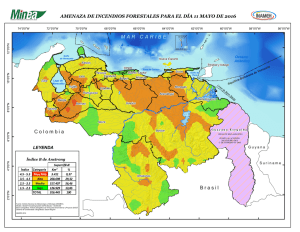
Lutjanus campechanus - Red Cubana de la Ciencia
Anuncio

Lutjanus campechanus 1. IDENTIDAD 1.1. Nomenclatura 1.11 Nombre válido: Lutjanus campechanus (Poey, 1860) Mesoprion campechanus Poey, 1860. En Mem. Hist. Nat. Isla de Cuba, 2(49):149 (Banco de Campeche, Key West). 1.12 Sinonimia Lutjanus campechianus Poey, 1875; Lutjanus blackfordii Goode & Bean , 1878 (Tomado de Allen, 1985). 1.13 Nombres comunes Nombres FAO: Español - pargo del golfo; Inglés - northern red snapper; Francés - vivaneau Campeche. Nombres locales: EUA -red snapper, sow snapper (individuos grandes), rat snapper (individuos pequeños, costa noroeste de la Florida), mule snapper (grandes), chicken snapper (individuos pequeños noreste de la Florida), Gulf red snapper, American red snapper (Moran y Morais, 1988); Pensacola snapper, Mexican snapper; México – guachinango (Duarte-Bello y Buesa, 1973, Allen, 1985). 1.2 Morfología 1.21 Larvas y juveniles Los huevos recién desovados de L. campechanus tienen un diámetro promedio de 0,82 mm (0,77-0,85). Son transparentes, esféricos y pelágicos, con un sólo glóbulo de aceite, con diámetro de 0,15-0,19 mm, vitelo claro y homogéneo. El período de incubación es de 20-27 horas a 23-25ºC (Richards, et al., 1994). En un desove provocado en laboratorio con gonadotropina coriónica humana, la mitosis se detectó a los 30 minutos de la fertilización. Minton et al., (1983) reportaron la eclosión a partir de las 20 horas después de la fertilización y aproximadamente el 50% de los huevos eclosionaron antes de las 25 horas, con temperaturas de 27ºC, mientras que Rabalais et al., (1980) la obtuvieron a 24-27 horas a 25°C. El embrión no presenta pigmentación hasta 21,5 horas después de la eclosión, cuando aparecen algunos melanóforos sobre los somites (Rabalais et al., 1980). El desarrollo larval y juvenil ha sido descrito por varios autores (Rabalais et al., 1980; Collins et al., 1980; Minton et al., 1983; y resumido por Drass et al. (2000). Las larvas presentan espinas en el preopérculo, opérculo, post- temporal y en el supracleitrum, sin aserraciones en las espinas de la cabeza. La flexión del notocordio ocurre a los 5 mm aproximadamente. Las aletas se desarrollan en la siguiente secuencia: caudal, pélvicas, pectorales, la dorsal y la anal. La pigmentación de las larvas y los detalles merísticos descritos por Drass et al. (2000) son diagnósticos. Los juveniles tempranos y juveniles tienen una mancha dorsolateral grande y son de color pálido con bandas difusas generalmente presentes. La 2da dorsal más pigmentada que las demás. Los juveniles de L. vivanus son similares pero tienen el iris amarino, desde los 50 mm LE. 1.22 Adultos Cuerpo alto, moderadamente comprimido, lomo elevado, ojo 5,5 a 6,0 veces en la longitud de la cabeza (Fig. 1). Preorbital 8-9% de la longitud estándar, parche de dientes vomerinos en forma de ancla, con una extensión en su línea media; lengua con dos parches de dientes granulares. Dorsal IX-X y 13-15, usualmente14; anal III-9 a veces 8, puntiaguda en ejemplares mayores de 50 mm LE; pectorales largas, pero sin llegar al nivel del ano, con 17 radios. Con 46-51 líneas de escamas oblicuas sobre la línea lateral, usualmente 47 ó 48. Espinas branquiales en el limbo superior del primer arco, 6 a 8 y 14 a 16 en el limbo inferior, total 21-24. Mandíbula inferior ligeramente proyectante, hocico algo puntiagudo. Color rojo cobrizo en la parte superior, rozado en la inferior, aletas rojizas, la dorsal con borde externo amarillento, la caudal puede tener un borde oscuro; con algunas líneas azules en la cabeza y a lo largo de las filas de escamas. Los ejemplares menores de 300-350 mm con una mancha oscura sobre la línea lateral; iris rojo (Resumido de: Rivas, 1966, Guitart, 1977, Allen, 1985 y Anderson, 2003). Según Robins et al (1986) se distingue de L. purpureus por tener 9 radios blandos en la anal (en vez de 8), menos escamas en una línea a lo largo del cuerpo (50-53 en L. purpureus) y menos escamas entre el inicio de la dorsal y la línea lateral (8-9). Una descripción detallada del desarrollo osteológico de esta especie fue presentada por Potthoff et al. (1988). Camber (1955) observó cierta diferencia entre los especímenes del Banco de Campeche y Bradley y Bryant (1975) la cuantificaron: en el 57% de los ejemplares, una línea recta, que se inicia en el hocico, pasa a través del extremo del opérculo y en un 34% de los ejemplares termina encima de la cola y en el 9% termina por debajo de la cola (N = 392). Fig. 1. Lutjanus campechanus (Poey, 1860) 1.3 Hibridación No hemos encontrado reportes de híbridos naturales de esta especie, a diferencia de otros pargos. Mediante ingeniería genética, sin embargo, se han logrado híbridos con Lutjanus griseus, con fines de cultivo (ver epígrafe 5.5). 2. DISTRIBUCIÓN Y HÁBITAT 2.1 Distribución Habita a lo largo de las plataformas continentales que bordean el Golfo de México desde Yucatán y el suroeste del Golfo hasta Key West y por la costa Atlántica hacia el norte hasta 151 Cabo Hatteras, Carolina del Norte (Fig. 2; Grimes et al., 1977b; Moran y Morais, 1988), aunque Anderson (2003) lo reporta hasta Massachusetts. En los muestreos realizados por el proyecto MARMAP (Marine Resources Monitoring Assessment and Prediction) en el este de la Florida ocupó el segundo lugar entre los pargos (Cuellar et al., 1996). No aparece registrado para las Bahamas por Böhlke y Chaplin (1993), ni por Rivas (1966; 1970) para el Caribe. En Cuba, sólo un ejemplar fue reportado por Guitart (1977) en el Golfo de Cazones, pero no hay evidencias de una población establecida. Fig. 2. Área de distribución de L. campechanus y principales zonas de pesca. 2.2 Hábitat 2.21 Juveniles Los juveniles, desde su asentamiento hasta que alcanzan 180-200 mm LT habitan preferentemente en fondos blandos (Grimes et al., 1977b; Szedlmayer y Shipp, 1994, Goodyear, 1995), aunque son más abundantes en microhábitats de bajo relieve, preferentemente en fondos arenosos con conchas de moluscos (Workman y Foster, 1994; Szedlmayer y Howe, 1997; Szedlmayer y Conti, 1999), donde no son abundantes los depredadores de gran talla (Gallaway y Cole, 1999a). Szedlmayer y Conti (1999) describieron, como una importante área de cría, el hábitat de fondo arenoso con conchas de moluscos localizado a 13 km al sur de Mobile Bay, Alabama. Generalmente son abundantes en aguas relativamente poco profundas (20-46 m). Nieland y Wilson (2003) indicaron que las plataformas de petróleo y gas en el norte del Golfo de México proveen importante sustrato duro y relieve vertical que sirve de hábitat a los juveniles de esta y otras especies. Según dichos autores, L. campechanus es reclutado a ese hábitat durante su segundo año de vida y permanece en el mismo durante varios años. La disponibilidad de hábitats adecuados para los juveniles puede ser un factor limitante del reclutamiento (Powles, 1977). Además, muchas áreas de cría son arrastradas por los chinchorros camaroneros y la mortalidad de los juveniles se estima en más de 20 millones de individuos por año en el Golfo de México. En Texas, los juveniles se mueven desde las aguas someras en el verano, hacia aguas más profundas (35-60 m) antes del invierno (Moseley, 1966; Bradley y Bryan, 1975). Esa migración en parte esta relacionada con el crecimiento, aunque algunos peces juveniles regresan durante la primavera a las aguas menos profundas. Nelson y Manooch (1982) sin embargo, no encontraron segregación de tallas entre las aguas costeras (<35 m) y profundas (> 35 m) en las afueras de las 152 Carolinas. Szedlmayer y Conti (1999), por su parte encontraron que la dispersión ocurre en todas direcciones después del asentamiento. De acuerdo con sus datos, los juveniles pueden mantenerse en los hábitats de asentamiento hasta cuatro meses. En aguas del sureste de Estados Unidos, los juveniles habitan en los arrecifes, pero emigran de estos cuando la temperatura del agua es menor de 16ºC (Parquer, 1990). Gallaway y Cole (1999a) concluyen que las mejores condiciones de hábitat para los juveniles se encuentran a profundidades de 18 a 64 m (preferentemente de 28 a 37) con temperaturas de 24 a 26°C, salinidad de 35 ppm (33-37 ppm según Moseley, 1966) y 5 mg/L de oxígeno disuelto. En muestreos realizados por Szedlmayer y Conti (1999) en las áreas de cría del nordeste del Golfo de México, encontraron salinidades de 30,5 a 35,4 ppt, oxígeno disuelto desde 3,7 hasta 7,9 ppm, con temperaturas que fluctuaron desde 17°C hasta 30°C. Franks et al. (2002) reportaron interesantes características de una población de L. campechanus (229-714 mm LT; 2-5 años), asociados a un refugio artificial en un estuario en Mississippi, Golfo de México. En algunos estudios se ha encontrado que los juveniles de L. campechanus son atraidos a los hábitats con refugios de bajo relieve (Szedlmayer and Howe 1997; Szedlmayer and Conti 1999). 2.22 Adultos Al alcanzar 180-200 mm LT, aproximadamente a los 18 meses de edad, L. campechanus empieza a asociarse con depresiones y estructuras de alto relieve, como arrecifes, rocas, refugios artificiales y plataformas de petróleo, abundantes en el norte del Golfo de México (Klima, 1976; Sonnier et al., 1976 Johnston et al., 1976; Stearns 1885; Moran y Morais, 1988, Gallaway y Cole, 1999a). Algunos autores sugieren que es más abundante alrededor de refugios aislados que en áreas con gran concentración de arrecifes (Sonnier et al., 1976; Smith, 1976; Clarke, 1986; Szeldlmayer y Shipp, 1994). Es abundante a profundidades de 40 a 110 m (Carpenter, 1965), aunque ha sido capturado desde 7 hasta 190 m (Moseley, 1966; Rivas, 1970; Parker y Mays, 1998). Según Grimes et al. (1977b) y Parker y Mays (1998) en la plataforma de Carolina del Norte y Carolina del Sur generalmente se encuentra entre 23 y 101 m, sobre fondos duros de bajo y alto relieve, mientras que en el Golfo de México es más abundante en 90 m. Parker et al., (1983) estiman que entre Pensacola, Florida y Texas-México, existen unos 2 780 km2, de hábitats con relieve irregular entre los 18 y 91 m de profundidad. De acuerdo con LGL y SAIC (1997), las plataformas de petróleo del golfo suministran unos 12 km2 adicionales de hábitat arrecifal en la región, ampliamente utilizados por L. campechanus. Szedlmayer y Shipp (1994) consideran que la existencia de una gran cantidad de refugios artificiales en las aguas de Alabama, hacen de esta región un hábitat óptimo para esta especie. Los refugios de bajo relieve probablemente también son importantes para los peces adultos en sitios donde no hay refugios artificiales. Los registros históricos de pesca apoyan este criterio, ya que indican que los adultos de L. campechanus eran frecuntemente capturados en las regiones norte-central y este del Golfo, antes del advenimiento de los refugios artificiales (Shipp 1999). Nieland y Wilson (2000) sugieren que L. campechanus es reclutado en las plataformas de petróleo del Golfo a edades de 1 y 2 años, ocupa ese hábitat durante unos pocos años y posteriormente emigra hacia otros hábitats. Los peces más viejos y grandes al parecer evitan las plataformas y permanecen en áreas más uniformes. Szeddlmayer y Schroepfer (2005) marcaron (con transmisores ultrasónicos) y rastrearon algunos guachinangos, en refugios artificales del noreste del Golfo de México. El 67% de los especímenes mostró una larga residencia (117-595 días), el 13% fueron residentes por 8-91 días y el 20% abandonaron el área poco después del marcaje (< 4 días). Los peces permanecieron en el mismo refugio artificial durante todas las estaciones (invierno, primavera, verano y otoño), sin cambios estacionales en la proximidad al refugio. Los autores concluyeron que L. 153 campechanus muestra una alta fidelidad durante largos períods de tiemo, lo cual sugiere que los refugios artificiales proveen de un hábitat adecuado para esa especie. L. campechanus ha sido capturado a temperaturas de 13 a 32°C (Rivas, 1970; Roe, 1976; Moran y Morais, 1988). Los límites de tolerancia parecen ser de 12,5°C, el inferior (Moore, 1973) y 33,5°C, el superior (Rivas, 1970). Salinidades de 60 ppm son letales para todos los ejemplares en experimentos de laboratorio, pero sobreviven a 45 ppm sin consecuencias graves (Huff y Burns, 1981). 3. CICLO DE VIDA 3.1 Reproducción 3.11 Sexualidad Lutjanus campechanus es una especie gonochórica (heterosexual). Tanto en las hembras como en los machos las gónadas son pares y de forma y tamaño semejantes, generalmente rodeadas de tejido adiposo. El sexo es diferenciable a partir de los 140-150 mm LH (Bradley y Bryan, 1975). 3.12 Maduración Varios autores (Camber, 1955; Futch y Bruger, 1976; Nelson y Manooch, 1982; Collins et al., 1996, Moran y Morais, 1988) coinciden en que L. campechanus tiene maduración sexual parcial desde que alcanza un año de edad, con 230 a 325 mm LH y maduración total al segundo año y aproximadamente 375 mm LH (Tabla 1). Al igual que otros pargos, los machos maduran a menor talla que las hembras (Grimes et al., 1977b). Tabla 1. Relación entre el largo máximo (mm) y la talla de maduración sexual de L. campechanus (después de Grimes, 1987) Región Sexo Largo Largo Talla Relación Relación Referencias mínimo de medio de máxima en Mi/Mx Me/Mx maduración maduración la población (Me) (Mi) (Mx) Banco de M, H 320 FL 778 0,41 Camber, 1955 Campeche Noroeste del Golfo M,F 190-300 LE Moseley, 1966 de México Noroeste del Golfo M 230 LH 850 0,27 Bradley y Bryan, 1975 H 255 LH 850 0,30 de México Suroeste de la M, H 334 LH, 941 0,35 Futch y Bruger, 1976; 2 años Florida U.S. South Atlantic M,H 325 LH 970 FL 0,33 Collins et al., unpub. Bight y Golfo de Cit por Grimes, 1987 México Sureste Atlántico H 287 LT 976 0,29 Allman et al., 2002b (1991-2000) 3 años. Golfo de México H 349 LT 934 0,37 (1998-2000) 3 años Alabama M,H 267 LH 275 916 0,29 0,30 Woods et al, 2003 Louisiana 292 LH 300 910 0,32 0,33 2 años 2 años Yucatán M 238 859 0,28 Brulé et al., 2002 H 247 0,29 Según Collins et al. (1996), en sus muestreos en el SE del Golfo de México, las hembras más pequeñas con oocitos hidratados tenían 344-400 mm LT. Otros autores (Bradley y Bryan, 1975; Brulé et al., (2002)) sin embargo, encontraron ejemplares maduros con tallas de maduración aún inferiores, aunque en la mayoría de los casos esta ocurre cuando los peces 154 alcanzan aproximadamente 30% de la talla máxima en la población (Tabla1). No hemos encontrado información sólida en relación con la talla media de maduración. 3.13 Gónadas Collins et al., (1987) encontraron, simultáneamente, oocitos en diferentes estadios de desarrollo en los ovarios, por lo que asumen que desova varias porciones durante el período de reproducción. A juzgar por los datos de Collins et al. (1996, 2001), la vitelogénesis es asincrónica y continua (tipo B en García-Cagide et al. 2001) y puede producir un elevado número desoves. Arnold et al. (1978) en mayo y junio de 1978 obtuvieron desoves múltiples de L. campechanus en cautiverio. Collins et al. (1996) estimaron de 20 a 35 desoves, con intervalos de 4 a 6 días. Según Collins et al. (2001) de acuerdo con el análisis de 36 hembras capturadas en 1998, la frecuencia de desoves fue aproximadamente 50% mayor en los peces de más de 6 años de edad que en aquellos de 3 a 5 años. Según Collins et al. (1996; 2001) el Índice Gonadosomático (IG) de L. campechanus (Fig. 3) presenta los valores más altos en los meses de mayo a julio. De mayo hasta noviembre encontraron gónadas en estadio III y de mayo a septiembre-octubre en estadio IV, pero preferentemente de junio a septiembre. El tamaño máximo promedio de los oocitos también se observo de junio a agosto, con 0,51 mm en junio, 0,67 en julio y 0,55 en agosto de 1992. Este parece ser el período de mayor intensidad de desove. Collins et al., 1996 realizaron el análisis histológico de 237 ovarios en estadios III y IV, colectados entre mayo y octubre de 1993. La apariencia de los oocitos en estadios I a IV fue idéntica a la descrita por Render y Wilson (1992) para una especie de Clupeidae que se caracteriza por el desarrollo asincrónico y continuo de los oocitos. En muchos ovarios en estadio IV se encontraron oocitos vitelógenos en reabsorción (atrésicos). Fig. 3. Variación del IG de L. campechanus en varias regiones del Golfo de México, durante el ciclo anual. A - en 1991-93 (Collins et al., 1996); B - en el período 1998-99 (Collins et al., 2001). 