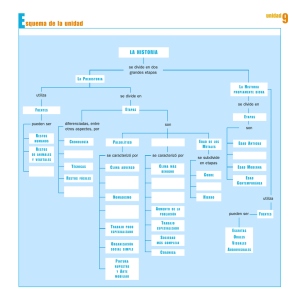
estudio de los macromamiferos de la cueva de santimamiñe (vizcaya)
Anuncio
")
KOBIE (Serle Paleoantropologia y C. Naturales) Bilbao Zientzietako Aldizkaria - Revista de Ciencias Bizkaiko Foru Aldundia - Diputación Foral de Vizcaya N.º XIV, 1984 ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMIÑE (VIZCAYA) Por Pedro M. Castaños * RESUMEN En este trabajo se estudia la fauna de Macromamíferos de la cueva de Santimamiñe (Vizcaya). Los 9.058 restos proceden de dos épocas distintas de Excavación: la más antigua realizada por T. de Aranzadi, J. M. de Barandiarán y E. Eguren entre 1918 y 1926. Posteriormente J. M. de Barandiarán vuelve a excavar entre 1960 y 1962. El yacimiento contiene niveles que van desde el Auriñaciense hasta la época Romana y quizás posteriores. Se han identificado 25 especies distintas: 6 domésticas, 8 Ungulados salvajes y el resto pertenece a Carnívoros y Lagomorfos. Hay una clara ev0lución faunística que pasa de una economía cazadora en los niveles más antiguos hasta la aparición de la domesticación en los más recientes. Se ofrece una primera parte que incluye un estudio conjunto de la fauna. Se valora la presencia relativa de cada grupo de especies con distinto significado económico. Se realiza un estudio de la proporción relativa de cada especie de Ungulados salvajes y de domésticos. Se completa el informe con el estudio particular de cada especie. En él se ofrecen tablas con los restos, medidas, resúmenes estadísticos y comparaciones biométricas con ejemplares diacrónicos y sincrónicos. SUMMARY In the present work the big Mammals fauna of Santimamiñe Cave (Vizcaya) is considered. The 9058 determinable bone remains come from two different periods of excavation: the oldest made by T. Aranzadi, J. M. Barandiarán and E. Eguren between 1918 and 1926. Later J. M. Barandiarán retook the excavation between 1960 and 1962. The site in question contains levels going from the Aurignacian to the Roman Age and a later ane perhaps. 25 different species have been identified: 6 tame ones, 8 wild Ungulate and the rest of the belong to Carnivorous and Lagomorphes. There is a clear evolution of the fauna which goes from a hunting economy at the oldest levels to the appearance of taming in the latest ones. We offer a first part which includes a point study of the faune. The relative presence in each group of species with a different economic meaning is valued. A study of the relative proportion of each specie of wild Ungulate and tame ones is made. The report es completed with the particular study of each specie in which tables with the remains, measures, statistical summaries and metric comparisons with synchronic and diachronic animals are offered. * l\(luseo Arqueológico, Etnográfico e Histórico Vasco. Bilbao. 236 PEDRO M. CASTAÑOS LABURPENA Lan honetan Bizkaiko Santimamiñeko leizeko Urgaztun Haundiak aztertzr;m dira. 9058 kutsuak bilaketak egin ziraneko aldi bikoak dira: lehenengoa 1918.etik 1926.era Aranzadi-tar T., Barandiaran-tar J.M. eta Eguren-tar E. Jaunak egindakoa: Geroago, 1960.etik 1962.era Barandiaran-tar J.M. Jaunak berriz bilaketak egiten dauz. Aztarnategiak Auriñaciensetik Erromatarren aldirainokoak eta baliteke ondorengoetarainokoak diran azalak izatea. 25 moeta ezberdinen ezagupideak egin dira, 6 etxe-abereenak, 8 ezi-bako apaxtunenak eta gainontzekoak Haragi-Zale eta Largomorfoenak izan diralarik. Aspaldiko denboratan izan eben ehiza-ekonomiatik heziketa azaldu arte abere-bilakakuntza egon zan. Abereekiko guztizko azterlana azaltzen deneko lehenengo zatia eskaintzen jatzue. Ekonomiaren aldetik esangura ezberdina daben moeta-talde bakoitzaren neurri bateko bertan izateari balioa ematen jako. Txostena moeta bakoitzaren azterlanagaz osatzen da. Hor kutsuak, neurriak, zehaztaketa-laburpenak eta lehengo eta oraingo aleen biometriazko alderaketak emanez taulak azaltzen dira. INTRODUCCION Los restos óseos objeto del presente trabajo, proceden de dos series de excavaciones realizadas con una gran distancia de tiempo en la Cueva de Santimamiñe (CortézubiVizcaya). Las primeras campañas se realizaron entre 1918 y 1926 por el equipo formado por Aranzadi, Barandiarán y Eguren. Posteriormente entre 1960 y 1962 J. M. de Barandiarán excava una zona marginal donde se conservaban testigos de la primera excavación. Los materiales óseos fueron depositados en ambas ocasiones en el Museo Arqueológico, Etnográfico e Histórico Vasco de Bilbao. Todos ellos conservaban la sigla en la que se indicaba el cuadro y profundidad. En los paquetes de la excavación antigua se añadió posteriormente un nivel con cifras romanas por el mismo J. M. de Barandiarán. Las referencias a la fauna son continuas a lo largo de la descripción de la excavación. No obstante, sólo se indica el nombre del hueso y especie en nombre vulgar así como su localización espacial. En la Memoria Segunda de las tres publicadas por el citado equipo, se dedica un capítulo a la descripción general de algunas de las especies halladas, acompañada de escasas medidas y algún dibujo original de T. de Aranzadi. l. BARANDIARAN (1967) y J. ALTUNA (1972) realizan un resumen de las especies citadas en cada nivel, con algunos porcentajes para los niveles VI y VIII que no parecen corresponder al material conservado. La relación de especies, relativa sólo a Macromamíferos, es la que sigue, utilizando la nomenclatura original de los autores: Superficial: Ganado vacuno Ciervo Perro 1-111: Marta Ciervo Corzo IV: Jabalí Cervus elaphus Capreolus capreo/us Rupicapra rupicapra Capra pyrenaica Zorro Gato montés Gamuza Cabra montés Ovis aries Oso Zorro Marta Garduña Gato montés V: Caballo (abundante) Gran Bóvido (abundante) Capra pyrenaica (menos) Rupicapra rupicapra (menos) Ciervo (muy abundante) Capreolus capreolus Sus scropha VI: G. Bóvidos Caballo Ciervo Cabra montés VIII: Ciervo Caballo Cabra montés Jabalí Oso Corzo 22 % 22% 18 % 15 % 20 % 12 % 8% 8% 6% 2% Ursus arctos Vulpes vulgaris Fe/is silvestris ve/ catus Me/es taxus Mustela nivalis Mustela erminea Gamuza o Rebeco Jabalí Oso pardo y rest. mamíferos G. Bóvido Gato montés Otras alimañas Aves Corneja Arrendajo 6% 9% 8% 44 % Po una parte, la falta de concreción de los primeros niveles, así como la vaguedad de indicaciones del nivel V, la ausencia de datos del nivel VII y el oscuro origen de los porcentajes de los niveles VI y VIII, convertían la fauna de este yacimiento en una muestra poco o nada utilizable. Por otro lado, la riqueza de fauna de Santimamiñe hacia necesario un estudio sistemático de todos los materiales ya procedieran de una u otra de las épocas de excavación. El presente trabajo es el informe del estudio completo de todos los restos determinables procedentes de los Macromamíferos. 1.-ESTRATIGRAFIA En las Memorias de excavación de las antiguas campañas se describen por un lado cinco estratos denominados por letras mayúsculas desde la A a la E. En otro lugar, se habla de hasta diez niveles identificados con cifras romanas. Los niveles IX y X tienen escasísimas piezas óseas y por tanto no se consideran en este estudio. La descripción somera de los restantes niveles es la que se expone a continuación. Las asignaciones culturales son obra de J. M. ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMlfiilE (VIZCAYA) de BARANDIARAN (1953) y no coinciden exactamente con las originales de las primeras publicaciones. Niveles 1-111: conjunto de niveles denominados también estrato A. Incluye todas las capas con cerámica. Podría identificarse cada nivel así: 1: Hierro-Romanización 11: Bronce-Eneolítico 111: Neolítico La fauna citada es común a los tres niveles. IV: denominado estrato B. Se asigna al Asturiense. V: constituye la parte superior del estrato C. Aziliense. VI: es la capa inferior del citado estrato C. Magdaleniense. VII: corresponde al estrato D. Solutrense. VIII: denominado estrato E. Auriñaciense'. Hay discrepancia entre los autores posteriores acerca de las culturas asignadas a cada nivel. l. BARANDIARAN (1967) coincide fundamentalmente con la descrita con algunas pequeñas variantes. J. M. APELLANIZ (1975) asigna a los cuatro primeros niveles culturas algo más tardías que van desde el Vascorromano (1) al Neolítico (IV). Un resumen de esta problemática se ofrece en la Carta Arqueológica de Vizcaya (MARCOS, 1982). El problema se complica aún más con las campañas realizadas entre 1960 y 1962. El propio Barandiarán nunca ha publicado la correspondencia entre los niveles de i.ina y otra excavación. En la Memoria de las campañas modernas da una estratigrafía que a efectos prácticos hemos agrupado y denominado de la siguiente forma: Nivel S: entre 500 y 640 cms. de profundidad. I': 640-700 cms. 11': 700-735 cms. 111': 735-745 cms. IV': 745-760 cms. V': 760-815 cms. Hay algunas asignaciones culturales del autor dentro del denominado nivel S que afectan sólo a ciertos sectores de la zona excavada. En el estudio del material óseo conjunto haremos algunas precisiones sobre posibles correspondencias entre grupos de niveles. 237 Estas últimas denominaciones con apóstrofe son nuestras y no tienen otro valor que la utilidad práctica a la hora de referirnos a niveles faunísticos procedentes de las campañas realizadas al inicio de los años sesenta. 2.-ESTUDIO DE LA MACROFAUNA EN CONJUNTO Este yacimiento ha proporcionado 9.058 fragmentos identificables cuya distribución por especies y niveles se ofrece en las Tablas 1a y 1b. En ellas se agrupan las especies en. tres categorías con distinta significación arqueológica: domésticos, Ungulados salvajes y restantes mamíferos. La Tabla 1a corresponde a los niveles 1 a VIII de la primera excavación ( 1918-1926). La Tabla 1b incluye los niveles que denominamos S a V' correspondientes a la excavación posterior (1960-1962). La excavación antigua con sus 8.117 restos, constituye frente a la moderna con tan sólo 941, la mayoría de la muestra. Como ya se indicaba anteriormente, J. M. de Barandiarán no pudo establecer una correlación eotre los niveles de cada una de las épocas de excavación. Si observamos la Tabla 2 con los porcentajes de domésticos, Ungulados salvajes y restantes mamíferos de cada nivel, podemos comprobar un gran paralelismo no entre los niveles considerados individualmente pero sí agrupándolos. En la excavación antigua puede apreciarse que en el nivel 1 el predominio de domésticos es total. A medida que avanzamos del nivel 11 al IV se establece un equilibrio entre ambos grupos que va resolviéndose poco a poco a favor de los Ungulados salvajes. En el nivel V la situación se ha invertido con relación al l. Y desde el nivel VI al VIII el predominio total de Ungulados y la ausencia de domésticos es la tónica dominante. Los restantes mamíferos en todo momento forman un grupo residual que no es demasiado indicativo. Este mismo proceso se puede observar en la excavación moderna. Según esto, resulta ilustrativo comparar ambas campañas asociando los niveles de la siguiente forma: el S con el conjunto formado por 11 a IV, el I' con el V y el grupo 11' a V' con el VI a VIII respectivamente. Para facilitar esta comparación se ha elaborado la Tabla 4. En ella se ha calculado el porcentaje promedio de los niveles agrupados. A la vista de la misma, parece razonable pensar que la excavación moderna representa en líneas generales la misma evolución faunística que la antigua. El único punto de divergencia sería la ausencia del estadio representado por el nivel 1 donde se observa una economía masivamente doméstica. 238 PEDRO M. CASTAi\JOS Ovis aries 3 Capa hircus 4 14 18 3 39 Ovis a/Capra h. 104 246 340 200 890 Sus domesticus 138 347 399 Canis familiaris 10 12 9 303 2 Felis catus 24 150 111 205 14 7 VII 11 148 12 Bos taurus IV VI V 8T VIII 12 Total 602 36 59 1.246 33 24 Equus ferus 1 16 16 9 21 136 77 56 332 Capra pyrenaica 1 14 3 7 17 122 28 56 248 Rupicapra rüp. 3 13 19 26 4 57 23 30 175 3 6 4 1 57 10 17 98 427 785 509 232 935 213 474 3.672 Grandes 'Bóvidos Cervus elaphus 97 Rangifer tarandus Capreolus capreolus 6 51 110 70 2 36 3 9 287 Sus ferus 5 37 51 26 4 86 3 22 234 3 10 2 21 7 57 Canis lupus Vulpes vulpes 18 6 Ursus sp. 3 2 5 4 11 8 8 13 2 Mustela erminea Martes sp. 1 2 20 Meles meles 5 Lutra lutra 1 Felis silvestris 4 Linx linx 6 6 29 3 17 1 9 12 4 32 2 2 4 8 4 9 Panthera leo Lupus sp. 1 4 20 3 A. domésticos 433 779 985 602 71 Ungulados sal. 113 561 990 651 281 1.432 359 665 26 37 22 7 35 14 10 200 572 1.377 1.997 49 1.302 359 1.467 373 675 8.117 Rest. Mamíf. TOTALES 2.875 5.047 Tabla 1a.- Distribución por niveles y especies de los restos de Macromamíferos de las campañas antiguas (1918-1926) de Santimamiñe. 239 ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMll\IE (VIZCAYA) Bos taurus s I' 13 19 11' 111' V' IV' Total 32 Ovis aries 8 Capa hircus 8 8 8 Ovis a./Capra h. 102 102 Sus domesticus 100 100 Equus ferus Capra pyrenaica 7 Rupicapra rupic. 3 12 4 1 17 29 25 2 6 2 27 18 21 Gran Bóvido Cervus elaphus 179 1 19 32 20 130 7 5 2 43 21 42 88 369 5 1 17 8 8 2 3 2 1 1 8 6 11 Rangifer tarandus 1 Capreolus capr. 18 Sus ferus' 6 Canis lupus 2 20 Ursus sp. Martes sp. 26 61 9 15 3 3 Meles meles Felis silvestris 1 Lupus sp. 3 A. domésticos 231 19 A. salvajes 213 70 101 73 90 6 2 5 11 7 450 91 106 84 97 Rest. Mamíf. TOTALES 251 113 659 114 941 32 Tabla 1b.- Distribución por niveles y especies de los restos de Macromamíferos de las campañas entre 1960-1962 de Santimamiñe. 11 111 IV V VI VII VIII s I' 11' 111' IV' V' A. Domésticos 75,7 56,5 49,3 46,2 19,7 51,3 20,8 Ungulados s. 19,7 40,7 49,5 50 78,2 97,6 96,2 98,5 47,3 76,9 95,2 86,9 92,7 99, 1 4,5 2,6 1,1 1,9 2,4 3,7 1,4 1,3 2, 1 4,7 13, 1 7,3 0,8 Rest. Mam. 3,7 Tabla 2.- Porcentajes de domésticos, Ungulados salvajes y restantes Mamíferos en cada nivel de Santimamiñe. 1 NR Bos taurus % 150 34,6 NMI % NR % 10 25,6 148 18,9 11 NMI 9 111 % NR % 17,3 205 20,8 IV NMI % 13 20 NR V % NMI 87 14,4 % NR % NMI % 1 16,6 10 22,2 12 16,9 Ovis aries 3 1 12 3 14 3 7 2 Capra hircus 4 25,6 2 30,7 14 34,9 3 51,9 18 37,7 4 47,6 3 34,8 1 41,2 Ovis/Capra 104 9 Sus domest. 138 31,8 9 23, 1 347 44,5 Canis fam. 246 10 2,3 2 5, 1 Felis cat. 24 6 15,3 TOTALES 433 5,5 39 12 779 21 1,5 340 15 28,8 399 40,5 1 52 1,9 9 0,9 985 24 200 19 29,2 303 50,3 2 65 3 2 0,3 602 16 15 33,3 1 45 59 83, 1 5 83,3 71 6 2,2 Tabla 3.- Número de restos (NR) y número mínimo de individuos (NMI), así como los respectivos porcentajes de las especies domésticas de Santimamiñe. 240 PEDRO M. CASTAl\IOS En definitiva, si la excavación moderna parece constituir un reflejo bastante fiel de la antigua y dado que su riqueza ósea es mucho menor, en el estudio conjunto de la fauna del yacimiento vamos a prescindir de ella para no caer en paralelismos nivel a nivel que pueden no ser válidos. No obstante, se mantendrá el material de dicha excavación en el estudio sistemático de cada especie y por supuesto en las tablas de medidas. A continuación se analizará por separado cada uno de los conjuntos de especies asociados en la Tabla 1a. 2. 1. -Animales domésticos: En la Tabla 3 se ofrece tanto el número de restos como el número mínimo de individuos representados por cada especie y en cada nivel. Se añade a la derecha de cada dato el porcentaje del mismo. En este apartado los cálculos se realizan considerando sólo las especies domésticas. En la citada tabla se han agrupado ovejas y cabras a la hora de calcular sus porcentajes. El nivel V refleja un momento de transición antes de la aparición de la domesticación. Aunque se han identificado en el mismo algunos restos de bovino y de cerdo como domésticos, el único criterio utilizado ha sido el del tamaño. Este criterio es muy relativo y por tanto por la proble·mática especial que comporta, dejaremos aparte este nivel en las consideraciones acerca de las especies domésticas. A la vista de la Tabla 3 se puede observar la escasa importancia del perro y del gato que no parecen haberse utilizado como aporte alimenticio. El buen estado de conservación de sus huesos apunta en esta dirección. Nos centramos por tanto en los tres grupos restantes: vaca, ovicapri- 11-IV V A. domésticos 75,6 50,7 19,7 Ungulados s. 19,7 46,7 78,2 4,5 2,5 1,9 s I' Rest. Mam. VI-VIII 97,4 2,5 11'-V' A. domésticos 51,3 20,8 Ungulados s. 47,3 76,9 93,5 1,3 2, 1 6,5 Rest. Mam. no y cerdo. En la Fig. 1 se ofrecen los diagramas con los porcentajes de cada especie doméstica y en cada uno de los niveles. No están representados ni el gato ni el perro. Tampoco se ofrece el diagrama del nivel V. En la parte superior, el cálculo se ha realizado a partir del número de restos, mientras que en los inferiores se ha partido del número mínimo de individuos. Las diferencias de porcentajes según se calculen a partir de un dato o de otro (NR o MNI) es más patente en el ganado ovicaprino. La sobrevaloración en el NMI de estas especies se debe al hecho de poder utilizar en ellas un criterio que en las otras no se da: la distinción en muchas piezas entre oveja y cabra. El líneas generales, se pueden señalar los siguientes hechos: -el ganado ovicaprino mantiene una proporción relativamente constante a lo largo de todos los niveles, proporción que oscila alrededor de un 30 % . -el ganado vacuno aumenta progresivamente desde los niveles más antiguos a los más recientes pasando de ser el más escasamente representado en el nivel IV a ocupar la primera posición en el nivel l. - el cerdo por el contrario disminuye gradualmente conforme se avanza hacia niveles más superficiales. El aumento del ganado vacuno coincide con las observaciones ya realizadas por J. ALTUNA (1980) al estudiar el proceso de domesticación en el País Vasco. La disminución del cerdo desde el Neolítico al Bronce coincide con lo observado por A.v.d. DRIESCH (1972) para yacimientos del Sur de la Península Ibérica. Tabla 4.- Comparación de porcentajes de niveles agrupados entre la excavación de 1918-26 y la de 196062 de Santimamiñe. 11 111 IV V VI VI VIII Equus ferus 0,8 2,8 1,6 1,4 7,4 9,4 21,4 8,4 Capra pyren. 0,8 2,4 0,3 1,1 6 8,5 8,4 Rupicapra r. 2,6 2,3 1,9 3,9 1,4 3,9 6,4 4,5 0,5 0,6 0,6 0,3 3,9 2,8 2,5 G. Bóvido Cervus elaph. 7,8 85,8 76, 1 79,3 78,2 82,5 65,3 59,3 71,2 Rangifer t. 0,2 0,5 Capreolus ,c. 5,3 9, 1 11, 1 10,7 0,7 2,5 0,8 1,3 Sus ferus 4,4 6,6 5, 1 1,4 6 0,8 3,3 3,9 O, 1 Tabla 5.- Distribución de los porcentajes de Ungulados salvajes en cada nivel y a partir del número de restos de Santimamiñe. ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMlf\JE (VIZCAYA) Número de restos e ..-- e - 50 - 40 o 30 e - V 20 -o ..f. ......o V -V o - V 10 Nivel 11 IV 111 V = Bos taurus O = Ovis aries + Capra hircus C = Sus domesticus Número mínimo de individuos 60 -o 50 40 - o ::!.. 30 -o - - .::!... V 20 - -e e e ~ -o - V 10 Nivel 11 Figura 1.- Histograma con los porcentajes del número de restos y del número mínimo de individuos de las principales especies domésticas de Santimamiñe. 