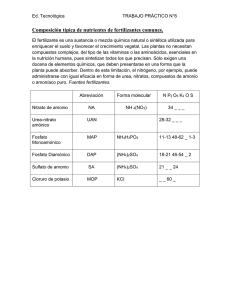
UNIVERSIDAD VERACRUZANA
Anuncio

UNIVERSIDAD VERACRUZANA FACULTAD DE CIENCIAS QUÍMICAS ZONA XALAPA PROGRAMA EDUCATIVO: INGENIERÍA AMBIENTAL “EVALUACIÓN DE LA CONDUCTA FISIOLÓGICA DE CONSORCIOS MICROBIANOS ANAEROBIOS EN LA DEGRADACIÓN DE COMPUESTOS NITROGENADOS DE AGUAS RESIDUALES.” TESIS Que para acreditar la experiencia educativa: experiencia recepcional Presenta: Arely Gómez Aquino Director Dr. Sergio Martínez Hernández Xalapa de Enríquez, Veracruz, Junio de 2014 I El presente trabajo se llevó a cabo en el Instituto de Biotecnología y Ecología Aplicada (INBIOTECA) de la Universidad Veracruzana - Campus Xalapa y contó con el apoyo financiero del proyecto PROMEP/103.5/12/4801- Evaluación de los aspectos fisiológicos y de la estructura poblacional durante la coexistencia de consorcios microbianos desnitrificantes y ANAMMOX en la eliminación simultánea de compuestos orgánicos y nitrogenados. II A mis padres por el esfuerzo realizado para poder alcanzar esta meta A Evelyn, Iris e Irving, por su cariño y comprensión III AGRADECIMIENTOS A Dios por llenarme de bendiciones durante el camino y permitirme cumplir este sueño. A mi asesor, el Dr. Sergio Martínez Hernández, por depositar su confianza en mí, por darme la oportunidad de participar en este proyecto tan bonito y escuchar mis opiniones y propuestas. Por apoyarme en la realización de los experimentos y el trabajo escrito. A mi tutor académico, el Dr. Epifanio Morales Zárate, por guiarme en el transcurso de la carrera, por escucharme, apoyarme y aconsejarme durante momentos difíciles y de ocio. Al Dr. Mario Artega Vázquez por brindarme su apoyo para realizar experimentos de biología molecular en su laboratorio y por sus acertadas asesorías. A PROMEP, gracias por la beca otorgada para la realización del proyecto y mi trabajo de tesis. A la Dra. María Teresa Leal Ascencio, por los comentarios y sugerencias que me permitieron enriquecer este trabajo. Gracias por su paciencia y por tomarse el tiempo de atender a mis dudas. A la Dra. Lorena De Medina Salas, por tener siempre la disposición para atenderme, por su preocupación por mejorar mi trabajo y sus recomendaciones. A mis padres por darme la oportunidad de desarrollarme profesionalmente. Por estar siempre al pendiente de mi, escuchar y motivarme a crecer y aprender cosas nuevas a lo largo de mi vida. Gracias a mi papá por los valores adquiridos y por enseñarme a dar siempre mi máximo esfuerzo, y gracias a mi madre por luchar siempre por mí y estar atenta a cada uno de mis pasos, por darme un hogar a donde siempre es grato volver. A mis hermanas Iris y Evelyn, por el apoyo y la confianza brindados durante tantos años, mismos que posibilitaron la conquista de esta meta. A pesar de sus IV múltiples distracciones gracias a ellas siempre tuve un lugar donde desestresarme. A mis compañeros del instituto. Alberto que compartió conmigo parte de este proyecto volviéndose un gran amigo, gracias por el tiempo brindado. David y Mariam, quienes me ayudaron a realizar las determinaciones, gracias por todas esas horas que pasamos juntos, volvieron el trabajo más ameno y sin su ayuda no habría podido terminar mis experimentos. A Karina y Marco, por darme un poco de su tiempo para poder resolver mis dudas, enseñarme tantos tips y ayudarme con todos los experimentos de biología molecular. A Magdiel y Luis por los consejos y por ayudarme a realizar mi gel de DGGE. A mis amigos por concederme el honor de su amistad durante estos años, donde juntos compartimos tantos sueños y aventuras, Andy, Angy, Dulce, Mayra, Yaline, Laura, Irving, Alejandro, Jorge, Marlon, Paco, Ricardo, gracias por su cariño y atenciones, los llevo en el corazón. “Un experto es alguien que conoce algunos de los peores errores que se pueden cometer en su especialidad, y cómo evitarlos.” ―Werner Karl Heisenberg “Siempre hay que saber cuándo una etapa llega a su fin. Cerrando ciclos, cerrando puertas, terminando capítulos; no importa el nombre que le demos, lo que importa es dejar en el pasado los momentos de la vida que ya se han acabado” ―Paulo Coelho V Contenido Introducción ............................................................................................................. 1 Capítulo I. Generalidades ........................................................................................ 3 1.1 Antecedentes ................................................................................................. 3 1.2 Planteamiento del problema ....................................................................... 5 1.3 Justificación ................................................................................................... 6 1.4 Objetivos ........................................................................................................ 7 1.4.1 Objetivo general .......................................................................................... 7 1.4.2 Objetivos específicos .................................................................................. 7 1.5 Hipótesis ........................................................................................................ 7 Capítulo II: Marco teórico ...................................................................................... 11 2.1 Normatividad en México acerca del nitrógeno en aguas residuales ............ 11 2.2 Compuestos nitrogenados en aguas residuales .......................................... 12 2.3 Operaciones y procesos para el control del nitrógeno ................................. 14 2.3.1 Métodos fisicoquímicos ......................................................................... 14 2.3.1.1 Separación del amoniaco por arrastre con aire (air stripping) ........ 14 2.3.1.2 Cloración al breakpoint ................................................................... 16 2.3.1.3 Intercambio iónico ........................................................................... 18 2.3.1.4 Procesos de precipitación ............................................................... 19 2.3.2 Métodos biológicos................................................................................ 20 2.3.2.1 Nitrificación ..................................................................................... 20 2.3.2.2 Desnitrificación ............................................................................... 25 2.3.3.3 Oxidación anaeróbica de amonio (Anamox) ................................... 30 Capítulo III: Materiales y métodos ......................................................................... 34 3.1 Ensayos anammox ...................................................................................... 34 3.1.1 Inóculo ................................................................................................... 34 VI 3.1.2 Matraces anammox ............................................................................... 34 3.1.3 Medio de cultivo .................................................................................... 35 3.1.4 Ensayos control ..................................................................................... 36 3.1.5 Ensayos en lote ..................................................................................... 36 3.2 Ensayos desnitrificantes .............................................................................. 37 3.2.1 Botellas serológicas .............................................................................. 37 3.2.2 Inóculo ................................................................................................... 38 3.2.3 Medio de cultivo .................................................................................... 38 3.2.4 Ensayos control ..................................................................................... 39 3.2.5 Ensayos en lote ..................................................................................... 39 3.3 Métodos analíticos ....................................................................................... 40 Tratamiento previo ......................................................................................... 40 3.3.1 Determinación de amonio ..................................................................... 40 3.3.2 Determinación de nitrito ........................................................................ 41 3.3.3 Determinación de nitrato ....................................................................... 41 3.3.4 Determinación de nitrógeno molecular .................................................. 42 3.3.5 Determinación de sólidos suspendidos volátiles. .................................. 42 3.4 Análisis molecular ........................................................................................ 43 3.4.1 Extracción de ácidos nucléicos ............................................................. 43 3.4.2 Amplificación de fragmentos de interés por PCR .................................. 43 3.4.3 Método de electroforesis en gel con gradiente desnaturalizante (DGGE) ....................................................................................................................... 44 Capítulo IV: Resultados y discusión ...................................................................... 45 4.1 Ensayos tipo anammox ................................................................................ 45 4.1.2 Obtención del inóculo estabilizado que consume nitrito y amonio para las cinéticas en lote ........................................................................................ 45 VII 4.1.2.1 Consumo de amonio ....................................................................... 45 4.1.2.2 Consumo de nitrito .......................................................................... 46 4.1.2.3 Velocidades específicas de consumo ............................................. 47 4.1.2 Ensayo sin nitrato .................................................................................. 48 4.1.3 Ensayo con nitrato................................................................................. 50 4.1.4 Comparación de consorcios anaerobios tipo anammox con y sin nitrato ....................................................................................................................... 53 4.2 Ensayos desnitrificantes .............................................................................. 56 4.2.1 Ensayos control ..................................................................................... 56 4.2.2 Ensayos desnitrificantes sin amonio ..................................................... 57 4.2.4 Comparación de ensayos desnitrificantes con y sin amonio ................. 62 4.2.4.1 Perfil de consumo de amonio en ensayos desnitrificantes con y sin amonio ........................................................................................................ 62 4.2.4.2 Perfil de producción de nitrógeno gaseoso en los ensayos desnitrificantes ............................................................................................ 63 4.2.5 Análisis molecular ................................................................................. 65 4.2.5.1 Extracción de DNA ......................................................................... 66 4.2.5.2 Amplificación de fragmentos de DNA por PCR............................... 69 4.2.5.3 Electroforesis en gel con gradiente desnaturalizante (DGGE)........ 71 Conclusiones ......................................................................................................... 74 Bibliografía ............................................................................................................ 76 VIII Índice de figuras Figura 1. Curva generalizada obtenida durante la cloración al breakpoint. Fuente: Metcalf y Eddy, 1979. ............................................................................ 17 Figura 2. Postulado del acoplamiento del proceso anammox a la membrana de la anammoxosoma. HH: hidracina-hidrolasa, HZO: enzima hidracinaoxidasa, NIR: enzima nitrito-reductasa Fuente: (van Niftrik y col., 2004)....... 33 Figura 3. Perfil de degradación de amonio por un consorcio estabilizado tipo anammox (Cada pico superior representa una nueva alimentación). ............ 45 Figura 4. Perfil de degradación de nitrito por un consorcio estabilizado para el proceso tipo anammox. (Cada pico superior representa una nueva alimentación). ...................................................................................................... 46 Figura 5. Velocidades específicas de consumo de amonio por un consorcio tipo anammox. ..................................................................................................... 47 Figura 6. Degradación de amonio y nitrito por un consorcio microbiano anaerobio tipo anammox. ................................................................................... 48 Figura 7. Conducta metabólica de un consorcio microbiano anaerobio, tipo anammox, en presencia de amonio, nitrito y nitrato. ....................................... 51 Figura 8. Conducta metabólica de un consorcio microbiano anaerobio, tipo anammox, durante la degradación de amonio y nitrito en presencia y ausencia de nitrato. ............................................................................................. 54 Figura 9. Conducta metabólica de un consorcio desnitrificante en presencia únicamente de nitrato. ........................................................................................ 57 Figura 10. Degradación anaerobia de nitrato a nitrógeno gaseoso por un consorcio desnitrificante. ................................................................................... 58 Figura 11. Degradación anaerobia de nitrato a nitrógeno gaseoso por un consorcio microbiano desnitrificante en presencia de nitrato y amonio. ...... 60 Figura 12. Degradación anaerobia de nitrato por un consorcio desnitrificante en presencia y ausencia de amonio. ................................................................. 62 Figura 13. Conducta metabólica de un consorcio desnitrificante durante la formación de nitrógeno molecular en presencia y ausencia de amonio. ....... 64 Figura 14. Visualización de la extracción de DNA de consorcios microbianos. .............................................................................................................................. 67 Figura 15. Amplificación de fragmentos de DNA por PCR .............................. 71 IX Figura 16. Comunidades microbianas de los diferentes ensayos separadas por DGGE. ............................................................................................................ 72 X Índice de tablas Tabla 1. Límites máximos permisibles de nitrógeno total para descargas a aguas y bienes nacionales. Fuente: DOF (1996) .............................................. 11 Tabla 2. Composición del lisier del ganado doméstico. Fuente: SeoánezCalvo, 2004........................................................................................................... 13 Tabla 3. Géneros representativos de bacterias nitrificantes. Fuente: Parés y col., 2002. ............................................................................................................. 21 Tabla 4. Factores de enriquecimiento para diferentes bacterias reductoras de 𝐍𝐎𝟑 − en un medio mínimo en condiciones anaerobias. Fuente: Parés y col., 2002. ..................................................................................................................... 27 Tabla 5. Grupos microbianos y su preferencia en donadores y aceptores de electrones, y fuentes de carbono. Fuente: Khanal, 2008. ................................ 35 Tabla 6. Composición química del medio mineral tipo anammox. ................. 35 Tabla 7. Composición química del medio de cultivo para bacterias desnitrificantes. ................................................................................................... 38 Tabla 8. Balance general de materiales y eficiencias de consumo de un consorcio microbiano anaerobio, tipo anammox, durante la degradación de amonio y nitrito. .................................................................................................. 49 Tabla 9. Balance general de materiales y eficiencias de consumo de un consorcio microbiano anaerobio, tipo anammox, en presencia de amonio, nitrito y nitrato. .................................................................................................... 52 Tabla 10. Velocidades específicas de consumo de un consorcio microbiano anaerobio, tipo anammox, durante la degradación de amonio y nitrito en presencia y ausencia de nitrato. ........................................................................ 55 Tabla 11. Balance general de materiales respecto al tiempo, eficiencia de consumo de nitrato y rendimiento de producción. .......................................... 59 Tabla 12. Balance de materiales respecto al tiempo, eficiencias de consumo y rendimiento de producción, en la degradación anaerobia de nitrato a nitrógeno molecular. ........................................................................................... 61 Tabla 13. Velocidad específica de consumo de un consorcio desnitrificante durante la degradación de nitrato en presencia y ausencia de amonio. ........ 63 Tabla 14. Velocidades específicas de producción de un consorcio desnitrificante durante la degradación o eliminación de nitrato en presencia y ausencia de amonio. ........................................................................................ 