Descargar - InvestigacionesHistoricaseuroAsiaticas
Anuncio

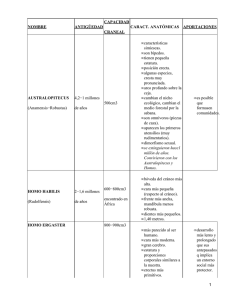
UN TRÁNSITO POR LA EVOLUCIÓN HUMANA. DE LOS HOMININOS ARCAICOS AL HOMBRE MODERNO Dr. Julio López Saco Escuela de Historia, UCV Doctorado en Ciencias Sociales, UCV Introducción Este ensayo desea servir de prolegómeno a una de las áreas de estudio más complejas a la que los investigadores deben asomarse: la paleo antropología. El estudio de la evolución humana ha progresado enormemente en las últimas décadas, aunque sigue siendo una de las disciplinas más dinámicas debido a los continuos nuevos hallazgos que van modelando un poco mejor el enrevesado tejido de la historia humana. No pretendemos aquí ni elaborar un pormenorizado resumen, ni proponer teorías de avanzada. Mucho menos, aspiramos a valorar y analizar cada hallazgo, ni siquiera referirnos a todas las especies. Nuestro interés se centrará en enfatizar los descubrimientos más recientes, algunos ya en este nuevo milenio, y ofrecer un aperitivo a los que deseen indagar en este proceloso y complejo escenario de la investigación científica y académica. El repertorio bibliográfico, de por sí ciclópeo y casi inabarcable, es buena prueba de lo mucho que se ha hecho. Sin embargo, queda mucho por descubrir y son numerosos los pasos que hay que seguir dando para obtener una radiografía más clara del arborícola esquema evolutivo del ser humano. Sirvan las siguientes líneas, por consiguiente, como una puesta al día y como un anzuelo para aquellos atrevidos que tengan como reto intelectual inmiscuirse en el todavía no muy claro ambiente de nuestros ancestros y de aquello que nos ha hecho humanos. Sahelanthropus, Orrorin y Ardiphitecus El primero de los posibles homininos más arcaicos conocidos es Sahelanthropus tchadensis, cuyos restos fueron descubiertos por Michel Brunet y su equipo desde al 2001, siendo datados, a través del método de cronología relativa denominado 1 biocronología, entre 7 y 6 millones de años. Estos vestigios fósiles fueron hallados en Toros-Menalla, en Chad. La evidencia geológica y paleontológica sugiere que Sahelanthropus vivía en un hábitat lacustre, de bosque bajo con herbáceas y en zonas ribereñas bordeadas de bosques. Se cuenta con un cráneo y dos mandíbulas. El cerebro de este espécimen era del tamaño del de un chimpancé. El segundo de los posibles homininos más arcaicos es Orrorin tugenensis, cuyo nombre proviene de los sedimentos de Tugen Hills, en el norte de Kenia, donde los fósiles fueron encontrados. Según la datación de potasio/argón, Orrorin tendía una antigüedad de 6 millones de años. El primer hallazgo se produjo en 1974, una corona molar, si bien ha sido recientemente, desde el año 2000, cuando han salido a la luz doce especímenes más. Sus descubridores fueron Martin Pickford y Brigitte Senut, paleo antropólogos del Collège de France en París. El fémur de tugenensis no es muy diferente del de los primates que se mueven entre los árboles, y la morfología de sus dientes es semejante a la de los simios. El tercero y el cuarto de los posibles homininos más antiguos corresponden al género Ardipithecus. El fósil más antiguo, datado entre 5.7 y 5.2 millones de años, corresponde a Ardipithecus kadabba, de la zona del Awash medio, en Etiopía. Los fósiles incluyen una mandíbula, algunos huesos pos craneales y dientes. Los caninos superiores, además de muchos otros aspectos del fósil, semejan los de los chimpancés. Los restos de Ardipithecus ramidus proceden del Awash medio y de Gona, ambas regiones etíopes. Se han datado entre 4.5 y 4 millones de años. La colección de fósiles consta de dientes, diversas mandíbulas, algunos