111 IV 241 242 PEDRO M. CASTAl'\JOS 2.2. - Ungulados salvajes: La Tabla 5 resume los porcentajes de cada especie de Ungulados salvajes en cada nivel y calculado dicho porcentaje a partir del número de restos. A la vista del mismo se destaca en primer término el dominio absoluto del ciervo a lo largo de todos los niveles. En los tres niveles más antiguos se sitúa entre el 60 % y el 70 % ya que la presencia del caballo alcanza hasta un 20 % . Pero en los cuatro niveles más superficiales el ciervo está siempre por encima · del 80 % . Este predominio tan notable del ciervo es un fenómeno observado frecuentemente en la mayoría de los yacimientos cantábricos que no se hallen situados en ecosistemas especializados de roquedo, donde la cabra montés suele aparecer como especie dominante. Otro dato observable es la escasa presencia del reno en los tres niveles más antiguos. Su representación es casi testimonial, pero interesante ya que demuestra una vez más que ,esta especie, aunque en escasa densidad, convivió con ciervo hasta el final del Würm en la cornisa cantábrica. También destacan por su baja proporción los Grandes Bóvidos que apenas llegan al 4 % en los niveles más antiguos. Su presencia a partir del nivel V es también residual, estando totalmente ausente en el nivel l. Las otras cinco especies de Ungulados mantienen entre sí unas proporciones que cambian totalmente de los niveles paleolíticos a los postpaleolíticos. Para hacer más intuitiva la comparación se ha elaborado la Fig. 2. En ella se observa que en los niveles VI a VIII las especies mejor representadas son caballo, cabra montés y sarrio en el orden descrito y que las más escasas son corzo y jabalí. A medida que avanzamos hacia los niveles superiores las tres primeras especies van sufriendo una notable disminución. Por el contrario, el corzo y el jabalí aumentan progresivamente hasta constituirse en las mejor representadas a partir del nivel IV. La inflexión se produce en el nivel V que también es el nivel a partir del cual se inicia la posible presencia de domesticación. el Para poder comparar la estructura faunística de los niveles paleolíticos se han elaborado las figuras 3 a 5. En ellas se ofrecen diagramas acumulativos de los porcentajes de las distintas especies de Ungulados salvajes. Este tipo de diagramas iniciado por DELPECH (1975) en el campo faunístico ha sido utilizado en la mayoría de los yacimientos cantábricos publicados en la última década. Permiten comparar y deducir aspectos climáticos o ecológicos entre distintos yacimientos e incluso entre distintos niveles dentro de un mismo yacimiento. Santimamiñe a lo largo de sus niveles, viene a corroborar en esencia lo que ya ALTUNA (1979) en un amplio estudio sobre las faunas de Ungulados del Tardiglaciar cantábrico indicaba. Una vez más se comprueba aquí un predominio total del ciervo que no necesariamente es indicativo de condiciones climáticas más templadas ya que mantiene su preponderancia tanto en períodos wurmienses asociados al reno y otras especies de hábitat más frío, como en épocas postwurmienses. También corrobora Santimamiñe la disminución, a partir del inicio del postglaciar, del caballo, cabra y sarrio asociado al aumento de corzo y jabalí así como la desaparición del reno. La amplitud y continuidad del sustrato arqueológico de Santimamiñe inclinan a pensar en una ocupación de la cueva durante todo el año. No obstante, para confirmar esta hipótesis se ha calculado la edad en meses de varios ejemplares juveniles de ciervo. Para ello, además de los trabajos ya clásicos, se han utilizado los criterios recientemente publicados por K. MARIEZKURRENA (1983). En los niveles con suficiente material, como el VI, los meses en que se abaten estas presas cubren las cuatro estaciones. En otros niveles la falta de piezas cobradas en algunos meses se puede deber más a la escasez de la muestra que a una ocupación estacional. De todas formas, cuando se consideran todos los niveles del yacimiento, la edad de las piezas indica actividad cazadora durante todos los meses del año. 2.3. - Restantes mamíferos: Este grupo está compuesto por diez especies de Carnívoros y un Lagomorfo. En ningún nivel alcanzan el 5 % de representación. Cabe por tanto suponer que no fueron objeto de alimento y caza sistemática. Su presencia en la cueva responde a otras causas. Hay que destacar la presencia de cuatro especies que no habían sido citadas en el yacimiento: Linx línx, Panthera leo, Lutra lutra y Lepus sp. No hemos podido confirmar la presencia de Mustela nivalis citada por los excavadores. ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMll\IE (VIZCAYAl 243 Figura 2.- Diagrama con la variación de porcentajes de Ungulados salvajes de Santimamiñe. % 90 80 70 60 Cervus Equus -· - · - · - Capra - - - - -· Rupicapra · · · · · · · · · · ·· Sus -··-··-··- Capreolus 30 20 10 .. ...... -·· ---··---··-··-- ..,.·, ... ·· ····· .. . .. .. . . -- . -- . . . . . . ....... :--· .- ---·"" ··~ .. "'""'-~--··-..- - - - - 11 111 • • IV _ . .·..- --·--·__ ............... ..... .. .. ·..... -- . . -~ :o--- • ._:,..· ,.,,,.. V •• - VI •• ....... ...:..·.··~···· ·-· ·-·· VII ·v111 Figura 3.- Diagrama con los porcentajes acumulados de los Ungulados del nivel VI de Santimamiñe. 100% /V / r, 90% 80% 70% 60% 50% 40% 30% ..... 20% 1 / 10% I :JJ m z o s: s: )> e --i :JJ z o("') m :JJ o z --i m en )> ("') )> / V / / ("') ("') )> OJ )> )> OJ :JJ )> r r o en )> :JJ :JJ o co o e en~~ ºo z< --i mo o )> r ("') m s: ("') m m m :e )> < o G> ("') m :JJ o en :JJ -< o :JJ e z --i z e en ("') o :JJ N o e_ )> OJ )> r Figura 4.- Diagrama con los porcentajes acumulados de los Ungulados del nivel VII de Santimamiñe. -- 100% 90% 80% 70% 60% 50% .. 40% .~ 30% / V / / I 10% ~ m )> o ~ z 1/ j 20% :JJ l.' e --l :JJ z o() m :JJ o z --l m en )> () )> () () )> )> )> :JJ )> ce r r o ce en )> :JJ :JJ o )> r () m ce o e - . :JJ en ce o ºo z< --l mo o ~ () m G> :JJ m :e m )> () m :JJ o en o< -< o :JJ e z --l z e en () o :JJ N o e_ )> ce )> r Figura 5.- Diagrama con los porcentajes acumulados de los Ungulados del nivel VIII de Santimamiñe. 100% ---- / 90% 80% 70% . 60% 50%1 40% 30% 1 ,I~ 20% 10% ~ J/ / ~ 1 ' / V :o m z o s: )> s: :o z --i :o e o(") m o z --i m (f) (") (") (f) )> )> )> )> (") CD CD )> )> r r o :o )> :o :o o -CD .(") C :o (f) CD ºo z< --i mo o 0 )> r (") m s: m G> )> (") m :o o(f) (") m :o < o m :e -< o :o e z --i z e(f) (") o :o N o (_ )> CD )> e ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMll\JE (VIZCAYA) 3.-ESTUDIO DE CADA ESPECIE EN PARTICULAR BOVINO Bos taurus Los 643 restos de ganado vacuno se distribuyen a lo largo de todos los niveles postpaleolíticos. Representan un total de 49 individuos: 31 adultos y 18 juveniles. La Tabla 6 ofrece la distribución de estos restos según las distintas partes del esqueleto. Las medidas aisladas y sus resúmenes estadísticos se reflejan en las Tablas 28 y 50 respectivamente. La mayoría de los valores entran dentro de los dados por ALTUNA J. (1980) para ejemplares postneolíticos del País Vasco. No hemos podido observar diferencias significativas entre el nivel 111 (supuestamente neolítico) y los dos primeros. Hay que señalar sin embargo, que la atribución a Gran Bóvido de los restos se ha hecho en estos niveles atendiendo al tamaño de los huesos. Por tanto, la peque- Cavija c. ñez de las medidas de vaca son también una consecuencia lógica de este criterio de discriminación. A partir de dos Metatarsos enteros se ha podido estimar la altura en la cruz de una hembra y de un macho. Estos valores calculados a partir de los factores de Fock (1966) y Matolcsi (1970) son los siguientes: Nivel 11 Medida Sexo LM (mm) Factor Alt. en la cruz (cm) m 193 5,55 106, 15 h 218 5,35 116,63 Estas alzadas entran dentro de las publicadas por el mismo autor citado para ejemplares del País Vasco. No obstante, la muestra es muy escasa para establecer afirmaciones demasiado concluyentes. IV 111 11 247 V s I' 3 1 2 2 Cráneo Maxilar Mandíbula D. aislados 2 115 Vértebras Esternón 1 4 8 11 101 138 5 7 2 75 3 5 9 446 2 17 4 2 Costillas 6 2 Escápula 2 3 12 2 6 3 Húmero Radio Total 4 2 3 3 Ulna 2 2 Carpo Metacarpo 3 6 2 2 Pelvis 3 Fémur 2 Tibia 3 Rótula Calcáneo 2 1 Astrágalo 2 4 Resto tarso 5 1 7 3 4 7 Metatarso 2 1 2 Metap. ind. 1 2 1 Falanges 18 15 30 2 4 TOTALES 150 148 205 87 12 10 9 13 10 NMI 5 5 74 13 19 634 3 3 49 3 31 Adultos 7 5 8 6 1 Juveniles 3 4 5 4 2 Tabla 6.- Distribución de los restos de Bos taurus de Santimamiñe. 18 248 PEDRO M. CASTAJ\IOS OVICAPRINO Ovis aries+ Capra hircus El ganado ovicaprino ofrece un total de 1.083 restos repartidos en los cuatro primeros niveles. Se han podido identificar 44 y 47 fragmentos .como pertenecientes a la oveja y cabra respectivamente. El resto queda sin atribución específica. Las Tablas 7 y 8 ofrecen la distribución de los restos de cada una de estas especies según las distintas partes del esqueleto a lo largo de los distintos niveles. Como se verá las proporciones· de cabra y de oveja son muy semejantes incluso considerando cada nivel. Según esto, parece razonable suponer que ambas especies se hallan representadas más o menos por igual. Las piezas dentarias y falanges que constituyen una proporción muy importante de la muestra son de atribución . insegura. Su distribución según los mismos criterios citados se resume en la Tabla 9. Las medidas de oveja, cabra Y ovicaprino (sin atribución específica) se ofrecen en las Tablas 29, 30 y 31 respectivamente. No se aprecian diferencias significativas entre los distintos niveles. Hay que notar, sin embargo, que la · muestra es relativamente escasa. En conjunto, estos valores coinciden fundamentalmente con los dados por ALTUNA (1980) para ejemplares de todo el País Vasco. En algunos casos las medidas de Santimamiñe son algo superiores a las citadas. Pudiera explicarse esta tendencia si se consi- 11 Clavija c. Cráneo Escápula Húmero 2 2 111 4 1 2 2 IV s 2 Radio Ulna 2 Adultos Juveniles 6 6 5 Clavija c. 1 Calcáneo 3 Astrágalo 2 Falanges 2 5 3 4 14 18 111 IV s Total 19 7 4 52 10 20 12 6 54 4 Cráneo 4 Húmero 6 Radio 2 Ulna 3 Carpo 11 2 2 2 2 111 IV 1 s 2 3 Total 6 6 2 2 2 Fémur Fémur 11 16 Como puede apreciarse, el número de animales inmaduros sacrificados iguala e incluso supera ligeramente el de adultos. Total Carpo TOTALES dera la posibilidad de que se hayan incluido dentro del ganado doméstico restos de cabras monteses jóvenes. Otro motivo que puede aducirse es el predominio que parece existir de individuos machos. Entre los restos de oveja se han podido identificar cuatro machos y ninguna hembra. En la cabra aparecen cinco machos frente a tres hembras. Este desequilibrio sexual ha podido inducir un ligero aumento en las medidas debido al dimorfismo que suelen presentar estas especies. Los resúmenes estadísticos de cabra y ovicaprino se resumen en las Tablas 51 y 52. No se conserva entero ningún hueso largo. Por tanto, no se ha podido obtener ninguna estimación acerca de la altura en la cruz de los ejemplares. La distribución de adultos y juveniles en cada nivel y tomando ambas especies en conjunto se resume así: 5 5 2 5 3 5 Rótula 4 Calcáneo 2 9 Astrágalo 2 2 2 2 2 4 5 Resto tarso 1 4 3 8 47 Falanges 2 NMI 2 3 4 2 12 TOTALES 12 14 7 8 44 Adultos 2 3 3 2 11 NMI 3 3 2 2 11 Adultos 2 2 2 2 Juveniles 1 9 2 Juveniles Tabla 7.- Distribución de los restos de Capra hircus de Santimamiñe. 3 Tabla 8.- Distribución de los restos de Ovis aries de Santimamiñe. 249 ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMll\JE (VIZCAYA) 1 Cráneo Maxilar Mandíbula D. aislados Vértebras 1 IV s Total 2 6 3 20 9 4 41 7 68 28 43 25 10 113 119 162 106 10 465 5 16 28 29 22 91 Sacro Costillas 111 11 1 5 7 2 Escápula 7 5 4 8 1 7 2 7 6 25 22 Húmero 5 6 7 1 Radio 1 4 3 Ulna 3 1 Carpo 1 Metacarpo 3 1 3 2 Pelvis 1 3 1 3 8 9 10 1 25 6 2 4 1 5 7 25 5 14 3 2 12 3 2 25 Fémur Tibia ' 7 Calcáneo Astrágalo 2 4 Resto tarso 1 11 3 1 5 Metatarso 5 9 5 Desgraciadamente no se puede comprobar si los machos son adultos o subadultos. Pero los datos señalados apuntan hacia un modelo de selección bastante común desde el Neolítico hasta épocas recientes. Se trataría de un sacrificio sistemático de la mayoría de los machos antes de que lleguen a su madurez sexual. Por el contrario, las hembras serían conservadas hasta edad avanzada con vistas a la reproducción y explotación lechera y lanar. Terminamos este apartado, señalando que las frecuentes marcas de cuchilla indican un método de despiece muy similar al descrito ya en numerosos lugares y que se mantiene casi hasta la actualidad sin grandes diferencias. CERDO Sus domesticus Entre los restos de este yacimiento se han identificado 1.641 fragmentos de súido. El amplio espacio de tiempo representado hace suponer que esta muestra incluye huesos procedentes de la forma doméstica y del agriotipo salvaje. El primer problema precisamente estriba en la distinción entre ambas formas. Se trata de una cuestión ya clásica, tratada por muchos autores y que habrá de ser dilucidada con distintos métodos. Distribución de edad y sexo en la muestra: Comenzaremos por indica'r la estructura poblacional de edades y sexos tomando toda la muestra en conjunto. Ofrecemos en primer término la distribución de adultos y juveniles correspondientes a cada nivel. Se consideran juveniles a todos los menores de dos años. 4 2 1 1 8 17 5 19 5 7 9 7 246 340 200 102 55 992 21 24 16 6 76 Adultos 1 4 11 5 9 Adultos 9 3 11 14 4 32 Juveniles 5 6 10 10 12 6 44 NMI 9 10 15 10 Juveniles s I' 1 Metapodio ind. Falanges TOTALES NMI 5 104 8 27 Tabla 9.- Distribución de los restos de Ovis aries+ Capra hircus de Santimamiñe. Por otra parte, se ha podido determinar a partir de las dentaduras las edades en meses de varios ejemplares menores de dos años. El resumen de esta determinación se ofrece en el siguiente cuadro: 0-3 meses 1 1 11 3-6 meses 6-12 meses 1 2 12-18 meses 1 5 18-24 meses 111 5 3 1 4 IV 4 s 1 11 3 4 10 3 1 1 8 11 1 Total 1 A la vista de los datos anteriores, se podría esbozar el patrón de crianza y consumo del ganado ovicaprino en los siguientes términos: - se sacrifica más de la mitad de la población antes de los dos años. · -la mayoría de los inmaduros son matados sistemáticamente desde el nacimiento hasta el año y medio. - hay un predominio claro de machos entre los animales sacrificados. VIII 2 Juveniles 2 3 5 NMI 4 8 Adultos 111 19 IV 6 V 3 VI 4 9 15 2 4 5 8 111' IV' 2 2 2 11' 2 1 3 3 VII v: Los niveles 1 a IV y el S que corresponden a época postpaleolítica presentan un claro predominio de individuos inmaduros. Por el contrario, en los niveles peleolíticos dicha relación está equilibrada. Esto nos induce a pensar que se trata de un modelo de utilización que implica un control de la población por parte del hombre. La mayoría de los ejemplares se sacrifican antes del año como demuestra la siguiente distribución en la que se incjica la edad de sacrificio de los individuos inmaduros en los niveles más modernos. 0-3 meses 11 3 111 2 IV S 4 6 3 5 2 9 4 3-6 meses 6-12 meses 2 12-20 meses 2 20-24 meses 2 Total 5 1 10 10 PEDRO M. CASTAJ\IOS 250 En segundo lugar, resumimos la distribución de sexos en cada nivel indicando si se trata de adultos o juveniles. El grado de discriminación sexual es menor que el de la edad pero nos da también una idea del tipo de selección realizado. 11 adultos 111 IV 3 V VI s 111' 3 machos juveniles 2 2 3 adultos 3 2 2 hembras juveniles A la vista del cuadro puede decirse que en los niveles modernos apenas se sacrifican hembras inmaduras. Por el contrario, la mayoría de los machos muertos lo son antes de llegar a la madurez. Esta situación se altera en los niveles VI y 111' que entran plenamente dentro del Paleolítico. Esto parece apuntar hacia una conservación de las hembras con fines reproductores tal como se observa en otros yacimientos peninsulares y continentales a partir del Neolítico. En resumen, hay diferencias notables en la distribución de sexos y edades entre los cinco primeros niveles y el resto. Esto puede interpretarse como resultado de una manipulación humana que sería indicativa de un predominio de domésticos en los niveles postpaeleolíticos. Criterios biométricos de diferenciación: La disminución de tamaño como consecuencia de la domesticación ha sido uno de los criterios más usados por los autores a la hora de diferenciar la forma salvaje de la doméstica. Este método tiene sus problemas ya que los in-. dividuos salvajes jóvenes no pueden diferenciarse de algunos adultos domésticos. A pesar de esta dificultad se logra al menos separar un buen número de restos que por su tamaño pertenecerían claramente al Jabalí. El problema se presenta cuando se trata de determinar los límites biométricos a partir de los cuales se puede diferenciar ambas formas en cada pieza anatómica. Para intentarlo hemos recogido los criterios utilizados por la mayoría de los autores que han trabajado sobre ejemplares de la Península Ibérica. No se incluyen datos procedentes de yacimientos europeos, a pesar de ser más amplios, ya que las variaciones debidas a la subespeciación podrían introducir otros factores de variación. Los criterios recogidos pertenecen a los siguientes autores y publicaciones: DRIESCH A.v.d. (1972 y 1976), LAUK H.D. (1976), ALTUNA J. (1980), HAIN F.H. (1982). En la Figura 6 se ofrecen varios histogramas con las frecuencias de medidas para las piezas anatómicas más frecuentes de Santimamiñe indicando el límite de discriminación elegido. En cada caso se adjuntan los límites, superior para el cerdo e inferior para el jabalí, utilizado por algunos de los autores citados. Las falanges presentan una problemática más compleja. En ellas, además de las diferencias específicas inciden otros factores individuales tales como la posición anterior o posterior de la pieza. La dificultad para determinar esta posición en cada pieza aislada es grande. Al realizar los histogramas para las falanges, se obtienen distribuciones claramente unimodales que no proporcionan ningún criterio de discriminación. Por esta razón, no se han incluido aquí. En su caso, utilizamos para la asignación específica, criterios estimativos aplicados por otros autores. Por último, hay que señalar, que para los niveles paleolíticos, hemos considerado todos los restos como pertenecientes a la forma salvaje. La distribución de los restos de la forma doméstica, tras la aplicación de los criterios anteriormente expuestos, se ofrece en la Tabla 10. Se hallan distribuídos según las distintas partes del esqueleto y en cada nivel. Se evidencia la escasez de huesos largos y el predominio de piezas dentarias, especialmente aisladas, así. como de falanges. En la Tabla 32 se ofrece el conjunto de medidas obtenidas a partir de esta muestra. La Tabla 53 incluye los resúmenes estadísticos de las medidas más frecuentes. En cada medida (excepto en el Maxilar) se agrupan los datos correspondientes a los niveles 1 y 11 por un lado, y por otro los incluidos en los niveles 111, IV y S. Esta separación tiene por objeto comparar los niveles atribuidos al Neolítico (111, IV y S) con los del Hierro y Bronce (1 y 11). A la vista de la Tabla, se observa que las medidas de ejemplares neolíticos son claramente superiores a las que corresponden a individuos más modernos. Este hecho puede tener más de una justificación. Puede interpretarse como prueba de la progresiva disminución de tamaño a lo largo del proceso de domesticación. En este caso, los ejemplares de los niveles 111, IV y S serían intermedios entre los salvajes y los postneolíticos. Por otro lado, el fenómeno puede deberse a la inclusión de ejemplares salvajes jóvenes entre los restos atribuídos al cerdo de forma que se eleve el promedio de las medidas de éste. Incluso es razonable pensar que en las primeras etapas de la domesticación, los cruces frecuentes entre domésticos y salvajes hayan producido una población biométricamente mayor que la posterior en la que estos cruces serían mucho más raros debido a la estabulación relativa del cerdo. Una o varias de estas causas pueden ser responsables de estas diferencias observadas. Las medidas de Santimamiñe superan en general a las publicadas por ALTUNA (1980) para ejemplares postneolíticos del País Vasco. También superan a las obtenidas por nosotros en el Poyo del Cid (1981) y Bílbilis (inédito), ambos yacimientos aragoneses de época romana. Son mayores que las de la mayoría de Jos yacimientos meridionales de la Península Ibérica. No se ha conservado ningún hueso largo entero. Esto nos impide estimar la altura en la cruz de los animales. Por tanto, no podemos comparar la alzada de estos ejemplares con los de ningún otro lugar. PERRO Canis familiaris Los 33 restos de perro se distribuyen entre los cuatro primeros niveles. La Tabla 11 ofrece su distribución según las distintas partes del esqueleto. En la Tabla 33 se reflejan las medidas obtenidas. El origen de la domesticación del perro es una cuestión ampliamente tratada por muchos autores. Es opinión ya común que se trata de la especie doméstica más antigua en Europa. Su origen se sitúa entre 9.000 y 10.000 años ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMlt\IE (VIZCAYA) 1 111 11 Cráneo IV 8 5 V s Total 13 Maxilar 3 16 31 28 2 5 85 Mandíbula 7 17 32 14 5 14 89 D. aislados 92 205 175 126 20 10 628 4 5 28 6 43 Vértebras Sacro Costillas 1 Escápula 1 Húmero 2 3 Radio 1 1 4 5 1 1 1 4 2 12 3 1 2 3 6 Ulna Carpo Metacarpo 3 Pelvis 4 17 5 3 4 3 1 1 Fémur 1 3 Tibia 1 2 Rótula 1 2 1 5 9 Astrágalo 2 9 10 8 Resto tarso 1 2 8 6 Metatarso 1 2 2 Metapodios ind. Falanges TOTALES 4 1 respectivamente los siguientes resultados: 49,06 cm. y 51, 18 cm. El promedio entre ambos valores da una media de 50,12 cm. GATO DOMESTICO Fe/is catus El primer problema que se nos plantea es la discriminación entre el gato doméstico y la forma salvaje que está presente en la casi totalidad de los niveles de Santimamiñe. El criterio generalmente utilizado por los autores es el del tamaño. La forma doméstica es menor que el gato montés. Los estudios de KRATOCHVIL (1974 y 1976) sobre muestras numerosas de ambas formas parecen confirmar la validez general del método. 