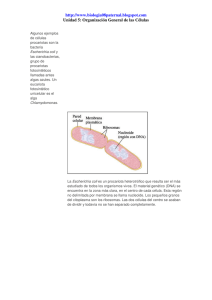
Bacteria - Instituto de Investigaciones Biotecnológicas
Anuncio

MICROBIOLOGIA GENERAL CURSO 2010 TEMA: Introducción a la Microbiología. DOCENTE: Dra. Nora Iñón Bibliografía recomendada: Brock Biología de los Microorganismos de Madigan, Martiniko y Parke, 10a edición (PEARSON, Prentice Hall). Bioquímica de Stryer, Berg y Tymoczko, 5ta edición (Editorial Reverté) Microbiology de Prescott, Harley y Klein 7ma edición (McGraw-Hill). Sitio web recomendado: http://www.ugr.es/local/eianez 1 Microbiología • Es el estudio de los microorganismos • Como ciencia básica permite conocer los procesos vitales en sistemas simples. • Como ciencia aplicada permite solucionar problemas prácticos, por ej. en medicina, agricultura, industria, etc. 2 Características de la célula • Las células microbianas son distintas a las células de animales o de plantas. • Crecen, se multiplican, generan energía. • A diferencia de las células de animales o de plantas, las células microbianas pueden vivir en forma aislada de otras células. 3 a y b: organismos pluricelulares, compuestos por muchos tejidos que forman órganos; c y d: microorganismos. (c) bacterias rojas, (d) cianobacterias. 4 La microbiología estudia • Las células vivas: como trabajan y como existen en vida libre. • La diversidad de los microorganismos. • La evolución de los microorganismos. • Los distintos procesos que llevan a cabo. 5 La microbiología es una ciencia biológica • Como ciencia básica es la herramienta mas simple para el estudio de procesos vitales. • Como ciencia aplicada se puede utilizar en distintos campos: para el estudio de la producción animal; fertilidad de la tierra; enfermedades humanas, de animales y de plantas; procesos biotecnológicos o “Biotecnología”, etc. 6 Los microorganismos como células • Es la unidad fundamental de la vida. • Posee membrana celular y en algunos casos pared. • En el citoplasma se encuentran sustancias químicas, estructuras subcelulares y una región nuclear o núcleo. • Las células contienen proteínas, ácidos nucleicos, lípidos y polisacáridos 7 Ancestro común • Se piensa que todas las células provienen de un ancestro común o ancestro universal que a lo largo de millones de años de evolución ha dado lugar a la gran diversidad observada hoy en día. 8 Localización de las macromoléculas en la célula a) Las proteínas se encuentran en toda la célula formando parte de estructuras celulares y de las enzimas. b) ácidos nucleicos. El DNA (verde) se encuentra en el nucleoide de las células procarióticas y en el núcleo de las células eucarióticas. El RNA (naranja) se encuentra en el citoplasma (mRNA y tRNA) y en los ribosomas (rRNA). (c) Los polisacáridos (amarillo) se localizan en la pared celular y, en ocasiones, en gránulos de reserva internos. (d) Los lípidos (azul) se localizan en la membrana citoplasmática, la pared celular y en gránulos de reserva. 9 Peso seco de una célula en crecimiento de Escherichia coli aproximadamente 2,8 x 10-13 g Peso total (70 % agua) = 9,5 x 10-13 g El 96% del peso seco de una célula es debido a las macromoléculas y, dentro de estas , las proteínas son las más abundantes. Las proteínas se encuentran por toda la célula y tienen funciones estructurales y catalíticas. Le sigue el ácido ribonucleico (RNA) presente en los ribosomas (junto con las proteínas), RNA mensajero y de transferencia. 10 - Las células tienen estructura y tamaño definidos. - Sufren cambios continuamente sustituyendo cada una de sus partes. - Son unidades dinámicas. - Son sistemas abiertos que cambian continuamente pero que a la vez permanecen dentro de ciertos límites. 11 Características de una célula • • • • • • Metabolismo Reproducción Diferenciación Comunicación Movimiento Evolución 12 Características de los sistemas vivos 1- Metabolismo. Toman sustancias químicas del medio y las transforman, conservan parte de la energía de dichas sustancias de modo que las células puedan usarla, y luego eliminan los productos de desecho. La célula es por lo tanto un sistema abierto. 2- Reproducción. Poseen la capacidad de autoduplicarse es decir dirigir una serie de reacciones químicas que conducen a su propia síntesis. Como resultado de los procesos metabólicos, una célula crece y se divide para formar dos células. 13 3- Diferenciación. Proceso por el cual se forman nuevas sustancias o estructuras. A menudo forma parte de un ciclo de vida en el cual la células forman estructuras especiales relacionadas con la reproducción, la dispersión o la supervivencia. 4Comunicación. Las células responden a señales químicas en su medio ambiente, tales como las producidas por otras células. Pueden comunicarse por medio de pequeñas moléculas (como la AHL o lactona de homoserina acilada), que se difunden y pasan entre células vecinas. Existen vías regulatorias que responden a densidad de población o Quorum sensing (percepción en quorum). 14 5- Movimiento. Algunos organismos vivos son capaces de moverse por autopropulsión. Hay diferentes mecanismos responsables de la movilidad en el mundo microbiano. 6- Evolución. A diferencia de las estructuras inertes, las células pueden evolucionar. A través del proceso de evolución las células pueden cambiar permanentemente sus características y transmitir las nuevas propiedades a su descendencia. 15 Las células como máquinas y como sistemas codificados - Como máquinas químicas: llevan a cabo transformaciones químicas. Convierten energía de una forma a otra, rompen moléculas, construyen moléculas. Las fuerzas conductoras son las enzimas o catalizadores que son proteínas capaces de acelerar la velocidad de reacciones químicas específicas. - Como sistemas codificados: son análogos a las computadoras, que guardan y procesan la información genética (DNA) que pasa finalmente a la descendencia durante la reproducción. El enlace entre estos dos atributos es el crecimiento, el cual debe estar coordinado y regulado para lograr que una célula se reproduzca con fidelidad. 16 Las células como dispositivos de codificación procesan la información que se transmite a su descendencia o se traduce de otra forma. Función codificante: transcripción DNA traducción RNA Proteína La célula es un depósito de información de secuencias de proteínas almacenadas en forma codificada. El código es el código genético almacenado en el DNA DNA: secuencia de nucleótidos. Cada aminoácido de una proteína está codificada por una secuencia de tres base o triplete. A pesar de la diferencia en la frecuencia del uso de un determinado codón para codificar por un aminoácido determinado, el código genético, con unas pocas excepciones, es el mismo para todos los organismos. 17 Síntesis de tres tipos de moléculas informacionales -Los genes están compuestos de ácido desoxirribonucleico (DNA). -La información guardada en el DNA codifica la secuencia de una proteína, y lo hace a través de una molécula intermediaria, el ácido ribonucleico (RNA). -La transferencia de la información al RNA se denomina transcripción. - La molécula de RNA que lleva la información que codifica una proteína se denomina RNA mensajero (mRNA). - Algunas regiones del DNA que se transcribe codifican también para otros tipos de RNA, tales como el RNA de transferencia (tRNA) y RNA ribosómico (rRNA). Gen se puede definir entonces como un segmento de DNA que codifica para una proteína, un rRNA o un tRNA. 18 Traducción. La secuencia específica de aminoácidos en cada proteína está determinada por una secuencia específica de bases en el mRNA. - Existe una correspondencia lineal entre la secuencia de bases de un gen y la secuencia de aminoácidos de un polipéptido. -Se necesitan tres bases del mRNA para codificar un solo aminoácido y cada uno de estos tripletes se denomina codón. -Este código genético se traduce a proteína por medio de la maquinaria para la síntesis de proteínas. Este sistema consta de ribosomas (compuestos a su vez de proteínas y rRNA), tRNA y varias enzimas. La transferencia de información de ácido nucleico a proteína es unidireccional. La secuencia de la proteína no indica la secuencia del ácido nucleico. Esta transferencia unidireccional de la información se conoce como dogma central de la biología porque es compartida por todas las formas de vida de nuestro planeta. 19 El código genético consiste de 64 codones. Tres de esos codones son reservados para “stops”, y uno (AUG) es reservado para iniciación. Hay un solo codon (UGG) para el amino ácido triptofano. Para el resto de los amino ácidos hay al menos 2, usualmente 4 y algunas veces 6. Por ejemplo hay 6 codones (UUA, UUG, CUU, CUC, CUA y CUG) para el aminoácido leucina. 20 Código genético y frecuencia de uso en Escherichia coli y en humanos. Los distintos codones que codifinan para un mismo aminoácidos no son usados por los diferentes organismos con la misma frecuencia. De los 4 codones para glicina, GGA es usado 26% del tiempo por genes estructurales humanos y aproximadamente 9% del tiempo para las proteínas codificadas por genes de la bacteria E. coli. Lo mismo ocurre con el uso de los codones stop. Codon Amino ácido Frecuencia de uso E. coli humanos GGA glicina 0,09 0,26 UGA stop 0,30 0,61 UAG stop 0,09 0,17 UAA stop 0,62 0,22 21 El DNA sufre dos transformaciones principales: 1- replicación 2- transcripción traducción. -El aparato de traducción es una maquinaria muy compleja y es el centro mismo de la función celular. - Es el atributo central y fundamental de la célula. - Con pequeñas variaciones es prácticamente el mismo en todos los organismos. - Ha sido probablemente una adquisición muy temprana en la historia de la Tierra y se ha mantenido sin grandes variaciones hasta ahora. 22 Teoría de Darwin • Durante el crecimiento celular y división debe haber una duplicación del código genético. La fidelidad de la copia debe ser muy alta. Los errores suelen dar mutaciones. En general las mutaciones son perjudiciales. • En raros casos las mutaciones son beneficiosas y, si el medio ambiente es adecuado, dan una ventaja selectiva permitiendo una selección natural. Esto forma parte del proceso de evolución enunciado en la Teoría de Darwin. Teoría de Darwin • mutación selección evolución 23 La evolución necesita reproducción, variación y presión selectiva El proceso evolutivo. Hay varios principios básicos comunes a los sistemas en evolución, tanto si son simples colecciones de moléculas como poblaciones de organismos en competición. 