Tema II.- Estructura de la célula
Anuncio

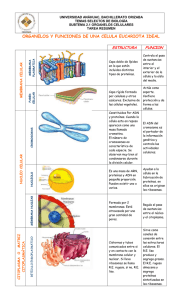
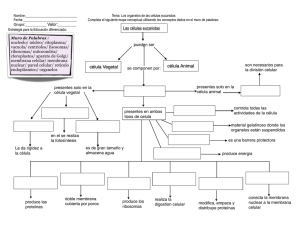
UNIDAD DIDÁCTICA II (1ª Parte): ESTRUCTURA DE LA CÉLULA 1.- INTRODUCCIÓN En el S.XVII Hooke y Van Leeuwenhoeck realizan las primeras observaciones de células al microscopio óptico. Desde entonces, los conocimientos de la célula han ido avanzado paralelamente al perfeccionamiento de, primero, el microscopio óptico y, posteriormente, el electrónico (que sustituye la luz por haces de electrones y permite una mejor percepción de los detalles). Asimismo, han tenido que ir apareciendo nuevas técnicas para realizar las preparaciones microscópicas, tanto de tinción y fijación como de corte (las láminas finas que se precisan se obtienen con micrótomos). 2.- TEORÍA CELULAR A principios del S. XIX Schleiden y Schwann postulan la teoría celular, según la cual, la célula es la unidad estructural y funcional de los seres vivos, capaz de mantener una existencia propia e independiente. Esta teoría fue completada por Virchow, Strasburger y Cajal..., hasta llegar a los postulados actuales, que se pueden resumir en: 1.- Todos los seres vivos están compuestos por una o más células (unidad estructural). 2.- Una célula realiza todas las reacciones químicas necesarias para el mantenimiento de la vida (unidad funcional). 3.- Toda célula procede de otra ya preexistente, lo que permite la transmisión de caracteres de generación en generación. 4.- La célula es la unidad de vida más pequeña que existe. 3.- TIPOS DE CÉLULAS: A.- Introducción: Existe una enorme diversidad de tipos celulares. Podemos establecer diferencias respecto al tamaño (las vegetales oscilan entre 20 y 50 µ m; las animales entre 10 y 20 µ m y las bacterianas en torno a 5 µ m aunque existen excepciones notables, como los óvulos de aves y reptiles, visibles a simple vista o los micoplasmas, de sólo 0,1 µ m), o a la forma (existe una estrecha relación entre forma y función, y de ahí que existan células esféricas, alargadas, fusiformes, estrelladas...); pero, en general, se considera que existen dos tipos básicos de organización celular: El procariota y el eucariota. B.- La célula procariota: Es la célula típica de las bacterias. Son mucho más pequeñas y sencillas que las eucariotas, presentando las siguientes características: • La membrana plasmática está rodeada de una cubierta rígida protectora o pared celular, cuya composición química es diferente a la de las células eucariotas. La membrana presenta invaginaciones o mesosomas, donde se localizan las enzimas necesarias para la respiración. A su vez, la pared está rodeada de una cápsula, en cuya superficie aparecen frecuentemente fimbrias o pelos y flagelos, que permite el movimiento de la célula. • El citoplasma contiene numerosos ribosomas, más pequeños que los de las células eucariotas. • El material genético está constituido por una sola molécula de ADN circular, que recibe el nombre de cromosoma procariota, pues su organización es diferente a la de los de los eucariotas. Dicho cromosoma se localiza en el citoplasma, en una región denominada nucleoide, pero no existe membrana celular, por lo que tampoco existe núcleo definido. También se pueden encontrar pequeños anillos de ADN, independientes del cromosoma, denominados plásmidos, que confieren a la célula la capacidad de conjugación (plásmido F) o codifican enzimas que proporcionan resistencia a las bacterias frente a antibióticos (plásmido R). 1 C.- La célula eucariota: a.- Características generales: Son más complejas y de mayor tamaño que las procariotas, presentando gran variedad de formas, en relación con su forma de vida (organismos unicelulares) o su función (organismos pluricelulares). La poseen los organismos de los reinos Protoctistas, Hongos, Metazoos y Metafitas. Se caracterizan por: • El material genético se encuentra Célula animal Célula vegetal aislado del resto de la célula por una Tamaño Entre 10 y 20 µm Entre 20 y 50 µm membrana, formando el núcleo. Matriz Pueden presentar No presentan • Existe un complejo sistema de extracelular membranas internas que aíslan Pared celular No presentan Presentan Poco desarrolladas Muy desarrolladas compartimentos u orgánulos, cada uno Vacuolas Plastos No presentan Presentan con una o varias funciones. Centríolos Presentan Las típicas no presentan Existen dos tipos morfológicos: la célula animal y la vegetal, cuyas diferencias aparecen en el cuadro. DIFERENCIAS ENTRE CÉLULAS PROCARIOTAS Y EUCARIOTAS CÉLULA EUCARIOTA Generalmente grandes (10-100 µm) Material genético en un núcleo con membrana Material genético formando cromosomas Orgánulos citoplasmáticos aislados por membranas Material genético extranuclear en mitocondrias y plastos Con citoesqueleto Ribosomas 80s Membranas celulares con colesterol y esteroides Pared celular de celulosa o quitina (no en animales) División por mitosis Enzimas respiratorios en mitocondrias En fotosintéticas los enzimas se localizan en cloroplastos Movimiento por cilios, flagelos o pseudópodos Organismos unicelulares y pluricelulares CÉLULA PROCARIOTA Generalmente pequeñas (1-10 µm) Material genético no rodeado por membrana Un solo cromosoma procariota, circular Sin sistemas endomembranosos que aíslen orgánulos Material genético extracromosómico en forma de plásmidos Sin citoesqueleto Ribosomas 70s Membranas celulares sin colesterol ni esteroides Si existe pared no es de celulosa (de mureína en ocasiones) División celular directa Enzimas respiratorios en mesosomas En fotosintéticas los enzimas se localizan en la membrana Movimiento por flagelos Organismos unicelulares b.- Envueltas celulares: Son todas aquellas estructuras que aíslan la célula de su entorno y separan el contenido celular del medio extracelular. Incluyen la membrana plasmática, presente en todo tipo de células, y las membranas de secreción, es decir, el glucocálix de las células animales, la pared celular de las vegetales y la pared de quitina de los hongos. a’.- Membrana plasmática: Es una finísima (75 a 100 Å) envuelta continua, que sólo puede observarse al microscopio electrónico. Se considera que de ella derivan el resto de las membranas internas celulares. Actúa como una barrera selectiva que permite que el contenido del interior celular sea distinto al del medio extracelular, regulando, al mismo tiempo, el intercambio célula – medio. a’’.- Composición química: Las membranas biológicas están constituidas mayoritariamente por lípidos y proteínas, junto a una pequeña proporción de glúcidos. a’’’.- Lípidos de membrana: Son principalmente fosfolípidos (que al ser moléculas con un polo hidrófilo y otro hidrófobo se disponen formando una doble capa o bicapa lipídica, que constituye la estructura básica de la membrana) y, en menor proporción, colesterol (se intercala entre los fosfolípidos) y glucolípidos (en la parte exterior de la membrana). Tienen una cierta capacidad de movimiento, mediante difusión lateral, rotación alrededor de un eje perpendicular a la membrana, flexión y flip-flop (salto de un lado al otro de la bicapa). 2 b’’’.