1 1 1 29 6 19 Mandíbula 1 2 D. aislados 2 6 Vértebras 2 4 1 11 111 IV Total 1 2 8 16 6 10 2 7 Húmero 1 3 Metacarpo 2 2 1 Calcáneo 2 251 . Fémur 8 23 3 36 Calcáneo 2 19 Metatarso 2 8 2 16 13 9 5 8 53 22 57 83 48 20 21 251 138 347 399 303 59 100 1.346 15 19 15 5 8 71 NMI 9 Adultos 4 5 9 6 3 3 30 Juveniles 5 10 10 9 2 5 41 7 2 1 2 Total 10 NMI 2 2 6 Adultos 2 2 6 12 9 2 33 Tabla 11.- Distribución de los restos de Canis familiaris de Santimamiñe. Tabla 10.- Distribución de los restos de Sus domesticus de Santimamiñe. Cráneo Maxilar 2 antes de nuestra era. Se suponen varios centros de domesticación distintos, tanto en Europa y Asia como en el Nuevo Mundo. Hay incluso autores (MUSIL R. 1974) que afirman tener motivos para reconocer cánidos domesticados en niveles Magdalenienses. En la Península Ibérica las citas más antiguas se remontan al Neolítico primitivo y Epipaleolítico (DRIESCH A.v.d., 1980) de la cueva de Nerja: ALTUNA (1980) cita restos de perro en el nivel 11 de Marizulo y en el nivel 111 de Arenaza. Este último datado en el 6. 0 milenio antes de Cristo. Con estos antecedentes no es de extrañar la presencia de una mandíbula y un fragmento de Fémur de perro en el nivel IV de Santimamiñe. Este nivel corresponde a un Preneolítico con matiz Asturiense. Coexiste con la forma salvaje. La longitud basal de cráneo estimada a partir de las medidas mandibulares y siguiendo los criterios de BRINKMANN (1924) oscila entre 150 y 151,8 mm. Corresponde a un perro de tamaño mediano (C. f. intermedius). Se trata de un tipo de perro bastante común en el resto de la Península Ibérica. La conservación completa de un Fémur ha permitido estimar la altura en la cruz del animal. Utilizando los factores de KOUDELKA (1885) y HARCOURT (1974) se obtienen Mandíbula Vértebras 7 Húmero 4 Radio 2 Ulna 2 Fémur 2 3 2 Total 24 NMI 6 Adultos 5 Juveniles Tabla 12.- Distribución de los restos de Fe/is catus de Santimamiñe. La aplicación de este criterio a Santimamiñe nos pone en evidencia diferencias biométricas significativas entre la mayoría de los restos del nivel 1 y los correspondientes al resto de los niveles posteriores. La Figura 7 elaborada a PEDRO M. CASTAfilOS 252 e Driesch Altuna e j 33 34,5 40 35,5 j 39 38,5 36,5 36 36,5 Driesch Lauk Altuna 1 ~,5 1 1 1 1 1 1 26 28 34 32 30 36 38 28 L M3 32 36 34 j 25' 27 25,3 27 29,3 25,5 24 22 e e 26 Driesch Lauk Altuna 30 28 32 40 42 50 52 54 41 40,5 44 48 46 Húmero Ad e j 32 30 Lauk Altuna 34 36 37,5 33 33 32 Driesch Lauk Altuna 28 j 45 42 44,5 43 42 40 Escápula LmC 34 36 40 38 e j 82,5 76 85,5 100,5 90 88 86 Tibia Ad 92 Astrágalo LMI 36 96 94 98 Calcáneo LM Driesch Lauk Altuna 34 38 L M3 Driesch Lauk Altuna 20 30 38 40 42 44 46 48 50 Figura 6.- Histograma de frecuencias de los valores de distintas medidas de Sus de Santimamiñe. 52 e j 42 42,5 42 44 43 44,5 100 ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMlr\IE (VIZCAYA) 9 o o 8 o 7 • • • •• o • 8 o o 8 o • ••• 7 17 18 19 Figura 7.- Diagrama de dispersión de las piezas dentarias inferiores de Felis de Santimamiñe. 20 21 22 253 254 PEDRO M. CASTAf\JOS partir de restos mandibulares que son los más numerosos, corrobora lo dicho. Excepción hecha de un ejemplar fragmentado del nivel V que presenta valores muy bajos, la aparición de dos conjuntos netamente diferenciados es clara. No parece razonable justificar estas diferencias de tamaño como resultado de dimorfismo sexual. En primer lugar porque son mucho mayores que las que se presentan en una población de gato doméstico entre machos y hembras. Los trabajos citados de KRATOCHVIL son muy elocuentes al respecto. Y su fiabilidad es grande ya que se han utilizado varias razas que presentan diferencias de tamaño entre sí. Además, resultaría un tanto casual que todos los ejemplares femeninos se hubieran concentrado en el nivel 1 y los masculinos en los niveles posteriores. Sin negar rotundamente esta posibilidad, hay que reconocer que con nueve ejemplares es bastante improbable. En resumen, podemos resaltar varios hechos: -existe un hiato métrico entre los valores del nivel 1 y posterior~s. -las medidas del nivel 1 entran dentro de las atribüídas al gato doméstico (KRATOCHVIL, o.e.). -las medidas de los niveles 11 a VIII corresponden al dominio de variación del gato montés. -apenas hay medidas en la zona de solapamiento. La explicación de esta distribución claramente bimodal puede deberse a alguna de las causas que a continuación se especifican. 1. -Se puede pensar que estamos ante individuos salvajes subadultos o inmaduros que utilizan la cueva de guarida cuando ésta es abandonada por el hombre. Pero en el esqueleto postcraneal predominan los huesos largos con ambas epífisis perfectamente soldadas, lo cual parece invalidar este primer supuesto. 2. - Pudiera también tratarse de un grupo de individuos salvajes en los que el tamaño haya sufrido una disminución a partir del Neolítico. Este fenómeno se ha comprobado en yacimientos europeos pero la disminución no es tan grande como la aquí presente. CLOT (1980) parece observar esta disminución en la relación entre la longitud de la carnicera inferior y la longitud P3-M1 entre un amplio conjunto de gatos monteses neolíticos daneses (DAGERBÓL, 1933) y ejemplares actuales (KURTEN, 1965). Pero los valores de estos últimos son siempre muy superiores a los del nivel 1 de Santimamiñe. Una disminución tan drástica de tamaño como la presente aquí ha sido precisamente usado como criterio de discriminación entre la forma doméstica y salvaje. 3. - Por tanto, si se trata de gato doméstico, se plantea una doble alternativa: ~Que este nivel 1 se halle mezclado con materiales muy posteriores y por tanto que se trate de ejemplares modernos. La posibilidad no puede rechazarse totalmente. No obstante, la coloración de la mayoría de los huesos de gato es similar a las de otras especies. Por otro lado, en la cueva de Lumentxa, relativamente próxima a Santimamiñe y que estamos estudiando, este hecho se repite y de forma más acentuada. Luego, parece un fenómeno más propio de la zona que de alguna circunstancia particular del yacimiento. -Parece más probable pensar que se trata de restos de gato doméstico presentes ya en el nivel l. Si el nivel 1 corresponde al Vascorromano, tal como la mayoría de los autores aceptan, se trataría de la primera o una de las primeras pruebas de la presencia de esta especie doméstica en el País Vasco en época tan temprana. También puede suponerse que este nivel 1 incluye materiales posteriores en el tiempo, lo cual implicaría aceptar algún tipo de ocupación, al menos esporádica en épocas medievales. No hay por ahora ninguna prueba arqueológica que apoye este supuesto, pero no puede tampoco descartarse. Aplicando los criterios de discriminación anteriormente citados, podemos atribuir a la forma doméstica 24 restos pertenecientes todos al nivel l. Parecen corresponder a un mínimo de 6 individuos distintos: 5 adultos y un juvenil. Su distribución según las distintas partes del esqueleto es la que se resumen en la Tabla 12. La Tabla 34 ofrece las medidas aisladas de esta especie. Hay tan pocas medidas de gato doméstico publicadas en el Norte de la Península, que resulta imposible cualquier comparación fiable. La cita con datación más segura en el País Vasco es la publicada por ALTUNA (1981b) entre los restos medievales del castillo de Aitzorrotz ( Guipúzcoa). Nosotros la hemos identificado .eh los niveles más superficiales de la cueva de Lumentxa ya citada y aún inédita. CABALLO Equus ferus Los 351 restos de équido pertenecen a un mínimo de 32 individuos distintos. La distribución de los restos según las distintas partes del esqueleto sé ofrece en la Tabla 13. El conjunto de medidas obtenidas a partir de esta muestra y sus resúmenes estadísticos se reflejan en las Tablas 35 y 54 respectivamente. La mayoría de los restos corresponden a niveles paleolíticos. Los pocos fragmentos de niveles más modernos no presentan diferencias morfológicas ni métricas respecto del resto. Parece razonable aceptar que se trata de un mismo tipo de caballo. Ya en el estudio de la fauna de Bolinkoba estudiada por nosotros (1983) se ofrecía un cuadro comparativo (Tabla 35) de las medidas dentarias y de las falanges de varios yacimientos peninsulares. Los restos de Santimamiñe coinciden en líneas generales con la mayoría de los allí citados. En algún caso presentan ciertas diferencias con Bolinkoba, yacimiento que paradójicamente se halla más cercano geográficamente. La morfología dentaria de los ejemplares de Santimamiñe es típicamente caballina. No aparecen caracteres arcaicos excepto en algunos huesos (5 sobre un total de 16) pertenecientes a la parte distal de metapodios. En ellos se observa una anchura en el límite entre la diáfisis y la epítasis mayor que en la propia superficie articular. Este carácter ha sido calificado por muchos autores como arcaico y asociado a la presencia de dedos laterales (GROMOVA, 1949). Esta afirmación ha sido puesta en duda por autores más recientes (SONDAAR P., 1968; FORSTEN A.M., 1975 y EISENMANN V., 1979). Acerca de la posición sistem~tica del caballo de Santimamiñe dentro del Paleolítico de Europa conviene hacer alguna observación. En primer lugar, la mayoría de los autores aceptan la existencia de dos formas de caballo: una de más talla y la otra de menor alzada que posee caracteres dentarios más evolucionados aunque con similar 255 ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMIÑE (VIZCAYA) robustez. Hay ciertas discrepancias respecto a su denominación taxonómica. Por ser más próxima a nosotros, mantenemos la nomenclatura de PRAT F. (1968) que identifica estas dos formas como E. c. germanicus y E. c. ga//icus respectivamente. El único criterio sólido se basa en los valores métricos. Si se comparan las medidas de Santimamiñe con las dadas por el citado autor, se observa cierta complejidad. En unos casos los valores coinciden con los atribuidos a E. c. germanicus. Otros, corresponden más al E. c. gal/icus. Incluso hay varias medidas que se sitúan en una posición claramente intermedia. Es difícil interpretar este hecho ya que inciden varios factores en el problema. Por un lado, el solapamiento que existe entre las medidas de ambas formas, puesto de manifiesto en muchos yacimientos. Por otro, la escasez de restos de esta especie en yacimientos peninsulares. La altura en la cruz calculada a partir del Radio y del Metatarso y utilizando los factores de KIESEWALTER (1888) tampoco ayuda mucho a esclarecer la cuestión. El Radio proporciona una alzada de. 147,9 cm. que parece situarse en los dominios del E. c. germanicus. Pero la estimación a partir del Metatarso es de 140 cm., claramente inferior y más cercana al E. c. gallicus, aunque también queda en la zona de solapamiento entre ambas. 11 111 IV V En resumen, la biometría no permite asignar con seguridad este conjunto a ninguna de las formas de caballo generalmente admitidas en esta época en toda Europa. Este problema es bastante frecuente en yacimientos peninsulares, acentuado además por la escasez de material. Sería muy conveniente un estudio sistemático de todos los materiales de caballo paleolítico obtenidos hasta el presente para ver si se confirma la presencia de ambas formas en la Península Ibérica. Actualmente hemos iniciado una revisión sistemática del material de caballo en la cornisa cantábrica fijándonos en la morfología y biometría dentaria. CABRA MONTES Capra pyrenaica Esta especie con un total de 378 restos está presente a lo largo de toda la ocupación del yacimiento aunque presenta una disminución en los niveles más superficiales. La distribución de los restos según las distintas partes del esqueleto se resume en la Tabla 14. La orografía que rodea a Santimamiñe justifica la presencia constante de esta especie aunque no en proporciones altas. El dominio del bosque y matorral de los aledaños es más favorable a especies como el ciervo, corzo o jabalí. Pero la proximidad de cimas que sobrepasan los 700 metros de altitud ha permitido a la cabra conservarse durante VII VI VIII Maxilar 4 1 Mandíbula 5 6 3 72 44 20 D. aislados 11 12 6 18 I' 11' 111' IV' V' Total 5 16 7 192 Vértebras 1 1 3 Costillas 2 2 5 1 2 3 Escápula 1 Húmero Radio 1 Carpo 2 11 7 2 2 Metacarpo 4 5 2 17 3 Pelvis 1 Fémur 5 3 5 Tibia 6 9 Rótula 1 Calcáneo 2 2 4 Astrágalo 4 Resto tarso 2 4 Metatarso 2 2 Metap. i. 14 8 Falanges 12 6 7 11 37 2 17 Sesamoideos TOTAL 4 16 16 9 21 136 77 56 12 NMI 3 2 2 2 7 6 3 2 32 Adultos 3 2 2 4 3 3 2 25 3 3 Juveniles Tabla 13.- Distribución de los restos de Equus ferus de Santimamiñe. 4 351 7 256 PEDRO M. CASTAl\IOS 11 111 IV V VI VI VIII s I' 11' V' IV' 111' Total Clavija c. 6 8 Cráneo 1 2 Maxilar 2 Mandíbula 6 D. aislados Vértebras 11 2 4 3 5 2 27 6 11 2 98 7 37 6 10 9 10 5 12 2 5 3 2 3 2 3 2 Costillas 5 Escápula 5 Húmero 5 4 2 Radio 7 3 5 3 2 2 31 12 2 1 10 3 27 9 2 Ulna 2 Carpo 5 2 Metacarpo 4 4 1 Pelvis 1 2 1 Fémur 3 4 2 2 4 1 3 2 Tibia 3 11 3 2 10 9 12 4 12 Rótula Calcáneo Astrágalo 2 2 13 Resto tarso 2 2 3 4 17 8 1 4 17 122 28 56 2 5 2 6 2 3 2 1 Metatarso Metap. ind. 7 1 TOTALES 14 3 Juveniles 2 4 3 11 17 29 25 378 4 3 32 4 20 4 4 39 4 2 2 2 2 3 26 2 2 2 2 Falanges NMI Adultos 6 7 7 6 42 2 13 Tabla 14.- Distribución de los restos de Capra pyrenaica de Santimamiñe. 1a V y S m el largo intervalo de ocupación de la cueva. En las Tablas 36 y 55 se ofrecen las medidas aisladas y los resúmenes estadísticos obtenidos a partir de esta muestra. Los valores de Santimamiñe son algo menores que los de algunos yacimientos cantábricos tales como los guipuzcoanos y Rascaño (ALTUNA, 1972 y 1980). Por otro lado coinciden con los de Bolinkoba (CASTAÑOS, 1983a) y Ekain (ALTUNA, 1984). Como ya comentábamos en el estudio de los materiales de Bolinkoba estas diferencias métricas pueden ser debidas a un predominio de hembras ya que esta especie presenta dimorfismo sexual. Para valorar la proporción de hembras y machos presentes en la muestra hemos aplicado los criterios dados por ALTUNA (1978) a cada pieza anatómica que presentara dimorfismo. El resultado, agrupando algunos niveles es el siguiente: h VI a VIII m h Clavija córnea 3 Escápula 3 Húmero 2 6 I' a V' m h 2 3 5 4 1 Radio Tibia Astrágalo 1 3 2 Calcáneo 1 Metacarpo 1 Metatarso 3 TOTALES 2 4 5 9 3 19 Como puede observarse, hay un ligero predominio en la ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMlr\IE (VIZCAYA) GRAN BOVIDO presencia de hembras que se manifiesta sobre todo en el nivel VI que es el más rico en material. Este dato podría explicar las diferencias métricas observadas respecto de algunos yacimientos citados, así como la coincidencia con Bolinkoba donde se produce un fenómeno similar. Bos primigenius + Bisan priscus . Los 141 restos de Gran Bóvido se reparten por casi todos los niveles de Santimamiñe. Faltan sólo en los niveles 1 y S que son los más superficiales de cada época de excavación. Representan un mínimo de 20 individuos distintos. 19 adultos y un juvenil. Llama la atención la escxasez de individuos inmaduros sobre todo si se compara con el bovino doméstico donde los juveniles sobrepasan el 30 % . Esta diferente estructura en la edad de la población pudiera interpretarse como argumento que prueba una intervención humana en el control y crianza de los animales. La Tabla 16 ofrece la distribución de los restos según las distintas partes del esqueleto. Las medidas aisladas y sus resúmenes estadísticos se reflejan en las Tablas 38 y 57 respectivamente. . En la muestra parecen estar presentes las dos especies. Siguiendo los criterios ya clásicos de diferenciación, hay piezas atribuibles al Bisonte y al Uro. La relación de piezas con atribución especifica es la siguiente: SARRIO Rupicapra rupicapra . El sarrio con 186 restos se reparte a lo largo de casi todos los niveles del yacimiento. En la Tabla 15 se resume la distrubución de estos restos según las distintas partes del esqueleto correspondientes a cada nivel. En esta especie se aprecia una progresiva disminución del número desde los niveles más antiguos a los más superficiales, disminución que prácticamente roza con la ausencia en los primeros niveles de cada campaña de la excavación. Las Tablas 37 y 56 reflejan las medidas aisladas y los resúmenes estadísticos respectivamente. Este conjunto métrico coincide fundamentalmente con los dados para otros yacimientos cantábricos (ALTUNA, 1972, 1976, 1981 Y 19084; CASTAÑOS, 1980, 1982 y 1983 a y b). 