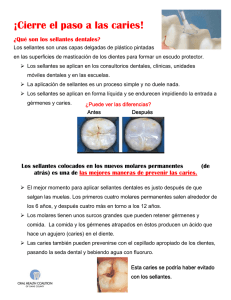
determinacion de polisacáridos extracelulares en cepas aisladas en
Anuncio

UNIVERSIDAD VERACRUZANA FACULTAD DE ODONTOLOGIA REGION POZA RICA -TUXPAN ACADEMIA DEL AREA DE FORMACION TERMINAL “DETERMINACION DE POLISACÁRIDOS EXTRACELULARES EN CEPAS AISLADAS EN NIÑOS DE LA ESCUELA PRIMARIA IGNACIO RAMÍREZ” TESIS PRESENTA: SAMUEL XOCHIHUA RAMIREZ DIRECTOR Q.F.B HERIBERTO DEL ANGEL CALDERON ASESOR C.D.M P ARACELI GARCIA ROCHA POZA RICA DE HGO., VER. NOVIEMBRE 2011 AGRADECIMIENTOS A DIOS: POR DARME LA VIDA, LA SALVACIÓN, EL AMOR, LA PAZ, LA TRANQUILIDAD, LAS FUERZAS Y POR LAS MUCHAS BENDICIONES COMO LO SON MIS PADRES Y MIS HERMANOS: A AQUEL QUE ES PODEROSO PARA GUARDAROS SIN CAÍDA Y PRESENTARNOS SIN MANCHA DELANTE DE SU GLORIA CON GRAN ALEGRÍA, AL ÚNICO Y SABIO DIOS, NUESTRO SALVADOR, SEA GLORIA, MAJESTAD, IMPERIO Y POTENCIA, AHORA Y POR TODOS LOS SIGLOS. AMÉN. JUDAS 1: 24-25 A MIS PADRES: POR SIMPRE APOYARME EN TODOS LOS SENTIDOS, POR SUS CONSEJOS, Y SU GUIANZA EN MI VIDA DIARIA. RESUMEN El presente trabajo piloto de investigación trata acerca de la producción de polisacáridos extracelulares por parte de la bacteria Estreptococo mutans, primeramente en el marco teórico se dividió de la manera siguiente, empezando por una visión general de los carbohidratos (tipos de carbohidratos, estructura química, usos, importancia) para conocer y comprender mejor los términos utilizados en el presente trabajo. Posteriormente se describe la placa dentobacteriana por es en este lapso o fase de proceso donde está involucrado el estreptococo mutans en la producción de polisacárido, se describe la definición de placa dentobacteriana, los tipos y el proceso de formación de esta. El siguiente apartado trata acerca de la caries, la importancia que juegan los microorganismos de la flora en la producción de esta, los procesos químicos para que se de la producción de ácidos por parte de las bacterias así como su relación con los polisacáridos extracelulares. En el último apartado se detalla el los tipos, el metabolismo, los tipos de enzimas que actúan para la formación de polisacárido extracelular soluble e insoluble. Se definen términos para comprender mejor el tema y ya que ultimo se utilizo un medio de cultivo para poder sembrar a la bacteria y que produjera polisacárido extracelular, se describe acerca de los medios de cultivos, (tipos, clasificación). Se describe después el procedimiento utilizado así para la elección de las colonias a sembrar, fueron 16 colonias de 4 pacientitos, previamente obtenidas por un proyecto de investigación antes realizado. Las cuales se sembraron y se llego al resultado esperado donde fue mayor la producción de polisacárido extracelular insoluble que el soluble en agua. (De las 16 muestras, 12 resultaron producir polisacárido extracelular insoluble y solo4 soluble) en porcentaje seria de 75% P. insoluble contra un 25% de polisacárido soluble. ABSTRACT This paper discusses research pilot production of extracellular polysaccharide by the bacteria Streptococcus mutans, primarily in the theoretical framework was divided as follows, starting with an overview of the carbohydrates (types of carbohydrates, chemical structure uses, importance) to know and understand the terms used in this work. Later described by plaque is in this period or phase of the process where Streptococcus mutans is involved in the production ofpolysaccharide, describes the definition of plaque, types and the formation of this. The next section deals with the decay, the importance played by micro flora in the production of this, the chemical processes so that acid production by bacteria and its relationship with extracellular polysaccharides. The final section details the types, metabolism, the types ofenzymes involved in the formation of soluble and insolubleextracellular polysaccharide. Terms are defined to better understand the issue and was being used as a culture medium togrow the bacteria and to produce extracellular polysaccharide, is described on the culture media (type, classification). It then describes the procedure used and the choice of plantingcolonies, 4 colonies were 16 little patients, previously obtained by a research project done before. Which were planted and where I get the expected result was higher extracellular polysaccharide production insoluble soluble in water. (Of the 16 samples, 12 wereproducing extracellular polysaccharide insoluble and solublesolo4) as a percentage would be 75% P. against 25% insoluble polysaccharide soluble. INDICE Agradecimientos Resumen Abstract Índice Lista de Figuras Lista de Gráficas Lista de Tablas CAPITULO I INTRODUCCION……………………………………………1 Planteamiento del problema……………………………….. .. 1 Justificación………………………………………….... …….2 Objetivo general…………………………………….….. …...3 Objetivos específicos Hipótesis………………………………………………..........3 Variables………………….………………………………..…4. Palabras clave CAPITULO II MARCO TEORICO Antecedentes históricos de la caries en relación con los azucares.5 Hidratos de carbono visión general Estructura química Tipos de glúcidos Monosacáridos Disacáridos Polisacáridos Placa dentobacteriana……………………………………………18 La saliva Composición de la saliva Función de la saliva Película adquirida Materia alba Tipos de placa dentobacteriana Matriz orgánica intracelular Metabolismo de la placa Caries dental………………………………………………………33 Etiología de la caries Carbohidratos en la dieta Teorías de la caries Flora oral en relación con la caries dental Bacterias proteolíticas en relación con la caries Localización de flora oral y caries Bioquímica de la caries Bioquímica enzimática extracelular del E. mutans Descripción del proceso carioso Polisacáridos extracelulares………………………………………59 Síntesis de glucanos extracelulares Funciones de los glucanos extracelulares Medios de cultivo en microbiología……………………………..67 Condiciones para el cultivo de microorganismos Clasificación de los medios de cultivo Morfología colonial CAPÍTULO III METODOLOGIA Tipo de estudio……………………………………………………….79 Análisis de la muestra Criterios de inclusión Criterios de exclusión Criterios de eliminación Infraestructura Recursos humanos Recursos financieros Material Procedimiento……………………………………………………82 CAPITULO IV RESULTADOS …………………………………………………….97 CAPITULO V CONLCUSIONES ………………………………………………….102 DISCUSIÓN ………………………………………………………...103 RECOMENDACIONES……………………………………….……104 BIBLIOGRAFIA……………………………………………………105 LISTA DE FIGURAS Fig. 1. Polisacáridos extracelulares Fig. 2. Glándulas salivales Fig. 3. Formación de biofilm Fig. 4. Proceso de formación de caries Fig. 5. Sistemas de ingreso de glucosa Fig. 6. Actividad de las glucosiltransferasas Fig. 7. Medios de cultivo Fig. 8. Morfología colonial superficie Fig. 9. Morfología colonial forma Fig. 10. Morfología colonial borde Fig. 11. Medio de cultivo utilizado (infusión cerebro corazón) Fig.12. Ingredientes del medio Fig.13. Sacarosa utilizada Fig.14. Agua destilada utilizada Fig.15. Matraz y balanza Fig.16. Matraz con medio utilizado Fig.17. sacarosa y balanza Fig.18. Agua destilada y medio mezclados Fig.19 Medio y mechero Fig.20. Medio totalmente homogenizado Fig.21. Tubos y medio BHI Fig.22. Vaso de precipitado con tubos y sacarosa Fig.23. Vaso de precipitado con medio Fig.24. Vaso de precipitado y autoclave Fig.25. Medio y estufa bacteriológica Fig.26. Colonias seleccionadas y tubos con medio Fig.27. Área de trabajo Fig.28. Cepas no. A1 Fig.29. Cepas no.B1 Fig.30. Cepas no.C1 Fig.31. Cepas no.D1 Fig.32. Tubo con colonia Fig.33. Tubos inoculado Fig.34. asa y mechero Fig.-35 colonia escogida Fig.-36 asa y tubo Fig.-37 asa y tubo 2 Fig.- 38 colonia en tubo Fig.- 39 colonia y fondo de tubo Fig. 40 tubos sembrados Fig.41 condiciones de anaerobiosis Fig.- 42 muestras A1 5 tubos Fig.- 43 muestra B1 2 tubos Fig.-44 muestra C1 2 tubos representativos Fig.-45 muestra D1 tubos solubles LISTA DE GRAFICAS 1.-Porcentaje de polisacáridos extracelulares producidos LISTA DE TABLAS 1.-Tabla de frecuencia de producción de producción de polisacáridos 2.- tabla de distribución de colonias en los tubos para la producción de polisacáridos extracelulares INTRODUCCIÓN Los hidratos de carbono, en especifico la sacarosa (disacárido formado por fructuosa y glucosa) están contenidos en una gran variedad de alimentos consumidos diariamente, los cuales poseen gran potencial cariogénico (se obtiene según la consistencia física del alimento, la cantidad, la frecuencia y el momento de consumo) Los hidratos de carbono participan entre otras cosas en la constitución de peptidoglucanos, que son sustancias que conforman las paredes celulares bacterianas, propiciando el desarrollo de placa dentobacteriana, otro factor de riesgo , el cual es reservorio a más bacterias cariogénicas ; el estreptococos mutans es la especie bacteriana considerada como la de mayor potencial cariogénico, produce enzimas que rompen los enlaces de sacarosa y une los residuos de glucosa entre sí para formar glucanos solubles e insolubles (polisacáridos) de los cuales estos últimos los insolubles sirven de matriz pegajosa para que se adhieran otras bacterias y sigan produciendo los ácidos en la cavidad bucal que conducen a la desmineralización de la superficie del esmalte y el inicio de las lesiones cariosas PLANTEAMINETO DEL PROBLEMA La caries dental es una de las enfermedades más antiguas de la humanidad, es un problema de alcance mundial, afecta a los países industrializados y cada vez con mayor frecuencia a los países en desarrollo, en especial entre las comunidades más pobres, constituye la causa principal de la pérdida dentaria y puede predisponer a otras enfermedades. Su etiología es multifactorial, y ya que es una enfermedad crónica termina con la destrucción de los tejidos del diente, esto es causa directa de los ácidos producidos por diversas bacterias de las cuales los grupos más representativos son el grupo de estreptococos mutans y lactobacilos. La fuente nutricional principal de estas bacterias son los hidratos de carbono, estas bacterias en especial estreptococo mutans, tiene la capacidad de que a partir de 1 carbohidratos generar polisacáridos extracelulares, en especial glucanos, que constituyen un factor etiológico grave, ya que le dan a la bacteria ventajas tales como la adhesividad al diente, la reserva de energía y la aportación de esta y como producto final tendríamos los ácidos productores que desmineralizarían al diente y generarían la caries dental. Los polisacaridos extracelulares por lo tanto representan un problema que no ha sido muy estudiado hasta la fecha, la determinación de este tipo de polisacáridos en pacientes de México es poca, no se ha dado la importancia que se requiere, ya que si conociéramos más acerca de la producción de polisacáridos por estreptococo mutans, así como de su metabolismo se podrían tomar medidas para disminuir la producción de este tipo de polisacáridos que como se he venido diciendo es un factor etiológico importantísimo para la producción de caries. Por lo cual surge la siguiente pregunta de investigación: ¿Cuál es la producción de polisacáridos extracelulares de niños de la escuela primaria Ignacio? ¿cuál es la producción de Polisacáridos extracelulares insolubles de niños de la escuela primaria Ignacio Ramírez? ¿Cuál es la producción de Polisacáridos extracelulares solubles de niños de la escuela primaria Ignacio Ramírez? JUSTIFICACION Existen polisacáridos extracelulares de los cuales los que sintetizan las bacterias son glucanos, fructanos y heteropolisacáridos. Los glucanos son uno de los componentes principales del biofilm dental, de los cuales se puede distinguir 2 tipos principales de glucanos extracelulares bacterianos, el dextrano (glucano soluble en agua) y el mutano (glucano insoluble en agua). Así, la habilidad del E. mutans de sintetizar mutano, es esencial para la colonización eficiente y el desarrollo de la caries. En este estudio de investigación se realizara una siembra de colonias de estreptococos de diferentes niños de la escuela primaria Ignacio Ramírez los cuales se sembraran en medio especial para el desarrollo de glucanos extracelulares y determinar la producción de glucanos insolubles así como solubles en agua, con esto se pretende resaltar la 2 importancia de los polisacáridos extracelulares en especial los glucanos, saber más acerca de su estructura, síntesis, metabolismo así como sus funciones, así como hacer recomendaciones para la aplicación de estos conocimientos en la prevención de la caries dental. Con este estudio de investigación se deja opción a que se sigan haciendo investigaciones acerca de la producción de polisacáridos en una población más grande así como su determinación y comportamiento en una diferente dieta. OBJETIVO GENERAL Determinar la producción de polisacáridos extracelulares en cepas aisladas de los niños de la escuela primaria Ignacio Ramírez. OBJETIVOS ESPECÍFICOS Determinar la producción de polisacáridos extracelulares solubles de las cepas aisladas de los niños de la escuela primaria Ignacio Ramírez Determinar la producción de polisacáridos extracelulares insolubles de las cepas aisladas de los niños de la escuela primaria Ignacio Ramírez HIPÓTESIS Existe una alta producción de polisacáridos extracelulares Existe una baja producción de polisacáridos extracelulares No existe producción de polisacáridos extracelulares 3 VARIABLES Dependiente La capacidad del Estreptococo mutans para producir polisacáridos La producción de polisacáridos insolubles y solubles Independiente La selección de la colonia a inocular PALABRAS CLAVES Polisacárido extracelular, glucano, mutano, dextrano, caries INDICADORES Morfología colonial característica de estreptococo mutans 4 CAPITULO II MARCO TEORICO ANTECEDENTES HISTÓRICOS: La historia de la caries discurre paralelamente a muchos cambios ocurridos con el tiempo en la práctica de la odontología. Así estudios antropológicos han confirmado la escasez relativa de casos de caries antes del inicio de este milenio, cuando aún no se disponía de azúcares simples y la odontología era prácticamente inexistente. Durante los 500 años siguientes, la incidencia de la caries fue aumentando a medida que se introdujeron los azúcares en la dieta. Con el establecimiento de las plantaciones de azúcar en el nuevo mundo a principios del siglo XVIII, y con la posterior proliferación en Europa del azúcar de remolacha durante el siglo XIX, apareció una pandemia de caries que aún persiste en la actualidad y que se caracteriza porque la población continúa consumiendo ciertos monosacáridos y disacáridos (sobre todo sacarosa) aunque antiguamente la presencia de caries en un diente se consideraba un signo de gangrena y el único tratamiento disponible era la extracción de la pieza enferma, durante el siglo XIX se introdujeron ya las técnicas de desbridamiento de la caries y la restauración. El papel protector del flúor frente a la caries dental fue descubierto hasta mediados de los años treinta, cuando se demostró que los niños que bebían aguas fluoradas presentaban muchas menos cavidades que los que bebían agua procedente de suministros con bajo contenido en flúor. Las pastas desnitrificas fluoradas se introdujeron en 1955 y ofrecen una importante protección adicional. Sin embargo, a pesar de la reducción espectacular de las tasas de caries dental durante la última mitad del siglo XX y el aumento simultáneo de la importancia de la odontología preventiva, la caries sigue afectando aún a un porcentaje muy importante de la población. 1 5 HIDRATOS DE CARBONO: Los glúcidos, carbohidratos, hidratos de carbono o sacáridos son moléculas orgánicas compuestas por carbono, hidrógeno y oxígeno. Son solubles en agua y se clasifican de acuerdo a la cantidad de carbonos o por el grupo funcional aldehído. Son la forma biológica primaria de almacenamiento y consumo de energía. Otras biomoléculas energéticas son las grasas y, en menor medida, las proteínas. El término "hidrato de carbono" o "carbohidrato" es poco apropiado, ya que estas moléculas no son átomos de carbono hidratados, es decir, enlazados a moléculas de agua, sino que constan de átomos de carbono unidos a otros grupos funcionales. Este nombre proviene de la nomenclatura química del siglo XIX, ya que las primeras sustancias aisladas respondían a la fórmula elemental Cn (H2O)n (donde "n" es un entero=De 3 en adelante; según el número de átomos). De aquí que el término "carbonohidratado" se haya mantenido, si bien posteriormente se vio que otras moléculas con las mismas características químicas no se corresponden con esta fórmula. Además, los textos científicos anglosajones aún insisten en denominarlos carbohydrates lo que induce a pensar que este es su nombre correcto. Del mismo modo, en dietética, se usa con más frecuencia la denominación de carbohidratos. Los glúcidos pueden sufrir reacciones de esterificación, aminación, reducción, oxidación, lo cual otorga a cada una de las estructuras una propiedad específica, como puede ser de solubilidad. 