- Ninguna Categoria
Funciones alométricas para el bosque mediterráneo chileno
Anuncio
Funciones alométricas
para el bosque mediterráneo
chileno
Modelos de Biomasa y Carbono
para las especies Quillay, Peumo, Espino y Litre
distribuidas en la zona de bosques
mediterráneos de Chile
San ago 2014
Fundación
undación para la
Innovación Agraria
Corporación
oración Nacional Forestal
Fores
Escuela d
de Ingeniería
í Forestal
UNIVERSIDAD MAYOR
Centro de
d EEstudios
t di de
d Recursos
R
Naturales
N t
Funciones alométricas
para el bosque mediterráneo
chileno
Modelos de Biomasa y Carbono
para las especies Quillay, Peumo, Espino y Litre
distribuidas en la zona de bosques
mediterráneos de Chile
3
San ago 2014
Autores:
Pablo Cruz
Alejandro Bascuñan
Juan Velozo
Colaboradores:
Daniela Prado
Jorge Neumann
Jesús Torralba
Javier Cano
Waldo Pérez
Paulina Vidal
Asesora estadística:
Marisol Rodríguez
Fundación
undación para la
Innovación Agraria
Corporación
oración Nacional Forest
Forestal
Escuela de Ingeniería Fo
Forestal
UNIVERSIDAD MAYOR
Centro de Estudios de Recursos Naturales
Natu
Índice
PRESENTACIÓN ...................................................................................................................... 7
1.
OBJETIVOS .................................................................................................................. 9
1.1. O
G
.................................................................................................. 9
1.2. O
............................................................................................ 9
2.
ESTADO DEL ARTE .................................................................................................... 11
3.
FASE DE CAMPO ...................................................................................................... 15
3.1. Z
............................................................................................. 15
3.2. S
............................................ 17
3.3. V
,
......................................................... 18
4.
ESTIMACIÓN DE CRECIMIENTOS DIAMÉTRICOS DEL DAP............................................... 21
4.1 M
....................................................................................................... 21
4.2 R
.......................................................................................................... 22
5.
RELACIÓN ENTRE PESO VERDE Y VARIABLES ALOMÉTRICAS MEDIDAS ............................. 25
5.1. Q
................................................................................................................ 25
5.2. P
................................................................................................................. 27
5.3. L
.................................................................................................................... 28
5.4. E
.................................................................................................................. 29
6.
PROCESAMIENTO Y ANÁLISIS DE RESULTADOS DE LABORATORIO.................................... 33
6.1. P
..................................................... 33
6.1.1 Determinación del contenido de humedad................................................ 33
6.1.2 Determinación de la densidad básica de la madera.................................. 33
6.1.3 Determinación del contenido de carbono .................................................. 33
6.1.4 Análisis estadísticos ................................................................................... 34
5
6.2. R
..................................................................... 34
6.2.1. Contenido de Humedad ............................................................................ 34
6.2.2. Densidad básica ........................................................................................ 35
6.2.3. Relación peso seco peso fresco (Ps/Pv). ................................................... 36
6.2.4. Contenido de carbono ............................................................................... 37
7.
ELABORACIÓN DE MODELOS ..................................................................................... 39
7.1 M
..................................................................... 39
7.1.1 Quillay ........................................................................................................ 40
7.1.2 Peumo ........................................................................................................ 44
7.1.3 Litre ............................................................................................................ 48
7.1.4 Espino ......................................................................................................... 50
7.1.5 Espino – Vástago ....................................................................................... 54
7.2 M
....................................................... 56
7.2.1. Quillay ....................................................................................................... 57
7.2.2 Peumo ........................................................................................................ 58
7.2.3 Espino ......................................................................................................... 58
8.
MODELOS PARA PREDICCIÓN DE CARBONO ................................................................ 59
9.
MODELOS DE BIOMASA Y CARBONO ÚTILES PARA TELEDETECCIÓN ................................ 61
10 REFERENCIAS BIBLIOGRÁFICAS .................................................................................. 61
Presentación
La Corporación Nacional Forestal (CONAF) está impulsando estudios sobre la creación de funciones
alométricas que permitan cuan ficar carbono en las
especies na vas de Chile. En este marco, se propuso
a OTERRA la confección de modelos de cuan ficación de carbono para especies del bosque mediterráneo de la zona central del país.
Las especies que componen estos bosques otorgan
posiblemente la mayor dificultad que puede encontrarse en Chile en relación a la confección de funciones de biomasa y carbono, debido principalmente
a dos factores: en primer lugar, el bosque ha sido
some do a una permanente intervención y destrucción teniendo como consecuencia que las formas
naturales de los árboles se hayan perdido, siendo
modificadas por crecimientos simpódicos, errá cos
de los más complejos de modelar con los tradicionales métodos de aproximaciones a sólidos regulares (cilindros, conos, paraboloides y otros (Gayoso
& Guerra, 2005). En segundo lugar, el agrupamiento de las especies y las interrelaciones entre ellas
no guarda un patrón natural, sino que el azar de la
destrucción que han sufrido, perdiéndose las fisionomías originales de los bosques y afectando con
ello el modelamiento normal que las copas y que el
árbol en general tenía.
La confección de funciones alométricas para es mar
biomasa y carbono (ByC) se desarrolló para cuatro
especies del Bosque Mediterráneo Chileno, quillay
(Quillaja saponaria Mol.), peumo (Cryptocarya alba
(Mol.) Looser), litre (Lithraea caustica Mol.) y espino (Acacia caven Mol.). Se abarcaron 19 zonas de
muestreo distribuidas entre las regiones V y VIII con
un total de 328 árboles procesados.
Finalmente, se realizó un análisis de crecimientos
para cada especie, y para complementar así el obje vo final que es la mejor aproximación a la determinación de capturas de carbono en este po de
bosque según su área de distribución.
7
1. Objetivos
1.1. Objetivo General
Confeccionar métodos de cuan ficación de
Biomasa y Carbono sobre el suelo en especies
del bosque mediterráneo chileno.
1.2. Objetivos específicos
•
Obtener datos empíricos de campo de
Biomasa y Carbono de especies seleccionadas para que sean la base de la búsqueda de relaciones entre variables alométricas.
•
Analizar la bondad de usar variables alométricas diferentes a las tradicionales
para las es maciones de Biomasa y Carbono, sobre la base de la fac bilidad de
capturar cada variable en terreno y las
bondades de los modelos construidos.
•
Estudiar opciones de es mación de Biomasa y Carbono agregada a unidades de
bosques y su ventaja respecto del uso de
funciones por especie.
9
2. Estado del Arte
El marco de referencia de este estudio es la generación de líneas bases de los contenidos de carbono
en ecosistemas forestales mediterráneos chilenos.
Así, es importante resaltar cual es la información
fundamental que se requiere y en los términos más
simples posibles. En primer lugar, es deseable conocer los montos de carbono que pueden contener
los pos de bosques mediterráneos existentes, vale
decir el conjunto de materiales orgánicos que lo
componen, dentro de los que son fácilmente reconocibles las partes aéreas de la vegetación, el suelo,
y las raíces de la vegetación. Un segundo cues onamiento es cuanto carbono acumulan en un período
de empo, cuya respuesta depende del crecimiento
del conjunto de componentes anteriores. Ambas temá cas resultarán tan complejas como complejos,
heterogéneos y diversos sean los sistemas que se
estudien, y dicha complejidad aumenta cuando se
considera que el contenido de carbono y el crecimiento son interdependientes.
Respecto del contenido de carbono, existen a su vez
dos temá cas que deben ser atendidas en procesos
diferentes (Picard, Saint-André, & Henry, 2012). La
primera es la evaluación del contenido de carbono
de cada uno de los componentes de la vegetación,
principalmente los componentes arbóreos y de gran
talla. En este ámbito, el bosque mediterráneo posee el 50% de las especies de la flora vascular de
los bosques del sur de Sudamérica (Armesto, Rozz,
Smith-Ramírez, & Arroyo, 1998), que implica un arduo trabajo de creación de modelos para cada una
de las especies. Obtenidos los modelos es necesario
entender cómo se organizan en la arquitectura boscosa en que par cipan, dando lugar a la vegetación
que compone el bosque mediterráneo, y en par cular a la frecuencia en que puede aparecer una u
otra especie. Si bien existe vasta información relava a la es mación de existencias en bosques, los
mediterráneos son aquellos de los más complejos
de cuan ficar, debido principalmente a dos factores
combinados: su riqueza florís ca y perturbaciones
de larga data y extremadamente heterogéneas respecto de sus efectos (Arroyo, Cavieres, Mar corena, & Muñoz, 1995; Holgrem, 2002). Schulze et al,
(2008) compararon diferentes pos de inventarios
de masas en bosques del po esclerófilo, con censos en las mismas áreas, y describieron errores de
es mación de hasta un 200%. Nótese que los inventarios con enen un error rela vo a la medición de
las frecuencias de cada una de las especies, sumado
a que dichas frecuencias posiblemente no tengan un
patrón posible de capturar con un muestreo, ya que
provienen de la casuís ca de las alteraciones antrópicas. Por tanto, la es mación de biomasa y carbono (ByC) a escala de bosque cons tuyen también un
desa o a la inves gación futura.
11
Pablo Cruz
Alejandro Bascuñan
Juan Velozo
Respecto de la acumulación de carbono, existen técnicas simples para medir los crecimientos del “cuerpo” de un individuo basándose en sus crecimientos
en grosor del tallo, el crecimiento en altura, y el
crecimiento de la copa, esta úl ma una estructura
difusa, dada su composición de hojas y ramas y ramillas. Los bosques mediterráneos también presentan par cularidades en estas tres variables. Los diámetros en bosques monopódicos pueden medirse
de forma estándar a un metro y treinta del suelo,
bajo el supuesto que esa sección del tronco ende
a aproximarse a un cilindro. Dado que los bosques
mediterráneos poseen la cualidad de rebrotar de tocón, el crecimiento se distribuye en varios pies, cuya
competencia provoca frecuentemente alteraciones
en la forma. Un caso extremo lo presenta Litrahea
caustica, que genera extensiones horizontales tan
cercanas al suelo que es imposible reconocer el
punto de origen de la cepa. Por otra parte, la existencia de varios pies sobre una misma cepa agrega
dificultad – y costo al inventario – por el hecho de
aumentar en varias mediciones la masa que estaría
contenida en un cuerpo, en comparación a una forma monopódica.
12
Respecto de la altura, hace más de un siglo fue propuesta la ley de Eichorn (1904), que indica que la
producción de masa para una especie está determinada casi completamente por el crecimiento en
altura (para una especie dada, en una región dada).
En los bosques mediterráneos también ocurren diferencias notorias en esta regla, que posiblemente
estén causadas por el cambio de fisonomía de los
árboles, provocando que se reduzca la relación de la
produc vidad con el crecimiento en altura.
La copa, como tercer componente, es un volumen
difuso que para una misma especie, podrá variar su
contenido de masa según la condición de competencia que tenga el árbol. Cruz (2013) propuso un
modelo de masa de copa para Quercus pyrenaica
Willd, con una alta e inversa relación con la densidad del bosque, según clase diamétrica, vale decir, a
mayor densidad de la vegetación, menor masa en la
copa del árbol, relación que es aparente a la vista si
se aprecia cómo cada árbol compite por un espacio
para su copa con sus vecinos. Los bosques mediterráneos y sobre todo aquellos con alteraciones, poseen
grandes espacios a su alrededor, de manera que los
tamaños de las copas están determinados por una
competencia que no pareciera ser la luz, sino que posiblemente la disponibilidad de agua en los ecosistemas. En defini va, es posible que los tamaños de las
copas no sea una buena variable de correlación con
el crecimiento, aunque si del contenido de ByC.
Los atributos descritos, descontadas y tratadas las
limitaciones comentadas, permi rán es mar la ByC
de un árbol al ser incorporadas en un modelo predictor llamado función alométrica (Chave, Andalo,
& Brown, 2005). El proceso de construcción básicamente consiste en probar estadís camente si las
variables alométricas están relacionadas con la variable dependiente y que se quiere predecir, en este
caso ByC. Es importante en este contexto discu r
dos aspectos: número de variables a escoger dentro
del modelo y el po de modelo.
El desarrollo tecnológico actual ha facilitado los
procesos y análisis que se requieren para construir
modelos, y ello ha derivado en la confección de modelos con más variables y más complejos (Prado &
Alarcón, 1991). CONAF (2013) describe un grupo de
29 variables independientes posibles de usar, entre
ellas algunas tan precisas como diámetro al inicio de
la copa sin corteza. Sin embargo, desde una perspecva opera va se recomienda priorizar la simpleza y
uso mínimo de variables ante las precisiones de los
modelos. Ello, porque el costo de recoger la información no ha cambiado con las tecnologías, y a la
fecha sigue siendo la fracción más costosa de la generación de información. Así, frente a dos modelos
es posible que siempre se escoja aquel que requiera
menos datos de terreno aunque se sacrifique precisión. Sobre todo con las consideraciones que se han
señalo rela vas a la heterogeneidad intrínseca del
bosque mediterráneo. Así también, es interesante
conservar modelos que se realizan con alguna variable con baja relación. Par cularmente el caso del
área de copa, por ejemplo, puede resultar de baja
FUNCIONES ALOMÉTRICAS PARA EL BOSQUE MEDITERRÁNEO CHILENO
potencia, pero relevante para proyectos de teledetección, que sólo cuentan con dicha variable para
predecir.
El tratamiento estadís co de los modelos está determinado por el po de relación entre las variables
independientes y aquella que se busca es mar, vale
decir, cómo cambia la variable dependiente ByC
respecto del cambio de la variable independiente.
Nótese por ejemplo que en general la altura ene
una relación lineal con el aumento de la biomasa.
Sin embargo, el diámetro relaciona con la biomasa
al cuadrado. Ambas sentencias explican que generalmente las relaciones mejores son del po DAP2 *
HT (Prodan, Peters, & Cox, 1997), y que merece dos
consideraciones. La primera es que aumentos lineales del diámetro provocarán aumentos mayores a
los lineales en el volumen/masa. Por ello, se deberá
recurrir a funciones no lineales para las es maciones. Un segundo aspecto y posiblemente de mayor
relevancia, es la heterocedas cidad que presentan
la mayoría de la variables usadas en mediciones de
volumen/masa (Pardo & Ruiz, 2005). Al respecto,
considérese lo siguiente, cuando el tamaño del cuerpo es pequeño, las diferencias entre valor real y valor es mado resultan muy pequeñas – piénsese en
diámetros de árboles pequeños -. En la medida que
los cuerpos son mayores la tendencia (la relación)
sigue verificándose, pero la variación entre valor
real/valor predicho puede ser mayor, debido a que
un árbol maduro posee un volumen afectado por
su historia de crecimiento – par cularidades de su
microsi o, su propia gené ca, los fenómenos climácos en su vida, por mencionar algunos (Prodan et
al., 1997). Este fenómeno se aprecia adecuadamen-
te en una gráfica de puntos para los datos empíricos
biomasa - DAP, si fuera el DAP la variable en análisis, donde la nube de puntos asemejará un cono
que se abre según crece la variable independiente.
Si bien siempre es posible construir un modelo que
es me la tendencia de dicha nube de puntos, la heterocedas cidad tendrá como consecuencia que las
es maciones de ByC será menos precisa mientras
mayores sean los individuos que a los que se quiere es mar su ByC. La heterocedas cidad no puede
repararse con modelos matemá cos y debe tenerse en cuenta al momento de usar alguna función.
Por ello, se recomienda observar gráficas de niveles
de confianza de los modelos de predicción que dan
una idea de la magnitud en los errores que pueden
tener las es maciones según los tamaños de los individuos.