- Proteínas: Se insertan en la bicapa lipídica Las regiones polares sobresalen en las superficies de la membrana, mientras que las apolares se encuentran en el interior hidrofóbico de éstas. Pueden moverse por difusión lateral y rotación, fundamentalmente. Se distinguen dos tipos: • Proteínas intrínsecas, integrales o transmembrana: Atraviesan por completo la membrana y sobresalen por ambas caras de ésta, estando íntimamente asociadas a los lípidos. Constituyen aproximadamente el 70% del total de proteínas de membrana. Pueden formar glucoproteínas. • Proteínas extrínsecas o periféricas: No atraviesan toda la membrana situándose habitualmente en su superficie. Pueden estar unidas a lípidos débilmente o a proteínas transmembrana. c’’’.- Glúcidos: Glucocálix: Son principalmente oligosacáridos que aparecen formando glucolípidos o glucoproteínas, y se sitúan siempre en la cara exterior de la membrana (distribución asimétrica), formando una capa periférica conocida como glucocálix, o glicocálix, que, en el caso de las células de los tejidos animales, se encuentra inmerso en una matriz extracelular, compuesta por proteínas y polisacáridos, siendo difícil establecer el límite entre ambas. La función de esta cubierta no se conoce con exactitud, pero se considera que protege a la superficie celular de daños mecánicos y químicos y que interviene en procesos de comunicación y reconocimiento celular (muchos receptores de membrana son glucoroteínas). Asimismo, las moléculas glucídicas actúan como antígenos sobre los que se fijan anticuerpos específicos que determinan el comportamiento inmunológico de las células. También intervienen como receptor de moléculas (hormonas), agentes patógenos (virus, bacterias) o sus toxinas. b’’.- Estructura: En 1972 Singer y Nicolson proponen el modelo de Mosaico fluido para las membranas celulares. Según éste, todas las membranas celulares serían como un mosaico formado por una solución de proteínas dispersas en la matriz de una bicapa lipídica fluida. Las proteínas intrínsecas atravesarían total o parcialmente la bicapa, y las extrínsecas estarían en la superficie de dicha bicapa. Tanto lípidos como proteínas se pueden desplazar lateralmente dentro de la bicapa. Las membranas son asimétricas en cuanto a la disposición de sus componentes moleculares c’’.- Funciones: La membrana plasmática de la célula es una estructura altamente diferenciada, cuyas principales funciones son: • Confiere a la célula su individualidad, al separarla de su entorno, manteniendo, al mismo tiempo, la integridad estructural de la célula. • Controla el intercambio de sustancias, actuando como una barrera semipermeable entre la célula y el medio extracelular. Es un filtro altamente selectivo que permite la entrada, salida y permanencia de ciertas moléculas esenciales, manteniendo constante el medio interno. De ahí la importancia de los mecanismos de transporte a través de la membrana, que estudiaremos en el tema siguiente. • Controla el flujo de información entre las células y su entorno, ya que contiene receptores específicos para los estímulos externos. A su vez, algunas membranas generan señales (químicas o eléctricas). Por ello las membranas juegan un papel central en las comunicaciones biológicas • Posibilitan el contacto y las uniones entre células, permitiendo la formación de tejidos y dando consistencia a los seres vivos. • Proporcionan el medio óptimo para el funcionamiento de las proteínas de membrana (enzimas, receptores y proteínas transportadoras). 3 d’’.- Diferenciaciones de la membrana: Las células a veces presentan especializaciones en su membrana, entre las que destacan las microvellosidades y las uniones intercelulares. a’’’.Microvellosidades: Son prolongaciones digitiformes (en forma de dedo de guante) del citoplasma, recubiertas por la membrana, situadas en la superficie libre de ciertas células (p.e. epitelio que tapiza el intestino delgado), cuya misión es aumentar la superficie de absorción. b’’’.- Uniones intercelulares: Sirven para unir entre sí las células de los organismos pluricelulares, así como para permitir las comunicaciones intercelulares. Son básicamente de 3 tipos: • Uniones herméticas o de oclusión: sellan las membranas de las células adyacentes, para impedir que las sustancias pasen a través de los espacios intercelulares. • Uniones de anclaje: Son puntos de contacto intercelulares (a modo de remaches) que mantienen firmemente unidas a las células, aumentando la resistencia y rigidez del tejido. Entre ellas destacan los desmosomas. • Uniones de comunicación o tipo gap: Son canales intercelulares que permiten el paso de iones y pequeñas moléculas entre células adyacentes. Un ejemplo serían las sinapsis b’.- Pared celular: Es una gruesa cubierta que rodea externamente a la membrana plasmática de las células vegetales. a’’.- Composición química y estructura: Varía de unas plantas a otras e, incluso dentro de la misma, según su estado de desarrollo y tejido del que forme parte. En general se distinguen una serie de capas: a´´´.- Lámina media: Formada fundamentalmente por pectina. Es la primera en formarse entre dos células que acaban de dividirse y permanecen unidas. Se puede impregnar de lignina en células de algunos tejidos de sostén o de conducción, con lo que adquiere mayor rigidez. b’’’.- Pared primaria: Está formada por celulosa dispuesta en forma reticular, y una matriz de hemicelulosa, pectinas y glucoproteínas. Se va formando durante el crecimiento de la célula entre la membrana plasmática y la lámina media, que se irá desplazando hacia fuera. c‘’’.- Pared secundaria: Formada por abundantes fibras de celulosa, dispuesta formando diversas capas (la celulosa se dispone con distinta orientación en cada una de ellas) y una matriz más escasa de hemicelulosa. Puede también impregnarse de lignina, siendo muy rígida y difícilmente deformable, por lo que sólo aparece en células especializadas de tejidos de sostén y conductores. 4 d’’’.- Especializaciones: La lámina media y la pared primaria son permeables al agua, gases y pequeñas moléculas hidrosolubles. Pero existen, además, estructuras que favorecen el paso de sustancias. • Punteaduras: Zonas en forma de disco donde la pared se adelgaza, quedando reducida prácticamente a la lámina media. • Plasmodesmos: Son canales entre células vecinas que atraviesan la pared de ambas. A través de ellos se prolongan las membranas plasmáticas y se comunican los citoplasmas, e incluso sacos del retículo. Pueden localizarse en las punteaduras o distribuirse por toda la pared celular. b’’.- Funciones: • Constituye un exoesqueleto que protege y da forma a la célula vegetal. • Es la responsable de que la planta se mantenga erguida. • Permite a las células vegetales vivir en un medio hipotónico. Como consecuencia de la entrada de agua, la célula se hincha y presiona contra la pared rígida. Esta presión (denominada turgencia) es la principal fuerza impulsora de la expansión celular durante el crecimiento y es la causa de gran parte de la rigidez de la planta joven. • La lignificación (impregnación de lignina) aumenta la resistencia mecánica de los tejidos. La suberificación y cutinización (impregnación de suberina y de cutina respectivamente) que tiene lugar en los tejidos protectores superficiales de la planta producen una impermeabilización que evita la pérdida de agua. • Constituye una barrera para el paso de sustancias y agentes patógenos. c.