3.14 Apareamiento Evidentemente el período de desove de L. campechanus resulta extenso, aunque probablemente existen variaciones no detectadas entre zonas e incluso interanuales. La Tabla 2 resume los datos aportados por varios autores. Collins et al. (2001) muestrearon toda el área del Golfo de México de los Estados Unidos, desde el oeste de la Florida hasta Texas en un corto período: 1998-1999. Según sus resultados el período de reproducción empieza y termina en los mismos meses al sur de Texas y Louisiana y en todo el noreste del Golfo: desde abril-mayo hasta septiembre-octubre. No obstante, a juzgar por los valores del IG, el desove pico parece ocurrir de junio a agosto, lo cual coincide con lo reportado por otros autores. Szedlmayer y Conti (1999) mediante retrocálculo de la fecha de desove a partir de las marcas diarias en los otolitos, encontraron sólo dos cohortes bien definidas, en los meses de junio y julio en 1994 y mayo y 155 junio en 1995 (Fig. 5). En Yucatán, Brule et al. (2002) encontraron un período de desove más largo, con dos momentos de máxima actividad (Tabla 2). Tabla 2. Períodos de reproducción de L. campechanus en varios ecosistemas del Atlántico occidental. Simbología: x - colecta de hembras maduras u otro indicio de desove; xx - meses de desove pico. Localidad E F M A M J J A S O N D Referencias Campeche xx xx x x Camber, 1955 Texas x x xx x Moseley, 1966 NE Golfo, Texas x xx xx x x x x x Bradley y Bryan, 1975 W Florida x xx xx x x Futch y Bruger, 1976 Carolina del Norte y x x x Grimes et al., Carolina del Sur 1977b NW Florida x x xx x x Collins et al., 1986 N Golfo x x x Render, 1995 NE Golfo x x x x x x Collins et al., 1996 N Golfo, de Florida a Texas x x xx xx xx x x Collins et al., 2001 Yucatán x x xx x x xx xx xx x Brulé et al., 2002 Fig. 4. Distribución de frecuencias de las fechas retrocalculadas de eclosión a partir de las marcas diarias en los otolitos de juveniles en edad 0+ de L. campechanus (tomado de Szedlmayer y Conti (1999). 3.15 Agregaciones y desove La información existente sobre las agregaciones de desove del L. campechanus es pobre y poco consistente. (Moe, 1963; Futch y Bruger, 1976; Bradley y Bryan, 1975; Beaumariage y Bullock, 1976). Siendo una especie abundante y sobre la cual recae una pesquería tradicional, resulta raro que no sean bien conocidas sus agregaciones de desove. Si bien se reporta incremento de la densidad de peces maduros en los sitios conocidos, no ha quedado bien definido en cada caso si se trata de sitios de desove o de tránsito. Por otra parte, la mayoría de las agregaciones de desove de otros pargos se produce en áreas pequeñas, muy localizadas, con características específicas (profundidad, tipo de fondo, temperatura, régimen de circulación, etc), mientras que 156 en este caso no parece existir un patrón común entre ellas. La estacionalidad de las capturas tampoco muestra un marcado incremento, directamente dependiente de las agregaciones de desove, como ocurre con otras especies de pargos (ver sección correspondiente sobre L. analis, L. synagris y otras especies). Aunque evidentemente durante la reproducción se producen agrupaciones más numerosas que las normales, parece que el desove ocurre con una mayor dispersión y en mayor diversidad de condiciones ambientales, a lo largo del área de distribución. Moe (1963) reportó dos áreas al sur de Panama City, Florida, entre 28 y 37 m de profundidad donde esta especie forma notables agregaciones, todos con las gónadas maduras. Esas áreas han sido pescadas por unos pocos años, por lo que no se conoce la frecuencia de dichas agregaciones (Futch y Bruger, 1976), ni se mantiene aún. Mediante encuestas a los pescadores, Lindeman et al. (2000) identificaron dos sitios en los Cayos de la Florida (al sur) en los cuales las agregaciones de desove son pescadas. Pescadores experimentados están convencidos de que, a diferencia de las agregaciones de L. analis y L. cyanopterus, las de L. campechanus continúan alimentándose durante ese período, lo cual enfatizan con el refrán - “el ultimo pez morderá el último anzuelo” (P. Glading, pers. com.). Según Bradley y Bryan (1975) L. campechanus desova lejos de los arrecifes. Beaumariage y Bullock (1976) reportaron desoves a profundidades de 18 a 37 m sobre fondos duros de arena con bajo relieve. Franks et al. (2002) reportaron un raro caso de una hembra en etapa final de maduración en un estuario. Collins et al., (2001) relacionaron seis áreas de agregación en el Norte del Golfo de México, localizadas a partir de informaciones de capitanes de embarcaciones comerciales y recreativas, sobre la presencia de hembras con las gónadas hidratadas. Las profundidades en esos sitios fluctúan desde 40 hasta 122 m y la temperatura en el fondo, de 20,6 a 29°C. Este amplio rango de temperaturas es llamativo, por cuanto en otras especies de pargos el desove parece estar vinculado con límites estrechos de temperatura. Otros autores han reportado para esta especie rangos de temperaturas durante la reproducción, 50% menor, pero también relativamente amplio: 23-25°C por Arnold et al, (1978), 25-27°C por Minton et al. (1983) y 24-27°C por Chestney y Filippo (1994). 3.2 Desarrollo temprano y asentamiento 3.21 Huevos y larvas Las larvas en áreas costeras prefieren las aguas medianamente profundas por el día y se mueven hacia arriba durante la noche, con una distribución más uniforme (Powles, 1977; Leis, 1987). 3.22 Transporte, asentamiento y cría El tránsito de la etapa larval planctónica a la forma de vida juvenil-bentónica ocurre cuando alcanzan 17-18 mm LE con unos 26 días de edad (Szedlmayer y Conti, 1999; Schirripa, 2000). No obstante, Workman y Foster (1994) encontraron post-larvas recién asentadas desde 12,7 mm LH. Según estos autores, este proceso ocurre generalmente con temperaturas en el fondo de 2224°C, comienza en junio y alcanza su pico en julio, agosto y septiembre y prácticamente cesa en este último mes (Schirripa, 2000). Holt y Arnold (1982) reportaron asentamientos principalmente en junio y julio, aunque pueden ocurrir de forma limitada hasta octubre. Durante el otoño los juveniles empiezan a emigrar hacia aguas más profundas donde pasan el invierno (Schirripa, 2000). 3.3 Alimentación Las investigaciones sobre la alimentación de L. campechanus se ven limitadas debido a que, por el cambio de profundidad durante la captura, la mayoría de los individuos vomita el contenido estomacal. En los juveniles se ha encontrado una mayor retención debido a que muchos han sido obtenidos de la pesca con chinchorro, el cual asciende lentamente, mientras que los peces 157 obtenidos con cordel y anzuelo (principalmente adultos) son ascendidos rápidamente (Bradley y Bryan, 1975). L. campechanus es un pez carnívoro oportunista bentónico, que utiliza una gran variedad de presas, como tunicados, celenterados, cangrejos, camarones y peces pequeños, aunque la proporción de peces en la dieta aumenta con el crecimiento (Grimes et al., 1977b). Futch y Bruger (1976) y Davis (1975) coinciden en que se alimenta en los fondos arenosos, conchíferos o fondos blandos cercanos a los arrecifes y otros fondos duros, más que en estos mismos. 3.31 Hábitos alimentarios de los pre-adultos En los estómagos de los juveniles capturados en el Golfo de México, los camarones se encuentran con una alta frecuencia durante todo el año (Camber, 1955; Bradley y Bryan, 1976; Tabla 3). Otros crustáceos (incluyendo cangrejos y squillas), peces y calamares se encontraron en el 2 al 10% de los estómagos. Por su volumen, sin embargo, los calamares, pulpos y camarones contribuyen con el mayor porcentaje. Szedlmayer y Lee (2004) reportaron que los pequeños guachinangos (< 60 mm LE) consumen principalmene chaetognatos, copépodos, camarones y calamares. Los de tallas algo mayores (50-280 mm LE) pasan a consumir peces, mayores cantidades de calamares y cangrejos y continúan alimentándose de camarones. Las dietas del guachinango se separan más por el hábitat que por la talla de los peces, lo cual se atribuye a una mayor caza sobre las presas de los arrecifes que sobre las presas de aguas abiertas. Así, los cambios de hábitat observados en los juveniles, se reflejan en su dieta y sugieren diferentes valores para el hábitat, basados no solo en la existencia de refugio para evitar la depredación, sino también en sus recursos alimentarios (Szedlmayer y Lee, op. cit.) 3.32 Hábitos alimentarios de los adultos Según los estudios de Camber (1955), Moseley (1966), Bradley y Bryan (1973), Futch y Bruger (1976) los peces constituyen el principal alimento de L. campechanus adulto (Tabla 3; Fig. 5). Aunque los invertebrados están representados con mayor frecuencia en los muestreos, los peces son más importantes por su peso. Concluyen también dichos autores que la mayoría de los invertebrados encontrados en los estómagos habitan en fondos arenosos o fangosos. Camber (1955) reportó que los adultos del Golfo de México consumen las siguientes presas en orden decreciente de ocurrencia: camarones, peces arrecifales pequeños, cangrejos y gasterópodos. Durante la primavera, se encuentran tunicados en sus estómagos. En especímenes asociados a refugios artificiales en un estuario en Mississippi, Franks et al. (2002) encontraron un predominio de peces (54% por su ocurrencia) en la dieta de ejemplares de 229 a 714 mm LT. Entre dichas presas predominaron Anchoa hepsetus y Arius felis y les siguieron en importancia los crustáceos (Portunide y Penaeidae). McCawley et al. (2003) en L. campechanus asociados a refugios artificiles en aguas afuera de Alabama, encontraron que los crustáceos en su conjunto, fueron dominantes en la dieta por su peso (Tabla 3). Los peces ocuparon un segundo lugar por su peso y por su Indice Relativo de Importancia (IRI), aunque los organismos planctónicos fueron predominantes por su frecuencia ((IRI = 30,9%) y número (30,2%). Las larvas de peces no identificadas de las familias Ophicthidae y Triglidae contribuyeron sustancialmente a la proporción de peces en la dieta. Entre los crustáceos braquiuros predominaron las jaibas (Portunidae) y otros cangrejos no identificados. El zooplancton pelágico estuvo formado principalmente por larvas de camarones mantis, anfípodos y pteropodos (Cavolinia sp.). 158 Fig. 5. Composición del espectro alimentario de L. campechanus, en el norte del Golfo de México. (N = 712; con contenido en el estómago: 187; tomado de Moseley, 1966). Concluyen los autores que L. campechanus se alimenta selectivamente de organismos no arrecifales, ya que en sus estómagos prevalecieron animales (tanto peces como invertebrados) que habitan en sustrados de fango o arena, adyacentes a los refugios artificiales muestreados. Muchas presas potenciales, que son abundantes en los arrecifes de la zona, no fueron consumidos por L. campechanus o en muy pequeña proporción. Lee (1998) sin embargo, reportó en juveniles también colectatos en Alabama, que la dieta de esta especie está asociada a las comunidades arrecifales, (peces tales como Halichoeres sp, Serranus sp, Centropristis sp.) camarones (Lysmata sp y Synalpheus sp,) y calamares. Tal diversidad de hábitos alimentarios caracterizan a esta especie como altamente oportunista. 3.33 Variaciones ontogénicas y estacionales de la alimentación Según los datos de Bradley y Bryan (1975), los juveniles muestreados en 1970 y 1971 consumieron camarones y otros crustáceos en mayor proporción durante el invierno y la primavera. En 1972, encontraron también el más alto consumo de camarones y crustáceos durante la primavera, un aumento del consumo de calamares durante el verano y de pulpos durante el otoño (Tabla 3). En los adultos, los peces constituyeron el alimento fundamental durante todo el año, pero consumieron mayor diversidad de presas durante el verano y la menor en invierno. En primavera aumentó el consumo de anguilas, camarones mantis y Sicyonia sp. En verano aumentó el consumo de Callinectes danae y Sicyonia dorsalis, mientras que en invierno se mantuvo un alto consumo de anguillas. McCawley et al. (2003), observaron que aunque los peces son consumidos durante todo el año, no constituyen el alimento principal en todas las estaciones. En el verano (31,4%) y en el otoño (36,3%) los cangrejos fueron más importantes por su peso, mientras que en invierno predominaron los camarones mantis adultos (33,3%) y zooplancton pelágico (39,1%). Bailey (1995) reportó similar composición de la dieta, también en aguas de Alabama. En relación con los cambios de alimentación durante la ontogenia, Moran y Morais (1988) resumen lo siguiente: los juveniles de hasta 150 mm LH se alimentan de pequeños organismos zooplantónicos, pero ya desde los 40-90 mm empieza una lenta transición de una dieta 159 dominada por zooplancton a otra dominada por macro-animales (Moseley, 1966; Bradley y Bryan, 1976; Fig. 5). No obstante aún en ejemplares de 120-160 mm se ha encontrado un abundante consumo de larvas de peces, camarones y gusanos-flecha (Krohnitta sp.) Según Ouzts y Szedlmayer (2003) los guachinangos pequeños (200-299 mm LE) se alimentan principalmente de presas que habitan los arrecifes y arenales, los medianos (300-399) lo hacen en similares partes del arrecife, en la arena y de presas de que utilizan ambos tipos de hábitat y los grandes (400-499 mm) lo hacen de presas de hábitats mezclados. Las dietas de los guachinangos pequeños están dominadas por peces, las de mediana talla por peces y tunicados y la de los grandes por cangrejos y peces. Considerando igual tamaño de los peces, el grado de repleción estomacal fue significativamente más bajo al oscurecer que durante el día. Los peces fueron las presas más importantes en todos los períodos (amanecer, por el día, al anochecer y por la noche). Dichos autores sugirieron que la clase de talla tiene mayor influencia en la dieta que el período del día. 3.4 Edad y crecimiento 3.41 Patrones de crecimiento Bortone y Hollingsworth (1980) compararon lecturas de edades en otolitos, escamas y vértebras de 46 ejemplares de L. campechanus capturados en las afueras de Alabama. En los tres casos, de 76 a 80% de las estructuras fueron legibles y se obtuvieron similares valores de largos medios para cada clase anual y fueron considerados con igual nivel de confiabilidad. Por su parte, Nelson y Manooch (1982) en las Carolinas también compararon las lecturas de escamas y otolitos con similares resultados, aunque prefirieron las escamas. González et al. (1994) en el Banco de Campeche, encontraron que en las escamas se forman dos anillos anuales. No obstante, dicho autor sólo pudo leer las de ejemplares menores de 620 mm LH, por lo que utilizó el análisis de frecuencia de tallas por el programa ELEFAN para calcular los parámetros de crecimiento. En su análisis incluyó peces de 247 hasta 825 mm LH (885 mm LT) obtenidos de las capturas comerciales de 1984-1985. En la región norte del Golfo de México, L. campechanus está sujeto a importantes cambios cíclicos de los parámetros ambientales tales como la temperatura y productividad, por lo cual los cambios en la tasa de crecimiento son marcados y se registran bien en las estructuras. Camber (1955) y Moseley (1966) plantearon que esta especie deja de alimentarse durante la reproducción, lo cual pudiera ser la causa de formación del anillo anual. Futch y Bruger (1979) analizaron los incrementos marginales en los otolitos y observaron que efectivamente, en aguas de la Florida se forma un anillo en los otolitos durante los meses de junio a octubre, coincidiendo con el período de reproducción. Aunque los datos de Nelson y Manooch (1982) apoyan esta hipótesis, la presencia de un anillo en peces de un año (inmaduros) puede indicar que un ritmo fisiológico innato, pudiera también estimular la formación de dicho anillo. Estos últimos autores plantearon que en las escamas de esta especie también se forma un anillo de crecimiento cada año en los meses de junio-julio en los peces del norte del Golfo y en abrilmayo en los del Atlántico. O sea, que la formación de marcas en las estructuras no parece estar asociada con las temperaturas mínimas del invierno. Manooch et al. (1997) precisaron, por el análisis de los incrementos marginales mensuales, que en el sureste Atlántico, el anillo en los otolitos se forma entre marzo y mayo, lo cual difiere de los resultados de Futch y Bruger (1979). Szedlmayer (1998) comprobó que en los otolitos de los juveniles se forma un anillo de crecimiento diario, con algunas variaciones dependientes de la tasa de crecimiento, por lo que esta técnica puede ser utilizada para estudiar el crecimiento durante las etapas tempranas de desarrollo. 