En Chile existe profusa información de funciones
alométricas para biomasa de las especies de los bosques templados (Gayoso, 2013a, 2013b, 2013c; Gayoso & Guerra, 2005; Picard et al., 2012; Schlegel,
2001). La reciente publicación “Compendio de funciones alométricas de biomasa de especies forestales presentes en Chile” (CONAF, 2013) recopiló 21
funciones de biomasa para espino, 3 para litre y 28
para quillay, aunque ninguna específica para peumo.
Así, las funciones elaboradas en este estudio aportarán la componente de carbono que es parte del contexto que da origen a estas líneas de inves gación,
cuidando par cularmente abarcar territorialmente
la mayor proporción de la distribución actual de las
especies seleccionadas.
13
3. Fase de Campo
3.1. Zonas de muestreo
Las especies obje vo cubren una amplitud la tudinal cercana a 7 grados (Figura 3.1), par cipando en
diversas formaciones forestales, y estados de conservación.
Figura 3.1 Diagrama de distribución de las especies estudiadas, cubriendo cerca de 7 grados de la tud.
Dada la diversidad de si os que forma la fisionomía
y topogra a de la distribución de las especies en el
territorio, fue necesario analizar la per nencia de
establecer localidades que permi era capturar las
diferencias morfológicas de las especies. Si bien no
existen estudios de diferenciaciones de razas y expresiones de plas cidad para la mayoría de las especies, fue recomendable considerar a lo menos una
estra ficación la tudinal, y longitudinal, separando
la cordillera de los Andes, la depresión intermedia y
la cordillera de la costa.
Una vez definidas las zonas, se ges onaron permisos
de ingresos a los predios con la colaboración de las
oficinas regionales y provinciales de CONAF, quienes
verificaron la existencia de planes de manejo.
Así, se logró abarcar zonas de muestreos comprendidas entre las Regiones V y VIII, de manera que se
cubrió la más amplia distribución geográfica posible
para el conjunto de especies (Cuadro 3.1) (Figura
3.2).
15
Pablo Cruz
Alejandro Bascuñan
Juan Velozo
Cuadro 3.1 Distribución geográfica del muestreo y número de árboles.
Especie
Quillay
Peumo
Litre
Espino
16
Región
Localidad
R.M
V
VI
VI
VI
VI
V
VII
VII
R.M
VI
VI
R.M
VII
VIII
R.M
R.M
V
V
VI
VII
VII
R.M
R.M
V
V
V
VI
VII
San Carlos de Apoquindo
Putaendo
Lolol
San Fco. Mostazal
Pumanque
Chépica
Cajón Magdalena
Pencahue
Batuco
Curacaví
Chépica
Paine
Cajón del Maipo
Sagrada Familia
Arauco
San Carlos de Apoquindo
Curacaví
Ñilhue
Cajón Magdalena
Los Tricahues
Pencahue
Batuco
Melipilla
Curacaví
Putaendo
Ñilhue
Cajón Magdalena
Chépica
Pencahue
N
1
24
9
17
3
6
3
11
6
17
28
11
13
2
11
28
6
19
5
9
11
2
23
2
3
7
15
22
14
Figura 3.2 Ubicación de puntos de muestreo de biomasa para las cuatro especies.
FUNCIONES ALOMÉTRICAS PARA EL BOSQUE MEDITERRÁNEO CHILENO
3.2. Selección de sitios para la extracción
de árboles
Para cada zona se escogieron áreas que cumplieran
con la definición legal de bosque, es decir, al menos
un 25% de cobertura de dosel. La selección incluyó tanto individuos monopódicos como simpódicos,
que pueden haber tenido su origen en tocones anteriores o de semilla. Así también se procuró cubrir
el mayor rango de estados de desarrollo, representados por los diámetros de los tallos. Es importante
señalar que el grupo de árboles de clases superiores
a 30 cm es extremadamente escaso en la zona de
distribución, y los pocos individuos encontrados no
estaban disponibles para el estudio. En consecuen-
cia en este trabajo se han construido modelos para
la muestra más frecuente y que corresponde a árboles de hasta 35 cm de diámetro.
Para el caso de las especies quillay y peumo la distribución diamétrica incluyó diámetros a la altura del
pecho (DAP) ≥ 5 cm y para el caso de litre y espino
diámetros a la altura de tocón (DAT) ≥ 5 cm (Figura
3.3 y 3.4).
Seleccionados los individuos se midió el conjunto de
variables alométricas (Cuadro 3.2) y las mediciones
se realizaron con instrumentos y métodos convencionales de inventarios para estos bosques.
Figura 3.3 Distribución de frecuencias diamétricas para quillay y peumo.
58
58
56
56
54
54
<
<
:
17
:
8
8
6
6
4
9 < 55 58 5; 64 68 6; 75 85 88
4
9
/0
54
59
64
78
84
89
/0
Figura 3.4 Distribución de frecuencias diamétricas para espino y litre.
58
58
56
56
54
54
<
<
:
:
8
8
6
6
4
9
54 59 64 69 74 79 84 89
/0
4
8
=
58 5= 68 6= 78 7= 88
/0
Pablo Cruz
Alejandro Bascuñan
Juan Velozo
Cuadro 3.2 Variables alométricas medidas.
Variable
alométrica
en campo
Detalle de medición, precisión y límites
Observaciones
Altura total
Distancia del inicio del tallo en el suelo y hasta la parte
más alta de su copa medida con hipsómetro.
Variable tradicional.
DAP
Diámetro a la altura del pecho a 1.3 mts. Medida con
forcípula, límite inferior 5 cm.
Variable tradicional.
DAT
Diámetro a la altura de tocón a 0.2 mts. Medida con
forcípula, límite inferior 5 cm.
Variable tradicional.
Numero de vástagos
Vástagos con más de 5 cm sobre una cepa, naciendo a
menos de la altura del DAP.
Usada para probar en modelos de
cuan ficación no tradicionales, en
que el modelo se aplicó a cepas.
Diámetro de Copa (DC)
Medidos los diámetros desde un punto de origen del
conjunto, mayor y menor con huincha de distancia y
precisión de 10 cm.
Usada para probar en modelos de
cuan ficación no tradicionales.
Altura de copa
Medido desde el suelo y hasta el inicio del follaje.
Usada en modelos de cuan ficación no tradicionales.
3.3. Volteo, pesado y extracción de
muestras
18
Los árboles fueron volteados con un corte a ras de
suelo, direccionados hacia una malla sobre el suelo
de manera de no perder follaje en la faena. Una vez
volteado se midió la altura total, altura de comienzo
de copas y fue separado en los siguientes componentes.
• Tallo: Sección leñosa entre el suelo y la clara generación de copa en varias bifurcaciones.
• Ramas: Desde el tallo y hasta donde la sección
transversal de la rama alcanza los 3 cm.
• Ramillas y hojas: De las anteriores todo el resto.
Cada componente fue pesado en terreno (Figura
3.5) teniendo como instrumental de pesado:
• Trípode de madera.
• Eslingas de amarre de 2.000 kg.
• Sistema de mosquetones para resistencia de la
carga.
• Pesa con capacidad de 1:1000 Kg para tallo y
1:200 kg para ramas y ramillas y hojas.
A cada componente se le extrajeron probetas con
tres repe ciones, las cuales fueron pesadas en terreno, empaquetadas y rotuladas con códigos de
iden ficación de cada árbol para el laboratorio.
Figura 3.5 Detalle del proceso de volteo y pesado de cada componente Tallo, Ramas y Ramillas y Hojas.
FUNCIONES ALOMÉTRICAS PARA EL BOSQUE MEDITERRÁNEO CHILENO
La distribución de peso verde por componente es
muy similar entre el quillay y el peumo y entre el
litre y el espino (Cuadro 3.3, Figura 3.6).
Cuadro 3.3 Relación porcentual de peso y coeficiente de variación por componente.
Especie
quillay
peumo
litre
espino
Componente
Tallo
Ramas
RyH
Tallo
Ramas
Ry H
Tallo
Ramas
Ry H
Tallo
Ramas
Ry H
Peso. (%)
66%
13%
21%
68%
12%
20%
49%
26%
25%
51%
22%
27%
C.V (%)
16%
33%
39%
14%
38%
41%
27%
37%
35%
24%
29%
40%
Figura 3.6 Distribución de peso por cada componente.
19
4. Estimación de crecimientos
diamétricos del DAP
Los crecimientos diamétricos descritos en este capítulo deben usarse como nacionales, a pesar que se
determinaron por macrozona. Ello porque el principal obje vo de este proyecto implicó un muestreo
destruc vo, que condicionó los si os en los cuales
fue posible trabajar. No obstante, sería conveniente
en el futuro precisar esta información con un análisis previo de zonificación de la especie, de manera
que cubriera sa sfactoriamente su distribución. En
este trabajo no fue posible debido a que la principal
tarea fue la obtención de muestras para las funciones de carbono, y ello limitó fuertemente los si os
a aquellos lugares en que se permi ó la corta de árboles.
La rodela fue secada, lijada y escaneada. El tamaño muestral quedó conformado por un total de 313
muestras (Cuadro 4.1). El conteo y medición de los
anillos de crecimiento se extrajo de la imagen, usando el so ware ArMap (Figura 4.1). El procesamiento y medición de anillos se realizó según el método
propuesto por Cruz (2011).
Cuadro 4.1 Número de muestras analizadas
para determinación de crecimientos.
Especie
N
quillay
80
peumo
82
4.1 Metodología
litre
69
Para la determinación de crecimientos, a cada árbol le fue extraída una rodela a la altura del DAT.
espino
82
Figura 4.1 Medición de ancho de anillos en las 4 especies estudiadas.
21
Pablo Cruz
Alejandro Bascuñan
Juan Velozo
4.2
Resultados
La distribución de muestras en el territorio se agrupó en las siguientes categorías:
• Macrozona 1 Extremo norte de la V región (Norte).
• Macrozona 2 Región metropolitana y extremo sur de la V (Centro - Norte).
• Macrozona 3 Región VI (Centro - Sur).
• Macrozona 4 Regiones VII y VIII (Sur).
/0
4,=
4,<
4,<
4,;
4,;
4,:
4,:
4,9
4,9
4,8
4,8
4,7
!(5
!(6
!(7
!(8
4,6
4,5
4
4
22
4,=
4,=
54
/0
64
74
84
/0
4,7
!(5
!(6
!(7
!(8
4,6
4,5
4
4
94
/0
5,8
54
/0
64
74
84
/0
5,6
4,<
4,;
5
4,:
4,<
4,9
4,:
4,8
4,7
!(5
!(6
!(7
!(8
4,6
4,5
!(5
!(6
!(7
!(8
4,8
4,6
4
4,4
4
54
64
74
/0
4
54
64
74
84
/0
Figura 4.2 Incrementos de DAP o DAT la muestra por especie según localidades o macrozonas.
La variación del ancho de anillos es rela vamente
baja, considerando que los coeficientes de variación
están en torno al 20 %. El espino es una excepción y
muestra un aumento de la variabilidad hacia el sur.
Estos comportamientos pueden relacionarse con la
variación ambiental en que están creciendo las especies. Así sería posible decir que los ambientes en
que está creciendo espino en el sur, son poco homogéneos (a lo menos aquellos donde se tomaron las
muestras).
Cuadro 4.2 Incrementos medios del diámetro anuales promediados para cada macrozona.
Especie
Macrozona
quillay
peumo
litre
espino
IMAt
Promedio
(cm año-1)
Coeficiente
de variación
IMAt
Promedio
(cm año-1)
Coeficiente
de variación
IMAt
Promedio
(cm año-1)
Coeficiente
de variación
IMAt
Promedio
Coeficiente
de variación
1N
0,48
19%
0,37
31%
0,47
22%
0,74
23%
2 C-N
0,46
20%
0,39
17%
0,44
27%
0,73
26%
3 C-S
0,40
18%
0,41
27%
0,41
35%
0,69
38%
4S
0,61
26%
0,33
15%
0,42
19%
0,58
30%
Promedio
0,49
26%
0,38
27%
0,44
26%
0,70
30%
Crecimiento diametral
0.8
0.7
cm año -1
0.6
0.5
0.4
0.3
0.2
23
0.1
0
1N
2 C-N
3 C-S
4S
Macrozonas
quillay
peumo
litre
espino
Figura 4.3 Incrementos medios anuales del DAP por macrozona.
El quillay presenta un significa vo aumento en su
crecimiento en la zona sur. El peumo en cambio pareciera ofrecer mejores crecimientos en la zona central de su distribución. Tanto el litre como el espino
presentan una leve tendencia a aumentar su crecimiento hacia el norte. Del grupo, el espino ofrece
las mayores tasas de crecimiento anual.
5. Relación entre peso verde
y variables alométricas medidas
En la siguiente sección se contrasta gráficamente las
tendencias de las potenciales variables alométricas
y los correspondientes pesos verdes para cada especie y cada componente, con el obje vo de observar
las tendencias que muestran los datos. Los cuadros
por debajo de las gráficas corresponden al coeficiente de correlación de Pearson1.
25
5.1. Quillay
Peso Verde (Kg) - DAP (cm)
Peso Tallo
Peso Ramas
Peso R y H
Peso Total
0.953*
0.835
0.787
0.947
*Coeficiente de correlación de Pearson.
1.
El coeficiente de correlación de Pearson expresa la relación lineal de dos variables aleatorias cuan ta vas y es independiente de la escala de medida de cada una de las
variables. Es ú l para evaluar una primera aproximación de la relación, aunque puede ser débil si las relaciones fueran altas pero no lineales.
Pablo Cruz
Alejandro Bascuñan
Juan Velozo
Peso Verde (Kg) - Altura total (m)
6444
744
5944
5444
944
4
4
9
54
59
744
694
694
644
644
594
594
544
544
94
4
64
4
9
54
59
6944 6444
5944
5444
944
94
4
64
4
9
54
59
4
4
64
9
54
59
64
Peso Tallo
Peso Ramas
Peso R y H
Peso Total
0.715
0.623
0.521
0.699
Peso Verde (Kg) - Diámetro copa (m)
6444 5944
5444
944
4
4
9
54
" !# !#
!
!
#
59
#
#
#
#
#
#
#
#
#
26
Peso Tallo
Peso Ramas
Peso R y H
Peso Total
0.655
0.657
0.555
0.662
Peso Verde (Kg) - Altura de copa (m)
744
6444
694
744
5944
644
544
94
4
4
6
8
:
<
5944
594
5444
544
944
4
4
9
54
6444
644
5444
594
6944
694
94
4
4
6
8
:
<
944
4
4
6
8
:
Peso Tallo
Peso Ramas
Peso R y H
Peso Total
0.511
0.533
0.413
0.514
En el quillay, las variables que presentan mejores correlaciones con peso verde son el DAP, altura total y diámetro
<
de copa, a diferencia de la variable altura de copa que
presenta muy baja correlación.
FUNCIONES ALOMÉTRICAS PARA EL BOSQUE MEDITERRÁNEO CHILENO
5.2. Peumo
Peso Verde (Kg) - DAP (cm)
694
5444 694
5944 <44
644
644
5644
:44
594
594
=44
844
544
544
:44
94
644
4
4
94
4
744
4
4 54 64 74 84 94
4 54 64 74 84 94
Peso Tallo
Peso Ramas
Peso R y H
Peso Total
0.934
0.849
0.771
0.955
4 54 64 74 84 94
4 54 64 74 84 94
Peso Verde (Kg) - Altura Total (cm)
5444 694
694
5844
<44
644
644
:44
594
594
<44
844
544
544
:44
644
94
94
4
4
9
54
4
59
4
9
54
5444
844
644
4
59
5644
4
9
54
4
59
4
9
54
59
Peso Tallo
Peso Ramas
Peso R y H
Peso Total
0.645
0.473
0.375
0.6
Peso Verde (Kg) - Altura de Copa (cm)
5444
694
694
5844
<44
644
644
:44
594
594
<44
844
544
544
:44
644
94
94
4
4
6
8
:
<
4
4
6
8
:
<
5644
5444
844
4
4
6
8
:
<
644
4
4
6
8
:
Peso Tallo
Peso Ramas
Peso R y H
Peso Total
0.189
0.041
0.04
0.128
<
27
Pablo Cruz
Alejandro Bascuñan
Juan Velozo
Peso Verde (Kg) - Altura de Copa (cm)
5444
694
694
5844
<44
644
644
:44
594
594
<44
844
544
544
:44
644
94
94
4
4
6
8
:
4
<
4
6
8
:
5444
844
4
<
5644
4
6
8
:
644
4
4
<
6
8
:
Peso Tallo
Peso Ramas
Peso R y H
Peso Total
0.189
0.041
0.04
0.128
<
Peso Verde (Kg) - DAT (cm)
5444
694
694
<44
644
644
:44
594
594
844
544
544
644
94
94
4
4
28
59
74
4
4
89
59
74
4
4
89
59
74
5844
5644
5444
<44
:44
844
644
4
4
89
59
74
Peso Tallo
Peso Ramas
Peso R y H
Peso Total
0.95
0.84
0.79
0.95
En esta especie la variable DAP, DAT y diámetro de copa
presenta las mejores correlaciones, aunque la altura total
89
presenta una correlación interesante a diferencia de la
altura de copa que no existe relación entre variables.