2 Sinónimos Carbohidratos o hidratos de carbono: Ha habido intentos para sustituir el término de hidratos de carbono. Desde 1996 el Comité Conjunto de la Unión Internacional de Química Pura y Aplicada (International Unión of Pure and 6 Applied Chemistry1 ) y de la Unión Internacional de Bioquímica y Biología Molecular (International Union of Biochemistry and Molecular Biology) recomienda el término carbohidrato y desaconseja el de hidratos de carbono. Glúcidos: Este nombre proviene de que pueden considerarse derivados de la glucosa por polimerización y pérdida de agua. El vocablo procede usarse para del griego "glycýs", que significa dulce. Azúcares: Este término sólo los monosacáridos (aldosas y cetosas) puede y los oligosacáridos inferiores (disacáridos). En singular (azúcar) se utiliza para referirse a la sacarosa o azúcar de mesa. Sacáridos: Proveniente del griego σάκχαρον que significa "azúcar". Es la raíz principal de los tipos principales de glúcidos (monosacáridos, disacáridos, oligosacáridos y polisacáridos). Estructura química: Los glúcidos son compuestos formados en su mayor parte por átomos de carbono e hidrógeno y en una menor cantidad de oxígeno. Los glúcidos tienen enlaces químicos difíciles de romper llamados covalentes, mismos que poseen gran cantidad de energía, que es liberada al romperse estos enlaces. Una parte de esta energía es aprovechada por el organismo consumidor, y otra parte es almacenada en el organismo. En la naturaleza se encuentran en los seres vivos, formando parte de biomoléculas aisladas o asociadas a otras como las proteínas y los lípidos. 7 Todos los carbohidratos son azucares pequeños solubles en agua (glucosa y fructuosa) o bien cadenas como el almidón y la celulosa que se elaboran enlazando subunidades de azúcar. 3 Tipos de glúcidos Si el carbohidrato se compone de una sola molecula de azúcar se le llama monosacárido, si se enlazan dos o más monosacáridos se forma un disacárido, o un polisacárido. Los carbohidratos como los azucares y almidones, son fuentes importantes de energía para casi todos los organismos. Consideremos un desayuno que incluya panqueques de moras, jarabe y jugo de naranja, los panqueques constituyen principalmente los carbohidratos almacenados en las semillas de trigo u otros cereales. El azúcar que endulza al jarabe las moras y el jugo de naranja también se almacena en las plantas como fuente de energía otros carbohidratos como por ejemplo la celulosa y las moléculas similares proporcionan soporte estructural a células individuales o incluso al cuerpo entero de organismos tan diversos como plantas, hongos bacterias e insectos. Los azucares se enlazan en forma de anillo para formar disacáridos y polisacáridos. Casi todos los carbohidratos pequeños son solubles en agua. 8 Monosacáridos La glucosa es la más común de los carbohidratos presentes en los organismos vivos, y es la subunidad de la que están formados casi todos los polisacáridos. La glucosa tiene 6 carbonos, así su formula química es C6 H 12 O6. Muchos otros organismos sintetizan otros monosacáridos que tienen la misma fórmula que la glucosa, pero una estructura ligeramente distinta, entre ellos está la fructosa ( el azúcar del maíz presente en la miel de maíz, y también la molecula que hace que el jugo de naranja sepa dulce) la galactosa ( parte de la lactosa, el azúcar de la leche), otros monosacáridos incluyen la ribosa y la desoxirribosa que tienen 5 carbonos, estas forman parte de las moléculas genéticas acido ribonucleico y acido desoxirribonucleico. Disacáridos Los monosacáridos en especial la glucosa y sus moléculas relacionadas tienen una vida relativamente corta en las células, casi todos se descomponen para liberar su energía química la cual se usa en diversas actividades celulares o se encadenan mediante síntesis por deshidratación para formar disacáridos o polisacáridos, los disacáridos se utilizan a menudo para reservar energía a corto plazo sobre todo en plantas, tal vez usted tomo café con leche y azúcar en el desayuno, entre los disacáridos más comunes están la sacarosa ( glucosa mas fructosa), que es lo que endulzo su café, la lactosa ( el azúcar de la leche) y la maltosa ( glucosa mas glucosa que se formara en su tracto digestivo cuando descomponga en almidón de sus panqueques. Si se requiere energía los disacáridos se dividen en subunidades de monosacárido mediante hidrólisis. Polisacáridos Pruebe masticar una galleta salada durante un largo plazo para que las enzimas de la saliva tengan tiempo de descomponerla en sus azucares componentes ¿sabe más dulce cuanto más tiempo se mastica? Así debería ser, los monosacáridos casi siempre la 9 glucosa se juntan para formar polisacáridos como el almidón en las plantas o el glucógeno en los animales, donde se almacena energía a largo plazo. El almidón comúnmente se forma en las raíces y en las semillas, en el caso de la galleta, se las semillas de trigo, el almidón puede presentarse como cadenas enrolladas no ramificadas de hasta 1000 subunidades de glucosa o más comúnmente cadenas ramificadas de hasta medio millón de monómeros de glucosa. El glucógeno almacenado como fuente de energía en el hígado y en los músculos de los animales entre ellos los seres humanos suele ser mucho más pequeño que en el almidón y tiene ramificaciones cada 10 a 12 subunidades de glucosa. Tener muchas ramas pequeñas probablemente facilita la separación de subunidades de glucosa cuando se necesita liberar energía rápidamente. Muchos organismos utilizan también polisacáridos como materiales estructurales, uno de los más importantes polisacáridos extracelulares estructurales es la celulosa, quien integra la mayor parte de las paredes celulares de las plantas y aproximadamente la mitad de la masa del tronco de un árbol. Si pensamos en los extensos campos y bosques que cubren gran parte de nuestro planeta no nos extrañara enterarnos que habría más celulosa en la tierra que en todas las demás moléculas orgánicas juntas.los ecólogos calculan que se sintetiza cerca de un billón de toneladas de celulosa cada año. Al igual que el almidón la celulosa consiste en pequeñas unidades de glucosa unidas o enlazadas. Sin embargo aunque la mayoría de los animales puede digerir fácilmente el almidón, solo unos cuantos microbios como los que habitan en el tracto digestivo de las vacas o de las termitas pueden digerir la celulosa. ¿Por qué es así, si tanto el almidón como la celulosa están formados de glucosa? Es porque la orientación de los enlaces entre las subunidades es diferente en los dos polisacáridos, en la celulosa cada segunda glucosa está literalmente de cabeza, esta orientación de los enlaces impide que las enzimas digestivas de los animales ataquen los enlaces de entre las subunidades de glucosa. 10 Las enzimas sintetizadas por ciertos microbios en cambio pueden romper esos enlaces, el resultado es que la celulosa sirve como alimento para esos microbios, en cambio para la mayoría de los animales la celulosa es fibra (material que pasa por el tracto digestivo sin digerirse). Las cubiertas externas duras de los insectos, cangrejos y arañas están formados de quitina, un polisacárido en el que las subunidades de glucosa han sufrido alguna modificación química por la adición de un grupo funcional nitrogenado. Resulta interesante que la quitina también hace rígidas las paredes celulares de muchos hongos, las paredes celulares de las bacterias contienen otros tipos de polisacáridos modificados, los mismos que los fluidos lubricantes de nuestras articulaciones y las corneas trasparentes de los ojos. Muchas otras moléculas incluido el moco, algunos mensajeros químicos llamados hormonas y muchas moléculas de la membrana plasmática se componen parcialmente de carbohidratos, quizá las más interesantes de esas moléculas son los ácidos nucleicos que llevan la información hereditaria. 2 11 Clasificación de los polisacáridos: Para la clasificación de los polisacáridos, se acude a uno de dos criterios, el funcional, que es el más difundido, o el químico, que se atiene a su estructura y composición. Según la función biológica, podemos clasificar los polisacáridos en los siguientes grupos: Bien como sustancias de reserva, o bien como moléculas estructurales. Los que realizan una función estructural presentan enlace b -glucosídico, y los que realizan una función de reserva energética presentan el enlace a -glucosídico . Polisacáridos de reserva: Almidón: El almidón es la sustancia con la que las plantas almacenan su alimento en raíces (yuca), tubérculos (patata), frutas y semillas (cereales). Pero, no sólo es una importante reserva para las plantas, también para los seres humanos tiene una alta importancia energética, proporciona gran parte de la energía que consumimos los humanos por vía de los alimentos. El almidón se diferencia de los demás hidratos de carbono presentes en la naturaleza en que se presenta como un conjunto de gránulos o partículas. Estos gránulos son relativamente densos e insolubles en agua fría, aunque pueden dar lugar a suspensiones cuando se dispersan en el agua. Suspensiones que pueden variar en sus propiedades en función de su origen. Almidón y estructura química: Desde el punto de vista químico el almidón es un polisacárido, el resultado de unir moléculas de glucosa formando largas cadenas, aunque pueden aparecer otros constituyentes en cantidades mínimas. 12 El almidón es una sustancia que se obtiene exclusivamente de los vegetales que lo sintetizan a partir del dióxido de carbono que toman de la atmósfera y del agua que toman del suelo. En el proceso se absorbe la energía del sol y se almacena en forma de glucosa y uniones entre estas moléculas para formar las largas cadenas del almidón, que pueden llegar a tener hasta 2000 o 3000 unidades de glucosa. El almidón está realmente formado por una mezcla de dos sustancias, amilasa y amilopectina, que sólo difieren en su estructura: la forma en la que se unen las unidades de glucosa entre si para formar las cadenas. Pero esto es determinante para sus propiedades. Así, la amilosa es soluble en agua y más fácilmente hidrolizable que la amilopectina (es más fácil romper su cadena para liberar las moléculas de glucosa). En realidad, la estructura del almidón es muy parecida a la de la celulosa, otro polisacárido que producen las plantas. Pero mientras el almidón es parte del alimento de muchos animales y se descompone fácilmente por acción de las enzimas digestivas, la celulosa es parte del tejido de sostén de las plantas y muy difícil de digerir, algo que la mayoría de los animales aprenden rápidamente. En los animales, el equivalente al almidón, como sustancia de reserva energética, es otra sustancia de estructura parecida que recibe el nombre de glucógeno. El almidón se puede identificar fácilmente gracias a que la amilosa en presencia de yodo forma un compuesto azul estable a bajas temperaturas. Usos: El almidón es importante porque forma parte de nuestra dieta. Se encuentra en las patatas, el arroz, los cereales, las frutas, etc. En una dieta sana, la mayor parte de la energía la conseguimos a partir del almidón y las unidades de glucosa en que se hidroliza. 13 El almidón también es muy utilizado en la industria alimentaria como aditivo para algunos alimentos. Uno más de los muchos utilizados. Tiene múltiples funciones entre las que cabe destacar: adhesivo, ligante, enturbiante, formador de películas, estabilizante de espumas, conservante para el pan, gelificante, aglutinante, etc. El problema surge porque muchas veces no se nos informa de su uso. Así, por ejemplo, se utiliza en la fabricación de embutidos y fiambres de baja calidad para dar consistencia al producto. Antiguamente, el almidón se utilizaba para "almidonar" la ropa. Cuando se lavaba la ropa se le daba un baño en una disolución de almidón para conseguir que después del planchado quedara tersa o con apresto y evitar que se arrugara, por ejemplo sábanas y camisas. También se utilizaba en mayor concentración, incluso para conseguir que la ropa quedara tiesa, como por ejemplo, los "can-can" que llevaban las mujeres debajo de las faldas para dar volumen. Hoy en día el almidón tiene otras muchas aplicaciones. Por ejemplo, es un excelente agente antiadherente en múltiples usos. Pero también puede utilizarse para todo lo contrario: como adhesivo. Una utilización muy interesante del almidón es la preparación de embalajes de espuma, una alternativa biodegradable a los envases de poliestireno. 4 Glucógeno: El glucógeno es la forma principal de almacenaje de carbohidratos en los animales, se encuentra en proporción mayor en el hígado (hasta 6%) y en el músculo, donde rara vez excede de 1%. Sin embargo, debido a su masa mayor, el músculo almacena tres a cuatro veces la cantidad de glucógeno que tiene el hígado como reserva. Al igual que el almidón, es un polímero ramificado de alfa-glucosa. En las células hepáticas, el glucógeno aparece en forma de grandes gránulos, constituidos por agrupaciones de simples moléculas, muy ramificadas, por lo que tiene un peso molecular muy elevado. A semejanza de la amilopectina, el glucógeno es un polisacárido de la D-glucosa con enlaces alfa 1-4, sin embargo, está más ramificado, y su molécula es más reducida que la amilopectina; las ramificaciones aparecen cada 8 a 12 residuos de glucosa. 14 El glucógeno puede aislarse de los tejidos animales digiriéndolos con disoluciones calientes de KOH en las que los enlaces no reductores alfa 1-4 y alfa 1-6 son estables. Importancia biomédica: La función del glucógeno muscular es actuar como una fuente de fácil disponibilidad de unidades de hexosa para la glucólisis dentro del propio músculo. El glucógeno hepático sirve en gran parte para exportar unidades de hexosa para la conservación de la glucosa sanguínea, en particular entre comidas. Después de 12 a 18 horas de ayuno, el hígado casi agota su reserva de glucógeno. El glucógeno muscular sólo disminuye de manera significativa después de ejercicio vigoroso prolongado. Puede inducirse un almacenaje mayor de glucógeno muscular con dietas ricas en carbohidratos después de la depleción por el ejercicio. Las "enfermedades por almacenamiento de glucógeno" son un grupo de trastornos hereditarios que se caracterizan por movilización deficiente del glucógeno y depósito de formas anormales del mismo, conduciendo a debilidad muscular e inclusive muerte. Funciones de las reservas de glucógeno: El glucógeno constituye la principal fuente de energía para la contracción del músculo esquelético. Dado que el hígado obtiene la mayor parte de su energía metabólica de la oxidación de los ácidos grasos, el glucógeno hepático tiene una función muy distinta, como fuente de glucosa sanguínea que se transporta a otros tejidos para su catabolismo. El hígado actúa fundamentalmente como un "glucostato", sensor de las concentraciones de glucosa sanguínea que ajusta en función de ello la síntesis y degradación de glucógeno; gran parte de esta regulación comporta el control de la glucógeno sintasa y la fosforilasa. 15 Para cumplir esta función, el hígado contiene unas reservas de glucógeno relativamente elevadas, desde un 2 a un 8% del peso del órgano. En el hígado, la velocidad máxima de síntesis y degradación de glucógeno son aproximadamente iguales, mientras que en el músculo la velocidad máxima de glucogenólisis supera a la síntesis de glucógeno en unas 300 veces. Aunque la enzimología de la síntesis y degradación del glucógeno es semejante en el hígado y el músculo, el control endocrino del hígado es bastante diferente. Las enzimas difieren también estructuralmente. 5 Polisacáridos estructurales Figura 1 Estructura de la celulosa. Se trata de glúcidos que participan en la construcción de estructuras orgánicas. Los más importantes son los que constituyen la parte principal de la pared celular de plantas, hongos y otros organismos eucarióticos osmótrofos, es decir, que se alimentan por absorción de sustancias disueltas. Éstos no tienen otra manera más económica de sostener su cuerpo, que envolviendo a sus células con una pared flexible pero resistente, contra la que oponen la presión osmótica de la célula, logrando así una solución del tipo que en biología se llama esqueleto hidrostático. 16 La celulosa es el más importante de los polisacáridos estructurales. Es el principal componente de la pared celular en las plantas, y la más abundante de las biomoléculas que existen en el planeta. Es un glucano, es decir, un polímero de glucosa, con enlaces glucosídicos entre sus residuos de tipo β(1→4). Por la configuración espacial de los enlaces implicados, los residuos de glucosa quedan alineados de forma recta, no en helicoide, que es el caso de los glucanos α(1→4), del tipo del almidón. Ésta es la regla en cuanto a la conformación de todos los polisacáridos estructurales de las paredes. Esas cadenas rectas se enlazan transversalmente, por enlaces de hidrógeno, en haces de cadenas paralelas. La quitina cumple un papel equivalente al de la celulosa, pero en los hongos, y además es la base del exoesqueleto de los artrópodos y otros animales emparentados. La quitina es un polímero de la N-acetil-2, D-glucosamina, un monosacárido aminado, que contiene por lo tanto nitrógeno. Siendo éste un elemento químico de difícil adquisición para los organismos autótrofos, que lo tienen que administrar con tacañería, la quitina queda reservada a heterótrofos como los hongos, que lo obtienen en abundancia. Otras funciones La mayoría de las células de cualquier ser vivo suelen disponer este tipo de moléculas en su superficie celular. Por ello están involucrados en fenómenos de reconocimiento celular (ejemplo: Complejo Mayor de Histocompatibilidad), protección frente a condiciones adversas (Ejemplo: Cápsulas polisacarídicas en microorganismos) o adhesión a superficies (ejemplo: la formación de biofilmes o biopelículas, al actuar como una especie de pegamento). 