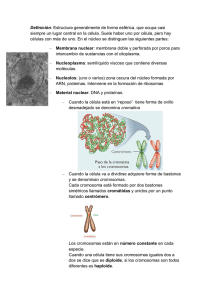
Material Biologia 101
Anuncio

CURSO DE AMBIENTACION UNIVERSITARIA (CAMU) BIOLOGIA UNIVERSIDAD NACIONAL DE JOSÉ C. PAZ CURSO DE AMBIENTACION UNIVERSITARIA (CAMU) Materia: BIOLOGIA Responsable: Lic. VIVIANA GRANADOS OBJETIVO DEL CURSO: Proporcionarle al ingresante elementos para la comprensión de la complejidad de la estructura y el funcionamiento de los seres vivos. Las múltiples interrelaciones, los mecanismos de la perpetuación de la vida, que permitirán en el desempeño de las carreras relacionadas con la salud, valorar en toda su amplitud el milagro de la vida. PROGRAMA: MODULO 1: “Introducción a la Biología” La Biología en el transcurso de la historia humana. Los seres vivos actuales (Dominios y Reinos). Características de los seres vivos (Niveles de Organización). Origen de la Vida. Método Científico. Unidades de Medidas. Importancia y Aplicaciones de la Biología. MODULO 2: “Biología Celular” Componentes químicos celulares. Componentes inorgánicos (Agua y otros). Compuestos orgánicos: Hidratos de Carbono, Lípidos, Proteínas, Ácidos Nucleicos. Difusión y Osmosis: transporte de sustancias a través de la membrana celular. Teoría Celular. Estructura celular: Procariotas y Eucariotas (Modelo E coli). Componentes celulares (Diferencias entre célula vegetal y animal). Tipos celulares (células nerviosas, sanguíneas, musculares, etc.). Respiración y Fotosíntesis. MODULO 3: “Genética” Reproducción celular: Mitosis, Meiosis. Conceptos de senescencia y apoptosis. Células haploides, diploides y poliploides. Experimentos de Mendel las Leyes en general. Herencia ligada al sexo. Ejemplos de Alteraciones cromosómicas (durante la mitosis y meiosis). MODULO 4: “Evolución y Diversidad Biológica” Conceptos generales sobre: Teoría de Darwin. Variación hereditaria. Selección Natural. Evolución de homínidos. TEMAS PÁGINAS MODULO 1 - INDICE • • • • • • • • • • • • • Biología en el transcurso de la historia 05 Viajes de Charles Darwin08 Los seres vivos actuales09 Características de los Reinos10 Características de los seres vivos13 Niveles de organización14 El Origen de la Vida16 Teoría de Oparín18 Teoría Endosimbiótica21 Método Científico 21 Unidades de Medidas 23 El microscopio24 Bibliografía28 MODULO 2 - INDICE • • • • • • • • • • • • • • • Componentes químicos celulares30 Componentes inorgánicos32 Compuestos orgánicos36 Lípidos39 Glúcidos40 Proteínas43 Ácidos Nucleicos48 Teoría Celular54 Estructura celular 59 Componentes celulares69 Difusión y ósmosis82 Respiración93 Fotosíntesis108 Tipos celulares114 Bibliografía119 TEMAS PÁGINAS MODULO 3 - INDICE • • • • • • • • • • • • • • Reproducción celular121 Mitosis123 Meiosis128 Gametogénesis132 Muerte celular135 Experimentos de Gregor Mendel135 1º Ley de Mendel138 2º Ley de Mendel145 Genética humana148 Herencia No Mendeliana150 Alelismo múltiple152 Herencia ligada al sexo153 Alteraciones cromosómicas154 Bibliografía163 MODULO 4 - INDICE • • • • • • • • • • • • Teoría de Lamarck164 Teoría de Charles Darwin165 Teoría Sintética de la Evolución169 Mecanismos de la Teoría Sintética169 El Origen de las Especies174 Especiación alopátrica174 Mecanismos de Aislamiento reproductivo176 Especiación simpátrica177 Evolución de los homínidos178 Modelo Multirregional179 Modelo del Arca de Noé180 Bibliografía183 MODULO Nº 1: INTRODUCCIÓN A LA BIOLOGÍA • LA BIOLOGÍA EN EL TRANSCURSO DE LA HISTORIA Es importante conocer los hechos, momentos y algunos nombres de personas que tuvieron un rol significativo en el curso de la historia de la biología, como así también comprender que antiguamente tantos las ciencias como el arte se desarrollaban principalmente por placer y curiosidad. Asimismo podemos decir que los hechos más comunes como los de nacimiento, crecimiento, muerte, salud y enfermedad, las plantas y animales que se utilizaban para alimento y abrigo, fueron considerados y estudiados por el Hombre puesto que eran necesarios para su supervivencia y su vida cotidiana. Estos mismos motivos son los que investiga la actual Biología Aplicada, como rama dentro de las ciencias biológicas. Probablemente la primera civilización que mostró interés por la Biología y de la que se guardan testimonios escritos sea la china, hace varios milenios antes de Cristo. Así, entre el cuarto y el tercer milenio a. C., ya se cultivaba el gusano productor de seda para la obtención de tejidos de dicha fibra. La antigua cultura china ya tenía los tratados de materia médica en los cuales se describen plantas y animales con propiedades terapéuticas, así como numerosos conceptos sobre fisiología humana en sus tratados de acupuntura. La antigua civilización hindú también hace referencia a los principios anteriores, aunque posiblemente sea debido a la influencia de la cultura china. Sin embargo la medicina hindú no presentaba un carácter mágico, sino que estaba basada en el pensamiento racional. Las culturas mesopotámicas también investigaron aspectos relacionados con la Biología, con la Medicina, y la Zootecnia. Por su parte, los egipcios tenían importantes conocimientos agrícolas, así como profundos conocimientos sobre la anatomía humana y animal, debido a las técnicas de embalsamamiento que realizaban. Ya en el Imperio Antiguo (2700-2200 a.C.) se desarrolla ampliamente la medicina y la cirugía, algunos de cuyos instrumentos y técnicas, convenientemente modificados, se siguen utilizando en la actualidad. Los egipcios recogían muestras vivas de plantas y animales de sus expediciones y desarrollaban jardines zoológicos y botánicos, lo que demuestra un gran interés por las Ciencias Naturales. Si se analiza la cultura occidental, el origen de la Biología se debe buscar en la antigua Grecia. El pueblo heleno estaba constituido por una serie de tribus, algunas de las cuales, como las de los jonios y los dorios, alcanzaron un gran desarrollo cultural. En la frontera entre ambas tribus estaba la isla de Cos, donde unos 600 años antes de Cristo se constituyó la primera institución científica reconocida: una escuela de medicina. Su figura más relevante fue Hipócrates (460-370 A.C.). Es considerado como el Padre de la biología científica y de la medicina; y que escribió toda una serie de tratados que conforman el Cuerpo Hipocrático, una enciclopedia médica que incluye en la medicina, temas de embriología, anatomía y fisiología de esa época. Su influencia llegó hasta el siglo XVII. Platón (428-347 A.C.) aportó su versión idealista del metabolismo: los alimentos tienen las sustancias necesarias para nutrir todas las partes del cuerpo; así la sangre se forma en el vientre, sede de la parte apetitiva del alma, por la acción del fuego y de allí se distribuye a todo el cuerpo. Fue Aristóteles (384-322 a. C.) uno de las personas más destacadas dentro de la biología. Nació en Estagira (Macedonia) y posteriormente se trasladó a Atenas, donde fue discípulo de Platón y maestro de Alejandro Magno. Escribió varios tratados sistemáticos sobre embriogénesis, anatomía y botánica, abordó el problema de generación de las plantas y de los animales. Observó la morfología y estudió el comportamiento de más de 500 especies de animales, además de crear una escuela de clasificación biológica. Se lo considera el padre de la Zoología. Aristóteles consideraba que las especies biológicas eran fijas y no podían cambiar, y además sugería que su origen no era casual, sino que seguía un plan predeterminado. Pensaba que el corazón era la sede de la inteligencia y que el cerebro era un simple órgano de refrigeración. Uno de sus más infortunados errores fue la racionalización del concepto de generación espontánea, que se admitió como verda5 dera durante tantos siglos e impidió el correcto desarrollo de la Biología. Su discípulo Teofrastos (372-287 a.C.) estudiaba botánica y en su Historia de las plantas se recogen algunas aportaciones originales como la observación de la germinación de la semilla. En el siglo III a. C se produce un importante desarrollo en el área de la medicina y la biología, en la ciudad de Alejandría. Posteriormente, en el siglo II d. C, el Imperio Romano desarrolla especialmente los campos de la zoología y botánica para aplicarlas en actividades de ganadería y agricultura. En Roma adoptan los conocimientos griegos para sus estudios en medicina pero no su método de disección en la enseñanza, por lo tanto las obras romanas eran principalmente filosóficas. En esta época la idea que predominaba era que la naturaleza existía para satisfacer las necesidades del Hombre. Este concepto perduró durante quince siglos y dominó la Historia Natural. Cabe mencionar a Galeno (129 – 199 d. C) médico griego, como una persona célebre dentro de la biología antigua. Establecido en Roma, realiza disecciones de animales vivos y muertos para sus estudios e investigaciones, pero no sobre cuerpos humanos porque en esa época se condenaba la disección de cadáveres con fines científicos. No era cristiano pero creía en la existencia de Dios. Creía que todo se había hecho con una finalidad, y así encontraba signos de la creación divina en todas partes del cuerpo. Esto concordaba con las creencias del cristianismo en expansión y explica la popularidad de Galeno en los siglos posteriores. El desarrollo de sus teorías sobre el cuerpo humano fue influyente hasta la época moderna. A partir del siglo XV, y dentro de la revolución científica que tuvo lugar en el Renacimiento, resurge el interés por los estudios anatómicos y fisiológicos. Como figuras importantes hay que destacar a Leonardo da Vinci (1452-1519), quien representa uno de los anatomistas más del Renacimiento. Éste realizó estudios sobre el cuerpo humano y su comparación con el de otros animales estableciendo “homologías” que posteriormente echarían las bases para las teorías de evolución. También realizó estudios sobre el vuelo de las aves y los dibujó, porque estaba muy interesado en diseñar una máquina que hiciera posible el vuelo del Hombre. El desarrollo de la biología no ha sido tan rápido como el de las ciencias físicas. A lo largo de los siglos XVIII y XIX, la aparición de restos fósiles reinició una polémica sobre el origen de los seres vivos que databa ya desde los filósofos griegos. Ante la presentación de pruebas de la existencia de organismos antiguos y diferentes, la ciencia del siglo XIX se dividió en dos posiciones extremas agrupadas bajo el nombre de Escuela Catastrofista y Escuela Evolucionista, que se desarrollaran más adelante. Se puede decir que es en esta época cuando surge el nacimiento de la biología como actividad científica, proporcionando un marco conceptual a los meros conocimientos empíricos. No obstante, pueden reconocerse desde el comienzo de la Modernidad un intento experimentador y sistematizador notable. Por ejemplo, en 1543, el mismo año en que Copérnico publica su obra maestra que sería el origen de la nueva física, Andreas Vesalio, anatomista belga, publica su obra De humani corporis fabrica (De la estructura del cuerpo humano), basado en un sistemático proceso de disecciones. Los naturalistas limitaban sus estudios a la flora y la fauna del lugar donde habitaban. Sin embargo sabían, por relatos de viajeros, que en otras regiones vivían seres vivos muy diversos pero no contaban con material para estudiarlos. Por tal motivo los grandes viajes, desde los de Marco Polo (1254 – 1323) a Asia hasta el descubrimiento de América por Cristóbal Colón (1451 – 1506), modificó la percepción de los naturalistas sobre la diversidad de los seres vivos y su distribución geográfica. Comenzaron a traer las primeras colecciones de organismos exóticos o raros. Posteriormente se fue reglamentando la forma de recolectar, observar y registrar las colecciones de seres vivos. Estas colecciones pasaban a formar parte de museos y herbarios en toda Europa. En estos viajes exploratorios, los naturalistas formaban también parte de la tripulación y algunos de ellos realizaron aportes muy valiosos para la biología. En 1753, Carl von Linneo (1707 – 1778), publicó Species Plantarum donde describió cada especie de planta conocida hasta ese momento y creó el sistema de clasificación jerárquica y el sistema de nomenclatura universal que actualmente se conoce como Taxonomía y se utiliza para nombrar a las especies. 