1- La propiedad fundamental de los sistemas en evolución es su capacidad para replicarse o reproducirse. Sin su capacidad de reproducirse, cada “especie molecular” que pueda aparecer está condenada a su extinción, tan pronto las moléculas individuales se degraden. Las moléculas capaces de replicarse seguirán representadas en la población, incluso si el tiempo de vida de cada molécula individual sigue siendo corto. 2- El segundo principio fundamental de la evolución es la variación. Los sistemas que se replican deben sufrir cambios. Si un sistema se replica siempre perfectamente, la molécula replicada será igual a las moléculas progenitoras y la evolución no podrá tener lugar. 24 3- El tercer principio básico de la evolución es la competición. Las moléculas que se replican compiten entre sí por los recursos disponibles, tales como los precursores químicos, y la competición permite que ocurra el proceso biológico de la evolución por la selección natural. La variación producirá distintas poblaciones de moléculas. Algunas de las variantes que surjan pueden estar, por casualidad, mejor adaptadas que las moléculas progenitoras para sobrevivir y replicarse en las condiciones predominantes. Las condiciones predominantes ejercen una presión selectiva que proporciona una ventaja a una de las variantes. Estas moléculas que son más capaces de sobrevivir y replicarse, aumentarán en concentración relativa. Por tanto surgen nuevas moléculas que son más capaces de replicarse en las condiciones de su entorno. El mismo principio es válido para los organismos actuales. Los organismos se reproducen, presentan variaciones entre organismos individuales y compiten por los recursos; las variantes con ventajas evolutivas se reproducirán con más éxito. 25 Fidelidad de la replicación del DNA y fenómeno de mutación La parte de arriba de la figura muestra un evento de replicación normal; la parte de debajo de la figura esquematiza la introducción de una mutación 26 Reproducción de una célula. Para que una célula se reproduzca debe disponer de un suministro adecuado de energía y de precursores para la síntesis de nuevas macromoléculas, el material genético debe duplicarse para que durante la división celular cada célula reciba una copia. Los genes deben expresarse (mediante los procesos de transcripción y traducción) para formar las cantidades requeridas de proteínas y macromoléculas necesarias para hacer la nueva célula. 27 Resumiendo -Una célula no está en equilibrio con su entorno - Es un sistema abierto que toma energía la cual utiliza en parte para mantener su propia estructura. -Si se destruye su estructura, la célula muere. -La estructura celular es la base de la función celular. -Las células se autoduplican. Las células reproducen las células. -Si la reproducción es correcta se origina una célula viva. 28 El origen de la vida La composición química promedio de una célula viva es muy distinta a la de la Tierra. Los elementos clave como por ej C, H, O, N, P y S son mucho mas abundantes en los organismos vivos que en la materia no viva como por ejemplo la corteza de la tierra. La célula es un sistema químico selectivo compuesto principalmente de C, H, O, N, P y S, (los elementos principales de la vida). Esto enfatiza la naturaleza especial y no al azar de una célula viviente. 29 Diferencias químicas entre un organismo vivo y la Tierra 30 Según el famoso patólogo celular alemán Rudolph Virchow: “Omnis cellula e cellula” que quiere decir: “Toda célula proviene de una célula”. La pregunta es entonces ¿De donde proviene la primera célula? Se piensa que proviene de una estructura procelular 31 En 1920 Oparin en la Unión Soviética y Haldane en Inglaterra sugirieron que hubo un período de evolución química prebiótica antes de la aparición de la primera célula Acción de la luz UV Rayos Calor volcánico Superficies inorgánicas (catalizadores) Concentración por períodos de congelamiento o ciclos de desecación Diversos compuestos orgánicos. Polimerización de estos caldos. Catalizadores proteicos. Confinamiento de un sistema autocatalítico en una célula rodeada por una membrana. Evolución desarrollando vías biosintéticas. 32 El planeta Tierra tiene aproximadamente 4.500 millones de años. Hay evidencias fósiles de que organismos que se asemejan morfológicamente a las bacterias modernas, existieron hace ya 3.500 millones de años. A partir de resultados de estudios programados y de descubrimientos accidentales se ha podido construir una vía evolutiva hipotética, pero plausible, desde el mundo prebiótico hasta el presente. Podemos imaginar que distintas etapas llevaron a la aparición de las especies vivas modernas. 1ra etapa. Producción inicial de algunas de las moléculas claves de la vida (ácidos nucleicos, proteínas, hidratos de carbono y lípidos) mediante procesos no biológicos. 2da etapa. Transición de la bioquímica prebiótica a sistemas capaces de replicarse. Con el paso del tiempo estos sistemas se hicieron cada vez más complejos, permitiendo la formación de las células vivas. Bioquímica prebiótica sistemas capaces de replicarse célula 33 3ra etapa. Evolucionaron los mecanismos para convertir la energía de las fuentes químicas y del Sol en formas que pudieran ser utilizadas para dirigir las reacciones químicas. Estos procesos de conversión de energía están interconectados con las vías para sintetizar los componentes de los ácidos nucleicos, las proteínas y otras sustancias clave, a partir de moléculas más sencillas. Con el perfeccionamiento de los procesos de transformación de la energía y de las vías de biosíntesis se desarrollaron una gran variedad de organismos unicelulares. 4ta etapa. Evolución de los mecanismos que permitían a las células ajustar su bioquímica a entornos diferentes y, a menudo cambiantes. Organismos con estas capacidades pudieron formar colonias compuestas por grupos de células capaces de interaccionar y algunos finalmente evolucionaron hacia organismos pluricelulares complejos. Organismos células capaces de interaccionar organismos complejos 34 Señalamos que la vida apareció aproximadamente 1000 millones de años después de la formación de la Tierra. Pero antes de que la vida pudiese existir fue necesario que otro proceso fundamental tuviese lugar. Esto es, la síntesis de moléculas orgánicas, necesarias para los sistemas vivos, a partir de moléculas sencillas presentes en el entorno. Muchos componentes de las macromoléculas bioquímicas se pueden producir en reacciones prebióticas sencillas. Entre las distintas teorías que compiten para explicar las condiciones del mundo prebiótico, ninguna es totalmente satisfactoria o está libre de interrogantes. Una teoría sostiene que la primera atmósfera de la Tierra era muy reductora, rica en metano, amoníaco, agua, e hidrógeno y que esta atmósfera estaba sometida a grandes cantidades de radiación solar y relámpagos. De momento supondremos que esas condiciones fueron las de la Tierra prebiótica. ¿Se pueden sintetizar moléculas complejas en esas condiciones? 35 En los años 50, Stanley Miller y Harold Urey elaboraron una respuesta a esta pregunta. Hicieron saltar una descarga eléctrica, simulando relámpagos, a través de una mezcla de metano, amoníaco, agua e hidrógeno. Asombrosamente, estos experimentos dieron una mezcla de compuestos orgánicos muy determinada, que incluía aminoácidos y otras sustancias fundamentales para la bioquímica. H20 agua NH3 amoníaco chispas eléctricas aminoácidos CH4 metano H2 hidrógeno 36 El proceso produce los aminoácidos glicina y alanina con un rendimiento aproximado del 2% y pequeñas cantidades de ácido glutámico y leucina. 37 El ácido cianhídrico (HCN), otro componente probable de la atmósfera primitiva, se condensa al exponerse al calor o a la luz para producir adenina, una de las cuatro bases de los ácidos nucleicos. Otras moléculas sencillas se combinan para formar el resto de las bases. Una gran cantidad de azúcares, incluyendo la ribosa, se pueden formar a partir del formaldehido en condiciones prebióticas. 38 ¿Cuál fue el origen de la información genética? ¿Cómo se originó el código genético?, es decir la posibilidad de almacenar información que pueda ser traducida en la formación de macromoléculas. Como un sistema primitivo de replicación y expansión de la información es probable que el RNA fuese la primera macromolécula portadora de información combinando las funciones de catalizadores y de moldes (templados) en la misma molécula. Recientemente Cech y Altman, en forma independiente, descubrieron que ciertas moléculas de RNA podían ser catalizadores eficientes. A estos RNA catalizadores se los denominó ribozimas. Las ribozimas cortan su propia cadena en lugares específicos y pueden aumentar la longitud uniendo los extremos generados en dichos cortes. El descubrimiento de las ribozimas ha sugerido la posibilidad de que las moléculas de RNA catalíticas pudieran haber desempeñado papeles esenciales en las etapas tempranas de la evolución de la vida. 39 La capacidad catalítica de las moléculas de RNA está relacionada con la posibilidad de adoptar estructuras específicas y complejas. La ribozima “cabeza de martillo” identificada por primera vez en un virus de plantas induce la hidrólisis de moléculas de RNA determinadas en lugares específicos. La ribozima, que necesita Mg++ y otros iones para que tenga lugar la hidrólisis, forma un complejo con la molécula de RNA sustrato, que puede adoptar entonces, una conformación reactiva. 40 Finalmente el almacenamiento de información habría pasado a ser el DNA (compuesto mas estable y menos versátil), mediante la transcripción inversa, mientras que la actividad catalítica habría sido asumida por las enzimas proteicas (mas flexibles y eficaces). No existe indicio alguno de cómo podría haber sido el mecanismo elemental de traducción previo al actual, en el cual participan mas de 100 componentes. 41 Potencial genético de una célula ¿Cuántos genes tiene una célula? Escherichia coli tiene aproximadamente 4,6 millones de pares de bases (4.600 kilobases). Si un gen en promedio tiene 1.1 kb, significa que la célula tiene aproximadamente 4.300 genes, si todo el DNA codificara para proteínas. Pero no todo el DNA codifica para proteínas. Se estima que E. coli tiene unos 3000 genes , de los cuales 1000-2000 son expresados en algún momento particular. Las células eucarióticas pueden tener muchos mas. Por ejemplo la célula humana tiene 1000 veces mas DNA que la célula de E. coli y alrededor de 7 veces su número de genes. La mayor parte del DNA en las células eucarióticas es no-codificante. 