pequeños huesos de pies y manos y la parte inferior de un cráneo. Sus caninos tienen una semejanza menor a la de los simios que los de kadabba. Ciertos rasgos vinculan a ramidus con los homininos, especialmente la posición del foramen magnum; sin embargo, tanto ramidus como kadabba eran similares a un chimpancé. De estos cuatro potenciales homininos, únicamente dos de ellos, Sahelanthropus tchadensis y Ardipithecus ramidus, tienen posibilidades reales de ser incluidos en la rama (clado) del árbol filogenético hominino. Austropitecinos más recientes: Kenyanthropus platyops y Australopithecus sediba El K. playtops es un homínido del este de África que apareció en 1999 en Lomekwi, al oeste del lago Turkana, entre rocas volcánicas del Mioceno. El cráneo KNM-WT 40000, datado en torno a 3.5. m.a., fue designado por el equipo de Leakey como Kenyanthropus, de la especie tipo platyops. Presenta un pequeño grado de prognatismo 2 subnasal, característico de los austrolopitecinos. Otros hallazgos vinculados corresponden a un maxilar (KNM-WT 38350) y a la presencia de algunos dientes. Aunque el tamaño y forma encajan con afarensis y africanus, el cráneo presenta algunas diferencias, mostrando rasgos cercanos a Homo rudolfensis (KNM 1470), lo que indicaría su posible ascendencia del género Homo. Algunos paleo antropólogos piensan, no obstante, en virtud de que una parte de la bóveda del cráneo sufrió una expansión diploica, con la presencia de materiales que expandieron el hueso, que estamos, en realidad, ante una variante afarensis. Este nuevo individuo pudiera ser, en definitiva, una variante de australopiteco, quizá un taxón hermano de afarensis, aunque también pudiera tratarse de un genus separado, quizá un ancestro de H. rudolfensis. El Australopithecus sediba u homínido de Malapa, fue recientemente descubierto (agosto del 2008), en Sudáfrica por Lee Berger, de la Universidad de Witwatersrand, pero fue dado a conocer en la revista Science en abril del 2010. El hallazgo se produjo en una cueva no lejos de los yacimientos clásicos sudafricanos de Sterkfontein, Swartkrans y Kromdraai. A. sediba tiene la cabeza y el tamaño del cerebro propio de un australopiteco, pero también posee rasgos derivados que lo acercan más al género Homo que ninguno de sus parientes, como son sus dientes reducidos y la nariz proyectada. Al mismo tiempo, no obstante, presenta brazos muy largos, como los de un orangután actual, de modo similar a los de otros australopitecos. Este nuevo homínido está representado por dos individuos (MH1 y MH2), bastante completos, un varón joven y una mujer adulta. Aparte de estos dos especímenes, se descubrieron otros dos individuos más, un infante y otra mujer adulta, aunque todavía no han sido publicados. La mujer se identificó gracias a la forma de su mandíbula y de la cadera. El análisis de los dientes probó que la edad de la fémina oscilaba entre los veinte y los treinta años al momento de su muerte. El varón tendría, por el contrario, unos doce. Se ha dicho que es probable que ambos fuesen conocidos e incluso parientes. Su estatura rondaba más o menos un metro veinte de altura. Se han conservado huesos del esqueleto pos craneal (no sólo del cráneo), aunque los restos más completos corresponden a un ejemplar inmaduro, lo que tiende a hacer gráciles los rasgos, dificultando con ello el análisis. Las piernas son largas y los tobillos parecen ser intermedios entre los primeros homínidos y los humanos modernos. La cadera y la pelvis se acercan más a la estructura del Homo erectus, según el propio Berger y sus colegas. Todo esto indica, en consecuencia, que A. 3 sediba podría caminar de forma