histología del tejido
Anuncio

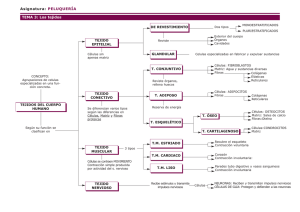
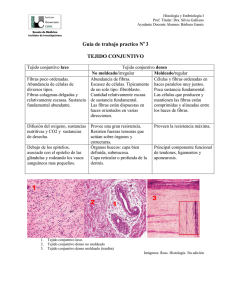
M. Ruth Álvarez Quintillán. HISTOLOGÍA DEL TEJIDO: Tejido epitelial, muscular, nervioso y conjuntivo. Con la denominación de “tejido”, se conoce a la agrupación de células morfológicamente iguales y con un trabajo en común. Dos o más tejidos forman unidades funcionales mayores, los órganos, que según su funcionalidad se agrupan formando aparatos y sistemas, cuya unión da lugar al individuo. Todos los tejidos se forman a partir de las tres capas primitivas de células del embrión, llamadas hojas embrionarias: la hoja externa o ectodermo, la media o mesodermo y la interna o endodermo. El estudio del tejido puede efectuarse según su origen, pero lo habitual es hacerlo basándose en su morfología y función. De esta manera, se distinguen cuatro tipos de tejido: -Tejido epitelial, cuya función es recubrir y segregar sustancias. Derivan de las tres hojas embrionarias. -Tejido muscular, que es el responsable de la movilidad de los órganos o del individuo. Derivan del mesodermo. -Tejido nervioso, que se encarga de la percepción de estímulos, transmitiéndolos y elaborando una respuesta frente a ellos. Deriva del ectodermo. -Tejido conjuntivo (o conectivo), une, envuelve y refuerza a los demás tejidos, además de transportar vasos y nervios, regular la temperatura e intervenir en la autodefensa. TEJIDOS EPITELIALES. El tejido epitelial es la variedad de tejido básico o primario constituido por agrupaciones de células situadas en forma adyacente, fuertemente adheridas entre si, con escasa matriz extracelular y relacionada con el tejido conjuntivo a través de la membrana basal. Carecen de vasos sanguíneos, linfáticos y de fibras nerviosas, siendo su origen embriológico a partir de cualquiera de las tres hojas embrionarias. Las poblaciones celulares epiteliales, presentan una renovación constante debido a lo cual encontramos en ellos células indiferenciadas que pueden actuar como células madres y células diferenciadas, especializadas en las funciones que le son propias. Es de destacar que cualquier tipo de célula epitelial, presenta dos propiedades que lo caracterizan la cohesión y la polaridad. La cohesión, está dada por la tendencia que tienen a estar fuertemente adheridos entre si, lográndose ello por las especializaciones de las superficies celulares, y la polaridad se manifiesta por la presencia de una superficie basal, adjunta al tejido conjuntivo y una superficie apical libre o secretora que da a la superficie ó a la luz de un órgano, destacándose la disposición particular y estable de los organitos citoplasmáticos y de las especializaciones de la superficie celular. Por la disposición, estructura y función de las células epiteliales, este tejido se divide en dos grandes grupos. El primero lo constituyen los epitelios o membranas epiteliales de cubierta y revestimiento, que son capas de células especializadas en funciones de protección, absorción e intercambio y el segundo los epitelios glandulares, que son masas o agrupaciones celulares, especializados en la secreción. Epitelios de cubierta y revestimiento: Los epitelios de cubierta y revestimiento presentan una o varias capas de células y diferenciaciones estructurales, como consecuencia de sus diversas especializaciones funcionales. Están constituidos, casi totalmente, por células poliédricas íntimamente unidad con escasa matriz extracelular. Están separados del tejido conjuntivo por una estructura denominada membrana basal, que no se colorea con la coloración de Hematoxilina y Eosina; sin embargo, se distingue fácilmente al M/O con la técnica de PAS (ácido peryódico de Schiff) y con impregnación argéntica. Al M/E esta membrana si es visible. Siempre están relacionadas con una capa de tejido conjuntivo subyacente que le ofrece soporte, sostén, nutrición, irrigación, drenaje y defensa. Reciben las sustancias nutritivas por difusión del líquido tisular proveniente de los vasos sanguíneos del tejido conjuntivo subyacente. Están inervados por terminaciones nerviosas libres provenientes también de las fibras nerviosas procedentes del tejido conjuntivo, las cuales atraviesan la membrana basal y cursan entre las células epiteliales. La clasificación morfológica de los epitelios se basa fundamentalmente en tres criterios. El primero de ellos atienden al número de capas que incluye: si presenta una sola capa de células el epitelio es simple, y si posee dos o más capas se clasifican, como estratificados. Cuando el epitelio tiene una sola capa de células, pero da la impresión de poseer más de una, se considera seudoestratificado, aunque en realidad es una variedad de epitelio simple. El segundo criterio que se utiliza para clasificar a los epitelios es la forma que presentan las células, siendo calificadas como, planas (pavimentosas), cúbicas y cilíndricas (prismáticas), según el aspecto que estas presentan en los cortes perpendiculares a la superficie de la membrana. El tercero es por la presencia de especializaciones en la superficie apical (microvellosidades, cilios), la presencia de células acompañantes(caliciformes) y la presencia o no, de queratina M. Ruth Álvarez Quintillán. (queratinizados o no queratinizados). Así distinguimos entre los distintos tipos de epitelio de revestimiento: -Epitelio simple plano, formado por una sola capa de células planas, que reviste internamente vasos (endotelios) y cavidades celómicas (mesotelios). -Epitelio simple cúbico, formado por una sola capa de células cúbicas, que recubre los conductos glandulares (salivares, pancreáticos). -Epitelio simple cilíndrico, formado por una sola capa de células cilíndricas, recubriendo gran parte del tubo digestivo, útero y trompas uterinas (en este caso, es además, ciliado) -Epitelio estratificado plano, las células de las capas más profundas son cúbicas y se aplanan a medida que suben a la superficie, como en la epidermis (queratinizado), epitelio vaginal y esofágico (no queratinizado). -Epitelio estratificado cúbico, solo se observa en los canales excretores de las glándulas