160 Goodyear (1994), precisó que el mínimo valor del incremento marginal, en los L. campechanus del Golfo de México, se observa en el mes de julio, por lo que considera que es el mes pico de formación del anillo anual y coincide con el periodo de reproducción. Allman et al. (2002b) sin embargo, encontraron que en mayo culmina la deposición del anillo opaco en esa región, mientras que en los especímenes del Atlántico ocurre en junio y julio. Wilson et al. (2001) realizaron un minucioso análisis de los incrementos marginales en los otolitos L. campechanus en el norte del Golfo, región de Louisiana, durante 1989-1992 y 19951998. Estos autores coinciden en que se forma un sólo anillo (hialino) anual, en el periodo entre junio y noviembre, cuando se observan los valores más bajos del incremento marginal (Fig. 6). Dichos autores encontraron ejemplares de 104 a 1 039 mm LT y de 17 a 22793 g de peso total, con edades de 0,5 a 52,6 años. Sus datos muestran que a partir del décimo año de vida, el crecimiento es casi asintótico, por lo que, a partir de esa edad, no se observa una clara correlación entre esta y el largo del pez. Concluyen dichos autores, que debido a la gran heterogeneidad de las edades a un largo o peso determinado, el uso de variables morfométricas para predecir la edad en esta especie presenta serias limitaciones. Allman et al. (2002a) también encontraron una gran variabilidad en la talla a cada edad. Por ejemplo, entre los peces de tres años se encontraron tallas desde 320 hasta 700 mm LT (300 hasta 655 LH) y en los de cuatro años varió de 350 a 780 mm LT (325 a 725 LH). Fig. 6. Variación anual del porcentaje de anillos opacos en el borde de los otolitos de L. campechanus (tomado de Wilson et al., 2001). Szedlmayer (1998) observó en condiciones naturales, que los juveniles marcados con oxytetraciclina, cuya tasa de crecimiento era rápida (0,63 mm LH por día), presentaron incrementos de anillos diariamente, pero en aquellos con crecimiento somático muy lento (0,2 mm LH por día) no se depositaban anillos cada día. En larvas cultivadas, los incrementos fueron diarios (0,99-1,03 por día) y la tasa de crecimiento fluctuó de 0,6 a 0,9 mm LH por día. Con tasas de crecimiento por encima de 0,3 mm por día, la deposición de anillos es diaria. Considerando que L. campechanus se reproduce a final de la primavera o en verano, cuando la temperatura del agua es más alta, es de es esperar altas tasas de crecimiento, por lo que dicho autor asume que el método de datación de anillos diarios es aplicable a esta especie. Actualmente se trabaja en el desarrollo de tecnologías más modernas para el estudio del crecimiento en los peces y en particular de L. campechanus. Patterson et al. (1998; 2001a) utilizaron marcadores bioquímicos naturales para estudiar las marcas de crecimiento en juveniles de edad 0+ e incluso identificar su área de cría en base a la composición microquímica 161 de los mismos. Las investigaciones actuales se orientan a la microquímica de los otolitos de los pargos adultos en los arrecifes exteriores para determinar en que áreas de cría fueron reclutados. Baker et al. (2001) utilizaron análisis radiométrico con Ra226 para validar las estimaciones de edad realizadas en otolitos seccionados L. campechanus en el norte del Golfo de México, obteniendo valores similares a los calculados por métodos convencionales. La actividad de Ra226 es inversamente correlacionada con el incremento de masa de los otolitos con la edad, validando la hipótesis de que la acumulación de Ra226 ocurre a una tasa proporcional a la del Ca. Este método ofrece una alternativa que elimina los probables errores de interpretación del lector de otolitos, aunque se requiere profundizar en las investigaciones y validar los resultados 3.42 Tasa de crecimiento La Tabla 4 presenta los parámetros de crecimiento de L. campechanus estimadas por varios autores. Los valores de la tasa de crecimiento K , fluctuaron de 0,07 a 0,33 con un valor promedio de 0,154 ± 0,04 DE para 24 estudios diferentes. Todos los valores de φ´, con excepción de los obtenidos por Fischer (2002) para Texas, caen dentro del intervalo de confianza al 95% de la media de φ´ = 3,059 ±0,2 DE, obtenida con todos los valores. Algunas investigaciones sobre el crecimiento de L. campechanus indicaban una tasa de crecimiento alta en el primer año de vida, aproximadamente 200-220 mm LE, mientras que otros estudios sugirieren un crecimiento lento en el primer año (Apéndice 1). Holt y Arnold (1982) concluyeron que en el primer año de vida alcanza 110-130 mm y 200-230 mm en el segundo. En algunos casos las diferencias pudieran ser consecuencia de imprecisiones o diferentes criterios en la definición del primer anillo anual. No obstante, la mayoría de los estudios son bastante consistentes, independientemente de algunas diferencias, tanto en cuanto a los largos retrocalculados como a los parámetros de crecimiento (Tabla 4). Tabla 4. Parámetros de crecimiento de L. campechanus en varios ecosistemas del Golfo de México. Abreviaturas: O- otolitos; E- escamas; FT- frecuencias de tallas; * -valores calculados a partir de los datos originales obtenidos de la literatura; LH -largo horquilla; LT -largo total; LE -largo estándar. Localidad Referencias Lmax, K To Método W∞ ϕ´* φ´* L∞ (cm) (cm) O 64,4 LH* 0,33* -0,29* 4228* 90,0 LT 0,907 3,22 Florida Futch y Bruger, 1976 69,2 LT* 117,0* 0,18* -0,21* 22572 0,791 3,113 Golfo de México Wakeman et al., 1979 125,5* E 97,0 LT 0,165 -0,01 17144 0,708 3,191 Carolinas (1974-78) Nelson y Manooch, 1982 E E E E E E 0 O 97,0 LT 97,5 LT 94,1 LT 95,0LT 94,1 LT 97,0 LT 92,5 LT 102,5 LT 0,155 0,160 0,170 0,175 0,170 0,162 0,14 0,15 -0,01 0,00 -0,1 -0,1 -0,10 -0,01 0,10 - 13416 15246 12015 13362 13593 15096 13714 15498 FT O O O O O O FT 116,6LT 95,5 LT 93,8 LH 96,9 LT 88,4 LH 87,3 LH 102,3 LH 86,0 LH 92,3 LT* 0,115 0,146 0,175 0,192 0,21 0,24 O,07 0,133 -0,59 0,182 12198 -0,530 0,020 0,63 11817* 0,73 12554* -2,92 -0,36 9631 93,7 91,6 91,3 84,6 0,680 0,696 0,711 0,726 0,711 0,700 0,621 0,684 3,164 3,182 3,178 3,198 3,178 3,183 3,078 3,198 0.307 0,649 0,722 0,773 0,782 0,836 1,353 0,598 2.384 3,124 3,187 3,256 3,215 3,262 3,865 3,054 162 E Florida (1976-78) SE Atlántico, EUA W Florida (1978-79) Louisiana (1978-79) G. México Todas las áreas unidas. Nelson et al., 1985 G. México, EUA Alabama, EUA (1989- Szedlmayer y Shipp, 1991) 1994. N Golfo de México Goodyear y Phares, 1990 SE Atlántico, EUA Manooch y Potts, 1997c NW Golfo de México Wilson et al., 2001 N Golfo de México Patterson et al., 2001 Alabama, G. Mexico Fischer, 2002 Louisiana, G.Mexico Texas, G. Mexico Yucatán (1984) González et al., 1994 FT E FT (BP) FT (SLCA) 97,4 LT 92,0LT 80,4LT 987,LT 85,0 LT 0,135 -0,76 11700 0,12 0,13 0,12 12200 0,14 12700 0,622 0,552 0,543 0,575 0,593 3,107 3,007 2,924 3,068 3,005 Yucatán (1985) Tamaulipas Tamaulipas Campeche Campeche Rodríguez Castro, 1992 Rodríguez Castro, 1992 Rodríguez Castro, 1992 Leonce-Valencia y Monroy, 1997 Nelson y Manooch (1982) encontraron diferencias pequeñas, aunque estadísticamente significativas entre las curvas de crecimiento obtenidas para aguas de Louisiana, el oeste de la Florida, el este de la Florida y las Carolinas (Tabla 4). Fischer (2002) también encontró mayor tasa de crecimiento en Alabama y Louisiana que en Texas, lo cual consideran, pudiera indicar diferencias biológicas en las poblaciones, que justifiquen el manejo a nivel sub-regional. Wakeman et al. (1979) estimaron el crecimiento a partir de peces de 1-2 años de edad mantenidos en cautiverio por un período de 300 días y calcularon un incremento diario en peso de 0,45% del peso del cuerpo, valor aproximado al estimado en poblaciones naturales. Szedlmayer y Conti (1999) a partir del análisis de los anillos diarios en juveniles, calcularon tasas de crecimiento de 0,4 a 1,1 mm por día (N = 857) para peces de edad 0+. Las tasas de crecimiento de las cohortes de junio (0,77 mm/día) y julio (0,71 mm/día) de 1994 fueron significativamente mayores que la de las cohortes de mayo (0,51 mm/día) y junio (0,67 mm/día) de 1995 (Fig. 4). Al parecer, esa diferencia se debe a una menor densidad de juveniles en 1994 (menor CPUE), lo que facilitó mayor tasa de crecimiento. No obstante, también puede haber contribuido a un menor crecimiento, las bajas temperaturas observadas en el mes de mayo de 1994. De cualquier forma, se evidencia una notable variabilidad interanual del crecimiento. Las tasas de crecimiento observadas en los juveniles estudiados por Szeldmayer y Conti (1999), no concuerdan bien con los largos retrocalculados para el primer año de vida por la mayoría de los autores. Tomando, por ejemplo, los valores mínimo (0,51 mm) y máximo (0,77 mm) de crecimiento diario reportados por dichos autores, los L. campechanus deberian alcanzar de 186 a 281 mm en su primer año. Por otra parte, a partir de la recaptura de peces marcados (Beaumariage, 1964; 1969; Beaumariage y Wittich, 1966), Futch y Bruger (1976) estimaron un crecimiento anual promedio de 78 mm LT y 62 mm LE para peces de 205 a 419 mm LE al momento de liberación. Moe et al., 1970 reportaron la recuperación de un pargo marcado después de 5,8 años, el cual creció de 307 hasta 765 mm LT, con un promedio anual de 68 mm. Futch y Bruger (1976) recapturaron otro ejemplar después de 10 años en libertad, que promedió 44 mm LT por año. Bradley y Bryan (1975) sugieren que la tasa de crecimiento fluctúa entre 60 y 90 mm (promedio 75 mm) en peces de más de un año de edad. Tales tasas de crecimiento de los peces marcados, son semejantes a las estimadas por Szeldmayer y Conti (1999) para los juveniles. En algunos estudios del crecimiento de L. campechanus, utilizando estructuras duras, se han obtenido valores semejantes a los estimados en las marcaciones mencionadas: según los datos de Wilson y Nieland (2000) alcanzan 204 mm LT (0,56 mm/día) en el primer año, según los de Fischer (2002) para Texas (245 mm LH≅264 mm LT ≅ 0,67 mm/día) y de acuerdo con González et al. (1994) a partir de análisis de las escamas, se obtiene 206 mm LH (222 mm LT≅0,61 mm/día). Por otra parte, resulta paradójico que las tasas de incremento en el primer año de vida resulten ser inferiores que las obtenidas por marcación en peces de mayor edad. Otros autores sin embargo han obtenido valores inferiores de las recapturas: Szedlmayer y Shipp (1994) estimaron una tasa de crecimiento de sólo 0,22 mm LT por día o sea, 80 mm por año (n = 132), que resulta similar a la estimada por ellos mediante el análisis de los otolitos para los peces menores de 10 años de edad (0,27 mm LT por día; n = 397). También a partir de las recapturas, dichos autores plantearon que el crecimiento durante los 10 primeros años es lineal y 163 puede ser expresado por la ecuación: LT = 97,7 Edad + 67,6 (N = 397, R = 0,87). Patterson et al. (2001c) obtuvieron una regresión lineal (∆LT= 2,34 mm + (0,238 mm/días en libertad), (F1,286 = 631,0; P < 0,001; R2 = 0,76) entre el cambio de tamaño y el tiempo en libertad de 561 ejemplares marcados en el norte del Golfo de México. O sea, que el crecimiento fue de 0,238 mm/día (LT), cifra muy parecida a la obtenida por los mismos autores a partir de lecturas de otolitos: 0,240 mm/día. Dicha cifra es también comparable con la tasa de crecimiento estimada por otros autores (Render, 1995; Szedlmayer y Shipp, 1994). No se encontraron diferencias significativas entre los parámetros de crecimiento para hembras y machos. El largo estimado a partir de los parámetros de crecimiento (Apéndice 1) en la gran mayoría de los estudios realizados fluctúa entre 100 y 170 mm LT (0,27-0,46 mm/día). Goodyear y Phares (1990) calcularon la ecuación invertida de von Bertalanffy para estimar la edad en función del largo del pez (en pulgadas): Edad = -0,47 log (1- LT/47,1)/ 0,11)). 3.43 Relaciones morfométricas durante el crecimiento La Tabla 5 resume las ecuaciones que representan la correlación entre el largo y el peso de L. campechanus, dadas por varios autores. Patterson et al. (2001c) no encontraron diferencias significativas entre sexos por lo unieron todos los ejemplares para una sola ecuación. La Tabla 6 presenta las correlaciones reportadas en la literatura, entre diferentes medidas de longitud, a los efectos de convertir las unidades. N Tabla 5. Relaciones largo-peso de L. campechanus en varios ecosistemas del Gran Caribe. Rango de Formula Unidade Localidad Referencias tallas s 90-190 LH Log10W = – 4,7921 + 3,01 Log10FL mm/g Campeche Camber, 1955 (Juv.) P = 1,614 x 10-5 (LH)3,01 290-750 F P = 0,302x10-5(LH)2,915 M P = 2,98 x 10-5(LH)2,917 242 251 240 Juv Juv 200-676 LH 722 90 232-745 143 142 177 462 512 175-940 108 4143 3610 200-890 Log10P =-3,93125 + 2,6051 Log10 LT P = 1,171 x 10-5 (LT)2,605 LT = 11,8517 P03347 Log10W = – 4,7724 + 2,9942 Log10 FL P = 1,68892x10-5 (LH)2,9942 Log10W = – 4,8539+3,0092 Log10 FL P= 1,40x10-5 (LT)3,0092 Log10W =– 4,483 + 2,885Log10 FL P = 0,3289x10-5 (LH)2885 P = 1,82 x 10-5 LT2,996 P = 1,36 x 10-5 LT3,017 P = 3,15 x 10-5 LT2,887 Log10W =– 4,7399 + 2,966 log10 FL P = 2,04 x 10-5 (LT)2,966 P = 5,728x10-6(LT)3,145 P = 6,07x10-5 (LE)2,89 P = 5,28x10-6(LH)2,819 P = 2,02x10-6(LH)2,969 P = 3,95x10-6 (LH)2,868 mm/g Norte del Golfo de Dawson, 1963 México mm/g Florida Futch y Bruger, 1976 mm/g Alabama Wade, 1981 mm/g Texas mm/g mm/g mm/g mm/g Wakeman et al., 1979 Nelson y Manooch, 1982 P = 0,000414LT3,07 LT = 11,851 (P)0,3347 P = 9,36x106(LH)3,0972 Pulg/lb P = 4,68 x 10-9 LT3,17 mm/kg ? Louisiana Panama City, Fl. Carolinas Todas las áreas unidas Texas Camber, 1984 Louisiana Dennis, 1984 W Flower, Garden Zastrow, 1984 Bank E Flower, Port Aransas, Texas Golfo de México, Goodyear, 1987 EUA Golfo de México, Cummings Parrack, EUA 1986 N Golfo de Patterson et al., México, 2001c N Golfo de México Wilson y Nieland, 2000 Golfo de México Goodyear, 1994 mm/g 3791 104-1039 P = 1,17 x 10-8(LT)3,04 mm/kg 2918 10-38 pulg P = 4,30x10-4 (LT)3,048 Pulg./Lb 164 777 247-825 280-7800 g 84 P = 3,9452x10-5 (LH)2,857 mm/g González et al., 1994 P = 1,5 x 10-8 (LT)2,99 Banco de Campeche mm/kg SE Florida P = 2,57x10-8(LH)2.94 Mm/g Manooch y Pots, 1997 Fischer, 2002 P = 1,54x10-8(LH)3.03 Mm/g Alabama, G. México Louisiana, G. Mëxico Tabla 6. Ecuaciones de correlación entre el largo horquilla (LH), el largo estándar (LE) y el largo total (LT) de L. campechanus. N Ecuación Coef. de correlación Referencias 21 100 180 265 171 171 240 203 203 1285 265 FL = 1,1585 SL + 13,3 TL = 1,0678 FL + 3,5 LT = 1,0712 FL + 1,7 LH = 0,9248 LT + 1,2746 LT = 1,0702 LH + 3,1185 LH = 1,1419 + 17,5348 LE = 0,8532 * LH - 6,1493 LT = 1,1873 LE + 34,0365 LE = 0,7989 LT - 9,5574 LT = - 3,21 + 1,08 LH LT = 1026 + 1,24 LE LH = 11,67 + 1,15 LE TL = 0,038 + 1,275 LE TL = 0,120 + 1,070 LH r = 0,9901 r = 0,9975 r = 0,99 r2 = 0,99 r2 = 0,99 r2 = 0,99 r2 = 0,99 r2 = 0,99 Futch y Bruger, 1976 Fuch y Bruger, 1976 Nelson y Manooch, 1982 Cummings Parrack, 1986 Manooch y Pots, 1997 Goodyear y Phares, 1990 3.44 Longevidad L. campechanus es, entre las especies de la familia Lutjanidae, la que mayor longevidad alcanza a juzgar por los datos reportados en la literatura: 39 años (Fitzhugh et al., 1999), 42 años (Szedlmayer y Shipp, 1994), 49 años (Wilson y Nieland, 2000) y 53 años (Render, 1995; Wilson et al., 2001), con longitudes de 900 a 1 000 mm LT. Sin embargo, en el sureste Atlántico la longevidad máxima reportada es de 25 años (Manooch y Potts, 1997) y en el Banco de Campeche es de sólo 22 años (González et al., 1994). 3.5 Fisiología 3.51 Dinámica de los indicadores morfo-fisiológicos y bioquímicos. No hay información 3.52 Metabolismo Moore (1973) determinó en condiciones experimentales la tasa de respiración y la velocidad de natación de L. campechanas a 12,5, 18 y 27,5ºC. La relación entre el consumo de oxígeno y la temperatura fue similar a la encontrada en otras especies, con valores de 112 mg/kg/hora, a 18°C. Wakeman et al. (1979) estimaron que en el metabolismo respiratorio de ejemplares de 1-2 años de edad, nadando a una velocidad de 0,5 km/hora se consume 89 mg O2-2/kg/hora a 20°C, mientras que para el metabolismo básico a la misma temperatura, se requiere de 62 mg O22/kg/hora. Para el mantenimiento del metabolismo de rutina esta especie requiere de una ración diaria mínima de 1,5% del peso de su cuerpo. Así mismo, dichos autores calcularon la eficiencia de conversión del alimento en 30%. 