilizó un programa de post-procesamiento de señales acústicas, principalmente para la disgregación de los valores 88 ecointegrados y grabación de ecos obtenidos durante el crucero. La discriminación se efectuó de acuerdo a los porcentajes de las capturas realizadas por los lances de arrastre pelágico e identificación de registros típicos de especies. Evaluación hidroacústica de anchoveta y otros pelágicos en invierno 2003 Castillo, Peraltilla y Segura Figura 6.- Distribución del bagre Figura 5.- Distribución de la samasa. Figura 8.- Distribución de la pota. Figura 7.- Distribución de la vinciguerria. Para delimitar el área de distribución o mapeo de los recursos se realizó la interpolación de datos con el método krigging. La estimación de biomasa se ejecutó por estratificación isoparalitoral. La formulación está descrita en MacLennan and Simmonds (1992). Las ecuaciones de fuerza del blanco (TS) utilizadas para la frecuencia de 38 kHz, fueron: Anchoveta: 89 TS = 20 Log L – 78,90 dB TS = 20 Log L – 77,35 dB para ejemplares mayores de 13,5 cm Samasa: TS = 20 Log L – 78,90 dB Informe IMARPE Vol. 35(2): Abril-Junio 2008 Bagre: TS = 20 Log L – 78,90 dB Vinciguerria: TS = 20 Log L – 82,00 dB Pota: TS = 31,2 Log L – 83,15 dB Múnida: TS = 20 Log L – 95,40 dB La relación longitud-peso para cada especie se determinó a partir de los datos biométricos de los lances de pesca (Chipollini et al.): Anchoveta: Samasa: Bagre: Vinciguerria: Pota: Múnida: w = 0,009 L 2,86 entre los 04° y 09° S w = 0,005 L 3,0 entre los 10° y 14° S Casma tuvo distribución continua y amplia, asociada a la presencia de las ACF; y desde Huarmey a Tambo de Mora su distribución fue discontinua y replegada hacia la costa, relacionada al acercamiento hacia la costa de las ASS. La mayor abundancia y concentración se localizó entre Pimentel y Salaverry desde las 15 a 75 mn de la costa, hacia el sur de Salaverry se encontró principalmente dispersa (Figura 2). Verticalmente, se detectó entre 3 y 70 m de profundidad; al norte de Mórrope alcanzó mayor profundidad y hacia el sur sólo hasta 40 m, sobre todo en las ACF. La biomasa estimada fue de 8.949.966 t con LC de 9,77%. El mayor valor (5.019.200 t) se registró entre Pimentel y Salaverry (7° a 8°30´S). Dentro de las 50-60 mn de la costa se halló el 24,25% de la biomasa (2.164.819 t) y dentro de 0-10 mn, el 23,2% (2.077.692 t) (Tabla 1). w = 0,0054 L 3,105 w = 0,023 L 2,7279 w = 0,0118 L 2,476 w = 0,0004 L 2,9364 w = 0,0005 L 2,8784 RESULTADOS Anchoveta, Engraulis ringens.Fue la especie dominante desde Talara a Tambo de Mora, desde 1,0 hasta 120 mn de distancia a la costa (entre Punta Chao y Chimbote). Desde Mórrope a Jurel, Trachurus murphyi.- Fue muy escaso y disperso; se detectó en pequeños cardúmenes aislados entre 3 a 45 m de profundidad, en áreas alejadas entre 55 y 92 mn de la costa entre Cerro Azul y Pisco (Figura 3). Caballa, Scomber japonicus.- Fue muy escasa, replegada a la costa de Supe hasta 10 mn (Figura 4). Samasa, Anchoa nasus.- Estuvo replegada hasta 28 mn de la costa, entre Talara a Chicama, y dispersa frente a Chancay. Su biomasa total fue 565.614 t, con LC 24,68%. La mayor concentración y abundancia se encontró entre Chérrepe y Chicama (258.419 t) (Figura 5, Tabla 2). Normalmente se registró mezclada con otras especies costeras, entre 3 y 48 m de profundidad. Bagre, Galeichthys peruvianus.Se halló en áreas aisladas frente a Sechura, Chérrepe, Chicama, Salaverry, Chimbote y sur del Callao (Chorrillos). Su distribución tuvo mayor amplitud, en Salaverry hasta 30 mn de la costa, y mayor concentración en pequeños núcleos frente a Chimbote y Salaverry (Figura 6), entre 3 y 65 m de profundidad. Disminuyó su población en comparación con el verano 2003. Su biomasa fue de 480.322 t, con límite de confianza de 25,34%. La mayor abundancia se registró en Salaverry y Chimbote (8°S) con 263.928 t (Tabla 3). Tabla 1.- Biomasa latitudinal y por distancia a la costa de la anchoveta TOTALES (t) TOTAL (%) 120-130 110-120 100-110 90-100 80-90 Distancia de la costa (mn) 70-80 60-70 50-60 40-50 30-40 20-30 10-20 0-10 Grado de latitud sur 03° 30 - 04° 00 03°30' - 04° 04° 00 - 04° 30 95 570 95 570 112 289 36 446 148 735 21 536 50 070 71 606 1 357 379 1 914 913 3 272 291 1 746 909 882 601 2 629 510 436 056 579 636 1 015 693 387 613 379 001 766 614 355 001 184 445 539 446 164 755 220 759 385 514 22 866 2 120 24 987 95 570 1.07 1.66 1 805 0.80 1 805 357 36.56 2 612 29 400 11.35 29 400 29 298 29 298 7 269 22 649 22 649 04° - 05° 05° 00 - 05° 30 36 446 05° 30 - 06° 00 148 735 05° - 06° 21 536 06° 00 - 06° 30 18 347 06° 30 - 07° 00 39 883 06° - 07° 435 034 233 158 185 678 93 315 386 526 23 311 07° 00 - 07° 30 284 072 479 495 280 087 401 845 325 084 144 331 07° 30 - 08° 00 357 719 107 712 653 465 764 495 160 711 610 162 866 729 028 383 775 172 760 133 822 44 784 117 262 08° 00 - 08° 30 43 084 78 695 64 374 22 353 5 567 2 855 9 457 656 217 08° 30 - 09° 00 43 084 241 561 793 403 2 612 29.38 7 269 04° 30 - 05° 00 95 570 112 289 167 641 07° - 08° 406 127 178 327 136 677 54 241 773 480 08° - 09° 113 200 58 026 29 449 27 072 47 330 36 020 12 207 5 971 370 47 712 09° 00 - 09° 30 813 22 633 51 160 146 457 143 858 190 067 24 086 533 30 114 014 80 659 80 608 173 529 191 188 226 087 36 292 6 504 401 241 096 8.57 19 326 220 026 19 326 461 122 28 714 47 712 09° - 10° 117 803 10° 00 - 10° 30 62 013 10° 30 - 11° 00 45 583 32 053 45 583 60 767 179 816 10° - 11° 77 891 61 883 215 228 11° 00 - 11° 30 46 384 6.03 09° 30 - 10° 00 124 274 4.31 29 561 9 735 15 237 91 444 9 735 15 237 298 755 11° - 12° 34 839 5 584 9 546 114 785 83 527 12° 00 - 12° 30 7 317 24 540 42 157 30 124 188 903 12° 30 - 13° 00 9 546 303 688 12° - 13° 456 22 411 13° 00 - 13° 30 2 120 0.28 11° 30 - 12° 00 2 576 13° 30 - 14° 00 22 411 13° -14° 14° 00 - 14° 30 14° 30 - 15° 00 14° -15° TOTALES Porcentaje 29 400 100.00 0.33 29 298 0.33 114 014 1.27 83 271 0.93 123 692 434 772 1.38 4.86 90 2 164 819 24.19 1 514 725 16.92 876 556 9.79 685 468 7.66 816 260 9.12 2 077 692 23.21 8 949 966 toneladas Evaluación hidroacústica de anchoveta y otros pelágicos en invierno 2003 Castillo, Peraltilla y Segura Vinciguerria, Vinciguerria lucetia.- Se encontró en núcleos dispersos, en diversas áreas alejadas de la costa. Sus registros fueron casi continuos de Talara a Pimentel y de Huarmey a Cerro Azul; y discontinuos de Chérrepe a Casma y de Puerto Pizarro a Punta Sal. Escasas áreas de alta concentración se ubicaron frente a: Punta Sal, Punta La Negra, Callao, Pisco y bahía Independencia (Figura 7). Compartió áreas con otros recursos mesopelágicos como mictófidos y salpas. Su distribución se relacionó con aguas de mezcla ACF-AES al norte de Punta La Negra, y ACF-ASS al sur de Pimentel. Verticalmente, abarca una amplia distribución de 3 a 380 m; fue superficial en las horas de la noche y alcanzó mayor profundidad en las horas del día. La biomasa fue de 1.392.846 t, límite de confianza 15,98% (máximo 1.615.434 t, y mínimo 1.170.258 t). La mayor abundancia se encontró a los 12°S (326.827 t) y 3°S (221.863 t). A 50 mn de la costa se hallaron 467.227 t ( 31,53 %) y a las 70 mn 1.269.736 t ( 29,15 %) (Tabla 4). Figura 10.- Área de distribución de la anchoveta por grado de latitud a partir del crucero 0208 de MPH. Calamar gigante, Dosidicus gigas.La distribución de la pota, jibia o calamar gigante fue principalmente dispersa, se encontró alejada de la costa; en muchas ocasiones mezclada con otros recursos mesopelágicos, especialmente con la vinciguerria. Sus áreas se localizaron entre Puerto Pizarro a Punta Gobernador, Chérrepe a Huacho y entre Pucusana a Tambo de Mora. Al igual que la vinciguerria su distribución se ubicó en las aguas de mezcla entre las AES-ACF y ASS-ACF. Por los registros observados, es posible que su distribución continúe más allá del área evaluada (Figura 8). Verticalmente, se localizó de 3 a 360 m, superficial durante la noche y a mayor profundidad en el día. La biomasa estimada fue de 357.243 t con límite de confianza ± 9,7%. La mayor abundancia se localizó a los 7°S (67.101 t). A la distancia de 60 mn de la costa se halló el 21,93% de la biomasa total; y entre 80 y 90 mn, el 21,52% (Tabla 5). Figura 11.- Distribución de la anchoveta registrada en los cruceros realizados en el 2003. 91 Informe IMARPE Vol. 35(2): Abril-Junio 2008 Múnida, Pleuroncodes monodon.Se encontró en aguas costeras frías (ACF) y con una menor biomasa que en años anteriores. Su distribución fue continua frente al área entre Chérrepe y Chimbote, y de Chancay a Pucusana; con concentraciones densas, especialmente frente a Chérrepe, Chicama, Salaverry y Callao. Se registraron otras áreas menores frente a Casma y Cerro Azul. Alcanzó hasta las 65 mn de la costa y una profundidad de 65 m; en muchas ocasiones se encontró mezclada con la anchoveta (Figura 9) y a veces, durante el día, por debajo de los cardúmenes de anchoveta. La biomasa estimada fue de 597.099 t, con límite de confianza de 18,75% (máximo 709.078 t; mínimo 485.121 t). El mayor valor se encontró a los 7°S (375.792 t; 62,94% de su biomasa total). Por distancia a la costa se encontraron que entre 20 a 30 mn (268.562 t; 44,98%; y 10 a 20 mn (178.732 t; 29,93% de la biomasa) (Tabla 6). Figura 12.- Abundancia latitudinal de anchoveta por grado de latitud según cruceros. Tabla 2.- Biomasa latitudinal y por distancia a la costa de la samasa TOTALES (t) TOTAL (%) 50-60 40-50 D istancia de la costa (mn) 30-40 20-30 10-20 0-10 03°30' - 04° 04° 00 - 04° 30 41 634 41 634 41 634 88 510 5 712 7.36 94 222 132 257 38 183 16.66 170 441 65 627 192 792 30.13 258 419 45.69 Con respecto a la distribución de la anchoveta, debe mencionarse que en el crucero MPH 0208-09, la distribución alcanzó hasta las 125 mn de la costa, preferencialmente dispersa. Otros cruceros de evaluación hidroacústica: 0209-11, 0302-04 y 0305-06 corroboraron la abundancia de anchoveta. En el crucero 0209-11, de Paita y San Juan su distribución estuvo limitada por TSM de 19 °C, ACF y aguas de mezcla ACF + ASS, y en el norte con las AES. En el crucero 0302-04 se distribuyó cercana a 04° 30 - 05° 00 41 634 04° - 05° 88 510 05° 00 - 05° 30 5 712 05° 30 - 06° 00 94 222 05° - 06° 132 257 06° 00 - 06° 30 06° 30 - 07° 00 172 2 796 35 215 172 2 796 167 472 06° - 07° 65 627 07° 00 - 07° 30 102 901 89 891 07° 30 - 08° 00 102 901 155 518 07° - 08° 08° 00 - 08° 30 08° 30 - 09° 00 08° - 09° 09° 00 - 09° 30 DISCUSIÓN El diseño de muestreo utilizado en este crucero (transectos perpendiculares a la línea de la costa y paralelos entre si) han sido favorables para la aplicación del método hidroacústico, especialmente en la intensidad del muestreo de barrido (separación entre transectos de 10 y 12 mn); sin embargo, en los muestreos de las capturas de comprobación de registros no fue ideal, quizás debido al tiempo previsto para la ejecución de este crucero. Grado de latitud sur 03° 30 - 04° 00 09° 30 - 10° 00 09° - 10° 10° 00 - 10° 30 10° 30 - 11° 00 10° - 11° 522 376 898 0.16 522 11° 00 - 11° 30 376 11° 30 - 12° 00 898 11° - 12° 12° 00 - 12° 30 12° 30 - 13° 00 12° - 13° 13° 00 - 13° 30 13° 30 - 14° 00 13° -14° 14° 00 - 14° 30 14° 30 - 15° 00 14° -15° TOTALES Porcentaje 100.00 la costa de Talara a Morro Sama. En el crucero 0305-06, entre Paita y Salaverry prevalecieron las aguas de mezcla (ACF y ASS) y la anchoveta se concentró frente a Talara y Pimentel con gran abundancia. En este crucero MPH (0308-09) las condiciones oceanográficas han propiciado que la anchoveta se desplace hacia a las ACF y aguas de mezcla ACF + ASS, especialmente frente al área entre Mórrope y Casma, hasta las 120 mn de distancia a la 92 172 105 696 459 745 0.03 18.69 81.28 565 614 toneladas costa. Hacia el norte de Punta Aguja, este recurso se localizó debajo de los 10 m en ACF. Hacia el sur de Casma fue costera en la capa superficial de los 40 metros. Sin embargo, estas variaciones en su distribución no han sido significativas, siempre se hallaron en la zona norte (Figuras 10 y 11). La captura de anchoveta en el 2002 fue de 7,90 millones de toneladas. En mayo y junio 2003 se capturaron 1.037.467 y 567.909 t, respectivamente, cifra decreciente por la veda temporal y por la Evaluación hidroacústica de anchoveta y otros pelágicos en invierno 2003 Castillo, Peraltilla y Segura dispersión del recurso debido a la ampliación de las ACF en la franja costera, propias de estos meses del año, época en que la anchoveta incrementa su área de distribución hacia las zonas mas alejadas de la costa. La estimación de anchoveta en el crucero 0302-04 fue de 7,4 millones de toneladas, y en el crucero 0305-06 resultaron 5,8 millones, a pesar que no se evaluó totalmente su distribución. En este crucero MPH 0308-09 se ha estimado en 8,9 millones de toneladas; se evaluó totalmente su abundancia, con un muestreo adecuado (intertransectos de 10 y 12,5 mn) en el cual se obtuvo 9,77 % de límite de confianza, es decir con un alto nivel de confiabilidad en el estimado de biomasa. Después del evento El Niño 9798, las mayores poblaciones de anchoveta se han encontrado en las zonas norte y centro, con altas concentraciones, especialmente después del límite hacia el norte de la presencia de la múnida (Figura 12). Tabla 3.- Biomasa latitudinal y por distancia a la costa del bagre. TOTALES (t) TOTAL (%) 50-60 40-50 D istancia de la costa (mn) 30-40 20-30 10-20 Grado de latitud sur 0-10 03° 30 - 04° 00 03°30' - 04° 04° 00 - 04° 30 04° 30 - 05° 00 04° - 05° 05° 00 - 05° 30 9 735 9 735 9 735 2.03 05° 30 - 06° 00 9 735 05° - 06° 06° 00 - 06° 30 483 483 483 18 221 18 476 0.10 36 697 112 599 151 329 7.64 263 928 64 607 54.95 64 607 13.45 06° 30 - 07° 00 483 9 551 06° - 07° 8 670 07° 00 - 07° 30 18 476 07° 30 - 08° 00 9 551 27 146 07° - 08° 52 518 20 110 39 970 08° 00 - 08° 30 13 481 38 158 99 690 08° 30 - 09° 00 66 000 58 268 139 661 08° - 09° 64 607 09° 00 - 09° 30 09° 30 - 10° 00 64 607 09° - 10° 10° 00 - 10° 30 10° 30 - 11° 00 10° - 11° 11° 00 - 11° 30 11° 30 - 12° 00 11° - 12° 104 871 104 871 12° 00 - 12° 30 12° 30 - 13° 00 104 871 21.83 104 871 12° - 13° 13° 00 - 13° 30 13° 30 - 14° 00 13° -14° 14° 00 - 14° 30 14° 30 - 15° 00 14° -15° TOTALES 100.00 Porcentaje 66 000 67 819 346 503 13.74 14.12 72.14 480 322 toneladas Tabla 4.- Biomasa latitudinal y por distancia a la costa de la vinciguerria. TOTALES (t) TOTAL (%) 221 863 221 863 15.93 D istancia de la costa (mn) 130-140 120-130 110-120 100-110 90-100 80-90 70-80 60-70 50-60 40-50 40 698 40 708 10 644 92 898 7 673 15 140 14 102 40 698 40 708 10 644 92 898 7 673 15 140 14 102 30-40 20-30 10-20 0-10 Grado de latitud sur 03° 30 - 04° 00 03°30' - 04° 04° 00 - 04° 30 11 703 11 703 9 538 180 348 0.84 189 886 26 219 9 313 13.63 35 532 3 941 7 541 2.55 11 482 15 188 31 519 0.82 46 707 485 34 987 3.35 35 471 3 442 37 321 2.55 40 763 26 958 73 174 2.93 100 132 275 720 51 108 7.19 326 827 13 587 152 535 23.46 166 123 206 357 11.93 206 357 14.82 7 992 1 473 1 827 411 7 992 1 473 1 827 411 04° 30 - 05° 00 3 967 4 228 1 343 05° 00 - 05° 30 814 05° 30 - 06° 00 04° - 05° 18 066 9 942 82 324 69 201 18 066 9 942 82 324 73 168 4 228 2 157 05° - 06° 1 064 9 586 10 720 4 848 06° 00 - 06° 30 2 793 06° 30 - 07° 00 2 029 2 321 4 199 3 385 13 785 10 720 7 641 06° - 07° 1 912 07° 00 - 07° 30 07° 30 - 08° 00 3 525 1 872 1 249 895 3 525 1 872 3 278 2 807 13 383 1 806 07° - 08° 08° 00 - 08° 30 22 526 6 373 1 634 985 35 909 8 179 1 634 985 08° 30 - 09° 00 08° - 09° 485 09° 00 - 09° 30 485 12 014 22 972 12 014 22 972 09° 30 - 10° 00 09° - 10° 2 712 731 10° 00 - 10° 30 10° 30 - 11° 00 22 378 8 446 6 497 22 378 11 157 7 228 14 746 6 697 10° - 11° 5 515 55 167 15 546 2 462 69 913 22 243 2 462 162 993 112 727 11° 00 - 11° 30 11° 30 - 12° 00 5 515 11° - 12° 12° 00 - 12° 30 23 686 17 271 9 658 493 23 686 180 264 122 385 493 12° 30 - 13° 00 3 878 9 675 136 914 15 621 3 878 146 589 15 621 34 12° - 13° 34 13° 00 - 13° 30 13° 30 - 14° 00 13° -14° 64 004 9 323 3 563 72 983 56 484 14° 00 - 14° 30 64 004 9 323 3 563 72 983 56 484 14° -15° 1 392 846 14° 30 - 15° 00 TOTALES Porcentaje 100.00 40 698 40 708 10 644 39 918 102 949 66 352 321 130 467 227 121 541 36 452 88 742 56 484 2.92 2.92 0.76 2.87 7.39 4.76 23.06 33.54 8.73 2.62 6.37 4.06 93 toneladas Informe IMARPE Vol. 35(2): Abril-Junio 2008 Tabla 5.- Biomasa latitudinal y por distancia a la costa de la pota. TOTALES (t) D istancia de la costa (mn) TOTAL (%) 120-130 110-120 56 504 56 504 41 993 7 027 15.82 49 020 6 208 13.72 100-110 90-100 80-90 70-80 60-70 50-60 40-50 30-40 20-30 10-20 1 753 6 263 27 052 6 518 4 950 3 511 935 2 135 522 2 864 1 753 6 263 27 052 6 518 4 950 3 511 935 2 135 522 2 864 3 613 31 251 6 278 851 4 964 1 427 636 8 577 32 677 6 914 3 115 1 753 1 340 0-10 Grado de latitud sur 03° 30 - 04° 00 03°30' - 04° 04° 00 - 04° 30 04° 30 - 05° 00 851 04° - 05° 05° 00 - 05° 30 05° 30 - 06° 00 6 208 1.74 3 115 1 753 1 340 05° - 06° 06° 00 - 06° 30 06° 30 - 07° 00 06° - 07° 39 458 27 643 67 101 449 34 520 18.78 34 969 2 333 1 986 9.79 4 319 10 241 11 876 1.21 22 117 13 781 6.19 57 57 1 012 25 004 7 924 4 862 10 262 4 595 7 924 5 874 35 265 18 038 55 394 08° 00 - 08° 30 08° 30 - 09° 00 9 913 19 360 3 499 1 045 704 9 913 19 360 3 499 1 100 1 097 170 227 288 985 605 207 973 806 378 1 200 1 094 13 443 07° 00 - 07° 30 07° 30 - 08° 00 07° - 08° 08° - 09° 09° 00 - 09° 30 09° 30 - 10° 00 985 605 262 262 9 888 09° - 10° 7 243 2 737 6 852 5 024 14 095 7 760 10° 00 - 10° 30 10° 30 - 11° 00 10° - 11° 3 894 11° 00 - 11° 30 11° 30 - 12° 00 13 781 3.86 9 888 3 894 5 726 26 553 15 337 5 726 26 553 15 337 11° - 12° 12° 00 - 12° 30 55 790 55 790 34 588 12 847 15.62 47 434 13.28 24 476 5 405 2 768 5 405 2 768 12° 30 - 13° 00 12° - 13° 10 112 13° 00 - 13° 30 12 847 13° 30 - 14° 00 37 323 10 112 13° -14° 14° 00 - 14° 30 14° 30 - 15° 00 14° -15° TOTALES 100.00 Porcentaje 57 12 043 27 084 76 892 20 203 78 359 54 875 35 905 39 334 8 776 3 715 0.02 3.37 7.58 21.52 5.66 21.93 15.36 10.05 11.01 2.46 1.04 357 243 toneladas Tabla 6.- Biomasa latitudinal y por distancia a la costa de la múnida. TOTALES (t) TOTAL (%) D istancia de la costa (mn) 80-90 70-80 60-70 50-60 40-50 30-40 20-30 10-20 0-10 Grado de latitud sur 03° 30 - 04° 00 03°30' - 04° 04° 00 - 04° 30 04° 30 - 05° 00 04° - 05° 05° 00 - 05° 30 05° 30 - 06° 00 05° - 06° 06° 00 - 06° 30 06° 30 - 07° 00 06° - 07° 200 006 175 786 16 087 375 792 78 835 31 692 62.94 110 527 12 669 27 18.51 12 697 2.13 142 416 41 504 07° 00 - 07° 30 19 538 2 548 13 789 65 853 70 222 3 835 19 538 2 548 29 876 208 268 111 726 3 835 4 433 8 671 32 505 33 226 535 1 126 20 879 9 151 535 5 559 29 550 41 656 162 1 345 942 162 1 345 969 07° 30 - 08° 00 07° - 08° 08° 00 - 08° 30 08° 30 - 09° 00 33 226 08° - 09° 10 221 27 09° 00 - 09° 30 09° 30 - 10° 00 10 221 09° - 10° 10° 00 - 10° 30 10° 30 - 11° 00 10° - 11° 11° 00 - 11° 30 15 924 15 924 56 197 2.67 56 197 22 157 9.41 22 157 3 806 3.71 3 806 0.64 11 867 4 058 11 867 4 058 11° 30 - 12° 00 11° - 12° 3 252 20 944 32 001 12° 00 - 12° 30 3 252 20 944 32 001 12° 30 - 13° 00 6 771 15 386 6 771 15 386 12° - 13° 13° 00 - 13° 30 13° 30 - 14° 00 13° -14° 3 806 14° 00 - 14° 30 14° 30 - 15° 00 TOTALES Porcentaje 100.00 3 806 20 073 8 270 67 542 268 562 178 732 53 921 3.36 1.38 11.31 44.98 29.93 9.03 94 14° -15° 597 099 toneladas Evaluación hidroacústica de anchoveta y otros pelágicos en invierno 2003 Castillo, Peraltilla y Segura Con respecto a otros recursos, la disminución de la pota y la vinciguerria que como es habitual se encuentran en áreas alejadas de la costa, preferencialmente en las aguas de mezcla de ASS+ACF y ACF+ATS y en áreas oceánicas. Los recursos jurel y caballa, después del evento El Niño 1997-98 han disminuido considerablemente, comparados la década de los años 80. CONCLUSIONES • La distribución de la anchoveta se localizó en las ACF y aguas de mezcla de las ACF + ASS, con mayor abundancia entre Pimentel y Salaverry desde 15 a 75 mn de la costa. • La biomasa de la anchoveta evaluada por el método de evaluación hidroacústica fue 8.949.966 t, con alta abundancia en los 7°S (3.272.291 t), 8°S (2.629.510) y 9°S (1.015.693 t). • La samasa (biomasa total de 565.614 t) se localizó costera principalmente desde Talara a Chicama, con mayor concentración entre Chérrepe a Chicama. • El bagre (biomasa total de 480.322 t) se encontró principalmente entre Pimentel y Chimbote en diversas áreas costeras. • La pota (357.243 t) y la vinciguerria (1.392.846 t) se localizaron principalmente en áreas alejadas de la costa, en las aguas de mezcla entre ACF-AES por el norte, y ACF-ASS hacia el sur de Pimentel, se detectaron hasta 380 m de profundidad. • La múnida (597.099 t) se encontró en aguas costeras frías y su presencia en esta zona de evaluación ha disminuido en relación a los años anteriores. REFERENCIAS Castillo R, Segura M, Gutierrez M, Ganoza F, Peraltilla S. 2002. Distribución y biomasa de los recursos pelágicos en el mar peruano entre setiembre y noviembre del 2002. Crucero BIC Olaya y BIC SNP-2, 0209-11. Tacna a Piura. (Informe interno, datos no publicados). Castillo R, Segura M, Gutiérrez M, Ganoza F, Peraltilla S. 2003 Localización y abundancia de los recursos pesqueros evaluados en el mar peruano durante el verano del 2003. BIC José Olaya Balandra y BIC SNP-2. Tacna a Tumbes. (Informe interno, datos no publicados). 95 Castillo R, Peraltilla S, Cornejo J. 2003. Distribución y biomasa de recursos costeros determinados por el método de evaluación hidroacústica entre mayo y junio del 2003. BIC SNP-2. Bahía Independencia a Tumbes. Inf. Inst. Mar Perú. XXX: xx-yy. (Informe interno, datos no publicados). Foote KG. 1987. Fish target strengths for use in echo integrator surveys. J. Acoustic Soc. A.m., 82, 981-7. IMARPE. 2003. Informes de Campo del Crucero 0308-09 de Evaluación de Biomasa Desovante de Anchoveta por el Método de Producción de Huevos, a bordo de los BICs Olaya y SNP-2. Informes Internos. IMARPE. 2003. Informe Ejecutivo del Crucero 0308-09 de Evaluación de Biomasa Desovante de Anchoveta por el Método de Producción de Huevos, a bordo de los BICs Olaya y SNP-2; caleta La Cruz a Punta Carretas. Informe Interno. MacLennan D, Simmonds J. 1992. Fishereis Acoustics. Fish and Fisheries Series 5. First edition. Chapman and Hall editors. London, UK. Mori J, PeÑa C, Ventosilla A. 2009. Características biológicas de la anchoveta peruana y otros recursos pelágicos durante agosto-setiembre 2003. Inf. Inst. Mar Perú 35(2): 97-101. Características biológicas de anchoveta y otros pelágicos en invierno 2003 Mori, Peña y Ventosilla CARACTERÍSTICAS BIOLÓGICAS DE LA ANCHOVETA PERUANA Y OTROS RECURSOS PELÁGICOS DURANTE AGOSTO - SETIEMBRE 2003 BIOLOGICAL CHARACTERISTICS OF PERUVIAN ANCHOVETA AND OTHER PELAGIC RESOURCES DURING AUGUST - SEPTEMBER 2003. Julio Mori1 Cecilia Peña2 Arturo Ventosilla1 RESUMEN Mori J, PeÑa C, Ventosilla A. 2008. Características biológicas de la anchoveta peruana y otros recursos pelágicos durante agosto - setiembre 2003. Inf Inst Mar Perú 35(2):97-101.- El crucero BIC Olaya y SNP2 0308-09, se realizó del 13 agosto al 17 setiembre del 2003, entre caleta La Cruz y punta Infiernillos. La captura total fue de 8.026,9 kg. La anchoveta Engraulis ringens constituyó el 71,78%; con LT 5,5 – 17,5 cm; modas 9,0 y 10,0 cm, en la fracción juvenil y 14,0 cm en adultos; el porcentaje de juveniles fue de 74,71% en número y 50,42% en peso. La amplia distribución cercana a la costa, entre Mórrope y Casma, se asoció al ingreso de aguas cálidas de la zona ecuatorial y ASS entre Huarmey y Tambo de Mora. Otras especies importantes fueron: samasa Anchoa nasus 8,95%; pota Dosidicus gigas 7,63%; múnida Pleuroncodes monodon 4,71% y vinciguerria Vinciguerria lucetia 1,20%. Palabras clave: anchoveta, invierno 2003, recursos pelágicos, mar peruano. ABSTRACT Mori J, PeÑa C, Ventosilla A. 2008. Biological characteristics of Peruvian anchoveta and other pelagic resources during August - September 2003. Inf Inst Mar Perú 35(2):97-101.- The RV Olaya and SNP2 030809 cruise was carried on from caleta La Cruz to punta Infiernillos, since August 13th to September 16th. The total catch was 8026.9 kg. The Peruvian anchoveta Engraulis ringens, represented 71.78% of the total, with a wide distribution between Mórrope and Casma; near to the coast because of equatorial waters and the presence of SSW between Huarmey and Tambo de Mora. The TL varied 5,5 to 17,5 cm, with modes in 9,0 and 10,0 cm in young individuals, and adult length mode in 14,0 cm. The juvenile percentage in TL was 74.71% and the weight, 50,42%. Other important species were the long nose anchovy, Anchoa nasus 8,95%; flying squid, Dosidicus gigas 7,63%; carrot lobster, Pleuroncodes monodon 4,71%; and light fish Vinciguerria lucetia 1,20%. Keywords: Peruvian anchoveta, winter 2003, pelagic resources, Peruvian sea. INTRODUCCIÓN MATERIAL Y MÉTODOS El método de la producción de huevos (MPH) es uno de los métodos directos para determinar la biomasa de anchoveta, y está basado en la estimación de la biomasa desovante. La evaluación se realiza cuando el recurso se encuentra cercano a su máxima actividad reproductiva, recolectando información de los valores del índice gonadosomático (IGS) y de la fracción de hembras desovantes. El crucero BIC Olaya y SNP2 0308-09 se llevó a cabo entre caleta La Cruz y punta Infiernillos, del 13 agosto al 17 setiembre 2003. Se colectaron muestras representativas de 103 lances de comprobación, para realizar muestreos biométricos y biológicos con el fin de determinar la estructura por tallas de las cinco especies en evaluación, proporción sexual, madurez gonadal, relación longitud-peso; la colección de otolitos permitió estudios de edad y crecimiento; y los estómagos sirvieron para evaluar la alimentación. 1 2 Para la catalogación macroscópica de madurez gonadal de anchoveta, se examinaron ejemplares sexualmente maduros; se empleó la escala de seis estadios de Einersson et al (1966). Para la determinación del índice gonadosomático se consideraron hembras a partir de 12,0 cm de LT. [email protected], cpeñ[email protected] 97 Para el muestreo biométrico, la selección de los individuos fue aleatoria, pero para el muestreo biológico se emplearon 10 individuos por tallas y por cada grado de latitud.. RESULTADOS Aspectos biolÓgico - pesqueros Composición por especies.- En los 103 lances de comprobación realizados se capturó 8.026,9 kg: anchoveta, Engraulis ringens, 71,8 %; samasa, Anchoa nasus, 8,95%; pota, Dosidicus gigas, 7,63%; múnida, Pleuroncodes monodon, 4,71%; y vinciguerria, Vinciguerria lucetia, 1,20%. Otras especies fueron mictófidos, esperlán plateado, Leuroglossus urotranus; bagre Galeichthys peruvianus; pampanito, Stromateus Informe IMARPE Vol. 35(2): Abril-Junio 2008 Tabla 1.- Captura de anchoveta (kg) según grado latitudinal y distancia a la costa. stellatus; calamar, principalmente Abraliopsis sp.; medusas, salpas. Millas náuticas de la costa En 74 lances positivos, la distribución de anchoveta fue costera entre 5 y 6°S; fue amplia y homogénea entre 7 - 11°S (Pimentel y Huarmey), con capturas hasta las 112 millas frente a punta Chao (9°S). Al sur del Callao, se presentó dentro de las 20 mn de la costa. La mayor captura (815 kg) se registró frente a Asia, entre 12 y 10 mn de la costa, y otras capturas obtenidas entre las 40 y 70 mn en 7°S, con promedio de 438 kg (Tabla 1). Latitud (°S) 10 05°00" 5 05°30" 62 06°00" 37 06°30" 106 07°00" 16 07°30" 388 08°00" 110 08°30" 412 09°00" 09°30" 10°00" 258 10°30" 350 11°00" 65 11°30" 40 12°00" 5 12°30" 815 13°00" 13°30" Total 2 669 Porcentaje (%) 43.0 20 112 59 30 7 23 64 48 7 3 0 1 0 40 50 60 70 450 3 300 400 55 1 600 8 50 5 10 85 50 0 660 31 80 90 100 110 120 Total 5 62 37 113 1 302 1 111 375 484 40 207 105 308 478 725 72 5 815 0 0 40 6 205 0,6 100,0 100 59 1 5 44 20 105 0 128 0 175 2,8 150 1 293 2,4 20,8 350 5,6 643 10,3 673 10,8 15 7 0,1 205 3,3 0 0,0 0 0,0 15 Juveniles 74.71% 10 Juveniles 50.42% Porcentaje (%) Porcentaje (%) % 0,1 1,0 0,6 1,8 21,0 17,9 6,1 7,8 3,3 1,7 5,0 7,7 11,7 1,2 0,1 13,0 0,0 0,0 100,0 5 0 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 Longitud total (cm) 10 Los adultos se hallaron principalmente entre 5-6°S y 10-13°S, y los ejemplares <12 cm se registraron entre 7-8°S (86,2%) (Figura 2). 5 0 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 Longitud total (cm) Figura 1.- Estructura de tallas de anchoveta en número (izquierda) y peso (derecha), ponderados a la biomasa acústica. Crucero MPH 0308-09. Latitud Sur (°S) Juveniles (%) Destacó 80% de incidencia de juveniles entre las 20 y 60 mn de la costa; los adultos se hallaron principalmente en dos zonas, una costera, dentro de las 10 mn (63%) y por fuera de las 60 mn (88%) (Figura Dist. Costa (mn) Juveniles (%) 1.0 5° 6° 0.8 7° 90.3 8° 9° 6 7 8 9 10 11 12 13 14 15 Longitud total (cm) 16 17 18 100 0.0 90 0.0 70 16.4 60 63.2 50 86.9 40 80.9 30 95.9 20 77.5 10 36.8 0.0 13° 0.0 34.0 10.1 12° 1.4 110 80 33.9 11° 120 79.3 31.6 10° 5 Estructura por tamaños de la anchoveta.- La LT tuvo rango 5,5 17,5 cm, estructura polimodal, moda principal 9,0 cm y las secundarias en 10,0 y 14,0 cm. Los juveniles alcanzaron 74,71%, que en peso representó 50,42% de la biomasa acústica total, importante grupo de juveniles, de 7 a 8 meses, provenientes principalmente del desove de verano del 2003 (Figura 1). 19 5 20 6 18.7 7 8 9 10 11 12 13 14 15 16 17 18 19 20 Longitud total (cm) Figura 2.- Estructura de tallas de anchoveta ponderada a la biomasa acústica, según grados de latitud. Crucero MPH 0308-09. 98 Figura 3.- Estructura de tallas de anchoveta ponderada a la biomasa acústica, según distancia a la costa (mn). Crucero MPH 0308 -09. Características biológicas de anchoveta y otros pelágicos en invierno 2003 Puerto Latitu Latitud d (°S) (°S) Distancia a costa (mn) 10 Paita 5 5 Pimentel 6 7 8 9 10 11 12 13 14 15 16 17 18 40 50 60 70 80 90 6 7 8 9 12 13 14 15 16 17 18 19 20 5 75% 07° 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 5 6 7 8 9 10 11 12 75% 08° 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 6 7 8 9 87% 13 14 15 16 17 18 6 7 8 9 12 13 14 15 16 120 17 18 19 20 99% 19 20 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 97% 5 10 11 81% 5 6 7 8 9 10 11 12 13 99% 10 11 12 13 14 15 16 17 18 19 20 5 6 7 8 95% 9 10 11 12 13 14 15 16 17 18 19 20 14 15 16 17 18 19 99% 205 6 7 8 9 10 11 12 96% 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 60% 13 14 15 16 17 18 19 54% 2% 20 5 6 7 8 9 10 11 12 13 95% 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 14 15 16 17 18 19 20 5 6 7 8 83% 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 9 10 11 12 13 14 15 16 17 18 39% 0% 5 6 7 8 9 62% 10 11 12 13 14 15 16 17 18 19 20 5 6 7 8 9 48% 1% 10 11 12 13 14 15 16 17 18 19 20 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 75% 0% 0% 1% 09° Huacho Huarmey 5 6 5 Callao 7 8 9 8 9 10 11 12 13 14 15 16 17 18 19 20 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 7% 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 5 7% 10 11 12 13 14 15 16 17 18 19 20 42% 12° 5 82% 5 6 7 8 9 6 7 8 9 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 0% 10 11 12 13 14 15 16 17 18 19 20 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 3% 10 11 12 13 14 15 16 17 18 19 20 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 25% 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 0% 13° Pisco 7 81% 10° 11° 6 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 14° Figura 4.- Crucero 0308-09. Estructura por tallas de anchoveta según latitud y distancia a la costa. Los números indican % de juveniles (sombreado rojo, izq. de TMC). Adultos (línea azul, der. TMC). 1 60 60 0 4 º - 14 º S Porcentaje (%) Porce ntaje (%) 70 50 40 30 20 1 0 50 04º - 10º S 40 30 20 10 0 0 V I V I VI II III IV V VI Estadio Figura 5.- Madurez gonadal de anchoveta Crucero MPH 0308-09 Presentó una amplia distribución, se le registró asociada a la captura de pota. 100 Porce ntaje (%) Samasa.- El rango de LT fue 6,0 y 15,5 cm; moda principal 13,0 cm. Se halló en la zona norte (5-7°S) dentro de las 20 mn de la costa. 110 1% 10 11 Estadio Otros Recursos 100 19 20 5 Análisis macroscópico de gónadas.- Según el análisis macroscópico de gónadas de anchoveta (n = 622) en la región norte-centro (caleta la Cruz - Bahía Independencia), se observó 64,5% en desove (estadio V) (Figura 5), índice gonadosomático (IGS) de 6,73 y un factor de condición (FC) de 0,6480. Por separado, en la región norte (410°S) hubo 29,7% de ejemplares en maduración (estadio III) y 51,85% de desovantes (estadio V); con IGS 5,76 y FC 0,6332%. En la región centro se registró 10,9% de madurantes y 82,9% de desovante, IGS 8,47 y FC 0,6847, indicando que el recurso se encuentra en su periodo de desove invierno-primavera (Figura 6). 30 1% 06° Aspectos reproductivos 20 1% 05° Chimbote 3). La distribución de ejemplares juveniles de anchoveta se observó principalmente dentro de las 60 mn en la zona comprendida entre Pimentel y Chimbote (7° - 9°S), área donde se presentaron las mayores densidades del recurso; además se registró otro pequeño núcleo en la zona costera (dentro de las 20 mn) entre los puertos de Huarmey y Callao (Figura 4). (referencia) Mori, Peña y Ventosilla 80 10º - 14º S 60 40 20 0 I II III IV V VI Estadio Múnida.- El rango de longitud del cefalotórax (LC) fue 7 - 23 mm, con estructura polimodal, modas principales en 8, 14 y 17 mm (Figura 8). Se distribuyó entre las 20 y 70 mn en los grados 7 y 8°S (Chérrepe y Punta Chao), presentándose como fauna acompañante de la anchoveta. Jurel.- Presentó un rango de LT de 23 a 34 cm, con frecuencia polimodal, moda principal 30 cm y las secundarias en 27 y 33 cm (Figura 12); la incidencia de Figura 6.- Madurez gonadal de anchoveta por regiones. Crucero MPH 0308-09 juveniles fue de 92,6%. El recurso se distribuyó a 90 mn frente a Asia. 12 10 Porcentaje (% ) Pota.- El rango de la longitud del manto (LM) fue 120 a 740 mm, moda principal 170 mm (Figura 7). Presentó una amplia distribución, por fuera de las 30 mn de distancia a la costa. 8 6 4 2 0 80 110 140 170 200 230 260 290 320 350 380 410 440 470 500 530 560 590 620 650 680 710 740 Longitud del manto (mm) Vinciguerria.- El rango de LT fue 3,0 a 7,0 cm, moda 5,0 cm (Figura 9). Figura 7.- Estructura de tallas de pota 99 Informe IMARPE Vol. 35(2): Abril-Junio 2008 nica (fuera de las 60 mn), y esta distribución de ejemplares adultos comprendió las áreas donde se registró la presencia de huevos de anchoveta. 35 20 Porcentaje (%) Porcentaje (%) 30 15 10 5 25 20 15 10 5 0 0 5 7 9 11 13 15 17 19 21 23 25 27 2 29 3 4 5 Longitud del cefalotorax (mm) 6 7 8 9 10 Longitud total (cm) Figura 9.- Estructura de tallas de vinciguerria Figura 8.- Estructura de tallas de múnida. 25 Juveniles : 92.6% Porcentaje (%) 20 15 10 5 0 10 12 14 16 18 20 22 24 26 28 30 32 34 36 38 40 Longitud total (cm) Figura 10.- Estructura de tallas de jurel 25 Juv Mo Cr.0208 16.3 % 13.0 Cr.0308-09 74.7 % 9.0, 10.0 y 14.0 Porcentaje (%) 20 15 10 5 0 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 Longitud Total (cm) Figura 11.- Comparación de la estructura de tallas de anchoveta. Cr. MPH 0208 (en rojo; 16,3% juveniles; moda 13 cm) y Cr. 0308-09 (en azul; 74,7% juveniles; modas 9, 10, y 14 cm). DISCUSIÓN Anchoveta.- Las mayores concentraciones de anchoveta se situaron entre 7 y 9°S. Al norte de los 7°S se presentó costera, en relación a la intromisión de Aguas Ecuatoriales Superficiales (AES) y Aguas Tropicales Superficiales (ATS), que alcanzaron anomalías de +5 °C (frente a Talara). Desde el sur de Chimbote hasta Callao, el ingreso de Aguas Subtropicales Superficiales (ASS) de la zona oceánica replegó el recurso dentro de las 20 mn. En la región norte, en comparación con la situación de la anchoveta durante agosto del 2002 (Crucero MPH 0208), cuando las mayores abundancias se presentaron al norte de los 7°S (entre Mórrope En el crucero MPH 0208 se halló 58,7% en estado de desove. En el crucero MPH 0308-09, la fracción de ejemplares desovantes de anchoveta en la región nortecentro (caleta la Cruz -Bahía Independencia), presentó 64,5% en estadio de desove (V). Los valores obtenidos del seguimiento del proceso reproductivo de la pesquería pelágica de la anchoveta para el 7 de setiembre del 2003, presentaron valores de 41,67% de ejemplares en estado de desove (estadio V), IGS de 6,355 y FC 0,6321, lo cual corrobora los resultados obtenidos en el crucero de invierno 2003. CONCLUSIONES • La anchoveta presentó una distribución continua y amplia entre 7 a 11°S, registrándose capturas hasta las 112 mn de distancia a la costa. • La mayor captura de anchoveta (815 kg) se registró frente a Asia, a 10 mn de la costa; también fueron importantes las capturas obtenidas en 7°S, entre 40 y 70 mn, con promedio de 438 kg. y Chérrepe), en el presente Crucero 0308-09 se observó un desplazamiento de las mayores concentraciones de anchoveta al sur de los 7°S (Pimentel y Chimbote), donde se observó una intromisión de ASS que causó la concentración del recurso en esta zona. En la región centro, en ambos años se observó una distribución escasa principalmente entre Cerro Azul y Bahía Independencia. • El rango de LT de anchoveta fue 5,5 a 17,5 cm, moda principal 9,0 cm y las secundarias 10,0 y 14,0 cm. El porcentaje de juveniles fue de 74,71% que, en peso, representó un 50,42%. En agosto del 2002, los adultos de anchoveta, tuvieron moda 13,0 cm; la proporción de juveniles alcanzó 16,3%, lo cual coincidió con una amplia distribución de huevos de anchoveta. En el crucero MPH 0308-09 se registraron principalmente juveniles (74,7%), adultos en la zona norte (5°-6°S) en la zona costera (10 mn) y en la zona oceá- • La distribución de juveniles de anchoveta se observó dentro de las 60 mn entre Pimentel y Chimbote (7° - 9°S), área con altas densidades del recurso. 100 • Los ejemplares juveniles de anchoveta, con 7 a 8 meses aproximadamente, provenían del desove de verano del 2003. • En la región norte se observó una buena fracción de ejemplares en maduración (29,7%) y de Características biológicas de anchoveta y otros pelágicos en invierno 2003 Mori, Peña y Ventosilla desovantes (51,9%). En la zona centro se registró menor porcentaje de madurantes (10,9%) y un aumento de desovantes (82,9%). El índice gonadosomático total (6,73) indicaba que el recurso se encontraba en el periodo de desove invierno –primavera. El análisis por regiones reflejó una mayor actividad del desove en la región centro (IGS: 8,47), y en la región norte fue menor (5,76). REFERENCIAS Chirichigno N, Vélez J. 1998. Clave para identificar los peces en el Perú. (2da edición). Publicación especial Inst. Mar Perú, diciembre 1998; 500 pp. Einarsson H, Flores LA, MiÑano J. 1966.El ciclo de madurez de la anchoveta peruana (Engraulis ringens J.) En: Seminario Latinoamericano sobre el Pacífico oriental, Universidad Nacional Mayor de 101 San Marcos , Lima, Perú: 128-135. Mori J, Goicochea C, PeÑa C, Ventosilla A. 2007. Aspectos biológicos pesqueros de los principales recursos pelágicos. Invierno 2002. Inf Inst Mar Perú 34(3):197-202. Vazzoler AE.1982. Manual y métodos para estudios biologicos de populacoes de Peixes. Reproducción y Crecimiento. CNP/PN2, 108 pp. Fitoplancton en el invierno 2003 Chang y Delgado FITOPLANCTON EN EL INVIERNO 2003 PHYTOPLANKTON IN WINTER 2003 Flor Chang1 Elcira Delgado RESUMEN Chang F, Delgado E. 2008.- Fitoplancton en el invierno 2003. Inf. Inst. Mar. Perú 35(2):103-107.- Se da a conocer las características de la comunidad fitoplanctónica en el periodo agosto-setiembre 2003. Los volúmenes de plancton superficial variaron entre 0,1 y 3,23 mL.m-3, localizados frente a Paita y Pisco, con promedio general de 0,66 mL.m-3. Este tipo de fitoplancton en superficie predominó en el 31% del área evaluada. A 10 m de profundidad la mayor concentración celular (2 966x103cel L-1) se registró en el perfil Callao con el predominio de Skeletonema costatum y Chaetoceros socialis. Ceratium breve indicador de AES y C. praelongum de ASS, denotaron aguas de mezcla desde Puerto Pizarro hasta Punta Falsa dentro de las 30 mn; el indicador de ACF Protoperidinium obtusum presentó una distribución normal al sur de Mórrope. Palabras clave: fitoplancton, invierno 2003, mar peruano, crucero de biomasa desovante. ABSTRACT Chang F, Delgado E. 2008.- Phytoplankton in winter 2003. Inf Inst Mar Peru 35 (2): 103-107.