6 El biólogo francés George Cuvier (1769 – 1832) dedicó su vida a clasificar y comparar las estructuras de diferentes animales y fósiles, convirtiéndose así en el padre de la anatomía comparada y paleontología. Entre los siglos XVIII y XIX, dos grandes viajeros marcaron los cambios que sentaron las bases de la biología moderna. El geógrafo y físico alemán Alexander von Humboldt (1769 – 1859) a partir de sus viajes desarrolla un nuevo modelo de ciencia natural al describir no solo la especie sino también el lugar donde eran halladas. Los viajes a Tenerife de Humboldt influyeron en el inglés Charles Darwin (1809 – 1882) quien se embarca en el Beagle como naturalista recorriendo la costa atlántica de Sudamérica. Exploró el interior del continente, observando los estratos geológicos, los variados organismos y tomando ejemplares para su colección de plantas y animales desconocidos en esa época. Así fue el primero en reunir una gran cantidad de evidencia que contribuía al pensamiento que los organismos evolucionan o cambian a lo largo del tiempo. El viaje de Darwin a bordo del Beagle representó un hecho fundamental en la historia de la ciencia. Dos décadas más tarde, la teoría de Darwin acerca de la evolución se convirtió en el marco teórico unificador más importante de la biología. (Se verá en detalle en el Módulo Nº 4) Viajes de Alexander von Humboldt: Viajó desde Europa a América del Sur, parte del actual territorio de México, Estados Unidos, Is. Canarias y Asia Central. El viaje fue de 5 años. 7 Charles Darwin y el Beagle: En principio el viaje iba a ser de un año y finalmente duró 5 años. El Beagle tenía una misión científica, la recolección de datos sobre corrientes marinas, geología, flora y fauna de los lugares que iban visitando. Mapa que muestra el recorrido de Darwin embarcado en el Beagle Como se vio en esta breve reseña sobre el nacimiento de la biología, ésta se fue modificando y especializando cada vez más, creando diversas áreas de investigación. Una de las más recientes en el campo de la biología es la rama de Ecología. La consideración del Hombre como parte de los ecosistemas estableciendo relaciones de interdependencia y ejerciendo una acción capaz de modificar los equilibrios naturales, la consideración que las agresiones a la naturaleza pueden desbordar la 8 capacidad de autorregulación de los ecosistemas amenazando con consecuencias imprevisibles a todos sus integrantes, y la conciencia de la realidad de destrucción y saqueo de la naturaleza y de Mapa del viaje, dibujado por Darwin a bordo del Beagle gradación de la biosfera por parte de la civilización industrial ha dado lugar a la denominada crisis ecológica, una nueva visión del Hombre respecto a sus propias acciones sobre la naturaleza. La ciencia es un proceso dinámico y aunque en biología se han realizado avances importantes, actualmente muchas preguntas no tienen respuestas. • LOS SERES VIVOS ACTUALES A través del tiempo, los naturalistas han clasificado a las diferentes formas de vida de la Tierra siguiendo distintos modelos, a fin de poner orden en el caos de animales y plantas que aumentaban su número día a día, a medida que nuevos viajeros traían de vuelta más ejemplares de seres desconocidos. La rama de la biología que actualmente clasifica la diversidad de organismos vivos es la Sistemática. La unidad básica de la clasificación biológica es la Especie, en latín significa “tipo”. Es importante definir qué es una especie para entender la clasificación de los seres vivos. En 1940 el biólogo evolutivo Ernst Mayr, propone una definición de especie biológica: “un grupo de poblaciones naturales cuyos individuos se cruzan entre sí exitosamente de manera real o potencial y 9 que están reproductivamente aislados de otros grupos”. Muchas especies están formadas por un conjunto de poblaciones con características genéticas, morfológicas o comportamentales diferentes, y se habla de Subespecies. Estas pueden llegar a formar una nueva especie biológica en el futuro si se presentan mecanismos de especiación como aislamiento reproductivo. La primera organización en Reinos se debe a Aristóteles que diferenciaba, a través de un método lógico, a todas las entidades de la naturaleza en dicotomías. Por ejemplo se preguntaba si eran seres vivos o no; si eran animales o no. Como se vio en el apartado anterior fue Carl von Linneo quien en el siglo XVIII diseñó un sistema de nomenclatura binomial. Es decir el nombre científico de un organismo tiene dos partes: el primero indica el género al que pertenece y el segundo a la especie. Por convención se escriben en letra cursiva y con mayúscula sólo el género. Ejemplo: Escherichia coli, Homo sapiens. Linneo adoptó una jerarquía de siete niveles: imperio, reino, clase, orden, género, especie y variedad. En esa época los naturalistas reconocían dos reinos: Vegetal y Animal. Posteriormente se fueron eliminando algunas categorías y agregando otras. Con el desarrollo del microscopio se descubrió una gran cantidad de microorganismos. Ernst Haeckel (1834 – 1919) evolucionista alemán, fue el primero en diferenciar los organismos unicelulares de los pluricelulares y propone a fines del siglo XIX, un tercer Reino formado por microorganismos que llama Protistas. Algunos de ellos carecían de núcleo celular y los llama Monera (bacterias). El reino de los hongos, Fungi, fue el último en crearse. En 1959, el fundador del reino Fungi, Robert Whittaker (1924 – 1980) ecólogo estadounidense, propone una clasificación general de los seres vivos en cinco reinos: Monera (bacterias), Protistas (protozoos), Fungi (hongos), Animalia y Plantae. En 1978, Whittaker y Margulis modifican esta clasificación, incluyendo dentro del reino Protista a las algas. Así hasta 1977, el Reino se consideraba la categoría sistemática más abarcativa, sin embargo los continuos avances de la ciencia y el énfasis en la filogenia, llevó a constituir un nivel más arriba en esta jerarquía: el Dominio. Fue propuesto por Carl Woese y colaboradores quienes construyen un árbol filogenético único en el cual diferencian tres linajes evolutivos principales. Surgen tres dominios: Bacteria, Archaea y Eukarya. Esta nueva clasificación de los seres vivos se basa en el orden de ramificación de los linajes durante la historia evolutiva. No todos los científicos están de acuerdo con esta nueva clasificación de Woese, existen varias hipótesis alternativas, como la que propone dos Imperios: el imperio Bacteria y el imperio Eukaryota con un total de ocho reinos. Como ya se mencionó, la ciencia es un proceso dinámico que se va modificando y descubriendo en forma contínua. 10 Diagrama: Modificación del Sistema de Clasificación de seres vivos Definiciones de algunas de las palabras más usadas en este cuadernillo Características generales de los tres dominios La clasificación taxonómica de los procariontes1 , denominados comúnmente bacterias, actualmente no se conciben como un grupo monofilético2 . Por tal motivo Woese divide al reino Mónera en dos Dominios y propone así tres linajes evolutivos o dominios diferentes, como ya fue expresado. De estos tres dominios solo Eukarya es eucarionte3 . Si bien las bacterias y archaea comparten características de tamaño y organización celular, poseen diferencias a nivel molecular de sus membranas y ARN ribosómico. 11 BACTERIA: Células procariontes. Organismos muy pequeños, libres o agregados, sin núcleo definido ni organelas. Gran variedad de estructuras y metabolismos. Incluyen organismos que pueden vivir a altas temperaturas y anaerobios. Pertenecen a este dominio las bacterias grampositivas, cianobacterias y bacterias rojas o proteobacterias. ARCHAEA: Células procariontes. Organismos muy pequeños, sin núcleo definido ni organelas. Poseen adaptaciones que les permiten habitar ambientes con condiciones extremas como en grietas volcánicas, pantanos, lagos muy salados (Mar Muerto). Pueden tolerar temperaturas superiores a 100 ºC o inferiores a 0 ºC, en ambientes muy salinos o con pH4 extremos. Pueden ser anaerobias o aerobias. EUKARYA: Células eucariontes. Pueden ser autótrofos, heterótrofos o ambas a la vez. La reproducción puede ser tanto sexual como asexual. Poseen citoesqueleto, membranas internas y ADN lineal. En esta categoría se encuentran todos los organismos de la naturaleza excepto los procarionte (Monera). Características generales de los Reinos Siguiendo la clasificación de Woese, incluidos en el dominio Eukarya, se encuentran los reinos: Protista, Fungi, Plantae y Animalia: PROTISTA: Células eucariontes. Dentro de este grupo se encuentran los protozoos, algas, mohos del limo, y otros organismos acuáticos y parásitos menos conocidos. Pueden unicelulares o coloniales. Poseen diversas formas de nutrición: fotosíntesis, absorción, ingestión y combinación de éstos. Ciclos reproductivos variados, sexuales o asexuales. FUNGI: Células eucariontes y primariamente multinucleados, los núcleos dispersos en un micelio5 ; raramente unicelulares (p. ej. levaduras). Sin pigmentos ni organelas fotosintéticas. Nutrición absortiva, son heterótrofos. Almacenan glucógeno y lípidos como sustancias de reserva, a diferencia de las plantas que almacenan almidón. Reproducción asexual o sexual. Pueden ser parásitos y simbiontes. Junto con las bacterias son los descomponedores principales de la materia orgánica. Algunas especies de hongos son perjudiciales para el Hombre, otras son esenciales para la producción de queso, pan, vino. También hay especies comestibles como los champiñones y otras que son fuente de antibióticos y medicamentos. PLANTAE: Organismos multicelulares (o pluricelulares), más raramente unicelulares. Células eucariontes con pared celular rígida y clorofila a y b en cloroplastos. Almidón como sustancia de reserva. Modo principal de alimentación: fotosíntesis (puede haber absorción). Diferenciación en órganos. Reproducción sexual y asexual. Pueden poseer semillas o no. Pueden ser arbustivas o no y tener o no flores. Ejemplos: helechos, briofitas, coníferas, palmeras, plantas con flores. ANIMALIA: Organismos eucariontes, heterótrofos y multicelulares. Células sin pared rígida ni plásticos con pigmentos fotosintéticos. Nutrición originalmente por ingestión con una cavidad 1 2 3 Procarionte: célula que carece de núcleo y organelas limitadas por membrana. Monofilético: conjunto de especies que incluye el ancestro común y todos sus descendientes. Eucarionte: célula que tiene un núcleo y organelas rodeadas por membrana y cromosomas en los que el ADN está combi- nado con proteínas histónicas. 4 pH: potencial de hidrógeno. Símbolo que indica la concentración de iones hidrógeno en una solución. Sus valores van desde 0 a 14. Cuanto más bajo es el valor, más ácida es la solución. 5 Micelio: es el conjunto de filamentos fúngicos o hifas. 