65 XI Tabla 15. Numeración y contenido de las muestras de consorcios anaerobios, usadas para biología molecular. ................................................... 66 Tabla 16. Cuantificación de DNA extraído y calidad obtenida. ....................... 68 XII Introducción El nitrógeno es un componente crucial de aminoácidos, proteínas, algunas vitaminas, nucleótidos (como el ATP) y ácidos nucléicos (Audesirk, 2011). Por ello se considera un nutriente esencial para el crecimiento de los seres vivos. Por otro lado, la atmósfera contiene alrededor del 78 % de nitrógeno gaseoso (Audesirk, 2011), éste es su principal suministro de nutrientes. Las distintas etapas de transformación del nitrógeno entre la atmósfera, suelo, plantas y animales dan lugar a un ciclo (Vega-de Kuyper, 2007). El ciclo biogeoquímico del nitrógeno incluye los procesos de fijación de nitrógeno (N2), nitrificación, desnitrificación, anammox, asimilación y amonificación. Este movimiento del nitrógeno a través de la biósfera determina en gran medida la productividad ecológica de los ambientes terrestres (Castillo-Rodríguez y col., 2005). Durante la fijación de N2 las bacterias descomponen los enlaces para formar amoniaco (NH3) (Audesirk, 2011). Éste puede ser oxidado a nitrito y nitrato por el proceso de nitrificación, o bien, reducirse a N2 a partir de amonio y nitrito mediante el proceso anammox (Castillo-Rodríguez y col., 2005). Existen organismos desnitrificantes, que reducen anaeróbicamente el nitrato a amonio (NH4 + ), o bien, forman óxidos de nitrógeno, terminando con la liberación de N2O o N2 (Vega-de Kuyper, 2007). El nitrógeno penetra en el medio acuático procedente de fuentes naturales y artificiales. Entre las naturales están: precipitación, polvo, escorrentía y fijación biológica (Metcalf y Eddy, 1979). Como fuentes artificiales se derivan: escorrentía en zonas urbanas, aguas residuales municipales, drenaje de tierras de cultivo y pastos, residuos industriales y filtraciones de las fosas sépticas (EPA, 2012). La química del nitrógeno es compleja. Éste puede tomar diferentes estados de oxidación que pueden ser inducidos por los organismos vivos dependiendo de las condiciones predominantes y los microorganismos presentes. Las formas de 1 interés en aguas residuales son: nitrógeno orgánico, amoniacal, nitritos y nitratos. Todas son formas interconvertibles bioquímicamente y componentes del ciclo del nitrógeno (Romero-Rojas, 2000). El presente trabajo pretende contribuir al estudio de la eliminación de compuestos nitrogenados, como amonio, nitrito y nitrato, presentes en aguas residuales, mediante procesos tipo anammox y desnitrificantes. Se enfatizó en estudiar la estabilización de un consorcio microbiano bajo condiciones anammox mediante el monitoreo de consumo de amonio, nitrito y las velocidades específicas de consumo de amonio. También se realizaron ensayos tipo anammox donde se evaluó la conducta de degradación de amonio y nitrito de un consorcio en presencia y ausencia de nitrato. Esto mediante la medición de eficiencias de consumo y velocidades específicas de consumo de sustrato en reactores por lotes. Para estudiar el proceso desnitrificante se evaluó en ensayos por lotes la degradación de nitrato y producción de nitrógeno molecular de un consorcio microbiano en presencia y ausencia de amonio. Esto mediante la medición de eficiencias de consumo de nitrato, eficiencias de producción de nitrógeno molecular, rendimientos de producción y velocidades específicas de consumo de sustrato. El trabajo también se apoyó en algunos métodos de biología molecular con el objeto de relacionar los aspectos de degradación de nitrato con la diversidad de la comunidad microbiana. 2 Capítulo I. Generalidades 1.1 Antecedentes La presencia de diversos componentes en el agua residual interfiere con la fisiología microbiana. Diferentes estudios se han llevado a cabo para conocer el efecto de compuestos tanto orgánicos como inorgánicos en los procesos biológicos que degradan compuestos nitrogenados. Van Dongen et al. (2001) realizaron estudios de la combinación de una nitrificación parcial por medio del proceso SHARON y Anammox. El proceso SHARON fue realizado en un reactor de continuo de mezcla completa (CSTR) de 10 L, con aireación y un tiempo de retención hidráulico (TRH) de un día. En esta fase el 53% de amonio se convirtió a nitrito, para pasar a la siguiente etapa. El proceso Anammox fue llevado a cabo en un reactor de lodo granular por lotes (SBR) convirtiendo más del 80% de amonio a nitrógeno gaseoso a una carga de 1.2 kgN/m3∙d. Dapena-Mora et al. (2004), estudiaron la estabilidad del proceso Anammox en un reactor gas-lift y uno SBR alrededor de 200 días. El reactor gas-lift trabajó con volumen de 7 L, una zona de sedimentación de 3 L, flujo másico de 2 g/L∙d, 0.9 gN-NH4 + /L, 1.1 gN-NO2 − /L, TRH= 1 día, pH controlado en 8 ± 0.1 y 30°C. Entre los resultados obtenidos se encontró una velocidad máxima específica de consumo de 1.15 g/g∙d, eficiencia de remoción de nitrógeno de 88% y nitrito completamente consumido. El reactor SBR trabajó a un volumen de 1 L, flujo másico de 0.75 g/L∙d 0.375 gN-NH4 + /L, 0.375 gN-NO2 −/L, TRH= 0.625 días, 35 °C, pH entre 7.8 y 8, 78 rpm. Entre los resultados obtenidos se encontró una velocidad máxima específica de consumo de 0.65 g/g∙d, eficiencia de remoción de nitrógeno de 78 % y nitrito completamente consumido. Güven et al. (2005) estudiaron la inhibición de cultivos anammox por enriquecimiento de metanol y etanol. Entre sus hallazgos está la inactivación de 3 anammox bajo la adición de metanol a una concentración de 0.5 mM. Por su parte, el etanol inhibe la reacción anammox en un 30% a 2mM. Martínez-Hernández (2007), analizó el efecto de una fuente fácilmente oxidable como el acetato sobre el consumo de tolueno por desnitrificación, utilizando un reactor anaerobio de flujo ascendente (UASB), donde los productos obtenidos de las reacciones fueron bicarbonatos y nitrógeno molecular, mostrando eficiencias de consumo de carbono de 95.9 ± 3.6% y de nitrato de 87 ± 0.5%. Chong-Jiang et al. (2009) Hallaron efectos negativos significativos de la materia orgánica sobre la eliminación anaeróbica de amonio en un reactor UASB de anammox, encontrando que la razón principal de la disminución del rendimiento de la eliminación anaerobia del amonio pudo ser la desnitrificación heterotrófica. Mathava y Jih-Gaw (2010) encontraron que la coexistencia de la desnitrificación y anammox es técnicamente factible y económicamente favorable cuando las aguas residuales contienen amonio y carbono orgánico. Bettazzi et al. (2010) observaron efecto de inhibición a corto plazo en la actividad Anammox con concentraciones a partir de 60 mgN-NO2 − /L. Asimismo, obtuvieron que las adiciones de compuestos intermediarios, como hidroxilamina e hidracina, restauran la actividad. En investigaciones realizadas por Duran (2013), con un reactor de lecho empacado, se estudió la conducta metabólica de un consorcio desnitrificante para la eliminación de fenol y nitritos, obteniendo evidencias de la capacidad metabólica para desnitrificar con nitrito, oxidando el fenol y obteniendo nitrógeno molecular . Téllez-Pérez (2013), realizó ensayos para demostrar la flexibilidad metabólica de consorcios microbianos (lodos) y cultivos axénicos. El lodo mostró capacidad para efectuar tres procesos respiratorios desnitrificación, metanización y sulfato reducción con rendimientos de producción de N2 (0.9±0.1), CH4 (0.1±0.01) y SH(0.4±0.1). 4 Martínez-Gutiérrez et al. (2013), examinó el consumo biológico del 2-clorofenol (2CF) en cultivos en lote, recurriendo a un lodo desnitrificante fisiológicamente estable. En estos ensayos pudo observar que las fuentes de carbono fácilmente oxidables promueven el consumo de compuestos recalcitrantes. Se utilizó fenol, acetato y glucosa como fuente de carbono, el acetato y 2-CF fueron eficientemente consumidos (Eacetato=100% y E2-CF=94.7±0.5.3) y mineralizados a bicarbonato (Y HCO3-=0.99). 1.2 Planteamiento del problema La descarga de nitrógeno en agua residual a cuerpos de agua provoca afectaciones en el ambiente. Entre ellas se encuentra la eutroficación, cuando junto con el fósforo, actúa como nutriente para el crecimiento de plantas y algas en las aguas. Esto provoca una baja en la entrada de luz y oxígeno, causando la muerte de peces y malos olores. 5 1.3 Justificación La elaboración del presente trabajo aportará información necesaria para comprender el metabolismo autotrófico y organotrófico de consorcios microbianos en el consumo de compuestos nitrogenados. Esta información es relevante y puede aplicarse a diversas áreas. En el área de la investigación cada vez se le da más importancia al estudio de la diversidad microbiana y a su dinámica en presencia de diversos compuestos contaminantes. Este tipo de información ayuda a comprender el mundo microbiano en su hábitat natural. En el área de ingeniería ambiental, esta investigación aporta conocimientos de cómo trabaja un reactor tipo anammox y desnitrificante, mismos que pueden ser utilizados para diseño de reactores, modelados matemáticos, escalamiento o como referencia para tratar agua residual contaminada por compuestos nitrogenados. Este tipo de estudios también es relevante para la industria. Muchas industrias necesitan tratar sus aguas residuales a bajo costo, algunas descargan compuestos nitrogenados aún después de un tratamiento convencional, por lo que los datos obtenidos en este trabajo pueden servir como soporte a nuevos estudios o aplicaciones. Además, abre el panorama de los ingenieros a no trabajar sólo con la parte técnica de bioreactores sino intentar involucrarse a la parte biológica mediante métodos de análisis molecular, mismos que están actualmente complementando las formas convencionales de seguimiento y control. 6 1.4 Objetivos 1.4.1 Objetivo general Evaluar en reactores en lote la degradación de compuestos nitrogenados en aguas residuales por consorcios microbianos anaerobios. 1.4.2 Objetivos específicos Estabilizar en reactores en lote, la degradación de compuestos nitrogenados por consorcios microbianos anaerobios. Analizar, en reactores en lote, la degradación de amonio y nitrito por un consorcio microbiano tipo anammox en presencia de nitrato. Analizar, en reactores en lote, la degradación de nitrato por un consorcio microbiano desnitrificante en presencia de amonio. Analizar, mediante técnicas de biología molecular, la dinámica de las poblaciones microbianas anaerobias en presencia de distintos compuestos nitrogenados. 1.5 Hipótesis La presencia de compuestos nitrogenados distintos a los sustratos del proceso biológico en estudio, tendrá un efecto en la degradación de éstos por los consorcios microbianos anaerobios. 7 Capítulo II: Marco teórico 2.1 Normatividad en México acerca del nitrógeno en aguas residuales De acuerdo a la normatividad en este país, el nitrógeno total es la suma de las concentraciones de nitrógeno Kjeldahl, de nitritos y de nitratos según DOF (1996). La NOM-001-SEMARNAT-1996, establece los límites máximos permisibles de contaminantes en las descargas de aguas residuales en aguas y bienes nacionales (DOF, 1996). Entre estos límites se encuentra el nitrógeno medido como nitrógeno total. La tabla 1 muestra los límites máximos permisibles de nitrógeno total, definido anteriormente, para descargas a aguas y bienes nacionales: Tabla 1. Límites máximos permisibles de nitrógeno total para descargas a aguas y bienes nacionales. Fuente: DOF (1996) Tipo de P.M. de P.D. de cuerpo nitrógeno nitrógeno receptor total (mg/L) total (mg/L) Uso en riego agrícola (A) 40 60 Uso público urbano (B) 40 60 Protección de vida acuática (C) 15 25 Embalses Uso en riego agrícola (B) 40 60 naturales y Uso público urbano (C) 15 25 Estuarios 15 25 Ríos Clasificación artificiales Aguas costeras P.D.=Promedio Diario P.M.= Promedio Mensual 11 (A), (B) y (C): Tipo de Cuerpo Receptor según la Ley Federal de Derechos. 2.2 Compuestos nitrogenados en aguas residuales Entre las aguas residuales con contenido de nitrógeno, como resultado de actividades humanas, se pueden considerar las domésticas. Este tipo de aguas sin tratar, presentan una concentración de nitrógeno total que varía entre 2080ppm (Henry y Heinke, 1999; Metcalf y Eddy, 1979; Villaseñor-Camacho, 2001), nitrógeno amoniacal que se encuentra entre 12-50 ppm (Metcalf y Eddy, 1979; Villaseñor-Camacho, 2001) y nitrógeno de nitrato que alcanza valores mayores a 30 mgN/L en efluentes de plantas de tratamiento (Jiménez-Cisneros, 2001; EPA, 2014). Las aguas residuales industriales, también desechan estos compuestos, éstas pueden dividirse de acuerdo al sector en la industria agraria y el resto de industrias. El sector agrario es responsable de una gran parte de la escorrentía de nitrato hacia los ríos (Baird, 2001). Se sabe que el cultivo intensivo de la tierra, por sí mismo, facilita la formación de nitrato en materia orgánica descompuesta en el suelo, por medio de aireación y humedad (Baird, 2001). Una hectárea de tierra cultivada aporta 400 kg de materias solubles por año, de las cuales aproximadamente 1 kg es N (Jiménez-Cisneros, 2001). La ganadería y las industrias que se derivan de ésta también producen grandes cantidades de compuestos nitrogenados. Entre sus residuos se encuentra el lisier, excreción de ganado. Estas excreciones corresponden a la mayor parte de residuos nitrogenados del ganado. La siguiente tabla muestra brevemente la composición del lisier del ganado doméstico: 12 Tabla 2. Composición del lisier del ganado doméstico. Fuente: SeoánezCalvo, 2004. Características Especie y naturaleza de las deyecciones Bovino mayor Cantidad N total N amoniacal (kg/día) (g/kg) (g/kg) Orina 15 7 - Sólido 30 4 0.4 Lisier completo fresco 45 4.5 0.5 Lisier líquido (20% agua) 55 3.5 1.7 Lisier pastoso ? 3.6 Estiércol ? 4.6 0.5 Bovino menor Lisier 5.8 1.8 0.25 Cerdos (100 Orina 6.7 4 - kg) Heces 3.3 5 - Lisier ? 4.5 3 Orina ? 12 - Heces ? 8 - Lisier 2.5 8 - Aves Lisier 0.2 13 - Conejos Lisier 0.17 12 - Ovino Como puede notarse, estas cantidades son muy grandes y multiplicadas por el flujo másico de lisier producido diariamente, puede observarse que las cargas de nitrógeno se elevan. La problemática de la industria agraria no acaba en eso, muchas industrias derivadas de productos primarios (como la alimentaria) pertenecen a esta categoría. Entre estas, las más conocidas por los desechos nitrogenados de sus efluentes sin tratamiento son: lecheras (Nk=15-20mg/L), azucareras de remolacha (Ntotal=20-80mg/L), fábricas de quesos (Nk=15-20mg/L), vinazas (por HI de alcohol, N=0.6 Kg), destilación de vinos (N=50-500mg/L), mataderos (N= 145mg/L) y 13 vertidos de feculerías (N=400-1000mg/L) (Seoánez-Calvo, 2004). Entre otras se encuentran: plantas de harina de pescado (N total=689mg/L; Namoniacal=568mg/L; Norgánico=121mg/L), planta de harina de papa (NH3-Libre=36mg/L; NH3=30mg/L; Norgánico=528mg/L), producción de almidón de arroz (N total=96mg/L; Namoniacal=15mg/L), producción de almidón de trigo (222-375mg/L), residuos del remojo de lino (Ntotal=7-109mg/L; NH3=3-19mg/L) (CEPIS, 1991). Entre otras industrias, se tienen las de fabricación de seda artificial (NH3>800mg/L), fábrica de cartón de paja (Ntotal=165mg/L; Norgánico=148mg/L; Namoniacal=17mg/L), residuos de textiles de lana y algodón (N orgánico=4-22mg/L; NH3=6mg/L), carbonización de antracita (NH3= hasta 10g/L, producción de tintes orgánicos, fundiciones de hierro (NH3=0-4.8 mg/L), producción de maquinarias y talleres mecánicos, fábricas de margarina (N=7mg/L) (CEPIS, 1991). 2.3 Operaciones y procesos para el control del nitrógeno Los métodos para el control de nitrógeno, de acuerdo a su naturaleza, pueden ser divididos en fisicoquímicos o biológicos. A continuación se describen brevemente algunos de estos. 2.3.1 Métodos fisicoquímicos 2.3.1.1 Separación del amoniaco por arrastre con aire (air stripping) El arrastre con aire del amoniaco es una modificación del proceso de aireación utilizado para la eliminación de gases disueltos en el agua. Los iones amonio se encuentran en el agua residual en equilibrio con el amoniaco, como lo muestra la ecuación (Metcalf y Eddy, 1979): 𝑵𝑯𝟑 + 𝑯𝟐 𝑶 ⇌ 𝑵𝑯𝟒 + + 𝑶𝑯− Ecuación 1 14 Cuando el pH del agua residual aumenta por encima de 7, el equilibrio se desplaza hacia la izquierda, y el ion amonio se convierte en amoniaco (Metcalf y Eddy, 1979). Para conseguir su alcalinización es necesario aumentar la alcalinidad del agua hasta pH=11, mediante el tratamiento con óxido de calcio y someter después a aireación el agua con lo que el nitrógeno se desprende como gas amoniaco (Contreras-López y Molero-Meneses, 2006). En la práctica, se utiliza una torre de arrastre equipada con un soplador de aire. La eficiencia de eliminación depende de la temperatura, tamaño y proporciones de la instalación, así como de la eficiencia del contacto aire-agua. La separación del amoniaco no es satisfactoria cuando la torre no ha sido proyectada correctamente, o bien se encuentra sobrecargada. En este caso, un volumen adicional de aire puede mejorar la operación (Metcalf y Eddy, 1979). En la mayoría de los casos en los que se ha utilizado el arrastre del amoniaco se han presentado problemas como la formación de incrustaciones de carbonato cálcico dentro de la torre y conducciones de alimentación, así como bajo rendimiento al operar en tiempo frío (Ramalho, 2003). La cantidad y naturaleza de las incrustaciones del carbonato cálcico variará según las características del agua y condiciones locales del medio ambiente. Cuando la temperatura disminuye, la cantidad de aire requerida aumenta significativamente para el mismo tipo de eliminación (Metcalf y Eddy, 1979). Aparte de las dificultades de funcionamiento, la descarga del amoniaco a la atmósfera cerca de grandes masas de agua puede crear problemas de contaminación de nitrógeno. En dichas situaciones es necesario un depurador ácido o la adopción de un método alternativo de eliminación de nitrógeno (Metcalf y Eddy, 1979). 