3.6 Conducta 3.61 Migraciones y movimientos locales 165 Existe consenso entre los pescadores comerciales y deportivos, en cuanto a que L. campechanus en verano se mueve hacia la costa y en invierno se aleja de esta (Moe, 1963). No obstante, muestra una fuerte tendencia a la residencia en determinados hábitat. Holt y Arnold (1982) plantearon que los ejemplares menores de 160 mm fueron raros en los muestreos con redes de arrastre en el noroeste del Golfo de México. Sus datos de marcaciones indican que los peces de esa talla aproximadamente, después de diciembre se mueven hacia aguas más profundas o sufren una alta mortalidad. Fable (1980) encontró pocos movimientos entre los 17 peces recuperados, de 299 marcados en aguas de 60 m de profundidad, cerca de las costas de Texas. Por su parte Szedlmayer y Shipp (1994) recapturaron el 57% de los ejemplares marcados, en el mismo lugar en que los soltaron y el 76% a una distancia menor de 2 km del punto de marcaje. El mayor movimiento fue observado en sólo un ejemplar, capturado a 32 km del sitio de marcaje. Similares resultados encontró Szedlmayer (1997) mediante marcación con transmisores ultrasónicos. Beaumeariage y Bullock (1976) también en base a marcación y recuperación, concluyeron que L. campechanus muestra una marcada tendencia a la residencia en los arrecifes profundos con limitados movimientos estacionales a las zonas de alimentación. Las recapturas de L. campechanus marcados en aguas someras (15 brazas = 27 m) sugieren que los peces se alejaron de los arrecifes después de ser marcados pero retornaron a los mismos arrecifes al año siguiente (Goodyear (1994). Los movimientos de aguas interiores-exteriores han sido ampliamente reportados y aparentemente están relacionados con patrones estacionales (Beaumariage y Bullock, 1976; Bradley y Brian, 1975; Moseley, 1966, Topp, 1964). Ocasionalmente se han observado movimientos de algunos individuos hasta distancias de 220 millas (354 km) en unos 3 años (Beaumariage, 1969). No obstante, los datos existentes indican que esta especie no realiza migraciones estacionales, al estilo de las pelágico-costeras. No obstante, sí se evidencia la existencia de movimientos de corto radio y no se descarta la posibilidad de una dispersión gradual desde los centros de abundancia. Tales movimientos explican la rápida colonización de muchos arrecifes artificiales por varias clases de edad. No obstante, al parecer la principal forma de dispersión de la población ocurre por el transporte de los huevos y larvas. Waterson et al. (1998) y Patterson et al. (2001b), en el norte del Golfo de México, analizaron el grado de fidelidad por determinado sitio y los movimientos de peces marcados, en ambos casos con resultados coincidentes. En el primer caso se documentó que, aunque los adultos son muy residentes, bajo la influencia de eventos climáticos de gran escala, como los huracanes, realizan importantes movimientos que facilitan la mezcla de la población. Patterson et al. (2001b) marcaron 2 932 ejemplares que fueron soltados, en unos casos, en lugares cercanos al lugar de captura, cerca de refugios artificiales y otros fueron liberados en lugares alejados. De las 561 recapturas, 235 se hicieron en el propio lugar de marcaje y 326 se recuperaron por la pesca recreativa o comercial. La distancia media recorrida fue de 29,6 km y la mayor distancia fue de 352 km. El tiempo medio en libertad fue de 404 días y el máximo 1501 días. La dirección resultante de todos los peces fue hacia el este. El 80% de los peces marcados eran menores de 380 mm LT (talla mínima de captura). De esos peces jóvenes, el 58% fue recapturado en un área de 20 km alrededor del sitio en que fueron soltados. Con el tiempo los movimientos de los adultos son mayores. Las migraciones observadas fueron más amplias que las reportadas previamente para los adultos y al parecer en ello influyó la ocurrencia de dos huracanes durante el periodo de liberación de los peces. Ingram y Patterson (2001) en un análisis posterior de los mismos datos, concluyen que L. campechanus muestra una fidelidad moderada hacia su área de hábitat Resulta evidente la existencia de movimientos estacionales de L. campechanus. Adams y Kendall (1891), Camber (1955), Moseley (1966) y Bradley y Bryan (1975) coinciden en que, en el Golfo de México, con el crecimiento se mueve de aguas someras a otras más profundas 166 antes del inicio del invierno, donde se mantiene con poco movimiento. Moseley (1966) no encontró relación con la disminución de la temperatura y sugirió que la migración puede estar relacionada con mayor disponibilidad de alimento en aguas más profundas. Bradley y Bryan (1975) plantearon que parte de esa población regresa a aguas someras durante la primavera y el verano. Por otra parte, dichos autores sugieren que los adultos durante el verano se mueven hacia aguas menos profundas, desovan y regresan a las aguas profundas donde se mantienen durante los meses más fríos. Esta especie muestra un marcado tigmotropismo. Workman y Foster (1994) plantearon que la mayoría de los L. campechanus se observan alrededor de estructuras u objetos sobre el fondo, las cuales al parecer son utilizadas como refugio o como punto de orientación. Cuando los juveniles, por ejemplo, se encuentran con un chinchorro camaronero, tienen la tendencia a nadar hacia dentro del mismo, en lugar de tratar de escapar. Rogers (1999) planteó que la complejidad del arte de pesca puede incrementar la retención de los juveniles, debido a que estos tienden a refugiarse en las cavidades, rugosidades y otros escondrijos que forman las redes. Por otra parte, Engaas y Foster (1999) argumentaron que muchos especímenes se posicionan en las áreas de menor flujo de agua y no tratan de escapar, lo cual aumenta su vulnerabilidad. Según Workman y Foster (1994) los L. campechanus tienden a escapar activamente por los orificios de exclusión cuando el flujo de agua se reduce a 0,2 y 0,5 m/seg. Por tanto, recomendaron diseñar mecanismos de exclusión de peces con bajos flujos de agua, que no afecten la producción de camarones. Engaas y Foster (1999), sin embargo, plantearon que el escape de los juveniles de L. campechanus por las aberturas diseñadas para ello, es generalmente bajo y errático durante el rastreo, no así para otras especies. La mayor proporción de estos escapa durante la recogida del chinchorro. 3.62 Gregarismo y conducta social Los adultos y juveniles grandes pueden formar cardúmenes cerca de los hábitats arrecifales (Davis, 1975, Beamuariage y Bullock, 1976) y forrajear de esa forma sobre los fondos blandos (Moseley, 1966; Futch y Bruger, 1976). Los sub-adultos (180 a 300 mm) fueron observados cerca de las plataformas de petróleo en cardúmenes de 30-100 individuos, mientras que los adultos tienden a ser solitarios o formar pequeños grupos (Render, 1995). Dichos cardúmenes tienden a dispersarse al oscurecer. Siendo esta una especie depredadora, la razón más factible para la formación de cardumen es el incremento de habilidad en la obtención de alimento y para reducir la depredación, aunque en el caso de los adultos, pudiera también facilitar la fertilización (Rogers, 1998). Los sub-adultos tienden a la asociación con las plataformas de petróleo en el Golfo de México, pero los adultos no parecen tener igual dependencia de estas (Render, 1995). Rogers (1998) al estudiar la conducta de los juveniles pequeños concluye que estos prefieren las estructuras arrecifales pero son ahuyentados de estas por la conducta agonística de los juveniles de mayor talla y de los adultos. Similares resultados obtuvo Bailey (1995) en condiciones de laboratorio. La conducta agresiva de los adultos parece ser una expresión de dominancia, o puede ocurrir que los peces más grandes se alimenten de los más pequeños (Rogers, 1998). Bailey (1995) observó que los individuos grandes de L. campechanus mantienen a los juveniles de un año de edad apartados de los arrecifes artificiales, con una mínima cantidad de agresiones, aunque no encontró evidencias de canibalismo. Una de las funciones de esta conducta territorial pudiera ser la de mantener alejados a los menor talla para aprovechar mejor los recursos del territorio. 167 En experimentos de laboratorio y de campo con juveniles de menos de un año de edad, Rogers (1998) comprobó que la reducción del flujo de agua, provoca en los juveniles una reacción territorial. Dicha autora también comprobó que el aumento en la densidad, incrementa la conducta agonística de los juveniles. Sin embargo, la actividad agonística disminuye en el tiempo cuando el número de pargos se mantiene constante. Así mismo, la presencia de un arrecife, en ausencia de flujo de agua también incrementa la agresividad, cuando uno o dos peces defienden un territorio. Por otra parte, la conducta territorial es mayor en áreas de reducido flujo que en áreas sin movimiento. En ausencia de refugios, o cuando es excluido de estos por los residentes, L. campechanus forma grupos jerárquicos de dominación (Rogers, 1998). 4. POBLACIÓN 4.1 Estructura 4.11 Proporción de sexos La mayoría de los autores han encontrado una tendencia a la equidad en la proporción de sexos (Tabla 7). Las variaciones locales no muestran una clara regularidad, aunque se requiere de muestreos con una mayor definición de clases de largos o de edades. Tabla 7. Características de la proporción de sexos en relación con la talla o la edad de L. campechanus (Después de Grimes, 1987). Localidad Relación H:M Relación con la longitud Referencias Banco de Campeche 1:0,9 (N = 555) Rango de tallas: 250-800 mm LH, Camber, 1955 1:1 hasta 600 mm después aumenta el % de hembras Norte del Golfo de México 1:1,3 (N = ? ) Rango de tallas: 200-400 mm LE, Moseley, 1966 predominan los machos Noroeste del Golfo de México 1:0,9 (N= 1,129) No hay datos Bradley y Bryan, 1975, South Atlantic Bight y Noreste del 1:0,9 (N= 1,245) Rango de tallas, 230-858 mm LH Collins et al,, Unpub, Golfo de México Golfo de México, Panama City, 1:0,7 (N = 484) En 1992 Collins et al,, 1996 1:1 (N = 649) En 1993 Florida Golfo de México desde Texas 1:1 (N = 1517) Todos con más de 500 mm LT, Collins et al,, 2001, hasta la Florida (1998-1999) Sureste Atlántico y Golfo de 1:1 (N = 733 y Allman et al., 2002b México 8322) Yucatán 1:0,95(N= 1280) Brulé et al., 2002 4.12 Composición por tallas y edades Según Grimes et al. (1977b) en la década del 70 en la pesca recreativa que se realizaba en las Carolinas, se obtenían L. campechanus de 1,9 a 35 libras (0,9 a 16 kg) con longitudes de 13 a 35,7 pulgadas (330-907 mm LT) y el peso promedio era de 17 libras (7,7 kg). Tal peso correspondería a ejemplares de aproximadamente 800 mm LT con edades superiores a los 6 años. En los años 1983-1984 el peso medio en las capturas comerciales era de sólo 1,8-5,3 kg. A partir de 1985 y en especial desde 1991 se observó un incremento (3-4 kg en 1992-1995) como resultado de las regulaciones de talla mínima adoptadas. Similar proceso ocurrió en la pesca recreativa (Manooch et al., 1998). En el norte del Golfo de México, al menos desde 1978 en las capturas comerciales se obtenían ejemplares de 228-890 mm (Fig. 7). En la pesca recreativa, las tallas eran inferiores que en las capturas comerciales siendo la mayoría de los ejemplares de 220 a 330 mm LT y los ejemplares de más de 400 mm eran muy pocos en las capturas (Goodyear y Phares, 1990). 168 Fig. 7. Composición por tallas de L. campechanus en el Golfo de México, en diferentes años. En capturas: AGoodyear y Phares (1990); B – Goodyear (1994); en capturas comerciales y recreativas: C – Fitzhugh et al., (1999). Los cambios en la composición por tallas durante ese período, tanto en las capturas comerciales como en las recreativas de esa región, evidenciaron una reducción en la proporción de los peces más viejos en la población. Aunque la tendencia resultó evidente en todas las regiones, fue mayor en La Florida, Louisiana y Texas que en Alabama y Mississippi (Goodyear, 1994). El establecimiento de una talla mínima de 305 mm en 1984 y otras regulaciones, evidentemente produjo un modesto incremento (Fig. 7A) en las tallas y edades, pero con un impacto positivo significativo al limitar el descenso de la talla media en las capturas (Goodyear y Phares, 1990). En los años siguientes se incrementó por pasos la talla mínima hasta 406 mm en 1998 (ver epígrafe 6.1). En consecuencia, también aumentó la talla media y la proporción de peces de edad avanzada en las capturas (Fig. 7B; Goodyear, 1995). Fitzhugh et al. (1999) reportaron un desplazamiento notable hacia tallas mayores en 1998, con una pequeña proporción de ejemplares con tallas inferiores a 350 mm y con mayor proporción de peces de mediana y gran talla, tanto en las capturas comerciales como en las recreativas (Fig.7C). Los L. campechanus de 3 años de edad fueron dominantes tanto en las capturas comerciales con cordel y anzuelo, como en el sector recreativo y al parecer es la edad 169 de reclutamiento a ambas pesquerías. A partir de los 8 ó 9 años de edad la proporción es generalmente inferior a 1%. En las capturas con palangre sin embargo, se captura una alta proporción de peces de mayor edad y sólo a partir de los 18 años la proporción es menor que 1% por clase de largo. En 2 793 ejemplares obtenidos en la pesca comercial en el norte del Golfo de México en 1995, Nieland y Wilson (2000) encontraron tallas de 320 a 880 mm LH (Fig. 8A). El largo modal fue de 360 mm y el 50% de los peces tenían menos de 420 mm. La edad de estos peces fluctuó de 2 a 48 años, sin embargo, el 97.2% tenían de 3 a 7 años. Entre los peces colectados en 1998 se observó mayor variabilidad de los largos, que en el período 1995-1998. Las edades fluctuaron de 2 a 39 años, con similar proporción por edades, sólo con una mayor proporción de peces de 3 años (Fig. 8B). Entre los peces obtenidos con explosivos en una torre de petróleo el 93% tenían 3 ó menos años de edad (Nieland y Wilson, 2000). Fig. 8. Composición por tallas y edades de L. campechanus en las capturas comerciales del Golfo de México, en diferentes períodos. A y B – tomado de Nieland y Wilson (2000); C y D – tomado de Wilson et al. (2001). Similares resultados reportan Wilson et al. (2001) en la década del 90. De 3 787 ejemplares muestreados en el norte del Golfo de México entre 1989-92 y 1995-98, aproximadamente el 90% tenía menos de 650 mm LT (Figs. 8C, 8D). Aunque en las capturas comerciales y recreativas están representados peces de 250 a 950 mm LT y de 2 a 52 años, la gran mayoría de los peces tenía de 2 a 5 años de edad y sólo el 1,2% tenía más de 15 años. No obstante, la clase modal fue mayor (400 mm) que la reportada por Nieland y Wilson (2000) en la misma zona y años (360 mm), pero sólo en la captura comercial. En la pesca recreativa las tallas fueron mayores, como resultado de la regulación de talla mínima de 14 pulgadas (380 mm) impuesta en 1994. Sin embargo, Fitzhugh et al. (1999) encontraron mayor proporción de peces viejos (más de 20 años), en las capturas comerciales, tanto con palangres como con cordel y anzuelo. Según Allman et al. (2002a) en 1998 los ejemplares de 3 años dominaban las capturas tanto comerciales como recreativas en el norte del Golfo de México. En 1999 los de cuatro años dominaron en la pesca comercial y los de 3-4 años en la recreativa. Sin embargo, la edad de reclutamiento total a la pesca en esos años se estimó en cinco años. La estructura por edades fue similar en el este y el oeste del Golfo, aunque en el oeste era mayor la proporción de peces más 170 viejos. En un análisis posterior Allman et al. (2002b) reportaron una edad media de 4 años en la pesca comercial, tanto en el sureste Atlántico como en el Golfo de México y que los especímenes de la primera región tienden a mayores tallas a una misma edad, que los de la segunda región. 