12 digestiva (existen variantes). Desarrollo de sistemas neuromotores y sensoriales. Reproducción sexual principalmente y también asexual. Gran diversidad de formas de vida y tipos morfológicos. Se han propuestos numerosas clasificaciones en los últimos 10 años, antiguamente se los agrupaba en “invertebrados inferiores” como las esponjas, medusas, anémonas de mar; “invertebrados superiores” como los insectos, moluscos, platelmintos y en “vertebrados” como anfibios, peces, reptiles, aves y mamíferos. • CARACTERÍSTICAS DE LOS SERES VIVOS Antes de exponer la Teoría sobre el Origen de la Vida, será oportuno analizar las condiciones que deben cumplirse para que un sistema sea considerado viviente. Forzosamente la definición será subjetiva y adolecerá de defectos y superposiciones: es una interpretación que se pretende imponer a un complejo fenómeno natural, pero la naturaleza no necesariamente responde a esa interpretación. Para que un sistema se considere viviente debe presentar las siguientes propiedades: a) Organización específica: Los sistemas vivos presentan una organización jerárquica de niveles crecientes de complejidad, como se verá más adelante. b) Homeostasis: Es la capacidad de un sistema de mantener la constancia de su medio interno (constancia estructural, de concentración de sustancias químicas, de variables físicas, etc.) frente a alteraciones de su ambiente (estímulos). Esta propiedad engloba los fenómenos necesarios para mantener esa constancia (nutrición, metabolismo6 , excreción, etc.). La homeostasis utiliza mecanismos de regulación. c) Reproducción: Los sistemas vivos son capaces de producir otros sistemas similares a ellos, dotados de esa misma capacidad. La reproducción conlleva un sistema de transmisión de la información: la herencia. Sin esta capacidad, no podrían persistir en el tiempo, generación tras generación. d) Sistemas termodinámicos obligatoriamente abiertos: Un sistema que intercambia materia y energía con su ambiente se denomina Abierto. Si sólo intercambia energía, se denomina Cerrado y si no realiza intercambio alguno con el ambiente, se llama Aislado. La mayor parte de los sistemas que observamos son abiertos, pero los sistemas vivos presentan la característica diferencial de que deben ser necesariamente abiertos. Por ejemplo, una piedra es un sistema abierto porque puede calentarse y enfriarse durante el día o según las estaciones del año y cede materia por erosión o acepta depósitos de materia; sin embargo si se la coloca dentro de un recipiente que la aísle del ambiente esto no hace que la pierda su característica específica de una piedra. En cambio, si un sistema vivo es aislado de su ambiente, no puede funcionar y se desorganiza, es decir pierde su característica de viviente. Estas condiciones deben cumplirse simultáneamente para que un grupo de sistemas se considere 6 Metabolismo: es el conjunto de reacciones químicas y de transformaciones de energía, incluidas la síntesis (o catabolismo) y la degradación (o anabolismo) de moléculas. 13 vivo. Un organismo particular puede no cumplir con alguna condición: por ejemplo, los machos de una especie con reproducción sexual no tienen capacidad por sí solos de producir individuos semejantes a ellos, pero no se discute que sean seres vivos. Por otra parte un sistema no vivo puede presentar, aisladamente, alguna de las características mencionadas: por ejemplo un animal recién muerto mantiene por cierto tiempo la misma organización compleja, aunque haya perdido las otras características. Además de estas condiciones indispensables, los seres vivos presentan otras características que no se dan necesariamente en todos: por ejemplo el Crecimiento, en muchos casos resultaría una fase final de la reproducción porque se detiene cuando el organismo llega a adulto. Se puede decir que en general, los organismos atraviesan un ciclo vital en el cual crecen, se desarrollan y reproducen. Si bien los seres vivos comparten muchas características, existen en una gran diversidad de formas y funciones y esto es una consecuencia del proceso evolutivo, y son detectables a largo plazo, es decir sobrepasa la vida del observador, como ocurre con la Adaptación en el sentido evolutivo darwiniano. Como se menciona en el ítem a), los seres vivos son sistemas muy organizados y complejos. A continuación se establece un nivel de organización desde lo más simple a lo más complejo y sus características generales. Subatómico: Es el nivel más simple de la materia en el que se encuentran los protones principalmente, los neutrones y electrones que constituyen los Átomos. Molecular: Este contiene los niveles atómicos y subatómicos y moléculas más complejas o macromoléculas formadas a partir de moléculas simples. Celular: En este nivel surge la propiedad más notable, la vida, en la forma de organismos unicelulares o multicelulares. Por lo tanto es el primer nivel con vida. Cabe aclarar que en ocasiones se encuentran organismos unicelulares agrupados formando colonias, pero no se pueden considerar organismos multicelulares porque a pesar de estar formados por más de una célula, cada una vive como un organismo independiente. Tejidos: Las células vivas especializadas se organizan en tejidos como por ejemplo: tejido nervioso, epitelial, muscular. En este nivel, las células especializadas realizan la misma función y tienen un mismo origen. Órganos: Los tejidos forman órganos que realizan una acción concreta y son altamente complejos. Ejemplo: hígado, cerebro, riñones, pulmones, etc. Sistemas: Son conjuntos de órganos parecidos pero que realizan acciones independientes. Ejemplo: sistema nervioso, sistema circulatorio, sistema respiratorio, etc. Individuo: Un individuo es un sistema biológico funcional. En este nivel de organización, las células, tejidos, órganos y sistemas, actúan coordinadamente para constituir un individuo. Este nivel de individuo lo encontramos también en una única célula con individualidad de ser vivo, pero en este caso el individuo multicelular posee un grado de complejidad mucho más elevado. Ejemplo: un lobo, un roble o mismo el Hombre. Población: Nivel en el cual los individuos de una misma especie ocupan un área determinada al mismo tiempo y pueden intercambiar material genético. Comunidad: Un conjunto de poblaciones que habitan en un ambiente común en un momento 14 determinado e interactúan unas con otras. Ejemplo: comunidad biótica del bosque, la comunidad de peces, caracoles, algas y plantas acuáticas de la laguna de Chascomús, etc. Ecosistema: Este nivel incluye las relaciones que existen entre el conjunto de organismos que se encuentran en un área determinada y su ambiente. Es la unidad funcional de la biosfera. Biosfera: Es el conjunto de todos los ecosistemas naturales. Es el ecosistema total de la Tierra. Los ecólogos estudian los niveles de organización: • Poblaciones • Comunidades • Ecosistemas Los anatomistas y fisiólogos estudian los niveles de organización: • Sistemas • Órganos • Tejidos • Celular Los químicos estudian los niveles: • Celular • Molecular Los físicos- químicos estudian los niveles: • Atómico • Subatómico Niveles de organización: desde átomos hasta biosfera 15 • EL ORIGEN DE LA VIDA Las teorías de la antigüedad clásica con respecto a la aparición espontánea (en medios apropiados) de organismos complejos, fueron refutados por una serie de experimentos llevados a cabo por un médico italiano Francesco Redi (1626 – 1697), el biólogo italiano Lázaro Spallanzani (1729 – 1799) y definitivamente el químico francés Louis Pasteur (1822-1895). Muchas de esas teorías, como se verán más adelante, se generaron sobre la base de observaciones insuficientes, defectuosas o prejuiciados por los conocimientos de la época. Pero el primer conjunto de hipótesis contrastables fue propuesto por el bioquímico ruso A. I. Oparín (1894 - 1980) y por el inglés J. B. Haldane (1892 – 1964), quienes trabajaban en forma independiente, cada uno en su país de origen y sin conocer uno el trabajo del otro. Sin embargo ambos llegaron a postular que el surgimiento de la vida fue precedida por un largo período que denominaron evolución química. Este conjunto de hipótesis no explica cómo se originó la vida en la Tierra, sino cómo pudo haber surgido. Pese a las numerosas lagunas en su exposición, esta teoría sigue siendo la más aceptada en biología aunque todavía generan muchas controversias que no han sido dilucidadas. Día a día se incorporan experimentos que aumentan su verosimilitud. No obstante, cabe destacar la posibilidad que dentro de un tiempo estas hipótesis entren en la categoría de olvido, como consecuencia de las nuevas tecnologías y el avance continuo de la ciencia. Mecanicistas vs. Vitalistas Durante los siglos XVII y XVIII coexistían dos líneas de pensamientos antagónicas acerca de la vida. Los mecanicistas sostenían que el cuerpo de los organismos trabaja esencialmente como máquinas. Por lo tanto todos los fenómenos biológicos podían interpretarlo en términos físico-químicos. Así el corazón funciona como una bomba, el estómago como un mortero, los pulmones como un fuelle, etc. Estos modelos simplistas servían para comprender el funcionamiento del cuerpo animal. Un destacado defensor de este pensamiento fue el filósofo francés René Descartes (1596 – 1650). Los vitalistas creían que todos los sistemas vivos poseían de un “espíritu vital” y esto los diferenciaba de los sistemas no vivos y que no alcanzaba con las leyes físicas y químicas para explicar un sistema vivo. Algunos sostenían incluso que había una fuerza natural que gobernaba a los seres vivos. Uno de los seguidores de esta corriente vitalista fue Louis Pasteur, quien decía que los cambios que se daban en el jugo de fruta cuando se formaba vino eran “vitales” y sólo podía ocurrir en células vivas, en este caso en las células de levadura. En 1898, los químicos alemanes Edgard y Hans Buchner mostraron que una sustancia extraída de las levaduras podía fermentar fuera de las células vivas. A esta sustancia se le dio el nombre de enzimas. Así se demostró que una reacción considerada hasta ese momento como “vital” era sólo una reacción química. Finalmente la discusión sobre el vitalismo se dejó de lado. La generación espontánea Durante la Edad Media y hasta el siglo XVI predominó la idea que la generación de los organismos siempre involucraba la intervención de un acto de creación divina. En el siglo XVII comenzó 16 a retomar fuerza una idea vinculada con mitos y observaciones de distintos pueblos de la antigüedad: la de que algunos seres vivos podían surgir por generación espontánea, a partir de sustancias inertes (sin vida) del medio ambiente. Esta interpretación fue objeto de un largo debate que duró unos 200 años. Desde la época de Aristóteles, la mayoría de los naturalistas aceptaba la idea que los seres humanos y los animales de gran tamaño, provenían del cuerpo de sus madres o de huevos puestos por ellas. Pero esto no resultaba tan claro con los seres vivos de menor tamaño como los gusanos, los insectos, las ranas y las salamandras; así pensaban que éstos se originaban repentinamente a partir de materia inorgánica o carne en descomposición. Este origen de la vida a partir de la materia inanimada se denominaba “generación espontánea”. El ejemplo clásico presentado como evidencia de generación espontánea era la aparición de larvas en la carne en descomposición. Parecía obvio que esos pequeños organismos, semejantes a gusanos se habían originado en dicha materia en descomposición. Casi todos los biólogos de esa época, aceptaban ese hecho. Hasta que en el siglo XVII, Redi decide poner a prueba esa hipótesis y en 1668 lleva a cabo un famoso experimento en el que pone carne en descomposición en un grupo de 8 recipientes con boca ancha, algunos con tapas, algunos cubiertos con una delgada tela y otros abiertos. Así demostró que aparecían larvas solamente en los frascos abiertos, en los que las moscas podían acceder a la carne para depositar sus huevos. A partir de este momento la biología podía haber abandonado el concepto de la generación espontánea pero con la llegada de la microscopía y las observaciones hechas por van Leeuwenhoek en el siglo XVIII, se reaviva con fuerza la creencia de la generación espontánea pero ahora con la aparición de microorganismos en lugar de gusanos o larvas. Sólo era necesario poner sustancias en descomposición en un lugar cálido y en poco tiempo surgían “bestias vivas” al observar con una lupa. En esa época se hablaba de una fuerza vital que provocaba la aparición de los microorganismos. Entonces Spallanzani hierve el contenido de los frascos, algunos de los cuales estaban sellados y otros no y demuestra que en los frascos