salivares, sudoríparas y pancreáticas. -Epitelio estratificado cilíndrico, las células profundas son cúbicas y las superficiales, cilíndricas, como en la uretra esponjosa. -Epitelio de transición o urotelio, formado entre dos a seis capas, de forma irregular, en las vías urinarias de los mamíferos. Está entre el epitelio cúbico y el plano estratificado: las células superficiales son grandes, redondas y binucleadas. -Epitelio pseudoestratificado, las células descansan en la membrana basal, pero sus núcleos están a diferentes alturas, como en el epitelio respiratorio. Epitelios glandulares: Definimos como glándula a la célula o conjunto de células cuya función es secretar una sustancia determinada, siendo la secreción un producto celular específico sintetizado a partir de biomoléculas simples (hidratos de carbono, aminoácidos, lípidos) obtenidas de la sangre. De acuerdo al "medio" al que se vierte la secreción, las glándulas pueden ser: -Endócrinas: si el producto tiene como destino la sangre -Exócrinas: si el producto tiene como destino el medio externo o una cavidad que contacta con el él. -Mixtas: si la misma glándula genera diferentes sustancias, algunas endócrinas y otras exócrinas. TEJIDOS MUSCULARES: El tejido muscular, es un tejido que está formado por las fibras musculares o miocitos. Compone aproximadamente entre el 40% y 45% de la masa de los seres humanos y está especializado en la contracción, lo que permite que se muevan los seres vivos. Como las células musculares están altamente especializadas, sus orgánulos necesitan nombres diferentes. La célula muscular en general se conoce como fibra muscular; el citoplasma como sarcoplasma; el retículo endoplásmico liso, retículo sarcoplásmico liso y en ocasiones las mitocondrias, sarcosomas. A la unidad anatómica y funcional se la denomina sarcómero. Debido a que las células musculares son mucho más anchas que largas, a menudo se llaman fibras musculares; pero por esto no deben ser confundidas con la sustancia intercelular forme, es decir las fibras colágenas, reticulares y elásticas; pues estas últimas no están vivas, como la célula muscular. Los tres tipos de músculo derivan del mesodermo. El músculo cardíaco tiene su origen en el mesodermo esplácnico, la mayor parte del músculo liso en los mesodermos esplácnico y somático y casi todos los músculos esqueléticos en el mesodermo somático. Consta de tres elementos básicos: fibras musculares, que suelen disponerse en haces o fascículos, una abundante red capilar y tejido conectivo fibroso de sostén con fibroblastos y fibras colágenas y elásticas. Éste actúa como sistema de amarre y acopla la tracción de las células musculares para que puedan actuar en conjunto. Además conduce los vasos sanguíneos y la inervación propia de las fibras musculares. De acuerdo con sus características morfológicas y funcionales se pueden diferenciar en los mamíferos tres tipos de tejido muscular: -Músculo estriado o esquelético: Está formado por haces de células muy largas (hasta de 30 cm.) cilíndricas y multinucleadas, con diámetro que varía de 10 a 100 µm., llamadas fibras musculares estriadas. Las fibras musculares están organizadas en haces envueltos por una membrana externa de tejido conjuntivo, llamada empimisio. De éste parten septos muy finos de tejido conjuntivo, que se dirigen hacia el interior del músculo, dividiéndolo en fascículos, estos septos se llaman perimisio. Cada fibra muscular está rodeada por una capa muy fina de fibras reticulares, formando el endominsio. El tejido conjuntivo mantiene las fibras musculares unidas, permitiendo que la fuerza de contracción generada por cada fibra individualmente actúe sobre el músculo entero, contribuyendo así a su contracción. Este papel del tejido conjuntivo tiene gran importancia porque las fibras generalmente no se extienden de un extremo a otro del músculo. También por intermedio del tejido conjuntivo la fuerza de contracción del músculo se transmite a otras estructuras como tendones ligamentos, aponeurosis y huesos. Los vasos sanguíneos penetran en el músculo a través de los septos del tejido conjuntivo y forman una red rica en capilares distribuidos paralelamente a las fibras musculares. Estas fibras se adelgazan en las extremidades y se observa una transición gradual de músculo a tendón. Estudios en esta región de M. Ruth Álvarez Quintillán. transición al microscopio electrónico reveló que las fibras de colágena del tendón se insertan en pliegues complejos del sarcolema presente en esta zona. Cada fibra muscular presenta cerca de su centro una terminación nerviosa llamada placa motora. La fibra muscular está delimitada por una membrana llamada sarcolema y su citoplasma se presenta lleno principalmente de fibrillas paralelas, las miofibrillas. Las miofibrillas son estructuras cilíndricas, con un diámetro de 1 a 2 µm. y se distribuyen longitudinalmente a la fibra muscular, ocupando casi por completo su interior. Al microscopio se observan estriaciones transversales originadas por la alternancia de bandas claras y oscuras. La estriación es debida a repetición de unidades llamadas sarcómeros. Cada unidad está formada por la parte de la miofibrilla que queda entre dos líneas Z y contiene una banda A. -Músculo cardíaco: Constituido por células alargadas, formando columnas que se anastomosan irregularmente. Estas células también presentan estriaciones transversales, pero pueden distinguirse fácilmente de las fibras musculares esqueléticas por el hecho de poseer solo uno o dos núcleos centrales. La dirección de las células cardíacas es muy irregular y frecuentemente se pueden encontrar con varias orientaciones, en la misma área de una preparación microscópica, formando haces o columnas. Esas columnas están revestidas por una fina vaina de tejido conjuntivo, equivalente al endomisio del músculo esquelético. Hay abundante red de capilares sanguíneos entre las células siguiendo una dirección longitudinal a éstas. La célula muscular cardiaca es muy semejante a la fibra muscular esquelética , aunque posee más sarcoplasma, mitocondrias y glucógeno. También llama la atención