- Ninguna Categoria
FRANCISCO CARDENAS RUIZ
Anuncio
UNIVERSIDAD VERACRUZANA FACULTAD DE MEDICINA VETERINARIA Y ZOOTECNIA PAPEL DE LA FSH EN EL CICLO ESTRUAL Y LA OVULACIÓN MÚLTIPLE EN LA HEMBRA BOVINA MONOGRAFÍA PROFESIONAL QUE PARA OBTENER EL TÍTULO DE: MÉDICO VETERINARIO ZOOTECNISTA PRESENTA: FRANCISCO CÁRDENAS RUIZ ASESORES: MPA. BERTHA CLEMENTINA HERNÁNDEZ CRUZ DR. FELIPE MONTIEL PALACIOS VERACRUZ, VER. 2009 DEDICATORIA Por su amor y su apoyo, su comprensión y en especial por creer en mi. A mis papás Antonio y Elvia (Q.D.E.P.) con todo cariño y a mis hermanos Chela, Marytoña, Elvia, Toño y Felipe con todo afecto. A Rocío, el amor de mi vida. A mis hijos Valeria, Fey y Paco, que son el latir de mi vida. A mis amigos Carlos Briseño, Santiago Mier, Marisela Hernández, Elsa Cortez, Inés Figuereo, que siempre están en las buenas y en las malas. AGRADECIMIENTOS A mis maestros de la Facultad de Medicina Veterinaria y Zootecnia, de la Universidad Veracruzana, que me forjaron y orientaron durante mi época de estudiante. Gracias: MVZ José Manuel Páez (Q.E.P.D.), MVZ. Sergio Castellanos, MVZ. Augusto Mancisidor. A mis asesores: MPA. Bertha Clementina Hernández Cruz, Dr. Felipe Montiel Palacios. A mi Honorable Jurado. Índice Página Resumen ..................................................................................................…….. 1 Introducción...................................................................................….......……… 2 Objetivo ……………………………………………………………………………….. 4 Justificación ....................................................................................….......……. 4 Metodología .............................................................................................……... 4 1. Hormona folículoestimulante (FSH)……………………………………….. 4 2. Papel de la FSH en el ciclo estrual..…………………………………………. 6 2.1 Ciclo estrual………………………………………………..……………… 6 2.2 Desarrollo folicular ………………………………….........……………… 8 2.3 Foliculogénesis …………………………………………..………………. 9 2.4 Ovulación ………………………………………………..…..…………… 21 3. Papel de la FSH en la ovulación múltiple……………………………….....… 25 3.1 Sincronización del celo …………….…..………………………………... 25 3.2 Tratamiento hormonal……………………………………….…………... 28 Conclusiones……………………………...….…………….………………………… 31 Literatura citada ……………………………………………………..………………. 32 1 RESUMEN Francisco Cárdenas Ruiz. Papel de la FSH en el ciclo estrual y la ovulación múltiple en la hembra bovina. Monografía profesional. Asesores: MPA. Bertha Clementina Hernández Cruz, Dr. Felipe Montiel Palacios. Facultad de Medicina Veterinaria y Zootecnia, Universidad Veracruzana, Veracruz, Ver. 2009. La hormona folículoestimulante (FSH) es producida en la hipófisis anterior y secretada en forma pulsátil a lo largo del ciclo estrual de la hembra bovina; es esencial para la función reproductiva normal, ya que regula el crecimiento y maduración de los folículos ováricos. Durante la fase folicular del ciclo estrual los niveles plasmáticos de FSH aumentan significativamente, dándose el desarrollo y maduración folicular. Durante la fase de selección, la mayoría de los folículos antrales inician su crecimiento bajo la acción de la FSH; en la fase de dominancia sólo uno de ellos se desarrolla, mientras los otros sufren atresia. La FSH se une a sus receptores en las células de la granulosa del ovario; esta respuesta se ve aumentada en hembras que reciben tratamiento para ovulación múltiple (OM). La OM es la inducción de más de una ovulación, mediante la aplicación de hormonas que estimulan los folículos produciendo un crecimiento intensivo, con el fin de que al momento de la inseminación artificial se produzca más de un embrión. Para realizar la OM se utiliza comúnmente FSH, que hace que en ambos ovarios se desarrollen varios folículos dominantes. Los tratamientos de inducción de OM con FSH comprenden inyecciones de la hormona en dosis decrecientes dos veces al día por 4 días, comenzando el día 10 del ciclo estrual. Aunque estos tratamientos en general resultan en un mayor número de folículos desarrollados, existe gran variación en la respuesta al tratamiento entre animales y en el mismo animal. Esta variabilidad puede deberse a diferencias en los tratamientos de OM, o a factores inherentes al animal y a su medio ambiente. Palabras clave: Bovino, ciclo estral, folículos ováricos, ovulación, sincronización. 2 INTRODUCCIÓN Las hormonas son sustancias químicas producidas por tejidos específicos, que se vierten directamente en el torrente sanguíneo en respuesta a determinados estímulos, provocando una respuesta específica que puede manifestarse de forma inmediata o mediata. Como resultado, la célula receptora del estímulo modifica su comportamiento a través de cambios metabólicos. Las acciones promovidas por las hormonas pueden ser modificación en la permeabilidad de las membranas o en los mecanismos de transporte, modificación de la síntesis proteica y/o de la actividad enzimática celular (Calandra y de Nicola, 1985). La producción hormonal es controlada por un sistema de retroalimentación negativa, en el que el aumento de la concentración de hormona origina una menor producción de la misma, usualmente mediante una interacción con el hipotálamo o hipófisis (Hafez, 2002). El eje hipotálamo-hipófisis-gónadas regulan la producción de hormona luteinizante (LH), FSH y esteroides ováricos, esenciales para la maduración folicular, ovulación, implantación y mantenimiento de la gestación (Ascoli y Segaloff, 1996). La producción de FSH inicia en el hipotálamo, con la liberación de hormona liberadora de gonadotropinas (GnRH), que es regulada por el núcleo arcuato ubicado en el hipotálamo medio basal. La GnRH es liberada en forma de pulsos por la vascularización portal, a través de la cual llega a la hipófisis anterior. Allí se une a los receptores de superficie de las células productoras de FSH y LH o gonadotropos, los que también las secretan en forma pulsátil (Rosell et al., 2004). En bovinos, se ha observado gran variabilidad en la respuesta del eje hipotálamohipósifis-gónada-órgano blanco al estímulo hormonal, ya sea fisiológico, o utilizando diferentes tratamientos para la inducción de ovulación múltiple (OM) (Mapletoft et al., 2002). Esto podría deberse a cambios estructurales en los genes de receptores hormonales, que podrían resultar no deseables en programas 3 reproductivos. Se ha comprobado que muchos trastornos cíclicos son de origen genético y afectan generalmente al ovario (Adashi y Hennebold, 1999). El desarrollo del sistema reproductivo inicia en el embrión. Después del nacimiento, se produce un estado de latencia hasta la pubertad, cuando el animal debe alcanzar el tamaño y peso adecuados para enfrentar un estado de futura madurez sexual o inicio del ciclo reproductivo, el cual es coordinado por las gonadotropinas FSH y LH, responsables de controlar la función ovárica. La FSH es esencial para la función reproductiva normal, es decir, para el desarrollo folicular en las hembras y la maduración sexual normal (Pearly et al., 2002). A nivel ovárico, la FSH actúa sobre los receptores en las células de la granulosa, regulando la maduración folicular (Dieidrich et al., 1998). La LH induce la ovulación y luteinización de las células de la granulosa y teca del ovario, en donde se liberan los estrógenos producidos por los folículos, y la progesterona secretada por el cuerpo lúteo (CL). Ambos regulan el ciclo reproductivo mediante mecanismos de retroalimentación que actúan sobre el hipotálamo y la hipófisis (Mukasa-Mugerwa, 1989). Esta revisión resume el papel de la FSH en el ciclo estrual y la OM en la hembra bovina, permitiendo conocer los factores que originan la variabilidad en la efectividad de los tratamientos de OM como parte de programas reproductivos. 4 OBJETIVO Recopilar y analizar información sobre el papel de la FSH en el ciclo estrual y la ovulación múltiple en la hembra bovina, a fin de generar un documento que permita el acceso al conocimiento de esta área. JUSTIFICACIÓN El conocimiento del papel de la FSH en el ciclo estrual y la ovulación múltiple en la hembra bovina permite conocer la fisiología y endocrinología que se ven involucradas en estos procesos, así como entender la respuestas a los tratamientos farmacológicos para OM, para aprovechar al máximo técnicas de mejoramiento genético como la inseminación artificial y la transferencia de embriones. METODOLOGÍA Se revisaron artículos publicados en revistas de arbitraje internacional y nacional relacionados con el tema el papel de la FSH en el ciclo estrual y la ovulación múltiple en la hembra bovina, publicados en los últimos 10 años, o anteriores cuando se consideró pertinente. Asimismo, se realizaron búsquedas relacionadas con el tema, utilizando bancos de datos electrónicos disponibles en bibliotecas y/o centros de investigación. 1. HORMONA FOLÍCULOESTIMULANTE (FSH) La FSH es una glicoproteína de alto peso molecular, secretada en pulsos de baja frecuencia desde los gonadotropos de la hipófisis, y es producto de la unión de la GnRH a los receptores acoplados a proteínas G en la hipófisis (Recabarren et al., 5 2006). La FSH está compuesta por dos subunidades no covalentes α y β, codificadas por genes ubicados en cromosomas diferentes (Graus et al., 2001). La subunidad α es común para todas las hormonas glicoproteicas, mientras que la subunidad β confiere la especificidad hormonal (Pierce y Parsons, 1981). Sin embargo, el control de su secreción depende en parte de la estimulación ejercida por la GnRH secretada por el hipotálamo, y en parte de la interacción de inhibinas, que inhiben su secreción, y de activinas, que la estimulan (Carroll et al., 1991). La FSH cumple diferentes funciones biológicas, como la proliferación y secreción de células de Sertoli, influyendo en la espermatogénesis en machos (Kilgour et al., 1998), así como reclutamiento y selección de folículos ováricos y síntesis de estradiol a partir de precursores androgénicos en hembras (Campbell et al., 2004). La FSH ejerce su acción a través de un mecanismo en el cual la hormonoa llega al interior de las células blanco al unirse a receptores específicos encontrados en la membrana plasmática (Pierce y Parsons, 1981). La expresión de estos receptores en las células de la granulosa depende del estadío de la diferenciación celular en el ovario (Donadeu y Ascoli 2005). Sin embargo, por diferentes razones algunos receptores en el órgano blanco no reconocen la señal hormonal. La FSH es capaz de unirse a su receptor en las células de la granulosa, respuesta que se ve aumentada en vacas con inducción de ovulación múltiple (OM) por la alta producción de óvulos (Mapletoff et al., 2002). Los receptores para FSH son expresados tanto en células maduras como inmaduras, y cumplen dos funciones principales: reconocer la hormona por medio de un acople o ligadura de ésta, e iniciar los eventos químicos que dan lugar a la acción biológica del sistema hormonal específico, en este caso la maduración folicular, por la combinación receptor-hormona (Wangsness et al., 1981). Los receptores FSHR o metabotrópicos, pertenecen a una familia común de genes acoplados a proteínas G (GPCRS). Estos receptores están formados por siete 6 hélices insertadas en el plasmalema celular (Shemesh, 2001). El receptor para FSH pertenece a una familia de proteínas homólogas que atraviesan siete veces la membrana. Estos receptores transmembranales regulan a las proteínas G y tienen la función de transducir información del espacio extracelular al interior de la célula, estimulando o inhibiendo sistemas de segundos mensajeros (Kroese et al., 2003). Una vez que el receptor es expresado en las células de la granulosa, se inicia la señalización en cascada. La FSH se une a su receptor con alta afinidad, y los eventos de unión inician un cambio conformacional en el receptor, el cual activa a las proteínas G encargadas de iniciar la respuesta intracelular (Simoni et al., 1997). 