- The characteristics of the phytoplankton community in the period August-September 2003 are annotated. Surface plankton volumes ranged between 0.1 and 3.23 mL.m-3, off Paita and Pisco, with 0.66 mL.m-3. general average. This type of phytoplankton in surface predominated in 31% of the area evaluated. At 10 m depth, the highest cell concentration (2966x103cel.L-1) was recorded at Callao profile with a predominance of Skeletonema costatum and Chaetoceros socialis. Ceratium breve, indicative of equatorial surface waters, and C. praelongum of subtropical superficial waters denoted mixing waters from Puerto Pizarro to Punta Falsa within 30 mn, while Protoperidinium obtusum, cold coastal waters indicator showed a normal distribution beyond to south Mórrope. Keywords: phytoplankton, winter 2003, Peruvian sea, spawning biomass cruise. INTRODUCCIÓN El Instituto del Mar del Perú, debido a la importancia del recurso anchoveta, ejecuta en el invierno (agosto-setiembre) de cada año el crucero de evaluación de la masa desovante por el método de producción de huevos (MPH), incluyendo el estudio de las condiciones ambientales. Por estas razones es indispensable el estudio del plancton, pues el éxito de la sobrevivencia de los estadios larvales de peces depende de la calidad y cantidad de alimento. En este crucero BIC Olaya y SNP2 0308-09, se estudió la comunidad planctónica, volúmenes de plancton, con énfasis en la distribución y composición de especies del fitoplancton, así como la concentración celular (cel L-1) a 10 m de profundidad y la distribución de los organismos indicadores de masas de agua. La carta de posiciones y el número de lances se describen en el primer trabajo de este número (AyÓn y BuitrÓn 2009) MATERIAL Y MÉTODOS Se colectó un total de 107 muestras para los análisis cualitativos y cuantitativos de fitoplancton. La metodología usada corresponde a la establecida por el IMARPE, y mencionada en trabajos anteriores, anotados en las Referencias. RESULTADOS Distribución de los volúmenes de plancton y composición de la comunidad fitoplanctónica en superficie.- Para agosto-setiembre del 2003 los volúmenes de plancton fluctuaron entre 0,11 mL m-3 (Est. 82) y 3,23 mL m-3 (Est. 705), donde el 81% de los volúmenes fueron menores a 1,0 mL/m3. El volumen promedio general fue de 0,66 mL m-3. Los rangos de temperatura superficial del mar (TSM) fluctuaron entre 14,7 y 24,1 ° C. La predominancia del fitoplancton fue en 31% de las estaciones, en dos áreas principales: la primera desde el sur de Punta Falsa hasta el norte de Punta Chao, dentro de 60 a 90 mn; y la segunda, entre Huarmey y Callao con máxima distribución de 70 mn frente a Huarmey. La predominancia del zooplancton fue 59% y la codominancia de fito y zoo fue 10% (Figura 1). Diatomeas.-Frente al área entre Punta Falsa y norte de Punta Chao hubo abundancia de diatomeas neríticas (Coscinodiscus perforatus, Skeletonema costatum, Actinocyclus Unidad de Producción Primaria. DIO. IMARPE. [email protected], [email protected] 103 Informe IMARPE Vol. 35(2): Abril-Junio 2008 84°W 82°W 80°W 78°W 76°W 74°W Pto. Pizarro 4°S 1 4°S Cabo Blanco 0.5 Talara Paita 6°S 6°S Pta. Falsa 1 Mórrope Pimentel 1.5 0.5 Pacasmayo Chicama 8°S 8°S Salaverry 0.5 Punta Chao 0.5 Chimbote 1 0.5 10°S 1 1.5 0. 5 12°S 10°S Huarmey Supe Huacho 0.5 1 Chancay 12°S Callao Pucusana 84°W 82°W 80°W 1 FITOPLANCTON ZOOPLANCTON FITOPLANCTON Y ZOOPLANCTON 14°S .5 Tambo de Mora 2 1 Pisco 14°S Bahía Independencia 78°W 76°W 74°W Figura 1.- Distribución de volúmenes de planecton (mLm-3). Cr. Biomasa Desovante de anchoveta 0308-09. BICs José Olaya B. y SNP-2 2.00 1.60 mL/m 3 1.20 0.80 0.40 14° - 15°S 13°-14° S 12°-13°S 11°-12° S 10°-11° S 09°-10° S 08°-09° S 07°-08°S 06°-07°S 05°-06°S 04° - 05°S 03°-04°S 0.00 Grados latitudinales 0-30 mn 30-60 mn >60 mn Figura 2.- Distribución latitudinal de volúmenes de plancton (mLm-3) Cr. Biomasa Desovante 0308-09. BIC’S Olaya y SNP-2 sp., Thalassiosira anguste-lineata, Thalassiosira sp. y Lithodesmium undulatum. Entre Huarmey y Callao, a las diatomeas anteriores se sumó un número de especies oceánicas: Thalassiosira partheneia, Planktoniella sol, Corethron hystrix, Fragilariopsis doliolus, Rhizosolenia bergoni, Proboscia alata f. indica distribuidas generalmente por fuera de las 55 mn. leonis, P. mendiolae, P. depressum y P. conicum. Los dinoflagelados termófilos ocuparon áreas más restringidos, entre Puerto Pizarro y Punta Falsa hasta muy cerca de la línea costera; y entre Huarmey y sur de Callao, por fuera de las 30 mn (Ornithocercus quadratus, Ceratocorys horrida, Podolampas bipes, Goniodoma polyedricum, Ceratium extensum y Amphisolenia bidentata). Dinoflagelados.- Los dinoflagelados cosmopolitas presentaron una amplia distribución (Ceratium tripos, Ceratium furca, Protoperidinium Dentro de las 60 mn hubo importantes registros. En el sur resaltó un núcleo >3,0 mL m-3 frente a Pisco (Est. 705), y otros núcleos de 104 2,0 mL m-3 entre Punta Falsa y Mórrope (C-6) y de Huarmey a Supe (Est. 352). En estos núcleos predominó el zooplancton (Figura 1). Distribución latitudinal de los volúmenes promedio de plancton.- En el invierno 2003 la distribución latitudinal de los volúmenes promedio de plancton presentó el esquema de la Figura 2. Dentro de las 30 mn, el volumen promedio más alto (1,85 mL/ m3) se presentó entre 13 y 14°S; a otras latitudes los valores fueron <1,0 mL/m3. Entre 30 y 60 mn se presentaron máximos de 1,19 entre 13-14°S; y 1,13 en latitud 4 - 5°S. Por fuera de las 60 mn también se presentaron volúmenes de 1,45 mL/ m3, entre 12-13°S y de 1,36 mL/m3 entre 3-4°S. Distribución de la comunidad fitoplanctónica a 10 m de profundidad (N°celx103l-1).- El análisis cuantitativo identificó un total de 117 especies (67 diatomeas, 31 dinoflagelados, 3 silicoflagelados, 11 cocolitofóridos y 5 fitoflagelados). Las concentraciones del total de fitoplancton fluctuaron entre 27x103 cel.L-1 y 2966x103 cel.L-1 ambas localizadas en el Callao con un promedio de 263lx103L-1 (Figura 3a). La distribución del fitoplancton se caracterizó por presentar áreas con concentraciones >500x103 cel L-1 en zonas costeras; con un núcleo máximo en Callao >1000x103 cel L-1. Las concentraciones <100x103 cel.L-1 se registraron lejos de la costa, excepto en los perfiles de Paita y Punta Falsa que se registraron dentro de la franja costera (Figura 3b). El nanoplancton fue el más representativo en el 81% de las estaciones, excepto en Callao (Est. 407 y 411), así como en Pisco donde destacó el microplancton. La máxima densidad del nanoplancton se alcanzó en Chimbote (393x103 cel L-1), por fuera de las 90 mn. Un grupo de monadas, y fitoflagelados (49x103cel L-1) en Callao (dentro de las 60 mn); los cocolitofóridos con Emiliania huxleyi (46x103cel L-1) en Paita, a 10 mn. Fitoplancton en el invierno 2003 Chang y Delgado 4°S 84°W 74°W 76°W 78°W 80°W 82°W 84°W 82°W 80°W Paita 6°S 10 4°S 10 Pta. Falsa Pta. Falsa 8°S Pacasmayo Chicama Punta Chao Chimbote Punta Chao Chimbote Supe Huacho 20 Chancay 0 500 0 Callao 10 Supe 00 Huacho Chancay 20 00 12°S 100 50 0 (a) (b) Tambo de Mora Pisco 20 82°W 80°W 78°W 84°W 74°W 76°W Pucusana 0 10 Bahía Independencia 84°W 82°W 80°W 12°S Callao 1000Pucusana 14°S 10°S Huarmey Huarmey 10 100 8°S Salaverry Salaverry 20 0 6°S Mórrope Pimentel Mórrope Pimentel Pacasmayo Chicama 10°S 74°W Paita 10 10 0 0 76°W Cabo Blanco Talara Cabo Blanco Talara 100 78°W Pto. Pizarro Pto. Pizarro 78°W Tambo de Mora Pisco Bahía Independencia 76°W 14°S 74°W Figura 3.- Fitoplancton total (a) y total de diatomeas (b) (Nro. cél x 10 /L. Cr. Biomasa Desovante de anchoveta 0308-09. BIC’s José Olaya B. y SNP-2 20 40 Similarity 60 80 100 G po ru 3 o4 up Gr po ru Gr up o2 G ru po 1 5 Grupo I, incluyó estaciones de áreas geográficas distantes observadas en los perfiles de Pisco dentro de las 30 mn (Est. 710); Paita y Callao por fuera de las 90 mn (Est. 595, 601 y 74). Este grupo G Índice de diversidad y análisis comunitario.Los indices de Shannon y Wiener (H’) estuvieron comprendidos entre 0,305 y 1,986 bits/individuos, con la mayor diversidad de especies (41 especies) a 10 mn frente al Callao y la menor frente a Punta Falsa por fuera de las 90 mn (6 especies). El análisis de clasificación numérica en modo Q al 42% de similitud permitió distinguir la presencia de 5 grupos (Figura 4). 407 104 86 114 125 411 51 81 462 B587 119 291 469 100 714 474 699 705 710 74 595 601 El microplancton solo estuvo presente en el 19% de las estaciones. Destacaron las diatomeas dentro de las 30 mn en el perfil Callao (2966x103cel L-1) predominando especies de la primera fase como Skeletonema costatum (2589x103cel L-1) y Ch. socialis (2520x103cel L-1), asociado a Guinardia delicatula (256x103cel L-1) y Dactyliosollen fragilissimus (39x103 cel L-1) especies de fases intermedias, así como en Pisco a 26 mn en donde sobresalió Guinardia flaccida con 87x103 cel L-1. Figura 4.- Dendrograma de Similaridad (10 m). Crucero de Biomasa Desovante. BIC Olaya y SNP2 0308-09. se caracterizó por la presencia de Navicula sp., fitoflagelados y el grupo de las Monadas. Grupo II, comprendió tres áreas geográficas distantes ubicadas en el perfil Chimbote dentro de las 60 mn (Est. 291, 474 y 469) y frente a los perfiles de Paita (Est. 100 y 119) y Pisco (Est. 714, 705 y 699) dentro de la franja costera y por 105 fuera de las 90 mn; caracterizado por la dominancia total del nanoplancton principalmente por los Fitoflagelados y Monadas. Grupo III, reunió las estaciones localizadas entre las 30-60 mn en los perfiles hidrográficos de Callao, Paita y Chimbote, que fue observado por fuera de las 90 mn (Est. 462). Este grupo al igual Informe IMARPE Vol. 35(2): Abril-Junio 2008 84°W 82°W 80°W 78°W 76°W 74°W Pto. Pizarro 4°S 4°S Cabo Blanco Talara Paita 6°S 6°S Pta. Falsa Mórrope Pimentel Pacasmayo Chicama 8°S 8°S Salaverry Punta Chao Chimbote 10°S 10°S Huarmey Supe Huacho Chancay 12°S 12°S Callao Pucusana 14°S Protoperidinium obtusum (ACF) (AES) Ceratium breve Ceratium praelongum, C. incisum (ASS) 84°W 82°W 80°W 78°W Tambo de Mora Pisco Bahía Independencia 76°W 14°S 74°W Figura 5.- Indicadores de masas de agua. Crucero de Biomasa Desovante anchoveta 0308-09 que el segundo se caracterizó también por la dominancia del nanoplancton destacando el grupo de las Monadas, Fitoflagelados y E. huxleyi, junto a especies de afloramiento como Chaetoceros socialis. Grupo IV, pequeño grupo referido sólo por dos estaciones localizadas en el perfil Punta Falsa por fuera de las 90 mn (Est. 125) y Callao dentro de la franja costera (Est. 411). Se careacterizó principalmente por la dominancia del grupo del nanoplancton con las especies mencionadas anteriormente junto a diatomeas de fase intermedias (Guinardia delicatula, Dactyliosolen fragilissimus y Leptocilindrus danicus). Grupo V, captó estaciones de los perfiles de Paita y Punta Falsa ubicadas entre las 30 – 60 mn, incluyendo la estación costera a 22 mn dentro de este último perfil. Este grupo se caracterizó también por la dominancia del grupo del nanoplancton (Monadas, Fitoflagelados y E. huxleyi ). Distribución de especies de fitoplancton indicadoras de masas de agua.- Ceratium breve, indicador de Aguas Ecuatoriales Superficiales (AES) y C. praelongum y C. incisum, indicadores de Aguas Subtropicales Superficiales (ASS) se distribuyeron ampliamente desde Puerto Pizarro hasta Punta Falsa, denotando aguas de mezcla cerca a la línea costera, con 23 °C y salinidad de 34,5 ups. Protoperidinium obtusum, indicador de Aguas Costeras Frías (ACF) estuvo distribuido en forma homogénea cerca de la línea costera entre Mórrope y sur de Pucusana, hasta 48 mn frente a Pimentel, relacionado a temperaturas de 14 a 17 °C y salinidades de 35,0 ups, las cuales son típicas de esta masa de agua. Cabe resaltar una gran riqueza de especies termófilas entre Puerto Pizarro-Punta Falsa y Huarmey-sur de Callao, en esta última por fuera de las 30 mn (Figura 5). DISCUSIÓN La variabilidad ambiental constituye un factor importante en la alimentación de los primeros estadios larvales de los peces planctófagos. En esta evaluación de invierno 2003, el volumen promedio de plancton superficial fue <1,0 mL.m-3, por tanto, las condiciones 106 del medio ambiente fueron adversas. Se registró intromisión de AES y ASS, disminución del fitoplancton, pero predominancia de las diatomeas centrales, que son alimento de óptima calidad; esto difiere con lo encontrado en invierno 2002 (Villanueva y Sánchez 2003), cuando el valor medio fue de 1,0 mL.m-3 y fue mayor la predominancia del fitoplancton. Con relación a las concentraciones de biomasa planctónica, se observaron diferencias entre los años 2002 y 2003. En el invierno 2002 el fitoplancton caracterizó a estos núcleos y en el 2003 fue el zooplancton, cuya predominancia estaría relacionada al fuerte pastoreo que esta comunidad ejerce sobre el fitoplancton. Latitudinalmente, el invierno 2003 tuvo 1,85 mL/m3 como valor medio máximo de 0 a 30 mn de la costa; en el 2002, la media máxima fue de 3,0 mL/m3, entre 30 a 60 mn. Cuantitativamente, a 10 m la mayor densidad (N°celx103 L-1) fue observada frente a Callao dentro de la franja costera coincidiendo con altos valores de clorofila-a (6,7 ug/L) (Flores, com. per.), dado principalmente por diatomeas de la primera fase de la sucesión ecológica. En el invierno 2002 se registraron concentraciones >1000x103 cel L-1, dentro de la franja costera de Paita y Chimbote. Este desplazamiento podría estar relacionado con la presencia de aguas de mezcla (AES y ASS), las cuales son pobres en nutrientes. También se observó cambio notorio en la composición especiológica. En 2003 sobresalió el nanoplancton y en el 2002 destacó el microplancton dentro de la franja costera; este hecho podría estar relacionado con el intenso afloramiento observado en mayo del 2002 (IMARPE 2002b). Existen estudios sobre alimentación de larvas de anchoveta (Muck et al. 1989) que incluyen en su dieta especies del nanoplancton (Monadas y Fitoflagelados). De acuerdo a Margalef (1968), los índices <2,5 bits/individuo corresponden a comunidades inmaduras. Blasco (1921) y Fitoplancton en el invierno 2003 Chang y Delgado Rojas de Mendiola y Estrada (1976), confirmaron que las comunidades del mar peruano se encuentran en esta fase. Los valores encontrados en el presente estudio, indicarían que la comunidad fitoplanctónica se encuentra en etapas iniciales. Con respecto a los indicadores de masas de agua, durante el mismo periodo estacional, en el 2001 (Sánchez et al. 2002) y el 2002 (Villanueva et al. 2003), se apreció una similar tendencia del desplazamiento de AES hacia la costa peruana hasta los 6°S, pero con cierta peculiaridad, pues hubo una fuerte intromisión de AES y ASS denotando aguas de mezcla (6°20’S) donde se determinó a C. breve y C. praelongum hasta cerca de la línea costera. Estas condiciones estuvieron acorde a los parámetros físicos (IMARPE 2003). En relación a las ACF, en el 2001 (Sánchez et al. 2002) el afloramiento fue casi cercano a lo normal, pues P. obtusum presentó una amplia distribución. Difiere del 2002 (Villanueva et al. 2003) cuando fue observado en zonas costeras, concentrado en áreas localizadas y presentándose una amplia distribución en el 2003. Para este período los organismos termófilos estuvieron mejor representados al norte de los 6°S hasta muy cerca de la costa, debido las masas de agua cálidas (ASS y AES). Durante el invierno del 2002 estuvieron dentro de las 30 mn a lo largo de la línea costera, zona que se vio influenciada por la intromisión de ASS. CONCLUSIONES • El promedio general del volumen de plancton y la predominancia porcentual del fitoplancton estuvieron por debajo de lo normal. El grupo de las diatomeas centrales registró abundancia, lo que estaría reflejando una buena disponibilidad de alimento óptimo para las especies consumidores de plancton. • El nanoplancton (Fitoflagelados y Monadas) destacó en el 81% de las estaciones, lo que indicaría disponibilidad de alimento para los primeros estadios larvales de anchoveta. • Como respuesta a las condiciones ambientales inusuales, principalmente en la región norte, para este período en estudio, los indicadores biológicos de masas de agua se distribuyeron hasta aproximadamente los 6,40°S, pues la presencia de Ceratium breve (AES) y C. praelongum (ASS) indicaría aguas de mezcla. Mientras que P. obtusum (ACF) presentó una distribución normal al sur de Mórrope asociados a temperaturas y salinidades propias de esta masa de agua. Agradecimiento.- Al personal del Área de Fitoplancton y Producción Primaria, por el apoyo brindado en el análisis de las muestras y elaboración de gráficos. REFERENCIAS Blasco D. 1971. Composición y distribución del fitoplancton en la región del afloramiento de las costas peruana. Inv. Pesq. 35(1):61-112. Cupp E. 1943. Marine plankton diatoms of the west coast of North America. Bull. Scripps Inst. Oceangr. 5: 1-237. Balech E. 1988. Los dinoflagelados del Atlántico Sudoccidental. Public. Espec. Inst. Español de Oceanog. España. 310 pp. Hasle GR Syvertsen EE.1996. Identifying Marine Diatoms and Dinoflagellates. Academic Press, Inc. 585 pp. Hasle Gr Syvertsen EE.1996. Identifying Marine Diatoms and Dinoflagellates. Academic Press, Inc. 585 pp. Hendey I. 1964. An introductory account of the smaller algae of British Coastal waters. Part. V. Bacillariophyceae (Diatoms). Her Majesy’s Stationery Office, London: 317 pp. Hustedt F. 1930. Die Kieselalgen Deutschlands. Osterreichs und der Schweiz mit Berücksichtigung der übrigen Länder Europas sowie der angerzenden Meeresgebiete. En: L. Rabenhorst’s 107 (ed.). Kryptogamen-Flora von Deutschland, Osterreich und der Schweiz. 1 Teil. Akad. Verlag. Leipzig, Reprint Johnson Rep. Goop, New York 1971: 920 pp. Licea S, Moreno L, Santoyo H. 1996a. Diatomeas del golfo de California Inst. de Ciencias del Mar y Limnología, México. 1-258 pp. Licea S, Moreno L, Santoyo H. 1996 b. Dinoflagelados del golfo de California Inst. de Ciencias del Mar y Limnología, México, 1- 258 pp. Margalef R., 1968. Perspectives in ecological theory . The University of Chicago Press. Chicago 111 pp. Muck P, Rojas de Mendiola B, Antonietti E. 1989. Comparative studies on feeeding in larval anchoveta (Engraulins ringens) and sardine (Sardinox sagax). The Peruvian upwelling ecosystem: dinamics and interactions. Eds. Pauly, Muck, Mendo, Tsukayama. IMARPE, ICLARM, GTZ: 86-96. Rojas de Mendiola B, Estrada M. 1976. El fitoplancton en el área de Pimentel. Verano de 1972. Inv. Pesq 40(2):463-490. Steidinger K, Tangen K. 1996. Dinoflagellates. In : Tomas C. (ed) Identifying Marine Diatoms and Dinoflagellate. Academic Press, Inc. San Diego. 387- 570 pp. SundstrÖm B. 1986. The Marine diatom genus Rhizosolenia. A new approach to the taxonomy. Lund, Sweden: 196 pp. Schiller J. 1971a. Dinoflagellate (Peridinae) in monographischer Behandlung. 2 Tell. En: L. Rabenhortst (ed). Kriptogamen-Flora von Deutschland, Osterreich und der Schweiz. Reprint by Johnson Repr. Corp. New York, 1971, Vol. 10 Section 3, Part 1: 617 pp. Schiller J. 1971b. Dinoflagellate (Peridinae) in monographischer Behandlung. 2 Tell. En: L. Rabenhortst (ed). Kriptogamen-Flora von Deutschland, Osterreich und der Schweiz. Reprint by Johnson Repr. Corp. New York, 1971, Vol 10 Section 3, Parte 2: 589 pp. Sournia A. 1967. Le genere Ceratium (Peridinien Planctonique) dans le Canal de Mozambique. Contribution a une revision mondiale. Vie et Milieu. 18 (2A-A): pp. 375-580. Villanueva P, Sánchez. 2003 Características de la comunidad Fitoplanctónica de Invierno en el Crucero de Estimación de Biomasa de la anchoveta 0208. Condiciones oceanográficas físicas en invierno 2003 Vásquez CONDICIONES OCEANOGRÁFICAS FÍSICAS EN INVIERNO 2003 PHYSICAL OCEANOGRAPHIC CONDITIONS IN WINTER 2003 Luis Vásquez Espinoza1 RESUMEN Vásquez L. 2008. Condiciones oceanográficas físicas en invierno 2003. Inf Inst Mar Perú 35(2):109-114.El crucero BIC Olaya y SNP2 0308-09 abarcó de Puerto Pizarro (3°30´S) a Punta Infiernillos (14°40’S), del 14 agosto al 16 setiembre 2003. Las condiciones fueron cálidas entre Puerto Pizarro y Pimentel (7°S); fueron normales de Pimentel a Infiernillos, pero se registraron frías en la zona oceánica al norte de Chimbote. La inusual proyección hacia el sur de aguas tropicales y ecuatoriales, ocasionó anomalías positivas de hasta +5°C frente a Talara-Paita. Al sur de Pimentel las aguas costeras frías tuvieron amplia distribución, y los vientos de intensidad moderada a fuerte contribuyeron a mantener activos los procesos de afloramiento, con excepción de la zona frente a Supe, donde se presentó una trasgresión de las aguas superficiales subtropicales (ASS) hacia la costa motivando que los procesos de afloramiento estuvieran restringidos a zonas muy costeras. Palabras clave: condiciones oceanográficas físicas, biomasa desovante, invierno 2003, mar peruano ABSTRACT Vasquez L. 2008. Physical oceanographic conditions in winter 2003. Inf Inst Mar Peru 35 (2): 109-114.The cruise BIC Olaya and SNP2 0308-09 covered Puerto Pizarro (3°30’S) to Punta Infiernillos (14°40’S) offshore, from August 14th to September 16th 2003. The conditions were warm between Puerto Pizarro and Pimentel (7°S), were normal south to Infiernillos, but were cold in the oceanic area north of Chimbote. The unusual southward projection of tropical and equatorial waters, resulting in positive anomalies of up to +5 °C off Talara to Paita. South to Pimentel the cold coastal waters were widespread, and the winds, moderate to strong, contributed to upwelling processes, except the area off Supe, where the subtropical surface water (SSW) toward the coast, motivated the coast upwelling processes were restricted to areas near the coast. Keywords: physical oceanographic conditions, spawning biomass, winter 2003, Peruvian sea. INTRODUCCIÓN Las condiciones del ambiente marino en julio 2003 se caracterizaron por las condiciones neutrales de El Niño Oscilación del Sur (ENOS) en el Pacífico tropical. Las anomalías negativas de TSM en el Pacífico tropical oriental continuaron debilitándose, pero una tendencia a enfriamiento ligero se observó en el Pacífico central en la última quincena. La capa superficial mostró tendencia a un calentamiento en el Pacífico tropical central y oriental, con signos evidentes de declinación en la primera semana de agosto. Los índices oceánicos y atmosféricos frente a la costa sudamericana para el invierno, indicaron características climáticas neutras, pero que se alteraron en agosto por el arribo de una onda Kelvin que, aunada al relajamiento de los vientos alisios, causaron un ambiente cálido al norte de Pimentel. Al sur se mantuvieron los afloramientos costeros favoreciendo y ampliando la distribución de ACF que sobrepasaron las 90 mn. Bajo este panorama se desarrolló el Crucero 0308-09, encontrando un ambiente cálido en la zona norte, con un fuerte desplazamiento de aguas ecuatoriales y condiciones casi normales al sur de los 8°S. En este trabajo se da a conocer las condiciones físicas registradas. MATERIAL Y MÉTODOS El crucero de evaluación de la biomasa desovante de anchoveta, BIC Olaya y SNP2 0308-09, por el método de la producción de Dirección de Investigaciones Oceanográficas. IMARPE. [email protected] 109 huevos (MPH), se realizó del 14 agosto al 18 setiembre del 2003, desde Puerto Pizarro (3°30’S) a Punta Infiernillos (14°40’S), cubriendo un área entre la costa y 120 mn de dominio marítimo. El estudio de la estructura térmica y halina en las secciones Paita, Punta Falsa, Chicama, Chimbote, Callao y Pisco se realizó sobre la base de lanzamientos de CTD, hasta 500 m de profundidad y de botellas Niskin a profundidades estándar, hasta 300 m, en el BIC Olaya y hasta 150 m en el BIC SNP2). Adicionalmente, los lanzamientos de CTD frente a caleta La Cruz y Supe llegaron hasta 300 m de profundidad. En la capa superficial del mar se registró temperatura, salinidad y viento (dirección y velocidad) cada 4-5 mn (durante las estaciones de plancton) (Figura 1). Informe IMARPE Vol. 35(2): Abril-Junio 2008 Zorritos 4 Zorritos 4° Pta. Sal 5 Cabo Blanco Talara Talara Paita Pta. Gobernador CRUCERO BIOMASA DESOVANTA .2003/08-09 5° CARTA DE POSICIONES 6° Parachique Pta. Falsa 6 Pta. Sal Cabo Blanco Crucero 2003/08-09 Paita Pta. Gobernador Parachique Pta. Falsa Mórrope Mórrope Pimentel 7 TEMPERATURA (°C) Pimentel 7° Chérrepe Pacasmayo Chérrepe Pacasmayo Chicama 8 Chicama 8° Trujillo Salaverry Guañape 25 9 Punta Lobos Supe 11 Bujama Cerro Azul Punta Cóndor Tambo de Mora Pisco Peninsula Paracas 13 BIC SNP 2 BIC OLAYA 78 77 76 Huacho Punta Salinas Chancay Ancón Callao Punta Hermosa v 15 Bujama Cerro Azul 14 Punta Cóndor Tambo de Mora 13 14° 75 Pisco Peninsula Paracas 12 Bahía Independencia 11 Punta Infiernillos 79 Supe 16 13° Bahía Independencia 80 Punta Bermejo 17 12° Callao Punta Hermosa 81 Huarmey 18 Chancay Ancón 82 Punta Lobos 19 11° Huacho Punta Salinas 12 Casma 20 Punta Bermejo 83 Chimbote 21 10° Huarmey 15 Punta Chao 22 Casma 14 Salaverry Guañape 23 9° Chimbote 10 Trujillo 24 Punta Chao 15° 74 Punta Infiernillos 10 Punta Caballas 83° Figura 1.- Carta de Posiciones. Crucero Biomasa Desovante 0308-09 82° 81° 80° 5° ANOMALIAS TERMICAS(°C) Paita Crucero 2003/08-09 SALINIDAD (ups) Parachique 6° Pta. Falsa Mórrope Mórrope Pimentel Pimentel 7° Chérrepe Chérrepe Pacasmayo Pacasmayo Chicama 5 Chicama 8° Trujillo Salaverry Guañape 4 35.2 35.1 35 34.9 34.8 34.7 34.6 34.5 34.4 34.3 34.2 34.1 34 33.9 33.8 33.7 33.6 33.5 83° 82° Punta Chao 9° 9° Chimbote 3 Casma Punta Lobos 2 10° Huarmey Punta Bermejo 11° 1 Supe 11° Huacho Punta Salinas 0 12° Chancay Ancón 12° Callao -1 Punta Hermosa v 13° Bujama -2 13° Cerro Azul Punta Cóndor Tambo de Mora -3 14° Pisco Peninsula Paracas -4 14° Bahía Independencia Punta Infiernillos 15° -5 83° 15° Punta Caballas 82° 81° 80° 79° 78° 77° 76° 75° 74° Figura 3.- Anomalías Térmicas (°C). Crucero Biomasa Desovante 0308-09 Trujillo Salaverry Guañape Punta Chao Chimbote Casma Punta Lobos Huarmey Punta Bermejo Supe Huacho Punta Salinas Chancay Ancón Callao Punta Hermosa v Bujama Cerro Azul Punta Cóndor Tambo de Mora Pisco Peninsula Paracas Bahía Independencia Punta Infiernillos Punta Caballas 81° 80° Paita Parachique 6° Pta. Falsa Pta. Falsa Mórrope Pimentel Pimentel 7° Chérrepe Chérrepe Pacasmayo Pacasmayo Chicama 15 Chicama 8° Trujillo Salaverry Guañape 14 Punta Chao 10° 13 Chimbote 12 Casma 12° 9° Punta Lobos 11 10° Huarmey 10 11° Punta Bermejo 9 Supe 8 Chancay Ancón 7 12° Callao 6 Punta Hermosa v Bujama 13° Cerro Azul 4 Punta Cóndor Tambo de Mora 3 14° 11° Huacho Punta Salinas 5 13° Pisco Peninsula Paracas 2 14° Bahía Independencia 1 15° 0 83° Punta Infiernillos Punta Caballas 82° 81° 74° TURBULENCIA (m3) Pta. Gobernador Mórrope 9° 75° Crucero 2003/08-09 Talara Parachique 8° 76° Pta. Sal 5° VIENTO (m/s) Paita Pta. Gobernador 7° 77° Cabo Blanco Crucero 2003/08-09 Talara 6° 78° Zorritos 4° Pta. Sal Cabo Blanco 5° 79° Figura 4.- Distribución de Salinidad (ups). Crucero Biomasa Desovante 0308-09 Zorritos 4° 74° Paita Pta. Gobernador Pta. Falsa 7° 10° 75° Pta. Sal Talara Parachique 8° 76° Cabo Blanco Crucero 2003/08-09 Pta. Gobernador 6° 77° Zorritos 4° Pta. Sal Cabo Blanco Talara 5° 78° Figura 2.- Distribución de Temperatura (°C). Crucero Biomasa Desovante 0308-09 Zorritos 4° 79° 80° 79° 78° 77° 76° 75° 15° 74° Figura 5.- Distribución del Viento (m/s). Crucero Biomasa Desovante 0308-09 Trujillo 4000 3800 3600 3400 3200 3000 2800 2600 2400 2200 2000 1800 1600 1400 1200 1000 800 600 400 200 0 83° Salaverry Guañape Punta Chao Chimbote Casma Punta Lobos Huarmey Punta Bermejo Supe Huacho Punta Salinas Chancay Ancón Callao Punta Hermosa v Bujama Cerro Azul Punta Cóndor Tambo de Mora Pisco Peninsula Paracas Bahía Independencia Punta Infiernillos Punta Caballas 82° 81° 80° 79° 78° 77° 76° 75° 74° Figura 6.- Distribución de la Turbulencia (m3/s3). Crucero Biomasa Desovante 0308-09 110 Condiciones oceanográficas físicas en invierno 2003 Vásquez Las muestras de salinidad se analizaron por el método de inducción con el Portasal 8410A Guildline (BIC Olaya) y el salinometro Kahlsico RS 10 (BIC SNP2). Zorritos 4° 5° RESULTADOS Temperatura 8° La temperatura superficial del mar (TSM) tuvo un rango de 14,02 a 24,14 °C. Los valores <16 °C, asociados al afloramiento costero se ubicaron dentro de las 20 mn entre Salaverry y Callao, y por fuera de las 90 mn amplió su distribución al sur del Callao. Las temperaturas >24 se localizaron frente a Zorritos relacionadas con Aguas Tropicales Superficiales (Figura 2). Anomalías térmicas Las anomalías térmicas de la superficie del mar (ATSM) se han registrado desde –1,45 °C (Chimbote) a +5,32 °C (Talara). Las condiciones ambientales fueron cálidas al norte de Pimentel, resaltando un núcleo muy cálido frente a Talara con anomalías > +5 °C. Al sur de Salaverry las condiciones fueron casi normales con anomalías entre –0,5 a +0,5 °C exceptuando la zona oceánica afuera de las 60 mn al norte de Chimbote, donde las condiciones fueron frías con anomalías negativas > –1,0 °C (Figura 3). Salinidad En la superficie del mar se registraron concentraciones de sales que variaron de 33,081 a 35,25 ups. Las concentraciones <34,00 ups se localizaron al norte de Cabo Blanco, asociadas a aguas del Golfo de Crucero 2003/08-09 PROFUNDIDAD DE LA ISOTERMA DE 15 C (m) Paita Pta. Gobernador Parachique Pta. Falsa 6° 7° En general, la distribución térmica presentó un comportamiento anómalo, observándose un fuerte contraste térmico al norte de Mórrope, asociado al desplazamiento de aguas cálidas del Golfo de Guayaquil hasta los 7°S, debido al arribo de una onda Kelvin. En el sur, la distribución fue homogénea apreciándose solo dos isotermas (16 y 17°C). Pta. Sal Cabo Blanco Talara Mórrope Pimentel Chérrepe Pacasmayo Chicama Trujillo Salaverry Guañape Punta Chao 9° Chimbote Casma Punta Lobos 10° Huarmey Punta Bermejo Supe 11° Huacho Punta Salinas Chancay Ancón Callao Punta Hermosa v Bujama Cerro Azul Punta Cóndor Tambo de Mora Pisco Peninsula Paracas 12° 13° 14° Bahía Independencia Punta Infiernillos 15° 83° 82° 81° 80° 79° 78° 77° 76° 75° 74° Figura 7.- Profundidad de la Isoterma de 15°C (m). Crucero Biomasa Desovante 0308-09 Guayaquil desplazadas por las ondas Kelvin, y el relajamiento de los vientos. Las concentraciones >35,20 ups, asociadas a aguas oceánicas, se localizaron a 30 mn formando un núcleo frente a Supe. Las concentraciones moderadas asociadas a procesos de afloramiento se distribuyeron ampliamente superando las 100 mn al norte de Chimbote y al sur de Cerro Azul (Figura 4). Vientos La intensidad de los vientos varió entre 1,1 a 15,1 m/s, predominando vientos moderados a lo largo del área evaluada. Algunos núcleos con vientos fuertes al sur del Callao, donde resalta un núcleo con intensidades >12 m/s y una zona próxima a la costa con bajas intensidades entre Cerro Azul y Pisco (Figura 5). Respecto a la dirección del viento, al norte de Talara se observó una predominancia de vientos del suroeste; y al sur de esta localidad, la tendencia dominante fue del sureste con excepción de la zona con vientos, débiles donde no se observó una dirección 111 predominante. Turbulencia En general los índices de turbulencia fueron bajos (<500 m3/s3) al norte de Casma, con excepción de las zonas comprendidas frente a Punta Falsa - Pimentel y entre Talara - Paita, donde los valores de turbulencia fueron entre 500 y 1000 m3/s3. Al sur de Casma mostraron un amplio rango de variación, con los valores más bajos en la zona costera. Por fuera de las 30 mn resaltaron tres áreas con valores moderados (1000 a 2000 m3/s3) y una con valores altos (>2500 m3/ s3) (Figura 6). DISTRIBUCIÓN SUBSUPERFICIAL Isoterma de 15 °C La profundidad de la isoterma de 15 °C varió de 6 a 136 m (Figura 7). Las profundidades >120 m se localizaron entre Talara y Paita y las menores, próximas a la costa entre Cerro Azul y Pisco. El comportamiento de las isobatas Informe IMARPE Vol. 35(2): Abril-Junio 2008 25 23 AES 17 15 AESS 13 11 7 21 ASS ACF 19 TEMPERATURA TEMPERATURA ASS ACF 19 23 21 19 TEMPERATURA ATS 21 9 25 25 23 17 15 AESS 13 11 CALETA LA CRUZ PAITA PUNTA FALSA AII 9 CHIMBORE 4 CHICAMA 7 5 33.2 33.4 33.6 33.8 34.0 34.2 34.4 34.6 34.8 35.0 35.2 35.4 SALINIDAD AII ACF 17 ASS 15 AESS 13 11 SUPE 9 CALLAO PISCO AII 7 5 33.2 33.4 33.6 33.8 34.0 34.2 34.4 34.6 34.8 35.0 35.2 35.4 5 33.2 33.4 33.6 33.8 34.0 34.2 34.4 34.6 34.8 35.0 35.2 35.4 SALINIDAD SALINIDAD Figura 8.- Distribución en la columna de agua de las Masas de Agua a) Norte, b) Centro y Sur. Cr. Biomasa Desovante 0308-09 SECCION CALETA LA CRUZ (16-17/08/2003) TEMPERATURA (°C) m 0 50 50 100 m secciones asociadas a temperaturas <15 °C, salinidades >34,9 ups; y SALINIDAD ( ups) 0 m 0 50 50 100 100 150 150 200 200 250 250 300 300 350 350 400 400 450 450 Aguas Antárticas Intermedias (AAI) con salinidad <34,7 ups. Solo se pudo observar el borde superior de esta masa de agua, ya que la profundidad de muestreo fue de 500 m. 100 150 150 200 200 250 300 SECCION PAITA (18-19/08/2003) TEMPERATURA (°C) SALINIDAD (ups) m 0 ESTRUCTURA VERTICAL 250 150 120 90 60 30 mn 300 150 120 90 60 30 mn 0 500 100 80 60 40 20 0 mn 500 100 80 60 40 20 0 mn Figura 9.- Distribución Vertical de a) Temperatura (°C) y b) Salinidad (ups), frente a caleta La Cruz. Cr. Biomasa Desovante 0308-09 Figura 10.- Distribución Vertical de a) Temperatura (°C) y b) Salinidad (ups), frente a Paita. Cr. Biomasa Desovante 008-09 indicó flujos de norte a sur desde la frontera norte hasta Chicama; al sur de esta localidad, la circulación marina mostró flujos hacia el norte, con la zona de convergencia entre Pacasmayo y Salaverry. >18°C y <34,8 ups. También se encontraron desplazadas más al sur de su ubicación normal (más de 120 mn), llegaron hasta Pimentel, y alcanzaron profundidades de 80 m frente a Paita, y 25 m frente a Punta Falsa. MASAS DE AGUA Aguas Subtropicales Superficiales (ASS) con salinidad >35,1 ups y temperaturas >18°C, estuvieron presentes entre los 6°30’S y 13°S en el borde exterior del área de estudio, proyectándose hasta las 15 mn frente a Supe, mostrando su máxima profundidad (50 m) frente a Chimbote, Supe y Callao. Frente a la costa peruana se presentaron las siguientes masas de agua (Figura 8): Aguas Tropicales Superficiales (ATS) con salinidad <34,0 ups y temperaturas >22 °C. Se ubicaron al norte de los 6°S dentro de las 70 mn y con un espesor de 20 m frente a Paita y 5 m frente a Punta Falsa; masas de agua que en mayo-junio se localizaron al norte de los 4°S frente a Punta Sal (Crucero 030506). Aguas Ecuatoriales Superficiales (AES) con índices termohalinos En la sección caleta La Cruz (Figura 9), dentro de las 100 mn, se observó un fuerte contraste térmico asociado a la convergencia de un flujo de aguas cálidas al sur, con otro de aguas frías hacia el norte. La isoterma de 15 °C se ubicó entre los 80 y 105 m de profundidad. Las masas de agua presentes fueron ATS hasta los 30 m de profundidad (a 60 mn), las AES ubicadas sobre los 60 m (a 30 mn) en la zona costera, ascendiendo sobre los 20 m en la zona oceánica (a 150 mn) y las ACF por debajo de las AES. Aguas Costeras Frías (ACF) asociadas a procesos de afloramiento se encontraron ampliamente distribuidas al sur de Chimbote y al sur de Cerro Azul. La sección Paita (Figura 10), dentro de las 100 mn, mostró un fuerte contraste térmico asociado a la convergencia de un flujo de aguas cálidas al sur, con otro de aguas frías hacia el norte. La isoterma de 15 °C se ubicó entre los 50 m (100 mn) y 150 m (costa) de profundidad. Las ATS se ubicaron sobre los 20 m de profundidad entre las 20 y 70 mn de la costa. Las AES se encontraron desde la superficie (a las 90 mn) hasta los 80 m en la zona próxima a la costa y las ACF por fuera de las 90 mn y debajo de las AES. Aguas Ecuatoriales Subsuperficiales (AESS). Se hallaron en la capa subsuperficial, en todas las Sección Punta Falsa (Figura 11); mostró una termoclina moderada que se debilita conforme se aleja de 112 Condiciones oceanográficas físicas en invierno 2003 Vásquez la costa y ubicando a la isoterma de 15 °C entre los 30 m (a 100 mn) y 120 m de profundidad cerca de la plataforma continental. Por otro lado, se observó una ligera presencia de ATS sobre los 5 m; AES dentro de las 60 mn, y hasta 20 m de profundidad cerca de la costa y ACF por debajo de las AES y mezcladas por fuera de las 80 mn. En la Sección Chicama (Figura 12) no se observó una temoclina definida debido a procesos de surgencia. La isoterma de 15 °C se ubicó entre los 55 y 75 m, estuvo ligeramente más profunda en la zona costera respecto a la zona oceánica. Las ACF predominaron en toda la columna de agua con excepción de una pequeña área por fuera de las 80 mn y sobre los 40 m donde se encontraron ASS. En la Sección Chimbote (Figura 13), se observó una termoclina muy débil que desapareció dentro de las 30 mn. La isoterma de 15 °C se localizó entre los 75 y 20 m de profundidad. En tanto la distribución halina muestra ASS por fuera de las 80 mn y sobre los 55 m de profundidad, el resto de la sección estuvo cubierta por ACF. La Sección Supe (Figura 14), presentó una distribución térmica homogénea no se observó una termoclina definida; la isoterma de 15 °C se localizó a los 75 m de profundidad entre las 70 y 50 mn ascendiendo rápidamente hasta los 50 m, manteniéndose hasta la costa en esta. La distribución halina indica la presencia de ASS sobre los 50 m y entre las 15 y 50 mn, el la zona costera y por fuera de las 50 mn se ubicaron aguas de mezcla por la interacción de las ASS y ACF. En la Sección Callao (Figura 15), la estructura térmica no presentó una termoclina definida, la isoterma de 15 °C se ubicó entre los 50 m (90 mn) y la superficie (10 mn de la costa), en tanto la estructura halina mostró la incursión de ASS entre las 30 y 50 mn, separando a las ACF en dos zonas, dentro de las 20 mn y por fuera de las 70 mn. En la Sección Pisco (Figura 16) tampoco se observó una termoclina. La isoterma de 15 °C SECCION PUNTA FALSA (21-22/08/2003) TEMPERATURA (°C) 0 m0 50 m SECCION CHICAMA (26-27/08/2003) TEMPERATURA (°C) SALINIDAD ( ups) SALINIDAD ( ups) m0 m0 50 50 50 100 100 100 100 150 150 150 150 200 200 200 200 250 250 250 250 300 300 300 300 350 350 350 350 400 400 400 400 450 450 450 450 500 100 80 60 40 20 0 mn 500 100 80 60 40 20 0 500 100 mn Figura 11.- Distribución Vertical de a) Temperatura (°C) y b) Salinidad (ups), frente a Punta Falsa. Crucero 0308-09 50 0 50 50 100 100 150 150 200 200 250 250 300 300 350 350 400 400 450 450 500 100 80 60 40 20 0 500 100 TEMPERATURA (°C) 60 40 20 0 50 50 100 100 150 150 200 200 250 250 0 Figura 13.- Distribución Vertical de a) Temperatura (°C) y b) Salinidad (ups), frente a Chimbote. Cr. Biomasa Desovante 0308-09 60 50 50 100 100 150 150 200 200 250 250 300 300 350 40 20 0 300 0 50 50 100 100 150 150 200 200 250 250 300 300 350 350 350 400 400 400 400 450 450 450 450 80 60 40 20 0 500 80 60 40 20 Figura 15.- Distribución Vertical de a) Temperatura (°C) y b) Salinidad (ups), frente a Callao. Crucero Biomasa Desovante 0308-09 113 40 20 0 SALINIDAD ( ups) TEMPERATURA (°C) 0 500 60 SECCION PISCO (14-15/09/2003) SALINIDAD (ups) 0 0 mn Figura 14.- Distribución Vertical de a) Temperatura (°C) y b) Salinidad (ups), frente a Supe. Cr. Biomasa Desovante 0308-09 SECCION CALLAO (11/09/2003) TEMPERATURA (°C) 0 50 SALINIDAD ( ups) 0 300 80 100 SECCION SUPE (06-07/09/2003) SALINIDAD ( ups) 0 500 Figura 12.- Distribución Vertical de a) Temperatura (°C) y b) Salinidad (ups), frente a Chicama. Cr. Biomasa Desovante 0308-09 SECCION CHIMBOTE (03-04/09/2003) TEMPERATURA (°C) 0 mn 0 500 80 60 40 20 0 500 80 60 40 20 0 Figura 16.- Distribución Vertical de a) Temperatura (°C) y b) Salinidad (ups), frente a Pisco. Crucero Biomasa Desovante 0308-09 Informe IMARPE Vol. 35(2): Abril-Junio 2008 se ubicó a los 40 m descendiendo ligeramente hasta los 55 m para luego ascender rápidamente alcanzando la superficie a las 30 mn. Las concentraciones de sales indican la presencia de ACF en toda la columna de agua. CONCLUSIONES Las condiciones oceanográficas que prevalecieron durante el desarrollo del crucero mostraron anormalidades positivas al norte de los 7°S. A inicios de agosto el arribo de una onda Kelvin que, sumado al relajamiento de los vientos alisios del sureste, contribuyeron al desplazamiento de las ATS, con el consecuente incremento térmico de hasta +5°C frente a Talara-Paita, así como la profundización de la termoclina en 47 m (Zuta 1990) frente a Punta Falsa. En general las condiciones ambientales encontradas en el presente crucero fueron ligeramente mas frías que las registradas en agosto del 2002, debido a una menor presencia de ASS en el área evaluada, exceptuando la zona entre caleta La Cruz y Pimentel donde las condiciones fueron más cálidas asociada a la presencia de aguas del Golfo de Guayaquil. Al sur las condiciones fueron casi normales con excepción de la zona oceánica (fuera de las 70 mn) entre Punta Falsa y Chimbote donde las condiciones fueron frías. 