el hecho de que en los músculos cardiacos, los filamentos ocupen casi la totalidad de la célula y no se agrupen en haces de miofibrillas. Una característica específica del músculo cardiaco es la presencia de líneas transversales intensamente coloreables que aparecen a intervalos regulares. Estos discos intercalares presentan complejos de unión que se encuentran en la interfase de células musculares adyacentes. Son uniones que aparecen como líneas rectas o muestran un aspecto en escalera. En la parte en escalera se distinguen dos regiones. La parte transversal, que cruza la fibra en línea recta y la parte lateral que va en paralelo a los miofilamentos. En los discos intercalares se encuentran tres tipos de contactos: la fascia adherens o zona de adhesión, mácula adherens o desmosome y uniones tipos gap (gap juntion). Las zonas de adhesión representan la principal especialización de la membrana y de la parte transversal del disco sirven para fijar los filamentos de actina de los sarcómeros terminales. Básicamente representa una hemibanda Z (media). Las máculas adherentes son desmosome que unen fibras musculares cardiacas, impidiendo que se separen por la actividad contráctil constante del corazón. Los desmosomas son estructuras complejas en forma de un disco constituidas por la yuxtaposición de dos regiones electrodensas que se hallan en las regiones contiguas de la membrana celular de dos células vecinas, en las cuales se insertan haces de tono filamentos. Las fibrillas tienden acumularse en el polo superior de la célula inmediatamente por debajo de la superficie celular, formando la trama terminal (citoesqueleto). En las partes laterales de los discos se encuentran uniones tipo gap, responsables de la continuidad iónica, entre células musculares próximas. Desde el punto de vista funcional, el paso de iones permite que las cadenas de células musculares se comportan como si fueran un sincito (célula simple con muchos núcleos), pues el estímulo de la contratación pasa como si fuera una onda de una célula a otra. Debido a la capa de tejido conjuntivo que reviste internamente el corazón existe una red de células musculares cardiacas modificadas localizadas dentro de la pared muscular del órgano. Tales células desempeñan un papel importante en la generación y conducción del estímulo cardiaco. El corazón recibe nervios tanto del sistema simpático con del parasimpático que forman plexos en la base del órgano. No existen en el corazón, terminaciones nerviosas comparables a la placa motora del músculo esquelético. Se admite que las fibras musculares cardiacas son capaces de autoestimulación independiente del impulso nervioso. Cada una de estas fibras tiene su ritmo propio, pero dado que están enlazadas en uniones tipo gap, que tienen un ritmo acelerado y conducen a todas las otras distribuyendo el impulso a todo el órgano. Las fibras del sistema generador y conductor del impulso son las del ritmo más rápido, pero las otras células del corazón pueden hacer que el órgano trabaje con un ritmo más lento, en el caso de que exista un fallo en el sistema conductor. Por lo tanto el sistema nervioso ejerce en el corazón una acción reguladora, adaptando el ritmo cardiaco a las necesidades del organismo como un todo. -Músculo visceral o liso: Esta formado por la asociación de células largas que pueden medir de 5 a 10 um. de diámetro por 80 a 200 µm. de largo. Están generalmente dispuestas en capas sobre todo en las paredes de los órganos huecos, como el tubo digestivo o vasos sanguíneos. Además de esta disposición encontramos células musculares lisas M. Ruth Álvarez Quintillán. en el tejido conjuntivo que reviste ciertos órganos como la próstata y las vesículas seminales y en el tejido subcutáneo de determinadas regiones como el escroto y los pezones. También se pueden agrupar formando pequeños músculos individuados (músculo erector del pelo), o bien constituyendo la mayor parte de la pared del órgano, como el útero. Las fibras musculares lisas están revestidas y mantenidas unidad por una red muy delicada de fibras reticulares. También encontramos vasos y nervios que penetran y ramifican entre las células. En el corte transversal el músculo liso se presenta como un aglomerado de estructuras circulares o poligonales que pueden ocasionalmente presentar un núcleo central. En corte longitudinal se distinguen una capa de células fusiformes paralelas. La fibra muscular lisa también está revestida por una capa de glucoproteína amorfa (glucálix). Frecuentemente los plasmalemas de dos células adyacentes se aproximan mucho formando uniones estrechas y gap. Esas estructuras no sólo participan de la transmisión intercelular del impulso, sino que mantienen la unión entre las células. Existe un núcleo alargado y central por célula. La fibra muscular lisa presenta haces de miofilamentos que cruzan en todas direcciones, formando una trama tridimensional. En el músculo liso también existen terminaciones nerviosas, pero el grado de control de la contracción muscular por el sistema nervioso varia. Es importante las uniones gap, en la transmisión del estímulo de célula a célula. El músculo liso, recibe fibras del sistema nervioso simpático y para simpático y no muestra uniones neuromusculares elaboradas (placas motoras). Frecuentemente los axones terminan formando dilataciones del tejido conjuntivo. Estas dilataciones contienen vesículas sinápticas con los neurotransmisores acetilcolina (terminaciones colinérgicas) o noradrenalina (terminaciones adrenérgicas). Una de las dificultades más comunes en la identificación de tejidos es la de distinguir el músculo liso del tejido conectivo. Las fibras musculares son celulares y suelen teñirse más intensamente con la eosina que las fibras colágenas. Los núcleos están situados dentro de las fibras, y pueden verse arrugados y son más grandes que los núcleos de los fibroblastos, que están situados entre las fibras colágenas. Con ciertas técnicas de tinción, por ejemplo la de Mallory y la de van Gieson, se pueden distinguir fácilmente estos dos tejidos. TEJIDO NERVIOSO: El tejido nervioso se desarrolla