erecta y con zancadas, forma muy eficiente de caminar y correr. Los Parántropos: Robustus y Boisei. ¿Un camino equivocado? El parántropo robustus (1.8 millones de años), fue un austrolopiteco sudafricano que desarrolló, como todos sus congéneres, una cara ancha, un poderoso aparato masticador y una cresta sagital, fruto de poderosos músculos mandibulares, necesarios para masticar vegetales muy duros y raíces, aunque no se puede descartar que comiera algunos animales. Su capacidad craneal apenas excedía de los 500 centímetros cúbicos. Algunos de los yacimientos en los que fueron hallados sus restos fósiles son Kromdraai, Gondolin y Swartkrans. El paleo antropólogo Louis Leakey, reportó el hallazgo, en 1959, de una nueva especie, denominada Zinjanthropus, en la garganta de Olduvai, en Tanzania. Este espécimen fue datado en 1,8 millones de años. Se trataba de un cráneo casi completo que fue bautizado como Dear Boy, debido a que correspondía a un individuo juvenil al que los terceros molares aún no le habían salido por completo. Otros hallazgos posteriores de esta especie se produjeron en el norte de Tanzania, en el norte de Kenia y en el sur de Etiopía. Convivieron con algunos Homo durante 1,5 millones de años. Descendiente de Paranthropus aethiopicus, su capacidad craneal no sobrepasaba los 500 c.c. Su cara estaba muy reforzada, con pequeños incisivos, pero con unos enormes molares y una cresta sagital a la que se unían grandes músculos masticadores. Su foramen magnum se encontraba más adelantado que en los australopitecos. El desarrollo de unas poderosas mandíbulas trajo como consecuencia su acceso a raíces y tallos gruesos, así como una especialización en ese tipo de alimento. No obstante, el resto de su estructura corporal no varió prácticamente nada con respecto a sus antecesores australopitecos. Los homínidos en transición: Homo hábilis y Homo rudolfensis Los primeros hallazgos de lo que hoy se conoce como Homo hábilis se llevaron a cabo en 1960 en la garganta de Olduvai, Tanzania, por parte de Louis y Mary Leakey. Estos primeros restos consistieron en algunos huesos de las manos, muchos de los pies y parte de una cima craneal. Desde un principio, hubo dudas al respecto de si estos vestigios pertenecían a una primitiva especie del género Homo o si correspondían a un hominino arcaico de mayor cerebro. En 1961 fue encontrado un cráneo un completo de un 4 adolescente, dientes y una mandíbula inferior. El cráneo no mostraba una cresta semejante a la de los cráneos de los individuos Paranthropus Boisei, en tanto que sus molares y premolares eran considerablemente más pequeños que los de Boisei. Estos nuevos hallazgos motivaron que Louis Leakey, el anatomista John Napier y Phillip Tobias argumentaran que se podía justificar el establecimiento de una nueva especie, Homo hábilis, dentro del género Homo, a pesar de que el cerebro de este último descubrimiento estaba por debajo (600-700 c.c.) de la medida considerada estándar para el género (750 c.c.), y a pesar también de que los fósiles desenterrados en Koobi Fora, en Kenia, parecían evidenciar una estimación de la longitud de sus miembros no diferente de la de aquellos de los Australopithecus afarensis. En términos generales existen pocas distinciones entre Homo hábilis y los homininos arcaicos australopitecinos. A través del tamaño de la mandíbula y de los dientes, que sirven para estimar el tamaño del cuerpo, se evidencia que hábilis es más similar a los australopitecos que a los Homo posteriores. Además, el esqueleto pos craneal de hábilis apenas difiere del de los australopitecos y de los parántropos. Si bien la presencia de huesos de la