42 ¿Cuántas proteínas hay en una célula? Una única célula de Escherichia coli contiene aproximadamente 1900 clases de proteínas diferentes y un total de 2,4 millones de moléculas de proteínas individuales, algunas de ellas en gran número (mas de 100.000) y otras en pequeño número (menos de 100). La célula tiene algunos medios para controlar la expresión (transcripción y traducción) de sus genes y debido a esto no todos los genes son expresados en la misma extensión o al mismo tiempo. Tanto las células procariotas como eucariotas controlan la expresión de sus genes de modo que no todos ellos se expresan con la misma frecuencia o al mismo tiempo. 43 Glosario Cromosoma: elemento genético que lleva genes esenciales para el funcionamiento de la célula. Citoplasma: contenido celular que se encuentra dentro de la membrana citoplasmática, excepto el núcleo (si existe). Genoma: el conjunto de genes de un organismo. Nucleoide: masa total del DNA que constituye el cromosoma de las células procarióticas. Núcleo: una estructura rodeada por una membrana que contiene los cromosomas en células eucarióticas. Plásmido: un elemento genético extracromosómico que no es necesario para el crecimiento. Ribosoma: partícula citoplasmática donde se realiza el proceso de la síntesis de proteínas. 44 Procariota: una célula que carece de un núcleo delimitado por una membrana nuclear y de otros orgánulos. Eucariota: una célula con un núcleo delimitado por una membrana nuclear y que en general presenta otros orgánulos, como las mitocondrias o los cloroplastos que contiene su propio DNA (normalmente en disposición circular, como en Bacteria) y sus propios ribosomas. Los organismos pertenecen al dominio Eukarya. Filogenia: relaciones evolutivas entre organismos. Dominio: el nivel mas elevado de la clasificación biológica (Bacteria, Archaea y Eukarya) 45 Estructura celular y relación estructura función Todas las células tienen una barrera llamada membrana citoplasmática o celular que separa el interior del exterior de la célula. En el interior se encuentra el citoplasma, donde se llevan a cabo las funciones de la célula. Los principales componentes del citoplasma son: agua, macromoléculas (proteínas y ácidos nucleicos), ribosomas, pequeñas moléculas orgánicas precursores de macromoléculas y varios iones inorgánicos. En los ribosomas se sintetizan las proteínas. Están compuestos por ácido ribonucleico (RNA) y proteínas que interaccionan con proteínas solubles y con el RNA mensajero. La pared celular se encuentra en el exterior, rodeando la membrana plasmática, es relativamente permeable y otorga rigidez estructural a las células. Los vegetales y la mayoría de los MOs poseen pared, la mayoría de las células animales carecen de ella pero tiene un citoesqueleto. 46 Células eucariotas y procariotas Se pueden distinguir por un análisis detallado de la estructura celular interna. Las células eucarióticas son generalmente mas grandes y estructuralmente mas complejas que las procarióticas presentando estructuras limitadas por membranas llamadas orgánulos. Los orgánulos comprenden el núcleo, las mitocondrias y los cloroplastos (presentes solo en las células fotosintéticas). Las mitocondrias y los cloroplastos desempeñan funciones específicas en la generación de energía, llevando a cabo la respiración y la fotosíntesis, respectivamente. Los microorganismos eucarióticos comprenden las algas, los hongos y los protozoos. Todos los metazoos (animales y plantas) están formados por células eucarióticas. 47 Estructura interna de las células microbianas a) Diagrama de un procariota b) Diagrama de un eucariota 48 49 50 Organización del DNA. Núcleo versus Nucleoide. Los genomas presentan una organización diferente en células procariotas y en células eucariotas. En procariotas, el DNA se encuentra como una molécula larga de dos cadenas formando el cromosoma bacteriano que se condensa para dar una masa visible llamada nucleoide. En la mayoría de los procariotas el DNA es circular, y en general, poseen un cromosoma único que contiene una sola copia de cada gen, por lo cual son genéticamente haploides. Pueden tener también pequeñas cantidades de DNA extracromosómico, llamados plásmidos que les confieren propiedades particulares no esenciales y en general requeridas para la supervivencia. 51 En eucariotas, el DNA se encuentra dentro del núcleo organizado en cromosomas. El número de cromosomas depende del organismo y a diferencia de los procariotas contienen dos copias de cada gen, por lo tanto son diploides. Saccharomices cerevisiae tiene 8 pares de cromosomas, células humanas tiene 23 pares de cromosomas Los cromosomas contienen, además de DNA, proteínas que favorecen el plegamiento y el empaquetamiento del DNA, así como proteínas necesarias para la expresión génica. Durante la división celular el núcleo se divide (luego de duplicarse el número de cromosomas) mediante el proceso de mitosis originándose 2 células hijas idénticas. 52 Empaquetamiento de DNA alrededor de un núcleo de histonas para formar un nucleosoma. Los nucleosomas se ordenan a lo largo de la cadena del DNA asemejando bolas en una cuerda. Esta conformación es típica del DNA en las células eucarióticas. Histonas centrales 53 Características que diferencian a procarióticas de los eucarióticas -Cromosoma circular único (nucleoide), en lugar de un núcleo con múltiples cromosomas. -Ausencia de membrana nuclear y aparato mitótico. -Ausencia de histonas verdaderas, nucleosomas. -Ausencia de intrones en los genes que codifican proteínas y escasa presencia de los mismos en los que codifican tRNA. -La transcripción se acopla con la traducción. El extremo del mRNA no está poliadenilado (excepto en arqueobacteria) -mRNA generalmente poligénico. -El ribosoma es 70S en lugar de 80S; ligeras diferencias de forma, longitud de moléculas de rRNA y número de moléculas proteicas. -Pared celular con compuestos exclusivos (por ejemplo: ácido. Murámico, D-aminoácidos, frecuentemente ácido diaminopimélico,..). 54 -Ausencia de esteroides y fosfatidilcolina en la membrana. -Ausencia de vesículas endocíticas. -Ausencia de retículo endoplásmico (aunque algunas bacterias autotróficas pueden presentar algunas invaginaciones de la membrana citoplasmática). -Ausencia de mitocondrias y otros orgánulos limitados por membranas (con algunas excepciones). -Ausencia de triglicéridos (en algunos grupos el carbono se almacena en forma de ácido poli-beta-hidroxibutírico -Movimiento por rotación de flagelos -Características metabólicas exclusivas: fijación de nitrógeno, respiración anaeróbica (con aceptores de electrones distintos al oxígeno), fotosíntesis sin producción de oxígeno. 55 Por muchos años se pensó que los organismos procarióticos carecían de un citoesqueleto que le permitiera tener un nivel de organización citoplasmática similar a los organismos eucarioticos. Recientemente se han identificado los tres elementos del citoesqueleto de eucariotes en Bacteria (microfilamentos, filamentos intermedios y microtubulos) y uno de ellos en Archaea. Los elementos del citoesqueleto de Bacteria son similares estructuralmente a su contraparte en eucariotas y tienen funciones similares: los mismos participan en la división celular, localizan proteínas en ciertos sitios de la célula y determinan la forma de la misma. 56 MreB (la contraparte en células eucarióticas es la actina). Se observa en muchas bacterias en forma de bastón. En Bacillus subtilis es llamada Mb1. MreB forma filamentos en espiral alrededor del interior de la célula determinando la forma de la misma y ayuda a mover cromosomas a polos opuestos de la misma. 57 Mb corresponde a megapares de bases. Todos los genomas procarióticos indicados han sido secuenciados. Todos los organismos del listado son unicelulares 58 a) Micrografía de células de bacterias b) Sección longitudinal de una célula bacteriana vista con microscopio electrónico. c) Interpretación artística en 3 dimensiones de una bacteria Gram negativa. 59 Micrografía electrónica de una sección de una célula del dominio Bacteria. Helicobacterium modesticaldum La célula mide 1x3 μm. El área clara representa el nucleoide, región de la célula que contiene el DNA. Corte longitudinal. 60 61 Micrografía electrónica de una célula de Saccharomyces cerevisiae (Dominio Eukarya). La célula mide 8 μm de diámetro Núcleo Membrana citoplasmática Pared celular Membrana interna Mitocondria 62 Estructura interna de células microbianas Pared celular a citoplasma b Mitocondria c d Nucleolo Procariote a) diagrama y b) microscopía electrónica Núcleo Eucariote c) diagrama y d) microscopía electrónica 63 Los virus -no son células -No son sistemas abiertos dinámicos -Son estructuras estáticas y muy estables -Incapaz de cambiar o reponer estructuras por si mismos -Necesitan de la maquinaria de la célula para replicarse y para sintetizar sus proteínas -No tienen metabolismo propio -No tienen aparato de traducción -Son mucho más pequeños que las células 64 Tamaño de los virus y estructura a) Partículas de adenovirus (tamaño 100 nm de diámetro) b) Tamaño de una partícula de virus y comparación con bacterias y células animales 65 Clasificación : -Se clasificaron primero como animales o plantas pero luego aparecieron los microorganismos. -Se descubrió luego que todos los organismos vivos se podían dividir en dos tipos celulares distintos (procariotes y eucariotes) con base en diferencias fundamentales en la estructura celular. Todos los animales y plantas son eucariotes, como lo son tres grupos importantes de microorganismos: algas, hongos y protozoos. 66 Relación evolutiva entre organismos Relación estructural no necesariamente implica relación evolutiva. Hoy en día se utiliza un método que compara la estructura del RNA ribosomal como una medida de las relaciones filogenéticas o evolutivas. De estos estudios surgen tres linages celulares evolutivamente distintos, dos de los cuales son procariotes en estructura y uno es eucariote. A estos grupos se les dio el nombre de Bacteria, Archaea y Eukarya. Todos estos grupos se piensa que vienen de un ancestro común o “ancestro universal”. 