11 111 IV 257 Bos primigenius: Dos fragmentos de clavija córnea en los niveles VI y VIII V VI VII VIII 11' 111' IV' 4 4 Cráneo Maxilar 1 Mandíbula 2 D. aislados Total 6 3 Clavija c. 14 5 Vértebras 4 7 5 15 4 7 34 2 2 5 1 7 5 9 Costillas 5 Escápula Húmero 4 Radio Ulna Metacarpo Pelvis 2 Fémur 1 Tibia 2 2 2 Calcáneo 1 3 Metatarso 2 1 3 Metap. ind. 2 3 1 Resto tarso 2 1 2 6 4 5 7 13 19 26 NMI 2 2 3 Adultos 2 2 2 TOTALES 3 3 8 2 3 Astrágalo Falanges 10 6 2 8 4 10 57 1 10 3 11 8 6 2 2 28 6 186 23 30 5 3 3 23 4 2 2 18 Juveniles Tabla 15.- Distribución de los restos de Rupicapra rupicapra de Santimiñe. 3 2 5 PEDRO M. CASTAÑOS 258 Cuatro M3 del nivel VI Un Axis del nivel VII Tres fragmentos distales de Húmero del nivel VI Un fragmento distal de Metacarpo del nivel VI Dos astrágalos de los niveles VIII y IV' Un fragmento distal de Metatarso del nivel 11 cas en los niveles más superficiales. Este conjunto óseo representa un mínimo de 176 individuos distintos: 119 (un 67 % ) adultos y 58 (32 % ) juveniles. La distribución de los restos según las distintas partes del esqueleto se resume en la Tabla 17. La muestra ha proporcionado un amplio conjunto de medidas. Lá Tabla 39 refleja la totalidad de las mismas. La disminución de talla de esta especie a partir de la última glaciación es un hecho repetidamente observado por muchos autores. La riqueza de material de Santimamiñe constituye una ocasión óptima para corroborar dicha observación en Vizcaya. Para ello hemos elaborado los resúmenes estadísticos agrupando los datos biométricos en dos conjuntos. Por un lado todos los correspondientes a niveles que se han atribuído al Paleolítico (V a Vllly I' a V'). Por otro, los niveles con atribución postpaleolítica (1 a IV y S). El resultado se ofrece en la Tabla 58. En esta última se pone de manifiesto la disminución producida en casi la totalidad de las medidas, diferencia que es estadísticamente significativa en la mayoría de los casos. Las tres excepciones corresponden a medidas con muy escaso número de datos (nunca más de 5) que pudieran deberse a efectos aleatorios. Esta especie es muy frecuente en la mayoría de los yacimientos cantábricos y su elevada proporción sólo disminu- Bisan priscus: Un fragmento de clavija córnea del nivel VI Tres M3 de los niveles IV, VI y VII Un fragmento de Ulna del nivel Vi Un Carpa! 11-111 del nivel VI Un fragmento distal de Metacarpo del nivel VI Un fragmento distal de Tibia del nivel VI Un Astrágalo del nivel IV Un fragmento distal de Metatarso del nivel VI El Bisonte está ausente en los niveles 1a 111, es abundante en el nivel VI donde coexiste con el Uro. Este, sin embargo, parece pervivir hasta el nivel 11, aunque también es más éjbundante en los niveles VI a VIII. CIERVO Cervus e/aphus El ciervo con sus 4.041 restos es con mucho la especie mejor representada a lo largo de todas las épocas de ocupación. Sólo se ve superada por algunas especies domésti11 111 IV V VI Clavija c. 3 Maxilar 3 Mandíbula 5 VII 11' 111' IV' V' Total 5 4 7 1 14 3 3 2 Costillas 6 2 I' 1 Vértebras D. aislados VIII 5 7 4 2 38 9 2 8 Escápula 2 Húmero 2 Radio Ulna 1 Carpo 3 Metacarpo 2 3 1 Pelvis 7 4 Fémur 2 Tibia 4 6 2 7 7 Calcáneo 2 Astrágalo Resto tarso Metatarsa 2 2 4 2 Falanges 1 2 TOTAL 3 6 Adultos 5 2 Metap. i. NMI 2 6 2 2 4 57 10 17 1 5 4 3 4 2 27 2 2 3 2 2 3 Juveniles Tabla 16.- Distribución de los restos de Gran Bóvido de Santimamiñe. 18 7 5 1 2 141 20 19 ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMlr'JE (VIZCAYA) ye en asentamientos con un hábitat especializado de tipo roquedo por ejemplo, donde la cabra y el sarrio pasan a ser dominantes. 259 obstante, en el cercano yacimiento de Lumentxa hemos encontrado también varios restos de metapodios y falanges, algunos en buen estado de conservación. Las únicas medidas obtenidas son: RENO Falange 2: LM 31,5 Ap 16,9 Rangifer tarandus Los 7 restos de reno pertenecen a niveles wurmienses. Son indicativos una vez más de la presencia del reno en el Norte de la Península Ibérica. La distribución de los restos por niveles es la siguiente: AD Ad 12,6 15,2 CORZO Cuerna VIII IV' Total VI VII 3 2 ·6 3 2 7 Falange 2 Totales Como se ve predominan los fragmentos de cuerna. No 1 11 10 111 IV V VI Capreolus capreo/us Los 313 restos de corzo se distribuyen a lo largo de todos los niveles del yacimiento. Representan un mínimo de 44 individuos distintos: 37 adultos y 7 juveniles. La Tabla 18 ofrece la distribución de estos restos según las distintas partes del esqueleto. Las medidas aisladas y sus resúmenes estadísticos se ofrecen en las Tablas 40 y 59 respectivamente. VII VIII s 21 31 36 3 1 1 32 1 Cráneo 1 1 3 1 11 8 4 1 Maxilar 5 38 22 6 32 8 16 Cuerno 11" I' 111" 1 3 15 21 288 24 913 1 1 331 21 49 26 14 58 16 57 7 2 7 3 D. aislados Vértebras 198 66 50 212 73 95 9 4 4 7 6 49 35 76 20 71 4 37 30 1 1 1 1 6 6 10 8 12 Escápula 2 7 3 21 Radio 1 9 Ulna Carpo Metacarpo Pelvis Fémur Tibia Resto. tarso Sesamoideos NMI 3 8 6 40 11 15 7 4 6 23 10 8 13 3 1 4 6 3 5 2 2 28 8 33 3 15 1 160 5 109 1 44 11 6 10 31 2 9 52 5 12 3 4 5 5 3 4 5 28 3 10 6 3 9 1 3 17 5 1 1 8 8 12 7 4 15 8 1 1 3 1 1 25 4 2 12 29 31 6 24 2 5 25 56 50 16 7 6 4 1 7 3 2 3 1 1 85 130 83 1 2 75 1 1 1 89 1 2 12 1 2 7 11 5 2 7 24 6 7 35 1 2 27 6 26 10 4 28 15 16 9 2 14 70 159 85 42 137 21 58 53 3 5 4 427 8 4 1 785 509 232 935 3 213 474 179 97 1 1 1 3 3 1 3 1 1 35 14 Metap. ind. TOTALES 5 8 Metatarso Falanges 5 4 11 1 Rótula Calcáneo Astrágalo 3 66 21 42 10 19 6 4 52 1 142 35 139 118 Húmero 5 5 7 4 3 Total 4 38 Costillas V' 1 Mandíbula Hioides IV' 2 3 126 1 3 5 2 7 1 4 18 21 21 42 88 9 1 6 4 14 31 28 9 31 9 4 4 13 21 7 24 7 6 4 3 10 21 14 2 Adultos 1 3 2 3 6 Juveniles 2 4 18 7 2 7 2 7 2 1 2 1 3 Tabla 17.- Distribución de los restos de Cervus elaphus de Santimamiñe. 210 54 101 130. 660 28 4.041 177 119 58 260 PEDRO M. CASTAÑOS Los primeros datos acerca del Género Capreolus son posteriores al Villafranquiense sin que se sepa nada acerca de sus antecesores pliopleistocenos (HEINTZ E., 1970). La única especie presente es Capreo/us capreolus Linneo, 1758 con tres subespecies: C. c. capreolus Linneo 1758, de Europa y Asia occidental, C. c. pygargus Pallas 1971 de Asia Central y C. c. bedfordi Thomas 1908 del Extremo Oriente (ELLERMAN Y MORRISON-SCOTT, 1951). En el Pleistoceno de Europa se cita Capreolus susenbornensis durante el Mindel de Mosbach (KAHLKE H.D., 1960) en el que se aprecian variaciones morfológicas de los premolares inferiores (BOUCHUD J., 1972) respecto de otros ejemplares wurmienses europeos. En la evolución de esta subespecie varios autores han demostrado una progresiva disminución biométrica que se hace muy patente a partir del Neolítico (LEHMAN E.v., 1957; BOUCHUD J., 1972 ... ). BOESSNECK (1963) ha demostrado que los ejemplares neolíticos son mayores que los actual,es. Todo ello parece corroborar que tal disminución ·de la talla ha continuado hasta nuestros días. Las causas de este fenómeno parecen aún no estar aclaradas. Las citas en la Península Ibérica siempre han sido escasas y las medidas publicadas relativamente fragmentarias. La cornisa cantábrica ha proporcionado más datos que el resto. Los estudios recientes de materiales excavados en los últimos años han aportado elementos que sin ser abundantes pueden aceptarse como inicialmente significativos en vistas a esclarecer las pautas que esta especie ha seguido en la Península desde el Würm hasta épocas históricas. 11 Cuerno 2 111 IV 2 5 V VI Por esta razón, hemos intentado recoger la mayoría de los datos tanto publicados como inéditos, relativos a citas y medidas de corzo del Norte de la Península. Ofrecemos pues una relación de los yacimientos en los que se cita esta especie agrupándolos en dos conjuntos: todos los preneolíticos peninsulares por un lado y los neolíticos unidos a los postneolíticos de la cornisa cantábrica por otro. En algunos autores la cita no va acompañada de medida alguna. No se incluyen en este inventario aquellos yacimientos del primer tercio del siglo en los que se cita la especie sin ningún dato morfológico o biométrico complementario. A este respecto ALTUNA J. (1972) y MORALES A. (1976) ofrecen en sus respectivas tesis doctorales catálogos más exhaustivos y que no parece útil volver a repetir aquí. Yacimientos preneolíticos: Parpalló, Valencia (SOS BAYNAT, 1942) Cueva de Toll (RAFOLS, de VILLALTA y FUSTER, 1957) Cueva de Mollet (RIPOLL y LUMLEY, 1964-1965) Lezetxiki, Aitzbitarte, Urtiaga, Ermittia y Marizulo (ALTUNA, 1972) Cueva Morín (ALTUNA, 1973) Los Casares (ALTU NA, 1973) Tito Bustillo (ALTU NA, 1976) Cava Negra (PEREZ RIPOLL, 1977) La Paloma (CASTAl\JOS, 1980) La Riera (AL TUNA, 1981) Cueto de la Mina (CASTAl\JOS, 1982) VII VIII I' 11" 111" IV' Total 14 4 16 11 1 1 Mandíbula 16 20 11 6 2 D. aislados 9 33 14 11 Maxilar s 3 34 4 61 2 72 2 5 2 3 15 Hioides Escápula 1 4 Húmero 2 5 Radio 1 Ulna Carpo 2 2 5 4 13 3 Metacarpo 3 3 9 3 2 17 Pelvis Rótula 2 1 Calcáneo 2 Astrágalo 2 Resto tarso 3 7 7 5 5 3 2 Metap. ind. Falanges 1 7 14 13 TOTALES 6 2 1 36 3 4 2 44 5 313 51 110 70 9 18 NMI 6 12 10 4 2 3 44 Adultos 6 8 7 4 2 3 37 4 3 Juveniles 2 Tabla 18.- Distribución de los restos de Capreo/us capreolus de Santimamiñe. 7 ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMlr\IE (VIZCAYA) Atxeta (CASTAÑOS, 1984) Abrigo de La Peña-Navarra (CASTAÑOS, inédito) Yacimientos neo y postneolíticos (área cantábrica y atlántica): Berroberría (M. de LORIANA, 1940) Castro de Peñas de Oro (ALTUNA, 1965) Las Pajucas (ALTUNA, 1967) Los Husos (ALTUNA, 1972) Castro de Zambujal (DRIESCH, 1976) Sacaojos (BOESSNECK y DRIESCH; 1980) Aitzorrotz (ALTUNA, 1981) . Abauntz (ALTUNA, 1983) Santa Ana (de MIGUEL y MORALES, 1983) Cueva Mayor de Atapuerca (CASTAÑOS, inédito) La progresiva disminución de tamaño señalada por varios autores a partir de ejemplares europeos parece confirmarse en la Península Ibérica con alguna matización. Para analizar la variación biométrica, hemos querido agrupar los datos según las distintas épocas. En la Tabla 60 se ofrece un resumen estadístico de las medidas más frecuentes de corzos preneolíticos. Para su elaboración se han utilizado los yacimientos siguientes: Lezetxiki, Aitzbitarte, Urtiaga, Ermittia, Marizulo, Cueva Morín, Los Casares, Cava Negra, La Paloma, Cueto de la Mina, Atxeta y el Abrigo de La Peña, todos ellos indicados en la relación precedente. En la Tabla 41 se recogen las principales medidas de niveles neo y postneolíticos publicados en los últimos 15 años. Los yacimientos concretos se indican en la misma tabla. La Tabla 61 ofrece un resumen estadístico de algunas de las medidas más frecuentes. Se han agrupado ejemplares neolíticos y posteriores ya que el número de datos procedentes de niveles neolíticos es muy escaso y no presentan diferencias significativas con el resto. La disminución de tamaño es muy clara en casi todas las partes del esqueleto. Las falanges son una excepción al respecto. En la Fig. 8 se pone de manifiesto gráficamente la variación y medias de cada conjunto comparándolas entre sí y con las obtenidas en Santimamiñe. Con los datos precedentes, parece posible afirmar que: -Hay diferencias significativas entre los ejemplares preneolíticos y los posteriores. - Esta disminución de tamaño parece haberse producido ya en el Neolítico y no a partir del mismo como parece observarse en Europa. - El proceso parece haberse realizado entre el final del Paleolítico y la llegada de la neolitización. A este respecto es interesante observar que los ejemplares de Santimamiñe en la mayoría de los casos ocupan una posición intermedia. Téngase en cuenta que casi todas las medidas de este yacimiento proceden del nivel IV asociado a industrias que unos autores califican de Asturiense y otros de Preneolítico. En cualquier caso situados en plena transición hacia el Neolítico. - Los ejemplares modernos son aún significativamente más pequeños. Esto nos induce a suponer que el proceso de reducción ha proseguido hasta nuestros días. - Las falanges no parecen haber seguido estas pautas y permanecen relativamente estables a diferencia del resto del esqueleto. JABALI Su ferus Los criterios de atribución de los restos al jabalí ya se 261 han expuesto al estudiar los restos de cerdo. Según lo dicho, pueden ser asignados a la forma salvaje 295 restos distribuídos a lo largo de todos los niveles. Su distribución según las distintas partes del esqueleto a las que pertenecen se ofrecen en la Tabla 19. Las medidas aisladas y sus correspondientes resúmenes estadísticos se reflejan en las Tablas 42 y 62 respectivamente. Cualquier comparación en esta especie queda relativizada por la problemática discriminación que se plantea con la especie doméstica. Habrá ejemplares de tamaño más reducido que han sido incluídos en la forma doméstica. Hecha esta observación se puede decir que los ejemplares de Santimamiñe coinciden básicamente con los de Guipúzcoa y con la mayoría de los restantes yacimientos tanto del Sur como del Centro de la Península Ibérica. LOBO Canis lupus El Género Canis aparece en la Península Ibérica ya en el Turoliense de Concud (CRUSAFONT, 1950), Los Mansuetos (ADROVER, 1963) citado como Canis cipio y en la Venta del Moro (MORALES y AGUIRRE, 1976) como Canis sp. La forma más extendida a lo largo del Pleistoceno inferior y medio es C. etruscus Forsyth Majar 1877, descrita en varios yacimientos europeos y en alguno peninsular como Bagur en Gerona (MARTI y VILLALTA, 1974). PRAT y THIBAULT (1976) crean ·1a forma C. etruscus mosbachensis en la que incluyen C. etruscus y C. mosbachensis denominación esta última de origen alemán y que parece sinónima de la anterior. BONIFAY (1971) describe la forma C. lupus lunil/ensis Bonifay de pequeño tamaño y que por aumento progresivo de la talla daría lugar a C. lupus wurmiense. CLOT (1980) recoge las medidas de los principales yacimientos franceses corroborando el aumento de tamaño del Mi ya señalado por M. F. BONIFAY (1971). Respecto al esqueleto postcraneal observa dos formas: una de tamaño mediano Y otra más robusta. Atribuye estas diferencias a dimorfismo sexual. El lobo se ha citado en varios yacimientos del Paleolítico peninsular aunque siempre su representación es escasa y las medidas obtenidas también. En los últimos años se han publicado algunos conjuntos métricos procedentes de varios yacimientos cantábricos: Lezetxiki, Aitzbitarte Urtiaga, Ermittia y Marizulo (ALTUNA, 1972), Cueva' Morín (ALTUNA, 1973), Sierra de Gibijo (ALTUNA 1974) Tito Bustillo (ALTUNA. 1976), La Paloma (CASTAÑOS Í980) Rascaño (ALTUNA, 1981) y Bolinkoba (CASTAÑOS: 1983). ALTUNA en los yacimientos guipuzcoanos parece corroborar la observación ya anotada por R. MARTIN ( 1968) de que la dentición presenta gran talla mientras que el. esqueleto postcraneal parece más bien pequeño o mediano. . P?r desgracia las medidas obtenidas de todos estos yac1m1entos son más bien escasas. A pesar de ello si se comparan las medidas de la carnicera inferior, se observa que los ejemplares de Vizcaya son netamente inferiores al resto, en especial a los guipuzcoanos. 262 PEDRO M. CASTAl\IOS s 34 13 19 33 33 PN 32 18 PN 31 12 s PN 28 s N 17 4 32 26 32 s 5 31 31 16 30 30 29 15 29 28 14 28 27 13 27 26 12 26 19 30 3 N 5 L M3 L PTP4 PN = N 2 ¡ 29 N Santimamiñe 26 N Húmero Ad Neo y postneolítico 35 33 67 32 16 PN 18 s 34 12 PN 29 j 27 PN 30 6 28 Escápula IMP s Preneolítico PN 22 66 2 33 s 7 33 s 31 3 31 29 ¡ 27 25 23 Radio Ap 30 N 4 29 t 28 t 65 32 64 31 N 63 5 30 N 2 t 27 26 Tibia Ad Figura 8.- Gráfica con la variabilidad y medias de lagunas medidas de Capreolus capreolus peninsulares de diversas épocas, comparadas con las de Santimamiñe. 62 29 N 61 2 60 + Calcáneo LM 28 27 Astrágalo LMI 263 ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMIÑE (VIZCAYA) 111 11 IV V Cráneo Maxilar 3 2 2 2 Mandíbula D. aislados VI VII VIII s I' 111' IV' V' Total 1 6 16 5 12 1 2 34 4 11' 1 6 8 2 5 63 Vértebras 1 2 Costillas 2 Escápula 2 3 8 Húmero 4 13 3 Radio 2 Metacarpo 4 Pelvis 2 6 Fémur 6 Tibia 2 1 Calcáneo 2 6 Astrágalo 7 16 6 11 2 2 4 3 2 Metatarso 1 1 Metap. ind. 2 2 Falanges 3 17 16 11 TOTALES 5 37 51 26 3 2 4 3 2 4 NMI Adultos 1· 3 3 8 22 4 4 2 4 2 23 4 Juveniles 86 18 2 44 2 2 1 6 5 5 2 2 20 17 8 8 3 3 2 2 84 2 295 2 2 Tabla 19.- Distribución de los restos de Sus ferus según las distintas partes del esqueleto en los distintos niveles de Santimamiñe. Lezetxiki L A Urtiaga 26,5 29,5 34 12,3 Ermitt 28,5 28,7 30 11,4 11,4 12,3 Sin embargo, las medidas postcraneales no presentan diferencias sensibles. Por tanto, no parece que se pueda generalizar una diferencia entre la robustez de la dentadura y la menor talla del cuerpo. Las diferencias pueden estar influídas por el azar al tratarse-~de tan pocos datos y al existir también cierto dimorfismo sexual tal como señala CLOT (1980). Será necesario replantear la cuestión cuando se posean conjuntos más amplios. Un dato que parece más claro es el menor tamaño del esqueleto postcraneal de los ejemplares peninsulares respecto a sus congéneres del Centro y Norte de Europa. Las medidas ofrecidas por MORALES (1976) procedentes de individuos daneses modernos son netamente superiores, no sólo a lobos peninsulares actuales, sino también a ejemplares wurmienses. Por último, la distinción entre la forma C. /. signatus y C. l. deitanus (CABRERA, 1914) que parece observarse en tiempos recientes, no hay forma de demostrar si tiene origen pre o protohistórico. Los datos procedentes del S.E. (área de distribución del C. l. deitanus) son más escasos y no hay posibilidad de comparación alguna. Mar 30,4 T.B. 30 12 22,2 8,4 Bolinkoba 22,5 30,2 28, 1 27,6 9,1 11,6 10,6 10,6 Gibijo 26,8 28 11,7 11,6 La Tabla 20 ofrece la distribución de los 30 restos de lobo según las distintas partes del esqueleto y por cada nivel. Las medidas aisladas de esta especie son objeto de la Tabla 43. ZORRO Vu/pes vulpes Los 57 restos de zorro se distribuyen a lo largo de casi todos los niveles del yacimiento. Parecen representar un mínimo de 9 individuos distintos: 7 adultos y 2 juveniles. Su distribución según las. distintas partes del esqueleto se ofrece en la Tabla 21. La Tabla 45 recoge las medidas aisladas de esta especie. Las medidas de Santimamiñe corresponden al zorro común (Vulpes vulpes). Ningún resto presenta valores o caracteres morfológicos de zorro polar (A/opex /agopus). Este dato observado en yacimientos guipuzcoanos (AL TUNA, 1972) y en Bolinkoba (CASTAÑOS, 1983) parece confirmarse de nuevo en Santimamiñe. Las pocas medidas obtenidas parecen apuntar hacia una disminución de tamaño de los ejemplares postwurmienses PEDRO M. CASTAl\JOS 264 respecto de los wurmienses. Este dato señalado por muchos autores se confirmaba ya en el material mucho más abundante de Bolinkoba. Tabla 23 resume la distribución de estos restos según las distintas partes del esqueleto. Las medidas obtenidas se reflejan en la Tabla 45 bis. Las falanges terceras indican por su morfología que se trata del Ursus arctos. El índice de robustez del Metatarsiano 3 presenta igualmente valores propios de la forma arctoide. El resto del material está constituído en su mayor parte por piezas dentarias aisladas. Las medidas y morfología del Mi y del M3 denotan de nuevo su pertenencia al oso pardo. Por este motivo, hemos atribuído toda la muestra a la forma Ursus arctos. OSO PARDO Ursus arctos Los 28 restos de úrsido se distribuyen a lo largo de la mayoría de los niveles de ocupación de Samtimamiñe. Sólo faltan en los niveles 1, 11 y S que son los más superficiales de cada una de las dos épocas de excavación. La 111 11 Mandíbula 1 D. aislados 2 IV VI s VIII 11 111 IV V Total 1 Vértebras 1 1 6 2 8 2 10 2 Costillas Escápula Ulna 1 Metacarpo 2 Pelvis Metatarso 1 Falanges 2 TOTALES 10 3 3 2 2 5 3 NMI 2 30 1 10 10 Adultos Tabla 20.