2. PAPEL DE LA FSH EN EL CICLO ESTRUAL 2.1 CICLO ESTRUAL En la hembra bovina, el ciclo estrual se divide en dos fases, folicular (proestro y estro, terminando con la ovulación) y lútea (metaestro y diestro, terminando con la luteólisis), con una duración promedio de 21 días (Whittier, 2003). El 84% de los ciclos duran entre 18 y 24 días, siendo la manifestación de celo aproximadamente de 12 a 16 h. La ovulación ocurre 10 a 15 h después del final del celo (Hafez, 2002). La fase folicular dura de 2 a 4 días y comienza al final del ciclo que la antecede con un aumento significativo y progresivo de los niveles circulantes de FSH, comprendiendo los estadíos de desarrollo y maduración folicular o foliculogénesis. La fase lútea dura aproximadamente 18 días (Ownby, 2004). logy La FSH secretada en ondas a lo largo del ciclo estrual desempeña un papel fundamental para el crecimiento de un grupo de folículos. Las hembras Bos taurus y Bos indicus presentan generalmente dos a tres ondas de crecimiento folicular en su ciclo estrual, variabilidad que no está relacionada con la raza, edad y nivel de fertilidad de las mismas. El número de ondas de crecimiento folicular durante el ciclo estrual es determinado por la longitud de la fase luteal (Gigli et al., 2006). 7 En los ciclos estruales de dos a tres ondas, la emergencia de la primera onda folicular ocurre generalmente alrededor del día de la ovulación. La emergencia de la segunda onda ocurre en el día 9–10 para los ciclos de dos ondas, y en el día 89 para los de tres ondas. En estos últimos, la onda emerge en el día 15–16. En las ondas foliculares anovulatorias, los foliculos sufren regresión sin ovular. El folículo dominante presente en el inicio de la luteólisis será el folículo dominante ovulatorio, y la emergencia de la próxima onda (primera del nuevo ciclo) se demorará hasta el momento de la ovulación. La primera, segunda y tercera onda folicular se observan durante las fases luteal temprana, luteal media y folicular, respectivamente (Ginther et al., 1997). En las ondas foliculares no ovulatorias, las concentraciones de estradiol circulante se mantienen bajas, pues a pesar de que el tamaño preovulatorio puede ser alcanzado, el folículo dominante no reúne las características funcionales preovulatorias, tales como la producción de estradiol. El folículo dominante (folículo estrógeno activo) de la última onda continúa su crecimiento y la biosíntesis de estradiol, hasta que se alcanza el nivel suficiente de estradiol circulante para producir la retroalimentación positiva sobre el área de control cíclico del hipotálamo que aumenta los pulsos de GnRH/ LH/ FSH, e induce el pico preovulatorio de LH y FSH, y por lo tanto, la ovulación. Después del pico de LH esta retroalimentación se interrumpe y las células de la granulosa sintetizan progesterona y se luteinizan. El mecanismo que regula la dinámica folicular está basado en las respuestas diferenciales a la FSH y LH (Johnson et al., 2004). Durante la fase de selección, la mayoría de los folículos antrales comienzan su crecimiento bajo la acción de la FSH; en la fase de dominancia sólo uno de los folículos seleccionados se desarrolla a expensas de los otros. Después de la ovulación, en la cavidad dejada por el folículo se forma una estructura similar a un coágulo, el cuerpo hemorrágico, que se transforma en CL hacia el día 5 del ciclo (día 0 = estro), iniciando la secreción de progesterona (P4). Hacia el día tres, las concentraciones plasmáticas de P4 son bajas (1 ng/mL), del día 7 al 18 aumentan 8 (6-10 ng/mL), y durante la luteólisis disminuyen en 24 a 36 h (Wright y Malmo, 1992). Hacia el día 15 del ciclo, la P4 declina bruscamente, desencadenando la secuencia de cambios hormonales que producen el estro y la ovulación; por lo tanto, el CL es el reloj biológico que controla la duración del ciclo estrual. El CL degenera si la vaca no resulta gestante, convirtiéndose en cuerpo albicans y permaneciendo visible en el ovario durante varios ciclos subsecuentes. Al mismo tiempo, la FSH estimula el desarrollo del folículo que ovulará en el siguiente ciclo (Hafez, 2002; Asprón, 2004). 2.2 DESARROLLO FOLICULAR El ovario bovino contiene un gran número de folículos primordiales al nacimiento, pero menos del 1% de éstos ovulan durante la vida reproductiva de la vaca (Eppig, 2003). La mayoría de los folículos que entran en el depósito de crecimiento están destinados a la atresia, aunque no se conoce el mecanismo por el cual ciertos folículos son elegidos para dejar la reserva de folículos primordiales y convertirse en folículos primarios continuando su desarrollo, obteniendose al final del ciclo sólo un folículo para ovular. Sin embargo, si se utilizan tratamientos para OM con gonadotropinas, éstas estimulan a más de un folículo a crecer a talla ovulatoria y ovular. La OM consiste en administrar durante el crecimiento folicular hormonas que provoquen el desarrollo de más folículos, no solamente de uno como normalmente sucede (Calva et al., 2001). Se necesitan alrededor de 60 días para que un folículo primordial llegue a preovulatorio o de Graaf, y alrededor de 47 días para que llegue de la etapa antral temprana (0.1 mm) a preovulatoria (13-16 mm) (Lussier et al., 1987). De esta manera, un folículo dominante se observa en cada fase. Durante el crecimiento del folículo dominante y cese del crecimiento de los otros folículos, aparece una onda folicular de emergencia después de que el folículo dominante cesó su crecimiento u ovuló (Hiroyuki et al., 1995). 9 En la fase ovulatoria o fase de atresia, el folículo dominante alcanza dimensiones preovulatorias o sucumbe a la degeneración, respectivamente. Al mismo tiempo, a lo largo del ciclo estrual se producen tres picos de estrógenos (E2), tanto en la circulación periférica como en las venas ováricas, que son coincidentes con los períodos de dominancia folicular (Perry, 2004). El reclutamieno cíclico tiene incio después de la pubertad y resulta del incremento de los niveles de FSH en la circulación sanguínea. No se conoce el mecanismo por el cual antes de la pubertad, un folículo primordial deja de formar parte de la reserva e inicia su crecimiento, aunque se han señalado algunos factores de crecimiento, ciertas neurotropinas y a la hormona inhibidora de los conductos de Muller involucrados en el proceso (Espinosa et al., 2007). 2.3 FOLICULOGÉNESIS 2.3.1 Fase preantral y antral La primera fase de la foliculogénesis se conoce como preantral o fase independiente de gonadotropinas, y es caracterizada por el crecimiento y diferenciación del ovocito. La segunda fase, llamada fase antral o fase dependiente de gonadotropinas, se caracteriza por un gran aumento en la talla del folículo, aproximadamente arriba de 25 mm (Erickson, 2003). En la fase preantral, los factores de crecimiento producidos por el ovocito juegan un papel crucial en la regulación de la foliculogénesis (Erickson y Shimasaki, 2000). La fase preantral es controlada por factores de crecimiento producidos localmente, que actuan a través de mecanismos paracrinos y autocrinos. La segunda fase es regulada por la FSH y LH, así como otros factores de crecimiento que pueden estimular la proliferación celular y modular la acción de las gonadotropinas. En este mecanismo, la interacción de hormonas gonadotrópicas, factores de crecimiento y hormonas esteroides que intervienen tanto en la fase preantral como 10 en la folicular regulan negativa o positivamente la foliculogenésis, ovulación y luteogénesis (Erickson, 2003). La fase preantral se divide en tres estadíos: folículo primordial, primario y secundario (Erickson, 2003). Después de que ocurre la formación del antro, la tasa de crecimiento folicular se acelera y un folículo dominante es seleccionado de un grupo de folículos de la fase luteal del ciclo. La atresia puede ocurrir en el folículo secundario, pero la mayor incidencia es en el grupo de pequeños y medianos folículos (Henao et al., 2000). El proceso de la foliculogénesis ocurre en la corteza del ovario. Estos eventos inician en el folículo primordial (Hafez, 2002). 2.3.2 Folículo primordial Todos los folículos primordiales se forman en el feto; como resultado, todos los ovocitos que una hembra tendrá en su vida reproductiva están presentes en el ovario al nacimiento. Este número de óvulos o folículos primordiales constituye la reserva ovárica (Hafez, 2002). El reclutamiento de folículos primordiales ocurre a una tasa relativamente constante en la vida productiva del animal. Las concentraciones plasmáticas de FSH actúan para acelerar la transición de primario a secundario. Un cambio en la forma de células escamosas a cuboidales y la adquisición del potencial mitótico en las células de la granulosa es seguido de la activación de ciertos genes y el subsecuente crecimiento del ovocito. Además, los mecanismos primarios que controlan el reclutamiento envuelven a las células de la granulosa y el ovocito, que es un tejido de respuesta de la activación primaria (Hafez, 2002). En esta etapa, a los activadores del reclutamiento folicular se les conoce como Kit de Ligandos de la Granulosa (Granulosa-Derived Kit Ligand; KitL) (Parrott y Skinner, 1999), formado por la proteína morfogenética derivada de la teca (Derived Bone Morphogenetic Protein) (Findlay et al., 2002), y altos niveles plasmáticos de FSH hipofisiaria (Fortune et al., 2000). En el KitL también se 11 encuentra la substancia mulleriana de inhibición (Müllerian Inhibiting Substance, MIS) que inhibe el reclutamiento folicular (Durlinger et al., 2002). En roedores, el KitL puede promover el crecimiento folicular (Packer et al., 1994), indicando que pudiera ser una parte del mecanismo de activación primaria del folículo, y el momento de su secreción estimula el crecimiento del ovocito; además, si esta proteína se encuentra ausente, no se desarrolla el folículo de Graaf y la hembra es infértil (Durlinger et al., 2002). Se ha observado que bajas concentraciones de FSH decrecen la tasa del KitL. Esto sugiere que una correcta concentración de FSH es crucial para la regulación de los factores paracrinos que promueven el desarrollo del ovocito, y que el KitL es hormonalmente regulado por FSH, y sin la presencia de FSH éste no se observa (Thomas y Vanderhyden, 2006). Se ha sugerido que el desarrollo del ovocito se daña cuando se inhibe la señalización de FSH, y que la comunicación entre el ovocito y las células de la granulosa se ve influida cuantitativamente por los FSHR (Thomas y Vanderhyden, 2006). Se ha demostrado que la FSH es capaz de regular las células de la granulosa para que se conecten con el ovocito (Combelles et al., 2004). 