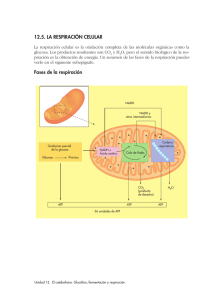
Balance de la descarboxilación del piruvato por
Anuncio

4. LA RESPIRACIÓN CELULAR AEROBIA La respiración celular aerobia es un proceso catabólico de oxidación total de la glucosa hasta CO 2 y H2O, siendo el O2 el último aceptor de los electrones, y cuya función es proporcionar energía para el trabajo celular y para la biosíntesis. La llevan a cabo las células eucariotas animales y vegetales en las mitocondrias, y muchas procariotas en el citosol y en la membrana celular. Previamente a la RCA se produce la glucólisis, que como ya hemos dicho produce dos moléculas de piruvato a partir de una de glucosa. Este piruvato sigue la vía respiratoria, que se divide en tres fases, que representan rutas conectadas: 1. Descarboxilación oxidativa del ácido pirúvico y formación del AcetilCoA. 2. Ciclo de Krebs, del ácido cítrico o de los ácidos tricarboxílicos. 3. Cadena de transporte electrónico y fosforilación oxidativa. 1. Formación del AcetilCoA En los organismos aerobios, el piruvato obtenido en la glucólisis (en el citosol) entra en la matriz mitocondrial y se descarboxila oxidativamente para formar AcetilCoA y CO 2, reacción catalizada por el complejo enzimático piruvato deshidrogenasa. Esta reacción se llama descarboxilación oxidativa, porque el ácido pirúvico libera una molécula de CO2 (descarboxilación) y es oxidado (oxidativa) para formar acetilo, el cual es transferido en la misma reacción a una molécula de coenzima A (HS-CoA) para formar acetil-CoA. NAD+ NADH+H+ Piruvato (3C) + CoASH Piruvato deshidrogenasa Como cada molécula de glucosa se rompe en dos de piruvato, esta reacción se multiplica por dos. Vemos que el esqueleto carbonado de la glucosa, con 6 carbonos, se ha oxidado a dos de CO2 y dos grupos acetilo, de dos carbonos, unidos al CoA. Esta oxidación del piruvato va acompañada de la reducción del NAD+, que pasa a NADH+H+. CO2 + AcetilCoA Balance de la descarboxilación del piruvato por molécula de glucosa: Dos AcetilCoA. 2 NADH + 2 H+. 2 C02. 2. Ciclo de Krebs Es la ruta oxidativa final de la glucosa y de la mayoría de los combustibles metabólicos (ácidos grasos y aminoácidos). Su función es oxidar totalmente el acetilo del AcetilCoA a CO2, al mismo tiempo que se producen transportadores reducidos (3 NADH y 1 FADH2), y GTP, que es equivalente en energía al ATP. Para ello se produce una cadena cíclica de reacciones catalizadas enzimáticamente. Se produce en la matriz mitocondrial. El ciclo se inicia cuando una molécula de AcetilCoA se combina con el oxalacetato, molécula de 4 carbonos, para formar citrato, de 6 carbonos, y liberándose el CoA, que puede volver a reutilizarse en el paso anterior. El citrato continúa con una serie de etapas que eliminan, en dos descarboxilaciones sucesivas los dos carbonos del acetilo en forma de dos moléculas de CO 2, regenerándose el oxalacetato, que puede volver a unirse a otro AcetilCoA. La ecuación global es: AcetilCoA + 3NAD+ + FAD + GDP + Pi + H2O 2CO2 + 3NADH +3H+ + FADH2 + GTP + CoASH Como cada molécula de glucosa origina dos de AcetilCoA, el ciclo de Krebs dará dos vueltas para quemar totalmente dicha glucosa. En el ciclo de Krebs se obtiene poca energía en forma de ATP (el GTP cede su grupo fosfato al ADP para formar ATP), pero sí en forma de coenzimas reducidos, que en la siguiente etapa entrarán en la cadena de transporte de electrones, donde se generará la mayor parte de la energía producida en la respiración celular. Pero hemos conseguido degradar toda la glucosa a CO 2. Balance del ciclo de Krebs por molécula de glucosa: 4 6 2 2 CO2. NADH + 6 H+. FADH2. ATP. El ciclo de Krebs es una ruta anfibólica, catabólica y anabólica a la vez, ya que se crean intermediarios que son precursores de las rutas biosintéticas. A partir de estos intermediarios se pueden fabricar aminoácidos, ácidos grasos, esteroles, bases nitrogenadas e incluso glucosa por neoglucogénesis. Este ciclo es por tanto la ruta central del metabolismo en las células aerobias. 