67 Los procariotas comprenden las Bacteria y las Archaea. Bacteria y Archaea comparten estructura celular, pero se diferencian mucho por su historia evolutiva o filogenia. Por estudios de evolución molecular en procariotas, específicamente las relaciones filogenéticas deducidas luego de comparar las secuencias de macromoléculas esenciales, en especial los RNAs ribosómicos, se llegó a la conclusión que Archaea y Bacteria no están relacionadas evolutivamente. Mas aún, las especies de Archaea están más relacionadas con los eucariotas que con las especies del dominio Bacteria. Este hecho ha sido ratificado por estudios comparativos realizados con otras macromoléculas de especies de los tres dominios. Por lo tanto, la diversificación evolutiva, a partir del ancestro común fue en dos direcciones, hacia Bacteria y hacia otra cosa que finalmente se diversificó dando origen a los dominios Archaea y Eukarya. 68 Árbol filogenético de la vida construido a partir de la comparación de las secuencias del RNA ribosómico. El árbol está formado por tres dominios de organismos: Bacteria y Archaea (que presentan células procarióticas) y Eukarya (células eucarióticas). El grupo en rosa son macroorganismos, el resto de los organismos son microorganismos. Como todas las células de los animales y plantas son eucarióticas, se deduce que los microorganismos eucarióticos fueron los precursores de los organismos pluricelulares. 69 Secuenciación del gen de RNA ribosómico y filogenia a) Se rompen la células; b) se aisla el gen que codifica el RNA ribosómico y se amplifica el número de copias por la técnica de PCR (reacción en cadena de la polimerasa); c) el gen es secuenciado; d) las secuencias obtenidas se alinean por computadora; Un programa informático realiza comparaciones por pares y genera un árbol (e) que refleja las diferencias en la secuencia del RNA ribosómico del organismo analizado. Si en el análisis se usa una muestra natural, los genes del RNA ribosómico aislados de los diferentes organismos de una muestra deben ser clonados antes de ser amplificados y secuenciados. 70 Micrografías electrónicas de secciones de células pertenecientes a cada uno de los tres dominios de organismos: a) Bacteria, b) Archaea y c) Eukarya. 71 estructura Los organismos se definen por estudios de composición genética comportamiento Los microbiólogos usan el sistema de nomenclatura binomial desarrollado por Linneo para plantas y animales. Serie de organismos relacionados se le da un nombre o “género” dentro de este género se separan las distintas “especies” u organismos de diferente tipo. Por ej. Escherichia coli, Brucella abortus, Brucella melitensis, Brucella suis, Agrobacterium tumefaciens, Agrobacterium rhizogenes, Bacillus anthracis, Bacillus subtilis, Sinorhizobium meliloti, etc. En general, los nombres derivan del latín o griego latinizado que describen alguna propiedad típica de la especie y se escriben en letras itálicas o cursivas. Por ej. Se han descripto más de 100 especies diferentes del género Bacillus, tales como Bacillus subtilis, Bacillus cereus y Bacillus tearothermophilus. Estos nombres específicos significan “fino”, “ceroso” y “amante del calor”. En general describen las características morfológicas, fisiológicas o ecológicas importantes de c/u 72 Los microorganismos y sus ambientes naturales Población: grupo de células relacionadas. Hábitat: ambiente natural en el que vive una población. Ecología: estudio de los organismos en sus ambientes naturales. Comunidades: conjuntos de poblaciones que viven juntas en un mismo hábitat. Ecosistema: es el conjunto de los organismos y de los componentes físicos y químicos del medio. Interacciones entre poblaciones beneficiosas perjudiciales Los MOs también interaccionan con el medio ambiente de su hábitat y pueden modificarlo. La comunidad en un hábitat determinado está definida en gran parte por las características físicas y químicas de ese medio. Estas características pueden cambiar con el tiempo debido al metabolismo propio de los organismos. Los MOs son parte importante de los ecosistemas. 73 Con el tiempo un ecosistema microbiano puede cambiar gradualmente, tanto desde el punto de vista físico como químico. El oxígeno es un buen ejemplo. El O2 es un nutriente vital para algunos MOs mientras que es venenoso para otros. La actividad de MOs aeróbicos al consumir en algunos casos el oxígeno puede cambiar el hábitat a condiciones anóxicas y permitir de este modo el desarrollo de otros organismos anaeróbicos que no lo podían hacer en un primer momento. 74 Ejemplos de comunidades microbianas a) Micrografía de una comunidad bacteriana que se desarrolla en las profundidades de un lago pequeño. Se muestran células de varios tamaños. b) Comunidad microbiana en una muestra de sedimentos de aguas residuales. La muestra se tiñó con una serie de colorantes, cada uno de ellos tiñe un grupo bacteriano diferente. 75 La importancia de la vida microbiana -Número total de células microbianas en la (procariotas) es aproximadamente 5x1030 células. Tierra -Cantidad total de C presente aproximadamente igual al de todas las plantas de la tierra (biomasa). -Contenido de N y P, 10 veces mayor que la de toda la biomasa vegetal. Implica que las células procarióticas constituyen la mayor porción de biomasa sobre la Tierra y son la reserva de nutrientes esenciales para la vida. La mayor parte de las células procarióticas se encuentra bajo la superficie en los ambientes oceánicos y terrestres. 76 El impacto de los microorganismo sobre el hombre Hay que conocer como trabajan los MOs para diseñar metodologías tendientes a aumentar los beneficios y disminuir los efectos perjudiciales . Aunque muchos sólo consideran a los MOs en el contexto de las enfermedades infecciosas, en realidad sólo unos cuantos causan enfermedades. Los MOs afectan muchos aspectos de nuestras vidas, además de ser los agentes etiológicos de enfermedades. Microorganismos y agricultura. Nuestros sistemas de agricultura dependen en general de las actividades microbianas como es el caso de las leguminosas que viven en estrecha asociación con bacterias específicas que forman estructuras llamadas nódulos en sus raíces donde fijan el N2 atmosférico el cual es utilizado por las plantas para crecer. 77 Los microorganismos (MOs) son también esenciales en el proceso digestivo de los rumiantes, como las vacas y las ovejas. En el órgano especial llamado rumen las bacterias realizan el proceso digestivo. Los MOs desempeñan funciones críticas en el reciclado de elementos importantes para la nutrición vegetal, en particular del carbono, nitrógeno y azufre. En el suelo y el agua los MOs convierten estos elementos en formas asimilables por las plantas. Los MOs también son causantes de enfermedades en plantas y en animales. 78 Microorganismos y alimentación Los productos agrícolas o los animales deben llegar a los consumidores con calidad sanitaria, de allí la importancia de los MOs en la industria alimentaria. El deterioro de alimentos por la actividad de los MOs ocasiona enormes pérdidas económicas. Las industrias de enlatado, congelado y desecado de alimentos tiene por finalidad evitar esto. También es importante las enfermedades que pueden ser transmitidas a través de los alimentos. Algunos alimentos requieren para su preparación la actividad de determinados MOs. Este es el caso de los productos lácteos que se manufacturan, como el queso, el yogurt o la manteca. Los alimentos de panadería se elaboran usando levaduras al igual que algunas bebidas alcohólicas. 79 Microorganismos, energía y medio ambiente Los MOs desempeñan funciones clave en lo que respecta a la energía. La mayor parte del gas natural (metano) es un producto bacteriano, derivado de las actividades de las bacterias metanogénicas. Los MOs fotótrofos pueden utilizar la luz como fuente de energía para la formación de biomasa, es decir la energía acumulada en forma de organismos vivos. La biomasa microbiana y los desechos, como la basura doméstica, los excedentes de cosechas y los residuos animales, se pueden convertir en “biocombustibles”, como el metano y el etanol, por las actividades degradativas de los MOs. Los MOs también pueden ayudar para eliminar la polución originada por las actividades humanas, un proceso que se denomina biorremediación. Este área está en pleno desarrollo en la actualidad. 80 Los microorganismos y el futuro El uso de MOs genéticamente modificados para poder sintetizar productos específicos de alto valor comercial dio origen a la Biotecnología. La biotecnología depende en gran medida de la ingeniería genética, una disciplina que trata de la manipulación artificial de genes y de sus productos. También se pueden hacer genes completamente artificiales. Una vez que se tiene el gen por síntesis o por selección a partir de una fuente natural, el mismo se puede insertar en un MO y se puede expresar allí originando el producto génico deseado. Este es el caso de la insulina humana, hormona que se encuentra en cantidades anormalmente bajas en sujetos con diabetes. Esta hormona se prepara hoy en día a partir del gen de la insulina humana expresado en un MO. 81 Influencia de los microorganismos en las actividades humanas 82 83 Los microorganismos como agentes etiológicos de enfermedades Se estudió la frecuencia de las diez causas de muerte mas usuales en USA en el año 1900 y en el año 2000 (Datos del Centro Nacional de Estadísticas de la salud de USA ). En el año 1900 las enfermedades infecciosas eran las principales causas de muerte en USA. Hoy en día son las minoritarias. Sin embargo los MOs pueden ser aún un riesgo importante para la supervivencia por ej. en pacientes inmunodeprimidos o con SIDA. Aún hoy las enfermedades microbianas constituyen la principal causa de muerte en los países subdesarrollados. La viruela se erradicó, pero aún continua la malaria, tuberculosis, cólera, enfermedad del sueño, síndromes diarreicos severos, etc. 84 Frecuencia de las 10 causas mas importantes de muertes en los EEUU 85 Conocimiento de los procesos de infección Control de Mejora de las prácticas sanitarias enfermedades Infecciosas Descubrimiento y uso de nuevos agentes antimicrobianos 86 FIN 87 Raíces históricas de la Microbiología Se sospechaba que había criaturas demasiado pequeñas para ser percibidas a simple vista. El desarrollo del microscopio permitió verlas. -1664 Robert Hooke describió los cuerpos fructíferos de mohos. -1684 Antonie van Leeuwenhoek diseñó un microscopio de lente simple y fue capaz de ver microorganismos tan pequeños como los procariotas. Lo comunicó en la Royal Society of London. En las figuras siguientes se muestran el microscopio utilizado por Robert Hooke, el microscopio diseñado por Leeuwenhoek y dibujos de algunos organismos observados por ambos. 