114 REFERENCIAS IMARPE. 2002. Informe Ejecutivo, Crucero de Evaluación Biomasas Desovante en el Invierno del 2002. IMARPE. 2003. Informe Ejecutivo, Crucero de Evaluación Hidroacústica de Recursos Pelágicos en el Verano del 2003. Reynolds RW. 1982. A monthly average climatology of Sea Surface Temperature Tecnical Report NWS31. National Meteorological Center, NOAA, Silver Spring MD. 1-35. Zuta S, Guillén O. 1970. Oceanografía de las Aguas Costeras del Perú, Bol. Inst. Mar Perú Vol. 2 (3). Zuta S. 1990. Variations of the Mass Field and Currents off the Peru Cost. Bol. No. 33, International Oceanographic Commission, UNESCO Biomasa desovante de la anchoveta peruana (MPH). Invierno 2004 Ayón y Perea BIOMASA DESOVANTE DE LA ANCHOVETA DEL STOCK NORTE CENTRO EN EL INVIERNO 2004 ANCHOVETA SPAWNING BIOMASS OF THE NORTH AND CENTRAL STOCK DURING WINTER 2004 Patricia Ayón1 Ángel Perea2 RESUMEN AyÓn P, Perea A. 2008. Biomasa desovante de la anchoveta del stock norte centro en el invierno 2004. Inf. Inst. Mar Perú 35(2):115-117.- El crucero BIC Olaya y SNP2 0408-09 tuvo el objetivo de estimar la biomasa desovante de la anchoveta (Engraulis ringens) del stock norte centro (4 a 15°S) durante el invierno 2004. Se utilizó el método de producción de huevos (MPH). El total estimado fue de 4,2 millones de toneladas. El peso promedio para las hembras fue de 18,2 g, la proporción sexual fue de 0,5; la fecundidad parcial estimada fue de 15.255 ovocitos/hembra, con una frecuencia de desove cuyo porcentaje fue 9,4% y una producción diaria de huevos de 16,506E+13 huevos / día. El parámetro con mayor variabilidad fue la frecuencia de desove con 22,9%; la producción diaria de huevos tuvo un CV de 3,5%. Palabras clave: biomasa desovante, anchoveta peruana, producción diaria de huevos. ABSTRACT AyÓn P, Perea A. 2008. Anchoveta spawning biomass of the north and central stock during winter 2004. Inf Inst Mar Peru 35 (2): 115-117.- The BIC Olaya and SNP2 0408-09 cruise aimed to estimate the spawning biomass of anchoveta (Engraulis ringens) in the north-central stock (4 to 15°S) during winter 2004, using the method of egg production (EPM). The estimated total biomass was 4.2 million tons. The average weight for females was 18.2 g, the sex ratio was 0.5, and the batch fecundity was 15,255 oocytes per mature female, with 9.4% spawning frequency and a daily egg production of 16.506 E +13 eggs / day. The parameter with greater variability was spawning frequency of 22.9%, while the daily egg production had a CV of 3.5%. Keywords: spawning biomass, Peruvian anchoveta, daily egg production. INTRODUCCIÓN Los cruceros para evaluar la biomasa desovante de la anchoveta por el método de producción de huevos (MPH), provee información de aspectos biológicos, para entender mejor el comportamiento de la especie. Asimismo este método permite trabajar cada variable del modelo de manera independiente, y se puede ajustar cada una de ellas sin afectar a las otras. En los últimos años los resultados obtenidos mediante este método han sido coherentes con los métodos acústicos así como los de análisis de población virtual. Sin embargo, la biomasa desovante estimada en el invierno 2003, resultó baja, hecho que mereció un análisis más profundo y explicar cuales factores podrían haber influido 1 2 para que esta biomasa no fuera similar a la obtenida por el método acústico. En el análisis de los datos biológicos, tanto reproductivos como la composición de tallas y la producción de huevos, se pudo concluir que esa diferencia se debía por un lado a que un alto porcentaje de la biomasa (más del 60%) correspondía a juveniles, asimismo durante todo el año 2003 la anchoveta no tuvo un periodo de reposo, por lo que el porcentaje de desovantes habría sido menor en el momento de la evaluación. Este informe presenta los resultados del estimado de la biomasa desovante de la anchoveta para el stock norte centro del mar peruano (4 to 15°S), durante el invierno 2004. Asimismo se da a conocer los valores de los parámetros involucrados en el modelo. Area de Evaluación de Producción Secundaria. [email protected] Laboratorio de Biología Reproductiva. [email protected] 115 MATERIAL Y MÉTODOS El crucero BIC Olaya y SNP2 0408-09, para estimar la biomasa desovante de la anchoveta en el invierno 2004, se efectuó del 6 de agosto y el 8 de setiembre del 2004, entre Cabo Blanco (4°S) y Punta Infiernillos (15°S). El BIC SNP2 trabajó en el área costera, es decir entre las 0,5 y 32 mn de la costa; el BIC Olaya, desde las 32 millas hasta una distancia promedio de 90 mn; frente al Callao se llegó hasta por fuera de las 200 millas. El diseño de muestreo empleado para los adultos fue similar al dado por Santander et al. (1984). Para la producción diaria de huevos se utilizó el diseño planteado por AyÓn (2008), en donde todos los perfiles no tienen igual longitud y se modifican de acuerdo con la distribución de los huevos de la Informe IMARPE Vol. 35(2): Abril-Junio 2008 de 2%. En la fecundidad parcial se obtuvo una regresión lineal entre la fecundidad y el peso sin gónada, para lo cual se utilizaron hembras con ovocitos hidratados con LT entre 12,5 y 16,5 cm. Esta regresión tuvo un coeficiente de correlación de 0,6; y se obtuvo una fecundidad parcial promedio de 15.255 ovocitos/hembra con un CV de 2,2%. Talara Paita Pta. Gobernador Parachique Pta. Falsa 6°S Mórrope Pimentel Chérrepe Pacasmayo Chicama 8°S Trujillo Salaverry Guañape Punta Chao Chimbote En la frecuencia de desove (F) se trabajó con 1175 hembras, provenientes de 47 calas, y se obtuvo una F equivalente a 0,094 es decir que el 9,4% de las hembras se encontraba desovando cada día. Casma Punta Lobos 10°S Huarmey Punta Bermejo Supe Huacho Punta Salinas Chancay Ancón 12°S Callao Punta Hermosa Bujama Cerro Azul Punta Cóndor Tambo de Mora Pisco Peninsula Paracas 14°S ProducciÓn diaria de huevos Bahía Independencia Punta Infiernillos Punta Caballas Los valores de abundancia de huevos (Po) de anchoveta por categoría de edad fueron: San Nicolás San Juan 82° W 80° W 78° W 76° W 74° W Figura 1.- Carta de Estaciones de muestra (red CalVET). anchoveta encontrados durante el desarrollo del crucero. La distancia entre los perfiles para este crucero fue de 15 mn, habiéndose recorrido un total de 49 perfiles (Figura 1). Se colectaron 958 muestras para determinar la producción diaria de huevos, que fueron obtenidos mediante una red CalVET jaladas de 70 a 0 m de profundidad. En los lugares donde la profundidad era menor que 70 m, se lanzó a 5 metros menos del fondo del lugar. los huevos se basó en el trabajo de Moser y Ahlstrom (1985), para la asignación de la categoría de edad y el cálculo de la producción diaria de huevos (Po) se siguió con Santander et al. (1984), donde se utiliza un sistema gráfico de Zweifel & Smith que considera el desarrollo embrionario, la temperatura del mar y la hora de la colección. Para el estimado de Po se hizo una iteración con 4 categorías de edad: A1, A2, B1 y B2. Se efectuaron 91 calas, considerándose válidas para el método 47 de ellas, y en el caso de la proporción sexual se trabajó con 43 calas. Para la regresión de la fecundidad se utilizaron 60 hembras hidratadas con rangos de LT entre 12,5 y 16,5 cm. Se aplicó una regresión lineal, entre la fecundidad parcial y el peso sin gónada, obteniéndose un coeficiente de correlación de 0,6. Los cálculos para los parámetros de peso, frecuencia de desove, proporción sexual y fecundidad se hicieron de acuerdo con Santander et al. (1984). Para el cálculo de la frecuencia de desove se consideró solamente los folículos del día 1. Las ecuaciones empleadas para el cálculo de los parámetros y el estimado de la biomasa desovante fueron las de Santander et al. (1984). Para el estimado de las covarianzas entre los parámetros se utilizó la metodología propuesta por Armstrong et al. (1988), en la que se consideraron regiones (AyÓn 2008). La asignación de los estadios de RESULTADOS Parámetros de adultos La proporción sexual (R) fue de 0,50 con un coeficiente de variación (CV) de 3%; el peso promedio (W) fue de 18 g por hembra, y un CV 116 A1 (0-20 h) A2 (12-32 h) B1 (24-44 h) B2 (36-56 h) : 11,90 E+13 : 7,67 E+13 : 4,80 E+13 : 3,54 E+13 Se obtuvo una producción diaria de huevos de 16,506 E+13 huevos/día, coeficiente de variación de 3,51% y tasa de mortalidad de 0,832 que significa que aproximadamente el 56,48% de los huevos están muriendo cada día. Biomasa de la anchoveta La biomasa desovante de la anchoveta para el área comprendida entre Cabo Blanco y Punta Infiernillos obtenida por el MPH fue de 4,2 millones de toneladas métricas, con CV de 3,52. En la Tabla 1 se presentan los valores obtenidos de todos los parámetros con sus respectivos estadísticos. DISCUSIÓN El estimado de la biomasa desovante de la anchoveta para el invierno 2004 (4.200.000 t) ha sido mayor que el año 2003 (1.500.000 t). En invierno 2003, al efectuar la evaluación, se observó que el proceso reproductivo había sido continuo y algunos indicadores mostraban que no se había dado Biomasa desovante de la anchoveta peruana (MPH). Invierno 2004 Ayón y Perea Tabla 1. Valores de los parámetros y biomasa desovante de la anchoveta. Invierno 2004. Parámetro R W E Promedio 0,50 18,20 15255,10 0,09 1,65E+14 4,22E+07 Varianza 0,00 0,13 115986,84 0,00 3,38E+25 2,20E+12 Desv. Est. 0,02 0,36 340,57 0,02 5,81E+12 1,48E+06 Coef. Variac. 3,03 2,00 0,02 22,89 3,52 3,52 el periodo de reposo reproductivo, común en esta especie en la estación de otoño. Por otro lado, el porcentaje de individuos juveniles en las capturas había sido del 74,71% equivalente al 50,42% de la biomasa total. Para el crucero 2004, la situación ha sido diferente. Por un lado, las condiciones previas al crucero mostraban un claro reposo reproductivo durante la estación de otoño, es decir el recurso aparentemente mostraba un comportamiento normal para la estación. En el inicio de la evaluación, el proceso reproductivo había mostrado un incremento; pero durante los muestreos de rutina y análisis microscópicos realizados a bordo, se pudo comprobar que éste aún no registraba valores significativamente altos (sobre el valor crítico 20%) excepto en la zona sur, pudiendo inclusive notar la grasa visceral o vestigios perfectamente reconocibles, indicando que estos individuos habrían tenido una buena alimentación previa al periodo de desove y que aún no gastaban toda esta energía acumulada en los procesos masivos de maduración y desove, como suele suceder durante los periodos importantes de reproducción. De acuerdo con los estimados encontrados para los parámetros de adultos se aprecia que los valores de peso, proporción sexual y frecuencia de desove son comparativamente muy similares a los del 2003; sin embargo, se nota que en relación a la fecundidad y la producción diaria de huevos estos se F Po B habrían incrementado notablemente. En el caso de la fecundidad parcial el incremento fue de casi un 30%, y la producción diaria de huevos se incrementó en casi 5 veces su valor. Lo interesante en este crucero 0408-09 es que los valores obtenidos para los coeficientes de variación han sido bastante bajos en comparación con cruceros anteriores. Lo más notorio es el caso de la producción diaria de huevos, que corresponde al CV más bajo de los que se ha determinado desde su aplicación. Las posibles causas de esta reducción pueden deberse a que el muestreo cubrió adecuadamente el área de distribución del recurso anchoveta. Solamente en el caso de la frecuencia de desove este coeficiente fue alto, debido al momento reproductivo encontrado durante la evaluación, caracterizado por no hallarse a la población en un periodo de máxima intensidad del desove, lo cual generó una alta variabilidad entre los cardúmenes; situación contraria a la que se hubiese observado si la evaluación se hubiese realizado en pleno pico de desove. El estimado de la biomasa desovante de la anchoveta ha sido menor al obtenido acústicamente, sin embargo como se menciona en AyÓn y BuitrÓn (2008) la biomasa acústica incluye toda la biomasa, sin distinguir la estructura de tallas, ni las condiciones reproductivas de los individuos. 117 CONCLUSIONES • La biomasa desovante de la anchoveta para el stock norte centro en el invierno 2004 fue estimada en 4,2 millones de toneladas. • El estimado de la biomasa desovante corresponde al momento en que el proceso reproductivo se encontraba recién en su fase de ascenso del máximo pico del desove. Agradecimiento.- A la señora Blga. Soledad Guzmán por la revisión del manuscrito y sus oportunos consejos. Agradecimiento al personal del Área de Evaluación de Producción Secundaria y Biología Reproductiva por el análisis de las muestras y apoyo desinteresado. REFERENCIAS Armstrong M, Shelton P, Hampton I, Jolly G, Melo Y. 1988. Egg production of anchovy biomass in the southern Benguela. Calcofi Rep. Vol 29:137-157. AyÓn P. 2008. Producción diaria de huevos de la anchoveta peruana en agosto-setiembre 2003. Inf. Inst. Mar Perú 35(2): 81-85. AyÓn P, BuitrÓn B. 2008. Biomasa desovante de la anchoveta peruana. Método de Producción de Huevos (MPH) invierno 2003. Inf. Inst. Mar Perú 35(2): 77-80. Moser H. y Ahlstrom E. 1985. Staging anchovy eggs. En: Reuben Lasker (ed.) An Egg production Method for Estimating Spawning Biomass of Pelagic Fish: Application to the Northern Anchovy, Engraulis mordax. NOAA Technical Report NMFS 36: 37-41. Santander H, Alheit J, Smith P. 1984. Estimación de la biomasa de la población desovante de anchoveta peruana Engraulis ringens en 1981 por aplicación del Método de Producción de Huevos. Bol. Inst. Mar Perú 8(6):209-250. Fitoplancton en invierno 2004 Sánchez y Jacobo EL FITOPLANCTON DURANTE EL INVIERNO 2004 PHYTOPLANKTON DURING WINTER 2004 Sonia Sánchez Nelly Jacobo1 RESUMEN Sánchez S, Jacobo N. 2008. El fitoplancton durante el invierno 2004. Inf. Inst. Mar Perú 35(2):119-122.- Se dan a conocer las principales características de la comunidad fitoplanctónica estudiadas durante el Crucero BIC Olaya y SNP2 0408-09. Los volúmenes de plancton variaron entre 0,06 y 8,07 mL/m3, promedio 0,69 mL/ m3. El rango de la TSM fue 13,9 a 22,3 °C. El fitoplancton fue dominante en el 54% del área evaluada. Ceratium breve, indicador de aguas ecuatoriales superficiales (AES) fue registrado frente a Paita; C. praelongum y C. incisum, indicadores de aguas subtropicales superficiales (ASS), entre Paita y Punta Falsa y entre Huacho y Pucusana. Protoperidinium obtusum indicador de Aguas Costeras Frías (ACF) se localizó en la zona costera desde punta Falsa hasta punta Mendieta. Palabras clave: fitoplancton, indicadores de masas de agua, invierno 2004, mar peruano ABSTRACT Sánchez S, Jacobo N. 2008. The phytoplancton during winter 2004. Inf. Inst. Mar Perú 35(2):119-122.This paper informs about the main characteristics of the phytoplankton community studied during the BIC Olaya and SNP2 0408-09 cruise. Plankton volumes varied between 0.06 and 8.07 mL/m3, average 0.69 mL/m3. The SST range was 13.9 to 22.3 °C. The phytoplankton was dominant in 54% of the area evaluated. Ceratium breve, indicator of equatorial surface waters (ESW) has been registered in Paita offshore, C. praelongum and C. incisum, indicators of subsurface equatorial waters (SEW), between Paita and Punta Falsa and between Huacho and Pucusana. Protoperidinium obtusum indicator cold coastal waters (CCW) was located in the coastal area from Punta Falsa until Punta Mendieta.. Keywords: phytoplankton, indicators of water masses, winter 2004, Peruvian sea. INTRODUCCIÓN MATERIAL Y MÉTODOS Las condiciones ambientales en invierno 2004 en la zona norte del litoral peruano (hasta los 6°S), tendieron a ser frías a inicios de agosto; luego pasaron a condiciones cálidas dentro de las 60 mn a fines de agosto, debido al desplazamiento latitudinal de la onda Kelvin (Vásquez 2004). En este panorama, la comunidad fitoplanctónica en general incrementó sus concentraciones, y llegó a ser dominante en el 54% de las estaciones analizadas, dentro de las 30 mn, y amplió su distribución por fuera de las 90 mn frente a Paita, Salaverry y Chimbote. El Crucero de estimación de biomasa desovante de anchoveta por el método de producción de huevos (MPH), BIC José Olaya Balandra y SNP2 0408-09, se realizó del 6 agosto al 8 setiembre 2004 con una cobertura máxima de 200 mn desde la zona costera frente a Talara (4°S) a Punta Mendieta (15°S). Este informe describe las características latitudinales de distribución de la comunidad planctónica en general (volúmenes de plancton), con énfasis en la distribución y composición por especies del fitoplancton de superficie, y la presencia de los organismos indicadores de masas de agua. 1 Se obtuvo un total de 88 muestras de superficie colectadas según el método establecido por el IMARPE, y fijadas siguiendo las pautas descritas en TrÖndsen (1978). Para la determinación de los volúmenes se han seguido las recomendaciones de la reunión del Programa del Plancton (UNESCO 1981). Para los análisis semicuantitativos se consideraron los principales grupos del plancton (fitoplancton – zooplancton), otorgando a los organismos más representativos valores convencionales de acuerdo a Unidad de Producción Primaria. DIO. [email protected] 119 su abundancia, de acuerdo la metodología establecida por el Área de Fitoplancton y Producción Primaria del IMARPE. Para la determinación taxonómica de la flora planctónica se consultaron los trabajos de Hasle y Syvertsen (1996), Steidinger y Tangen (1996), Balech (1988), SundstrÖm (1986), Schiller (1971), Sournia (1967), Hendey (1964), Cupp (1943) y Hustedt (1930). RESULTADOS DistribuciÓn de los volÚmenes de plancton y composiciÓn de la comunidad fitoplanctÓnica en superficie Durante agosto – setiembre 2004 los volúmenes de plancton fluctuaron entre 0,06 y 8,07 mL/m3; el 90% de los volúmenes fueron menores a 1,0 mL/m3; el promedio fue 0,69 mL/m3, asociados a rangos de temperatura superficial del mar Informe IMARPE Vol. 35(2): Abril-Junio 2008 80°W 78°W 76°W 74°W 84°W 72°W 82°W 80°W 4°S Pta. Sal 4°S Paita Pta. Gobernador Paita Pta. Gobernador 6°S Pta. Falsa 6°S Pimentel Chicama 8°S Salaverry Chicama 8°S Punta Chao Chimbote Casma 10°S Huarmey 10°S Supe Huacho Chancay Chancay 12°S Callao Pucusana 12°S Pucusana Cerro Azul Cerro Azul 16°S Punta Caballas San Juan Chala 16°S Atico 18°S 84°W 82°W 80°W Punta Caballas San Juan Chala 16°S Ilo 78°W 76°W 74°W 18°S Figura 1.- Distribución de los volúmenes de plancton en superficie (mL/m3). Crucero de Estimación de biomasa Desovante 0408-09. (TSM) entre 13,9 y 22,3 °C. El fitoplancton fue dominante en 54% de las estaciones, dentro de 30 mn, pero ampliando su distribución por fuera de las 90 mn frente a Paita, Salaverry y Chimbote. El zooplancton predominó en el 38% y la codominancia de fitoplancton y zooplancton fue del 8% (Figura 1). Se registraron núcleos con altas concentraciones de biomasa planctónica. El primer núcleo fue de 8,0 mL/m3, localizado al sur de Punta Gobernador y dentro de las 30 mn. Otros dos núcleos >3,0 mL/m3, uno entre Huarmey y Chancay donde el fitoplancton fue dominante, y el otro frente a Punta Falsa por fuera de las 50 mn, donde dominó el zooplancton. Así mismo se presentaron dentro de las 30 mn otros núcleos >1 mL/m3 uno frente a Chicama - Salaverry y el otro entre Pisco y Punta Mendieta (Figura 1). Al norte de Punta Falsa se determinaron especies de fases intermedias en la sucesión como las diatomeas Guinardia flaccida, Thalassiosira subtilis, Detonula confervacea, Proboscia alata v. indica y Licmophora abbreviata, asociada a especies de etapas tempranas en 82°W 80°W 78°W Ilo 18°S 76°W 74°W 72°W Figura 3.- Distribución de indicadores de masas de agua. Cruceros de Estimación de biomasa Desovante 0408-09. la sucesión como Chaetoceros compressus, Ch. didymus, Ch. debilis, Ch curvisetus. Al sur de Punta Falsa el fitoplancton se caracterizó por la presencia de Lithodesmiun undulatum, Actinocyclus sp., Coscinodiscus perforatus, Thalassiosira angulata, Th. anguste-lineata, Pleurosigma sp., Ch. debilis, Ch. affinis, Ch. curvisetus, entre otras. Los dinoflagelados estuvieron representados por especies como Ceratium furca, C. tripos, C. dens, Protoperidinium leonis, P. pentagonum, P. mendiolae y Gonyaulax polygramma, las cuales destacaron por su abundancia entre Callao y Cerro Azul. Ceratium dens se localizó frente a Talara, dentro de las 30 mn. Las diatomeas y dinoflagelados termófilos se localizaron principalmente frente a Punta Falsa y entre Supe - Pucusana, destacando Goniodoma polyedricum, Ceratium lunula, C. gravidum, C. massiliense, Ornithocercus quadratus, Pyrocystis lunula y P. fusiformis, Durante el desarrollo del crucero se observó la ocurrencia de una marea roja, ocasionada por Messodinium rubrum frente a Pimentel, aproximadamente a 30 mn de la costa. 120 Quilca Matarani ACF : Protoperidinium obtusum AES : Ceratium breve ASS : Ceratium incisum, C. praelongum 18°S 84°W 72°W 16°S Atico Quilca Matarani Fitoplancton Zooplancton Fitoplancton y Zooplancton 14°S Pta. Mendieta DistribuciÓn latitudinal de los volÚmenes de plancton Latitudinalmente la mayor concentración de biomasa planctónica promedio (3,12 mL/m3), se localizó a los 5 – 6 °S (dentro de las 30 mn, línea verde), y la menor (0,06 mL/ m3) en dos áreas: a los 5 – 6°S (30 – 60 mn, línea roja) y a los 8 – 9°S (por fuera de las 60 mn, línea azul) (Figura 2). En la latitud de 5-6°S, dentro de las 30 mn de la costa, se reportaron los valores máximos de biomasa planctónica promedio (3,12 mL/ mL/m3 6’ - 7’ 1 Pisco 5’ - 6’ 14°S Pta. Mendieta 14°S 4’ - 5’ 1 Pisco 14°S 12°S Callao 15’ - 16’ 12°S 10°S Huarmey Supe 3 Huacho 14’ - 15’ 10°S 8°S Salaverry Punta Chao Chimbote Casma 13’ - 14’ 1 11’ - 12’ 0.5 1 12’ - 13’ 3 6°S Pta. Falsa Pimentel 8°S 4°S 10’ - 11’ 1 72°W Talara 8’ - 9’ 6°S 74°W Pta. Sal Talara 8 76°W Pto. Pizarro Pto. Pizarro 4°S 78°W 9’ - 10’ 82°W 7’ - 8’ 84°W Figura 2.- Distribución latitudinal de los volúmenes de plancton (mL/m3). Crucero de biomasa desovante 0408-09. Fitoplancton en invierno 2004 Sánchez y Jacobo m3). Entre 30 – 60 mn los volúmenes promedio fueron <1 mL/m3. Por fuera de las 60 mn, se registró un valor máximo de 1,31 mL/m3 entre 6–7°S. Al norte y sur de esta latitud los valores promedio fueron descendiendo. DistribuciÓn de organismos indicadores de masas de agua Ceratium breve, indicador de Aguas Ecuatoriales Superficiales (AES) tuvo una distribución débil frente a Paita (5°S); C. praelongum y C. incisum, indicadores de Aguas Subtropicales Superficiales (ASS) se distribuyeron en dos zonas: la primera entre Paita y Punta Falsa por dentro de las 90 mn, donde se registraron TSM >20 °C. La segunda zona se ubicó entre Huacho y Pucusana por fuera de las 60 mn asociada a TSM >17 °C y concentraciones salinas superiores a 35,300 ups. Protoperidinium obtusum, indicador de Aguas Costera Frías (ACF) se encontró distribuido en forma homogénea dentro de la franja costera entre Punta Falsa y Punta Mendieta, dentro de las 40 mn, relacionado a TSM desde 13,9 hasta 17,7 °C y salinidades típicas de esta masa de agua. DISCUSIÓN La concentración y distribución de la biomasa planctónica (mL/m3) se encontró influenciada, en gran parte, por la variabilidad de los parámetros ambientales, lo cual afecta la oferta alimentaria para el recurso pelágico, en especial para los estadios tempranos. La intensidad de los vientos constituye un factor determinante en esta variabilidad, los mismos que oscilaron entre 1,2 a 12,2 m/s, con vientos moderados al norte de Punta Falsa, vientos débiles entre Chicama y Punta Falsa y vientos entre moderados a intensos al sur de los 8°S (Vásquez 2004). Estas características estarían condicionando la distribución de la comunidad planctónica en especial del fitoplancton. Así tenemos, concentraciones en promedio relativamente más altas localizadas al norte de los 8°S; concentraciones en promedio relativamente más bajas estuvieron localizados al sur de esta latitud en parte debido a la dispersión del fitoplancton. A diferencia de lo reportado por Chang y Delgado (2004), durante el invierno del 2003, cuando la predominancia del fitoplancton en general fue menor. Se conoce, por estudios anteriores, que los máximos volúmenes promedio de plancton se observan principalmente dentro de las 30 mn, tal como ocurrió en los inviernos 1999, 2000, 2003. Esta tendencia fue la misma para el 2004, pero en este año dicho valor promedio fue mayor que el de invierno, a pesar del ingreso de ASS en el norte ocasionando una fuerte zona de mezcla con ACF asociadas a temperaturas de 17 y 18 °C (Vásquez 2004). Entre las especies halladas en la presente evaluación se halló organismos microplanctónicos ampliamente distribuidos, con especies abundantes como algunos Chaetoceros, Thalassiosira y Thalassionema nitzschioides; especies que según Rojas de Mendiola (1966), son componentes de la dieta de la anchoveta en etapas adultas. Con respecto a los indicadores de masas de agua, las evaluaciones en este mismo periodo de los inviernos 2001 (Sánchez et al. 2002), 2002 (Villanueva et al. 2003) y 2003 (Chang et al. 2004), destacan de manera general una tendencia muy similar. Estas características han estado aunadas al desplazamiento de las ASS hacia nuestras costas hasta los 6°S y a una débil intromisión de AES en Paita. Adicionalmente a este desplazamiento de masas de agua se ha determinado zonas de mezcla entre AES y ASS, llegando hasta las 90 mn de la costa, frente a Paita, condiciones que correspondieron a los parámetros físicos. De manera semejante para el indicador de ACF, su distribución a lo largo de la costa, estaría determinando la recuperación del afloramiento, casi de manera normal para este periodo en estudio. 121 CONCLUSIONES • Durante el invierno 2004 se ha incrementado el promedio del volumen de plancton y la predominancia del fitoplancton en comparación con el invierno del 2003. La composición por especies estaría reflejando una buena oferta alimentaria principalmente para los ejemplares adultos. • Como respuesta a las condiciones ambientales inusuales en la región norte, los indicadores biológicos de AES y ASS se distribuyeron hasta aproximadamente los 6°S, indicando aguas de mezcla. Protoperidinium obtusum por su parte, presentó una distribución normal asociada a condiciones típicas de las ACF. Agradecimientos.- Los autores expresan su agradecimiento al personal del Área de Fitoplancton y Producción Primaria por el apoyo brindado en el análisis de las muestras. REFERENCIAS Balech E. 1988. Los dinoflagelados del Atlántico Sudoccidental. Public. Espec. Inst..Español de Oceanog. España. 310 pp. Cupp E. 1943. Marine plankton diatoms of the west coast of North America. Bull. Scripps Inst. Oceangr. 5: 1-237. Chang F, Delgado E. 2004. El fitoplancton de invierno en el crucero de estimación de biomasa de anchoveta (13 agosto – 16 setiembre 2003). Informe interno del Area de Fitoplacnton y Producción Primaria. Delgado E, Chang, F, Villanueva P. 2004. Composición, abundancia y distribución del fitoplancton frente a la costa peruana en el verano 2004. Cr. BIC J. Olaya y SNP-2 0402-03. Informe Interno del Área de Fitoplancton y Producción Primaria. Hasle G, Syvertsen EE. 1996. Marine diatoms. In: Tomas C. (ed.). Identifying.Marine Diatoms and Dinoflagellates. Academic Press, Inc. San Diego. 1-383pp. Hendey I. 1964. An introductory account of the smaller algae of British Coastal waters. Part. V. Ba- Informe IMARPE Vol. 35(2): Abril-Junio 2008 cillariophyceae (Diatoms). Her Majesy’s Stationery Office, London: 317 pp. Hustedt F. 1930. Die Kieselalgen Deutschlands. Osterreichs und der Schweiz mit Berücksichtigung der übrigen Länder Europas sowie der angerzenden Meeresgebiete. En: L. Rabenhorst’s (ed.). Kryptogamen-Flora von Deutschland, Osterreich und der Schweiz. 1 Teil. Akad. Verlag. Leipzig, Reprint Johnson Rep. Goop, New York 1971: 920 pp. GirÓn M, Chang F, AyÓn P. 1996. Larvas de anchoveta (Engraulis ringens) y su relación con el fitoplancton como alimento disponible durante el crucero 9608-09 entre Paita y Tambo de Mora. Informe interno de AEPS-IMARPE. Rojas de Mendiola B. 1966. Relación entre la cosecha estable de fitoplancton, el desove y la alimentación de la anchoveta (Engraulis ringens J.). En : I Seminario Latinoamericano sobre el Oceáno Pacífi- co Oriental, p:60-69 Bol. Extraord. Inst Mar Perú: 33-40. Sánchez S, Villanueva P, Fernández C. 2002. Fitoplancton de invierno durante el Crucero de Estimación de Biomasa Desovante de anchoveta 0108-09. Informe interno del Área de Fitoplancton y Producción Primaria. Schiller J. 1971a. Dinoflagellate (Peridinae) in monographischer Behandlung. 2 Tell. En: L. Rabenhortst (ed). Kriptogamen-Flora von Deutschland, Osterreich und der Schweiz. Reprint by Johnson Repr. Corp. New York, 1971, Vol. 10 Section 3, Part 1: 617 pp. Sournia, A. 1967. Le genre Ceratium (Peridinien Planctonique) dans le Canal de Mozambique. Contribution a une revision mondiale. Vie et Milieu. 18 (2A-A): 375-580 pp. Steidinger, K. y K. Tangen. 1996. Dinoflagellates. In: Tomas C. (ed.). Identifying Marine Diatoms and Dinoflagellates. Academic Press, Inc. San Diego. 387-570pp. 122 SundstrÖm, B. 1986. The Marine diatom genus Rhizosolenia. A new approach to the taxonomy. Lund, Sweden: 196 pp. ThrÖndsen, J. 1978. Preservation and storage. En: A. Sournia (Ed.) Phytoplankton manual. UNESCO, París:69-74. UNESCO. 1981. Programa de plancton para el Pacífico Oriental. Informes de la UNESCO sobre ciencias del mar. Inst. Mar Perú, Callao 11:2526. UtermÖhl, H. 1958. Zur Vervollkomnung der Quantitativen Phytoplankton methodik Mitt. Inter. Ver. Limnol. 9: 1-38. Villanueva, P. y S. Sánchez 2003. Características de la comunidad fitoplanctónica de invierno en el crucero de Estimación de Biomasa da la anchoveta 0208 (en prensa). Vásquez, L. 2004. Informe de la Componente Oceanográfica Física del Crucero de Biomasa Desovante 2004-0809. Informe Interno de DIO. Características oceanográficas físicas en invierno 2004 Vásquez CARACTERÍSTICAS OCEANOGRÁFICAS FÍSICAS EN INVIERNO 2004 PHYSIC OCEANOGRAPHIC CHARACTERISTICS IN WINTER 2004 Luis Vásquez Espinoza1 Vásquez L. 2008. Características oceanográficas físicas en invierno 2004. Inf Inst Mar Perú 35(2): 123128.- Las observaciones se realizaron durante el crucero BIC Olaya y SNP2 0408-09, para evaluación de la biomasa desovante de anchoveta. La distribución térmica indicó predominio de condiciones ambientales casi normales (ATSM +0,5 y -0,5 °C), con zonas cálidas al norte de Paita (+3,0 °C, AES) y frente al Callao (>+0,5 °C, ASS), y una zona fría (ATSM >-1, °C) entre Paita y Pimentel. La salinidad indicó presencia inusual de aguas superficiales subtropicales (ASS) al sur de Pimentel. El ambiente cálido al norte de Paita mostró desplazamientos de aguas ecuatoriales hasta 6°S (>18 °C, >34,8 ups), aproximación de aguas oceánicas hasta 30 mn de la costa y ACF dentro de las 30 mn, ampliando su distribución al sur de Pisco. Los vientos SE y SO variaron en intensidad de 1,2 a 12 m/s, y fueron moderados a intensos al sur de 8°S; de intensidad moderada al norte de punta Falsa, y débiles entre Chicama y punta Falsa. La topografía de la isoterma 15 °C indicó flujos de norte a sur de cabo Blanco a punta Falsa (> 90 m) asociados a la ESCC, los cuales convergieron con los flujos de sur a norte al sur de punta Falsa. Palabras clave: temperatura, vientos, anomalías, invierno austral, mar peruano. Vásquez L. 2008. Physic oceanographic characteristics in winter 2004. Inf Inst Mar Perú 35(2): 123128.- The observations were made during the BIC Olaya and SNP2 0408-09 cruise to evaluate the anchoveta spawning biomass. The predominance of thermal distribution indicated almost normal environmental conditions (temperature surface anomalies +0.5 and -0.5 °C) with warm areas north of Paita (+3.0 °C) with equatorial surface waters (ESW), off Callao (>+0.5 °C), with subtropical surface waters (SSW) and a cold zone (ATSM> -1 °C) from Paita to Pimentel. The salinity indicated the unusual presence of ASS south of Pimentel. The warm north of Paita had equatorial water shifts up to 6 °S (>18 °C, >34,8 psu), approximation of ocean waters up to 30 nm offshore and CCW within 30 nm, extending its distribution in southern Pisco. SE and SW winds varied in intensity from 1.2 to 12 m/s, and were moderate to intense south of 8°S; of moderate intensity north of Punta Falsa, and weak between Chicama and Punta Falsa. The topography of the 15 °C isotherm indicated flows from north to south (>90 m) of Cabo Blanco to Punta Falsa, associated with Southern Extension of Cromwell Current (SECC), and converged south of Punta Falsa with the other flows from south to north. Keywords: temperature, winds, anomalies, Austral winter, Peruvian sea. INTRODUCCIÓN El crucero Evaluación de la Biomasa Desovante de la Anchoveta BIC Olaya y SNP2 0408-09 se ejecutó, como todos los años, con el fin de determinar los niveles de abundancia, distribución de huevos, larvas y estructura poblacional de este recurso, además de conocer las características del ambiente marino y su relación el comportamiento de esta fracción de la población. En el mes de julio 2004, las anomalías de la superficie del mar (ATSM) en el océano Pacífico occidental y central (Niño 1+2) mostraron valores entre -1,1°C y -0,1°C, que fueron similares en agosto, pero en la última semana del mes arribaron ondas cálidas que se propagaron de norte a sur, pegadas a la costa, produciendo un ligero incremento de la anomalía, que pasó de -1,3°C a -1,5°C Los modelos numéricos y estadísticos de la NOAA (numérico acoplado del NCEP, acoplado del sistema de Pronóstico CFS), así como estadísticos de: Correlación Canónica CCA, Analógico Construido CA y Markoviano MKV, junto con el consolidado de todos ellos, pronosticaban condiciones dentro de lo normal a cálidas en el Pacífico central (área Niño 3.4: 120°W – 170°W y 5°S - 5°N) para los próximos tres meses. Resultados similares se obtuvieron con las observaciones del mes de agosto. Bajo este panorama se desarrolló el Crucero 2004/08-09, encontrándose un ambiente cálido en la zona [email protected] 123 costera al norte de Paita, condiciones casi normales al sur de los 5°S, con desplazamiento de aguas ecuatoriales hasta los 6°S y un acercamiento de las aguas oceánicas hasta las 30 mn a la costa. En este informe se da a conocer los resultados más relevantes de las condiciones físicas durante el presente crucero. MATERIAL Y MÉTODOS Entre el 6 agosto y el 8 setiembre 2004, el crucero Olaya y SNP2 0408-09, tuvo una cobertura desde frente a Cabo Blanco hasta Punta Infiernillos, con una amplitud máxima de 200 mn. El estudio de la estructura térmica y halina en las secciones Paita, Informe IMARPE Vol. 35(2): Abril-Junio 2008 84 4 83 82 21 5 6 81 Pta. Sal 80 79 78 77 76 75 74 84 83 82 81 Pta. Sal 80 Cabo Blanco Cabo Blanco Talara Talara Paita Pta. Gobernador Paita Pta. Gobernador Parachique Pta. Falsa Parachique Pta. Falsa Mórrope 9 28 Punta Chao 26 Casma 2 Punta Bermejo Huacho Punta Salinas 18 Supe Chancay Ancón 0 Callao Punta Hermosa Bujama Cerro Azul Punta Cóndor Tambo de Mora Pisco Peninsula Paracas -1 Bahía Independencia Punta Caballas San Nicolás San Juan 80 79 78 77 76 75 82 81 Pta. Sal 80 79 78 77 76 74 75 San Nicolás San Juan -2 84 83 82 4 74 80 79 78 77 76 75 74 Pta. Sal Cabo Blanco Talara 5 Talara Paita Pta. Gobernador Paita Pta. Gobernador VIENTO Parachique Pta. Falsa 6 SSM Parachique Pta. Falsa 6 81 Figura 2.- ATSM (°C). Crucero Biomasa Desovante 0408-09 Cabo Blanco 5 14 Punta Infiernillos Punta Caballas 81 13 Bahía Independencia -1.5 Punta Infiernillos 82 12 Callao Punta Hermosa -0.5 Bujama Cerro Azul Punta Cóndor Tambo de Mora Pisco Peninsula Paracas Figura 1.- TSM (°C). Crucero Biomasa Desovante 0408-09 Mórrope Mórrope Pimentel 7 Pimentel 7 Chérrepe Pacasmayo Chérrepe Pacasmayo Chicama 8 36.0 35.7 35.4 35.1 34.8 34.5 34.2 33.9 33.6 33.3 33.0 32.7 32.4 32.1 31.8 31.5 31.2 30.9 30.6 30.3 30.0 9 10 11 12 13 14 15 84 83 Chicama 8 Trujillo Salaverry Guañape Punta Chao 15 14 13 12 11 10 9 8 7 6 5 4 3 2 1 0 9 Chimbote Casma Punta Lobos 10 Huarmey Punta Bermejo Supe 11 Huacho Punta Salinas Chancay Ancón 12 Callao Punta Hermosa Bujama Cerro Azul Punta Cóndor Tambo de Mora Pisco Peninsula Paracas 13 14 Bahía Independencia 35.0 Punta Infiernillos Punta Caballas 15 San Nicolás San Juan 82 81 80 79 78 77 76 75 84 Figura 3.- Salinidad (ups). Crucero Biomasa Desovante 0408-09 83 Trujillo Salaverry Guañape Punta Chao Chimbote Casma Punta Lobos Huarmey Punta Bermejo Supe Huacho Punta Salinas Chancay Ancón Callao Punta Hermosa Bujama Cerro Azul Punta Cóndor Tambo de Mora Pisco Peninsula Paracas Bahía Independencia Punta Infiernillos Punta Caballas San Nicolás San Juan 82 81 80 79 4 4 Pta. Sal 75 74 Pta. Sal 5 5 TURBULENCIA Parachique Pta. Falsa Paita Pta. Gobernador 6 6 7 7 8 8 1000 9 9 0 10 10 -1000 11 -2000 Mórrope Pimentel Chérrepe Pacasmayo Pimentel Chérrepe Pacasmayo Chicama Chicama 2000 INDICE DE AFLORAMIENTO Parachique Pta. Falsa Mórrope Trujillo Salaverry Guañape Punta Chao 1750 Chimbote Trujillo Salaverry Guañape Punta Chao Chimbote Casma Casma Punta Lobos 1500 Huarmey Punta Lobos Huarmey Punta Bermejo Punta Bermejo 1250 Supe 11 Huacho Punta Salinas Chancay Ancón 1000 750 Bujama Cerro Azul Punta Cóndor Tambo de Mora Pisco Peninsula Paracas 500 13 13 14 14 15 15 Punta Caballas San Nicolás San Juan 79 78 77 76 75 Huacho Punta Salinas Chancay Ancón -3000 Callao Punta Hermosa -4000 Bujama Cerro Azul Punta Cóndor Tambo de Mora Pisco Peninsula Paracas Bahía Independencia -6000 Punta Infiernillos 80 Supe -5000 Bahía Independencia 250 12 12 Callao Punta Hermosa 81 76 Talara Paita Pta. Gobernador 82 77 Cabo Blanco Talara 0 83 78 Figura 4.- Distribución del Viento (m/s). Crucero Biomasa Desovante 0408-09 Cabo Blanco 84 11 Huacho Punta Salinas Chancay Ancón 10 83 10 Huarmey 0.5 Supe 12 84 4 Punta Lobos Punta Bermejo 14 83 9 Chimbote 1 Huarmey 16 84 Punta Chao Casma Punta Lobos 18 15 8 Trujillo Salaverry Guañape 1.5 Chimbote 22 14 7 TSM Chérrepe Pacasmayo Trujillo Salaverry Guañape 24 13 74 4 Chicama 30 12 75 6 TSM 8 20 76 5 Chicama 11 77 Pimentel Chérrepe Pacasmayo 10 78 Mórrope Pimentel 7 79 84 74 Figura 5.- Distribución de la turbulencia (m3/s3) Cr. Biomasa Desovante 0408-09 Punta Infiernillos Punta Caballas -7000 83 82 San Nicolás San Juan 81 80 79 78 77 76 75 74 Figura 6.