15 2.3.1.2 Cloración al breakpoint La cloración al breakpoint es un método alternativo de eliminación de nitrógeno. Consiste en la adición de una cantidad suficiente de cloro para oxidar el nitrógeno amoniacal en disolución a gas nitrógeno y otros compuestos estables (Metcalf y Eddy, 1979). Con un adecuado control y homogenización del caudal, puede reducirse a cero la totalidad del nitrógeno amoniacal presente en el agua residual. Una ventaja adicional es que al mismo tiempo se consigue la desinfección del agua residual (Metcalf y Eddy, 1979). La ecuación que representa la reacción global es la siguiente (Metcalf y Eddy, 1979): 𝟐𝑵𝑯𝟑 + 𝟑𝑯𝑶𝑪𝒍 → 𝑵𝟐 + 𝟑𝑯𝟐 𝑶 + 𝟑𝑯𝑪𝒍 Ecuación 2 La relación estequiométrica de masas entre el cloro expresado como Cl 2 y el amoniaco como N, es 7.6:1. En la práctica, se ha comprobado que la relación varía entre 8:1 y 10:1, aproximadamente (Metcalf y Eddy, 1979). Es importante cuidar esta relación estequiométrica debido a que en caso de relaciones molares de cloro a amoniaco menores que 1, se formarán monocloramina y dicloramina (Metcalf y Eddy, 1979): 𝑵𝑯𝟑 + 𝑯𝑶𝑪𝒍 → 𝑵𝑯𝟐 𝑪𝒍 𝒎𝒐𝒏𝒐𝒄𝒍𝒐𝒓𝒂𝒎𝒊𝒏𝒂 + 𝑯𝟐 𝑶 Ecuación 3 𝑵𝑯𝟐 𝑪𝒍 + 𝑯𝑶𝑪𝒍 → 𝑵𝑯𝑪𝒍𝟐 𝒅𝒊𝒄𝒍𝒐𝒓𝒂𝒎𝒊𝒏𝒂 + 𝑯𝟐 𝑶 Ecuación 4 Entre el punto B y el breakpoint (ver figura 1) algunas de las cloraminas se convierten en tricloruro de nitrógeno, mientras que las cloraminas restantes se oxidarán a óxido nitroso y nitrógeno (Metcalf y Eddy, 1979). 𝑵𝑯𝑪𝒍𝟐 + 𝑯𝑶𝑪𝒍 → 𝑵𝑪𝒍𝟑 𝒕𝒓𝒊𝒄𝒍𝒐𝒓𝒖𝒓𝒐𝒅𝒆𝒏𝒊𝒕𝒓ó𝒈𝒆𝒏𝒐 + 𝑯𝟐 𝑶 Ecuación 5 16 Figura 1. Curva generalizada obtenida durante la cloración al breakpoint. Fuente: Metcalf y Eddy, 1979. El pH óptimo para la reacción es entre 6 y 7. Fuera de este rango se necesita más cloro para alcanzar el breakpoint y la velocidad de reacción es más lenta. La temperatura no parece tener un efecto importante en el proceso, caso contrario a algunas sustancias que interfieren en la reacción (Metcalf y Eddy, 1979). Algunas sustancias fácilmente oxidables que se encuentran en el agua residual son: Fe+2, Mn+2, H2S y la materia orgánica, éstas reaccionan con el cloro reduciendo la mayor parte de él a ion cloruro (Metcalf y Eddy, 1979). Para optimizar el rendimiento de este proceso y minimizar el coste de los equipos e instalaciones, generalmente es necesario hacer una regulación del caudal a tratar. Asimismo, a causa de los problemas de toxicidad potencial que pueden crearse por la descarga de compuestos clorados al medio ambiente, es necesario hacer una decloración del efluente. Para ello se incluyen columnas de carbón activado (Metcalf y Eddy, 1979). 17 2.3.1.3 Intercambio iónico Es un proceso unitario en el cual los iones de una especie dada son desplazados de un material insoluble de intercambio por otros iones de una especie diferente que se encuentra en la solución. Las operaciones de intercambio iónico pueden ser de tipo continuo o discontinuo (batch). En un proceso discontinuo, la resina se agita simplemente con el agua que se ha de tratar en un reactor hasta que se complete la reacción. La resina agotada se extrae por sedimentación y, seguidamente, se regenera y reutiliza. En un proceso continuo, el material de intercambio se coloca en un lecho o una torre y se hace pasar por ella el agua a tratar (Metcalf y Eddy, 1979). La química del proceso de intercambio iónico puede representarse por las siguientes ecuaciones de equilibrio (Metcalf y Eddy, 1979). Reacción: 𝑹𝑯 + 𝑵𝒂+ ⇌ 𝑹𝑵𝒂 + 𝑯+ Ecuación 6 𝑹𝑵𝒂𝟐 + 𝑪𝒂+𝟐 ⇌ 𝑹𝑪𝒂 + 𝟐𝑵𝒂+ Ecuación7 𝑹𝑵𝒂 + 𝑯𝑪𝒍 ⇌ 𝑹𝑯 + 𝑵𝒂𝑪𝒍 Ecuación 8 Regeneración: 𝑹𝑪𝒂 + 𝟐𝑵𝒂𝑪𝒍 ⇌ 𝑹𝑵𝒂𝟐 + 𝑪𝒂𝑪𝒍𝟐 Ecuación 9 Donde R representa la resina. Las ecuaciones anteriores representan las reacciones que tienen lugar en la eliminación de los iones de sodio y calcio del agua, utilizando una resina sintética de intercambio catiónico. Las reacciones de separación mostradas se completarán hasta un punto que depende del equilibrio que se establezca entre los iones en la fase acuosa y los de la fase sólida (Metcalf y Eddy, 1979). 18 Aunque se dispone de resinas de intercambio iónico naturales y sintéticas, no son estas últimas las más utilizadas en razón de su durabilidad. No obstante, algunas resinas naturales (zeolitas) han encontrado aplicación en la eliminación de amoniaco del agua residual. El ion amonio separado de la zeolita se convierte en amoniaco debido al elevado pH. Al llegar a este punto, la solución regenerante se hace pasar a través de una torre de arrastre para la separación de amoniaco (Metcalf y Eddy, 1979). Entre las desventajas de este proceso se encuentra la formación de precipitados de carbonato cálcico dentro del hecho de intercambio de la zeolita, así como en la torre de arrastre y en las conducciones. Otro problema que resulta en desventaja, es el aglutinamiento de la resina causado por la materia orgánica residual presente en el efluente procedente de un tratamiento biológico. Este último problema se puede resolver filtrando previamente el agua residual o utilizando resinas intercambiadoras eliminadoras antes de su aplicación a la torre de intercambio (Metcalf y Eddy, 1979). El intercambio iónico se utiliza para remover altas concentraciones de compuestos nitrogenados (Liu y Lipták, 1997). Algunas empresas utilizan resinas de intercambio aniónico para la eliminación de nitratos. La resina se regenera con salmuera y se acondiciona mediante tratamiento con ácido y metanol. Puede requerirse el pretratamiento mediante filtración en los procesos de intercambio iónico, con lo cual se incrementan los costes (Ramalho, 2003). 2.3.1.4 Procesos de precipitación En lo que se refiere a la eliminación de nitrógeno, los métodos de precipitación son bastante ineficaces, suponiendo menos del 30% de eliminación del nitrógeno total (Ramalho, 2003). Las tentativas de utilizar resinas, electrodiálisis, precipitación electroquímica en forma de MgNH4PO4 o la cloración, no han dado de momento resultados suficientemente satisfactorios (Orozco-Barrenextea y col., 2011). 19 2.3.2 Métodos biológicos 2.3.2.1 Nitrificación La degradación de la materia orgánica nitrogenada acaba generando amonio. La nitrificación consiste en la oxidación del amonio a nitrato (Castillo-Rodríguez y col., 2005). Este proceso se lleva a cabo, principalmente, en suelos bien aireados y de pH neutro por la acción de un número muy limitado de bacterias autótrofas, aerobias o anerobias, denominadas nitrificantes (Parés y col., 2002). La conversión de amonio en nitrato requiere de la actividad metabólica de dos tipos de microorganismos (Castillo-Rodríguez y col., 2005). Al primer grupo se le conoce como grupo nitroso, y sus representantes se encargan de catalizar la transformación del amonio en nitrito. El nitrito es a su vez el sustrato de microorganismos del segundo grupo, denominado nitro, que lo transforman en nitrato (Parés y col., 2002). Aunque cada una de las oxidaciones la lleva a cabo una población bacteriana distinta, los dos procesos están muy relacionados y no se produce acumulación de nitrito (Castillo-Rodríguez y col., 2005). 2.3.2.1.1Las bacterias nitrificantes Ambos procesos son exergónicos y las bacterias nitrificantes son gramnegativas y estrictamente aerobias. Se incluyen dentro de la familia Nitrobacteriáceas y son quimiolitótrofas, es decir, utilizan la energía derivada de la nitrificación para asimilar el CO2 (Castillo-Rodríguez y col., 2005). 2.3.2.1.1.1 El grupo nitroso Microbiología del proceso Los géneros y especies de bacterias pertenecientes a este grupo se reseñan en la tabla 3. Dentro del grupo, Nitrosomonas europea es la especie caracterizada. No obstante, especies del género Nitrosolobus parecen ser las principales responsables de la nitrificación en los suelos agrícolas (Parés y col., 2002). 20 Tabla 3. Géneros representativos de bacterias nitrificantes. Fuente: Parés y col., 2002. Morfología Motilidad Hábitat ± Suelo, aguas dulces, Grupo nitroso Nitrosomonas Bacilos saladas o residuales Nitrosospira Espirales + Suelo Nitrosococcus Cocos ± Suelo Nitrosolobus Pleomórficos + Suelo Bacilos cortos ± Suelo, aguas dulces, Grupo nitro Nitrobacter aguas saladas Nitrospina Bacilos largos y finos - Aguas saladas Nitrococcus Cocos grandes + Aguas saladas Nitrospira Células - Aguas saladas helicoidales a vibrioides Bioquímica del proceso Los electrones producidos en la oxidación del amonio hasta nitrito no se transfieren en un único paso, sino en tres etapas de dos electrones cada una (Castillo-Rodríguez y col., 2005). En la primera de ellas, catalizada por la amonio monooxigenasa, se incorpora uno de los átomos de oxígeno del O 2 atmosférico a la molécula de amonio, formándose hidroxilamina (Castillo-Rodríguez, 2005); no se produce energía en este proceso (Parés y col., 2002). 𝑵𝑯𝟑 + 𝑶𝟐 + 𝟐𝑯+ + 𝟐𝒆− → 𝑵𝑯𝟐 𝑶𝑯 + 𝑯𝟐 𝑶 Ecuación 10 Los dos pasos siguientes están catalizados por la hidroxilamina óxido-reductasa. En primer lugar, la hidroxilamina se oxida con una molécula de H2O a nitrosilo (que permanece unido al sitio activo), y otro sistema enzimático que oxida al 21 intermediario hipotético (NOH) el cual se oxida finalmente a NO2 − (Castillo- Rodríguez y col., 2005; Parés y col., 2002): 𝑵𝑯𝟐 𝑶𝑯 + 𝑯𝟐 𝑶 → 𝑵𝑶𝑯 → 𝑵𝑶𝟐 − + 𝟓𝑯+ Ecuación 11 La amonio monooxigenasa es una enzima integral de la membrana plasmática, mientras que la hidroxilamina óxido-reductasa se encuentra en el periplasma (Castillo-Rodríguez y col., 2005). En cuanto a la obtención de energía, hay que considerar que los electrones de los compuestos nitrogenados entran en una cadena de transporte de electrones, y el flujo a través de ella establece un potencial de membrana y una fuerza protón motriz suficiente para producir ATP (Castillo-Rodríguez y col., 2005). En la primera etapa del proceso de transformación del amoniaco en nitrito, es decir, la formación de hidroxilamina, se aportan dos electrones al sistema (Parés y col., 2002). Estos electrones son aportados por el proceso de oxidación de la hidroxilamina, que genera cuatro electrones. Es decir, de cada 4 electrones generados en la oxidación de amoniaco a nitrito, sólo dos entran en la cadena respiratoria. Debido a que los potenciales de óxido-reducción de las parejas NH4 + /NH2OH (+899 mV) y NO2 − /NH2OH (+66 mV) son demasiado positivos como para reducir NAD+ (Parés y col., 2002), es necesario un flujo inverso de electrones para generar el poder reductor a partir de ATP. Dicho flujo se deriva de la cadena respiratoria y es necesario para fijar el CO2 (Castillo-Rodríguez y col., 2005). Pero, en función del potencial redox del par NO2 −/NH2OH, los electrones entran a nivel del citocromo c, con lo que solamente puede utilizarse un punto de fosforilación en la cadena respiratoria establecida (Parés y col., 2002). 2.3.2.1.1.2 El grupo nitro Este grupo incluye las bacterias que oxidan el nitrito hasta nitrato (Parés y Col., 2002). Las bacterias del grupo nitro emplean una nitrito oxidasa para oxidar el nitrito a nitrato en un solo paso (Castillo-Rodríguez y col., 2005). Esta óxido22 reductasa es una molibdoproteína localizada en la parte interna de la membrana (Castillo-Rodríguez y col., 2005). Los electrones son transportados al oxígeno mediante una cadena que incluye a los citocromos a1, c y aa3 (Parés y col., 2002). Sin embargo, al contrario de lo que pudiera pensarse, el nitrito no es oxidado directamente por el O 2, sino por el H2O, que se utiliza como donador de H2 para la reducción concomitante del NAD + (Castillo-Rodríguez y col., 2005). El O2 sólo sirve como aceptor terminal de los electrones. La reacción global es (Castillo-Rodríguez y col., 2005). 𝟏 𝑵𝑶𝟐 − + 𝟐 𝑶𝟐 → 𝑵𝑶𝟑 − (△ 𝐺°′ = −74.1 𝑘𝐽𝑚𝑜𝑙 −1 ) Ecuación 12 Debido al alto potencial redox del par NO2 −/NO3 − (E’0=+0.42 V), la cadena de transporte de electrones que se establece es muy corta (Parés y col., 2002). Por lo anterior, aunque la energía liberada en la oxidación sería suficiente para la fosforilación de dos moléculas de ADP, parece que sólo se sintetiza una molécula de ATP (Castillo-Rodríguez y col., 2005). La generación de la fuerza protón motriz que dirige la síntesis de ATP ocurre por acción de la oxidasa terminal, el citocromo aa3 (Castillo-Rodríguez y col., 2005). Debido al bajo rendimiento energético de su oxidación, es necesario oxidar hasta cinco moléculas de nitrito para conseguir la reducción de una NAD +, mediante el transporte inverso de electrones (Castillo-Rodríguez y col., 2005). Por ello, el tiempo de duplicación de los Nitrobacter es de unas 18 horas (Parés y col., 2002). Microbiología del proceso Las bacterias del grupo nitro usualmente se encuentran asociadas a las bacterias del grupo nitroso en sus hábitats naturales. El género más característico es Nitrobacter, siendo N. winogradski probablemente la especie más estudiada (Parés y col., 2002). Pero la nitrificación no es un proceso exclusivo de los géneros Nitrosomonas y Nitrobacter, ya que hay especies de los géneros 23 Nitrosospira, Nitrosococcus, Nitrosovibrio y Nitrosolobus, que pueden oxidar amoniaco a nitrito, e incluso algunas especies de Nitrosolobus que juegan un papel destacado en la nitrificación de suelos agrícolas. Por su parte, la oxidación de nitrito a nitrato se ha descrito en diferentes miembros de los géneros Nitrococcus, Nitrospira y Nitrospina. Cualquier tipo de estrés puede afectar gravemente al proceso de nitrificación, especialmente al segundo paso, por lo que en condiciones estresantes puede producirse una acumulación de nitrito (CastilloRodríguez y col., 2005). En los miembros del grupo nitro el quimiolitotrofismo no tiene un carácter tan estrictamente obligado como en el grupo nitroso (Parés y col., 2002). Nitrobacter agilis, además de crecer aerobiamente a expensas del nitrito, también es capaz de crecer, en cierta medida, mixotróficamente e incluso heterotróficamente (Parés y col., 2002). Nitrobacter reduce nitratos aerobiamente, interviniendo en esta reducción el citocromo c y un componente del citocromo a1. El nitrito producido puede ser utilizado como sustrato de la cadena respiratoria. Un exceso de nitrito no inhibe la ulterior reducción de más nitrato. El oxígeno tampoco la inhibe, lo cual contrasta con otros sistemas de nitrato reductasas. En cambio, un exceso de nitrato en el medio inhibe el consumo de oxígeno (Parés y col., 2002). N. agilis también contiene nitrito reductasa e hidroxilamina reductasa, las cuales se utilizan, junto a la nitrato reductasa, para asimilar el nitrato. Las dos primeras enzimas son solamente activas en condiciones anaerobias (Parés y col., 2002). Se ha demostrado que algunos eucariotas (Fabaceae) llevan a cabo un proceso nitrificante sui generis, consistente en la oxidación de aminoácidos a compuestos nitroalifáticos (3-nitropropionato y 3-nitropropanol), que pueden liberar el grupo nitro como nitrito y nitrato. Cuando esto sucede en plantas fijadoras, se produce un compendio del ciclo del nitrógeno en un solo sistema bacteria-planta (CastilloRodríguez y col., 2005). 