5.3. Litre
Peso Verde (Kg) - DAT (cm)
694
644
594
544
94
4
4
54
64
74
584
564
544
<4
:4
84
64
4
4
54
64
74
584
564
544
<4
:4
84
64
4
944
844
744
644
4
54
64
74
544
4
4
54
64
Peso Tallo
Peso Ramas
Peso R y H
Peso Total
0.821
0.753
0.715
0.874
74
FUNCIONES ALOMÉTRICAS PARA EL BOSQUE MEDITERRÁNEO CHILENO
Para el litre la variable alométrica con mayor pernencia es el diámetro de tocón de cada vástago,
debido a la infac bilidad de medir claramente cepas
individuales (Figura 5.1), y a que la conducta de cada
vástago es tender a un crecimiento horizontal que
impide la medición de la altura predominante, la altura de copas y diámetros de copa.
Figura 5.1 Crecimiento caracterís co en litre. El po de crecimiento
de múl ples vástagos y de forma
rastrera impide la correcta medición de algunas variables alomértricas.
5.4. Espino
Peso Verde (Kg) - DAT (cm)
794
594
594
;44
:44
944
844
744
644
544
4
744
694
644
594
544
94
4
4
54
64
74
84
544
544
94
94
4
4
54
64
74
84
4
4
54
64
74
29
4
84
54
64
74
Peso Tallo
Peso Ramas
Peso R y H
Peso Total
0.910
0.852
0.814
0.922
84
Peso Verde (Kg) - Altura Total (m)
794 594
644
744
694
944
844
544
594
94
544
94
4
6
8
:
<
644
94
4
4
744
4
4
6
8
:
<
:44
594
544
644
;44
4
6
8
:
<
544
4
4
9
Peso Tallo
Peso Ramas
Peso R y H
Peso Total
0.545
0.549
0.282
0.520
54
Pablo Cruz
Alejandro Bascuñan
Juan Velozo
Peso Verde (Kg) - Altura Diámetro de Copa (m)
844
594
744
644
;94
:44
594
544
644
894
544
94
544
4
4
9
4
4
54
744
94
9
4
54
594
4
4
9
4
54
9
54
Peso Tallo
Peso Ramas
Peso R y H
Peso Total
0.772
0.831
0.763
0.838
Peso Verde (Kg) - Número de vástagos (V)
844
594
/0
744
644
544
644
94
544
4
4
30
6
8
:
594
:44
544
844
94
4
<
<44
4
6
8
:
644
4
4
<
6
8
:
<
4
4
6
8
:
Peso Tallo
Peso Ramas
Peso R y H
Peso Total
0.340
0.309
0.337
0.357
<
Peso Verde (Kg) - Altura de copa (m)
844
594
744
644
544
644
94
544
4
4
5
6
7
8
4
4
5
6
7
8
<44
594
:44
544
844
94
644
4
4
5
6
7
8
4
4
5
6
7
Peso Tallo
Peso Ramas
Peso R y H
Peso Total
0.092
0.084
0.065
0.077
8
FUNCIONES ALOMÉTRICAS PARA EL BOSQUE MEDITERRÁNEO CHILENO
El diámetro de tocón y el diámetro de copa muestran la
mayor correlación con el peso verde, a diferencia de lo
que ocurre con la altura de copas y número de vástago
donde no existe relación con los pesos. Además es notorio que algunas variables de alta relación, no enen
un comportamiento lineal (diámetro de tocón) y otras sí
(diámetro de copas). La altura en espino en par cular, se
hace insensible en árboles de mayor tamaño.
metro del suelo, DAM). Al establecer las correlaciones por
vástagos, fue necesario realizar las mediciones para cada
vástago en par cular. Además, el peso del tallo inferior se
asignó proporcionalmente a cada vástago en función del
diámetro relativo de cada uno.
En campo se observó que esta especie en par cular forma un sólo tallo en los primeros cen metros desde el
suelo, ya sea de individuos de semilla o de rebrote. Esta
sección alcanza regularmente 40 a 100 cm desde el suelo,
para luego ramificarse en pocos brotes secundarios (de
3 a 6 máximos). Sobre 160 cm ocurre una segunda ramificación en que los broten podrían cons tuir las ramas
verdaderas.
• De las variables ensayadas, el diámetro a la altura de
un metro, presenta la mayor relación, respecto de las
tradicionales.
• No es heterocedas ca hacia los grandes diámetros (Figura 5.2). Como se discu ó (capítulo 2), la mayoría de
los modelos de predicción de peso y volumen, deben
lidiar con puntos que se “abren” hacia los diámetros
mayores. Así, el dam puede considerarse una potente variable de es mación sobre todo en los rangos en
que los modelos pierden potencia por heterocedas cidad.
Por esta razón, se tomó una variable alométrica adicional,
correspondiente al diámetro de cada vástago, es decir,
después de la primera ramificación y que está regularmente a menos de un metro desde el suelo (diámetro a 1
Al modelar la relación de esta nueva variable con el peso
verde total se encontraron las siguientes bondades:
El modelo resultante se presenta en el capítulo 7.
31
"
/0
844
"!"#
744
>C4+=8
644
544
4
4
54
64
74
/0
Figura 5.2 Ajuste de variable auxiliar: Diámetro por vástago a la altura de 1 metro (DAM). Comportamiento del crecimiento
de vástagos.
6. Procesamiento y análisis
de resultados de laboratorio
6.1.2 Determinación de la densidad básica de la
madera
6.1. Procesamiento de muestras en
laboratorio
6.1.1 Determinación del contenido de humedad
Para la determinación del contenido de humedad
en base peso seco, se u lizó la norma chilena NCH
176/1-Of1984. Las muestras comprendieron rodelas
del tallo tomadas a la altura del tocón, a 1 y a 1,5 m
(de dos a cuatro cen metros de espesor), secciones
de ramas de primer orden con diámetros de 2 a 5
cm; y además, una muestra compuesta de hojas y
ramillas menores a 1,5 cm de diámetro. Para los tres
componentes del árbol se determinó el contenido
de humedad por el promedio de tres repe ciones
de cada componente. El peso fresco fue obtenido
en terreno con una balanza granataria digital portá l, con una sensibilidad de 0,01 g. La masa seca
de las muestras fue obtenida por medio de sacado
en una estufa de ro forzado a 105 °C; las muestras
se mantuvieron por 72 horas hasta alcanzar peso
constante, la masa fue medida en laboratorio en
una balanza con sensibilidad de 0.01 g. La fórmula
de cálculo para CH fue:
Para la determinación de la densidad básica, se u lizó como referencia la norma chilena NCH176/21986. Se usaron probetas saturadas, y procedimientos para determinar la densidad de probetas
irregulares. La es mación del volumen de las probetas se realizó mediante la inmersión de las probetas
y la medición del desplazamiento volumétrico por
diferencia de pesadas (se usó una balanza con sensibilidad de 0.01 g). Las densidades se determinaron
sobre probetas en forma de cuña, las que comprendieron desde el duramen hasta la albura. Las probetas se obtuvieron de rodelas del tallo, tomadas a
la altura del tocón, a 1 y a 1,5 m y, en secciones de
ramas de primer orden con diámetros de 2 a 5 cm
(se usó las mismas muestras del contenido de humedad). La fórmula de densidad básica fue:
Densidad Básica (t m3 -1) =
Peso Seco
volumen verde
[2]
6.1.3 Determinación del contenido de carbono
Contenido de Humedad (%) =
Peso verde – Peso Seco
Peso Seco
* 100
[1]
La determinación de carbono se realizó mediante el
método Dumas con un equipo TruSpec CN, Leco®.
Este equipo ocupa la técnica basada en la completa e instantánea oxidación de la muestra mediante
33
Pablo Cruz
Alejandro Bascuñan
Juan Velozo
combus ón completa con oxígeno puro a una temperatura de aproximadamente 950 ºC. El producto
de esta combus ón se presenta como CO2, H2O y
N2, los cuales son transportados por helio, y posteriormente separados en columnas específicas. Finalmente pasan de forma separada por detectores; el
carbono se mide mediante un sensor de absorción
IR. El detector emite una señal proporcional a la
concentración de carbono contenido en la muestra.
Se tomaron muestras representa vas de cada componente, para cada clase diamétrica y especie (13 a
23 repe ciones por componente analizado en cada
especie). De cada muestra se tomó una sub-muestra de 140 mg de masa seca (la que fue previamente molida homogéneamente). Esta sub-muestra se
ubica en una lámina de estaño que actúa como catalizador en el proceso de combus ón inicial. Para
determinar el contenido de carbono se u lizaron estándares trazables NIST.
34
No obstante que las funciones elaboradas en este
estudio usaron los contenidos de carbono es mados como se ha indicado, es importante señalar
que el valor u lizado en los Inventarios de Gases de
Efecto Invernadero ha sido considerado por defecto
como 0,5.
6.1.4 Análisis estadísticos
Se aplicó el test de normalidad Anderson-Darling, a
los datos de contenido de humedad, la razón peso
seco/peso fresco y el contenido de carbono. La existencia de diferencias se es mó con la prueba Kruskal Wallis, para probar la hipótesis nula de que las
muestras provienen de poblaciones con medianas
iguales. Luego para determinar las diferencias entre
los componentes se aplicó un test de comparación
múl ple con α = 0,05. En el caso de la densidad básica, la normalidad de los datos se evaluó mediante
el test de Anderson-Darling. Luego las diferencias
se probaron con la prueba no paramétrica U de
Mann-Whitney, que u liza rangos de datos muéstrales de dos poblaciones independientes. Si las muestras de densidad se distribuían de manera normal se
aplicó un análisis de varianza.
6.2. Resultados variables biométricas
6.2.1. Contenido de Humedad
La diferencia en el CH fue estadís camente significava entre componentes en todas las especies (Cuadro 6.1).
Cuadro 6.1 Pruebas de comparación múltiple entre componentes para el contenido de humedad.
Componente
quillay
peumo
litre
espino
p
n
p
n
p
n
p
n
Hojas y Ramillas
a
229
a
229
a
187
a
231
Ramas
b
234
b
234
b
188
b
305
Tallo
c
238
c
238
c
186
c
301
Total
701
701
561
837
Cuadro 6.2 Contenido de Humedad según tipo de muestra.
Componente
Contenido de Humedad (%)
Coeficiente de variación (%)
quillay
peumo
litre
espino
quillay
peumo
litre
espino
Hojas y Ramillas
106,6
107,6
99,9
93,2
21
27
26
41
Ramas
93,2
72,4
63,9
73,9
15
17
15
21
Tallo
87,9
69,3
58,8
65,2
17
17
17
19
Total
92,5
77,3
70,4
74,7
* Total: Promedio ponderado por el peso de cada componente
FUNCIONES ALOMÉTRICAS PARA EL BOSQUE MEDITERRÁNEO CHILENO
En general el contenido de humedad ene una variación estrecha en todas las especies para los componentes tallo y rama – coeficientes de variación
menores al 21% -. La componente hoja y ramilla
es más variable en todas las especies, debiéndose
posiblemente a que el material de medición es intrínsecamente heterogéneo (Cuadro 6.2). En efecto,
se trata de una mezcla de ramillas y hojas, éstas úlmas también de tamaño variable, siendo extremo
en el caso de espino, con un 41%. Es posible que la
variación en el espino se deba a que la muestra se
recogió en un periodo de 6 meses aproximadamente y el resultado se haya visto afectado por el ciclo
fenológico de la hoja.
Del grupo de especies, el quillay es la que con ene
más humedad y con la menor variación en toda la
muestra. Así también el espino y el litre arrojaron los
menores contenidos de humedad y en niveles similares, cons tuyéndose una segunda similitud entre
estas especies (Figura 6.1).
6.2.2. Densidad básica
El quillay y el espino, no enen una distribución
normal de la densidad básica (p < 0,01, de Anderson-Darling), mientras que el peumo y el litre si. Por
lo tanto, el análisis de la densidad por componente se hizo con la prueba U Mann-Whitney para las
Contenidos de humedad (%)
% de Humedad
120
100
80
60
40
20
0
Hojas y Ramillas
Ramas
Tallo
Total
35
componente
quillay
peumo
litre
espino
Figura 6.1 Contenido de humedad (%) según componente para las especies estudiadas.
Cuadro 6.3 Tamaño muestral para la densidad básica, por componente y total.
Componente
quillay
peumo
litre
espino
Ramas
182
82
85
203
Tallo
201
66
135
259
Total
383
148
220
462
Cuadro 6.4 Densidad básica según tipo de muestra.
Densidad básica (t m3 -1)
Coeficiente de variación (%)
Componente
quillay
peumo
litre
espino
quillay
peumo
litre
espino
Ramas
0,64
0,63
0,71
0,74
9,5
10,5
9
10,2
Tallo
0,63
0,63
0,74
0,74
7,0
9,2
5,4
7,8
Promedio
0,63
0,63
0,73
0,74
* Total: Promedio ponderado por el peso de cada componente.
Pablo Cruz
Alejandro Bascuñan
Juan Velozo
Densidad básica
0.75
(tm3 - 1)
0.70
0.65
0.60
0.55
0.50
Ramas
Tallo
promedio
componente
quillay
peumo
litre
espino
Figura 6.2 Densidad básica de las especies estudiadas por cada componente.
primeras y ANDEVA para las segundas (Cuadro 6.3 y
Cuadro 6.4).
36
Ni quillay ni el peumo registraron diferencias significa vas de la densidad básica entre componentes,
por tanto para estas especies es posible afirmar que
la densidad básica corresponde a 0,63 y 0,62 t m3 -1
respec vamente. El espino y el litre, en cambio resultaron con densidades estadís camente dis ntas
entre componente. Por ello, la forma correcta de
usar estos valores sería tomar los datos separados
por componente. Además estas dos especies son
consistentemente más pesadas que quillay y peumo.
Dentro de la muestra de cada componente, la variación es estrecha en todas las especies (Figura 6.2).
6.2.3. Relación peso seco peso fresco (Ps/Pv).
La variable Ps/Pv, permite valorar la proporción de
materia seca contenida por una unidad de masa
fresca dada. Está variable es rela vamente estable
para una especie y por componente, bajo condiciones de crecimiento homogéneas. El aumento
de este parámetro respecto a los niveles estándar
de la especie, indica condiciones de estrés hídrico
o algún factor asociado a éste. En nuestro estudio
por ejemplo, las especies que enen mayor densidad de madera presentan mayor relación Ps/Pv, lo
cual, probablemente esté asociado al mayor contenido de fibra (densidad básica) en estas maderas.
Sin embargo en el análisis de hojas y ramillas, para
espino el valor aparece muy alto respecto al valor
promedio de las otras especies. Lo anterior se debe
a que en el espino coexiste una porción de follaje
muerto con el ac vo; de esta forma al tomar una
muestra compuesta se observa un valor mayor a lo
normal. Finalmente cabe señalar que variable Ps/Pv
ene la ventaja de presentar una varianza más estable que la función inversa por la cual se determina el
volumen de agua, produciéndose un menor error de
es mación empírico.