88 Fotografía de una copia del microscopio de Leeuwenhoek 89 Replica en bronce del microscopio de Leeuwenhoek. Se muestra también como se sostiene el mismo. 90 Dibujos de van Leeuwenhoek de bacterias publicados en 1684 a las que llamó “animálculos”. podemos reconocer varios tipos morfológicos de bacterias comunes. A, C, F y G formas bacilares; E formas esféricas o cocos; H grupos de cocos. 91 92 Dibujo que representa un moho azul creciendo sobre la superficie de un trozo de cuero. Microscopio utilizado por Robert Hooke 93 Recién en el siglo XIX se contó con una amplia distribución de microscopios mejorados. La microbiología como ciencia se desarrolló a fines del siglo XIX con el desarrollo del microscopio y de técnicas básicas de laboratorio para el estudio de microorganismos. -Microscopios mejorados -Distintas técnicas de tinción de microorganismos -Esterilización -Cultivos de enriquecimiento o cultivos selectivos -Medios sólidos con gel o agar -Cajas de Petri -Tapones de algodón 94 Preguntas básicas que se debían contestar 1) ¿Se da la generación espontánea? Es decir los organismos vivos se pueden generar de materia inorgánica. 2) Se sabía que las enfermedades infecciosas se transmitían de un individuo a otro, pero no se conocían los mecanismos de esa transmisión. Las preguntas fueron contestadas por el químico francés Louis Pasteur y por el fundador de la microbiología médica Robert Koch respectivamente. 95 El botánico Ferdinand Cohn fundó la bacteriología (el estudio de las bacterias). Descubrió las endosporas estudiando el género Bacillus. Cohn describió el ciclo de vida completo de Bacillus célula vegetativa endospora célula vegetativa Descubrió que por ebullición solo morían las células vegetativas. Cohn aportó las bases experimentales para un esquema de clasificación de bacterias y fundó una revista científica importante. Ideó métodos simples para evitar la contaminación de medios de cultivo estériles, como el uso de matraces y algodón para cerrar los tubos. Estos métodos fueron utilizados posteriormente por Koch permitiéndole aislar y caracterizar varias bacterias causantes de enfermedades. 96 Pasteur y el fin de la generación espontánea Comida fresca A se deja a la la comida de donde vienen intemperie se pudre los MOs B Observación microscópica En A no se observan MOs En B se observan MOs Se originan espontáneamente Sustancia inanimada 2 hipótesis célula viva Provienen de semillas o gérmenes que entran a los alimentos provenientes del aire (hipótesis apoyada por Pasteur) 97 Pasteur demostró primero que existen en el aire estructuras muy similares a las encontradas en las sustancias putrefactas. Concluyó que los microorganismos se originan a partir de MOs presentes en el aire. Postuló que dichas células en suspensión se depositaban constantemente sobre todos los objetos. Si esto era correcto, entonces no debería estropearse un alimento tratado de tal modo que todos los organismos que lo contaminaran fueran destruidos. Pasteur empleó calor para eliminar los contaminantes. Tomó un matraz de vidrio con los nutrientes, lo selló, lo calentó a ebullición y observó que nunca se descomponía el medio de cultivo. Los defensores de la generación espontánea criticaron el experimento argumentando que hacía falta aire fresco y que el aire caliente se modificaba. Pasteur entonces modificó el experimento utilizando un matraz de vidrio de cuello de cisne. 98 Experimento de Pasteur con matraces de cuello de cisne Esterilización del contenido del matraz. 99 Si el matraz se mantiene en posición vertical no hay crecimiento microbiano 100 Si los microorganismos atrapados en el cuello alcanzan el líquido estéril, crecen rápidamente. Los principios de Pasteur se utilizaron en el envasado y conservación de alimentos. Pasteur desarrolló vacunas para enfermedades como el carbunco, el cólera aviar y la rabia (1880-1890). 101 Koch y la teoría microbiana de las enfermedades infecciosas. Siglo XIX Se pensaba que una persona enferma A → transmitía “algo” “contagio” →persona sana B → persona B enferma. Se sospechaba que los MOs podrían ser los responsables de enfermedades, pero faltaban las pruebas definitivas. MOs → ¿causa o efecto de las enfermedades? 102 La teoría microbiana de las enfermedades infecciosas fue probada por el médico Robert Koch (1843-1910). Koch estudió el carbunco, enfermedad causada por la bacteria Bacillus anthracis formadora de endoesporas. Sus estudios dieron origen a los postulados de Koch que demostraron que un tipo concreto de MO es el agente etiológico de una enfermedad específica. Por estudios cuidadosos de microscopía demostró que: 1- La bacteria B. anthracis estaba presente en la sangre de todos los animales enfermos y no así en los sanos, pero la mera asociación de la bacteria con la enfermedad no demostraba que la bacteria fuera la causa de la enfermedad, por el contrario, podría ser un efecto de la enfermedad. 103 Koch demostró que la enfermedad podía ser transmitida de un animal enfermo a uno sano inyectando una pequeña cantidad de sangre. Esto lo podía repetir en otro ratón y así sucesivamente. En todos los casos podía demostrar por microscopía en muestras de sangre de los animales enfermos la presencia de las bacterias formadoras de esporas características. Koch pudo además cultivar el MO fuera del animal en caldos nutritivos e incluso luego de numerosas resiembras o transferencias de cultivo, la bacteria podía causar aún la enfermedad cuando se reinoculaba a un animal. Es decir, la bacteria procedente de un animal enfermo y la mantenida en cultivo inducían los mismos síntomas de enfermedad tras su inoculación. Basándose en éste y en otros experimentos, Koch formuló los siguientes criterios, conocidos en la actualidad como postulados de Koch para demostrar que un tipo concreto de MO es el agente etiológico de una enfermedad. 104 Postulados de Koch 1- El organismo debe estar siempre en los animales que sufren la enfermedad y no en individuos sanos. 2- El organismo debe cultivarse en cultivo axénico o puro fuera del cuerpo del animal. 3- Cuando dicho MO se inocula a un animal susceptible, debe iniciar en él los síntomas característicos de la enfermedad. 4- El organismo debe aislarse nuevamente de estos animales experimentales y cultivarse nuevamente en el laboratorio, tras lo cual debe mostrar las mismas propiedades que el MO original. Los postulados se resumen en la siguiente figura. Usando estos postulados como guía otros investigadores revelaron la causa de muchas enfermedades importantes para el hombre y para los animales. 105 106 Medios sólidos, la placa de Petri y los cultivos puros Koch fue el primero que cultivó bacterias en medios de cultivo sólidos. Empleó gelatina como agente solidificante de diversos caldos nutritivos que usaba para cultivar bacterias patógenas. Fue reemplazado mas tarde por el agar (polisacárido derivado de algas rojas) que es mas versátil y permite utilizar temperaturas mas elevadas de incubación. El primer uso del agar como agente solidificante se debe a Walter Hesse. La técnica utilizando agar fue adoptada por Koch para sus estudios clásicos sobre el aislamiento de la bacteria Mycobacterium tuberculosis, agente etiológico de la tuberculosis. En 1887 Richard Petri publicó un trabajo corto describiendo una modificación de las técnicas de láminas horizontales de Koch. La mejora de Petri resultó ser enormemente útil. 107 Las ventajas de las cajas de Petri: -Fácilmente almacenadas y esterilizadas independientemente del medio. - Luego de añadir el medio fundido a la mas pequeña de las dos tapaderas circulares, la de mayor tamaño podía usarse como tapa para evitar contaminaciones. -Las colonias que se formaban en la superficie del agar quedaban expuestas al aire y podían ser fácilmente manipuladas para su estudio. Las cajas de Petri ya sea de vidrio o de plástico son utilizadas en la actualidad y constituyen una de las principales herramientas en los laboratorios de microbiología, junto con los microscopios, las técnicas de tinción, los medios de enriquecimiento, los tapones de algodón, los cultivos axénicos, etc. 108 A B Transferencia aséptica (A) y Método para hacer una extensión en placa para obtener cultivos axénicos o puros (B). Colonias de Micrococcus luteus cultivadas en placas de agar sangre. 109 110 111 Variaciones en la morfologia de una colonia vista a simple vista 112 Koch observó que sobre el medio sólido expuesto a contaminantes se desarrollaban colonias con formas diferentes (diferían en color, forma, tamaño u otros caracteres) y que tales colonias se podían perpetuar y diferenciar entre sí por sus características particulares. Las células de diferentes colonias diferían microscópicamente y a menudo también en sus temperaturas óptimas de crecimiento o en sus requerimientos nutricionales. Koch señaló que “toda bacteria que mantenga las características que la diferencian de otras, cuando se cultivan en el mismo medio y bajo las mismas condiciones, debería ser designada como especie, variedad, forma o cualquier otra designación adecuada”. Los trabajos de Koch permitieron que la microbiología fuera aceptada como una ciencia biológica independiente. 113 Koch y la tuberculosis. El mayor logro de Koch en la bacteriología médica está relacionado con la tuberculosis. Para aislar el agente causante de la enfermedad Koch empleó todos los métodos que se habían desarrollado previamente: microscopía, tinción de tejidos, aislamiento en cultivo puro e inoculación en animales. Diseñó un método de tinción en muestras de tejido usando azul de metileno alcalino y un segundo colorante (marrón de Bismark) que teñía solo el tejido. El método de Koch fue el precursor del método de Ziehl-Nielsen usado hoy para teñir bacterias ácido-alcohol resistentes como el Mycobacterium tuberculosis. Para la obtención de cultivos de M. tuberculosis se utilizó suero de sangre coagulada. Koch pudo obtener cultivos de este MO a partir de diversas fuentes de origen humano y de animales. Con el MO aislado pudo demostrar que era el causante de la tuberculosis. Koch recibió el Premio Nobel de fisiología y medicina en 1905. 