a partir del ectodermo embrionario. Es un tejido formado por dos tipos celulares: neuronas y glía, y cuya misión es recibir información del medio externo e interno, procesarla y desencadenar una respuesta. Es también el responsable de controlar numerosas funciones vitales como la respiración, digestión, bombeo sanguíneo del corazón, regular el flujo sanguíneo, control del sistema endocrino… Las células del sistema nervioso se agrupan para formar dos partes: el sistema nervioso central que incluye el encéfalo y la médula espinal, y el sistema nervioso periférico formado por ganglios, nervios y neuronas diseminados por el organismo. Y en cuanto a estas células, también son de dos tipos: neuronas o células nerviosas y neuroglia o células de sostén. -Neuronas: Son las células funcionales del tejido nervioso. Ellas se interconectan formando redes de comunicación que transmiten señales por zonas definidas del sistema nervioso. Los funciones complejas del sistema nervioso son consecuencia de la interacción entre redes de neuronas, y no el resultado de las características específicas de cada neurona individual. En cada neurona existen cuatro zonas diferentes: el pericarion que es la zona de la célula donde se ubica el núcleo, las dendritas que son numerosas y aumentan el área de superficie celular disponible para recibir información desde los terminales axónicos de otras neuronas, el axón que nace único y conduce el impulso nervioso de esa neurona hacia otras células ramificándose en su porción terminal (telodendrón) y uniones celulares especializadas llamadas sinapsis, ubicadas en sitios de vecindad estrecha entre los botones terminales de las ramificaciones del axón y la superficie de otras neuronas. Cada zona de las células nerviosas se localiza de preferencia en zonas especializadas del tejido nervioso. Los cuerpos celulares, la mayor parte de las dendritas y la arborización terminal de una alta proporción de los axones se ubican en la sustancia gris del SNC y en los ganglios del SNP. Los axones forman la parte funcional de las fibras nerviosas y se concentran en los haces de la sustancia blanca del SNC; y en los nervios del SNP. Según el número y la distribución de sus prolongaciones, las neuronas se clasifican en: bipolares, que además del axón tienen sólo una dendrita; se las encuentra asociadas a receptores en la retina y en la mucosa olfatoria; seudounipolares, desde las que nace sólo una prolongación que se bifurca y se comporta funcionalmente cono un axón salvo en sus extremos ramificados en que la rama periférica reciben señales y funcionan como dendritas y transmiten el impulso sin que este pase por el soma neuronal (es el caso de las neuronas sensitivas espinales) y multipolares desde las que, además del axón, nacen desde dos a más de mil dendritas lo que les permite recibir terminales axónicos desde múltiples neuronas distintas. La mayoría de las neuronas son de este tipo. Un caso extremo do lo constituye la célula de M. Ruth Álvarez Quintillán. Purkinje que recibe más de 200.000 terminales nerviosos. -Células de sostén: En el tejido nervioso del SNC, por cada neurona hay entre 10 a 50 células de neuroglia y que a diferencia de las neuronas retienen su capacidad de proliferar. Existen 4 clases de células de neuroglia: -Astrocitos: son las mayores células de la neuroglia y se caracterizan por la riqueza y dimensiones de sus prolongaciones citoplasmáticas que se dirigen en todas direcciones. Los astrocitos poseen núcleos esféricos y centrales. Entre sus prolongaciones muchas aumentan de grosor en sus porciones terminales, formando dilataciones que envuelven la pared endotelial de los capilares sanguíneos. Estas dilataciones se llaman pies vasculares de la neuroglia. Los astrocitos orientan sus prolongaciones en el sentido de la superficie de los órganos del SNC donde forman una capa. Los astrocitos protoplasmáticos poseen citoplasma abundante y con prolongaciones no tan largas como los astrocitos fibrosos. Estas prolongaciones son muy ramificadas y gruesas. Se localizan sólo en la sustancia gris de SNC. Algunos astrocitos de pequeño tamaño se sitúan cerca de las neuronas formando las células satélites. Los astrocitos fibrosos presentan prolongaciones largas lisas y delgadas que no se ramifican con frecuencia. Están en la sustancia blanca del encéfalo y médula espinal. Los astrocitos mixtos se encuentran en la zona de transición de la sustancia blanca y la sustancia gris, presentan en la misma célula prolongaciones fibrosas, protoplasmáticas, las fibrosas se dirigen a la sustancia blanca y las protoplasmáticas a la sustancia gris. -Oligodendrocitos: Son más pequeños y con menos prolongaciones que la astroglía. Su núcleo es rico en heterocromatina y su citoplasma contiene ergastoplasma, polirribosomas libres, un aparato de Golgi desarrollado y un alto contenido n microtúbulos, tanto en el citoplasma que rodea al núcleo como en sus prolongaciones. Su función más notable es la formación de la mielina, que rodea a los axones del SNC. En la sustancia blanca los oligodendrocitos se disponen en hileras entre las fibras mielínicas. -Células ependimarias: Forman un tipo de epitelio monostratificado que reviste las cavidades internas del SNC que contienen al líquido cefalo raquídeo (ventrículos y conducto del epéndimo). Se unen entre sí por complejos de unión similares a los epiteliales pero carecen de zona de oclusión, de modo que el liquido cefaloraquídeo se comunica con los espacios intercelulares existentes entre las células nerviosas y las glías. Presentan además largas prolongaciones en su zona basal que se asocian a las prolongaciones de la astroglia y en su superficie apical presenta microvellocidades y cilios. -Microglía: Se caracterizan por ser pequeñas, con un denso núcleo alargado y prolongaciones largas y ramificadas. Contienen lisosomas y cuerpos residuales. Si bien se la clasifica generalmente como célula de la neuroglia ellas presentarnel antígeno común leucocítico y el antígeno de histocompatibilidad clase II, propio de las células presentadoras de antígeno. En el tejido nervioso del