- Distribución de los restos de Canis lupus de Santimamiñe. 11 IV V VI 1 2 6 VII VIII Mandíbula 2 D. aislados Vértebras Total 1 Cráneo 2 2 2 2 10 9 13 12 Costillas 2 Húmero Radio Ulna 1 Pelvis 2 Fémur 1 Tibia 2 Peroné Astrágalo 1 2 Metatarso 6 2 Metap. ind. 2 Falanges TOTALES NMI 1 18 6 2 3 2 4 Adultos Juveniles Tabla 21.- Distribución de los restos de Vu/pes vulpes de Santimamiñe. 11 8 2 7 57 9 7 2 ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMIÑE (VIZCAYA) 11 IV 111 VI V VIII 265 I' Total 4 Mandíbula D. aislados 1 1 Vértebras 5 7 Escápula 1 Húmero 2 2 4 Radio 2 Ulna 3 Metacarpo 3 Tibia 1 2 Metatarso 3 3 2 Metap. ind. TOTALES 9 4 2 12 4 33 2 NMI 9 Adultos 8 Juveniles Tabla 22.- Distribución de los restos de Fe/is sílvestris de Santimamiñe. ARMll\JO Mustela erminea Esta especie ya citada por los excavadores sólo está representada por un resto perteneciente al nivel VII. Se trata de un Fémur izquierdo cuya longitud máxima es de 36,3 mm. MARTA Martes sp. Los 32 restos de este mustélido se sitúan sobre todo en los niveles postpaleolíticos. Parecen representar un mínimo de 8 individuos distintos: 6 adultos y dos juveniles. La Tabla 24 ofrece la distribución de los mismos según las distintas partes del esqueleto. Las pocas medidas obtenidas se resumen en la Tabla 44. Martes martes y Martes taina son especies más bien escasas en los yacimientos peninsulares. Hasta el momento casi todas las citas proceden de niveles pre y protohistóricos del Norte de la Península. HARLE (1909) la cita en Furninha (Portugal) y BATALLER (1918) en la cueva de Serinya (Gerona). En ambos casos se atribuyen los restos a la foina. Nosotros hemos hallado M. taina en Aldehuela (Teruel) que pueden proceder de algún ejemplar moderno por el estado de conservación de los restos. ALTUNA (1972) señala 7 restos en los tres primeros niveles de Marizulo (Guipúzcoa) que parecen pertenecer a la 111 IV Cráneo 1 D. aislado 3 V marta y restos de tr,es individuos (1974) en la Sierra de Gibijo (Alava) identificados como de marta también. El mismo autor cita también la Marta en Abauntz (Navarra), 1983). Nosotros hemos encontrado restos en el Abrigo de la Peña (Marañón-Navarra), en la Cueva Mayor de Atapuerca (Burgos) (ambos inéditos), sin que hayamos podido especificar por tratarse de escasos restos postcraneales. En Vizcaya ha sido identificado Martes sp, en Aldeacueva y Las Pajucas (ALTUNA, 1967). Martes foina ha sido determinada por nosotros en la cueva de Urratxa 111 en niveles revueltos del Paleolítico y Bronce. Los primeros excavadores de Santimamiñe señalaron la presencia de marta en los niveles 1 a 111 y de foina en el IV. Si atendemos a la distancia entre los foramina mentoniana resulta que de las cinco mandíbulas conservadas, cuatro corresponden a marta y una a foina, ésta efectivamente en el nivel IV. Las otras dos piezas con posibíiidad de especificación son dos maxilares en los que se conserva el P4 • Ambos presentan caracteres atribuíbles a la marta y pertenecen a los niveles 11 y IV respectivamente. El esqueleto postcraneal no proporciona elemento alguno de diagnosis. En resumen, la mayoría de los restos de Santimamiñe parecen corresponder a Martes martes. También aparece algún resto atribuíble a Martes taina. Esto implica la coexistencia de ambas especies al menos en el nivel IV. VII 11' 111' IV' Total 8 6 19 Carpo 1 Metatarso 2 Falanges TOTAL 2 2 2 8 2 Tabla 23.- Distribución de los restos de Ursus sp. de Santimamiñe. 5 8 6 28 PEDRO M. CASTAÑOS 266 NUTRIA IV 11 VI s Total 1 Cráneo Maxilar 1 Mandíbula 3 1 5 .1 1 D. aislados Vértebras 5 6 Húmero 3 4 Radio 2 3 Ulna 1 Metacarpo Fémur Tibia 2 2 3 2 1 Peroné Metatarso TOTAL 3 2 2 20 6 32 3 NMI 3 2 8 Adultos 1 2 6 Juveniles 2 2 Lutra lutra El único resto presente de esta especie es una carnicera inferior cuyas medidas son: Long. 13 Anch. 6, 1 Los restos de esta especie son muy escasos en la Península Ibérica y todas son postpaleolíticas. La de Santimamiñe procede del nivel 11. La relación somera de todas las citas de nutria peninsulares es la siguiente: - Marizulo (Guipúzcoa) (ALTUNA, 1972) en niveles que van desde el Mesolítico al Bronce. -Cerro de la Virgen (DRIESCH v.d., 1972) en nivel Campaniforme. -Cerro de la Encina (LAUK, 1976) en el Argar. - Cueva Mayor de Atapuerca (inédito) en Bronce Medio (determinado por nosotros). Existe una cita en el Asturiense de Cueto de la Mina sin especificación ni medida alguna. La carnicera de Santimamiñe tanto por sus medidas como por su morfología coincide con la de Atapuerca. GATO MONTES Fe/is silvestris Tabla 24.- Distribución de los restos de Martes sp. de Santimamiñe. 11 111 Los 33 restos de gato montés se reparten a lo largo de casi todos los niveles de ocupación del yacimiento. Repre- IV V VI s Total 1 Mandíbula D. aislados 2 Húmero 2 2 5 3 Radio 1 Ulna 2 Metacarpo 2 2 Calcáneo Metatarso 1 TOTAL 5 2 6 3 18 Tabla 25.- Distribución de los restos de Me/es me/es de Santimamiñe. TEJON Me/es me/es Los 18 restos de Tejón se distribuyen por los niveles 1 a VI. Su distribución según las distintas partes del esqueleto se refleja en la Tabla 25. Las medidas obtenidas se ofrecen en la Tabla 46. Estas medidas coinciden básicamente con las dadas por ALTUNA (1971) para ejemplares subfósiles de Guipúzcoa. También entran dentro de las distribuciones publicadas por ZABALA J. (1980) para ejemplares actuales de la Sierra de Aralar. Por último, son semejantes a las obtenidas por nosotros en el yacimiento vizcaíno de Atxeta (1983). Esta especie presenta una gran variabilidad. Sus costumbres fosoras inciden a veces en la determinación de su antigüedad. sentan un mínimo de 9 individuos distintos: 8 adultos y un juvenil. Su distribución según las distintas partes del esqueleto se refleja en la Tabla 22. En el nivel 1 coexiste con la forma doméstica. Respecto a la problemática acerca de la discriminación entre ambas formas, ya ha sido tratada al hablar del gato doméstico. Las medidas obtenidas a partir de esta muestra se ofrecen en la Tabla 47. Estos valores son inferiores a los de Urtiaga y Lezetxiki publicados por AL TUNA (1972) que se caracterizan por su robustez. Coinciden fundamentalmente con los del Abrigo de La Peña (Navarra) estudiados por nosotros. Entran dentro del dominio de variación de los gatos monteses actuales que son más pequeños que los prehistóricos. ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMIÑE (VIZCAYA) 267 cos. Esta variedad del lince se cita por vez primera en el yacimiento. LINCE NORDICO Linx linx Los 8 restos de lince parecen corresponder a tres individuos adultos distintos. La Tabla 27 ofrece la distribución de los mismos según las distintas partes del esqueleto. La Tabla 48 resume las medidas aisladas obtenidas. La mayoría de los restos de lince hallados en la península Ibérica proceden de la forma Linx pardina, Temminck. Hace menos de un lustro que aparece por vez primera el lince boreal Linx linx, L. en la sima de Pagolusieta en Vizcaya (ALTUNA, 1980) y en Rascaño (ALTUNA, 1981). Su repetida presencia en la vertiente norte de los Pirineos hacía bastante probable su aparición a este lado de la cordillera. Todos los restos de Santimamiñe, tanto por sus medidas como por algunos caracteres morfológicos, parecen corresponder al Linx linx. La carnicera inferior presenta sobre el cíngulo basal un dentículo típico de esta especie y que rara vez aparece en la especie ibérica. Las medidas coinciden. con las del ejemplar de Pagolusieta y son muy superiores a las obtenidas en Bolinkoba. Por tanto, Santimamiñe confirma la presencia del lince nórdico que convive con el lince ibérico presente en otros yacimientos vasVI IV LEON Panthera leo En el nivel IV aparece una Falange primera de un gran félido cuyas medidas son las siguientes: Falange 1: LM 47 Ap 18, 1 AD 11,9 Estas medidas son superiores a las de Panthera pardus e inferiores a las de Panthera spelaea. Parecen coincidir con las dadas por ALTUNA (1972) para el león le Urtiaga. No abstante, hay que recordar que existe un gran solapamiento entre la pantera y el león de Urtiaga en cuanto a las falanges. La de Santimamiñe supera a las dos que PEREZ RIPOLL (1977) atribuye a Panthera leo en el Musteriense de Cava Negra. Sin embargo, los valores más altos de la Panthera pardus de Bolinkoba son similares a esta de Santimamiñe. Provisionalmente atribuíamos la pieza a Panthera leo. Su aparición en el nivel IV corrobora una vez más la Vil 11 111 Total 1 Vértebras Escápula 1 Húmero 2 Radio 2 Ulna 1 Metacarpo 2 Pelvis 2 2 Fémur Tibia Metatarso 1 2 2 3 2 Calcáneo 1 Falanges TOTALES 4 NMI 1 4 9 Adultos 1 4 3 23 2 8 2 7 Juveniles Tabla 26.- Distribución de los restos de Lepus sp. de Santimamiñe. 111 IV VI Total pervivencia tardía de grandes félidos en la Península Ibérica. Mandíbula LIBRE D. aislados Metacarpo 3 4 4 8 Metatarso Falanges TOTALES 1 2 2 1 NMI 3 Adultos 3 Tabla 27.- Distribución de los restos de Linx linx de Santimamiñe. Lepus sp. Los, 23 restos le liebre corresponden a un mínimo de 8 individuos distintos: 7 adultos y un juvenil. Su distribución según las distintas partes del esqueleto se resumen en la Tabla 26. La Tabla 49 ofrece las medidas aisladas procedentes de esta especie. . Estas medidas coinciden fundamentalmente con las publicadas por ALTUNA ( 1972) para ejemplares de la provincia de Guipúzcoa y con las obtenidas por nosotros en Bolinkoba (1983). Son claramente supefiores a las publicadas por varios autores para yacimientos mericlTonáies de la Península Ibérica. No hay restos craneales que permitan reconocer I~ p~esencia de Lepus timidus. Por todo ello, podemos atribuir estos restos a Lepus capensis europaeus. 268 PEDRO M. CASTAl'ilOS BIBLIOGRAFIA Altuna J. 1967: Cuevas sepulcrales de Vizcaya. Estudio paleontológico de los mamíferos hallados en los estratos arqueológicos. Munibe, 19. 1971: Los Mamíferos del yacimiento prehistórico de Morín (Santander). Publicac. Patronato Cuevas Prehistóricas. Santander, 6, 367-399. 1972: Fauna de Mamíferos de los yacimientos prehistóricos de Guipúzcoa, Munibe, 24, 1-464. 1973: Fauna de Mamíferos del yacimiento prehistórico de Los Casares (Guadalajara). Exc. Arq. en España, n.º 76, 97-116. 1974: Hallazgo de un Uro (Bos primigenius Boj.) en la Sierra de Gibijo (Alava), estudio de su esqueleto y de la fauna asociada al mismo, Munibe, 26. 1976: Los Mamíferos del yacimiento prehistórico de Tito Bustillo (Asturias). En: Maure Romanillo y Cano Herrera. Excavaciones en la Cueva de Tito Bustillo (Asturias). Trabajos de 1975. lnst. de Est. Asturianos, 149-154. 1978: Dimorfisme sexuel dans le squelette postcephalique de Capra pyrenaica pendant le Würm Final, Munibe, 30, 201-214. 1980: Historia de la domesticación animal en el País Vasco desde sus orígenes hasta la Romanización, Munibe, 32. 1981: Alimentación de origen animal de los habitantes del Castillo de Aitzorrotz ( Escoriaza, Guipúzcoa), Munibe, 33. 1984: El yacimiento prehistórico de la Cueva de Ekain (Deba, Guipúzcoa), Soc. de Estudios Vascos. Aranzadi T., Barandiarán J.M. y Eguren E. 1925: Exploraciones de la Caverna de Santimamiñe (Basondo-Cortézubi): 1. - Figuras rupestres. Bilbao. 1931: Exploraciones de la Caverna de Santimamiñe ( Basondo-Cortézubi): 2. - Los niveles con cerámica y el conchero. Bilbao. 1935: Exploraciones de la Caverna de Santimamiñe ( Basondo-Cortézubi): 3. - Exploraciones en la Caverna de Lumentxa (Lequeitio). Bilbao. Apellániz J.M. 1975: El Grupo de Santimamiñe durante la Prehistoria con cerámica, Munibe, 27. Barandiarán l. 1967: El Paleomesolítico del Pirineo occidental. Bases para una sistematización tipológica del instrumental óseo paleolítico. Zaragoza. Barandiarán J.M. 1953: El hombre prehistórico vasco. Buenos Aires. 1962: Santimamiñe. Excav. Arq. en España, 7. Bataller J.R. 1918: Mamifers fóssils de Catalunya. Trébals de la lnstitució Catalana d'Hist. Nat., 111-272. Bouchud J. 1972: Les grands herbivores rissiens des «Abimes de la Fage» en Correre, Nouv. Arch. Mus. Hist. nat., 10, Lyon. Boessneck J. 1963: In Boessneck, Jéquier, Stampfli. Seeber Burgaschisee Süd. Teil 3. Die tierreste. Acta Bernensia, 11. Bern. 1968: Vid. Driesch A.v.d. Bonifay M.F.: 1966: Les Carnivores.En Lavocat R. Faunes et Flores préhistoriques de l'Europe Occidentale. París. 1971: Carnivores quaternaires du Sud-Est de la France. Mem. Mus. Nat. Hist., P.aris, Ser. C, 21, 43-337. Brinkmann A. 1924: Canidenstudien 5-6. Bergens Mus. Aarbock 19231924. Naturvidensk. Raekke, 7, 1-57. Cabrera A. 1924: Fauna ibérica. Mamíferos. Madrid. Castaños P.M. 1980: La Macrofauna de la Cueva de La Paloma (Pleistoceno terminal de Asturias). En: Hoyos Gómez L. La Cueva de la Paloma, Excav. Arq. en España, n.º 116, 65-100. 1982: Los Macromamíferos de Cueto de la Mina (Asturias). Bol. del lnst. de Est. Asturianos. 1983a: Estudio de los Macromamíferos de la Cueva de Bolinkoba, Kobie, XIII. 1983b: Estudio de los Macromamíferos de la Cueva de Atxeta, Kobie, 13. Clot A. 1980: La grotte de la Garriere (Gerde, Htes. Purénées) Stratigraphie et Paléontologie des Carnivores, These 3 Cycle, Toulouse. Couturier M.A.J. 1962: Le Bouquetin des Alpes. Grenoble. Crusafont M. 1961: El cuaternario español y su fauna de mamíferos. Ensayo de síntesis. Speleon, 13: 181-199. Dagerbol M. 1933: Danmarks Pattedyr i fortiden, Copenhague. Delpech F. 1975: Les faunes du Paléolitique Supérieur dans le Sudouest de la France. Bordeaux. 3 tomos. Dubois A. y Stehling H.G. 1931-1933: La grotte de Contencher, station mousterienne. Mem. Soc. Paléontol. Suisse, 52, 1-178 y 53, 1979292. Driesch A.d. y Boessneck J. 1968: Die Fauna del «Cabezo Redondo» bei Villena (Alicante), Studien über frühe Tierknochenfunde van der lberischen Halbinsel, 1, München. 1972: Osteoarchaeologische Untersuchungen auf der lberischen Halbinsel. Studien über frühe Tierknochenfunde van der lberischen Halbinsel, 3, München. 1976: Das Vermessen vor Tierknochen aus vof und fruhgeschichtliche Siedlungen. Munchen. 1980: Tierknochenfunden aus vier Südspanischen Hohlen. Studien über frühe Tierknochenfunde van der lberischen Halbinsel, 7, München. Eisenmann V. 1979: Les métapodes d'Equus sensu lato (Mammalia, Perissodactyla). Geobios, 12 (6), 863-886. Fock J. 1966: Metrische Untersuchungen an Metapodien einiger europaischer Rinderrasen. Dissertaton Tierarztl, Fakultat der Unibersitat München. Forstén A.M. 1975: The fossil Horses of the Texas Gulf Coastal Plain: a revision. The Pearce-Sellards Ser. (22), 1-86. ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMIÑE (VIZCAYA) Harcourt R.A. 1974: The Dog in Prehistoric and Early Historie Ritain. Jornal of Arch. Science, 1, 151-175. Harlé E. 1909: Faune de la Grotte a Hyenes rayées de Furninha et d'autres grottes de Portugal. Bull. Soc. Céolog. de France, 4 ser., 9.85. Hein F.H. 1982: Kupferzaitliche Tierknochenfunde aus Valencina de la Concepción (Sevilla). Studien über frühe Tierknochenfunde van der lberischen Halbinsel, 8, München. Heintz E. 1970: Les cervidés Villafranchiens de France et d'Espagne. Mem. Mus. Hist. Nat. París. Kahlke H.D. 1960: Die Cervidenreste aus den Altpleisrozanen Sander van Mosbach. Abhand. der dent. Akad. wiss. zu Berlín. Berlín. Kiesewalter L. 1888: Skelettmessugen an Pferden als Beitrag zur theoretiscen Grundiage der Beurteilungslehere des Pferdes. Dissertation. Leipzig. Koby F.Ed. 1958: Ostéologie de la Chevre fossile des Pyrénées (Capra pyrenaica Schins). Eclogai geologicae Helvetiae, 51, 475-480. 1959: Les renards Magdalénien de la Vache. Bull. Soc. Préhistorique de I' Ariege, 14, 26-34. Kormos Th. 1916: Die Felsuische van Pilisszántó. Mitteilungen aus dem Jahrbucg der Kgl. Ungarischen geolog. Rechsanstalt, 23, 6. Koudelka F. 1885: Das Verhaltnis der ossa longa zur Skeletthode bei den Saugethieren. Verhandl. d. Naturforsch. Ver. Brunn., 24, 127-153. Kratochvil Z. 1974: Die Wertrilarionen von Schadelmerkmalspaaren als taxonomiche Kriterien Fe/is s. s!'lvestris und F.s.f. catus (Mammalia) Zoologicke Listy, 24, 13-29. 1976: Die Unterscheidung postkranialer merkamlspaare bei Fe/is s. si/vestris und F. lybica F. catus (Mammalia), Folia Zoologica, 26. Kurtén B. 1965: On the evolution of the european wildcat (Fe/is si/bestris Schreber), Acta Zoologica Fenica 111, 1-26. 1966: A Late-Glacial find of Artic Fox (Alopex /agopus L.) from southwestern Finland. Comentationes Biologicae, 29, 1-7. Lauk H.D. 1976: Tierknochenfunde aus brozezeitkicen Siedlugen bei Monachil und Purullena (Provinz. Granada). Studien über frühe Tierknochenfunde von der lberischen Halbinsel), 6, München. Lehmann U. 1957: Kie Heterogeneitat des auropi:iischen Rehs Zeitschrift f. Jagdwiss., 3, 53-63. Loriana Marqués de 1940: Excavaciones arqueológicas realizadas en la gruta y covacho de Berroberría término de Urdax (Navarra) y sus inmediaciones, Atlantis, 15, 91-102. Mariezkurrena K. 1981: Vid. Altuna J. 1983: Contribución al conocimiento del desarrollo de la 269 dentición y el esqueleto postcraneal de Cervus alaphus, Munibe, 35, 149-202. Martín R. 1968: Les mammiferes fossiles du gisement de Villereversures (Ain). Utude des Carnivores, Cervidés et des Equidés. Documents des Laboratoires de Géologie de la Faculté des Sciences de Lyon, 27. Martín C.S. y Villalta J.F. 1974: Un yacimiento Mindeliense en las cercanías de Bagur provincia de Gerona, Acta Geol. Hisp., 9, 4-9. Marcos J.L. 1982: Carta arqueológica de Vizcaya. Yacimiento en cueva. Cuadernos de Arqueología de Deusto. Bilbao. 40-45. Miller A. 1976: Catalogue of the Mammals of Western Eurpe. Britisk Museum (Nat. Hist.) Londres. Matolcsi J. 1970: Historische Erforschung der Korpergrosse des Rindes auf Grund von ungarischen Knochenmaterial. Zeitschrift fur Tierzuchtung und Zuchtungsbiologie, 87, 2, 89-137. Hamburg und Berlín. Morales A. 1976: Contribución al estudio de las faunas mastozoológicas asociadas a yacimientos prehistóricos españoles. Tesis doctoral. Madrid. Morales J. y Aguirre E. 1976: Los Carnívoros (Carnívora, Mammalia) de Venta del Moro (Valencia). Trabajos sobre Neógeno-Cuaternario, 5. Musil R. 1974: Tiergesellschaft der Kniogrotte, in: Feustel RI, Die Kniegrotte, eine Magdalénien-station in Thuringen, Weimar, 39-95. Nobis G. 1971: Van Wildpferd zum Hauspferd. Fundamenta B., 6, Koln. Pérez Ripoll M. 1977: Los Mamíferos del yacimiento musteriense de Cova Negra (Játiva, Valencia). Servicio de Investigación Prehistórica, 53, Valencia. Prat F. 1968: Recherches su les Equidés pleistocénes en France. Bordeaux. Rafols J.S., Villalta J.F. y Fustá M. 1957: Exposición monográfica sobre la cueva de Toll (Moiá). Exposiciones en el Palacio de la Virreina. Barcelona. Ripoll E. y Lumley H. de 1964-1965: El Paleolítico Medio en Cataluña, Ampurias XXVI-XXVII, 1-42, Barcelona. Reichenau W. von 1906: Beitrage zur naheren Kenhtnis der Carnivoren aus den Sander van Mauer und Mosbach. Abhand. d. grossherzoglich hessinschen Landeraustalt zu Darmstadt, 4, 189-314= 14 pi. Sondaar P. 1968: The osteology of the manus of fossil and recent Equidae. Verhandeligen der Kan nederl Akad. von weternscjappen Afd. Natuurkunde: Eerste reeks deel XXV n. 0 1, Amsterdam. Stehling H.G. 1933: Vid. Dubois A. Zabala J. 1980: Biometría del tejón (Meles meles L. 1758) en la sierra de Aralar, Munibe, 32, 302-315. 