2.3.3 Folículo primario El folículo primario presenta células cuboidales en la granulosa, las cuales son organizadas en una sola capa rodeando al ovocito. En el folículo primario ocurren eventos como la expresión de los receptores de FSH en las células de la granulosa, lo cual sucede por acción de la activina producida por las células de la granulosa, a través de mecanismos autocrinos y paracrinos. Otro evento es el crecimiento y diferenciación del ovocito (Erickson, 2003). Se ha observado en animales con altos niveles de FSH plasmática, que a pesar de no encontrarse en una fase dependiente de gonadotropinas, ésta sí puede acelerar el desarrollo del folículo primario, lo que sugiere la posibilidad de que la FSH pudiera afectar eventos en el folículo primario (Fortune et al., 2000). 12 El mecanismo de acción de la activina es controlado por la folistatina (FSRP, follistatin-related protein), una proteína que une a esta última y a la inhibina con una elevada afinidad (Knigth y Glister, 2001). Se ha sugerido que la activina puede jugar un papel importante como regulador local en el folículo ovárico, porque ambos (folículo y ovario) producen y responden a ésta (Erickson, 2003). La inducción del FSHR en las células de la granulosa es una etapa crítica en la fisiología reproductiva, ya que el desarrollo de grandes folículos es estimulado y depende de la FSH. La influencia estimulatoria de la activina sobre los FSHR es un mecanismo por el cual el folículo preantral llega a tener respuesta a la FSH (Woodruff et al., 1988), lo que aunado a la interacción de la señalización en cascada de péptidos y hormonas esteroides en el ovario, producen el crecimiento folicular, ovulación y luteinización (Richards et al., 2002). Posteriormente, la FSH estimula la expresión de receptores para LH y éstos a su vez la de receptores para P4, al tiempo que estimulan la producción de folistatina (proteína unidora de activina), la cual suprime el nivel de activina en las células de la granulosa, y por tanto su efecto sobre la expresión de los FSHR (Kishi et al., 1998). Los mecanismos locales de retroalimentación terminan la acción de la activina después de un corto período. Esta reducción de la actividad de la activina inducida por la FSH puede ser importante para el crecimiento normal de cada folículo (Tano et al., 1995). Conforme las células de la granulosa continúan su proliferación y el folículo va completando la formación de su antro, las concentraciones crecientes de estrógenos incrementan la síntesis de inhibina y folistatina, disminuyendo las concentraciones efectivas en el folículo, lo cual permitirá la expresión de los LHR en células de la granulosa y la teca; el incremento en la secreción de inhibinas permitirá la síntesis de novo de andrógenos, principalmente androsteniona, y la expresión de LHR en las células de la teca, además de disparar la progresión 13 meiótica en el ovocito detenido en diploteno I y de regular la síntes hipofisiaria de FSH a través de mecanismos de retroalimentación negativa (Findlay et al., 2002). Las inhibinas y activinas son producidas por las células de la granulosa del folículo dominante; están formadas por la unión de las subunidades α y β. La unión de la subunidad α con la subunidad βA, forma la inhibina A. La unión de la subunidad α con la βB, forma la inhibina B. En cambio la unión de dos subunidades αA o dos subunidades βB dan lugar a la hormona activina (activina A ó B de acuerdo a que subunidades la conforman). Tanto las inhibinas como las activinas actúan en forma parácrina, inhibiendo o estimulando el crecimiento de los folículos, respectivamente (Glister et al., 2001) La inhibina es una hormona glicoproteíca secretada por las células de la granulosa que actúa como inhibidor de la producción de FSH (Montaño y Ruiz, 2005). Ambos, inhibina y activina durante el proestro provocan mediante un sistema de retroalimentación negativa clásico entre el folículo y la hipófisis, disminución en los niveles periféricos de FSH, lo que bloquea el reclutamiento de nuevos folículos (Hafez, 2002). Las activinas también participan en la expresión de aromatasa en las células de la granulosa, inhibiendo a la vez la expresión de los LHR en las células de la granulosa y de la teca (Gigli et al., 2006). La inhibina y los estrógenos producidos por el folículo dominante son responsables del estímulo atrésico en los folículos subordinados, promueven la expresión de LHR en las células de la granulosa del folículo dominante y ejercen un estímulo inhibitorio sobre la liberación de FSH de la adenohipófisis, haciendo que el crecimiento del folículo dominante sea dependiente de LH hasta alcanzar el tamaño preovulatorio (Fortune et al., 2000). En este proceso se involucran también los factores de crecimiento semejantes a insulina (IGFI)-I y II, los que actúan como reguladores hipofisiarios del crecimiento folicular potencializando la acción de las gonadotropinas en el ovario por 14 mecanismos que no son del todo entendidos (Knight y Glister, 2006). La unión funcional de las vías de señalización entre FSH e IGF-1 se basan en la observación de que IGF-1, el receptor de IGF-1 (Igf-1r), y FSHR se localizan en las células de la granulosa de los folículos pequeños y preovulatorios (Velásquez y Mendieta, 2005). Las interacciones potenciales que ocurren en la transición de folículo primario a secundario involucran el KitL y factores básicos de crecimiento de los fibroblastos (bFGF) secretados por las células pregranulosas y ovocito, respectivamente, y tienen efectos estimulantes mutuos sobre el ovocito y las células de la granulosa. También promueven el reclutamiento de las células de la teca de la población de células intersticiales estromales. Las células intersticiales estromales y las células de la teca secretan BMP-4 y BMP-7, que promueven la activación y supervivencia folicular. El GDF-9 y/o BMP-15 secretados por el ovocito del folículo activado, promueven la proliferación de las células de la granulosa, la expresión del kitL y la formación de la teca. Al mismo tiempo, las células de la granulosa del fóliculo secretan AMH que al parecer actua como un freno en el reclutamiento de folículos primordiales (Knight y Glister, 2006). Un evento importante en el desarrollo del folículo primario es el desarrollo de uniones en hendidura (Gap junctions) entre el ovocito y las células de la granulosa (Eppig, 1994), que permiten el intercambio de información y moléculas. Los canales intercelulares, también llamados canales de las uniones en hendidura (CUH), son permeables a iones y moléculas pequeñas (menores de 1, 2 kDa) y comunican el citoplasma de células adyacentes, permitiendo la coordinación de respuestas (metabólicas y eléctricas) de un conjunto de células frente a un determinado estímulo, tanto fisiológico como fisiopatológico (Eppig, 2003). Cada CUH lo constituyen dos hemicanales o conexones, cada uno es aportado por una de las dos células en contacto; a su vez, cada hemicanal está compuesto por seis subunidades proteicas denominadas conexinas (Cxs). Las Cxs son 15 codificadas por una familia de genes compuesta por al menos 20 genes diferentes en el humano y se denominan con el prefijo Cx seguido de un número que indica el peso molecular teórico aproximado (Brañez et al., 2004). Éstas se acoplan a células adyacentes permitiendo la difusión de iones, metabolitos y señales potenciales de moléculas tales como AMP cíclico y calcio (Saez et al., 2003). Después del reclutamiento folicular, la conexina 37 (Cx37) es sintetizada por el ovocito y forma uniones de comunicación alrededor de las células de la granulosa. Su interrelación en esta etapa es importante para la foliculogénesis y la fertilidad femenina en animales de laboratorio. Se cree que las células de la granulosa se conectan al ovocito vía conexina 37, y que mediante sus uniones de comunicación los materiales pasan de las células nodrizas de la granulosa hacia dentro del ovocito (Eppig, 1994). Estas células derivan materiales que son moléculas reguladoras y nutrientes, y que son requeridos para el crecimiento del ovocito y para la adquisición del potencial de reiniciar la meiosis (Webb et al., 2002). Los ovocitos con crecimiento completo rara vez reinician la meiosis durante la foliculogénesis (Thomas y Vanderhyden, 2006). Esto ha conducido al concepto de que existe un mecanismo de inhibición meiotica. Se sugiere que las altas concentraciones de AMP cíclico inhiben la maduración meiótica. La hipótesis es que al inducir FSH el AMPc en las células de la granulosa, éste se difunde dentro del ovocito a través de la Cx37, y su unión provoca la inhibición de la continuación de la meiosis (Webb et al., 2002). 