114 Dibujos de Koch de células de Mycobacterium tuberculosis a) Sección transversal de un tubérculo en tejido pulmonar. Las células de Mycobacterium tuberculosis se tiñen en azul mientras que las del tejido pulmonar se tiñen en marrón. b) Células de M. tuberculosis en esputo de paciente tuberculoso c) Crecimiento en cultivo puro de M. tuberculosis en placa de vidrio con suero sanguíneo coagulado. d) Se tomó una colonia de la placa anterior y se observó al microscopio a 700 aumentos. Las células aparecen formando largas “cuerdas”. a b c d 115 Fotografía coloreada a mano de colonias formadas sobre agar, realizada por Walter Hesse, colaborador de Robert Koch. Las colonias corresponden a hongos (mohos) y bacterias que se obtuvieron en estudios iniciados por Hessen sobre el contenido microbiológico del aire de Berlín, Alemania en 1882. 116 117 118 Diversidad microbiana: el holandés Martinus Beijerinck y el ruso Sergei Winogradsky fueron microbiólogos pioneros en el estudio de las bacterias del suelo y del agua y se destacaron por sus importantes contribuciones al conocimiento de la diversidad microbiana. Beijerinck: formulación de cultivos de enriquecimiento o cultivos selectivos para aislar microorganismos específicos relacionados fisiológicamente a partir de muestras naturales. Aisló los primeros cultivos puros de muchos MOs del suelo y del agua (bacterias aerobias fijadoras de N2 como el Azotobacter, reductoras del SO4=, oxidantes del azufre, bacterias fijadoras de N2 en los nódulos radiculares, especies de Lactobacillus, algas verdes, etc). 119 Dibujo realizado por la hermana de Beijerinck mostrando células de Azotobacter chroococcum, bacteria aerobia fijadora de N2. Este dibujo era usado para dar clases en una época en que no existían transparencias, diapositivas y cañones de proyección con computadora Azotobacter vinelandii: a) células vegetativas b) cistos 120 Beijerinck describió los principios básicos de la virología. En sus estudios sobre la enfermedad del mosaico en el tabaco puso de manifiesto, mediante el uso de técnicas de filtración selectiva, que el agente infeccioso (un virus) no era una bacteria y que se incorporaba a las células de la planta y necesitaba que ésta estuviera viva para reproducirse. Winogradsky logró aislar por primera vez varias bacterias importantes del suelo, en particular las implicadas en los ciclos del nitrógeno y del azufre, por ej. Las bacterias nitrificantes (oxidación de NH3 a NO3-), bacterias oxidantes del azufre. Postuló el concepto de quimiolitotrofía, es decir, la oxidación de compuestos inorgánicos acoplada a la liberación de energía utilizable. Demostró que las bacterias nitrificantes obtenían su carbono del CO2 del aire, es decir que eran autótrofos. 121 Dibujos coloreados a mano de células de bacterias fotosintéticas rojas del azufre incluidas en la monografía Microbiology du Sol, de Sergei Winogradsky. Se muestran células del género Chromatium, como C. kenii (Fig. 3 y 4) y de C. vinosum (Fig 5-8). Micrografía de campo claro de bacterias rojas del azufre Chromatium okenii 122 123 124 125 126 127 128 129 130 131 132 Subdisciplinas más importantes de la microbiología. Microbiología médica e inmunología Microbiología agrícola Citología Microbiología industrial Bioquímica microbiana Microbiología acuática Genética bacteriana Ecología microbiana Biología molecular Sistemática bacteriana Bacteriólogos (virus) Fisiología bacteriana Biotecnología 133 Desarrollo de las subdisciplinas más importantes de la microbiología. Desarrollo rápido de la microbiología → Básica y aplicada 1- Trabajos de Koch → expansión de la microbiología médica y de la inmunología (en la primera parte del siglo) -Descubrimiento de muchas bacterias patógenas nuevas. -Establecimiento de los principios por los cuales estos patógenos infectan el cuerpo y se hacen resistentes a sus defensas. 2-Avances prácticos en el campo de la microbiología agrícola -Impulsores fueron Beijerinck y Winogradsky -Comprensión de los procesos microbianos en el suelo. Son beneficiosos o perjudiciales para el crecimiento de las plantas. 134 3- Se abrió el uso de los MOs para la obtención de productos industriales (síntesis de antibióticos, etc) y dio origen a la microbiología industrial luego de la segunda guerra mundial. 4- Microbiología acuática. Tratamiento de aguas residuales, suministro de agua potable para uso humano. 5- Ecología microbiana. Biodiversidad, actividades de los MOs en su hábitat. Los conocimientos de los principios básicos de la función microbiana permitió descubrir y clasificar muchas clase de MOs nuevos dando origen a la extensión de la 6-Sistemática bacteriana. 135 7- Fisiología microbiana. Estudio de los nutrientes que requieren los MOs y productos que se originan. 8- Citología. Conocimiento de la estructura física y química de los MOs. 9- Bioquímica. Metabolismo , enzimas y reacciones que llevan a cabo los MOs. 10- Genética bacteriana. Herencia, variación genética, intercambio genético. Conocimiento del DNA, RNA, síntesis de proteínas. 11- Bacteriófagos. Estudio de los virus. Analogía con la transferencia genética. 12- Biotecnología 13- Análisis genómico. Relaciones evolutivas. 14- Microbiología industrial. 136 137 Opciones metabólicas para la obtención de energía 138 Diversidad fisiológica de los microorganismos Energía y carbono Todas las células requieren energía, la cual se puede obtener de tres modos: a partir de compuestos orgánicos, a partir de compuestos inorgánicos o a partir de la luz. La oxidación de compuestos orgánicos o inorgánicos produce adenosina trifosfato (ATP) en los organismos quimiotróficos, mientras que la conversión de energía solar en energía química (también en forma de ATP) se produce por los organismos fototróficos. Los MOs aerobios son los que pueden obtener energía del compuesto en presencia de oxígeno. Otros obtienen energía en ausencia de oxígeno (anaerobios). Otros utilizan los compuestos orgánicos tanto en presencia como en ausencia de oxígeno. 139 Los organismos que obtienen energía a partir de compuestos orgánicos se los llama quimioorganotrofos y los que lo hacen a partir de compuestos inorgánicos se llaman quimiolitotrofos y al metabolismo empleado quimiolitotrofía. Este tipo de metabolismo energético se encuentra solo en procariotas y está ampliamente distribuido tanto en especies de Bacteria como de Archaea. El rango de compuestos inorgánicos que se pueden utilizar es amplio, sin embargo un determinado procariota suele especializarse en la utilización de uno o de un grupo de compuestos inorgánicos relacionados. Esta cualidad le da la ventaja de no competir con los organismos quimioorganotrófos y poder utilizar además productos de desecho de estos últimos como el H2 y el H2S. Los MOs fototróficos contienen pigmentos fuertemente coloreados que les permiten usar la luz como fuente de energía. 140 No existe competencia de los organismos fototróficos con los quimiotrófos por la energía luminosa. Fuente de carbono Todas las células requieren carbono como nutriente principal. Las células microbianas son heterotróficas si requieren uno o más compuestos orgánicos como fuente de carbono, o autótrofas si la fuente de carbono es el CO2. Los quimioorganotrofos son también heterótrofos. Por el contrario, muchos quimiolitotrofos y prácticamente todos los fotótrofos son autótrofos. Los autótrofos se denominan también productores primarios porque sintetizan materia orgánica a partir de CO2, tanto para su propio beneficio como para el de los quimioorganototrofos. Estos últimos se alimentan directamente de los productores primarios o viven a expensas de los productos que ellos excretan. 141 Resumen: Todas las células necesitan fuentes de energía y de carbono. Organismo Fuente de energía fotótrofo Luz quimiolitótrofo Compuestos inorgánicos quimioorganotrofo Compuestos orgánicos 142 Organismo Fuente de carbono fotótrofo Prácticamente todos utilizan CO2 como fuente de C, (autótrofos). quimiolitótrofo En la mayoría de los casos utilizan CO2 como fuente de C. Pueden ser autótrofos o heterótrofos. quimioorganotrofos Utilizan compuestos orgánicos como fuente de C. Son heterótrofos. 143 Tolerancia a condiciones ambientales extremas Los microorganismos, en particular los procariotas tienen la capacidad de poder vivir en hábitat caracterizados por uno o más parámetros extremos en las condiciones ambientales, por ejemplo aguas termales a ebullición, en el hielo, en aguas de elevada salinidad, y en suelos y aguas que tienen un pH inferior a 0 o tan alto como 12. En ocasiones no es que sean meramente tolerantes a esas condiciones extremas, sino que realmente las requieren para crecer. Estos procariotas se denominan extremófilos. En los hábitat que presentan condiciones extremas, los procariotas son siempre mayoritarios. 144 El microorganismo citado en cada categoría es el que tiene el récord actual en cuanto a requerir una condición extrema particular para crecer. Picrophilus oshimae también es un termófilo, con crecimiento óptimo a 60oC. Natronobacterium gregoryi es un halófilo extremo, con crecimiento óptimo a NaCl 20%. La cepa MT41 aún no tiene nombre oficial de género ni especie y también es un psicrófilo, crece mejor por debajo de 10oC. 145 FIN 146 La Tierra primitiva y el origen de la vida Se cree que la tierra tiene una edad de 4600 millones de años; la primera evidencia de vida microbiana se observa en rocas de 3860 millones de años. La Tierra primitiva era anóxica y mucho mas caliente que en la actualidad. Los primeros compuestos bioquímicos se formaron por síntesis abiótica y esto estableció las bases para el origen de la vida. Si bien no conocemos como era el primer organismo capaz de autorreplicarse, a partir de lo que sabemos de las formas microbianas actuales podemos afirmar que las entidades autorreplicativas más simples necesitaban un mecanismo para obtener energía y alguna forma de herencia para hacer copias de si mismas. Podría extrapolarse desde el presente hacia el pasado y postular que los organismos primitivos se parecían a las células modernas, aunque con muy pocos genes con limitadas capacidades para la transcripción y la traducción. Tras el descubrimiento de que ciertos tipos de ácido ribonucleico (RNA) son catalíticos, muchos científicos actuales creen que las formas de vida más primitivas carecían totalmente de DNA, contenían tan sólo unas cuantas, si es que alguna, proteína, y consistían principalmente de RNA; era la edad de la vida del RNA, donde el RNA era tanto agente catalizador como el código genético. 