- Índice de afloramiento. Crucero Biomasa Desovante 0408-09 124 Características oceanográficas físicas en invierno 2004 Vásquez Punta Falsa, Chimbote, Callao y Pisco se realizó sobre la base de lanzamientos de CTD, hasta 500 m de profundidad; y de botellas Niskin hasta 300 m, en el BIC Olaya; y hasta 150 m en el BIC SNP2. Para el estudio de la capa superficial y subsuperficial del mar se obtuvieron datos de temperatura del aire y velocidad y dirección del viento (solo en el BIC Olaya) y muestras de agua para el análisis de salinidad cada 4 a 5 mn (durante las estaciones de plancton). Las muestras de salinidad fueron analizadas por el método de inducción con el Portasal 8410A Guildline (BIC Olaya) y el salinometro Kahlsico RS 10 (BIC SNP-2). Con la información de vientos se calculó Índice de Turbulencia (IT= v3, m3/s3) que se obtiene de la intensidad del viento al cubo y los índices de afloramiento (IA) usando la ecuación: 3 -1 IA = E /cos (a), m *s *km -1 Donde: E : esfuerzo de viento y a : ángulo de la línea de costa con la dirección del viento. RESULTADOS Temperatura La temperatura en la superficie del mar (TSM) presentó rango de 14,0 a 20,4 °C; valores <16 °C, asociados al afloramiento costero, se registraron dentro de las 20 mn entre Mórrope y Callao, y al sur de Pisco ampliaron su distribución más allá de 90 mn. Las temperaturas >20°C se localizaron al norte de Paita relacionadas con aguas ecuatoriales (Figura 1). En general la distribución térmica fue homogénea predominando las isotermas de 16° y 17°C en casi toda el área evaluada, con excepción a la zona norte donde se observó un mayor contraste térmico asociado al desplazamiento norte-sur de las aguas cálidas del norte (Golfo de Guayaquil) hasta los 5°S, en cambio en el extremo sur se observó la zona más fría asociada a un mayor afloramiento. Anomalías térmicas Las anomalías térmicas han presentado valores entre –1,67 °C (130 mn de Paita) y +3,60 °C (en la zona costera frente a Talara). Las condiciones ambientales predominantes fueron casi normales con anomalías entre –0,5 °C a +0,5°C, destacándose dos zonas cálidas: una dentro de las 60 mn al norte de Paita y la otra por fuera de las 60 mn del Callao. También se pudo observar una amplia zona fría entre Chicama y Paita (fuera de las 100 mn) donde las condiciones presentaron anomalías negativas hasta de –1,5 °C (Figura 2). Salinidad En la superficie del mar se registraron concentraciones de sales que variaron de 34,04 a 35,38 ups (Figura 3). Las concentraciones <34,8 ups se hallaron al norte de Punta Gobernador, asociadas a aguas del golfo de Guayaquil desplazadas al sur por la onda Kelvin y el relajamiento de los vientos. Los valores >35,30 ups, asociados a aguas oceánicas, se localizaron más allá de las 60 mn de costa y destacó un núcleo frente al Callao. Las Aguas Ecuatoriales Superficiales (AES) con índices termohalinos >18 °C y <34,8 ups, se encontraron desplazadas más al sur de su ubicación normal y llegaron hasta punta Falsa (4 y 5 setiembre 2004). Las Aguas Subtropicales Superficiales (ASS) con salinidad >35,1 ups y temperaturas >17 °C, estuvieron presentes atípicamente por fuera de las 30 mn entre Pimentel y Punta Infiernillos. Las Aguas Costeras Frías (ACF) asociadas a procesos de afloramiento se encontraron dentro de las 30 mn ampliando su distribución al sur de Pisco. Vientos La intensidad de los vientos varió de 1,2 a 12,2 m/s; predominaron vientos moderados a intensos al sur de 8°S. Entre Chicama y Punta Falsa los vientos fueron débiles y al norte de Punta Falsa, vientos mostraron una intensidad moderada. Con referencia a la dirección del viento se observó 125 una predominancia de vientos del segundo cuadrante (sureste SE) y tercer cuadrante (suroeste SO) (Figura 4). Turbulencia Los índices de menor turbulencia, con valores promedio <500 m3/s3, se registraron al norte de Chicama, lo que estaría favoreciendo la concentración de organismos planctónicos en esta zona. Al sur de esta localidad, el índice de turbulencia fue moderado (500 a 1000 m3/s3) y con algunos núcleos intensos (> 1000 m3/s3) principalmente al sur de Punta Bermejo, produciendo un elevado proceso de mezcla, que a su vez provoca una dispersión de alimento (plancton) en el agua y larvas (Figura 5). Índice de afloramiento El área geográfica en estudio está fuertemente condicionada por la forma de la línea de costa, batimetría, velocidad y dirección de los vientos (posición del anticiclón del Pacífico sur). Los índices de afloramiento indicaron procesos de surgencia permanente que coincidieron con la dirección de los vientos alisios del SE. Los promedios son mayores a -3000 m3s-1km-1, y los índices más bajos se hallan al sur de Pisco y entre Pimentel y Punta Falsa, asociados a vientos débiles. Por otro lado no se observaron hundimientos o apilamiento en las áreas (Figura 6). Profundidad de la isoterma de 15 °C La profundidad de la isoterma de 15 °C varió de 11 a 97 m. Las mayores profundidades (mayores a 90 m) se localizaron entre Talara y Punta Falsa, así como también por fuera de las 100 mn frente al Callao; Por otro lado las profundidades menores a 20 m se ubicaron próximas a la costa frente a Chimbote y Callao. La distribución de las isobatas mostró flujos de norte a sur en la zona costera (dentro de las 60 mn) entre Cabo Blanco y Punta Falsa asociados a la ESCC, en cambio al sur de esta localidad los flujos fueron de sur Informe IMARPE Vol. 35(2): Abril-Junio 2008 4 Chicama, presentó una termoclina débil que desapareció dentro de las 50 mn, asociada a moderados procesos de afloramiento. La isoterma de 15 °C se localizó entre los 20 m (litoral) y 70 m (100 mn) de profundidad. Las concentraciones de sales indicaron ACF dentro de las 40 mn, aguas de mezcla entre las 50 y 100 mn sobre los 50 m de profundidad, interrumpida esta distribución por las ASS entre las 60 a 80 mn (Figura 10). Pta. Sal Cabo Blanco Talara 5 Paita Pta. Gobernador ISOTERMA DE 15C Parachique Pta. Falsa 6 Mórrope Pimentel 7 Chérrepe Pacasmayo Chicama 8 150 140 130 120 110 100 90 80 70 60 50 40 30 20 10 0 9 10 11 12 13 14 15 84 Trujillo Salaverry Guañape Punta Chao Chimbote Casma Punta Lobos Huarmey Punta Bermejo Supe Huacho Punta Salinas Chancay Ancón Callao Punta Hermosa Bujama Cerro Azul Punta Cóndor Tambo de Mora Pisco Peninsula Paracas Bahía Independencia Punta Infiernillos Punta Caballas San Nicolás San Juan 83 82 81 80 79 78 77 76 75 74 Figura 7.- Profundidad de la Isoterma de 15 °C (m). Crucero Biomasa Desovante 0408-09 SECCION PAITA (02-03/09/2004) 0 TEMPERATURA SALINIDAD -100 -60 50 50 100 100 150 150 200 200 250 250 300 300 350 350 400 400 450 450 500 superficie (80 mn) hasta los 50 m en la zona próxima a la costa y las ASS se localizaron sobre los 10 m de profundidad por fuera de las 100 mn, formando un pequeño núcleo en la capa subsuperficial (Figura 8). -20 0 150 100 50 0 500 150 100 50 0 Figura 8.- Distribución vertical de la Temperatura (°C, izq.) y Salinidad (ups, der.) frente a Paita. Cr. 0408-09 a norte; convergiendo los flujos antes mencionados al sur de Punta Falsa (Figura 7). Estructura vertical La secciÓn Paita mostró, dentro de las 80 mn, un contraste térmico asociado a la convergencia de un flujo al sur de aguas cálidas con otro hacia el norte de aguas frías. La isoterma de 15 °C se ubicó entre los 60 m (100 mn) y 90 m de profundidad (zona costera). Las AES se encontraron desde la Frente a punta La Negra, la distribución térmica presentó una termoclina no muy definida por fuera de las 50 mn. La isoterma de 15 °C se localizó entre los 50 m (zona costera) y 75 m (100 mn). Las concentraciones de sales indican un predominio de las ACF en la columna de agua, exceptuando la capa de 0 a 50 m fuera de las 60 mn, donde se presentaron aguas de mezcla (ACF y ASS). Después de 4 a 5 días se volvió a realizar la sección Punta Falsa, y se apreciaron cambios dentro de las 70 mn por el desplazamiento de aguas cálidas del norte; la temperatura aumentó en 1 °C y también se notó la presencia de AES dentro de las 40 mn. Resaltó una gran capa de mezcla (AES, ACF y ASS) sobre 130 mn. En la capa subsuperficial se observó la profundización de las isotermas e isoxígenas indicando una intensificación de los flujos al sur, asociados a la ESCC (Figura 9). La secciÓn vertical frente a 126 En la secciÓn vertical frente a Chimbote, se observó una termoclina muy débil que desapareció dentro de las 30 mn. La isoterma de 15°C se localizó entre los 75 y 10 m de profundidad. En tanto la distribución halina indica la presencia de ASS por fuera de las 30 mn y sobre los 50 m de profundidad; Por otro lado entre las 30 y 10 mn de costa se localizaron aguas de mezcla y en la zona muy costera y por debajo de los 100 m de profundidad se encontraron las ACF (Figura 11). Frente al Callao, se presentó una termoclina, débil que desapareció dentro de las 50 mn, asociada a moderados procesos de afloramiento. La isoterma de 15 °C se localizó entre los 20 (en la zona costera) y 80 m (100 mn). Las concentraciones de sales indicaron que las ACF estuvieron dentro de las 20 mn, aguas de mezcla entre las 20 - 50 mn y las ASS por fuera de las 50 mn en una capa de 70 m de profundidad. Después de 20 días se volvió a repetir la sección Callao encontrándose un cambio en la estructura termohalina, debido al avance de las aguas oceánicas que en agosto llegaron a 50 mn, en setiembre las encontramos hasta las 5 mn de las costa, también el espesor de esta masa de agua pasó de 50 a 80 m de profundidad. En la estructura térmica se notó incremento térmico de 1 °C dentro de las 70 mn, la termoclina aunque débil estuvo mejor definida ubicándose alrededor de los 90 m, con una gran napa (capa de mezcla) sobre ella (Figura 12). La estructura termohalina en la secciÓn Pisco no presentó una termoclina, ubicándose la isoterma de 15 °C entre la superficie (a 50 mn) Características oceanográficas físicas en invierno 2004 Vásquez y los 100 m de profundidad (a 100 mn), en tanto la salinidad mostró una distribución homogénea con valores próximos a 35,0 ups, encontrándose ACF dentro de las 70 mn y aguas de mezcla hacia fuera debido a una ligera influencia de las ASS (Figura 13). A 150 mn de la costa se observó una termoclina débil, más superficial frente a Paita, con profundidad límite inferior cerca de los 60 m (Paita) y 75 m (Punta Falsa). Por debajo de la termoclina la distribución fue homogénea con un gradiente térmico de 1,0 °C/100 m. Las masas de agua presentes fueron las ASS sobre los 10 m de profundidad, aguas de mezcla entre los 10 y 150 m y aguas de la corriente costera por debajo de los 150 m de profundidad. En esta sección fue evidente el efecto de la onda Kelvin, lo atípico fue la gran capa de mezcla presente (Figura 14). SECCION PUNTA FALSA (04-05/09/2004) SECCION PUNTA LA NEGRA ((30-31/08/2004) TEMPERATURA TEMPERATURA SALINIDAD SALINIDAD 0 0 0 0 50 50 50 50 100 100 100 100 150 150 150 150 200 200 200 200 250 250 250 250 300 300 300 300 350 350 350 350 400 400 400 400 450 450 450 450 500 100 50 500 100 0 50 500 0 150 100 50 500 0 35.1 150 SECCION CHICAMA (26-27/08/2004) 0 TEMPERATURA 50 0 SECCION CHIMBOTE (21/08/2004) TEMPERATURA SALINIDAD 0 35.1 SALINIDAD 0 0 50 50 50 100 100 100 100 DISCUSIÓN 150 150 150 150 Las condiciones ambientales frías predominantes en la zona norte del litoral peruano, en los últimos días de agosto cambiaron rápidamente a cálidas dentro de las 60 mn al norte (6°S), asociadas a una onda Kelvin, cuyos efectos se reflejaron en el desplazamiento de aguas cálidas del golfo de Guayaquil, provocando anomalías térmicas positivas de hasta +3,6°C frente a Talara. El nivel del mar mostró incrementos en su nivel medio entre +0,6 y +0,9 cm (Informe Técnico, agosto ENFEN). Los procesos de afloramiento de carácter permanente que ocurrían en la zona norte, se interrumpieron o se redujeron rápidamente. 200 200 200 200 250 250 250 250 300 300 300 300 350 350 350 350 400 400 400 400 450 450 450 450 Las distribución de las masas de agua también cambió entre fines de agosto y comienzos de setiembre. En la Estación Fija Paita el 19 agosto 2004, se encontró una fuerte influencia de ASS hasta la 100 Figura 9.- Distribución Vertical de Temperatura (°C) y Salinidad (ups), frente a Punta La Negra y Punta Falsa. Cr. Biomasa Desovante 0508-09 50 En la capa subsuperficial se notó la intensificación de los flujos hacia el sur, asociados a la ESCC, la cual hasta la tercera semana de agosto se encontraba replegada al norte de los 5°S, pero en la primera semana de setiembre alcanzó los 6°S. 35.0 500 100 75 50 25 0 500 100 75 50 25 0 Figura 10.- Distribución Vertical de Temperatura (°C) y Salinidad (ups), frente a Chicama. Cr. Biomasa Desovante 0408-09 500 100 TEMPERATURA 0 50 50 100 100 150 150 200 200 250 250 300 300 350 350 400 400 450 450 500 500 100 50 0 0 50 25 0 500 TEMPERATURA 100 75 50 25 SALINIDAD 50 100 100 150 150 200 200 250 250 150 100 50 0 300 150 100 50 0 Figura 12.- Distribución vertical de temperatura (°C) y salinidad (ups), frente a Callao. Crucero Biomasa Desovante 0408-09 127 0 0 50 300 75 25 SECCION CALLAO (07-08/09/2004) SALINIDAD 100 50 Figura 11.- Distribución Vertical de Temperatura (°C) y Salinidad (ups), frente a Chimbote. Cr. Biomasa Desovante 0408-09 SECCION CALLAO (14-15/08/2004) 0 75 0 Informe IMARPE Vol. 35(2): Abril-Junio 2008 SECCION PISCO (09-10/08/2004) TEMPERATURA REFERENCIAS SECCION PARALELA A 150 mn DE LA COSTA SALINIDAD 0 0 0 PAITA PUNTA FALSA 17 100 50 50 200 100 100 150 300 400 150 200 0 250 TEMPERATURA 500 200 250 PAITA PUNTA FALSA 35.1 100 300 300 350 200 300 350 400 400 400 SALINIDAD 500 450 450 500 100 75 50 25 0 500 100 75 50 25 0 Figura 13.- Distribución vertical de temperatura (°C) y salinidad (ups), frente a Pisco. Crucero Biomasa Desovante 0408-09 costa; en setiembre su influencia decreció, y esta masa de agua mezclada con AES y ACF, quedó dentro de las 100 mn Por lo antes comentado se puede deducir que el arribo de la onda Kelvin modificó el ambiente marino principalmente en la zona norte, pero estas modificaciones fueron de menor magnitud que las generadas por la onda Kelvin de agosto del 2003, cuando las alteraciones alcanzaron +5 °C de anomalías térmicas y AES se desplazaron hasta Pimentel (7°10’S). En general, las condiciones ambientales encontradas en el presente crucero de estimación de la Figura 14.- Distribución vertical de temperatura (°C) y Salinidad a 150 mn de la costa. Cr. Biomasa Desovante 0508-09 biomasa desovante de la anchoveta, 0408-09, fueron más frías que las registradas en el año anterior 030809. A pesar de haberse detectado una mayor presencia de ASS en el 2004, esto se debió a que las ASS estuvieron asociadas a relativas bajas temperaturas (17 – 18 °C) y a una menor presencia de AES en la zona norte. CONCLUSIONES La distribución térmica indica un predominio de condiciones casi normales, con zonas cálidas al norte de Paita (anomalías > +3°C) y frente al Callao (anomalía > +0,5°C), y otra fría (anomalías > -1,0°C) entre Paita y Pimentel. La distribución halina indica una presencia inusual para la época de ASS al sur de Pimentel. 128 Bakun A. 1973. Costal upwelling indices, west coast of North America; 1967-73. U.S. Dept. NOAA Tech. Rep. NMFS SSRF-671.103 pp. BOLETIN. 2004. El Niño/Southern Oscillation (ENSO), Climate Prediction Center E N S O , augut 5, 2004 ENFEN. 2004. Condiciones Ambientales durante el mes de julio. Informe Técnico-julio ENFEN (Web IMARPE) ENFEN. 2004. Condiciones Ambientales durante el mes de agosto. Informe Técnico-agosto ENFEN (Web IMARPE) IMARPE. 2003. Condiciones Ambientales durante el Crucero de Biomasa desovante 2003-0809. Informe Interno, Instituto del Mar del Perú. IMARPE. 2002. Condiciones Ambientales durante el Crucero de Biomasa desovante 2002-0809. Informe Interno, Instituto del Mar del Perú. Reynolds R. y W. 1982. A monthly average climatology of sea surface temperature. Technical Report NWs31. National Meteorological Center, NOAA, Silver Spring MD. 1-35. Zuta S, Urquizo W. 1970. Temperatura promedio de la superficie del mar frente a la costa peruana. Periodo 1928-1969. Bol. Inst. Mar Perú 2(8): 459-520. Zuta S. 1988. Variations of the mass field and currents off the Peru Coast. IOC (UNESCO). Time Series of Ocean Measurements, 33. Vol. 4: 55-75. Evaluación hidroacústica de especies pelágicas en invierno 2004 Gutiérrez y Peraltilla EVALUACIÓN ACÚSTICA DE LA DISTRIBUCIÓN Y ABUNDANCIA DE ESPECIES PELÁGICAS Y MESOPELÁGICAS EN INVIERNO 2004 ACOUSTIC ASSESSMENT OF DISTRIBUTION AND ABUNDANCE OF PELAGIC AND MESOPELAGIC SPECIES DURING WINTER 2004 Mariano Gutiérrez Salvador Peraltilla RESUMEN Gutiérrez M, Peraltilla S. 2008.- Evaluación acústica de la distribución y abundancia de especies pelágicas y mesopelágicas en invierno 2004. Inf. Inst. Mar Perú 35(2):129-140.-.El crucero BIC Olaya y SNP2. 0408-09 se realizó del 8 agosto al 5 septiembre 2004, entre Punta Infiernillos (15°S) y Cabo Blanco (4°40´S), desde 1 a 100 mn de la costa, efectuando 49 transectos. Para obtener información oceanográfica se efectuaron perfiles hidrográficos de 200 mn frente a Callao y Paita. La anchoveta, Engraulis ringens, presentó amplia distribución y abundancia desde Talara hasta Punta Infiernillos, con cierta discontinuidad frente al área entre Chimbote y Cerro Azul, donde los mayores núcleos se ubicaron por fuera de las 25 mn. La múnida, Pleuroncodes monodon, mostró una amplia distribución costera, desplazó a la anchoveta en ciertas zonas, donde alcanzó sus mayores índices de abundancia. Las especies mesopelágicas evaluadas fueron: vinciguerria, Vinciguerria lucetia, que mostró una abundancia inusual para la estación como producto de la marcada influencia de aguas subtropicales superficiales (ASS), especialmente en el sur; y calamar gigante, Dosidicus gigas, distribuido en dos amplias zonas, en el norte y en el sur. Palabras clave: recursos pelágicos, evaluación hidroacústica, distribución, abundancia, mar peruano. ABSTRACT Gutiérrez M, Peraltilla S. 2008.- Acoustic assessment of the distribution and abundance of pelagic and mesopelagic species during winter 2004. Inf. Inst. Mar Perú 35(2):129-140.- The cruise BIC Olaya and SNP2 0408-09 was held on 8 August to 5 September 2004, between Punta Infiernillos (15°S) and Cabo Blanco (4°40’S), from 1 to 100 nautical miles from the coast, making 49 transects. For oceanographic information, hydrographic profiles were made of 200 nm off Callao and Paita. The anchoveta, Engraulis ringens, showed extensive distribution and abundance from Talara to Punta Infiernillos, with a discontinuity compared to the area between Chimbote and Cerro Azul, where the largest nuclei were located outside the 25 nm. The munida, Pleuroncodes monodon, showed large coastal distribution shifted to the anchovy in certain areas, where it reached its greatest abundance indices. Mesopelagic species evaluated were: vinciguerria, Vinciguerria lucetia, which had unusual abundance for the season as a result of the strong influence of subtropical surface waters (SSW), especially in the south; and giant squid, Dosidicus gigas, spread at two broad areas, north and south. Keywords: pelagic resources, hydroacoustic assessment, distribution, abundance, Peruvian sea. INTRODUCCIÓN El Programa de Seguimiento de Pesquerías Pelágicas comprende la ejecución de una serie de actividades orientadas a conocer las variaciones estacionales en los parámetros poblacionales de las especies que son objeto de pesca intensiva, como en el caso de la anchoveta. El Crucero 0408-09 para evaluación de la biomasa desovante de la anchoveta, por el método de producción de huevos (MPH), fue planificado para estudiar, con énfasis, el proceso reproductivo invernal de anchoveta, las variables referidas a procesos biológicos tales como la alimentación, crecimiento, desarrollo larval, distribución y abundancia, tanto de la anchoveta como de otras especies que, ya sea por su abundancia o importancia ecológica o pesquera, hayan podido encontrarse accesibles durante el período de evaluación. De este modo ha sido posible estimar, desde el enfoque de la investigación hidroacústica, tanto la distribución como la abundancia de dos especies pelágicas: anchoveta, Engraulis ringens, y múnida, Pleuroncodes monodon; y de dos especies mesopelágicas: vinciguerria, Vinciguerria lucetia, y calamar gigante o pota, Dosidicus gigas. El presente informe presenta los principales resultados de dicho estudio. MATERIAL Y MÉTODOS Itinerario El plan del Crucero BIC Olaya y SNP2 0408-09 de Evaluación de 129 Biomasa Desovante se llevó a cabo del 8 agosto al 5 de septiembre del 2004. El BIC SNP-2 evaluó el área costera en transectos de 1 a 30 mn de extensión entre Punta Infiernillos y Lobitos; el Olaya prospectó trayectos desde 30 hasta 200 mn de la costa, entre Punta Infiernillos y Paita. Equipamiento acÚstico El mismo tipo de ecosonda científica (Simrad EK500 V5.30) fue utilizado en ambos barcos para la detección y ecointegración operando a las frecuencias de 38 y 120 kHz. La cobertura del muestreo vertical a 120 kHz fue de 0 a 150 m; y para 38 kHz, de 0 a 500 m. La colección de los datos Sv de las ecosondas se efectuó a Informe IMARPE Vol. 35(2): Abril-Junio 2008 través de redes LAN conectadas a computadores personales empleando un programa de registro digital (Echolog V2.5). Tabla 1.- Parámetros y resultados de las calibraciones acústicas de las ecosondas científicas de los BIC Olaya y SNP-2 antes de la ejecución del Crucero 0408-09 BIC SNP-2 38 kHz 120 kHz 0.0100 0.0380 1508 1508 2000 1000 -20.90 -20.50 -33.60 -40.40 25.53 22.86 26.59 23.21 14.20 17.50 0.0397 0.0126 1.0000 1.0000 3.8 1.2 7.0 7.2 W, Q, E, TSW W, Q, E, TSW 72 30 10 3 0.99 0.97 Transductores Frecuencias Coeęciente de absorcion (dB/m) Velicidad del sonido (m/s) Potencia de trnasmisión (W) Angulo equivalente (dB re 1 esteradian) TS medido (dB) Ganancia Sv (dB) Ganancia TS (dB) Profundidad de la esfera (m) Longitud de onda (m) Longitud de pulso (ms) Ancho de banda (kHz) Angulo nominal (grados sexagesimales) Ecogramas registrados Tamaño de muestra ecograma Q (cm) Tamaño de muestra ecograma W (cm) Constante NASC(teorico)/NASC(leido) Calibraciones Las calibraciones acústicas de las ecosondas se realizaron con anterioridad al inicio del crucero (Tabla 1). La intercalibración de las ecosondas y transductores de ambos barcos se efectuó durante la prospección (Figura 1), donde se presenta la compatibilidad entre los valores NASC o SA medidos (Fernández y MacLennan 2000). Las calibraciones e intercalibraciones se efectuaron de acuerdo a Foote et al. (1987) y MacLennan y Simmonds (1992). Grilla de crucero La grilla de muestreo comprendió transectos paralelos, separados 15 mn entre sí y perpendiculares a línea de la costa; el intervalo de las unidades básicas de muestreo acústico (UBM) fue de 1 mn. En total se muestrearon 5611 UBM en 49 perfiles; 3622 UBM correspondieron al BIC Olaya y 2035 al SNP2. Se efectuaron 91 lances de comprobación de ecotrazos y para muestreo biológico. Para análisis de los datos acústicos se utilizó un programa de postprocesamiento (Echoview 2.51.1), cuyo módulo virtual facilita la Intercalibración Acústica 120 kHz BIC Olaya - BIC SNP-2 Intercalibración Acústica 38 kHz BIC Olaya - BIC SNP-2 1200 Sa Olaya 1800 1000 NASC (m2/mn2) 200 73 70 67 64 61 58 55 52 49 46 43 40 37 34 31 28 25 22 0 1 61 57 53 49 45 41 37 33 29 25 21 17 9 13 5 1 0 400 73 Sa SNP2 69 Sa Olaya 200 65 400 19 600 600 16 800 13 1000 7 1200 Sa SNP2 800 10 1400 4 1600 NASC (m2/mn2) extracción de la información correspondiente a los cardúmenes o enjambres de las especies en estudio (Higgimbottom et al. 2000). La discriminación de ecotrazos (cardúmenes o enjambres) se efectuó de acuerdo a registros típicos o en función a los porcentajes de las capturas realizadas por especies. Análisis de datos 2000 Intervalos navegados Intervalos navegados Correlacion 120 kHz Sa BIC Olaya - BIC SNP2 Correlacion 38 kHz Sa BIC Olaya - BIC SNP2 1800 1200 1600 1000 1400 1200 Sa BIC SNP Sa BIC SNP BIC Olaya 38 kHz 120 kHz 0.0100 0.0380 1508 1508 2000 1000 -20.50 -20.80 -33.60 -40.40 25.36 25.13 25.78 24.91 19.41 22.10 0.0397 0.0126 1.0000 0.3000 3.8 12.0 6.9 7.2 W, Q, E, TSW W, Q, E, TSW 72 30 10 3 0.99 0.99 1000 800 600 400 y = 1.0959x - 1.8258 R2 = 0.9851 200 800 600 400 y = 1.0132x - 9.8887 R2 = 0.9806 200 0 0 0 200 400 600 800 1000 1200 1400 1600 1800 0 200 400 600 800 1000 1200 Sa BIC Olaya Sa BIC Olaya Figura 1.- Resultados de las intercalibraciones de las ecosondas cientificasEK500 llevadas a cabo el día 22 de Agosto entre los BIC Olaya y SNP-2. Las dos figuras a la izquierda muestra la intercalibración a 38 kHz, y las de la derecha presentan la de 120 kHz. En ambos casos los resultados son coherentes, y los valores colectados por los dos barcos, compatibles. 130 Evaluación hidroacústica de especies pelágicas en invierno 2004 Gutiérrez y Peraltilla La carta de trayectos y lances ejecutados se aprecia en la Figura 2. Para delimitar las áreas de distribución y concentración se utilizó un programa de interpolación de datos (Surfer v8.0) por el método de krigeo (Gutiérrez 1997). El análisis de las variables acústicas colectadas se efectuó a través de un programa de estadística multivariada (S-Plus v6.0). La estimación de límites de confianza se realizó de acuerdo con Bazigos (1976). EstimaciÓn de la abundancia La estimación de abundancia de anchoveta fue realizada a través de dos métodos: el método convencional de estratificación por áreas isoparalitorales (Gutiérrez y Peraltilla 2000); y el método de estratificación por transectos (Jolly y Hampton 1990; Hewitt et al. 2003). Este último método presenta la ventaja de permitir la incorporación al análisis de mayor cantidad de información biométrica, a fin de obtener estimados de abundancia para cada UBM, diferenciando por tallas los valores NASC colectados. En primer término, se calculan las proporciones (p) por lance (i) y clase (j) de talla (L) de los lances efectuados durante la prospección, considerando la talla, el número de individuos (n) y el peso individual (w) como factores de ponderación. En el caso de las tallas es conveniente efectuar la ponderación según el cuadrado de las tallas, ya que la magnitud de los ecos están aproximadamente relacionados con el cuadrado del tamaño del pez (Foote 1987, MacLennan 1998): pij = L2j .wij .nij ∑ L .w .n 2 j ij Figura 2.- Carta de trayectos y lances efectuados por los BIC Olaya y SNP-2. C F Paralelamente se debe determinar el Factor de Conversión (CF), que es definido como el cociente del peso medio del pez (w) y su correspondiente área de sección transversal de retrodispersión (σ). Ambos valores se obtienen considerando toda la distribución de especímenes colectados durante los muestreos con red de arrastre: 4.π .10 T S ∑ CF j .10 / 10 j 6 wij = ai Lj bi ; (g) ; (t/m2) =1 , donde: σ j = 4.π .10 T S j / 10 ; (m2) La Fuerza de Blanco (TS) en la clase j es (Foote 1987): T S ij = ij wij j = A.Log ( L j ) − B20 ; (dB) Dos de los factores de las ecuaciones anteriores (CFij y w(Lij)) se calculan para cada lance i y clase de talla j. A y B son factores que dependen de la reflectividad de la especie en estudio. En el caso del peso individual wij (gramos), éste puede obtenerse directamente de la determinación del peso de la muestra de cada clase j, o bien a través de la relación talla peso, donde a y b son constantes: 131 La densidad en el transecto k, UBM l (ρkl) , lance i y clase j es calculada obteniendo previamente un factor de corrección K para cada lance i y clase j: K ij = C F ij pij ; (t/m2) ρ klij = NASC kl .K ij ; (t/mn2) Para poder aplicar la ecuación anterior se debe decidir cuál lance servirá para estimar la abundancia en cada transecto; los transectos, formados por un número discreto de UBMs, pueden subdividirse en caso exista más de un lance sobre un mismo transecto. La biomasa B en el transecto k, UBM l, es en realidad la suma de las abundancias por talla, dependiendo de la separación E entre transectos: Informe IMARPE Vol. 35(2): Abril-Junio 2008 84°W 82°W 80°W 78°W 76°W 74°W 4°S 4°S Pto. Pizarro Zorritos Pta. Sal Cabo Blanco Talara 6°S 35.0 ups 35.1 ups 35.2 ups Parachique Pta. La Negra 6°S Vinciguerria: TSj = 4.16 Log (Lj) – 81.056 ; (dB) LEYENDA: Paita Pta. Gobernador de las especies mesopelágicas fueron las siguientes (tomadas de Gutiérrez, 2002): Mórrope Pota: TSj = 20.0 Log (Lj) – 87.200 ; (dB) Pimentel RESULTADOS 8°S 8°S Chérrepe Pacasmayo Chicama Trujillo Salaverry Punta Chao Chimbote Casma 10°S 10°S Punta Lobos Huarmey Punta Bermejo Supe 1000 Huacho 250 100 14°S 50 10 5 0.1 84°W 12°S Chancay Callao Pucusana Cerro Azul Tambo de Mora Pisco Bahía Independencia Punta Infiernillos Area cubierta por la distribución de anchoveta: 35, 612 mn2 82°W 80°W 14°S 500 Valores ecointegrados 12°S 750 Punta Caballas San Juan N klij = E.ρ klij wij Fuerza de Blanco ₍TS₎ Las ecuaciones de TS utilizadas para la frecuencia de 120 kHz, que La anchoveta tuvo amplia distribución dentro del área prospectada. Aunque la prospección se llevó a cabo en la estación invernal, cuando la que dispersión de la especie suele ser mayor, se detectaron dos grandes núcleos de concentración: uno en el norte (de Huacho a Punta La Negra) y otro en el sur (Callao a Punta Caballas). Los núcleos de alta densidad detectados tuvieron menor magnitud, y la distribución tendió a ser más dispersa. Anchoveta: TSj = 30.258 Log (Lj) - 89.353 ; (dB) Múnida: TSj = 40.700 Log (Lj) – 114.42 ; (dB) La agregación de anchoveta en grandes núcleos, se dio bajo cuatro condicionantes: Las ecuaciones de fuerza del blanco (TS) utilizadas para la frecuencia de 38 kHz, empleadas en la estimación de la abundancia (1) existencia de núcleos costeros de alta densidad donde la salinidad fue <35,0 ups (Aguas Costeras Frías (ACF); 76°W 74°W 2 Figura 3.- Distribución de anchoveta según valores ecointegrados NASC (m /mn ) y su relación con la salinidad (masas de aguas). Para determinar el número de individuos en el transecto k, UBM l, lance i y clase j se debe utilizar la siguiente relación: Anchoveta (Engaulis ringens) En la Figura 3 se representa la distribución de anchoveta de acuerdo con los valores ecointegrados (NASC) y en función a las masas de aguas (salinidad). Las isohalinas de la figura aparecen paralelas a la línea de costa, de allí que la estructura física de las masas de agua haya condicionado la existencia de un patrón de distribución bastante claro. 78°W 2 Bkl = E.∑ ρ klij ; (t) DistribuciÓn y abundancia relativa de los recursos evaluados fue empleada para la estimación de abundancia de las especies pelágicas, fueron las siguientes (tomadas de Gutiérrez, 2002) : Figura 4.- Ecograma de anchoveta registrado a bordo del SNP-2 en las inmediaciones de Sechura. La distribución de anchoveta en grados y continuas agregaciones es usual durante las estaciones de primavera y verano. La ocurrencia de ondas Kelvin en julio y agosto podría haber influenciado en la formación de éstas agregaciones. 132 Evaluación hidroacústica de especies pelágicas en invierno 2004 Gutiérrez y Peraltilla Figura 5.- Resultados del análisis exploratorio GAM sobre la base de datos de cardúmenes de anchoveta. En el presente crucero es la clarra la existencia de tendencias latitudinales y horarias en el nivel de concentración y distribución vertical. (2) núcleos ubicados entre 20 y 40 mn de la costa en aguas de mezcla entre ACF y Aguas Subtropicales Superficiales (ASS); (3) grandes núcleos ubicados sobre el límite de las ASS, con salinidades de 35,1 a 35,2 ups; y (4) pequeños y dispersos núcleos ubicados en salinidades >35,2 ups. Estos últimos, sin embargo, no fueron explorados sino cuando la mayor extensión de los perfiles hidrográficos de Callao, Chicama y Punta Falsa permitió ampliar el estudio acústico más allá de las 100 mn inicialmente programadas. Por ello, la distribución y abundancia de anchoveta se encuentra, por lo menos, ligeramente subestimada. En la Figura 4 se presenta un ecograma con importantes ecorregistros de anchoveta, en forma de cordones, el cual ha sido obtenido en las cercanías de Sechura. Esta clase de distribución, que fue frecuente durante el crucero, es típica en primavera y verano, pero no en invierno. El arribo de por lo menos dos ondas Kelvin a las costas peruanas, entre julio y agosto habría propiciado este tipo de agregación que probablemente aumentará la disponibilidad y capturabilidad de anchoveta durante la temporada de pesca que se iniciará a comienzos de octubre de 2004. Verticalmente, la anchoveta se presentó generalmente distribuida desde la superficie hasta 40 m de profundidad. Los cardúmenes las cuatro especies en evaluación, fueron detectados a fin de crear una base de datos, con las cuales analizar ciertos parámetros de su estructura espacial durante el crucero. Para ello se recurrió a análisis GAM (Modelos Aditivos Generalizados) a fin de explorar las tendencias espacio-temporales y geomé133 tricas que permitan caracterizar la distribución y abundancia relativa. La Figura 5 presenta algunos de los principales resultados del uso de la estadística multivariada GAM en el caso de anchoveta: - La sección (a) muestra que el recurso se distribuyó en aguas con temperatura superficial entre 14 y 20 °C, con la mayor proporción estuvo entre 16 y 18 °C; - El panel (b) presenta el rango de salinidad superficial, donde se muestra que la especie de distribuyó entre 34,4 y 35,4 ups, con las mayores concentraciones entre 34,9 y 35,1. Debe hacerse notar que los extremos de la distribución (donde la salinidad fue <34,8 o >35,2 ups) corresponde a una distribución subsuperficial de anchoveta; - La sección (c) muestra que la especie se distribuyó hasta 200 mn mar afuera, pero que la Informe IMARPE Vol. 35(2): Abril-Junio 2008 Figura 6.- Variación de la magnitud de las áreas (a) y biomasas (b) latitudinales en la distribución de anchoveta (mn2) entre 2002 y 2004. mayor proporción lo hizo sólo hasta 50 m; a disminuir en horas nocturnas; - En (e) se aprecia un efecto inverso para la extensión de los cardúmenes, que tienden a alcanzar mayor longitud - El panel (d) muestra que la altura de los cardúmenes fue siempre mayor hacia las 10:00 h, y tendió 80°W Pto. Pizarro Zorritos 78°W 76°W 74°W 4°S 82°W 4°S 84°W Pta. Sal Cabo Blanco Talara 47 Paita 46 Pta. Gobernador Pimentel 41 40 39 38 Chérrepe Pacasmayo 37 8°S Chicama 36 35 Trujillo Salaverry 34 33 32 31 30 29 1 3 5 7 9 11 Punta Chao Chimbote 28 27 26 10°S a a a a a a 6°S 0 1 3 5 7 9 Mórrope 8°S 42 Casma Punta Lobos 25 24 23 10°S 6°S Leyenda : Parachique Pta. La Negra 43 Huarmey 22 Punta Bermejo 21 Supe 12°S 20 19 Huacho 18 17 16 Chancay 12°S 45 44 Callao 15 Pucusana 12 11 9 13 Tambo de Mora Pisco 8 14°S 7 Bahía Independencia 6 Punta Infiernillos 5 4 3 84°W 82°W 80°W - En (f) se aprecian las tendencias en la distribución vertical, donde es claro que la especie se profundiza moderadamente en horas de luz, para ascender a la superficie en horas nocturnas. La misma Figura 5 presenta otros seis paneles (g - l) donde se presentan otras relaciones geométricas conectados a factores espacio-temporales. En casi todos los casos la deviancia (95% de confianza) tiende a ser mayor en comparación con las primeros seis secciones (a - f). En este caso resaltan la asimetría (“skewness”), rugosidad y área transversal de los cardúmenes en relación con la hora del día, ya que en los tres casos son parámetros que tienden a disminuir a lo largo del día para luego incrementarse durante la noche. Por último, es de destacar que el área transversal de los cardúmenes tendió a ser mayor en el sur, a pesar de que la mayor abundancia de anchoveta se dio en el norte (sección k). Cerro Azul 10 14°S 14 en horas sin luz, es decir, los cardúmenes se contraen en horas de luz, sacrificando longitud a cambio de mayor altura; 78°W Punta Caballas 2 1 San Juan 76°W 74°W Figura 7.- Distribución de anchoveta según el número de individuos por UBMs y transectos. Los números que aparece a la izquierda de cada transecto indican la codificación correlativa (sur a norte) de los transectos. 134 El área de distribución de anchoveta ha sufrido modestas variaciones durante los dos últimos años. Existe una relativa estabilidad en la magnitud de las áreas de distribución entre 6 y 18°S; pero en las latitudes de 3 a 5°S la variación en las reducidas áreas cubiertas por anchoveta sufren amplias variaciones estacionales como producto Evaluación hidroacústica de especies pelágicas en invierno 2004 Gutiérrez y Peraltilla 3.E+10 1,400,000 7.00E+10 1,200,000 6.00E+10 240,000 1,000,000 4.00E+10 600,000 3.00E+10 2.E+10 200,000 Biomasa (t) 800,000 Biomasa (Ton) 5.00E+10 2.E+10 160,000 120,000 1.E+10 400,000 2.00E+10 80,000 Biomasa Número de individuos 200,000 1.00E+10 Numpeces Biomasa 40,000 197 190 183 176 169 162 155 134 148 127 141 120 113 99 92 106 85 78 71 64 57 50 43 36 29 8 22 0.E+00 1 .0 11 .5 12 .0 12 .5 13 .0 13 .5 14 .0 14 .5 15 .0 15 .5 16 .0 16 .5 17 .0 17 .5 18 .0 11 5 .0 .5 0 9. 10 5 9. 10 5 0 0 5 7. 8. 0 6. 8. 5 6. 7. 0 5. 0 Talla (cm) Distancia a la costa (mn) Figura 8.- Abundancia de anchoveta por tallas. 1,500,000 Figura 9.- Abundancia de anchoveta según distancia a la costa -4 1.E+11 Biomasa Numpeces -5 1.E+11 1,000,000 8.E+10 750,000 6.E+10 500,000 4.E+10 250,000 2.E+10 0 0.E+00 -6 Porcentaje latitudinal de biomasa 1,250,000 Numpeces Biomasa (t) 5.E+09 0 5. 0.00E+00 15 Numpeces 280,000 Número de individuos 8.00E+10 -7 -8 -9 -10 -11 -12 -13 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 -14 18 Latitud 8 Figura 10.- Abundancia latitudinal de anchoveta de la proyección o retracción de las Aguas Ecuatoriales Superficiales (AES) que condicionan la presencia de la especie en la zona, como se muestra en la Figura 6, sección (a). En la sección (b) de la Figura 6 se aprecia el resultado de similar tipo de análisis, que muestra que la variabilidad de la abundancia de anchoveta es menor en las latitudes de la costa central (8 a 12°S), y los cambios en la abundancia han sido mayores hacia los dos extremos de la distribución (norte y sur). La estimación de la abundancia de anchoveta se realizó utilizando dos métodos de estratificación: por transectos y por áreas isoparalitorales. Ambos alcanzaron resultados bastante similares. El método por transectos arrojó una abundancia de 7,51x106 t (34,4% de confianza), y el de áreas isoparalitorales, 7.49x106 t (12,85% de confianza). La amplia diferencia en la estimación de los límites de confianza se debe a que mientras el método de isoparalitorales se limita a las zonas donde hay presencia de anchoveta (Tabla 2), el de transectos en cambio se proyecta a la totalidad del área evaluada (Tabla 2). 8.5 9 9.5 10 10.5 11 11.5 -15 12 12.5 13 13.5 14 14.5 15 15.5 16 16.5 17 17.5 18 Figura 11.-Abundancia latitudinal de anchoveta según tallas. La Figura 7 presenta el total de individuos por UBM y transectos. El método de estratificación por transectos, además de medir la abundancia, permite ponderar las estructuras de tallas obtenidas en los lances a los valores ecointegrados (NASC) por UBM, generando estimados por número de peces por tallas para utilización directa en modelos de dinámica poblacional. La abundancia de individuos adultos (>12 cm) se estimó en 6,62x106 t (88,11% del total), y los juveniles en 0,89x106 t (11,89% del total). En término de individuos, la cantidad de adultos fue estimada en 80,31% de un total de 4,63x1011 peces, y los juveniles se estimaron en 19,69% (Figura 8). Según distancia a la costa la mayor fracción de biomasa se distribuyó dentro de la franja comprendida en las primeras 70 mn de la costa (Figura 9). La mayor abundancia latitudinal se registró en los grados 7, 9, 13 y 5°S (Figura 10). De acuerdo a la estructura de la biomasa por tallas, la mayor cantidad de adultos correspondió a tallas >13 cm, entre 8 y 15°S. La 135 fracción más importante de los juveniles estuvo entre 6 y 11°S; fue prácticamente nula la presencia de juveniles en los 5 y 15 (Figura 11). Múnida, Pleuroncodes monodon La múnida tuvo una amplia distribución latitudinal, y se halló en gran parte de la zona estudiada (Punta Infiernillos –14°30´S- a Punta La Negra -6°S-). Por lo general la múnida se distribuyó en aguas con valores halinos <35,1 ups en superficie (Figura 12). En su distribución destacan dos grandes zonas: (1) la primera ubicada entre Punta Infiernillos y Huacho, donde la presencia de la especie no se extiende más allá de las 30 mn, y donde la presencia de núcleos de alta densidad es discontinua; y (2) la zona entre Huacho y Punta La Negra, donde las zonas de alta densidad acústica son prácticamente continuas. En esta última zona las zonas de afloramiento ubicadas entre Chimbote y Mórrope Informe IMARPE Vol. 35(2): Abril-Junio 2008 84°W 82°W 80°W 78°W 76°W 74°W 4°S 4°S Pto. Pizarro Zorritos Pta. Sal Cabo Blanco Talara LEYENDA: Paita Pta. Gobernador 35.1 ups 6°S 6°S Parachique Pta. La Negra Mórrope Pimentel 8°S 8°S Chérrepe Pacasmayo Chicama Trujillo Salaverry Punta Chao Chimbote Casma 10°S 10°S Punta Lobos Huarmey Punta Bermejo Supe Huacho 250 100 14°S 50 10 12°S 500 Chancay Callao Pucusana Cerro Azul Tambo de Mora Pisco Bahía Independencia 14°S 750 Valores ecointegrados 12°S 1000 Punta Infiernillos 5 Punta Caballas 1 San Juan 84°W 82°W 80°W 78°W 76°W 74°W 2 Figura 12.- Distribución de múnida según valores ecointegrados NASC (m /mn2) y su relación con la salinidad (masas de aguas). han alejado la distribución de múnida por fuera de las 15 mn en promedio, aunque alcanzan áreas hasta 65 mn de la costa. Los datos acústicos colectados permitieron crear una base de datos sobre enjambres de múnida, la cual ha sido analizada recurriendo a los modelos exploratorios GAM. En la Figura 13 puede observarse que: - los paneles a y b presentan la relación de la presencia subsuperficial de múnida respecto a parámetros físicos superficiales como temperatura y salinidad; - la sección c muestra que la múnida presentó una ligera tendencia declinante en tanto la distancia a la costa aumenta; - los paneles d y e muestran que la altura y la extensión de los enjambres presentan tendencias similares, es decir que ambas dimensiones se incrementan en horas diurnas y disminuyen durante la noche; - el panel f presenta los desplazamientos verticales de Figura 13.- Resultados del análisis exploratorio GAM sobre la base de datos de emjambres de múnida 136 Evaluación hidroacústica de especies pelágicas en invierno 2004 Gutiérrez y Peraltilla Tabla 2.