24 2.3.2.2 Desnitrificación El nitrato, además de usarse cómo fuente de nitrógeno para el crecimiento, puede ser utilizado por una amplia variedad de microorganismos como aceptor final de electrones en ausencia de oxígeno. Este proceso de respiración anaeróbica se denomina desnitrificación, da lugar a la formación de diversas especies reducidas de nitrógeno y va acompañado de la oxidación simultánea de materia orgánica (Castillo-Rodríguez y col., 2005). El rendimiento energético de la reducción desasimiladora (respiración) de nitrato es superior al que se obtiene de la fermentación de los correspondientes compuestos orgánicos (Castillo-Rodríguez y col., 2005). Existen dos tipos de organismos desnitrificantes, si la reducción anaeróbica del nitrato termina en NH4 + (ocurre fundamentalmente en enterobacterias), o si transcurre con formación de diversos óxidos de nitrógeno, además del nitrito, y que termine con la liberación de N2 (Castillo-Rodríguez y col., 2005). Dentro del primer tipo, se encuentran las plantas y algunas bacterias, donde hay una reducción asimiladora del nitrato, el cual pasa a nitrito que luego se reduce a amoniaco, pasando por hidroxilamina (Parés y col., 2002): 𝑵𝑶𝟑 − → 𝑵𝑶𝟐 − → 𝒙 → 𝑵𝑯𝟐 𝑶𝑯 → 𝑵𝑯𝟒 + Ecuación 13 El amoniaco es incorporado al material celular. El sistema enzimático implicado en el proceso asimilador del nitrato incluye flavina, molibdeno, NAD y NADP reducidos como dadores de hidrógeno y electrones (Parés y col., 2002). El nitrato puede sustituir al oxígeno en algunas bacterias fermentativas, las cuales entonces pueden utilizar para el crecimiento sustratos no fermentables. En estos casos puede ocurrir otra reducción desasimiladora del nitrato parecida a la reducción asimiladora y que lleva a la excreción de NH4 + (Parés y col., 2002). También puede tener lugar la acumulación de nitritos. Se ha pensado que se trata de una respiración incidental del NO3 − que permitiría la eliminación de hidrógeno y una mayor disponibilidad de NAD+, con evidencias de un mayor rendimiento 25 energético. Esta ruta es característica de las enterobacteriáceas y también de algunos miembros de los géneros Bacillus y Clostridium (Parés y col., 2002). Por otro lado, la desnitrificación se refiere a la reducción desasimiladora de los óxidos de nitrógeno iónicos (nitrato: NO3 − y nitrito: NO2 − ) que llevan a cabo algunas bacterias aerobias obligadas en ausencia de oxígeno (Parés y col., 2002). En condiciones anaerobias no hay crecimiento en ausencia de NO3 − o de NO2 − . Los productos de reducción son óxidos de nitrógeno gaseosos (NO y N 2O), los cuales pueden ser reducidos a N2, su producto final más importante (Parés y col., 2002). La ecuación que describe el proceso antes mencionado es la siguiente: 𝑵𝑶𝟑 − → 𝑵𝑶𝟐 − → 𝑵𝑶 → 𝑵𝟐 𝑶 → 𝑵𝟐 Ecuación 14 La desnitrificación necesita condiciones anóxicas o de presiones de oxígeno bajas, aunque en algunas ocasiones puede darse en ambientes aeróbicos si éstos contienen microhábitats anóxicos (Castillo-Rodríguez y col., 2005). Los procesos de nitrificación y desnitrificación en el suelo ocurren a menudo en el mismo lugar, de manera que una parte del nitrato formado en el primero se difunde hacia la zona de desnitrificación anaeróbica, donde se reduce a N 2 (Castillo-Rodríguez y col., 2005). Por una parte se encuentra Paracoccus denitrificans que puede oxidar H2 (Parés y col., 2002): 𝟓𝑯𝟐 + 𝟐𝑵𝑶𝟑 − → 𝑵𝟐 + 𝟒𝑯𝟐 𝑶 + 𝟐𝑶𝑯− Ecuación 15 Paracoccus denitrificans puede reducir NO3 − tanto con H2 como con compuestos orgánicos (Parés y col., 2002). Las fuentes de carbono utilizadas lactato, piruvato y formiato, pero también puede utilizar el glicolato como precursor de ácidos grasos C4 dicarboxílicos, sin pasar por intermediarios con coenzima A (Parés y col., 2002). La tabla 4 muestra diversos compuestos que se utilizan como factores de enriquecimiento para las bacterias reductoras de nitrato: 26 Tabla 4. Factores de enriquecimiento para diferentes bacterias reductoras de 𝐍𝐎𝟑 − en un medio mínimo en condiciones anaerobias. Fuente: Parés y col., 2002. Factor de enriquecimiento Bacteria Con NH3 - Trazas de peptona y etanol o Pseudomonas aeruginosa propionato - Glucosa - Tartrato, succinato o malato - Ácidos extracto Pseudomonas fluorescens orgánicos, de Pseudomonas stutzeri etanol carne con y alta Bacillus licheniformis − concentración de NO3 (5-12%) - Trazas de extracto de levadura y Paracoccus denitrificans atmósfera de H2 Sin NH3 - Lactato y piruvato Enterobacterias Por otra, Thiobacillus denitrificans, oxida azufre y tiosulfato con NO3 − en sustitución del O2 (Parés y col., 2002): 𝟓𝑺 + 𝟔𝑵𝑶𝟑 − + 𝑯𝟐 𝑶 → 𝟓𝑺𝑶𝟒 −𝟐 + 𝟑𝑵𝟐 + 𝟒𝑯+ Ecuación 16 Estos casos se incluyen dentro de bacterias desnitrificantes (Parés y col., 2002). Para el aislamiento de bacterias que usan el NO3 − como oxidante se utilizan medios de enriquecimiento apropiados para cada tipo, dada la gran diversidad taxonómica de las especies que presentan esta propiedad (Parés y col., 2002). 2.3.2.2.1Bioquímica de la desnitrificación La primera fase de la reducción respiratoria de nitrato consiste en su conversión a nitrito (Castillo-Rodríguez y col., 2005). Esta reacción está catalizada por una nitrato reductasa, desasimiladora. Dicha enzima es una molibdoproteína, con un 27 cofactor de molibdeno (MGD), asociada a la membrana. Está formada por tres subunidades diferentes, no se inhibe por amonio y su síntesis se induce por nitrato y se reprime por el oxígeno molecular. La subunidad catalítica (α, NarG) es la mayor (112-140 kDa) y contiene el cofactor MGD, mientras que en la subunidad β (NarH; 52-64 kDa) hay un centro de 3Fe-4S y tres 4Fe-4S, y en la subunidad γ (NarI; 19-25 kDa), que es una proteína integral de membrana, existe una agrupación bihemob que oxida al ubiquinol (Castillo-Rodríguez y col., 2005). Las subunidades α y β se encuentran unidas a la cara citosólica de la subunidad γ y se pueden solubilizar por tratamiento con detergentes o por calor (Castillo-Rodríguez y col., 2005). Los electrones para la reducción proceden de la formiato deshidrogenasa, a través de una cadena transportadora que implica citocromos bc1 y las quinonas de la membrana (Castillo-Rodríguez y col., 2005). La fuerza protón motriz para la producción de ATP se genera por un mecanismo en el que la oxidación de las quinonas por el hemo b de Narl localizado en la cara periplásmica de la membrana provoca la liberación de 2H+ al periplasma. La transferencia de electrones tiene lugar primeramente entre los grupos hemo b de Narl, de bajo y alto potencial, localizados en caras opuestas de la membrana (Castillo-Rodríguez y col., 2005). A través de los centros Fe-S de NarH pasan a NarG donde, con la participación del cofactor MGD, permiten la reducción de nitrato, reacción en la que se consumen además 2H+ citoplásmicos (Castillo-Rodríguez y col., 2005). Debido al potencial de reducción del par NO3 −/NO2 − (+0.43 V), el rendimiento energético de la respiración anaeróbica con nitrato es inferior al que se obtiene cuando se emplea oxígeno como aceptor terminal de los electrones ( 1/2 O2/H2O, +0.82V) (Castillo-Rodríguez y col., 2005). Las bacterias desnitrificantes reducen el nitrito en el periplasma a óxidos de nitrógeno y N2 como producto final. Una nitrito reductasa periplásmica, reduce el nitrito a óxido nítrico (NO), que es bastante tóxico, por lo que su concentración debe mantenerse por debajo de concentraciones nanomolares (Castillo-Rodríguez y col., 2005). Esto se consigue mediante el acoplamiento adecuado con la óxido 28 nítrico reductasa. La producción del óxido nitroso (N2O) tiene lugar en su cara periplásmica (Castillo-Rodríguez y col., 2005). El último paso consiste en la reducción del N2O a N2 catalizada por la óxido nitroso reductasaperiplásmica (NosZ) (Castillo-Rodríguez y col., 2005). En el proceso de transferencia de electrones se encuentran implicados el citocromo bc1 de la membrana y un citocromo c555 periplásmico. Durante el mismo se produce una fuerza protón motriz, con producción de ATP adicional, debido a que la NO reductasa de membrana está ligada a la extrusión de protones (Castillo-Rodríguez y col., 2005). Todas las enzimas respiratorias de este sistema se inducen en anaerobiosis por sus sustratos (NO3 − , NO2 − , NO y N2O), independientemente del balance C/N (Castillo-Rodríguez y col., 2005). Las reductasas de los óxidos de nitrógeno se hallan reprimidas durante el crecimiento aerobio (Parés y col., 2002). En condiciones anaerobias, la fase de desrepresión en presencia de NO3 − es de unas 3 horas (Parés y col., 2002). El control se lleva a cabo a nivel trascripcional mediante reguladores tipo FNR, factor de transcripción que detecta aumentos en la pO2, IHF y NarXL/NarPQ, sistema de dos componentes (sensor/regulador) que se fosforila en respuesta a cambios en la concentración de nitrato o nitrito (Castillo-Rodríguez y col., 2005). En los organismos desnitrificantes se produce una red compleja de interacciones entre diversos estímulos ambientales (presencia de oxoaniones nitrogenados y O2) y proteínas de tipo FNR, que controlan la transcripción de los genes nar, nir, nor y nos (Castillo-Rodríguez y col., 2005). La N2O reductasa es la que resulta más severamente inhibida por la presencia de oxígeno, por lo que su presencia provoca la acumulación de N 2O (Castillo-Rodríguez y col., 2005). 2.3.2.2.2Microbiología del proceso La diversidad taxonómica de las bacterias desnitrificantes es muy grande, ya que entre los microorganismos que llevan a cabo la desnitrificación se encuentran organótrofos (Pseudomonas, quimiolitótrofos (Rhizobium, (Thiobacillus, Azospirillum), Alcaligenes, Thiomicrospira, Bacillus, Agrobacterium), Nitrosomonas), fotótrofos (Rhodopseudomonas), diazótrofos arqueobacterias 29 (Halobacterium), y otros (Castillo-Rodríguez y col., 2005). La mayoría pueden reducir NO3 − a N2, pero algunas bacterias carecen de una o varias de las enzimas que participan en el proceso global, por lo que sólo pueden llevar a cabo la reducción a partir de NO2 − o sólo pueden llegar a alguno de los productos intermedios (Castillo-Rodríguez y col., 2005). Cada vez son encontradas más características distinguibles de la microbiología desnitrificante. Paracoccus es una bacteria desnitrificante aerobia, comúnmente encontrada en procesos simultáneos de nitrificación- desnitrificación (Yuan y col., 2014). Algunas bacterias como Comamonadaceae y Rhodocyclaceae del grupo Beta-proteobacteria y Rhodobacteraceae del grupo Alpha-proteobacteria son encontradas cuando se utiliza el acetato como el sustrato de carbono (Ginige y col., 2005). Cuando se utiliza metanol como fuente de carbono se detectan Methylophilaceae y Hyphomicrobiaceae (Osaka, 2006). También han sido detectadas bacterias del grupo taxonómico γ-proteobacteria, como Pseudomonas halodenitricans en digestores termofílicos que tratan agua residual de cerdos (Patil y col., 2010). 2.3.3.3 Oxidación anaeróbica de amonio (Anamox) Se han encontrado bacterias capaces de llevar a cabo la formación de N 2 a partir de amonio y nitrito, bajo condiciones anaerobias (Castillo-Rodríguez y col., 2005). Dichas bacterias son conocidas como anamox (o “anammox”, de “anaerobic ammonia oxidation”). Las bacterias anammox convierten amonio con nitrito (como aceptor de electrones) a nitrógeno gaseoso con hidracina (N2H4) e hidroxilamina (NH2OH) como intermediarios. Esta reacción es llevada a cabo 15 veces para fijar una molécula de dióxido de carbono con el nitrito como donador de electrones que conduce a la producción anaerobia de los nitratos en el anabolismo (Strous y col., 1998). Catabolismo (15x): 𝑵𝑯𝟒 + + 𝑵𝑶𝟐 − → 𝑵𝟐 + 𝟐𝑯𝟐 𝑶 Ecuación 17 30 Anabolismo: 𝑪𝑶𝟐 + 𝟐𝑵𝑶𝟐 − + 𝑯𝟐 𝑶 → 𝑪𝑯𝟐 𝑶 + 𝟐𝑵𝑶𝟑 − Ecuación 18 La ecuación general queda de la siguiente manera: 𝑵𝑯𝟒 + + 𝟏. 𝟑𝟐𝑵𝑶𝟐 − + 𝟎. 𝟎𝟔𝟔𝑯𝑪𝑶𝟑 − + 𝟎. 𝟏𝟑𝑯+ → 𝟏. 𝟎𝟐𝑵𝟐 + 𝟎. 𝟐𝟔𝑵𝑶𝟑 − + 𝟎. 𝟎𝟔𝟔𝑪𝑯𝟐 𝑶𝟎.𝟓 𝑵𝟎.𝟏𝟓 + 𝟐. 𝟎𝟑𝑯𝟐 𝑶 Ecuación19 Como es observado en la ecuación 18, el hábitat de la bacteria anammox requiere la presencia simultánea de amonio y nitrito, cerca de la interfase aeróbicaanaeróbica de sedimentos en cuerpos de agua. El amonio es producido por la degradación anaeróbica de materia orgánica, amonificación y reducción desasimilatoria de nitrito y/o nitrato. El nitrato se origina de la reducción de nitrato, originada por organismos desnitrificantes o por la bacteria anammox en presencia de compuestos orgánicos como formato, acetato o propionato (Kuenen, 2008). El descubrimiento de este proceso es relevante desde el punto de vista ambiental, ya que se pensaba que el amoniaco era estable en medios anóxicos, pero ahora se abren perspectivas en campos como el tratamiento de aguas residuales anóxicas, para eliminar el amoniaco y las aminas. Aunque estas bacterias son litoautotróficas, se ha visto que pueden utilizar simultáneamente amonio y acetato como donadores de electrones para la reducción de NO3 − a N2, mediante una combinación de dos vías diferentes (Castillo-Rodríguez y col., 2005). Este proceso también se puede dar acoplado a la producción de nitrato por bacterias nitrificantes. En este caso, el nitrito se obtiene de la oxidación de amoniaco por las bacterias desnitrificantes. Ambos grupos de bacterias oxidadoras de amoniaco, aerobias y anaerobias, pueden coexistir en ambientes ricos en dicho compuesto (como son las aguas residuales y otras de desecho), ya que en ellos se encuentran partículas en suspensión que presentan zonas óxicas y anóxicas (Castillo-Rodríguez y col., 2005). En contraste con otros procariotas conocidos, los Planctomycetos típicamente, poseen compartimentos subcelulares con membrana, cuya función es estudiada. Los estudios de microscopía electrónica indican que la bacteria anammox 31 contiene un compartimento membranal anammox-específico, al cual le dieron el término de „anamoxosoma‟ (Kuenen, 2008). Se ha observado la existencia de organismos anamoxótrofos oceánicos mediante la detección de laderanos (lípidos que rodean los anamoxosomas, como compartimentos, en los planctomicetos de los digestores) y secuenciación de DNA (Castillo-Rodríguez y col., 2005). Se han encontrado las membranas de laderanos sólo en bacterias anammox. Esto plantea el significado funcional de los lípidos de laderanos (van Niftrik y col., 2004). 2.3.3.3.1 Microbiología Entre las bacterias que realizan el proceso anammox, se encuentran Brocadia anammoxidans, Kuenenia sturrgartiensis, Candidatus Anammoxoglobus propionicus y Scalindua sorokinii (Kuenen, 2008; Kuenen y Jetten, 2001; Jetten y col., 2001; Schmid y col., 2000). Estas bacterias son cocos con diámetro menor a 1 μm. Tienen un “tiempo de generación” de 10-30 días y son fisiológicamente distintas a otros planctomicetos debido a que son anaerobios quimiolitoautótrofos (van Niftrik y col., 2004). 2.3.3.3.2 Modelo bioquímico para la acumulación de una fuerza protón motriz y la síntesis de ATP Una de las enzimas clave de la reacción anammox, la enzima hidracina-oxidasa, se purificó y mostró que estaba situada en el anammoxosoma (Lindsay y col., 2001). Esto indica que el catabolismo anammox se lleva a cabo dentro del anammoxosoma. Se ha propuesto un modelo bioquímico en el cual amonio e hidroxilamina están combinados por la hidracina-hidrolasa (HH), la enzima formadora de hidracina. La hidrazina se oxida por una enzima hidracina-oxidasa (HZO). HZO ha demostrado tener cierta similitud con hidroxilamina-oxidoreductasa (HAO) de Nitrosomonas europea (Jetten y col., 1998). La oxidación, que tiene lugar en el interior de la anammoxosoma, resulta en nitrógeno gaseoso, cuatro protones y cuatro electrones. Estos cuatro electrones se utilizan junto con cinco 32 protones del riboplasma por una enzima nitrito-reductasa (NIR) para reducir el nitrito a hidroxilamina (Kuenen y Jetten, 2001). Figura 2. Postulado del acoplamiento del proceso anammox a la membrana de la anammoxosoma. HH: hidracina-hidrolasa, HZO: enzima hidracinaoxidasa, NIR: enzima nitrito-reductasa Fuente: (van Niftrik y col., 2004). En el modelo, la reacción ANAMMOX establece un gradiente de protones por el consumo efectivo de protones en el riboplasma y la producción de protones dentro de la anammoxosoma, un mecanismo conocido como separación de cargas. Esto resulta en un gradiente de protones electroquímico dirigido desde la anammoxosoma al riboplasma. Este gradiente contiene energía química potencial - El gradiente químico de protones resulta en una diferencia de pH (△pH), donde el riboplasma es alcalino comparado con el anammoxosoma- y un potencial eléctrico de energía –el gradiente eléctrico de protones resulta en una diferencia de cargas (△ᴪ), donde el riboplasma está cargado negativamente comparado al anammoxosoma. Ambos, △pH y △ᴪ, tienen un tener una fuerza base sobre los protones desde el interior al exterior de la anammoxosoma: la fuerza motriz de protones (△p). Esto podría ser utilizado para conducir la síntesis de ATP catalizada por las adenosintrifosfatasas ligadas a la membrana (ATPasas), ubicadas en la membrana de la anammoxosoma (figura 2). Los protones fluirían pasivamente de nuevo en el riboplasma (con el gradiente electroquímico de protones=cuesta abajo) a través de los poros de los protones formados por las 33 ATPasas, de una manera análoga a lo conocido en los sistemas de fosforilación oxidativa de respiración y la fosforilación fotosintética. Las ATPasas ligadas a la membrana de la anammoxosoma se ubicarían con su globular, la de dominio hidrofílico sintetizando ATP en el riboplasma y su dominio hidrofóbico traslocando protones en la membrana de la anammoxosoma. El ATP sintetizado sería entonces lanzado en el riboplasma (van Niftrik y col., 2004). 2.4 Ingeniería de procesos anaerobios Para poder realizar pruebas con procesos desnitrificantes y anammox, es necesario conocer el fundamento de éstos en términos de reactores. 2.4.1 Bioprocesos anaerobios Los procesos anaeróbicos están definidos como procesos biológicos en los cuales es metabolizado material biodegradable en un ambiente libre de oxígeno disuelto o sus precursores (como H2O2). Los procesos anaerobios están clasificados en fermentación anaerobia o respiración anaerobia de acuerdo al tipo de aceptor de electrones (Khanal, 2008). 2.4.1.1 Fermentación anaerobia En este la materia orgánica es catalizada en ausencia de un aceptor de electrones externo por anaerobios estrictos o facultativos a través del balance de reacción de óxido-reducción bajo condiciones de oscuridad. El producto generado durante el proceso acepta los electrones liberados durante el desglose de materia orgánica. Entonces la materia orgánica actúa como donador y aceptor de electrones. El sustrato es sólo parcialmente oxidado y sólo una pequeña cantidad de la energía contenida en él es conservada. La mayor parte de ATP o energía es generada por fosforilación nivel sustrato (Khanal, 2008). 