270 PEDRO M. CASTAl\JOS ANEXO Este Anexo incluye las tablas 28 a 62 en la que se ofrece todas las medidas con sus resúmenes estadísticos y comparaciones con otros conjuntos faunísticos. Las medidas se han tomado siguiendo la metodología de A.v.d. DRIESCH (1976), trabajo que se ha hecho ya clásico en los últimos años. En las medidas dentarias de caballo se sigue el método descrito por F. PRAT (1968) en su trabajo sobre Equidos de Plio-Pleistoceno. El grado de desgaste se señala con una, dos o tres cruces para describir un nivel incipiente, medio o avanzado respectiva mente. Las abreviaturas utilizadas son las siguientes: ACO AFM AS Ad Adsa ADe AmV Alt. Ap APacd APC AS ASc AScr ASp ASd AT D Da-pM DmT DIT Dt E EC Ep Ed Anch. cóndilos occipitales Anch. foramen magnum Anch. del Acetabulum Anch. distal Anch. distal subarticular Anch. del dens epistropheus Anch. mínima de la vértebra Altura Anch. proximal Anch. sobre proc. art. caudales Anch. sobre proc. coronoides Anch. superf. articular Anch. superf. caudal Anch. superf. craneal Anch. superf. proximal Anch. superf. distal Anch. de tróclea Diámetro Diám. antera-post. máx. Diám. mín. tróclea Diám. lateral tróclea Diám. transverso Espesor Esp. caput Esp. proximal Esp. distal El EmO EPA Esa L LA Ldo LD LDS LM LmC LMI LMm LMP LMpe LO LPr LS M m SDI NMI NR s so/o X Esp. lateral Esp. mín. olécranon Esp. proceso articular Esp. superf. articular Longitud Long. acetabulum Long. dorsal Long. diastema Long. diagonal de la base Long. máxima Long. mín. cuello Long. máx. lateral Long. máx. mesial Long. máx. proc. articular Long. máx. periférica Long. olécranon Long. protocono Long. superf. articular Máximo Mínimo Long. serie dentaria inferior Número mínimo de individuos Número de restos Desviación standard Coeficiente de variación Media (estadística) Mandíbula: L M1-M3 Axis: 111 82 91,5 Atlas: AM 11 81 82,5 AScr Alt M 11 108,5 80 72 A Ser Alt M MJ aislados: L M3 11 37,5 36 Escápula: LMP LS AS LmC Entrotarsal: AM 65,5 59 111 58,5 37,5 37,5 35,5 36 111 33 30,5 36,5 38 11 81 68 56 65,5 El Ad 38 65 55 37 36 s 65,5 60 36 41 57 31 27,5 31 61,5 30 24,5 28 p. 35 39 32 36 34,5 34,5 76,5 70 111 45,5 126,5 49 Metacarpo: Ap Ad Ed 138 46 57,5 35 40,5 57 52 32,5 39,5 57 33,5 42 111 63 58 35 43 Ap AD Ad 51,5 28 24 27 49 26 22 26 52 27,5 23,5 25,5 51 27,5 22 26 a. a. a. a. LMpe Ap AD Ad 49,5 24,5 21 23 a. 50 24,5 21 23,5 f p. 111 49,5 24,5 21 23 p. Ap AD Ad AD Ad 39,5 28,5 23 24,5 36 26 20,5 23 38,5 27,5 22 24 a. a. Ldo 59,5 29 23,5 27 p. 39,5 41 60 28,5 24,5 28,5 p. a. 37 25,5 19,8 21,5 32,5 ~5 ~5 ~ ~5 20 23,5 a. 22,5 27 a. 23,5 24,5 20,5 22,5 a. a. 34,5 20 20 a. I' 38,5 30,5 24 25,5 p. 36 27,5 21 23,5 p. 64 50,5 57 44,5 IV 59,5 61 32 52 11 111 218 47 26 50 IV 49 26 23 26 61 29,5 24 27 a. p. 54 111 52 24 20,5 24 p. 54 25,5 21 24 p. p. a. 37,5 29,5 23,5 24 p. 37 26 20,5 21,5 p. 36,5 25 19,7 21,5 p. 67 53 63,5 46,5 111 71 55 72,5 54 Tabla 28.- Medidas aisladas de Bos taurus de Santimamiñe. 62,5 52 V 64,5 48 IV 111 48 26,8 54 11 Falange 3: LSD 57,5 93 Ad 53 27 22,5 26,5 111 LM Ap Rótula: AM 11 AD 57 30,5 25,5 28 a. Falange 2: LM 68,5 V 62,5 32 26,5 29,5 a. 37,5 Radio: Ap 11 a. 37 49 Metatarso: LM1 Ap Falange 1: LMpe 38,5 I' s Calcáneo: LM AM 35,5 Fémur: EC 51 11 63 58 35 38,5 33,5 AT 48 51,5 38 Húmero: Ad 71,5 61,5 49 36 IV 111 Astrágalo: LMI LMm 37 33 111 36,5 ~ ~ 19,9 24 a. 24 25 a. 40,5 29 23 24 p. 49,5 24 19,8 24 a. 49,5 PEDRO M. CASTAi\JOS 272 Tabla 29.- Medidas aisladas de Ovis aries de Santimamiñe. Húmero: Ep s 111 11 Radio: 42,5 11 111 Ap 28,5 34,5 26,5 32 Ad 23,5 29,5 28,5 ASp AT 22 28,5 28 Ad Fémur: 11 41 42 LMI 27,5 EC Ad 19,9 19,6 LMm 27 El 15,5 Ad 17,4 Calcáneo: 41 42 s 111 LM 65,5 57,5 AM 21,5 18 Centrotarsal: AM 29 . 111 Ap 31,5 20,5 IV Ulna: 27 Astrágalo: 111 s IV EPA 25 EmO 21 APC 17, 1 IV Rótula: 31,5 31,5 LM 26 30,5 29,5 AM 17,4 17 18,2 20,5 20,5 111 Falange 3: 26,5 19,5 11 LSD 26 Ldo 20 Tabla 30.- Medidas aisladas de Capra hircus de Santimamiñe. Escápula: 11 Húmero: 111 LMP 43 40,5 30,5 30 Ad LS 35,5 23,5 AT 28 33 27,5 23,5 AS LmC 19, 1 19,6 26 25,5 18,2 16 Radio: Ap ASp 11 30,5 28 s Ulna: 26 EPA EmO 25,5 APC 11 Astrágalo: Fémur: 111 25 20 14,5 IV 111 31,5 30,5 31 31 31 30,5 31,5 33 LMm 29,5 29 30 29 28,5 31 31 El 30 17,7 17,5 17,8 17,6 16,9 17,5 17,4 19 Ad 21,5 21 19,9 20,5 20,5 20,5 20,5 21,5 LM 76,5 74 AM 29 26,5 69,5 26 59 19,5 s 11 42 28,5 29,5 29,5 31 39,5 27 29 28,5 11 111 46 19,4 39 38,5 19,7 19,8 EC Ad s 111 32 Ap LML Calcáneo: 11 1 30,5 29 40,5 s IV 37,5 Rótula: AM IV 18,6 273 ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMIFiJE (VIZCAYA) Tabla 31.- Medidas aisladas de Ovis a./Capra h. de Santimamiñe. Maxilar: 11 L M1-M3 39,5 L P2-P4 L P2-M3 IV 111 48,5 45 20,5 66,5 Mandíbula: L P2-M3 80 L P2-P4 26,5 L M1-M3 55 L M3 45,5 22,5 41,5 21,5 63,5 48 20,5 22 11 26,5 21,5 22 21,5 111 71,5 22,5 50 23 46,5 23 50 48 22,5 71,5 21,5 53 s IV L M1-M3 L P2-P4 24 L P2-M3 L M3 23 M3 aislados: L 20,5 24 69,5 24,5 50,5 45,5 18,7 23 22 27,5 31 71 22 48 21,5 67 21 25 17,9 22 11 23 23 22,5 20,5 22 25,5 21 23,5 27,5 23,5 22 24,5 23 28 22 22 IV 22 111 L Axis: LCDe AScr APacd AmV Tibia: Ad Ed F~lange 1: LMpe Ap AD Ad Falange 2: LM Ap AD Ad 66 18,5 46,5 23,5 23,5 22 11 60 51 32,5 25 41 24 1 23,5 s IV 111 60 39 31,5 29,5 23 24,5 18,7 24,5 26 18,3 20,5 41,5 . 43 46 s 11 24,5 22 23 Atlas: ASer ASed Centrotarsal: .AM 22,5 25,5 11 111 22 23 25,5 42 25 24 44 45 40,5 44,5 111 24,5 s s 19,7 24 22,5 22,5 22 23 11,9 11,4 10,5 11,9 10,7 11 8,4 7,8 7,3 8,5 7,2 8,4 9,2 9,4 7,7 8,2 9,3 21 111 38 36,5 36 41 39 37 36,5 38 38,5 39 35 35 13,4 13,3 12,3 11,8 12 11,4 12,8 12,4 11,6 13 11,4 10,7 10, 1 10,4 9,4 9,4 9,6 8,8 9 9,2 9,5 8,4 12,7 12,4 12 11,3 11,6 10,9 12,2 11 10,7 11,9 10,2 21,5 21,5 12, 1 12,2 8,5 8,4 9,8 9,9 23,5 11 111 11 21,5 10,7 7,8 12,8 11 11,3 42 44,5 IV 41 45 274 PEDRO M. CASTAl'\IOS Tabla 32.- Medidas aisladas de Sus domesticus de Santimamiñe. Maxilar: L M3 A M3 L M1-M3 11 27,5 17,4 24,5 12, 1 33,5 19 27 16,7 33 21 71,5 31,5 18,6 28 16,6 35,5 15,8 70 35 32 16 37,5 18,4 92 34,5 14,7 37 16,8 69,5 37 17,7 38,5 17,6 Mandíbula: L M3 A M3 L M1-M3 72 L M3 A M3 33,5 15,5 Calcáneo: LM AM Metacarpiano 3: LM Ap Ad Astrágalo: LMI LMm 36,5 18,9 37 18 11 47,5 50 54,5 59,5 35,5 17,5 28,5 14,7 37 35 15 33 18,6 s IV Atlas: LM Alt. M ASer ASed 111 33 18,7 Axis: AScr Espápula: LmC 20,5 Tibia: Ad 11 111 32,5 31,5 11 s 44,5 55,5 IV 29,5 Radio: Ap 111 Fémur: Ad 11 52,5 56 66 s 111 86,5 24 88 24,5 24,5 26 25 s IV 17 17 2:5 Metacarpiano 2: LM 88 25 111 72 17 17,5 l 20,5 23 39 36,5 40,5 37 40,5 37,5 42,5 39 s 71,5 22 IV 111 42 38,5 66,5 Metacarpiano 5: LM 11 34,5 32,5 s 60,5 41 37,5 43,5 40,5 40,5 38,5 41,5 38,5 42,5 37,5 41 38,5 62 275 ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMll\IE (VIZCAYA) Tabla 32. (continuación) 11 Metacarpiano 4: LM Ap Ad Falange 1: LMpe Ap AD Ad 15 1 37,5 16 13,9 15,4 14,2 21,5 13,5 15,7 17,6 Ad IV 111 36,5 16,6 13,6 14,9 37 16,6 13 15,7 15,4 36,5 15,4 12,8 14,9 24,5 18,7 16 16 24,5 14,8 12,4 13, 1 111 24,5 18,3 15 15 LSD Ldo 33 30,5 34,5 32 LSD Ldo 33,5 33,5 35,5 35 LM Ap AD 15 14,6 16,6 17 V 36 15,6 12,4 15 36,5 15, 1 24,5 17,5 14,8 16,8 20,5 15,5 12,2 14,2 14,6 23,5 16,2 13,2 13,9 21 16,2 13,3 14,6 24 19,2 16,4 17,7 22 16,2 13,7 14,5 24,5 18,6 15,7 17,5 s IV 23 18,4 14,6 14,6 25 19,6 25 18,8 15,5 18,4 24 17,8 13,8 16 24 17,5 13,7 15, 1 23 17 14 15,8 23,5 17,8 13,5 16, 1 22 16,5 14,4 15,6 25 19 15,6 17,2 11 Falange 3: 35 34 33 31 29,5 27 28 26 36 36 34 34,5 31,5 34,5 33 34 36 35,5 111 36 35,5 IV LSD Ldo 35 33,5 36,5 34,5 LSD 36,5 36 36,5 34,5 Ldo s 11 11 Falange 2: LM Ap AD Ad 1 80 14,3 15,4 Metatarsiano 3: LM Ap IV 64 34 32 35,5 34,5 34 32 29 26 29 28,5 33,5 33,5 37 34,5 36,5 35,5 35,5 36,5 32 32,5 35,5 35 32 32 37 38 38 35,5 37 34 s V 37,5 37,5 36,5 36,5 32,5 32 35 36 37,5 37 34 31 34 33 37 35 37,5 36,5 276 PEDRO M. CASTAÑOS Tabla 33.- Medidas aisladas de Canis familiaris de Santimamiñe. Maxilar: L p4 11 Húmero: Ep 11 16,8 A p4 Metacarpo 5: 1 LM 36 1 46 9 L M1 A M1 13 18,6 Mandíbula: L cónd. - infradent. L proc. ang. - infradent. 11 L cónd. - alv. C L proc. ang. - alv. C L serie molariformes 'L serie premolares L serie molares L Ml A Ml Alt. post. Ml 19,9 8,2 163 33 16,7 EC Ad 109,5 69 36 34 20,4 7,9 30 Calcáneo: 31 1 43 LM Metatarso 5: 21,5 11,5 AM IV Fémur: LM Ap IV 123 125,5 107,5 57 LM Tabla 34.- Medidas aisladas de Fe/is catus de Santimamiñe (nivel 1). Maxilar: L serie molariformes L serie premolares L p4 Mandíbula: L cónd. - infradent. L proc. ang. - infradent. L cónd. - alv. C L proc. ang. - alv. C Alt. ramus 21, 1 19,3 10,2 19,2 17, 1 8, 1 9,2 50 49 44,5 59 58 51,5 51 24 11,3 55 54,5 47,5 48,5 26 43 Alt. tras M1 L serie P3-M1 L serie P3-P4 19, 1 9, 1 16 9,9 A proceso artic. L P4 L carnicera AM corpus 11 5, 1 6,7 4,6 13 6,9 5,4 Húmero: LM Ep Ad Fémur: Ep EC 87 17,9 16 98 19,6 20 9,6 15,6 7,8 17,5 16,6 15,2 48,5 54 54 43 48 47,5 23,5 9,9 10,3 18,3 9,7 18,2 9,4 18,3 11,4 13,4 6 6,7 5,6 11,5 11,2 17,8 11 6, 1 6,7 4,8 5,8 6,8 4,5 5,9 6,7 4,9 Radio: LM Ap 81,5 7,2 Ulna: LM EPA 10,9 10,6 AD 11,4 EmO APC 9,2 9,3 2Tl ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMll\IE (VIZCAYA) Tabla 35.- Medidas aisladas de Equus ferus de Santimamiñe. Maxilar: LP 2 - P4 : VI 101,5 11 L A 32,5 25.5 VI V 35,6 24,7 34 24,5 +++ +++ L A Pr M1-2: 36,5 25 36,5 25 37,3 23,8 37,5 25 37 25,8 +++ + + ++ +' ++ VI V 11 27,4 27,2 15,8 57,6 29,3 26,9 14,8 50,5 26,6 26 12,7 47,7 + + + IV 25, 1 29,8 27,4 24 13,4 13 53,3 43,9 27,9 26,2 12 43 + L 1 Mandíbula: LM1-Ma: L VIII 29,7 29,4 31 32,8 35,9 30 32,3 29,2 28,3 27,7 27 26,4 13,3 15,6 15,2 13,2 12, 1 12,5 40,7 42,5 42,9 47,5 42,3 44 + 30,9 27,7 16,4 27,3 25,2 13,5 49,4 53 + 111' 29,6 26,8 13,3 + VI VII I' 26,4 25,2 25,1 25,9 23,7 26,8 26,9 25,9 25,4 28,4 25,7 24,9 27,5 25,4 25 26,4 27,7 27,4 26,4 26, 1 24,6 28,8 26,6 26,9 26,8 29,2 25,8 25,9 28,4 25,5 27,5 28 12,7 13,8 13,2 14,4 11,8 12,2 11,9 11,2 15,3 12,8 13,1 12,2 15,6 14,4 11,7 11,4 48, 1 54,7 52,5 55,5 49,7 45,5 44,2 43,2 60,2 45 52,4 48,9 56,7 56,6 46,8 43, 1 + L A Pr. x 100 I=--- VII + V A Pr VII 41 23 + + + + +++ ++ +++ + + + ++ + IV V VI 31 24,4 30,4 24,3 27,3° 24,2 31 23,8 30,2 24,3 31 22,6 28 22,7 30 23,3 + + +++ ++ + ++ +++ + 37 14,7 V 33 18, 1 33 18,8 33 18,6 30,5 15 24 15,7 33,3 17 33,3 18,8 30,9 15 34, 7 17,5 + ++ +++ ++ + + ++ +++ + + + ++++++ VII VI 105,5 IV L A L A VI VII L 31 13,9 11 111 31,5 30,5 28,5 24,5 14, 1 17,3 15,8 14,4 IV V VI 28 17 30 16,1 27 31,5 28,5 28,5 27 27,5 29 26 26 30,5 27 17,2 17,5 19,3 18,4 17,4 15,3 17,4 14,5 14,6 18,1 17,9 +++ + + + + + VII A VIII 28,5 19,2 ++ + ++ + VIII 29 18,5 24,3 14 27,5 16,5 25,7 15, 1 27,4 16,6 27 16,6 26,5 15,8 + +++ + + + + + + + + + + 278 PEDRO M. CASTAl'\JOS Tabla 35. (Continuación) Mi-2: L A IV VI 24,2 17 24 18,3 22,7 18 L A Axis: Escápula: LS AS 26 14,6 26 17,4 26,8 16,8 23,2 14,7 25,5 14, 1 + + ++ + ++ + 11 IV V 26,5 13,4 29,5 12,7 30,5 13,7 28,5 15,6 32,8 13,7 33,5 14,8 30,6 12 + + + + + +++ ++ VI Radio: 11' 58,5 50,5 AIM AM ASd LmT 73 VI 114,5 IV VI 67,5 70 59,5 69,5 VI LM Ap 263 52,3 49 48 38, 1 34 12,9 97,9 Ads.a. Ed AD ¡, 12 ++ + 27,5 14,5 59 Rótula: V LM 67 67 AM IV' I= 78 AD. 100 LM 82 85 Centrotarsal: AM V 11 31,5 12,3 42 79,5 VII 34,8 12,3 VIII 44 113,5 61,5 64,5 53 63,5 Metatarso: Ad 1 90 82 45,5 56 VI VI 341 86,5 75,5 41 78 AD Astrágalo: 42,5 11 ASp LM VI 40,2 56 84,5 66 Ed 111 29 15,8 94,5 Ad Ap Calcáneo: 58,5 Tibia: 39,7 I' VIII 26,6 16, 1 VI EC Ad LM Ad VII Fémur: 87,5 Carpa! 3: AM I' 31 16 VI AScr VIII 26 15,2 +++ +++ +++ MJ: VII VI Tarsal 3: 61,5 65 56,5 64 AM li= AD. 100 LM 57 53 60 VI 49 50 12= Ad.s.a .. 100 Ad ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMIÑE (VIZCAYA) 279 Tabla 35. (Continuación) Metapodios: Ad I= VI V 52,5 51,6 51,7 52,2 Ad.s.a. 51,2 50,3 Ed 38,1 39,4 97,5 97,4 48,5 52,8 Ad.s.a. 47,5 50,4 Ed 38,9 97,9 38,8 Ad.s.a .. 100 100,9 53,8 54,3 VII 52,4 52,9 49,9 48,8 48,4 47,7 55,5 55 49,9 50,4 39,6 40,4 38,5 40, 1 103, 1 101,2 95,2 95,2 96,9 49,6 97,7 49,8 53,9 49,2 51,5 55, 1 37 40,5 39,5 103,4 102,2 99, 1 50,2 38,3 Ad Falange 1.ª: VIII Ad VI LM 48,8 37,2 95,4 86,6 83 Ap 54,5 54,8 55,4 59,4 ASp 48, 1 48,9 55,2 33,8 35,2 Ad 38,3 48,6 50 34,2 ASd 46,5 44,3 AD I= VIII 83,9 44 46,2 AD. 100 43, 1 38,9 41,9 41,2 LM Falange 2. ª: I= VI LM Ap 50,7 50,5 50 50,6 54,5 53,9 55,7 58,6 Falange 3. ª: AD 44,2 53,6 47 51,2 47,8 43,8 47,3 AM VI VII 81 Ad 48,8 50,6 48,6 51,9 AS a 49,5 52, 1 AD. 100 87, 1 96,6 86,5 92,3 ESa 31 33,8 LM Tabla 36.- Medidas aisladas de Capra pyrenaica de Santimamiñe. Clavija córnea: CbC DMB Mandíbula: VI 105 112 Maxilar: 118 38,5 L P2-P4 h h h L P2-M3 VI 22,5 21 L M1-M3 26 29,5 IV' L P2-M3 L P2-P4 L M1-M3 s 75 23 21,5 32,5 26 53 26,5 Axis: 24,5 I' 80,5 70 24 23 57 29 47 21,5 VIII 80,5 AScr 50,5 24 AmV 31,5 55,5 V' 31,5 VIII L P2-M3 L M3 26 33,5 L P2-P4 IV' 44,5 33,5 V VI L M1-M3 46 54 22,5 23,5 50 24 67 77 72,5 11' 111' 76 22,5 23 21,5 52,5 23,5 Atlas: A Ser 26 VI 38,5 PEDRO M. CASTAÑOS 280 Tabla 36. (Continuación) VI Escápula: LMP LS AS LmC 36,5 31,5 25,5 22,5 Húmero: Ad AT 44 41,5 36,5 30,5 27,5 23 111 46,5 41,5 Pelvis: AS 35,5 Astrágalo: LM1 LMm El Ad Falange 1: LMpe Ap AD Ad 48 44 11' 36 33,5 Falange 2: LM Ap AD Ad V' 43,5 37 28 Radio: Ap ASp Ad 11 34 32,5 29,5 VIII 38,5 45,5 33,5 39 36 22,5 27,5 37,5 34,5 20,5 25 33,5 31 34,5 32 40,5 39,5 41 39 30,5 37 35,5 s VII 21,5 21 Calcáneo: LM AM V 70 23 V 30,5 28,5 17,8 19,4 36 34,5 19,8 22,5 33 30,5 18,5 21 27,5 13,6 10,5 11,6 11 28 14, 1 10,6 11,4 . 15,4 15,3 15,5 17,5 17,4 18,5 25,5 14,8 11,6 12,4 30,5 14,4 9,6 11, 1 IV' 111' VI 29 15,2 11,9 27,5 33 16,8 12,5 12,9 77,5 27,5 VIII 35,5 33,5 19,7 22,5 15,6 20 77,5 29 39,5 36,5 21,5 26,5 40,5 37 22,5 26,5 IV I' 14,5 VIII 28 IV' VIII 71,5 24 VI 30 28 17, 1 20 16,3 16,6 39 37,5 Centrotarsal: AM 27,5 45 51 17,7 14,6 16,2 38 36,5 40,5 IV 15,9 VIII 37,5 35 38 36,5 40 39 11 15,4 VIII 37 35,5 V' IV' 38,5 36,5 40,5 38,5 VII 44,5 42,5 VII 39,5 38,5 37 35 111' 40,5 37 VI 11 37,5 34,5 20,5 23,5 VI VI 37,5 36 Fémur: EC Ad IV s LMpe Ap AD Ad 11' 35,5 29 23 V 41,5 39,5 I' Ad AT Tibia: Ap Ad 33,5 29 22 23,5 IV' 32 14,3 9,6 10,9 25,5 15,5 11,4 12,4 s 111' V' 32 31 18,6 21,5 38 36 20,5 26 38 35 20,5 24 VI 43,5 15 11 12,6 14 VIII 17 15,7 16,7 ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMll\IE (VIZCAYA) 281 Tabla 36. (Continuación) Falange 3: LSD Ldo 11 VI VII 37 32,5 47 37 40,5 30,5 Metacarpo: Ad VI 34 35 Metatarso: 111 V Ad 33 37 11' 39,5 33 111' IV' 45,5 37,5 VII VIII V' 41 30,5 36 36 34 37,5 VIII s V' 38 32,5 31 179 25,5 15,2 37,5 VI 36,5 40,5 32,5 46 39 44 LM Ap AD Tabla 37.- Medidas aisladas de Rupicapra rupicapra de San!imamiñe. AM sb cóndilos AFM Maxilar: LP2p4 VI VIII 21,5 40,5 62 V 19,4 IV VI LP2-MJ Húmero: Ad AT Escápula: LMP LS AS LmC 30,5 28,5 29,5 Atlas: VII Axis: 36,5 42,5 AScr AScd VII 43 47,5 17,3 17,9 18,4 35 32,5 30,5 29,5 34 32,5 31.5 27,5 23,5 IV' 41,5 21,5 48 70 17,8 32,5 31 31 30,5 32 30 IV VIII 32 26 22,5 19,8 VIII 17,2 VIII 25,5 19,8 21 AScr 17,6 VI 29,5 25 23 19,7 21,5 18,9 Dt base 1 36 26 21,2 19, 1 VIII Dapbase VI 18,5 40 61 17 LP2P4 LM1-MJ LM3 39 32,5 23 LM 1-M 3 LP2-M 3 Mandíb.ula: Clavija: IV Cráneo: 32 27,5 23 19 35,5 29,5 20,5 21,5 VI 37,5 PEDRO M. CASTAl\IOS 282 Tabla 37. (Continuación) Radio: Ap ASp VI 34,5 32,5 IV 32,5 30 VIII AA VI 111 111 66,6 22,5 Centrotarsal: 111 21 31,5 26 El Ad LMpe Ap AD Ad 20,5 68 22 66 21,5 IV 1 VI 22 26,5 27 1 31 29 17 19,5 11 28 26,5 15,7 17,9 36,5 11,3 8,5 10,6 39,5 11,7 9 10,9 49,5 14,4 9 13,3 Falange 2. ª: LM Ap AD Ad 28 26,5 14,9 17,8 111 27,5 26,5 15,6 18,5 26 25 14,9 16,9 36,5 11,7 8,8 10,5 111 48,5 12,9 8,4 12,2 IV' 44 14,6 13,5 20,5 10,8 7,4 8,3 IV 22 9,4 6,6 7,6 11 23 10,6 7,8 9 39 12,5 9,2 11,4 VI 30 12,7 10,4 IV 25 26 20,5 67,5 21,5 68 22 Metatarso: LM Ap Ad 11 VIII 1) 2) 3) 4) 29,5 11 28 Ad VI 71,5 23,5 67 24 Falange 1.ª: 1) 2) 3) 4) 29 Tibia: 44 IV' 66,5 22 11 111 25,5 23,5 VI 29 AD Astrágalo: LML LMm VIII 23 21 VIII Fémur: Ap EC 11 25 23,5 Calcáneo: LM AM AM Metacarpo: Ap Ad 29,5 Ad Pelvis: LA 11' 28,5 27 IV 28,5 26,5 16 18,2 V 30,5 VI 31,5 17,2 21 VII 31,5 29,5 18,2 19,8 IV' 30 28,5 17,3 19,4 47,5 13,9 10,2 13 VI 41 11,8 9,4 11, 1 IV 38,5 11,6 8,8 10,6 12,4 45 14,5 10, 1 13,4 VI 178,5 22,5 30,5 29 30 15 12 VIII 37,5 12,8 8,9 11 21 283 ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMll\IE (VIZCAYA) Tabla 37. (Continuación) Falange 3.ª: LSD Ldo 111 IV 26,5 28,5 22 22 V 37,5 31,5 VI 36,5 30,5 Tabla 38.- Medidas aisladas de Gran Bóvido de Santimamiñe. Mandíbula: IV V L P2-M3 L P2-P4 L M1-M3 L M3 A M3 Axis: Ascr A dens 161 21,5 46 19,2 54,5 116,5 39,5 16,9 + ++ +++ VII 76,5 35 IV' 52. Tibia: Ad Ed Centrotarsal: AM 150 54,5 93 38 14,6 56,5 45 19,3 37 16,4 + +++ Astrágalo: LM1 LMm El 11' 54,5 LM Ap 81 49,5 VI VII 57 172,5 50 26,5 93,5 ++ Húmero: Fémur: IV VIII 86,7 83,7 48,5 57,5 65,6 60,5 38 11' 50 43,5 11' VI Ad AT 84,5 79,5 88,5 83,4 DmT DIT 53 40,3 51,8 40 VIII 58 59,5 VI 58,5 11 66 EC 43 45,8 Ad 43 19,2 15,3 VI Carpal 11-111: VI 95 69 38 89 71 56 LS AS L A 111' VII Escápula: LMP 61,5 Metacarpo: AD Ad 39 14,5 76 Ulna: AMP VI IV' 65 59,5 68,5 64 36 46,5 38 94,5 91,5 56 45,5 62 Metatarso: 11 Ad 82,5 1 VI 80,5 116, 1 72,2 51,8 I' 59,5 284 PEDRO M. CASTAl'iJOS Tabla 39.