sellados no hay presencia de microorganismos. Sus oponentes justifican el hecho diciendo que al hervir el contenido había destruido la fuerza vital. Fue en el año 1864 con Louis Pasteur quien ideó nuevos experimentos para mostrar que los microorganismos aparecían sólo como resultado del ingreso se aire contaminado con gérmenes y no “espontáneamente” como pensaban en esa época. Para ello usó matraces con cuello de cisne que permitían la entrada de oxígeno (considerado necesario para la vida) pero impedía la entrada de bacteria, esporas de hongos y otros tipos de microorganismos. Pasteur demuestra que si se hervía el contenido del matraz, no aparecía ningún microorganismo. Con estos experimentos de Pasteur, la idea de la generación espontánea queda definitivamente refutada. Recién 50 años más tarde, Oparín y Haldane proponen un marco teórico para interpretar el origen de la vida. Origen extraterrestre Unos años antes de la Teoría de Oparín sobre el origen de la vida, un físico y químico sueco Svante Arrhenius (1859 – 1927), en el año 1906 postula una hipótesis sobre el origen extraterrestre de la vida. Plantea que los primeros gérmenes de la vida (esporas o bacterias) habrían llegado del espacio en meteoritos desprendidos de un planeta en el que ya habría vida. Estas ideas o hipótesis, aún hoy siguen surgiendo, sin embargo no contribuyen a comprender o explicar realmente el origen de la vida. Cabe destacar que actualmente existen evidencias de la presencia de aminoácidos en un meteori17 to encontrado en 1969 en Australia. Más recientemente, en 1996, se encontró otra evidencia que la vida podría haberse originado en otros planetas: en un meteorito proveniente de Marte hallado en la Antártica aparecieron posibles señas de organismos. Pero hasta el momento, los estudios astronómicos y las exploraciones espaciales, indican que entre los planetas de nuestro sistema solar, la Tierra es sólo la que tiene vida. Teoría de Oparín Publica su Teoría en el año 1924. Como fue mencionado más arriba, Oparín propone una primera etapa en su teoría a la cual denomina: evolución química. En este momento la Tierra primitiva tendría las siguientes condiciones: a) Por un lado había muy poca cantidad de oxígeno libre (o molecular) en la atmósfera b) Estaban presentes los cuatro elementos básicos que constituyen el 95% de todos los tejidos vivos, hidrógeno, carbono, nitrógeno y oxígeno; disponibles en alguna de sus formas en la atmósfera y aguas de la Tierra primitiva. Además de estas dos condiciones, Oparín propone una gran cantidad de energía en forma de rayos, calor, radioactividad y radiación proveniente del Sol. Bajo estas condiciones se habrían formado moléculas orgánicas. Estas moléculas en ciertos microambientes como por ejemplo en un lago posteriormente a su desecación, se habrían concentrado y reaccionado formando a su vez moléculas más grandes. A su vez éstas se pudieron combinar y formar pequeños sistemas plurimoleculares. Así propone la finalización de la etapa química y la aparición de una segunda etapa a la cual llamó evolución prebiológica. En esta etapa, Oparín postula que los sistemas plurimoleculares más estables químicamente en la Tierra primitiva, fueron los que perduraron en el tiempo y favorecieron a un aumento en la complejidad que llevaría a la adquisición de un metabolismo simple, punto de partida de todo el mundo viviente. Oparín experimentó su hipótesis utilizando un modelo al que llamó coacervados. Los coacervados son sistemas constituidos por macromoléculas en suspensión que se habrían formado en la Tierra primitiva en un medio acuoso. Estas condiciones externas especiales propuestas por Oparín, no ocurren en la actualidad, por lo tanto bajo las condiciones que tiene nuestra Tierra no es posible que se repita este proceso porque las moléculas orgánicas se degradarían en presencia de oxígeno y por otro lado la capa de ozono (O3) filtra y disminuye las radiaciones ultravioletas necesarias, de acuerdo a la teoría de Oparín, para la formación de sistemas plurimoleculares. Podemos decir que los seres vivos modificaron la atmósfera primitiva, los mares y cada rincón de la Tierra, lo que impidió, a su vez, la formación posterior de nueva vida a partir de sustancias inorgánicas. Oparín decía: “…por paradójico que ello pueda parecer, debemos admitir que la causa principal de la imposibilidad de la aparición de la vida en las condiciones naturales actuales reside en el hecho de que ya existe”. En 1952, el bioquímico Stanley Miller realizó los primeros experimentos que demostraban que la etapa química propuesta por Oparín era posible. Se han postulado desde entonces modelos alternativos como el propuesto por Sydney Fox hacia el año 1960, quien encontró que se podían encadenar experimentalmente, aminoácidos en cadenas de proteínas bajo las mismas condiciones propuestas por Oparín para la Tierra primitiva. 18 Atmósfera terrestre primitiva Fotografía del experimento de Miller Esquema del experimento de Miller: Stanley Miller simuló en su laboratorio las condiciones que habría tenido la Tierra primitiva. Esta fue la primera evidencia experimental de la teoría de Oparín. Organismos Heterótrofos y Autótrofos. Procariotas y Eucariotas Para poder entender la Teoría Endosimbiótica, la cual explica el origen de algunas organelas y que se verá a continuación, es necesario comprender algunos conceptos. 19 Se llaman organismos heterótrofos, aquellos que incorporan moléculas orgánicas del ambiente exterior y degradan para obtener su energía. Dentro de este grupo se encuentran los animales, hongos y muchos unicelulares. Otros organismos son capaces de sintetizar moléculas orgánicas ricas en energía a partir de sustancias inorgánicas simples, a este grupo se los llama autótrofos. Son autótrofos todas las plantas y varios tipos de protistas. Existen dos tipos celulares distintos a) Células procariotas: carecen de núcleo y organelas limitadas por membrana. El material genético es una molécula grande y circular de ADN a la que están asociadas débilmente diversas proteínas. Como ya se vio el ítem de Dominios; Archaea y Bacteria incluyen a organismos procariontes unicelulares o coloniales. b) Células eucariotas: poseen núcleo cuya envoltura es una doble membrana. Organelas rodeadas por membranas y cromosomas en los que el ADN lineal está combinado fuertemente a proteínas. Los organismos incluidos en el dominio Eukarya, son eucariontes. Organismo Procarionte: esquema de Esche- Organismo Eucarionte: esquema de Chlamydo- richia coli, un procarionte heterótrofo muy monas, un organismo unicelular eucarionte estudiado. y fotosintético. 20 Teoría Endosimbiótica Hace unos 30 años, la investigadora estadounidense Lynn Margulis propuso la Teoría Endosimbiótica7 para explicar el origen de principalmente dos organelas eucariotas: mitocondrias8 y cloroplastos9 . (Se verán más en detalle en el Módulo II) Varias evidencias apoyan a esta teoría. Por un lado las mitocondrias poseen ADN propio y circular (como las bacterias) y diferente al ADN nuclear. También muchas de las proteínas de las membranas de las bacterias se encuentran en las membranas de las mitocondrias. Según la teoría, hace aproximadamente de 2.500 millones de años, cuando la atmósfera era rica en oxígeno, ciertas organismos procariontes (bacterias) habrían sido fagocitadas (o ingeridas) por células de mayor tamaño. Algunas de estas asociaciones simbióticas habrían sido favorables y por lo tanto podrían haber conquistado diferentes ambientes. De forma similar, los cloroplastos se incorporaron cuando ciertos procariontes fotosintéticos fueron ingeridos por células eucariontes no fotosintéticas de mayor tamaño y que ya tenían núcleo y mitocondrias. • MÉTODO CIENTÍFICO A lo largo del siglo XVIII, se configuraron dos modos de hacer ciencia que hoy conocemos como método cualitativo-inductivo instaurado por Francis Bacon (1561 – 1626) en el cual el científico recoge datos empíricos y a partir de estos datos llega a una generalización; y el método matemático-deductivo (o hipotético-deductivo), desarrollado por Galileo y sobretodo por Descartes (su obra “Discurso del Método”) en el que el razonamiento va de lo general a lo específico. De esta forma se plantean hipótesis que a su vez hacen una serie de predicciones que se pueden probar mediante experimentos con sus correspondientes controles. Podemos decir que el objetivo de toda ciencia es lograr explicaciones para los hechos observados, y a partir de esos datos establecer determinados principios básicos que permitan prever o predecir las relaciones entre hechos diversos. Las descripciones, las conclusiones y los principios generales que conforman las ciencia de la biología, han sido logrados utilizando una u otra (o ambas) de las herramientas más importantes 7 Endosimbiosis: relación en la cual dos organismos se benefician mutuamente. Ej: los líquenes que están formados por algas y hongos. 8 9 Mitocondrias: organela en la cual ocurre el Ciclo de Krebs, transporte de electrones y fosforilación oxidativa. Cloroplastos: organela en la cual ocurre la fotosíntesis 21 de la investigación científica: la observación y medida directa de fenómenos naturales y el experimento controlado. En la observación, como lo indica la palabra, el investigador no interviene sino sólo observa. En la segunda herramienta, se trata de identificar y medir los factores de estudio en determinadas condiciones conocidas y variando cada factor por separado, manteniendo los otros constantes. Recordar los experimentos de Redi con la carne en descomposición y los frascos con o sin tapa. El experimento científico controlado debe ser planeado en etapas definidas, con testigos válidos, con métodos de medición y análisis elaborados y réplicas numerosas. Una hipótesis es una respuesta a una pregunta que surge, una suposición que debe ser probada a través de la observación y experimentación o de ambas. Puede probarse en forma rápida o puede llegar a necesitar mucho tiempo, como por ejemplo determinar las interacciones que forman la estructura de una selva tropical. En este caso se necesitan tomar muchos datos. En el caso de las hipótesis sobre la organización celular no pudieron ponerse a prueba sino hasta que se fabricó el microscopio electrónico. Cuando una hipótesis importante y fundamental, ha sido puesta a prueba exitosamente en forma Pregunta que surge sobre una cuestión Formulaci ón de una Hipótesis Comp robaci ón de la Hipótesis Ob servaci ón Experime ntaci ón Recolecci ón e interpretación de os l datosobtenidos Hipótesis la Hipótesis Formulaci ón de una Teoría independiente por muchos científicos de diferentes países, pasa a la categoría de teoría. Cuando una teoría ha resistido repetidas pruebas pasa a ser un principio o ley. Entonces las ideas de la ciencia se organizan en distintas categorías que en orden creciente en cuanto a su alcance, se denominan: hipótesis, teorías y principios o leyes. 22 Independientemente del método que se utilice, si se obtienen resultados precisos, éstos pueden llevar a establecer las bases de una generalización, como dijimos, esto es una teoría científica. Sin embargo en biología no hay un “método científico” único, sí hay una gran cantidad de métodos y metodologías que deben usarse según cada caso y que están relacionadas con la pregunta que se quiera responder. Al decir que existe una relación entre hipótesis, comprobación y teoría, esto no implica una realidad universal porque muchas veces en las fases iniciales de una investigación, surgen hechos que obligan a un replanteo de la hipótesis (como se puede ver en el esquema de arriba cuando los datos no confirman la Hipótesis). También es frecuente el descubrimiento por azar de un hecho durante el desarrollo experimental que puede cambiar el curso de la investigación. Como le ocurrió al médico húngaro Ignaz Semmelweis (1819 – 1865), considerado una figura pionera en antisepsia y prevención de la infección nosocomial (u hospitalaria). En esa época un médico amigo se lastima con el bisturí mientras estaba realizando una autopsia: al cabo de pocos días comenzó a mostrar los mismos síntomas que las mujeres que padecían la fiebre posparto. Entonces Semmelweis afirmó: “la materia cadavérica que introdujo el bisturí en la sangre de su amigo, es la causa de su infección”.Al seguir investigando, descubrió que los mismos médicos eran los portadores de la materia cadavérica que enfermaban a sus pacientes. Observó que luego de realizar una autopsia, los médicos revisaban a las madres que habían dado a luz, habiéndose lavado las manos en forma superficial. Ante esto, Semmelweis colocó carteles en la puerta de la sala que decían: “¡Prohibida la entrada a todo doctor que no haya lavado sus manos con solución clorada!”