- Citosol o Hialoplasma: Es el medio celular donde se encuentran inmersos los orgánulos citoplasmáticos. Se trata de un sistema coloidal muy heterogéneo, tanto por la diversidad de sus componentes como por las variaciones que sufre según el momento de la fisiología celular que se considere. Está formado por una elevada cantidad de agua (80% aproximadamente), en la que se encuentran en disolución o dispersión coloidal toda clase de biomoléculas (aminoácidos, glúcidos, nucleótidos, ATP, sales, iones, enzimas y otras proteínas...) y gran cantidad de compuestos intermediarios biológicos, ya que es aquí donde se desarrollan la mayoría de las reacciones del metabolismo celular (glucólisis, gluconeogénesis, glucogenolisis, biosíntesis de proteínas y de ác. grasos...). Asimismo, regula el pH intracelular. Se puede presentar en estado viscoso (citogel) o fluido (citosol), pudiendo pasar de un estado a otro en función de las necesidades metabólicas de la célula. A estos cambios se debe, p.e. el movimiento ameboide. En el citosol pueden almacenarse sustancias en forma de gránulos, denominadas Inclusiones, como, p.e., depósitos de glucógeno, gotas de grasa, o muchos pigmentos. Por último, en el medio acuoso del citosol se encuentran inmersos los orgánulos celulares y el citoesqueleto. d.- Citoesqueleto y sus derivados: El citoesqueleto es una red compleja de filamentos proteicos que se extiende por todo el citoplasma, estando implicado en la morfología celular, su organización interna y sus movimientos. Está formado por tres tipos de componentes: Microfilamentos, Microtúbulos y Filamentos intermedios. 5 a’.- Microfilamentos: Son los filamentos más finos. Están constituidos por una proteína, la actina (aunque también los puede haber en menor cantidad de otras proteínas, como la miosina), que se presenta en dos formas, la actina G o globular, que va unida a otra proteína, la profilina, que evita su polimerización; la otra forma es la actina F, constituida por grupos de dos moléculas de actina G que se unen formando polímeros en forma de hebras helicoidales. Las funciones que realizan son diversas: • Contráctil: En las células musculares, los filamentos de actina se asocian con fibras de miosina, permitiendo el deslizamiento entre ambos y provocando la contracción muscular. • De soporte: En las microvellosidades aparecen filamentos paralelos de actina asociados a los de miosina, confiriendo rigidez a las microvellosidades. • Permitir el movimiento ameboide: Los pseudópodos son evaginaciones de la membrana plasmática que contienen filamentos de actina asociados a miosina. • Formación del anillo contráctil que divide el citoplasma en dos durante la división celular. • Refuerzo de la membrana plasmática. b’.- Filamentos intermedios: Tienen un tamaño de unos 10 nm, y su composición y funciones varían según el tipo celular. Desempeñan funciones estructurales, como p.e. los filamentos de queratina que proporcionan fuerza mecánica a las células epiteliales, los neurofilamentos que dan resistencia a los axones de las neuronas o los filamentos de la lámina nuclear, que forman un armazón interno, debajo de la envoltura del núcleo. c’.- Microtúbulos: Tienen forma de cilindro hueco, con longitud variable y diámetro de unos 24 nm. Están constituidos por la polimerización de una proteína globular, la tubulina, que al polimerizarse forma los denominados protofilamentos. Un microtúbulo está formado por 13 protofilamentos situados en círculos y dejando un eje central hueco. 6 Sus funciones son diversas: • Formación del huso mitótico, responsable del movimiento de los cromosomas durante la división celular. • Intervienen en la organización de todos los filamentos del citoesqueleto. • Determinan la organización y distribución del retículo endoplasmático y del aparato de Golgi. • Determinan la forma de la célula. • Dirigen el transporte de orgánulos (mitocondrias, vesículas...) por el citoplasma. Pueden aparecer dispersos por el citoplasma, como parte del citoesqueleto, o bien formar estructuras estables, entre los que tenemos: a’’.- Centríolos y Centrosoma a´´´.- Centríolo: Un centríolo es una estructura cilíndrica formada por 9 grupos de 3 microtúbulos cada uno (triplete), formando la denominada estructura 9 + 0. Los 3 microtúbulos de un triplete están estrechamente asociados los unos a los otros, y, a su vez, cada triplete queda unido al siguiente mediante una proteína (nexina). Podemos encontrarlos en dos estructuras diferentes: en el centrosoma y en la base de cilios y flagelos. b´´´.- Centrosoma: Es una estructura sin membrana, presente en todas las células animales susceptibles de dividirse. Salvo algunas excepciones, no existe en células vegetales. Consta de un cuerpo central, formado por dos centríolos, dispuesto uno perpendicular al otro (formando el “Diplosoma”), rodeado por el “material pericentriolar”, electrónicamente denso y amorfo. Al conjunto de estos dos componentes se le llama “Centro organizador de microtúbulos” (COMT), ya que de él derivan todas las estructuras formadas por microtúbulos, incluido el huso mitótico. Los microtúbulos citoplasmáticos tienden a irradiar en todas direcciones a partir del centrosoma. b’’.- Cilios y flagelos: Son prolongaciones de la membrana plasmática dotadas de movimiento. Los cilios son cortos y muy numerosos, mientras que los flagelos son largos y escasos (frecuentemente sólo existe uno), pero ambos poseen la misma estructura: Un “Tallo o Axonema”, una “Zona de transición” y el “Corpúsculo basal”. • Axonema o Tallo: Está rodeado por la membrana plasmática y en su interior tiene 2 microtúbulos centrales y 9 pares o dobletes de microtúbulos periféricos (estructura 9 + 2), todos ellos orientados paralelamente al eje principal del cilio o flagelo. De cada doblete al contiguo parten fibras de una proteína (dineína), y las parejas de microtúbulos adyacentes están unidas por nexina. 7 • Zona de Transición: Es la base del cilio o flagelo. Desaparece el par de túbulos centrales y aparece la “placa basal”, de naturaleza proteica, que conecta la base del cilio o flagelo con la membrana plasmática. • Corpúsculo basal: Su estructura es idéntica a la del centríolo (9 + 0) de manera que dos de los microtúbulos periféricos de cada triplete procede del axonema. Está situado justo por debajo de la membrana plasmática. La función de cilios y flagelos está relacionada con el movimiento, ya que permite el desplazamiento de las células en un medio líquido (caso de algunos protozoos o de los espermatozoides, p.e.), o bien provoca que sea el medio extracelular situado sobre la superficie ciliar el que se mueva (células de las trompas de Falopio o epiteliales ciliadas, p.e.). e.- Ribosomas: Son pequeñas partículas sin membrana, observables solamente con el microscopio electrónico. En general existen en todas las células. a’.- Estructura: Los ribosomas se componen de ARNr y proteínas. Están constituidos por dos subunidades, una grande y otra pequeña, de distinto tamaño en el caso de células procariotas y eucariotas, lo que hace que también los ribosomas posean distinto tamaño. Los de células procariotas tienen un coeficiente de sedimentación de 70S y los de eucariotas 80S. El ARNr se sintetiza en el nucléolo de la célula y las proteínas en el citosol, emigrando posteriormente al nucléolo. Las dos subunidades salen al citoplasma, y es allí donde se ensamblan para formar el ribosoma. b’.