2.4.1.2 Respiración anaerobia La respiración anaerobia por otro lado requiere aceptores de electrones externos para la disposición de electrones liberados durante la degradación de materia 34 orgánica. El aceptor de electrones en este caso puede ser 𝑁𝑂3 − , como sucede en bacterias desnitrificantes, 𝑁𝑂2 − , en bacterias anammox, etc. Tanto la fosforilación nivel sustrato como la fosforilación oxidativa generan energía (o ATP). La energía liberada bajo tal condición es mejor que la fermentación anaerobia (Khanal, 2008). La tabla 5 presenta algunas bacterias anaerobias junto a sus respectivos donadores y aceptores de electrones y fuente de carbono: Tabla 5. Grupos microbianos y su preferencia en donadores y aceptores de electrones, y fuentes de carbono. Fuente: Khanal, 2008. Microbios Aceptor de Donador de Fuente de electrones electrones carbono Acetotróficas (o acetoclásticas) Acetato Acetato Acetato Hidrogenotróficas CO2 H2 CO2 NO3 − , NO2 − Carbono Carbono orgánico orgánico NO3 − , NO2 − S° o H2 CO2 Acetotróficas (o acetoclásticas) SO4- Acetato Acetato Hidrogenotróficas SO4- H2 CO2 NH4+ CO2 Bacterias metanogénicas Bacterias reductoras de nitrito/ nitrato Desnitrificantes heterotróficas Desnitrificantes autótrofas Bacterias reductoras de sulfato Bacterias anaeróbicas oxidadoras de NO2 − amonio La afinidad del microorganismo por el aceptor de electrones está en el siguiente orden: 𝑶𝟐 > 𝑵𝑶𝟑 − > 𝑴𝒏𝑶𝟐 > 𝑭𝒆𝑶𝑯 > 𝑺𝑶𝟒 −𝟐 > 𝑪𝑶𝟐 Ecuación 20 35 2.4.2 Consideraciones para el tratamiento de aguas residuales Para el tratamiento de aguas residuales es importante examinar algunos factores importantes que gobiernan el proceso de bioconversión anaeróbica. Éstos incluyen velocidad de carga orgánica, rendimiento de biomasa, velocidad de utilización de sustrato, tiempo de retención hidráulico, tiempo de retención de sólidos, tiempo de arranque, microbiología, factores ambientales y configuración del reactor (Khanal, 2008). 2.4.2.1 Velocidad de carga orgánica volumétrica Los procesos anaerobios están caracterizados por altas velocidades de carga orgánica volumétrica (VOLR, por sus siglas en inglés), esto indica que más agua residual puede ser tratada por unidad de volumen del reactor. VOLR está dada por la siguiente expresión (Khanal, 2008): 𝑽𝑶𝑳𝑹 = 𝑪𝒊 𝑸 𝑽 Ecuación 21 Donde 𝐶𝑖 es la concentración de carbono orgánico disuelto (COD, mg/L) biodegradable del afluente, Q el caudal (m3/día) y V el volumen del reactor (m3). 2.4.2.2 Rendimiento de la biomasa Es una medición cuantitativa de crecimiento celular en un sistema por un sustrato dado. El término comúnmente usado para representar rendimiento de la biomasa es el coeficiente de rendimiento (Yx/s), el cual se expresa como (Khanal, 2008): △𝒙 𝒀𝒙/𝒔 = △𝑺 Ecuación 22 Donde △ 𝑥 es un incremento en la concentración de biomasa (mgSSV/L) y △ 𝑆 como decremento (consumo) de la concentración de sustrato (mg/L). △ 𝑥 puede sustituirse por la formación de producto que también está dada por un incremento (YP/S), de acuerdo a la siguiente ecuación (Scragg, 2009): 𝒀𝑷/𝑺 = △𝑷 △𝑺 Ecuación 23 36 2.4.2.3 Actividad biológica específica Indica la habilidad de la biomasa para utilizar el sustrato. Usualmente es reportado como velocidad específica de consumo (q): 𝐪= 𝐤𝐠 𝐫𝐞𝐦𝐨𝐯𝐢𝐝𝐨 𝐤𝐠𝐒𝐒𝐕∙𝐝í𝐚 Ecuación 24 Donde los kgremovido son el consumo de sustrato y puede expresarse en kg, mg, Mol o mM, y kgSSV, son la biomasa expresada como sólidos suspendidos volátiles (Khanal, 2008). 2.4.2.4 Tiempo de retención hidráulica y tiempo de retención de sólidos El tiempo de retención hidráulica (TRH) y el tiempo de retención de sólidos (TRS) son dos parámetros de diseño importantes en procesos de tratamiento biológico. El TRH indica el tiempo en que se remueve el contaminante en el reactor en contacto con la biomasa. Este valor es dependiente de la velocidad del metabolismo microbiano. El TRS, por otro lado, controla la biomasa en el reactor para alcanzar un determinado grado de estabilización de residuos (Khanal, 2008). 2.4.2.5 Tiempo de arranque Es el periodo inicial de puesta en marcha durante el cual el proceso es conducido a un punto donde el desempeño normal del sistema de tratamiento biológico puede ser conseguido con continua alimentación de sustrato. El tiempo de arranque es una de las mayores consideraciones en procesos anaerobios debido a la baja velocidad de crecimiento de los microorganismos anaerobios y a su susceptibilidad a los cambios en factores ambientales. Los sistemas de tratamiento anaerobios a menudo necesitan un largo periodo de arranque, el cual puede debilitar su competitividad con sistemas de tratamiento aerobio que tienen un periodo de tiempo de arranque corto de 1 o 2 semanas. Este periodo se puede reducir considerablemente si el cultivo microbiano del residuo en cuestión es usado como inóculo. El tiempo de arranque además depende de la concentración inicial de biomasa. Velocidades de carga y factores ambientales como pH, disponibilidad de nutrientes, temperatura, y el potencial de óxido-reducción (ORP) 37 son mantenidos dentro del confort microbiano durante el tiempo de arranque (Khanal, 2008). 2.4.2.6 Microbiología Un proceso anaerobio es un proceso multipaso en el cual un grupo diverso de microorganismos degrada la materia orgánica en orden secuencial resultando en una acción sinérgica. La estabilidad de un sistema de tratamiento es a menudo debatida, debido principalmente a la naturaleza frágil de microorganismos ante los cambios de las condiciones ambientales como pH, temperatura, ORP, disponibilidad de nutrientes/metales traza y toxicidad. Cuando un sistema de tratamiento anaerobio falla debido a la falta de condiciones ambientales adecuadas o al lavado de biomasa del reactor, puede tomar varios meses para que el sistema retorne a las condiciones normales de operación debido a la extremadamente lenta velocidad de crecimiento (Khanal, 2008). Trabajos recientes utilizan consorcios microbianos para el tratamiento anaerobio. Un consorcio microbiano consiste de varias bacterias y/u hongos expuestos a un rendimiento superior de biodegradación a cepas individuales por medio de la combinación de actividades enzimáticas de degradación inherentes a los miembros individuales del consorcio (Mikesková, 2012). Diversos estudios demuestran la viabilidad de la biodegradación mediante cultivos microbianos mixtos. 2.4.2.6 Factores ambientales Los procesos anaerobios son severamente afectados por los cambios en las condiciones ambientales. Algunos factores ambientales importantes son descritos a continuación: 2.4.2.6.1Temperatura Los procesos anaeróbicos son más sensibles a los cambios de temperatura que los aerobios. Algunos tienen su eficiencia más alta en condiciones mesofílicas (entre 35-40 °C) o en condiciones termofílicas (sobre 55 °C) (Khanal, 2008). 38 2.4.2.6.2 pH de operación El objetivo principal de controlar esta variable es por el efecto que puede ocasionar sobre las actividades de las enzimas y las consecuencias que tendría en la ruta enzimática del proceso. El pH considerado como óptimo es el cercano a la neutralidad. 2.4.2.6.3 Toxicidad e inhibición Los microorganismos anaerobios son inhibidos por sustratos presentes en el agua residual y por subproductos metabólicos de microorganismos (Khanal, 2008). Entre otros factores se encuentran la presencia de nutrientes y metales trazas potenciales de óxido-reducción y la configuración del reactor. 2.4.3 Balance de masa para biorreactores Los reactores pueden operar de varias formas, las más comunes son (Mansi y col., 2007): Batch o por lotes, donde F entrada=Fsalida=0; por lo que el volumen es constante. Continuo, donde Fentrada=Fsalida≠0, por lo que el caudal es constante. Fed-batch (o semibatch), donde Fentrada≠0 y Fsalida=0, por lo que el volumen se incrementa. Para comprender mejor esto es necesario conocer la dinámica del balance de masa para el sustrato, productos metabólicos, constituyentes de la biomasa y la biomasa. La base para el balance general de masa dinámico es la siguiente ecuación (Mansi y col., 2007): 𝐀𝐜𝐮𝐦𝐮𝐥𝐚𝐜𝐢ó𝐧 = 𝐞𝐧𝐭𝐫𝐚𝐝𝐚 – 𝐬𝐚𝐥𝐢𝐝𝐚 + 𝐠𝐞𝐧𝐞𝐫𝐚𝐜𝐢ó𝐧 – 𝐜𝐨𝐧𝐬𝐮𝐦𝐨 Ecuación 25 La ecuación anterior sufre modificaciones de acuerdo al tipo de reactor utilizado. 39 2.4.3.1 Reactor batch La operación de este tipo de reactor es muy utilizada ya que puede llevarse a cabo en una forma experimental relativamente simple. Los experimentos batch tienen la ventaja de que pueden operar de forma sencilla y es posible llevar a cabo un gran número de experimentos a la par. La tasa de dilución es cero para un reactor batch y el balance de masa para la biomasa y el sustrato limitante toma la siguiente forma (Mansi y col., 2007): 𝒅𝒙 = 𝝁𝒙 𝒅𝒕 𝒙 𝒕 = 𝟎 = 𝒙𝟎 Ecuación 26 Donde 𝑥 representa la concentración de la biomasa, 𝑥0 indica la concentración inicial de la biomasa, la cual es obtenida inmediatamente después de la 𝑑𝑥 incubación, 𝑑𝑡 es la variación de la concentración respecto al tiempo y 𝜇 representa el crecimiento de la biomasa. 𝒅𝑪𝑺 𝒅𝒕 = −𝒓𝒔 𝒙 𝑪𝒔 (𝒕 = 𝟎) = 𝑪𝒔,𝟎 Ecuación 27 Donde 𝐶𝑠 indica la concentración del sustrato, 𝐶𝑠,0 es la concentración inicial del sustrato, 𝑑𝐶𝑆 𝑑𝑡 es la variación de la concentración del sustrato respecto al tiempo y −𝑟𝑠 representa el consumo de sustrato. La ecuación anterior también puede representar la formación de un nuevo producto cuando se considera a 𝐶𝑆 como el producto y −𝑟𝑠 se toma sin el signo negativo. En reactores batch el sustrato puede ser consumido como: Ecuación 28 △ 𝑺 =△ 𝑺 𝒂𝒔𝒊𝒎𝒊𝒍𝒂𝒄𝒊ó𝒏 +△ 𝑺 𝒆𝒏 𝒍𝒂 𝒃𝒊𝒐𝒎𝒂𝒔𝒂 𝒂𝒔𝒊𝒎𝒊𝒍𝒂𝒄𝒊ó𝒏 𝒆𝒏 𝒖𝒏 𝒑𝒓𝒐𝒅𝒖𝒄𝒕𝒐 𝒆𝒙𝒕𝒓𝒂𝒄𝒆𝒍𝒖𝒍𝒂𝒓 +△ 𝑺 𝒆𝒏𝒆𝒓𝒈í𝒂 𝒅𝒆 +△ 𝑺 𝒄𝒓𝒆𝒄𝒊𝒎𝒊𝒆𝒏𝒕𝒐 𝒆𝒏𝒆𝒓𝒈í𝒂 𝒅𝒆 𝒎𝒂𝒏𝒕𝒆𝒏𝒊𝒎𝒊𝒆𝒏𝒕𝒐 40 Capítulo III: Materiales y métodos 3.1 Ensayos anammox 3.1.1 Inóculo Fue necesario realizar la estabilización del lodo para los ensayos anammox. El inóculo se obtuvo de sedimentos de los lagos de la USBI. De esta manera se aseguró obtener un inóculo anaerobio o facultativo, como los de interés. Después se realizaron lavados del inóculo con solución fisiológica (8.5 g NaCl/L) a fin de prevenir efectos osmóticos y daño consecuente en las células microbianas. Se mantuvo una concentración de nitrógeno constante y la relación NO2-/NH4+ fue de 1.32. La fuente donadora de electrones fue amonio a una concentración de 90-120 mgNH4 + /L, mientras que la fuente aceptora fue nitrito a una concentración de 118158 mg/L, lo correspondiente a la relación NO2 − /NH4 + de 1.32. Posteriormente, se incubó bajo condiciones anammox haciendo cambio de medio cada 8 días al inicio, reduciendo este periodo de acuerdo a las mediciones de amonio y nitrito obtenidas. Los lodos fueron lavados antes de cada ensayo. 3.1.2 Matraces anammox Los ensayos cinéticos en lote se realizaron en matraces Erlenmeyer con volúmenes de 250 mL. Cada matraz se inoculó con 5.8 ± 0.2 gSSV/L, después se adicionó medio de cultivo hasta alcanzar un volumen de trabajo de 200 mL manteniendo un espacio de cabeza de 80 mL. Los matraces fueron cerrados con tapones de algodón forrados con gasa y después con capuchones de papel aluminio. Debido a que el cierre no fue completamente hermético no se tomaron muestras del espacio de cabeza para medición de gases, entre estos el nitrógeno molecular. 34 Para desplazar el oxígeno, se utilizó una corriente de helio por 8 minutos. Los matraces fueron incubados a 30°C con una agitación de 150 rpm. Cada ensayo cinético se realizó por duplicado. 3.1.3 Medio de cultivo Para los ensayos en lote se utilizó un medio de cultivo litoautotrófico cuya composición se muestra en la siguiente tabla (mg/L): Tabla 6. Composición química del medio mineral tipo anammox. Reactivo Cantidad CaCl2∙H2O 100 Na2PO4∙H2O 57.5 MgSO4∙7H2O 200 NaHCO3 4000 Solución A de elementos traza: 1.0 mL/L (mg/L) FeSO4 5000 EDTA 5000 Solución B de elementos traza:1.0 mL/L EDTA 1500 ZnSO4∙7H2O 430 CoCl2∙6H2O 240 MnCl2 629 CuSO4∙5H2O 250 Na2MoO4∙2H2O 220 NiCl2∙6H2O 190 Na2SeO4∙10H2O 210 H3BO3 14 NaWO4∙2H2O 50 35 Además de los nutrientes anteriores, se adicionó como fuente donadora de electrones cloruro de amonio y como fuente aceptora, nitrito de sodio, siguiendo la relación estequiométrica de la ecuación. 𝑵𝑯𝟒 + + 𝟏. 𝟑𝟐𝑵𝑶𝟐 − + 𝟎. 𝟎𝟔𝟔𝑯𝑪𝑶𝟑 − + 𝟎. 𝟏𝟑𝑯+ → 𝟏. 𝟎𝟐𝑵𝟐 + 𝟎. 𝟐𝟔𝑵𝑶𝟑 − + 𝟎. 𝟎𝟔𝟔𝑪𝑯𝟐 𝑶𝟎.𝟓 𝑵𝟎.𝟏𝟓 + 𝟐. 𝟎𝟑𝑯𝟐 𝑶 Ecuación 19 3.1.4 Ensayos control Se corrieron dos ensayos control con la finalidad de observar si existía consumo de las fuentes de electrones o aceptoras adicionadas por separado. Un experimento control contenía el inóculo, medio mineral y la fuente aceptora de electrones, pero sin la fuente de electrones. En otro control, se puso el lodo anammox, medio mineral y la fuente de electrones, pero sin la fuente aceptora. 3.1.5 Ensayos en lote Se realizaron dos series de ensayos en lote. En la primera serie de experimentos se adicionó amonio a una concentración de 50 mgNH4 + /L, mientras que la fuenteaceptora fue nitrito a una concentración de 66 mgNO2 − /L.En la segunda serie de experimentos, se mantuvo una concentración de nitrito constante, similar a la de la primera serie de experimentos, 66 mgNO2 − //L. La fuente de electrones, el amonio, se agregó a una concentración de 50 mgNH4 + /L adicionando 20 mgNH4 + /L de amonio extra. Además se agregó nitrato, a una concentración de 30 mgNO3 −/L. 3.1.6 Variables de respuesta El proceso de estabilización de lodos fue evaluado a través del consumo simultáneo de nitrito y amonio, característico del sistema anammox y la constancia en las velocidades específicas de consumo de sustrato (q). 36 Los ensayos tipo anammox con y sin nitrato fueron evaluados a través del consumo simultáneo de nitrito y amonio, eficiencias de consumo de nitrito, amonio y consumo total representadas en % (ENIT, EAM y ECT, respectivamente) y velocidades específicas de consumo de amonio y nitrito (q amonio y qnitrito, respectivamente). 3.2 Ensayos desnitrificantes 3.2.1 Botellas serológicas Los ensayos cinéticos en lote se realizaron en botellas serológicas con volúmenes de 120 mL. Cada botella fue una unidad experimental independiente y después de su muestreo se descartó. El volumen de lodo utilizado correspondió a 1.7 ± 0.5 gSSV/L, después se adicionó medio de cultivo hasta alcanzar un volumen de 100 mL en un matraz volumétrico. Dicha mezcla de traspasó al frasco serológico obteniendo un volumen de trabajo de 100 mL y un espacio de cabeza de 20 mL. Las botellas serológicas, también conocidos como viales de prueba, se sometieron a corriente de gas helio durante 5 minutos. Después de dicho periodo, fueron cerrados con tapones de hule y sellados con arillos metálicos para mantener condiciones herméticas. Para desplazar el nitrógeno y el oxígeno del espacio de cabeza, se utilizó una corriente de helio por 5 minutos más utilizando dos agujas de jeringa. Una aguja se conectó al gas helio mientras que la segunda funcionó como la salida de gas del vial. Lo anterior es para promover condiciones anaeróbicas en los frascos. Al inicio de cada ensayo, se tomó una muestra gaseosa de 100 μL para la cuantificación de N 2. Las botellas fueron incubadas a 30°C con una agitación de 150 rpm. Cada ensayo cinético se realizó por duplicado. 37 3.2.2 Inóculo El inóculo se obtuvo de sedimentos de los lagos de la USBI tomando lodo de la parte más profunda del lago. Después se realizaron lavados del inóculo con solución fisiológica (8.5 g de cloruro de sodio/L) a fin de prevenir efectos osmóticos. Posteriormente, se incubó bajo condiciones desnitrificantes haciendo recambio de medio de cultivo cada 8 días. Los lodos también fueron lavados antes de cada ensayo. 3.2.3 Medio de cultivo Para los ensayos en lote se utilizó un medio de cultivo organotrófico cuya composición se muestra en la siguiente tabla (g/L): Tabla 7. Composición química del medio de cultivo para bacterias desnitrificantes. COMPUESTO CONCENTRACIÓN (g/L) KH2PO4 4 CaCl2∙H2O 0.3 NaMoO4 0.06 MgSO4 0.6 FeCl3 0.1 CuSO4∙5H2O 0.06 Se adicionó la fuente de carbono y nitrógeno siguiendo la relación estequiométrica de la ecuación. Dicha relación C/N es de 1.4. Las fuentes utilizadas para carbono y nitrógeno, fueron acetato de sodio y nitrato de sodio, respectivamente. 38 3.2.4 Ensayos control Se corrieron dos ensayos control con la finalidad de observar si existía consumo de las fuentes carbonadas o nitrogenadas adicionadas por separado. Un experimento control contenía el inóculo, medio mineral y la fuente de nitrógeno, pero sin la fuente de electrones. En otro control medio mineral y la fuente de carbono. 3.2.5 Ensayos en lote Se realizaron dos series de ensayos en lote. En la primera serie de experimentos se mantuvo una concentración de nitrógeno constante y la relación C/N de 1.4. La fuente de nitrógeno fue nitrato a una concentración de 50 mgN-NO3 −/L, mientras que la fuente de carbono fue acetato a una concentración de 70 mg/L.En la segunda serie de experimentos, se mantuvo una concentración de acetato constante, similar a la de la primera serie de experimentos, 70mg/L. La fuente de nitrógeno, el nitrato, se agregó a una concentración de 50 mgN-NO3 − /L adicionando un 30% de nitrato extra. Además se agregó una fuente alternativa de electrones, el amonio, a una concentración de 20 mgN-NH4 + /L. 3.2.6 Variables de respuesta El proceso fue evaluado a través del consumo de nitrato, característico del sistema desnitrificante, eficiencias de consumo de nitrato representadas en % (E C), rendimientos de producción de nitrógeno molecular (YP/S) y velocidades específicas de consumo de nitrato (qnitrato). 39 3.3 Métodos analíticos Tratamiento previo Las muestras fueron previamente filtradas por una membrana de nylon de 13 mm de tamaño de diámetro y tamaño de poro de 0.45 μm, posteriormente se realizaron las respectivas determinaciones. 