2.3.4 Folículo secundario La foliculogénesis continúa en el folículo preantral y la estructura del folículo comienza a cambiar. El mayor cambio ocurre durante la transición e incluye un acúmulo de un número aumentado de células de la granulosa y la adquisición de la teca. El desarrollo de un folículo primario a uno secundario de crecimiento completo resulta en la activación de mecanismos endocrinos y paracrinos que 16 regulan el proceso y que envuelven factores de crecimiento producidos por el ovocito (Espinoza et al., 2007). La transición de folículo primario a secundario inicia con la adquisición de una segunda capa de células de la granulosa (Pineda y Dooley, 2003). El estado primario y secundario es una etapa crítica regulatoria en el proceso de la foliculogenésis; el crecimiento y desarrollo folicular en el estadío primario en ratas y ovejas en ausencia de GDF-9 and BMP-15 se detiene (Chang, 2002). Estos factores derivados del ovocito son obligatorios para la transición de folículo primario a secundario y presumiblemente tienen la habilidad de estimular la mitósis y su patrón de arreglo (Espinoza et al., 2007). El desarrollo de un folículo secundario se encuentra también caracterizado por el desarrollo tecal (Erickson et al., 1985). En el momento de la transición de folículo primario a secundario, numerosas capas de células aparecen alrededor de la lámina basal (Gigli et al., 2006). En el desarrollo del folículo dos capas primarias de teca aparecen, teca interna que se diferencia en células de la teca intersticial, y teca externa que se diferencia en células de músculo liso. El desarrollo de la teca está acompañado por la formación de numerosos vasos presumiblemente a través de una angiogénesis por la acción de los factores de crecimiento angiogénicos que estimulan la vascularización (Antezak y Van Vlerkom 2000). Consecuentemente, la sangre circula alrededor del folículo trayendo nutrientes y gonadotropinas y sacando productos y desechos del folículo en desarrollo. Cuando se completa la fase antral de la foliculogénesis, un folículo secundario de desarrollo completo contiene cinco distintas unidades estructurales: un ovocito de desarrollo completo rodeado de la zona pelúcida, aproximadamente nueve capas de células de la granulosa, una lámina basal, teca interna, teca externa y una red de capilares en el tejido tecal (Erickson, 2003). 17 Cuando la teca se diferencía y se forma el antro, se da la expresión de una serie de genes que permite a las células intersticiales de la teca tener la capacidad de producir androstendiona. Esto pudiera indicar que la teca de todos los folículos de Graaf expresan este estado diferenciado (McNatty et al., 1979). En esta etapa, el folículo posee una cavidad o antro conteniendo fluido folicular, el cual es un exudado de plasma y está condicionado por los productos secretorios de las células de la granulosa. En este medio residen las células de la granulosa y el ovocito y las moléculas regulatorias entran y salen al microambiente (Brañez et al., 2004). El crecimiento del folículo se produce como respuesta a la acción de la FSH, la cual induce mitosis en las células de la granulosa, durante la cual se da una alta proliferación de células, característica del desarrollo del folículo dominante. Durante la fase folicular del ciclo, el número de células de la granulosa aumenta de 1 a 50 x 106 células en el estado preovulatorio. Cada folículo dominante tiene como destino completar la transición de pequeño (1-6 mm), mediano (7-11 mm), grande (12-17 mm), al estadío preovulatorio (18-23 mm). Un folículo atrésico falla de pasar del estado pequeño al mediano (1-10 mm) (Erickson, 2003). La teca interna contiene grandes células intersticiales típicas de células activas productoras de esteroides, en las cuales el citoplasma se llena de gotitas de lípidos. Las células de la teca intersticial poseen receptores para LH e insulina. En respuesta a su estimulación se producen altos niveles de andrógenos, mayormente androstendiona (Erickson, 1993). La teca interna está altamente vascularizada por una red capilar que rodea el foliculo de Graaf durante su crecimiento (Clark et al., 2004). 2.3.5. Folículo de Graaf En el folículo de Graaf, las células de la granulosa y ovocito están distribuidos en una masa de células de forma y posición precisa, la cual da cuatro subtipos de células de la granulosa: la membrana, el área periantral, el cumulus oophorus y la 18 corona radiada. Todas las células de la granulosa expresan receptores de FSH durante el desarrollo del folículo de Graaf, sin embargo, cada grupo de células de la granulosa está influenciada por su posición para un estado de diferenciación específica en respuesta a la estimulación de FSH. Por ejemplo, las células de la membrana de la granulosa expresan P450aromatasa y receptores de LH, mientras que el área periantral, cumulus y corona radiata, no (Erickson, 2003). Al final de la fase luteal, el folículo dominante es seleccionado de una cohorte de folículos clase 5 y la mitosis de la granulosa parece incrementarse. A medida que la fase folicular prosigue, el folículo dominante crece rápidamente, lo que no sucede con el crecimiento de otros folículos de Graaf de la cohorte, en los cuales el crecimiento procede más lentamente. El mecanismo de selección envuelve el pico secundario de FSH plasmática, por lo que los niveles de FSH permanecen elevados a través del primer pico de la fase folicular del ciclo. Se cree que el decremento de estradiol e inhibina activados por el CL son la causa del pico secundario de FSH en la selección del folículo dominante (Simoni et al., 1997). El pico secundario de FSH produce un progresivo aumento en los niveles del fluido folicular en el microambiente del folículo dominante, lo que provocaría la entrada de FSH en el fluido folicular para permitir una obligatoria inducción de la selección. Sin embargo, existe la interrogante de cuál es el mecanismo por el cual un folículo tiene la capacidad de concentrar altos niveles de FSH dentro de su microambiente (Erickson, 2000). En la selección del folículo dominante, la FSH juega un papel obligatorio que ningún otro ligando posee por sí mismo. El mecanismo primario por el cual la FSH controla la selección es por la estimulación del receptor y por las vías de transducción de la señal en las células de la granulosa. Aunque la LH no es esencial para la selección, tiene cierta importancia en la regulación de la formación del folículo dominante a través de su capacidad para estimular la expresión del sustrato de aromatasa androstendiona (Marsters et al., 2003). 19 La mitosis caracteristíca de esta etapa, es un periodo sostenido de alta proliferación de las células de la granulosa y es característico del desarrollo del folículo dominante. Sin embargo, existen numerosos factores de crecimiento que también lo estimulan (Tamura et al., 1995). A medida que el folículo dominante crece, la granulosa adquiere el potencial de adquirir grandes aumentos de estradiol. La inducción de FSH produce la expresión en las células de la granulosa de P450aromatasa, quen causa la adquisición del potencial de estrógenos en el folículo. La P450aromatasa es detectada en el folículo dominante; su actividad incrementa progresivamente, alcanzando niveles muy altos en la célula de la granulosa del folículo preovulatorio en la fase folicular (Inkster y Brodie, 1991). El tipo I 17ß-hidroxiesteoride dihidrogenasa (17ß-HSD) es expresado en las células de la granulosa en folículos del estadío primario al preovulatorio (Zhang et al., 1996). Por la expresión de P450aromatasa y 17ß-HSD, las células de la granulosa se vuelven altamente activas para convertir androstediona a estradiol. El progresivo aumento en el nivel de la expresión del gen CYP19 y P450aromatasa hace posible la expresión de receptores a LH. Durante la fase folicular, las células de la granulosa también adquieren el potencial de incrementar el aumento de la progesterona. Existen numerosos procesos operativos envueltos en la adquisición del potencial de luteinización (Erickson, 1994). La contínua estimulación de las células de la granulosa por la FSH está involucrada en este proceso progresivo. Cuando ocurre la luteinización, las células de la granulosa expresan grandes aumentos de proteína reguladora aguda de esteroides (Steroid Acute Regulatory Protein, StAR). A pesar de que el aumento progresivo en el potencial de luteinización ocurre, éste parece suprimirse justo antes de la ovulación. La inhibición es causada por el ovocito e inhibidores de luteinización presentes en el fluido folicular (Erickson y Shimasaki, 2000). El tipo I 20 17ß-HSD, parece ser expresado en las células de la granulosa en folículos del estadío primario al preovulatorio (Zhang et al., 1996). La cascada de eventos del proceso folicular completo involucra mecanismos de las células de la granulosa en los cuales éstas sufren un proceso de diferenciación completo que incluye la adquisición de los receptores para FSH (FSHR) en el estado temprano de crecimiento folicular. El folículo dominante contiene altos niveles de RNAm para receptores de gonadotropinas, principalmente para el receptor de hormona luteinizante (LHR), así como receptores de estrógenos, y proteínas y enzimas involucradas en la síntesis de andrógenos y P4. El aumento del receptor LHR en las células de la granulosa del folículo dominante incrementa la actividad aromatasa en respuesta a las gonadotropinas, presentándose en ellos un aumento en la producción de algunos productos génicos que aparecen en los folículos secundarios como resultado de la estimulación de FSH, siendo del RNAm para la enzima P450 aromatasa, responsable de la biotransformación de andrógenos en estradiol, (Montaño y Ruiz, 2005), lo cual provoca un cambio de dependencia de FSH a LH). Esto aumenta o mantiene la capacidad del folículo dominante para producir más estradiol que los folículos subordinados (Fortune et al., 2000). El pico de LH permite que el folículo ovulatorio responda a su estímulo inductivo, debido a que la ovulación envuelve la expresión de receptores de LH en las células de la granulosa. La FSH juega un papel vital en la inducción del receptor de LH en las células de la granulosa. Al igual que los factores antes mencionados, la expresión de los receptores de LH permanencen suspendidos hasta la última etapa de la fase folicular. Hay evidencia en animales de laboratorio de que los inhibidores derivados del ovocito inhiben también la inducción de los receptores para LH por parte de la FSH (Simoni et al., 1997). El mecanismo fisiológico por el cual el folículo dominante produce estradiol se llama concepto de dos células-dos gonadotropinas. La llegada de la LH a la teca 21 intersticial permite la síntesis y secreción de androstendiona, y el aumento en la secreción de andrógenos refleja la presencia de moléculas regulatorias incluyendo insulina, IGF-I, lipoproteinas, activina e inhibina (Erickson, 1993). En esta etapa están presentes los factores de crecimiento que modulan la teca como la insulina, IGF-I, lipoproteina, activina, inhibina, GDF-9, y BMP-15 (Yamamoto et al., 2002), FGF (Factor de crecimiento de fibroblastos) e IGFBP (Factor de crecimiento similar a la insulina unido a proteínas) (Knight y Glister, 2006). Se ha observado, que el folículo dominante adquiere bajas concentraciones intrafoliculares de IGFBP-15 y aumento de estradiol lo que sugiere que IGFBP-15 hace que aumente el IGF-1 libre y modifique las respuestas a FSH lo que confiere una ventaja al folículo elegido (Yamamoto et al., 2002). En el ovario, el IGF-I estimula la proliferación de células de la granulosa, promueve la esteroidogénesis, la folículogénesis, la ovulación, la fertilización, la implantación y el desarrollo embrionario. El receptor tipo I está en estas células en folículos saludables o atrésicos, ya sea de estados primarios o preovulatorios (Spicer et al., 2002). El IGF-I influye en la función reproductiva promoviendo la síntesis de receptores de FSH y de LH seguido por la esteroidogénesis y la producción de inhibina en las células foliculares. En la vía esteroidogénica, el IGF-I estimula el sistema aromatasa, e incrementa la sensibilidad de las células foliculares a FSH y LH (Spicer et al., 2002). El IGF-I puede mediar el efecto estimulador de FSH sobre la proliferación de las células de la granulosa, produciendo un aumento gradual del crecimiento folicular hasta la formación del antro (Henao et al., 2000). 