SNP, tanto las neuronas, en los ganglios, como los axones ubicados en las fibras nerviosas, están rodeadas por de células de sostén. -Celulas de Schwann: se originan de la cresta neural y acompañan a los axones durante su crecimiento, formando la vaina que cubre a todos los axones del SNP desde su segmento inicial hasta sus terminaciones. Ellas son indispensables para la integridad estructural y funcional del axón. -Celulas satélites o capsulares: son células pequeñas localizadas en los ganglios, alrededor del pericaron las dendritas y terminales axónicos. Están rodeadas por lámina basal y separan a las células nerviosas del estroma fibrocolagenoso presente en lel tejido propio del SNP. -Sustancia blanca y sustancia gris: En el SNC se distinguen la sustancia blanca y la sustancia gris. La primera formada por fibras mielínicas, oligodendrocitos, astrocitos fibrosos y células de microglía. En la sustancia gris hay cuerpos de las neuronas, fibras amielínicas en gran cantidad y algunas fibras mielínicas, astrocitos protoplasmáticos, oligodendrocitos y células de la microglia. El color característico de la sustancia blanca es debido a la riqueza de fibras con mielina, sustancia lipidíca blanquecina. La disposición de las dos sustancias varía según la parte del sistema nervioso considerada. En cortes transversales de la médula espinal, la sustancia blanca se localiza externamente y la gris internamente adoptando la forma de H. El trazo horizontal de la H presenta un orificio, corte del canal central de la médula revestido por la células ependimarias. Este canal representa la luz del tubo neural embrionario. La sustancia gris de los trozos laterales de la H forma los cuernos M. Ruth Álvarez Quintillán. anteriores que contienen neuronas motoras, cuyo axones da origen a las raíces centrales de los nervios raquídeos. Forma también los cuernos posteriores que reciben las fibras de las neuronas situadas en los ganglios de las raíces de los nervios espinales (fibras sensitivas). Las neuronas de la médula son multipolares y voluminosas, principalmente las neuronas motoras de los cuernos anteriores alcanzan grandes dimensiones. En el cerebelo se distinguen dos hemisferios unidos por una parte central el vermis. En la superficie del cerebelo se extienden depresiones perpendiculares al vermis, que divide al órgano en lóbulos. En cada lóbulo hay pliegues formados por una parte superficial de la sustancia gris y un eje central de sustancia blanca. Además de constituir la corteza (capa superficial) cerebelosa, la sustancia gris también está presente formando núcleos en la sustancia blanca. La corteza del cerebelo tiene tres capas que de dentro hacia afuera son las siguientes:capa granulosa, capa de células de Purkinje y capa molecular. Las células de la capa granulosa son las neuronas de menor tamaño del cuerpo humano con estructura atípica. La capa de células de Purkinje está formada por una única hilera de éstas células que son muy grandes y poseen dendritas que se subdividen profusamente en un mismo plano. La capa más externa de la corteza cerebelosa es la capa molecular que contiene pocas neuronas y muchas fibras nerviosas amielínicas. TEJIDO CONECTIVO: EL TEJIDO CONJUNTIVO. El tejido conectivo es el principal constituyente del organismo. Se le considera como un tejido de sostén puesto que sostiene y cohesiona a otros tejidos y órganos, sirve de soporte a estructuras del organismo y proteje y aisla a los órganos. Además, todas las sustancias que son absorbidas por los epitelios tienen que pasar por este tejido, que sirve de puente de comunicación entre distintos tejidos y órganos, por lo que generalmente se le considera como el medio interno del organismo. Bajo el nombre de conectivo se engloban una serie de tejidos heterogéneos pero con características compartidas. Una de estas características es la presencia de células embebidas en una abundante matriz extracelular, la cual representa una combinación de fibras colágenas y elásticas y de una sustancia fundamental rica en proteoglucanos y glucosamicoglucanos. Las características de la matriz extracelular son precisamente las responsables de las propiedades mecánicas, estructurales y bioquímicas del tejido conectivo. La clasificación del tejido conectivo en distintos subtipos depende de los autores pero generalmente se agrupan de la siguiente forma: conectivo propiamente dicho (mesenquimático, mucoso o gelatinoso, reticular, elástico, laxo o areolar y denso) y conectivo especializado (adiposo, cartilaginoso, óseo y sanguíneo). Sus funciones, también, son la de unir y reforzar a los demás tejidos, el transporte de vasos y nervios, ayudar a regular la temperatura corporal e intervenir en la autodefensa, mediante la reparación tisular y la fagocitosis. En los órganos se llama estroma las porciones ocupadas por tejido conjuntivo propiamente dicho, colaborando con su arquitectura, y parénquima a la porción funcional, generalmente constituida por tejido epitelial. Aunque hay muchas variantes de este tipo de tejido, todos tienen en común la presencia de células conectivas y material extracelular, formado por una matriz orgánica denominada sustancia fundamental, en cuyo seno se encuentran las fibras conectivas. Existen 2 variedades de células conjuntivas: células estables, las que se originan en el mismo tejido y que sintetizan los diversos componentes de la matriz extracelular que las rodea, y población de células migratorias, originadas en otros territorios del organismo, las que llegan a habitar transitoriamente el tejido conjuntivo. Las células llamadas estables o de sostén corresponden a un grupo de células diferenciadas cuyo principal rol es producir la matriz intercelular propia de cada tipo de tejido conjuntivo. Ellas se forman a partir de células mesenquimáticas localizadas en los sitios del organismo en que van a formar al tejido conjuntivo. Estas células se caracterizan por encontrarse en proceso de activa diferenciación para sintetizar a la matriz extracelular que caracteriza al tipo de tejido conjuntivo que corresponda. Ellas pueden diferenciarse como: -Fibroblastos, producen los tejidos conjuntivos fibrosos cuya matriz extracelular está constituida por fibras colágenas y fibras elásticas, asociadas a glicosaminoglicanos, proteoglicanos y glicoproteínas. - Condroblastos, producen el tejido cartilaginoso, cuya matriz extracelular se caracteriza por la presencia de una cantidad importante de proteoglicanos asociados a ácido hialurónico y a microfibrillas de colágeno tipo II. Al quedar totalmente rodeados por la matriz cartilaginosa ellos pasan a llamarse condrocitos. - Osteoblastos, producen el tejido óseo, sintetizando el componente orgánico de la matriz extracelular ósea que se caracteriza por un alto contenido en colágeno tipo I, glicosaminoglicanos y gliproteínas. Al quedar totalmente rodeados por la matriz ósea pasan a llamarse osteocitos. -Lipoblastos, producen el tejido adiposo. Ellas se diferencian a células almacenadoras de grasa , sintetizan su matriz extracelular y se rodean de una lámina basal . Ellos pasan así a formar los adipocitos o células adiposas. M. Ruth Álvarez Quintillán. Se caracterizan, al microscopio de luz, por su basofilia citoplasmática . Poseen un retículo endoplásmico rugoso bien desarrollado, un aparato de Golgi definido y escasas vesículas de secreción, organelos que se relacionan con la síntesis de moléculas precursoras del colágeno, elastina, proteoglicanos y glicoproteínas de la matriz extracelular. Cuando estas células se hallan en fase de relativa quiescencia en la matriz extracelular que han formado se las llama: fibrocitos, osteocitos y condrocitos. Los lipoblastos se diferencian posteriormente a células adiposas cuyo principal rol es almacenar grasas. Las células conjuntivas libres se originan en la médula ósea hematopoyética y usan la circulación sanguínea como un medio de transporte hacia los tejidos conjuntivos, donde realizan sus principales funciones. Entre ellas se encuentran las células cebadas(o mastocitos) y los macrófagos (o histiocitos) que son componentes estables del tejido conjuntivo al que llegan. Un grupo distinto lo forman las células plasmáticas, los linfocitos y los granulocitos polimorfonucleares, todos ellos células de vida media relativamente corta y que tienden a concentrarse en las zonas de tejido conjuntivo en que ocurren reacciones relacionadas con la defensa. La matriz extracelular es una red organizada, formada por el ensamblaje de una variedad de polisacáridos y de proteínas secretadas por las células estables, que determina las propiedades físicas de cada una de las variedades de tejido conjuntivo. El análisis microscópico de la estructura del material intercelular muestra: elementos fibrilares bien estructurados llamados fibras conjuntivas, que pueden ser colágenas, reticulares o elásticas, un material poco estructurado al que se denomina sustancia fundamental amorfa y en los sitios en que la matriz extracelular conjuntiva se asocia a células de otros tejidos se observan las láminas basales. -Fibras conjuntivas: los principales tipos corresponden a fibras elásticas y a las estructuras formadas por fibrillas colágenas. Las fibras elásticas se estiran fácilmente y recuperan su longuitud original cuando la fuerza deformante ha desaparecido. Al microscopio electrónico aparecen formadas por un componente amorfo rodeado y penetrado por microfibrillas de unos 10 nm de diámetro.El principal componente de las fibras eláticas es la elastina, material proteico muy insoluble. Se caracteriza por un alto contenido en aminácidos apolares como prolina y valina y tiene un alto contenio en aminácidos no cargados como la glicina. Contiene además dos aminácidos exclusivos: desmosina e isodesmosina. Las microfibrillas tienen como principal componente una glicoproteína llamada fibrilina, rica en aminoácidos hidrofílicos y que contiene residuos de cistina los cuales pueden formar puente disulfuro, estabilizando la estructura de las microfibrillas. En las fibras elásticas juegan un rol estructural, formando una especie de vaina alrededor de la elastina, permitiendo así la formación de estructuras fibrilares elásticas. En ausencia de microfibrillas la elastina se deposita formando láminas elásticas, como ocurre por ejemplo en la pared de las arterias. Las fibrillas colágenas forman estructuras que se caracterizan por su gran resistencia a las fuerzas de tracción y el modo en que ellas se asocian entre sí varía de un tejido conjuntivo a otro. Ellas son el principal componente de: las fibras colágenas de los tejidos conjuntivos fibrosos, las fibras reticulares y las laminillas colágenas del hueso. Cada fibrilla colágena está formada por la asociación de moléculas de colágeno, proteínas fibrosas de varios tipos, que constituyen más del 25% de la masa proteica total de los mamíferos. Las moléculas de colágeno se asocian formando polímeros que corresponden a las fibrillas colágenas visibles al microscopio electrónico. El diámetro de las fibrllas varía entre 10 y 300 nm, según el tipo de colágeno de que se trate. Las moléculas de colageno de tipo I, II y III, son capaces de interactuar entre sí, asociándose lateralmente en forma estable por uniones covalentes entre residuos de lisina. -Variedades del colágeno: -Colágeno tipo I: Se encuentra abundantemente en la dermis, el hueso, el tendón y la córnea. Se presenta en fibrillas estriadas de 20 a 100 nm de diámetro, agrupándose para formar fibras colágenas mayores. Es sintetizado por fibroblastos, condroblastos y osteoblastos. Su función principal es la de resistencia al estiramiento. -Colágeno tipo II: Se encuentra sobre todo en el cartílago, pero también se presenta en la córnea embrionaria y en la notocorda, en el núcleo pulposo y en el humor vítreo del ojo. En el cartílago forma fibrillas finas de 10 a 20 nanómetros, pero en otros microambientes puede formar fibrillas más grandes, indistinguibles morfológicamente del colágeno tipo I. Es sintetizado por el condroblasto. Su función principal es la resistencia a la presión intermitente. M. Ruth Álvarez Quintillán. -Colágeno tipo III: Abunda en el tejido conjuntivo laxo, en las paredes de los vasos sanguíneos, la dermis