. La mortalidad comenzó a disminuir y en el año 1848 había descendido a un 1,2%. En 1861, Semmelweis escribe el libro “La Etiología, los conceptos y la profilaxis de la fiebre puerperal”. Fallece a los 47 años de edad como consecuencia de una septicemia, provocada por una herida al realizar una autopsia en un hospital para enfermos mentales. Posteriormente Pasteur publicaría la hipótesis microbiana y Joseph Lister extendería la práctica quirúrgica higiénica al resto de las especialidades médicas. • UNIDADES DE MEDIDAS La enorme multitud de especies que constituyen la biosfera abarcan desde el tamaño de un virus, que se mide en nanometros (1nm = 10-9 m), pasando por bacterias o protozoos del orden de micrometros (1μm = 10-6 m), hasta las especies que representan los límites superiores del tamaño orgánico: las ballenas (30 m), los alerces (100 m), etc. En la siguiente figura se indica el tamaño de las diferentes células, bacterias, virus y moléculas en escala logarítmica y se los compara con los límites de la resolución del ojo humano, el microscopio10 óptico y el electrónico. El ojo humano puede resolver o discriminar dos puntos separados por más de 0,1 mm (100 μm). La mayoría de las células son mucho menores y necesitan del microscopio óptico (0,2 μm) para ser estudiadas. La mayor parte de las estructuras celulares son todavía más pequeñas y necesitan del poder de resolución del microscopio electrónico. 10 Microscopio: palabra griega que significa “para ver lo pequeño” 23 Cuadro comparativo de distintos ejemplos de niveles de organización a nivel microscópico. El microscopio Como se expresó, la microscopia es fundamental para el estudio de la biología, incluso desde el nivel de organización de órganos, se necesita del poder de resolución de un microscopio óptico o de una lupa. A continuación una breve reseña del origen de la microscopia y su importancia en el campo de la biología. Ya los antiguos sabían que los espejos curvos y las esferas de cristal llenas de agua aumentaban el tamaño de las imágenes. En las primeras décadas del siglo XVII se iniciaron experiencias con lentes a fin de lograr el mayor aumento posible. Para ello se basaron en otro instrumento con lentes que obtuvo gran éxito, el telescopio, usado por primera vez con fines astronómicos por Galileo en 1609. Así la biología por primera vez ampliaba su campo gracias a un mecanismo que llevaba el sentido de la vista humana más allá de sus límites naturales. El naturalista holandés Jan Swammerdam (1637 – 1680) observó insectos con el microscopio e hizo dibujos de los detalles que observaba. Descubrió también que la sangre no es un líquido uniformemente rojo, como parecía a simple vista, sino que contenía numerosos corpúsculos pequeños que le daban ese color y hoy llamamos glóbulos rojos. El anatomista holandés Regnier de Graaf (1641 – 1673) realizó estudios similares en animales. Examinó la fina estructura de los testículos y los ovarios. Describió en particular ciertos elementos del ovario que desde entonces se conoce con el nombre de folículos de Graaf. Pero uno de los descubrimientos más importantes fueron los realizado por el fisiólogo italiano Marcello Malpighi (1628 – 1694), quien además de estudia plantas e insectos, observó los pulmones de las ranas y vio una compleja red de vasos sanguíneos. También comprobó que los vasos pequeños se unían a vasos mayores que eran venas en una dirección y arterias en la dirección opuesta. Se completaba así la teoría de la circulación de la sangre. Propuesta primariamente por Harvey en 1660. Sin embargo no fue Malpighi quien impuso el microscopio, sino que fue un comerciante holandés Antón van Leeuwenhoek (1632 – 1723) para quien la microscopia era un pasatiempo y fue el primero en observar los espermatozoides y los protozoos en agua estancada. Se hizo evidente que no solo existían objetos demasiados pequeños para poder ser vistos a ojo desnudo sino que también había organismos vivos de esa dimensión. Nace así la microbiología. Posteriormente observa 24 organismos todavía más pequeños que los protozoos y que más tarde serían llamadas bacterias. Uno de los estudios de esa época que tendrían una importancia significativa en biología, fueron realizadas por el científico inglés Robert Hooke (1635 – 1703), quien en 1665 publica su libro “Micrographia” en el cual dibuja sus observaciones al microscopio. Dentro de ellas, la más importante fue la de un delgado trozo de corcho el cual ve que estaba constituido por una fina trama de celdillas rectangulares a las que llamó células. Aunque la Teoría Celular aún tardaría más de un siglo en formularse, este hecho daría inicio a la “citología”. Durante el siglo XVIII, el uso del microscopio decae. Uno de los defectos de los microscopios primitivos era que sus lentes descomponían la luz blanca y las observaciones se veían rodeadas de anillos de color (aberración cromática). Recién alrededor de 1820 se perfeccionan y surgen los microscopios acromáticos. Durante el siglo XIX el uso del microscopio permitió abrir el camino hacia nuevos progresos en el campo de la biología. Regnier de Graaf • Antón van Leeuwenhoek Microscopio óptico Esquema de un microscopio óptico El microscopio óptico nunca puede resolver detalles menores a 0,2mm, la medida de una pequeña bacteria. Esta resolución es limitada por la longitud de onda de la luz visible usada para iluminar la muestra. Los microscopios ópticos o de luz pueden aumentar efectivamente alrededor de 1000 a 1500 veces el tamaño de la muestra real; si se incrementase el aumento la imagen proyectada sería borrosa. La mayoría de las mejoras en microscopia de luz del comienzo de este siglo ha involucrado nuevos métodos para aumentar el contraste. Sin estas técnicas sería imposible para el ojo humano el conocimiento del mundo celular. 25 • Microscopio electrónico El electrónico utiliza un haz de electrones para obtener una imagen aumentada y utiliza lentes electromagnéticas. La mayoría de los microscopios electrónicos modernos pueden lograr una resolución de 0,2 nm, unas 1000 veces mejor que la obtenida con el microscopio óptico. Los biólogos usan el término ultraestructura celular para referirse a la anatomía celular cuando se resuelve por un microscopio electrónico. Comparación entre un Microscopio Óptico y uno Electrónico Característica IMAGEN D ADA MECANISMO: Microscopio Óptico M icroscopio Electrónico POR De i nterferencia d e rayos luminosos De dispersión de electrones Al v acío con g ran diferencia de potencial TUBO Simple con aire FUENTE Luz (fotones) Filamento de t (electrones) ELEMENTOS Lentes: Ocular, objetivo y condensador Bobinas electromagnéticas ESTUDIAN Células vivas o muertas Células muertas OBSERVACIÓN IMAGEN Distintas tonalidades de g ris. En l a actualidad s e ven imágenes " sombreadas" electrónicamente. 3 Å a 5 Å (teórico) DE L A Coloreadas o no LIMITE DE RESOLUCIÓN ungsteno 0,25m 10 Å (en la práctica AUMENTO (el signo significa aumento) Diferencias entre Microscopio Óptico y Microscopio Electrónico X 500 X a 1500 X 30.000 X a 1.000.000 X Nivel microscópico: Nivel submicroscópico: ESTRUCTURA ULTRAESTRUCTURA NIVEL DE OBSERVACIÓN 26 Hay dos tipos de microscopio electrónico: el microscopio electrónico de transmisión (MET) y el microscopio electrónico de barrido (MEB): El MET permite develar la ultraestructura de las células y de la matriz extracelular. El haz de electrones es desviado por un campo electromagnético (que actuaría como lente). En este tipo de microscopia se requiere que las muestras tengan un grosor de 100 nm. El MEB es especialmente útil para estudios de superficie del espécimen. El haz de electrones explora la superficie de la muestra la que usualmente se recubre con una película de oro. El haz excita a los electrones sobre la superficie de la misma muestra y estos electrones secundarios se recolectan y enfocan sobre una pantalla. Esto forma una imagen de la topografía del espécimen. Un importante atributo del MEB es su gran profundidad de campo, la cual resulta en una imagen tridimensional. El ME revela muchas organelas que son imposibles de resolver con MO. Pero el MO ofrece muchas ventajas especialmente para el estudio de la célula viva. Una desventaja de ME es que los métodos químicos y físicos usados para preparar la muestra, no sólo matan a las células sino que puedan introducir artefactos, estructuras peculiares vistas en micrografías que no existe en una célula viva. Fotografías de observaciones con: microscopio óptico (Nº 1) y microscopio electrónico (Nº 2 y 3) 27 Microfotografía electrónica de células de Escherichia coli. BIBLIOGRAFÍA Alberts, B; Watso, J; Bray, D; Lewis, J; Raff, M; Roberts, K. Biología Molecular de la Célula; 3ª Edición; Ediciones Omega S.A. Barcelona. 1996. Asimov, I. Breve Historia de la Biología. 3ª Edición. Editorial Universitaria de Buenos Aires. 1975. Avers, Ch. Biología celular. Grupo Editorial Iberoamérica. 1991. Castro, R; Hande, M; Rivolta, G. Actualizaciones en Biología. 8ª Edición. Editorial Universitaria de Buenos Aires. 1990. Curtis, H; Barnes, N. Biología. 7ª Edición. Editorial Médica Panamericana. Buenos Aires. 2008. Curtis, H; Barnes, N. Invitación a la Biología. Editorial Médica Panamericana. Buenos Aires. 1987. De Robertis, E; Hib, J. Fundamentos de Biología Celular y Molecular. 4ª Edición. Editorial El Ateneo. Buenos Aires. 2004. De Robertis, E; Hib, J. Biología Celular y Molecular. 12ª Edición. Editorial El Ateneo. Buenos Aires. 1996. Dickerson, R. La Evolución Química y el origen de la Vida. Investigación y Ciencia Nº 26. No28 viembre 1978. Holtzman, E; Novikoff, A. Estructura y Dinámica Celular. 3ª Edición. México. Editorial Interamericana. 1986. Karp, G. Biología Celular y Molecular. Editorial Mc Graw Hill Interamericana. México. 1998. Lehninger, A. Bioquímica. 2ª Edición. Editorial Omega S.A. México. 1985. Lehninger, A; Nelson, D; Cox, M. Principios de Bioquímica. 4ª Edición. Editorial Omega S.A. España. 2005. Moreno Azorero, R; Schwartzman, G. Principios de Biología Celular. Editorial El Ateneo. Buenos Aires. 1986. Solomon, E; Ville, C; Davis, P. Biología. 4ª Edición. Editorial Mc Graw Hill Interamericana. México. 1998. 