4.2 Abundancia y densidad A juzgar por las capturas comerciales y recreativas (ver epígrafe 6) el pargo L. campechanus es la segunda especie de esta familia con mayor densidad poblacional y abundancia en el Atlántico occidental, después de L. purpureus. Klima (1976) estimó una biomasa total de 65000 TM para el Banco de Campeche y una captura potencial de 6 800 a 13 600 TM, en base a una mortalidad total de 0,2-0,4. Estas cifras, sin embargo parecen ser muy optimistas, ya que las capturas en la zona, aún con un intenso esfuerzo pesquero nunca alcanzaron las 4 000 TM en el área. Monroy García et al. (2002) plantearon que en el Banco de Campeche, es la segunda especie en importancia entre los peces demersales. Dichos autores estimaron que la biomasa de L. campechanus disminuyó de 32 957 TM en 1984 a 16 877 en 1999, es decir en un 51%. También calcularon un rendimiento máximo sostenible (CMS) de 1 271 TM/año. Sin embargo, en 1992 se registró una captura de 3 083 TM, o sea 1 850 TM por encima de la CMS. En los últimos cinco años se estimó una captura promedio de 1 384 TM. Es decir, que en todo el período 19841999 la captura siempre excedió la CMS y en consecuencia la biomasa presenta un fuerte decremento. La abundancia de L. campechanus en las costas de Alabama parece ser muy superior a la existente en todas las otras áreas del Golfo de México y el SE Atlántico, lo cual se deduce a partir de la captura por unidad de esfuerzo (Darnell et al., 1987; Szedlmayer y Shipp, 1994). Strelcheck et al (2005) encontraron que L. campechanus era la especie más abundante en las evaluaciones de CPUE con cordel y anzuelo en las afueras de Alabama, computando el 83,2% de todas las especies de peces. En los censos visuales, esta especie representó el 73,8% de todos los peces observados. Ello se debe en parte, a la gran cantidad de refugios artificiales existente en la zona (más de 6 000), a pesar de que la zona tiene pocas áreas cubiertas por estructuras arrecifales naturales y prevalecen vastas áreas de arena y fondos blandos (Shultz et al., 1987). No obstante, Szedlmayer y Shipp (1994) concluyeron que la alta productividad pesquera en aguas de Alabama, no es resultado de un sólo factor, sino de la combinación de varios, tales como el rápido crecimiento de la especie, tallas y edades mayores, alta residencia y una abundancia mayor que en otras áreas. La poca existencia de hábitats arrecifales pero con un número extremadamente alto de refugios artificiales evidencian no sólo un mecanismo simple de atracción, sino más bien de alta productividad. 4.3 Natalidad y reclutamiento 4.31 Tasas de reproducción Debido a que el pargo L. campechanus es un desovador parcial, el cálculo de la fecundidad es poco exacto, ya que por lo general sólo pueden ser contados los oocitos vitelógenos. No obstante, Collins et al. (1996; 2001) utilizaron el método propuesto por Hunter y Macewics (1985) para calcular la fecundidad annual (AFE), mediante una estimación previa del número de porciones (SFE) y a partir de la fecundidad porcional (BFE) estimada por el conteo de oocitos hidratados en las gónadas maduras. Asimismo, dichos autores establecieron la correlación entre la fecundidad porcional y la fecundidad annual (AFE = (5,401 * AFE previa) + 2,054*105); r2 = 0,956), con la cual es posible recalcular los estimados previos de otros autores, asumiendo que la frecuencia de desoves es estable. En los muestreos realizados en 1998 a lo largo de todo el norte del Golfo, Collins et al. (2001) encontraron valores extremos de fecundidad parcial desde 13 (en un ejemplar de 417 mm 171 LT y 4 años de edad) hasta 3,4 millones (en un pez de 851 mm LT y 11 años de edad). Tanto el largo como la edad mostraron una relación exponencial con la fecundidad parcial, pero la edad resultó mejor para el pronóstico (Tabla 8). Según los mencionados autores, las hembras de edades entre 6 y 35 años desovan aproximadamente un 50% más de veces que las de edades de 3 a 5 años. Goodyear (1994) sin embargo, estimó el valor reproductivo relativo de cada clase anual de L. campechanus, en base al aporte de cada clase a la fecundidad total (Fig. 9). Sus datos muestran que, aunque algunas hembras maduran desde los 3-4 años de edad, su contribución individual al desove anual es muy pequeña, comparado con los peces de más de 7-8 años de edad. Las hembras de 11 años producen mayor número de huevos que cualquier otra clase anual. Los datos presentados por el autor reflejan cierta senescencia en las edades mayores. Tabla 8. Fecundidad individual y relativa de Lutjanus campechanus en el Golfo de México, LH – largo horquilla; LT – largo total; P – peso total del pez; t – edad (años), N 66 59 Rango de Tallas, Pesos o edades No, de oocitos x 103 (min-max) 420605LH 110012485 g 349-820 LT 600-9100 g _?-12 años 359-901 LT 770-8080 g 4-11 años BFE= 44-9320 AFE=25850543* BFE=0,0133400 AFE=20599180* 506 LH (237-916) 916 509 LH (282-913) BFE= 0,7-1903 BFE=1,42236-7980 Relación F/Talla , peso o edad Localidad Referencias South Atlantic Bight y Gulf of México Collins et al,, 1987 BFE= (1,949 x 103LT) – 8,055 x 105 BFE= (1,363 x 105 P) – 1,315 x 105 BEF=(1,278 x 105(t) – 4,375 x 105 SE Golfo de México Collins et al,, 1996 BFE= (8,0 x 10-6LT) + (2,2x10-3LT) EU Golfo de México Collins et al,, 2001 F = 1,02x10-7 LT5,87; LT (en pulg,) Golfo de México, EUA Alabama Goodyear, 1987 Woods et al., 2003 234-4048* BFE=0,4581700 148 1020 F relativa Prom. (No. oocitos/g peso) 0,262275* BFE = 753,63 (Log edad) – 776,63 (R2 = 0,52) BFE = 3,8049 (Log LH) – 11,6; (R2 = 0,407) BFE = 936,14 (Log edad) – 976,44 (R2 = 0,58) BFE = 5,7478 (Log LH) – 24,5; (R2 = 0,687) Louisiana BFE = Fecundidad porcional estimada (Hunter y Macewics, 1985); AFE – Fecundidad anual estimada; * – AFE estimada por nosotros a partir de la ecuación: AFE = (5,401x AFE previa) + 2,054x105; Collins et al, 1996). Woods et al. (2003) demostraron la existencia de diferencias demográficas entre las poblaciones de L. campechanus en Alabama y Louisiana (Tabla 8) tanto en cuanto a la fecundidad porcional (BEF) como en la frecuencia de desoves (SFE = 3,4 y 4,2 respectivamente, equivalentes a 44 y 36 desoves). El Índice de Importancia Reproductiva (IRI) contra la edad, indicó que los especímenes de 4 años de edad en Alabama tienen una mayor contribución a la población de desove que los de Louisiana, mientras que en esta última localidad los de 8 años tienen mayor contribución que los de Alabama. Concluyeron dichos autores que tales diferencias pueden ser debidas a diferencias ambientales, aunque con mayor probabilidad son consecuencia de una compensación al incremento de la mortalidad por mayor presión pesquera en Alabama. 172 Fig. 9. Valor relativo del aporte a la fecundidad total por clases de largo de L. campechanus (tomado de Goodyear, 1994). 4.32 Reclutamiento Según Bradley y Bryan (1975), debido al prolongado período de reproducción, el reclutamiento a la pesca se produce durante casi todo el ciclo anual. Según ellos, la menor clase fue de 210230 mm LH. Los datos de Nelson y Manooch (1982) mostraron que la edad de reclutamiento total a la pesca a fines de la década de los 70s, fluctuaba de 2 años en Louisiana a 6 años en las Carolinas. La edad media en la captura total fue de 2,4 años en Louisiana y de 4,1 años en el oeste de la Florida. Tal diferencia se debió a una mayor presión pesquera en el área de Louisiana. En el sureste Atlántico Manooch et al. (1998) observaron un incremento en el rendimiento por recluta entre los períodos 1986-1991 y 1992-1995, evidentemente resultado del establecimiento de una talla mínima de captura. Goodyear y Phares (1990) en base a las frecuencias de tallas en las capturas en la década del 80, estimaron que L. campechanus en el Golfo de México de EU era completamente reclutado a la pesca a las 10 pulgadas (220 mm) LT, que correspondería con una edad inferior a los dos años. Fitzhugh et al., (1999), en muestreos realizados en 1998, estimaron que la edad de reclutamiento total era de 3 años (aproximadamente 350 mm). Esta diferencia probablemente fue resultado de la implantación de la talla mínima de captura y su incremento posterior hasta llegar a 16 pulgadas (352 mm) en 1998. Según Goodyear (1994) los datos del monitoreo que realiza desde 1972 el Laboratorio de Pascagoula del National Marine Fisheries Council (NMFC) en el norte del Golfo de México, sobre la captura de juveniles (41-190 g de peso) con redes de arrastre, indican una disminución notable de la abundancia y por tanto del reclutamiento a mediados de 1980, que alcanzó su valor más bajo en 1987, aunque con un notable incremento en 1990. De acuerdo con los datos de Goodyear y Phares (1990), existe una fuerte relación estadística entre los indicadores de reclutamiento de los años 1978-1987 y la captura total estimada de las clases de 254 y 279 mm por la pesca recreativa. Por tanto, los datos de dicho monitoreo pueden ser empleados para predecir el reclutamiento a la pesca al año siguiente (Goodyear, 1994). Debe considerarse que en tales muestreos están presentes dos clases de edad (0 y 1 año) por lo que es necesario descomponer la captura por clases de edad para hacer el pronóstico. Schirripa (2000) también encontró una correlación negativa entre la proporción de juveniles en los arrastres de los camaroneros y las capturas en los años siguientes. 173 Patterson et al. (1998) utilizaron los otolitos de juveniles de edad 0+ para discriminar su área de reclutamiento y cría a partir de la composición microquímica de los mismos. Diez de los 12 elementos detectados en los otolitos mostraron diferencias significativas entre las tres áreas de cría analizadas, aunque sólo ocho (Al, As, Ca, Fe, Mg, Mn, Se y Zn) fueron empleadas en el análisis multivariado para identificar su afinidad con las áreas, lo cual se logró con un 90% de confiabilidad. Las investigaciones actuales se orientan a la microquímica de los otolitos de los pargos adultos en los arrecifes exteriores para determinar, de forma retrospectiva, el área de reclutamiento y cría original. 4.4 Mortalidad y morbilidad 4.41 Tasas de mortalidad La Tabla 9 resume los datos de la literatura a nuestro alcance sobre las tasas de mortalidad de L. campechanus. Nelson y Manooch (1982) encontraron notables diferencias en las tasas instantáneas de mortalidad total en diferentes regiones del Golfo de México. La mortalidad por pesca parece ser la principal causa de la diferencia en la mortalidad total entre áreas, ya que la mortalidad natural es muy similar en todos los casos. El valor de F fue mayor en Louisiana y el este de la Florida, en parte debido a que los arrecifes, donde se concentra la pesca, están más cerca de la costa y por tanto más accesibles a la pesca comercial y recreativa, lo cual se manifiesta en valores más altos del coeficiente de explotación (E). Las áreas ubicadas al oeste de la Florida y en las Carolinas, están más alejadas de la costa y por tanto menos accesibles, por lo que están sometidas a menor presión de pesca. Dichos autores consideran que las diferencias significativas en la mortalidad y estructura por edades de las poblaciones de las cuatro áreas de pesca de L. campechanus en EU, tiene mayor impacto sobre los modelos de manejo que las diferencias, también significativas, en el crecimiento. Nelson y Manooch (1982) estimaron la mortalidad natural de L. campechanus entre 0,18 y 0,20, utilizando la ecuación de Pauly (1980; Tabla 9). Tabla 9. Valores de mortalidad total (Z), mortalidad natural (M) y por pesca de L. campechanus en varios ecosistemas del Gran Caribe. Error estándar entre paréntesis, Localidad Z M F E (F/Z) Observaciones Referencias 3 2 0,58 0, 74* Datos de 1978- Nelson y Manooch, Louisiana 0,78 (0,11) 0,20 79 1982 0,3841* 0,23 0,55* 1978-79 Oeste de la Florida 0,423 (0,05) 0,192 0,3621* 3 Este de la Florida 0,50 (0,07) 0,31 0,62* 1976-78 0,192 0,3241* Carolinas 0,393 (0,05) 0,54* 1974-78 0,182 0,3491* 0,15-0,30 0,31-0,48 1986-1991 Manooch et al., SE Atlántico 0,483 0,763 0,50-0,69 1992-1995 1998 0,769 1982-91 Goodyear, 1994 Norte del Golfo de 0,877 0,15 0,2231* México 4 Yucatán 0,44 González et al, 0,31462 0,1851 0,1653 1984 0,25495 1994, 0,2691* 4 0,31462 0,1777 0,1414 1985 Yucatán 0,4751 0,29745 0,2741 1 M calculada por la ecuación de Ricker: M= -0,0666+2,52 K (Ralston, 1987); 2 M calculada por la ecuación de Pauly (1980): ln m = -0,0152 -0,279 ln l∞ + 0,6543 ln k + 0,463 ln tº; 3 Valores de Z calculados por la curva de captura (Ricker, 1975); 4 Calculada por ecuación de Gulland, 1969; 5 M calculada por la ecuación de ArreguínSánchez y Chávez, 1985; *valores calculados por nosotros a partir de los datos originales 174 Manooch et al. (1998) utilizando diferentes métodos estimaron valores de 0,15 a 0,30 en el sureste Atlántico en dos períodos posteriores: 1986-1991 y 1992-1995. Consideran dichos autores que el valor más confiable se encuentra entre 0,20 y 0,30. También calcularon mortalidades totales mayores en esos períodos que para los años 1978-1979. Los valores promedios de mortalidad por pesca resultaron mayores en el período 1992-1995 que en 1986-1991, debido evidentemente a que en el primero las capturas se basaban en peces de 1 a 12 años, mientras que en el segundo periodo eran de 6 a 12 años, gracias al establecimiento de una talla mínima de captura. Para fines comparativos, Goodyear (1994) sugirió utilizar M = 0,15. Las estimaciones a partir de la ecuación de Ricker (Ralston, 1987) en base a los valores de K en la Tabla 9, resultan más altas que las estimadas por Nelson y Manooch (1982) y Goodyear (1994) y menores que las calculadas por González et al. (1994) para Yucatán. No obstante, debido a la gran reducción de los stocks en el norte del Golfo de México, la precisión en el valor de M no altera mucho las estimaciones que se realizan para el manejo (Goodyear, 1994). Los resultados obtenidos por Goodyear (1988) mediante técnicas de simulación, a partir de la composición por edades en el periodo 1984-86, evidencian que la mortalidad por pesca aumenta rápidamente a medida que los juveniles son reclutados a la pesca, alcanzando a los tres años de edad un valor de 0,75 y disminuyendo después hasta 0,3 a los ocho o nueve años de edad, para una mortalidad natural M = 0,2. En una posterior evaluación, Goodyear (1994) también mediante técnica de VPA estimó la mortalidad en la captura incidental de camarones para el periodo 1982-1993, asumiendo una mortalidad natural de 0,5 para la edad 0 y 0,3 para la edad 1. Se observó un claro incremento de la mortalidad acumulativa (F) con el incremento del esfuerzo de los camaroneros. La mortalidad media acumulativa, provocada por la captura de juveniles en los arrastres camaroneros, para el periodo 1984-1990 fue de F = 1,79 que implica una probabilidad de supervivencia de 0,17 Los estimados de mortalidad acumulativa fluctuaron de 1,97 para la clase anual de 1982 a 1,65 para la clase de 1985. Si la capturabilidad de los peces de edad 1+ es la misma en los muestreos de verano y otoño, la relación entre sus abundancias relativas brinda un estimado de la supervivencia. Así, para el periodo 1982-1991 se estimó un valor promedio de 0,42, que corresponde con una mortalidad instantánea total de 0,853. Mediante la acumulación de las medías mensuales de junio a noviembre, se obtiene un valor muy semejante de F = 0,769 y Z = 0,877 (Goodyear, 1994). González de la Rosa et al. (1994) obtuvieron valores bajos de mortalidad total y por pesca en la Península de Yucatán (Tabla 9) con los cuales estimaron una tasa de explotación de sólo un 15% de la población, por lo que asumieron que el recurso estaba sub-explotado en los años 1984-1985. 4.42 Factores que afectan la mortalidad Ya desde la década de los 70s, Bradley y Bryan (1975) y posteriormente en la de los 90s Goodyear (1994) demostraron una clara relación entre el incremento de la flota camaronera, las capturas incidentales de pargo L. campechanus en los chinchorros y el decrecimiento de la captura con cordel y anzuelos (Figs. 10 y 11). Esa captura incidental sobre los peces juveniles es responsable de una alta proporción de la mortalidad y por tanto uno de los principales factores que mantienen la situación de sobrepesca de L. campechanus en el Golfo de México (Schirripa, 1999; 2000).Ya desde que alcanzan 50 mm LT son capturados por dicho arte (Szedlmayer y Shipp, 1994; Goodyear, 1995) aunque no son completamente vulnerables hasta que alcanzan 100 mm LT ó más (Goodyear, 1995; Gallaway y Cole, 1999b). La frecuente demolición de plataformas de petróleo en el norte del Golfo es una fuente potencial de mortalidad para esta 175 especie, aunque Gitschlag et al. (2003) demostraron que la misma es insignificante en comparación con la causada por los arrastres camaroneros. Aunque en menor medida, la pesca comercial con cordel y anzuelo también provoca mortalidad de peces juveniles (tallas inferiores a la mínima legal). Tales peces son devueltos al mar, pero aproximadamente el 64% están ya muertos o en muy mal estado (Baker et al., 2002). La vulnerabilidad de esta especie a la pesca se incrementa en los meses de invierno, cuando los peces permanecen con poco movimiento en hábitats arrecifales (Duffy, 1970). Fig. 10. Capturas de L. campechanus con cordel y anzuelos (A); capturas incidentales en los chinchorros camaroneros (B); incremento de la flota camaronera (C) y captura por unidad de esfuerzo con cordel y anzuelo (D) en aguas de Texas, de 1950 a 1971 (datos de Bradley y Bryan, 1975). Fig. 11. Capturas incidentales de L. campechanus (A), esfuerzo (B) y captura por unidad de esfuerzo (C) de L. campechanus en los chinchorros camaroneros del Norte del Golfo de México (modificado de Goodyear, 1994). 