2 17 Según la composición Se distinguen dos tipos de polisacáridos según su composición: 1. Homopolisacáridos: están formados por la repetición de un monosacárido. 2. Heteropolisacáridos: están formados por la repetición ordenada de un disacárido formado por dos monosacáridos distintos (o, lo que es lo mismo, por la alternancia de dos monosacáridos). Algunos Heteropolisacáridos participan junto a polipéptidos (cadenas de aminoácidos) de diversos polímeros mixtos llamados peptidoglucanos, mucopolisacáridos o proteoglicanos. Se trata esencialmente de componentes estructurales de los tejidos, relacionados con paredes celulares y matrices extracelulares. 6 PLACA DENTOBACTERIANA La placa dentobacteriana es una masa blanda, tenaz y adherente que presenta en su composición distintos tipos de colonia bacterianas, se adhieren a la superficie de los dientes, encía, lengua y otras superficies de la boca. Esta llega a formarse por una mala higiene bucal y es un factor importantísimo en la etiología de la caries dental, la formación de tártaro y la enfermedad periodontal. Otro término que se le da es de una película trasparente sin color que se adhiere al diente compuesta por bacterias y células descamadas dentro de una matriz de mucoproteinas y mucopolisacáridos. Así como la película que la origino la placa dentobacteriana es traslucida y por eso es poco visible a menos que haya deposito de minerales o hemoglobina procedentes de la ruptura de capilares gingivales. Si es delgada se logra observar por medio de un colorante que logre pigmentarla o con uno fluorescente que se ilumina con luz ultravioleta. La placa se puede eliminar con un cepillado vigoroso la película no. 18 Para conocer el mecanismo de la placa dentobacteriana primero es necesario conocer antes la función de la saliva así como las características de la película adquirida y la materia alba LA SALIVA La saliva es un líquido de procedencia orgánica producido por las glándulas salivales, como son 2 glándulas parótidas, dos submaxilares, dos sublinguales así como otras distribuidas de manera aislada de la mucosa bucal. 7 Glándula parótida Es una glándula tubuloacinosa que es sólo serosa, en el ser humano es la de mayor tamaño, está rodeada por una gruesa cápsula de tejido conectivo desde donde parten tabiques de tejido conectivo hacia el interior de la glándula que la dividen en finos lóbulos. El conducto excretor principal o conducto de Stenon o parotídeo desemboca a nivel del segundo molar superior. Tamaño más o menos de 6 cm de longitud y de 3 a 4 cm de ancho. Peso varía de 15 a 30 g. Está situada en la cara lateral de la fosa retromandibular.Las glándulas salivales están localizadas alrededor de la boca y producen la saliva que humedece los alimentos para ayudar con la masticación y la deglución. Las glándulas salivales producen saliva algunas veces llamada esputo - y la segregan hacia la boca a través de aberturas llamadas ductos. La saliva ablanda los alimentos, lo que ayuda a masticarlos y tragarlos. Ayuda a digerir la comida. También limpia la boca y contiene anticuerpos que pueden matar gérmenes. Los problemas de las glándulas salivales pueden irritar e inflamar las glándulas. Eso provoca síntomas, tales como: 19 Mal sabor en la boca Dificultad para abrir la boca seca Dolor en la cara o la boca Inflamación de la cara o el cuello figura 2 Glándula submaxilar La glándula submaxilar una glándula salival que tiene una forma irregular y un tamaño parecido a una nuez. Se localiza en la parte posterior del piso de la boca. Esta glándula produce una secreción musinosa acuosa, llamada mucoserosa, a través del conducto de Wharton. Glándula sublingual Es la más pequeña en volumen y peso (representa un tercio aproximadamente de la submandibular). Ubicada en el surco alveololingual, subyacente a la mucosa con un borde craneal que produce una elevación denominada eminencia sublingual. Su forma es elipsoidal y está aplanada transversalmente, con un eje mayor de dirección ventromedial, y mide 3 cm de longitud aproximadamente.La saliva contiene enzimas que comienzan el proceso de digestión y ayudan a limpiar la boca, eliminando bacterias 20 y partículas de alimentos. Al mantener la boca húmeda, la saliva ayuda a conservar las dentaduras postizas, retenedores u otros aparatos ortodóncicos en su lugar. Composición de la saliva Agua: Representa un 99% de su volumen, en la que se disuelven el 1% restante formado por sales minerales como iones de sodio, potasio, cloruro, bicarbonato y fosfatos. El agua permite que los alimentos se disuelvan y se perciba su sabor en el sentido del gusto. Iones cloruro: Activan la amilasa salival o ptialina. Bicarbonato y fosfato: Neutralizan el pH de los alimentos ácidos y de la corrosión bacteriana. Moco: Lubrica el bolo alimenticio para facilitar la deglución y que pueda avanzar a lo largo del tubo digestivo, sin dañarlo. Lisozima: Es una sustancia antimicrobiana que destruye las bacterias contenidas en los alimentos, protegiendo en parte los dientes de la caries y de las infecciones. Enzimas: Como la ptialina, que es una amilasa que hidroliza el almidón parcialmente en la boca, comenzando la digestión de los hidratos. 21 Estaterina: Con un extremo amino terminal muy ácido, que inhibe la precipitación de fosfato cálcico al unirse a los cristales de hidroxiapatita. Además, también tiene función antibacteriana y antifúngica. Otras sustancias: Como inmunoglobulinas específicas, transferrina, lactoferrina.Tiene una composición viscosa y lubrica el estomago y las tráquea para cuando llega el alimento Calcio: Que ayuda a digerir el alimento. Es inodora como el agua (sin olor). 8 Función de la saliva La saliva se produce por lo general como respuesta a estímulos del sistema nervioso autónomo. La estimulación parasimpática origina secreción acuosa y abundante, en cambio la estimulación simpática con frecuencia se debe a estrés y origina volúmenes menores de una secreción viscosa, con lo cual proporciona la sensación de resequedad bucal. Las funciones son las siguientes: 1.- Proporcionar un medio protector para la mucosa y los dientes Enjuaga la boca al arrastrar consigo partículas de alimentos y desechos celulares. Además contiene inmunoglobulinas A, G Y M, las cuales proporcionan protección a la boca contra la invasión microbiana. 22 El moco de la saliva mantiene la flora bacteriana en una condición constante, lo hace al trasportar las sustancias antibacterianas a las zonas donde se requiere neutralizar a los agentes patógenos. Las glucoproteínas se relacionan con la adhesión de algunas bacterias. Amortigua la acidez normal de la boca Funciona como protector contra la disolución del fosfato de calcio en los tejidos duros ( dentina, cemento y esmalte expuestos) esto lo hace por medio de amortiguadores salivales así como la conservación de una concentración saturada de iones calcio y fosfato. Los amortiguadores salivales mantienen a la saliva en un pH de entre 5.6 y 6.2 pero si existe una estimulación potente puede incrementarlo a 7 u 8, esto debido a un aumento en la concentración de bicarbonato. El fosfato y las proteínas son importantes amortiguadores en la placa dentobacteriana. Contiene antibacterianos específicos (bacteriostáticos, bactericidas y aglutinantes) los cuales actúan como mecanismo de defensa. Por ejemplo las opsoninas vuelven susceptibles las bacterias al romper los enlaces entre Nacetilmuramico y N- acetilglucosamina de las paredes celulares y leucocitos. La cantidad de leucocitos en una boca saludable varia de 100 a 1000 000/ml y puede llegar hasta 11 000 000/ml en una boca con problemas inflamatorios. La lactoferrina une iones Fe suprimiendo este elemento esencial para algunas bacterias bucales y a la lactoperidasa que hace la unión del tiocinato y el peróxido de hidrogeno produzca sustancias toxicas para muchas bacterias. 23 2.- humedece y lubrica la mucosa bucal y los labios Esta humidificación es continua debido a la evaporación y la deglución de la saliva. La lubricación se lleva a cabo por medio de glucoproteínas llamadas mucinas de las cuales depende la viscosidad de la saliva, ya que son capaces de unir moléculas de agua con sus grupos hidroxilo. Ayuda en la digestión, ya que humedece los alimentos ingeridos para dar la consistencia semisólida y facilitar su deglución. Además contiene enzimas muy importantes como la amilasa alfa, esta hidroliza las dextrinas, disminuye la capacidad de los genes de almidón y ayuda a eliminar desechos de hidratos de carbono de los dientes. También contiene a la amilasa beta que desdobla las moléculas. También contiene aliesterasas las cuales hidrolizan los esteres de ácidos grasos , lipasas que desdoblan glicéridos de ácidos grasos así como enzimas de trasferencia como la catalasa la peroxidasa y la hexocinasa que catalizan reacciones en las cuales trasfieren un grupo químico de un compuesto a otro 7 Película adquirida El esmalte del diente que se encuentra en erupción está cubierto por una ligera capa de proteínas llamada lámina basal o cutícula del esmalte, la cual es producto final del metabolismo del ameloblasto y desaparece con rapidez para así permitir el contacto directo del diente con el medio bucal. 24 Composición Tiempo después se forma una nueva película, la película adquirida, esta se une con firmeza a la superficie dental, tiene menos de una micra de espesor y se compone de proteínas salivales como fosfoproteínas y glucoproteínas, así como de enzimas e inmunoglobulinas que posteriormente se desnaturalizaran. La composición de la película varía de un sujeto a otro, sin embargo las cargas eléctricas de sus moléculas orgánicas son distintas a las de los cristales minerales de la hidroxiapatita adamantina y ello favorece su fuerte fijación en grietas, fisuras y la superficie del esmalte. Es importante decir que la película adquirida no se elimina con el cepillado, solo se quita con un abrasivo fuerte, pero vuelve a formarse inmediatamente al contacto con la saliva, a los 90 minutos ya están integradas sus primeras capas y a las 3 o 4 horas como máximo esta ya formada en su totalidad. Características Su aspecto es claro y translucido aunque puede pigmentarse con el consumo de tabaco o en sitios donde existen muchos polvos de cobre, níquel, cadmio o hierro. Suele considerarse una estructura simple sin embargo Meckel la divide en tres capas: Película subsuperficial: es una red de fibrillas que se introduce y adhiere a las irregularidades microscópicas del esmalte. Tiene de 2 a 3 micras d espesor 25 Película superficial: es una capa de material amorfo y mide de 0.02 a 5 micras de espesor. Película suprasuperficial o manchada: aquí se encuentran algunas ocasiones algunos microorganismos y productos finales de su metabolismo. Funciones Se le han atribuido funciones tanto protectoras como perjudiciales y están incluyen: Retrasar la desmineralización del esmalte al actuar como barrera para la difusión de los ácidos desde la placa dentobacteriana hacia la superficie adamantina. Retrasar la difusión de iones calcio y fosfato desde el área de desmineralización y de ese modo intensificar el proceso de remineralización. Actuar como matriz inicial a la cual se le adhieren las bacterias bucales para iniciar la formación de la placa dentobacteriana. Materia alba Está compuesta por masas de bacterias, restos alimenticios, células epiteliales descamadas así como leucocitos. Tiene una característica muy especial esta ligeramente adherida a los dientes por los cual es posible eliminarla incluso con una jeringa de agua, pero es distinta a la placa dentobacteriana. 26 TIPOS DE PLACA DENTOBACTERIANA Según su localización, la placa dentobacteriana puede ser supragingival, subgingival, de fosas y fisuras, proximal y radicular. Placa dentobacteriana supragingival Esta se extiende desde el margen libre de la encía hasta la corona del diente, su composición es variada pero en general está constituida por microorganismos y matriz orgánica celular. Microorganismos La formación de placa dentobacteriana supragingival se inicia con la colonización primaria, es decir primeramente con la adherencia de microorganismos aerobios grampositivos en colonias aisladas. Esta colonización es selectiva, porque al parecer la película adquirida tiene receptores para las bacterias. El primer colonizador es Streptococcus sanguis y después Actinomyces viscosus y otros Streptococcus. Estas bacterias unen a la película adquirida mediante enlaces débiles. Después de esto se agregan estreptococos de las especies Mitis, crista y otras bacterias como de Neisseria y Corynebacterium matruchotii). Este tipo de placa tiene metabolismo aerobio, las bacterias aerobias facultativas se adaptan, con excepción de las especies veillonella, las cuales sobreviven a partir del lactato elaborado por otros microorganismos y poseen mecanismos de resistencia al oxigeno. Las especies de Provotella, porphyromonas y fusobacterium conforman el 0.2 % de la colonia bacteriana y son microorganismos anaerobios estrictos. 27 En el trascurso de las primeras 48 horas las colonias crecen y confluyen es decir se unen unas con otras. Por medio del microscopio electrónico es posible observar al principio imágenes en granos de maíz por que predominan los cocos, mas tarde las típicas mazorcas con formas filamentosas recubiertas de cocos. En la fase de colonización primaria algunas placas dentobacterianas supragingivales no son cariogenicas ya que tienen estreptococos mutans y lactobacilos pero poseen poco poder de adhesión. La colonización secundaria comienza entre los 3 a 5 días posteriores, algunas bacterias aumentan en número y otras disminuyen y otras más se agregan, como hay competencia por el consumo de oxigeno las mas aerobias van siendo sustituidas por anaerobias y anaerobias facultativas. Por medio del microscopio electrónico es posible ver un aumento en las formas bacilares sobre todo de especias de Actinomyces, la agregación de bacilos sobre bacilos da lugar a acumulaciones pilosas.los microorganismos se distribuyen en las capas externas y los anaerobios en las más profundas, los estreptococos son aun los mas abundantes y se localizan en cualquier lugar. La velocidad de crecimiento de la placa dentobacteriana supragingival es rápida durante la primera semana y disminuye en las dos siguientes mientras alcanza su maduración. Partir de este momento puede aumentar o disminuir dependiendo de los hábitos de higiene, dieta y flujo salival. Cuando las capas más profundas ya no tienen oxigeno ni nutrimentos los productos de desechos se acumulan y van muriendo los microorganismos. Matriz orgánica intracelular Constituye aproximadamente el 30% de la placa dentobacteriana y está formada por: Glucoproteínas Proteínas 28 Hidratos de carbono Compuestos inorgánicos Agua proveniente de dieta, saliva y bacterias Los compuestos inorgánicos varían depende de esto el esmalte, los minerales y la edad pero entre ellos tenemos: Sodio Potasio Calcio Fosfato inorgánico Magnesio Hierro Fluor Agua (70 a 80%) Los hidratos de carbono provienen en su mayoría de la dieta así como de las glucoproteínas salivales, aunque hay intracelulares, extracelulares y capsulares. Estos pueden ser Glucanos (polímeros de glucosa) Fructanos (polímeros de fructosa) Heteroglucanos Los hidratos de carbono de la dieta para ser asimilados por las bacterias requieren de enzimas especiales como la amilasa alfa de la saliva, las oxidoreductasas y las deshidrogenasas. Las enzimas bacterianas (dextranasas, fructanasas, neuraminidasas, glucosidasas, glucógeno fosforilasas, entre otras) tienen una función fundamental. Las consecuencias del metabolismo de los hidratos de carbono son: 1. Cuando las glucoproteínas son atacadas por las bacterias, se separan residuos terminales de acido sìalico, se vuelven menos solubles y se depositan con facilidad alrededor de los microorganismos. 29 2. Se obtienen materiales de reserva factibles de ser degradados y movilizados en cualquier momento. 3. Los polisacáridos extracelulares, en especial los glucanos insolubles, facilitan la adhesión, la agregación y la coagregacion intermicrobiana en el esmalte. 4. Al formarse el tártaro dental, los hidratos de carbono dificultan el acceso de saliva y la salida de productos tóxicos. 