3.3.1 Determinación de amonio La detección y cuantificación de amonio se realizó por espectrofotometría uv-vis. El método es una adaptación del ASTM Manual of Water and Environmental Technology, D1426-92, Método de Nessler. La reacción entre el amonio de las muestras y los reactivos del kit provoca una coloración amarilla en la muestra al formarse un complejo que es detectable a una longitud de onda de 420 nm. El equipo utilizado fue un espectrofotómetro de HANNA Instruments Modelo HI 83225. Para esta técnica se utilizó el kit para amonio en alto rango con las siguientes especificaciones: Rango de 0 a 100 mg/L Resolución de 1 mg/L Precisión ±1 mg/L ±4 % de lecturas a 25 °C Fuente de luz: lámpara de tungsteno a una longitud de onda de 420 nm. El equipo estaba calibrado y estandarizado por HANNA, por lo que no arrojaba datos de absorbancia sino los valores equivalentes de amonio. Para comprobar la respuesta del equipo, se analizaron en diferentes ocasiones estándares de amonio, uno equivalente a una concentración alta (50 mgNH 4+/L) y otro equivalente a una concentración baja (10 mgNH 4+/L) encontrándose que la precisión de las lecturas del equipo es aceptable. 40 3.3.2 Determinación de nitrito Este método se basó en la reducción de nitrito a óxido nitroso en medio ácido de sulfato ferroso. Los iones ferrosos se combinan con el óxido nitroso para formar un complejo de color verde/marrón. Su intensidad es proporcional al nitrito presente en la muestra de agua siguiendo la ley de Beer. La reacción que ocurre es la siguiente (HACH, 2014): 𝟐𝑭𝒆+𝟐 + 𝟒𝑯+ + 𝟐𝑵𝑶𝟐 − → 𝟐𝑭𝒆+𝟑 + 𝟐𝑵𝑶 + 𝟐𝑯𝟐 𝑶 Ecuación 29 𝑵𝑶 + 𝑭𝒆𝑺𝑶𝟒 → 𝑭𝒆𝑺𝑶𝟒 • 𝑵𝑶 Ecuación 30 El equipo utilizado fue un espectrofotómetro de HANNA Instruments Modelo HI 708. Para esta técnica se utilizó el kit para nitrito en alto rango con las siguientes especificaciones: Rango de 0 a 150 mg/L Resolución de 1 mg/L Precisión ±3 mg/L ±5 % de lecturas Fuente de luz: fotodiodo a una longitud de onda de 575 nm. El equipo estaba calibrado y estandarizado por HANNA, por lo que no arrojaba datos de absorbancia sino los valores equivalentes de nitrito. Para comprobar la respuesta del equipo, se analizaron en diferentes ocasiones estándares de amonio, uno equivalente a una concentración alta (100 mgNO2-/L) y otro equivalente a una concentración baja (10 mgNO 2-/L) encontrándose que la precisión de las lecturas del equipo era aceptable. 3.3.3 Determinación de nitrato La cuantificación del nitrato se realizó empleando el electrodo de ion selectivo perfectION™ comb NO3CombinationElectrode de METTLER TOLEDO. Este 41 electrodo combinado, cuenta con una membrana que responde selectivamente al ion antes mencionado, donde al entrar en contacto con la muestra, ocurre un intercambio iónico. La diferencia de potencial eléctrico en la membrana, se mide por electrodos de referencia, las variaciones de concentración propician esta diferencia de voltaje con la que se cuantifica este ion (Skoog, 2002). Se realizó una curva estándar para la cuantificación de nitrato, preparando cinco puntos o concentraciones de 1,10, 40, 70 y 100 ppm. 3.3.4 Determinación de nitrógeno molecular Esta determinación se realizó por cromatografía de gases. Para el análisis se empleó un cromatógrafo de gases de la marca Perkin Elmer® con un detector de conductividad térmica. Se utilizó una columna capilar Carboxen™ Plot 1006, de 30 m x 0.32 mm. El gas de arrastre fue helio de pureza al 99.99% a un flujo de 20mL/min. Las temperaturas programadas fueron: horno, 35°C; detector, 230°C y puerto de inyección, 230°C. La muestra del gas producido en el reactor se obtuvo de la parte superior del frasco serológico, mediante una jeringa para gases de 100 μL. El volumen de inyección fue de 100 μL, con Split 5:1. El volumen de gas generado se calculó considerando la presión atmosférica de la ciudad de Xalapa y la temperatura del sistema. Se calculó la producción volumétrica de N 2, el número de moles (mol N2/L) y por último se convirtieron a miligramos (mgN2/L). Se realizó una curva de calibración con concentraciones de 0, 1, 5, 20, 50 y 100% de nitrógeno. 3.3.5 Determinación de sólidos suspendidos volátiles. Se realizó la medición de sólidos suspendidos volátiles (SSV) como forma de lectura de biomasa, siguiendo el método estándar establecido por APHA (1985). Para la determinación la muestra es secada y evaporada a 110°C durante 24 horas, posteriormente es pesada. Para cuantificar los sólidos suspendidos fijos, la 42 muestra es calcinada durante una hora a 550°C. La cantidad de SSV presentes en la muestra es obtenida por diferencia de peso entre los sólidos suspendidos totales y los sólidos suspendidos fijos y es considerada como la biomasa presente en el lodo. 3.4 Análisis molecular 3.4.1 Extracción de ácidos nucléicos La extracción de ácidos nucléicos se realizó empleando un kit comercial, el kit Powersoil® DNA Isolation Kit, siguiendo las instrucciones del fabricante. El tamaño del DNA se analizó por electroforesis en gel de agarosa (1%) que contenía 1 μL de bromuro de etidio (10 mg/mL) a 90 voltios (V) por 1 hora y se visualizó en un transiluminador de UV. Posteriormente, fue medida su concentración inyectando 2 μL en un nanodrop. 3.4.2 Amplificación de fragmentos de interés por PCR Se utilizó la reacción en cadena de la polimerasa (PCR) ya que esta técnica permite obtener en gran cantidad un fragmento específico de DNA (CastilloRodríguez y col., 2005). Para esta técnica se utilizaron los primers específicos para bacterias. La región V6-V8 de genes bacterianos del rRNA 16S fue amplificada usando los iniciadores bacterianos 968-f(5‟-GAACGCGAAGAACCTTACC-3‟) con grapa y 1401-r(5‟CGGTGTGTACAAGACCC-3‟). El mix de reacción de PCR se realizó con 1 µL de cada primer, 200 ng/µL de ADN, 10 µL de GoTaq (Buffer + Mg + Taqpolimerasa + contaminante) y agua estéril hasta obtener un volumen final de 20 µL. 43 La amplificación se llevó a cabo en un termocicladorBio-Rad® por medio del siguiente programa: predesnaturalización a 94°C por 2 min., 30 ciclos con una desnaturalización a 94°C por 45 s, una hibridación a 60°C por 30 s y una elongación a 72°C por 90 s., y una post-elongación a 72°C por 10 min. El tamaño del fragmento se analizó por electroforesis en gel de agarosa (1%) que contenía 1 μL de bromuro de etidio (10 mg/mL) a 90 voltios (V) por 1 hora y se visualizó en un transiluminador de UV. 3.4.3 Método de electroforesis en gel con gradiente desnaturalizante (DGGE) Los productos de amplificación fueron separados por DGGE en un equipo DCode™ Universal Mutation Detection Systems (Bio-RAD Laboratories, Hercules, USA). La electroforesis se corrió a 60°C, primero a un voltaje de 200 V por 5 minutos y después a 60 V durante 960 minutos, utilizando un gel de poliacrilamida y un gradiente desnaturalizante de 40 a 54%. Los geles de DGGE fueron teñidos con bromuro (10 mg/L) y escaneados mediante un fotodocumentador. 44 Capítulo IV: Resultados y discusión 4.1 Ensayos tipo anammox 4.1.2 Obtención del inóculo estabilizado que consume nitrito y amonio para las cinéticas en lote 4.1.2.1 Consumo de amonio El reactor batch bajo condiciones anammox presentó los perfiles mostrados en la figura 3. Puede apreciarse que durante los primeros días, el tiempo necesario para degradar el amonio era de al menos 5 días. Posteriormente, el tiempo de consumo se redujo en 4 días, después a 2 días y finalmente, a partir del día 40 de monitoreo, se observó que el amonio era completamente consumido de un día para otro. Concentración (mg/L N-Amonio 80 60 40 20 0 0 10 20 30 40 50 Días Figura 3. Perfil de degradación de amonio por un consorcio estabilizado tipo anammox (Cada pico superior representa una nueva alimentación). Kuenen (2008) muestra una gráfica de consumo de amonio de un reactor piloto conteniendo amonio, sulfuro, compuestos orgánicos no especificados y nitrato. La concentración de inicial de amonio fue de 80-120 mgNH4 + /L. Durante los primeros 60-80 días de monitoreo no se observa consumo de amonio mientras que después 45 comenzó a observarse su degradación (concentración final entre 80-100 mgNH4 + /L). Conforme transcurrió el tiempo, la concentración de amonio de salida fue cercana a cero. Estos datos son compatibles con los obtenidos en esta gráfica, ya que se observa la misma tendencia de disminución de amonio. 4.1.2.2 Consumo de nitrito En la figura 4 puede apreciarse el consumo de N − NO2 − . Estas mediciones, corresponden a los últimos días de los 51 evaluados en total. Puede observarse una estabilización en el consumo de nitrito, similar al consumo de amonio mostrada anteriormente. Los resultados encontrados del consumo simultáneo de amonio y nitrito, muestran características típicas de un proceso anammox. Concentración (mg/L) 70 60 50 40 30 N-Nitrito 20 10 0 0 5 10 15 Días Figura 4. Perfil de degradación de nitrito por un consorcio estabilizado para el proceso tipo anammox. (Cada pico superior representa una nueva alimentación). Kuenen (2008) menciona que en el proceso anammox las bacterias utilizan nitrito como aceptor de electrones para la oxidación de amonio y como donador de electrones para la reducción de dióxido de carbono. Esta puede ser una explicación del consumo de nitrito representada en la figura 4. 46 4.1.2.3 Velocidades específicas de consumo En la Figura 5, se aprecian las velocidades específicas de consumo de amonio (mg N-NH4 + /gSSV∙d) durante 51 días de monitoreo, obtenidas durante los ensayos de estabilización de consumo de amonio. Se puede observar que comienza con una q menor a 1, este valor va incrementándose conforme pasa el tiempo hasta permanecer constante entre valores que van de 1.85 a 2.05, estos resultados dieron muestra una constancia en las velocidades de consumo de este sustrato. Velocidad específica de consumo 2.5 2 1.5 1 mgN-Amonio/SSV*D 0.5 0 0 -0.5 5 10 15 20 25 30 35 40 45 50 Días de monitoreo Figura 5. Velocidades específicas de consumo de amonio por un consorcio tipo anammox. Bettazzi et al. (2010) estudiaron la reactivación de un reactor anammox utilizando un reactor SBR (reactor secuencial por lotes, por sus siglas en inglés) de 40 L enriquecido con 30 L de lodo anammox, 35 °C, pH entre 7.5 y 7.8, afluente: 319.3 ± 51.4 mgN-NH4 + /L y 310.8 ± 58.5 mgN-NO2 − /L, relación N-NO2 − /N-NH4 + =0.98 ± 0.12. Entre sus resultados muestran que cuando la velocidad específica de consumo de amonio es constante, el reactor trabaja de forma estable, esto se asemeja a los últimos días de monitoreo observados en la figura 5. Mientras que 47 en los primeros días de rehabilitación la velocidad de consumo no era constante, tal y como sucede en este trabajo durante los primeros 38 días. El lodo microbiano generado bajo estas condiciones, fue utilizado para inocular los cultivos en lote para los ensayos tipo anammox posteriores. 4.1.2 Ensayo sin nitrato La figura 6, ilustra el ensayo anammox, donde se agregó 22.52 ± 0.43mg NNO2 − /L como fuente aceptora de electrones y 35 ± 1.09 mg N-NH4 + /L como fuente donadora de electrones. Adicionalmente, se le agregó carbono inorgánico en forma de bicarbonato, 4 g NaHCO3/L, es decir, el medio fue enriquecido en condiciones litoautotróficas. En los resultados, se encontró que el consorcio llevó a cabo el consumo completo de los substratos nitrogenados en un periodo de 25 horas, teniendo una concentración final de 0 ± 0 mg N-NO2 − /L y 3.11 ± 1.09 mg N-NH4 + /L. Figura 6. Degradación de amonio y nitrito por un consorcio microbiano anaerobio tipo anammox. 48 La figura 6 concuerda con los resultados obtenidos por Martínez-Hernández et al. (2013), quienes estudiaron la inhibición de la actividad anammox por toluenonitrito. En sus experimentos batch de 160 mL agregaron 1.25 gSSV/L, 100 mL de medio mineral, 2.7 mM de amonio, 3.57 mM de nitrito y diferentes concentraciones de tolueno que fueron de 0.1 a 3 mM. En sus resultados se observa el consumo simultáneo de nitrito y amonio en los ensayos control, presentando una tendencia similar observada en la figura 6. La tabla 8 muestra el balance general de la cinética anammox. En ella pueden observarse las concentraciones iniciales de N-NO2 − y N-NH4 + (22.52± 0.43 y 35± 1.09, respectivamente), según la relación estequiométrica NO2 − /NH4 + =1.32 (ecuación 21). Se observan también las concentraciones finales de N-NO2 − y NNH4 + (0 ± 0 y 3.11 ± 1.09, respectivamente) y las eficiencias de consumo. Al final de las 25 horas se lograron eficiencias de consumo de nitrito de 100 %, de amonio de 88.64 % y totales de ECT= 94.59 %. Estos datos indican que el sustrato limitante fue el nitrito pues al término de la cinética fue totalmente consumido. Tabla 8. Balance general de materiales y eficiencias de consumo de un consorcio microbiano anaerobio, tipo anammox, durante la degradación de amonio y nitrito. Tiempo Concentración Concentración ENIT EAM ECT (horas) inicial (mg/L) final (mg/L) (%) (%) (%) 100 88.64 94.5 Inicial 0 Final N-𝐍𝐎𝟐 − N-𝐍𝐇𝟒 + N-𝐍𝐎𝟐 − N-𝐍𝐇𝟒 + 25 22.52 ± 35± 0± 3.11 ± 0.43 1.09 0 1.09 9 ENIT; eficiencia de consumo de nitrito, EAM; eficiencia de consumo de amonio y ECT; eficiencia de consumo total (nitrito y amonio). Es importante mencionar que de acuerdo a la estequiometria de proceso, el producto principal es el nitrógeno molecular; sin embargo de acuerdo a las condiciones bajo las cuales se corrieron estos experimentos no fue posible medir este compuesto. No obstante, por las condiciones y composición química del 49 medio de cultivo hacen pensar que el proceso biológico predominante fue el anammox. Dapena-Mora et al. (2004), estudiaron la estabilidad del proceso Anammox en un reactor gas-lift. El reactor fue monitoreado por 200 días y se alimentó con flujo másico de 2 g/L∙d, 0.9 gN-NH4 + /L, 1.1 gN-NO2 − /L, TRH= 1 día, pH controlado en 8 ± 0.1, 30°C. Entre los resultados obtenidos se encontró una eficiencia de remoción de nitrógeno de 88% y nitrito completamente consumido. Esos resultados concuerdan con los obtenidos en estas cinéticas pues la remoción fue de 88.64 % para el amonio y 100 % para el nitrito. Los resultados indican que el lodo mostró una actividad tipo anammox, donde las condiciones experimentales resultaron favorables para degradar amonio y nitrito a concentraciones estequiométricas. 4.1.3 Ensayo con nitrato La Figura 7, ilustra el ensayo anammox, donde se agregó 23.74 ± 1.29 mg NNO2 − /L como fuente aceptora de electrones, 36.94 ± 1.65 mg N-NH4 + /L como fuente donadora de electrones y 30 mgNO3 − /L. Este ensayo fue llevado a cabo para evaluar la actividad anammox del inóculo bajo la presencia de otro compuesto nitrogenado como el nitrato. En los resultados se encontró que el lodo llevó a cabo el consumo completo de amonio y nitrito a las de 25 horas de iniciada la experimentación, obteniendo concentraciones finales de 0 ± 0 mg NNO2 − /L y 1.56 ± 1.09 mg N-NH4 + /L. 50 Figura 7. Conducta metabólica de un consorcio microbiano anaerobio, tipo anammox, en presencia de amonio, nitrito y nitrato. Kartal et al. (2006) investigaron la posibilidad de la reducción desasimilatoria de nitrato a amonio por la bacteria anammox. Utilizaron células en suspensión con 80-90 % de K. stuttgartiensis (bacteria anammox) a 6.6 ± 0.4x109, incubadas con 15 NO3 − como aceptor y 13 C-formato como donador de electrones, y se obtuvieron 15 N15N y 13CO2 como productos. La degradación comenzó después de una fase de 2 horas y el principal producto de la reducción de nitrato fue el nitrito. Este proceso se llevó a cabo a una velocidad de 0.3 ± 0.02 mol/cel∙d. Se estudiaron los posibles intermediarios de 15 N-marcado con células en suspensión de K. stuttgartiensis. El cultivo se incubó con formato y 15 NO3 − en presencia de uno de los compuestos no marcados, NO2 − y NH4 + (2mM y 10 o 2 mM, respectivamente). En presencia de nitrito la mayor parte de que en presencia de amonio, el 15 15 NO3- se convirtió en nitrito, mientras N se convirtió parcialmente a NH4 + (2-10 %). 15 Estos datos indican que con una fuente orgánica de carbono, como el formato, la reducción de nitrato por bacterias anammox en presencia de nitrito produce 51 principalmente nitrito. En la fig. 7 se observa que en las primeras horas se mantiene constante la concentración de nitrito mientras que la concentración de amonio sí disminuye, esta información puede ser comparada con los datos obtenidos por Kartal et al. (2006) sugiriendo que aunque la fuente de carbono no es orgánica es posible el consumo de nitrato. La tabla 9 muestra el balance general de la cinética anammox. En ella pueden observarse las concentraciones iniciales de N-NO2 − y N-NH4 + (23.74± 1.29 y 36.94± 1.65, respectivamente), las concentraciones finales de N-NO2 − y N-NH4 + (0 ± 0 y 1.56 ± 1.09, respectivamente) y las eficiencias de consumo. Es conveniente indicar que debido a problemas técnicos, no fue posible medir nitrato. Al final de las 25 horas logró consumirse prácticamente en su totalidad el nitrito y el amonio, obteniendo eficiencias de nitrito de 100 %, de amonio de 95.79% y totales de 98.19 %. Tabla 9. Balance general de materiales y eficiencias de consumo de un consorcio microbiano anaerobio, tipo anammox, en presencia de amonio, nitrito y nitrato. Tiempo Concentración Concentración ENIT EAM ECT (horas) inicial (mg/L) final (mg/L) (%) (%) (%) 100 95.79 98.19 Inicial 0 Final 25 N-𝐍𝐎𝟐 − 23.74 ± 1.29 N-𝐍𝐇𝟒 + 36.94 ± 1.65 N-𝐍𝐎𝟐 − 0± 0 N-𝐍𝐇𝟒 + 1.56 ± 1.09 ENIT; eficiencia de consumo de nitrito, EAM; eficiencia de consumo de amonio y ECT; eficiencia de consumo total (nitrito y amonio). Molinuevo et al. (2009) encontraron eficiencias de consumo de amonio similares en pruebas con lodos anaerobios y anammox utilizados para tratamiento de agua residual. En sus pruebas utilizaron un reactor UASB operado de forma semicontinua, 40 mL de lodo anaerobio granular, 40 mL de lodo anammox, 37 °C y un flujo de 120 mL/d. El reactor fue alimentado con agua sintética similar a la utilizada en estos experimentos, con 0.26 gNaNO 2/L, 0.19 g(NH4)2SO4/L y 1.22 52 gNaNO3/L, más el efluente de un digestor de oxidación parcial de agua residual de estiércol de cerdo, ésta última a concentraciones de 2 y 3% (V/V). Encontró eficiencias de consumo de 92 ± 4.9% para 2% y 80 ± 7.8% para 3%, valores menores a los 95.79 % obtenidos en este experimento. El consumo de estos substratos indica la prevalencia de un consorcio tipo anammox a pesar de la presencia de nitrato. Pudo observarse que las eficiencias de amonio y nitrito fueron similares en presencia y ausencia de nitrato, indicando un nulo efecto de este último sustrato sobre el metabolismo de estos micoorganismos. 