4.43 Parásitos, enfermedades y anormalidades 176 Varios parásitos han sido reportados en L. campechanus. Williams (1979) reportó la presencia de cuatro ejemplares de Tracheolobdella lubrica en las branquias de un ejemplar capturado en Mobile Bay, Golfo de México. Suarez-Morales y Ju-shey (1994) reportaron la presencia del copépodo parásito Lernaeolophus sultanus en los arcos branquiales de dos especímenes de L. campechanus capturados en la porción sur del Golfo de México, en las afueras de Veracruz. 4.44 Biotoxicidad Los especímenes de L. campechanus capturados en las cercanías de los pozos de petróleo no mostraron señales de contaminación en sus tejidos, aunque otras 13 especies del área lo estaban (Middleditch et al., 1979). Scott et al. (1980) tampoco encontraron evidencias de efectos tóxicos en los testes de cinco ejemplares capturados en dichas áreas. Al parecer la concentración de hidrocarburos clorinados en la carne de los peces esta correlacionada con el contenido de grasas en estos. Stout (1980) encontró menores concentraciones en los músculos de L. campechanus que en especies con niveles de lípidos superiores a 3%. En los músculos de esta especie dicho autor encontró como promedio 0,039 ppm de DDT y sus metabolitos, cifra muy inferior al máximo legal en Estados Unidos – 5 ppm. Sólo en uno de nueve ejemplares se encontró un nivel detectable de dieldrín y endrín. No se han reportado casos de toxicidad por ciguatera en L. campechanus. 4.5 Dinámica de las poblaciones Las grandes diferencias en la longevidad de L. campechanus entre el sudeste Atlántico y el norte del Golfo de México, resultan en notables diferencias en los estimados de mortalidad natural y por pesca en ambos ecosistemas. Similar diferencia parece ocurrir con la población del Golfo de Campeche. En el sudeste Atlántico, aunque se mantiene la tendencia al decrecimiento de las capturas y los rendimientos, los datos de Manooch et al. (1998) indican que el establecimiento de una talla mínima de captura provocó un efecto positivo al incrementar el rendimiento por recluta, la talla media en la captura y un aumento importante de la Tasa Potencial de Reproducción (TPR). Dichos autores concluyen que las poblaciones de L. campechanus en la región se encuentran en etapa de transición. Es decir, que su estado no es el deseable pero parece responder positivamente a las acciones de manejo. En el norte del Golfo de México la población de L. campechanus esta sometida a otros impactos. El reclutamiento de juveniles presenta una tendencia a la reducción desde 1970, aunque con grandes fluctuaciones. La mortalidad provocada por la pesca incidental de los arrastres camaroneros reduce en aproximadamente 82% la supervivencia a la pesca directa (Goodyear, 1994). La mortalidad por pesca directa también resulta demasiado alta, todo lo cual determina la situación de sobrepesca existente en el norte del Golfo de México. González de la Rosa et al. (1994) en el Golfo de Campeche estimaron niveles máximos de rendimiento por recluta de 954g con una edad de primera captura de cuatro años, para F(0,1) = 0,37. Tales rendimientos coinciden con el mayor nivel de incremento de biomasa individual. Estos datos sugirieron la posibilidad de incrementar la tasa de explotación aunque, precisaron dichos autores, se requiere de un estudio mas detallado sobre el esfuerzo pesquero. Gold et al. (1994, 1997) y Gold y Richardson (1998) mediante el análisis de ADN mitocondrial, encontraron una notable homogeneidad genética entre los L. campechanus muestreados en ocho localidades del norte del Golfo y en aguas del Banco de Campeche. Estos resultados evidencian la existencia de una sola población, sin barreras genéticas. Varios autores coinciden con el criterio de que en el norte del Golfo existe una población relativamente homogénea de L. campechanus (Goodyear, 1995: Patterson et al., 2001c). Gold 177 et al. (1994, 1997) y Gold y Richardson (1998) opinan que la naturaleza residente tanto de los juveniles como de los adultos, sugiere que el intercambio genético fluye a través del transporte hidrodinámico de los huevos y larvas. Heist y Gold (2000), aunque apoyan la hipótesis anterior, plantean que aún existen algunas dudas y que para definir con seguridad la estructura de la población en el norte del Golfo de México se requiere de muestreos más amplios e investigar las características de la reproducción y el reclutamiento a nivel local. Saillant y Gold (2002) plantearon que algunos resultados sugieren una divergencia genética significativa entre el L. campechanus de la parte noroccidental y el de la parte nororiental del golfo, aunque no hay suficiente consistencia en los resultados. Los estudios de marcaciones y de genética poblacional también muestran que hay una mezcla entre poblaciones adyacentes en esa región del Golfo. Se sabe que los juveniles son relativamente estacionarios, pero los adultos se encuentran más alejados de la costa y realizan notables migraciones (Beaumariage, 1969; Goodyear, 1995). Patterson et al. (2001b) detectaron notables migraciones entre los adultos y sugirieron que los frecuentes huracanes en el Golfo de México juegan un importante papel en la dispersión y mezcla de las poblaciones de L. campechanus. Debido a que estos peces pueden alcanzar hasta 50 o más años de edad, están expuestos al efecto de varios huracanes durante su vida (Patterson et al., 2001b; Render, 1955; Waterson et al., 1998). Sin embargo, Arreguín-Sánchez y Manickchand-Heileman (1998) a partir de modelos tróficos de balance de masas (empleando ECOPATH) y simulando perturbaciones con crecientes niveles de mortalidad, sugirieron la existencia de más de una población de L. campechanus en el sur del Golfo de México. Dichos autores encontraron diferencias en las respuestas ante crecientes mortalidades por pesca, entre las dos poblaciones residentes en el Banco de Campeche: la de Yucatán y la del suroeste del Golfo. Concluyen dichos autores que desde el punto de vista pesquero los pargos de esos dos ecosistemas deben ser manejados como poblaciones discretas e independientes. 4.6 La población en la comunidad y en el ecosistema 4.61 Competidores Es muy pobre la información concreta sobre este aspecto en L. campechanus. Probablemente los juveniles son excluidos de las aguas interiores de algunas localidades por Lutjanus griseus (Smith, 1976). Todos los depredadores piscívoros que habitan en el mismo ecosistema son potenciales competidores de L. campechanus, incluso los tiburones. 4.62 Depredadores Bradley y Bryan (1976) encontraron un L. campechanus juvenil en el estómago de un por pez lagarto (Synonontidae) y otro de 340 mm en el estómago de un Coryphaena hippurus. Los tiburones parecen ser importantes depredadores de L. campechanus. Bradley y Bryan (1976) reportan que a veces estos son atacados por tiburones durante la pesca con cordel y anzuelos. Señala dicho autor que cuando los tiburones abundan en un área, disminuye la captura de L. campechanus. 4.63 Interrelaciones en la comunidad L. campechanus puede ser considerado como un depredador tope en el ecosistema. Debido a su abundancia y actividad de depredación puede ser un importante competidor por los recursos del ecosistema e incluso excluir o reducir la presencia de otras especies. El decrecimiento de sus poblaciones por tanto también puede provocar serios cambios en la estructura de las comunidades. Arreguín-Sánchez y Manickchand-Heileman (1998) a partir de modelos tróficos de balance de masas (empleando el modelo ECOPATH) calcularon niveles tróficos de 4,2 y 4,6 para las 178 poblaciones del suroeste del Golfo de México y el norte de Yucatán, así como tasas de producción / biomasa de 0,35 y 0,46 y tasas de consumo / biomasa de 1,075 y 1,418 respectivamente. La composición de las dietas de L. campechanus en ambos ecosistemas muestra aproximadamente la misma proporción pero con un espectro más alto de presas bentónicas en el norte de Yucatán, probablemente como consecuencia de diferencias estructurales entre ambos ecosistemas. La magnitud de varios indicadores tróficos indica que las relaciones tróficas de L. campechanus en el suroeste del Golfo son más complejas que en Yucatán. Concluyen dichos autores que los impactos de las perturbaciones simuladas sobre la mortalidad de ambas poblaciones se propagaron a través de la trama alimentaria con mayor nivel de impacto en el ecosistema del suroeste del Golfo que en Yucatán. El primero muestra mayor estabilidad a las perturbaciones por pesca sobre la población de esta especie. 5. EXPLOTACIÓN L. campechanus se considera como la especie más apreciada del complejo pargos-meros a lo largo de la costa SE y el Golfo de México de los Estados Unidos (Manooch y Potts, 1997) y es también una de las especies más importantes en el Banco de Campeche, razón por la cual ha sido sometida a una intensa explotación pesquera al menos durante los últimos 100 años. 5.1 Artes de pesca En el Golfo de México y el sureste de la Florida, las capturas comerciales de esta especie tradicionalmente se han realizado con líneas (cordel y anzuelos encarnados), levadas a mano o con winches eléctricos o hidráulicos (de 2 a 40 anzuelos por línea) y en pequeña proporción, con palangres de fondo, sobre todo en la década del 80. Las capturas con este ultimo arte alcanzaron su máximo en 1984 (514 TM), para disminuir hasta casi desaparecer en 1994 (Goodyear, 1994; Ehrhardt, 1996) debido al establecimiento en 1990 de una regulación prohibiendo su uso en aguas de menos de 50 brazas (90 m). Una pequeña proporción se obtenía con chinchorros para peces aunque esta pesca fue disminuyendo hasta su total desaparición en 1990. El 99% de la captura con chinchorros se obtiene como parte de la captura incidental de camarones. Las capturas con nasas son prácticamente nulas Goodyear (1994) planteó que la gran mayoría de la captura se obtiene con cordel y anzuelo y la CPUE con dicho arte depende fundamentalmente de la rapidez con que el arte se tira y recoge. Por otra parte, debido a la conducta territorial del L. campechanus, dicho arte puede provocar una mortalidad por pesca muy alta. Por tanto, considera que la CPUE con dicho arte no es un buen indicador del tamaño del stock. Se utilizan varios tipos de carnadas, entre las cuales las más comunes son los calamares y el banano (Elops saurus) y en menor medida: sierra (Scomberomorus spp), cojinúa (Caranx crysos), lisas, (Mugil spp) sardinas (menhaden) y camarones (Futch y Torpey, 1966). En Carolina del Sur en la década del 70 se capturaba con trawl y cordel y anzuelo. En las capturas con trawl L. campechanus constituía el 38% de la captura (Ulrich et al, 1977). En la pesca con palangres las tallas eran mayores, pero con la misma tendencia a la disminución, que en las capturas con cordel y anzuelo. Goodyear y Phares (1990) observaron que con este último tipo de arte, hay una tendencia a capturar peces más pequeños que cuando se usa el carrete eléctrico o winche hidráulico. En el Banco de Campeche se utilizan tres métodos principales: alijos, carretes de línea larga y bicicleta. Este último arte produce el mayor CPUE (366 kg/viaje de pesca). El 12% de la flota yucateca utiliza este arte y captura el 45% del total de L. campechanus (Monroy et al., 2002b). 5.2 Áreas de Pesca 179 L. campechanus se pesca a todo lo largo de su área de distribución, aunque las principales zonas de pesca se ubican en el norte del Golfo de México y en el Banco de Campeche (Fig. 2). Esta última región fue la principal zona de pesca de la flota del oeste de la Florida hasta la década del 50 (Camber, 1955) y sigue siendo la principal para los Estados Unidos de México. También fue relativamente importante para la flota Cubana en las décadas del 70 y 80. Con el establecimiento de la zona económica exclusiva de 200 millas por México, la flota de Estados Unidos inició su retiro de esa región y la de Cuba siguió pescando por convenio desde 1976, conforme a una cuota límite y fuera de las 12 millas de aguas territoriales. Ya desde 1973 más del 87% de las capturas de esta especie por la flota de EUA se obtenía en aguas nacionales (Moran y Morais, 1988) y actualmente casi el 100% (Fig. 12). Las principales zonas de pesca comercial de ese país se localizan en el norte del Golfo de México entre Panama City, Florida y Galveston, Texas. Los mayores volúmenes de captura se obtienen en la región de Louisiana (Moran y Morais, 1987; Goodyear, 1990; Goodyear, 1994), pero al parecer en aguas de Alabama la productividad es mayor (Szedlmayer y Shipp. 1994). En el norte del Golfo se obtiene más del 90% de la captura total de EUA y menos del 10% en el este de la Florida (Keihly y Roberts, 1999). De esta última (desde Cabo Hatteras hasta Key West), la mayor parte de las capturas se obtienen al norte de La Florida y en las Carolinas (Parker y Mays, 1998). Tanto la pesca comercial como la recreativa centran su actividad pesquera en los arrecifes exteriores del Golfo, a profundidades de 40 a 110 m, preferentemente alrededor de los 90 m. En aguas de las Carolinas parece ser más abundante entre 50 y 90 m (Grimes et al., 1977b). Fig. 12. Capturas comerciales de L. campechanus por EUA en el N del Golfo de México y en el Banco de Campeche (elaborado a partir de los datos de Goodyear, 1994 y Fisheries of the United States, 1975-2001). 5.3 Épocas de pesca Moe (1963) y Duffy (1970) indicaron que las mayores capturas de L. campechanus se obtenían en los meses de invierno, en las aguas más profundas, donde los peces se mantienen con poco movimiento. Bradley y Bryan (1975) sin embargo, plantearon que en aguas de Texas las mayores producciones se obtienían en Marzo-Abril, al parecer debido al aumento de la actividad alimentaria antes de la reproducción y posteriormente otro pico en agosto, que pudiera ser reflejo de otro incremento de la actividad alimentaria después del desove, sumado al incremento 180 de la población por el reclutamiento a la pesca de la generación nacida el año anterior. Las capturas más bajas se obtienen en invierno, aunque la productividad (CPUE) es mayor en ese periodo. Bradley y Bryan (1975) afirman que durante el verano los peces más jóvenes son capturados en aguas someras y a medida que la estación progresa, los peces se mueven a aguas más profundas. Según datos más recientes (Goodyear, 1994), L. campechanus se captura durante todo el año en el Golfo de México, aunque a partir de 1984 se observó una tendencia a la disminución de las capturas en los meses invernales. Ello esta determinado por las capturas con cordel y anzuelo, que son las predominantes, las cuales tienen su mínimo en diciembreenero, por las inclemencias del tiempo. La pesca recreativa, realiza su mayores capturas en los meses de junio a octubre y su valor mínimo en invierno (Goodyear, 1994). En el Banco de Campeche, Monroy et al. (2002b) reportaron los rendimientos más altos de enero a marzo y los valores más bajos de agosto a octubre. 5.4 Operaciones pesqueras y resultados 5.41 Esfuerzo pesquero El esfuerzo pesquero sobre L. campechanus se incrementó rápidamente desde inicios del siglo 19 y provocó, ya desde la década de los 60s una declinación de los rendimientos y las capturas (Bullis y Jones, 1976). Desde la década del 50 se produjo un incremento del esfuerzo pesquero de los barcos camaroneros en el Golfo de México y como consecuencia, ocurrió un incremento en las capturas incidentales de L. campechanus con ese arte y se produjo una disminución en las capturas y los rendimientos de la pesca con cordel y anzuelo (Figs. 10 y 11; Bradley y Bryan, 1975; Goodyear, 1994). Desde la década de los 60s se incrementó rápidamente el esfuerzo de la pesca recreativa (en más de un 35% de 1965 a 1975), que parece ser una de las causas de la disminución de las capturas comerciales en la Florida y los estados del Golfo durante los 70s (Futch y Bruger, 1976). En general, en el norte del Golfo de México el esfuerzo pesquero directo se mantiene por encima del nivel aceptable (Goodyear, 1994), lo cual es una de las causas de la situación de sobrepesca de ese recurso. 