5. Los ácidos producidos reducen el pH y de ese modo facilitan la desmineralización del esmalte. Por último las proteínas procedente de la saliva y de la dieta proporcionan nitrógeno y aminoácidos esenciales para la vida microbiana a su vez el amoniaco resultante es perjudicial para el huésped. Placa dentobacteriana subgingival Esta se localiza a partir del margen gingival en dirección apical, su formación se favorece cuando el pH del surco es más alcalino que el de la saliva y el líquido gingival tiene mayor cantidad de sales. En este tipo de placa existe poca matriz intercelular, salvo en las zonas adheridas al diente, por lo cual las fuentes nutricias son endógenas por ejemplo del liquido gingival o interbacteriana. Los microorganismos existentes dependen de la profundidad a la que se encuentren, por ejemplo cerca del margen dontogingival predominan los microorganismos grampositivos, en la porción apical el potencial de oxidorreduccion es más bajo lo cual permite el desarrollo de microorganismos anaerobios facultativos, bacilos gramnegativos anaerobios y bacterias anaerobias estrictas. Placa dentobacteriana fisural Este tipo de placa se forma en fosetas y fisuras apenas tiene matriz extracelular y contiene abundantes restos de alimentos. En ella abundan los cocos grampositivos sobre 30 todo estreptococo sanguis y S. salivarius, también se desarrollan lactobacilos, Corynebacterium matruchotii, especias de veillonella y estreptococo mutans, el cual puede construir el 40% de la colonización bacteriana cuando hay caries activa Placa dentobacteriana proximal Está situada en los espacios interproximales en dirección apical, aquí predominan Actinomyces viscosus y Actinomyces naeslundii. Pero también se detectan estreptococo sanguis, Actinomyces israelii, especias de veillonella y algunos bacilos gramnegativos anaerobios estrictos como las especias de selenomonas, Porphyromonas, Provotella y Fusobacterium. En las caries abundan estreptococo mutans y lactobacilos. Placa dentobacteriana radicular Esta se desarrolla cuando el cemento radicular se expone al microambiente bucal, ya sea por retracción gingival en edad avanzada o por enfermedades del periodonto, también se forma en áreas interproximales y a lo largo de la unión cemento esmalte. 7 Estadios de la formación de la placa La formación de la placa dental la podemos dividir en 3 estadios: En la primera fase las glucoproteínas de la saliva son absorbidas en la superficie externa del esmalte dentario, produciendo una película orgánica, delgada, acelular y carente de estructura, conocida como película adquirida. El segundo estadio de formación de la placa comprende la colonización selectiva, de la película por bacterias adherentes específicas, aunque las bacterias pueden en algunos casos iniciar la formación de la placa en ausencia de la película adquirida, con mayor frecuencia, existe una capa que separa la superficie del diente de la capa más profunda de microorganismos. 31 El paso último de la formación de la placa a veces es conocido como maduración de la placa, engloba la multiplicación y el crecimiento de más bacterias sobre las que están inicialmente. El cuerpo de la placa dentobacteriana, contiene muchas cepas de bacterias, es mantenido unido por adherencias interbacterianas en gran medida por los glucanos extracelulares. (9) FIGURA 3 Metabolismo de la placa Para las bacterias de la placa la principal fuente de energía son alimentos que contengas altos índices de carbohidratos, así la placa metaboliza carbohidratos fermentables (sacarosa) con la resultante formación de varios ácidos organismos como subproductos y una consiguiente caída del pH. Estos ácidos son los responsables de iniciar con la caries dental. 32 Sin embargo no todas las bacterias de la placa metabolizan carbohidratos. Algunas tienen capacidades proteolíticas y utilizan proteínas como fuentes de energía con la formación final de bases. La formación de estos materiales básicos y los valores de pH altos pueden favorecer la enfermedad periodontal. Y promover la precipitación de calcio y fosfato en la placa como tártaro dental. Sabemos que como se ha venido estudiando estreptococo mutans en presencia de sacarosa en la dieta forma polisacáridos extracelulares, pero también se sabe que varias cepas bacterianas sintetizan polisacáridos semejantes al glucógeno dentro de las células bacterianas. Estos polisacáridos se denominan polisacáridos intracelulares, no funcionan como componentes de la matriz de la placa sino que pueden servir como fuente de energía para las bacterias durante los periodos en que no hay aporte de carbohidratos a la dieta. Así los pacientes cuya placa contiene estos microorganismos formaran ácidos aun cuando estos estén ayunando, la producción de ácidos en la placa durante las horas de sueño cuando los mecanismos de defensa de la boca (salivación, movimientos de los labios, lengua y carillos) están en reposo es particularmente peligrosa desde el punto de vista de la formación de caries. 10 INTRODUCCION (CARIES DENTAL) Las enfermedades bucodentales, como la caries dental es un problema de salud de alcance mundial que afecta a los países industrializados y, cada vez con mayor frecuencia, a los países en desarrollo, en especial entre las comunidades más pobres, ha afirmado hoy la Organización Mundial de la Salud (OMS). Al anunciar las conclusiones del informe mundial sobre salud bucodental, la OMS ha declarado que se estima que cinco mil millones de personas en el planeta han sufrido caries dental. 11 33 La caries dental es una enfermedad que se caracteriza por una serie de complejas reacciones químicas y microbiológicas que traen como resultado la destrucción final del diente si el proceso avanza sin restricción. Se acepta casi universalmente que esta destrucción que avanza hacia adentro desde la superficie dentaria externa es el resultado de ácidos producidos por bacterias en el medio ambiente inmediato del diente. Clínicamente la caries dental se caracteriza por un cambio de color, perdida de translucidez y descalcificación de los tejidos afectados. A medida que el proceso avanza se destruyen tejidos y se forman cavidades, este estadio del proceso se denomina periodo de cavitación, Etiología de la caries Son varios los factores que desempeñan algún papel en la formación de caries por lo que se dice que la caries es una enfermedad multifactorial. Keyes ha representado diagramáticamente los 3 factores principales requeridos para el desarrollo de la caries como 3 círculos que se superponen parcialmente, Un circulo representa el agente (microorganismo), el otro el medio ambiente (sustrato) y el otro el huésped (diente) Newbrun he agregado un cuarto circulo, es decir el tiempo, lo que significa que para que se produzca caries es necesario que exista un periodo de tiempo. 34 Así los microorganismos criogénicos deben actuar sobre un sustrato criogénico para crear un ambiente que llegue a la caries, que se extienda sobre un periodo en el diente susceptible. Hidratos de carbono en la dieta: figura 3 El factor ambiental más importante de la caries dental es la presencia de hidratos de carbono en la dieta, existe una gran cantidad de evidencias tanto de estudios sobre animales como de ensayos epidemiológicos que indican fuertemente que en ausencia de hidratos de carbono fermentables la caries dental no se desarrolla. El estudio clínico más notable sobre la relación de la dieta y la caries dental en humanos es el clásico estudio de Vipehom. Aquí la actividad de caries en pacientes adultos que residen en un instituto de enfermos mentales en Suecia fue seguida durante varios años mientras se controlaba muy de cerca la composición de la dieta y los horarios de comidas. Se observo que el nivel de caries se relacionaba mucho mas con la frecuencia de ingesta de sacarosa que con la cantidad de ingesta total de sacarosa ingerida. Además las formas solida retentivas de azúcar resultaron mas cariogenicas que las liquidas.12 35 Otro estudio que comprendía una población distinta, también de internados mostro que la restricción del azúcar y de otros hidratos de carbono refinados de la dieta de los niños traía como resultado una marcada reducción en la caries dental. Estos hallazgos sugieren una disminución en la ingesta de hidratos de carbono fermentables, aun con una mala higiene bucal se reducirán los niveles de caries. A medida que ha avanzado la investigación odontológica durante la última década, se han conocido mejor los mecanismos por medio de los cuales los hidratos de carbono de la dieta contribuyen al proceso carioso estos pueden resumirse de la siguiente manera: 1. Los hidratos de carbono ingeridos son convertidos por las bacterias a polisacáridos extracelulares adhesivos. Estos llevan a la adhesión de colonias bacterianas entre si y a la superficie dentaria (formación de placa) 2. Las bacterias de la placa usan los hidratos de carbono de la dieta como fuente de energía, el resultado de este proceso metabólico es la formación de ácidos orgánicos que disuelven a los minerales del diente. 3. Los hidratos de carbono de la dieta pueden ser convertidos a polisacáridos de almacenamiento extracelular. Estos polisacáridos cuya estructura recuerda la de la aminopectina pueden ser usados como fuente de energía durante el tiempo en que no hay carbohidratos exógenos, como resultado de este almacenamiento se incrementa el periodo durante el cual los ácidos son producidos por microorganismos. 4. El hidrato de carbono en la dieta puede ser también metabolizado en polisacáridos de reserva extracelular, estos polisacáridos también pueden ser utilizados por los microorganismos durante los periodos de ayuno y aumentar el tiempo durante el cual se forman ácidos en la placa. 10 36 Teoría de la etiología de caries dental: Son dos las teorías, endógenas y exógenas, y dependen de la dirección de los fluidos: Endógenas HIPOCRATES 456 a. C. Afirmaba que los fluidos afectados (causa dentro del organismo), iba de adentro hacia fuera, es decir, primero por la pulpa, dentina y esmalte. GALENO 130 d. C. (Inflamatoria) Afirmaba que había un desequilibrio encefálico y hacía producir fluidos a la pulpa, ésta se inflamaba y se dirigía al esmalte. JOURDAIN (1734-1816) Afirmaba que alteraciones metabólicas producían inflamación del odontoblasto y ésta odontitis, comprometía descalcificación dentinaria y producía caries. Exógenas Teoría quimioparasitaria (Acidógena) Miller realizó sus experimentos en el laboratorio de Koch. Demostró que una cantidad de microorganismos bucales tenían esta propiedad y que el ácido láctico era uno de los principales medios. Esta teoría propone la producción de ácidos en la superficie dentaria o cerca de ella mediante la fermentación microbiana de los hidratos de carbono, los ácidos disuelven los cristales de apatita. 37 Teoría proteínica o proteólisis Sugerido por Gothreb en 1944. Según esta teoría la matriz orgánica sería atacada antes que la fase mineral del esmalte. La teoría propone que las enzimas proteolíticas liberadas por las bacterias bucales podrían destruir la matriz orgánica del esmalte, con el resultado de un aflojamiento de los cristales de apatita y colapso del tejido. Teoría proteólisis - quelación Originada por Schatz y Martin en 1955, propone que algunos de los productos de la acción microbiana sobre el esmalte, la dentina, los alimentos y los componentes de la saliva pueden tener la propiedad de formar complejos o quelatos con el calcio, los quelatos pueden formarse con valores de pH neutros o alcalinos, la teoría sugiere la posibilidad de que la desmineralización del esmalte pueda surgir sin formación de ácido. Teoría organotrópica Leungreber dice que la caries no es una destrucción local de los tejidos, sino una enfermedad de los órganos dentales. Considera el diente como parte de un sistema biológico compuesto de pulpa, tejidos duros y saliva. Todo agente capaz de destruir enlaces polares o de valencia romperá el equilibrio produciendo caries. 38 Flora oral en relación con caries dental Microbiología oral.- El origen de la microbiología oral coincide con el descubrimiento de las bacterias. Lienwen Hock observó en su saliva y en material depositado en los dientes que denominó "MATERIA ALBA" (animaliculos) y así los comunicó. En 1880 Miller en su libro "The microorganism of the human mouth"; expone su teoría quimioparasitaria, en virtud de la cual los microorganismos acidógenos, actúan sobre los carbohidratos de la dieta acumulados en la boca producirían ácidos que desmineralizarían el esmalte y la dentina. En 1897, Williams encontró en las zonas con caries incipientes acumulaciones de bacterias adheridos al esmalte cariado pensando que la producción de ácidos en dichas zonas era la responsable del proceso. En 1898 Black describió sobre la superficie de los dientes acumulaciones bacterianas especialmente constituido por los que hoy se conoce como Estreptococos, acuñó el término "placa" para definir estos depósitos. En 1944 Stephan mostró que no todas las placas producen caries administrando carbohidratos fácilmente fermentables y mediante unos electrodos especiales, medio seguidamente el pH intraplaca. En zonas con caries el pH descendía rápidamente cosa que no ocurría en las zonas con caries inactivas, demostrando la importancia de los ácidos y el descenso de pH en la desmineralización del esmalte buscándose desde el principio de bacteria o bacterias causantes. Durante los años 30 y 40 de este siglo los lactobacilos fueron especialmente implicados. 39 En 1924 se demostró la importancia de otra bacteria descubierta por Clart, Estreptococos mutans, es un agente importante en la caries dental, ésta es una enfermedad multifactorial en la que influye al menos una dieta, el hospedador, el factor tiempo y los microorganismos no solo sino muchos. Una indicación de que la flora oral era esencial fue que el diente no desarrollaba caries mientras no emergiera. Una vez que había emergido y se había puesto en contacto con la flora oral se hacía susceptibles a la caries dental. Los lactobacilos son un grupo característico de bacterias orales aunque numéricamente constituyen una fracción mínima. Sus cantidades son variables, pero es probable que algunas estén presentes en todas las cavidades orales humanas inmediatamente después del nacimiento, aún cuando probablemente no alcancen una proporción significativa. Son acidúricos con un pH óptimo de generalmente 5,5 a 5,8. La fermentación de los carbohidratos por los lactobacilos es variable con las especies, aunque generalmente es bastante activa. Las diferencias en la acción de los lactobacilos orales sobre la glucosa los divide en especies homofermentativas, las cuales producen principalmente ácido láctico (más de 65%) y un considerable número de otros productos terminales (principales ácido acético y etano) incluyendo Gas (generalmente bióxido de carbono). Aunque estas características son cariogénicas y estas bacterias presentan poca afinidad por la superficie del diente, lo que difícilmente lo implica en el inicio de caries dental en superficies lisas, no obstante son los primeros microorganismos en el frente de avance 40 del proceso carioso en la dentina. Los lactobacilos se encuentran en mayor frecuencia como agentes transitorios en la boca de infantes, representan el 1% de la flora oral. El Lactobacilo casei y Lactobacilo fermenteum son las especies orales más comunes, su hábitat es la dentina de las lesiones cariosas profundas. Relación de los estreptococos con la caries Los estreptococos facultativos forman el grupo más grande en la cavidad oral promediando casi en la mayoría casi la mitad de las cuentas variables de saliva y dorso de la lengua, y aproximadamente 1/4 de las cuentas variables de la placa y el surco gingival. Se ha calculado que los estreptococos son aproximadamente mil veces más numerosos que los lactobacilos de la flora microbiano oral. Son igualmente abundantes en las cavidades de dientes de niños así como de adultos. Los estreptococos han sido aislados más frecuentemente de la placa precariosa, transaccional y cariosas sobre el esmalte que cualquier otra especie de bacteria. Se encuentra presente de manera consistente en caries de fisuras, en la interproximal y en diferentes etapas de la dentina cariada. Los estreptococos pueden invadir hacia delante de los que considera el frente de avance de la caries dentinal profunda, tal como lo indica el hecho de ser el invasor más frecuentemente de la pulpa vital de los dientes cariados, siendo su ruta de invasión a lo largo o entre los túbulos dentinales. Otra característica de los estreptococos orales relacionados con su rango de crecimiento y producción de ácido, observándose que exceden a los de cualquier otro microorganismo oral. De los estreptococos el más implicado en el inicio y desarrollo de la caries es el estreptococo del grupo mutans. La producción de polisacáridos extracelulares a partir de 41 la sacarosa y concretamente de glucanos insolubles, desempeña un papel fundamental en la colonización y crecimiento de estreptococos del grupo mutans sobre el diente. Esta gran afinidad por las superficies dentales se debe a fenómenos de adhesión, agregación y congregación. La síntesis de polisacáridos intracelulares por Estreptococos mutans y su capacidad de metabolizarlos son factores de virulencia, ya que proporcionan a la célula un substrato de donde obtener la energía y mantener la producción de ácido durante largos períodos de tiempo. Además producen dextranasas y fructonasas, enzimas capaces de metabolizar los polisacáridos extracelulares, sobre todo los glucanos solubles, favoreciendo la producción de ácido y constituyendo un substrato en los períodos en que disminuye el aporte exógeno. A partir del metabolismo de la sacarosa, estos microorganismos producen principalmente ácidos lácteos, que son fundamentales en la virulencia debido a que aparentemente es el ácido más potente que interviene en la desmineralización del diente. Además de acidógenos, los estreptococos del grupo mutans son acidófilos o tolerantes al ácido, propiedad que le es muy necesaria para sobrevivir y desarrollarse con un pH bajo, y acidúricos o capaces de seguir produciendo ácido con un pH bajo. Otra característica es su corto efecto post pH, que puede definirse como el tiempo necesario para recuperar su actividad de crecimiento habitual cuando, tras estar sometidos a un pH bajo, este vuelve a la normalidad. Estas especies bacterianas son las que consiguen alcanzar más rápidamente el pH crítico de 4,5 necesario para iniciar el proceso de desmineralización. 