- Localización: Pueden localizarse libres en el citoplasma, ya sean aislados o unidos entre sí formando polisomas (o polirribosomas), que son agregados de hasta 100 ribosomas unidos por una cadena de ARNm. En otras ocasiones aparecen adheridos a la cara externa de las membranas del retículo endoplasmático rugoso y del núcleo. Por último, existen ribosomas 70s libres en el interior de mitocondrias y cloroplastos. 8 c’.- Funciones: Los ribosomas intervienen en la síntesis de proteínas, uniendo los aminoácidos en un orden determinado. La subunidad mayor tiene como función primordial catalizar la formación de enlaces peptídicos. La menor se une a moléculas de ARNt y ARNm. Las proteínas sintetizadas por ribosomas libres quedan en el citosol. Las sintetizadas en los adheridos a la membrana del retículo pasan al interior del retículo, para incorporarse a otros orgánulos intracelulares o ser secretadas al exterior celular. f.- Sistema de endomembranas y sus derivados: El sistema de endomembranas es un complejo sistema de vesículas y sacos aplanados, revestidos por membranas y extensamente comunicados y relacionados entre sí (unas estructuras se forman a expensas de otras), que ocupan gran parte del citoplasma en todas las células eucariotas. Básicamente está formado por el retículo endoplasmático y el Aparato de Golgi, aunque también se suele incluir en él la membrana nuclear (que estudiaremos con el núcleo), y algunos orgánulos derivados de los dos primeros, como las vacuolas, lisosomas y peroxisomas. Las endomembranas presentan la misma estructura y composición que la membrana plasmática. a’.- Retículo endoplasmático (RE): Representa aproximadamente más de la mitad de la membrana de la célula. Está constituido por una serie de cavidades membranosas de forma irregular (sacos o cisternas, vesículas y tubos aplanados) e interconectadas entre sí que se extienden por todo el citoplasma. La membrana del RE delimita un espacio central o lumen, que establece comunicación con la envoltura nuclear y los sacos del Aparato de Golgi. Se distinguen dos zonas bien diferenciadas: a’’.- Retículo endoplasmático rugoso(RER): Lleva adheridos ribosomas en su cara externa. Sus funciones son: • Síntesis, almacenamiento y transporte de proteínas: Se sintetizan en los ribosomas adheridos a la membrana del RER. Pueden quedarse en la membrana como proteínas transmembrana, o pasar al lumen para ser exportadas a otros destinos, incluido el exterior celular. • Glucosilación de proteínas: La mayor parte de las proteínas sintetizadas y almacenadas en el RER, antes de ser transportadas a otros orgánulos citoplasmáticos, a la membrana o al exterior celular, deben ser glucosiladas (uniéndose a un oligosacárido) para convertirse en glucoproteínas. Este proceso se inicia en el lumen del RER y se termina en el Aparato de Golgi. b’’.- Retículo endoplasmático liso(REL): Carecen de ribosomas adheridos en su cara externa. Su membrana forma una fina red de túbulos, que se conecta con las cisternas del RER. Suele ser escaso, pero abunda en células musculares estriadas, en las especializadas en el metabolismo lipídico, como las de la corteza suprarrenal y en los hepatocitos. Sus funciones son: • Síntesis, almacenamiento y transporte de lípidos: Sobre todo de los de membranas (fosfolípidos y colesterol ) y de esteroides (a partir de colesterol). 9 • Procesos de detoxificación: Metabolizando sustancias tóxicas liposolubles (pesticidas, conservantes, carcinógenos...) y compuestos perjudiciales producidos por el metabolismo, para evitar que se acumulen en las membranas celulares, y transformándolos en compuestos hidrosolubles que pueden abandonar la célula y ser eliminados por la orina. Esta función se realiza en los hepatocitos, y también en piel, pulmones, riñones e intestino. • Almacenamiento de Ca++: Tiene lugar en las células musculares estriadas, lo que le permite intervenir en el proceso de contracción muscular. Además de las funciones específicas de ambos tipos de retículo, en general, el RE proporciona membranas para la renovación de los otros sistemas membranosos y tiene un papel estructural, como soporte mecánico, proporcionando un sostén adicional al citoplasma. b’.- Aparato de Golgi: Es un complejo membranoso cuya localización y desarrollo varían en los distintos tipos celulares, y con el estado funcional de cada célula. Así, está muy desarrollado en células animales secretoras (glándulas), y muy poco en células vegetales. Suele situarse cerca del núcleo y, en las células animales, alrededor del centrosoma. Está constituido por una o varias unidades estructurales, denominadas dictiosomas, cada una de las cuales está constituida por la agrupación de varios (mínimo 4) sáculos aplanados, con forma de disco bicóncavo ensanchado por los extremos, de donde se van desprendiendo vesículas de distinto tamaño. Los dictiosomas presentan dos caras bien diferenciadas: • Cara cis: La más cercana al RE, del cual se desprenden vesículas (de transición), que pasan a fusionarse con los sacos del dictiosoma de esta cara, incorporándose, por tanto, compuestos del RE al Aparato de Golgi. • Cara trans: Opuesta a la anterior. Por ella se liberan vesículas de secreción que contienen los productos de la actividad del Aparato de Golgi y que se dirigen a la membrana plasmática (con la que se unen), se quedan dentro de la célula, constituyendo lisosomas, o forman gránulos de almacenamiento o de secreción que vierten al exterior el contenido que transportan. Sus principales funciones son: • Glucosilación de lípidos y proteínas: Puede realizarse entera o tan sólo completarse. Los lípidos y proteínas proceden del RE, llegan a través de vesículas de transición a la cara cis, y emigran, también mediante vesículas, de sáculo en sáculo (sufriendo transformaciones en cada uno), hasta la cara trans, saliendo finalmente en el interior de vesículas liberadas por esta cara. • Sulfatación y fosforilación de cadenas glucídicas, es decir, se añaden grupos sulfatos y fosfatos a determinados monosacáridos de glucoproteínas. • Renovación y mantenimiento de las membranas. Así, pe., la membrana plasmática se renueva a partir de vesículas liberadas en la cara trans, que se unen a la superficie celular. 10 c’.- Lisosomas: a’’.- Estructura: Son vesículas rodeadas por una membrana, presentes en todas las células eucariotas, y cuyo interior contiene gran cantidad de enzimas hidrolíticas (hidrolasas), capaces de digerir la mayoría de las macromoléculas biológicas, por lo que la función general de los lisosomas va a ser realizar la digestión celular, bien de moléculas captadas del exterior, bien de moléculas obsoletas de la propia célula. Las hidrolasas de los lisosomas son activas en medio ácido (pH cercano a 5), por lo que existe una “bomba” de hidrogeniones en la membrana de los lisosomas, que transporta H+ procedentes del citosol hacia el interior, gastando ATP, ya que se trata de un transporte activo. b’’.- Tipos: Según su origen y función existen distintos tipos de lisosomas: a´´´.- Lisosomas Primarios: Se originan a partir del Aparato de Golgi y todavía no han intervenido en ningún proceso de digestión. Poseen un contenido homogéneo. Pueden fusionar su membrana con la plasmática (exocitosis) y verter sus enzimas al exterior celular, para que digieran material extracelular, o pueden fusionarla, dentro de la célula, con otras vesículas portadoras de materiales exógenos o endógenos y convertirse en lisosomas secundarios. b´´´.