Cuadro 6.5 Pruebas de comparación múltiple y tamaño muestreal para las cuatro especies.
Componente
quillay
peumo
litre
espino
p
n
p
n
p
n
p
n
Hojas y Ramillas
a
230
a
230
a
187
a
236
Ramas
b
235
b
223
b
188
b
305
Tallo
c
238
c
228
c
186
c
302
Total
703
681
561
843
FUNCIONES ALOMÉTRICAS PARA EL BOSQUE MEDITERRÁNEO CHILENO
Cuadro 6.6 Relación peso seco / peso verde por componente.
Relación peso seco/peso fresco (%)
Coeficiente de variación (%)
Componente
quillay
peumo
litre
espino
quillay
peumo
litre
espino
Hojas y Ramillas
49
49
51
74
11,5
12,5
13,0
23,8
Ramas
52
59
61
93
9,0
9,9
6,2
9,7
Tallo
54
59
63
77
8,2
7,1
6,8
8,0
Total
53
57
59
80
* Total: Promedio ponderado por el peso de cada componente.
Relación Ps/Pv (%)
Relación ps/pv (%)
60
45
30
15
0
Hojas y Ramillas
Ramas
Tallo
Total
componente
quillay
peumo
litre
espino
37
Figura 6.3 Relación Ps/Pv por componente para las cuatro especies.
El contenido de materia seca (DM, siglas del inglés
“dry ma er”) es la relación de peso seco y el peso
fresco de la planta. Se expresa en kg (peso seco) kg–1
(peso fresco).
La relación Ps/Pv es estadís camente diferente entre
componentes para todas las especies (Cuadro 6.5).
La relación Ps/Pv es considerablemente homogénea – bajos coeficientes de variación - en las cuatro
especies y en todos los componentes (Cuadro 6.6).
Hojas y ramillas es el componente con más variación
debido probablemente a las razones ya discu das y
especialmente en el espino, que en este componente vuelve a ofrecer las mayores variaciones.
Cuadro 6.7 Pruebas de comparación múltiples y tamaño muestral.
Componente
quillay
peumo
litre
espino
p
n
p
n
p
n
p
n
Hojas y Ramillas
a
13
a
14
a
12
a
13
Ramas
b
16
b
18
b
13
b
17
Tallo
b
19
b
22
b
17
c
23
48
54
42
53
Pablo Cruz
Alejandro Bascuñan
Juan Velozo
6.2.4. Contenido de carbono
El quillay, el peumo y el litre con enen los mismos
porcentajes de carbono en ramas y tallos. El espino,
en cambio arrojo una leve reducción del contenido
en las ramas, respecto del tallo, no obstante los análisis indicaron que es estadís camente significa va.
Así también, para todos los casos, la componente
hoja y ramillas ene estadís camente mayores contenidos de carbono respecto del resto (Cuadro 6.8).
Es destacable el nivel constante que presenta el
contenido de carbono en todos los componentes
en todas las especies, que oscila entre 48 y 52 %
(Figura 6.4).
Contenidos de carbono (%)
Contenido de carbono %
55
53
51
49
47
45
Hojas y Ramillas
Ramas
componente
quillay
peumo
Tallo
litre
Total
espino
38
Figura 6.4. Contenido de carbono en las cuatro especies, por componente. Los valores de la ordenada se han
restringido al rango 45 – 55 % para acentuar las diferencias entre especies, en atención a la similitud que han
presentado.
Cuadro 6.8 Contenido de carbono por componente y por especie.
Contenido de Carbono (%)
Coeficiente de variación (%)
Componente
quillay
peumo
litre
espino
quillay
peumo
litre
espino
Hojas y Ramillas
50
52
51
51
1,9
1,6
2,7
2,1
Ramas
48
49
48
48
1,4
1,3
1,2
1,8
Tallo
49
49
48
49
2,2
1,3
1,8
1,6
Total
49
50
49
49
* Total: Promedio ponderado por el peso de cada componente.
FUNCIONES ALOMÉTRICAS PARA EL BOSQUE MEDITERRÁNEO CHILENO
7. Elaboración de modelos
Se construyeron dos pos de modelos. Los primeros
son los más sencillos de dos variables – dependiente
y una independiente – y los segundos con el conjunto de variables independientes con el fin de comprobar si éstos mejoran sustan vamente a los primeros.
Con fines prác cos, si la calidad de un modelo de
dos variables es similar a uno de múl ples variables,
razonablemente debería recomendarse aquel más
sencillo, debido principalmente al beneficio de reducir la captura de información en terreno. En los
casos que los niveles de correlación no superen el
65% (R2), los modelos simplemente no se escribieron como una forma de impedir que fueran usados.
Dado que recurrentemente se debe aludir a estadís cas que respaldan los resultados, se han incorporado notas explica vas en cuadros para las explicaciones de la interpretación de dichos valores
estadís cos.
R2. En este capítulo se hace alusión frecuentemente a R2 o R cuadrado, y que puede tener
diferentes definiciones según la estadís ca que
se esté usando. Cuando se hacen pruebas de correlación entre variables, R2 se indica como coeficiente de correlación de Pearson, que es una
prueba para evaluar si dos variables están relacionadas, y puede aplicarse cuando las variables
cumplen con condiciones de normalidad. Si no
fuera lineal, la prueba de correlación que debiera
hacerse es la de Spearman, y el equivalente a R2
es “rho” de Spearman (ρ). En el caso que se está
evaluando la calidad de un modelo predictor, se
llamará coeficiente de determinación (Pardo &
Ruiz, 2005).
7.1 Modelos simples de dos variables
Se analizó la relación entre la variable dependiente
y cada una de las variables independientes, con el
obje vo de proponer modelos de los más sencillos
posibles. El primer análisis corresponde a pruebas
de normalidad para definir el po de análisis de correlación a evaluar. Los análisis estadís cos fueron
realizados en el paquete SPSS ver 17.
Pruebas de normalidad. Es necesario evaluar si
el comportamiento de las variables cumplen con
las tres pruebas de normalidad clásicas: a) si la
distribución de frecuencias es similar a una curva
de distribución normal. Básicamente es una comparación de ambas curvas, la de los datos y una
con distribución normal. Esta prueba es llamada
Kolmorogov-Smirnov (KMS) y define que si no
hay diferencias significa vas en la prueba – los
datos no hacen diferencia significa va con una
curva normal – el “p” valor de la prueba será mayor a 0,05. b) Homogeneidad de varianza u homocedas cidad, que básicamente indica que la
variación en los datos en un rango de la variable
es similar en todos los otros. En caso contrario
exis rá heterocedas cidad. La prueba recomendada para evaluar homocedas cidad es la Prue-
39
Pablo Cruz
Alejandro Bascuñan
Juan Velozo
ba de Levene. c) independencia de las unidades
muestrales que básicamente indica que resultados en una variable no dependen de otros anteriores. En este caso la prueba estadís ca es Durbin Watson y aplica de la misma forma de KMS.
Si las pruebas indican que no existe normalidad
en alguna variable, se debe recurrir a estadís ca
no paramétrica. Casi la mayoría de los modelos
en es maciones de biomasa arrojan los mejores
ajustes con modelos no paramétricos (Pardo &
Ruiz, 2005).
Se ha incorporado como variable única la relación
DAP2HT ya que es una combinación que refleja una
dimensión volumétrica y que así combinada puede
tratarse como una variable independiente más. En
el capítulo siguiente volverá a aparecer al combinar
cada una - DAP y HT – como variables por separado.
La mayoría de las variables independientes usadas
muestran alguna correlación sa sfactoria en algún
modelo (Rho˃0,7) (Cuadro 7.3). Con estos resultados, se evaluó la calidad de los modelos no lineales
tradicionales de manera de elegir aquellos con mejor coeficiente de determinación.
A pesar que el modelo que mejor ajusta para altura,
DAP y diámetro de copa es el cúbico, cae a valores
nega vos hacia el origen (la izquierda). Así, parece
recomendable elegir el modelo de potencia, sacrificando levemente precisión.
Nótese que el análisis de varianza (ANDEVA) indica que los coeficientes son significa vos, es decir,
aportan a la explicación del comportamiento de la
biomasa. El peor modelo es consistentemente el de
diámetro de copa (R2 = 0,65). Los coeficientes del
cuadro 7.4 fueron usados para la confección de los
modelos que se presentan a con nuación.
7.1.1 Quillay
40
Nótese que sólo altura y diámetro de copa enen
cercanía entre la media y la mediana, indicando una
distribución cercana a la normal (Cuadro 7.1).
Los análisis de correlación indican que la altura y el
DAP presenta comportamiento normal (KMS > 0,05),
y no así diámetro de copa ni la variable DAP2HT
(Cuadro 7.2). Por tanto las correlaciones se evaluaron tanto con Pearson como Spearman.
ANDEVA. Si bien cada modelo puede evaluarse con R2, la aplicación de una ANDEVA, permite evaluar el aporte de cada coeficiente en el
modelo, según se señala en el cuadro. La lógica de aplicación consiste en demostrar que H0
se cumple y que indicaría que ambas variables
no enen relación (nivel F˃0,05), y lo mismo
para cada uno de los coeficientes del modelo.
Cuadro 7.1 Estadística descriptiva para quillay, para una muestra de 80 árboles.
Estadístico
Altura
(m)
DAP
(cm)
D. Copa
(m)
DAP2 HT
Peso Seco
(Kg/arb.)
Media
6,9
16,7
3,2
3141,5
119,7
Mediana
6,0
15,0
2,9
1611,75
67,3
Mínimo
2
5
1
72
5,1
Rango
14
36
10
26824
1105,4
Máximo
16
41
11
26896
1110,4
Cuadro 7.2 Prueba de Kolmogorov-Smirnov para una muestra de 80 árboles de quillay.
Altura
(m)
DAP
(cm)
D. Copa
(m)
DAP2 HT
Desviación Típica
3,1
7,7
1,6
4436,5
Z de Kolmogorov -Smirnov
1,2
1,1
1,4
2,2
Sig. Asintótica (bilateral)
0,12
0,17
0,04
0,00
Estadístico
FUNCIONES ALOMÉTRICAS PARA EL BOSQUE MEDITERRÁNEO CHILENO
Cuadro 7.3 Coeficiente de determinación (R2) para los modelos tradicionales
posibles de probar para cada una de las variables. La variable dependiente
en todos los casos es Biomasa. En negrita los R2 mayores entre los modelos.
Variables
Altura
DAP
D. Copa
DAP2 HT
Lineal
0,57
0,73
0,57
0,93
Logarítmica
0,42
0,53
0,39
0,53
Inversa
0,27
0,34
0,21
0,13
Cuadrá ca
0,71
0,93
0,65
0,94
Cúbica
0,78
0,96
0,68
0,96
Compuesta
0,71
0,91
0,48
0,61
Potencia
0,74
0,95
0,51
0,93
S
0,65
0,86
0,39
0,56
Crecimiento
0,71
0,91
0,48
0,61
Exponencial
0,71
0,91
0,48
0,61
Logís ca
0,71
0,91
0,48
0,61
Modelos
Cuadro 7.4 Coeficiente de determinación (R2), valores de cada coeficiente
mediante análisis de varianza y valor de los coeficientes para construir los
modelos seleccionados.
Variables
Modelos
Tipo de modelo
2
R corregido
Altura
DAP
D. Copa
DAP2 HT
Potencia
Potencia
Cuadrá co
Cúbico
0,73
0,96
0,65
0,96
0,0003
0,000
0,000
0,628
0,00
0,000
0,000
0,00
ANDEVA
b0
(Constante)
b1
0.00
b2
b3
Coeficientes para los modelos
b0 (Constante)
1,494
0,150
39,404
3,21
b1
2,066
2,251
-11,357
0,048
9,198
-2,5E-6
b2
b3
8,3E-11
41
Pablo Cruz
Alejandro Bascuñan
Juan Velozo
M O D E L O S Q U IL L A Y
MODELOS
M O D E LQUILLAY
O S Q U IL L A Y
MAltura
ODELO
S Q U IL L A Y
Total
MODELOS Q
U IL L A Y
5644
""
#
<44
844
4
4
9
54
59
64
Parámetros de cada modelo
Variables y unidades
N muestral = 80
Rango HT = 2 – 16 m
= 0,74
R2
B = Biomasa (kg)
HT = Altura total (m)
42
DAP
5644
"
#
<44
844
4
4
54
64 74
Parámetros de cada modelo
N muestral = 80
Rango DAP = 5 - 41 cm
= 0,95
R2
84
94
Variables y unidades
DAP = Diámetro a la
altura del pecho (cm)
FUNCIONES ALOMÉTRICAS PARA EL BOSQUE MEDITERRÁNEO CHILENO
Diámetro de Copa
[5]
"
#
5644
<44
844
4
4
6
8
:
<
54
56
#!
Parámetros de cada modelo
N muestral
Rango DC
R2
Variables y unidades
= 80
= 1-10,5 (m)
= 0,65
DC = Diámetro de copa(m)
43
DAP2 * HT
[6]
5644 "
#
<44
844
4
4
54444
-
Parámetros de cada modelo
N muestral = 80
Rango DAP = 5 – 41cm
Rango HT = 2- 16 m
R2
= 0,96
64444
74444
Variables y unidades
=
Relación
DAP2HT
diámetro al cuadrado por
Altura Total
Pablo Cruz
Alejandro Bascuñan
Juan Velozo
7.1.2 Peumo
Sólo altura y diámetro de copa tienen cercanía entre
la media y la mediana, indicando una distribución
cercana a la normal. Efec vamente, los análisis de
correlación indican que sólo la altura y el DAP ene
un comportamiento normal (KMS ˃ 0,05) (Cuadro
7.5 y Cuadro 7.6).
La mayoría de las variables independientes usadas muestran correlación bastante sa sfactoria
(Rho˃0,7) (Cuadro 7.7) en algún modelo. Con estos
resultados, se evaluó la calidad de los modelos no
lineales tradicionales de manera de elegir aquellos
con mejor coeficiente de determinación.
La variable que ofrece los menores coeficientes es la
altura. En consecuencia, no será construido.
Cuadro 7.5 Estadística descriptiva para peumo, para una muestra de 82 árboles.
Estadístico
44
Altura
(m)
DAP
(cm)
DAT
(cm)
D. Copa
(m)
DAP2HT
Peso Seco
(Kg)
Media
8,2
14,3
16,3
3,5
2.397
99,4
Mediana
8,0
13
15.0
3,3
1.362
55,3
Mínimo
3
5
6
1
75
4
Rango
10
32
36
8,3
16.353
800
Máximo
13
37
42
9,3
16.428
805
Cuadro 7.6 Prueba de Kolmogorov-Smirnov para una muestra de 82 árboles de peumo.
Estadístico
Altura
(m)
DAT
(cm)
D. Copa
(m)
DAP
(cm)
DAP2 HT
Peso Seco
(Kg)
Desviación Típica
2,2
6,6
1,8
7,4
3373,8
141,6
Z de Kolmogorov -Smirnov
0,98
1,61
1,5
1,6
2,4
2,6
Sig. Asintótica (bilateral)
0,29
0,01
0,02
0,11
0,00
0,00
FUNCIONES ALOMÉTRICAS PARA EL BOSQUE MEDITERRÁNEO CHILENO
Cuadro 7.7 Coeficiente de determinación (R2) para los modelos tradicionales posibles de probar
para cada una de las variables. La variable dependiente en todos los casos es Biomasa. En negrita
los R2 mayores entre los modelos.