42 Relación de los actinomices con caries dental Actinomyces predomina en la placa que cubre las lesiones de la superficie de la raíz en los dientes humanos, aunque su papel en el inicio de las mismas es difícil de demostrar. Es interesante destacar que además de su poder acidógeno, posee fimbrias que están implicadas en su capacidad de adhesión y de congregación. El papel que desempeñan los polisacáridos extracelulares es más nutricional que adherencia. Factores de cariogenicidad de Actinomyces: Poder acidógeno. Poseen fimbrias. Actividad proteolítica moderada. Se ha mostrado 5 especies en la flora oral: Anaeróbicos: A. Bovis A. israelii Anaeróbicos: A. viscosus A. naeslundii A. odontolyticus 43 Bacterias proteolíticas y caries dental Las proteínas y sus productos de degradación orales y en la placa, sobre todo los lactobacilos y estreptococos acidogénicos. La saliva completa para ser deficiente en las proteínas y aminoácidos requeridos por las bacterias orales acidogénicas. Las bacterias proteolíticas orales pudieran tener función en el medio oral para proporcionar proteínas y aminoácidos requeridos por la flora oral. En general, las proteínas de la dieta no están disponibles en cantidades importantes para las bacterias de la placa, debido a su corta permanencia en la boca y la falta de enzimas proteolíticas en la saliva que fluye libremente. Por otra parte, la placa contiene proteasas las glucoproteínas, células epiteliales, leucocitos, bacterias y alimentos impactados, para proporcionar proteínas y aminoácidos necesarios para las bacterias cariogénicas de la placa. Localización de la flora oral relacionada con caries 44 Una evaluación cuidadosa a cerca de caries indica que diferentes organismos muestren selección de la superficie del diente que atacan y también sugiere que existe por lo menos 4 procesos relacionados con ella. Caries de las hendiduras y fisuras Esta es la más común de las lesiones cariogénicas encontradas en el hombre. Muchos organismos pueden colonizar en las fisuras, las cuales proporcionan una retención mecánica para las bacterias. Hay una enorme variedad de microorganismos capaces de iniciar las caries en las hendiduras y fisuras. Flora de las hendiduras y fisuras Es muy poco lo que se sabe a cerca de la microbiología de las lesiones de fisuras en el hombre y esto se debe principalmente a las dificultades que se presentan para tomar muestras de ellas en la base de la fisura. La gran variación existente en la microflora presente en dichos lugares indica que cada fisura representa un sistema ecológico independiente. En general, los cocos grampositivos predominan en la microflora (70% a 90% de la flora), esencialmente el S. sanguis, mientras que los fusiformes, Spirillae y espiroquetas no aparecen en ella. Frecuentemente se ha aislado S. mutans en número considerable, en la placa de las lesiones presentes en la superficie lisa y fisuras. Caries radicular 45 Se ha asociado a los bastoncillos filamentosos grampositivos con esta lesión, incluso (actinomyces). Las cepas de Nocardia, S. mutans, S. sanguis, además de ocasionar la caries del esmalte, pueden producir también caries radicular. Puede dividirse de acuerdo con su función en productores de ácidos licueficantes, proteolíticos y productores de pigmento; el que los estreptococos y los lactobacilos eran los más abundantes de las especies acidogénicas residentes y que los lactobacilos eran los más acidúricos, Howe y Hatch fueron los primeros en postular que los lactobacilos pueden intervenir en la fase descalcificante de la caries dental. En 1922 dos grupos de investigadores, Rodríguez y McIntosh y Lazaruz Barlow encontraron lactobacilos en las lesiones de caries y demostraron su alto potencial de producción ácida y de su capacidad de sobrevivir a los ácidos que producen. Los estreptococos están relacionados con la caries dental, Miller (1890) encontró estreptococos en la cavidad oral; de 1900 en adelante, los estreptococos han recibido una atención considerable como agente causal de la caries dental. Sieberth 1901 aisló los estreptococos primero a partir de dentina cariada. Niedergesan, en 1915, confirmó esta observación. Gordon, 1905, y Kligler y Gies, 1915, encontraron que el estreptococo era el microorganismo predominante de la boca. Dichos postulados se basaron principalmente en la abundancia de estreptococos en la cavidad oral, su presencia en la caries dentinal profunda y la consistencia como un agente causal de pulpitis acompañando a la caries dentinal profunda, sin exposición de la pulpa. La determinación del papel de los estreptococos en la caries dental fue aclarada enormemente por una serie de investigaciones destinadas a establecer el potencial productor de la caries de una sola capa o especie bacteriana, primero en ratas blancas gnotobióticas y después en hamsters, mediante estudios de las causas de la variabilidad de la caries dental en animales de experimentación y por el establecimiento de un agente 46 transmisible, al menos en la caries dental, en animales de experimento y probablemente en la caries humana. Flora de la caries radicular La caries radicular es una pequeña lesión y progresiva que se presenta en la superficie de la raíz e incluye la invasión microbiana y de placa. Es diferente a la erosión, abrasión y resorción idiopática, afecciones que también atacan la superficie de la raíz. Se le conoce con diversos nombres: Caries cemental, caries cervical, caries radicular e incluso caries senil. Por lo general, este tipo de caries tiene una ligera profundidad (menos de 2mm) no se ve claramente, es suave al tacto, con frecuencia es una zona decolorada y se caracteriza por la destrucción del cemento con penetración de la dentina subyacente. A medida que la lesión progresa, se extienden en forma de circunferencia en lugar de hacerlo en profundidad. Algunos organismos presentes en la caries radicular son diferentes a los que se encuentran en lesiones de otras superficies lisas debido a que la lesión inicial es en el cemento y la dentina y no en el esmalte. Las muestras bacteriológicas de la placa que cubre la caries en la raíz producen predominantemente Actinomyces viscosus. Las muestras microbianas tomadas de la dentina de personas con caries radicular, revelaron también la presencia de otras especies del género actinomyces (A. naeslundii, A. odontolyticus, A. eriksonii) así como también Rothia dentocariosa, Nocardia y S. mutans. 47 Caries en la superficie lisa de los dientes Se ha probado que un número limitado de organismos pueden colonizar en la superficie lisa en cantidad suficiente para causar deterioro dental en animales de experimentación. En este aspecto el S. mutans es de gran significado. Caries de la dentina profunda Debido a que el medio presente en las lesiones en la dentina profunda es diferente a aquel existente en estas zonas no es de sorprenderse que la flora que hay en ellas sea también distinta. El organismo que predomina en este tipo de caries es el Lactobacillus, con frecuencia se encuentran bastoncillos y filamentos anaerobios grampositivos aislados, tales como Arachnia, Bífido bacteria, Embacteria y Propioniabacteria. Los actinomyces, Rothia y Bacillus también se encuentran en la parte frontal de las lesiones de la dentina profunda. La frecuencia con que aparecen los cocos facultativos grampositivos es reducida. Bioquímica de la caries La caries es una enfermedad de naturaleza esencialmente bioquímica, desencadenada por reacciones que ocurren sobre los tejidos dentarios bajo condiciones ecológicas bucales específicas que clínicamente determinan lo que se conoce como 48 Riesgo Cariogénico. Lehner en 1983 define a la caries como "El resultado final de una compleja secuencia de procesos bioquímicos bacterianos que terminan en la producción de ácidos que atacan la superficie del diente causando caries". Todos los aspectos de la caries -etiopatogénicos, ambientales y sus repuestas defensivas- están sustentadas por reacciones bioquímicas. Una discusión de la caries debe comprender al menos el metabolismo bacteriano relacionado con la formación de la placa, producción de ácidos y la regulación ácidobase de la misma, sin olvidar la importante participación del fluido salival. Bioquímica enzimática extracelular del estreptococo mutans El germen con mayor actividad cariogénica, tanto en animales como en humanos, es el Estreptococo mutans, el que realiza sus principales actividades metabólicas extracelulares en función de dos enzimas cuyo substrato son moléculas de carbohidrato. Las dos enzimas extracelulares del mutans son la Dextranosacarosa y la Glucosiltransferasa. La dextranosacarosa ensambla las unidades de glucosa en polímeros lineales mediante enlaces del tipo alfa 1-6. Posteriormente, y dependiendo de la disposición de substratos del medio, entra en actividad enzimática la Glucosiltransferasa. Esta última ensambla las unidades lineales preexistentes en una forma ramificada, resultando una macromolécula insoluble y adherente que forma gran parte de la matriz extracelular de la placa bacteriana. Diversos investigadores han discutido sobre los enlaces de este poliglucano insoluble. Enlaces alfa 1-3, alfa 1-4 e incluso alfa 1-5 han sido propuestos y cuestionados. Cabe recordar que en ocasiones el desacuerdo ha sido originado por las diferentes y numerosas formas estequiométricas espaciales utilizadas por los autores para representar tanto las macromoléculas de sacarosa como las de glucosa. Por su parte la glucosa -al igual que la sacarosa- cuando se encuentra en solución salival, presenta diferentes configuraciones diasteroisómeras. Los enlaces ramificados confieren principalmente adherencia a la matriz extracelular o a la placa. Para lograrlo, las cepas deben tener la capacidad genética de desdoblar su ADN para llevar a cabo la síntesis de DS y GTF. Además deben tener la capacidad de transportar dichas enzimas a través de la membrana y pared celular hasta 49 el medio externo. Luego, bajo condiciones ambientales favorables, serán estas enzimas responsables en gran medida de la colonización estreptocócica primaria de la superficie dentaria. Recientemente, el hallazgo de anticuerpos específicos para los marcadores de superficies del Estreptococo mutans en la película adquirida (Reinholdt 1987), sugieren que su presencia puede jugar un papel importante en la selectividad inicial de la colonización estreptocócica primaria. Si las condiciones ambientales otorgan un excedente de glucosa, ésta es utilizada por el Estreptococo mutans como nutriente y para ello debe ingresarla al protoplasma celular. Ingreso de azúcar al estreptococo mutans La mayoría de los nutrientes bacterianos aumentan abruptamente su concentración en el medio extracelular durante los períodos de ingesta alimenticia. En especial el azúcar aumenta su concentración rápidamente creando una gradiente de difusión hacia el medio intracelular. Previo a la ingestión alimenticia, el azúcar extracelular disponible se encuentra en una concentración inferior respecto al protoplasma bacteriano. En la cavidad bucal suelen haber largos períodos sin aporte exógeno de nutrientes bacterianos. Los microorganismos viven bajo condiciones de abundancia y escasez debiendo estar preparados para ello. El transporte de azúcar a través de la membrana celular desde el ambiente externo al interior del citoplasma requiere de unas proteínas específicas denominadas genéricamente Portadoras (P). Si la concentración externa de azúcar es muy elevada, estas proteínas facilitan el transporte de azúcar sin consumo de energía. Cuando la concentración externa del azúcar es baja y se alcanza un nivel más alto en la célula, el transporte requiere de energía. Los Estreptococos utilizan la energía almacenada en la membrana celular en forma de gradiente electroquímica de protones. Cuando la concentración extracelular de glucosa es baja, el azúcar debe ingresar en contra de una gradiente de concentración. La energía se obtiene acoplando la entrada de un azúcar a la de un protón que va por gradiente electroquímica montado en una enzima portadora. Este sistema de ingreso de azúcar bacteriano se denomina Sistema Permeasa. El Estreptococo mutans utiliza dos sistemas de ingreso de glucosa; uno de baja afinidad que funciona a altas concentraciones con una constante de saturación de 100mM de glucosa (Ks=100mM), y otro con alta afinidad por la glucosa que funciona 50 cuando las concentraciones son menores. Este segundo mecanismo de ingreso de glucosa al protoplasma bacteriano es el Sistema Fosfotrasferasa-Fosfoenolpiruvato que posee una alta afinidad por la glucosa con una Ks de sólo 5mM, por tanto funciona a bajas concentraciones de glucosa y está reprimido en concentraciones altas de azúcar y a bajo pH. Por el contrario, en condiciones muy excesivas de glucosa se activan rápidas vías metabólicas de eliminación de desechos a fin de evitar acúmulos tóxicos de intermediarios de la glucolisis en el protoplasma bacteriano. Los principales sistemas de ingreso de glucosa en el Estreptococo mutans aparecen diagramados en la Fig. 4.1. Figura 4 Fig. 4.1: Principales sistemas de ingreso de glucosa a través de la membrana del Estreptococo mutans. La gradiente electroquímica aparece parcialmente esquematizada: 1. Corresponde al sistema permeasa acoplado a un protón. 51 2. Representa el sistema fosfoenoltransferasa-fosfoenolpiruvato que confiere al mutans una alta afinidad por la glucosa (Ks=5mM) 3. Existe un ingreso pasivo de glucosa común en condiciones de ingesta de azúcares. 4. Intermediario de la glicolisis. 5. Vía de conversión de piruvato a lactato por acción de la deshidrogenasa láctica (DL). Ingresada la molécula de glucosa al protoplasma bacteriano, la enzima 1-6 glucosidasas (4) la modifica a glucosa 6 fosfato. La glucosa 6 fosfato puede ir a constituir amilopectina de reserva energética por acción enzimática de sintetasas de glucógeno. La amilopectina puede a su vez degradarse y parte de ella quedar como dihidroxiacetona-P, pigmento de importancia en la coloración de la caries. Lo más frecuente es que sea utilizada por el estreptococo mutans como un catabolito. Bioquímica protoplásmica del estreptococo mutans Las reacciones enzimáticas intracelulares del Estreptococo mutans y de otras cepas con capacidad cariogénica son parte del ciclo metabólico bacteriano. El ciclo metabólico en todos los organismos vivos posee dos grandes vías: por una parte el catabolismo que degrada macromoléculas para obtener energía de los enlaces interatómicos, la que luego será utilizada en la otra gran vía, la anabólica, encargada de construir y ensamblar los componentes esenciales de la materia viviente. Las enzimas catabólicas del Estreptococo mutans necesarias para metabolizar energéticamente aparecen enumeradas en la tabla 4.1. El catabolismo bacteriano busca finalmente obtener energía liberándola de los enlaces carbono-carbono de la glucosa. Teóricamente la máxima liberación de energía se obtiene por la oxidación completa de la glucosa hasta anhídrido carbónico y agua. Las enzimas intracelulares del mutans sólo le permiten oxidar hasta dos triosas (dos moles de ácido láctico obteniendo dos moles de ATP). Así las enzimas de la tabla 4.1 52 catalizan la degradación energética de la glucosa terminando en la producción de ácidos pirúvico y láctico (Fig. 4.2). TABLA 4.1: ENZIMAS GLUCOLÍTICAS REACCIÓN ENZIMA 1 Amilo-(alfa-1,4-alfa-1,6)-transglucosilasa 2 Sintetasa de glicógeno 3 Fosforilasa 4 Amilo-1,6-glucosidasa 5 UDPG profosfolilasa 6 Fosfoglucomutasa 7 Hexocinasa 8 Fosfoglucoisomerasa 9 Fosfohexocinasa 10 Aldolasa 11 Isomerasa de triosafosfato 12 Deshidrogenasa de alfa-glicerofosfato 13 Deshidrogenasa de fosfato de gliceraldehido 14 Fosfoglicerilcinasa 15 Fosfoglicerilmutasa 16 Enolasa 17 Cinasa de piruvato 53 18 Carboxilasa de piruvato 19 Deshidrogenasa de alcohol 20 Deshidrogenasa de lactato Enzimas bacterianas necesarias para que una cepa metabolice enérgicamente la glucosa. La tabla considera desde la Amilo (alfa1-4 - alfa 1-6) transglucosilasa, enzima que utiliza el mutans cuando no dispone de carbohidrato exógeno y debe utilizar su amelopectina intracelular de reserva. Cuando las bacterias utilizan directamente la glucosa ingresada a la célula, la vía enérgica suele comenzar por la acción de las enzimas número 7-8-9 o 10 de esta tabla En bioquímica se utilizan los términos Glucolisis, Fermentación o Glicolisis para denominar un esquema general de reacciones enzimáticas similares que ocurren en células bacterianas y/o animales por medio de las cuales se produce la conversión de la glucosa en ácido pirúvico y/o láctico con obtención de energía. El factor que determina la mayor producción de ácido pirúvico o láctico es la presión de oxígeno (PO2) del medio en que se desarrolle la vía metabólica. A menor presión de oxígeno, la ecuación se decía hacia la formación de grandes cantidades de ácido láctico. En una placa bacteriana cariogénica y madura, las zonas más profundas adosadas al esmalte son anaerobias; entonces la obtención de energía por las cepas allí residentes (principalmente Estreptococos mutans) se efectúa por la vía de la glicolisis anaerobia, cuyo catabolito final es el ácido láctico responsables molecular directo de la caries dental. Los términos lactato y piruvato son sinónimos respectivos de ácido láctico y pirúvico. Se utilizan expresamente en bioquímica