5.42 Selectividad Las capturas tanto comerciales como recreativas de L. campechanus se realizan principalmente con cordel y anzuelo, artes que se consideran bastante selectivos. Por otra parte el establecimiento de tallas mínimas legales relativamente altas, al menos en EUA, obliga al uso de anzuelos de gran tamaño, ya que los anzuelos menores pueden capturar tallas juveniles. Baker et al. (2002) reportaron que en la pesca con multi-anzuelo Abandit, el 17,3% de la captura en peso es descartada por tallas inferiores a la mínima. Los ejemplares descartados tenían una talla media de 316 mm LH y dos años de edad. Por otra parte, resulta inevitable la captura incidental de juveniles por los chinchorros camaroneros, aún con el establecimiento de mecanismos de selección, para reducir la mortalidad de los peces jóvenes. El Servicio Nacional de Pesquerías Marinas (NMFS) de la NOAA trabaja desde hace años en el desarrollo de modificaciones de las redes de arrastre camaroneras para reducir la pesca incidental de juveniles de L. campechanus (Workman y Foster, 1994; Render y Wilson, 1996; Schirripa, 1999). Los mecanismos de reducción de la captura incidental (Bycatch Reduccion Device -BRD) son aditamentos instalados en la parte superior del chinchorro camaronero, que facilitan el escape de los peces, reteniendo la captura de camarones. En mayo de 1998 entró en vigor una resolución elaborada por el NMFS que establece la obligatoriedad del uso de aditamentos de reducción de la BRD en los chinchorros camaroneros, para reducir la mortalidad 181 de los L. campechanus juveniles. El NMFS aprobó el uso de tres tipos de BRD: 1- Gulf Fisheye BRD, 2- el Jones Davis BRD y 3- el Fisheye BRD (Fletcher, 1998). Workman y Foster (1994), considerando la conducta de L. campechanus ante la presencia de los chinchorros, recomendaron crear mecanismos de escape en los artes que faciliten la salida de los juveniles una vez dentro de este. Según dichos autores los juveniles de L. campechanus tienden a escapar activamente por los orificios de exclusión cuando el flujo de agua se reduce a 0,2 y 0,5 m/seg. Por tanto, recomendaron diseñar mecanismos de exclusión de peces con bajos flujos de agua, que no afecten la producción de camarones. Sin embargo, Rogers (1999) planteó que la eficiencia de los mecanismos de selección de los chinchorros camaroneros, es limitada por algunos factores conductuales. Por ejemplo, los paneles de los BDR que deben facilitar el escape de los juveniles disminuyen el flujo de agua en algunas partes del arte, provocando una reacción territorial de algunos juveniles que se ubican en ese lugar y atacan a los que se acercan, limitando el acceso de estos a los orificios de escape. Por otra parte, según dicha autora, la tendencia de los juveniles a refugiarse en las cavidades, pliegues y cualquier estructura que semeje un escondrijo, aumenta la retención de estos dentro del chinchorro, por lo que recomendó diseños simples que no brinden refugios en el interior del arte, tales como el sistema de exclusión Fisheye. Engaas y Foster (1999), sin embargo, plantearon que el escape de los juveniles de L. campechanus por las aberturas diseñadas para ello, es generalmente bajo y errático durante el rastreo, no así para otras especies. La mayor proporción de estos escapa durante la recogida del chinchorro. Watson et al., (1999) plantearon que los aditamentos experimentados (Jones Davis y Fisheye 2,6 y 3,8 m) logran reducciones de la tasa de mortalidad entre 52 y 70%. Sin embargo, Gallaway y Cole (1999b) plantearon que el máximo potencial de exclusión de juveniles de L. campechanus con el uso de dichos mecanismos de reducción de la captura incidental es de sólo 25-27% y no el 50% (de las edades 0+ y 1 año) estimado para lograr la recuperación de las poblaciones. Independientemente de estos datos, la aplicación de estas regulaciones ha sido tema de litigio por las asociaciones de pescadores de camarones, quienes cuestionan la efectividad de la medida (Wilson y Fletcher, 1998). Por otra parte, aun no existe suficiente información sobre la supervivencia de los juveniles que logran escapar del arte, por lo cual resulta difícil predecir la eficiencia real de tales medidas, ni se conoce aún hasta que punto los cambios ocurridos en los hábitas han contribuido a la reducción de las poblaciones de L. campechanus y pudieran limitar la recuperación hasta los niveles históricos (Gallaway y Cole, 1999b). 5.43 Capturas L. campechanus es una de las especies de mayor importancia comercial entre los pargos del Atlántico occidental por el volumen de sus capturas, sólo superada por L. purpureus. En el norte del Golfo de México ha sido tradicionalmente la especie que más contribuye a las pesquerías demersales (Carpenter, 1965; Klima, 1976; Cato y Prochaska, 1978) y la segunda en importancia en el Banco de Campeche (González et al., 1994). Tomando en consideración los desembarques en EUA (SE Atlántico y Golfo de México) más los de Campeche, en el período 1987 a 1997 las capturas totales de L. campechanus fluctuaron de 7 000 a 11 400 TM, con un promedio anual de 8 500 TM aproximadamente (FAO, 2002; Fisheries of the United States, 1985-2001). Estados Unidos de América (EUA). Los datos de desembarques presentados por varios autores muestran una clara tendencia al decrecimiento de las capturas y ofrecen suficientes elementos para considerar a la especie como fuertemente sobre-explotada (Goodyear, 1995; 182 Ehrhardt y Legault, 1996; Render y Wilson, 1996; Keihly y Roberts, 1999; Schirripa, 1999; 2000; Allman et al., 2002). Hasta la década del 70 las capturas comerciales de L. campechanus por EUA sobrepasaban las 5 000 TM anuales, aunque del 35 al 45% se obtenían en el Banco de Campeche (Carpenter, 1955). En 1964 la flota de EUA inició su desplazamiento hacia el Mar Caribe y ya en 1968 la producción en dicha región era mayor (Fig. 12) que la que se obtenía en el Banco de Campeche (Allen y Tashiro, 1976) y desde fines de la década de los 70s se reubicó casi completamente en aguas nacionales. Las capturas comerciales en aguas de Estados Unidos, ya desde principios del siglo 19 mostraron disminución de los rendimientos en varias zonas de pesca (aunque aumentaba la captura), como resultado del constante incremento del esfuerzo pesquero (Jarvis, 1935; Camber, 1955; Carpenter, 1965; Moe, 1963; ver resumen en Moran y Morais, 1987). Las capturas comerciales domésticas fueron relativamente estables entre 1964 y mediados de los 70s, alrededor de 7 millones de libras (3 178 TM), para después declinar hasta un mínimo de 4.5 millones de libras (2 043 TM) en 1979. Aunque con una recuperación temporal en 1983, las capturas se han mantenido a niveles bajos en los últimos 20 años (Fig. 13). De 90 a 95% de las capturas de L. campechanus en aguas de Estados Unidos se obtienen en el norte del Golfo de México y el resto en el SE de la Florida (Keihly y Roberts, 1999). En el SE Atlántico (Carolina del Norte hasta los Cayos de la Florida) las capturas comerciales disminuyeron de 272 TM promedio en el período 1951-1985 (con máximo de hasta 454 TM), a sólo 83 TM promedio en el período 1986-1995 y no sobrepasan las 90 TM desde 1990. La reducción de las capturas en los últimos años se atribuye a las regulaciones de talla y cuota límite impuestas desde 1991 (Manooch et al., 1998). Las capturas de la pesca recreativa se estimó en 135 TM como promedio en el período 1981-1995, aunque probablemente cierta cantidad de R. aurorubens está incluida en esas cifras. Desde 1985 a 1995 el promedio fue de 89 TM (Manooch et al., 1998). Ambas pesquerías muestran igual tendencia al decrecimiento. Fig. 13. Capturas comerciales y recreativas de L. campechanus en el Golfo de México de E.U.A (elaborado a partir de los datos de Schirripa, 2000 y Fisheries of the United States. 1998-2001). Las capturas comerciales en el Golfo de México norteamericano muestran tendencia a la sobrepesca desde la década de los 60s (Ehrhardt y Legault, 1996; Scott-Denton y Nance, 1996). La captura total (comercial + recreativa) en dicha región sobrepasó los 10 millones de libras (4 540 TM) en 1979-80 y cayó a menos de 5 millones (2 270 TM) de 1981 a 1984 (Fig. 13). A partir de 1983 las capturas de la pesca recreativa también disminuyeron sensiblemente (Bennett, 183 1993), aunque desde 1994 se produjo un incremento y se mantiene entre 4 y 5,5 millones de libras (1816-2497 TM; Goodyear, 1994; Schirripa, 1998, 2000; NMFS, 2002). La disminución en 1990-1992 en parte fue producto de las regulaciones para proteger el recurso (ver epígrafe 6.1). Aunque se mantiene el estado de sobreexplotación de este recurso, las capturas totales (comercial más recreativa) promediaron 3 260 TM entre 1990 y 1998 (Patterson et al., 2002). En ese período se incrementó la pesca recreativa, en respuesta al incremento de las regulaciones a la pesca comercial. La Fig. 14 muestra las variaciones de la pesca recreativa en los estados del Golfo. La mayor parte de esas capturas se obtienen en las aguas exteriores de la zona económica exclusiva (Goodyear, 1994; Shirripa, 1998). Fig. 14. Capturas anuales de L. campechanus (1979-1997) por la pesca recreativa en los estados del Golfo de México (datos tomados de Goodyear, 1994 y Schirripa, 1998). En el SE Atlántico la captura total de L. campechanus (comercial y recreativa) disminuyó de 918 TM en 1985 hasta sólo 58 en 1992 y aunque con fluctuaciones posteriores, se mantiene a niveles relativamente bajos (Parker y Mays, 1998). Banco de Campeche. En la década del 60 y 70 la captura anual de L. campechanus por las flotas de EUA, México y Cuba en aguas del Banco de Campeche era de unas 3 000 TM anuales (Klima, 1976). No se observaba entonces ningún indicio de reducción del stock. Klima (1976) estimó una captura máxima sostenible de pargos en el área de 4 800 TM, de las cuales aproximadamente el 93% (4 460) corresponderían a L. campechanus. En el norte de Yucatán, González de la Rosa et al. (1994) y Monroy-García et al. (1996) plantearon que, durante los 80s, L. campechanus se encontraba sub-explotado y la pesquería se encontraba en proceso de crecimiento, cuando las capturas anuales de México crecieron de 1 800 a 4 500 TM. Según datos de FAO (2000) las capturas totales de esta especie se mantienen entre 3 000 y 3 500 TM anuales. 5.5 Acuacultura Arnold et al. (1978) en junio de 1978, por primera vez obtuvieron un desove espontáneo de L. campechanus mantenidos en cautiverio desde aproximadamente un año antes, sólo con manipulación del fotoperíodo y la temperatura. Un mes antes los peces mostraron una coloración más acentuada, aumentó espontáneamente su actividad y los machos con frecuencia embestían a las hembras por su vientre. No se observó conducta agresiva o territorial entre los machos. Al menos seis desoves se observaron en un período de 35 días, cada uno consistente en 184 unos pocos miles de huevos transparentes (0,8 mm de diámetro). El desove ocurrió con fotoperíodo de 15 horas de luz y 9 de oscuridad, temperatura del agua 23-25°C y salinidad de 31-34 ppm. El primer intento de obtener el desove inducido de L. campechanus se realizó en 1980 por Minton et al. (1983). Las hembras fueron inyectadas con Gonadotropina Coriónica Humana (GCH) a razón de 1,1 IU/g de peso del cuerpo. En los casos necesarios se inyectó posteriormente 0,55 IU/g. La ovulación ocurrió entre 42 y 56 horas después. Los oocitos fueron extraídos húmedos y posteriormente fertilizados con el esperma de dos machos. La mitosis inicial se detectó a los 30 minutos post-fertilización. La eclosión se produjo a las 20 horas con temperatura del agua de 27°C (±1°C), pero los embriones no sobrevivieron. Los ejemplares adultos de esta especie se adaptan rápidamente al cautiverio en estanques y aceptan tanto dietas frescas como preparadas comercialmente (Watanabe et al., 2001). No obstante, resultan muy susceptibles al stress provocado por el manejo para inducir la maduración gonadal y su conducta territorial ha dificultado el logro de métodos eficientes de producción. Sin embargo, se han logrado desoves exitosos con adultos maduros capturados en las agregaciones en condiciones naturales. Con inyecciones simples o múltiples de GCH (5001,500 IU/kg de peso del cuerpo) y LHRH-a (100 mu g/kg) se ha inducido el desove de hembras maduras, pero las tasas de fertilización han sido muy variables (Watanabe et al., 2001). Las experiencias han demostrado que las larvas de esta especie son difíciles de cultivar en sistemas intensivos, pero en cultivos semi-intensivos utilizando copépodos como primer alimento, se han logrado obtener pequeñas cantidades de larvas para criarlas hasta estadios juveniles. Si las tecnologías de cultivo continúan desarrollándose, pronto se podrá contar con protocolos de producción de juveniles (Watanabe et al., 2001). La producción actual de juveniles de L. campechanus depende del cultivo extensivo de zooplancton para su alimentación durante el desarrollo larval temprano. El zooplancton se produce mediante un sistema semi-continuo de bombeo de agua estuarina rica en plancton en estanques cerrados y cultivando aproximadamente la mitad del volumen del estanque en intervalos de 48 horas. Debido a la variabilidad en la productividad del plancton y el gran espacio requerido para este tipo de cultivo, se trabaja actualmente en el desarrollo de métodos intensivos de larvicultura, por investigadores del Instituto Oceánico (Oceanic Institute), en colaboración con el Laboratorio de Investigaciones Costeras del Golfo (Gulf Coast Research Laboratory), Mississippi (Shields et al., 2001). La Estación de Acuacultura de la Universidad Estatal de Louisiana evalúa el papel de la calidad de los gametos en la producción de embriones e híbridos de L. campechanus con Lutjanus griseus. La utilización de esperma criopreservado para el desove permitirá concentrar el esfuerzo en el mantenimiento de un stock de hembras reproductoras, monitorear el desarrollo de los ovarios e incrementar la eficiencia durante el proceso de desove. El esperma criopreservado, en los primeros experimentos produjo una tasa de fertilización de los huevos menor (32 ± 28%) que la obtenida con esperma fresco (58 ± 17%) pero ambos fueron igualmente efectivos en la fertilización de los huevos y producción de larvas (Riley et al., 2001). Similares resultados obtuvieron Chesney et al., (2001) quienes señalan que en el caso de los híbridos logrados de esas dos especies, la supervivencia fue mayor y crecieron mucho mejor que las larvas de L. campechanus en idénticas condiciones de cultivo. En dicho Laboratorio se ha logrado cultivar larvas de L. campechanus y en dos ocasiones fueron liberados los juveniles en aguas costeras. Las larvas se cultivaron en estanques oscuros colocados a la sombra en un invernadero abierto por los lados. Las larvas de un día de nacidas fueron sembradas a razón de 10/L en agua con salinidad de 30 ppt, con ligera aireación. Al segundo día se inició la alimentación con nauplios de copépodos calanoides producidos en 185 tanques de 100 m2 utilizando un método de cultivo con aguas turbias. Los peces cultivados solamente con copépodos alcanzaron mayores tallas y supervivencia que aquellos alimentados con Artemia durante algún tiempo. Los mejores resultados se obtuvieron con poca aireación y sin mantenimiento diario de los tanques. En los días 19-21 después de la eclosión ocurrió la mayor mortalidad de las larvas, aproximadamente el 50% del total en el primer y segundo años. Los juveniles producidos el primero fueron alimentados con varios tipos de alimento fresco y artificial. Estos peces tuvieron una alta mortalidad por el manejo, mostraron señales de deformaciones y mortalidad crónica. Los juveniles del segundo año se alimentaron con una dieta nombrada Mahi-Mahi, desarrollada en el Instituto Oceánico y no mostraron esos problemas. La mortalidad por conducta agresiva durante la transición de alimento vivo a pellets se redujo en el tercer año mediante la trasferencia temprana a los tanques exteriores de crecimiento. Un total de 13 146 ejemplares fueron cultivados con éxito, de los cuales 1 200 fueron marcados y soltados en las costas de Mississippi (Ogle et al., 2001). El Programa de rehabilitación de Poblaciones Marinas del Golfo de México norteamericano (USGMSEP) trabaja en el desarrollo de tecnologías de acuacultura como herramienta para la restauración de las poblaciones de L. campechanus. El programa ha desarrollado: 1- marcadores microsatélite y mitocondrial para el análisis de las poblaciones, de los reproductores y de los juveniles; 2 – un método para el cultivo de copépodos que permite la producción masiva de L. campechanus; y 3 – un procedimiento manejable y repetible para el cultivo de grandes cantidades de larvas (Blaylok et al., 2000). Con el fin de conocer la eficiencia de la rehabilitación de las poblaciones, Blackburn et al. (2002) liberaron 851 juveniles criados en cultivo, en tres tipos de hábitat naturales y artificiales, en las costas de la Florida. En las primeras 24 horas se estimó una mortalidad de 50% y en la primera semana se produjo otro 50% de mortalidad de las restantes, manteniéndose posteriormente en equilibrio durante 120 días. En un experimento posterior, aclimataron los juveniles durante 72 horas en jaulas ubicadas en el propio medio natural, lográndose una notable supervivencia de los peces liberados. Concluyen dichos autores que la aclimatación in situ puede ser una valiosa herramienta para los futuros esfuerzos de rehabilitación de las poblaciones de L. campechanus. 