2.4 OVULACIÓN Para prepararse para la ovulación, los pequeños folículos deben madurar hasta el estado preovulatorio, tiempo durante el cual el ovocito, células de la granulosa y células de la teca adquieren características funcionales específicas. Así, después 22 de un prolongado periodo de descanso, el ovocito en estado preovulatorio reinicia la meiosis, las células de la granulosa adquieren la habilidad de producir estrógenos y responder a la LH vía los receptores de LH, y las células de la teca comienzan a sintetizar más andrógenos que sirven como sustrato para la enzima aromatasa en las células de la granulosa (Richards, 1994). Durante la secuencia ovulatoria, el núcleo del ovocito o la vesícula germinal tienen una serie de cambios que incluyen la ruptura de la vesícula geminal (GVBD), y por consecuencia, la progresión de la meiosis a la segunda metafase meiótica o etapa de primer cuerpo polar (Sato et al., 2001). La meiosis se detiene aquí y proseguirá hasta que el óvulo sea fertilizado debido a las altas concentraciones de AMPc dentro del ovocito, las cuales pueden inhibir la maduración meiótica. Así, el pico LH/FSH de alguna manera causaría que el nivel de AMPc fallara en el ovocito, debido en parte a la desensibilización y regulación hacia abajo (down regulation) de los receptores a LH y FSH en la granulosa. Durante la maduración meiótica, las células del cumulus sufren una mucificación resultado del dramático aumento en la secreción de mucopolisacáridos hacia el espacio extracelular, seguida de una expansión. El pico preovulatorio de FSH provoca la expansión del cumulus (Zhou et al., 2001). Las células que rodean al ovocito y las células de la matriz del cumulus están formadas por tres componentes principales: ácido hialurónico (HA) y al menos dos proteínas unidoras de HA, llamadas Factor de Necrosis Tumoral, (tumornecrosis factor-stimulated gene) y Gen supresor de tumor (TSG)-6 (tumor suppressor gene) (Yoshioka et al., 2000). La expansión del cumulus es inducida como consecuencia del pico de LH, y es dependiente de la inducción de genes específicos (Hess et al., 1999). Quizás el cambio más dramático durante la ovulación es la formación de la abertura en la superficie del ovario, la mácula pelúcida o estigma. Durante este proceso, el folículo produce proteasas que degradan el tejido ovárico. El pico de 23 LH está directamente involucrado en la formación del estigma, ya que induce genes específicos como el de ciclooxigenasa-2 (COX-2), la cual interviene en la síntesis de prostaglandinas (Sirois et al., 1992). La prostaglandina E2 (PGE2) se produce en respuesta al pico de LH; el folículo ovulatorio produce progesterona, PGE2 y receptores de PGE2; ambas son necesarias para el desarrollo del estigma en animales de laboratorio, ya que la hembra que carece de genes de receptores para progesterona o ciclooxigenasa no ovula y es infértil (Lydon et al., 1995). Una vez que se produce la ovulación disparada por el pico preovulatorio de LH, el cual a su vez se induce por la elevada concentración circulante de estrógenos producidos por el folículo preovulatorio, se inducen marcados cambios en el folículo que incluyen tanto el reinicio de los procesos de maduración del ovocito, como alteraciones funcionales en el complejo ovocito-células del cumulus, y modificaciones en la expresión génica de las células de la granulosa que la llevarán hacia otros cambios que provocarán la ruptura de la pared folicular (Hess et al., 1999). De esta manera es posible explicar el mecanismo en cascada de la ovulación: el pico de LH de la mitad del ciclo estimula la expresión del receptor de progesterona (PR) y la producción de progesterona, los ligandos de progesterona interactúan con PR en las células del folículo, las cuales a su vez inducen el gen que codifica para COX-2 que actúa sobre el gen prostaglandin sintasa-2, el cual codifica la biosíntesis de prostaglandinas y genes asociados con el complejo cumulo, las cuales interactuan con receptores específicos en las células de la superficie epitelial del estigma presuntivo y activan las vías de señalización que permiten la liberación de enzimas lisosomales (proteolíticas), las cuales degradan las capas internas del tejido y causan o provocan que la superficie epitelial se separe de la base de la membrana, lo que resulta en la formación del estigma del complejo ovocito-cumulus (Lydon et al., 1995). 24 Después del proceso de ovulación, se produce un aumento de FSH mientras la LH continúa alta debido a su lento metabolismo en el hígado. Primeramente se forma un coágulo de fibrina donde se localizaba el antro y el líquido folicular, en el cual se pierde tejido conectivo y hay invasión de células sanguíneas. Las células que forman el CL provienen de la membrana de la granulosa, la teca interna y externa y tejido sanguíneo; por su parte, las células de la granulosa logran un gran tamaño y se transforman en células luteínicas de la granulosa que contienen ésteres de colesterol en el citoplasma. Las células luteínicas de la granulosa tienen una alta capacidad de producir progesterona y estradiol (Chappell et al., 1997). En esta etapa, la LH es requerida para el mantenimiento de la esteroidogénesis por las células luteinizadas de la granulosa, las cuales tienen un potente efecto luteotrópico sobre la producción de progesterona (Carr et al., 1982). La alta afinidad de los receptores de LH de las células luteales permite que el CL se mantenga activo pese a las concentraciones bajas de esta hormona durante el diestro. De esta manera, si la implantación no ocurre, el CL degenera por el proceso de luteolísis. El útero es el órgano que determina la regresión del CL a través de la secreción de la hormona luteolítica prostaglandina F2α (PGF2α) (Hafez et al., 2002). Los estrógenos tienen un efecto importante, siendo necesario que la progesterona haya actuado previamente sobre las células endometriales. Los estrógenos tienen una retroalimentación positiva sobre ellos mismos al aumentar sus propios receptores en el endometrio; además, producen el aumento de los receptores para oxitocina. En rumiantes, el CL está formado por células de la granulosa y de la teca interna. Posee receptores para oxitocina y para estradiol, sugiriendo que los estrógenos tendrían una acción directa sobre la lisis del CL (Erickson, 2003). El primer signo de la muerte del CL es la reducción de las células luteinizadas de la granulosa; eventualmente la luteólisis destruye todas las células del CL y se 25 transforma histológicamente en cuerpo albicans que es un nódulo de tejido conectivo denso (Erickson, 2003). Para que el desarrollo de la foliculogénesis ocurra ininterrumpidamente hasta la ovulación, es necesario el éxito en la unión hormona-receptor. La presencia de alteraciones en la interacción FSH-FSHR, provocadas por cambios en la estructura del gen que regula a FSH, pueden afectar la habilidad reproductiva, especialmente en hembras y mujeres (Gromoll et al., 1996). La FSH y su receptor en el folículo constituyen la unidad estructural y funcional de los ovarios, permitiendo la maduración folicular (Gigli et al., 2006). La influencia de FSH en la fase independiente de gonadotropinas puede afectar el mecanismo de los factores autocrinos y paracrinos entre el ovocito y las células de la granulosa, los cuales permiten el crecimiento de folículos en los primeros estadíos de su desarrollo (Thomas y Vanderhyden, 2006). 3. PAPEL DE LA FSH EN LA OVULACIÓN MÚLTIPLE 3.1 SINCRONIZACIÓN DEL CELO Uno de los aspectos más importantes y que probablemente tiene mayor peso en los parámetros reproductivos en la ganadería tropical es la deficiente detección de celos. Porcentajes del 36% en la detección de calores reportados en México, contrastan con datos de explotaciones ubicadas en climas templados, cuyos porcentajes varían entre un 40 y un 65% (Favero et al., 1995). Entre las herramientas de manejo reproductivo que permiten eficientizar la inseminación artificial (IA) se encuentran la sincronización de celos y la sincronización de ovulaciones. Los métodos hormonales de sincronización se basan en el efecto luteolítico de la prostaglandina F2 alfa o el efecto lúteo de los progestágenos. La prostaglandina sólo funciona en animales ciclando y con CL, y son abortivas en animales gestantes. El empleo de progestágenos (progesterona 26 o sus análogos) se fundamenta en la acción inhibitoria de la progesterona para la manifestación del celo. Al administrarlos en dispositivos vaginales (durante 7 días), implantes subcutáneos (9 días), o por vía oral (14 días), actúan como un CL artificial, y mientras ejercen su acción la vaca no manifiesta estro. Al retirarlos, permiten la presentación del celo. En animales que están en anestro logran inducir el estro, y en animales ciclando funcionan en cualquier etapa del ciclo estrual. Se puede emplear IA a tiempo fijo después de terminar la administración del progestágeno, pero se obtiene mejor fertilidad si se insemina a celo detectado (AM-PM) (Asprón, 2004). Los animales sometidos a inducción de OM son sincronizados previamente al tratamiento de OM. Si se aplican correctamente, dentro de las limitantes fisiológicas debidas a sus mecanismos de acción, las técnicas de sincronización de celos son altamente exitosas. Sin embargo, se pueden presentar variaciones en la dinámica de las ondas foliculares que dificultan controlar de manera precisa el momento del celo y de la ovulación (Mapletoft et al., 2002). La detección del celo en la donadora es un importante factor a tomar en cuenta en un programa de transferencia de embriones (TE). Diversos estudios han determinado una amplia variación en la duración del estro (3 a 26 h), atribuida entre otros factores a la raza y la época del año. Contrariamente, la variación de la duración del ciclo estrual es menor, y