mano sugieren que hábilis fue capaz de poseer ciertas destrezas manuales necesarias para la manufactura y uso de herramientas simples de piedra, también tal habilidad es aplicable a Australopithecus afarensis y a Paranthropus robustus. Lo cierto, sin embargo, es que cráneos, mandíbulas y dientes de Homo hábilis son más variables de lo que cabría esperar en una simple especie. Es por eso que muchos investigadores dividen los restos en dos especies: Homo hábilis propiamente dicho, y Homo rudolfensis. Este último posee un cerebro más grande, en torno a 700-800 c.c. y una cara más alargada, lo que sugiere que su dieta pudo haber diferido de la de hábilis. Los grandes “Homo”: ergaster, heidelbergensis y antecessor Homo ergaster (1.8 m.a.), fue descubierto en 1975 por Bernard Ngeneo, C. Groves y V. Mazak. Aunque algunos paleo antropólogos creen que corresponde a un antiguo erectus africano, es probable que ergaster sea un vínculo directo con los hombres modernos. Entre los especímenes más relevantes se encuentran el KHM-HEU 3733 y el fósil KNM-WT 15000, llamado Niño Nariokotome o Niño de Turkana. Este último fue hallado por K. Kimeu en 1984, en la localidad de Niriokotome, Kenia. Su primera descripción apareció en la revista Nature un año después, siendo considerado como un 5 Homo erectus. Se trata de un joven de unos 13 años con una clara habilidad para desplazarse bípedamente, a pesar de algunas semejanzas cercanas a los austropitecinos. Homo heidelbergensis es un individuo alto y fuerte, con un cráneo de gran capacidad, en torno a los 1300 c.c. Presenta una gran abertura nasal y mandíbulas salientes. Se le considera una especie intermedia entre el Homo antecessor y el Neandertal. Cronológicamente, sus fósiles datan entre 600000 y 250000 años, tanto en Europa como en África. El resto fósil que le ha dado nombre es una mandíbula hallada en Mauer. Otros restos significativos son los de Bodo, en Etiopía, Kabwe en Zambia, Dali en China y Swascombe en el Reino Unido, entre otros. En el yacimiento de Terra Amata, en Francia, se hallaron restos de refugios construidos durante la época de esta especie. Homo antecessor (800000 años), fue descubierto en 1997 por el equipo compuesto por Bermúdez de Castro, J.L. Arsuaga, E. Carbonell, A. Rosas, I. Martínez y M. Mosquera, en el yacimiento de Atapuerca, Burgos, concretamente en la Gran Dolina. Esta especie presenta rasgos muy arcaicos que recuerdan otras tan antiguas como el Homo hábilis, diferenciándose claramente de los nuestros. Sin embargo, sus huesos faciales son similares a los de Homo sapiens, poseyendo, en consecuencia, una cara muy moderna. Con una capacidad craneal en torno a los 1000 c.c, puede considerarse el eslabón que une al Homo ergaster y enlaza con las formas más cercanas a nosotros. Si aceptamos el origen africano de nuestra especie hace unos 150000 o 200000 años, debemos también aceptar un origen africano para esta especie, muy probablemente a partir de poblaciones pertenecientes a H. ergaster. Del mismo modo es sumamente probable una continuidad evolutiva de H. antecessor en África, que culminaría con la aparición de las poblaciones humanas modernas. Una rareza: Homo floresiensis Homo floresiensis es una especie descubierta en la caverna denominada Liang Bua, en la isla de Flores, en Indonesia, en el año 2003. El fósil tiene apenas un metro de altura y es bípedo, siendo el volumen de su cráneo de apenas unos 400 centímetros cúbicos, semejante al de un australopiteco o un chimpancé. Su cronología se estima entre 40 y 18 mil años, aunque algunos paleo antropólogos creen que pudo vivir en la zona hace unos cien mil. Asociado con el fuego y