147 La vida con RNA. Las moléculas de RNA habrían funcionado con el fin exclusivo de replicarse a sí mismas y, probablemente, habrían llevado a cabo un número mínimo de reacciones catalíticas necesarias para esta finalidad. Estudios derivados de varias ribozimas catalíticas de RNA han demostrado que pueden catalizar varias reacciones, tales como la síntesis de nucleótidos. Así, un período de vida con RNA podría haber precedido a la vida celular. Estas formas de vida con RNA podrían haber evolucionado hacia las primeras formas de vida celulares, cuando el RNA autorreplicativo quedara encerrado en vesículas de lipoproteína. Estas estructuras parecidas a las células podrían haber surgido mediante la agregación espontánea de moléculas de lípidos y de proteínas para formar estructuras membranosas en cuyo interior quedaron atrapados los RNA y otros precursores de biomoléculas clave. Esta etapa pudo haber ocurrido un sinnúmero de veces en la Tierra primitiva para volver a extinguirse, pero finalmente, los constituyentes apropiados y las circunstancias adecuadas coincidieron y surgió un organismo celular primitivo. Aunque carecía de DNA y procesaba tan sólo proteínas producidas abióticamente, esta forma de vida celular se asemejaría en otros aspectos a una célula moderna. A medida que estas formas de vida se extendían, la selección natural las conduciría hacia su posterior desarrollo evolutivo. 148 Escenificación hipotética de la evolución de las formas de vida celulares a partir de las formas de vida con RNA. Los RNAs autorreplicativos podrían haber llegado a ser entidades celulares mediante su integración estable en vesículas de lipoproteína. Con el tiempo, las proteínas reemplazaron las funciones catalíticas del RNA y el DNA reemplazó sus funciones de codificación 149 Las proteínas muestran un grado de especificidad catalítica mucho mayor que el que poseen los RNAs catalíticos. A medida que los organismos primitivos se hicieron bioquímicamente mas complejos, se produjo una presión evolutiva hacia la utilización de las proteínas como principales agentes biocatalíticos. La célula moderna: DNA → RNA → Proteína El establecimiento del DNA como genoma de la célula pudo ser una consecuencia de la presión evolutiva hacia una mayor eficacia y fidelidad en la replicación de la información genética (las DNA polimerasa son mas precisas que las RNA polimerasas). Además, al guardar toda la información genética en un lugar en la célula y procesar solamente la que se necesita bajo unas determinadas condiciones (al regular la expresión génica), las células habrían ahorrado energía, lo que incrementaría su adecuación competitiva. De alguna manera, en los estadios tempranos de la evolución microbiana, el sistema tripartito DNA-RNA-proteína quedó fijado en las formas de vida celulares como la mejor solución al procesamiento de la información biológica. A juzgar por las células actuales, este sistema fue un éxito evolutivo: todas las células conocidas contienen estos tres tipos de macromoléculas. 150 Metabolismo de los organismos primitivos La vida es un proceso altamente ordenado y requiere de energía para mantener su estructura. Para satisfacer los requerimientos energéticos los microorganismos primitivos sólo podían recurrir a mecanismos generadores de energía que tienen lugar en condiciones anóxicas ya que hasta la aparición de las cianobacterias, el oxígeno molecular no existía en cantidades significativas en la Tierra. Una simple reacción química en la que participa el ión ferroso (abundante en la Tierra primitiva) pudo haber sido un proceso primitivo de obtención de energía utilizado por los primeros organismos. ΔGo´= -42kj / reacción se La reacción FeS + H2S → FeS2 + H2 produce exotérmicamente con liberación de energía. Esta reacción también produce H2, y se ha propuesto que este H2 pudo haber sido utilizado por las células primitivas para originar una fuerza motriz de protones a través de una membrana, de la que una ATPasa primitiva podría haber extraído energía químicamente útil como ATP. Con H2 como el donor de electrones, se requiere su aceptor, que bien podría haber sido azufre elemental, So. Esta sencilla reacción acoplada habría requerido pocas enzimas y podría haber constituido un suministro ilimitado de energía en tanto existiera FeS accesible. 151 Esquema de un hipotético sistema generador de energía de las células primitivas. La formación de piritas conduce a la producción de H2 y la reducción de So, los cuales alimentan una ATPasa primitiva. Nótese que el SH2 tiene una función catalítica. Los sustratos netos serían FeS y So. 152 Es sorprendente que muchas Archeae hipertermófilas (que son los organismos actuales más cercanos a los organismos primitivos de la Tierra) pueden llevar a cabo este tipo de reacción. Los organismos primitivos pudieron haber obtenido carbono de varias fuentes, tales como carbono orgánico de síntesis abiótica e incluso CO2 , un gas que era abundante en la Tierra primitiva. La utilización de CO2 continuó evolucionando hacia la autotrofía, el proceso en el cual el dióxido de carbono se convierte en todos los componentes orgánicos de la célula. La hipótesis de una “autotrofía primitiva” se sustenta en los proyectos de secuenciación de genomas microbianos, donde se ha puesto de manifiesto que la autotrofía se produce en un número de hipertermófilos que contienen un genoma pequeño que se ramifican cerca de la raíz del árbol evolutivo de la vida. 153 Oxigenación de la atmósfera La evolución de la fotosíntesis oxigénica en las cianobacterias constituye un hito en la historia de la Tierra. Estos organismos aparecieron probablemente hace 3000 millones de años, pero el O2 que produjeron no se acumuló en la atmósfera debido a la gran cantidad de sustancias reductoras (como el FeS) todavía presentes que reaccionaban espontáneamente con el O2 para formar H2O. Es altamente probable que las cianobacterias evolucionaran a partir de protótrofos oxigénicos mediante el desarrollo de un fotosistema que pudiera utilizar el H2O como un donante de electrones para la reducción fotosintética del CO2, liberando O2 como subproducto (CO2 + H2O →CH2O + O2). La evolución de la fotosíntesis oxigénica tuvo enormes consecuencias sobre el ambiente de la Tierra ya que, a medida que se acumuló el O2, la atmósfera cambió de anoxigénica a oxigénica. Al existir abundancia de O2 como aceptor de electrones pudieron evolucionar los organismos aeróbicos. Estos organismos eran capaces de obtener más energía de la oxidación de compuestos orgánicos que los anaeróbicos, lo que permitió alcanzar densidades de poblaciones más altas e incrementó las posibilidades de evolución de nuevos organismos y esquemas metabólicos. 154 Existe buena evidencia , procedente del registro fósil, de que cuando la atmósfera de la Tierra se hizo oxidante, la velocidad de la evolución se incrementó enormemente, lo que condujo a la aparición de microorganismos eucarióticos con orgánulos, y, a partir de ellos, a la rápida diversificación de los metazoos (organismos pluricelulares) y, finalmente, a la aparición de plantas y animales superiores. Otra consecuencia importante de la aparición del O2 fue la formación de ozono (O3), una sustancia que suministra una barrera que impide a la intensa radiación ultravioleta del Sol alcanzar la Tierra. Cuando el O2 se somete a radiación UV de onda corta se convierte en O3, el cual absorbe fuertemente hasta longitudes de onda de 300 nm. Hasta que se formó el escudo de O3, la evolución pudo haber continuado en lugares protegidos de la radiación directa del Sol, como los océanos o debajo de las rocas, ya que la intensa radiación UV habría causado daño letal al DNA. Sin embargo, después de la producción fotosintética de O2 y el posterior desarrollo de la capa de O3, los organismos pudieron expandirse por la superficie de la Tierra, permitiendo la aparición de una mayor diversidad de organismos vivos. A continuación se muestra un resumen de etapas que pudieron producirse en la evolución biológica y en la oxigenación de la Tierra. 155 Principales acontecimientos de la evolución biológica. La posición de los estadios de la escala temporal son aproximados. Nótese como la oxigenación de la atmósfera, debida al metabolismo de las cianobacterias, fue un proceso gradual a lo largo de un período de unos 2000 millones de años. Nótese también que durante la mayor parte de la historia de la Tierra, solo existieron formas de vida microbiana. Aunque no existe un registro microfósil, la prueba microquímica de las células eucarióticas se remonta a 2700 millones de años. 156 Resumen Las primeras “formas de vida” pueden haber sido RNA autorreplicativos. Los primeros organismos celulares probablemente emplearon una estrategia simple para la obtención de la energía. El metabolismo primitivo fue anaerobio y posiblemente quimiolitotrófico, explotando las abundantes fuentes de FeS y H2S presentes. La fotosíntesis oxigénica condujo al desarrollo de un ambiente óxico y a una gran explosión evolutiva. 157 FIN 158 Orgánulos y eucariotas: endosimbiosis ¿Cómo surgió la célula eucariótica actual con su estructura interna característica: el núcleo rodeado por una unidad de membrana y los orgánulos (por ejemplo las mitocondrias y los cloroplastos)? Origen del núcleo. Las células eucarióticas primitivas eran estructuralmente simples y carecían de mitocondrias, cloroplastos y núcleo rodeado por membrana. A medida que las células en la línea eucariótica de descendencia se hicieron mayores, aparecieron el núcleo y el aparato mitótico, a la vez que el DNA, se fraccionó en unidades discretas (cromosomas). Los cromosomas pueden haber surgido para asegurar la replicación y la partición ordenada del DNA a medida que los genomas primitivos se hicieron mas grandes, hasta llegar a un punto en el que su replicación en forma de una única molécula (como en procariotas) no era viable. El desarrollo del núcleo eucariótico también facilitó la aparición de los enormes genomas que necesitaban los eucariotas microbianos de mayor tamaño (y mucho más tarde los organismos pluricelulares), e hizo posible la recombinación de genomas mediante la reproducción sexual. 