- Lisosomas Secundarios: Resultan de la fusión de un lisosoma primario con vesículas que contienen material de naturaleza variable. Están implicados en la digestión celular. Según el material con el que se fusionan, existen 3 tipos: • Fagolisosomas: Se originan en células especializadas en fagocitosis (neutrófilos o macrófagos, p.e.). La sustancia fagocitada (fagosoma) se fusiona con un lisosoma primario, dando lugar al fagolisosoma, en el cual se procede a la digestión intracelular. Así se alimentan, por ejemplo, muchos protoctistas, y también llevan a cabo una función defensiva, pues es un mecanismo de destrucción de organismos patógenos (bacterias, p.e.). • Autofagolisosomas: Se forman tras la unión de un lisosoma primario con un autofagosoma, que, a su vez, se ha originado al envolver el RE a un orgánulo o resto celular que ha de ser digerido. Así se elimina el exceso de macromoléculas, o se destruyen zonas lesionadas o envejecidas. También es el mecanismo mediante el que se producen determinados procesos de la metamorfosis de insectos o anfibios. La autofagia asegura tanto el recambio de los componentes celulares como la nutrición de la célula en condiciones de ayuno, en que consume sus propios materiales. • Endolisosomas: Formado por la fusión de un lisosoma primario con vesículas procedentes de la endocitosis, que contienen sustancias que se han incorporado desde el exterior celular En cualquiera de los casos anteriores, terminada la digestión, los productos atraviesan la membrana lisosomal y son aprovechados por la célula. Si la digestión es incompleta, queda una pequeña formación con sustancias no digeridas, denominada cuerpo residual, que puede permanecer en el interior celular o ser eliminado por exocitosis. 11 d’.- Vacuolas: Son orgánulos característicos de las células eucariotas vegetales, aunque no exclusivos de ellas. Se presentan como vesículas de gran tamaño, llenas de agua con sustancias disueltas y rodeadas de una membrana denominada tonoplasto. En las células jóvenes se forman pequeñas vesículas derivadas del RE que, conforme la célula madura y se diferencia, se unen entre sí, hasta formar una o varias vacuolas que ocupan casi todo el citoplasma de las células adultas. Sus funciones son variadas: • Almacén de sustancias: de reserva (almidón, proteínas, grasas...), productos de desecho que resultarían tóxicos en el citosol (esencias, nicotina, opio, resinas, cristales de sustancias como el oxalato...), pigmentos antocianos en las células de los pétalos de las flores... Las moléculas nocivas se eliminan cuando una planta es comida o dañada, proporcionando un sistema de defensa contra los depredadores. • Mantenimiento del volumen y la turgencia celular: La presión osmótica en el interior de las vacuolas es muy alta, debido a la elevada concentración de sustancias. El agua tiende a penetrar en las vacuolas por ósmosis, para equilibrar la presión osmótica, por lo que la célula se mantiene turgente. • Crecimiento celular: El aumento de tamaño de las células vegetales se debe, en gran parte, a la acumulación de agua en sus vacuolas, lo que supone un sistema muy económico para el crecimiento celular. • Vacuolas contráctiles: Características de protoctistas unicelulares de agua dulce. Al ser el medio hipotónico, el agua entra constantemente en la célula por ósmosis, pudiendo llegar a estallar. La vacuola contráctil se encarga de eliminar este exceso de agua: se va llenando de agua del citoplasma y cuando alcanza un cierto tamaño, se fusiona con la membrana plasmática de la célula, y vierte su contenido al exterior, volviendo nuevamente a iniciarse la fase de llenado. e’.- Peroxisomas: Presentes en todas las células eucariotas, se tratan de vesículas de menor tamaño que los lisosomas, que contienen enzimas de oxidación (peroxidasas y catalasas). Las catalasas a menudo están cristalizadas. Su membrana proviene del REL, del que se desprende por estrangulación, incorporándose posteriormente los enzimas que se sintetizan en polirribosomas libres del citosol. Entre sus funciones están: • Las Peroxidasas, al oxidar sustratos, reducen el oxígeno a peróxido de hidrógeno (H2O2 = Agua oxigenada) RH2 + O2 R + H2O2 Posteriormente, la catalasa descompone el agua oxigenada (que es muy tóxica para las células) en agua y oxígeno, o bien la utiliza para oxidar a diferentes sustancias como fenoles o alcoholes: H2O2 + AH2 2 H2O2 2 H2O + A 2 H2O + O2 12 • Debido a su actividad enzimática, intervienen en numerosos procesos metabólicos como: - Biosíntesis de lípidos, como el colesterol (también sintetizado por el REL). - Fotorrespiración de las células vegetales (mediante la que las plantas obtienen glúcidos consumiendo O2 y liberando CO2). - Procesos de detoxificación que tiene lugar en algunos tejidos, como la degradación del etanol en el hígado de vertebrados, transformándolo en acetaldehído. - β – oxidación de ácidos grasos que se produce en semillas en germinación, transformando lípidos en azúcares. Estos peroxisomas se denominan Glioxisomas. g.- Mitocondrias: Son orgánulos, normalmente de forma cilíndrica, aunque son plásticos y se deforman, que existen en todas las células eucariotas en nº elevado (de varios cientos a varios miles), distribuidas por el citoplasma. a’.- Estructura: Una mitocondria presenta los siguientes elementos: • Membrana externa: Muy permeable, pues presenta unos canales formados por la proteína “porina”, que permite el paso de sustancias desde el citosol. De • Espacio intermembranoso: composición similar al citosol (debido a la permeabilidad de la membrana externa), y con menos enzimas que la matriz. • Membrana interna: Presenta repliegues hacia el interior denominados “Crestas mitocondriales”, que suelen disponerse perpendiculares al eje mayor de la mitocondria y que aumentan la superficie de la membrana. Se trata de una membrana prácticamente impermeable a las sustancias polares e iones, y muy especializada, compuesta por fosfolípidos especiales (cardiolipinas), y de proteínas transportadoras, tanto de los distintos compuestos que son necesarios en la matriz (ADP, piruvato, ácidos grasos...) o en el exterior (ATP...), como las que forman la llamada “Cadena respiratoria” • Partículas F: Estructuras globosas con una especie de tallito anclado en el seno de la membrana interna, en la que se han aislado enzimas ATP - sintetasas. • Matriz mitocondrial: Es un sistema coloidal complejo; entre sus componentes, además de agua, destacan: Enzimas del ciclo de Krebs y de la β-oxidación de ácidos grasos; ADN, en forma de doble cadena cerrada sobre sí misma, que contiene la información genética necesaria para la síntesis de ARN y proteínas mitocondriales; Ribosomas 70s, similares a los de las bacterias y responsables de la síntesis de las proteínas mitocondriales; Enzimas que regulan y controlan la replicación, transcripción y traducción del ADN mitocondrial; otro tipo de compuestos, como nucleótidos o iones. b’.Función: Las mitocondrias son los orgánulos encargados de llevar a cabo la respiración celular, es decir, la oxidación de moléculas orgánicas para obtener energía en forma de ATP. De todas las fases de la 13 respiración, en las mitocondrias se produce la β-oxidación de ácidos grasos, el ciclo de Krebs, la cadena respiratoria y la fosforilación oxidativa, procesos que estudiaremos en el siguiente tema. h.