- Biomasa (toneladas) por grado de latitud de las especies evaluadas. Crucero 0408-09 Grado de latitud sur 03° 30 - 04° 00 03°30' - 04° 04° 00 - 04° 30 04° 30 - 05° 00 04° - 05° 05° 00 - 05° 30 05° 30 - 06° 00 05° - 06° 06° 00 - 06° 30 06° 30 - 07° 00 06° - 07° 07° 00 - 07° 30 07° 30 - 08° 00 07° - 08° 08° 00 - 08° 30 08° 30 - 09° 00 08° - 09° 09° 00 - 09° 30 09° 30 - 10° 00 09° - 10° 10° 00 - 10° 30 10° 30 - 11° 00 10° - 11° 11° 00 - 11° 30 11° 30 - 12° 00 11° - 12° 12° 00 - 12° 30 12° 30 - 13° 00 12° - 13° 13° 00 - 13° 30 13° 30 - 14° 00 13° -14° 14° 00 - 14° 30 14° 30 - 15° 00 14° -15° 15° 00 - 15° 30 15° 30 - 16° 00 15° -16° 16° 00 - 16° 30 16° 30 - 17° 00 16° -17° 17° 00 - 17° 30 17° 30 - 18° 00 17° -18° 18° 00 - 18° 30 18° -18°30' TOTALES Anchoveta (t) Jurel (t) Caballa (t) Samasa (t) Bagre (t) 84,..786 84,786 109,571 532,635 642,206 246,924 383,172 630,096 529,526 947,031 1,476,557 508,707 385,754 894,461 1,070,504 432,916 1,503,420 257,194 218,138 475,332 238,211 158,473 396,684 103,113 266,240 369,353 460,399 198,412 658,811 140,208 189,665 329,873 28,474 Munida (t) 3,394 50,270 53,664 101,277 64,029 165,306 153,446 183,863 337,309 69,959 60,658 130,617 53,055 21,553 74,609 73,358 16,904 90,262 54,268 103,863 158,131 66,880 42,328 109,208 42,715 13,454 56,169 Pota (t) 811 118 929 0 7,052 7,052 3,302 8,498 11,800 11,887 38,961 50,848 33,133 27,595 60,728 4,706 969 5,675 493 287 780 4,397 4,206 8,603 5,989 11,337 17,327 35,943 12,649 48,591 27,838 34,874 62,713 22,942 22,942 28,474 7,490,053 Vinciguerria (t) 49,357 536,850 586,207 157,942 218,868 376,811 3,701 17,190 20,892 5,422 407,004 412,425 336,044 470,806 806,850 243,586 125,830 369,416 131,132 595,419 726,551 353,345 497,315 850,661 465,673 952,830 1,418,503 1,789,513 1,274,196 3,063,709 529,494 385,824 915,318 876,768 876,768 0 múnida, que se profundiza durante el día y asciende durante la noche en rangos que pueden alcanzar hasta 60 u 80 m. 0 0 0 1,175,275 275,047 10,424,111 - los paneles g, h, i corresponden a dimensiones geométricas de los enjambres respecto a las horas del día, donde el coeficiente de asimetría se incrementa hasta estabilizarse en horas del día, para luego caer en horas nocturnas, la rugosidad de los enjambres aumenta notoriamente hasta las 15:00 h para luego declinar; el área de sección transversal de los enjambres se mantiene bastante uniforme durante el ciclo diurno-nocturno. el incremento de la rugosidad de los enjambres en 12°S, y la existencia de dos zonas (los grados 8 y 13°S) donde los enjambres alcanzaron sus máximas áreas de sección transversal coincidiendo con las zonas más frías de la región prospectada, lo que demostraría la preferencia de múnida por las bajas temperaturas. ción por áreas isoparalitorales, en 1,7x106 t, con límites estadísticos de confianza en 11,88%. Latitudinalmente las mayores abundancias se presentaron en los grados 7 y 8°S en el norte, y 12 y 13°S en el sur (Tabla 2). - el mismo parámetro (área transversal de los enjambres) se muestra en la sección l, donde se aprecia que estos valores disminuyen con la distancia a la costa, debido posiblemente a las mayores temperaturas existentes mar afuera. El calamar gigante se registró distribuido en dos grandes zonas por fuera de las 30 mn en promedio, con salinidad superficial >35,1 ups, y sobre zonas con profundidades >100 brazas (Figura 14). - Las secciones j y k de la Figura 13 presentan, respectivamente, La biomasa de esta especie fue estimada por el método de estratifica137 Pota, calamar gigante o jibia, Dosidicus gigas La zona del norte presenta los mayores índices de abundancia, especialmente por fuera de Punta Chao, Salaverry, Chicama y Mórrope. Informe IMARPE Vol. 35(2): Abril-Junio 2008 84°W 82°W 80°W 78°W 76°W 74°W 4°S 4°S Pto. Pizarro Zorritos Pta. Sal Cabo Blanco Talara LEYENDA: Paita Pta. Gobernador 6°S 6°S 35.1 ups Límite de las 100 Brazas Parachique Pta. La Negra Mórrope Pimentel 8°S 8°S Chérrepe Pacasmayo Chicama Trujillo Salaverry Punta Chao Chimbote Casma 10°S 10°S Punta Lobos Huarmey Punta Bermejo Supe Huacho 1000 100 14°S 50 10 5 Callao Pucusana Cerro Azul Tambo de Mora Pisco Bahía Independencia 14°S 250 Valores ecointegrados 12°S 500 12°S Chancay 750 Punta Infiernillos Punta Caballas 1 San Juan 84°W 82°W 80°W 78°W 76°W 74°W Figura 14.- Distribución de calamar gigante según valores ecointegrados NASC (m2/mn2) y su relación con la salinidad (masa de aguas). 84°W 82°W 80°W 78°W 76°W 74°W 4°S 4°S Pto. Pizarro Zorritos Pta. Sal Cabo Blanco Talara LEYENDA: Paita Pta. Gobernador 6°S 6°S 35.1 ups Límite de las 100 Brazas Parachique Pta. La Negra Mórrope Pimentel 8°S 8°S Chérrepe Pacasmayo Chicama Trujillo Salaverry La zona del sur, de similar magnitud en términos del área de distribución, presenta núcleos más reducidos, que se ubican por fuera de la Península de Paracas y Punta Caballas. La abundancia fue estimada en 0,29x106 t, y límites estadísticos de confianza en 10,18% (Tabla 2). Vinciguerria lucetia La vinciguerria presentó una abundancia inusual para la estación invernal, en gran parte del área evaluada al sur de Punta Bermejo como producto de la influencia de ASS, con salinidad hasta de 35,1 ups. La llegada de dos ondas Kelvin durante los meses de julio y agosto ha significado que las masas de aguas subtropicales se acerquen a la costa en magnitudes mayores a lo que es común observar en el invierno, favoreciendo la presencia y abundancia, de especies mesopelágicas en la zona,. El aumento del volumen de ACF entre Punta Bermejo y Sechura ha significado que en dicha zona, sea menor la abundancia latitudinal de este pez, cuya distribución longitudinal se ve limitada por la proyección de la plataforma, donde la isóbata de las 100 bz aparece como el límite de su distribución (Figura 15). La biomasa de vinciguerria fue estimada en 10,42x106 t, con límites estadísticos de confianza en 7,02 %; la mayor abundancia de esta especie fue detectada en 12 y 13°S (Tabla 2). Punta Chao Chimbote DISCUSIÓN Casma 10°S 10°S Punta Lobos Huarmey Punta Bermejo Supe Huacho 5000 1000 750 500 14°S 250 100 50 10 12°S 2000 Chancay Callao Pucusana Cerro Azul Tambo de Mora Pisco Bahía Independencia 14°S 3000 Valores ecointegrados 12°S 4000 Punta Infiernillos 5 Punta Caballas 0.1 San Juan 84°W 82°W 80°W 78°W 76°W 74°W Figura 15.- Distribución de vinciguerria según valores ecointegrados NASC (m2/mn2) y su relación con la salinidad (masas de aguas). 138 La anchoveta Desde el punto de vista de seguimiento de las pesquerías, los resultados sobre anchoveta muestran que las estructuras de tallas determinadas en el Crucero 0402-03, de evaluación de recursos pelágicos, tuvieron una progresión positiva, es decir que en el verano, predominaron individuos con LT <10 cm (rango 3 a 17 cm). En el crucero 0408-09 se obtuvo una estructura mayoritariamente de adultos con LT 12 cm (rango 10 a 17 cm). Evaluación hidroacústica de especies pelágicas en invierno 2004 Gutiérrez y Peraltilla La abundancia determinada, si bien es menor que durante el verano (11,2x106 t), es concordante con aquélla, considerando los desembarques en el período abril - julio 2004. De otro lado, la cobertura del crucero demostró que, al menos en algunas áreas, la distribución y por lo tanto la abundancia de anchoveta sería mayor a la determinada en esta ocasión. Un ejemplo de ello fue el perfil Callao, donde se detectó anchoveta hasta las 180 mn de la costa; algo parecido ocurrió en el perfil Pimentel, pero la rigidez del itinerario impide extender los transectos más allá del período de tiempo programado. Además, la zona al sur de los 15°S no ha sido prospectada, por lo que se puede afirmar que la cifra dada como resultado de la abundancia es conservadora. En cuanto a la distribución latitudinal, la anchoveta presentó una estructura espacial que, a seis años de El Niño 1997-98, podríamos definir como “estándar”, ya que con la excepción del débil Niño de 2002 las principales zonas de abundancia han sido siempre aquellas ubicadas entre los 6 y 10°S, y así ha sido en el caso presente. El resto del litoral prospectado presenta abundancias significativas, pero menores a las latitudes mencionadas, incluyendo la extensión de la distribución de anchoveta hacia el norte, hasta 4°S y en ocasiones hasta el 3°S, desde la primavera 2000, pero en todo caso dentro de un patrón general bastante bien definido, al menos para los últimos años, la mencionada distribución “estándar”. Longitudinalmente, y aceptando que habría zonas con anchoveta que no han sido evaluadas, la distribución hallada, desde un punto de vista estacional, está más de acuerdo con una estación de primavera, con una aproximación a la costa de las ASS, o una retracción de las ACF que genera usualmente vastas zonas de aguas de mezcla igualmente ideales para la distribución de la especie. En el caso presente, la ocurrencia entre julio y agosto de sucesivas ondas Kelvin que se propagaron sobre el pacífico sudeste, se tradujeron en un ligero calentamiento y el acercamiento a la costa de las ASS, con concentraciones de 35,1 y 35,2 ups a 40 y 50 mn de la costa, en promedio. Esta es la razón por la cual la anchoveta se congregó sobre la costa y mostró los altos índices de densidad acústica que han sido determinados, y es además la explicación para la alta e inusual abundancia (considerando la estación invernal) que ha sido determinada para vinciguerria. Los análisis GAM realizados para anchoveta contribuyen a la descripción del efecto espacio-temporal de las condiciones oceanográficas sobre los cardúmenes de la especie. Entre estos resultados destacan la determinación de la variación horaria en la dimensión y ubicación vertical de los cardúmenes, que son aspectos aún en estudio, pero la claridad de las relaciones halladas permite vislumbrar su utilidad futura en un manejo dinámico de las labores extractivas. Por ejemplo, durante el crucero los cardúmenes con la mayor área de sección transversal se ubicaron entre Callao y bahía Independencia, no obstante la mayor abundancia de anchoveta en el norte. El objetivo de esta investigación, que se describe brevemente, es conocer cómo la anchoveta responde a diversos escenarios y condiciones. Así como desde el Crucero 0402-03 los análisis GAM se han incorporado como herramienta de análisis de la información acústica, en el caso presente se ha incorporado el método de estratificación por transectos como medio de validación del método tradicional (por áreas isoparalitorales); además, la determinación de las proporciones acústicas por tallas (K) permitirá mejorar la evaluación acústica en el sentido de que se determina la abundancia por clases y se utiliza en mucho mayor volumen la cuantiosa información que proviene de los muestreos biológicos post-lance de pesca. Otros recursos que mostraron una distribución y abundancia importantes fueron múnida, pota y vinciguerria. La vinciguerria es la especie que incrementó su presencia en la zona prospectada, 139 en comparación con lo observado en similares períodos (1999-2003), como producto de la inusual influencia estacional de ASS. La múnida es una especie que aparece endémica de la Corriente Peruana desde su presencia en 1994; sin embargo, su distribución en el mar peruano es cíclica, según lo descrito por Schweigger (1964). Los estudios acústicos desarrollados en los últimos 10 años sobre múnida muestran que presenta una variabilidad estacional respecto a su abundancia (más en invierno, menos en verano), pero que existe una progresión norte-sur en su distribución latitudinal conforme nos adentramos en el ciclo frío interdecadal definido como La Vieja (Chavez 2003). De otro lado, no se tiene aún el resultado específico acerca de los hábitos alimentarios de múnida en aguas peruanas, pero es muy posible que su dieta esté compuesta, entre otros, de huevos de anchoveta, como se ha determinado en Chile (Dr. Leonardo Castro, Universidad de Concepción, en comunicación personal). En efecto, existe cierta sobreposición en la distribución de anchoveta y múnida (esta última se distribuye siempre en relación con ACF, y por lo general muy cerca de la costa), aunque solo en horas nocturnas aparecen mezcladas. En horas diurnas existe una clara estratificación vertical entre las dos especies (anchoveta por lo general más cerca de la superficie). Este y otros aspectos de la morfología de los enjambres de múnida han sido también estudiados recurriendo a los análisis GAM. De esta manera se ha podido mostrar (Figuras 5 y 13), por ejemplo, que la distribución vertical horaria de anchoveta (profundidad de cardúmenes, paneles “f” en ambas figuras) se da en un rango de unos pocos metros, pero que la de múnida ocurre en un rango medio de 30 m. Asimismo, otros paneles muestran que los parámetros geométricos de los enjambres de múnida son bastante más estables –latitudinalmente- que los de los cardúmenes de anchoveta, lo que indicaría el bienestar de la múnida en el ecosistema costero. Informe IMARPE Vol. 35(2): Abril-Junio 2008 En el caso del calamar gigante, la asignación de valores de ecointegración NASC que se emplean en la determinación de la abundancia, ha dependido por lo general de las proporciones de captura durante los lances efectuados debido a la dificultad de discriminar sus registros de las de otras especies (los calamares en general poseen una baja reflectividad); sólo en altos volúmenes de densidad el calamar gigante es acústicamente distinguible, todo lo cual constituye una fuente constante de incertidumbre en la estimación de su abundancia (Gutiérrez 2002). La distribución de la especie estuvo dentro de los rangos esperados: salinidades mayores de 35,1 ups, sobre profundidades también mayores a 100 bz. Es conveniente comentar que el calamar gigante tiene una distribución muy grande en el Pacífico sudeste, al igual que vinciguerria, por lo que las cifras de abundancia que han sido calculadas están obviamente subestimadas, y corresponden únicamente a la zona que ha sido prospectada. CONCLUSIONES • La distribución y abundancia determinadas para anchoveta y vinciguerria guardan concordancia con las condiciones ambientales existentes al momento de efectuarse el Crucero 0408-09. • La abundancia y estructura demográfica determinadas para anchoveta corresponden con la estructura y biomasa estimadas para la especie en el verano (Crucero 0402-03); considerando el efecto de la pesquería, más las zonas no evaluadas (al sur y al oeste), la diferencia en la abundancia estimada durante los dos cruceros es coherente. • Se estimó la biomasa de anchoveta en 7,49x106 toneladas, con límites estadísticos de confianza de 12,85%. El 11,47% de la biomasa corresponde a individuos hasta de 12 cm LT. • La múnida mostró una amplia distribución costera, desplazando a la anchoveta en ciertas zonas, dentro de las cuales alcanzó sus mayores índices de abundancia. La biomasa de esta especie ha sido estimada en 1,17x106 t, con límite estadístico de confianza en 11,88 %. • Pota se distribuyó en dos amplias zonas, en el norte y en el sur, alcanzando una biomasa de 0,29x106 t, con límite estadístico de confianza en 10,18%. • Vinciguerria mostró una abundancia inusual para la estación, como producto de la marcada influencia de ASS especialmente en el sur de la zona evaluada. Su abundancia fue estimada en 10.42x106 t, con límite estadístico de confianza en 7,02%. REFERENCIAS Bazigos G. 1976. Applied Fishery Statistics. FAO Fish. Tech. Pap. (135): 164 pp. Bertrand A, Cordova J, Barbieri MA, Leiva F, Gerlotto F. 2004. Opposite day and night ecological preferenda on jack mackerel (Trachurus murphyi) in the Humboldt Current system: a ‘unique’ behavioural adaptation to cyclic availability of preys. ICES Journal of Marine Science. In press. Chavez F, Ryan J, Lluch-Cota S, Ñiquen M. 2003. From anchovies to sardines and back: multidecadal change in the Pacific Ocean. Science Vol 299: 217-221 Foote KG. 1987. Fish target strength for use in echointegrator surveys. Journal of the Acoustical Society of America, 82:981-987 Foote KG, Knudsen HP, Vestnes G, MacLennan DN, Simmonds EJ. 1987. Calibration of acoustic instruments for fish density estimation: a practical guide. Cooperative Research Report, Internacional Council for the Exploration of the Sea, 144, 57 pp. Gutiérrez M, Peraltilla S. 1999. 140 Aplicación de un Sistema de Información Geográfica y de la carta electrónica isoparalitoral en las evaluaciones hidroacústicas de la biomasa de recursos pesqueros en el mar peruano. Inf. Inst. Mar Perú 146: 25-29 Gutiérrez M. 1997. Aplicación de software de interpolación en las evaluaciones hidroacústicas de la biomasa y distribución de recursos pelágicos. Inf. Prog. Inst. Mar Perú 67: 21-30. Gutiérrez M. 2002. Determinación de la Fuerza de Blanco (TS) de las principales especies del mar peruano. Tesis (¿con qué fin?). Universidad Nacional Federico Villarreal. 81 pp. Higgimbottom I, Pauly T, Heatley D. 2000. Virtual echograms for visualization processing of multiplefrequency echosounder data. Proceedings of the Fifth European Conference on Underwater Acoustics, ECUA 2000 (Ed. M.E. Zakharia), 1497-1502. Jolly D, Hampton I. 1990. A stratified random transect design for acoustic surveys of fish. Can. J. Fish. Aquat. Sci. 47: 1282-1291. MacLennan DN, Fernandez P. 2000. Definitions, Units and Symbols in Fisheries Acoustics. FASTICES Working Group Meeting. St. Johns´Newfoundland, Canada, April 1999 (Mimeo). 6 pp. MacLennan DN. 1998. Revisión de los resultados obtenidos durante el Crucero 9803-05 de Evaluación Hidroacústica de Recursos Pelágicos. Inf. Inst. Mar Perú N°135: 191-201. MacLennan D, Simmonds J. 1992, Fisheries Acoustics. Chapman and Hall, London. Hewitt R, Kim S, Naganobu M, Gutierrez M, Kang D, Takao Y, Quinones J, Lee Y-H, Shin H-C, Kawaguchi S, Emery JH, Demer DA, Loeb VJ. 2003. Variation in biomass density and demography of Antarctic krill in the vicinity of the South Shetland Islands during the 1999/2000 austral summer. En prensa (Deep Sea Research). Schweigger E. 1964. El Litoral Peruano. Fondo Editorial de la Universidad Nacional Federico Villarreal, Lima. 455 pp. Biomasa desovante de la anchoveta peruana (MPH). Invierno 2005 Ayón y Perea BIOMASA DESOVANTE DE ANCHOVETA EN LA ZONA NORTE CENTRO DEL MAR PERUANO (4O36,5’ – 14O40’S) A FINALES DEL INVIERNO AUSTRAL 2005 THE ANCHOVY SPAWNING BIOMASS IN THE NORTH CENTRAL ZONE OF PERUVIAN SEA (4O36,5’ – 14O40’S) AT THE LATE AUSTRAL WINTER 2005 Patricia Ayón1 M. Ángel Perea2 RESUMEN AyÓn P, Perea MA. 2008.- Biomasa desovante de anchoveta en la zona norte centro del mar peruano (4°36,5’ – 14°40’S) a finales del invierno austral 2005. Inf. Inst. Mar Perú 35(2): 141-144.- La biomasa desovante entre Punta Infiernillos (14O40’S) y Paita (4°36,5S) fue estimada en 6,1 millones de toneladas (límites de confianza del 95% de 42,29%), y un coeficiente de variación de 21,15%. El peso promedio de las hembras (W) fue 16,27 g, la fecundidad parcial (E), 11.386 ovocitos/hembra, con una proporción sexual (R) de 0,51 y una frecuencia de desove de 0,0984. La producción diaria de huevos fue de 2,16E+14 huevos/día. Se hizo un análisis de las variables y los resultados se compararon con los obtenidos por el método acústico. Palabras clave: MPH, Engraulis ringens, anchoveta peruana, biomasa desovante, fines de otoño 2005. ABSTRACT AyÓn P, Perea MA. 2008.- Spawning biomass of the anchovy in the north central zone of Peruvian sea (4°36,5’ – 14°40’S) at the end of Austral winter 2005. Inf. Inst. Mar Perú 35(2): 141-144.- The spawning biomass between Punta Infiernillos (14O40’S) and Paita (4°36,5S) was calculated in 6.1 million tons with 95% confidential limits of ±42,29% and a variability coefficient of 21,15%. The female individuals average weight (W) was 16,27 g; the batch fecundity (E) was 11.386 ovocytes/female, with a sex ratio (R) of 0,51 and a spawning frequency of 0,0984. The daily egg production was 2,16E+14 egg/day. Analysis of variables and comparison with results obtained by acoustical method was done. Keywords: MPH, Engraulis ringens, Peruvian anchovy, spawning biomass, late autumn 2005. INTRODUCCIÓN La anchoveta peruana, Engraulis ringens, constituye uno de los peces marinos de mayor importancia económica para el Perú, lo cual motiva al Ministerio de la Producción, por intermedio del IMARPE, a realizar cruceros para evaluar el estado de dicho recurso. En la actualidad el IMARPE cuenta con varios métodos para estimar su biomasa. El Método de Producción de Huevos (MPH) es bastante confiable, por lo cual el IMARPE lo utiliza como un método directo que permite estudiar el estado biológico de la población así como sus niveles de abundancia. La aplicación del MPH ha permitido conocer el estado y comportamiento de la especie en el momento de su reproducción y las diferentes estrategias que la 1 2 especie utiliza en relación con su entorno ambiental. En el verano del 2005, la biomasa de la anchoveta se estimó en 12,7 millones de toneladas distribuida desde Cabo Blanco en el norte hasta Los Palos en el sur; y a una distancia máxima de 90 mn por fuera de Salaverry y Chimbote. En general, con un promedio de 50 mn cuando las condiciones ambientales mostraban la influencia de las AES hacia el norte, y las ASS en el extremo sur (IMARPE 2005). El rango de LT fue de 3,0 a 18,0 cm, con modas en juveniles de 9,0 - 9,5 cm, y de 13,5 y 15,0 cm en adultos. En la zona entre Pimentel y Callao se registró un importante contingente juvenil con modas desde 7,0 a 11,0 cm de LT, cuyas las tallas menores se acercaban a la costa. En esta zona, el stock norte-centro mostró gran éxito de reclutamiento, en Area de Evaluación de Producción Secundaria, [email protected] Biología reproductiva, [email protected] 141 abundancia y extensión, y la fracción de juveniles alcanzó el 67% en número y el 39% en biomasa. Con estos antecedentes del estado del recurso en el verano 2005, se realizó el crucero anual a fines de otoño, para estimar la biomasa desovante de anchoveta utilizando el Método de Producción de Huevos (MPH). MATERIAL Y MÉTODOS El crucero BIC Olaya, SNP2 e IMARPE VI 0508-09 se ejecutó entre el 15 de agosto y el 24 de setiembre del 2005. Cubrió el área entre Talara (4°36,5’S) y Punta Infiernillos (14°40’S). La trayectoria del crucero se hizo en 54 transectos perpendiculares a la línea de costa, con distancia promedio entre ellos de 13 millas. El BIC Olaya trabajó en la zona comprendida entre 40 y 90 mn de la costa entre Informe IMARPE Vol. 35(2): Abril-Junio 2008 5°S 12 los primeros 800 g. Para el cálculo de fecundidad de las hembras se hizo una regresión lineal a partir de 155 hembras, y se utilizó el promedio de la fecundidad en las calas donde se encontraron las hembras hidratadas. Los cálculos estadísticos siguieron el método de Santander et al. (1984). 13 11 10 9 8 7 7°S 6 5 312 La producción diaria de huevos se determinó de acuerdo a AyÓn (2003), adaptado de Santander et. al (1984). La estimación de la biomasa en estado de reproducción se obtuvo mediante la ecuación general (Parker 1980, Stauffer Picquelle 1980): 4 C34 C31 C30 534/C28 474/C 25 9°S C 29 C27 C26 C23 C20/C21 C 24 C22 C18 C19 C 17 C23 C20 C 17 C16C15 B=PoW/FER C 14 C13 11°S C 15 C13 105-C11 102-C9 C12 103-C10 En donde B: biomasa reproductora, Po: producción de huevos diaria; W: peso promedio; F: frecuencia de desove; E: fecundidad; R: proporción sexual. C9 C7 C6 C11 C8 C5 13°S 75/C7 58/C6 38/C4 34/C3 C3 24/C2 3C-1 15°S 82°W 80°W 78°W 76°W 74°W Figura 1.- Carta de distribución e las calas efecutadas durante el Crucero Olaya, SNP2 e IMARPE VI 0508-09. Punta Infiernillos y Chimbote, salvo algunos transectos que se prolongaron más allá de las 100 mn y otros efectuados dentro de las 40 mn. Al norte de Chimbote dicho barco cubrió tanto la parte costera como la oceánica, extendiéndose en algunos transectos hasta 170 millas de la costa (Figura 1). El SNP2 exploró la parte costera hasta 40 mn de la costa entre Punta Infiernillos y Chimbote. El IMARPE VI apoyó en las faenas de pesca en el área comprendida entre Pimentel y Paita. Se ejecutaron 70 lances; sólo 33 de ellos fueron válidos para el MPH (Figura 1). La colecta de adultos y de huevos estuvo de acuerdo con la metodología descrita por Santander et al. (1984). De manera paralela al muestreo de red CalVET, se colectaron muestras con el equipo CUFES a fin de complementar la información de la presencia de huevos de anchoveta in situ. La temperatura se midió a 3 m de profundidad, dato que se utilizó en el cálculo de las categorías de edad de los huevos, para estimar su producción diaria. Entre Talara y Pimentel, más allá de las 20 mn, debido a problemas de logística, sólo se realizaron observaciones acústicas, sin efectuar lances de comprobación. Sólo fue factible ejecutar calas en la zona costera, con la IMARPE VI. En el cálculo de los parámetros de peso (W) y frecuencia de desove (F) se utilizaron las primeras 25 hembras de cada cala; en el caso de la proporción sexual (R), se utilizaron Para el estimado del coeficiente de variación se empleó la ecuación descrita por Parker (1980), en donde los términos de las covarianzas son considerados a su mínimo valor equivalente a cero, por lo que finalmente el Coeficiente de Variación es: CV (B) = V (CV Po)2 + (CV W)2 +(CV E)2 + (CV F)2 + (CV R)2 RESULTADOS Parámetros de adultos El peso promedio de hembras (W) fue de 16,27 g con un CV equivalente al 4,1%. La fecundidad parcial (E), es decir el número de ovocitos hidratados, fue de 11.386 ovocitos/hembra con un coeficiente de variación de 0,3%. La proporción sexual (R) fue de 0,51 con un coeficiente de variación del 3,5%. La frecuencia de desove Tabla 1.- Parámetros de adultos Promedio Varianza Desv. CV R 0,518 0,00035 0,018 0,035 142 W 16,270 0,0000 0,000 0,041 E 11386 1125 33,537 0,003 F 0,0984 0,0003 0,0175 0,178 Po 2,16E+14 4,62E+26 2,15E+13 0,10059 Biomasa desovante de la anchoveta peruana (MPH). Invierno 2005 fue de 0,0984 con un coeficiente de variación del 17,8% (Tabla 1). 4° S ProducciÓn diaria de huevos Para estimar este parámetro se utilizaron las categorías de edad A1, A2, B1 y B2 (Tabla 2). La producción de huevos de anchoveta se calculó en 2,16E+14 huevos/día, con una tasa de mortalidad de 1,06E+14, que equivale a decir que 64% de los huevos estaban muriendo diariamente. Tabla 2.- Producción diaria de huevos Categoría de edad Abundancia (huevos/día) A1 A2 B1 B2 1,4228 E+14 0,8526 E+14 0,4044 E+14 0,3773 E+14 Pto. Pizarro Zorritos Pta. Sal Cabo Blanco Talara 4° S N° huevos Paita Pta. Gobernador 6° S 1 a 50 50 a 100 100 a 1000 1000 a 2000 Parachique Pta. La Negra Mórrope Pimentel Chérrepe Pacasmayo Chicama Trujillo Salaverry 8° S 750 500 250 100 Punta Chao Chimbote 50 Casma Punta Lobos Huarmey 10° S 10 El presente estimado de biomasa desovante de la anchoveta por el método de producción de huevos a fines de otoño 2005 (6.100.000 t), ha sido uno de los más altos después del otoño 1985, cuando la evaluación llegó a 8.500.000 t, oportunidad en que la población estaba constituida principalmente por individuos de LT entre 9,5 y 19 cm, con modas en 16 y 17 cm (Santander 1985, Informe interno). En la presente evaluación la LT tuvo un rango de 7 a 18 cm, con una moda en 13 cm (IMARPE 2005). En cuanto a la variación de los parámetros, el peso y proporción sexual fue baja (<4%); la frecuencia de desove (17,8%) fue la variación más alta; y la fecundidad ha sido mayor que las registradas anteriormente. Esto pudo estar relacionado con el hecho de haber realizado calas donde la fecundidad era muy baja (4.462 6° S 8° S 10° S 0.1 Chancay 12° S 12° S Callao Pucusana Cerro Azul Tambo de Mora Pisco 14° S Bahía Independencia Biomasa desovante DISCUSIÓN 5 Punta Bermejo Supe Huacho 14° S El estimado de la biomasa entre Punta Infiernillos y Paita del 15 de agosto al 24 de setiembre fue de 6,1 millones de toneladas, con límites de confianza al 95% de ± 42,29%, y un coeficiente de variación de 21,15%. 1000 Valores ecointegrados (m²/mn²) Ayón y Perea Punta Infiernillos Punta Caballas San Juan Chala 16° S 83° W Atico 81° W 79° W 77° W 16° S 75° W Figura 2.- Distribución y abundancia de huevos y adultos de anchoveta. Crucero 0508-09, ovocitos/hembra), debido a que los ejemplares eran muy jóvenes (9,5 a 12 cm), con moda en 11 cm y todos en condición de desovantes. En este crucero, la biomasa desovante calculada a partir del MPH, ha tenido valores semejantes a las 7.500.000 t obtenidas por el método acústico, para la misma época y área de muestreo. Estos cálculos han ocasionado 2 interrogantes: (1) ¿La biomasa desovante ha sido muy alta para este periodo en comparación con los juveniles? Para este cuestionamiento tenemos evidencia que ejemplares pequeños, <12 cm, han mostrado alto porcentaje de actividad reproductora, incluso algunos individuos de 9,5 cm se encontraron en estado reproductivo (BuitrÓn y Perea, comunicación personal), tamaños que corresponden a ejemplares en condición de juveniles. (2) ¿Es posible que se haya subesti143 mado la biomasa total por el método acústico?. En este caso, cuando sobreponemos la carta de distribución de adultos con la de los huevos, se nota áreas donde sólo se ve la presencia de huevos y no la de adultos (Figura 2). Sin embargo, en esta zona con otro aparejo (CUFES que opera a 3 m de profundidad) se encontraron adultos en las muestras. Asimismo, la edad de los huevos determinada en esta zona correspondía en muchos casos a huevos recién desovados, lo cual indicaba directamente que los padres se encontraban en la zona y probablemente localizados en capas más superficiales. Agradecimientos.- Los autores queremos expresar nuestro profundo agradecimiento al grupo del Área de Evaluación de Producción Secundaria y Biología Reproductiva por el apoyo en el análisis de las muestras. Informe IMARPE Vol. 35(2): Abril-Junio 2008 REFERENCIAS AyÓn P. 2008. Producción diaria de huevos de anchoveta peruana en agosto - setiembre 2003. Inf. Inst. Mar Perú 35(2): 81-84 IMARPE. 2005. Crucero 0502-04 de Evaluación Hidroacústica de Recursos Pelágicos (Tumbes a Tacna) del 20 febrero al 04 de abril del 2005. Informe Ejecutivo. Parker K. 1980. A direct method for estimating northern anchoveta, Engraulis mordax, spawning biomass. Fishery Bulletin 78(2): 541-544. Parker K. 1985. Biomass model for the Egg Production Method. Reuben Lasker (Ed.). An Egg Production Method for estimating Spawning Biomass of Pelagic Fish: Application to the Northern Anchovy, Engraulis mordax. NOAA Technical Report NMFS 36. Santander H, Alheit J, Smith P. 1984. Estimación de la biomasa de la población desovante de anchoveta peruana Engraulis ringens en 1981 por aplicación del Método de Producción de Huevos. Bol. Inst. Mar Perú. 8(6):209-250. 144 Santander H. 1985. Evaluación de las poblaciones desovantes de anchoveta y sardina por el método de producción de huevos. Crucero BIC Humboldt 8508-09 y operaciones en bolicheras 28 agosto-5 octubre 1985. Stauffer GD, Picquelle SJ. 1980. Estimates of the 1980 spawning biomass of the central subpopulation of northern anchovy. Southwest Fisheries Center. Adm. Rep. LJ 80-09. Estimación hidroacústica de especies pelágicas en invierno 2005 Castillo, Peraltilla y Gutiérrez BIOMASA DE ANCHOVETA Y OTROS RECURSOS PELÁGICOS ESTIMADOS POR EL MÉTODO HIDROACÚSTICO A FINALES DEL INVIERNO 2005 PERUVIAN ANCHOVY AND OTHER PELAGIC RESOURCES ESTIMATED BY THE HYDROACOUSTIC METHOD AT LATE WINTER 2005 Ramiro Castillo Salvador Peraltilla Mariano Gutiérrez RESUMEN Castillo R, Peraltilla S, Gutiérrez M. 2008. Biomasa de anchoveta y otros recursos pelágicos estimados por el método hidroacústico a finales del invierno 2008. Inf. Inst. Mar Perú. 35 (2): 145-152.- El crucero BIC Olaya, SNP2 e Imarpe VI 0508-09 de evaluación de biomasa desovante de anchoveta se ejecutó entre el 15 de agosto y el 24 de setiembre del 2005, de Punta Infiernillos a Talara. Se determinó la distribución y biomasa de la anchoveta y otros recursos pelágicos. La especie más importante fue la anchoveta (Engraulis ringens), con amplia distribución entre Punta La Negra y Chimbote (hasta 150 mn frente a Salaverry) y costera entre Pucusana y Bahía Independencia; su distribución estuvo limitada por la isoterma superficial de 17,5 °C, y por la isohalina de 35,12 ups, mostró características discontinuas, especialmente hacia el sur de Salaverry. La biomasa estimada por el método de áreas isoparalitorales fue 7,66 x 106 t con un intervalo de confianza de 17,61%; en la estimación por transectos fue relativamente menor. El jurel (273.629 t) y la caballa (176.842 t) se localizaron principalmente en el sur. Cerca de la costa se detectaron samasa (96.894 t), bagre (28.487 t) y múnida (965.474 t). La pota presentó una importante biomasa de 433.400 t. La vinciguerria fue más abundante en la zona sur. Palabras clave: anchoveta, recursos pelágicos, otoño 2005, evaluación hidroacústica, mar peruano. ABSTRACT Castillo R, Peraltilla S, Gutiérrez M. 2008. Peruvian anchovy and other pelagic resources estimated by the hydroacoustic at late winter 2008. Inf. Inst. Mar Peru. 35 (2): 145-152 .- The BIC Olaya, SNP2 and Imarpe VI Cruise 0508-09 to evaluate the spawning biomass of anchovy was executed from August 15th to September 24th, 2005, from Punta Infiernillo to Talara. The distribution and biomass of the anchovy and other pelagic resources was determined. The most important species was the anchovy, with wide distribution between Punta La Negra and Chimbote (up to 150 mn in front of Salaverry), and near to coastal Pucusana and Bahía Independencia; its distribution was limited by the surface isotherm 17.5 °C, and the isohaline of 35.12 psu, and showed discontinuous nature, especially at south of Salaverry. Biomass estimated by the method of isoparalitoral areas was 7.66 x 106 t with a confidence level of 17.61%; in the estimation of transects was relatively minor. Jack mackerel (273,629 t) and chub mackerel (176,842 t) were located mainly in the south. Near the coast were detected samasa anchovy (96,894 t), catfish (28,487 t) and múnida (965,474 t). The giant squid keeps important biomass of 433,400 t. The vinciguerria was more abundant in the south. Keywords: anchovy, pelagic resources, Autumn 2005, hydroacoustic evaluation, Peruvian sea. INTRODUCCIÓN El crucero BIC Olaya, SNP2 e Imarpe VI 0508-09 de evaluación de biomasa desovante de anchoveta, por el método de la producción de huevos (MPH) se ejecutó entre el 15 de agosto y el 24 de setiembre del 2005, de Punta Infiernillos (14O40’S) a Paita (4°36,5S). En este crucero realizado en el 2004, la anchoveta tuvo una distribución amplia y mostró dos grandes zonas de agregación: al norte de Huacho y al sur de Callao. Los mayores núcleos de abundancia se ubicaron por fuera de las 25 mn, con una biomasa de 7,49 x106 t, con límites estadísticos de confianza de 12,85%. El 11,47% de la biomasa correspondió a individuos con LT <12 cm. Posteriormente, se hallaron comportamientos diferentes. En la evaluación hidroacústica de recursos pelágicos realizada en la primavera 2004 mostró cierta discontinuidad en su distribución; y en el verano 2005, fue casi continua, de norte a sur, con una biomasa total de 12.713.668 t, con la mayor abundancia en los 8°S (2,54 x106 t) y 7°S (2,12 x106 t),. Esta biomasa fue la mas alta estimada en los últimos años. También se han estimado las biomasas y áreas de distribución de otras especies pelágicas: jurel, ca145 balla, samasa, bagre, múnida, pota y vinciguerria. MATERIAL Y MÉTODOS El estudio se llevó a cabo entre el 16 de agosto al 26 de septiembre de 2005. Las plataformas de investigación fueron los BIC José Olaya Balandra, SNP-2 y la LIC IMARPE VI. El área evaluada fue entre Paita (Talara, 04°36`’S) y Punta Infiernillos (14°40’S.). El BIC SNP2 prospectó la zona costera en transectos hasta las 40 mn; y el BIC Olaya, entre 40 y 90 mn, y en algunos casos más allá de las 100 mn; y otros dentro de las 40 mn. Informe IMARPE Vol. 35(2): Abril-Junio 2008 82° W 80° W 78° W Pto. Pizarro Zorritos Pta. Sal Cabo Blanco TRAYECTOS Talara 4° S Paita Pta. Gobernador 76° W 74° W 84° W 4° S 4° S 6° S Crucero MPH 0508-09 8° S BIC OLAYA 8° S Chimbote Casma Punta Lobos Huarmey 10° S Casma Punta Lobos Huarmey 10° S Punta Bermejo Supe Huacho 12° S Pucusana Cerro Azul Tambo de Mora Pisco 14° S 14° S Bahía Independencia Punta In ernillos Punta Caballas San Juan Atico 82° W 80° W 78° W 76° W 16° S 74° W Biomasa Numpeces Cerro Azul Tambo de Mora Pisco 100 2.E+11 1,386,259 2.E+11 Punta In ernillos Punta Caballas 10 San Juan 0.1 Atico 82° W 80° W 1.E+11 792,148 8.E+10 594,111 4.E+10 198,037 2.E+10 Numpeces Biomasa 12 13 14 15 16 17 18 8. 0 11 7. 0 10 6. 0 9 5. 0 8 4. 0 7 3. 0 6 2. 0 5 En ambos barcos se utilizó la ecosonda científica Simrad EK-500 para la detección e ecointegración de la reflexión de peces, que operaron en las frecuencias de 38 y 120 kHz. La cobertura del muestreo vertical a 120 kHz fue de 0 a 150 m; y para 38 kHz, de 0 a 500 m. La calibración de los equipos acústicos fue previa al inicio del crucero; y la íntercalibración entre ambos 1,200,000 1,000,000 400,000 2.E+10 200,000 0.E+00 0 Talla (cm) Latitud Figura 3.- Abundancia latitudinal de anchoveta. Crucero MPH 0508-09. En total, se muestrearon 6588 mn (4575 por el BIC Olaya y 1013 mn por el BIC SNP2), en 54 transectos de 0,5 a 90 mn en promedio de extensión. El diseño de la grilla de muestreo fue sistemático con trayectos paralelos y perpendiculares a línea de la costa; la separación entre transectos fue de 15 mn. El intervalo del muestreo fue de 1 mn (UBM). 1.E+11 600,000 1. 0 4 1,400,000 4.E+10 0.E+00 3 1.E+11 800,000 0. 0 0 74° W 6.E+10 6.E+10 396,074 76° W 8.E+10 Numpeces 990,185 78° W 16° S Figura 2.- Distribución de la anchoveta. Crucero MPH 0508-09. 1.E+11 1.E+11 14° S Bahía Independencia 50 2.E+11 1,584,296 1,188,222 12° S Pucusana 250 84° W Numpeces Biomasa (Ton) Callao 500 Chala 16° S Figura 1.- Carta de trayectos y lances. Crucero MPH 0508-09. 1,782,334 Chancay 750 5 Chala 16° S 1000 9. 0 10 .0 11 .0 12 .0 13 .0 14 .0 15 .0 16 .0 17 .0 18 .0 19 .0 20 .0 14° S Valores ecointegrados (m²/mn²) 12° S Callao 84° W 10° S Punta Bermejo Supe Huacho Chancay 12° S 8° S Punta Chao Chimbote 10° S 17.5 ° C Trujillo Salaverry BIC SNP.2 Punta Chao 6° S 35.12 ups Chérrepe Pacasmayo Chicama L/P IMARPE VI Trujillo Salaverry 74° W 4° S Mórrope Pimentel BIC SNP.2 Chérrepe Pacasmayo Chicama 76° W DISTRIBUCION DE ACHOVETA Parachique Pta. La Negra 6° S BIC OLAYA Mórrope Pimentel 78° W Paita Pta. Gobernador Crucero MPH 0508-09 8° S 80° W Pto. Pizarro Zorritos Pta. Sal Cabo Blanco Talara Y LANCES EJECUTADOS Parachique Pta. La Negra 6° S 82° W Biomasa (Ton) 84° W Figura 4.- Abundancia de anchoveta por tallas. Crucero MPH 0508-09. equipos de detección de los BIC se realizaron durante la evaluación. Los análisis fueron realizados empleando el programa S-Plus 6,0. Se utilizó un programa de postprocesamiento (Echoview), utilizando los módulos de detección de cardúmenes y de variables virtuales, lo que permitió el tratamiento de datos acústicos para la discriminación de ecotrazos, teniendo en cuenta los registros típicos de las especies y de los resultados de las capturas realizadas durante el crucero. Se efectuaron 70 lances de pesca entre las embarcaciones (34 del Olaya, 23 del SNP-2 y 13 de IMARPE VI). (Figura 1). La estimación de abundancia de anchoveta fue realizada a través de dos métodos: (1) el método convencional de estratificación por áreas isoparalitorales; y (2) el método de estratificación por transectos (Jolly y Hampton 1990; Hewitt et al. 2003, Gutiérrez 2004). Para delimitar el área de distribución o mapeo de los recursos, se utilizó el programa de interpolación de datos Surfer 8, a través del método krigging. Anchoveta: TS= 20 Log L - 76.25 dB 146 Las ecuaciones de fuerza del blanco (TS) utilizadas para la frecuencia de 120 kHz, fueron: Anchoveta: TS = 30.258 Log L - 89.353 dB Jurel: TS = 20 Log L - 71,9 dB Estimación hidroacústica de especies pelágicas en invierno 2005 Castillo, Peraltilla y Gutiérrez Caballa: TS = 20 Log L - 71,9 dB 6.E+09 4.E+09 3.E+09 50.000 Biomasa Número de individuos Bagre : 4TS = 20 Log L – 74,1 dB 1.E+09 0.E+00 1 6 11 16 21 26 31 36 41 46 51 56 61 66 71 76 81 86 91 96 101 106 111 116 121 126 131 136 141 146 151 156 161 166 171 176 181 186 191 196 Distancia a la costa (mn) Figura 5.