Variables
Altura
DAP
DAT
D. Copa
DAP2HT
Lineal
0,36
0,82
0,83
0,67
0,93
Logarítmica
0,28
0,61
0,62
0,43
0,58
Inversa
0,19
0,36
0,37
0,20
0,11
Cuadrática
0,53
0,91
0,92
0,80
0,93
Cúbica
0,60
0,91
0,92
0,80
0,93
Compuesta
0,62
0,88
0,88
0,67
0,67
Potencia
0,60
0,95
0,94
0,58
0,96
S
0,54
0,84
0,86
0,37
0,52
Crecimiento
0,63
0,88
0,86
0,67
0,67
Exponencial
0,63
0,88
0,86
0,67
0,67
Logística
0,63
0,88
0,86
0,67
0,67
Modelos
Cuadro 7.8 Coeficiente de determinación (R2), valores de cada coeficientes mediante análisis
de varianza y valores de cada coeficiente para construir los modelos seleccionados.
45
Variables
DAP
DAT
D. Copa
DAP2HT
Potencia
Potencia
Cuadrá co
Potencia
0,95
0,94
0.80
0,96
(Constante)
0,0
0,0
0,03
0,00
b1
0,0
0,0
0,008
0,00
Modelos
Tipo de modelo
2
R corregido
ANDEVA
b0
0,00
b2
Coeficientes
para los modelos
b0
(Constante)
0,136
0.096
65,474
0,076
b1
2,352
2.366
-39,993
0,92
b2
11,153
Para todos los modelos propuestos las constantes y cada uno de los coeficientes son significa vos
para predecir la biomasa (Cuadro 7.8).
Pablo Cruz
Alejandro Bascuñan
Juan Velozo
MODELOS
PEUMO
MODELO
S PEUMO
MODELO
MODELO
S PEUMO
S PEUMO
DAP
[7]
5644
"
#
=44
:44
744
4
4
9
54
59
64
69
74
79
84
Parámetros de cada modelo
N muestral
=
82
Rango DAP =
5 – 37 cm
R2
0,95
=
Variables y unidades
DAP = diámetro a la altura
del pecho (cm)
46
DAT
5644
[8]
"
##
=44
:44
744
4
4
54
Parámetros de cada modelo
N muestral
=
82
Rango DAT
=
6 – 42 cm
R2
=
0,94
64 74
84
94
Variables y unidades
DAT = diámetro a la altura
del tocón (cm)
FUNCIONES ALOMÉTRICAS PARA EL BOSQUE MEDITERRÁNEO CHILENO
Diámetro de Copa
5644
=44
[9]
"
#
:44
744
4
4
6
8
:
#!
<
54
Parámetros de cada modelo
Variables y unidades
N muestral = 82
DC = Diámetro de Copa(m)
Rango DC =
1 – 9,3 m
R2
0,80
=
47
DAP2*HT
5644
[10]
"
#
=44
:44
744
4
4
9444
Parámetros de cada modelo
N muestral = 82
Rango DAP = 5 – 37 cm
Rango HT = 3 – 13 m
R2
=
0,96
54444
6
59444
64444
Variables y unidades
DAP2HT = Relación diámetro
al cuadrado por altura total
Pablo Cruz
Alejandro Bascuñan
Juan Velozo
7.1.3 Litre
Esta especie sólo pudo ser descrita con la variable
DAT, como ya se indicó, debido a que su forma de
crecimiento rastrero impide medir correctamente
alturas y DAP, y la agrupación de sus vástagos dificulta la determinación de una cepa de otra. Nótese
que el DAT presenta cercanía entre la media y la mediana, indicando una distribución cercana a la normal (Cuadro 7.9). Los análisis de correlación indican
que el DAT ene un comportamiento normal (KMS ˃
0,05) (Cuadro 7.10).
La variable DAT muestra una correlación sa sfactoria, evaluándose la calidad de los modelos no lineales tradicionales de manera de elegir el que presente
mayor coeficiente de determinación (Cuadro 7.11).
Para el modelo propuesto las constantes y los coeficientes son significa vos para predecir la biomasa
(Cuadro 7.12).
Cuadro 7.9 Estadística descriptiva para litre, para una muestra de 80 árboles.
48
Estadístico
DAT
(cm)
Peso seco
(Kg/arb.)
Media
12,2
47,0
Mediana
12,0
34,9
Mínimo
4
3,1
Rango
21
225,1
Máximo
23
228,2
Cuadro 7.10 Prueba de Kolmogorov-Smirnov para una muestra de 80 árboles de litre.
Estadístico
DAT
(cm)
Peso Seco
(Kg/arb.)
Desviación Típica
4,9
45,62
Z de Kolmogorov -Smirnov
1,037
1,500
Sig. Asintótica (bilateral)
0,233
0,022
FUNCIONES ALOMÉTRICAS PARA EL BOSQUE MEDITERRÁNEO CHILENO
Cuadro 7.11 Coeficiente de determinación (R2) para los modelos tradicionales posibles de probar para cada una de
las variables. La variable dependiente en todos los casos es Biomasa. En negrita el R2 mayor entre los modelos.
Variable
Modelos
DAT
Lineal
0,68
Logarítmica
0,59
Inversa
0,46
Cuadrática
0,73
Cúbica
0,73
Compuesta
0,84
Potencia
0,87
S
0,82
Crecimiento
0,84
Exponencial
0,84
Logística
0,84
Cuadro 7.12 Coeficiente de determinación (R2), valores de cada coeficientes mediante análisis de varianza
y valores de cada coeficiente para construir el modelo seleccionado.
Variable
Modelos
DAT
Tipo de modelo
Potencia
R2 corregido
0,87
ANDEVA
b0
(Constante)
0,000
b1
0,000
Coeficientes
para los modelos
b0
(Constante)
0.196
b1
2,091
49
Pablo Cruz
Alejandro Bascuñan
Juan Velozo
M O DELO LITRE
M O DELO
MODELO
LITRE LITRE
DAT
744
"
#
644
544
4
4
9
54 59
64
Parámetros de cada modelo
69
Variables y unidades
DAT = diámetro a la altura
del tocón (cm)
N muestral = 80
Rango DAT = 4 - 23 cm
R2
= 0,87
7.1.4 Espino
50
Nótese que sólo altura y diámetro de copa enen
cercanía entre la media y la mediana, indicando una
distribución cercana a la normal (Cuadro 7.13). Los
análisis de correlación indican que el DAT y diámetro
de copa tiene comportamiento normal (KMS>0,05)
(Cuadro 7.14).
La mayoría de las variables independientes usadas
muestran correlación sa sfactoria en algún modelo (Cuadro 7.15). Con estos resultados, se evaluó la
calidad de los modelos no lineales tradicionales de
manera de elegir aquellos con mejor coeficiente de
determinación.
Cuadro 7.13 Estadística descriptiva para espino, para una muestra de 86 árboles.
Altura
(m)
DAT
(cm)
D. Copa
(m)
DAT2 HT
Peso Seco
(Kg/arb.)
Media
3,7
15,1
3,4
1134
65.9
Mediana
3,5
14,0
3,3
662
38.1
Mínimo
1,5
5
10
38
3.8
Rango
5,5
27
7,8
5009
360
Máximo
7,0
32
8,8
5046
363.8
Estadístico
Cuadro 7.14 Prueba de Kolmogorov-Smirnov para una muestra de 86 árboles de espino.
Altura
(m)
DAT
(cm)
D. Copa
(m)
DAT2 HT
Peso Seco
(Kg/arb.)
Media
3,7
15,1
3,4
1134
65,9
Desviación Típica
1,1
1,1
1,6
1205
72,7
Z de Kolmogorov -Smirnov
1,6
1,3
1,2
1,9
1,8
Sig. Asintótica (bilateral)
0,01
0,05
0,13
0,00
0,00
Estadístico
FUNCIONES ALOMÉTRICAS PARA EL BOSQUE MEDITERRÁNEO CHILENO
Cuadro 7.15 Coeficiente de determinación (R2) para los modelos tradicionales posibles de probar para cada una de
las variables. La variable dependiente en todos los casos es Biomasa. En negrita los R2 mayores entre los modelos.
Variables
Altura
DAT
D.Copa
DAT2HT
Lineal
0,40
0,82
0,77
0,89
Logarítmica
0,37
0,68
0,59
0,68
Inversa
0,28
0,50
0,34
0,21
Cuadrática
0,40
0,89
0,84
0,89
Cúbica
0,40
0,89
0,84
0,90
Compuesta
0,49
0,88
0,77
0,71
Potencia
0,53
0,92
0,80
0,92
S
0,50
0,84
0,69
0,53
Crecimiento
0,49
0,88
0,77
0,71
Exponencial
0,49
0,88
0,77
0,71
Logística
0,49
0,88
0,77
0,71
Modelos
Cuadro 7.16 Coeficiente de determinación (R2) y valor de los coeficientes para construir los modelos seleccionados.
Variables
Modelos
Altura
DAT
D. Copa
DAT2HT
Potencia
Potencia
Cuadrá ca
Potencia
0,53
0,92
0,84
0,92
(Constante)
0,00
0,00
0,76
0,000
b1
0,00
0,00
0,62
0,000
Tipo de modelo
R2 corregido
ANDEVA
b0
0,000
b2
b3
Coeficientes
para los modelos
b0
(Constante)
0,967
0,071
4,408
0,068
b1
2,90
2,395
-3,838
0,972
b2
5,258
b3
La variable que ofrece el menor coeficiente de determinación (R2<0,65) es la altura, de manera que el
modelo no fue aceptado.
El ANDEVA indica que en todos los modelos escogidos, los coeficientes aportan a la explicación de la
biomasa. Ello se verifica para todas las variables a
excepción de diámetro de copa (Cuadro 7.16).
51
Pablo Cruz
Alejandro Bascuñan
Juan Velozo
MODELOS
MODELOS
ESPINO
ESPIN O
MODELOS
ESPINO
MODELOS
ESPINO
DAT
:44
"
#
844
644
4
4
9
54
59
64
69
74
79
Parámetros de cada modelo
N muestral
Variables y unidades
= 86
DAT = diámetro a la
Rango DAT = 5 - 32 cm
R2
altura del tocón (cm)
= 0,92
Diámetro de Copa
52
:44
[13]
"
#
844
644
4
4
6
8
:
#!
Parámetros de cada modelo
N muestral = 86
Rango DC = 1-8,75 (m)
R2
= 0,84
<
54
Variables y unidades
DC
=
diámetro
copa (m)
de
FUNCIONES ALOMÉTRICAS PARA EL BOSQUE MEDITERRÁNEO CHILENO
DAT2*HT
[14]
:44
"
#
844
644
4
4
6444
Parámetros de cada modelo
N muestral =
86
Rango DAT =
5 – 32 cm
Rango HT
= 1,5 – 7 m
R2
= 0,92
-
8444
:444
Variables y unidades
DAT2HT = Relación diámetro al
cuadrado por altura total
53
Pablo Cruz
Alejandro Bascuñan
Juan Velozo
7.1.5 Espino – Vástago
La variable DAM presenta una cercanía entre la media y la mediana, indicando una distribución cercana
a la normal. Los análisis de correlación indican que
la variable DAM ene un comportamiento normal
(KMS ˃ 0,05) (Cuadro 7.18).
La nueva variable DAM ofrece un interesante coeficiente de determinación (Cuadro 7.19) (R2 = 0,94).
Para el modelo propuesto las constantes y los coeficientes son significa vos para predecir la biomasa
(Cuadro 7.20).
Cuadro 7.17 Estadística descriptiva para vástago en espino, para una muestra de 97 vástagos.
Estadístico
DAM
(cm)
Peso Seco
(Kg/arb.)
Media
12,1
46,2
Mediana
11,0
35.3
Mínimo
5
4,1
Rango
20
192,8
Máximo
25
196,9
Cuadro 7.18 Prueba de Kolmogorov-Smirnov para una muestra de 97 vástagos de espino.
Estadístico
54
DAM
(cm)
Peso Seco
(Kg/arb.)
Desviación Típica
5,2
41,3
Z de Kolmogorov -Smirnov
1,07
1,52
Sig. Asintótica (bilateral)
0,19
0,02
Cuadro 7.19 Coeficiente de determinación (R2) para los modelos tradicionales posibles de probar para cada una de
las variables. La variable dependiente en todos los casos es Biomasa. En negrita el R2 mayor entre los modelos.
Variable
Modelos
DAM
Lineal
0,89
Logarítmica
0,78
Inversa
0,62
Cuadrática
0,93
Cúbica
0,93
Compuesta
0,88
Potencia
0,94
S
0,90
Crecimiento
0,88
Exponencial
0,88
Logística
0,88
FUNCIONES ALOMÉTRICAS PARA EL BOSQUE MEDITERRÁNEO CHILENO
Cuadro 7.20 Coeficiente de determinación (R2), valores de cada coeficiente mediante análisis de varianza y valor
de los coeficientes para construir los modelos seleccionados.
Variables
Modelos
DAM
Tipo de modelo
Potencia
R2 corregido
0,94
ANDEVA
b0
(Constante)
0,00
b1
0,00
Coeficientes
para los modelos
b0
(Constante)
0,177
b1
2,142
55
Pablo Cruz
Alejandro Bascuñan
Juan Velozo
MODELO VÁSTAGO
MODELO VÁSTAGO
MODELO VÁSTAGO
MODELO
VÁSTAGO
MODELO
VÁSTAGO
DAM
844
"
#
744
644
544
4
4
9
54
59
64
69
74
Parámetros de cada modelo
N muestral
= 97
Rango DAM = 5 – 25 cm
DAM
R2
del metro (cm)
= 0,94
Es destacable la baja heterocedas cidad que resulta
de usar DAM como variable independiente (nótese
en la gráfica la cercanía de los puntos de mayores
diámetros a la línea de tendencia del modelo).
56
Cada vástago de la muestra con ene la proporción
de biomasa de su cepa, de manera que para usar
el DAM en inventario, bastaría medir todos los vástagos de la correspondiente parcela, independiente
de las cepas. Considerando su ajuste homocedas co, el modelo resultante es maría mejor en diámetros mayores, condición que no enen los otros modelos construidos y, posiblemente, sea más simple
de medir en campo que los DAT.
7.2
Variables y unidades
B = Biomasa (kg)
Modelos complejos de múltiples
variables
En este capítulo se analizan modelo complejos para
las especies del estudio, cuyo fin es mejorar las precisiones de es mación. La creación de funciones de
predicción de volumen es una materia de larga data
en el sector forestal – el volumen es la base de la
predicción de biomasa -, y ha tenido como consecuencia el descubrimiento de modelos que normalmente son los que mejor ajuste entregan. Así, para
realizar un análisis que aproveche de mejor forma la
información recogida en terreno, se ha hecho una
= Diámetro a la altura
revisión de aquellos modelos que han sido indicados como los de mejor ajuste para un gran número
de especies (Cuadro 7.21). (Cano, Corral, Nájera, &
Abel, 2008; Diéguez et al., 2003; Drake, Emanuelli, &
Acuña, 2003; Prodan et al., 1997).
Estos modelos representan las formas matemá cas
generales que se han publicado, como aquellos que
frecuentemente ajustan mejor. El uso de DAP y HT,
se debe a que la relación DAP2HT, genera una base
cúbica a par r de la cual los demás términos de la
ecuación ajustan sobre la base de dicho cubo. Por
ello, se recomienda agregarla como una variable
independiente, como en el capítulo anterior. Nótese que al ser probada en un modelo lineal, queda
expresada semejante al modelo 1 del cuadro 7.21,
pero con una constante b0. Así, sin la constante, el
modelo toma la forma “factor de forma constante”,
que es un modelo clásico que se determina y se valora en la forma que está escrito, vale decir, la relación de, HT y sólo un coeficiente b1.
No se consideró finalmente la creación de modelos
lineales donde se agregan y relacionan variables
como altura de copa y número de vástagos debido
a que presentan muy bajas relaciones. Los modelos
con diámetro de copa se agregan en un capítulo especial, dado su valor para estudios de teledetección.