- Plastos: Son orgánulos citoplasmáticos exclusivos de las células eucariotas vegetales. Su tamaño, forma y número son muy variables, aunque, por lo general, son de mayor tamaño que las mitocondrias y su número suele oscilar entre 20 y 40, en los casos en que son más abundantes (a veces existe uno tan sólo). a’.- Tipos: Se clasifican en dos grandes grupos: a’’.- Leucoplastos: Carecen de pigmentos y en la mayoría de los casos almacenan diversas sustancias de reserva, como almidón (amiloplastos), grasas (oleoplastos) y proteínas (proteoplastos). Se localizan en las células vegetales que forman los cotiledones, los esbozos foliares del tallo y ciertas zonas de la raíz. b’’.- Cromoplastos: Son plastos que llevan en su interior un pigmento que les da color. Así, los que contienen carotenos son de color rojo y amarillos los que contienen xantofilas. A ellos se deben los colores anaranjados o rojos de flores, raíces y frutos. Los que almacenan clorofila constituyen el grupo principal de plastos, los cloroplastos, que merecen capítulo aparte. b’.- Estructura de los Cloroplastos: Los estudios al microscopio electrónico han revelado que los cloroplastos están constituidos por tres componentes: • Envoltura: Constituida por una membrana externa muy permeable, y una interna, mucho menos permeable, con proteínas de transporte especiales. Entre ambas queda un espacio intermembranoso. • Tilacoides: Son sacos aplanados que se disponen apilados e interconectados entre ellos, formando una red interna membranosa. Cada sáculo está constituido por una membrana (membrana tilacoidal), que rodea un compartimiento (espacio tilacoidal). La membrana interna del cloroplasto se continúa con la de los tilacoides únicamente durante el desarrollo del orgánulo; en el cloroplasto maduro desaparece esta continuidad. Los tilacoides pueden presentarse como pequeños sacos lenticulares, denominados tilacoides de grana, porque se agrupan en pilas denominadas grana, o como vesículas alargadas (tilacoides de estroma), que atraviesan el estroma e interconectan los grana. Mediante técnicas especiales se ha deducido la ultraestructura molecular de estas membranas de los tilacoides. Además de la bicapa lipídica y las proteínas de cualquier membrana, existen una serie de partículas que sobresalen ligeramente de la superficie, y de las que existen 2 tipos: unas esféricas más pequeñas llamado Fotosistema I (Fs I) y otras ovoides más grandes o Fotosistema II (Fs II). Los dos fotosistemas, aso-ciados a proteínas, contienen las moléculas de pigmentos implicados en captar la energía luminosa (clorofila α y 14 β y carotenoides). Por último, formando parte de la membrana tilacoidal existen proteínas de la cadena fotosintética de transporte electrónico y partículas F semejantes a los de las crestas mitocondriales, que también en este caso contienen ATP - sintetasa. • Estroma: Queda delimitado por la membrana interna del cloroplasto, y es equivalente a la matriz mitocondrial. Contiene una disolución concentrada de enzimas, una o más copias de ADN circular, ARN y ribosomas 70s, que intervienen en la síntesis de algunas proteínas del cloroplasto (la mayoría se sintetiza en el citosol). Entre las enzimas están las responsables del Ciclo de Calvin, las responsables de la reducción y asimilación de nitratos y sulfatos y las que regulan y controlan la replicación, transcripción y traducción del ADN del cloroplasto. También pueden encontrarse sustancias insolubles como granos de almidón o glóbulos lipídicos. c’.- Función de los Cloroplastos: Las principales funciones que realizan son: • Fotosíntesis: Este proceso tiene lugar en dos fases: la fase luminosa, dependiente de la luz y que se produce en membrana tilacoidal, mediante la que se obtiene ATP y NADPH, y la oscura, independiente de la luz y que se produce en el estroma, que emplea la energía producida en la fase luminosa en la fijación de CO2 y en la formación de glúcidos, grasas y aminoácidos. Ambas fases se estudiarán en el tema siguiente. • Biosíntesis de ácidos grasos, utilizando los glúcidos, el NADPH y el ATP sintetizados. • Reducción de nitratos a nitritos. Los nitritos, a su vez, se reducen a amoniaco (NH3), que es la fuente de nitrógeno para la síntesis de aminoácidos y de los nucleótidos. • Almacenamiento temporal de almidón: En el estroma del cloroplasto se almacena glucosa en forma de granos de almidón. • Síntesis de algunas proteínas del cloroplasto. La información se encuentra en el ADN del cloroplasto, y la síntesis tiene lugar en los ribosomas 70s del orgánulo. i.- Núcleo: a´.- Características generales: Puede considerarse como el orgánulo principal de la célula eucariótica, ya que en él reside el material genético (ADN), por lo que es responsable de la transmisión del mismo a través de las generaciones, y desde él se controla toda la actividad celular. Se encuentra inmerso en el citoplasma, separados por la envoltura nuclear. Fisiológica y estructuralmente se reconocen en el núcleo dos estados: núcleo en reposo o interfásico, que es el estado que presenta entre divisiones, y núcleo en división o mitótico, en el que se han estructurado los cromosomas. b’.- Núcleo interfásico: a’’.- Características generales: • Forma: Por lo general, es esférica, aunque puede serlo también ovoide (células prismáticas) o polilobulado (macrófagos, p.e.). • Posición: Suele estar situado en posición central, aunque hay numerosas excepciones, como las células adiposas que lo presentan lateral o las de los epitelios secretores que lo tienen basal. • Número: Habitualmente existe un núcleo por célula. En ocasiones son dos (paramecios, p.e.) o muchos (sincitios resultantes de la fusión de varias células uninucleadas como sucede con las fibras musculares esqueléticas, o plasmodios que proceden de divisiones sucesivas del núcleo sin que se divida el citoplasma); los glóbulos rojos de mamíferos son anucleados. 15 • Tamaño: Oscila entre 5 y 25 µm de diámetro, aumentando considerablemente antes de realizarse la división celular, debido a la duplicación del material genético. La relación existente entre el volumen nuclear y el citoplasmático se expresa mediante un coeficiente, que viene definido por la fórmula: Vnúcleo K= Vcélula - Vnúcleo Cuando k alcanza un valor mínimo, constante para cada tipo celular, la célula se divide. b’’.- Componentes: En la estructura del núcleo interfásico se distinguen las siguientes partes: a´´´.- Envoltura nuclear: Aísla al núcleo del citoplasma y está formada por dos membranas, la externa hacia el citoplasma, y la interna hacia el núcleo, separadas por un espacio intermembranoso. La externa presenta adheridos a su superficie ribosomas, y tiene continuidad con las cisternas del RE. Las dos membranas se fusionan en algunos puntos, originando unos orificios conocidos como Poros nucleares, cuyo nº varía en función de la actividad transcriptora de ADN. La cantidad media en mamíferos es de 3.000. Al microscopio electrónico se comprueba que los poros no son simples orificios, sino estructuras más complejas, constituidas por un anillo o estructura cilíndrica formada por 8 proteínas globulares, un gránulo en el centro del complejo (ribosomas recién formados o sustancias en tránsito) y fibrillas proteicas uniendo todas las estructuras. Los poros nucleares permiten el intercambio núcleocitoplasma, pero no de forma indiscriminada. b´´´.Nucleoplasma: Se denomina así al medio interno nuclear. Se trata de un sistema coloidal formado por agua y numerosas sustancias, principalmente nucleótidos, aminoácidos y proteínas, de las cuales unas forman un entramado de fibras, que tienen un papel similar al citoesquelo, manteniendo al nucléolo y la cromatina, y otras son enzimas relacionadas con la síntesis de los ácidos nucleicos, ya que en este medio se realizan la replicación y la transcripción. c´´´.