en promedio dura 21 días (Góngora y Hernández, 2006). El celo se caracteriza por la receptividad sexual de la vaca y la aceptación de la monta, debido a la concentración de estradiol presente en esta etapa del ciclo, siendo éste el responsable de la inducción del comportamiento del celo. El celo es más corto y menos evidente en los animales Bos indicus, comparados con Bos taurus, en los cuales la duración del celo varía entre 3 y 26 h, con un promedio de 14 h (Rae et al., 1999), mientras que en los animales cebú el rango es de 2 a 22 h, con un promedio de 7 h (Hansen, 2004). La duración del ciclo estrual en animales 27 con tres ondas de crecimiento folicular parece ser más corta en el ganado cebú (<21 días) que en las razas europeas y en el ganado híbrido Bos taurus x Bos indicus (22 a 25 días, Portillo, 2005). Los genotipos Bos indicus poseen una capacidad reducida en la secreción de LH. De igual manera, el pico de LH y la ovulación ocurren en forma mas temprana en relación con el estro en este tipo de ganado comparado con animales Bos taurus (Richards, 1988). Los ciclos de corta duración ocurren normalmente en la primera ovulación espontánea posparto en la mayoría de las vacas de carne (Day et al., 1990). La liberación prematura de PGF2α por el útero (Cooper et al., 1991), como resultado de los efectos de la baja progesterona (Zollers et al., 1993) y estradiol (Mann y Lamming, 1999) antes de la ovulación en los receptores de oxitocina y progesterona endometriales, parecen ser la causa de la temprana regresión del CL. La progesterona producida durante la fase luteal corta puede ser necesaria para el establecimiento de una función luteal normal en el siguiente ciclo estral (Mihm, 1999). El comienzo de la actividad de celo sigue diferentes patrones, con la mayoría de la actividad durante las últimas horas de la tarde, a lo largo de la noche, y en las primeras horas de la mañana. Las investigaciones muestran que más del 70% de la actividad de monta toma lugar entre las 7:00 de la noche y las 7:00 de la mañana (Sepúlveda y Rodero 2003). Tanto en el estro natural como en el inducido hormonalmente, se ha detectado que la presencia en el hato de más de una hembra en celo desencadena una interacción sexual de montas y otras conductas estimulantes del reclutamiento e intervención sexual de otras vacas, con la consiguiente formación transitoria y móvil de grupos homosexuales o heterosexuales, conocidos como grupos sexualmente activos (Orihuela, 2000). 28 Debido a esta variabilidad en la presencia de celo, la inducción de OM incluye la inyección de PGF2α para la regresión del CL y presentación del celo posterior a la OM, para reducir la variabilidad entre la inyección y la presencia de estro (Seidel, 1981). Existen sin embargo diversos factores que pueden afectar la presencia de celo aún con tratamientos de inducción de OM, como son el efecto del estrés calórico y nutricional (Cobanov y Schnitkey 2003). El estrés calórico y nutricional inhiben el desarrollo del folículo dominante durante el periodo preovulatorio en vacas. Como consecuencia se reducen también las concentraciones séricas de estradiol y se retrasa la declinación de la fase luteal tardía lo que incrementa la presencia de tres ondas de crecimiento folicular (Gilad et al., 1993). 3.2 TRATAMIENTO HORMONAL El conocimiento de la fisiología del ciclo estrual permite la utilización sistemática de hormonas para mejorar la reproducción y la eficiencia reproductiva de los animales. De esta manera es posible utilizar tratamientos hormonales para inducir la OM cuando se utilizan programas de TE, los que permiten utilizar de manera intensiva a hembras genéticamente superiores. La tecnología de la TE en bovinos requiere de la selección y el manejo, tanto físico como farmacológico, de las donadoras y las receptoras, y también de la recolección y transferencia de los embriones dentro de un periodo de tiempo corto y especifico después del estro (Mapletoft, 2002). En la actualidad, las investigaciones están orientadas al estudio en la reducción de la alta variabilidad reproductiva de la TE, especialmente en lo que se refiere a la respuesta individual a los tratamientos para OM, existiendo diversos protocolos de tratamientos hormonales mediante gonadotropinas para inducir la OM, los cuales no han podido disminuir la amplia variabilidad que se presenta en el número de ovulaciones entre animales (Cushman et al., 1999). 29 De una hembra bovina bien alimentada durante toda su vida reproductiva se espera obtener aproximadamente 10 crías nacidas. Con la técnica de OM se pueden obtener aproximadamente 15 embriones cada "mes" de una sola hembra, y con la técnica de TE éstos se pueden transferir a varias receptoras para obtener hasta 10 becerros por "mes" del mismo animal (Calva et al., 2001). La variación en el número de folículos presentes en el ovario capaces de responder al inicio del tratamiento de OM señala la diferencia en la respuesta. Estudios in vitro han demostrado que la FSH puede acelerar el rango de desarrollo de los folículos preantrales (Gutiérrez et al., 2000), permitiendo mejorar la fertilidad e incrementar la respuesta a la OM (Lucy et al., 1992). La OM es la inducción de más de una ovulación, mediante la aplicación de hormonas que estimulan los folículos antrales produciendo un crecimiento intensivo, con la finalidad de que al momento de la IA, se produzca más de un embrión (Armstrong, 1993). Se utilizan FSH ó LH, las cuales al contrarrestar los efectos de los estrógenos hacen que en ambos ovarios de la vaca se desarrollen varios folículos dominantes. Actualmente, los tratamientos de inducción de OM utilizan inyecciones de gonadotropinas exógenas en el diestro, en la mitad de la fase lútea (día 8 a 14), para reclutar folículos extra cerca del final de esta fase para que sean ovulados en la siguiente fase folicular. Para que el tratamiento sea exitoso, se deberá tomar la segunda oleada folicular, que es cuando hay mayor número de folículos en crecimiento; ésta ocurre el día 9 ó 10 del ciclo en aquéllos que tienen dos oleadas, y entre el día 8 a 9 en los que tienen tres (Mapletoft et al., 2002). Cuando se utiliza FSH, la aplicación inicia en el día 10 despúes de la detección de celo, y debido a que la vida media biológica de la FSH suprafisiológica en la vaca es de 5 horas ó menos, ésta debe ser inyectada dos veces al día para que la inducción sea exitosa (Mapletoft et al., 2002). Las preparaciones de FSH generalmente se aplican en series de 8 inyecciones a dosis decrecientes por 4 30 días, comenzando cerca del día 10 del ciclo estrual; con este método se ha reportado mayor producción de embriones transferibles que con otras hormonas, como la gonadotropina coriónica equina (eCG) (Crister et al., 1980). Los tratamientos para inducción de OM en algunos casos causan un dramático aumento en el número de folículos que desarrollan los ovarios, así como en el número de óvulos disponibles para ser liberados al momento de la ovulación (Mapletoft et al., 2002). Se ha reportado también que la raza influye en los resultados (Rodríguez et al., 2007). El alto grado de variación en la respuesta crea problemas que afectan tanto a la eficiencia como al beneficio de los programas de TE (Hasler, 1992). Mapletoft et al. (2002), demostraron que existe variabilidad entre animales y también en el mismo animal entre un tratamiento y otro, por lo que parece lógico pensar que la variabilidad inexplicable proviene de diferencias genéticas entre animales que determinan cómo ellos responderán a dosis suprafisiológicas de gonadotropinas. La variabilidad en la respuesta ovárica ha sido relacionada a diferencias en los tratamientos de OM, tales como la preparación de gonadotropina utilizada, la dosis total, la duración y tiempo del tratamiento y el uso de hormonas adicionales en el esquema de tratamiento. Sin embargo, existen factores adicionales que pueden ser fuente importante de la variabilidad y son inherentes al animal y a su medio ambiente, e incluyen estatus nutricional, historia reproductiva, edad, estación del año, raza y estado ovárico al momento de aplicar el tratamiento, así como el efecto de OM repetidas, entre otros (Mapletoft et al., 2002). Numerosos hatos están expuestas a resultados negativos; sin embargo, los resultados pueden indicar la presencia de cambios en la actividad genética. Esto significa que algunos animales parecen ser resistentes a los tratamientos y algunos no, y que la selección de animales resistentes a los tratamientos (con la posibilidad de pruebas genéticas) es la estrategia para eliminar este problema en 31 los hatos, lo que permitiría la selección de hembras con alta respuesta a la aplicación de FSH (Nett et al., 2007). Moor et al. (1984) sugirieron que tanto la tasa de ovulación como el número de embriones viables producidos son relativamente consistentes dentro de los mismos individuos. Aunque hay gran variabilidad entre vacas, se ha visto que el número de folículos es similar en los ovarios de la misma vaca. Además, el número de folículos >1.7 mm de diámetro en un ovario se correlacionó positivamente con la respuesta de OM a los tratamientos con gonadotropinas. Singh et al. (2004) demostraron que el número de folículos presentes en el momento de la emergencia de la onda folicular permite predecir la respuesta a la OM. En conjunto, esa información puede sugerir que algo de la variabilidad reside en la estructura genética o fisiológica del animal, más que en factores exógenos. Las vacas y novillonas seleccionadas por producción de gemelos tienen mayores respuestas de OM que otras hembras (Mapletoft et al., 2002). También, el microambiente folicular puede afectar la calidad del ovocito, su habilidad para ser fertilizado y el subsecuente desarrollo embrionario, lo que demuestra que la dinámica endocrina folicular se ve influida por el balance endocrino y los perfiles hormonales de la donadora (Greve y Callesen, 2005). La variabilidad individual ha sido un factor prevalente en todos los estudios de OM, demostrándose que la raza puede ser otro factor a considerar (Alkemade et al., 1993). Por otro lado, se ha demostrado que las temperaturas del invierno pueden se factores de estrés para el ganado Bos indicus (Mapletoff et al., 2002). CONCLUSIONES La FSH tiene un papel primordial en la reproducción de la hembra bovina, ya que interviene en el crecimiento y desarrollo de los folículos ováricos. La FSH, por tanto, es usada para inducir OM en estos animales, con el fin de obtener un número de folículos mayor al que se presenta de manera normal en un ciclo, y con 32 esto lograr la ovulación de más de un óvulo por ciclo estrual por vaca, resultando en la obtención de más de un embrión por hembra cada vez. LITERATURA CITADA 1. Adashi, E. y Hennebold, J. (1999). Mutaciones de un solo gen que llevan a la disfunción reproductiva femenina. New. Eng. J. Med. 340(9):709-718. 