6. PROTECCIÓN Y MANEJO 6.1 Regulaciones específicas Estados Unidos de América. Sólo a partir de 1984 fueron establecidas, por el Consejo de Administración Pesquera del Golfo de México de EUA, algunas regulaciones efectivas para la conservación de L. campechanus. Las regulaciones existentes incluyen cuotas de captura, talla mínima, áreas vedadas y restricciones en el uso de los artes de pesca (Ehrhardt, 1996). Estas fueron reforzadas por la Enmienda 1, que proveyó de mecanismos para especificar la Captura Total Permisible (TAC) y ajustar las regulaciones sobre una base anual (Goodyear, 1984). A partir de 1991 se establecieron vedas al cumplimentarse la cuota anual. En 1992 la temporada fue reabierta poco después de la veda, con límites en la captura por viaje, debido a presiones sociales y económicas (Goodyear, 1994). En 1991 se estableció una cuota restringida de 3,1 millones de libras (1 407 TM) que se redujo a 2,04 (926 TM) en 1991 y se impuso una veda en agosto de ese año. A partir de entonces se ha utilizado esta medida, cambiando la cuota de acuerdo a la situación del recurso y a factores socio-económicos. A partir de 1998 también se produjo una reducción de la cuota percápita para la pesca recreativa y se cerró por primera vez la pesca cuando se estimó que se habían alcanzado los 4 468 millones de libras (2 028 TM). Ya en 1994 Goodyear (1994) reportó 186 un incremento de las capturas y de la talla mínima de la pesca recreativa, como resultado de la imposición de esas regulaciones. Las tallas mínimas, tanto para la pesca comercial como recreativa, se han ido incrementando a partir de 1990, desde 12 pulgadas (305 mm LT) hasta 16 (406 mm) en el año 2000 para la comercial y recreativa. La cuota de captura por persona en esta última modalidad se redujo de 7 ejemplares/persona en 1991 a 4 en el 2000 para el Golfo y sólo dos por persona en el Atlántico. Así mismo, los días de pesca se han reducido de 365 a 200 para la recreativa y 60 para la comercial (Patterson et al., 2002), con una veda del 1ro de noviembre al 14 de abril en el Golfo. En el SE Atlántico, a partir de 1992 se estableció una talla mínima de 20 pulgadas (508 mm LT; Parker y Mays, 1998) lo cual parece haber provocado una favorable respuesta de la especie (Manooch et al., 1997). Las medidas de manejo establecidas desde 1990 parecen tener resultados positivos (Schirripa y Legault, 1997). Tanto el reclutamiento como las capturas y las tallas de los peces muestran una tendencia a la recuperación (Schirripa y Legault, 1997), pero aún insuficientes y en algunos casos existen discrepancias no resueltas. Goodyear (1995) analizó el efecto de la talla mínima y su relación con la captura por recluta en función de la mortalidad por pesca y con diferentes valores de mortalidad natural. De acuerdo con sus resultados, todas las tallas límites utilizadas provocan un incremento en el rendimiento por recluta, mayor que el producido sin límites de talla. No obstante, sugirió que límites de talla altos pueden provocar consecuencias indeseables para el stock. Considerando que el crecimiento de un individuo tiene una base genética (Parna y Deriso, 1990), dicho autor considera que la selección impuesta por las tallas límites evaluadas, provocarían una selección por la pesca de los individuos de menor tasa de crecimiento para la próxima generación. Por otra parte, el establecimiento de una talla mínima legal conlleva que muchos peces son capturados con tallas inferiores y deben ser liberados. La práctica de desinflar la vejiga natatoria de los últimos, ha sido recomendada (Sport Fishing Institute, 1991) para reducir la mortalidad. En el caso de los juveniles capturados con chinchorros, la medida puede ser efectiva, ya que estos se elevan con cierta lentitud y trabajan a menor profundidad. No obstante, Render y Wilson (1996) argumentaron que este método no es efectivo para los peces capturados con cordel y anzuelo, generalmente a mayor profundidad, ya que en la mayoría de los casos el daño provocado por la inflación es irreversible. Los resultados obtenidos al evaluar la supervivencia de tales peces resultan positivos pero aún muy variables. Parker (1985) no encontró mortalidad inmediata al liberar en superficie 30 ejemplares capturados a 5,5 brazas (10 m) cerca de Texas. El mismo trabajo describe un experimento en que se reportó una mortalidad del 21% de los especímenes capturados a 4 brazas (7,3 m) los cuales fueron colocados en cajas de mallas metálicas a la misma profundidad de captura. En un estudio similar, ejemplares capturados a 5,5 brazas (10 m) tuvieron una mortalidad de 11%. Render y Wilson (1996) observaron una mortalidad de 20% en L. campechanus capturados a 20 brazas (36 m) y liberados en superficie dentro de una caja de 30 m de profundidad, pero tampoco observaron reducción importante de la mortalidad mediante la deflación de la vejiga natatoria. Gitschlag y Renauld (1994) realizaron experimentos de campo mediante dos métodos: 1liberándolos en cajas y 2- liberándolos en la superficie. Los peces capturados a 50 m de profundidad en el nordeste de las costas de Texas fueron colocados en cajas, bajados a 35 m de profundidad y monitoreados a intervalos irregulares de tiempo por buzos, durante 10-15 días. La supervivencia en dicho experimento fue de 64%. No se observó diferencias en la supervivencia entre las tallas (peces menores y mayores de 300 mm, N = 55, P = 0,59) ni entre peces cuya vejiga natatoria fue o no proyectada hacia la boca. (N = 45, P = 0,13). No se observó 187 depredación sobre los ejemplares liberados en superficie. Las tasas de supervivencia fueron 99%, 90% y 56% para los peces capturados a 21-24 m, 27-34 m y 37-40 m respectivamente. Las tasas de supervivencia mostraron una relación inversa entre la supervivencia y la profundidad de captura. Wilson et al. (2003), en aguas de Louisiana estudiaron el retorno o supervivencia de 4 839 ejemplares capturados entre 9 y 85 m y encontraron que la mayoría de estos no sobreviven: el 16% nadaron vigorosamente, el 15% se mantuvo en superficie por un momento antes de sumergirse, el 36% no se sumergieron y el 33% fueron retornados ya muertos, o sea que solo un 30 % fueron retornados con posibilidades de sobrevivir. El 86% de los peces retornados tenía más de 12 pulg. (30,5 cm) LT y dos años de edad. La liberación de los peces vivos por la pesca recreativa aumentó de menos de 3% antes de 1984, hasta alcanzar casi 60% en 1990 y se mantuvo fluctuando entre 40 y 60%, al menos hasta 1997, en que llegó a 62% (Schirripa, 1998). La liberación de tales peces se estimó que reduciría la mortalidad en un 33% (Goodyear, 1993). No obstante, el número de peces liberados en los últimos años parece no ser muy alto. Más de la mitad de los L. campechanuss liberados en 1996 y 1997 lo fueron porque tenian tallas inferiores a la mínima de captura establecida (Schirripa, 2000). En 1998, también se estableció una cuota máxima de captura comercial de L. campechanus de 9 millones de libras (4 086 TM) y se redujo la cuota de captura recreativa de 5 a 4 peces por pescador por día (Wilson y Fletcher, 1998; Patterson, 2002). Además se inició ese año una vigilancia estricta para el cumplimiento de esa cuota: por primera vez la pesca recreativa se cerró cuando se estimó que había alcanzado el limite establecido de 4 468 000 libras (2 028 TM). Goodyear (1994) estimó que para lograr la cuota máxima asignada para la pesca recreativa, es necesario reducir la cuota de pesca a 3 ejemplares por pescador/dia. De acuerdo con el Plan de Manejo para los peces arrecifales del Golfo de México de Estados Unidos, para el alcance una captura óptima y evitar la sobrepesca, se requiere que la Tasa Potencial de Reproducción (TPR) alcance al menos el 20%. La TPR es equivalente a la relación biomasa desovadora por recluta del stock pescado / biomasa desovadora del stock no pescado. Goodyear (1994) mediante modelo de simulación, estimó que la TPR para 1984 fue equivalente al 1% del nivel no pescado, cifra muy inferior al mínimo aceptable de 20% adoptado por el Consejo de pesquerías para el Golfo. Para 1993, gracias a medidas de conservación introducidas, dicho valor alcanzó 2%. Schirripa (2000) opina que el manejo de L. campechanus en el Golfo de México resulta muy controvertido. Las pesquerías de esta especie están sometidas a numerosas regulaciones debido al constante incremento de la presión pesquera sobre este recurso. A pesar de esas medidas de manejo, las poblaciones se mantienen con bajos niveles de abundancia y una Tasa Potencial de Reproducción (TPR) poco mayor de uno porciento (Goodyear, 1993). El Plan de manejo para las Pesquerías Arrecifales, desarrollado por el Consejo para el Manejo de las pesquerías en el Golfo de México (GMDMC), emitió como mandato que la TPR para esas especies debe elevarse al menos hasta el 20% para el año 2007. Esa meta, sin embargo, probablemente no sea alcanzable si no se logran reducciones grandes de la mortalidad de juveniles en la pesca incidental de camarones (GMFMC, 1993; Schirripa, 1999; 2000). Ehrhardt y Legault (1996) a través de la técnica de "bootstrap" modelaron la influencia anual de la captura acompañante de los arrastres de camarón sobre el stock de L. campechanus. Las matrices resultantes de captura por edad correspondientes al periodo 1984-1991, muestran que el efecto de las capturas incidentales de L. campechanus presionan a la población hasta un punto en que la abundancia disponible para la pesquería directa es tan baja que ninguna restricción impuesta a la pesquería directa puede brindar resultados satisfactorios para la recuperación del 188 recurso, sometido a tan severa sobrepesca. Esa situación contrasta dramáticamente con la abundancia estimada para las cohortes bajo la condición de cero pesca incidental en la pesca de camarón. Shirripa (1999) planteó que los valores calculados de captura máxima sostenible (CMS) difieren notablemente para cada valor de mortalidad por pesca incidental y por tanto dichos valores se han reducido sistemáticamente por los incrementos de la mortalidad de juveniles en dicha pesquería. En tales condiciones, es poco probable mantener una mortalidad por pesca (F) por debajo de la correspondiente a la CMS y al mismo tiempo mantener un nivel de TPR igual a 20%, a menos que la mortalidad por pesca incidental se reduzca en 90% aproximadamente o más. Si como se calcula, los mecanismos existentes de selección sólo alcanzan a reducir como máximo en 50% la captura incidental de juveniles, sólo se lograría elevar la TPR a 10% (Schirripa, 2000). Mediante simulación Goodyear (1994) analizó ocho alternativas de manejo que parten del criterio de mantener una captura máxima de seis millones de libras (2 724 TM) con diferentes variantes, en relación con la reducción de 50% de la pesca incidental en los arrastres camaroneros, con inicio en diferentes años. Todas las alternativas parten del criterio de que la mortalidad asociada a la pesca recreativa es de 49% y la comercial es de 51%. Sólo las dos alternativas que consideraban una reducción de la mortalidad por pesca incidental de 50% a partir de 1994 tenían un 95% de probabilidad de alcanzar una TPR = 0,20 para el año 2009. Considerando que tal reducción de la mortalidad no se ha iniciado, dicho autor concluye que resulta muy poco probable que se logre la meta trazada. 6.2 Recomendaciones para el manejo Considerando que las medidas directas de manejo no pueden por si solas garantizar la CMS mientras exista la alta mortalidad indirecta de la pesca incidental (Goodyear, 1995; Ehrhardt y Legault 1996; Schirripa, 1999; 2000) debe buscarse un punto de equilibrio en el manejo de ambas pesquerías (la de L. campechanus y la de camarones) para lograr el mayor beneficio con el menor costo ecológico. En tal caso, las medidas y regulaciones existentes (reducción del esfuerzo, perfeccionamiento de los BRD, liberación de los peces capturados por la pesca recreativa, mantenimiento de la talla mínima actual, maximizar el rendimiento por recluta) contribuirán de forma más efectiva a la rehabilitación del recurso. El uso de refugios artificiales de bajo relieve pudiera contribuir a disminuir la mortalidad de los juveniles en las capturas de camarón. Estas estructuras pueden atraer a los juveniles fuera del área de arrastre (Workman et al., 1998a;b). No obstante, tal opción debe ser estudiada, ya que tales estructuras también atraen a peces mas grandes que pueden ser depredadores de los juveniles de L. campechanuss o competir con estos (Rogers, 1998). Galaway et al. (1999) sugieren que los planes de manejo deben incluir la identificación y descripción de los hábitat esenciales para esta especie, así como de los impactos adversos sobre estos, incluyendo los de la pesca, así como acciones para la conservación y rehabilitación de los mismos. L. campechanus muestra una gran atracción hacia los refugios artificiales, estructuras ampliamente utilizadas en el Golfo de México, principalmente por la pesca recreativa. Las estructuras sumergidas de las plataformas de explotación de petróleo en dicha región constituyen importantes sitios de concentración de esta especie (Moran y Morais, 1988; Render, 1995; Gallaway y Cole, 1999a; Nieland y Wilson, 2000). No obstante, la atracción de dichas estructuras puede incrementar la vulnerabilidad de los especimenes de 4 a 6 años de edad a la mortalidad por pesca, ya que estas edades están asociadas a dichas estructuras en mayor proporción que en las poblaciones naturales (Nieland y Wilson, 2003). En las costas de 189 Alabama, donde no existen muchos arrecifes naturales, se han instalado grandes cantidades de refugios artificiales. De acuerdo con Szedlmayer y Shipp (1994) en esta zona L. campechanus se caracteriza por alcanzar tallas y edades mayores (hasta 42 años) y presentar un alto nivel de residencia, aún a través de largos períodos de tiempo. El conjunto de estos factores sugieren que la alta producción pesquera en la zona es indicativa, no sólo de la atracción que propician los refugios artificiales, sino de una alta productividad pesquera (Szedlmayer y Shipp. 1994), lo cual sugiere el uso de los mismos como un mecanismo más de manejo. 190 Tabla 3. Composición de la dieta de Lutjanus campechanus en varios ecosistemas y épocas del año. Ver -verano; inv - invierno; prim - primavera. Localidad Golfo de Oeste de la Noroeste del Golfo de México (Texas) México Florida, Golfo de México Presas Presencia* Número PISCES CRUSTACEA Decapoda Brachiura Portunidae Majidae Otros bentónicos Anomura + Palinura Copepoda Peneidea Caridea Peneidea + Caridea +otros camarones Stomatopoda Otros crustáceos MOLLUSCA Gastropoda Pelecípoda Cephalopoda Zooplancton pelágico Otros organismos o no identificados Carnada (peces y calamares) No. peces muestreados No. estómagos llenos xxx xxx xxx xx x 32 22 15 10 4 3 2 1 xx x xx 1 2 xx 4 xxx x x xx 2 1 1 3 %V Ver. 20 25,7 %V Otoño 15,6 33,8 %V Inv 14,4 77,9 9,1 16,6 %V Prim 86,7 6,6 9,3 26,9 20,6 33,8 21,1 41,3 45,0 12,2 5,6 11,1 %V Ver 33,6 45,6 %V Otoño 77,6 16,4 39,2 6,0 6,4 10,4 %V Inv 96,9 Norte del Golfo de México (Alabama) %P %V Prim 75,6 281 46,5 271 41,7 2,51 7,7 171 33,3 11,7 13,3 6,6 20,8 6,0 3,1 231 9,6 8,8 40 213 96 Tallas Referencias Juveniles Adultos Moran y Futch y Bradley y Bryan, 1975 Morais, 1988 Bruger, 1976 * xxx – varios taxones, alta frecuencia; xx – taxones importantes ; x - otros taxones representados; 1-% con relación al peso de organismos identificados; organismos identificables; 3 estómagos con organismos no identificables o con carnada. 191 2 9,6 656 2682 1263 207-913 LH McCawley et al. (2003) -estómagos con 192