159 No existe una razón obvia por la que los organismos primitivos necesitaran orgánulos tales como las mitocondrias y los cloroplastos, los cuales aparecieron probablemente más tarde. En realidad, incluso actualmente, se conocen microorganismos eucarióticos que carecen de mitocondrias y cloroplastos, lo que indica que estos orgánulos no son esenciales para la función de la célula eucariótica. Endosimbiosis. Existen pruebas concluyentes a favor de la teoría de que la célula eucariótica moderna (con orgánulos) evolucionó en etapas mediante la incorporación estable de simbiontes quimioorganotróficos y fototróficos del dominio Bacteria. Esta teoría, llamada “teoría endosimbiótica” de la evolución eucariótica, postula que una bacteria aeróbica estableció su residencia estable dentro del citoplasma de un eucariote primitivo y le suministró energía, a cambio de un ambiente protegido y un aporte constante de nutrientes. Este simbionte fue el precursor de la mitocondria moderna. De igual manera, la adquisición, por endosimbiosis de un fotótrofo oxigénico habría conferido propiedades fotosintéticas a un eucariota primitivo, de manera que ya nunca dependería de compuestos orgánicos para el suministro de energía. El endosimbionte fototrófico fue el precursor del cloroplasto. 160 Algunas células eucarióticas nunca incorporaron endosimbiontes, o si lo hicieron, luego por alguna razón lo perdieron. Pruebas que respaldan la teoría de la endosimbiosis. -Las mitocondrias como los cloroplastos contienen ribosomas que son claramente de tipo procariótico. -Los ribosomas de estos orgánulos poseen secuencias del RNA ribosómico características de determinadas Bacteria, y su función es inhibida por los mismos antibióticos que afectan la función del ribosoma de Bacteria de vida libre. -las mitocondrias y los cloroplastos también contienen una pequeña cantidad de DNA dispuesto en una forma circular covalentemente cerrada, típica de procariotas. 161 Origen de los eucariotas actuales por endosimbiosis. Nótese que los orgánulos se originan a partir de Bacteria y no de Archeae. Es poco probable que la endosimbiosis fuera un acontecimiento único y probablemente ocurriera en varios tipos celulares de la línea nuclear. Nótese como algunos procariotas primitivos nunca experimentaron endosimbiosis, o bien, perdieron sus simbiontes, pero mantuvieron las propiedades básicas de las células eucarióticas. El incremento en el tamaño de las células en la línea nuclear, condujo a la evolución de genomas de mayor tamaño y probablemente también a la evolución del núcleo permitiendo la replicación ordenada y la repartición de dichos genomas. 162 Revisión de conceptos El núcleo eucariótico y el aparato mitótico surgieron probablemente como una necesidad para asegurar la partición ordenada del DNA en los organismos con genomas grandes. Las mitocondrias y los cloroplastos, los principales orgánulos productores de energía de los eucariotas, surgieron por asociación simbiótica de procariotas del dominio Bacteria en el interior de células eucarióticas, en proceso llamado endosimbiosis. Evolución biológica y escala del tiempo. Los metazoos han dejado un registro fósil considerable y muy diverso que permite un buen conocimiento de la evolución biológica desde su aparición. Sin embargo, en las 5/6 partes de la historia de la Tierra se desarrollaron los procariotas de los cuales no se tiene un buen conocimiento evolutivo a través de registros fósiles. Esto cambió con la utilización de métodos moleculares. 163 Relaciones evolutivas entre los microorganismos Filogenia de los microorganismos deducida de los estudios moleculares de los ácidos nucleicos. Comparación de secuencias del RNA estructural de los ribosomas. Cronómetros evolutivos Se ha demostrado convincentemente que la distancia evolutiva entre dos organismos puede determinarse por las diferencias en la secuencia de aminoácidos o nucleótidos de macromoléculas homólogas aisladas de cada una de ellas. Esto se debe a que el número de diferencias en la secuencia de macromoléculas es proporcional al número de cambios mutacionales estables fijados en el DNA que codifica esa molécula en ambos organismos. La evolución se produce a medida que las mutaciones quedan fijadas en las diferentes poblaciones, siendo la biodiversidad el resultado final. 164 Elección del cronómetro evolutivo -La molécula debe estar distribuida universalmente en el grupo elegido para el estudio. -Debe ser funcionalmente homóloga en cada organismo, o sea, tener idéntica función. - Las moléculas deben poder ser alineadas en forma apropiada a fin de identificar regiones tanto con homología como con variaciones de secuencia. -- La secuencia de la molécula elegida debería cambiar con una velocidad proporcional a la distancia filogenética que se va a determinar. Y, de hecho, cuanto mayor sea la distancia filogenética a determinar, menor será la velocidad de cambio de la molécula; demasiado cambio tiende a enturbiar el registro evolutivo. 165 Se han evaluado muchas moléculas como cronómetros moleculares y con ellas se han realizado estudios de comparación de secuencias a fin de generar árboles filogenéticos. Estas moléculas son: varios citocromos, proteínas de hierro y azufre tales como las ferredoxinas, y genes de otras proteínas y de los RNAs ribosómicos. Sin embargo, los genes que codifican los RNAs ribosómicos, componentes clave del sistema de traducción; la ATPasa, complejo enzimático de membrana que puede sintetizar e hidrolizar ATP; y la RecA, proteína requerida para la recombinación genética, son los que han proporcionado la información genética más significativa sobre los microorganismos. Todas estas moléculas eran probablemente esenciales incluso para las células más primitivas y así , la variación de la secuencia en los genes que las codifican, nos permiten profundizar en el registro evolutivo. 166 RNAs ribosómicos como cronómetros evolutivos Debido a la probable antigüedad de la maquinaria sintetizadora de proteínas, los RNAs ribosómicos son moléculas excelentes para poder discernir las relaciones evolutivas entre seres vivos. Los RNAs ribosómicos son funcionalmente constantes, están universalmente distribuidos y su secuencia está moderadamente bien conservada a través de amplias distancias filogenéticas. También, dado el elevado número de diferentes secuencias posibles de moléculas grandes, tales como los RNAs ribosómicos, la similitud de dos secuencias indica alguna relación filogenética. Sin embargo, es el grado de semejanza de las secuencias de RNA ribosómico de dos organismos lo que indica su parentesco evolutivo relativo. A partir del análisis comparativo de secuencias , se pueden construir las genealogías moleculares que conducen a los árboles filogenéticos. Estos muestran la posición evolutiva más probable existente entre los organismos. Recordemos que hay 3 moléculas de RNA ribosómico, que en procariotas tienen 5S, 16S y 23S. Los rRNAs bacterianos 16S y 23S contienen varias regiones de secuencia altamente conservada que resulta útil para obtener alineamientos de secuencia apropiados. 167 Tienen también la variabilidad de secuencia suficiente en otras regiones de la molécula para servir como excelentes cronómetros filogenéticos. En eucariotas se secuencia el homólogo del rRNA 16S, es decir, el rRNA 18S de los ribosomas 80S. Hay base de datos en Internet que contienen más de 24.000 secuencias de RNAs alineadas (16.000 secuencias de 16S alineadas y 8.000 secuencias de 18S alineadas). 168 RNA ribosómico a) Micrografía electrónica de ribosomas 70S de E. coli. b) Partes del ribosoma; 5S, 16S y 23S. c) Estructura primaria y secundaria del rRNA ribosomal 16S de E. coli (Bacteria). En general el rRNA 16S de Archeae es similar en cuanto a estructura secundaria, pero presenta numerosas diferencias en cuanto a estructura primaria. La molécula correspondiente en eucariotas es el rRNA18S presente en los ribosomas citoplasmáticos. 169 FIN 170 Comparación de la transferencia de información genética en procariotas y eucariotas. (a) Procariotas. Una única molécula de RNA contiene a menudo más de una región codificadora (tal mRNA se llama policistrónico). (b) Eucariotas. Las regiones no codificantes (intrones) se eliminan del transcripto primario antes de la traducción. Los mRNA de los eucariotas son casi siempre monocistrónicos. Los dos tipos celulares no están dibujados a escala. 171 PCR. La reacción en cadena de la polimerasa puede multiplicar moléculas de DNA hasta miles de millones de veces proporcionando grandes cantidades de genes para su clonación, secuenciación o mutagénesis. La PCR hace uso de la DNA polimerasa que copia moléculas de DNA. La técnica de PCR requiere que se conozca la secuencia nucleotídica de una región del gen deseado para poder diseñar oligonucleótidos iniciadores cortos o primers, complementarios de secuencias presentes en el gen o genes de interés. Las etapas de amplificación son las siguientes: 1- En un sintetizador de nucleótidos se fabrican los dos oligonucleótidos iniciadores que flanquean al gen diana, y se añaden en un gran exceso al gen diana desnaturalizado por calor. 2- Cuando se ha enfriado la mezcla, el exceso de iniciadores relativos al DNA diana asegura que la mayor parte de las cadenas diana hibriden con iniciadores y no entre sí. 3- La DNA polimerasa alarga los iniciadores usando las bandas diana como moldes. 4Después de un período de incubación adecuado, se calienta de nuevo la mezcla para separar las cadenas. Luego se enfría la mezcla para permitir que los iniciadores se hibriden con las regiones complementarias del DNA recién sintetizado, y se repite el proceso. 172 Así, cada “ciclo” de PCR implica las siguientes etapas: 1- desnaturalización por calor del DNA bicatenario diana. 2- enfriamiento para permitir el acople de iniciadores específicos con el DNA diana 3- extensión de los iniciadores por la DNA polimerasa. Los productos de la extensión de un iniciador pueden servir de molde para el otro iniciador en el siguiente ciclo. Cada ciclo de PCR duplica literalmente el contenido original de DNA diana. En la práctica suelen hacerse unos 20 o 30 ciclos, lográndose así incrementar la secuencia diana entre 106 y109 veces. 173 174 175