2. Antezak, M. y Van Vlerkom, J. (2000). The vascullarcharacter of ovarian follicular granulosa cells. Phenotypic and functional evidence for an endothelial-like cell population. Hum. Reprod. 15(11):306-2318. 3. Alkemade, S., Murphy, B., y Mapletoft, R. (1993). Superovulation in the cow; Effects of biological activity of gonadotropins. Theriogenology, 2003, 13 (2):315-320. 4. Armstrong, D. (1993). Recent advances in superovulation of cattle Theriogenology, vol. 39 (1) 724. 5. Ascoli, M. y Segaloff, D. (1996). Hormonas adenohipofisarias y sus factores liberadores hipotalámicos” Sección XIII. Cap.55 pp 1447-1467 En: Goodman & Gilman (1996) “Las bases farmacológicas de la terapéutica” Volumen 1. 9ª Edición Editorial Panamericana. 6. Asprón, M. (2004). Manejo Reproductivo del Ganado Bovino. Curso de Actualización. International Veterinary Information Service, Ithaca NY. 7. Brañez, M., Sáez, A., Villalón, M. y Sáez, J. (2004). Uniones en hendidura y su papel funcional en el tracto reproductor femenino. Rev. Chil. Obstet. Ginecol., 69 (1): 60-66. 8. Calandra, R. y De Nicola, A. (1985). “Endocrinología molecular” 2º edición. Editorial El Ateneo. En: Echeverras, J. Endocrinología reproductiva: Prostaglandina F2 en vacas. Revision bibliografía. Revista Electrónica de Veterinaria REDVET. 9. Calva, R., Cortés, F., Aja, G., Ortiz, C. y Cevenini, R. (2001). La transferencia de embriones. Una técnica de mejoramiento animal en ganado de lidia. Cir Ciruj 69: 129-134. 10.Campbell, B, Telfer, E., Webb, R. y Baird, D. (2004). Evidence of a role for follicle-stimulating hormone in controlling the rate of preantral follicle development in sheep. Endocr. 145:18701879. 11.Carroll, R., Kowash,P., Lofgren,J., Schwall,R. y Chin, W. (1991). In vivo regulation of FSH síntesis by inhibin and activin. Endocr. 129: 3299-3304. 12.Carr, B., MacDonald, P. y Simpson, E. (1982). The role of lipoproteins in the regulation of progesterone secretion by the human corpus luteum. Fertil and Sterility 38:303-11 13. Clark, L., Irving-Rodgers., H. Dharmarajan. A. y Rodgers, J. (2004). Theca Interna: The Other Side of Bovine Follicular Atresia. Biol of Reprod: 71 (4):1071–1078. 14. Chang, H., Brown, C. y Matzuk,. M. (2002). Genetic analysis of the mammalian transforming growth factor-ß superfamily. Endocrine Rev. 23:787-23 (26). 15.Chappell, P. Lydon, J. Conneely, O., O'Malley, B. y Levine, J. (1997). Endocrine defects in mice carrying a null mutation for the progesterone receptor gene. Endocrinology 138:4147-4152 16.Combelles, C., Carabatsos, M., Kumar, T., Matzuk, M. y Albertini, D. (2004). Hormonal control of somatic cell oocyte interactions during ovarian follicle development Mol. Reprod. Dev.69:347355. 17.Cooper, D., Carver, D., Villeneuve, P., Silvia, W. and Inskeep, E. (1991) Effects of progestogen treatment on concentrations of prostaglandins and oxytocin in plasma from the posterior vena cava of post-partum beef cows. J. Reprod. Fertil. 91, 411-442. 18. Cobanov, B, and Schnitkey, G. (2003). Economic Losses from Heat Stress by US Livestock Industries1. J. Dairy Sci. 86: (E. Suppl.):E52–E77. 19.Crister, J., Rowe, R., del Campo, M. y Ginther, O. (1980). Embryo transfer in catlle: factors affecting superovulatory response, number of transferable embryos, and length of posttreatment estrous cycles. Theriogenology, 13: 397-406. 33 20. Cushman, R., De Souza, J., Hedgpeth, V. y Britt, J. (1999). Superovulatory Response of One Ovary Is Related 69.to the Micro- and Macroscopic Population of Follicles in the Contralateral Ovary of the Cow. Biol. Reprod. 60: 349-354. 21.Day, M., Dyer, R., Wilson, G. y Pope, W. (1990). Influence of estradiol on duration of anestrus and incidence of short estrous cycles in postpartum cows. Dom. Anim. Endocrinology 7, 19-25. 22.Diedrich, A., Ram Sairam, M., Monaco, l., Fimia G., Gansmuller, A., LeMeur, M., Sassone-Corsi, P. (1998). Impairing follicle-stimulating hormone (FSH) signaling in vivo: Targeted disruption of the FSH receptor leads to aberrant gametogenesis and hormonal imbalance. Cell. Biol. 95: 13612 - 13617. 23.Donadeu, F. y Ascoli, M. (2005). The differential effects of the gonadotropin receptors on aromatase expression in primary cultures of immature rat granulosa cells are highly dependent on the density of receptors expressed and the activation of the inositol phosphate cascade. Endocrinology. 146 (9):3907-3916. 24. Durlinger, A., Visser, J. y Themmen, A. (2002). Regulation of ovarian function: the role of antiMüllerian hormone. Reproduction 124:601-609. 25.Eppig, J. (1994) Oocyte-somatic cell communication in the ovarian follicles of mammals. Seminars in Developmental Biology 5:51-59 26.Eppig, J. (2001). Ocyte control of ovarian follicular development and fuction in mammals. Reproducción 122: 829-838. 27. Eppig, J. (2003). Growth and development of the mammalian oocyte-granulosa cell complex in culture. Chapter 17. In: Biology and pathology of the oocyte: its role in fertility and reproductive medicine / edited by Alan O. Trounson, Roger G. Gosden. Cambridge, UK ; New York : Cambridge University Press. 28. Erickson, G. (1993). Normal regulation of ovarian androgen production. Seminars in Reproductive Endocrinology 11:307-312. 29. Erickson, G. (1994). Polycystic Ovary Syndrome: Normal and Abnormal Steroidogenesis. In ovarian Endocrinopathies: Proceedings of the 8th Reinier deGraaf Symposium. 103-115. Schats R. and Schoemaker J (eds): Ovarian Endocrinopathies:New York. Parthenon Publishing. 30. Erickson, G. (2000). The graafian follicle: A functional definition. In: Adashi EY (ed) Ovulation: Evolving Scientific and Clinical Concepts. Springer-Verlaag, New York. 31-48. 31.Erickson, G. (2003) Morphology and physiology of the ovary. In: Rebar, R. Ed. Female reproductive endocrinology morphology and physiology of the ovary. Chapter 2. 32.Erickson, G. y Shimasaki, S. (2000). The role of the oocyte in folliculogenesis. Trends in Endocrin. and Metabol. 11:193-198. 33.Erickson, G., Magoffin, D., Dyer, C. y Hofeditz. (1985). The ovarian androgen producing cells: A review of structure/function relationships. Endocrin. Rev. 6:371-399. 34. Espinoza, J., Ortega, R., Palacios, A., Valencia, J. y Arechiga, C. (2007). Crecimiento folicular ovárico en animales domésticos. Una revisión. Interciencia 32: 93-99. 35.Favero, R., Faulkner, D., Nash, T. y Kesler, D., (1995). Effect of Norgestomet treatment after insemination on the calving rate of postpartum suckled beef cows. J. Anim. Sci. 73: 3230 – 3234. 36.Findlay, J., Drummond, A., Dyson, M., Baillie, A., Robertson, D., Palacios, A., y Ethier J. (2002). Recruitment development of the follicle; the roles of the transforming growth factor-ß superfamily. Mol. and Cell. Endocrin. 191:35-43. 37.Fortune, J., Cushman, R., Wah, C., y Kito, S. (2000). The primordial to primary follicle transition. Molecular and Cellular Endocrinology 163:53-60. 38.Gigli., I., Russo, A. y Agüero A. (2006). Consideraciones sobre la dinámica ovárica en equino, bovino y camélidos sudamericanos. InVet. 8, núm. 1. 39.Gilad, E., Meidan, R., Berman, A., Graber, Y., y Wolfenson, D. (1993). Effect of heat stress on tonic and GnRH-induced gonadotrophin secretion in relation to concentration of oestradiol in plasma of cyclic cows. J. Reprod. Fertil. 99 (2): 315-21. 40.Ginther, O., Kot, K., Kulick, L. y Wiltbank, M. (1997). Sampling follicular fluid without altering follicular status in cattle: oestradiol concentrations early in a follicular wave. J Reprod Fertil 109 (2):181-186. 34 41.Glister, C., Tanneta, D., Groome, N. y Knigth, P. (2001). Interactions between follicle-stimulating hormone and growth factors in modulating secretion of steroids and inhibin-related peptides by nonluteinized bovine granulosa cells. Biol. Reprod. 65(4): 1020-1028. 42. Góngora, A. y Hernández, A. (2006). Comportamiento sexual, duración del estro y del ciclo estral en novillas criollas sanmartineras y brahman del piedemonte llanero Colombiano. Liv. Res. Rural Develop. 18 (1):58 43.Graus, S., Naor, Z. y Seger, R. (2001). Intracellular signaling pathways meadited by the gonadotropin-releasing hormone (GnRH) receptor. Arch.. Med Res. 32, 499-509. 44. Greve, T. y Callesen, H. (2005). Embryo technology: implications for fertility in Cattle. Rev. Sci. Tech. 24 (1), 405-412. 45.Gromoll, J., Simoni, M., Nordhoff, V., Behre, H., De Geyter. C. and Nieschlag, E. (1996). Functional and clinical consequences of mutations in the FSH receptor. Mol. Cell. Endocrin. 125(1-2):177-82. 46.Gutierrez, C., Ralph, J., Telfer, E., Wilmut, I. y Webb, R. (2000). Growth and antrum formation of bovine antral follicles in long-term culture in vitro. Biol. Reprod. 62:1322–1328. 47. Hafez, E. (2002). Reproducción e Inseminación Artificial en Animales. McGraw-Hill. 7ª edición. México. 48. Hansen, P., (2004). Physiological and cellular adaptations of cebu cattle to thermal stress. An. Reprod. Sc. 82-83, 349-360. 49.Hasler, J. (1992). Current Status and Potential of Embryo Transfer and Reproductive Technology in Dairy Cattle. J. Dairy Sci. 75: 2857-2879. 50. Henao, G., Trujillo, L. y Vásquez, J. (2000). Cambios en la dinámica folicular en vacas cebú anéstricas sometidas a suspensión temporal de la lactancia. Rev. Col. Cienc. Pec. Vol. 13:2, p.121-129. 51. Hess, K., Chen L., Larsen W. (1999). Inter- -inhibitor binding to hyaluronan in the cumulus extracellular matrix is required for optimal ovulation and development of mouse oocytes. Biol. Reprod. 61: 436–443. 52.Hiroyuki, K., Hisashi, K., Gen, W., Kazuyoshi, T., Shuji, S., y Yoshihisa, H. (1995). Changes in Plasma Concentrations of Immunoreactive Inhibin, Estradiol and FSH Associated with Follicular Waves during the Estrous Cycle of the Cow. J. Reprod. Develop. 41 (4): 311-320. 53.Inkster, S. y Brodie, A. (1991). Expression of aromatase cytochrome P-450 inpremenopausal and postmenopausal human ovaries: an immunocytochemical study. J. Clin. Endocrin. Metabol. 73:717-26. 54.Johnson, J., Canning, J., Kaneko, T., Pru, J. y Tilly, J. (2004). Germline stem cells and follicular renewal in the postnatal mammalian ovary. Nature 428: 145-150. 55.Kilgour, R., Pisselet, C., Dubois, M. y Courot, M. (1998). Ram lambs need FSH for normal testicular growth, Sertoli cell numbers and onset of spermatogenesis. Reprod. Nutr. Dev. 38: 539-550. 56.Kishi, H., Minegishi, T., Tano, M., Kameda, T., Ibuki, Y. y Miyamoto, K. (1998). The effect of activin and FSH on the differentiation of rat granulose cells. FEBS Lett 422:274-278. 