diversas herramientas pétreas, cazaba pequeños elefantes, endémicos de la región, así como roedores. El tipo de herramientas con las 6 que aparece vinculado han aparecido en depósitos de cronologías semejantes y confeccionadas por Homo erectus. Hoy se cree que es una forma enana de Homo erectus. En un principio, se creyó que los vestigios de Homo floresiensis correspondían a un ser deforme, enano o enfermo de microcefalia. Pero tras diferentes y concienzudos estudios, se probó que floresiensis corresponde a una especie nueva, y no a un individuo enfermo. También se pensaba que había evolucionado directamente de australopiteco en virtud de lo arcaico de sus huesos corporales y por su diminuto cráneo. Sin embargo, los análisis morfométricos del fósil LB1, comparado con los de los Homo sapiens, Homo sapiens insulares, como el hombre de Minatogawa, con Homo sapiens con microcefalia, Homo hábilis, Australopithecus africanus, Homo erectus asiático y cráneos neolíticos, han demostrado la cercanía de Homo floresiensis con Homo erectus. No es similar a ningún sapiens conocido y está bastante alejado de los australopitecos. El fósil LB1 consiste en un cráneo bastante completo y un esqueleto parcial, con presencia de partes de las piernas, pelvis, manos y pies. Corresponde a una mujer adulta de unos treinta años. Una breve historia del hombre moderno Los primeros hallazgos de homininos anatómicamente modernos se produjeron en Europa. Se trata del esqueleto de la Dama Roja, en una caverna en la península Gower, al oeste de Swansea, en Gales (1822-1823) y del Cromagnon de Les Eyzies, en la Dordogne francesa (1868). A estos hallazgos siguieron otros fuera del continente europeo: los restos en las cuevas del Monte Carmelo, en Palestina y las herramientas de piedra asociadas a humanos, en Kenia y en Egipto, descubiertas por los esposos Leakey y Gertrude C. Thompson, respectivamente. Sin embargo, los hallazgos de vestigios más antiguos se produjeron en África, en Sudáfrica y Etiopía. El más famoso es el del río Klasies, en Sudáfrica, y en 1968, donde se encontraron fragmentos de cráneo de unos 120 mil años de antigüedad. Otros relevantes fueron los de la localidad etíope de Kibish en la región de Omo, cuya datación (por medio de la biocronología) fue también de 120 mil años (aunque dataciones recientes del cráneo Omo I usando isótopos ha arrojado una datación más arcaica, en torno a 200 mil años), y la colección de fósiles de Herto, también en Etiopía. 7 El estudio pionero que aplicó los métodos biológicos moleculares fue el conformado en 1987 por Rebecca Cann, Mark Stoneking y Allan Wilson, biólogos moleculares de la Universidad de Berkeley, quienes se focalizaron en el ADN mitocondrial porque en él las mutaciones ocurren de manera mucho más rápida que en el ADN nuclear. El estudio comparó el ADN mitocondrial de casi ciento cincuenta humanos modernos (46 de Europa, África del norte y Próximo Oriente; 20 del África subsahariana; 34 asiáticos, 26 de Nueva Guinea y 21 australianos), encontrando ciento treinta y tres diferentes versiones de ADN mitocondrial. Con esos datos conformaron un árbol que presentaba un muy profunda rama africana y una segunda que contenía las variantes del ADN mitocondrial encontradas en las personas fuera del África subsahariana. Las variaciones son muchas más dentro de la rama del árbol en África subsahariana que en el resto del mundo junto. Esto significa que la mayoría de las variantes de ADN mitocondrial pudieron haber sido de origen africano. Con estos resultados, llegaron a las siguientes conclusiones: 1º los humanos modernos han estado en África más tiempo que en cualquier otra