4.1.4 Comparación de consorcios anaerobios tipo anammox con y sin nitrato La figura 8 ilustra una comparación del perfil de consumo de nitrito y amonio en los ensayos anammox en presencia y ausencia de nitrato. De acuerdo a la gráfica, se observa un perfil metabólico similar de los sustratos en ambos experimentos durante el periodo de 25 horas. Bajo estas condiciones, se obtuvieron datos de velocidades específicas de consumo, las cuales se indican a continuación. 53 Figura 8. Conducta metabólica de un consorcio microbiano anaerobio, tipo anammox, durante la degradación de amonio y nitrito en presencia y ausencia de nitrato. En la tabla 10, se puede observar que la cinética sin nitrato (N-NO2 − + N- NH4 + + HCO3 − ) y prueba con nitrato (N-NO2 − + N- NH4 + + N-NO3 − + HCO3 −) tuvieron una velocidad específica de consumo parecida, tanto para nitrito (qNIT) como para amonio (qAM). Puesto que en la primera, qNIT, tuvo un valor de 0.003698 mgNO2 −/mgSSV∙d mientras que en la segunda, resultó ser de 0.003898 mgNO2 −/mgSSV∙d. Respecto a qAM, en el primer ensayo se obtuvo una velocidad específica de consumo de amonio de 0.00523641 y en el segundo fue de 0.00581113. 54 Tabla 10. Velocidades específicas de consumo de un consorcio microbiano anaerobio, tipo anammox, durante la degradación de amonio y nitrito en presencia y ausencia de nitrato. Ensayo qNIT qAM − (mg𝐍𝐎𝟐 / (mg𝐍𝐇𝟒 + / mgSSV∙d) mgSSV∙d) Sin nitrato 0.003698 0.00523641 Con nitrato 0.003898 0.00581113 Langone et al. (2014) estudiaron en reactores batch el efecto de pH en la actividad anammox. Realizaron pruebas de pH con variaciones entre 6.5 y 9, 4 mM de NNH4 + y concentraciones de 1.5 mM de N-NO2 − . Encontraron pH óptimos entre 7.8 y 8, y una velocidad específica de consumo de amonio 19.68 mgNNH4 + /mgSST∙d, velocidad mucho más elevada que los 0.0052 mgN- NH4 + /mgSSV∙d obtenidos en este trabajo. Aunado, se observa diferencia en una unidad, lo que eleva más este valor. La diferencia de velocidades puede deberse a muchos factores, uno de ellos, el pH. En estos experimentos se mantuvo entre 8.6 y 8.9; más elevado que los valores recomendados por otros autores (Kuenen, 2008). Langone et al. (2014) evaluó la desnitrificación vía nitrito realizando pruebas sin nitrito y con nitrito. En las pruebas sin nitrito utilizó 200 mgN-NO3 −/L y 2000 mgCOD/L como acetato de sodio, pH= 7.5 y 10 mL de biomasa, responsable de procesos de nitrifitrifación-anammox; mientras que los experimentos con nitrito contenían 200 mgN-NO2 − /L, 2000 mgCOD/L como acetato de sodio y las mismas características de pH y biomasa. Se obtuvieron velocidades específicas de consumo de 0.15 mgNO3 − /mgSST∙d y 83.28 mgNO2 − /mgSST∙d, respectivamente. En la prueba con nitrato hubo una acumulación de nitrito las primeras 5 horas, hasta su completo consumo después de 24 horas. Aunque en los ensayos de este 55 trabajo no puede observarse el consumo de nitrato, el consumo simultáneo de nitrito y amonio arrojan una posible reducción de nitrato a nitrito y su consumo. Esto debido a que no se observa amonio residual y el nitrito presente era menor al necesario para el consumo de amonio. No obstante, debe señalarse que aunque las condiciones utilizadas por Langone son muy parecidas, la fuente de carbono es orgánica y esa es una diferencia importante. 4.2 Ensayos desnitrificantes 4.2.1 Ensayos control Antes de establecer los ensayos desnitrificantes, se corrieron ensayos control con la finalidad de evaluar alguna conducta ajena a factores biológicos. Estos resultados, se muestran a continuación. En la figura 9 se observa el perfil fisiológico de un consorcio microbiano en presencia únicamente de nitrato, es decir, sin el acetato como fuente de carbono. Se observa que no existe variación importante en las concentraciones de nitrato durante un periodo de 18 horas. Los datos indican que el consorcio no fue capaz de reducir o consumir el nitrato en ausencia de un donador de electrones. Una conducta similar (datos no mostrados) fue observada en un ensayo bajo las mismas condiciones pero sin el consorcio microbiano, indicando que el nitrato no fue eliminado por algún factor abiótico, como por ejemplo la reducción química. Esto indica que no existe eliminación de nitrato por oxidación, reducción, volatilización, etc. 56 Figura 9. Conducta metabólica de un consorcio desnitrificante en presencia únicamente de nitrato. 4.2.2 Ensayos desnitrificantes sin amonio La figura 10 ilustra el ensayo desnitrificante en presencia de nitrato (60.74 ± 4.15 mg N-NO3 − /L) y acetato (85 mg CH3COONa/L) como fuente de carbono, estas concentraciones fueron basados en la estequiometría del proceso con una relación C/N de 1.4. Se observó que el lodo desnitrificante llevó a cabo el consumo completo de nitrato después de 16 horas, teniendo una concentración final de 5.97 ± 1.2 mg NNO3 − /L en cuanto a la producción de nitrógeno se registraron valores de 33.38 ± 1.03 mg N-N2/L. El acetato, adicionado como fuente orgánica, no fue medido en este trabajo. 57 Figura 10. Degradación anaerobia de nitrato a nitrógeno gaseoso por un consorcio desnitrificante. La tabla 11 muestra el balance general de la cinética desnitrificante. En ella pueden observarse las concentraciones iniciales de N-NO3 − y N-N2 (60.74 ± 4.15 y 1.25 ± 0.13, respectivamente), las concentraciones finales de N-NO3 − y N-N2 (5.97 ± 1.2 y 33.38 ± 1.03, respectivamente) y la eficiencia de consumo de nitrato (EC), cuyo valor calculado fue de 90.16 %. Observando esta eficiencia de consumo y el balance de materiales, se aprecia que la remoción de nitrato fue alta. Sin embargo, los rendimientos de N 2 fueron de 0.60, lo que indica que no todo el nitrato consumido se fue a N 2, canalizándose a otra ruta que pudo haber sido a la producción de biomasa, necesaria para el crecimiento microbiano. Al respecto, Schuler et al. (2002) señalan algunos posibles usos, como asimilación por la biomasa, asimilación como un producto extracelular, energía de crecimiento o mantenimiento. 58 Tabla 11. Balance general de materiales respecto al tiempo, eficiencia de consumo de nitrato y rendimiento de producción. Concentración Concentración final inicial (mg/L) (mg/L) en t=0 en t=16 N-𝐍𝐎𝟑 − N-N2 N-𝐍𝐎𝟑 − N-N2 60.74± 1.25± 5.97 ± 33.38 ± 4.15 0.13 1.2 1.03 EC (%) YP/S 90.16 0.60 El inóculo utilizado en este experimento había sido anteriormente estudiado. Ramírez-Sánchez (2013) analizó el efecto de la temperatura en el proceso desnitrificante utilizando reactores batch del mismo volumen que estos experimentos, 1 gSSV/L, 46.19-59.39 mg N-NO3 − /L, temperaturas de 20, 30 y 40 °C y 150 rpm. Ramírez-Sánchez obtuvo eficiencias de consumo de nitrato de 99 % en todos sus tratamientos y rendimientos de producción (YP/S) de 0.65, 0.86 y 0.84 a 20, 30 y 40 °C, respectivamente. Siendo el tratamiento más parecido al observado aquí, el de 20 °C, puesto que el rendimiento de producción no es tan alto. Téllez-Pérez (2013) estudio la flexibilidad de un consorcio microbiano para desnitrificar, metanizar y reducir sulfato en reactores por lotes. Para el tratamiento desnitrificante utilizó 50 mgN-NO3 − /L y 100 mgC-Acetato/L, 200 rpm y pH=6.9. Obtuvo eficiencias de consumo de nitrato de 59.7 ± 1.2 % y rendimientos de producción de 0.61 ± 0.01. El rendimiento de producción es casi idéntico al obtenido en este trabajo (0.6) mientras que la eficiencia de consumo de nitrato es menor, pues en los ensayos desnitrificantes reportados aquí, es mayor (90.16). 59 4.2.3 Ensayos desnitrificantes con amonio La figura 11 ilustra el ensayo desnitrificante donde se agregaron 77.26 ± 3.78mg N-NO3 − /L, 85 mg CH3COONa/L como fuente orgánica de electrones y 27 ± 0 mg N-NH4 + /L como fuente inorgánica de electrones. En los resultados se encontró el consumo parcial de nitrato y la generación de nitrógeno molecular. Sin embargo, bajo estas condiciones el amonio permaneció prácticamente constante a lo largo de la experimentación. Figura 11. Degradación anaerobia de nitrato a nitrógeno gaseoso por un consorcio microbiano desnitrificante en presencia de nitrato y amonio. Bajo estas condiciones se realizó el balance general de materiales, cuyos resultados se muestran en la Tabla 12. Pueden observarse las concentraciones iniciales de N-NO3 − , N-NH4 + y N-N2 (77.26± 3.78, 27± 0 y 0 ± 0.22, 60 respectivamente), las concentraciones finales de N-NO3 − , N-NH4 + y N-N2 (23.59 ± 0.15, 27 ± 1.41y 38.06 ± 2.42, respectivamente), la eficiencia de consumo de nitrato de 69.46 % y rendimientos de producción de nitrógeno molecular de 0.70.De acuerdo a estos datos, se encontró que no todo el nitrato fue consumido y que de éste sustrato consumido, solo el 70% de fue a la generación de nitrógeno molecular. Es posible que el resto de nitrato consumido se haya dirigido hacia la formación de biomasa. Estos resultados indican que a pesar que el amonio es una fuente también nitrogenada, el consorcio microbiano no fue capaz de consumir este sustrato. Esta conducta podría ser atribuida a diversos factores, entre estos la falta de adaptación previa a este sustrato, como lo fue para el nitrato, y con ello la falta de síntesis de las enzimas necesarias para la degradación del amonio. Otros factores podrían ser las condiciones de cultivo. Éstas estaban adecuadas sólo para el desarrollo de microorganismos desnitrificantes y no para microorganismos consumidores de amonio. Tabla 12. Balance de materiales respecto al tiempo, eficiencias de consumo y rendimiento de producción, en la degradación anaerobia de nitrato a nitrógeno molecular. Concentración inicial Concentración final EC (mg/L) (mg/L) en t=32 (%) YP/S en t=0 N-NO3 − N-NH4 + N- N-NO3 − N-NH4 + N-N2 3.78 27±0 N-NO3 − NH4 + N2 77.26 ± N- 0± 23.59 ± 27± 38.0± 0.22 0.15 1.41 2.42 0 69.46 0.7 61 4.2.4 Comparación de ensayos desnitrificantes con y sin amonio 4.2.4.1 Perfil de consumo de amonio en ensayos desnitrificantes con y sin amonio La figura 12 ilustra los dos ensayos desnitrificantes realizados, uno de ellos en presencia de amonio. Se puede observar que en ausencia de amonio, el nitrato fue consumido prácticamente en su totalidad en un tiempo de 16 horas. No obstante, en presencia de amonio, el nitrato fue consumido parcialmente en un 70% durante un periodo de 32 horas de experimentación. Figura 12. Degradación anaerobia de nitrato por un consorcio desnitrificante en presencia y ausencia de amonio. Bajo estas condiciones se obtuvieron las velocidades específicas de consumo de nitrato, las cuales son mostradas en la Tabla 13. 62 Tabla 13. Velocidad específica de consumo de un consorcio desnitrificante durante la degradación de nitrato en presencia y ausencia de amonio. Ensayo qNIT (mg𝐍𝐎𝟑 −/mgSSV∙d) Sin amonio 0.038 Con amonio 0.028 En la tabla 13se puede observar que la cinética sin amonio (nitrato + acetato) tuvo una mayor velocidad específica de consumo (qNIT) que la prueba con amonio (nitrato + acetato + amonio). Puesto que en la primera, qNO3-, tuvo un valor de 0.038 mgNO3 −/mgSSV∙d mientras que en la segunda, resultó ser de 0.028 mgNO3 −/mgSSV∙d. Bajo esta condiciones se realizó un análisis estadístico de comparación de tratamiento mediante una prueba de t, encontrando diferencias significativas con un valor de α de 0.0106.En cuanto a la eficiencia de consumo, el ensayo sin amonio tuvo una mayor eficiencia de consumo (EC=90.16 %) en comparación con el ensayo con amonio (EC= 69.45 %), bajo estas condiciones también se encontraron diferencias significativas de consumo entre los dos tratamientos, con un valor de α = 0.0027. 4.2.4.2 Perfil de producción de nitrógeno gaseoso en los ensayos desnitrificantes La figura 13 ilustra la generación de nitrógeno molecular en los dos ensayos desnitrificantes, uno de ellos en presencia de amonio. Se puede observar una cinética de producción similar en ambos experimentos hasta las 16 horas, bajo este tiempo las velocidades de producción de ambos ensayos no resultaron con diferencias estadísticas significativas. 63 Figura 13. Conducta metabólica de un consorcio desnitrificante durante la formación de nitrógeno molecular en presencia y ausencia de amonio. En la tabla 14 se puede observar que la cinética sin amonio (que contiene nitrato y acetato) tuvo una velocidad específica de producción (qN-gas) de 0.03807834 mgN2/mgSSV∙d mientras que en la segunda (que contiene nitrato, amonio y acetato), resultó ser de 0.037 mgN2/mgSSV∙d, puede notarse que ambas velocidades de consumo fueron prácticamente similares. 64 Tabla 14. Velocidades específicas de producción de un consorcio desnitrificante durante la degradación o eliminación de nitrato en presencia y ausencia de amonio. Ensayo qN-gas (mgN2/mgSSV∙d) Sin amonio 0.038 Con amonio 0.037 Ramírez-Sánchez (2013) obtuvo velocidades específicas de producción de nitrógeno gaseoso de 0.0576 mgNO3 − /mgSSV∙d en sus cinéticas a 30 °C descritas anteriormente. Estos valores experimentales están bajo condiciones similares a las de estos ensayos por lo que se puede deducir que aunque no haya diferencia entre los ensayos con o sin amonio, la velocidad de producción de nitrógeno molecular en ambos es menor comparada con otros ensayos. 4.2.5 Análisis molecular Un aspecto importante relacionado con el control de procesos biológicos en el tratamiento de aguas residuales es el conocimiento de la diversidad microbiana involucrada de la degradación de los contaminantes. En este sentido, se realizaron algunos ensayos, apoyados con técnicas de biología molecular, con el fin de conocer la dinámica de las poblaciones microbianas involucradas en los ensayos desnitrificantes. Estos trabajos se basaron principalmente en la extracción de DNA, amplificación de fragmentos del gen 16S DNAr usando primers o cebadores universales de bacterias y el corrimiento de la técnica de Electroforesis en Gel con Gradiente Desnaturalizante (DGGE), cuyo fundamento se basa en que cada banda detectada en el gel, corresponde a una población bacteriana, dado que esta técnica es capaz de diferenciar cambios hasta en un par de bases nucleotídicas 65 en los fragmentos de DNA sometidos a esta técnica. A continuación se muestran los resultados. 4.2.5.1 Extracción de DNA La extracción de DNA se realizó usando un kit comercial (PowerSoil® DNA Isolation Mo-Bio). Se tomaron muestras de lodos de los ensayos desnitrificantes a tiempo iniciales y finales. Las muestras analizadas, así como su etiquetado para identificación se muestran en la Tabla 15. Tabla 15. Numeración y contenido de las muestras de consorcios anaerobios, usadas para biología molecular. Número de muestra Ensayo 1 Control, fuente de inóculo 2 desnitrificante, tiempo inicial (TI) 3 desnitrificante, Tiempo final (TF) 4 desnitrificante con amonio, TI 5 desnitrificante con amonio , TF M Marcador Una vez realizada la extracción de DNA, los productos obtenidos fueron sometidos en electroforesis en gel al 2 y 4% de agarosa teñido con 1 μL bromuro de etidio (10 mg/mL), y después visualizados en un transiluminador de UV. En la figura 14 se visualiza el DNA extraído de la muestras, de izquierda a derecha se observa el DNA de los ensayos y en el carril extremo derecho (M) un marcador de DNA. El marcador utilizado es de 100 pares de bases, Gene 66 RulerTM100 bp DNA Ladder. En general se puede observar una buena extracción de DNA pues todas las muestras presentan bandas anchas, un mayor tamaño que el marcador y son de un color intenso (color rosa). 1 2 3 4 5 M Figura 14. Visualización de la extracción de DNA de consorcios microbianos. Una vez que se realizó la visualización del DNA, se analizó su concentración mediante el equipo de nanodrop. Este equipo mide la absorbancia del DNA y moléculas contaminantes que determinan la calidad y pureza, obteniendo resultados que se muestran en la Tabla 16. 67 Tabla 16. Cuantificación de DNA extraído y calidad obtenida. Muestra Contenido CAN A260 A280 260/280 260/230 152.8 3.055 1.615 1.89 1.7 98.2 1.965 1.036 1.9 1.92 93.1 1.862 0.98 1.9 1.97 65.3 1.306 0.681 1.92 2 102.8 2.056 1.096 1.88 2.15 (ng/µL) 1 Control fuente de inóculo 2 Cinética desnitrificante sin amonio, t=inicial 3 Cinética desnitrificante sin amonio, t=final 4 Cinética desnitrificante con amonio, t=inicial 5 Cinética desnitrificante con amonio, t=final En la tabla 16 se presentan absorbancias a 260 nm y a 280 nm, en las que los ácidos nucléicos y las proteínas tienen una absorbancia máxima, respectivamente. La relación A260/ A280 permite determinar si el DNA está contaminado por la presencia de compuestos aromáticos. Una buena calidad del DNA muestra una relación mayor a 1.8; si el valor es mayor que 2.1 tiene considerable presencia de RNA; mientas que menor que 1.6 contaminación por proteínas o fenoles. La relación A260/A230 es una medida adicional para determinar la pureza del DNA. A una longitud de onda de 230 nm las sales y carbohidratos presentan una absorbancia máxima. Cuando la relación está en el rango de 1.5 – 2.2 el DNA es puro, mientras que menor a ese rango la muestra está contaminada. 