para poner de manifiesto que a pH de la célula en ácido está predominantemente disociado. De este modo, en la glicolisis anaeróbica de cepas cariogénicas la disociación del ácido láctico proporciona los 54 hidrogeniones (H+) suficientes que acumulados en la superficie del esmalte dan inicio a la enfermedad. Es interesante señalar que las reacciones de esta naturaleza no requieran oxígeno. Regulación de la glicolisis La glicolisis realizada por el Estreptococo mutans en condiciones normales no sólo libera ácido láctico extracelular, sino también ácido fórmico, etanol y acetato. Degradada la glucosa hasta piruvato, una parte es excretada como lactato por la deshidrogenasa láctica (DL). (Fig. 4.1). Los otros metabolitos de desecho son el etanol y el formato cuando se pasa por la vía de acetil-CoA y el acetato por vía CoA. Las vías del Estreptococo mutans para eliminar formato, etanol y acetato son más lentas ya que requieren una mayor y más compleja participación enzimática que de pirúvico a láctico. Las condiciones de nutrientes bacterianos son alternantes. En períodos de ingesta de alimentos, las concentraciones de azúcar pueden aumentar 10 mil veces. Los microorganismos sufren un período rápido de ingreso de glucosa al protoplasma, se producen niveles tóxicos de intermediarios de la glicolisis y las células mueren. Paradójicamente, los azúcares de la dieta son en grandes concentraciones una amenaza para las bacterias: se llama "azúcar asesino" y al proceso en general "muerte acelerada por substrato". Las células se auto protegen del azúcar asesino activando la vía glucosa-piruvatolactato. A su vez se abre la "Puerta del Lactato" aumentando la velocidad de la glicolisis y la síntesis de glucógeno intracelular de reserva. Con la apertura de la puerta del lactato se logra una mayor velocidad de la degradación del piruvato especialmente por la deshidrogenasa láctica. Además se producen grandes cantidades de ácido láctico, fórmico, acético y etanol que drenan abundante y rápidamente por la puerta de lactato. La conversión de piruvato por la vía lactato deshidrogenasa parece ser una propiedad importante de algunas bacterias en protección contra alzas tóxicas de substrato. La eficacia de esta "puerta del lactato" podría deberse a que la DL es una enzima constitutiva para los Estreptococos mutans y Lactobacilos. Ante exceso tóxico de azúcares la puerta parece ser una clave característica de muchas bacterias relacionadas con la caries dentinaria. Por la apertura de la puerta, los ácidos se forman 55 rápidamente en cantidades tan altas que el fosfato de calcio mineral se disuelve rápidamente y se inicia la caries a nivel molecular. Descripción del proceso carioso En la teoría quimioparasitaria aceptada en la actualidad (Miller, 1890), el mecanismo de la caries consiste en que los microorganismos de la superficie del diente producen ácidos orgánicos, incluyendo en particular el ácido láctico fuerte, el cual disuelve el mineral del diente. Hay evidencia directa de este mecanismo a partir de una disminución en el pH en las lesiones de caries. Se han propuesto otras teorías, las cuales ya no apoyan que sea el mecanismo predominante. En la teoría proteolítica se pensaba que el ataque inicial se debía a enzimas proteolíticas producidas por las bacterias que disolvían la matriz orgánica del esmalte. Aun cuando la eliminación enzimática de proteínas ya no es de primordial importancia, sí lo son las bacterias que producen enzimas proteolíticas, que afectan la matriz del esmalte, las cuales son la causa primaria de la eliminación de colágeno de la dentina en una etapa posterior. También se ha propuesto la quelación del Ca++ a pH neutro por aniones de ácido carboxílico o derivados de carbohidratos que producen las bacterias como un mecanismo para eliminar la fracción inorgánica. A pesar de que un componente marginal de la disolución mineral puede deberse a la quelación, ahora no se piensa que sea un mecanismo importante en la formación de la caries. La primera evidencia de caries en una superficie lisa es la formación de una "mancha blanca" que puede verse después que se seca la superficie. Esta aparición se debe al efecto óptico del aumento en la porosidad. Esta porosidad se origina de la disolución de una parte del esmalte que realizan los ácidos que se difunden a su interior a partir de la placa dental, la cual se adhiere a la superficie del esmalte. Este patrón de ataque se mantiene hasta que los ácidos en difusión y la porosidad resultante llegan a la unión amelodentinaria; después de ésta se extiende una desmineralización más rápida de la dentina hacia la pulpa y en forma lateral por debajo del esmalte sano bajo la lesión inicial. En esta etapa, el esmalte de la superficie se encuentra intacto, y una lesión de caries no se detecta mediante el uso de una exploración dental; sin embargo, podría detectarse por radiografía. No obstante, después de una mayor desmineralización, el esmalte y la dentina se debilitan y la superficie del esmalte se rompe de manera que las 56 bacterias tienen un acceso directo al esmalte más profundo, y en ocasiones a la dentina. Las bacterias, principalmente los lactobacilos, parecen ser capaces de crecer a lo largo de los conductos dentinales más profundos en áreas que continúan estando muy mineralizadas. Algunas veces, si no se ataque el proceso, la lesión llega hasta la cavidad de la pulpa y las bacterias la infectan. La dentina y la pulpa, a diferencia del esmalte, son tejidos celulares vivos, que responden a las lesiones y al ataque de las caries. Las respuestas incluyen: mayor mineralización dentro de los tubos dentinales (esclerosis tubular), formación de dentina como reacción en la superficie pulpa-dentina, e inflamación de la pulpa. Algunas veces el ataque de caries se detiene, en parte como resultado de estas respuestas; y en parte debido a la actividad bacteriana reducida, en cuyo caso se forma una "lesión de caries detenida". MECÁNICA DE PROCESO La caries dentaria comienza con la desmineralización del esmalte, que resulta de la acción de los ácidos orgánicos producidos localmente por las bacterias. Además de la desmineralización, las bacterias también destruyen el contenido proteico del diente (especialmente la dentina). Por tanto, se trata de una enfermedad bacteriana que se transmite de un miembro a otros de la misma especie. En el desarrollo de esta afección, ciertas bacterias de la cavidad bucal se establecen en colonias en las superficies de los dientes, adoptando la forma de "películas" gelatinosas adherentes que reciben el nombre de placas dentarias; además de bacterias, las placas también contienen mucus, células descamadas y restos de comida. Las bacterias principalmente vinculadas con la descomposición dentaria en el ser humano son los estreptococos anaerobios llamados Estreptococos mutans. Gran cantidad de estos microorganismos habitan en la placa dentaria; puede hallárselos en bocas con dientes naturales o artificiales, y su número es marcadamente inferior antes de la erupción de los dientes y después que éstos se pierden. Además del Estreptococo mutans, el Lactobacilo acidophilus probablemente desempeña un papel menor en la producción de ácido en la placa. La viscosidad de la placa se debe al dextran, que se produce por la acción del Estreptococo mutans sobre la sacarosa de la alimentación. El dextran forma aproximadamente el 10% del peso y alrededor del 33% del volumen de 57 la placa, e incluye en su contexto a todo tipo de microorganismos bucales. Las bacterias de la placa, particularmente el Estreptococo mutans, actúa sobre la fructosa de los alimentos produciendo ácido láctico, que ocasiona descalcificación de esmalte (a un pH de 5,5 o menor). Durante la abundancia de carbohidratos en el medio bucal, las bacterias de la placa pueden acumular polisacáridos intracelulares (amilopectina), y, durante los períodos de carencia, pueden actuar sobre esta reserva para continuar produciendo ácido láctico. Por tanto, la placa y los carbohidratos de los alimentos tienen importancia en la iniciación de la caries del esmalte. Además, la velocidad con que comienza y se extiende la caries depende de la susceptibilidad del esmalte. En consecuencia, para que haya una lesión cariosa deben satisfacerse las tres condiciones, que son: existencia de una placa, tipo de alimentación y susceptibilidad del esmalte. Además de la formación de ácido, algunas de las bacterias de la placa producen enzimas proteolíticas que destruyen las porciones orgánicas del esmalte y dentina. Este mecanismo desempeña un papel importante en la descomposición de la dentina, que posee una considerable cantidad de componentes orgánicos. Sin embargo, la destrucción proteolítica también reviste importancia en la caries, pues permite que la caries progrese a través de las laminillas del esmalte y otros defectos ocupados por proteínas. 13 58 POLISACÁRIDOS EXTRACELULARES Estructura de los glucanos extracelulares bacterianos Los polisacáridos extracelulares sintetizados por las bacterias incluyen glucanos (aquellos formados por polimerización de unidades de glucosa), fructanos (los constituidos por unidades de fructuosa) y heteropolisacáridos, entre otros.11 Los glucanos constituyen uno de los componentes principales del biofilm de la placa dental. Se pueden distinguir 2 tipos fundamentales de glucanos extracelulares bacterianos: uno similar al dextrano y el denominado mutano. Ambos están formados por la polimerización de unidades de alfa D-glucosa unidas por enlaces o-glicosídicos alfa (1-3) y alfa (1-6) con diferente grado de ramificación.12 Dextrano En su estructura predominan las uniones alfa (1-6), su aspecto es gelatinoso y es soluble en agua. Se considera que puede servir de base para la síntesis de mutano.12,13 Mutano El mutano está constituido por un 67 % de uniones alfa (1-3), en un esqueleto contiguo al restante 33 %, que se presenta con uniones alfa (1-6), posiblemente como residuos lineales que se extienden desde los puntos de ramificación alfa (1-6). 59 De los residuos, 14 % son puntos de ramificación que aparecen como promedio cada 10 unidades de glucosa y la relación de residuos lineales alfa (1-3) en el esqueleto: residuos alfa (1-6) en la cadena lateral es de 5:2. El predominio de uniones alfa (1-3) y el alto grado de ramificación, le confiere aspecto fibrilar y lo hace insoluble en agua. Así, la habilidad del E. mutans de sintetizar mutano, es esencial para la colonización eficiente y el desarrollo de la caries.14 Síntesis de los glucanos extracelulares bacterianos Este proceso se produce por la acción de enzimas extracelulares sintetizadas por las bacterias, denominadas glucosiltransferasas (EC.2.4.1.5), que constituyen un factor de virulencia significativo en el inicio de la caries dental.8 Tipos de glucosiltransferasas (GTFs) Una clasificación para las GTFs se basa en las características de solubilidad en agua del glucano que producen. 15 Según este criterio el E. mutans produce 3 formas diferentes de glucosiltransferasas, cada una de las cuales sintetiza un polímero de glucano a partir de la sacarosa. 16 Estas son: 1. La GTF-I (GTFB) produce al glucano insoluble en agua denominado mutano, rico en uniones glicosídicas alfa (1-3). 2. La GTF-S (GTFD) produce un glucano similar al dextrano, pues es soluble en agua y rico en enlaces alfa (1-6). 3. La GTF-SI (GTFC) produce una mezcla de glucanos solubles e insolubles en agua.13, 14,15 60 Los símbolos que aparecen entre paréntesis (GTFB, GTFC y GTFD) corresponden a otra nomenclatura, basada en el nombre que recibe el gen que codifica a cada una de las enzimas. La GTFB y la GTFC, se localizan en la superficie de la célula, mientras que la GTFD es liberada hacia el espacio extracelular.17 Las GTFs son proteínas muy grandes de aproximadamente 1,300-1,700 residuos, que presentan gran homología de secuencias. La enzima puede dividirse en 2 dominios: un dominio catalítico (CAT) en el tercio N-terminal, que une e hidroliza a la sacarosa y un dominio de unión al glucano (GLU), en el tercio C-terminal, que contiene múltiples secuencias repetitivas. 2, 8, 12,18 Las predicciones sobre la estructura secundaria sugieren que las GTFs son miembros de la superfamilia alfa-amilasa y como tal contienen una estructura de barril-(alfa/beta). Después de una secuencia señal conservada, seguida de una región no conservada de aproximadamente 200 aminoácidos, el dominio catalítico de aproximadamente 1 000 aminoácidos, contiene aminoácidos conservados necesarios para la hidrólisis de la sacarosa. El tercio carboxilo-terminal consiste en una serie de secuencias repetitivas que participan en la unión al glucano.15, 19 Además del E. mutans, otros estreptococos bucales producen glucosiltransferasas y sintetizan glucanos a partir de la sacarosa. Entre ellos se encuentran el E. oralis, E. gordonii, E. sanguis, quienes producen un solo tipo de GTF que es capaz de sintetizar glucanos de diferentes proporciones de enlaces alfa (1-3) y alfa (-6). Por otra parte, el E. sobrinus produce 3 GTF-S y una GTF-I, mientras el E. salivarius produce 2 GTF-S y 2 GTF-I. Sin embargo, aún no se ha dilucidado el papel biológico de los glucanos sintetizados por estas bacterias. 61 Reacción que catalizan las glucosiltransferasas bacterianas Las GTFs hidrolizan a la sacarosa en sus unidades de glucosa y fructuosa componentes, los residuos de glucosa resultantes son polimerizados originando a los glucanos.8 La enzima continúa transfiriendo residuos de glucosa al polímero, a partir de la sacarosa e introduce ramificaciones. La reacción general puede representarse como: figura 5 La actividad de algunas GTF depende o es incrementada por la presencia de moléculas de glucano pre-xistentes, aceptoras de los residuos de glucosa. De tal forma, estas 62 enzimas catalizan la transferencia de un residuo de glucosa desde la sacarosa hacia una cadena de glucano en crecimiento.15 En los organismos que como el E. mutans, producen múltiples enzimas GTFs, los dextranos alfa (1-6) sintetizados por las GTF-S, pueden funcionar como aceptores para las GTF-I. Así, una especie bacteriana puede producir gran variedad de productos glucanos, y la acción cooperativa de las 3 enzimas es esencial para la adhesión del microorganismo dependiente de sacarosa.15,17 Los parámetros cinéticos identificados para la GTF-I de E. mutans son: 8 Km = 5,7 ± 1,4 mM Vmax = 12, 4 ± 1, 3 ìM/min/mg Genes que codifican a las glucosiltransferasas y regulación de su expresión Se han identificado los genes que codifican a las GTFs en el E. mutans.17 Estos son: 1. gtfB: tiene como producto a la GTFB (GTF-I). 2. gtfC: tiene como producto a la GTFC (GTF-SI). 3. gtfD: tiene como producto a la GTFD (GTF-S). El análisis de su secuencia reveló que estos genes poseen regiones altamente conservadas como son: 17 a) Una secuencia señal 5', b) Un dominio catalítico 5', y c) Un dominio 3' de unión al glucano. 63 La actividad de las enzimas GTFs puede ser modulada por condiciones ambientales tales como el pH, la concentración de iones, y el potencial de oxidación-reducción. Por otra parte, se creía inicialmente que la expresión de los genes gtf era constitutiva, pero se ha identificado que varias señales ambientales como pH, disponibilidad de glúcidos, y fase de crecimiento, tienen profundos efectos en la expresión de estos genes. Se ha demostrado que cuando se somete al E. mutans a diferentes condiciones, los niveles de expresión relativos son: gtfB > gtfD > gtfC. La sacarosa incrementa la expresión de gtfD, pero reduce la de gtfB y gtfC. El análisis cuantitativo demostró además, coincidencia entre la relación de expresión de cada gen gtf y la relación óptima para la adhesión del E. mutans dependiente de sacarosa in vitro.17 Como los genes gtfB y gtfC están muy próximos en el genoma, se creyó originalmente que su expresión estaba vinculada. Sin embargo, ahora se conoce que no es así. Los sistemas de transducción de señales de 2 componentes desempeñan un papel importante en la expresión genética bacteriana en respuesta a una variedad de estímulos.2 Forman parte de la red regulatoria esencial para la adaptación, supervivencia y virulencia bacteriana. Funcionan como "interruptores moleculares" en la modulación de la expresión genética en respuesta a los cambios en el medio externo. Estos sistemas consisten en un sensor quinasa y un efector, o regulador de la respuesta (RR), quien generalmente es una proteína de unión al ADN que modula la expresión de determinados genes. 24,25 Un ejemplo de ello es el sistema de transducción de señales de 2 componentes CovR/S, un regulador global de la expresión de genes de virulencia en los estreptococos de los grupos A y B. El RR CovR regula alrededor del 15 % de los genes en los estreptococos 64 del grupo A, muchos de los cuales están involucrados en la producción de enfermedades. El análisis transcripcional ha revelado que CovR reprime al menos 2 importantes genes: gtfD y gtfC, en el E. mutans, es decir, ambos genes son regulados por CovR al nivel transcripcional. 28,29 La proteína CovR se une directamente con grandes regiones de los promotores de gtfB y gtfC, cerca de los sitios de inicio de la transcripción, y de esta manera, CovR regula negativamente la expresión de dichos genes. Otros sistemas de transducción de señales de 2 componentes pueden estar también involucrados en la regulación de la expresión de estos genes.28En realidad, se considera que hay múltiples mecanismos reguladores involucrados en la expresión de los genes que codifican a las enzimas productoras de polisacáridos extracelulares bacterianos. Función que desempeñan los glucanos extracelulares bacterianos Los glucanos tienen 2 funciones fundamentales: mediar la adherencia bacteriana y servir como fuente nutricional. 