3. Transporte de electrones y fosforilación oxidativa Los electrones y protones producidos en las fases anteriores se transfieren desde el NADH y FADH2 hasta el O2 (último aceptor), que se reduce para formar H2O, a través de una cadena de transporte de electrones llamada cadena respiratoria. Este proceso libera gran cantidad de energía libre que se emplea en la formación de ATP, proceso llamado fosforilación oxidativa. La finalidad del transporte de electrones en esta cadena es la regeneración de los coenzimas oxidados (NAD+ y FAD). En eucariotas, la cadena respiratoria se localiza en la membrana interna de las mitocondrias, concretamente en las crestas mitocondriales. En las células procariotas aerobias, se localiza en los mesosomas de la membrana plasmática. La fosforilación oxidativa es el principal medio de fabricación de ATP en los organismos heterótrofos a expensas de la energía generada en el transporte de electrones a través de la cadena respiratoria. Se dice por tanto que la síntesis del ATP a partir del ADP + Pi, está acoplada al transporte de electrones desde el NADH o el FADH2 hasta el O2. Se realiza mediante el complejo ATP-sintetasa. Por cada 2 electrones transferidos desde el NADH hasta el O2 se obtienen 3 moléculas de ATP, mientras que a partir del FADH2 se obtienen 2 ATP. El NADH cede sus electrones (reoxidándose a NAD+) a los distintos transportadores proteicos, liberándose la energía de forma gradual. Igual ocurre con el FADH2. Los electrones fluyen por la cadena mediante reacciones de oxidación-reducción de forma espontánea desde compuestos de mayor nivel energético a otros con menor nivel energético. Todas las proteínas transportadoras se reúnen en cuatro grandes complejos (I, II, III y IV), cada uno con sus coenzimas. El complejo I es el complejo de la NADH-deshidrogenasa, que contiene como coenzima el FMN, y transporta simultáneamente electrones y protones (H+), los cuales se los cede el NADH. El complejo II es el complejo llamado Ubiquinona, cuyo coenzima es el Coenzima Q, y que al igual que el anterior, transporta simultáneamente electrones y protones, los cuales pueden provenir del complejo I o del FADH2, por eso el rendimiento energético de esta cadena es mayor para el NADH que para el FADH2, ya que éste se incorpora más tarde a la misma. El complejo III es el complejo de citocromos b-c (citocromo b, citocromo c1 y citocromo c), que solo transporta electrones, los cuales los recibe del complejo II. El complejo IV es el complejo de citocromos a-a3, que al igual que el anterior sólo transporta electrones, que recibe del complejo III. El aceptor final de los electrones desde este complejo es el O2, que junto con los protones captados de la matriz mitocondrial, dará lugar a H2O. A continuación del complejo IV se sitúa el complejo de la ATP-sintetasa, que está formado por una base hidrófoba que se ancla en la membrana, un pedúnculo o región F0, y una zona esférica, o región F1, que es donde se catalizan las reacciones de síntesis de ATP. Balance de la cadena respiratoria por molécula de glucosa: 6 H2O. 10 NADH x 3 ATP = 30 ATP. 2 FADH2 x 2 ATP = 4 ATP. Se regeneran el NAD+ y el FAD. La teoría más aceptada para explicar cómo se transfiere la energía liberada en la reoxidación de los coenzimas para la síntesis de ATP es la teoría quimiosmótica, según la cual, la energía liberada en el transporte de electrones se emplea en bombear protones desde la matriz mitocondrial hacia el espacio intermembrana, produciéndose un gradiente electroquímico entre ambos (+ en el espacio intermembrana, con menor pH, y – en la matriz). Este gradiente es una situación desequilibrada que tiende a recuperar el equilibrio. La vuelta de los protones hacia la matriz, a favor de dicho gradiente, se realiza a través del complejo ATP-sintetasa obteniéndose ATP a partir de ADP + Pi. Los electrones del NADH y del FADH2 son transferidos al O2 que se une a protones de la matriz mitocondrial para formar H2O. Ecuación global y balance energético de la respiración aeróbica de la glucosa La ecuación global de la respiración celular aerobia de la glucosa es: Glucosa (C6H12O6) + 6 O2 6 CO2 + 6 H2O + Energía (38 ATP) El balance energético en número de moléculas de ATP que se forman en la oxidación completa de la glucosa hasta CO2 y H2O es: Glucólisis: se forman 2 ATP y 2 NADH Paso a AcetilCoA: 2 NADH Ciclo de Krebs: 2 GTP, 2 FADH2 y 6 NADH (2 + 2x3) = 8 ATP. (2x3) = 6 ATP. (2+ 2x2 + 6x3) = 24 ATP. Se obtienen 34 ATP en la oxidación de los transportadores de electrones mediante fosforilación oxidativa, y 4 ATP mediante fosforilación a nivel de sustrato. En total, 38 ATP.