68 Con esta información se comprueba la calidad del DNA extraído. Puesto que todas las muestras presentan un cociente 260/280 mayor que 1.8 y menor que 2.1, y un cociente 260/230 entre 1.7 y 2.15. 4.2.5.2 Amplificación de fragmentos de DNA por PCR Para la amplificación de los fragmentos de DNA se usó la técnica de la reacción en cadena de la polimerasa. Para lo anterior, se utilizaron primers específicos para bacterias amplificando la región V6-V8 de genes bacterianos del DNAr 16S. La amplificación fue llevada a cabo usando los primers 968-f con grapa GC y 1401rusados ampliamente en distintos estudios de comunidades microbianas (Hirooka et al., 2009; Ziembinska, 2007). Previo al uso de los primers, se resuspendieron con agua libre de DNAasas para alcanzar una concentración de 100 pmol/µL (se corroboró usando la aplicación resuspension calculator del fabricante) y se elaboraron alícuotas de 50 µL con una concentración final de 20 pmol/µL. El DNA obtenido durante la extracción se llevó a una concentración de 200 ng/µL para tener una concentración homogénea en todas las muestras. El rango recomendable para realizar PCR va de 100 a 500 ng/µL (Cano-Medina y BañoMata, 2011). Para realizar la PCR, cada mix de reacción se preparó con DNA a una concentración de200ng/µL de las muestras de cada tratamiento desnitrificante y el inóculo control, 10 µL de GoTaq® Green Master MixGoTaq, 1 µL de cada primer y se aforaron a un volumen de 20 µL con agua libre de DNAasas. Adicionalmente se prepararon dos mix de reacción a la par. Al primero se le agregaron 10 μL de Gotaq®Green Master Mix GoTaq, 1 µL de cada primer y se aforaron a un volumen de 20 µLcon agua libre de DNAasas. Este es un control blanco sin DNA. 69 Al segundo control se le agregaron 10 μL de Gotaq® Green Master Mix GoTaq, 200 µL DNA y se aforaron a un volumen de 20 µL con agua libre de DNAasas. Este es un control sin DNA. Este es un control blanco sin primers. La amplificación se llevó a cabo en un termociclador Bio-Rad® con siguiente programa: predesnaturalización a 94°C por 2 min., 30 ciclos con una desnaturalización a 94°C por 45 s, una hibridación a 60°C por 30 s y una elongación a 72°C por 90 s., y una post-elongación a 72°C por 10 min. Finalizada la PCR, el tamaño del fragmento de cDNA, se verificó por electroforesis como se mencionó anteriormente. Esta técnica es muy sensible por lo que se deben evitar contaminantes, ya que es posible que el DNA no deseado se amplifique y se obtenga un DNA que no es de las muestras objetivo. Cano-Medina y Baño-Mata (2011) recomiendan utilizar controles blanco para tener mayor seguridad de haber obtenido la amplificación de interés. En la imagen 15 se observan las muestras amplificadas, éstas van del 1 al 5. Todas las muestras amplificaron y de acuerdo al marcador se encontraron entre 400 y 450 pares de bases (pb) lo cual indica que se amplificó la región deseada, pues se esperaba una banda en 433pb. Los carriles 6 y 7 contienen los controles blanco. El control blanco sin DNA (carril 6) no amplificó, esto demuestra que los primers y el agua no estaban contaminados con DNA o inhibidores. El control blanco sin primer (carril 7) tampoco amplificó, demostrando que hubo una buena manipulación en la preparación del mix para la PCR. 70 Contenido 1 - Control, fuente de inóculo 400pb 2 - Cinética desnitrificante sin amonio, t= inicial 3 - Cinética desnitrificante sin amonio, t= final 450pb 4 - Cinética desnitrificante con amonio, t= inicial 5 - Cinética desnitrificante con amonio, t= final 6 - Control blanco sin DNA 7 - Control blanco sin primer Figura 15. Amplificación de fragmentos de DNA por PCR 4.2.5.3 Electroforesis en gel con gradiente desnaturalizante (DGGE) Los productos de PCR fueron separados por electroforesis en gel con gradiente desnaturalizante (DGGE) en un equipo DCode™ Universal Mutation Detection Systems (Bio-RAD Laboratories, Hercules, USA). Se preparó el gel de poliacrilamida con gradiente desnaturalizante de 40 a 54% de urea-formamida. La solución buffer para realizar la electroforesis se mantuvo a 60°C, primeramente se le aplicó un voltaje de 200 V por 5 minutos. Posteriormente se cargó el producto de PCR y se aplicaron 60 V durante 960 minutos. Finalmente el gel fue teñido con bromuro de etidio (10 mg/L) y escaneados mediante un fotodocumentador. La figura 16 muestra el resultado del corrimiento del DGGE. En esta técnica, el DNA empieza a migrar en la electroforesis y conforme los fragmentos del mismo tamaño ya entrando en el gradiente desnaturalizante, estos se van separando de acuerdo a su composición nucleotídica, de tal manera que aquellos fragmentos 71 que contienen mayor cantidad de bases de adenina y timina (unidos por dos 1 van 2 3 4 5en las áreas de menor puentes de hidrógeno) se deteniendo desnaturalización. Se utilizó el DGGE para determinar la diversidad bacteriana en muestras de los tratamientos desnitrificantes con o sin amonio y del inóculo para contrastarlo como control. Contenido 1 – Control, fuente de inóculo 2 – Cinética desnitrificante sin amonio, t= inicial 3 – Cinética desnitrificante sin amonio, t= final 4 - Cinética desnitrificante con amonio, t= inicial 5 - Cinética desnitrificante con amonio, t= final a – banda 1 b – banda 2 c – banda 3 d – banda 4 e – banda 5 f – banda 6 g – banda 7 Figura 16. Comunidades microbianas de los diferentes ensayos separadas por DGGE. De acuerdo a estos criterios, este tipo de análisis considera que cada banda en el gel es considerada como una especie distinta en la comunidad y la intensidad de las bandas es tomada como un reflejo de la abundancia de esa secuencia en la comunidad (Rosas y col, 2004). Considerando lo anterior, se puede observar en el gel un perfil de bandas prácticamente similares entre los carriles de la fuente de inóculo y los ensayos desnitrificantes con y sin amonio a los tiempos inicial y final. Todos los tratamientos presentaron el mismo número de bandas, que son 7 bandas 72 fácilmente distinguibles acotadas de la a la g. También se puede observar que éstas migraron a la misma altura en el gel. Bajo estas consideraciones se podría indicar de manera general que las diferencias encontradas en las eficiencias de consumo de nitrato y sus velocidades específicas de consumo fueron atribuidas a factores metabólicos más que a cambios en la diversidad de las poblaciones microbianas observadas en el gradiente 40-54% con los primers antes especificados. Esto debido a que a través de los distintos tratamientos desnitrificantes con o sin amonio, no cambia la diversidad de las poblaciones bajo las condiciones mencionadas. Es interesante el seguimiento de estos resultados mediante la secuenciación, tal y como lo han hecho otros estudios de desnitrificación. 73 Conclusiones En base a los resultados obtenidos en el presente trabajo se puede concluir que: El reactor batch alimentado bajo condiciones anammox, consume amonio y nitrito simultáneamente, a una velocidad específica de consumo constante, por lo que se consideró estable. Los ensayos anammox muestran una alta eficiencia de remoción de amonio (88.64 %) y nitrito (100 %). La presencia de nitrato a concentraciones bajas (30 mgNO3-/L) no parece tener efecto negativo en la degradación de amonio y nitrito en la presencia de un consorcio microbiano tipo anammox. Bajo condiciones de desnitrificación (60.74 ± 4.15 mg N-NO3-/L, 85 mg CH3COONa/L), el inóculo utilizado alcanza una alta eficiencia de consumo de nitrato (90.16 %) pero no de producción de nitrógeno molecular (YP/S=0.60). La presencia de amonio afectó negativamente la velocidad específica de consumo de nitrato por consorcios microbianos desnitrificantes obteniendo una disminución de 0.01 mgNO3-/mgSSV∙d. Los consorcios microbianos desnitrificantes no degradaron el amonio bajo las condiciones establecidas en estos experimentos. La presencia de amonio no tiene efecto en la diversidad microbiana observada en el gel desnaturalizante con 40-54% de formamida-urea y los primers 968-f con grapa GC y 1401r. En base a los resultados obtenidos en el presente trabajo es posible realizar las siguientes recomendaciones: Para comprender mejor el ensayo tipo anammox con nitrato, es necesario repetir el experimento con mediciones de nitrato. De esta forma se confirmará si existe una tolerancia hacia él o su consumo. 74 Se puede precisar la influencia de amonio en el proceso desnitrificante realizando el experimento a diferentes concentraciones de amonio. Esto para saber bajo qué concentración existe un efecto negativo en las velocidades de consumo de nitrato. Para comprender mejor por qué la presencia de amonio no tiene efecto en la diversidad microbiana estudiada, se deben cortar las bandas obtenidas en el DGGE, purificar y secuenciar. Debido a que existen organismos que son más tolerantes a la presencia de diversos compuestos, o desnitrificantes que son capaces de degradar amonio. Esta información puede ser útil para probar con nuevos compuestos, con características similares o incluso más tóxicos. También es interesante hacer el estudio de la comunidad del proceso anammox para confirmar la presencia de éstas bacterias y estudiar su diversidad en los experimentos con y sin nitrato. 75 Bibliografía APHA, (1995).Standard methods for examination of water and wastewater. 19 ed. American Public Health Association, Washington D. C. pp 2-56, 2-57. Audesirk, T. (2011).Biología: la vida en la tierra con fisiología. Pearson. Novena edición, pp. 640. Baird, C. (2001). Química ambiental. Reverté, Barcelona, España, 622 pp. Bettazzi E., Caffaz S., Vannini C., Lubello, C. (2010). Nitrite inhibition and intermediates effects on Anammox bacteria: A batch-scale experimental study. ProcessBiochem. 45, 573-580. Cano-Medina, J. y Baño Mata, R. (2011). Identificación microbiológica mediante técnicas de biología molecular.Fesitess Andalucía, Málaga. Castillo-Rodríguez, F. & colaboradores (2005). Biotecnología ambiental. Tébar, Madrid, España, 592pp. CEPIS (1991). Manual de disposición de aguas residuales: origen, descarga, tratamiento y análisis de las aguas residuales. Centro Panamericano de Ingeniería Sanitaria y Ciencias del Ambiente. Lima. Chong-Jiang Tang, Ping Zheng, Cai-hua Wang, Qaisar Mahmood (2010).Suppression of anaerobic ammonium oxidizers under high organic content in high-rate Anammox UASB reactor.BioresourceTech. 101, 17621768. Contreras-López, A. y Molero-Meneses, M. (2006). Ciencia y tecnología del medio ambiente. Madrid: Universidad Nacional de Educación a Distancia (UNED). Dapena-Mora, A., Campos, J., Mosquera-Corral, A., Jetten, M. y Mendez R (2004). Stability of the ANAMMOX process in a gas-lift reactor and a SBR.Biotech. 159-170. 76 Díaz y Rodarte. (2003). Métodos para el estudio de comunidades microbianas en alimentos fermentados. Revista Latinoamericana de microbiología. Vol. 45, No. 1 pp. 30-40. DOF (1996). NOM-001-SEMARNAT-1996. Diario Oficial de la Federación, 1996. Duran, S. (2013). Conducta metabólica de un consorcio desnitrificante para la eliminación de fenol y nitrito, empleando un reactor de lecho empacado. Tesis de especialidad. Universidad Autónoma Metropolitana, Iztapalapa, México.39 pp. EPA (2009). National Primary Drinking Water Regulations.Environmental Protection Agency (EPA 816-F-09-0004). [Consultado en línea: Mayo 03, 2014]. Disponible en: http://water.epa.gov/drink/contaminants/upload/mcl- 2.pdf EPA (2012). Nitrogen and Phosphorus Pollution Data Access Tool [en línea]. http://www2.epa.gov/nutrient-policy-data/nitrogen-and-phosphorus-pollutiondata-access-tool. 5/05/14. Ginige, M. P., Keller, J. &Blackall, L. L. (2005).Investigation of an acetate-fed denitrifying microbial community by stable isotope probing, full-cycle rRNA analysis, and fluorescent in situ hybridization-microautoradiography.Applied and Environmental Microbiology. 71. 8683–8691. Guven, D.(2005)Propionate oxidation by and methanol inhibition of anaerobic ammoniumoxidizing bacteria.Appl. Environ. Microbiol. 71, 1066–1071. HACH (2014). Nitrogen, nitrite. Manual técnico. 2 pp. Henry, J., y Heinke, G. (1999). Ingeniería ambiental. Person Educación. Jiménez-Cisneros, B. (2001).La contaminación ambiental en México: causas, efecto y tecnología apropiada. México: Limusa, pp. 240-260. 77 Kartal B., Kuypers M. M. M., Lavik G., Schalk J., Op den Camp J. M., Jetten M. S. M., Strous M. (2007).Anammox bacteria disguised as denitrifiers: nitrate reduction to dinitrogen gas via nitrite and ammonium. Environmental Microbio. 9 (3), 635-642. Khanal S. (2008).Biotechnology for Bioenergy Production: Principles and Applications. Wiley-Blackwell. P. 8-12. Kuenen J. G. (2008). Anammox bacteria: from discovery to application. Nature Pub. 6, 320-326. Langone M., Yan Y., Haaijer S. C. M., Op den Camp H. J. M., Jetten M. S. M., Andreottola G. (2014). Coexistence of nitrifying, anammox and denitrifying bacteria in a sequencing batch reactor.Frontiers in Microb. 5, 1-12. Lindsay M. R. y col. (2001). Cell compartmentalisation in planctomycetes: novel types of structural organisation for the bacterial cell. Arch. Microbiol. 175, 413-429. Mansi y col., (2007).Fermentation Microbiology and Biotechnology.Second edition.Ed. Taylor and Francis Group. 16-25. Martínez-Hernández Sergio (2007). Análisis ecofisiológico de la desnitrificación en presencia de acetato y tolueno. Doctor en Biotecnología. Unidad Iztapalapa, Universidad Autónoma Metropolitana, Cd. De México, México, 141 pp. Martínez-Hernández, S., Sun, W., Sierra-Alvarez, R., Field, J. A. (2013). Toluenenitrite inhibition synergy of anaerobic ammonium oxidizing (anammox) activity.Process Biochem. 48, 926-930. Mathava K. y Jih-Gaw L. (2010). Co-existence of anammox and denitrification for simultaneous nitrogen and carbon removal – Strategies and issues.Journal of HazardousMaterials. 9. Metcalf, & Eddy. (1979). Ingeniería sanitaria: tratamiento, evacuación y reutilización de aguas residuales.Mc Graw Hill. 78 Mikesková H., Novotný C., Svobodová K. (2012). Interspecific interactions in mixed microbial cultures in a biodegradation perspective. Appl. Microbiol. Biotechnol. 95, 861-870. MO BIO Laboratories, Inc. (2013) PowerSoil® DNA Isolation Kit Instruction Manual. Version: 11212013 Osaka T, Yoshie S., Tsuneda S., Hirata A., Iwami N., Inamori Y. (2006). Identification of acetate- or methanol- assimilating bacteria under nitrate reducing conditions by stable-isotope probing.MicrobialEcology. 52, 253266. Parés, R. y col. (2002). Bioquímica de los microorganismos. Reverté, Sevilla, España. Patil S. S., Kumar M. S., Ball A. S. (2010). Microbial community dynamics in anaerobic bioreactors and algal tanks treating piggery wastewater.Appl. MicrobiolBiotechnol. 87, 353-363. PROMEGA.(2012). GoTaq® Green Master Mix.Part# 9PIM712.Revised 3/12 Ramalho, R.S. (2003). Tratamiento de aguas residuales (ed.Rev.). España: Editorial Reverté S.A, pp. 66-77. Ramírez-Sánchez J. A. (2013). Reducción biológica de nitrato a nitrógeno molecular por un consorcio microbiano anaerobio. Análisis del efecto de la temperatura. Lic. En Ingeniería Química. Facultad de ciencias químicas, Universidad Veracruzana, Xalapa, México. 67. Romero-Rojas, J. A. (2000). Tratamiento de aguas residuales por lagunas de estabilización. Modelos de diseño (López, R., Ed.). Alfaomega, D. F., p. 141-165. Rosas, I. y col. (2004). Microbiología ambiental. Instituto Nacional de Ecología (INE-SEMARNAT). 200 pp. 79 Schmid M., Twachtmann U., Klein M., Strous M., Juretschko S., Jetten M. (2000).Molecular evidence for genus level diversity of bacteria capable of catalyzing anaerobic ammonium oxidation.SystApplMicrobiol. 23, 93-106. Schuler M. L. y Kargi F. (2002). Bioprocess Engineering basic concepts. Secondedition, Prentice Hall. 23-24. Scragg A. H. (2009). Biotecnología para ingenieros. Horwood. 11. Seoánez-Calvo, M. ( 2004). Aguas residuales urbanas. Tratamientos natura les de bajo costo y aprovechamiento. Colección ingeniería medio ambiental. Mundi-prensa.368 pp. Skoog, D. (2002). Introducción a la química analítica. Reverté, Barcelona. Strous M., Heijnen JJ, Kuenen JG, Jetten MSM (1998). The sequencing batch reactor as a powerful tool to study very slowly growing microorganisms.ApplMicrobiolBiotechnol. 50, 589-596. Téllez-Pérez S. K. (2013). Evidencias de la flexibilidad metabólica de un consorcio microbiano y cultivos axénicos para desnitrificar, metanizar y reducir sulfato. Van Dongen, U., Jetten, M.S.M. y Van Loosdrecht, M.C.M.(2001) The SHARONANAMMOX process for treatment of ammonium rich wastewater.Water Science and Technology. 4 (1), pp. 153-160. Van Niftrik, L. A., Fuerst, J. A., Sinninghe-Samsté, J. S., Kuenen, J. G., Jetten, M. S. M., Strous, M. (2004). The anammoxosome: an intracytoplasmic compartment in anammox bacteria. FEMS Microbiol.233, 7-14. Vega-de Kuyper, J. C.(2007). Química del medio ambiente. Alfaomega, Madrid, 236 pp. Villaseñor Camacho, J. (2001). Eliminación biológica del fósforo en aguas residuales urbanas. Recuperado el 16 de Septiembre de 2012, de http://books.google.com.mx/books?id=8Vlu05kqFEgC&pg=PA16&lpg=PA16 80 &dq=caracteristicas+de+aguas+residuales&source=bl&ots=NJD6ByUBy8&s ig=_6IFVpmjhS5xCoxWJYuJzQl8Le0&hl=en&sa=X&ei=spVWUOLOCMWR qgHU3ICQBw&ved=0CDcQ6AEwAg#v=onepage&q=caracteristicas%20de %20aguas Yuan C., Li W., Fang M., Ji-Xian Y., Shan Q. (2014).Tracking composition of microbial communities for simultaneous nitrification and denitrification in polyurethane foam. Water Sc. & Tech. 69, 1788-1797. 81