29 MÓDULO Nº 2: BIOLOGÍA CELULAR 1. COMPONENTES QUÍMICOS CELULARES De todos los elementos que constituyen la materia del Universo, sólo cuatro: Carbono (C), Oxígeno (O), Hidrógeno (H) y Nitrógeno (N), conforman el 98% de la materia viva. Sin embargo estos cuatro elementos se distribuyen en forma muy diferente en la corteza terrestre, así por ejemplo aproximadamente cerca del 50 % de los átomos de los organismos vivos son átomos de hidrógeno mientras que en la corteza terrestre (mundo inanimado) se encuentra en un porcentaje inferior al 1 %. El átomo de carbono tampoco es abundante en la corteza terrestre, representa sólo el 0,08 % en peso, pero resulta fundamental en la composición de los seres vivos. En la siguiente Tabla se observa que sólo doce (12) elementos constituyen el 99,81 % de la materia viva: ELEMENTO % MEDIO EN PESO 76,00 10,50 10,00 2,50 0,30 0,30 0,10 0,04 0,02 0,02 0,02 0,01 Oxígeno Carbono Hidrógeno Nitrógeno Fósforo Potasio Cloro Sodio Calcio Magnesio Azufre Hierro El restante 0,19 % está constituido por elementos que se encuentran presentes en cantidades inferiores a 0,01 % y se llaman oligoelementos aunque son también esenciales, tales como: Cobre, Cobalto, Manganeso, Fluor y Zinc. El agua, como se verá a continuación en el ítem 2, es la sustancia más abundante en las células vivas y constituye aproximadamente el 70 % del peso de las células. A parte del agua, la gran mayoría de las otras moléculas de una célula son compuestos de carbono y forma parte de la Química Orgánica. Dentro de este grupo encontramos monosacáridos, polisacáridos, aminoácidos, proteínas, lípidos, nucleótidos y ácidos nucleicos. Estos no son los únicos compuestos orgánicos que existen pero sí son la mayoría. Estos representan aproximadamente el 30 % de la composición química de los seres vivos. El 70 %, como se menciona arriba, lo constituye el Agua. También - 30 - encontramos algunos iones como: Na (sodio), Fe (hierro), Ca (calcio), K (potasio), etc. pero en proporciones muy pequeñas. El carbono se destaca entre todos los elementos de la Tierra por su capacidad de formar grandes moléculas con enlaces covalentes fuertes. Se puede unir a través de estos enlaces, a otros átomos de carbono formando cadenas carbonadas y anillos, generando así moléculas grandes y complejas. Los otros átomos abundantes de las células, H, O y N también son pequeños y capaces de formar enlaces covalentes muy fuertes. Los átomos de carbono pueden unirse con otros átomos de carbono y con muchos de otro tipo para formar una gran variedad de compuestos orgánicos. (La muscona, es el atrayente sexual del venado almizclero del Tibet.) Composición química de los seres vivos Todas las células están gobernadas por los mismos principios físicos y químicos de la materia inerte. Si bien dentro de las células encontramos moléculas que usualmente no existen en la materia inanimada, en la composición química de los seres vivos encontramos desde sencillos iones inorgánicos, hasta complejas macromoléculas1 orgánicas siendo todos igualmente importantes para constituir, mantener y perpetuar el estado vivo. 1 Macromoléculas: término que se usa para designar a moléculas de “gran tamaño”, sin embargo no define que se entiende por gran tamaño, muchas macromoléculas son polímeros (polisacáridos, ácidos nucleicos, proteínas) pero otros no (esteroides, triglicéridos, etc.) - 31 - Esquema de la composición química de la materia viva Composición porcentual de la materia viva Compuesto Agua Porcentaje de peso total 70 Macromoléculas: * Proteínas 15 Constituido aproximadamente por 1% de ADN y 6% de ARN. Ácidos Nucleicos 7* ** Polisacáridos 3 Lípidos 2 Moléculas orgánicas pequeñas Iones inorgánicas Incluyen los bloque de construcción para generar macromoléculas y otras moléculas en los procesos de síntesis o degradación 2** 1 2. COMPONENTES INORGÁNICOS Agua Como se mencionó en el capítulo anterior, el agua es la sustancia más abundante e importante puesto que la química de la vida ocurre en ella. De hecho, las células contienen entre un 70 a un 90 % de agua, y todas las reacciones que ocurren en el citoplasma de una célula tiene lugar en un medio acuoso. El agua es el solvente biológico ideal. Sin embargo el agua no solo es el medio en el que se desarrollan las reacciones químicas sino que también en muchos casos participa activamente de ellas ya sea como reactivo o producto de una reacción. - 32 - La molécula de agua consta de dos átomos de hidrógeno y uno de oxígeno, unidos covalentemente. Debido a la diferencia de electronegatividad entre los mismos se crea una distribución asimétrica de cargas lo que llega a la formación de una molécula polar. Esa polaridad permite la aparición de los puentes de hidrógeno entre las moléculas de agua. Figura: (a) Estructura de la molécula de agua. (b) Las moléculas de agua en disolución interactúan entre si a través de los puentes de hidrógeno (líneas punteadas) Muchas de las propiedades del agua se explican debido al gran número de puentes de hidrógeno que existen entre sus moléculas. El agua es líquida en un amplio intervalo de temperaturas que va desde los 0 °C a los 100 °C, lo que indica que no solo debemos entregarle calor para que ocurra el cambio de estado sino para poder romper los puentes de hidrógeno. El calor de evaporación del agua es muy superior al de otros líquidos por lo que muchos organismos utilizan esta propiedad para el mantenimiento de la temperatura corporal. El hielo flota en el agua, es decir que el agua al estado sólido es más liviana que al estado liquido, (debido a que por la acción de los puentes de hidrógeno en el estado sólido se forma una verdadera red cristalina por lo que el agua al congelarse se dilata, disminuyendo así su peso específico). Esta propiedad permite que la capa de hielo que cubre un río o un lago, flote sirviendo entonces como aislante, permitiendo que la vida acuática continúe. El agua actúa como disolvente para moléculas polares, principalmente para aquellas con las que puede formar puentes de hidrógeno. La alta polaridad del agua favorece también a la célula porque fuerza a las sustancias no polares a agregarse y permanecer juntas, contribuyendo así a la estructura de las membranas. Como se verá más adelante - 33 - las membranas biológicas están constituidas principalmente por sustancias no polares (lípidos) los cuales se agregan y cumplen una función de barrera selectiva. Propiedades químicas del agua: Moléculas hidrofóbicas y estructuras acuosas Las moléculas que son no polares y no pueden formar enlaces de hidrógeno (por ej. Hidrocarburos) sólo tienen una limitada solubilidad en agua, y se denominan hidrofóbicas. Cuando estás moléculas se encuentran con agua, las moléculas de agua se disponen a su alrededor de manera ordenada, como bloques de hielo. Estas estructuras son más ordenadas que el agua libre. En la figura de arriba, se muestra estas estructuras de agua (gris) rodeando al hidrocarburo (negro) en el centro. Ácidos y bases Un ácido es una molécula que, en solución, cede un ión H+ (protón). Ejemplo: Una Base es una molécula que, en solución, acepta un ión H+ (protón). Ejemplo: - 34 - El agua por si misma tiene una débil tendencia a ionizarse, actuando tanto como ácido débil y como base débil. Cuando actúa como ácido débil libera un protón, generando un ión hidroxilo. Como base débil acepta un protón formando ión hidronio. En solución acuosa la mayoría de protones están como iones hidronio. Ósmosis Si dos soluciones acuosas están separadas por una membrana que únicamente permite el paso de las moléculas de agua, dichas moléculas pasaran hacia la solución que contiene la mayor concentración de moléculas solubles, denominándose a dicho proceso ósmosis. (Se volverá a ver en mayor detalle en el capítulo de Difusión y ósmosis). pH Este pasaje del agua desde una solución hipotónica a una hipertónica, puede provocar un aumento de la presión hidrostática en el compartimiento hipertónico. Cuando las dos soluciones se equilibran, teniendo concentraciones idénticas de solutos, se dice que son isotónicas. (También se verá en mayor detalle en el capítulo de Transportes a través de la membrana). Minerales Como ya vimos, del total de los componentes de la célula, un 75 a 85% corresponde a H2 O; entre el 2 y el 3% son sales inorgánicas y el resto son compuestos orgánicos. Ciertos minerales se encuentran en forma no ionizada. Así ocurre con el calcio, que en el hueso y dientes, se encuentra unido al fosfato y carbonato bajo la forma de cristales. - 35 - Otro ejemplo es el hierro que en la hemoglobina, la ferritina, los fotocromos y en varias enzimas se halla ligado por uniones carbono-metal. Para mantener la actividad celular normal son indispensables pequeñas cantidades de manganeso, cobre, cobalto, yodo, selenio, níquel, molibdeno y cinc. Casi todos estos elementos son necesarios para la actividad de ciertas enzimas. El yodo es un componente de la hormona tiroidea. El magnesio es necesario para el normal funcionamiento del músculo. El flúor es un componente de los dientes. La mayoría de estos elementos están presentes en la dieta común y/o en el agua de consumo. 3. COMPUESTOS ORGÁNICOS Los compuestos orgánicos se basan en el carbono combinado con otros átomos de carbono y con otros elementos como el hidrógeno, el nitrógeno, el azufre, etc. Formando grandes estructuras con distinta complejidad y diversidad. El átomo de carbono puede unirse a otros átomos de carbono formando largas cadenas las cuales pueden ser lineales, ramificadas o bien cíclicas. (Ver el capítulo 1 “Componentes químicos celulares”). Se han aislado miles de compuestos de carbono de varios sistemas biológicos. Algunos ejemplos: - 36 - Recordando brevemente qué es un Grupo Funcional: Si bien en esta materia, no se verá Química Orgánica, es necesario e importante recordar algunos conceptos que nos servirán para comprender los temas que se desarrollarán en los capítulos siguientes. Se puede decir que las propiedades químicas específicas de una molécula orgánica derivan principalmente de los grupos de átomos conocidos como grupos funcionales. Estos grupos están unidos al esqueleto de carbono, reemplazando a uno o más de los hidrógenos que estarían presentes en un hidrocarburo. Un grupo -OH (hidroxilo) es un ejemplo de un grupo funcional. Un compuesto con un grupo hidroxilo que reemplaza a uno o más de los hidrógenos de un hidrocarburo, se conoce como alcohol. En la siguiente Tabla, se indican los grupos funcionales de mayor interés para la comprensión de los sistemas vivos. Al conocer los grupos funcionales se puede predecir sus propiedades. Por ejemplo, los alcoholes, con sus grupos hidroxilos polares, tienden a ser solubles en agua. Los grupos aldehídos, como el formaldehído, tienen olores desagradables. Grupos Funcionales Grupo Clase de Fórmula estructura funcional compuestos Hidroxilo Ejemplo Descripción Compuesto polar porque el oxígeno electronegativo capta electrones de átomo covalentes Alcoholes Etanol Amino Iónico, el grupo amino actúa como base. Aminas Aminoácido Carboxilo Iónico, el hidrógeno puede disociarse como hidrogenión Ácidos carboxílicos (orgánicos) Aminoácido Estér Relacionado con el grupo carboxilo, pero tiene un grupo en lugar del hidrógeno del hidroxilo; polar Esteres Metilacetato - 37 - Carbono de carbonilo enlazado con al menos un átomo de hidrógeno; polar Aldehídos Formaldehído Grupo carbonilo enlazado con otros dos átomos de carbono; polar Carbonilo Cetonas Acetona No polar Metilo Componente de muchos compuestos orgánicos. Metano Fosfato Fosfatos orgánicos Ester de fosfato Sulfidrilo Forma disociada del ácido fosfórico, el ión fosfato se enlaza en forma covalente, por medio de uno de sus átomos de oxígeno, con uno de los átomos de carbono; iónico. Ayuda a estabilizar la estructura interna de las proteínas Tioles Cisteína Monómeros, polímeros y macromoléculas Los monómeros son unidades moleculares sencillas agrupadas de acuerdo a sus propiedades químicas; podemos mencionar a los: monosacáridos, aminoácidos y los nucleótidos. Cuando los monómeros de un mismo grupo se unen entre sí en forma covalente obtenemos un polímero que es una molécula de gran tamaño. Los polímeros pueden ser cadenas relativamente sencillas de unidades monoméricas idénticas o pueden ser en extremo complejos. Las cadenas pueden ser ramificadas o sin ramificaciones. Los compuestos en los organismos son utilizados de distintas formas, para almacenar o liberar energía, para actuar como catalizadores, para guardar información, para construir estructuras que se requieren, etc. La mayoría de las biomoléculas que utilizan los organismos en su funcionamiento son polímeros. El término macromolécula, como se vio en el capítulo 1, se usa para designar a moléculas de “gran tamaño”. Se han identificado en los sistemas vivientes unos 30.000 - 