FUNCIONES ALOMÉTRICAS PARA EL BOSQUE MEDITERRÁNEO CHILENO
Cuadro 7.21 Modelos más importantes para estimar biomasa y volumen. Los coeficientes en las funciones corresponden a b0 (la constante), b1, b2 y b3. Las variables para los modelos son B: biomasa en kg, DAP: diámetro a la altura
del pecho en cm y h: altura en m.
Tipo de modelo
1. Factor de forma constante
Referencias
Modelo
Modelo de Spurr
B = b1 *DAP2 * HT
B = b0 + b1 * HT + b2 *DAP2+ b3 *DAP
2. Variable combinada generalizada completa
3. Alométrica o logarítmica sin intercepto
Schumacher-Hall
B = b1 * DAPb2 * HTb3
Newnham
B = b0 + b1 * DAPb2 * HTb 3
4. Logarítmica con intercepto
5. Variables transformadas
B=
Honer
DAP2
b
b0 + 1
HT
7.2.1. Quillay
Los ajustes de los modelos probados entregan un
coeficiente de determinación cercano a 0,95 (Cuadro 7.22).
cada una de las variables. La variable dependiente
en todos los casos es Biomasa. En negrita los R2 mayores entre los modelos.
Cuadro 7.22 Coeficiente de determinación (R2) para
los modelos tradicionales posibles de probar para
Cuadro 7.22 Coeficiente de determinación (R2) para los modelos tradicionales posibles de probar para Quillay. La
variable dependiente en todos los casos es Biomasa. En negrita los R2 mayores entre los modelos.
Modelo
R2 Corregida
Expresión
Factor de forma constante
0,93
B = 0,036 * DAPT2 * HT
[16
Variable combinada generalizada completa
0,94
B = 49,427 - 10,41HT+0,115* DAP2 + 0,033 *DAPT2* * HT
[17
Alométrica o logarítmica sin intercepto
0,94
B = 0,014 * DAP2,870 * HT0,188
[18
Logarítmica con intercepto
0,95
B = 29,129 + 0,002 * DAP3.528 * HT0.034
[19
Variables transformadas
0.93
B=
DAP2
0,25 + 24,594
HT
[20
57
Pablo Cruz
Alejandro Bascuñan
Juan Velozo
7.2.2 Peumo
7.2.3 Espino
Los ajustes de los modelos probados son similares
en su desempeño, ya que todos entregan un coeficiente de determinación en torno a 0,93 (Cuadro
7.23)
Los ajustes de los modelos probados entregan un
coeficiente de determinación cercano a 0,92 (Cuadro 7.24).
Cuadro 7.23 Coeficiente de determinación (R2) para los modelos tradicionales para Peumo. La variable dependiente
en todos los casos es Biomasa. En negrita los R2 mayores entre los modelos.
Modelo
R2 Corregida
Expresión
Factor de forma constante
0,93
B = 0,042 * “DAP2 HT
[21]
Variable combinada generalizada completa
0,93
B = 30,271 - 3,129 * HT - 0,158* DAP2 + 0,056 *DAP2*HT
[22]
Alométrica o logarítmica sin intercepto
0,93
B = 0,024 * DAP1,939 * HT1,316
[23]
Logarítmica con intercepto
0,94
“B = 9,323 + 0,012 * DAP2,032 * HT1,455
[24]
Variables transformadas
0,93
B=
DAP2
-0,678 + 31,918
HT
[25]
58
Cuadro 7.24 Coeficiente de determinación (R2) para los modelos tradicionales posibles pra Espino. La variable dependiente en todos los casos es Biomasa. En negrita los R2 mayores entre los modelos.
Modelo
R2 Corregida
Expresión
Factor de forma constante
0,899
B = 0,058 *DAT * HT
[26]
Variable combinada generalizada completa
0,915
B = 9,748 - 4,886*HT+0,092* DAT2 + 0,043 *DAT2*HT
[27]
Alométrica o logarítmica sin intercepto
0,917
B = 0,048 * DAT2,272 * HT0,565
[28]
Logarítmica con intercepto
0918
B = 3,589 + 0,033 * DAT2,372 * HT0,586
[29]
Variables transformadas
0,912
B=
DAP2
1,341 + 10,78
HT
[30]
FUNCIONES ALOMÉTRICAS PARA EL BOSQUE MEDITERRÁNEO CHILENO
8. Modelos para predicción
de carbono
En este capítulo se extractan, para cada especie, los
mejores modelos conseguidos con los análisis realizados. Con ellos se construyen las funciones alómetricas de carbono finales, ponderando los modelos
por el valor constante de carbono obtenido. Los criterios de selección han sido su potencia predictora y
su sencillez. Para cada especie se han escogido modelos de dos variables y de más variables en el caso
que estos úl mos sean significa vamente mejores
que los primeros (Cuadro 8.1).
Para el quillay, el peumo y el espino se han dejado
dos modelos, uno de dos y otro de tres variables.
Para el espino se presenta el modelo con la variable
DAM, que se propone en este estudio como nueva
variable Alométrica. En litre sólo se contó con la variable DAT.
59
Cuadro 8.1 Modelos propuestos para estimar contenido de carbono (C) en especies del Bosque
Mediterráneo Chileno.
Especie
Tipo de modelo - R2
Modelo
Quillay
Potencia - 0,95
Logarítmica con intercepto – 0,95
C = 0,073598 * DAP2,251
C = 14,27321+0,00098 * DAP3,528 * HT0,034
Potencia - 0,95
C = 0,067456 * DAP2,352
Logarítmica con intercepto – 0,93
C = 4,624 + 0,00595 * DAP2.032 * HT1.455
Litre
Potencia - 0,87
C = 0,09638 * DAT2,0912
Espino
Potencia - 0,92
Logarítmica con intercepto – 0,92
C = 0,034954 * DAT2,3949
C = 1,76938 + 0,01627 * DAT2,372 * HT0,586
Potencia - 0,94
C = 0,08711 * DAM2,1416
Peumo
Espino
vástago
9. Modelos de biomasa y carbono
útiles para teledetección
Si bien el diámetro de copa (DC) resultó ser un predictor de menor potencia que las otras variables
para la es mación de biomasa y carbono, se agregan a este informe las funciones con DC considerando que pueden tener u lidad como aproximaciones
cuando se es man ByC sin datos de campo y sólo
con imágenes.
No fue posible determinar una función para litre, debido a su forma de crecimiento, que impidió la medición de diámetros de copa. Los pobres resultados
con litre podrían mejorarse si se inves garan modelo de medición de masa dentro del bosque y no por
árboles individuales, como pude ser Hart-Becking,
entre otros.
61
Cuadro 9.1 Modelos propuestos para estimar contenido de biomasa y carbono aplicando métodos de teledetección.
DC corresponde a Diámetro de copa, medido en metros.
Especie
Quillay
Peumo
Espino
Modelo
R2
B = 39,404 – 11,357 * DC + 9,198 * DC 2
C = 32,475 – 19,836 * DC + 5,532 * DC 2
0,65
B = 39,404 – 11,357 * DC + 9,198 * DC 2
C = 19,308 – 5,565 * DC + 4,507 * DC 2
0,80
B = 4,408 – 3,839 * DC + 5,258 * DC 2
C = 2,173 – 1,893 * DC + 2,592 * DC 2
0,84
10. Referencias bibliográficas
Cano, M., Corral, E., Nájera, L., & Abel, J. (2008). Ajuste
de ecuaciones de volumen en Pinus durangensis en
la comunidad las Flechas, Durango. Revista de las
Sedes Regionales. h p://www.redalyc.org/ar culo.
oa?id=66615066012
Chave, J., Andalo, C., & Brown, S. (2005). Tree allometry
and improved es ma on of carbon stocks and balance in tropical forests. Oecologia, 145(1), 87-99.
CONAF. (2013). Compendio de funciones alométricas para
la estimación de la biomasa de especies forestales presentes en Chile. San ago, Chile, Chile: Corporación Nacional Forestal.
Cruz, P. (2011). Aplicaciones de ArcInfo para inves gación
en Silvicultura: Análisis fustal y Área Potencial Aprovechable. Encuentro de Usuarios Esri Chile 2011. h p://
www.usuarios.cl/agenda.html
Cruz, P. (2013). Análisis del estado de conservación de
bosques. Caso del Quercus pyrenaica en la comarca del Bierzo. h p://www.academia.edu/4638887/
Analisis_del_estado_de_conservacion_de_bosques._
Caso_del_Quercus_pyrenaica_en_la_comarca_del_
Bierzo#
Diéguez, U., Barrio, M., Castedo, F., A, R., Álvarez, M.,
Álvarez, J., & Rojo, A. (2003). Dendrometría. Madrid,
España: Mundiprensa.
Drake, F., Emanuelli, P., & Acuña, E. (2003). Compendio de
funciones dendrométricas del Bosque Nativo. San ago, Chile: GTZ.
Eichhorn, F. (1904). Beziehungen zwischen Bestandshöhe
und Bestandsmasse. Allgemeine Forst- und Jagdzeitung(80), 45-49.
Gayoso, J. (2013a). Funciones alométricas para la determinación de existencias de carbono forestal para la
especie Araucaria araucana (Molina) K. Koch (ARAUCARIA). San ago, Chile.: Unidad de cambio climá co,
Corporación Nacional Forestal-Programa Bosques
PROCARBONO, Universidad Austral de Chile.49 p.
Gayoso, J. (2013b). Funciones alométricas para la determinación de existencias de carbono forestal para la
especie Nothofagus oblicua (Mirb.) Oerst. (ROBLE).
San ago, Chile.: Unidad de cambio climá co, Corporación Nacional Forestal-Programa Bosques PROCARBONO, Universidad Austral de Chile. 41 p.
Gayoso, J. (2013c). Funciones alométricas para la determinación de existencias de carbono forestal para la
especie Nothofagus pumilio (peopp. et Endl) Krasser
(LENGA). San ago, Chile: Unidad de cambio climá co, Corporación Nacional Forestal-Programa Bosques
PROCARBONO, Universidad Austral de Chile.32 p.
Gayoso, J., & Guerra, J. (2005). Contenido de carbono en
la biomasa aérea de bosques na vos de Chile. Bosque,
26(2), 6.
Pardo, A., & Ruiz, M. (2005). Análisis de datos con spss 13.
Madrid: McGraw-Hill.
Picard, N., Saint-André, L., & Henry, M. (2012). Manual de
construcción de ecuaciones alométricas para estimar
el volumen y la biomasa de los árboles. Paris: FAO.
Prado, J., & Alarcón, C. (1991). Funciones de biomasa de
Eucalyptus globulus en la región costera central. Ciencias e investigaciones forestales, 5(1), 59-70.
Prodan, M., Peters, R., & Cox, F. (1997). Mensura Forestal.
San José: Ins tuto Iberoamericano de cooperación
para la agricultura (IICA).
Schlegel, B. (2001). Es mación de biomasa y carbono en
bosques del po forestal siempreverde. Vaidivia, Chile: Simposio Internacional Medición y Monitoreo de la
Captura de Carbono en Ecosistemas Forestales. 13 p.
Schulze, C., Urru a, J., & P., H. (2008). Generación de valor agregado y encadenamiento produc vo para la especie Peumo (Crytocarya alba) (pp. 44): U Mayor.
63
Pablo Cruz
Alejandro Bascuñan
Juan Velozo
64
Anexos
65
Pablo Cruz
Alejandro Bascuñan
Juan Velozo
66
FUNCIONES ALOMÉTRICAS PARA EL BOSQUE MEDITERRÁNEO CHILENO
67
68
"#%
"&!
# !
#&
$$
'%!
! %
!'
! %
!!&'
'%"
! &
! ! ! !!'$
!$!
!&'!
" "#
"$&
# &
#&"
"#%
"&"
# #
#&!
$$"
%%
'&
&
'!
## &$ '%"
!!
!"
! !#
!"
!$ !'!
" #&
"$'%
# !
#&$'
$$ #
%#
&#!&
"#%
"&"
# $
#&#
$$$
%%"
' &%
! #$!
&%$
! "$
!$&
"&!
'%#
!!
!#
! !%
!""
!$ #
!'
" $'
"%
# "&
#&' $$#!
%#"#
&#&
'%% ! $%$
""%
$!%
"#&
"&#
# %
#&$
$$&
%%$
'$
' !&
#% &&%
! $
!$'%
" "
"%&&
#"##
$ %$"
& '%# "#&
!! "&#
!% # %
! !' #&%
!"% $%
!$! %%'
!'$ ''
" %%
'$
"% !"
# $!
#%'
#'
&'$
$$%% ! % %#%" !%
&$$ " '&" "&'
&! #"&
%!% $ "
""&' %
$"#" &$!
&$"& '!&
&$ !! '%#
!! !&
! "
!"
!$!"
!' " &"
"%! # %#
#' $
$$'$
%#''
&$"$
'&#
%
%&&
"##
$# #
&%!
&
!&' $&&
!$
"#&
"&$
# &
#&&
$%
%&
' ''
!&
#&#
'"
! &
!% !
" !$
"& %
##
$ $$
% '
&'%
'%&
!%'
%%
!%
"%%'
# &
#&'
$% %& ' "
! !
#'
'
! '
!%!"
" "'
"&" ## $ &&
%##
& &
' !
"'
%#"
! #
"&"
$$%
&#!$
$!"
'
#!%"
&!"
!'
! " !" !$!%
!' $
" '
"%"
# &$
#'"
$%"
%$ &$% '&& $"
&!!
"$!
$#&%
&&!
$"
!'&&
$'&'
! &#
!!&' !%& &
" "$%#$
#%&"
#% #
"!
! $
"%#"
#'#
#!"
'$
#'$!
! '%
$%"!
!%"!
%$#$
" $
&%$
"&#$
''!#
##!$ ! &
$!& ''
%%& "$' &## $$' ' "# &' "
"#$
""
%'#
""&
! % %% "&'! !"'!
$$$& !" &
&$# !&'"
% " "'
''% "%'
#"% # #$
&" #%$ $
!% $!#%
!"' $'&#
!%" $ %$$"!
"'%% &!'%
""%%! '$'
"&& ! '''&$
#!!% && #%% # & '
& !&%&#
"'''$
" '$
"%"%
# '#
#'# $% '
%$!'
&$'#
''
'&
&%!
"$"'
$$" &&$%
!!&
"% %&$
!!'#
!"%
!%'$'
" $&
"$'! #'&
#%"!
$!!
$'$
%$!%'
&!$ %
'!%"
''$!'
&""!
%&%
%
! '
#''
' !!"
!%#
" %
"&$&
###
$! #
%''
&%'
' %!
"&&
&!!
!!"
"'" $% !
&$$$
%&
!%"
###%
& !%
! !
!" $
!%###
"'
""' '
"&''!
#!! !
#%' %
$ &&
$&#
%!"'&
"!#
"%$
#! #'%!
$%##
%$%
&%!"
''#&
!##
'"
"%!
$%!$
&'%$
"$"
" %
% #
!#&!
!" !
!& '
" #!%
"% !"
# !$
#%&!
$!%"
%$%
%$&& &"&
''%'
!
'$#
&#'"
&$%
!' $
#"%"
!! $!
' %
!!
!%#'
" &
"&%'
##$"
$!"
% &
& ' '&
#%
&$$
!!#!
"'&$
$%%!
&% &"!
!""
#$!#
&! "
! !"!!!
!%$% " "&
"#%
"'"%
#!"'
#&
$!%
$& %!%! Pablo Cruz
Alejandro Bascuñan
Juan Velozo
"%$#
#!'
#'& $%$$
%$&#
&%# ''%'
!&
'%
"%$#
$%%%
' !
#'
" &
%! !
!$$#
!"! !&!!
" $##
"%!$#
# "$ #%'$$
$!&'"
% $$
%% &"" ' "!
#&'
'"&
&'!$
&'%'
!'$!
#'
Continuación Quillay.
!!#
$$
'!
!!$
!%$$
" &&
"&'
##%$
$!#$
% !#
& '! #""
&'%
!!&&
# $
$&&
&%%!