parte del mundo, y el tamaño de las poblaciones de humanos modernos en ese continente fue mucho mayor que en el resto de los continentes combinados; 2º las diferencias entre las poblaciones modernas subsaharianas y no sub saharianas tomaron unos 200 mil años en acumularse. En consecuencia, predijeron que los humanos modernos se originaron en África en torno a 200 mil antes del presente; 3º la distribución de las variantes de ADN mitocondrial sugiere que cuando los humanos modernos abandonaron África, no se hibridaron con ninguna población arcaica, sino que debieron “encontrarse” al trasladarse a otras regiones del mundo. En consecuencia, los homininos arcaicos en otras partes del mundo no contribuyeron al moderno genoma humano. Otros estudiosos intentaron repetir los resultados de esta investigación usando métodos moleculares de vanguardia y técnicas estadísticas muy rigurosas. Lograron ciertos diferentes resultados que han modificado un tanto las conclusiones previas. Siguieron apuntando a África como el origen de un sustancial conjunto de las variaciones del ADN mitocondrial de los humanos modernos, si bien diversos de sus estudios sugirieron que hubo evidencia de que el hombre pre moderno fuera de África 8 contribuyó al genoma de ADN mitocondrial humano moderno. Los estudios se galvanizaron en torno al cromosoma Y masculino. Parte de este cromosoma del ADN es como el mitocondrial, excepto en que se transmite de una generación a la siguiente a través de los varones y no las hembras. Los resultados fueron como los del estudio del ADN mitocondrial: 21 de 27 variantes del cromosoma Y se originaron en África y hubo más variaciones en el cromosoma Y de los africanos que en los de las demás personas de las otras partes del mundo. Los estudios de los genes nucleares, no obstante, probaron evidencia de cierta mezcla entre genotipos humanos arcaicos y modernos. El mensaje primordial de estas investigaciones de ADN, sea mitocondrial, el cromosoma Y o el genoma nuclear regular, es que la mayoría, aunque no todos, los genes humanos modernos se originaron en el continente africano, y que en los dos últimos millones de años parece haber habido un conjunto de pulsiones de evolucionismo humano novedoso. El primero fue la emigración de Homo ergaster, luego la de Homo heidelbergensis, y en tercer lugar, quizás, diversas olas migratorias de humanos modernos con diferentes capacidades y habilidades culturales. Hoy en día se está generalmente de acuerdo en afirmar que los humanos modernos derivan de una relativamente reciente (50-45 mil años) migración desde el este de África. Ahora bien, los nuevos genes pudieron alcanzar el resto de los continentes de dos modos distintos: a través de la gente que emigra, o ser transmitidos por hibridación o mestizaje. Este último mecanismo incluiría a africanos biológicamente mestizados con personas en regiones adyacentes. Dicho tipo de transmisión de genes, que modifica un tanto las hipótesis establecidas por el equipo de Berkeley a fines de los años ochenta, se denomina hipótesis de la onda de difusión. Actualmente, hay casi un consenso en relación a tal planteamiento. 