1. Adherencia. Los variados grados de ramificación del mutano y el predominio de enlaces alfa (1-3), ocasionan que este polisacárido presente alto grado de adhesividad.15 De esta forma, los glucanos adhesivos como el mutano, median la unión de las bacterias a la superficie del diente, así como a otras bacterias. Por lo tanto, promueven la adherencia y coadherencia, así como la permanencia y maduración de la placa dental. Constituyen así 65 elementos críticos en el incremento de las proporciones del E. mutans en la placa y de su cariogenicidad, es decir, su capacidad de producir caries dental. 2. Fuente nutricional. Los polisacáridos extracelulares pueden también ser utilizados por las bacterias como fuente de nutrientes, gracias a la síntesis de enzimas glucanohidrolasas como la dextranasa [alfa-(1-6) glucanasa; EC 3.2.1.11], y la mutanasa [alfa-(1-3) glucanasa; EC 3.2.1.59], por parte de las propias bacterias. Se conoce que la dextranasa puede ubicarse en la superficie de la pared o ser liberada al espacio extracelular. En ambos casos, su acción permite la utilización de los glucanos como fuente nutricional. La dextranasa unida con la pared, se considera que también participa en el control de las propiedades adhesivas del glucano extracelular. En su anclaje a la pared celular bacteriana interviene una proteína denominada sortasa.33 Ambas glucanohidrolasas, presentes en el biofilm de la placa dental, pueden influir sobre la síntesis y estructura de los glucanos formados por las glucosiltransferasas a partir de la sacarosa. Las glucanohidrolasas, incluso en presencia de las GTFs, influyen en la síntesis de glucanos, el remodelado de sus enlaces polimerizantes y sus ramificaciones, lo que puede tener un impacto en la formación, maduración, propiedades físicas y sitios de unión bacteriana de los polisacáridos de la matriz de la placa dental. 14 66 Los medios de cultivo en microbiología Figura 6 Uno de los sistemas más importantes para la identificación de microorganismos es observar su crecimiento en sustancias alimenticias artificiales preparadas en el laboratorio. El material alimenticio en el que crecen los microorganismos es el Medio de Cultivo y el crecimiento de los microorganismos es el Cultivo. Se han preparado más de 10.000 medios de cultivo diferentes. Para que las bacterias crezcan adecuadamente en un medio de cultivo artificial debe reunir una serie de condiciones como son: temperatura, grado de humedad y presión de oxígeno adecuadas, así como un grado correcto de acidez o alcalinidad. Un medio de cultivo debe contener los nutrientes y factores de crecimiento necesarios y debe estar exento de todo microorganismo contaminante. La mayoría de las bacterias patógenas requieren nutrientes complejos similares en composición a los líquidos orgánicos del cuerpo humano. Por eso, la base de muchos medios de cultivo es una infusión de extractos de carne y Peptona a la que se añadirán otros ingredientes. El agar es un elemento solidificante muy empleado para la preparación de medios de cultivo. Se licúa completamente a la temperatura del agua hirviendo y se solidifica al enfriarse a 40 grados. Con mínimas excepciones no tiene efecto sobre el crecimiento de las bacterias y no es atacado por aquellas que crecen en él. Figura 7 La Gelatina es otro agente solidificante pero se emplea mucho menos ya que bastantes bacterias provocan su licuación. 67 En los diferentes medios de cultivo se encuentran numerosos materiales de enriquecimiento como hidratos de carbono, suero, sangre completa, bilis, etc. Los hidratos de Carbono se adicionan por dos motivos fundamentales: para incrementar el valor nutritivo del medio y para detectar reacciones de fermentación de los microorganismos que ayuden a identificarlos. El suero y la sangre completa se añaden para promover el crecimiento de los microorganismos menos resistentes. También se añaden colorantes que actúan como indicadores para detectar, por ejemplo, la formación de ácido o como inhibidores del crecimiento de unas bacterias y no de otras (el Rojo Fenol se usa como indicador ya que es rojo en pH básico y amarillo en pH ácido. La Violeta de Genciana se usa como inhibidor ya que impide el crecimiento de la mayoría de las bacterias Gram-positivas). Condiciones generales para el cultivo de microorganismos El desarrollo adecuado de los microorganismos en un medio de cultivo se ve afectado por una serie de factores de gran importancia y que, en algunos casos, son ajenos por completo al propio medio. 1- disponibilidad de nutrientes adecuados Un medio de cultivo adecuado para la investigación microbiológica ha de contener, como mínimo, carbono, nitrógeno, azufre, fósforo y sales inorgánicas. En muchos casos serán necesarias ciertas vitaminas y otras sustancias inductoras del crecimiento. Siempre han de estar presentes las sustancias adecuadas para ejercer de donantes o captadores de electrones para las reacciones químicas que tengan lugar. Todas estas sustancias se suministraban originalmente en forma de infusiones de carne, extractos de carne o extractos de levadura. Sin embargo, la preparación de estas sustancias para su aplicación a los medios de cultivo provocaban la pérdida de los factores nutritivos lábiles. Actualmente, la forma más extendida de aportar estas sustancias a los medios es utilizar peptona que, además, representa una fuente fácilmente asequible de nitrógeno y carbón 68 ya que la mayoría de los microorganismos, que no suelen utilizar directamente las proteínas naturales, tienen capacidad de atacar los aminoácidos y otros compuestos más simples de nitrógeno presentes en la peptona. Ciertas bacterias tienen necesidades nutritivas específicas por lo que se añade a muchos medios sustancias como suero, sangre, líquido ascítico, etc. Igualmente pueden ser necesarios ciertos carbohidratos y sales minerales como las de calcio, magnesio, manganeso, sodio o potasio y sustancias promotoras del crecimiento, generalmente de naturaleza vitamínica. Muy a menudo se añaden al medio de cultivo ciertos colorantes, bien como indicadores de ciertas actividades metabólicas o bien por sus capacidades de ejercer de inhibidores selectivos de ciertos microorganismos. 2- consistencia adecuada del medio Partiendo de un medio líquido podemos modificar su consistencia añadiendo productos como albúmina, gelatina o agar, con lo que obtendríamos medios en estado semisólido o sólido. Los medios solidificados con gelatina tienen el gran inconveniente de que muchos microorganismos no se desarrollan adecuadamente a temperaturas inferiores al punto de fusión de este solidificante y de que otros tienen la capacidad de licuarla. Actualmente los medios sólidos son de uso universal, por su versatilidad y comodidad, pero hay también gran cantidad de medios líquidos cuyo uso está ampliamente extendido en el laboratorio. 3- presencia (o ausencia) de oxígeno y otros gases Gran cantidad de bacterias pueden crecer en una atmósfera con tensión de oxígeno normal. Algunas pueden obtener el oxígeno directamente de variados sustratos. Pero los microorganismos anaerobios estrictos sólo se desarrollarán adecuadamente en una atmósfera sin oxígeno ambiental. En un punto intermedio, los microorganismos microaerófilos crecen mejor en condiciones atmosféricas parcialmente anaerobias (tensión de oxígeno muy reducida), mientras los anaerobios facultativos tienen un metabolismo capaz de adaptarse a cualquiera de las citadas condiciones. 69 4- condiciones adecuadas de humedad Un nivel mínimo de humedad, tanto en el medio como en la atmósfera, es imprescindible para un buen desarrollo de las células vegetativas microbianas en los cultivos. Hay que prever el mantenimiento de estas condiciones mínimas en las estufas de cultivo a 35-37ºC proporcionando una fuente adecuada de agua que mantenga la humedad necesaria para el crecimiento de los cultivos y evitar así que se deseque el medio. 5- Luz ambiental La mayoría de los microorganismos crecen mucho mejor en la oscuridad que en presencia de luz solar. Hay excepciones evidentes como sería el caso de los microorganismos fotosintéticos. 6- pH La concentración de iones hidrógeno es muy importante para el crecimiento de los microorganismos. La mayoría de ellos se desarrollan mejor en medios con un pH neutro, aunque los hay que requieren medios más o menos ácidos. No se debe olvidar que la presencia de ácidos o bases en cantidades que no impiden el crecimiento bacteriano pueden sin embargo inhibirlo o incluso alterar sus procesos metabólicos normales. 7- Temperatura Los microorganismos mesófilos crecen de forma óptima a temperaturas entre 15 y 43ºC. Otros como los psicrófilos crecen a 0ºC y los termófilos a 80ºC o incluso a temperaturas superiores (hipertemófilos). En líneas generales, los patógenos humanos crecen en rangos de temperatura mucho más cortos, alrededor de 37ºC, y los saprofítos tienen rangos más amplios. 8- Esterilidad del medio Todos los medios de cultivo han de estar perfectamente estériles para evitar la aparición de formas de vida que puedan alterar, enmascarar o incluso impedir el crecimiento microbiano normal del o de los especímenes inoculados en dichos medios. El sistema 70 clásico para esterilizar los medios de cultivo es el autoclave (que utiliza vapor de agua a presión como agente esterilizante) La evolución de los medios de cultivo Podemos decir que la microbiología empieza su verdadero desarrollo como ciencia en el momento en que se descubre el microscopio y comienza la observación de los primeros microorganismos, pero es indudable que la puesta a punto de los medios de cultivo y la utilización del agar como solidificante, marcan dos importantes puntos de inflexión en su evolución. La primera noticia de la utilización de medios de cultivo nos llega del micólogo Brefeld, que consiguió aislar y cultivar esporas de hongos en medios sólidos realizados a base de gelatina. Sin embargo este sistema no era adecuado para las bacterias (por su menor tamaño) y no fue hasta el año 1878 cuando Lister popularizó un método enfocado al cultivo puro basado en diluciones seriadas en un medio líquido. Koch realizó sus investigaciones utilizando en un primer momento rodajas de patata como soporte nutritivo sólido, pero no tardó en recurrir al caldo de carne líquido, diseñado por Loeffler, al que, en 1881, añadió gelatina, logrando un medio sólido transparente ideal para la observación de la morfología macroscópica de las colonias microbianas. En el año 1882 tiene lugar uno de los grandes avances de la microbiología en relación con los medios de cultivo: el médico alemán Walter Hesse introduce el agar-agar (polisacárido extraído de algas rojas) como solidificante. En 1887 un ayudante de Koch llamado Petri, comienza a utilizar placas de cristal planas, que se llaman desde entonces placas de Petri, para sustituir a las clásicas bandejas de vidrio cubiertas con campanas que se usaban hasta entonces. Beijerinck y Winogradsky, que desde de 1888 realizaron sus investigaciones sobre las bacterias quimioautótrofas (utilización de nitrógeno y azufre sobre todo) tuvieron gran importancia en el desarrollo de los medios selectivos y de enriquecimiento. Diseñaron este tipo de medios de tal forma que su especial composición química favorecía el crecimiento de ciertos tipos de microorganismos que, en función de sus procesos 71 metabólicos, eran los únicos capaces de utilizar para su desarrollo ciertos nutrientes del medio. En 1892 Würtz impulsó el uso de los medios diferenciales, incorporando indicadores de pH a la composición de ciertos medios con lo cual se podía observar la producción de ácidos en la fermentación en ciertos microorganismos. 15 Clasificación de los medios de cultivo Los medios de cultivo pueden clasificarse según diferentes criterios, pero los más importantes son aquellos que se basan en: a) su consistencia b) su utilización c) su composición d) su origen Según su estado físico (consistencia). 1) medios líquidos: Son los que se presentan en este estado, denominándose por esta razón caldos. El medio líquido más utilizado es el llamado caldo nutritivo, compuesto principalmente de extracto de carne, peptona y agua. Se utiliza fundamentalmente cuando se pretende la obtención de una suspensión bacteriana de una determinada concentración. 2) medios sólidos: Se preparan a partir de los medios líquidos, agregándoles un agente gelificante. Los más utilizados son la gelatina y el agar. Gelatina: Es una proteína animal obtenida de los huesos. Tiene el inconveniente de que es hidrolizada por muchas bacterias, y además su uso está muy limitado porque su punto de fusión es bajo (licúa a temperatura ambiente) razón por la que no puede utilizarse para cultivos a 37ºC, que es la tempera óptima de crecimiento para muchos microorganismos. Agar-agar: Es un 72 polímero de azúcares obtenido de algas marinas. Se trata de una molécula insoluble enagua pero soluble en agua caliente; una solución al 1,5% p/v forma un gel firme entre 32 y 39ºC y no se funde por debajo de 85ºC. Funde a 90ºC y solidifica una vez fundido alrededor de los 45ºC. Tiene el inconveniente de que introduce compuestos orgánicos indefinidos que pueden falsear los resultados de las necesidades nutritivas de un microorganismo. Aunque el agar es una mezcla de polisacáridos, Araki, en 1937, los dividió en dos grupos:- agarosa, con poco sulfato, libre de ácido pirúvico, yagaropectina, con mucho sulfato y gran cantidad de cenizas. La proporción de agarosa a agaropectina en el agar varía según el alga de origen. El agar puede utilizarse para diferentes fines, aunque los más importantes son: agar comercial para la industria alimenticia, agar bacteriológico, agares purificados y agarosa, utilizada para electroforesis en gel. Aproximadamente la mitad de todo el agar que se produce, se utiliza en trabajo microbiológico, por lo que es importante la ausencia de metales tóxicos. 3) medios semisólidos: Se preparan a partir de los medios líquidos, agregando a éstos un agente solidificante en una proporción menor que para preparar medios sólidos. Uno de sus usos es la investigación de la movilidad de las bacterias Según su utilización. 1) Medios comunes: Son aquellos que poseen los componentes mínimos para que pueda producirse el crecimiento de bacterias que no necesiten requerimientos especiales. El medio más conocido de este grupo es el agar nutritivo o agar común, que resulta de la adición de agar al caldo nutritivo. Otros representantes de este grupo son el agar tripticase de soja, el agar Columbia, etc. 16 73 2) Medios de enriquecimiento: Son aquellos que, además de las sustancias nutritivas normales, incorporan una serie de factores indispensables para el crecimiento de microorganismos exigentes. Este enriquecimiento se hace por adición de sangre u otros productos biológicos (sangre, suero, leche, huevo, bilis, etc.) que aportan dichos factores. En ocasiones es posible añadir suplementos artificiales a los medios para producir un enriquecimiento del mismo (p. ej. Polivitex, Isovitalex, etc.) El gonococo, por ejemplo, necesita cistina y cisteína para su crecimiento. Estas sustancias son aportadas por la sangre calentada adicionada al medio de cultivo (agar chocolate). 3) medios selectivos: Son medios utilizados para favorecer el crecimiento de ciertas bacterias contenidas en una población polimicrobiana. El fundamento de estos medios consiste en facilitar nutricionalmente el crecimiento de una población microbiana específica. Un ejemplo de medio selectivo es el caldo selenito, que se utiliza para favorecer el crecimiento de salmonellas y frenar el del resto de enterobacterias. 4) medios inhibidores: Cuando las sustancias añadidas a un medio selectivo impiden totalmente el crecimiento de una población microbiana, se denomina inhibidor. Los medios inhibidores podrían considerarse como una variante más restrictiva de los medios selectivos. Los medios inhibidores se consiguen habitualmente por adición de sustancias antimicrobianas o de cualquier otra que inhiba completamente el desarrollo de una población determinada. Un medio inhibidor es el MacConkey que permite el crecimiento de los gérmenes Gram negativos e impide el crecimiento de los Gram positivos. 5) Medios diferenciales: Se utilizan para poner en evidencia características bioquímicas que ayuden a diferenciar géneros o especies. La adición de un azúcar fermentable o un sustrato metabolizable se utilizan para este fin. El medio MacConkey es un medio diferencial porque permite distinguir los gérmenes que fermentan la lactosa de aquellos que no lo hacen. También lo son el C.L.E.D. (lactosa +/lactosa -), el agar sangre (tipo de hemólisis), el SS (que es doblemente diferencial), etc. 74 6) Medios de identificación: Son los destinados a comprobar alguna cualidad específica que puede servirnos para reconocer la identidad de un microorganismo. Estos medios han de poseer los elementos necesarios para asegurar el crecimiento de los microorganismos, el sustrato específico que vaya a ser metabolizado y el indicador que nos muestre el resultado. El agar Kligler, el medio de Simmons y en general, cualquier medio al que se le haya añadido un elemento diferencial de un microorganismo, son medios utilizados en identificación. Actualmente están apareciendo en el mercado una gran cantidad de medios específicos de identificación para ciertos microorganismos, con lo cual se consigue simultáneamente abaratar el costo y reducir el tiempo de identificación. Son ejemplos de esto último los medios CPS ID3 o Uriline ID (Biomerieux), utilizados para identificar los gérmenes urinarios más importantes a partir de la placa de cultivo gracias a la utilización de sustratos cromogénicos específicos. 