'
! '
#%%
&"!
!!&
!""!
!%%&
"!$$
"#''
"' &'
#!$"#
#& %'
$! $&"'
%!'"$
"%%
#! #
#''
$%%%
%$'&
&%$&
'''&
"!
''&
"%'%
$&#
'$%
#%
"!!&
%!'
!%"
!""&
!&"
" %$!
"%"&$
# #'%
#&$
$"$
%"#
%%!"
&"$"!
' "&$
&#$
'%%
' # '! !
"!
#!"
!!%
$'
'!#
!! !%% " '$
"&''
##&&
$!$'
% #
& !'
'!"!
#$'
' $
!" #$!
$&$
&& '#"
! $&
#%%!
&"%%
!!&'
!"# !%&%'
""%#
"#!&
"'"'
#!%&&
#&"!"
$!!$&
$&$
%"""
$%&%
%%
&%& $
" #
! "
"& %
$&#
'&
$%
"!' %"#
!&
!""&&
!&"''
" &$!
"%#'&
# % !
#& ##
$" "
%$ %%"'
&"&"'
' % !
"
'#"$
'$" "!"'
#$&'
!! #
!%%&
"!!
"'&
##'&
$!& % $#
& #%
'!$!
#' '# !"#
#'%
$&''
&&$#
!
!! !
#&!"
&#"#
!"$#
!"$"
!%'%
"#%$
"#" '
"'#"
#!' #&#%&
$!# "
$&%%
%"! %
""#
!"&
"&##
$&&!
'"$
$$
"""!
%#'
!&%$
!"#$ !&#&!
" '#%
"%%!
# &"
#&!&#
$"!#&
%%%'
%%$$%
&#"
' ' !
!!!
' '& ''!'
"$%
# !&
'& % ""'
%#$!
!'!&
!"$!
!&$$
"!"#
"%& # '"'
#&#%
$""'!
%' &
%%&!
&# '! #"'
# &
%%
! &
"'%"
# !$$
#$&
$!'!
% %'
& %!
'!& $"
'%%
!"%'
# '
$'!#
&'$
"'
!!%#
#&'
&$'
!#!$
!"$& !&#%
"$%
"##! "'$#!
#""!
#&% $!$%
$&' '
%""''
'!''
$!"
!#$
#$
$'$'
&'""
' !" !
#'"#
&$$&
!$ !"%##
!&!%
"%#'
"#$ '
"'%#'
#"#'
#&&!&
$!&%
$'%%
%"$#'
%$"
!''$
!"$'%
!&%!#
"! &
"%&'"
#!#!
#&$ $"$ %$&
%%'&#
&#!' '!!%
%#!
%#
!&
!"&
" #'
# $%"
!#!
#&&
%
&'&
!!
!"$&
#''$
&% #
!$$"
!"& !
!& !
"&" "#% "'&#'
#" $%
#&'#$
$!'!$
$' %
%"&
!"%#&
!&&#
"! $
"%'& #!#
#&%!
$"%"
% %&!
&### '!"&!
'"#
'#'
#"%
!% '
"# &
# '$#
%! '"
%
!#
$""
&%%&
!% !
!"&&&
!& &"
"' "#&%
"''#!
#"!%
#'$&
$"#%
$'!"'
%"'#!
"&$#
#! "!
#&&!!
$"&#"
%! %
%& %
&#%"
'!$#
%
#&
%$!
!'$"
"%&!
#! "
!## $&'
&& &
!%%'
!"'#
!&!# "''#
"#&&&
#" #""$&
#'%"
$"%!
$'"%"
%#&&
FUNCIONES ALOMÉTRICAS PARA EL BOSQUE MEDITERRÁNEO CHILENO
69
Pablo Cruz
Alejandro Bascuñan
Juan Velozo
%&
70
/
0
1
2
3
+*
++
+,
+-
+.
+/
+0
+1
+2
+3
,*
,+
,,
,-
,.
,/
,0
,1
,2
,3
-*
-+
-,
--
-.
-/
-0
-1
-2
-3
.*
.+
.,
.-
..
./
%&
%&
,"2
.",
/"3
1"3
+*"-
+-"+
+0"-
+3"2
,-"1
,2"*
-,"1
-1"2
.-"-
.3"-
//"0
0,".
03"1
11".
2/"/
3."+
+*-",
++,"1
+,,"1
+--",
+.."+
+//"/
+01".
+13"3
+3,"1
,*0"+
,,*"*
,-."/
,.3".
,0."2
,2*"1
,31",
-+.",
--+"1
-.3"1
-02"-
-21".
/"0
2"/
+,"*
+0",
,+"+
,0"2
--",
.*".
.2"-
/1"+
00"1
11"+
22".
+**"/
++-"/
+,1".
+.,",
+/1"3
+1."/
+3,"+
,+*"0
,-*"*
,/*".
,1+"2
,3."+
-+1".
-.+"2
-01"+
-3-".
.,*"2
..3"+
.12"/
/*3"*
/.*"/
/1-"*
0*0"0
0.+"-
011"*
1+-"3
1/+"2
13*"2
%&
%&
%&
-"3
/"2
2"-
++",
+."0
+2"/
,,"3
,1"2
--"-
-3".
.0"*
/-",
0+"*
03".
12"-
21"3
32"+
+*3"*
+,*".
+-,"/
+./"-
+/2"1
+1,"2
+21"/
,*,"3
,+3"*
,-/"2
,/-"-
,1+"/
,3*"-
-*3"3
--*",
-/+",
-1,"3
-3/".
.+2"0
..,"/
.01"+
.3,"0
/+2"1
/./"0
*"1
+"*
+".
+"3
,"/
-",
."*
."2
/"2
0"3
2"*
3"-
+*"0
+,"+
+-"0
+/"-
+1"+
+3"*
,*"3
,-"+
,/"-
,1"0
-*"+
-,"0
-/"-
-2"+
.+"*
.."*
.1",
/*"/
/-"3
/1".
0+"+
0."3
02"2
1,"2
11"*
2+",
2/"1
3*",
3."3
+"+
+"0
,"-
-"+
."*
/"+
0"-
1"1
3",
+*"2
+,"1
+."1
+0"2
+3"+
,+"0
,.",
,1"*
-*"*
--",
-0"/
.*"*
.-"1
.1"0
/+"0
//"3
0*"-
0."3
03"1
1."1
13"3
2/"-
3*"3
30"1
+*,"1
+*2"3
++/"-
+,+"2
+,2"0
+-/"0
+.,"2
+/*",
%& %&
1"-
++"*
+/"0
,+"*
,1".
-."1
.-"*
/,"-
0,"1
1."*
20"/
+**"*
++."0
+-*"-
+.1",
+0/",
+2.".
,*."2
,,0"-
,.3"+
,1-"*
,32",
-,."1
-/,".
-2+"-
.++"0
..-"+
.1/"3
/+*"+
/./"/
/2,"-
0,*".
0/3"3
1**"1
1.,"3
120"/
2-+".
211"2
3,/"/
31."1
+#*,/",
+"-
,"*
,"2
-"2
."3
0",
1"1
3".
++",
+-",
+/"/
+1"3
,*"/
,-"-
,0"-
,3"/
--"*
-0"0
.*"/
.."/
.2"2
/-"-
/2"+
0-"*
02",
1-"0
13",
2/"+
3+",
31"/
+*."+
++*"3
++2"*
+,/"-
+-,"2
+.*"0
+.2"1
+/1"*
+0/"/
+1."-
+2-"-
%&
%&
,",
-"-
."1
0".
2"-
+*"/
+-"*
+/"2
+3"*
,,".
,0",
-*"-
-."1
-3"/
.."0
/*"*
//"2
0,"*
02"/
1/".
2,"1
3*"-
32"-
+*0"1
++/"/
+,."0
+-.",
+.."+
+/.".
+0/",
+10"-
+21"2
+33"2
,+,",
,,."3
,-2"+
,/+"1
,0/"2
,2*",
,3/"+
-+*".
+*"2
+0"-
,-"*
-+"+
.*"0
/+".
0-"1
11"/
3,"2
+*3"1
+,2"+
+.2"+
+03"2
+3-"+
,+2"+
,.."2
,1-",
-*-".
--/"-
-03"*
.*."/
..+"3
.2+"*
/,,"+
/0/"*
0*3"2
0/0"/
1*/"+
1//"1
2*2",
20,"1
3+3",
311"1
+#*-2",
+#+**"1
+#+0/",
+#,-+"2
+#-**"/
+#-1+",
+#..."+
+#/+3"*
FUNCIONES ALOMÉTRICAS PARA EL BOSQUE MEDITERRÁNEO CHILENO
71
$#
&
$& !'
%! #
%% $$
&! &!
&' (
'$ ! !"#
!$(
(
%
!#
"#
#$
$&
& $'
%"
&
&&
'$
(#
$
%
!'
!% %"
"% %&
# &
$$ &&
& '$
$' &(
&' ''
((
((
&( '(
!$ !""
$
!! !"$
%
!&
"#
!!'
" !%' ""
" !
$$
!$% !& "% $!
"$(
&'
!'' #" "#& &!
# !#
"!! % "(! (#
#%&
!" "$( &' ## ! '
$!&
!%!
"(( (' #(! !##
$("
!(#
$#' !&!
%%"
"!(
%' "!
&"'
"%%
%& """
' &
#$
&"( "%%
( ##&
' # ((
#( ''# #"'
'"
$"&
(%" #&&
' $'%
#$ $ '
!'#
%"&
"(!
%(
$#
&#%
%! '#
&#"
'%#
'%(
(!&
!
((!
! "%
$(
!!&&
!(
!#!"
! ''
!!!
!%!
"'
"$(
# %
#&(
$#&
%! &
&'$
'&%
(&!
&#
'!
!($
# #
$"(
%&
'%
(#'
!(%
!!$
!#(
!$&$
!&#%
!(!!
" $
"!("
"#'&
"%'&
"'("
("
"
$"
&'
!%
!"&
!& "'
"#&
"'(
#"#
#'!
$""
$'%
%#!
& &%"
'!'
'(%
(%%
"(
%
($
!&&
"% ##(
$"(
%""
&!(
'!'
("
!(#
"#'
#(
#&&
$$ %"!
& (
' "
( #
!!
"%
!$&
"'$
$!
%% ' (%$
! !&
!!($
!#& !%$"
!'#!
""'
"!# "#$ "%%'
"'( # !!
#"$(
#%"
#'$$
$ "
#%
&"
!"
!"%
!&"
" "
"$&
#"
#$"
$&
$%"
%!"
%'&
&$#
'!#
'(&
(&#
$#
"'
!!$
" %
#(
$&
%&
& ' (
(!(
!##
! % !!'"
!#&
!$"$
"( #%!
$#
%!&
&! '!"
(""
$
&%
"(
#$
$((
&$%
(! !(#
!!&$
!#%"
!%%
!'%$
"&&
"!('
"$!%
"&%"
#&
#!%
#$! #&'(
$%%
$"$ $%#"
$(##
%!$"
%$&
%'($
&!!'
&$%(
&( (
(#
!!(
!%'
" "$&
#'
#%!
$! $'"
%#(
& (
&("
'& ($"
"'
!'
!! " (
#!
$!%
%"$
&#(
'%%
('&
! !
!!# !"&$
!$ !
!%$"
!&('
!(#&
" "!$'
"# (
"$'#
"&$"
"(!%
$ %
%&
&%
' #
(" $'
("
""&
#(
%$!
'!"
!#
! ("
!"(!
!$((
!' %
"#!
"!&&
"$! "&&$
#"&
#"(
#$(
#''
$ &(
$#''
$'%
% ""
%#%(
%' #
& %(
&$""
&(&
'!'(
'%' ('"
!$%
" "$
##
#%!
$!#
$( %%"
&"(
' (
(#
((#
'&
'%
!'(
"(%
$'
%!$
&#%
'&!
!!
! "%
!!&%
!#!
!$%'
!&! !'&(
"# "!'
""&(
"$$$
"&"$
"(!
# #"$
#$#
&'(
( !
#$
''
"# $$
%&(
'%"
!$&
!!%!
!#&&
!&!
!("'
" '#
"##
"&&
"('#
#!& #$%(
#'&'
$ (&
$$!%
$'%%
%! %
%$&&
%(#'
&"!(
&&!!
' !#
'$"'
'(% ("(%
('# !(%
"( #$!
$ '
$'(
%%$
&#%
'"!
(!#
!
! !!'
"#
#$&
$&(
&%
'"'
(&$
! '
!!%%
!# (
!$&&
!&#
!('
"'!
"!% "##$
"%"#
"'!(
#!'
#!""
###"
#%$(
#'&(
$ $
'&$
"
%!
"!"
#($
%&(
'&#
!' !"
!$!(
!&& "!#
"!'(
"$%$
"'$"
# $"
##%#
#&'&
$ !!
$#%(
$'!&
% (&
%$&(
%(&!
&"&&
&&(#
'!!"
'%%#
( %
($' $&
$#$
#$
$$%
#"#
$!
$&%
%$%
&# '""
(!(
"!
#
!$#
"&#
#((
%" &%'
( !$(
!! #
!"&#
!$#
!& !
!''(
"&"
"!%!
"#$&
"%$'
"'%$
#&&
#!(%
#$!
#&$
#('%
$!!'
$#&%
$&"
!'#
#%"
%$$
'%
!&&
!"&
!$$ !'&
"&%
""$'
"%$"
"(% #!' #% $
#(%!
$"!!
$%($
%' %#'
%'(!
&" &
&&$$
'!&
'%& ( #(
(%#
##
%% ( &"$
!!( !'% %"&
&!$
'! (!!
"
##
!%$
"(!
$!$
%%$
' (%#
! !"
!!''
!#%
!%"(
!'!#
" $
"! "
"# &
"%!'
"'#$
#%(
#!((
#$"%
#&'
$"
$!'%
$$#(
$' '
%(#
%"&&
#(
%'
'!
!#%
!!'&
!$# !' "("
""( "&"
#!(
#"%(
#&!#
$("
$#&%
$'&#
%!'%
%& "
& $#
&% ''
'$%#
(%"
($&&
$
%#'
!$
&&&
!"%"
!(%#
"$&(
#! 72
%((
&(&
(!
$
"#
!%
"("
$"#
%' '"%
(('
! %%
!"#!
!$!$
!& $
!( "
" &
""!(
"$#&
"&&"
#%
#!#%
##(#
#&#(
$ $!'
$$$%
$'"(
% "
%#!'
%&""
&#%
Pablo Cruz
Alejandro Bascuñan
Juan Velozo
FUNCIONES ALOMÉTRICAS PARA EL BOSQUE MEDITERRÁNEO CHILENO
%&
/
0
1
2
3
+*
++
+,
+-
+.
+/
+0
+1
+2
+3
,*
,+
,,
,-
,.
,/
,0
,1
,2
,3
-*
-+
-,
--
-.
-/
-0
-1
-2
-3
.*
.+
.,
.-
..
./
%& %&
%&
%&
%&
%&
%& %& %&
-"*
0"*
+*"0
."+
*"2
+"+
0"3
+"-
,".
."0
3",
+0"-
0"-
+",
+"1
+*"0
,"+
-"0
0"0
+-",
,-".
3"*
+"1
,"/
+/",
-"*
/",
3"*
+2"+
-,"*
+,"-
,".
-".
,*"2
."+
1"+
++"2
,-"3
.,"-
+0",
-"+
."/
,1"/
/".
3".
+/",
-*"0
/."+
,*"2
."*
/"2
-/",
0"3
+,"+
+3"*
-2"-
01"1
,0"*
/"*
1"-
.."+
2"0
+/"+
,-"-
.1"*
2-"+
-+"3
0"+
2"3
/."+
+*"/
+2"/
,2"+
/0"1
+**"-
-2"0
1".
+*"2
0/"-
+,"1
,,".
--"/
01"/
++3".