38 - compuestos orgánicos diferentes, sin embargo un pequeño grupo de ellas son las que cumplen funciones principales en los organismos. Figura: Esquema de polímero Existen cuatro grandes grupos de biomoléculas: Lípidos, Glúcidos (Hidratos de Carbono o Azúcares), Proteínas y Ácidos Nucleicos. I. Lípidos Constituyen un grupo de compuestos muy heterogéneo, cuya única característica común es la insolubilidad en agua y otros solventes polares. Son solubles solamente en solventes no polares como el éter, benceno, cloroformo, etc. Entre las biomoléculas, los lípidos son los únicos que en general no forman polímeros, aunque algunos pueden considerarse macromoléculas. Aunque existen otras, una manera sencilla de clasificarlos es la siguiente: Sus funciones biológicas son muy diversas. A continuación se muestran algunas de las principales funciones de los lípidos en los seres vivos: - 39 - Lípidos Función Biológica Ácidos Grasos Combustibles celulares Grasa y Aceites Principalmente reserva de energía en forma más eficiente que los glúcidos. Actúan en la termorregulación (aislante térmico), especialmente en animales que habitan zonas frías. Forman cubiertas protectoras en piel, pelos, plumas. En vegetales se encuentran formando una película en las hojas y frutos. Componente principal de las membranas biológicas. Forman parte de la vaina de mielina que recubre los axones de las células nerviosas. Igual que los fosfolípidos y el colesterol, forman parte de la estructura de las membranas biológicas. El más conocido es el colesterol que se encuentra en las membranas biológicas de todas las células excepto en bacterias. Los esteroides poseen diversas funciones de acuerdo a los grupos químicos que se unen a su estructura. Ceras (ej: colesterol) Fosfoglicéridos Glucolípidos Esteroides II. Glúcidos La mayor fuente de glúcidos, se encuentra en los vegetales, los cuales a través del proceso de fotosíntesis combinan el dióxido de carbono (CO2) y el agua (H2O) para dar las moléculas hidrocarbonadas que son los glúcidos. Estas moléculas proporcionan a las plantas y a los animales que se alimentan de ellas, la energía necesaria para los procesos metabólicos (se verá luego en Fotosíntesis) A excepción de la vitamina C, los glúcidos no son esenciales en la dieta, ya que el organismo mediante procesos metabólicos intracelulares puede sintetizar los azúcares necesarios a partir de otras moléculas, como los lípidos y aminoácidos. Clasificación: Los glúcidos se clasifican en primer lugar, teniendo en cuenta el número de unidades constitutivas de los mismos en: - 40 - Monosacáridos: constituidos por un azúcar simple. Oligosacáridos: Resultantes de la unión de 2 a 10 unidades de monosacáridos. Polisacáridos: formados por cadenas compuestas de muchas unidades de monosacáridos (más de 10). Estas cadenas pueden ser lineales o ramificadas. Algunos ejemplos de Monosacáridos: Las funciones biológicas de los monosacáridos, especialmente la glucosa, es ser la principal fuente de energía celular. También forman parte de moléculas más complejas. Por ejemplo la ribosa y desoxirribosa, componentes de los ácidos nucleícos. Oligosacáridos: De todos ellos los más importantes fisiológicamente son los disacáridos, como la sacarosa o azúcar común formada por la unión de glucosa y fructosa, la lactosa o azúcar de la leche (Galactosa + glucosa), o la maltosa o azúcar de malta formada por la unión de dos glucosas. Función biológica de los oligosacáridos: son formas de transporte en los vegetales y en algunos animales. Forman parte de moléculas más complejas como las glucoproteínas y glucolípidos. Intervienen en la estructura de la membrana plasmática, participando en el reconocimiento celular. Polisacáridos: Están constituidos por un gran número de monosacáridos unidos mediante enlaces glucosídicos, constituyendo largas cadenas. - 41 - Los polisacáridos más importantes presentes en la naturaleza son el almidón, el glucógeno y la celulosa. Almidón: Su función es la de ser la principal reserva de energía en las plantas. Glucógeno: Constituye una importante reserva de energía para los animales y se almacena principalmente en el hígado y en los músculos. Figura: Representación esquemática del glucógeno Celulosa: Es el polisacárido estructural más abundante y se lo encuentra formando las paredes celulares de los vegetales. Está constituida por cadenas lineales de b-glucosa. Figura: Celulosa Además de estos polisacáridos, existen otros que también cumplen funciones estructurales importantes, como por ejemplo la mureína de las paredes celulares de bacterias o la quitina del exoesqueleto de los insectos - 42 - III. Proteínas Las proteínas son las macromoléculas más abundantes en las células animales y constituyen alrededor del 50% de su peso seco. Dentro de las células se las encuentra en formas muy variadas: como constituyente de las membranas biológicas, como catalizadores de reacciones metabólicas (enzimas), interactuando con los ácidos nucleicos (histonas) o con neurotransmisores y hormonas (receptores), etc. Prácticamente, no existe proceso biológico en el que no participe por lo menos una proteína. Se las considera como el grupo de compuestos que mayor cantidad de funciones desempeñan en los seres vivos. Estas moléculas son polímeros de aminoácidos unidos por enlaces peptídicos. Las proteínas pueden ser simples o conjugadas. Las simples sólo están formadas por aminoácidos. Las conjugadas contienen además de la o las cadenas polipéptidicas, grupos no proteicos, denominados grupos prosteicos, por ejemplo la hemoglogina o las lipoproteínas. Para entender los aspectos estructurales y las características químicas de las proteínas, es fundamental analizar primero la de sus monómeros. Aminoácidos Como su nombre lo indica, cada aminoácido está formado por un grupo amino y un grupo ácido carboxílico, unidos a un átomo de carbono central o carbono α (carbono alfa) , el que además tiene unido siempre un átomo de hidrógeno y una cadena lateral de características variables. Figura: Fórmula general Como muestra la fórmula, el carbono central se encuentra unido a un grupo variable o resto (R). Es en dichos grupos R, donde las moléculas de los veinte aminoácidos que forman parte de las proteínas se diferencian unas de otras. En la glicina, el más simple de los ácidos, el grupo R se compone de un único átomo de hidrógeno. En otros aminoácidos el grupo R es más complejo, conteniendo carbono e hidrógeno, así como oxígeno, nitrógeno y azufre. - 43 - De acuerdo con la naturaleza del “R” podemos clasificar a los aminoácidos en polares (con y sin carga) y aminoácidos no polares. Aminoácidos Esenciales: La síntesis proteica requiere de un constante aporte de aminoácidos. Los organismos heterótrofos sintetizan gran parte de estos aminoácidos a partir de esqueletos carbonados. Los que requieren ser incorporados por la ingesta, no pudiendo ser sintetizados, se denominan aminoácidos esenciales, y son producidos por plantas y bacterias. Aminoácidos no esenciales y esenciales para el Hombre No esenciales Esenciales Glutamato Isoleucina Glutamina Leucina Prolina Lisina Aspartato Fenilalanina Asparagina Metionina Alanina Treonina Glicina Triptofano Serina Valina Tirosina Histidina Cisteína Arginina ( sólo en lactantes) Cuando una célula viva sintetiza proteínas, el grupo carboxilo de un aminoácido reacciona con el grupo amino de otro, formando un enlace peptídico, el producto de esta unión es un dipéptido. El grupo carboxilo libre del dipéptido reacciona de modo similar con el grupo amino de un tercer aminoácido, y así sucesivamente hasta formar una larga cadena. Los oligopéptidos contienen un número indefinido pero pequeño de aminoácidos, mientras que los péptidos y polipéptidos constan de un número mayor. Muchas moléculas de importancia biológica con acción hormonal e incluso gran parte de los neurotransmisores son oligopéptidos y péptidos, como se observa en las siguientes Tablas: - 44 - Péptidos con función hormonal Nombre Oxitocina Nº de aminoácidos Órgano productor Órgano blanco Función Contracción del Útero, glándula músculo liso uterino mamaria. y eyección de leche Antidiurética y Riñón, vasos. vasopresora Acción Crecimiento generalizada Promueve la síntesis Testículos y de los esteroides ovarios andrógenos y estrógenos, etc. Crecimiento de tubos Testículos y seminíferos y ovarios desarrollo folicular. Glándula Estimula la secreción mamaria de leche 9 Hipotálamo 9 Hipotálamo 191 Hipófisis Hormona luteinizante (LH) 200 Hipófisis Hormona Folículo estimulante (FSH) 200 Hipófisis Prolactina (PRL) 191 Hipófisis Hormona adrenocorticotrofa (ACTH) 39 Hipófisis Corteza adrenal Estimula la secreción de corticoesteroides Tirotropina (TSH) 220 Hipófisis Tiroides Estimula secreción de tiroxina Vasopresina (ADH) Hormona del crecimiento (GH) Algunos Péptidos con función neurotransmisora Nombre Nº de aminoácidos Efecto Sustancia P 11 Dolor Angiotensina II 8 Ansiedad Encefalinas (ENK) 4 Control del dolor Colecistoquinina(CCK) 8 Regulación del apetito Beta-endorfina 31 Placer y analgesia Cuatro niveles de la Estructura Proteica Estructura Primaria: Es la secuencia ordenada y única de los aminoácidos en la cadena polipeptídica, la cual está determinada genéticamente. La estructura primaria es fundamental para la forma tridimensional que tendrá la proteína. Cualquier modificación en la secuencia de aminoácidos podría ocasionar un cambio en la estructura tridimensional y afectará la función biológica de la proteína. Por - 45 - ejemplo la enfermedad humana hereditaria Anemia Falciforme, origina profundos cambios biológicos por la sustitución de un solo aminoácido en la molécula de hemoglobina. Estructura Secundaria: Es la organización espacial de aminoácidos que se encuentran próximos en la cadena peptídico. Algunas regiones pueden tener una estructura en forma de cilindro, o estructura en α hélice, en la cual la cadena peptídico se enrolla alrededor de un cilindro imaginario y queda estabilizada por puentes de hidrógeno. En la estructura en hoja β plegada, los aminoácidos adoptan la conformación de una hoja de papel plegada y su estructura se estabiliza también por puentes de hidrógeno. Otros segmentos de la proteína no muestran estas uniones transversales y adoptan una configuración al azar, denominada ad random. Figura: Esquema de una proteína presentando regiones con estructura secundaria en αHélice, en Hoja β-Plegada y regiones con enroscamientos aleatorios (ad random). - 46 - Estructura Terciaria: Es la relación tridimensional de los segmentos de aminoácidos que pueden estar muy separados entre sí en la secuencia lineal. En muchas proteínas la estructura terciaria le brinda a la proteína una forma globular, como por ejemplo en las enzimas, que son proteínas con función catalítica. Otras proteínas tienen estructura terciaria fibrosa y suelen tener largas hélices o extensas hojas plegadas. Estas proteínas fibrosas suelen tener función estructural como el colágeno. Estructura Cuaternaria: Es la disposición de subunidades proteicas en proteínas complejas formadas por dos o más de estas subunidades. Por ejemplo la molécula de hemoglobina está compuesta por cuatro cadenas polipeptídicas, dos de ellas denominadas α y dos β. La molécula de hemoglobina puede disociarse por acción de la urea en dos α y dos β. Cuando se retira la urea del medio, se vuelven a asociar. Son uniones altamente específicas que se da entre dos hemimoléculas. Figura: Estructura de la molécula de hemoglobina (estructura cuaternaria). Formada por dos cadenas de α-hemoglobina y dos cadenas de β-hemoglobina. Cada cadena transporta una molécula de oxígeno. - 47 - Funciones biológicas de las proteínas es la de dirigir la totalidad de los procesos celulares, incluso su propia síntesis. Las funciones de mayor importancia de las proteínas en los seres vivos son: Función estructural: Como el colágeno, la tubulina de los microtúbulos, las