9 Ilustraciones Esqueleto y cráneo de Australopithecus sediba 10 Cráneo LB1 y esqueleto de Homo floresiensis Posible esquema del linaje de Homo floresiensis Cráneo KNM-WT 40000 de K. playtops. KNM ER 1470, de H. rudolfensis. Koobi Fora, Kenia 11 Bibliografía Aguirre, E. (edit.), Atapuerca y la evolución humana, Fundación Ramón Areces, Madrid, 1998 Alcázar Godoy, J., “Origen y evolución del Homo Hábilis”, Revista de Arqueología, nº 56, pp. 18-23, 1985 Arsuaga, J.L., & Martínez, I., La especie elegida. La larga marcha de la evolución humana, ed. Temas de Hoy, Madrid, 2001 Arsuaga, J.L, El collar del neandertal. En busca de los primeros pensadores, Plaza y Janés edit., Barcelona, 2002 Balter M., “Skeptics question whether Flores hominid is a new species”, Science, nº 306, p. 1116, 2004 Baur, M. & Ziegler, G., La aventura del hombre: todo empezó en África, edit. Maeva, Madrid, 2003 Berger, L.R., & Milton-Barber, B., Tras las huellas de Eva. El misterio de los orígenes de la humanidad, edic. B., Barcelona, 2001. __________,et al., «Australopithecus sediba: A New Species of Homo-Like Australopith from South Africa», Revista Science, 9 de abril de 2010, Vol. 328. no. 5975, pp. 195-204 Bernabeu, J., & Aura, J.E., & Badal, E., Al oeste del Edén, ed. Síntesis, Madrid, 1993 Brown P. & Sutikna T. & Morwood M. & Soejono R.P. & Jatmiko, Saptomo E.W., et al., “A new small-bodied hominin from the late Pleistocene of Flores, Indonesia”, Nature, nº 431, pp. 1055-61, 2004 Carbonell, E. & Sala, R., Planeta humano, edit. Península, Barcelona, 2000. 12 Cavalli-Sforza, L. y F., ¿Quiénes somos?. Historia de la diversidad humana, edit. Crítica, Barcelona, 1999. ___________________., Genes, pueblos y lenguas, ed. Crìtica, Barcelona, 2000. Chaline, J., Del simio al hombre. Una familia poco común, ed. Akal, Madrid, 1997. ________., Un millón de generaciones. Hacia los orígenes de la humanidad, edit. Península, Barcelona, 2002 de Facchini, F., Los orígenes del hombre y la evolución cultural, edic. San Pablo, Madrid, 2007 de Lumley, H., El primer hombre, ed. Cátedra, Madrid, 2000 de Turbón, D., La evolución humana, edit. Ariel Barcelona, 2006 Dirks et al., «Geological Setting and Age of Australopithecus sediba from Southern Africa», Revista Science, 9 de abril de 2010, Vol. 328. no. 5975, pp. 205-208 Falk D. & Hildebolt C. & Smith K. & Brown P. & Jatmiko, Saptomo E.W. et al., “The Brain of LB1, Homo floresiensis”. Sciencexpress, 03 March 2005, 1, 2005 Fernández Martínez, V.M., Prehistoria. El largo camino de la humanidad, Alianza editorial, Madrid, 2007 Fullola i Pericot, J.M. & Nadal Lorenzo, J., Introducción a la prehistoria. La evolución de la cultura humana, edit. UOC, Barcelona, 2005 Hublin, J.-J., & Tillier, A.-M., (coords.), Homo Sapiens. En busca de sus orígenes, edit. F.C.E., México, 1999. Leakey, R., & Lewin, R., Nuestros orígenes. En busca de lo que nos hace humanos, ed. Crítica, Barcelona, 1999 Leakey, M.G. & F. Spoor & F.H. Brown & P.N. Gathogo & C. Klarle & L.N. Leakey & I. McDougall, "New hominin genus from eastern Africa shows diverse middle Pliocene lineages", Nature, vol. 410, 2001, pp. 433-440 13 Lieberman, D.E., "Another face in our family tree." Nature, vol. 410, 2001, pp. 419-420 López Saco, J., Introducción al origen del hombre y sus primeras creaciones culturales, edit. Comala.com, Caracas, 2007 Michael B., «Candidate Human Ancestor From South Africa Sparks Praise and Debate», Revista Science, 9 de abril de 2010, Vol. 328. no. 5975, pp. 154-155 Morwood M. & Soejono R.P. & Roberts R.G. & Sutikna T. & Turney C.S.M. & Westaway K.E. et al., “Archaeology and age of a new hominin from Flores in eastern Indonesia”, Nature, nº 43, pp. 1087-91, 2004 Ramírez Goicoechea, E., Evolución, cultura y complejidad: la humanidad que se hace a sí misma, edit. Universitaria Ramón Areces, Madrid, 2009 Reichholf, J.H., La aparición del hombre, ed. Crítica, Barcelona, 2001 Schwartz, J.H. & T.D. White, "Another perspective on hominid diversity", Science, vol. 301, 2003, pp. 763-764 White, T., "Early hominids - Diversity or distortion?", Science, vol. 299, 2003, pp. 1994-1997 www.modernhumanorigins.net www.talkorigins.org 14