7) Medios de multiplicación: Sirven para obtener una gran cantidad de células a partir de un microorganismo ya aislado. Se emplean en la obtención de vacunas, en la investigación y en la industria. Los medios más adecuados para la multiplicación suelen ser líquidos. El caldo-infusión cerebro-corazón (BHI), es un ejemplo típico de estos medios. 8) Medios de conservación: Se utilizan para conservar una cepa que, por diversas razones nos interese mantener. Fundamentalmente se utilizan como controles de calidad de las pruebas y reactivos utilizados en el Laboratorio de Microbiología. En el laboratorio se pueden conservar las cepas de tres formas: a) haciendo pases periódicos de placa a placa, b) mediante liofilización de una suspensión bacteriana, y c) congelando las cepas en leche descremada estéril al 0,1%. 75 9) medios de transporte: Se usan para el transporte de muestras clínicas que no pueden sembrarse inmediatamente. Su utilización debe hacerse introduciendo la torunda con la que se obtuvo la muestra en el interior del medio (generalmente en un tubo). Son ejemplos típicos de este grupo los medios de Stuart-Amies, Cary-Blair, etc. Atendiendo a su composición. Según las sustancias que entren a formar parte en su composición, los medios de cultivo pueden ser clasificados en: 1) medios complejos: Fueron los primeros utilizados, y los más empleados se preparan a partir de tejidos animales, y más raramente de vegetales. Su composición no es exactamente definida, y por consiguiente no es rigurosamente constante. Esto puede tener ciertos inconvenientes en condiciones experimentales, donde la reproductibilidad no podrá ser exacta. En la práctica corriente estos medios dan excelentes resultados y son los más empleados. 2) medios sintéticos: Son aquellos que contienen en su composición exclusivamente sustancias químicas conocidas y disueltas en agua destilada en proporciones determinadas, resultando un medio de composición perfectamente definida. 3) medios semisintéticos: El gran número de factores de crecimiento exigidos para ciertos gérmenes hace que la fabricación de un medio sintético para estos gérmenes sea imposible o demasiado cara. En este caso se aportan los factores de crecimiento bajo la forma de un extracto orgánico complejo (extracto de levadura, extracto de tejidos, etc.).Ciertos gérmenes no crecen en ningún medio por muy enriquecido que esté éste, haciéndolo exclusivamente en células vivas con unas características determinadas. Ejemplos de este tipo son, aparte de los virus, las Chlamydias, Rickettsias, etc. 76 Según su origen: a) naturales: son los preparados a partir de sustancias naturales de origen animal o vegetal como ser extractos de tejidos o infusiones y cuya composición química no se conoce exactamente. b) sintéticos: son los medios que contienen una composición química definida cuali y cuantitativamente. Se utilizan para obtener resultados reproducibles. c) semisintéticos son los sintéticos a los que se les añaden factores de crecimiento bajo una forma de un extracto orgánico complejo, como por ejemplo extracto de levadura.16 Morfología colonial Generalmente se reporta la morfología colonial tal como uno la percibe, por ejemplo si se ves que las colonias bacterianas son como puntitos, pues reportan que son puntiformes, si son lisas pues reportan que son lisas, si observas que están como moco, pues reportan que están mucosas, si su color es amarillento pues reportan que son de color amarillento, y así sucesivamente., también puedes agregar características que consideres importante para identificarlas, por ejemplo el olor., Aun así checa las siguientes imágenes para orientarte.17 77 figura 8 Morfología colonial bacteriana superficie. figura 9 Morfología colonial bacteriana forma. figura 10 Morfología colonial bacteriana borde. 17 78 CAPITULO 3 METODOLOGIA Tipo de estudio: estudio piloto de tipo observacional, descriptivo, transversal no probabilístico. Análisis de la muestra: se escogieron 16 colonias aisladas de estreptococo mutans obtenidas previamente de un proyecto de investigación. Selección de las colonias La selección de las colonias se llevo a cabo por medio de su morfología característica, a continuación muestro una foto donde se describirán por medio de que características se seleccionaron las colonias de Estreptococo mutans. Figura 11 Aquí se puede observar las colonias de estreptococo mutans, sus características generales para su selección fue: 79 Características: crecen manifestando coloración azul intenso o negruzca, a veces lisas o prominentes, con mucha frecuencia firmemente adheridas al agar, algunas veces presenta una gota brillante de polisacárido extracelular en la parte superior o a un lado. Forma colonias circulares en su mayoría, características de borde que sean redondeado u ondulado y características de superficie ya sean convexas o acuminadas, en cuanto a su consistencia pueden ser duras o mucosas. Criterios de inclusión: Colonias que tuvieran las características morfológicas principales de estreptococo mutans. Colonias que estuvieran separadas para ver correctamente su morfología colonial. Criterios de exclusión: Sectores donde haya tanto crecimiento de colonias de estreptococo mutans que fuera imposible describir su morfología colonial. Colonias que no cumplieran con las características morfológicas de estreptococo mutans. Criterios de eliminación: Colonias contaminadas con algún otro tipo de crecimiento bacteriano. Infraestructura: Laboratorio de microbiología de la Universidad Veracruzana campus Poza Rica – Túxpan. Recursos humanos: Alumno investigador: Samuel Xochihua Ramírez Director de tesis: Heriberto del Ángel Calderón Asesor de tesis: Araceli García Rocha 80 Maestra de experiencia educativa: Alma Luz San Martín López Recursos financieros: Autofinanciable Material: Autoclaves Tripie Tubos de ensaye Esterilizador Medio infusión cerebro Cepas aisladas de Refrigerador corazón Estreptococo mutans Estufa bacteriológica Sacarosa Asas bacteriológicas Mechero bunsen Matraces Gasas Balanza Pipetas Vaso de precipitado 81 Figura 12 PROCEDIMIENTO Se preparo una solución del medio llamado Infusión Cerebro Corazón (un medio especial para el crecimiento de bacterias como Estreptococos, en este caso Estreptococo mutans), se leyeron las instrucciones de cómo prepararlo, las instrucciones son las siguientes para 37 gramos se disuelven en un litro de agua destilada, ya que solo se preparara 50 ml de agua destilada se hacen los cálculos necesarios para sacar los gramos necesarios: Los cálculos se realizaron por medio de Medio de cultivo infusión cerebro corazón Una regla de 3: 37gr-----1000ml X---------50ml = 1.85 gr del medio deshidratado Después de hecho el cálculo para obtener los gramos necesarios del medio, al medio se le agregara sacarosa (como agente enriquecedor), la sacarosa estará al 15%. 82 Entonces para 100 mililitros se necesitan 15 gramos de sacarosa, pero como solo se prepararon 50 ml solo se necesitaran la mitad de sacarosa: 7.5 gramos. Se pesa la sacarosa para después vertirla en el medio ya calentado. Figura 13 Sacarosa Figura 14 Agua destilada figura 15 Por lo tanto se necesitaron 1.85 gramos del medio, 50 ml de agua destilada y 7.5 gramos de sacarosa (al 15%), ya que se tuvieron los cálculos se procedió a pesar para 82 preparar el medio primeramente el matraz para después adicionarle el medio, el matraz peso 74.45 Matraz figura 16 El matraz peso 74. 45 y a este se le agrego 1.85 gramos del medio lo que nos dio un total de 76.3 que nos dio en el cálculo. Matraz y medio de cultivo figura 17 83 Para pesar la sacarosa se peso en un matraz más pequeño (de 40ml) este peso 27.2 gramos mas 7.5 de sacarosa da un total de 34.7, La sacarosa al 15% se agregara al medio después de que este se halla calentado. Sacarosa y balanza Figura 18 Se agrega a 50ml de agua destilada los 1.85 gr del medio de cultivo. Figura 19 Agua destilada y medio de cultivo 84 Se calienta el medio para disolver los grumos hasta que la mezcla quede totalmente homogénea. Medio y mechero figura 20 Mezcla totalmente homogénea figura 21 85 Se le agrega los 7.5 gr de sacarosa al 15% y queda de esta manera. Hay un ligero cambio de color. Medio con sacarosa figura 22 Después de preparado el medio se vierte en tubos, son 3 ml por tubo, y son 50 ml de muestra en total, se prepararon 16 tubos con una concentración de sacarosa al 15% y sobraron 2 ml. 86 Figura 23 Tubos con medio BHI figura 24 Se pusieron los tubos en un vaso de precipitado para esterilizarlos en la autoclave. 87 Medio con sacarosa al 15% 16 tubos figura FIGURA 25 Se esteriliza durante 15 min. A 121 grados centígrados. 88 Autoclave figura 26 Estufa bacteriológica figura 27 Después de esterilizar los 16 tubos se introducen en la estufa bacteriológica 24 hrs. Para que pasen prueba de esterilidad, los tubos que a las 24 hrs. Presenten turbidez se desechan. 89 Se muestra las cajas Petri que contienen a las colonias previamente seleccionadas, que se sembraran en los tubos. Colonias de estreptococo aisladas figura 29 Aquí se muestra el área de trabajo para el inoculado en los tubos Mechero y tubos figura 30 90 A1 B1 Colonias aisladas A1 y B1 Figura 31 Colonias aisladas B1 Figura 32 91 Colonias aisladas C1 y D1 D1 C1 Figura 33 Con un asa bacteriológica se calienta al fuego para esterilizarla y después se toma con ella una colonia escogida. Figura 34 asa y mechero 92 Después de calentar al machero el asa se toca la colonia seleccionada con la punta del asa. Figura 35 colonia escogida Se lleva al tubo de ensaye, para depositarla en el medio bajo condiciones de seguridad, ya que la punta del asa toca el medio se le da vuelta para que las bacterias queden en el medio y se retira el asa. FIGURA 36 asa y tubo. 93 Figura 37 asa y tubo 2 Figura. 38 Colonia en tubo El asa se vuelve a pasar por el fuego para esterilizarla. Esto se hace sucesivamente con las 16 colonias escogidas depositándolos en los 16 tubos. 94 Tubo y colonia Figura 39 Tubos sembrados Figura.- 40 tubos sembrados Ya que están sembrados la totalidad de las colonias en los tubos se introducen en la estufa bacteriológica para encubarlos y que se desarrollen los polisacáridos, se incuban a 37 grados centígrados bajo condiciones de anaerobiosis, durante 24 hrs. 95 Figura.- 41 Condiciones de anaerobiosis 96 CAPITULO VI RESULTADOS Los resultados fueron exitosos, se logro responder a las preguntas del presente trabajo de investigación. La hipótesis que se corroboro fue la de trabajo ya que se la producción de polisacáridos insolubles fue mayor que los solubles. En la primera tabla llamada frecuencia de producción de polisacáridos, se describen los resultados obtenidos, se escogieron 16 cepas las cuales se sembraron en 16 tubos de los cuales en 12 tubos se dio la producción de polisacáridos extracelulares insolubles, y en los 4 restantes se dio la producción de polisacáridos extracelulares insolubles. Tabla no. 1 “TABLA DE FRECUENCIA DE PRODUCCIÓN DE PRODUCCIÓN DE POLISACÁRIDOS” Numero de cepas 16 Numero de tubos 16 Polisacáridos Polisacáridos insolubles solubles 12 4 En esta tabla se muestra la distribución de colonias de estreptococo sembrado en los tubos, por ejemplo observamos que para la muestra A1 se escogieron 5 colonias características de Estreptococo mutans las cuales se sembraron en 5 tubos los que resultaron positivos a la producción de polisacárido extracelular insoluble. 97 Tabla no. 2.- distribución de colonias en los tubos para la producción de polisacáridos extracelulares Muestras colonias Tubos P. insoluble P. soluble. sembrados A1 5 5 Positivos B1 4 4 Positivos C1 3 3 Positivos D1 4 4 negativos En esta segunda grafica que se llama porcentaje de polisacáridos producidos se expone el porcentaje del total de las muestras estudiadas, 25 % corresponden a la producción de polisacáridos solubles y 75% corresponde a la producción de polisacáridos solubles. "Porcentaje de polisacaridos extracelulares producidos" 25% polisacaridos solubles 75% polisacaridos insolubles 98 Aquí se presenta fotos de la producción de polisacáridos insolubles: MUESTRAS A1 5 TUBOS Figura 42 Ahora se voltea el tubo para ver la característica de los polisacáridos producidos por la bacteria “estos le dan una adhesividad muy grande a la bacteria” para pegarse al diente como lo hace al fondo de este tubo: MUESTRA B1 (1 TUBO REPRESENTATIVO) 99 Figura 43 muestra B1 MUESTRA C1 (2 TUBOS REPRESENTATIVOS) Figura 44. 100 Muestra D1 (4 tubos representativos) polisacáridos solubles) Figura.-45 101 CAPITULO V CONCLUSIONES Conocer más acerca de los polisacáridos extracelulares ayuda a ampliar el conocimiento acerca de su papel en el desarrollo de caries, estos como se había dicho proporcionan extrema adhesividad al diente, le sirve de reservorio de nutrimentos y cohadesion para otras bacterias (en especial los polisacáridos insolubles), por lo tanto resulta de vital importancia conocer más acerca de las pruebas para la producción de polisacáridos extracelulares. Se logro dar respuesta a las preguntas de investigación así como a los objetivos de la investigación. Se pudo corroborar la hipótesis de trabajo donde se produjeron en más alta frecuencia polisacáridos extracelulares insolubles que solubles. De un total de 16 tubos con 16 cepas sembradas se obtuvieron 12 muestras con producción de polisacáridos extracelulares insolubles, y solo 4 con producción de polisacáridos extracelulares solubles. Expresado en porcentaje de un total del 100% el 75 % fue de producción de polisacárido extracelular insoluble mientras que el 25% para la producción de polisacárido extracelular soluble. 102 DISCUSIÓN AUTOR INVESTIGACIÓN MARIA JESUS Estudio de streptococcus De las 30 cepas sembradas al CABRONERO mutans en un 10% de sacarosa, el 82% FERNÁNDEZ Modelo experimental. corresponden a polisacárido Madrid, 1991. Aportaciones insoluble y el 18% a Etiopatogenicas. polisacárido soluble en agua. Xochihua Ramírez Samuel “Determinación de De las 16 cepas sembradas al Químico: Heriberto del polisacáridos 15% de sacarosa el 75% fueron Ángel Calderón extracelulares en cepas polisacáridos insolubles, el 25% Dra. Araceli García Rocha aisladas” fueron polisacáridos solubles en México 2011 RESULTADOS agua. 103 RECOMENDACIONES El cirujano dentista debe interesarse más acerca de estudiar de los factores etiológicos que hay detrás de la caries, en el sentido de estudiarlos no solo teóricamente sino llevar a cabo investigaciones, proyectos, pruebas para la detección de estos, como en este caso se pudo conocer el metabolismo, síntesis, y producción de polisacáridos extracelulares por parte del estreptococo mutans, ya que cuando se comprenda de una manera mejor el comportamiento de los polisacáridos y de las bacterias que los forman se podrán incorporar medidas preventivas para reducir la producción de estos tipos de polisacáridos que son dañinos a la pieza dental. Que el cirujano dentista conozca a fondo la importancia que juega el fluor en el desarrollo de la producción de polisacáridos extracelulares, ya que este inhibe el ciclo para la formación de este tipo de polisacáridos. Para que las bacterias como estreptococo mutans pueda producir polisacáridos extracelulares necesita nutrimentos en especial carbohidratos, en este país no se puede pedir que se deje de comer carbohidratos en especial por el tipo de dieta que consume el mexicano, sin embargo si se puede exhortar a que si la dieta que llevan es alta en azucares, también la limpieza dental sea con la frecuencia y la técnica correcta. 104 Bibliografía 1.- http://www.elsiglodetorreon.com.mx/noticia/244449.su-salud-bucal-la-caries-en-elsiglo-xxi.html 2.- Teresa Audesirk, Gerald Audesirk y Bruce E. Byers, Biología la vida en la Tierra, editorial Pearson Educación, México 2003 paginas 980. 3.- http://es.wikipedia.org/wiki/Gl%C3%BAcido 4.- entros5.pntic.mec.es/ies.victoria.kent/Rincon-C/Curiosid/Rc-58.htm 5.- http://www.ferato.com/wiki/index.php/Gluc%C3%B3geno 6.- http://es.wikipedia.org/wiki/Polisac%C3%A1rido 7.- Higashida Bertha., Odontología Preventiva, McGraw- Hill Interamericana, México DF 2001, 304 pp. 8.- es.wikipedia.org/wiki/Saliva 9.- http://www.soledadguerrero.com.ar/placadental 10.- Katz Simon, McDonald James y Stookey George, Odontología preventiva en Acción, editorial medica Panamericana, tercera edición, 2002 México DF, 375 pp. 11.- www.who.int/mediacentre/releases/2004/pr15/es/ 12.- http://www.monografias.com/trabajos40/caries-dentales/caries-dentales2.shtml 13.- http://members.fortunecity.es/jacflgb/Opera1.htm 14.- http://bvs.sld.cu/revistas/est/vol45_3-4_08/est103_408.htm 15.- http://www.microinmuno.qb.fcen.uba.ar/SeminarioMedios.htm 16.- http://perso.wanadoo.es/sergioram1/medios_de_cultivo.htm 17.- http://microbitos.wordpress.com/2010/06/14/morfologia-colonial-bacteriana/ 105