- Abundancia de anchoveta según distancia a la costa. Crucero MPH 0508-09. Vinciguerria: TS = 20 Log L – 81,056 dB 3 Pota: TS = 31,2 Log L – 83,15 dB 6 4 5 7 Porcentaje latitudinal 8 9 10 11 12 13 14 RESULTADOS 15 16 DistribuciÓn y biomasa de anchoveta ₍Engraulis ringens₎ En general, el rango térmico superficial fue 15 a 18 °C, y la salinidad, de 34,8 a 35,2 ups; en estos casos, los núcleos densos correspondieron a áreas con los mayores patches de zooplancton, probablemente eufáusidos, detectados con frecuencia al norte del Callao. La distribución vertical de anchoveta en la zona costera, estuvo circunscrita a los primeros 25 m. En la zona oceánica, o alejada de la costa, por lo general entre 15 a 30 m. 17 20 19.5 19 18.5 18 17.5 17 16.5 16 15.5 15 14.5 14 13.5 13 12 11.5 11 10.5 10 9.5 9 8.5 8 7.5 7 7 6 5.5 5 4.5 4 3.5 3 2.5 2 1.5 1 0.5 18 0 Se registraron dos áreas de distribución. (1) La primera, muy amplia, limitada por la isoterma superficial de 17,5 °C e isohalina de 35,12 ups, situada entre Punta La Negra y Chimbote, alcanzó las 150 mn frente a Salaverry; donde se registraron los núcleos más densos. (2) Una zona costera con una población dispersa, entre Pucusana y Bahía Independencia, como consecuencia del acercamiento de las Aguas Subtropicales Superficiales (ASS). Los niveles de mayor abundancia y concentración se encontraron principalmente entre Mórrope y Salaverry (entre 30 a 100 mn de la costa), que coincidió con la proyección hacia el oeste de las Aguas Costeras Frías (ACF) (Figura 2). 2.E+09 0 Las ecuaciones de fuerza del blanco (TS) utilizadas para la frecuencia de 38 kHz, fueron: Los límites de confianza de los estimados de biomasa fueron determinados por Bazigos 1976, con un nivel de significancia de 5%. Número de individuos 5.E+09 100.000 12.5 Múnida: TS = 20 Log L – 95,40 dB 7.E+09 Biomasa (t) Samasa: TS = 20 Log L - 76,25 dB 150.000 Tallas (cm) Número de individuos Abundancia (t) Figura 6.- Abundancia latitudinal de anchoveta según tallas. Crucero MPH 0508-09. La biomasa estimada por el método de áreas isoparalitorales fue 7.661.212 t y por el método de estratificación por transectos fue de 6,77x106 t, con el 17,61% y 10,32% de límites de confianza, respectivamente. Las mayores biomasas se presentaron entre los 6° y 8°S. Con el método de áreas isoparalitorales el total fue 4,75 x106 t; y con el método de transectos, 4,16 x106 t. Similar característica muestra el número de ejemplares detectados. En el área de Punta La Negra a Chimbote se registró una mayor concentración de peces y alta densidad. Estas altas biomasas se localizaron en áreas con altos valores de integración (Tabla 1 y Figura 3). La abundancia por tallas en base al número de individuos (modas) se sitúa en la LT de 13 cm. La abundancia de individuos adultos (>12 cm) se estimó en 5,29 x106 t (78,17% del total); los juveniles alcanzaron una abundancia de 1,47 x106 t (21,83% del total) (Figura 4). 147 Según distancia a la costa, la mayor abundancia de anchoveta se registró dentro de las 75 mn. En esta distribución horizontal, entre las 28 y 37 mn de distancia de la costa hubo una notoria disminución de ecotrazos de anchoveta (Figura 5). La estructura de la biomasa por tallas, referida a los grados de latitud, mostró que la fracción juvenil estuvo distribuida principalmente entre los 4 y 7°S, con una pequeña fracción en los grados 13 y 14°S; y los individuos de mayor tamaño se ubicaron entre los 8 y 12°S y en el grado 15°S, principalmente ejemplares de 15 cm de LT (Figura 6). DistribuciÓn y biomasa de jurel ₍Trachurus murphyi₎ Se encontró principalmente en el sur del área evaluada, entre 60 y 160 mn de la costa (frente Punta Infiernillos y San Juan), en masas de agua con SSM de 35,1 y 35,2 ups. En el norte se detectaron entre 40 y 45 mn de Informe IMARPE Vol. 35(2): Abril-Junio 2008 Tabla 1.- Biomasa de las especies evaluadas por grado de latitud. Crucero MPH 0508-09. Tabla 1.- Biomasa de las especies evaluadas por grado de latitud. Crucero MPH 0508-09 Grado de latitud sur 03° 30 - 04° 00 03°30' - 04° 04° 00 - 04° 30 04° 30 - 05° 00 04° - 05° 05° 00 - 05° 30 05° 30 - 06° 00 05° - 06° 06° 00 - 06° 30 06° 30 - 07° 00 06° - 07° 07° 00 - 07° 30 07° 30 - 08° 00 07° - 08° 08° 00 - 08° 30 08° 30 - 09° 00 08° - 09° 09° 00 - 09° 30 09° 30 - 10° 00 09° - 10° 10° 00 - 10° 30 10° 30 - 11° 00 10° - 11° 11° 00 - 11° 30 11° 30 - 12° 00 11° - 12° 12° 00 - 12° 30 12° 30 - 13° 00 12° - 13° 13° 00 - 13° 30 13° 30 - 14° 00 13° -14° 14° 00 - 14° 30 14° 30 - 15° 00 14° -15° 15° 00 - 15° 30 15° 30 - 16° 00 15° -16° 16° 00 - 16° 30 16° 30 - 17° 00 16° -17° 17° 00 - 17° 30 17° 30 - 18° 00 17° -18° 18° 00 - 18° 30 18° -18°30' Anchoveta (t) Jurel (t) 7,365 7,365 691,475 36,977 728,452 473,525 1,424,864 1,898,389 532,883 843,644 1,376,527 918,315 560,480 1,478,795 326,861 69,219 396,079 469,711 28,676 498,387 150,133 61,104 211,237 196,018 442,393 638,410 92,716 92,899 185,614 73,344 157,557 230,901 1,446 9,608 11,054 Caballa (t) Samasa (t) Bagre (t) 1,410 1,410 20,683 74,801 95,484 46,386 46,386 Pota (t) Vinciguerria (t) DistribuciÓn y biomasa de caballa ₍Scomber japonicus₎ 995 995 Se observó por fuera de las 30 mn de la costa, coincidiendo con recursos demersales, y también con el incremento de anchoveta en zonas alejadas de la costa. Solamente se encontraron pequeños núcleos de alta concentración en áreas aisladas frente a Mórrope, Pacasmayo y entre Chimbote y Huarmey (Figura 8). La distribución vertical de la caballa varió entre 3 y 22 m. La biomasa estimada fue de 176.842 t, con mayor abundancia en los 7°S (56.991) y 10°S (59.752 t) (Tabla 1). 4,548 2,193 6,741 56,991 56,991 7,761 7,761 9,786 9,786 2,763 2,300 11,594 13,894 59,572 5,479 32,281 37,760 Munida (t) 2,763 59,572 823 584 1,407 57,177 57,177 135,753 34,741 170,494 8,197 177,770 147,066 324,836 92,565 122,956 215,521 49,932 17,932 67,865 15,205 16,220 31,425 619 5,818 6,437 18,270 71,723 89,993 7,064 60,333 67,397 141,467 19,539 161,006 10,425 32,999 43,424 2,306 3,567 5,873 42,438 42,438 71,863 71,863 5,568 5,568 107,825 107,825 47,199 47,199 DistribuciÓn y biomasa de samasa ₍Anchoa nasus₎ 22,662 22,662 8,197 TOTALES 7,661,212 84° W 82° W 273,629 176,842 80° W 78° W Pto. Pizarro Zorritos Pta. Sal Cabo Blanco Talara 4° S 28,458 76° W 965,474 346,862 74° W 0 84° W 4° S 82° W Crucero MPH 0508-09 Parachique Pta. La Negra Solamente se encontró en el norte, entre Punta La Negra y Pimentel, en una área costera de 17 mn, con pequeño núcleo de alta concentración frente a Pimentel (Figura 9). Verticalmente se le registró hasta los 22 m de profundidad. La biomasa estimada fue de 96.894 t, 80° W 4° S 6° S 8° S Punta Chao Chimbote Chimbote 10° S Casma Punta Lobos Huarmey 10° S Chancay 750 Callao 12° S 12° S Pucusana Cerro Azul 250 Tambo de Mora Pisco 100 14° S 14° S Bahía Independencia 50 Punta In ernillos Punta Caballas 10 San Juan 5 16° S 82° W 80° W 78° W 76° W Valores ecointegrados (m²/mn²) 1000 500 10° S Punta Bermejo Supe Huacho 0.1 84° W 6° S 8° S Trujillo Salaverry Punta Chao Casma Punta Lobos Huarmey 16° S Crucero MPH 0508-09 Chérrepe Pacasmayo Chicama 8° S Trujillo Salaverry Valores ecointegrados (m²/mn²) 14° S 4° S Mórrope Pimentel Punta Bermejo Supe Huacho 12° S 74° W DISTRIBUCION DE CABALLA Parachique Pta. La Negra 6° S Chérrepe Pacasmayo Chicama 10° S 76° W Paita Pta. Gobernador Mórrope Pimentel 8° S 78° W Pto. Pizarro Zorritos Pta. Sal Cabo Blanco Talara DISTRIBUCION DE JUREL Paita Pta. Gobernador 6° S 96,894 la costa (Figura 7). Verticalmente se detectó hasta los 26 m de profundidad, mayormente en horas diurnas. La biomasa estimada fue de 273.629 t, con mayor abundancia entre 15 y 14°S (Tabla 1). 16° S Chancay 750 Callao 12° S Pucusana 500 Cerro Azul 250 Tambo de Mora Pisco 100 14° S Bahía Independencia 50 Punta In ernillos Punta Caballas 10 San Juan 5 16° S 0.1 84° W 74° W Figura 7.- Distribución del jurel. Crucero MPH 0508-09. 1000 82° W 80° W 78° W 76° W 74° W Figura 8.- Distribución de la Caballa. Crucero MPH 0508-09. 148 Estimación hidroacústica de especies pelágicas en invierno 2005 Castillo, Peraltilla y Gutiérrez 84° W 82° W 80° W 78° W Pto. Pizarro Zorritos Pta. Sal Cabo Blanco Talara 4° S 76° W 74° W 84° W 4° S 82° W 4° S DISTRIBUCION DE SAMASA Paita Pta. Gobernador 78° W 6° S 4° S Crucero MPH 0508-09 8° S Chérrepe Pacasmayo Chicama 8° S Trujillo Salaverry 8° S Punta Chao Chimbote Chimbote 10° S 10° S Casma Punta Lobos Huarmey 10° S 1000 Chancay 750 Callao 12° S 12° S Pucusana 500 Cerro Azul 250 Tambo de Mora Pisco 100 14° S Bahía Independencia 50 14° S Punta In ernillos Punta Caballas 10 San Juan 5 Chala 0.1 Atico 82° W 80° W 78° W 76° W 16° S 74° W cuyo 98,54% (95.484 t) se registró en 6°S. Según distancia de la costa, el 75,86% de la biomasa se halló entre 10 y 20 mn de la costa, y el resto, más cerca. El bagre se encontró en diversas áreas aisladas y cercanas a la costa: frente a 80° W 78° W Pto. Pizarro Zorritos Pta. Sal Cabo Blanco Talara 4° S Chancay 750 Callao 76° W Tambo de Mora Pisco 100 Punta In ernillos Punta Caballas 10 San Juan 5 0.1 84° W 82° W 80° W 74° W 84° W 4° S 82° W Presentó varias zonas de distribución, todas costeras, en las cuales su densidad se relacionó con la intromisión de ASS hacia la costa. Una área densa se encontró entre Tambo de Mora y Bahía Independencia cerca de la costa; entre Chérrepe y Casma fue abundante 80° W 84° W 74° W 4° S DISTRIBUCION DE POTA Crucero MPH 0508-09 Parachique Pta. La Negra 6° S 6° S Chérrepe Pacasmayo Chicama 8° S 8° S 8° S Trujillo Salaverry Punta Chao Punta Chao Chimbote Chimbote 10° S Casma Punta Lobos Huarmey 10° S Callao 12° S 12° S Pucusana Cerro Azul 250 Tambo de Mora Pisco 100 14° S 14° S Bahía Independencia 50 Punta In ernillos Punta Caballas 10 San Juan 5 Chala 0.1 Atico 78° W 76° W 16° S Valores ecointegrados (m²/mn²) Chancay 500 10° S Punta Bermejo Supe Huacho 750 80° W 76° W Mórrope Pimentel 1000 82° W 78° W Pto. Pizarro Zorritos Pta. Sal Cabo Blanco Talara 4° S 6° S Casma Punta Lobos Huarmey 16° S 74° W Paita Pta. Gobernador Crucero MPH 0508-09 Trujillo Salaverry Valores ecointegrados (m²/mn²) 14° S 76° W DistribuciÓn y biomasa de la mÚnida (Pleuroncodes monodon) Punta Bermejo Supe Huacho 12° S 78° W Figura 10.- Distribución del bagre. Crucero MPH 0508-09. Chérrepe Pacasmayo Chicama 10° S 16° S Atico Mórrope Pimentel 8° S 14° S Bahía Independencia 50 Paita Pta. Gobernador 6° S Cerro Azul 250 DISTRIBUCION DE MUNIDA Parachique Pta. La Negra 12° S Pucusana 500 Mórrope, entre Pacasmayo y Salaverry, frente a Chimbote y en pequeños núcleos entre Huacho y el Callao. Más frecuente fue frente a Chicama, entre 26 y 48 mn de la costa (Figura 10). Verticalmente se presentó hasta los 65 m de profundidad y mayormente cerca del fondo en áreas muy someras. La biomasa estimada para el bagre fue de 28.458 t (9.876 t en los 8°S, y 7761 t en los7°S) (Tabla 1). DistribuciÓn y biomasa del bagre ₍Galeichthys peruvianus₎ 82° W 1000 Chala 16° S Figura 9.- Distribución de la samasa. Crucero MPH 0508-09. 84° W 10° S Punta Bermejo Supe Huacho Valores ecointegrados (m²/mn²) Valores ecointegrados (m²/mn²) Punta Bermejo Supe Huacho 84° W 8° S Trujillo Salaverry Punta Chao Casma Punta Lobos Huarmey 16° S 6° S Mórrope Pimentel Chérrepe Pacasmayo Chicama 14° S 74° W DISTRIBUCION DE BAGRE Parachique Pta. La Negra 6° S Mórrope Pimentel 12° S 76° W Paita Pta. Gobernador Crucero MPH 0508-09 Parachique Pta. La Negra 6° S 80° W Pto. Pizarro Zorritos Pta. Sal Cabo Blanco Talara Callao 12° S Pucusana 500 Cerro Azul 250 Tambo de Mora Pisco 100 14° S Bahía Independencia 50 Punta In ernillos Punta Caballas 10 San Juan 5 0.1 84° W 74° W Chancay 750 Chala 16° S Figura 11.- Distribución de la múnida. Crucero MPH 0508-09. 1000 Atico 82° W 80° W 78° W 76° W 16° S 74° W Figura 12.- Distribución de la pota. Crucero MPH 0508-09. 149 Informe IMARPE Vol. 35(2): Abril-Junio 2008 84° W 82° W 80° W 78° W 76° W 84° W 74° W 4° S 82° W 4° S 4° S Crucero MPH 0508-09 Parachique Pta. La Negra 6° S 8° S 8° S Chimbote Casma Punta Lobos Huarmey 84° W Chimbote Casma Punta Lobos Huarmey Punta Bermejo Supe Huacho Chancay 3000 Callao 2000 12° S 12° S Pucusana 1000 Cerro Azul 750 Tambo de Mora Pisco 500 250 14° S 14° S Bahía Independencia 100 Punta In ernillos Punta Caballas 50 10 San Juan 5 Chala 0.1 Atico 80° W 78° W 10° S Punta Bermejo Supe Huacho 4000 82° W Punta Chao 10° S 76° W 16° S 16° S 74° W DistribuciÓn y biomasa de la pota ₍Dosidicus gigas₎ La distribución de la pota ha sido muy dispersa; generalmente se halló en áreas de acercamiento de las ASS hacia la costa. Entre Chérrepe y Huarmey fue esporádica y en núcleos pequeños. Otras áreas se han ubicado por fuera de las 30 mn entre Chancay y Cerro Azul, así como por fuera de las 70 mn de Bahía Independencia (Figura 12). Verticalmente alcanzó hasta los 460 m. La biomasa se determinó en 346.862 t, ± 23,89 %. La mayor abundancia se localizó en los 12°S (107.825 t) y en 13°S (47.199 t) (Tabla 1). 1000 Chancay 750 Callao 12° S Pucusana 500 Cerro Azul 250 Tambo de Mora Pisco 100 14° S Bahía Independencia 50 Punta In ernillos Punta Caballas 10 San Juan 5 Chala 0.1 84° W Figura 13.- Distribución de la vinciguerria. Crucero MPH 0508-09. por fuera de las 5 mn de la costa. Al realizar una sobreposición de la distribución de anchoveta con la múnida, se observó que las zonas más densas de ambas especies no coinciden (Figura 11). Verticalmente la distribución llegó hasta los 45 m de profundidad. La biomasa se determinó en 965.474 t, con un límite de confianza de 16,28%; los valores más altos correspondieron a 7°S (324.836 t) y 8°S (215.521 t) (Tabla 1). Valores ecointegrados (m²/mn²) Valores ecointegrados (m²/mn²) 16° S 10° S 8° S Trujillo Salaverry Merluza juvenil (14-20 cm de longitud total) en concentraciones densas a 40 mn frente a Chérrepe Punta Chao 14° S 6° S Chérrepe Pacasmayo Chicama Trujillo Salaverry 5000 Crucero MPH 0508-09 Mórrope Pimentel Chérrepe Pacasmayo Chicama 12° S 74° W 4° S Parachique Pta. La Negra 6° S Mórrope Pimentel 10° S 76° W DISTRIBUCION DE RECUSOS DEMERSALES Paita Pta. Gobernador Paita Pta. Gobernador 8° S 78° W Pto. Pizarro Zorritos Pta. Sal Cabo Blanco Talara DISTRIBUCION DE VINCIGUERRIA 6° S 80° W 200 Brz 100 Brz Pto. Pizarro Zorritos Pta. Sal Cabo Blanco Talara Atico 82° W 80° W 78° W 76° W 16° S 74° W Figura 14.- Distribución de recursos demersales. Crucero MPH 0508-09. DistribuciÓn de la vinciguerria (Vinciguerria lucetia) La vinciguerria fue registrada desde Salaverry hasta Punta Caballas en diversas áreas a partir de las 28 mn de la costa hacia afuera, zona en la cual hubo un acercamiento hacia la costa de las ASS. Los núcleos más densos se ubicaron entre Huarmey y Punta Bermejo, Callao a Pucusana y Cerro Azul a Pisco. Verticalmente se observó hasta profundidades de 400 m durante el día, y en horas nocturnas, permaneció por lo general debajo de los 15 m (Figura 13). DistribuciÓn de recursos demersales Se realizó adicionalmente una detección de recursos demersales, de los cuales se capturó en un lance 100% de merluza juvenil en un área densa ubicada frente a Chérrepe (Figura 14). Otras especies acompañantes se encontraron también entre Talara y Punta La Negra, registradas principalmente en la isobata de las 100 brazas (182 metros). 150 DISCUSIÓN A fines del invierno 2005, la distribución de la anchoveta ha sido amplia en la zona norte, característica propia de esta especie durante el invierno. En los dos años anteriores, 2004 y 2003, la mayor concentración se presentó en el área frente a Mórrope a Huarmey, donde se registró también en los cruceros de verano. Así mismo, la anchoveta continúa siendo la especie de mayor abundancia en la zona costera, y su distribución depende de las condiciones oceanográficas (temperatura, salinidad y disponibilidad de alimento (Figura 15). Este comportamiento latitudinal registrado en invierno del 2003 a 2005, al norte de los 10°S presentó la mayor frecuencia de los registros y de los cardúmenes de anchoveta. En los histogramas se muestran algunas características principales de los cardúmenes que fueron más abundantes entre los grados 10 a 7°S. En relación a la energía acústica, los cardúmenes del 2003 y 2005 presentaron el mismo rango, pero en el 2004 tuvieron un ligero incremento (Figura 16). Estimación hidroacústica de especies pelágicas en invierno 2005 Castillo, Peraltilla y Gutiérrez 2004 4° S 2005 Pto. Pizarro Zorritos Pta. Sal Cabo Blanco Talara 4° S Paita Pta. Gobernador Parachique Pta. La Negra 6° S 6° S 35.12 ups Mórrope Pimentel 17.5 ° C Chérrepe Pacasmayo Chicama 8° S 8° S Trujillo Salaverry Punta Chao Chimbote Casma Punta Lobos Huarmey 10° S 10° S 12° S 14° S 16° S Valores ecointegrados (m²/mn²) Punta Bermejo Supe Huacho 1000 2003 Chancay 12° S Callao 750 Pucusana 500 Cerro Azul 250 Tambo de Mora Pisco 100 14° S Bahía Independencia 50 Punta In ernillos Punta Caballas 10 San Juan 5 Chala Atico 0.1 16° S Figura 15.- Distribución de la anchoveta durante los cruceros MPH 0308-09, 0408-09 y 0508-09 Con respecto al tope medio de los cardúmenes, el rango fue de la superficie hasta 30 m. En 2003 y 2004 los cardúmenes se registraron muy superficiales, y en 2005 hubo ligero incremento de estabilidad hasta 20 m. Con respecto al periodo horario se observó que entre las 15:00 h y las 02:00 h del día siguiente, existe un incremento que se debería al rompimiento de las estructuras de cardúmenes y formación de estratos durante las horas diurnas (Figura 17). Los resultados observados durante este crucero en las áreas exploradas, y algunas características conocidas de los cardúmenes de anchoveta, revelan que existen patrones de comportamiento estacional en este recurso. Esta situación podría deberse a que durante los últimos años, durante el invierno, al norte de los 10°S se produce una franja de anomalías negativas de temperatura, de salinidad y de menor contenido de oxígeno disuelto, que caracteriza un intenso proceso de surgencia desde la costa hasta un amplio sector oceánico, formando una lengua de agua fría hacia el oeste, que propicia condiciones ambientales favorables para la anchoveta. La distribución y abundancia de la múnida, es de especial interés, porque en muchas ocasiones comparte el área con la anchoveta, y de alguna manera su abundancia influye en la distribución de los cardúmenes de anchoveta. Este hecho se muestra con la sobreposición de áreas de distribución de estas especies en diversos cruceros, observándose una relación de espacio-abundancia, es decir en las áreas libres de distribución de anchoveta cerca de la costa, se encuentran las mayores concentraciones de múnida. El jurel y la caballa se mostraron durante esta estación, aunque en núcleos reducidos. Otros recursos costeros como el bagre y la samasa tuvieron presencia reducida. CONCLUSIONES • La anchoveta se presentó ampliamente distribuida, llegando hasta las 150 mn de la costa, las mayores capturas se efectuaron en el 8°S (70%) y dentro de las 20 mn con el 68%. • La anchoveta continúa siendo la especie dominante en el ecosistema costero. Su distribución fue amplia frente a Pacasmayo y Sa151 laverry, área en la cual se registraron las mayores concentraciones. La biomasa de anchoveta estimada por el método hidroacústico en áreas isoparalitorales de 10x30´ fue 7.661.212 t, con mayores valores en los grados 6, 8 y 7°S. Otras especies que tuvieron una abundancia significativa fueron la múnida (965.474 t), y la pota (433.401 t). • El jurel y la caballa se detectaron en núcleos reducidos y dispersos. El jurel se encontró principalmente en el extremo sur (Punta Infiernillos y San Juan), y la caballa fue observada por fuera de las 30 mn de la costa entre Mórrope y Huarmey. • La múnida presentó varias zonas de distribución costera, con mayor abundancia entre Chérrepe y Casma. • Otras especies pelágicas, como la samasa y bagre se encontraron en áreas aisladas de la zona norte. • La pota y la vinciguerria se encontraron dispersas, su distribución está subestimada debido a que estas especies se encontrarían en áreas más alejadas de la costa. 500 1000 1500 2003 0 0 400 800 Informe IMARPE Vol. 35(2): Abril-Junio 2008 -14 -12 -10 Latitud Sur -8 1 2 3 4 5 500 1000 1500 2004 0 0 500 10001500 Valores integrados -14 -12 -10 Latitud Sur -8 -6 -4 0 2 Valores integrados 4 6 1000 2005 0 0 500 1000 1500 -16 -14 -12 -10 Latitud Sur -8 -6 0 1 2 3 Valores integrados 4 5 6 0 0 200 400 600 400 800 1200 Figura 16.- Histogramas de las caracteristicas de los cardúmnes de anchoveta inviernos 2003, 2004 y 2005: Relación latitudinal y rango de valores integrados. 20 Tope medio 30 40 0 5 10 15 20 15 20 15 20 Hora 0 0 400 800 1200 1000 2000 3000 10 10 20 30 Tope medio 40 50 60 0 5 10 Hora 0 0 400 400 800 800 0 10 20 30 40 0 5 Tope medio 10 Hora Figura 17.- Histogramas de las caracteristicas de los cardúmnes de anchoveta inviernos 2003, 2004 y 2005: Relación de tope medio y comportamiento horario. REFERENCIAS Bazigos G. 1976. Applied Fishery Statistics. FAO Fish. Tech. Pap. 135: 164 pp. Foote KG. 1987. Fish target strength for use in echointegrator surveys. Journal of The Acoustical Society of America, 82:981-987 Foote KG, Knudsen HP, Vestnes G, MacLennan DN, Simmonds EJ. 1987. Calibration of acoustic instruments for fish density estimation: a practical guide. Cooperative Research Report, International Council for the Exploration of the Sea, 144, 57 pp. Gutiérrez M, Peraltilla S. 1999. Aplicación de un Sistema de Información Geográfica y de la carta electrónica isoparalitoral en las evaluaciones hidroacústicas de la biomasa de recursos pesqueros en el mar peruano. En Inf. Inst. Mar Perú 146: 25-29 Gutiérrez M. 2002. Determinación de la Fuerza de Blanco (TS) de las principales especies del mar peruano. Tesis. Universidad Nacional Federico Villarreal. 81 pp. Higgimbottom I, Pauly T, Heatley D. 2000. Virtual echograms for visualization processing of multiple-frequency echosounder data. Proceedings of the Fifth European Conference on Underwater Acoustics, ECUA 2000 (Ed. M.E. Zakharia), 1497-1502. Jolly D, Hampton I. 1990. A stratified random transect design for acoustic surveys of fish. Can. J. Fish. Aquat. Sci. 47: 1282-1291. Imarpe. 2005. Informe Ejecutivo del Crucero de Biomasa Desovante 152 de Anchoveta 0508-09. Informe interno. Imarpe. 2005. Informes de Campo del Crucero de Biomasa Desovante de Anchoveta 0508-09. Informes internos. MacLennan D, Fernández P. 2000. Definitions, Units and Symbols in Fisheries Acoustics. FASTICES Working Group Meeting. St. Johns´Newfoundland, Canada, April 1999 (Mimeo). 6pp. MacLennan DN. 1998. Revisión de los resultados obtenidos durante el Crucero 9803-05 de Evaluación Hidroacústica de Recursos Pelágicos. Inf. Inst. Mar Perú 135: 191-201. MacLennan D, Simmonds J. 1992, Fisheries Acoustics. Chapman and Hall, London. Fitoplancton en el invierno 2005 Delgado y Chang FITOPLANCTON DEL MAR PERUANO A FINES DEL INVIERNO 2005 PERUVIAN SEA PHYTOPLANKTON DURING LATE WINTER 2005 Elcira Delgado1 Flor Chang1 RESUMEN Delgado E, Chang F. 2008. Fitoplancton del mar peruano a fines del invierno 2005. Inf. Inst. Mar Perú. 35(2): 153-157.- La biomasa planctónica, a 10 mn de la costa, presentó un promedio general de 0,71 mL/ m3, con rango entre 0,05 mL/m3 (frente a Salaverry) y 8,92 mL/m3 (frente al sur de Punta Gobernador). El fitoplancton de red fue abundante y predominante en el 52%; se distribuyó principalmente en tres áreas: (1) entre Pisco y Chancay desde la franja costera hasta las 100 mn, (2) entre Huacho y Pimentel hasta la 60 mn y (3) entre norte de Pimentel y Punta Gobernador dentro de las 30 mn. Las diatomeas de afloramiento y las neríticas fueron las predominantes (Skeletonema costatum, Thalassiosira angulata, Actinocyclus sp., Chaetoceros lorenzianus, Ch. didymus, Coscinodiscus perforatus, Thalassiosira subtilis, T. rotula y Ditylum brightwellii). Los dinoflagelados representativos en la franja nerítica fueron Protoperidinium depressum, P. crassipes, Ceratium azoricum, Gonyaulax polygramma y Prorocentrum micans. Protoperidinium obtusum, indicador de ACF, estuvo ampliamente distribuido desde Punta Infiernillos hasta Paita dentro de las 60 mn, con distancias máximas frente a Punta Falsa (90 mn), Callao (90 mn) y Punta Infiernillos (120 mn). Ceratium praelongum indicador de aguas subtropicales superficiales (ASS), sólo fue encontrado en Chimbote aproximadamente a 70 mn de la costa. Palabras clave: Fitoplancton, composición y distribución, indicadores biológicos, invierno 2005. ABSTRACT Delgado E, Chang F. 2008. Peruvian marine phytoplankton in late winter 2005. Inf Inst Mar Peru. 35 (2): 153-157.- The plankton biomass 10 nm off the Peruvian coast, had an overall average of 0.71 mL/m3, with a range between 0.05 mL/m3 (off Salaverry) and 8 92 mL/m3 (off south of Punta Governor). The phytoplankton net was abundant and 52% dominant, was distributed in three main areas: (1) 100 nm off Pisco and Chancay; (2) 60 nm off Pimentel and Huacho; and (3) 30 nm north Pimentel to Punta Gobernador. Upwelling neritic diatoms had the highest prevalence (Skeletonema costatum, Thalassiosira angulata, Actinocyclus sp. Chaetoceros lorenzianus, Ch. didymus, Coscinodiscus perforatus, Thalassiosira subtilis, and T. patella, Ditylum brightwellii). The most representative dinoflagellates in the neritic area were Protoperidinium depressum, P. crassipes, Ceratium azoricum, Gonyaulax polygramma and Prorocentrum micans. Protoperidinium obtusum indicator of CCW, was widely distributed from Punta Infiernillo (until 120 nm offshore) to Paita (until 90 nm offshore. Ceratium praelongum indicator of superficial subtropical waters (SSW), was found only in Chimbote approximately 70 nm offshore. Keywords: Phytoplankton, composition and distribution, biological indicators, winter 2005. INTRODUCCIÓN En el invierno 2002, Villanueva y Sánchez (2003) registraron el plancton con un volumen promedio de 0,46 mL/m3, con sólo 10% de fitoplancton, lo que indicaba escasa disponibilidad de alimento para la anchoveta. En el invierno 2003, Chang y Delgado (2004) registraron 0,66 mL/m3, con 31% de fitoplancton, con predominio de diatomeas centrales, lo que aumentó disponibilidad de alimento para las especies planctófagas. En el invierno 2004 (Sánchez y Jacobo 2004), los promedios llegaron a 0,69 mL/m3 y 54%, respectivamente. El fitoplancton se distribuyó dentro de las 30 mn, ampliando su distri- 1 bución por fuera de las 90 mn frente a Paita, Salaverry y Chimbote. aproximadamente 170 mn frente a la costa (Figura 1). En este trabajo se comunican los resultados sobre la distribución de los volúmenes de plancton, composición específica del fitoplancton de red y finalmente la distribución de los indicadores de masas de agua durante el invierno del 2005. Se colectaron 96 muestras, con una red estándar, en arrastres horizontales de 5 minutos y a velocidad constante de 3 nudos. El volumen del plancton se determinó centrifugando cada muestra a 2400 rpm durante 5 min y aplicando la siguiente relación: MATERIAL Y MÉTODOS V = Vc/K Donde: V = volumen de plancton El crucero de estimación de la biomasa desovante de anchoveta BIC Olaya, SNP-2 e Imarpe VI 0508-09 se realizó del 15 de agosto al 24 de setiembre 2005. El área de estudio comprendió desde Punta Infiernillos a Talara, hasta Vc = volumen de plancton centrifugado (mL) K = 6,6273 m3, constante que indica el agua filtrada por la red. Área de Fitoplancton y Producción Primaria [email protected], [email protected] 153 Informe IMARPE Vol. 35(2): Abril-Junio 2008 84°W 82°W 80°W 78°W 76°W 74°W 72°W Pto. Pizarro 4°S 4°S Pta. Sal Talara 774 762 767771 777 Paita C13Pta. Gobernador C11 740 Pta. Falsa 737 734 C10 C9 730 C8 724 C7C6 C5 Pimentel C1 C4 C32 C34 C33 Chicama C31 C 29 C30 C27 C26 Salaverry C23 534/C28 C 24 C20/C21 Punta Chao 474/C 25 C22 389 Chimbote 396 C18 393 6°S 8°S 6°S 8°S Casma C 17 366 C-22 C-21 362 C20 Huarmey C16C15 C 17 141/C18 131/C16 C 14 Supe C13 C 15 Huacho 118 C 12 113 102-C9 C12 99 C10 Chancay 180 Callao C11 C9 183 187 191 Pucusana 95 C4 198 91 C8 77Cerro Azul 136 73 69 59 130 54 50 39Pisco 118 34/C3 30C3 Pta. Mendieta 76 20 Punta Infiernillos 70 41 Punta Caballas 63 3 8 9 San Juan 34/C1 17 Chala 25 Atico 29 10°S 12°S 14°S 16°S 84°W 12°S 14°S 18°S 80°W 78°W 76°W 74°W 72°W Figura 1.- Carta de posiciones. Crucero de estimación de la biomasa desovante de anchoveta BIC Olaya, SNP-2 e Imarpe VI 0508-09. 84°W 82°W 80°W 78°W 76°W 74°W 72°W Pto. Pizarro 4°S 4°S Pta. Sal Talara 0.5 0.5 Paita Pta. Gobernador 8.0 0.5 Pta. Falsa 0.3 6°S 6°S 1 Pimentel 5 0. Chicama 8°S 8°S Salaverry 0.5 Punta Chao Chimbote Casma 1 0.3 2 0.3 1 0.3 Supe Huacho 0.3 1 12°S 1.5 2 1 Chancay 1 Cerro Azul 0.3 18°S 0.3 0. 5 1 16°S 12°S Callao Pucusana 0.3 0.5 14°S 10°S Huarmey 5 1. 10°S Pisco 14°S Pta. Mendieta Punta Infiernillos Punta Caballas San Juan Chala 16°S Atico Quilca Matarani FITOPLANCTON ZOOPLANCTON FITO Y ZOOPLANCTON 84°W 82°W 80°W Ilo 18°S 78°W 76°W 74°W Para la determinación taxonómica se ha utilizado los trabajos de Hustedt (1930), Cupp (1943), Hendey (1964), Licea et al. (1966 a y b), Sournia (1967), Schiller (1971), SundstrÖm (1986), Balech (1988), Heimdal (1993), ThrÖndsen (1993), Hasle y Syvertsen ₍1996), Steidinger y Tangen (1996). RESULTADOS Ilo 82°W Las muestras fueron preservadas con formalina neutralizada (ThrÖndsen 1978). 16°S Quilca Matarani OLAYA SNP - 2 IMARPE VI 18°S 10°S Los resultados se expresan en mililitros de plancton por metro cúbico de agua de mar filtrada (mL/ m3). Las muestras fueron analizadas semi-cuantitativamente, y se ha considerando los componentes del plancton (fitoplancton y zooplancton), según metodología del IMARPE. 72°W Figura 2.- Distribución de volúmenes de plancton (mL/m3). Crucero de estimación de la biomasa desovante de anchoveta BIC Olaya, SNP-2 e Imarpe VI 0508-09. 154 VolÚmenes de plancton (mL/m3) y composiciÓn horizontal del fitoplancton de red El 86% de los valores fue <1,0. El promedio general fue de 0,71. El rango fue de 0,05 (muestra tomada a 10 mn frente a Salaverry), a 8,92 (muestra tomada a 10 mn al sur de Punta Gobernador). La comunidad de fitoplancton fue hallada con el 52% de predominancia, distribuida principalmente frente a tres áreas: (1) entre Pisco y Chancay, desde la franja costera hasta las 100 mn; (2) entre Huacho y Pimentel hasta la 60 mn; y (3) desde el norte de Pimentel a Punta Gobernador hasta la 30 mn (Figura 2). Al sur de Punta Gobernador (5°S, Estación 11), se ubicó el núcleo con el máximo valor (8,92 mL/m3), donde destacó el fitoplancton, con las diatomeas Chaetoceros debilis, Detonula pumila, Pseudonitzschia pungens. Núcleos con 1,0 mL/m3 se ubicaron en Cerro Azul (13°S, Estación 136) y Punta Chao (9°S, Estación C-18), con abundantes diatomeas de aguas cálidas. En Punta Falsa (6°S, Est. 737), un núcleo de 1,0 mL/m3 tuvo predominancia de zooplancton. Frente al Callao (12°S, Estación 187) Fitoplancton en el invierno 2005 Delgado y Chang se localizó un núcleo con 2,0 mL/ m3 con dominancia de diatomeas neríticas y de afloramiento. 3,00 Entre Pisco (14°S) y Chancay (11°30’S) se determinó la abundancia de diatomeas de afloramiento y neríticas como Skeletonema costatum, Thalassiosira angulata, Actinocyclus sp., Chaetoceros lorenzianus, Ch. didymus, Coscinodiscus perforatus, Thalassiosira subtilis, T. rotula y Ditylum brightwellii, A 100 mn de la costa destacaron diatomeas oceánicas como Chaetoceros concavicornis, Lioloma delicatulum, Thalassiosira partheneia y Planktoniella sol. De los dinoflagelados sobresalieron Protoperidinium depressum, P. crassipes, Gonyaulax polygramma y Prorocentrum micans cerca a la costa. 2,00 Entre Huacho (11°S) y Pimentel (7°S) la predominancia de fitoplancton incluyó a las especies antes mencionadas, y además Thalassiosira mendiolana, Lithodesmium undulatum, Chaetoceros affinis, Ch. debilis, Roperia tesellata y el dinoflagelado Ceratium azoricum. En el área entre el norte de Pimentel (7°S) y Punta Gobernador (5°S) dentro de las 30 mn predominaron las diatomeas C. perforatus, Ch. debilis, Pseudonitzschia pungens y Detonula pumila y el dinoflagelado Ceratium furca. mL/m3 2,50 1,50 1,00 0,50 16°-17° 15°-16° 14°-15° 13°-14° 12°-13° 11°-12° 10°-11° 9°-10° 8°-9° 7°-8° 6°-7° 5°-6° 0,00 Grados latitudinales 0-30 mn 30-60 mn >60 mn Figura 3.- Distribución latitudinal de los volúmenes de plancton (mL/m3). Crucero de estimación de la biomasa desovante de anchoveta BIC Olaya, SNP-2 e Imarpe VI 0508-09. 84°W 82°W 80°W 78°W 76°W 74°W 72°W Pto. Pizarro 4°S 4°S Pta. Sal Talara Paita Pta. Gobernador Pta. Falsa 6°S 6°S Pimentel Chicama 8°S 8°S Salaverry Punta Chao Chimbote Casma 10°S 10°S Huarmey Supe Huacho Chancay 12°S 12°S Callao Pucusana Cerro Azul DistribuciÓn latitudinal de los volÚmenes promedio de plancton (mL/m3) En la franja de la costa hasta las 30 mn, el máximo promedio de la biomasa planctónica (2,49) se registró en la latitud de 5 a 6°S; y el mínimo (0,25) entre 8-9°S. Entre las 30-60 mn el máximo (1,23) estuvo entre 12-13°S y el mínimo (0,32) entre 12-13°S. Por fuera de las 60 mn se registraron valores medios menores a 1,0 mL/m3 (Figura 3). Pisco 14°S 16°S 16°S Atico Quilca Matarani 18°S Ilo Protoperidinium obtusum (ACF) (ASS) Ceratium praelongum 84°W 82°W 18°S 80°W 78°W 76°W 74°W 72°W Figura 4.- Indicadores de masas de agua. Crucero BIC Olaya, SNP-2 e Imarpe VI 0508-09. Destacaron los dinoflagelados (Figura 4): 60 mn, con distancias máximas en el norte (Punta Falsa, 6°S, a 90 mn de la costa), centro (Callao, 12°S, a 100 mn), y sur (Punta Infiernillos, 15°S, a 120 mn). También se observó muy restringido (a 15 mn) frente a Punta Gobernador y entre Punta Falsa y Pimentel. • Protoperidinium obtusum, indicador de ACF, desde Punta Infiernillos hasta Paita dentro de las • Ceratium praelongum indicador de ASS, sólo fue encontrado frente a Chimbote (9°S), aproximada- DistribuciÓn de los indicadores biolÓgicos de masas de agua 14°S Pta. Mendieta Pta. Infiernillos Punta Caballas San Juan Chala 155 mente a 70 mn de la línea costera relacionado a 18,1 °C y 35,2 ups. Especies termófilas como Goniodoma polyedricum, Ceratium gibberum var. dispar, C. geniculatum y C. inflatum, sobresalieron por su frecuencia, distribuidos desde Punta Caballas hasta Salaverry asociados a P. obtusum por fuera de las 40 mn frente a Punta Chao, Chimbote y Huacho; a 30 mn fue localizado frente a Cerro Azul. Informe IMARPE Vol. 35(2): Abril-Junio 2008 DISCUSION Comparando los promedios de la biomasa planctónica de invierno, del 2002 al 2005, se ha observado un incremento desde 0,46 mL/m3 en el 2002 (Villanueva y Sánchez, 2003), a 0,66 en el 2003 (Chang y Delgado, 2003), a 0,69 en el 2004 (Sánchez y Jacobo, 2005) y 0,71 mL/ m3 en el 2005 (0,71). Sin embargo, estos promedios son menores a los considerados normales (>3,0 mL/m3) (>3,0 mL/m3) por Rojas de Mendiola et al. (1985) para mar peruano. La predominancia del fitoplancton, también se incrementó en el periodo mencionado: en el 2002 fue 10%, en el 2003, 31%, y para el 2004 y 2005 los valores fueron >50%. En cuanto a la composición específica del fitoplancton, durante el invierno 2002 estuvo conformado principalmente por organismos termófilos. En el 2003 (Chang y Delgado 2003) y el 2004 (Sánchez y Jacobo 2005), los principales componentes fueron las diatomeas neríticas (al norte del Callao) y las de alta de reproducción (con mayor incidencia al norte de Punta Chao). Características similares fueron encontradas en este crucero 0508-09, pero con una distribución diferente, a pesar de las condiciones ambientales que fueron casi normales al sur de Chimbote, y que al norte fueron frías en relación al patrón mensual (IMARPE 2005), esta composición fitoplanctónica estaría reflejando una buena disponibilidad de alimento óptimo para las especies consumidoras de fitoplancton. En lo referente a la distribución latitudinal, los máximos volúmenes de plancton (>1,0 mL/m3) fueron registrados dentro de las 30 mn, situación observada desde 1999 (Delgado et al. 2000) hasta el 2004 (Sánchez y Jacobo 2005), resultados que coinciden con el valor de 2,49 mL/m3 obtenido en este crucero 0508-09 entre 5-6°S, donde las condiciones fueron frías, con anomalías mayores a -2 °C por fuera de las 50 mn (Imarpe 2005). A distancias mayores de las 30 mn los promedios disminuyeron, coincidiendo con lo obtenido en el crucero 0408-09 (Sánchez y Jacobo 2005). El dinoflagelado Protoperidinium obtusum, indicador de las ACF, presentó una distribución bastante similar a los inviernos 2001 (Sánchez et al. 2002), 2002 (Villanueva y Sánchez 2003), 2003 (Chang y Delgado 2004) y 2004 (Sánchez y Jacobo 2005), localizado dentro de las 48 mn, lo cual difiere con el presente estudio, pues la distribución alcanzó 120 mn frente a Punta Infiernillos, asociado a especies termófilas con temperatura y salinidad de aguas de mezcla (ACF y ASS). Ceratium praelongum, indicador de las aguas subtropicales superficiales (ASS), durante los años 2002, 2003, 2004 fue localizado al norte de los 6°S, área que se caracterizó por la intromisión de ASS, con fuertes proyecciones frente a Punta Falsa (15 mn de la costa). En el presente estudio sólo fue determinado a 70 mn frente a Chimbote; sin embargo, se determinaron organismos termófilos (Ceratium massiliense, C. macroceros y Pyrophacus steinii) desde Huarmey hasta Cerro Azul, asociados a temperatura y salinidad propias de ASS. CONCLUSIONES • La biomasa planctónica tuvo un rango entre 0,05 y 8,92 mL/m3, localizada frente a Salaverry y al sur de Punta Gobernador, ambos valores a 10 mn de la costa, registrándose un promedio general de 0,71 mL/m3. • La comunidad de fitoplancton fue relativamente abundante con el 52% de predominancia. Se distribuyó principalmente en tres áreas: (1) entre Pisco y Chancay (0-100 mn), (2) entre Huacho y Pimentel (0-60 mn) y (3) entre el norte de Pimentel y Punta Gobernador (0-30 mn). La composición fitoplanctónica refleja una buena disponibilidad de alimento óptimo para las especies consumidoras de fitoplancton. • Protoperidinium obtusum, indicador de ACF, estuvo distribuido desde Punta Infiernillos hasta 156 Paita dentro de las 60 mn, con distancias máximas frente a Punta Falsa, Callao y Punta Infiernillos a 90, 100 y 120 mn de la costa, respectivamente. Ceratium praelongum indicador de ASS, sólo fue encontrado en Chimbote aproximadamente a 70 mn de la línea costera. Agradecimientos.- Al personal del Área de Fitoplancton y Producción Primaria por el apoyo brindado en el análisis de las muestras y elaboración de gráficos. REFERENCIAS Balech E. 1988. Los dinoflagelados del Atlántico Sudoccidental. Public. Espec. Inst. Español de Oceanog. España . 310 p. Cupp E. 1943. Marine plankton diatoms of the west coast of North America. Bull. Scripps it. Oceangr. 5: 1-237. Chang F, Delgado E. 2008. Fitoplancton en el invierno 2003. Inf. Inst. Mar Perú 35(2): 103-107 Delgado E, Chang F, Villanueva P, C. Fernández. 2000. Fitoplancton en el Invierno 1999 en un área seleccionada (7°- 9°S). Prospección BIC José Olaya Balandra 9908-09. Inf. Inst. Mar Perú 154:23-41. IMARPE 2005. Informe Ejecutivo del Crucero 0508-09 de evaluación de biomasa desovante de anchoveta. Hasle G y Syvertsen E. 1996. Marine diatoms. In: Tomas C. (ed.). Identifying Marine Diatoms and Dinoflagellates. Academic Press, Inc. San Diego. 1-383pp. Heimdal B. 1993. Modern Coccolithophorids. In Tomas C. (ed.). Marine Phytoplankton a guide to naked flagellates and coccolithophorids, Cap. 3. Academic Press, Inc. San Diego. 147-235 pp. Hendey I. 1964. An introductory account of the smaller algae of British Coastal waters. Part. V. Bacillariophyceae (Diatoms). Her Majesty’s Stationery Office, London: 317 pp. Hustedt F. 1930. Die Kieselalgen Deutschlands, Osterreichs und der Schweiz mit Beucksichtigung der ubrigen Lander Europas sowie der angerzenden Meerresgebiete. En: L. Rabenhorst (ed) Kryptogamenflora von Deutschland, Osterreich und der Schweiz. 1 Teil. Fitoplancton en el invierno 2005 Delgado y Chang Akat. Verlagsges. Leipzig, Reprint Johnson Rep. Goop, New York 1971: 920 pp. Licea S, Moreno L, Santoyo H. 1966a. Diatomeas del Golfo de California. Inst. de Ciencias del Mar y Limnología, México, pp 1-258. Licea S, Moreno L, Santoyo H. 1966b. Dinoflagelados del Golfo de California. Inst. de Ciencias del Mar y Limnología, México, pp 1-258. Rojas de Mendiola B, GÓmez O, Ochoa N. 1985. Efectos del fenómeno El Niño sobre el fitoplancton. En: Arntz, W., A. Landa y J. Tarazona (Eds.). El Niño, su impacto en la fauna marina. Bol. Inst. Mar Perú, Vol. Ext.: 33-40. Sánchez S, Villanueva P,. Fernández C. 2002. Fitoplancton de Invierno durante el Crucero de estimación de biomasa desovante de anchoveta 0108-09. Informe interno del Area de Fitoplancton y Producción Primaria. Sánchez S. Jacobo N. 2008. El fitoplancton durante el invierno 2004. Inf. Inst. Mar Perú 35(2): 119-122. Schiller J. 1971. Dinoflagellatae (Peridinea) in monographischer Behandlung.1 Teil. En: L. Rabenhorst (ed) Kriptogamenflora von Deutschland, Osterreich und der Schweiz., Vol. X Flagellatae, Section III3, Part. I. Reprint by Johnson Repr. Corp. New York, London: 617 pp. Sournia A. 1967. Le genre Ceratium (Peridinien Planctonique) dans le Canal de Mozambique. Contribution a une revision 157 mondiale. Vie et Milieu. 