57.Knigth, P. y Glister, C. (2001). Potential local regulatory functions of inhibins, activins and follistatin in the ovary. Reproduction 121: 503-512. 58.Kroeze, W., Sheffler, D. y Roth, B. (2003). "G-protein-coupled receptors at a glance". J Cell Sci 116: 4867–4869. 59.Lucy, M., Savio, J., Badinga, L., De La Sota, R. y Thatcher, W. (1992). Factors that affect ovarian follicular dynamics in cattle. J. Anim. Sci. 70: 3615-3626. 60.Lussier, J., Matton, P. y Dufour, J. (1987). Growth rates of follicles in the bovine ovary. J. Reprod. Fertil. 81: 301-307. 125. 61.Lydon, J., DeMayo F., Funk, C., Mani, S., Hughes, A., Montgomery, C., Shyamala, G., Conneely, O. y O'Malley, B. (1995). Mice lacking progesterone receptor exhibit pleiotropic reproductive abnormalities. Genes and Develop. 9:2266-78. 62. Mann, G., and Lamming, G. (1999) The influence of progesterone during early pregnancy in cattle. Reproduction of Domestic Animals. 34, 269-274. 63.Mapletoft, R., Bennett, K. y Adams, G. (2002). Superovulation in Perspective Western College of Veterinary Medicine, University of Saskatchewan, Saskatoon, Saskatchewan, S7N 5B4, and 35 BIONICHE Animal Health, Belleville, ONJ2. Canada http://www.bionicheanimalhealth.com/Uploads/Superovulation%20in%20Perspective 64.Marsters, P., Kendall, N. and Campbell, B. (2003). Temporal relationships between FSH receptor, type 1 insulin-like growth factor receptor, and aromatase expression during FSHinduced differentiation of bovine granulosa cells maintained in serum-free culture.Mol Cell Endocrinol. 30;203(1-2):117-127. 65.McNatty, K., Moore-Smith, D., Osathanondh, R., y Ryan, K. (1979). The human antral follicle: Functional correlates of growth and atresia. Anns. de Biol. Anim. Bioch. Biophys. 19:1547-1558. 66. Mihm, M. (1999). Delayed resumption of cyclicity in postpartum dairy and beef cows. Reprod. Domes. Anim. 34: 276-284. 67. Montaño,, E., y Ruiz Z. (2005). ¿Por qué no ovulan los primeros folículos dominantes de las vacas cebú posparto en el trópico colombiano? Rev. Col. Cienc. Pec. Vol. 18:2, 2- 127. 68.Moor, R., Kruip, A. y Green, D. (1984). Intraovarian control of folliculogenesis: limits to superovulation? Theriogenology 21: 103-116. 69. Mukasa-Mugerwa E. (1989). A review of a productive performance of female Bos indicus (zebu) cattle. ILCA Monograph6. ILCA, Addis Ababa, Ethiopia. 70.Nett, T., Sawyer, H. y Niswender, G. (2007). Reproductive performance indomestic ruminants project COL00220. Colorado AES Projects 2006-2007 Biomedical Sciences 71. Orihuela A. 2000. Some factors affecting the behavioral manifestation of oestrus in cattle: A review. Appl. Anim. Behaviour Sci. 70: 1-16. 72. Ownby, Ch. (2004.). Female Reproductive System. En: http://www.cvm.okstate.edu/instruction/mm_curr/histology/fr/HiFRp15.htm 73.Packer, A., Hsu, Y., Besmer, P. y Bachvarova, R. (1994). The ligand of the c-kit receptor promotes oocyte growth. Developmental Biology 161:194-205. 74. Parrott, J. y Skinner, M. (1999). Kit-ligand/stem cell factor induces primordial follicle development and initiates folliculogenesis. Endocrinology 140:4262-4271 75.Pearly, S., Buchan, A., Hsueh, A., Ho Yuen, B. y Leung P. (2002). Intracellular calcium mobilization in response to the activation of human wild-type and chimeric gonadotropin receptors. Endocrinology 143: 1732-1740, 76. Perry, G. (2004). The bovine estrous cycle. College of Agriculture & Biological Sciences, South Dakota State niversity. http://agbiopubs.sdstate.edu/articles/FS921A.pdf 77.Pierce J., y Parsons T. (1981). Glycoprotein hormones: structure and function. Annu. Rev. Biochem. 50, 465–495. 78.Pineda, M. and Dooley, M. (2003). McDonald's veterinary endocrinology and reproduction. Ames, Iowa: Iowa State Press. 79.Portillo, G. (2005). Fisiología reproductiva y diferencias reproductivas entre el ganado europeo y cebú. Manual de Ganadería Doble Propósito. 415-418 http://www.avpa.ula.ve/docuPDFs/libros_online/manual-ganaderia/seccion6/articulo2-s6.pdf 80.Rae, D., Chenoweth, P., Giangreco, M., Dixon, P. y Bennett, F. (1999). Assessment of estrus detection by visual observation and electronic detection methods and characterization of factors associated with estrus and pregnancy in beef heifers. Theriogenology 51: 1121-1132. 81.Recabarren, S., Muñoz, P., Lobos, C., Vilches, C., Parilo, J. (2006). Análogo de GnRH disminuye la secreción de hormona folículo estimulante (FSH) en ovejas prepúberes. Arch. Med. Vet. 38,(1):39-45. 82.Richards, M., Geisert, R., Rice, L., Buchanan, D. y Castree, J. (1988). Influence of synchromate-B and breed composition on estrous response and pregnancy rate in spring and fallbreed Brahman crossbred beef cows. Theriogenology. 29:951-960 83. Richards, J. (1994). Hormonal control of gene expression in the ovary. Endocr. Rev. 15: 725751. 84.Richards, J., Russel, l., Ochsner, S., Hsieh, M., Doyle, K., Falender, E., Lo, K., Sharma, S. (2002). Novel Signaling Pathways That Control Ovarian Follicular Development, Ovulation, and Luteinization. Recen.t Prog. Horm. Res. 57: 195-220 85.Rodríguez, J., Giraldo, C., Castañeda, S., Ruiz, T. y Olivera, M. (2007). Análisis Multifactorial de las Tasas de Preñez en Programas de Transferencia de embriones en Colombia. Rev. MVZ Córdoba 12(2):978-84 36 86.Rosell, R., Rodríguez, R., Grimón, M., Ceiro, S. y Miranda, G. (2004. Eficiencia de la transferencia de embriones. Facultad de Medicina Veterinaria. www.veterinaria.org, revista electrónica de veterinaria redvet 7(4) s/p 87.Saez, J., Berthoud, V., Branes, M., Martinez, A. y Beber, E. (2003). Plasma membrane channels formed by connexins: their regulation and functions. Physiol Rev; 83(4):1359-400. 88.Sato, H., Kajikawa, S., Kuroda, S., Horisawa, Y., Nakamura, N., Kaga, N., Kakinuma, C., Kato, K., Morishita, H., Niwa, H. and Miyazaki, J.(2001). Impaired fertility in female mice lacking urinary trypsin inhibitor. Biochem. Biophys. Res. Commun. 281: 1154–1160. 89.Seidel G. Jr. (1981). Superovulation and embryo transfer in cattle. Science 211: 351-358. 90. Sepulveda, N. y Rodero, E.(2003). Comportamiento sexual durante el estro en vacas lecheras. INCI. vol. 28, no.9, p.500-503. 91.Simoni, M., Gromoll, J. y Nieschlag, E. (1997). The Follicle-Stimulating Hormone receptor: biochemistry, molecular biology, physiology, and Pathophys. Endocrin. Rev. 18 (6): 739-773. 92.Singh, J., Dominguez, M., Jaiswal, R., (2004). A simple ultrasound test to predict superstimulatory response in cattle. Theriogenology; 62:227-243. 93.Shemesh, M., Mizrachi, D., Gurevich, M., Stram, Y., Shore, L. y Fields. M. (2001). Functional importance of bovine myometrial and vascular LH receptors and cervical FSH receptors. Seminars Reprod. Med. 19:87-96. 94.Sirois, J., Simmons, D. y Richards J. (1992). Hormonal regulation of messenger ribonucleic acid encoding a novel isoform of prostaglandin endoperoxide H synthase in rat preovulatory follicles. J Biol Chem 267: 11586–11592. 95.Spicer, L., Chamberlain, C. y Maciel, S. (2002). Influence of gonadotropins on insulin- and insulin-like growth factor-I (IGF-I)-induced steroid production by bovine granulosa cells. Domest. Anim. Endocrinol. 22: 237-254. 96.Tamura, M., Sasano, H., Suzuki, T., Fukaya, T., Funayama, Y., Takayama, K., Takaya, R. y Yajima, A. (1995). Expression of epidermal growth factors and epidermal growth factor receptor in normal cycling human ovaries. Hum Rep 10:1891-1896 97.Tano, M., Minegish,i T., Nakamura, K., Nakamura, M., Karino, S., Miyamoto, K. y Ibuki, Y. (1995). Regulation of follistatin messenger ribonucleic acid in cultured rat granulosa cells. Mol. Cell. Endocrinol.109:167-174. 98.Thomas, F. y Vanderhyden, B. (2006). Oocyte-granulosa cell interactions during mouse follicular development: regulation of kit ligand expression and its role in oocyte growth. Reprod. Biol. Endocrin. 4:19. 99.Velásquez, D. y Mendieta, M. (2005). Factores que regulan el desarrollo folicular II: folículos antrales. Encuentros en la Biología 101: 4-6. 100.Wangsness, P., Acker, W., Burdette, J., Krabill, L. y Vasilatos, R. (1981). Effect of fasting on hormones and metabolites in plasma of fast-growing, lean and slow-growing obese pigs. J. Anim Sci. 52(1):69-74. 101.Webb, R., Marshall, F., Swann, K., y Carroll, J. (2002). Folliclestimulating hormone induces a gap junction-dependent dynamic change in [cAMP] and protein kinase A in mammalian oocytes. Dev. Biol. 246(2): 441-454. 102.Whittier, J. (2003). Reproductive Anatomy and Physiology of the Cow. University of MissouriColumbia. County and regional extension centers. http://muextension.missouri.edu/explore/agguides/ansci/g02015.h 103.Wilson, S., Kirby, C., Koenigsfeld, A., Keisler, D and Lucy, M. (1998) Effects of Controlled Heat Stress on Ovarian Function of Dairy Cattle. 2. Heifers1. J Dairy Sci 81:2132–2138. 104.Woodruff, T., D'Agostino, J., Schwartz, N. y Mayo, K. (1988). Dynamic changes in inhibin messenger RNAs in rat ovarian follicles during the reproductive cycle. Science 239:1296-1299. 105.Wrigth, P., y Malmo, J. (1992). Pharmacology manipulation of fertility. Vet. Clin. North. Am.: Food Anim. Pract. 8 (1): 57-89. 106.Yamamoto, N., Christenson, L., McAllister, J. and Strauss, J., (2002). Growth differentiation factor-9 inhibits 3'5'-adenosine monophosphate-stimulated steroidogenesis in human granulosa and theca cells. J. of Clin. Endocrin. and Metabo.l 87:2849-2856. 107.Yoshioka, S., Ochsner ,S., Russell, D. Ujioka, T., Fujii, S., Richards, J. y Espey, L. (2000). Expression of tumor necrosis factor-stimulated gene-6 in the rat ovary in response to an ovulatory dose of gonadotropin. Endocrinology. 141: 4114–4119. 37 108.Zhang, Y., Word, R., Fesmire, S., Carr, B. y Rainey, W. (1996). Human ovarian expression of 17 ß-hydroxysteroid dehydrogenase types 1, 2, and 3. J Clin. Endocrin. Metabo. 81:3594-35. 109.Zhou, L., Yoneda, M., Zhao, M., Yingsung, W., Yoshida, N., Kitagawa, Y., Kawamura, K., Suzuki, T. y Kimata K. (2001). Defect in SHAP-hyaluronan complex causes severe female infertility: a study by inaction of the bikunin gene in mice. J. Bio.l Chem. 276: 7693–7696. 110.Zollers, W., Garverick, H. Smith, M., Moffatt, R., Salfen, B. and Youngquist, R. (1993). Concentrations of progesterone and oxytocin receptors in endometrium of post partum cows expected to have a short or normal oestrous cycle. J. Reprod. Fertil. 97, 329-337.
 0
0
Anuncio
Documentos relacionados



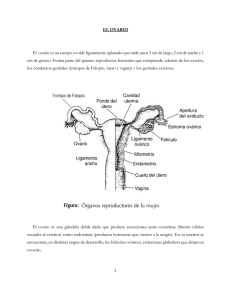

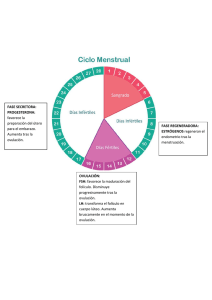


Descargar
Anuncio
Añadir este documento a la recogida (s)
Puede agregar este documento a su colección de estudio (s)
Iniciar sesión Disponible sólo para usuarios autorizadosAñadir a este documento guardado
Puede agregar este documento a su lista guardada
Iniciar sesión Disponible sólo para usuarios autorizados