./"3
2"2
+,"2
11"1
+/"+
,0"0
-3".
13".
+.*"/
/."*
+*"-
+/"+
3+".
+1"2
-+"-
./"2
3,".
+0-"/
0,"2
+,"*
+1"0
+*0".
,*"1
-0".
/,"3
+*0"0
+22"0
1,"/
+-"3
,*",
+,,"1
,-"3
.,"*
0*"/
+,+"3
,+/"1
2,"3
+/"2
,-",
+.*"-
,1"-
.2"+
02"1
+-2".
,./"*
3."+
+2"*
,0"-
+/3"-
-+"*
/."0
11"/
+/0",
,10".
+*0",
,*"-
,3"1
+13"2
-/"*
0+"0
20"3
+1/"+
-+*"*
++3"+
,,"2
--"-
,*+"0
-3"-
03"+
30"3
+3/".
-./"2
+-,"3
,/".
-1"+
,,."3
.-"2
11"+
+*1"0
,+0"3
-2-"3
+.1"/
,2",
.+",
,.3"1
.2"0
2/"0
++2"3
,-3"2
.,."-
+0-"*
-+",
./"0
,10"*
/-"1
3."0
+-*"3
,0-"3
.01"+
+13"/
-."-
/*"+
-*-"3
/3",
+*."+
+.-"0
,23".
/+,",
+30"2
-1"0
//"*
---",
0."3
++.",
+/0"3
-+0"-
//3"2
,+/"+
.+"+
0*"+
-0."+
1*"3
+,."2
+1*"3
-.."0
0*3"2
,-."-
.."2
0/"/
-30"1
11",
+-/"3
+2/"0
-1.",
00,"-
,/."/
.2"0
1+"+
.-*"2
2-"3
+.1"0
,*+"*
.*/"-
1+1",
,1/"0
/,"1
11"*
.00"/
3*"2
+/3"2
,+1"+
.-1"1
11."1
,31"1
/0"3
2-",
/*."*
32"+
+1,"1
,-."*
.1+"1
2-."2
-,*"1
0+"-
23"0
/.-"*
+*/"1
+20"+
,/+"/
/*1"+
231".
-.."2
0/"3
30"-
/2-"2
++-"0
,**"*
,03"2
/.."*
30,"1
-03"3
1*"1
+*-".
0,0",
+,+"3
,+."0
,22"3
/2,".
+#*-*"0
-30"*
1/"1
++*"0
01*".
+-*"/
,,3"1
-*2"1
0,,",
+#+*+"-
.,-"+
2*"3
++2",
1+0".
+-3"/
,./".
-,3",
00-"1
+#+1."0
./+"-
20"-
+,0"+
10."*
+.2"1
,0+"2
-/*"/
1*0"0
+#,/*"0
.2*"/
3+"3
+-."-
2+-"/
+/2".
,12"1
-1,"0
1/+"+
+#-,3".
/+*"2
31"0
+.,"1
20."1
+02"-
,30"-
-3/".
131",
+#.+*"3
/.,"+
+*-"0
+/+"/
3+1"2
+12"1
-+."/
.+3"+
2.."3
+#.3/"-
/1."/
+*3"2
+0*"/
31,"1
+23".
---"-
..-"/
23.",
+#/2,"/
0*2"*
++0",
+03"3
+#*,3".
,**".
-/,"1
.02"2
3./"+
+#01,"0
0.,"0
+,,"3
+13"0
+#*22"*
,++"2
-1,"2
.3."2
331"0
+#10/"/
012"-
+,3"1
+23"/
+#+.2".
,,-"0
-3-"/
/,+"1
+#*/+"1
+#20+"-
1+/",
+-0"1
,**"*
+#,+*"2
,-/"1
.+."2
73
Pablo Cruz
Alejandro Bascuñan
Juan Velozo
74
FUNCIONES ALOMÉTRICAS PARA EL BOSQUE MEDITERRÁNEO CHILENO
%& %& %& %&
%&
%&
%& %&
%&
%&
.
+"1
-"0
0"*
+"3
*"3
*"1
-"+
+"/
+".
/
,"2
/"1
3"0
-"+
+"/
+"+
."3
,".
,"-
0
."+
2"-
+."*
."/
,",
+"1
1"+
-"/
-"-
1
/"0
++"/
+3"-
0",
-"*
,"-
3"2
."3
."0
2
1"/
+/",
,/"0
2",
-"3
-"*
+-"*
0"/
0"+
3
3"/
+3".
-,"1
+*"/
/"+
-"3
+0"1
2"-
1"2
+*
++"3
,.",
.*"2
+-"+
0"-
."2
,*"2
+*"-
3"1
++
+."/
,3"0
.3"2
+0"*
1"1
/"3
,/"-
+,"0
++"2
+,
+1".
-/"/
/3"1
+3"+
3",
1"+
-*".
+/"+
+.",
+-
,*"0
.+"3
1*"0
,,"0
+*"3
2".
-/"3
+1"3
+0"2
+.
,."*
.2"3
2,".
,0".
+,"1
3"2
.,"*
,*"3
+3"0
+/
,1"2
/0"/
3/",
-*"/
+."1
++"-
.2"/
,."+
,,"0
+0
-+"2
0."1
+*2"3
-."3
+0"2
+,"3
//"/
,1"0
,/"3
+1
-0"+
1-"/
+,-"1
-3"1
+3"+
+."1
0-"*
-+"-
,3".
+2
.*"0
2,"2
+-3".
.."1
,+"/
+0"0
1+"*
-/"-
--"+
+3
./"/
3,"1
+/0"*
/*"+
,."+
+2"/
13"/
-3"/
-1"+
,*
/*"1
+*-",
+1-"1
//"1
,0"2
,*"0
22"/
.."*
.+"-
,+
/0"+
++."-
+3,".
0+"1
,3"1
,,"3
32"*
.2"1
./"1
,,
0+"2
+,/"3
,+,"*
02"*
-,"1
,/",
+*2"*
/-"1
/*".
,-
01"3
+-2",
,-,"1
1."0
-/"3
,1"0
++2"/
/2"3
//"-
,.
1.",
+/+"+
,/."-
2+"0
-3"-
-*",
+,3"0
0.".
0*".
,/
2*"2
+0."/
,11"*
22"3
.,"2
-,"3
+.+"+
1*"+
0/"2
,0
21"1
+12"0
-**"1
30"/
.0".
-/"1
+/-",
10"+
1+".
,1
3."3
+3-"-
-,/".
+*.".
/*"-
-2"1
+0/"1
2,".
11"-
,2
+*,".
,*2"0
-/+"+
++,"0
/.",
.+"1
+12"2
22"3
2-".
,3
++*",
,,.".
-11"2
+,+",
/2".
.."3
+3,"/
3/"0
23"1
-*
++2"-
,.*"3
.*/"0
+-*"+
0,"0
.2",
,*0"0
+*,"1
30"-
-+
+,0"1
,/2"*
.-.".
+-3"-
01"+
/+"0
,,+"-
++*"*
+*-",
-,
+-/".
,1/"1
.0.",
+.2"3
1+"1
//"+
,-0".
++1"/
++*",
--
+..".
,3."+
.3/"*
+/2"2
10"/
/2"2
,/,",
+,/"-
++1"0
-.
+/-"1
-+-"*
/,0"3
+03"*
2+".
0,"0
,02".
+--".
+,/"+
-/
+0-"-
--,"0
//3"3
+13"0
20"/
00"/
,2/",
+.+"1
+--"*
75
Pablo Cruz
Alejandro Bascuñan
Juan Velozo
76
FUNCIONES ALOMÉTRICAS PARA EL BOSQUE MEDITERRÁNEO CHILENO
77
78
! !'
"$
#$
$$
%'
'!
' $#
$' !" %&
& " '"
'$ "& !
!"
#'
&&
#'
" !#
'
&!
$'
"
! &
"#'
#'&
%""
&'&
$
'
"$
#' %&#
'&%
!'%
#"& #$
%! !"%
' " "" ""!
&''
''#
#$
#%
%" !# &'
"!& %' #'% %&%
!% $'
!''
""%
$" !# % "
!& $# "$! % &#
!#%
& $"#
" #'" !!$ $#'
" $
!&
#"
# #% '% #" $ $
&&# ' '
"&
"' ! #""
%!% &#$
&
!' '! ""
& &'
!" $& !&"
''$
&'$
& %"
$! ##%
"&%
" !
!$#
"'
"" !'#
!# ! %"
"
'
&
&%
'%%
&%"
%%&
$&'
$$
#!
"$
!'$
!!&
$!
#"
'# "# !!
'!$
&$
%&&
%'
$##
#'!
#!$
"& "!
!&"
!"
''
$
%
'#
$$
"
&
'&
&
$#
# "
!!
"$#
"!
!$$
! &
""
'
%'
#
$
"
&#
$'
##
"!
& !&
#'' & "& &%
" '%
# &&! ' &
% # &#
#%# %%$
"!! %$
'' $"
% #%&
#" #'
'" &!'
%" $# #%
"'"
" #
!$ !$
##
% !'
&&
''%
%&!
#%%
!&
'!
#
&"$
$&#
#!!
!&'
#"
$
%
&'$
%' $'$
$&
# %
"#!
!&#
! #
%
"
!$''
''"
'
&!
%#$
$&#
$&
###
"'%
"" !'
!"!
!
$
!
'
$
!"
'
& "
%"
#&&
"%&
!% %
%"
&
% #' !"##
#%$ !"## %!
"%! !
& !%! !' "&" !
%&
&$
''
#
! &#
!'
'&
$ !
#
&"
$%
"
!
'
'
%#
$
"'
!'
! &"
$"
#! #
'
&"
#
!
''
&
$"
!
$'
!'
#$
"$
!%
!
"
'!
%#
$ "
& '
#! %#
% $ # # &$
%
#&
$ $
%
# ##
#
&&%
%%'
$%'
#&&
##
" '
"!%
!&"
!!#
'
"'
%&
!$
'$%
&
#'
&'" " % !""
$%! !&
"$! "
!%'
%&
'&&
&&
$!%
"%%
! %
&$
#"
'!
&&
!'! '!' "#$
"'
"!%
!$%! & !&& '" "&!
!" # $&& !$' %&" !&%
!&$ #% !!$% $$ !#"
'#% "#& ! # #" ! &%
%!' !#
#! "%
!! "'
$ #
% && &' '! &! %' &"&
% % ##& %$&
$#$ "# $'!
#' $ $ # % & ##$
"$' ! "'#
""
!$"
!&
%#
!$
$'
"
$
" #$
'$
%'
$ &
"%#
!!
'$
$'
'#
&"
%!'
$""
##&
"%'
"&
!"!
&$
!#
##
!
&%%
%"$
$ #
!&$
%%
%!
%"
'&
&'
&%
% &
$#"
#&"
# "#'
"!
"#&
" %&
!'&&
!%
!""!
!&&
'"#
% "'
&
&
&'!
%"
#"$
!&'
"
'!!
&"#
%$ $&#
$ #""
"&
&
#&
'
'$$
& '
$'&
#% "# !!%
"
$
!
'%"
"%&
""$$
"$!
!&%!
!#'"
!! &
!%"
&!
$
!&
% '%#
%&'
$!
""'
!#%
#!
''
%% $"
##
!'$
& %"
%
'%"
&& %'#
%"
Pablo Cruz
Alejandro Bascuñan
Juan Velozo
FUNCIONES ALOMÉTRICAS PARA EL BOSQUE MEDITERRÁNEO CHILENO
" " "
"
" " " "
!
! ' !
!
!
/0
/0
/0
/0 /0 '/0
/0 #/0 #/0 #/0
9
5+:
7+7
9+<
5+=
4+;
4+;
7+6
5+6
5+8
:
6+:
9+6
=+4
7+4
5+5
5+5
8+=
5+=
6+6
;
7+;
;+9
57+4
8+7
5+:
5+:
;+5
6+;
7+6
<
9+5
54+7
5;+=
9+=
6+6
6+7
=+;
7+<
8+8
=
:+;
57+;
67+;
;+<
6+=
7+4
56+=
9+4
9+<
54
<+;
5;+:
74+9
54+4
7+;
7+=
5:+:
:+8
;+9
55
54+=
66+5
7<+7
56+:
8+:
8+=
64+<
<+5
=+8
56
57+8
6;+6
8;+6
59+9
9+;
:+4
69+:
=+=
55+:
57
5:+7
77+4
9;+5
5<+<
:+=
;+7
75+5
56+4
58+4
58
5=+8
7=+8
:<+6
66+9
<+7
<+;
7;+5
58+8
5:+;
59
66+=
8:+9
<4+9
6:+9
=+<
54+6
87+<
5;+4
5=+<
5:
6:+;
98+6
=8+4
74+=
55+8
55+=
95+5
5=+<
67+5
5;
74+=
:6+;
54<+:
79+<
57+6
57+<
9=+5
66+=
6:+;
5<
79+9
;5+=
568+:
85+4
59+5
59+<
:;+;
6:+7
74+:
5=
84+8
<5+=
585+<
8:+;
5;+6
5<+4
;;+5
6=+=
78+<
64
89+:
=6+:
5:4+7
96+<
5=+8
64+8
<;+6
77+<
7=+7
65
95+7
548+4
5<4+6
9=+7
65+<
66+=
=<+4
7<+4
88+6
66
9;+7
55:+7
645+8
::+7
68+8
69+:
54=+9
86+9
8=+8
67
:7+<
56=+8
668+5
;7+;
6;+6
6<+9
565+<
8;+6
99+4
68
;4+:
587+7
68<+5
<5+;
74+5
75+9
578+=
96+7
:4+=
69
;;+=
59<+4
6;7+:
=4+4
77+6
78+<
58<+<
9;+;
:;+5
6:
<9+9
5;7+9
744+9
=<+=
7:+8
7<+6
5:7+8
:7+8
;7+<
6;
=7+:
5<=+=
76=+4
54<+7
7=+=
85+<
5;<+=
:=+8
<4+;
6<
546+6
64;+6
79<+=
55<+5
87+9
89+:
5=9+5
;9+;
<<+5
6=
555+5
669+8
7=4+8
56<+9
8;+7
8=+:
656+6
<6+7
=9+<
74
564+9
688+9
867+8
57=+7
95+7
97+<
674+6
<=+7
547+=
75
574+7
6:8+8
89<+4
594+;
99+9
9<+6
68=+4
=:+:
556+8
76
584+:
6<9+7
8=8+5
5:6+:
9=+=
:6+<
6:<+;
548+6
565+7
77
595+8
74;+5
975+=
5;9+5
:8+9
:;+:
6<=+6
556+6
574+9
78
5:6+:
76=+=
9;5+7
5<<+4
:=+7
;6+:
754+:
564+9
584+6
79
5;8+7
797+:
:56+8
645+:
;8+7
;;+<
777+4
56=+5
594+7
Nota: El cálculo del contenido de biomasa seca total y carbono total se realizó con las funciones seleccionadas para cada
especie y descritas en el documento, la biomasa seca de cada componente se determinó según su porcentaje de peso seco, a
su vez, la biomasa verde de cada componente fue representada asignándole su contenido de humedad de cada componente
y la biomasa verde total como la suma de sus componentes.
Para fijar la altura máxima en las tablas de dos entradas se aumentó el rango de altura en un 30%.
79
 0
0
Anuncio
Documentos relacionados





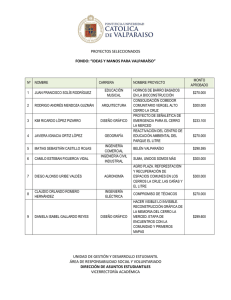



Descargar
Anuncio
Añadir este documento a la recogida (s)
Puede agregar este documento a su colección de estudio (s)
Iniciar sesión Disponible sólo para usuarios autorizadosAñadir a este documento guardado
Puede agregar este documento a su lista guardada
Iniciar sesión Disponible sólo para usuarios autorizados