- Medidas aisladas de Cervus e/aphus de Santimamiñe. Cráneo: ACO AFM Maxilar: LP2-P4 IV V 70,5 30,5 80,5 39 43 VI 81,5 33 111 36 43,5 87 29,5 74 34 VII 74 31,5 44 46,5 V 73 76 72,5 70,5 75,5 74 70 LP2-P4 42,5 47 43,5 LP2-M3 LM3 LM3 63,5 72 71,5 V' 47,5 67 66 111 71 74,5 11 48,5 LM1-M3 4) 71,5 70 40,5 73 IV' VIII Mandíbula: 1) 2) 3) 4) 73 46,5 LP'-M 3 LP 2-M 3 VI 49,5 46 73 VII 85,5 28,5 78 30 IV 41 LM'-M 3 ,Lp2_p4 82,5 31,5 27,5 42,5 73 114,5 28 83,5 141,5 32,5 +++ + 44 80 122,5 54 76 67 83 33,5 23,5 35 ++ 33 31 30 34,5 28,5 31 ++ + + + + + 35,5 29 68,5 107 37 50,5 44 +++ +++ 31,5 28,5 + ++ 111 1) 2) 3) 4) 42,5 +++ 29 29 29 30 26 32,5 31 + +++ ++ +++ ++ ++ ++ 72,5 28,5 28,5 27,5 28 28 29 ++ ++ ++ + +++ + IV 1) 2) 4) 45,5 46,5 40,5 27 + V 2) 4) 74 29,5 30 77 29,5 ++ + ++ 32 33 31 +++ +++ 70,5 ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMIÑE (VIZCAYA) 285 Tabla 39. (Continuación) VI 1) 2) 4) 4) 52,5 48 79,5 30,5 47 80 31,5 + + + 46 79 32 47,5 47,5 48,5 84 34,5 76,5 31,5 28,5 ++ + ++ 30,5 34,5 + 30 32 31,5 35,5 31,5 32,5 28,5 + + ++ ++ + + + ++ + 47 81 128,5 31,5 46,5 47 31 31,5 31 82 79 78,5 32 33,5 34 32 33 + ++ +++ ++ + 28 31 30 + + VII 1) 2) 3) 4) ++ +++ +++ ++ VIII 1) 2) 3) 4) 45,5 71,5 70 82,5 44,5 78,5 124 66,5 VIII 1) 2) 4) 50,5 48 34 73 27,5 ++ + 76 84,5 32,5 75 30 29 30 ++ + + + ++ Axis: LMDe AScr AmV AScd A De 30 36 ++ 44,5 32 28 34 29,5 32,5 ++ + ++ ++ ++ IV' 1) 2) 3) 4) 34 41,5 111' 11' I' 1) 4) 75 32 53,5 72 V' 48,5 51,5 78 12,95 35,5 32,5 47 51,5 81,5 33 ++ ++ 11 111 106 99 64,5 64 61 41,5 37,5 35,5 IV VI 58,5 65 70 66 77 28 37 35,5 27,5 35,5 VII VIII 34 32,5 36,5 ++ ++ +++ 31,5 29,5 + + 32,5 31,5 ++ ++ 28 30 + PEDRO M. CASTAl'ilOS 286 Tabla 39. (Continuación) Atlas: 111 AScr A Sed LM Escápula: 1) 2) 3) 4) LMP LS As AmC 80,5 1 64,5 49,5 44,5 35,5 70 VIII S 77 79 108 61,5 68 11 59 60 46,5 47,5 41 40 33,5 58,5 47,5 42 44 38 36 V IV 111 51 40,5 53 49 42 42,5 37,5 36,5 30,5 59 47 42 59,5 47,5 39,5 61,5 47,5 44 64 55 46 53 40,5 36,5 32 60 46 43 35,5 65,6 50 48 37 60 47,5 40,5 34 66 54 52,5 41 62,5 48 43 VI 1) 2) 3) 4) 58 44,5 41,5 35,5 56 43 39 31 60 49 40,5 38 62 66 50,5 48 45,5 48,5 38,5 42,5 1) 2) 3) 4) 60 46,5 42,5 36 59,5 46,5 43,5 32 65,5 52 47 40,5 57,5 63 44 49 41,5 40,5 35,5 1) 2) 3) 4) 55,5 59 41 46 39 41,5 59 48 41,5 32 66,5 61,5 55 47,5 49 43 42,5 36,6 56 60 44,5 47 38 43,5 33,5 37,5 60 44 39,5 36,5 VI 42 32 65,5 49,5 51,5 44,5 49,5 39,5 58 45 41 35 66,5 52,5 55,5 43 VIII 58 46,5 43,5 41 55 51 55,5 52 49,5 52 58,5 54 54,5 51 67 60 49 51 41,5 48 38,5 36,5 47,5 44,5 50 44 55 48,5 51 48,5 49,5 46,5 48 45 58,5 53,5 111' 65 50,5 46 38 Húmero: 1) 2) 3) Ep AT 55,5 53,5 68 61 Ep 84,5 77 Ad 11 1) 2) 3) 66,5 58 Ad AT 50,5 46,5 45,5 43,5 50 48,5 48,5 46 52 44,5 111 1) 2) 3) 74 75 74 73 53,5 50 60 51 67 59 50 45,5 81,5 80 60 53 52,5 49,5 49,5 46,5 287 ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMlt\JE (VIZCAYA) Tabla 39. (Continuación) IV 1) 2) 3) 73,5 2) 3) 66 60,5 2) 3) 58 53,5 V 52,5 50 66,5 62,5 62,5 58 54 58 54 66 60,5 59,5 54,5 69 61,5 62,5 57 70 59 56,5 52,5 67,5 60 58 54 60 55 60 54,5 61 52 60,5 57 59 53,5 61 56,5 66,5 61,5 52 48 47 69 63 53 VI VI 57 51 58 58,4 55 65,5 60,5 60,6 55 55,5 51 58,5 52,5 59 54 56,5 53 57 53 56 50,5 53 58 53,5 VIII 111' 78 51,5 46,5 51,5 48 47 44,5 Radio: LM Ap Asp AD Ad 1) 2) 3) 4) 5) 56,5 52 VII s 1) 2) 3) 64,5 59 61,5 55 46 44 56 52,5 111 11 58,5 53,5 59,5 V 60,5 55,5 VI 69,5 63,5 55,5 48 44 45 44 326 67 62 37 57 55,5 52 65,5 58,5 65,5 61 63,5 59,5 VI 2) 3) 4) 5) 61,5 56,5 50 60 VII 65,5 62,5 64 60 55,5 55 51,5 54,5 47,5 45,5 50 47,5 s VIII 2) 3) 4) 5) Ulna: EPA EmO APC LO 66,5 60 62 59 60 IV 36 46 53,5 52,5 31,5 59,5 32,5 30,5 VI 35 29,5 61,5 58 58,5 54,5 VII 34 53 48 VIII 54,5 27 45,5 42,5 34 75,5 49,5 40,5 29 76,5 s 47 41,5 33,5 27 288 PEDRO M. CASTAl'ilOS Tabla 39. (Continuación) 11 Metacarpo: LM Ap AD Ad Ed 44 30 41,5 28 39 42 49,5 31,5 44 29 IV 111 38 25,5 39,5 27 280 50 26 49 44 42,5 48 47 VI 47 44 36 49 46 263 46,5 25,5 46,5 277 47 29,5 48 43,5 29 40 28,5 VI Ap Ad Ed 48,5 48,5 41,5 Ap AD Ad Ed 43,5 38,5 42 29,5 27 29,5 Ad Ep 11' 38,5 47,5 26 29,5 41 27,5 11 54 47 111 62,5 52 LMI LMm El Ad 62,5 58 35 37,5 54,5 51,5 31 38 LMI LMm El Ad 51,5 49 28,5 31,5 53,5 64 49,5 58,5 29,5 36,5 34,5 39 LMI LMm El Ad 55 49 27,5 30,5 32 37 LA AA 45· 53 47 42 43 VII VI Pelvis: 44,5 42 42 48,5 41,5 28,5 s VIII 46 27,5 41,5 28 43 53,5 33,5 43,5 62 52,5 63,5 54,5 40 36,5 58 55 33 34,5 56 50 33 36 49 45 42,5 61 55 55 48,5 44 43 45 40,5 111' 59,5 52,5 37.5 25,5 36,5 27 50 48 27,5 32 53,5 50 28,5 32,5 V' IV' 44,5 44 30,5 30,5 VII IV s VIII 61,5 53,5 59,5 55 1 Astrágalo: 61 56,5 54,5 49,5 58 54 34,5 29,5 30 40,5 34 36 51,5 54,5 51 53 47 48,5 48,5 51 27,5 29,5 27,5 30 35,5 33,5 33 32 11 65,5 60,5 35,5 40 57,5 54 31,5 36 55 51,5 30,5 34,5 53 49,5 52,5 51,5 51,5 47 49 47,5 48 48 47,5 46 44 29,5 27,5 29,5 28,5 28 26,5 27,5 32,5 32,5 29,5 29,5 34 31,5 31 111 50,5 53 47 47,5 49,5 44 25,5 27,5 30,5 29,5 29,5 35,5 55 52 48,5 51 29 28 34,5 32,5 54,5 50,5 29 34,5 49 44,5 27,5 31,5 49 27 51,5 49 28,5 34,5 53,5 50,5 29 33,5 57 53 32 39 38,5 39,5 27 26,5 36,5 25,5 ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMll\IE (VIZCAYA) Tabla 39. (Continuación) LMI LMm El Ad 49,5 48,5 26,5 31 51 48 27,5 31,5 50,5 46,5 28 32,5 54 51 31 33 51,5 46,5 27,5 33 48 46 27 30 111 50,5 48,5 27,5 33,5 55 50 30 34 51 48 29 33 52,5 49.5 48,5 30 29,5 33,5 32 61 57 32 39 49 47 28 31 50 47 28 32 53 49,5 29 33 50 42 47 39 27,5 23 32,5 27,5 111 53,5 55 47,5 50,5 49,5 51,5 43,5 30 27 28 30 35,5 34,5 31,5 32 54 51 31 35 52 48,5 28 31,5 LMI LMm El Ad 50 47 28 33 .LMI LMm El Ad 49 45,5 27 30 55 54 49,5 51 30,5 30,5 37 36 52,5 49,5 49,5 46 29,5 26,5 33 31 LMI LMm El Ad 52,5 49 27,5 31 51,5 49 29 34 55,5 51 31 36 49,5 50,5 51,5 46,5 48,5 50 28 27,5 29 30,5 32 32,5 LMI LMm El Ad 52 49 28,5 33,5 52,5 50 29 34 53 49 50 48,5 49,5 49 45,5 47 47,5 47 29,5 25,5 27 27 27 35 29 29,5 30,5 32 55 50,5 29 34 111 48 50 43,5 47 27 27,5 31,5 29,5 52 50 49 47,5 28,5 27 32,5 33 49,5 49 26,5 31 47 44 26 28 50 54 55 55 48,5 52 50,5 51,5 26,5 29,5 30,5 30 31,5 34 32,5 34 52 50 29 33 50 49 49 55,5 48 47 46 47 52 45 28,5 27 ,5 27 31,5 26,5 33,5 31,5 32,5 34 31,5 53 50 29,5 33 53,5 50,5 29,5 34,5 IV 53 49,5 29 34 IV 54 50,5 29 34 52,5 54,5 48,5 49,5 28,5 29 33 34 52,5 49 28,5 34 47,5 45 26 30 49 47 28 31 47 43 26 30,5 50 48 29 33 53 51 30 35 53 49,5 29,5 34 59 58 31 53,5 33,5 39 IV LMI LMm El Ad 53,5 51 29 32 59 53,5 32 38,5 50 45 28 32 LMI LMm El Ad 51,5 48 30 33 54 57 52 32 35 LMI LMm El Ad 55,5 48,5 49,5 46,5 30,5 27 32,5 29,5 29 32,5 53 50 29,5 34 54 50 30 35 52,5 50 47,5 47 29,5 28 32,5 32,5 50 48,5 26,5 31,5 52,5 49,5 29,5 31,5 52 49 28,5 32,5 48,5 52 51 44,5 48,5 48,5 27,5 29,5 27,5 32 31,5 33 54,5 50 30 34 52,5 48,5 28,5 33,5 V 53,5 55 50,5 51,5 30,5 31 34 35,5 V 53,5 52,5 51 30 28,5 34,5 53,5 49,5 29,5 33 57 61 55,5 54,5 56,5 31,5 32,5 32 37,5 41 37,5 60 56 32,5 38,5 50,5 46,5 27,5 29 54 61,5 60,5 51,5 57,5 56 29,5 33,5 33,5 34,5 39,5 41,5 52,5 47 29,5 34,5 289 PEDRO M. CASTAt\JOS 290 Tabla 39. (Continuación) 1 VI Astrágalo: LMI LMm El Ad 58,5 55 32 37 54,5 52 30 35,5 51,5 55,5 48 51 29,5 30,5 36,5 33 53 51 29,5 34,5 56,5 60 54,5 56 30,5 32 39,5 37 53,5 49 29 32,5 56,5 52 30,5 34,5 69,5 63,5 38 41,5 56 54 30 34 52,5 56 54 50 28 30 34,5 30,5 55 49,5 31 36,5 62 51,5 58 35 30 35,5 41,5 55 51 30 37 48,5 44,5 26,5 32,5 55 51,5 30,5 36 54 48,5 30,5 34 58,5 55 31,5 37,5 57,5 53,5 48 52 52 47 31,5 29 27,5 31,5 36 35 61,5 57 34 40 60,5 61,5 58,5 55,5 56,5 55 32,5 32,5 33 39 38 40 VIII VII LMI LMm El Ad s LMI LMm El Ad 56 IV' 11' 50,5 53,5 58,5 47,5 49,5 53,5 27,5 29 32 32,5 35 37 Centrotarsal: 57 53 31,5 36 53,5 52 49,5 42 39 39,5 41 AM 43 V 37,5 59 56,5 32,5 37 111 35 40,5 11 AM V' 50 47,5 27 32,5 IV 44,5 36,5 36 44 44 38,5 39,5 39 52,5 37,5 VI 53,5 52 47 48,5 49 43 VIII 42 49 44 49,5 50,5 39 40,5 45,5 48 44,5 45,5 s AM 48 Metatarso: 11 Ap AD AD 1 43 38,5 38,5 41,5 46,5 41,5 45 V IV 111 38,5 VI 40,5 27 41 48,5 40 43 46 42,5 42 44,5 49 44 42 48 VI Ap AD Ad VII 42 28 45,5 48,5 43,5 47,5 43,5 50 49 47 48,5 44 45 43,5 42,5 52 54 51 VIII Ap Ad 44,5 I' 37 V' 39 45,5 45,5 43 43 44 44 44,5 45,5 42 44 43,5 47 53 45 43 291 ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMll\JE (VIZCAYA) Tabla 39. (Continuación) Fémur: 1) AP 2) EC 3) Ad 11 39,5 IV 111 74,5 41,5 38 71,5 69,5 30,5 42,5 44 65,5 39 36,5 70,5 VI 2) 3) V 69,5 29,5 66 68 VII 32 36 40 35 38 VI 96 36 74 s VIII 33,5 36,5 79 11 LM Ap AD Ed 425 86 43,5 57,5 55,5 34 45,5 41 Ap Ad 88,5 Ad Ed 51 54 11 54,5 43 54 41 114 36 106 35 LM AM 79,5 50 37,5 49 41,5 48 35,5 78 48 42 57 43,5 52 55,5 40,5 65 52 39,5 Calcáneo: LM AM VIII 91,5 83 95,5 55 62,5 52 s 51,5 60 IV V 46 54,5 46 64 111 53 56 58 IV' 47 34,5 46 43 45,5 34,5 32,5 34 58,5 44 57,5 52 VI 55,5 53,5 51 57 V' 52 39,5 VIII 48 31,5 49 44 59,5 51 11 111 119 110,5 109 115,5, 118 123 112 118 120 112,5 119,5 104 40 34 37,5 37,5 38,5 37,5 33 34,5 36,5 32,5 37,5 34 111 102 31 VI' IV 111 VIII Rótula: 85 63 VI 82 93 78 37 43 72 Tibia: 76 IV LM AM 119 112,5 38,5 34 LM AM 105,5 114,5 110,5 107 114,5 109 108,5 108 33,5 33,5 35,5 38,5 37,5 34,5 39,5 38 36 37 LM AM 122,5 122,5 121 114 40 40,5 39 40 125 109,5 114,5 44 34 37 104 32 111 102,5 108 113,5 105,5 110 101,5 35 34,5 34,5 39,5 35 31,5 35 IV VI 41 32 111 114,5 34,5 35,5 36,5 VII 121,5 115,5 121 122,5 123,5 116,5 124,5 40,5 37,5 40,5 42,5 43 35 52,5 55 292 PEDRO M. CASTAl\IOS Tabla 39. (Continuación) s VIII LM AM 117 119 38,5 38,5 135 140,5 118 111,5 109,5 120 43,5 47,5 35,5 38,5 35,5 37,5 44 Falange 1: LMpe Ap AD Ad 111 11 63 25,5 19,6 24 21 58,5 22,5 17,5 21 53,5 25 21 19,5 22,5 24,5 23 21 23,5 19,6 18,9 20 19,6 22 26 21 111 LMpe Ap AD Ad 62 20,5 19,8 24 49,5 18,3 15 18,8 IV 52,5 18,7 21 52 18,8 14,7 17,9 20,5 18,5 22,5 18,6 17,8 17,6 24 18,9 23,5 22,5 22 16,8 19,2 17,5 21,5 20 21 20,5 24,5 24 VI Ap Ad 30,5 23,5 LMpe 57 21,5 21,5 21,5 24 21,5 20,5 18,4 19,7 19,7 20,5 21 20 21,5 VI Ad 23,5 24,5 21,5 23,5 23 20,5 60 23,5 18,4 24 64,5 23,5 18,2 23,5 19, 1 19, 1 VII LMpe Ap AD Ad 51 18,7 23,5 16, 1 18 20,5 18,8 22 24 25,5 22 VIII 21,5 20,5 23 21,5 21 18,2 23,5 24,5 24,5 21 21,5 21,5 21 20 20,5 s Ap Ad 19,2 20,5 22,5 IV' V' 19 58,5 21,5 LMpe 20,5 22 Falange 2: LM Ap AD Ad 21,5 21 19,6 20,5 19 17,3 Falange 1: Ap 20,5 21 11 45 39 43,5 23,5 17,2 22 23,5 17,3 14,3 16,4 14,5 21 21 19,2 16,7 18,5 37,5 40 36,5 40,5 18,9 19,2 13,2 23,5 21,5 13,8 13,5 13,3 16,3 15,7 15,3 16,5 15,9 21 18,8 23,5 16,6 21 44,5 22 17,2 19,9 19,2 18, 1 17,9 24 18,6 21,5 111 23 23,5 ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMlf\IE (VIZCAYA) 293 Tabla 39. (Continuación) IV LM Ap AD Ad 21 17,7 37 39 40 20,5 19,6 19,3 15,4 14 14,6 18,4 15,3 16,2 21,5 V 22,5 19, 1 17,2 24 24 23,5 48,5 46,5 23,5 24,5 16,7 17 20,2 20,2 40,5 44,5 20,4 24 15,5 16,7 18,2 20,2 VI LM Ap AD Ad 46,5 46,5 25,5 24 17,8 16,3 20,5 20 40,5 44,5 46,5 46 45 43 19,6 21 20 23 24 23 18,6 18,3 17,7 15,6 14,5 16,7 17,5 17,6 19,5 19,8 19,8 23 22 24 22 21,5 19,8 17,9 17,7 20,5 s LM Ap AD Ad 20 20,5 18, 1 43,5 46,5 21,5 21,5 23 17,4 17,7 17,2 18,8 19,4 44 44 20,5 23,5 23 18,8 16,6 18,6 19,7 20 I' 16,7 18 19,7 19,6 24 24 20,5 V' 11 LSD Ldo 57 43,5 47,5 49 56 45 52 39 42 45,5 51,5 41 47 43 LSD Ldo 46 47 49,5 39,5 43,5 42 45 42,5 44,5 38 42,5 40 43 47,5 41,5 42 36,5 49,5 52,5 48 45,5 48,5 56 41 42,5 39 37 32,5 46 47,5 44,5 44,5 45,5 50 50 42,5 50,5 52 44 41,5 44 45,5 45 53 57 48,5 39,5 46,5 48 51 45 37,5 45,5 42,5 42 47,5 111 42 51,5 45 49 45,5 42 46,5 52,5 46,5 36 45,5 40,5 44,5 44 38,5 43 47,5 45 LSD Ldo 48;5 54 45 44 51,5 43 LDS Ldo 111 45,5 44 51 43 38,5 49,5 40 45,5 43,5 46 42 47 53 50,5 48 43 42 45,5 39,5 34,5 48,5 38,5 42,5 44,5 42,5 39 43,5 47,5 47,5 41 LDS Ldo 44,5 43 47,5 41,5 49,5 45,5 44 48,5 46,5 45 48 42 38,5 43 37,5 44,5 43 40 46 46 41 44,5 55 50 46 43,5 46,5 48 42 41,5 40,5 45 51 47 47 46 111 48 47 23 s 48 24 38,5 20 14, 1 17,5 40 20 14,7 18,4 17 Falange 3: 22 VIII VII LM Ap AD Ad 21,5 40 41,5 48 52 46,5 36 36 42,5 47 42 37,5 19, 1 14,4 16,9 PEDRO M. CASTAl\IOS 294 Tabla 39. (Continuación) 111 LDS Ldo 44 49 40 38,5 45,5 47 40 45,4 37 45 44 LDS Ldo 47 43 47 46,5 44 43 41 41 46 47 ,5 41,5 49 44,5 44,5 42 46 42,5 50,5 46,5 43 44,5 38 44,5 39 40,5 41 44,5 39 46 42,5 43 44,5 39,5 38 41 43 36,5 36 111 39 40,5 42,5 35 41 42,5 39,5 45 43,5 40,5 41 37 42 33 37,5 38 38 111 LDS Ldo LSD Ldo 36,5 46 44 49,5 46,5 51 34 44,5 43,5 40,5 42,5 48 38 36 48 55 43,5 51,5 52,5 44,5 51 43,5 40,5 43 44 51,5 40 46,5 48,5 39 47 41,5 39 38 41 41,5 39 37 37 49 49,5 47 36 37,5 35,5 33 34,5 46 44,5 42,5 58 46,5 52 46 51,5 49 50,5 52 52 48 50,5 53 46 46,5 40 46,5 47 47 47 47,5 46 46,5 IV LSD Ldo 46,5 48 47,5 46 49 44,5 48 48 46 47 45 47 42 44 45 43,5 44 43,5 43 42,5 44,5 42,5 42,5 46 43 46,5 45,5 39,5 36,5 38,5 41 42,5 40 37 36,5 36,5 IV LSD Ldo 44 48 41,5 51 48 43 49 40,5 38,5 46 43 44,5 40 47 46,5 38,5 45 36 34,5 44 42 46,5 40 50 48,5 49,5 41 51 38 43,5 37 45,5 44 47,5 36,5 46 V LDS Ldo 52 51,5 51 46 47,5 45 LDS Ldo 46 46 51 47,5 48 47 51 52 47 45 49 43,5 46,5 45 48,5 47,5 45,5 42 43 50,5 40,5 41,5 49 41 46,5 38 45 39,5 37,5 33,5 28,5 45,5 VI 49 . 48 39 46 60 54,5 57 57,5 56,5 53 54,5 47 46 47 36,5 44,5 55 50,5 51,5 51,5 48,5 47 51,5 43 VI LDS Ldo 50 57,5 56 55,5 46,5 45 51,5 51,5 50 44,5 VII 53 48,5 45,5 54,5 50,5 51 . 53 52,5 41,5 55,5 57 47,5 46,5 42 50,5 47 48,5 49 50 33 52,5 52 54 48 45 58,5 56,5 48 52,5 41 53 52 45 46,5 VIII LDS Ldo LDS Ldo 53 57 48,5 51,5 54 51,5 57,5 52 46 55,5 44,5 49 46,5 49,5 50,5 47 55 55,5 45 43 39 37 44,5 42 41 51 48 41,5 40,5 35 34,5 41 40,5 37 s 11' IV' 47,5 49 45 45 46,5 41 47 42 58,5 45 52 55 61 45,5 46 42,5 44 43 36,5 47 37,5 53 44,5 49,5 49,5 54 V' 52 52,5 47 47 295 ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMll\IE (VIZCAYA) Tabla 40.- Medidas aisladas de Capreo/us capreo/us de Santimamiñe. Maxilar: LP2_p4 LM'-M 3 LP2-M 3 Mandíbula: 1) LP2-P4 2). LM1-M3 3) LP2-M3 4) 33 28,5 26 34,5 31,5 33,5 55,5 60 33 IV 111 11 28 35 60 28,5 30 56 26,5 31 56 28,5 28 27,5 32 58 29 30,5 VI VIII 34 30 11 30,5 28,5 26 31 28 111 28,5 LM3 39 38,5 38,5 15 14,2 29,5 41,5 70,5 15 16,7 16, 1 16,8 17,6 28,5 28,5 28 Escápula: LMP LS AS LmC 40,5 15 11 32 25,5 23 20 AT 16. 14,3 16,8 16,4 16,9 16,3 16,5 14,5 VI 26,5 29 39 68,5 15,5 2~,5 30 30 23,5 22,5 18,3 15, 1 15 17,5 s VIII 41 16,4 VI 32 25 23 19, 1 IV' 29,5 30,5 42,5 72 17,4 IV 111 Húmero: Ep Ad 27 ' 27,5 28,5 30 27,5 42,8 IV 1) 2) 3) 4) 15,6 IV 111 1) 2) 3) 4) 39,5 43,5 40 16,8 16,4 16,3 17,3 Radio: 28,5 22,5 20,5 18, 1 ASp IV V 27,5 25,5 28 26 28 Ad VI V 111 11 111 31,5 30,5 11 Ap s VII 11' 38,5 32,5 31 32 29 29,5 26 28,5 26,5 28,5 25 29 27 30,5 29 30 28,5 30,5 28 31 28 28 26 28,5 25,5 29 26 296 PEDRO M. CASTAf\IOS Tabla 40. (Continuación) Ulna: EPA EmO APC 26,5 23,5 28 24 17,2 Metacarpo: Ad 11 23,5 23 Centrotarsal: AM Calcáneo: LM AM IV Falange 1.ª: LMpe Ap AD Ad Falange 2. ª: LM Ap AD Ad Falange 3. ª: LSD Ldo VIII 28 23,5 28,5 24,5 18,4 Pelvis: 111 23,5 24,5 23 25 26,5 111 25 25 26,5 66,5 22 65,5 22 31 29,5 17,6 18,5 28,5 27,5 16 17,6 s VI 24,5 111' 25,5 64,5 22,5 63,5 21 65 21,5 29 27 16, 1 18,3 32,5 31 17,9 19,9 23 IV 65,5 21 66 28,5 27 16, 1 18,4 32 21,5 VI 17,9 20,2 64,5 20,5 71 23 VIII 73 24 65 21,5 s IV 111 36 11,4 7,9 9,4 36 10,8 7,8 9,4 35,5 12 8,5 10,3 11 25 11 7,5 7,8 23 11,6 7,6 8,2 111 24 10,3 7,2 7,4 1 26,5 28 26 25,5 27,5 26,5 28 28 111 27 IV 12,6 11,9 12,5 10,4 IV 24 10,4 7,2 7,7 11 27 27 27,5 11,4 8,2 8,8 29 12,7 8,8 8,9 25 10,9 7,4 7,6 26 25,5 24 22,5 25 11,3 8 7,7 111 28,5 27,5 28 27 26,5 27 27,5 25,5 26,5 25 IV LSD Ldo Metatarso: Ad VI 11 31,5 30 17,5 19,5 111 29 25,5 LA AA 111 70 22,5 Astrágalo: LMI LMm El Ad VI 28 25 IV 28 25,5 26 27 25,5 24 28 26,5 29,5 25,5 27,5 27 25,5 27,5 25,5 26 s VII 24,5 23 28,5 29,5 27 27 27 26,5 111' 27,2 27,2 297 ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMll\IE (VIZCAYA) Tabla 41.- Resumen de algunas medidas de Capreolus capreolus procedentes de yacimientos neolíticos y postneolíticos de la Península Ibérica. Maxilar: P. Oro Sacaojos L P2-M3 L P2-P4 58 27 29 L M1-M3 33,3 31 Mandíbula: P. Oro 33,5 CMI L P2-M3 68 L P2-P4 28 L M1-M3 Vega C. 56,5 37,2 Munig. C. Cerdeña Ch aves 64,8 65,5 26,4 28 36 38 16,6 28 41 14,2 L M3 Escápula: S. Ana LMP LS 17 AS (19) LmC 14 Zambuj. Húmero: 26 27,5 27,4 27,9 27,6 21 AT 24 24,5 21,4 24,3 24, 1 18,8 18 ASp 25 25 25,5 Munig. Monach. Falange 1: Sacaoj. 40,3 C. l'Or Monach. LM 61,5 AM 20 Astrágalo: Valenc. 29,8 27,9 29 26,9 28,2 26,6 28 El 17 17,3 16, 1 16,2 16,5 Ad 19 15,9 18,9 18,5 19,2 Covalta C. l'Or 38,8 38,2 32,2 39 36,8 Ap 12, 1 13,5 12,7 13,3 12, 1 12,3 11,7 13,5 AD 9 8,5 9,2 9, 1 8,8 8,8 8, 1 9,2 Ad 10,5 Falange 2: Monach. 11,2 C. L'Or Valen c. Nerja LM 28 27,5 22 27,4 22,2 27,6 21,8 Ap 11,5 12 9 18,8 10 12,3 10, 1 AD 8 8,5 7 9, 1 8,5 7,9 Metacarpo: Zambuj. Monach. Valen c. 24,5 18,5 7,4 C. l'Or 15,5 Ad 22,5 20 27,3 38,5 13,2 60,2 30 28,5 39, 1 Ep C. l'Or LMm 39 AD 28 C. l'Or 38,2 11 Covalta Munig. LMI 26 Zambuj. Calcáneo: 23,8 LMpe Ap c. l'Or Covalta Ad 28 28 Zambuj. 26,5 26,5 42,5 Sacaoj. 22 20,5 Zambuj. Ap 14,8 28,5 26,5 Ad 30 23,5 Munig. Tibia: Zambuj. Ep Ap Radio: Sarsa 11,5 21,9 23,5 34,9 39,4 37 12,7 40 13,4 11,7 11,5 12, 1 9,2 9,5 8,3 8,8 8,9 PEDRO M. CASTAl\IOS 298 Tabla 41. (Continuación) Metatarso: LM Zambuj. Munig. Valenc. S. Ana Abauntz Sarsa 25,5 25,5 165 183 Ap 19,5 AD Ad 12,3 20,5 24,7 26 13 12 21 22,6 Tabla 42.- Medidas aisladas de Sus ferus de Santimamiñe. Maxilar: 11 35 37 A M3 19, 1 18,5 36,5 LMl-M3 L M3 Atlas: 36,5 37,5 20 21 73 Mandíbula: A M3 IV 111 L M3 111 40 18,7 IV V 40 18,7 39 39 41,5 22 18,3 19,4 111 37,5 VI 33,5 36,5 32,5 19 22,5 18,9 21 18,9 78,5 78 s VI IV' 36 41 44 16,6 17,7 18 Escápula: IV IV' 11' LMP 50 LM Alt. M 49,5 52 89 91,5 LS AM 51 50 AS 30 34 LmC 27,5 34,5 A Ser 56,5 53 47 AScd 59,5 59,5 54 Radio: Ap s Tibia: Ad Ad 111 s I' 111' 35,5 35 36,5 29 355 11 41,5 41,5 Fémur: 35,5 111' 36,5 11 111 59 56,5 IV VI s Ad 54,5 49,5 44,5 51,5 50 47,5 42,5 50,5 40 AT 44,5 41 34 39,5 37,5 38 37 42,5 32,5 92,5 93,5 97,5 92 92,5 93 28 26,5 27 25,5 101 29,5 26 27,5 Húmero: 11 111 1 Calcáneo: 92 91,5 AM 28 24,5 27,5 Astrágalo: s 111 11 LM 49 LMm 44,5 46 41,5 44 48,5 44 27 111 11 LMI I' 46 42,5 53,5 49,5 46 48 47,5 44,5 51 45 48,5 48,5 45,5 41 44,5 44,5 40 45 41,5 44,5 299 ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMll\IE (VIZCAYA) Tabla 42. (Continuación) LMI LMm 50 45,5 47 43,5 47,5 41,5 48 42,5 111 48 44,5 46 41,5 46 42,5 LMI LMm 50,5 48,5 45,5 43,5 49 44,5 IV 46,5 43 46 41,5 50,5 47 53 47 49,5 45,5 46,5 41 48,5 43,5 49 45,5 I' 41,5 37,5 s LMI LMm Metacarpiano 3: LM Ap Ad Falange 1: LMpe Ap Falange 2: LM Ap AD Ad 50 46 Metacarpiano 4: LM Ap Ad 11 81,5 18,5 18,4 11 40,5 15,8 39,5 18,2 44,5 VI 37 14,9 44 40,5 49,5 46 V 51,5 47,5 46,5 42 50 46,5 48,5 43,5 49,5 45 VI 41,5 39,5 38,5 37 41,5 38 VIII 49,5 45 52 47 I' 80 14,9 16,7 VI 83,5 14,2 16,2 Metatarsiano 3: LM Ap VI 80,5 14,5 s 19,7 18,8 21 11 26,5 18,3 15 16 27 18,8 15,8 18,6 27 20 16,5 18,8 LM Ap AD Ad 27 16,8 14 14,6 28 18,9 15,5 16 LM Ap AD Ad 22,5 15, 1 12 14 23,5 17,4 14,6 16,4 Falange 3: LSD Ldo 1 39 39 40 38 VI 27 18 15, 1 15,9 26,5 20,5 15,9 17,8 29 19,6 17 17,2 28 18,2 14,9 15,8 25,5 18,2 14,9 16 28,5 19,2 15,4 16,6 27 19,6 16,2 18,9 29,5 19,5 15,6 16,2 IV 27 20,5 16,6 19, 1 26 17,8 15 16,8 25,5 18,8 15,5 17,9 26 19,6 16, 1 17,5 11' 22 16 13 14,6 39,5 37 39,5 37,5 44 43 40,5 39 IV 40,5 40 41,5 40,5 27 16, 1 13,7 14,3 26 19 16,3 18,7 26,5 18,3 15,4 18,6 28 18,8 15,8 16 28 20,5 17 19,3 30 20,5 17,5 20 111 26,5 19,2 16,7 18,5 27,5 18,2 14,4 15,5 VIII 30 18,7 16 16,4 28,5 18,4 15 16 25,5 18,7 14, 1 16,8 26,5 16,8 14, 1 14,5 38,5 36,5 39 38,5 38 38 40 39,5 V 11 111 VIII 17, 1 PEDRO M. CASTAl\IOS 300 Tabla 42. (Continuación) V VIII VI LSD 40 40,5 39 39,5 38,5 38 37,5 36,5 35 36 38,5 Ldo 39,5 41 37,5 37,5 38,5 36 35 34,5 34 35 36,5 LSD 38 34 34 37 40,5 40,5 40,5 41,5 35 Ldo 35,5 31 33 35,5 38,5 38,5 39,5 39,5 34 s I' Tabla 43.- Medidas aisladas de Canis lupus de Santimamiñe. Maxilar: Mandíbula: 11 111 11 L P4 16,8 L M1 19,9 21 A P4 9 A M1 8,2 8 13 L M2 A M1 18,6 A M2 AS s LM 38 34,3 Ap 12 11,8 Ad 9,8 7,7 11 s Pelvis: 5 2 LA 21,5 LM 75 80,5 AA 21,5 Ap 14 9,5 Ad 12 12,7 Metacarpiano: EPA 33,8 EmO 27, 1 IV' IV Falange 1: VI Ulna: 25,5 10,6 12 L M1 Escápula: 27,6 11' IV Metatarsiano: 31 4 Ap 8,4 9,9 Tabla 44.- Medidas aisladas de Martes sp. de Santimamiñe. Maxilar: 11 IV Mandíbula: L C-M1 29 32,5 L cónd.