18 (2AA): 375-580 pp. Steidinger K, Tangen K. 1996. Dinoflagellates. In: Tomas C. (ed.). Identifying Marine Diatoms and Dinoflagellates. Academic Press, Inc. San Diego. 387-570pp. SundstrÖm B. 1986. The marine diatom genus Rhizosolenia. A new approach to the taxonomy. Lund, Sweden:196 pp. Throndsen J. 1978. Preservation and storage. En: A. Sournia (Ed.) Phytoplankton manual. UNESCO, París:69-74. Throndsen J. 1993. The Planktonic Marine Flagellates. In Tomas C. (ed.). Marine Phytoplankton a guide to naked flagellates and coccolithophorids, Cap. 2. Academic Press, Inc. San Diego. 7-145 pp. Aspectos biológicos de recursos pelágicos en invierno 2005 Peña, Goicochea y Mori ASPECTOS BIOLÓGICOS DE LOS PRINCIPALES RECURSOS PELÁGICOS PERUANOS A FINALES DEL INVIERNO 2005 BIOLOGICAL ASPECTS OF THE MAJOR PERUVIAN PELAGIC RESOURCES AT LATE WINTER 2005 Cecilia Peña1 Carlos Goicochea2 Julio Mori3 RESUMEN PeÑa C, Goicochea C, Mori J. 2008. Aspectos biológicos de los principales recursos pelágicos peruanos a finales del invierno 2005. Inf. Inst. Mar 35(2):159-166.- El estudio se hizo durante el crucero de biomasa desovante de anchoveta BIC Olaya, SNP2 e Imarpe VI 0508-09, de Punta Infiernillos (15°S) a Paita (5°S) del 15 agosto al 24 setiembre 2005. Se estudió principalmente la anchoveta, múnida, pota y bagre. Se anotó la distribución, concentración, estructura por tamaños, determinación de edad, y el estado reproductivo de anchoveta, especie dominante en el ecosistema pelágico, conformado por individuos provenientes de los desoves del invierno 2004 y verano 2005 con edades de 0,5 y 1,0 años; LT de 7 a 18 cm, moda 12 cm, y 37% de juveniles. Por el método hidroacústico se estimó una biomasa de 7.600.000 t; 52% distribuida en el área de 5 a 7°S, con 44% de juveniles. Entre los 8 a 12°S se encontraron adultos, con modas de 12,0 a 14,0 cm, donde la anchoveta estaba desovando y su biomasa algo reducida llegó al 42% del total. Se observó un marcado gradiente latitudinal N a S en la estructura por tamaños, con una progresión de la talla modal que varía desde los 10,0 cm (5°S) hasta los 14,0 cm (12°S), situación asociada al ingreso de aguas cálidas (ASS) en la zona centro, donde se presentó un núcleo importante de anomalías térmicas positivas. Al norte las condiciones fueron frías, lo que favoreció a los juveniles distribuidos en el área entre 7 y 5°S. Palabras clave: recursos pelágicos, invierno 2005, anchoveta peruana, mar peruano. ABSTRACT PeÑa C, Goicochea C, Mori J. 2008. Biological aspects of the major Peruvian pelagic resources in late winter 2005. Inf Inst Mar 35 (2): 159-166.- The study was conducted during the cruise of the anchovy spawning biomass BIC Olaya, SNP2 and Imarpe VI 0508-09, in the area from Punta Infiernillos (15°S) to Paita (5°S) during 15th August to 24th September 2005. Four species were studied: the Peruvian anchovy, múnida, giant squid and catfish, including their distribution, concentration, structure, size, age determination and reproductive status of anchovy. The anchovy was the dominant species in the pelagic ecosystem, 37% juveniles; composed of individuals from winter 2004 and summer 2005 spawning; the ages were 0,5 and 1,0 years; with LT, 7 to 18 cm; moda 12 cm. The anchovy biomass, estimated with the hydroacoustic method was 7.6 million tons, with 52% distributed into the 5 to 7°S area, where juveniles reached to 44%. The adults were found between 8 to 12°S, with modes of 12 and 14 cm, they were a spawning biomass reduced to 42% of the total. It was noted a marked latitudinal gradient from north to south in the structure size, with a modal length from 10 cm (5°S) to 14 cm (12°S); this was a situation associated with the entry of warm water (ASS); in whose center existed a core of positive thermal anomalies. In the north it was cold, good conditions for juveniles distributed into the area between 7 to 5°S. Keywords: pelagic resources, winter 2005, Peruvian anchovy, Peruvian sea. INTRODUCCIÓN Durante 1990 al 2003 la captura total acumulada de las principales especies pelágicas marinas (anchoveta, sardina, arenque, jurel y caballa) alcanzó 412 millones toneladas, lo que representó el 42% de la captura mundial, y la anchoveta peruana ocupó el 11% (FAO 2005). Debido a los altos rendimientos y gran variabilidad de la anchoveta en la costa peruana, es necesario realizar una permanente evaluación de su abundancia. 1 2 3 En el Perú, desde los años 90, la anchoveta (Engraulis ringens) es la especie dominante en la pesquería pelágica, y el método hidroacústico el más usado para estimar su abundancia. Sin embargo, a partir de 1994, se viene desarrollando el método de evaluación directa de biomasa desovante, mediante el estimado de cinco variables biológicas obtenidas independientemente, a partir de colección de huevos y adultos, según un diseño de muestreo que se aplica durante Unidad de Dinámica Poblacional de Recursos Pelágicos [email protected] Unidad de Edad y Crecimiento [email protected] Unidad de Seguimiento de Recursos Pelágicos [email protected] 159 el periodo de desove (Guzmán 1995), de relevante importancia en el cálculo de las proyecciones de pesca para las temporadas de actividad extractiva. Frente a la costa peruana se consideran dos stocks de la población de anchoveta: (1) el stock norte - centro (4-15°S) que posee las mayores concentraciones, y (2) el stock sur Perú -norte Chile (16-24°S) (Tsukayama 1983). Como este recurso está sometido a la extracción de grandes tonelajes, Informe IMARPE Vol. 35(2): Abril-Junio 2008 es necesario realizar constantes estudios sobre las variaciones en su comportamiento reproductivo, reclutamiento y pesquería. Se conoce que en el stock norte-centro el periodo de mayor intensidad de desove se registra en agosto – setiembre. El presente informe muestra los aspectos biológicos de los principales recursos pelágicos, con énfasis en la anchoveta durante su periodo de desove principal de invierno – primavera 2005. MATERIAL Y MÉTODOS El Crucero de Estimación de la Biomasa Desovante de Anchoveta 0508-09, estudió el área comprendida desde Punta Infiernillos (14°S) hasta Paita (5°S), del 15 de agosto al 24 de setiembre del 2005. Se ejecutaron 70 lances, 34 por el BIC Olaya, 23 por el BIC SNP2 y 13 por la LIC IMARPE VI. Se obtuvieron los siguientes cinco parámetros: peso promedio de hembras, producción diaria de huevos, fecundidad parcial, proporción sexual y fracción de hembras desovantes; y, simultáneamente, se determinó la biomasa total de anchoveta utilizando el método hidroacústico. En cada lance de comprobación positivo se determinó la composición por especies, con la clave Chirichigno y Vélez (1998) para peces y Okutani (1980) para cefalópodos. Además, se realizaron muestreos de las principales especies capturadas para conocer la estructura por tamaño, peso individual por grupos de longitud y sexo. La madurez gonadal de pelágicos pequeños (anchoveta), se determinó mediante la escala de 6 estadios de Einersson et al. (1966) y para pelágicos mayores (jurel y caballa), con la escala de 8 estadios de Johanssen (1924). Para el Índice Gonadosomático (IGS) se consideraron solo hembras de anchoveta a partir de 12 cm de longitud total. Se seleccionaron al azar y por grupo de talla, cada dos grados de latitud, 55 individuos de anchoveta, para realizar las lecturas de otolitos, utilizando un estereoscopio Nikon SMZ 800 con 10 X 2 aumentos. Tabla 1.- Captura (kg) por especies según embarcaciones durante el Crucero 0508-09, de evaluación de biomasa desovante de la anchoveta. Captura (kg) por embarcación Especie Olaya 5 547,4 508,1 0,0 119,6 29,4 6 204,5 Anchoveta Múnida Samasa Pota Otros Total SNP-2 215,6 146,0 0,0 26,8 6.3 394,7 Los otolitos fueron colocados en un otolitero negro con alcohol. La iluminación de las muestras se hizo con luz reflejada en forma oblicua en plano horizontal; para clasificar el material hialino y opaco que se deposita en el borde del otolito se aplicó el mismo criterio de Dannevig (1933) citado por Aguayo (1976). La periodicidad de formación de los anillos (2 formaciones hialinas por año) se determinó de acuerdo a los resultados de Chirinos y Chuman (1968) para la zona norte – centro, y de Chuman (1969) para la zona sur, ambos citados por Aguayo (1976). La diferencia en estructura de edades se determinó mediante el test de chi cuadrado. Imarpe VI 158,3 0,0 283,8 0,0 20,8 462,8 Total (kg) (%) 5 878,0 654,1 283,8 146,3 56.5 7 018,7 83,7 9,3 4,0 2,1 0,8 99,9 respectivo valor de biomasa, obteniéndose las estructuras por tamaños ponderadas a la biomasa, así como valores ponderados de moda y porcentaje de juveniles. RESULTADOS ComposiciÓn por especies Se efectuaron 70 lances de comprobación, con una captura total de 7.062 kg. Las principales especies fueron: anchoveta, Engraulis ringens con 84%; múnida, Pleuroncodes monodon con 9%; samasa, Anchoa nasus con 4% y pota, Dosidicus gigas con 2%. Otras especies importantes fueron: bagre, Galeichthys peruvianus; esperlán plateado, Leuroglossus urotranus; camotillo, Normanichthys crockeri; cachema, Cynoscion anales; caballa, Scomber japonicus (Tabla 1). Con los resultados de biomasa por área isoparalitoral (áreas tomadas cada medio grado latitudinal y cada 10 millas paralelas de distancia a la costa) obtenidos por el método hidroacústico, se procedió a ponderar la estructura por tamaño por medio grado latitudinal al La anchoveta tuvo una distribución amplia al norte del Callao. En el grado 8°S llegó a 100 mn de la costa; fue la latitud donde se lograron las Tabla 2.- Captura (kg) de anchoveta por área isoparalitoral durante el Crucero 0508-09. Latitud sur (°S) 0 5 3 0 6 3 0 7 3 0 8 3 0 9 3 0 10 3 0 11 3 0 12 3 0 13 3 0 14 3 Captura (k) Porcentaje (%) 10 20 20 43 31 8 19 15 30 Distancia a la costa (mn) 50 60 70 8 120 38 37 8 80 40 217 4 5 43 15 23 9 6 43 97 44 3560 60.6 200 21 160 100 18 70 1.2 52 0.9 347 5.9 45 350 350 147 100 1 32 41 60 370 6.3 946 16.1 4 386 6.6 90 14 243 3093 66 95 20 5 80 0 0.0 147 2.5 Captura Porcentaje (kg) (%) 63 1.1 0 0.0 50 0.9 37 0.6 8 0.1 408 6.9 3481 59.2 642 10.9 135 2.3 277 4.7 133 2.3 43 0.7 224 3.8 0 0.0 121 2.1 70 1.2 0 0.0 43 0.7 97 1.6 44 0.7 5878 100.0 100.0 Aspectos biológicos de recursos pelágicos en invierno 2005 Peña, Goicochea y Mori 20 20 Juveniles: 37% a) Porcentaje (%) 15 10 10 5 Las capturas de múnida se efectuaron de 7 a 14°S dentro de las 40 millas de distancia a la costa; pero la más abundante fue entre 7 y 9°S. La pota se capturó principalmente en los 12 a 14°S, entre 20 y 70 mn de la costa. La mayor captura de samasa (257 kg) ocurrió en los 6°S a 11 millas de la costa. Estructura por tamaÑos Anchoveta, Engraulis ringens El rango de la LT fue de 7 a 18 cm, con moda en 12 cm. La LT de juveniles (<12 cm) ponderada a la biomasa fue de 37%, que en peso representó un 26% (Figura 1). Latitudinalmente, el mayor porcentaje de adultos se localizó de 8°30’ a 13°S con moda en 13,5 cm; y el mayor porcentaje de juveniles se presentó de 5 a 7°S y de 13°30’ a 14°30’S. En las latitudes 12°30’ a 13°30°S (Callao a Tambo de Mora) se observó una moda secundaria en 15 cm (Figura 2). Espacialmente, entre 7°30’ a 13°S se observó una amplia distribución de adultos y los ejemplares juveniles, se registraron de 5°30’S a 6°30’S dentro de las 40 mn. En el 14°S, se registraron juveniles (<12 cm) dentro de las 10 mn (Figura 2). 5 0 0 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 Longitud total (cm) Longitud total (cm) Figura 1.- Estructura por tamaños de anchoveta ponderada a la biomasa durante el Crucero 0508-09. a) En número y b) En peso. Lat. sur 05°00'S JUV 69% 05°30'S 70% 06°00'S 77% 06°30'S 52% 07°00'S 65% 07°30'S 5% 08°00'S 16% 08°30'S 6% 09°00'S PAITA 5°S CRUCERO BIOMASA DESOVANTE 0508-09 PARACHIQUE 1% 10°00'S 2% 10°30'S 0% 11°00'S 1% 11°30'S 0% 12°00'S 0% 12°30'S 0% 13°00'S 6% 13°30'S 16% 14°00'S 43% 14°30'S 70% 5 6 DISTRIBUCION ESPACIAL DE LAS CALAS POSITIVAS PARA ANCHOVETA 6°S SANTA ROSA 7°S Juveniles CHICAMA 8°S 0 - 10% 10 - 50% SALAVERRY Mayores de 50% COIHSCO SAMANCO 9°S 0% 09°30'S CULEBRAS 10°S SUPE CARQUIN 11°S CHANCAY CALLAO 12°S PUCUSANA CERRO AZUL 13°S T. DE MORA PISCO 14°S Ba. INDEPE INFIERNILL 7 8 9 10 11 12 13 14 15 16 17 18 19 20 Longitud total (cm) Figura 2.- Estructura por tamaños de anchoveta ponderada a la biomasa y distribución espacial de los tamaños de anchoveta. Crucero 0508-09. Distancia a la costa Juveniles 3% 100 mn Según distancia a la costa, los juveniles se hallaron dentro de las 20 mn. A partir de las 30 mn, se registraron principalmente adultos (Figura 3). Excepcionalmente, a las 40 mn se observó una estructura con moda en 12.0, ésta corresponde a un muestreo efectuado en la zona frente a Santa Rosa (6°S). Pota, Dosidicus gigas El rango de la longitud del manto fluctuó de 170 a 780 mm, con moda principal en 230 mm (Figura 4). Juveniles: 26% b) 15 Porcentaje (%) mayores capturas (70,2% del total). De 13-14°S se localizó dentro de las 10 mn cercanas a la costa. De acuerdo a la distancia a la costa, el 68% de la captura total se realizó dentro de las 20 millas (Tabla 2). 70 mn 1% 60 mn 3% 50 mn 9% 40 mn 45 % 30 mn 1% 20 mn 29 % 10 mn 5 6 7 28 % 8 9 10 11 12 13 14 15 16 17 18 19 20 Longitud total (cm) Figura 3.- Estructura por tamaños de anchoveta ponderada a la biomasa según distancia a la costa durante el Cr.0508-09. 161 Informe IMARPE Vol. 35(2): Abril-Junio 2008 25 N = 145 individuos Porcentaje (%) Porcentaje (%) 20 15 10 5 18 16 14 12 10 8 6 4 2 0 N = 638 individuos 5 0 80 130 180 230 280 330 380 430 480 530 580 630 680 730 7 9 11 13 780 15 17 19 21 23 25 27 29 Longitud del cefalotorax (mm) Longitud del manto (mm) Figura 5.- Estructura por tamaños de munida durante el Cr.0508-09. Figura 4.- Estructura por tamaños de pota. Cruicero 0508-09. 16 100 N = 498 individuos 80 4 60 40 20 0 0.5 0 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 Longitud total (cm) Figura 6.- Estructura por tamaños de samasa. Crucero 0508-09. 5° - 6° 80 Porcentaje (%) 1 1.5 2 2.5 60 40 60 ÁREA A 50 5º - 6º LS 40 30 20 10 Edad (Años) 0 Figura 8.- Estructura por edades de la anchoveta de toda el área prospectada. Crucero 0508-09 Múnida (Pleurocondes monodon) 100 70 CR. 0508-09 El rango de longitud del cefalotórax fue de 10 a 27 mm, con moda principal en 18 mm y secundarias en 14 y 16 mm (Figura 5). 80 70 Porcentaje (%) 8 80 Porcentaje (%) Porcentaje (%) Porcentaje (%) 12 60 ÁREA B 50 7º - 14º LS 40 30 20 10 0 0.5 1 1.5 2 Edad (Años) Figura 9.- Estructura por edades de la anchoveta de las áreas A y B. Crucero.0508-09 20 Samasa (Anchoa nasus) 0 100 7° - 9° Porcentaje (%) 80 60 40 DeterminaciÓn de edad en anchoveta 20 0 100 10° - 12° Porcentaje (%) 80 60 40 20 0 100 13° - 14° 80 Porcentaje (%) El rango de la LT fue de 5.0 a 15.5 cm, con moda principal en 13 cm y secundarias en 8,5 y 6 cm (Figura 6). 60 40 20 0 0.5 1 1.5 2 Edad (años) Figura 7.- Estructura por edades de anchoveta según áreas. Crucero 0508-09. De la lectura de 616 pares de otolitos obtenidos a partir de los muestreos biológicos de anchoveta se elaboraron claves talla-edad con las cuales se pudo determinar la estructura por edades de la especie en cuatro áreas delimitadas según la similitud de la estructura por tallas. Se encontró que en el área de 5 - 6°S estuvo representada por individuos de 0,5 (54,5%) y 1 año (43,9%). En el área de 7° - 9° la fracción más importante correspondió a los individuos de 1 año (91,4%) al igual que el área 10° - 12° donde también dominó el grupo de 1 año (74,8%) con un aumento de los ejemplares de 1,5 años con 2,4%. En el área de 13 14°S, predominaron los individuos de 1 (47,5%) y 0,5 años (38%). 162 Estos resultados indican que dicha población proviene de los desoves del invierno 2004 y verano 2005, con una marcada ausencia de ejemplares mayores de 1,5 años de edad (Figura 7). De la estructura por edades obtenidas durante este crucero 0508-09 se determinó que el grupo predominante fue el de un año de edad (86%), y siguieron individuos de 0,5 años (8,3%) y de 1,5 años (5,7%) (Figura 8). Igualmente, se observó una segregación latitudinal de las edades, con la presencia de dos áreas bien definidas. Los resultados obtenidos de la comparación de las estructuras por edad de las áreas A y B descritas anteriormente, mediante la prueba del chi-cuadrado, confirman que ellas son significativamente diferentes, pues los ejemplares de 0,5 y 1,0 año de edad estuvieron en los 5-6°S, mientras que los adultos >1,0 año se presentaron en los 7-14°S (Figura 9). Aspectos biológicos de recursos pelágicos en invierno 2005 Peña, Goicochea y Mori DeterminaciÓn macroscÓpica De un total de 710 ejemplares hembras, se observó que: 1.- En las áreas situadas entre 4-6°S, el mayor porcentaje de individuos se encontraban en los estadios II y III, con 34% en su mayoría ejemplares juveniles y jóvenes maduros provenientes del desove de verano del 2005 y del desove de invierno-primavera 2004. 2.- En el área entre 7°-14°S, se presentó un mayor porcentaje de individuos en el estadio V, con 61% de adultos provenientes del desove invierno-primavera del 2004 (Figura 10). El índice gonadosomático (IGS) fue de 6,82, el factor de condición (FC) fue 0,6589 y el 50,6% se encontró desovando (estadio V). Latitudinalmente, se observó que los mayores valores de IGS se presentaron entre 7 y 12°S, y los menores entre 13 y 14°S. Es necesario mencionar que no se obtuvo datos entre los 5 y 6°S debido a que no se pudo registrar valores de pesos individuales (Figura 11). El desove de anchoveta (estadio V), determinado por la observación macroscópica de la madurez gonadal de anchoveta, presentó un comportamiento similar al IGS (Figura 12). DISCUSIÓN En los Cruceros de Evaluación de Biomasa Desovante efectuadas durante los años 2004 y 2005, se han registrado estimaciones similares de biomasa de anchoveta, pota y múnida obtenidas por el método acústico (Tabla 3). Sin embargo, en el 2005 se registró una estimación de 177.000 t de caballa (zona norte) y 274.000 t de jurel (zona sur) (Gutiérrez et al. 2008), asociada a un mayor acercamiento a la costa de las aguas de mezcla, que dio lugar a una mayor disponibilidad de estos recursos. En el presente crucero, 050809, se encontró una distribución 40 70 35 60 30 50 25 40 20 30 15 20 10 10 5 0 0 II III IV V VI II III IV V VI 7º- 14º S 4º- 6º S Figura 10.- Frecuencia porcentual de los estadios de madurez gonadal de anchoveta. Crucero 0508-09 10 8 I.G.S. Madurez gonadal de anchoveta 6 4 2 0 5° 6° 7° 8° 9° 10° 11° 12° 13° 14° Grado Latitudinal (°S) Figura 11.- IGS de anchoveta según grados latitudinales. Crucero 0508-09. Indice Gonadosomático Madurez Gonadal 50.0 8 .0 7 .0 40.0 6 .0 5 .0 30.0 4 .0 20.0 3 .0 2 .0 10.0 1 .0 0 .0 4º -6º 7 º - 9º 1 0º -1 2 º 13 º -1 4 º 0.0 4º -6º 7º -9º 10º -12º 13º -14º Figura 12.- IGS y madurez gonadal de anchoveta (estadio V) según grados latitudinales. Crucero.0508-09. Tabla 3.- Biomasa (millones de toneladas) por especie pelágica. Invierno 2004 y 2005 2004 2005 Anchoveta 7,49 7,66 Jurel SR 0,3 Caballa SR 0,2 Pota 0,3 0,4 Múnida 1,2 1,0 * SR: Sin registro acústico para la zona explorada. irregular de anchoveta de norte a sur, lo cual se vio reflejado en la estructura por tamaños según medio grado de latitud. En la Figura 13 se presenta una secuencia de gráficos relacionando ambiente, distribución y tamaños de anchoveta durante el invierno y su comparación con el 2004. En la Figura 13, para el año 163 2005 se puede observar: (1) en la estructura por tamaños, una marcada gradiente latitudinal de tamaños de anchoveta, con mayor presencia de juveniles en el 5°S y de adultos en los 12°30’S, con una progresión de la talla modal que varía desde los 10,0 cm (5°S) hasta los 14,0 cm (12°S); (2) que la mayor concentración (mayor biomasa) Año 2004 Año 2005 164 16°S 14°S 12°S 10°S 8°S 6°S 4°S 84 82°W -2 -2 -1 -1 0 1 1 2 2 82°W 83 -2 -1.5 -1 -0.5 0 0.5 1 1.5 2 83 80°W 81 78°W 79 80°W 76°W 77 4°S 80 80°W 81 80 78°W 79 76 76°W 74°W h v÷ 16°S 12° S 10° S 8° S 6° S 76°W 74°W 84° W h 16° S v÷ 14° S 0.5 5 10 50 0.1 5 10 50 100 250 500 750 1000 100 250 500 750 1000 82° W 82° W 82° W 82° W Valores ecointegrados 80° W Pto. Pizarro Zorritos 80° W 76° W 76° W 76° W 74° W 78° W 76° W Chala 74° W San Juan Punta Infiernillos Punta Caballas Bahía Independencia Tambo de Mora Pisco Cerro Azul Pucusana Callao Chancay Punta Bermejo Supe Huacho Casma Punta Lobos Huarmey Chimbote Punta Chao Trujillo Salaverry 15° 14° S 14° 13° 12° S 12° 11° 0 16° S 15° 14° 13° 14° S 12° 12° S 11° 10° 10° S 09° 8° S 08° 07° 6° S 06° 04° 4° S 05° 74° W Atico Distribución de la anchoveta 78° W 78° W San Juan Punta Caballas Punta Infiernillos 0 10° S 10° 09° 8° S 08° 07° 6° S 06° 04° 05° 4° S 74° W Bahía Indepen dencia Tambo de Mora Pisco Cerro Azul Pucusana Callao Chancay Huacho Supe Punta Be rmejo Huarmey Punta Lobos Casma Chimbote Punta Chao Trujillo Salaverry Chérrepe Pacasmayo Chicama Chérrepe Pacasmayo Chicama Mórrope Pimentel Parachique Pta. La Ne gra Paita Pta. Gobernador Pto. Pizarro Zorritos Pta. Sal Cabo Blanco Talara 80° W 80° W 78° W Distribución de la anchoveta Pimentel Mórrope Parachique Pta. La Negra Paita Pta. Gobernador Talara Pta. Sal Cabo Blanco 1000 1500 1000 1500 BIOMASA (Miles de t) 500 BIOMASA (Miles de t) 500 5 7 8 9 10 11 12 13 14 15 16 17 31% 6 14°30'S 14°00'S 13°30'S 8 9 1% 6 14°30'S 7 Longitud total (cm) 10 11 12 13 14 15 16 17 18 19 20 70% 16% 43% 13°30'S 6% 0% 0% 0% 14°00'S 13°00'S 12°30'S 12°00'S 11°30'S 11°00'S 2% 0% 10°00'S 10°30'S 0% 1% 09°30'S 6% 09°00'S 08°30'S 5% 16% 08°00'S 07°30'S 65% 52% 07°00'S 77% 20 06°30'S 70% 69% 43% Juveniles 18 19 0% 6% 3% 3% 9% 06°00'S 05°30'S 05°00'S 04°30'S Longitud total (cm) 1% 12°30' S 13°00'S 0% 23% 12°00'S 11°30'S 36% 39% 11°00'S 10°30'S 10°00'S 09°30'S Latitud sur 5 2% 26% 7% 09°00'S 08°30'S 10 % 2% 08°00'S 07°30'S 07°00'S 9% 16% 06°30'S 4% 1% Juvenile s 1% 06°00'S 05°30'S 05°00'S 04°30'S Latitud sur (°S) Figura 13.- Secuencia ambiente, distribución, biomasa y tallas de anchoveta durante los inviernos 2004 y 2005. 78°W 84° W 4° TES 74°W 75 84° W 14° S 12° S San Nicolás San Juan wGp wGj zGuç 35.0 zGq j wGi z o wGz 35.0 j hù j wGo 8° S 6° S Punta Caballas 35.0 Punta Infiernillos 84° W 74 4° S 10° S Bahía Independencia Pisco Peninsula Paracas wGp wGj zGuç zGq j 80°W 76°W 77 75 Bujama Cerro Azul Punta Cóndor Tambo de Mora SALINIDAD (ups) 78 76 Callao Punta Hermosa Chancay Ancón Huacho Punta Salinas Supe Punta Bermejo Huarmey 77 SSM Punta Lobos Casma Chimbote Punta Chao Trujillo Salaverry Guañape Chicama Chérrepe Pacasmayo 78 tù w jï w j { z n÷ wGj j j wGs o w wUGm Gw wUGn 79 Pimentel Mórrope Parachique Pta. Falsa Pta. Gobernador Paita wUGz jGi G{ 82 Pta. Sal Talara Cabo Blanco 81 i jGh¡ wGjù {GGt w GwGw ióGp 82°W 35.0 34.9 82°W 83 36.0 35.7 35.4 35.1 34.8 34.5 34.2 33.9 33.6 33.3 33.0 32.7 32.4 32.1 31.8 31.5 31.2 30.9 30.6 30.3 30.0 82 i jGh¡ wGjù {GGt w GwGw 14°S ióGp 12°S 10°S 8°S 6°S 2005 74°W 75 74 84 15 San Nicolás San Juan Punta Caballas Punta Infiernillos 76 wGi z o wGz j hù j wGo 78°W 13 13 12 12 11 11 10 10 9 9 8 8 7 7 6 6 5 5 74 84 83 4 4 14 Bahía Independencia14 ANOMALIA TERMICA (ºC) 78 75 Bujama Cerro Azul Punta Cóndor Tambo de Mora Pisco Peninsula Paracas Callao Punta Hermosa Chancay Ancón Huacho Punta Salinas Supe 76 TSM 77 Punta Bermejo Huarmey Punta Lobos Casma Chimbote Punta Chao Trujillo Salaverry Guañape Chicama 78 tù w jï w j { z n÷ wGj j j wGs o w wUGm Gw wUGn 80 79 Chérrepe Pacasmayo Pimentel Mórrope Parachique Pta. Falsa Paita Pta. Gobernador Talara Cabo Blanco 81Pta. Sal 80 wUGz jGi G{ 82 82 Valores ecointegrados (m²/mn²) 84 Informe IMARPE Vol. 35(2): Abril-Junio 2008 Aspectos biológicos de recursos pelágicos en invierno 2005 Peña, Goicochea y Mori 10 8 60 I.G. 40 20 0 Porcentaje (%) 100 15 14.5 14 13.5 13 12.5 12 11.5 11 10.5 10 Talla media IGS CR.0408-09 80 6 4 2 80 CR. 0508-09 60 0 40 5° 6° 7° 8° 20 9° 10° 11° 12° 13° Talla media (cm) Porcentaje (%) 100 14° Grado Latitudinal (°S) 0 0.5 1 1.5 2 2.5 Figura 15.- Comparación latitudinal entre los valores de IGS y talla media de anchoveta. Invierno 2005. Edad (Años) Figura 14.- Estructuras poblacionales por edad de anchoveta a finales de los inviernos 2004 y 2005. 10 2004 y mayor área de distribución de anchoveta se presenta entre los 6 y 8°S. Esta situación estuvo asociada al ingreso de aguas cálidas, ASS, en la zona centro, del área entre Bahía Independencia a Huarmey, donde se presentó un núcleo importante de anomalías térmicas positivas; pero al norte de Huarmey, las condiciones fueron frías, principalmente por fuera de las 50 mn frente a Paita donde las anomalías negativas fueron mayores a -2°C (Vásquez 2008) lo que originó condiciones favorables para los ejemplares juveniles distribuidos de 5 al 7°S. Durante el 2004, se observó una clara diferencia en cuanto a la distribución de las tallas, respecto a lo descrito en el 2005, destacando que la progresión en la talla modal se observó de manera discontinua en 3 ó 4 zonas del área evaluada. Esto quiere decir que hubo efectos ambientales y biológicos que alteraron el patrón de continuidad en las tallas modales. En cuanto a los factores ambientales, podemos mencionar que las Aguas Ecuatoriales Superficiales (AES) con índices termohalinos >18°C y < 34,8 ups, se encontraron desplazadas más al sur de su ubicación normal llegando hasta Punta Falsa (Vásquez com. pers.) lo cual ocasionó que una baja biomasa, compuesta por solo ejemplares adultos, estuviera presente en esta zona, mientras que, principalmente, de Chimbote a Callao se presentó un ingreso de Aguas Subtropicales Superficiales (ASS) con salinidades >35,1 ups, temperaturas >17°C y anomalías térmicas po- I.G.S. 8 2005 6 4 2 0 5° 6° 7° 8° 9° 10° 11° 12° 13° 14° Grado Latitudinal (°S) Figura 16.- Comparación de los valores de IGS latitudinal durante 2004 y 2005. sitivas (de 0°C a +0.5°C) en Paita, Chicama y frente a Chancay ocasionando que los procesos de afloramiento, propios de las (ACF), se encontraron dentro de las 30 mn, con lo cual ejemplares juveniles estuvieron presentes dentro de esta área. Por otro lado, al comparar la estructura poblacional de la anchoveta por edad, durante los cruceros 0408-09 y 0508-09 (Figura 14), se puede observar que en el invierno 2004 estuvo compuesta por 5 grupos de edad (0,5 a 2,5 años de edad), y fue más estable que la del 2005, sustentada por solo 3 grupos de edad (0,5 a 1,5 años de edad) constituida por adultos jóvenes. La diferencia significativa entre los grupos de edad de la anchoveta, registrada en las áreas A y B (Figura 9), puede explicarse por la influencia de aguas frías y de baja salinidad (ACF), encontradas atípicamente para la fecha en el área A, lugar donde se presentó el mayor porcentaje de jóvenes, tal como lo explica el informe oceanográfico de Vásquez (2008). En la Figura 15 se observa que la principal área de desove ocurrió en la región sur, donde se 165 presentaron las mayores modas de la LT de anchoveta (cm), lo cual significaría que este desove correspondíó, en su mayoría, a ejemplares de 12 a 14 cm, lo cual concuerda con lo registrado por Perea y Roque (2008), quienes mencionan que individuos <14,0 cm son los que habían alcanzado los mayores valores de fracción desovante (Figura 14). En una comparación del IGS de los años 2004 y 2005, se halló que fue mayor en el 2005 (Figura 16). Los máximos valores del IGS durante el 2005 se registraron entre los 11 y 12°S, y entre los 8 y 9°S coincidiendo con valores mayores al 20% de la fracción desovante de anchoveta localizados en los 12°S y de los 10° a 8°S (Perea y Roque 2008). Por otro lado, en el 2004 los valores de IGS por grado de latitud son bajos en toda el área explorada, coincidiendo con valores menores al 20% (valor crítico) de la fracción desovante estimada. CONCLUSIONES • La anchoveta fue la especie dominante en el ecosistema pelágico, con el 84% de la captura total durante el crucero de biomasa desovante 0508-09. Informe IMARPE Vol. 35(2): Abril-Junio 2008 En la zona centro, se efectuaron capturas de jurel y caballa en áreas más distantes a la costa, situación no observada en años anteriores. • La distribución de la anchoveta fue amplia, llegando hasta las 100 mn de la costa, aunque las mayores capturas se efectuaron dentro de las 20 mn (68% de total), y latitudinalmente en los 8°S (70% del total). • El rango de la LT de anchoveta fue de 7 a 18 cm, con una moda ponderada a la biomasa de 12 cm y un porcentaje de juveniles de 37%, que en peso representó un 26%. • La población de anchoveta estudiada en este crucero de finales del invierno 2005, proviene de los desoves del invierno del 2004 y verano del 2005 con edades de 1 y 0,5 años. • La anchoveta se encontró desovando desde los 8 hasta los 12°S, y durante la última semana se detectó el inicio del proceso de declinación de este periodo, desde 5 a 7°S. REFERENCIAS Aguayo M. 1976. Edad y crecimiento de la anchoveta (Engraulis ringens Jenys) del Norte de Chile (Arica-Iquique). Instituto de Fomento Pesquero Chile. Boletín cientifico.55 pp. Chirichigno N, Vélez J. 1998. Clave para identificar los peces marinos del Perú. (2da. Edición). Publicación Especial. Inst. Mar. Perú. 500 pp. Einersson H, Flores L,. MiÑano J. 1966. El ciclo de madurez de la anchoveta peruana (Engraulis ringens J.) En: Seminario Latinoamericano sobre el Pacífico Oriental. Universidad Nacional 166 Mayor de San Marcos, Lima: Perú: 128-135. FAO. 2005. FISHSTAT Plus: Universal software for fishery statical time series. Version 2.3. FAO Fisheries Department, Fishery Information, Data and Statistics Unit. Guzmán S, AyÓn P, Pizarro L. 1995. Biomasa Desovante de la anchoveta y condiciones oceanográficas Crucero 9408-10 BIC SNP-1 y bolicheras. Inf. Inst. Mar Perú 115. Johanssen A. 1924. On the Summer and Autumn spawning herring on the north sea. Medd. Forumm. Ha Vunderg Serv. Fisheri, Bd. Vi 5. Okutani T. 1980. Useful and latent cuttlefish and squids of the world. In: National Cooperative Association of Squid processors for the 15th anniversary of its foundation. Tokio, Japan.. Tsukayama I. 1983. Recursos pelágicos y sus pesquerías en Perú. Rev. Com.Perm. Pacífico Sur 13: 25-63. Parámetros reproductivos de anchoveta en invierno 2005 Perea y Peña PARÁMETROS REPRODUCTIVOS DE ADULTOS DE ANCHOVETA PERUANA A FINALES DEL INVIERNO 2005 REPRODUCTIVE PARAMETERS OF ADULT PERUVIAN ANCHOVY AT THE END OF WINTER 2005 Ángel Perea de la Matta1 Cecilia Roque García2 RESUMEN Perea A, Roque C. 2008.- Parámetros reproductivos de adultos de anchoveta peruana a finales del invierno 2005. Inf. Inst. Mar Perú 35(2): 167-171.- Se presentan los resultados del estudio sobre valores de la frecuencia de desove (0,088), fecundidad parcial (7,699), proporción sexual (0,51) y peso promedio de hembras (15,2 g) de la anchoveta Engraulis ringens. Estos parámetros fueron utilizados para la estimación de la biomasa desovante de anchoveta por el método de producción de huevos (MPH), llevada a cabo durante el crucero BIC Olaya, SNP2 y LIC Imarpe VI, entre el 15 de agosto y el 24 de septiembre del 2005. Se discute la fluctuación de estos parámetros y el estado de madurez gonadal en el momento de la evaluación. Palabras clave: Reproducción, anchoveta peruana, desove, fecundidad, madurez gonadal, peso de hembras. ABSTRACT Perea A, Roque C. 2008.- Reproductive parameters of adult Peruvian anchovy at the end of winter 2005. Inf Inst Mar Peru 35 (2): 167-171.- We present the results of the obtained values on the frequency of spawning (0.088), partial fertility (7.699), sex ratio (0.51) and average females weight (15.2 g) of the anchovy Engraulis ringens. These parameters were used to estimate the spawning biomass of anchovy by the method of egg production (MEP), carried out during the cruise BIC Olaya, SNP2 IMARPE and LIC and VI 0508-09, from August 15th to September 24th of 2005. We discuss the fluctuation of these parameters and the status of gonadal maturity at the time of evaluation. Keywords: Reproduction, Peruvian anchoveta, spawning, fecundity, gonad maturity, of females weight. INTRODUCCIÓN El Método de Producción de Huevos (MPH) usado para la estimación de la biomasa desovante de anchoveta, ha sido diseñado en base al reconocimiento de los productos del desove y de la condición reproductiva de los adultos, con lo que se retrocalcula el tamaño del stock parental (BuchananWollaston 1926). Además de la producción de huevos y larvas, se requiere un conocimiento de la biología desovante y fecundidad de los individuos adultos (Lasker 1984), pues, a excepción de la proporción sexual, son altamente variables los parámetros de adultos como son el peso promedio de hembras, frecuencia de desove y sobre todo la fecundidad; más aún, en una especie como la anchoveta, cuya población está caracterizada por vivir y responder rápidamente a los cambios en el ambiente (Hunter y Macewicz 1985). 1 2 La frecuencia de desove indica la periodicidad y la fecundidad, el volumen de los productos en cada evento reproductivo. Estos parámetros, sumados al tamaño del stock parental, determinan la producción de huevos en un momento dado. En especies como la anchoveta, esta producción de huevos toma gran interés debido a su relación con el reclutamiento, pues a ella se le atribuye más importancia que a la que existe entre el stock y el reclutamiento, debido a que este último suele ser muy débil e incierto (Claramunt et al. 1994). Durante los últimos años se ha venido utilizando con éxito el MPH para estimar la biomasa desovante de diferentes especies con reproducción parcial (engraúlidos y clupéidos) (Armstrong et al. 1988, Clarke 1992, Palomera y Pertierra 1993, Bentley et al. 1996, Sánchez et al. 1996). Laboratorio de Biología Reproductiva 4-297630 anexo 250 [email protected] Laboratorio de Biología Reproductiva 4-297630 anexo 250 [email protected] 167 El presente trabajo se efectuó en ocasión del Crucero de Evaluación de la Biomasa Desovante de Anchoveta, 0508-09, realizado entre el 15 de agosto y el 24 de setiembre del 2005. Estudia el estado reproductivo de anchoveta, distribución de cardúmenes desovantes, análisis de la estructura de tallas, además de un anexo donde se realiza el cálculo de tallas de primera madurez y tallas de primer desove. Se discute así mismo el momento reproductivo y sus implicancias. MATERIAL Y MÉTODOS Estado reproductivo de anchoveta según el análisis microscópico El método de colecta, el análisis de los resultados y el tratamiento estadístico fue efectuado de acuerdo a la metodología descrita por Santander et al. (1984). Las gónadas se procesaron usando la Informe IMARPE Vol. 35(2): Abril-Junio 2008 Zonas de desove 60 Frecuencia relativa (%) 50 Fracción de hembras desovantes (= cuyos ovarios presentan folículos post-ovulatorios y ovocitos hidratados) /número de hembras adultas. Valorcrítico crÍtico Valor indicador de indicador de desove desove 40 30 20 10 0 12 °S 11 °S 10 °S 9 °S 8 °S 7 °S 6 °S 5 °S Grados latitudinales Fraccion desovante ponderada Fraccion atresica ponderada Figura 1.- Fracción desovante y atrésica de anchoveta Engraulis ringens por grados latitudinales. Crucero de Biomasa Desovante 0508-09 4 Pta. Sal Cabo Blanco Talara 5 Paita Pta. Gobernador Distribucion de cardúmenes desovantes de anchoveta Parachique Pta. Falsa 6 Crucero de Biomasa desovante 0508-09 Mórrope Se estimó la proporción sexual por peso, y la proporción sexual en número por horas del día. Para estimar la fecundidad parcial y relativa se utilizaron 273 hembras en condición de hidratadas, las cuales no presentaron signos de haber desovado. Se utilizó la metodología de Hunter y Goldberg (1980) y Hunter et al. (1985), utilizando una balanza digital Sartorius BP 3100 S y una balanza analítica And HR-200. Pimentel 7 Chérrepe Pacasmayo Chicama 8 Trujillo Salaverry Guañape Punta Chao 9 Chimbote Casma Punta Lobos 10 Huarmey 12 13 14 15 84 Fracción de hembras desovantes Punta Bermejo 11 Supe > 50 Huacho Punta Salinas Chancay Ancón 45 Callao Punta Hermosa 35 Bujama Cerro Azul Punta Cóndor Tambo de Mora Pisco Peninsula Paracas 20 15 RESULTADOS Por grados latitudinales Bahía Independencia 5 Punta Infiernillos Punta Caballas San Nicolás San Juan 1 83 82 81 80 79 78 77 76 75 74 Figura 2.- Distribución de cardúmenes desovantes de anchoveta. Crucero de Biomasa Desovante 0508-09 técnica de infiltración en parafina (Humason 1979), un micrótomo de rotación American Optical Co., y cortes por congelación en un criostato (Leica 1997). Las láminas se tiñeron con un coloreador automático Leica Autostainer XL y se analizaron con un microscopio compuesto Nikon Eclipse E 400. Se cuantificó la frecuencia de cada uno de los estadios de madurez gonadal por lance. Estos estadios fueron identificados según la escala de BuitrÓn et al. (1997), basada en el desarrollo ovocitario y la terminología usada para la anchoveta del norte Engraulis mordax (Hunter et al. 1980). Para conocer la distribución espacial de los cardúmenes desovantes se calculó la fracción de hembras desovantes por cada lance de acuerdo a la relación que se anota a continuación (Hunter y Macewicz (1985b), la que luego se ponderó a la distribución de tallas de la captura: 168 En un muestreo aleatorio y estratificado por tallas (LT de 10,5 a 17,5 cm) se colectaron 997 ovarios de anchoveta adulta en toda el área prospectada. (5 a 14°S) provenientes de 45 lances positivos. Los ovarios fueron procesados y coloreados para su análisis microscópico. En base al desarrollo ovocitario se calculó la fracción desovante y porcentaje de atresia por cada lance, grado latitudinal y grupo de talla. Adicionalmente, utilizando los datos de los análisis microscópicos, se estimaron las tallas de primera madurez y del primer desove, por grupos de tallas de la población. En los resultados obtenidos sobre la fracción desovante y atrésica por grados latitudinales ponderados a la captura (Figura 1) destacaron los 10°S (35,2%) y 9°S (35,1%) como las áreas de reproducción más importantes; en el grado 11°S se registró intromisión de ASS. La fracción atrésica tuvo valores cercanos a cero en todas estas zonas, lo que demostró que la anchoveta se encontraba dentro de un periodo importante de desove. Desde los 7° hasta los 5°S los valores fueron bajos e inferiores al valor crítico (20%) indicador de los periodos de desove. En el 6°S se registró el valor de atresia más alto (18,2%). Mediante el estimado de la fracción desovante (Figura 2), se pue- Parámetros reproductivos de anchoveta en invierno 2005 Perea y Peña En las zonas más sureñas, desde los 14°S, hasta el Callao los valores de fracción desovante han sido bastante pobres. Probablemente, debido a una fuerte intromisión de ASS hasta zonas muy cercanas a la costa estando los núcleos de cardúmenes desovantes limitados por salinidades superiores a 35,1 ups; sin embargo, en el resto del área explorada las condiciones térmicas han sido de normales a frías Por rango de tallas Los individuos <14 cm alcanzaron los mayores valores, sobre todo en 9 y 10°S, zonas de desove más importantes de toda la evaluación. Esto es inverso a lo normal, pues los individuos >14 cm por lo general tienen mayor actividad que los más pequeños (Figura 3). Fecundidad La fecundidad parcial promedio de la anchoveta durante el invierno-primavera del 2005 fue estimada en 7.699 ovocitos por tanda de desove. La ecuación de regresión que relacionó las variables fecundidad parcial y peso eviscerado fue una de tipo lineal: 705,66 * peso sin gónada – 3012,4 (R2 = 0,6921)(Figura 4). Frecuencia de desove: 0,88, con un coeficiente de variación de 0,188. Proporción sexual: fue estimada en 0,514 (CV =0,039). Peso promedio de hembras: 15,179 g (CV= 0,046). Talla de primera madurez y desove.- Dada la variada estructura de tallas registrada en cada grado latitudinal, se procedió a estimar las tallas de primera madurez por zonas, bajo el criterio de grupos modales similares. Las zonas 70 35000 Menor 14 cm 60 Mayor 14 cm 50 30000 número de ovocitos Fracción desovante (%) de tener una visión panorámica de la distribución espacial de estos cardúmenes desovantes. Los más importantes se encontraron por fuera de las 40 mn frente a Ancón, Huarmey y Punta Chao. Los núcleos más costeros se ubicaron desde Salaverry a Chimbote, Casma a Huarmey, y frente al Callao. 