de la piel y el estroma de varias glándulas. Parece un constituyente importante de las fibras de 50 nanómetros que se han llamado tradicionalmente fibras reticulares. Es sintetizado por las células del músculo liso, fibroblastos, glía. Su función es la de sostén de los órganos expandibles. En cuanto a la sustancia fundamental, corresponde a la matriz traslúcida que rodea a células y fibras conjuntivas y que no presenta una organización estructural visible al microscopio de luz. Este componente constituye un gel semisólido, bioquímicamente complejo y altamente hidratado. Las principales macromoléculas que forman la sustancia fundamental corresponden a polisacáridos de la clase de los glicosaminoglicanos, que pueden existir como tales o pueden unirse covalentemente a proteínas, formando macromoléculas más complejas llamadas proteoglicanos. Y las láminas basales, son estructuras laminares que corresponden a una forma especializada de la matriz extracelular, que actúan como interfase entre las células de los tejidos epiteliales, músculares y nerviosos en los sitios en que ellas se asocian al tejido conjuntivo. Al microscopio electrónico aparece formada por 2 capas laminares: la que está vecina a la célula es poco densa a los electrones y se denomina lamina rara (o lúcida), la segunda capa es densa a los electrones y se denomina lamina densa , a ella se asocia una tercera capa que contiene fibrillas colágenas llamada lamina fibroreticular y que es continua con la matriz conjuntiva adyacente. Entre las funciones que desempeña la lámina basal se destacará en este momento sólo que ella constituye una interfase the adhesión de modo que las células poseen mecanismos de adhesión para fijarse a la membrana basal y ella a su vez está anclada en la matriz extracelular conjuntiva a las fibrillas colágenas. Las membranas basales están formadas por 4 tipos de moléculas predominantes: colágeno tipo IV, perlecan (proteoglicano de heparan-sulfato), laminina y entactina además de la fibronectina y del colágeno VII que la asocia a las fibrillas de colágeno III de la lámina fibroreticular. -Variedades del tejido conectivo/conjuntivo: -Tejido conjuntivo laxo: Se caracteriza por la presencia de una población relativamente alta de células residentes, ya sea propias como fibroblastos y adipocitos o migratorias como macrófagos y células cebadas, separadas por la matriz extracelular formada por fibras colágenas y elásticas, laxamente dispuestas en una sustancia fundamental bastante fluída. Las fibras colágenas que predominan están formadas por microfibrillas de colágeno I asociadas a cantidades variables de microfibrillas de colágeno III pero en menor cantidad. Estas fibras colágenas le confieren resistencia a la tracción. Las fibras elásticas, aunque menos abundantes que las de colágeno, cumplen un rol importante ya que le otorgan al tejido la capacidad de retomar su organización original luego de ser traccionado. La sustancia fundamental, que ocupa los espacios entre las fibras, tiene como principal componente a grandes moléculas de ácido hialurónico, el cual estabiliza una importante cantidad de agua, adoptando la estructura de un gel poco consistente, de modo que existen innumerables espacios intermoleculares que sirven de reservorio al líquido intersticial ("fluido tisular"). Este fluído facilita la difusión del oxígeno y nutrientes desde los capilares del conjuntivo hacia células de otros tejidos (epitelios, cartíago o músculo) y de los productos de desecho del metabolismo en sentido inverso. La capacidad de la sustancia fundamental de acumular líquido es la base del proceso llamado edema. Debido a la variedad de células que lo habitan y a la capacidad de acumular líquido proveniente del plasma sanguíneo, el conjuntivo laxo juega un rol muy importante en la iniciación del proceso de defensa orgánica a través de la generación de las respuestas inflamatoria e inmune. -Tejido conjuntivo fibroso denso: Presenta un contenido relativamente bajo de células, las que corresponden principalmente a fibroblastos. Su matriz extracelular es muy abundante, y su principal componente son gruesas fibras colágenas. La sustancia fundamental es relativamente escasa, predominando proteoglicanos de dermatan-sulfato. De acuerdo a la forma en que se disponen las fibras de colágeno, se distingue: -Tejido conjuntivo denso desordenado. Las fibras colágenas forman una red tridimensional lo que le otorga resistencia en todas las direcciones. Asociada a esta red colágena existen fibras elásticas. Entre las fibras colágenas y elásticas se ubican las células, principalmente fibroblastos y se encuentra por ejemplo en la dermis y formando la cápsula de órganos como los ganglioslinfático y el hígado. -Tejido conjuntivo denso ordenado o regular. Las fibras de colágeno se disponen en un patrón definido que refleja una respuesta al la direccción del requerimiento mecánico predominante. Las fibrillas de colágeno tipo I paralelas se asocian entre sí por interaccion con moléculas de colágeno tipo XII, lo que le otorga una mayor firmeza . Los fibroblastos son las únicas células presentes y se ubican entre los haces de fibras formando largas líneas paralelas. Estas células poseen escaso citoplasma y presentan un cuerpo celular alargado, con M. Ruth Álvarez Quintillán. núcleos son alargados y de cromatina condensada. -Tejido conjuntivo elástico: Es una variedad de tejido fibroso denso en el cuál las fibras conjutivas presentes corresponden a fibras o láminas elásticas dispuestas en forma paralela. Los espacios entre las fibras elásticas están ocupados por una fina red de microfibrillas colágenas con unos pocos fibroblastos. El tejido conjuntivo elástico forma capas en la pared de los órganos huecos sobre cuyas paredes actúan presiones desde adentro, como es el caso de los pulmones y de los vasos sanguíneos y forma algunos ligamentos como los ligamentos amarillos de la columna vertebral. -Tejido conjuntivo reticular: Es una variedad de tejido conjuntivo especializado que forma una malla tridimensional estable, que otorga un soporte estructural a las células migratorias