- Nucléolos: Son estructuras siempre presentes, de gran tamaño (se observan a microscopía óptica) y bien diferenciados del resto de los componentes del núcleo interfásico. Se presentan en nº variable, aunque en células diploides lo más habitual es que haya 1 ó 2. Su función es la síntesis del ARNr y el ensamblaje de los ribosomas. Estructuralmente, y a microscopía electrónica, se distinguen dos zonas: 16 • Zona fibrilar: Ocupa la parte central del nucléolo. Contiene cromatina con los genes ribosómicos, es decir, aquellos que codifican el ARN de los ribosomas (y que se conocen como organizadores nucleolares) y las moléculas de ARNr resultantes de la transcripción de dichos genes. • Zona granular: Ocupa la parte más periférica. Está constituida por gránulos de ribonucleoproteínas, precursoras de los ribosomas, en distintas fases de maduración. Al iniciarse la división nuclear los nucléolos desaparecen, constituyendo las constricciones secundarias de los cromosomas. d´´´.- Cromatina: El ADN de las células eucariotas no se encuentra libre, sino asociado con proteínas. Esta asociación de ADN y proteínas recibe el nombre de cromatina que, por tanto, es un complejo de nucleoproteínas. Las proteínas pueden ser de dos tipos: • Histonas, proteínas de carácter básico, debido a la abundancia de los aminoácidos arginina y lisina, ambos cargados positivamente. Son pequeñas y de carácter básicamente estructural. • No Histonas: Muy heterogéneas, y con funciones varias, entre las que destacan las enzimas encargadas de la replicación, transcripción y regulación del ADN. Existen también proteínas contráctiles, probablemente relacionadas con el proceso de condensación de la cromatina en cromosomas y la movilidad de éstos durante el proceso de división. Se distinguen dos clases de cromatina: • Eucromatina: Es la más abundante en la interfase, y es la cromatina menos compactada. En ella se realizan los procesos de transcripción activa. Equivaldría al cromosoma funcional. • Heterocromatina: Es la de mayor grado de compactación y aparece en la interfase como agrupaciones teñidas: Los cromocentros. No se transcribe. Estudios realizados mediante microscopía electrónica, difracción de rayos X y ensayos bioquímicos han permitido conocer la ultraestructura de la cromatina. Está formada por una serie de unidades repetitivas, en forma de cuentas de rosario, denominadas Nucleosomas. Cada uno de ellos está constituido por 8 unidades de histonas iguales 2 a dos, formando un núcleo alrededor del cual se dispone la doble hélice de ADN, dando 1 vuelta y 3/4. Las unidades se enlazan entre sí por fragmentos de la hebra de ADN (de unión). Este filamento (de 10 nm), a su vez se pliega. En un primer paso los nucleosomas se unen por su cara interna, mediante una histona, alcanzando un diámetro de 30 nm. Esta fibra, a su vez, se puede espiralizar sucesivamente, hasta llegar a la superespiralización en el momento de iniciar la mitosis, en el que la cromatina forma los cromosomas. Por tanto, cromatina y cromosoma son el mismo material con distinto grado de empaquetamiento, y se corresponden con dos momentos o estados funcionales de la célula: la interfase y la división celular, respectivamente. c’.- Núcleo mitótico: Durante la división celular el núcleo sufre una serie de cambios destinados a que las células hijas resultantes posean la misma dotación genética. El nucléolo y la membrana celular desaparecen, al tiempo que la cromatina se va condensando hasta formar los cromosomas. De esta manera, la única estructura característica que queda en el núcleo en división son los cromosomas. a’’.- Cromosomas: El estudio de los cromosomas suele hacerse durante la metafase de la mitosis, ya que es entonces cuando alcanzan el mayor grado de condensación, adquiriendo el aspecto de bastón cilíndrico, con una forma y tamaño característico de cada uno. 17 a´´´.- Estructura: En un cromosoma se distinguen las siguientes partes: • Cromátidas: Cada una de las dos subunidades en que aparece dividido longitudinalmente un cromosoma, y que están constituidas por una copia del ADN duplicado al final de la interfase. Las dos se sitúan en paralelo y separadas entre sí, excepto a nivel del centrómero, por donde permanecen unidas. • Centrómero o constricción primaria: Divide al cromosoma en dos brazos de igual o de diferente tamaño. Ocupa una posición variable a lo largo del cromosoma, pero fija para cada cromosoma. Contienen heterocromatina. A ambos lados del centrómero, y sobre cada una de las dos cromátidas se localiza una estructura de naturaleza proteica, denominada cinetocoro, que constituyen los puntos de unión de los microtúbulos del huso al cromosoma, e intervienen en la separación de los cromosomas durante la anafase. Por cada cromátida suele aparecer un único cinetocoro. • Constricciones secundarias: También conocidos como organizadores nucleolares. Son zonas más estrechas identificables en los brazos, y en las que se localizan los genes que se transcriben como ARNr, por lo que permiten la formación del nucléolo al final de cada mitosis y de los ribosomas. En ellas nunca aparece cinetocoro. • Telómeros: Localizados en los extremos de los cromosomas, se corresponden con los extremos de la molécula de ADN en los que se repiten una determinada secuencia de ADN. Confieren estabilidad al cromosoma, impidiendo que los extremos de los cromosomas se adhieran entre sí y sufran cambios estructurales. También son esenciales en la duplicación del cromosoma, y protegen a los cromosomas contra las nucleasas (enzimas que digieren el ADN). • Satélites: Es una porción esférica situada en un extremo y unida al brazo por un fino filamento (trabécula). Es constante en tamaño y forma para cada cromosoma que lo presenta (no todos lo poseen). • Bandas: Son segmentos de cromatina que se colorean con diferente intensidad, y que permiten una identificación inequívoca de los cromosomas homólogos. b´´´.- Tipos: Según la posición del centrómero se distingue entre los siguientes tipos: • Metacéntricos: El centrómero está más o menos en el centro, por lo que los dos brazos son casi iguales. • Submetacéntricos: Uno de los brazos es algo mayor que el otro. • Acrocéntricos: Los brazos presentan muy diferente longitud: Uno muy corto y otro muy largo, debido a que el centrómero se sitúa muy cerca del extremo del cromosoma. • Telocéntricos: El centrómero está tan próximo a uno de los telómeros que prácticamente sólo existe un brazo. En el cariotipo humano no existen cromosomas de este tipo. • Cromosomas gigantes: Son casos excepcionales, de cromosomas de gran tamaño. Se conocen dos tipos diferentes: los plumosos y los politénicos. c´’´.