-infrad. L s. molarif. Alt. M s IV 11 50,4 52,2 52,9 29,2 23,5 26,6 L P4 7,9 8 L s. molarifor. 30 29,5 A P4 5, 1 5,9 L s. premolar. 17,4 17,3 16, 1 8,8 L s. molares 13, 1 12,9 13,4 9, 1 9,4 4,6 5,7 A M1 AM paladar 29, 1 Am paladar 17,9 Asobre C 19,7 L M1 DFM 24,3 29,3 8,5 5',2 19 11,2 3,9 5,3 301 ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMlr\IE (VIZCAYA) Tabla 44. (Continuación) Húmero: 11 Axis: Radio: 11 LcDe Alt. M 18,6 LM 64,2 70,3 LM 17,2 Ep 11,6 13,2 Ap AScr 13,9 AD 3,9 4 4,4 Ad Ad 12,9 13,5 Fémur: Ad 13,2 Metatarsiano: 11 2 13,5 LM 22,5 LM Tibia: LM 86 Ap 14 Ad 9,6 Tabla 45.- Medidas aisladas de Vulpes vulpes de Santimamiñe. Cráneo: Atlas: L acrocranion - prostion 137 LM 22,5 L cónd. occip. - prostion 134,5 AScr 25,5 L basion - prostion 129 AScd 19,3 AFM 14,2 L arco dorsal AM sobre cóndilos 25,5 mediano A otion - otion 45 A zigion - zigion 73,3 A entorbitale - entorbitale 27 L serie molariformes 49,5 L serie premolares 24,5 L serie molares 25 L P4 11,6 11 50,8 51,3 14 Metacarpiano: 11 s 54 7 9,2 Húmero: Ad Radio: Ad 9, 1 11 20,2 VI 15,2 L M1 7,6 A M1 10,5 L M2 4,4 EPA 19 A M2 7,8 EmO 15,6 Ulna: VI VI 11 2 3 4 32,9 37, 1 38,4 PEDRO M. CASTAl'ilOS 302 Tabla 45. (continuación) Mandíbula: L cónd. - infradent. V L proc. ang. - infradent. L cónd. - alv. C L proc. ang. - alv. C Alt. ramus Alt. ante P3 AA 13,6 Metatarsiano: LM 11 4 57,8. 61 36 24,5 14,3 61,5 34,5 25,5 VIII 37 12,3 16,5 13,9 52 32,5 24,5 13,6 5 7,5 L carnicera A carnicera AM corpus 14,4 VII 1 oo 99,5 Alt. tras M1 A proceso articular L serie molariformes L serie premolares L serie molares Pelvis: LA VI 103,5 102 24 Fémur: Ap EC 34 16 15,6 1 VII Astrágalo: 14,8 16 LM Tibia: 24,5 11,6 IV VII VIII 2 61 5 59 2 62,2 Ad Falange 1: LM 18 IV 25 Tabla 45 bis.- Medidas aisladas de Ursus arctos de Santimamiñe. Canino sup: A Metatarsiano: P4: 14 L A IV V LM A mit D 3 92,5 15,2 2 73,5 Ap AD Ad 14,5 22,6 12,2 16,6 111' 14,4 9,4 111' M1: L A 22 12,4 MJ: 26,8 14,6 L A 11' 24,4 16,6 111' 23 15,7 ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMlfilE (VIZCAYA) 303 Tabla 46.- Medidas aisladas de Me/es me/es de Santimamiñe. Mandíbula: L cónd. - infradent. L 11-M2 L C-M2 L P1-P4 L M1-M2 s Húmero: LM 112 103 58 Ep 28 26 44 AD 10,4 18,6 Ad 33 16,5 Radio: A M1 7,8 Ap LM 9,2 31 33,3 25 L M1 1 11 31,5 20 Calcáreo: IV 111 88 Ulna: 11 13,8 Metacarpiano: LM 111 LM 11 111 2 4 28,2 29,1 1 116 Metatarsiano: LM VI 116,5 11 V 4 5 35,7 33 Tabla 47.- Medidas aisladas de Fe/is silvestris de Santimamiñe. Mandíbula: L cónd. - infradent. L proc. ang. - infradent. L cónd. - alv. C 111 59 50,5 L proc. ang. - alv. C Alt. ramus IV V VIII Escápula: 62,8 LM 61,8 LPA 10,3 -- 16,4 55 LS 14,7 53 AS 10,6 LmC 13,5 27,6 Alt. ante P3 1-11 79 11,3 9,3 Alt. tras M1 10,5 12,2 L serie P3-M1 21,5 21,3 17,7 21,4 L serie P3-P4 Húmero: IV 11 12,9 12,9 10,9 13,5 LM 115,5 95 95,5 L P4 6,2 6,3 6,2 6,2 Ep 24,5 20,2 19,8 L carnicera 8,4 8,1 6,8 7,8 Ad 22 17,4 16,8 AM corpus 5 6,3 11 IV Radio: LM Ap 111 108 9,4 AD 6 Ad 14,8 Tibia: Ad VI IV 94,5 7,8 12,7 Ulna: LM EPA EmO APC 5,6 11 LM VI Metacarpiano: 127 12,2 10,3 10,2 9,6 10 10,3 9 10,6 11,9 IV Metatarsiano: 13,3 111 127 2 4 5 53,2 67 54,4 LM 3 5 39, 1 25,5 304 PEDRO M. CASTAl\IOS Tabla 48.- Medidas aisladas de Linx linx de Santimamiñe. Mandíbula: L P3-P4 Alt. a. P3 L P3 A P3 L P4 A P4 L M1 A M1 IV Metacarpiano: 111 23 20 5 LM Ap AD 10, 1 4,5 11,7 10 Ad 5,5 15,8 6,9 VI 2 65,8 9,6 7,2 3 75 10,6 7,9 11,9 12,6 4 8,7 VI Falange 1: IV LM Ap 4 102,7 9,3 LM Ap 39 14, 1 Ad 8,8 AD Ad 8,4 12,5 Metatarsiano: Tabla 49.- Medidas aisladas de Lepus sp. de Santimamiñe. Escápula: LMP AS VI VI Pelvis: LA AA Metacarpiano: LM 13,3 12 11 2 26,3 I' Húmero: 13,3 .11,6 LM 103,5 Ap Ad 19,6 12,2 Fémur: 13,3 12, 1 Ap Metatarsiano: LM VI Radio: LM Ap 111 9,5 11,6 12,6 Ad IV Falange 1: IV 24,5 LM 19,8 1 3 54,5 305 ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMll\IE (VIZCAYA) Tabla 50.- Resumen estadístico de las medidas más frecuentes de Bos taurus de Santimamiñe. n var. X s s % M3: 27 30,5-39 36,03 LMP 4 65 -81 70,7 LS 4 55 -68 60,8 AS 2 48 -56 52 LmC 3 49 -65,5 55,1 LMI 4 57 -65,5 62,1 LMm 6 52 -60 El 6 32,5-36 Ad 6 38,5-43 L 1,99 5,5 57,08 2,45 4,3 34,5 1, 15 3,3 40,7 1,49 3,6 8, 1 Escápula: Astrágalo: Falange 1 ant.: LMpe 10 49 -62,5 52,8 4,3 Ap 24 -31 27,7 2,59 9,3 AD 10 10 19,8-27,5 2,32 9,9 Ad 10 23 -31 23.4 26,6 2,27 8,5 Falange 1 post.: LMpe 9 49,5-61 55,5 Ap 9 24 -30 26,9 2,24 AD 9 20,5-24,5 22,5 1,56 6,9 Ad 9 23 -28,5 25,7 1,97 7,6 32,5-41 36,9 2,76 7,5 25 -30,5 27,9 1,82 6,5 19,9-24 21,8 1,59 7,3 20 -27 23,9 1,71 7, 1 3,8 45, 1 8, 1 8,3 Falange 2 ant.: LM Ap AD Ad 10 10 11 11 Falange 2 post.: LM 6 36 -40,5 37.4 1,45 Ap 6 25 -29,5 27,08 1,71 6,3 AD 6 19,7-23,5 21,2 1,48 7,01 Ad 6 21,5-24 22,6 1, 17 5,2 LSD 8 57 -72,5 65,2 4,6 7,05 Ldo 8 44,5-55 50,4 3,52 6,9 Falange 3: PEDRO M. CASTAl\IOS 306 Tabla 51.- Resumen estadístico de las medidas de Capra hircus de Santimamiñe. Escápula: LMP LS AS LmC Húmero: Ad AT Fémur: Ap EC Calcáneo: LM AM Astrágalo: LMI LMm El Ad n var. X 4 4 4 4 30 -43 23,5-35,5 18,1-28 16 -26 36 28,8 24,8 21,4 6 6 28,5-42 27 -39,5 32 30,6 3 3 38,5-46 19,4-19,8 41, 1 19,6 4 4 59 -76,5 19,5-29 69,7 25,2 8 8 8 8 30,5-33 28,5-31 16,9-19 19,9-21,5 31,2 29,7 17,6 20,7 s s% 4,6 4, 1 14,3 13,4 0,75 0,86 0,56 0,51 2,4 2,9 3, 1 2,5 s s% Tabla 52.- Resúmenes estadísticos de las medidas de Ovis a./Capra h. de Santimamiñe. Maxilar: LP 2-M 3 LP2-p4 LM 1-M 3 Mandíbula: LP2-M3 LP2-P4 LM1-M3 LM3 n var. X 2 5 6 63,5-66,5 20,5-22,5 39,5-48,5 65 21,4 44,6 0,8 3,24 3,7 7,2 7 21 11 36 66 -80 17,9-31 44,5-55 20,5-27,5 70,9 23,2 48,8 22,9 4,22 2,89 3,06 1,68 5,9 12,4 6,2 7,3 6 5 38 -44 44,5-45 41,2 44,7 1,82 4,4 5 3 24 -26 18,3-20,5 24,7 19, 1 0,67 2,7 11 12 11 13 35 -41 11,4-13,4 8,4-10,7 10,2-12,7 37,4 12,31 9,5 11,4 1,78 0,68 0,65 0,7 4,7 5,5 6,8 6, 1 8 9 9 7 19,7-24 10,5-12,2 7,2- 8,5 7,7- 9,9 22,08 11,3 8,03 9,07 1, 18 0,63 0,49 0,75 5,3 5,5 6, 1 8,3 Atlas: AScr AScd Tibia: Ad Ed Falange 1: LMpe Ap AD Ad Falange 2: LM Ap AD Ad 307 ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMll\IE (VIZCAYA) Tabla 53.- Resumen estadísticos de las medidas de Sus domesticus del yacimiento de Santimamiñe. Nivel 3 : 1-111 AM 3 : 1-111 LM 1-11 LMJ: 111-IV-S 1-11 AMJ: 111-IV-S s so/o 3, 19 2,44 1,77 2,66 10,7 13,9 5 7,5 1,4 8,2 24,5-33,5 12,1-21 32 -37,5 28,5-38,5 14,7-18,4 14,7-18,9 X 29,7 17,5 34,9 35,3 16,2 17,03 3 5 86,5-88 24 -26 87,5 24,8 0,67 2,7 6 6 6 6 34,5-42,5 40,5-43,5 32,5-39 37,5-40,5 39,8 41,6 36,8 38,5 2,64 1,02 2, 11 1 6,6 2,4 5,7 2,5 5 6 4 5 36 -37 15, 1-16,6 12,4-13,6 14,6-15,7 36,5 15,7 12,9 15,02 0,31 0,59 0,86 3,7 0,36 2,4 6 18 6 19 6 17 6 17 20,5-24,5 22 -25 14,8-19,2 14,7-19,6 12,2-16,4 12,1-15,5 13,1-17,7 12,9-18,4 22,6 23,6 16,5 17,5 13,8 14, 1 15, 1 15,8 1,59 0,85 1,43 1,36 1,44 1, 11 1,58 1,56 7 3,6 1,6 7,7 10,4 7,9 10,4 9,9 n 8 8 5 11 4 10 var. Calcáneo: LM 111-IV-S AM 111-IV-S Astrágalo: 1-11 LMI 111-IV-S 1-11 LMm 111-IV-S Falange 1: LMpe 111-IV-V Ap 111-IV-V AD 111-IV-V Ad Falange 2: · 111-IV-V 1-11 LM 111-IV-S Ap 1-11 llHV-S AD 1-11 111-IV-S Ad 1-11 111-IV-S Tabla 54.- Resumen estadístico de las medidas de Equus ferus del yacimiento de Santimamiñe. p2: L A p3-4: L A I= Pr x 100 L Pr n var. X s s '?fr 9 9 32,5-41 23 -25,8 36,4 24,7 2,24 0,8 6, 15 3,2 15 15 25, 1-35,9 24 -32,5 29,5 27,2 2,5 1,81 8,5 6,6 15 14 12 -16,4 40,7-57,6 13,7 47,02 1,35 4,92 9,9 10,4 308 PEDRO M. CASTAÑOS Tabla 54. (Continuación) Ml-2: L A Pr 16 16 16 16 23,7-28,4 24,6-29,2 11,2-15,6 43, 1-60,2 25,9 26,9 12,9 50, 1 1, 10 1,22 1,32 5,26 4,2 4,5 10,2 10,5 M3: L A 8 8 27,3-31 22,6-24,4 29,8 23,6 1,33 0,67 4,4 2,8 P2: L A 11 11 30,5-37 13,9-18,8 33,06 16,6 1,77 1,75 5,3 10,5 PH: L A 25 25 24,3-31,5 14 -19,3 27,8 16,5 1,89 1,54 6,8 9,3 Ml-2: L A 9 9 22,7-26,8 14, 1-18,3 24,9 16,2 1,35 1,5 5,4 9,2 M3: L A 13 13 26,5-33,5 12 -16, 1 30, 1 14,06 2,48 1,44 8,2 10,2 2 3 2 5 86,5-90 75,5-82 41 -45,5 73 -85 88,25 78,5 43,2 79,5 4,02 5,06 5 39,7-44 41,6 1,56 3,7 4 56 -59 57,3 1,93 2,56 1,05 2,69 3,7 5,05 2,7 2,7 Radio: Ap Asp AD Ad Cp. 111: AM Fémur: EC Calcáneo: LM Astrágalo: HM AM ASd LmT Centrotarsal: AM Tarsal 111: AM Meta podios: 1- Ad Ad.s.a Ed Ad.s.a. 100 Ad 2 113,5-114,5 3 3 3 3 61,5-67,5 64,5-70 53 -59,5 63,5-69,5 63,5 66,5 56,3 65,6 3 53 -60 56,6 2 49 -50 49,5 48,5-54,3 47,5-55,3 37 -40,5 95,2-103,4 51,2 50,8 38;8 98,7 17 15 14 15 114 309 ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMIÑE (VIZCAYA) Tabla 54. (Continuación) Falange 1: LM Ap ASp AD Ad ASd I= AD. 100 3 4 4 4 3 3 3 83 -86,8 54,5-59,4 48, 1-55,2 33,8-38,3 44 -48,6 43, 1-46,5 38,9-41,9 84,5 56,02 50,5 35,3 46,2 44,6 40,6 5 5 5 4 4 50 -51,2 53,6-58,6 43,8-47,8 48,6-51,9 86,5-95,6 50,6 55,2 46,02 49,9 90,3 2 2 49,5-52, 1 31 -33,8 50,8 32,4 LM Falange 2: LM Ap AD Ad I= AD. 100 0,38 1,81 1,67 0,7 3,2 3,6 s s % LM Falange 3: AS a ES a Tabla 55.- Resúmenes estadísticos de las medidas de Capra pyrenaica de Santimamiñe. X n var. 3 3 105-118 33,5-38,5 111,6 35,1 5 4 3 22,5-31,5 44,5-54 67 -77 25,5 48,6 72,1 3,2 12,5 12 5 5 9 21 -26 47 -57 70 -80,5 21,5-32,5 23,04 53 76,4 26,6 1,36 3,42 3,91 3, 12 5,9 6,5 5,6 11,7 5 5 5 3 33,5-43,5 29 -37 22 -28 22,5-23,5 37,1 31,4 25,2 23 3,38 2,95 2,37 9 9,4 9,4 32,5-48 29,5-44 39,1 36,9 3,67 3,41 9,3 9,2 Cuerna: CbC DMB Maxilar: LP2_p4 LM 1-M 3 LP2-M 3 Mandíbula: LP2-M4 LM1-M3 LP2-MJ LM3 Escápula: LMP LS AS LmC Húmero: Ad AT 22 22 PEDRO M. CASTAtiJOS 310 Tabla 55. (Continuación) Astrágalo: LMI LMm El Ad 13 13 13 13 30 -39 28 -37 17, 1-22,5 19,4-26,5 35,9 33,5 20 23,5 3,34 2,9 1,6 2,54 9,3 8,6 8 10,8 4 4 70 -77,5 23 -29 74, 1 25,8 3 10 2 11 43,5-51 15 -20 11 -14,6 14 -17 46,5 16,9 12,8 15,3 1,48 8,7 1,21 7,9 8 8 7 8 25,5-33 13,6-16,8 9,6-12,5 10,9-12,4 28,8 14,8 10,8 11,8 2,6 0,93 0,99 0,65 9,09 6,3 9, 1 5,5 8 8 37 -47 30,5-39 42,5 34,9 3,37 2,94 7,9 8,4 5 30,5-41 35,3 3,4 9,6 35 2,35 6,7 Calcáneo: LM AM Falange 1.ª: LMpe Ap AD Ad Falange 2.ª: LM Ap AD Ad Fala·nge 3.ª: LSD Ldo Metacarpo: Ad Metatarso:, Ad 9 ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMll\IE (VIZCAYA) 311 Tabla 56.- Resumen estadístico de las principales medidas de Rupricapra rupricapra de Santimamiñe. n var. 2 5 2 8 18,5-21,5 40 -48 61 -70 17 -19,4 6 7 7 6 X s so/o 44 3,2 7,2 65,5 17,8 0,72 4 29,5-36,5 25 -29,5 19,8-23,5 19 -21,5 32,8 26,7 21,9 20,01 2,4 1,43 1,31 0,92 7,3 5,3 6 4,6 7 8 30,5-35 28,5-32,5 32,2 30,5 1,62 1,34 5 4,4 3 3 28,5-34,5 27 -32,5 31,8 29,8 Ad 4 26 -31,5 29 Ad 3 25 -28 26,3 8 8 66 -71,5 21,5-24 67,6 22,3 1,6 0,85 2,3 3,8 10 8 9 9 26 -31,5 25 -29,5 14,4-18,2 16,9-21 29,2 27,2 16,2 18,7 1,8 1,45 1, 17 1, 18 6, 1 5,3 7,2 6,3 4 21 -27 24,1 7 21 -30,5 26,9 3,37 12,5 12 14 11 12 36,5-49,5 11,3-14,6 8,4-10,2 10,5-13,4 41,9 12,8 9, 1 11,6 4,5 1, 12 0,55 1,03 10,9 8,7 6, 1 8,9 4 4 3 4 20,5-30 9,4-12,7 6,6- 7,8 8,3-10,4 23,8 10,8 7,2 8,8 4 4 26,5-37,5 22 -31,5 32,2 26,5 Mandíbula: LP2-P4 LM1-M3 LP2-M3 LM3 20 Escápula: LMP LS AS LmC Húmero: Ad AT Radio: Ap ASp Metacarpo: Tibia: Calcáneo: LM AM Astrágalo: LMI LMm El Ad Centrotarsal: AM Metatarso: Ad Falange 1: LMpe Ap AD Ad Falange 2: LM Ap AD Ad Falange3: LSD Ldo PEDRO M. CASTAÑOS 312 Tabla 57.- Resumen estadístico de las medidas más frecuentes de Gran Bóvido de Santimamiñe. Mandíbula: L P2-M3 L P2-P4 L M1-M3 L M3 Húmero: Ad AT Fémur: EC Astrágalo: LMI LMm El Ad Falange 1 ant.: LMpe Ap AD Ad Falange 2 ant.: LM Ap AD Ad Falange 3: LSD Ldo n var. X s so/o 2 3 2 9 150-161 54,5-56,5 93 -116,5 37 -46 155,5 55, 1 104,7 41,9 4,6 11,2 2 3 84,5-88,5 79,5-116, 1 86,5 92,6 5 58 -66 60,3 2,9 4,8 5 5 5 5 65 -94,5 59,5-91,5 36 -56 43,5-62 76,04 71,8 43,3 51 12,2 13, 1 7,71 7,34 16 18,2 17,8 14,4 4 4 4 4 60,5-75 29 -41,6 25,5-37,7 27,5-43, 1 69,6 36,6 33,6 37,6 5 5 5 4 37,5-52 26,5-48,8 21,8-40,7 23 -41,9 47,9 37,7 30,5 30,9 8, 17 9,04 7,35 17 23,9 24, 1 4 4 79 -95 58 -74,5 88,3 67 Tabla 58.- Resúmenes estadísticos de las medidas más frecuentes de Cervus e/aphus de Santimamiñe. n Cráneo: ACO AFM Maxilar: L P2-P4 L M1-M3 Mandíbula: L P2-P4 L M1-M3 L P2-M3 L M3 8 8 var. Paleolítico X s s % 74 -87 28,5-34 80,3 32,1 4,51 3,09 5,6 9,6 4 18 46,5-49,5 63,5-76 48 71,2 3, 13 4,4 21 24 2 58 41,5-53,5 66,5-84,5 124 -128,5 27,5-36,5 47,8 72,2 126,2 31,5 2,8 4,43 5,8 5,7 2,09 6,6 n Postpaleolítico var. X s so/o 70,5 30,5 7 2 36 -46,5 73 42,8 73 3,28 7,6 15 8 4 29 29 -54 67 -83,5 107 -141,5 23,5-35 43,7 75,4 121,3 29,6 5,74 5,88 13, 1 7,8 2,6 8,7 313 ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMll\JE (VIZCAYA) Tabla 58. (Continuación) Axis: 4 4 65 -77 27,5-37 69,5 33,8 32 35 37 27 55,5-67 40,5-55 38 -55 31 -43 61, 1 48, 1 43,8 36,6 3,7 3,67 4, 15 3,33 33 35 46 -70 44 -62,5 61,06 55,6 16 16 15 55,5-69,5 52 -63,5 44 -60 9 Ap 3 Ad 35 AScr AD e 4 58,5-64,5 62 28 6,06 7,6 9,4 9, 1 9 11 10 51 -64,5 40,5-49,5 37,5-44 30,5-36,5 58,4 46,2 41, 1 34,4 3,87 2,71 2,23 2,2 6,6 5,8 5,4 6,3 4,9 3,99 8,02 7, 1 32 31 45,5-74 43,5-73 54,2 50,2 6,64 5,99 12,2 11,9 62,3 57,9 53,2 4,07 3,56 4,33 6,5 6, 1 8, 1 7 4 4 47,5-60,5 45,5-55,5 44 -55,5 54,6 49, 1 48,1 4,61 8,4 27 -35 31,7 2,57 8, 1 2 27 -36 31,5 46 -47 37,5-53,5 46,5 44,2 3,67 8,3 4 17 44 -50 36 -49,5 47,2 42,4 4,48 10,5 40 -63,5 36,5-53,5 57,5 51, 1 7,94 6,61 13,8 12,9 5 5 45 -62,5 40,5-52,5 55, 7 48, 1 6,38 4,32 11,4 8,9 32 -43 72 -85 36,8 77,3 3,44 4, 14 9,3 5,3 7 29,5-44 63 -71,5 38,5 67,7 4,92 2,81 12,8 4,1 78 -95,5 51 -60 87,3 54,05 5,99 3,79 6,8 7,02 2 15 79,5-86 43 -57,5 82,7 51,2 5,77 11,2 48 -64 31,5-56 56,08 46,4 5,74 7,93 10,2 17, 1 4 46. -54,5 41 -46 52,2 43,3 6,84 3,07 5,6 7,6 41 44 111,8 35,8 5,79 2,57 5, 1 7, 1 7,3 6,9 7,03 8,2 129 128 132 131 47 -65,5 43,5-60,5 25,5-36,5 27,5-40,5 52,3 48,8 28,8 33, 1 3,4 2,95 1,98 2,36 6,5 6,04 6,9 7, 1 Escápula: LMP LS AS LmC 5 Húmero: Ad AT Radio: Ap ASp Ad Ulna: APC Metacarpo: Pelvis: LA 6 AA 6 Fémur: EC 14 Ad 6 Ap 7 17 5 Tibia: Ad Rótula: LM AM Calcáneo: LM AM Astrágalo: LMI LMm El Ad Centrotarsal: AM Metatarso: Ap Ad 6 6 3 16 18 114 -124,5 35 -47,5 45 42 43 43 48 -69,5 44,5-63,5 26,5-38 29,5-41,5 56,04 4, 12 52,4 3,65 30,6 2, 15 35,8 2,94 20 37,5-53,5 45,9 4, 18 9, 1 22 35 -53,5 42 5,3 12,6 39 -47 42 -54 42,5 45,7 2,65 3,08 6,2 6,7 3 40,5-41,5 38,5-48,5 41 43,8 3,28 7,5 6 40 121,9 40, 1 6 101,5-125 31 -44 PEDRO M. CASTAÑOS 314 Tabla 58. (Continuación) Falange 1: LMpe Ap AD Ad 20 36 19 54 51 -65,5 16,8-30,5 16,1-21 18 -24,5 60,8 22,4 18,5 22,03 3,63 2,42 0,98 1,71 5,9 10,8 5,3 7,7 7 23 6 34 49,5-63 18,2-25,5 14,7-19,6 16,7-26 55,8 21, 1 17,5 20,1 4,89 2,07 2,01 2,23 8,7 9,8 11,5 11, 1 17 32 17 25 38,5-48,5 19,6-25,5 14, 1-18,8 16,7-20,5 44,6 22,5 16,7 18,9 2,67 1,52 1,38 1,23 6 6,7 8,3 6,5 10 31 12 12 36,5-44,5 16,7-24 13,3-17,2 15,3-21 39,6 20,5 15, 1 18,01 2,56 2, 18 1,28 2, 18 6,4 10,6 8,4 12, 1 72 72 37 -61 34,5-55,5 50,7 46,7 5,31 4,56 10,4 9,7 202 199 35 -58 32,5-53 45,7 42,4 4,33 4,04 9,4 9,5 Falange 2: LM Ap AD Ad Falange 3: LSD Ldo Tabla 59.- Resumen estadístico de las medidas de Capreo/us capreo/us del yacimiento de Santimamiñe. Maxilar: LP2_p4 LM 1-M 3 LP 2-M 3 n var. X s s% 9 12 6 26 -29 30 -34,5 55,5-60 27,8 32,3 57,5 0,94 1,67 1,88 3,3 5, 1 3,2 20 12 3 27 26 -30,5 38,5-43,5 68,5-72 14,2-17,6 28,5 40,5 70,3 16,05 1,29 1,63 4,5 4,03 0,98 6,1 5 4 4 4 28,5-32 22,5-25,5 20,5-23 18, 1-20 30,5 24,1 22,2 18,8 1,34 4,3 13 13 28 -32,5 25 -31 29,8 27,3 1,36 1,66 4,5 6, 1 3 3 27,5-31,5 25,5-30,5 29 27,3 4 4 2 26,5-28,5 23,5-24,5 17,2-18,4 27,7 23,8 17,8 8 23 -25,5 24,06 0,88 3,6 Mandíbula: LP2-P4 LM1-M3 LP2-M3 LM3 Escápula: LMP LS AS LmC Húmero: Ad AT Radio: Ap Asp Ulna: EPA EmO APC Metacarpo: Ad ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMIÑE (VIZCAYA) 315 Tabla 59. (Continuación) Calcáneo: LM AM Astrágalo: LMI LMm El Ad 63,5-73 20,5-24 66,6 21,9 2,86 0,93 4,2 4,2 7 6 7 7 28,5-32,5 27 -31 16 -17,9 17,6-20,2 30,4 28,6 17,01 18,9 1,59 1,57 0,83 0,88 5,2 5,4 4,8 4,6 5 23 -26,5 25,2 1,28 5, 1 3 6 3 4 35,5-36,5 10,8-12,6 7,8- 8,5 9,4-10,4 36 11,8 8,06 9,8 0,62 5,2 8 8 8 8 23 -29 10,3-11,4 7,2- 8,8 7,4- 8,9 25,3 11,2 7,7 8,01 1,85 0,71 0,52 0,52 7,3 6,3 6,7 6,5 26 25 24 -29,5 22,5-29,5 27 26,1 1,24 1,48 4,6 5,7 12 12 Ce ntrota rsa 1: AM Falange 1: LMpe Ap AD Ad Falange 2: LM Ap AD Ad Falange 3: LSD Ldo Tabla 60.- Resumen estadístico de medidas de Capreo/us capreolus preneolíticos de la Península Ibérica. n var. X s so/o 3 5 29,8-32,5 30,7 35,9 2,9 8, 1 12 28 27,6-31 11,3-17,6 29,5 16, 1 0,95 1,25 3,2 7,7 4 4 4 27 -31,3 22 -24,2 21,7-23,2 29,2 23 22,3 22 16 27 -33,4 25 -30,9 30,7 28,09 1,86 1,58 6 5,6 Ad 6 25 -29,2 27,4 1,5 5,7 Ad 2 29,9-31,5 30,7 Maxilar: LP2_p4 LM 1-M 3 Mandíbula: LP2-P4 LM3 Escápula: LMP LS AS Húmero: Ad AT Radio: Tibia: PEDRO M. CASTAl'\JOS 316 Tabla 60. (Continuación) Astrágalo: 18 22 22 19 29,5-34,2 28 -32,8 16, 1-19, 1 18,7-22,3 32,08 30,7 17,9 20,6 1,2 1,08 0,76 1,05 3,7 3,5 4,2 5, 1 16 59,2-73,5 67,3 3,9 5,8 AD 24 31 23 Ad 44 32,5-42 10,6-14, 1 7,4-10,2 9,5-12, 1 36,9 12, 1 8,7 10,5 2,45 0,83 0,8 0,79 6,6 6,8 9,2 7,5 16 17 14 23,7-30,3 9,7-13 7,8- 9,2 27,1 11,4 8,3 2,4 1,05 0,37 9, 1 9,2 4,4 7 6 29,4-28,7 22,9-28,5 26,2 25,1 1,42 2,2 5,4 8,8 LMI LMm El Ad Calcáneo: LM Falange 1: LMpe Ap Falange 2: LM Ap Ad Falange 3: LSD Ldo Tabla 61.- Resumen estadístico de medidas de Capreo/us capreo/us neo y postneolítico de la Península Ibérica. n var. X 2 2 3 56,5-58 27 -29 31 -33,5 57,25 28 32,6 3 5 4 3 64,8-68 26,4-30 36 -41 14,2-16,6 66,1 28,08 38,05 15,2 2 3 3 3 26,5-28,5 17 -23,5 19 -21 14 -18,8 27,5 20,8 20, 1 16,9 6 5 26 -28 21,4-24,5 27,4 23,6 4 3 23,8-28 25 -25,5 26,2 25, 1 s so/o Maxilar: LP 2-M 3 LP2-p4 LM 1-M 3 Mandíbula: LP2-M3 LP2-P4 LM1-M3 LM3 1, 14 4,06 0,66 1, 14 2,4 4,8 Escápula: LMP LS AS LmC Húmero: Ad AT Radio: AP ASp ESTUDIO DE LOS MACROMAMIFEROS DE LA CUEVA DE SANTIMAMll'ilE (VIZCAYA) 317 Tabla 61. (Continuación) Metacarpo: 3 21,9-23,5 22,6 Ad Calcáneo: 2 26 -28 27 LM AM 2 2 60,2-61,5 20 60,8 20 5 5 5 5 27,3-30 26,6-28,5 16, 1-17,3 15,9-19,2 28,8 27,6 16,6 18,3 1,05 0,74 0,46 1,22 3,6 2,7 2,7 6,6 13 13 13 3 32,2-40 11,5-13,5 8, 1- 9,5 10,5-11,2 37,7 12,5 8,8 10,9 2,06 0,69 0,37 5,4 5,5 4,2 7 7 7 3 21,8-28 9 -18,8 7 - 9, 1 5 - 9 25,2 11,9 8,05 7,6 2,79 3 0,66 11 25,2 8,2 Ad Tibia: Astrágalo: LMI LMm El Ad Falange 1: LMpe Ap AD Ad Falange 2: LM Ap AD Ad Tabla 62.- Resumen estadístico de medidas más frecuentes de Sus ferus del yacimiento de Santimamiñe. n var. X s 9 8 4 32,5-37,5 18,5-22,5 73 -78,5 36,1 20 75,8 1,47 1,3 4,07 6,5 8 8 36 -44 16,6-22 40,06 18,6 2,15 1,47 5,3 7,9 2 2 3 3 39 -50 32 -41,5 25 -34 27,5-34,5 44,5 36,7 29,6 29,8 Ad AT Fémur: 8 9 40 -54,5 32,5-44,5 48,5 39,1 4,2 3,84 8,6 9,8 Ad 2 56,5-59 57,7 Ad 6 29 -41,5 35,5 3,64 10,2 so/o Maxilar: LM 3 AM 3 LM 1-M 3 Mandíbula: LM3 AM3 Escápula: LMP LS AS LmC Húmero: Tibia: PEDRO M. CASTAi\JOS 318 Tabla 62. (Continuación) Calcáneo: LM AM 9 11 91,5-101 24,5-29,5 93,9 20 3 1,29 3, 1 4,8 45 44 39,5-53,5 37 -48,5 47,8 43,5 2,87 2,52 6 5,8 4 7 3 3 37 -44,5 14,9-19,7 11,7-15 14,6-17,4 40,3 18,24 13,2 15,6 36 37 36 36 22,5-30 15, 1-20,5 12 -17,5 14 -20 26,8 18,5 15,2 16,7 1,72 1,26 1, 13 1,56 6,4 6,8 7,4 8,8 28 28 35 -41,5 34 -43 39, 1 37,9 1,94 2,14 4,9 5,6 Astrágalo: LMI LMm Falange 1: LMpe Ap AD Ad 2,01 11 Falange 2: LM Ap AD Ad Falange 3: LSD Ldo