de órganos relacionados directamente con los leucocitos de la sangre como son el bazo, los ganglios linfáticos y la médula ósea hematopoyética. Las células reticulares, que corresponden a fibroblastos especializados, que secretan las microfibrillas de colágeno III las que se asocian en manojos formando las fibras reticulares. Estas se disponen formando una malla fibrilar fina a lo largo de la cual se ubican las células reticulares, cuyos procesos envuelven las fibras reticulares y a la escasa sustancia fundamental. El sistema trabecular formado por la asociación de fibras y células genera matriz con características de esponja en la cual células y fluídos pueden desplazarse. -Tejido adiposo: Es un tejido conjuntivo especializado en el que predominan las células conjuntivas llamadas adipocitos. Los lipoblastos, células precursoras de adipocitos producen cantidades importantes de colágeno I y III, pero los adipocitos adultos secretan muy bajas cantidades de colágeno y pieden la capacidad de dividirse. Los adipocitos almacenan energía en forma de triglicéridos. Debido a la baja densidad de estas moléculas y su alto valor calórico, el tejido adiposo es muy eficiente en la función de almacenaje de energía. Los adipocitos diferenciados pierden la capacidad de dividirse; sin embargo, son células de una vida media muy larga y con capacidad de aumentar la cantidad de lípidos acumulados. Además, el tejido adiposo postnatal contiene adipocitos inmaduros y precursores de adipocitos residuales a partir de los cuales pueden diferenciarse adipocitos adicionales. Estos mecanismos se hacen operativos cuando la ingasta calórica aumenta exageradamente. Existen dos tipos principales de tejido adiposo: -Tejido adiposo blanco: que forma hasta el 20% del peso corporal total en varones adultos normales y bien nutridos y hasta el 25% en las mujeres. Se distribuye por todo el organismo, sobre todo en las capas profundas de la piel. Es un gran depósito de energía y sirve también como aislamiento térmico bajo la piel y como amortiguador de los choques mecánicos en lugares como la zona perirrenal. -Tejido adiposo pardo: muy especializado, se encuentra en los mamíferos recién nacidos y en algunos animales que hibernan, en los que desempeña un papel muy importante en cuanto a regulación de la temperatura corporal. En los adultos humanos la cantidad es muy escasa, y aunque antes se creía que contribuía poco a la termorregulación existen pruebas de que al menos en algunos individuos, puede desempeñar un papel en el gasto de exceso de energía, evitando así la obesidad. -Cartílago y hueso: Son tejidos conjuntivos que se caracterizan porque sus células están rodeadas de una matriz intercelular sólida y relativamente rígida. La matriz intercelular del cartílago es deformable y puede crecer por depósito de nuevo material en su interior, mientras que la matriz intercelular ósea es más bien rígida y puede crecer sólo por depósito de nuevo material sobre las superficies óseas. El cartílago está formado por una abundante matriz extracelular en la cuál los condrocitos se ubican en espacios llamados lagunas. Los condrocitos sintetizan y secretan los componentes orgánicos de la matriz extracelular que son básicamente colágeno, ácido hialurónico. proteoglicanos y glicoproteínas, y según las características de la matriz se distingue cartílago hialino y fibroso. Existe además el cartilago elástico en el cuál la elastina forma parte de la matriz extracelular. Los vasos sanguíneos no penetran a la matriz cartilaginosa y los condrocitos se nutren con material que difunde desde de los capilares sanguíneos del tejido conjuntivo adyacente. Cada placa o lámina de tejido cartilaginoso está rodeada por el pericondrio que corresponde a tejido conjuntivo denso en el cuál se distingue una capa externa fibrosa y una capa interna celular en la cual se ubican las células que pueden dar origen a los condroblastos, que corresponden a precursores de los condrocitos y que difieren de ellos solo en su edad y en su mayor actividad de la sintesis de componentes de la matriz intercelular cartilaginosa. Los condroblastos presentan un ergastoplasma y un aparato de Golgi muy desarrollados y presentan vesículas y granos de secreción, lo cuál guarda relación con su rol de sintetizar y secretar los distintos componentes de la M. Ruth Álvarez Quintillán. matriz extracelular cartilaginosa que está formada principalmente por colágeno de tipo II, proteoglicanos de condroitin y keratansulfato, ácido hialurónico y glicoproteínas. Al disminuir su actividad de síntesis disminuye el desarrollo tanto del ergastoplasma como del aparato de Golgi, acumulan glicógeno y lípidos en su citoplasma y se les llama condrocitos El tejido óseo es una variedad de tejido conjuntivo que se caracteriza por su rigidez y su gran resistencia tanto a la tracción como a la compresión. Está formado por la matriz ósea, que es un material intercelular calcificado y por células, que pueden corresponder a: -Osteoblastos: encargados de sintetizar y secretar la parte orgánica de la matriz ósea durante su formación. Se ubican siempre en la superficie del tejido óseo ya que este sólo puede crecer por aposición. -Osteocitos, responsables de la mantención de la mátriz ósea, que se ubican en cavidades o lagunas rodeadas por el material intercelular calcificado. La nutrición de los osteocitos depende de canalículos que penetran la matriz ósea y conectan a los osteocitos vecinos entre sí y con canales vasculares que penetran al hueso o que se ubican en las membranas conjuntivas que revisten la superficies del hueso (periostio y endostio). -Osteoclastos, células responsables de la reabsorción del tejido óseo, que participan en los procesos de remodelación de los huesos y pueden encontrarse en depresiones superficiales de la matriz ósea llamadas lagunas de Howship. REFERENCIAS BIBLIOGRÁFICAS: -Wheater´s Histología Funcional, Ed. Elsevier, Churchill Livingstone. -Histología, texto y atlas en color con biología celular y molecular, Ross Pawlina, Ed. Médica panamericana. -webs.uvigo.es/mmegias/ -www.infermeravirtual.com