40 30 20 y = 705.66x - 3012.4 R2 = 0.6921 25000 20000 15000 10000 5000 10 0 0 12 11 10 9 8 7 6 5 0 10 20 30 40 50 peso sin gónada (g) Grados latitudinales S Figura 3.- Fracción desovante de anchoveta por grupos de tallas. Crucero de Biomasa Desovante 0508-09 Figura 4.- Relación entre la fecundidad y el peso sin gónada (g) de la anchoveta. Crucero de Biomasa Desovante 0508-09 consideradas de acuerdo a la estructura de tallas se representan en la Figura 5. desovante inferiores a los >14 cm. Durante este Crucero 0508-09 se halló que este comportamiento se había invertido, pues los responsables del evento reproductivo registrado fueron los individuos <14 cm, tanto por su condición reproductiva como por su abundancia. De acuerdo a la estructura de tallas encontrada, es probable que este comportamiento reproductivo se deba a la ausencia de hembras grandes en la población, sobre todo en las áreas de mayor abundancia, donde el stock adulto estuvo representado por un solo grupo modal (12,5 cm). Todos estos aspectos han tenido una implicancia en el inicio de la madurez de anchoveta que, por lo general se da a los 12 cm; sin embargo, cálculos realizados dan cuenta de una maduración temprana registrada en la presente evaluación. DISCUSIÓN Se halló que la anchoveta estuvo desovando entre los 12 a 8°S, confirmado por la casi ausencia de individuos con atresia ovocitaria (en recuperación). Desde 7 a 5°S la intensidad de desove fue inferior al valor crítico. Inclusive, durante los muestreos de rutina se verificó a simple vista que los individuos tuvieron muy poco peso, reflejando un grado de bienestar muy bajo. Bajo estas condiciones, fue muy poco probable que los individuos tuvieran la suficiente energía para madurar y desovar. Debe considerarse la estructura del stock adulto (1 a 1,5 años de edad) y el tiempo de duración de la evaluación, que fue mayor a 4 semanas. Por lo general, los adultos jóvenes tienen un periodo de desove más estrecho que los de mayor talla. Es muy probable que, al final de la evaluación, los individuos localizados desde 7 a 5S estuvieran en el inicio de la declinación de su periodo de desove, cuyo indicador claro fue la presencia de individuos en recuperación, o con atresia ovocitaria, que en los 6S fue de 18%, valor considerado muy alto. Normalmente, los individuos <14 cm registran valores de fracción En todas las zonas consideradas, excepto entre 13 y 14°S, la talla de primera madurez fue inferior a 12 cm. Estos casos de maduración temprana en peces tienen varias razones. Ya se mencionó la ausencia de hembras grandes en la población. En otros casos, una disminución de la población adulta suele producir un stress que estimula la maduración y desove de los individuos más pequeños. Los otolitos estudiados, indican que la población estuvo representada por individuos de 0,5 y 1,0 años de edad, es decir, individuos de corta edad (Tabla 1). Tabla 1.- Biomasa (t) juvenil, adulta y desovante (BD) de anchoveta estimada por el método hidroacústico. Crucero 0508-09 Juveniles Adultos BD 5-6°S 261 125 2 373 081 638 366 169 7-9°S 104 3251298 3 230 907 10-12°S 2 952 1 345 082 1 341 095 13-14°S 231 219 196 350 196 350 TOTAL (t) 495 400 7 165 812 5 406 718 Informe IMARPE Vol. 35(2): Abril-Junio 2008 5 a 6ºS 1.00 1.00 Frecuencia acumulada Talla de primera madurez = 10,5 0.50 Talla de primer desove = 12,5 cm 0.50 0.00 16.5 17 17.5 16.5 17 17.5 16 15.5 15 14.5 14 13.5 13 12.5 12 11.5 11 10.5 10 9.5 9 8.5 8 7.5 7 6.5 6 17.5 17 16.5 16 15.5 15 14.5 14 13.5 13 12.5 12 11.5 11 10.5 10 9.5 9 8.5 8 7.5 7 6.5 6 5.5 5 5.5 5 0.00 Longitud total Longitud total 7 a 9ºS 1.00 1.00 11,0 cm Frecuencia acumulada Frecuencia acumulada 10,0 cm 0.50 0.00 0.50 16 15.5 15 14.5 14 13.5 13 12.5 12 11.5 11 Longitud total 10.5 10 9.5 9 8.5 8 7.5 7 6.5 6 5.5 5 17.5 17 16.5 16 15.5 15 14.5 14 13.5 13 12.5 12 11.5 11 10.5 10 9.5 9 8.5 8 7.5 7 6.5 6 5.5 5 0.00 Longitud total 10 a 12ºS 1.00 1.00 12, 2 cm Frecuencia acumulada Frecuencia acumulada 11,5 cm 0.50 0.50 0.00 17.5 17 16.5 16 15.5 15 14.5 14 13.5 13 12.5 12 11.5 11 Longitud total 10.5 10 9.5 9 8.5 8 7.5 7 6.5 6 5.5 5 17.5 17 16.5 16 15.5 15 14.5 14 13.5 13 12.5 12 11.5 11 10.5 10 9.5 9 8.5 8 7.5 7 6.5 6 5.5 5 0.00 Longitud total 13 a 14ºS 1.00 12,5 cm 0.50 0.00 17.5 17 16.5 16 15.5 15 14.5 14 13.5 13 12.5 12 11.5 11 10.5 10 9.5 9 8.5 8 7.5 7 6.5 6 5.5 5 Longitud Figura 5.- Tallas de primera madurez (cm, izquierda) y primer desove (cm, derecha), determinadas por grados de latitud. Crucero de biomasa desovante de la anchoveta peruana 0508-09. 170 Parámetros reproductivos de anchoveta en invierno 2005 Perea y Peña Estos resultados muestran que la biomasa la anchoveta estuvo conformada, en gran parte por individuos adultos. La biomasa de juveniles apenas llegó a 495.400 t, en una biomasa desovante de 5.406.718 t. CONCLUSIONES 1.- Los parámetros de adultos, utilizados para la estimación de la biomasa desovante de anchoveta Engraulis ringens por el método de producción de huevos (MPH), fueron: frecuencia de desove = 0.088 (CV = 0,0,188); fecundidad parcial = 7,699 (CV = 0,064).); proporción sexual = 0,514 (CV =0,039); peso promedio de hembras = 15,179 g (CV = 0,046). 2.- Las estimaciones de primera madurez y desove, han permitido conocer cual es el tamaño del stock adulto y cual es la biomasa desovante estimada por el método hidroacústico. Se determinó que la biomasa la anchoveta estuvo conformada en su gran parte por individuos adultos. REFERENCIAS Armstrong M, Shelton P, Hampton I, Jolly G, Melo Y. 1988. Egg production estimates of anchovy biomass in the Southern Benguela system. CalCOFI Report 29:137157. Bentley P, Emmett R, Lo N, Moser G. 1996 Egg production of Pacific sardine (Sardinops sagax) off Oregon in 1994. Sardine Symposium 1995. CalCOFI Report 37: 193-200. Buchanan-Wollaston H. 1926. Plaice-egg production in 1920-21, treated as a statistical problem, with comparison between the data from 1911, 1914 and 1921. Fish. Invest. Ser. II, 9(2). 45 pp. BuitrÓn B, Perea A. 1996. Frecuencia de desove de la anchoveta peruana Engraulis ringens en agosto y setiembre 1995. Inf. Inst. Mar Perú 119: 27-33. BuitrÓn B, Perea A, Alvarado S. 1998. Madurez sexual, frecuencia de desove y fecundidad de la anchoveta peruana durante invierno-primavera 1997. Inf Inst Mar Perú 130: 65-69. Claramunt G, Herrera G, Pizarro P. 1994. Producción potencial anual de huevos por tallas en Sardinops sagax (Jenyns, 1842) del norte de Chile. Rev. Biol.. Mar. Valparaíso, 29 (2): 211-233. Clarke T. 1992. Egg abundance and spawning biomass of the hawaiian anchovy or nehu, Encrasicholina purpurea, during 1984-1988 in Kanehoe Bay, Hawaii. Pacific Science 46(3): 325-343. Humason GL. 1979. Animal tissue techniques. Fourth edition. WH Freeman Co. San Francisco. 661 pp. Hunter J, Goldberg S. 1980. Spawning incidence and batch fecundity in northern anchovy, Engraulis mordax. Fish. Bull. U.S. 77: 641-652. Hunter J, Macewicz B. 1985. Rates of 171 atresia in the ovary of captive and wild northern anchovy, Engraulis mordax. Fish. Bull. U.S., 83:119136. Lasker R. 1984. An egg production method for estimating spawning biomass of pelagic fish: Application to the northern anchovy (Engraulis mordax). Administrative Report LJ-84-37 330 pp. Leica Mycrosystems Nussloch GmbH. 1997. Manual de instrucciones Leica CM 1100. Nussloch, Alemania. 34 pp. Palomera I, Pertierra P. 1993. Anchovy biomass estimate by the daily egg production meted in 1990 in western Mediterranean sea. Scientia Marina 57(2-3):243251. Perea A, BuitrÓn B. 1996. Valores críticos en ovarios desovantes y en recuperación de anchoveta (Engraulis ringens) y sardina (Sardinops sagax) en las épocas de desove. Inf. Prog. Inst. Mar Perú 44: 3-17. Sánchez R, Pájaro M, Macchi G. 1996. The application of the daily egg production method to the assesment of the SW Atlantic anchovy (Engraulis anchoita), spawning biomass off Argentina. ICES CM 196/H:29. Santander H, Alheit J, Smith P. 1984. Estimación de la biomasa desovante de anchoveta peruana Engraulis ringens en 1981 por aplicación del Método de producción de Huevos. Bol. Inst. Mar Perú 8 (6): 213-250. Características oceanográficas físicas en invierno 2005 Vásquez CONDICIONES AMBIENTALES FÍSICAS DEL MAR PERUANO A FINES DEL INVIERNO 2005 PERUVIAN MARINE PHYSICAL ENVIRONMENTAL CONDITIONS DURING LATE WINTER 2005 Luis Vásquez Espinoza1 RESUMEN Vásquez L. 2008.- Condiciones ambientales físicas del mar peruano a finales del invierno 2005.- Inf. Inst. Mar Perú 35 (2) 173-180.- El estudio se realizó frente al área de Talara a Punta Infiernillos, del 15 agosto al 24 setiembre 2005, durante el crucero 0508-09. Al norte, entre Talara y Huarmey, las condiciones fueron frías; y hacia el sur, entre Huarmey y Punta Infiernillo fueron normales, a excepción de una pequeña área calida frente a Bahía Independencia. Se encontró una fuerte proyección de las aguas frías hacia el norte y oeste debido a la intensificación de la corriente de Humboldt y a los vientos que favorecieron los procesos de afloramiento, lo que permitió que las Aguas Costeras Frías tuvieran una amplia distribución principalmente al norte del Callao. Al sur, las ACF estuvieron restringidas por la trasgresión de las Aguas Subtropicales Superficiales hacia la costa motivando que los procesos de afloramiento estuvieran restringidos a zonas muy costeras. Por otro lado, se observó una gran zona de mezcla en la zona sur asociada a la presencia de las Aguas Templadas de las Subantártica. Palabras clave: condiciones oceanográficas, invierno 2005, mar peruano. ABSTRACT Vásquez L. 2008 .- Peruvian marine physical environmental conditions in late winter 2005.- Inf Inst Mar Peru 35 (2): 173-180 .- The study was carried out off Talara to Punta Infiernillo, during August 15th to September 24th 2005, during the 0508-09 cruise. To the north, and between Talara to Huarmey, conditions were cold; and south, until Punta Infiernillo were normal, except for a small warm area off Bahía Independencia. There was a strong projection of the cold waters north and west due to the intensification of the Humboldt Current and the winds that helped the process of upwelling, which allowed cold coastal waters (CCW) have a wide distribution mainly north of Callao. To the south, the CCW were restricted by the transgression of the Subtropical Surface Water to the coast causing that upwelling processes were restricted to coastal zones. On the other hand, there was a large mixing zone in the south associated with the presence of the temperate waters of the Subantarctic. Keywords: oceanographic conditions, winter 2005, Peruvian sea. INTRODUCCIÓN El modelo numérico CFS NCEP – NOAA, indicó una distribución espacial de ligeras anomalías positivas confinadas en el sector occidental del Pacífico ecuatorial y negativas (-0,3°C) en el sector central y oriental hasta octubre. El resumen del IRI –International Research Institute- indica el incremento del número de modelos (5) que pronostican condiciones ligeramente frías en el Pacífico central, para el período de agosto a octubre, sin embargo, sigue habiendo una mayoría (12 de 20), que pronostican condiciones ligeramente sobre lo normal. El promedio general da un valor de +0,2°C para el Pacífico central (área Niño 3.4) durante el periodo indicado. 1 La TSM frente a la costa peruana, en la segunda semana de julio, registró el arribo del pulso cálido originado en el Pacifico ecuatorial central que incrementó la temperatura en la costa norte (Informe Técnico julio 2005 – ENFEN), especialmente en las estaciones de Talara y Paita, observándose en la estación de Paita la anomalía positiva más significativa que alcanzó el valor de +3,0° C durante cuatro días; en la costa central, este pulso fue registrado muy débilmente, lo cual no influyó sobre las aguas frías en el resto de la costa. Entre el 2 y 13 de agosto, durante la operación MOPFEN (Pizarro et al. 2005) se encontró un predominio de Aguas Costeras Frías (ACF) asociadas a temperaturas <17 °C Dirección de Oceanografía Física; [email protected] 173 hasta las 30 mn de la costa al frente de Chérrepe a Paita, ubicándose las Aguas Subtropicales Superficiales (ASS) asociadas a temperaturas <18°C por fuera de las 25 mn de Chimbote y Salaverry y 30 mn de la costa frente a Punta La Negra. Bajo este panorama se desarrolló el Crucero 0508-09. Se encontró un ambiente de normalidad al sur de Chimbote, pero al norte las condiciones fueron frías. En este informe se da a conocer los resultados más relevantes de las condiciones físicas durante el presente crucero. MATERIAL Y MÉTODOS Entre el 15 agosto y el 24 de setiembre del 2005, el crucero Informe IMARPE Vol. 35(2): Abril-Junio 2008 82°W 80°W 78°W 76°W 74°W Callao, proyectándose hasta las 30 mn frente a Huarmey. 4°S G{ Gw En general, las isotermas se distribuyeron en forma paralela a la línea de costa, observándose un fuerte contraste térmico (gradiente de 1°C/10 mn) frente a Huarmey, asociado a la proyección de aguas cálidas del oeste. Al sur de Pisco la distribución fue homogénea, pero con las temperaturas más bajas de toda el área evaluada (Figura 2). wUGm 6°S w j 8°S z j 10°S Anomalías térmicas o Han variado entre –2,62 °C (50-60 mn de Paita) y +1,20 °C (Bahía Independencia). Al sur de Huarmey, entre –0,5 °C a +0,5 °C, exceptuando una pequeña área cálida frente a Bahía Independencia. Al norte de Huarmey las condiciones fueron frías, principalmente por fuera de las 50 mn frente a Paita (>-2 °C) (Figura 3). o 12°S j jGh¡ w 14°S BIC OLAYA BIC SNP 2 zGq Salinidad 16°S h 82°W 80°W 78°W 76°W 74°W Figura 1.- Carta de Posiciones. Crucero Biomasa Desovante 0508-09 de Evaluación de la Biomasa Desovante BIC Olaya y SNP2, tuvo una cobertura desde la zona marina frente a Talara hasta Punta Infiernillos, cubriendo un área variable con una amplitud máxima de 200 mn (Figura 1). El estudio de la estructura térmica y halina en las secciones de: Paita, Punta Falsa, Chimbote, Callao y Punta Infiernillos se realizó sobre la base de los lanzamientos de CTD, hasta una profundidad de 500 m y de botellas Niskin a profundidades estándar (hasta 300 m, en el BIC Olaya y hasta 150 m en el BIC SNP 2). Adicionalmente se hicieron lanzamientos de CTD frente a Chicama y Punta Bermejo hasta los 300 m de profundidad. Para el estudio de la capa superficial y subsuperficial del mar se obtuvieron datos de temperatura del aire y velocidad y dirección del viento y muestras de agua para el análisis de salinidad cada 4-5 mn (durante las estaciones de plancton). Con la información de vientos se calculó Índice de Turbulencia (IT= v3, m3/s3) que se obtiene de la intensidad del viento al cubo, el transporte Ekman es calculado según Bakun (1973) y los índices de Afloramiento (IA) usando la ecuación: IA = E /cos (a), m3*s-1*km-1 Donde: E esfuerzo de viento y a ángulo de la línea de costa con la dirección del viento. RESULTADOS Estructura horizontal La SSM varió de 34,870 a 35,338 ups. Las concentraciones <34,9 ups se localizaron frente a Talara, con ligera influencia de AES; las altas concentraciones asociadas a ASS se localizaron en promedio por fuera de las 60 mn de costa, destacando dos fuertes proyecciones hacia la costa, una frente a Huarmey y otra entre Bujama y Bahía Independencia. Las ACF asociadas a procesos de afloramiento, se registraron dentro de las 30 mn, ampliando su distribución al sur de Pisco y al norte de Chicama, hasta 70 mn frente a Punta Falsa. Frente a Bahía Independencia una amplia franja con aguas de mezcla se proyectó hacia el noroeste, asociada a la interacción de las ASS, ACF y a Aguas Templadas de la Subantártica (ATSA), las que contribuyeron a los bajos tenores de salinidad en esta área (Figura 4). Temperatura La TSM tuvo rango de 14,6 a 18,6 °C, promedio 16,5°C. Valores <16°C, asociados al afloramiento costero, ubicados dentro de las 20 mn entre Salaverry y Callao, ampliaron su distribución al norte de Chicama y al sur del Pisco. Temperaturas más altas (>18°C) se localizaron entre Chicama y 174 Vientos La intensidad de los vientos fluctuó entre 1,0 a 12,4 m/s, predominando vientos moderados (4 a 8 m/s) en toda el área evaluada; los vientos intensos (>8 m/s) se localizaron formando núcleos por fuera de las 20 mn entre Pimentel y Punta Infiernillos; los vientos Características oceanográficas físicas en invierno 2005 Vásquez 82°W 4°S 80°W 78°W 76°W 74°W 82°W 4°S wUGz jGi G{ Gw wUGn w wUGm 6°S 8°S 10°S 12°S 14°S 16°S 30 29 28 27 26 25 24 23 22 21 20 19 18 17 16 15 14 13 12 11 10 82°W tù w jï w j { z n÷ wGj j j wGs o 2 10°S 12°S 1 i jGh¡ wGjù {GGt w GwGw ióGp 0 14°S -1 wGp wGj zGuç zGq j -1 -2 16°S h v÷ h v÷ -2 80°W 80°W 34.9 78°W 76°W 74°W 78°W 76°W 74°W 82°W SALINIDAD (ups) w wUGm 78°W 76°W 74°W Figura 3.- Anomalías Térmicas (°C). Cr. Biomasa Desovante 0508-09 82°W 4°S 80°W 78°W 76°W 74°W wUGz jGi G{ 8°S 10°S 12 11 10 wGp wGj zGuç 35.0 zGq j 16°S wGi z o wGz j hù j wGo 13 i jGh¡ wGjù {GGt w GwGw ióGp 14°S 15 tù w jï w j { z n÷ wGj j j wGs o 14 12°S h v÷ VIENTOS w wUGm 6°S wGi z o wGz 35.0 j hù j wGo 12°S 80°W Gw wUGn tù w jï w j { z n÷ wGj j j wGs o 10°S wGi z o wGz j hù j wGo 1 wGp wGj zGuç zGq j wUGz jGi G{ 8°S ANOMALÍA TÉRMICA (ºC) 2 i jGh¡ wGjù {GGt w GwGw ióGp 74°W tù w jï w j { z n÷ wGj j j wGs o 8°S wGi z o wGz j hù j wGo 82°W 35.0 76°W w wUGm 6°S Gw wUGn 6°S 78°W Gw wUGn TEMPERATURA (ºC) Figura 2.- Temperatura (°C). Cr. Biomasa Desovante 0508-09 4°S 80°W wUGz jGi G{ i jGh¡ wGjù {GGt w GwGw ióGp 9 8 14°S 7 6 wGp wGj zGuç zGq j 5 4 3 16°S 2 h v÷ 1 0 82°W 80°W 78°W 76°W 82°W 74°W Figura 4.- Distribución de Salinidad (ups). Cr. Biomasa Desovante 0508-09 80°W 78°W 76°W 74°W Figura 5.- Distribución del Viento (m/s). Cr. Biomasa Desovante 050809 175 Informe IMARPE Vol. 35(2): Abril-Junio 2008 82°W 4°S 80°W 78°W 76°W 82°W 74°W 4°S wUGz jGi G{ Gw wUGn w wUGm 6°S 10°S 100 80 12°S 70 50 14°S 40 10°S 20 16°S 1400 1300 1200 12°S 1100 1000 900 i jGh¡ wGjù {GGt w GwGw ióGp 800 700 14°S 600 500 400 wGp wGj zGuç zGq j 300 200 100 16°S 0 h v÷ 82°W 0 82°W TURBULENCIA h v÷ 10 80°W 78°W 76°W Turbulencia En general los índices de turbulencia fueron bajos (<500 m3/s3) al norte de Pimentel. Al sur de esta localidad predominaron índices moderados (500 a 1000 m3/s3) en la zona oceánica y bajos en la zona costera. Los índices más altos se presentaron formando pequeños núcleos por fuera de las 30 mn al sur de Pimentel (Figura 6). Profundidad de la Isoterma de 15 °C.- Varió de 0 a 89 m. Las profundidades >80 m se localizaron frente a Salaverry, Huarmey y Paita. Las <20 m, se ubicaron próximas a la costa entre Punta Chao y Pisco. La 80°W 78°W 76°W 74°W 74°W Figura 6.- Distribución de la turbulencia (m3/s3). Crucero de Biomasa Desovante 0508-09 débiles (<4 m/s) se localizaron principalmente en la zona costera entre Punta Bermejo y Pisco. Con respecto a la dirección del viento se observó una predominancia de vientos del segundo y tercer cuadrante (sureste SE y suroeste SO). Normalmente al norte de Paita los vientos presentan componente norte, lo que no ocurrió en el presente crucero lo cual viene favoreciendo los procesos de surgencia en esa zona (Figura 5). 74°W wGi z o wGz j hù j wGo 1500 wGp wGj zGuç zGq j 30 76°W tù w jï w j { z n÷ wGj j j wGs o 8°S i jGh¡ wGjù {GGt w GwGw ióGp 60 w wUGm 6°S wGi z o wGz j hù j wGo 90 78°W Gw wUGn PROFUNDIDAD DE LA ISOTERMA DE 15 ºC tù w jï w j { z n÷ wGj j j wGs o 8°S 80°W wUGz jGi G{ Figura 7.- Distribución de la profundidad (m) de la isoterma de 15°C. Crucero de Biomasa desovante 0508-09 distribución de las isóbatas indicó flujos hacia el sur hasta Punta Falsa, asociados a la ESCC; luego, desde el sur de esta localidad fueron hacia el norte; convergiendo con los antes mencionados. Por otro lado, se observó un remolino antihorario entre Salaverry y Supe (Figura 7). ESTRUCTURA VERTICAL Paita La temperatura, dentro de las 80 mn tuvo distribución homogénea en toda la columna de agua, con tres isotermas (14, 15 y 16 °C) en la capa 0 - 180 m. El comportamiento de la isoterma de 15 °C indicó la presencia de un flujo al sur dentro de las 40 mn. Las ASS se localizaron sobre los 90 m de profundidad por fuera de las 70 mn. Las ACF se ubicaron dentro de las 50-40 mn. El oxígeno disuelto no presentó una oxiclina definida debido a la proyección hacia la superficie de todas las isolineas (2, 3 y 4 mL/L), tampoco se pudo observar el tope superior de las capa mínima de oxígeno (0,5 mL/L) en los primeros 100 m de profundidad (Figura 8). 176 Punta Falsa No se presentó una termoclina, hubo distribución homogénea con solo tres isotermas (14°, 15° y 16°C) entre 0 y 160 m. La estructura halina, mostró ASS por fuera de las 60 mn, ACF en la zona costera y una gran capa con aguas de mezcla. En la distribución de oxígeno se observó una oxiclina débil por fuera de las 60 mn que desapareció conforme se acerca a la costa; la proyección a la superficie de las isoxígenas, al igual que las isotermas, indican moderados procesos de afloramiento dentro de las 50 mn (Figura 9). Chicama Presentó una termoclina débil que desapareció dentro de las 50 mn asociada a moderados procesos de afloramiento. La isoterma de 15 °C se localizó entre los 40 m (litoral) y 80 m (a 100 mn). Las concentraciones de sales indicaron ACF dentro de las 30 mn, aguas de mezcla entre las 30 y 60 mn y ASS sobre los 40 m, por fuera de las 60 mn (Figura 10). Características oceanográficas físicas en invierno 2005 Vásquez SECCION PAITA (20-21/09/2005) TEMPERATURA SECCION PUNTA FALSA (18-19/09/2005) SALINIDAD 0 0 0 20 20 20 40 40 40 60 60 60 80 80 80 100 100 100 120 OXIGENO 0 TEMPERATURA 0 140 140 160 160 160 20 20 20 40 40 40 60 60 60 80 80 80 100 100 100 120 120 120 140 140 140 160 80 60 40 20 0 180 80 60 40 20 0 80 60 40 20 160 80 0 Figura 8.- Distribución Vertical de a) Temperatura (°C), b) Salinidad (ups) y c) Oxigeno (mL/L), frente a Paita. Cr. Biomasa Desovante 0508-09 60 40 20 0 0 50 50 100 100 150 200 250 40 20 0 80 60 40 20 0 SECCION CHIMBOTE (01-02/09) TEMPERATURA 0 0 150 200 60 Figura 9.- Distribución Vertical de a) Temperatura (°C), b) Salinidad (ups) y c) Oxigeno (mL/L), frente a Punta Falsa. Cr. Biomasa Desovante 0508-09 SALINIDAD 0 160 80 SECCION CHICAMA (09-10/09/2005) TEMPERATURA 300 OXIGENO 0 120 120 140 180 SALINIDAD SALINIDAD 0 50 50 50 100 100 100 150 150 150 200 200 200 250 250 250 300 300 300 350 350 350 400 400 400 450 450 450 OXIGENO 250 150 100 50 0 300 150 100 50 0 Figura 10.- Distribución Vertical de a) Temperatura (°C) y b) Salinidad (ups), frente a Chicama. Cr. Biomasa Desovante 0508-09 Chimbote Hubo una termoclina muy débil que desapareció dentro de las 60 mn. La isoterma de 15°C se localizó entre los 90 y 5 m de profundidad. Por fuera de 50 mn, una termoclina moderada sobre 80 m de profundidad. La distribución halina indicó la presencia de ASS por fuera de las 50 mn y sobre los 70 m de profundidad, las ACF se ubicaron dentro de las 40 mn y por debajo de las masas de agua antes mencionadas. El oxígeno disuelto mostró una oxiclina sobre los 100 m de profundidad, ubicándose el tope superior de la capa mínima de oxigeno alrededor de los 125 m (Figura 11). Punta Bermejo Presentó una débil termoclina que desapareció dentro de las 40 mn. La isoterma de 15 °C se ubicó entre 60 m (85 mn) y 0 m de profundidad (5 500 100 80 60 40 20 0 500 100 80 60 40 20 0 500 100 80 60 40 20 0 Fig. 11 Distribución Vertical de a) Temperatura (°C), b) Salinidad (ups) y c) Oxigeno (mL/L), frente a Chimbote. Cr. Biomasa Desovante 0508-09 mn). La distribución halina mostró ASS sobre los 40 m y ACF por fuera de las 40 mn. Dentro de las 30 mn hubo interacción de ambos tipos de agua (Figura 12). Callao No presentó una termoclina. La isoterma de 15 °C se localizó entre los 0 m (en la zona costera) y 60 m (a 100 mn). Las concentraciones de sales indicaron ACF dentro de las 30 mn, aguas de mezcla entre las 30 - 60 mn y las ASS por fuera de las 60 mn en una capa de 50 m de profundidad. La distribución de oxígeno disuelto presentó una oxiclina moderada que se proyectó rápidamente a la superficie dentro de las 30 mn. El tope superior de la capa mínima de oxígeno (0,5 mL/L) se localizó entre 25 y 180 m de profundidad (a 100 mn) (Figura 13). 177 Punta Infiernillos En la estructura térmica no se nota una termoclina definida, ubicándose la isoterma de 15 °C entre los 75 m (a 200 mn) y 0 m de profundidad (20 mn). La distribución halina indica la presencia de ASS sobre los 60 m y por fuera de las 95 mn, ACF dentro de las 50-60 mn, quedando un amplia franja de aguas de mezcla entre las 60 y 95 mn por interacción de las ACF, ASS y Aguas Templadas Subantárticas (ATS) cuyo núcleo principal se localizó alrededor de los 100 m de profundidad y a 120 mn. El oxígeno disuelto presentó una fuerte oxiclina entre los 50 y 100 m de profundidad, conformada por las isolineas de 1 a 5 mL/L. El tope superior de la capa mínima de oxígeno (0,5 mL/L) se localizó entre los 100 y 150 m de profundidad (Figura 14). Informe IMARPE Vol. 35(2): Abril-Junio 2008 SECCION CALLAO (24-25-08-2005) SECCION PUNTA BERMEJO (29-08-2005) 0 0 50 50 100 100 150 150 200 200 SALINIDAD TEMPERATURA SALINIDAD TEMPERATURA OXIGENO 0 0 0 50 50 50 100 100 100 150 150 150 200 200 200 250 250 250 300 300 300 350 350 350 400 400 400 450 450 450 250 250 300 80 60 40 20 0 500 100 300 80 60 40 20 80 60 40 20 0 0 Figua 12.- Distribución Vertical de a) Temperatura (°C) y b) Salinidad (ups), frente a Punta Bermejo. Cr. Biomasa Desovante 0508-09 500 100 SALINIDAD 0 0 50 50 50 100 100 100 150 150 150 200 200 200 250 250 250 300 300 300 350 350 350 400 400 400 450 450 450 150 100 50 0 200 150 100 50 La Figura 15 muestra la estructura térmica a 30 y 50 mn de la costa. Se observa que las mayores temperaturas (17 °C) se encontraron a 50 mn entre Chicama y Callao. Al norte y sur de dichas localidades la TSM tuvo distribución similar en ambas secciones. Se aprecia también que la isoterma de 15 °C mantiene un comportamiento similar al sur de Chimbote. En la zona norte (Paita y Punta Falsa) esta isoterma se profundiza más a 30 mn que a 50 mn; esto se debió a que dentro de las 40 mn predominaron flujos al sur, en cambio hacia afuera se observó frente a Paita y Punta falsa flujos al norte. 20 0 100 80 60 40 20 0 0 200 DISCUSIÓN 150 100 50 0 Figura 14.- Distribución Vertical de a) Temperatura (°C), b) Salinidad (ups) y c) Oxigeno (mL/L), frente a Punta Infiernillos. Cr. Biomasa Desovante 0508-09 Secciones Paralelas 40 esta masa de agua se localizó a las 120 mn, y además por la dinámica de las masas de agua, procesos de afloramiento y régimen de vientos que contribuyeron a un fuerte proceso de mezcla en estas zonas. OXIGENO 0 200 60 Fig.ura 13.- Distribución Vertical de a) Temperatura (°C), b) Salinidad (ups) y c) Oxigeno (mL/L), frente a Callao. Cr. Biomasa Desovante 0508-09 PUNTA INFIERNILLOS (16-17/08/2005) TEMPERATURA 80 La salinidad, a 30 mn de la costa (Figura 16a), indicó distribución homogénea, predominio ACF en toda la columna de agua; una ligera influencia de ASS al norte de Chimbote y posiblemente AES frente a Paita. A 50 mn de la costa (Figura 16b), se observó mayor influencia de ASS, entre el Callao y Chicama donde llegaron a 5060 m de profundidad. Debemos mencionar que en estas figuras no se observa la presencia de las ASS al sur de Pisco debido a que estas secciones están hechas con la información obtenida de los lanzamientos del CTD en las secciones verticales. Tampoco se observa claramente la presencia de las ATSA frente a Punta Infiernillo, debido a que el núcleo principal de 178 Los valores de anomalías térmicas obtenidos respecto a los promedios mensuales, dados por Zuta y Urquizo (1970) y Reynolds (1982), destacan la presencia, de valores bajos indicando anomalías negativas en la mayor parte del área de estudio, formando una franja atípica de baja temperatura que se localiza al norte de Chimbote hasta Talara. Esta lengua de bajas temperaturas, delimitada por las isotermas de 16 y 17 °C y que se extiende hacia el norte, indica un fortalecimiento de la Corriente de Humboldt, que al trasportar aguas frías provoca el ascenso de las isotermas, ubicándose la isoterma de 15 °C al norte de los 9 °C (Chimbote) 20 m más arriba de su ubicación promedio (Zuta 1988). Por otro lado, la gran área con anomalía negativa de la temperatura, relativa baja salinidad y bajo contenido de oxígeno principalmente en la zona norte, también se debería al intenso y constante proceso de surgencia, debido a que los vientos se presentaron favorables durante el crucero en toda la franja costera del Perú, lo cual provocó que el área fría abarcara un amplio sector oceánico. Características oceanográficas físicas en invierno 2005 Vásquez SECCION PARALELA A 30 MN DE LA COSTA 0 PAITA PTA FALSA CHICAMA CHIMBOTE PTA BERMEJO CALLAO SECCION PARALELA A 30 MN DE LA COSTA PTA INFIERNILLOS 0 CHICAMA PTA INFIERNILLOS CHIMBOTE PTA BERMEJO CALLAO 50 50 100 100 150 150 TEMPERATURA 200 SALINIDAD 200 SECCION PARALELA A 50 MN DE LA COSTA SECCION PARALELA A 50 MN DE LA COSTA 0 0 PAITA PTA FALSA PAITA PTA FALSA CHICAMA CHIMBOTE PTA BERMEJO CALLAO PAITA PTA FALSA CHICAMA PTA INFIERNILLOS CHIMBOTE PTA BERMEJO CALLAO PTA INFIERNILLOS 50 50 100 100 150 150 TEMPERATURA 200 SALINIDAD 200 Figura 15.- Distribución Vertical de Temperatura (°C) a) Sección Paralela a 30 mn de la costa y b) Sección Paralela a 50 mn de la costa. Cr. Biomasa Desovante 0508-09 Figura 16.- Distribución Vertical de Salinidad (ups) a) Sección Paralela a 30 mn de la costa y b) Sección Paralela a 50 mn de la costa. Cr. Biomasa Desovante 0508-09 84° W 82° W 80° W 78° W 76° W 74° W Pto. Pizarro Zorritos 4° S 4° S Pta. Sal Cabo Blanco Talara 0508-09 0308-09 0408-09 Paita Pta. Gobernador Parachique Pta. La Negra 6° S Mórrope Pimentel Chérrepe Pacasmayo Chicama 8° S Trujillo Salaverry Valores ecointegrados 6° S 8° S Punta Chao Chimbote Casma Punta Lobos 10° S 10° S Huarmey 0208-09 Punta Bermejo Supe Huacho 0208-09 Chancay 12° S 12° S Callao Pucusana Cerro Azul Tambo de Mora Pisco 14° S Bahía Independencia 14° S Punta Infiernillos Punta Caballas San Juan 84° W 82° W 80° W 78° W 76° W 74° W Pto. Pizarro Zorritos 4° S 0508-09 0308-09 4° S Pta. Sal Cabo Blanco Talara 0408-09 Paita Pta. Gobernador Parachique Pta. La Negra 6° S Mórrope Pimentel Chérrepe Pacasmayo Chicama 8° S Trujillo Salaverry Punta Chao Chimbote Valores ecointegrados 6° S 8° S Casma Punta Lobos 10° S 10° S Huarmey Punta Bermejo Supe Huacho 0208-09 Chancay 12° S 12° S Callao Pucusana Cerro Azul Tambo de Mora Pisco 14° S Bahía Independencia 14° S Punta Infiernillos Punta Caballas San Juan 84° W 82° W 80° W 78° W Figura 17.- Distribución de la Temperatura (°C) y Salinidad (ups) de los años 2002, 2003, 2004 y 2005 durante los cruceros sobre Biomasa Desovante. 179 76° W 74° W Informe IMARPE Vol. 35(2): Abril-Junio 2008 La proyección de las ATSA al interactuar con las ASS y ACF dieron lugar a la formación de una gran zona de mezcla frente a Punta Infiernillos, conservando temperaturas relativas bajas pero incrementando ligeramente sus concentraciones de sales, lo cual hace difícil medir el grado de influencia de esta masa de agua, al tener la mezcla valores similares a los de las ACF. Indirectamente se puede decir que las ATSA tuvieron una fuerte influencia a pesar de la atípica presencia de ASS en la zona sur. En general, las condiciones ambientales obtenidas en el crucero 0508-09 fueron más frías que las registradas en los últimos tres cruceros de Biomasa Desovante (020809, 0308-09 y 0408-09), asociadas a una menor presencia de aguas ecuatoriales en la zona norte, y aguas oceánicas en la zona norte y centro ya que en la zona sur se tuvo una fuerte presencia de ASS (Figura 17). CONCLUSIONES • La distribución térmica indica condiciones casi normales al sur de Chimbote, en cambio al norte las condiciones fueron frías. • La distribución halina indica una presencia inusual para la época de ASS frente a Huarmey y al sur de Bujama. REFERENCIAS Bakun A. 1973. Costal upwelling indices, west coast of North America; 1967-73. U.S. Dept. NOAA Tech. Rep. NMFS SSRF-671.103 p. Pizarro L, Tenorio J. 2005. Condiciones ambientales durante la operación MOPFEN 2005-08. Informe Interno, Instituto del Mar del Perú. Reynolds RW. 1982. A monthly average climatology of sea surface temperature. Technical Report NWs31. National Meteorological Center, NOAA, Silver Spring MD. 1-35. 180 Vásquez L. 2004. Condiciones ambientales durante el Crucero de Biomasa desovante 2004-0809. Informe Interno, Instituto del Mar del Perú. Vásquez L. 2003. Condiciones ambientales durante el Crucero de Biomasa desovante 2003-0809. Informe Interno, Instituto del Mar del Perú. Vásquez L. 2002. Condiciones ambientales durante el Crucero de Biomasa desovante 2002-0809. Informe Interno, Instituto del Mar del Perú. Zuta S, Urquizo W. 1970. Temperatura promedio de la superficie del mar frente a la costa peruana. Periodo 1928-1969. Bol. Inst. Mar Perú 2(8): 459-520. Zuta S. 1988. Variations of the Mass field and Currents off the Peru Coast. IOC (UNESCO). Time Series of Ocean Measurements, 33. Vol. 4: 55-75. INSTRUCCIONES A LOS AUTORES. INF. INST. MAR PERÚ El INFORME DEL INSTITUTO DEL MAR DEL PERÚ (INF. INST. MAR PERÚ) (ISSN 0378-7702) (IIMP) es la publicación trimestral del Instituto del Mar del Perú, que da a conocer los resultados preliminares o finales de una operación o actividad programada dentro de un campo específico de la investigación científica y tecnológica y que requiere difusión inmediata. En principio, sus páginas están abiertas a las investigaciones realizadas por el personal técnico y científico del propio Instituto del Mar del Perú (IMARPE), pero podrán aceptarse trabajos realizados en otras instituciones o laboratorios, autorizados por la Dirección Científica, previa calificación por Revisores Especializados y aceptación del Comité Editorial y del Editor. Además, publica notas científicas, notas técnicas, reseñas bibliográficas, y obituarios referidos a destacados investigadores marinos destacados, peruanos y de otras nacionalidades. Manuscritos originales La solicitud para publicación de un trabajo en el IIMP deberá ser remitida a la sede del Instituto del Mar del Perú, dirigida al Director Científico. Los manuscritos deberán presentarse impresos en original, a una columna, y dos copias fotostáticas (incluyendo ilustraciones y tablas) en una sola cara en papel tamaño A4, a doble espacio y con márgenes amplios. Se acompañará además el disquete o CD. Los textos deberán seguir estrictamente la estructura adoptada en el último volumen del IIMP, o serán devueltos a los autores. La primera página incluirá: (1)TÍTULO DEL TRABAJO (explícito, descriptivo y tan corto como sea posible), debajo, irá el título traducido al inglés; (2) EL NOMBRE COMPLETO DEL AUTOR O LOS AUTORES [(a) El autor debe decidir una forma definitiva de expresar su nombre, es decir no utilizar a veces los dos nombres de pila y los dos apellidos; otras veces sólo un nombre y un apellido; otras veces poner sólo iniciales para el segundo nombre y el segundo apellido; si esto cambia, en los registros y en las bases de datos, ya se trataría de autores diferentes; (b) se aconseja a los autores que generalmente utilizan sus dos apellidos, los unan mediante un guión]; también debe incluirse la dirección institucional, el domicilio y el correo electrónico, en notas al pie de página); En las páginas siguientes, deberá ir un RESUMEN en español, incluyendo palabras clave y un ABSTRACT en inglés, incluyendo Keywords (Resúmenes en otros idiomas son opcionales). También debe incluirse el título abreviado sugerido para los encabezamientos de páginas impares. Las notas, reseñas bibliográficas, etc. no tendrán resúmenes. Los textos deberán ser concisos, claros y directos, e incluirán usualmente una INTRODUCCIÓN, MATERIAL Y MÉTODOS, RESULTADOS, DISCUSIÓN (podría ser un solo capítulo, RESULTADOS Y DISCUSIÓN) Y CONCLUSIONES, además de AGRADECIMIENTOS Y REFERENCIAS DE LITERATURA. Los nombres científicos deberán ser escritos en cursivas e irán acompañados por autor y familia cuando sean citados por primera vez en el texto. Los nombres científicos deben ser corroborados con cuidado. Las medidas y sus abreviaturas deben corresponder al Sistema Internacional de Unidades (SI). En las citas bibliográficas en el texto, los nombres de autores irán en letras VERSALES, seguidos por el año de publicación, p.ej. (GUEVARA-CARRASCO 2004), ESPINOZA Y ZEBALLOS (2004) o ESTRELLA et al. (2006). También pueden hacerse las citas utilizando un número arábigo escrito en superíndice, que corresponda a la relación de referencias numeradas al final del artículo, de acuerdo al orden en que aparecen citadas en el texto. En las REFERENCIAS se incluirán únicamente los artículos mencionados en el texto, y se seguirá estrictamente el formato del IIMP. Se ordenarán alfabéticamente por autor y cronológicamente para cada autor. Los títulos de publicaciones periódicas irán abreviados. Las referencias a libros o capítulos de libros deberán ser completas, incluyendo ciudad, editorial y paginación. En todos los casos, los nombres del autor principal y los colaboradores, en la relación de Referencias, se anotarán por el apellido y la inicial del nombre de pila, sin punto, solamente separado por coma; año de publicación; Título del artículo, Revista y número de páginas. Ejs.: DELGADO E, SÁNCHEZ S, CHANG F, VILLANUEVA P. 2001.Título artículo---Revista----Vol. (Núm.): pp. El tipo de letra que se recomienda es el “arial”, y su tamaño en el artículo variará de la siguiente manera: Título en español, 14 NEGRA ALTA (= mayúsculas) centrado. Título en inglés, 12 NEGRA ALTA centrado. Nombre de los autores, 12 cursiva blanca (a y b), centrado. Direcciones de los autores, 9 blanca, centrado.- RESUMEN y CAPÍTULOS 12 negra, alta. Subcapítulo, 12 negra alta y baja (mayúscula y minúscula), alineado a la izquierda. Texto en general 11 blanca, inicio de párrafos con sangría. Referencias de Literatura, en tipo 10 blanca. Ilustraciones y tablas Las fotografías, dibujos, gráficos y mapas serán considerados como FIGURAS, numeradas consecutivamente con guarismos arábigos. Se recomienda remitir las ilustraciones y tablas como imágenes digitalizadas contenidas en archivos electrónicos (separadas del archivo conteniendo el texto). Si se tratara de dibujos, normalmente deberán ser efectuados con tinta china sobre papel Canson y montados sobre cartulina blanca. Las fotografías (a color, o blanco y negro) impresas en papel brillante (no mate) deberán ser nítidas y contrastadas, y serán montadas en cartulina blanca; no se recomienda la remisión de diapositivas. Cuando sea pertinente, la ilustración llevará una indicación del grado de aumento o reducción con respecto al original; se recomienda utilizar una barra en escala métrica e indicar el tamaño final con que deberá aparecer en la publicación (tomando en cuenta el tamaño de página del IIMP (A4). Toda ilustración deberá estar claramente identificada con nombres de autores, título del manuscrito y número de figura, anotando estos datos al reverso de la misma. Las leyendas para las ilustraciones, numeradas consecutivamente, irán en página aparte, luego del texto del artículo. Las TABLAS deberán ir numeradas consecutivamente, con guarismos arábigos, e impresas en páginas aparte. Sus leyendas, numeradas consecutivamente, irán en página aparte, luego del texto del artículo. 25 sobretiros (o separatas) serán entregados gratuitamente a los autores; si se desea más sobretiros, el exceso les será cobrado. Nota.- Debemos resaltar dos aspectos importantes: 1.- En el Sistema Internacional de Unidades (SI): Algunos de los símbolos actuales aceptados son: m (metro), cm (centímetro), mm (milímetro), km (kilómetro), g (gramo), ha (hectárea), mn (milla náutica), L (litro), mL (mililitro), W (vatio), kW (kilovatio), Hz (hertzio), kHz (kilohertzio). 2.- En la escritura de cantidades: - No debe usarse el punto decimal, sino la coma decimal. Ej. Un sol con cincuenta céntimos será S/. 1,50 (está mal si se escribe 1.50). Ver también a continuación. - Las cantidades de miles y millones, deben separarse por puntos, cada tres dígitos: Ejs. Oncemil quinientos soles con cincuenta céntimos será: S/. 11.500,50 (estará mal si se escribe 11,000.50) - quince millones trescientos cincuenta mil y veinticinco céntimos se deberá escribir 15.350.000,25 - Cuando en un texto se tratan cifras con muchos ceros puede escribirse en letras. Ej. 1.000.000.000 (mejor escribir mil millones), o en vez de 100.000 escribir cien mil. - (Cualquier información: a [email protected])