- Cariotipo: Se denomina así al conjunto de todos los cromosomas metafásicos de una célula, y viene caracterizado por su número, tamaño y forma. El cariotipo es característico de cada especie. Se suele representar mediante Idiogramas, en los que los cromosomas se ordenan por parejas de mayor a menor, permitiendo una fácil identificación de cualquier anomalía cromosómica en las células de un individuo. Recordemos, por último, que una especie es diploide (2n) si presenta sus cromosomas iguales dos a dos (cromosomas homólogos) y haploide (n) si todos sus cromosomas son diferentes. 18 4.- EVOLUCIÓN CELULAR: Actualmente se piensa que todos los organismos y, por tanto, todas las células descienden de un tipo de célula primitivo. Ahora bien, probar cuál ha sido el origen de esta célula y cuáles los pasos evolutivos desde el principio hasta la actualidad es complicado, por lo que, en realidad, sólo poseemos algunas hipótesis tan difíciles de comprobar como de refutar. Se cree que al enfriarse la Tierra, se formó una atmósfera reductora en la que abundaban compuestos inorgánicos, como el CO2, NH3 o el H2, que se disolverían en el agua de mares y océanos. Estos compuestos, mediante descargas eléctricas procedentes de relámpagos y de otros fenómenos producidos en la atmósfera primitiva, formarían moléculas orgánicas tales como aminoácidos, nucleótidos, azúcares y ácidos grasos. Para que surgiera la primera célula viva fue necesario que apareciera una membrana mediante el ensamblaje de fosfolípidos que aislara a las moléculas formando los denominados “coacervados”, dentro de los cuales se producirían diversas reacciones químicas en condiciones más favorables que en el exterior. Si en este medio existían moléculas de ARN, se autorreplicarían, y también dirigirían la síntesis de pequeños polipéptidos. En algún momento el material genético se transferiría a ADN, mucho más estable. Este tipo de célula primitiva (procariota ancestral) debió aparecer hace unos 3.800 ó 4.000 m.a., y sería procariota, heterótrofa y anaerobia (en el caldo primitivo abundaba la materia orgánica y escaseaba el oxígeno). De la procariota ancestral derivarían tres tipos de células: • La procariota fotosintética, capaz de aprovechar las grandes cantidades de CO2 producidas en la respiración (y otros gases como el N2) para fabricar materia orgánica. Las primeras aparecerían hace unos 3.700 m.a., pero hasta hace unos 2.000 m.a. no se empezó a acumular grandes cantidades de O2 liberado en la fotosíntesis, lo que debió llevar a la extinción de muchos anaerobios, que no pudieron adaptarse a la nueva composición de la atmósfera (el O2 es muy tóxico para los anaerobios). Otros debieron refugiarse en medios sin oxígeno y otros evolucionaron, apareciendo la respiración aerobia, que permitió que la composición de la atmósfera se equilibrase hace unos 1.500 m.a. Todos estos grupos de procariotas constituyen ahora las “Eubacterias”. • Algunos procariotas ancestrales mantuvieron sus características primitivas, y han llegado hasta nuestros días formando el grupo de las “Arqueobacterias”. • Otros procariotas anaerobios primitivos poseían una membrana plasmática flexible capaz de invaginarse y plegarse y, por tanto, pudieron formar compartimentos internos que dieron lugar al sistema de endomembranas. El núcleo se originaría de un plegamiento interno de la membrana que arrastró al ADN. Además, podrían incorporar partículas del exterior por fagocitosis. Se tendría así el tipo “eucariota ancestral”, anaerobio y heterótrofo, hace unos 2.000 m.a. En 1967 Lynn Margulis postuló que las mitocondrias y cloroplastos proceden de procariotas (aerobio en el primer caso y fotosintético en el segundo) que establecieron una relación simbiótica con una célula eucariota ancestral, cuando ésta la fagocitó para obtener alimento. Primero se produjo la endosimbiosis con el aerobio. La ventaja que obtendría la célula sería grande ya que el aerobio (futura mitocondria), gracias al oxígeno sería capaz de sintetizar ATP. De esta forma celular derivarían las formas pluricelulares de los animales, hongos y protozoos. Posteriormente, se establecería la relación con el procariota fotosintético, que pasaría a transformarse en cloroplastos, que suministra a la célula materia orgánica (a cambio de protección). De este modo aparecería la célula eucariota fotosintética, a partir de la cual evolucionaron las algas, y de estas últimas, las plantas. Las pruebas de estas endosimbiosis son cada vez más claras, tanto por el parecido entre bacterias y mitocondrias y cloroplastos (tamaños similares, reproducción por división binaria, ribosomas 70s, fragmento circulares de ADN ...), como por el descubrimiento de que existe intercambio genético entre estos orgánulos y el núcleo, lo que les hace depender totalmente unos del otro. 19 EVOLUCIÓN CELULAR Caldo caliente (CO2, NH3, H2) Descargas eléctricas M. O. sencilla (Aa, nucleótidos, azúcares, ác. grasos Cél.procariota ancestral (Heterótrofa, anaerobia) Coacervado s ADN Cél.procariota ancestral (Heterótrofa, anaerobia) (3.800 – 4.000 m.a.) ARQUEOBACTERIAS (ctcas. primitivas) Cél. procariota, fotosintética, anaerobia (CO2, N2 M.O.) (3.700 m.a.) Sistema de endomembranas Aislamiento del núcleo Fagocitosis Cél.eucariota ancestral (Heterótrofa, anaerobia) (2.000 m.a.) ARN Celula procariota, Heterótrofa, aerobia Endosimbiosis con Cél.eucariota (Heterótrofa, aerobia) procariota aerobio Cél. procariota, fotosintética, aerobia (2.000 m.a.) EUBACTERIA S Endosimbiosis con Cél.eucariota (Autótrofa, aerobia) procariota fotosintético ANIMALES HONGOS PROTOZOOS ALGAS PLANTAS UNIDAD DIDÁCTICA II (1ª Parte): ESTRUCTURA DE LA CÉLULA 1.- INTRODUCCIÓN 2.- TEORÍA CELULAR 3.- TIPOS DE CÉLULAS A.- Introducción B.- La célula procariota C.- La célula eucariota a.- Características generales b.- Envueltas celulares: a’.- Membrana plasmática: a’’.- Composición química: a’’’.- Lípidos de membrana b’’’.- Proteínas: c’’’.- Glúcidos: Glucocálix b’’.- Estructura c’’.- Funciones d’’.- Diferenciaciones: a’’’.- Microvellosidades: b’’’.- Uniones intercelulares b’.- Pared celular: a’’.- Composición química y estructura: a´´´.- Lámina media b’’’.- Pared primaria c‘’’.- Pared secundaria d’’’.- Especializaciones b’’.- Funciones c- Citosol o Hialoplasma d.- Citoesqueleto y sus derivados: a’.- Microfilamentos b’.- Filamentos intermedios c’.- Microtúbulos: a’’.- Centríolos y Centrosoma: a´´´.- Centríolo b´´´.- Centrosoma b’’.- Cilios y flagelos e.- Ribosomas: a’.- Estructura b’.- Localización c’.- Funciones f.- Sistema de endomembranas y sus derivados: a’.- Retículo endoplasmático: a’’.- Retículo endoplasmático rugoso b’’.- Retículo endoplasmático liso b’.- Aparato de Golgi c’.- Lisosomas: a’’.- Estructura b’’.- Tipos: a´´´.- Lisosomas Primarios b´´´.- Lisosomas Secundarios d’.- Vacuolas e’.- Peroxisomas g.- Mitocondrias: a’.- Estructura b’.- Función h.- Plastos: a’.- Tipos: a’’.- Leucoplastos b’’.- Cromoplastos b’.- Estructura de los Cloroplastos c’.- Función de los Cloroplastos i.- Núcleo: a´.- Características generales b’.- Núcleo interfásico: a’’.- Características generales b’’.- Componentes: a´´´.- Envoltura nuclear b´´´.- Nucleoplasma c´´´.- Nucléolo d´´´.- Cromatina c’.- Núcleo mitótico: a’’.- Cromosomas: a´´´.- Estructura b´´´.- Tipos c´’´.- Cariotipo 4.- EVOLUCIÓN CELULAR Idiograma femenino Idiograma masculino