de las cápsides virales, etc. Función reguladora: Como las ciclinas que controlan el ciclo celular y los factores de transcripción que regulan la expresión de los genes. Función motora: Actina y miosina del músculo. Función de transporte: Globulinas en general, hemoglobima, mioglobina y las lipoproteínas son algunos ejemplos. Función de reserva: La ovoalbúmina, componente principal de la clara de huevo o la gliadina del trigo. Función de receptores: Como las proteínas receptoras de membrana. Función enzimática: La enzimas catalizan todas las reacciones metabólicas Función de defensa: Los anticuerpos son proteínas simples globulares y son sintetizadas por las células plasmáticas (linfocitos B activados), son también conocidas como inmunoglobulinas o gamaglobulinas. . Estas proteínas presentan gran diversidad ya que cada anticuerpo es específico para un determinado antígeno. Función de mensajeros químicos: La mayor parte de las hormonas son proteínas o glucoproteínas. También ciertos aminoácidos, derivados de aminoácidos y oligopéptidos son neurotransmisores en el sistema nervioso. IV. Ácidos Nucleicos Todas las células contienen la información necesaria para realizar distintas reacciones químicas mediante las cuales las células crecen, obtienen energía y sintetizan sus componentes. Está información está almacenada en el material genético, el cual puede copiarse con exactitud para transmitir dicha información a las células hijas. Sin embargo estas instrucciones pueden ser modificadas levemente, es por eso que hay variaciones individuales y un individuo no es exactamente igual a otro de su misma especie (distinto color de ojos, piel, etc.). De este modo, podemos decir que el material genético es lo suficientemente maleable como para hacer posible la evolución. La información genética o genoma, está contenida en unas moléculas llamadas ácidos nucleicos. Existen dos tipos de ácidos nucleicos: ADN y ARN. El ADN guarda la información genética en todos los organismos celulares, el ARN es necesario para que se exprese la información contenida en el ADN; en los virus podemos encontrar tanto ADN como ARN conteniendo la información (uno u otro nunca ambos). - 48 - Estructura de los ácidos nucleicos Los ácidos nucleicos resultan de la polimerización de monómeros complejos denominados nucleótidos. Un nucleótido está formado por la unión de un grupo fosfato al carbono 5’ de una pentosa. A su vez la pentosa lleva unida al carbono 1’ una base nitrogenada. Figura: Estructura del nucleótido monofosfato de adenosina (AMP) Las bases nitrogenadas son moléculas cíclicas y en la composición de dichos anillos participa, además del carbono, el nitrógeno. Estos compuestos pueden estar formados por uno o dos anillos. Aquellas bases formadas por dos anillos se denominan bases púricas (derivadas de la purina). Dentro de este grupo encontramos: Adenina (A), y Guanina (G). Si poseen un solo ciclo, se denominan bases pirimidínicas (derivadas de la pirimidina), como por ejemplo la Timina (T), Citosina (C), Uracilo (U). - 49 - Nucleótidos de importancia biológica ATP (adenosin trifosfato): Es el portador primario de energía de la célula. Esta molécula tiene un papel clave para el metabolismo de la energía. La mayoría de las reacciones metabólicas que requieren energía están acopladas a la hidrólisis de ATP. Figura: ATP (Adenosin trifosfato) AMP cíclico: Es una de las moléculas encargadas de transmitir una señal química que llega a la superficie celular al interior de la célula. segundo mensajero) NAD+ y NADP+: (nicotinamida adenina dinucleótido y nicotinamida adenina dinucleótido fosfato). Son coenzimas que intervienen en las reacciones de oxidoreducción, son moléculas que transportan electrones y protones. Intervienen en procesos como la respiración y la fotosíntesis. FAD+: También es un transportador de electrones y protones. Interviene en la respiración celular. Coenzima A: Es una molécula que transporta grupos acetilos, interviene en la respiración celular, en la síntesis de ácidos grasos y otros procesos metabólicos. - 50 - Polinucleótidos Existen dos clases de nucleótidos, los ribonucleótidos en cuya composición encontramos la pentosa ribosa y los desoxirribonucleótidos, en donde participa la desoxirribosa. Los nucleótidos pueden unirse entre sí, mediante enlaces covalentes, para formar polímeros, es decir los ácidos nucleicos, el ADN y el ARN. ADN – Ácido desoxirribonucleico Los ácidos nucleicos fueron aislados por primera vez en 1869, sin embargo no fue hasta mucho después que se conoció su función. A principio de siglo los científicos que querían explicar como se transmitía y se almacenaba la información genética se enfrentaron a un problema, era el ADN o las proteínas de los cromosomas los que portaban la información genética. Se sabía que el ADN constaba de solo cuatro tipo de monómeros, frente a los 20 aminoácidos que se encuentran formando parte de las proteínas, de manera que se pensaba que era demasiado sencillo como para guardar la información, por lo cual se le asignaba una función estructural. La evidencia que ha servido para esclarecer la función del ADN, ha procedido, por un lado, del hecho que la cantidad de ADN de una especie es constante, sin importar la edad, sexo, factores nutricionales o ambientales. Por otra parte, la cantidad de ADN tiene mayoritariamente una relación directa con la complejidad del organismo, así como también se observa que las gametas de los individuos con reproducción sexual poseen solo la mitad del ADN que posee cualquier de sus células somáticas. (Se verá en detalle en el Módulo Nº 3) Sin embargo esto por si solo no confirmó la función del ADN. Por ello se llevaron a cabo una serie de experimentos que lo demostraron en forma concluyente. En 1928, Griffith experimentó con distintas cepas de bacterias, una de ellas era la forma llamada lisa (L), rodeada de una cápsula de polisacáridos y causante de neumonía en los ratones. En contraste, las cepas rugosas ( R ), no contenía el polisacárido y no era virulenta. Griffith experimentó con ratones. A unos inyectándoles cepas lisas muertas por calor, a otras cepas rugosas vivas y a otros, una mezcla de cepa R viva con cepa L muertas por calor, en este último caso los ratones morían de neumonía, es decir que las células rugosas se habían transformado en cepas virulentas. En 1944 se demostró que ese principio transformador era el ADN y no las proteínas. - 51 - Figura: Experimento de Griffith Otra serie de experimentos realizados en 1952 por Hershey y Chase, demostraron en forma indiscutible que el ADN es el material genético. Así se sabía la función del ADN pero no su estructura. No fue sino hasta el año 1953 en que Watson y Crick propusieron el modelo de doble hélice, para esto se valieron de los patrones obtenidos por difracción de rayos X de fibras de ADN, y de los postulados enunciados por Chargaff que estableció que la cantidad de adenina de una molécula de ADN era igual a la cantidad de timina de la misma molécula y que la cantidad de guanina era igual a la cantidad de citosina, es decir que el contenido de purinas era igual al de pirimidinas. El modelo de Watson y Crick, describe a la molécula del ADN como una doble hélice, enrollada sobre un eje, como si fuera una escalera de caracol y cada diez pares de nucleótidos alcanzan para dar un giro completo. Excepto en algunos virus, el ADN siempre forma una cadena doble. - 52 - Figura: Pares de bases del ADN: La formación específica de enlaces de hidrógeno entre G y C y entre A y T genera los pares de bases complementarias a b Figura: (a) Modelo de la doble hélice de ADN, (b) Representación abreviada de un segmento de ADN - 53 - Funciones biológicas del ADN es ser el portador de la información genética y a través de ella pode controlar, en forma indirecta, todas las funciones celulares. “Debemos recordar aquí que las enzimas son proteínas que catalizan todas las funciones biológicas y se sintetizan en las células de acuerdo a la información genética. Vale decir que a la información genética la podemos comparar con un recetario, donde están las recetas de todas las proteínas del organismo.” Encontramos ADN en el núcleo de las células animales y vegetales, en los organismos procariontes, en organoides como los cloropastos y mitocondrias, como así también en algunos virus, a los que llamamos ADN - virus. ARN – Ácido ribonucleíco El ácido ribonucleíco se forma por la polimerización de ribonucleótidos. En general los ribonucleótidos se unen entre sí, formando una cadena simple, excepto en algunos virus, donde se encuentran formando cadenas dobles. La cadena simple de ARN puede plegarse y presentar regiones con bases apareadas, de este modo se forman estructuras secundarias del ARN, que tienen muchas veces importancia funcional, como por ejemplo en los ARNt (ARN de transferencia). Se conocen tres tipos principales de ARN y todos ellos participan de una u otra manera en la síntesis de las proteínas. Ellos son: El ARN mensajero (ARNm), el ARN ribosomal (ARNr) y el ARN de transferencia (ARNt). Función de los ARN: Un gen está compuesto, como hemos visto, por una secuencia lineal de nucleótidos en el ADN, dicha secuencia determina el orden de los aminoácidos en las proteínas. Sin embargo el ADN no proporciona directamente de inmediato la información para el ordenamiento de los aminoácidos y su polimerización, sino que lo hace a través de otras moléculas, los ARN. Todo el proceso que se lleva a cabo para la síntesis de proteínas se verá detalladamente en el Módulo Nº 3. 4. TEORÍA CELULAR “La célula es la unidad de vida más pequeña. Es la unidad anatómica y fisiológica de todos los seres vivos.” A continuación se hará un breve repaso de los contenidos vistos en el Módulo Nº 1, ítem 6 “Unidades de Medidas” y “Microscopia”. Alrededor de 1660, Antón van Leeuwenhoek construye un sistema de lentes que denomina microscopio. - 54 - En 1665 Robert Hooke utiliza un microscopio para observar que el corcho no era una sustancia homogénea, sino que estaba constituido por una serie de cavidades regularmente dispuestas a las que llamó Células cuyo significado es “habitaciones pequeñas”. El libro “Micrografía” (1665) de Robert Hooke contiene algunos de los primeros dibujos nítidos de células vegetales, basados en las observaciones de algunas secciones finas de “corcho” (corteza o cubierta exterior de cualquier planta leñosa). Pero Hooke nunca llegó a imaginar el verdadero significado de aquellas células; solamente había percibido su estructura, su esqueleto. Sin embargo el término célula se ha conservado en toda la literatura posterior. No sería hasta mediados del siglo XIX que dos científicos alemanes, Schleiden y Schwann, descubrirían la naturaleza celular de la materia viva. Cómo surge la Teoría Celular Mathias Jakob Schleiden (1804 – 1881), botánico alemán, rechazaba el vitalismo (posición según la cual los organismos vivos poseen una fuerza o sustancia vital especial, que no se puede encontrar en la materia inerte) en busca de una explicación mecanicista de la vida, y de carácter evolucionista. Su objetivo era hacer de la botánica una ciencia verdadera y exacta. En 1833, el botánico inglés R. Brown descubrió en diferentes células vegetales un “granito” (el núcleo). Schleiden se esforzó por demostrar que las células se forman de este núcleo; que del plasma viviente al principio, se separa el núcleo y que a su alrededor se forman células que van creciendo, hasta que sus paredes se tocan y por una especie de cristalización nace el tejido celular. Gran confusión originó la circunstancia de no establecer diferencia alguna entre el núcleo, las vacuolas y los granos de almidón, - 55 - creyendo que todos representaban células embrionarias. Schleiden, tampoco vio claro cómo estaban compuestas las plantas unicelulares. La doctrina de Schleiden de la evolución del tejido partiendo de células fue ampliada a los animales por Schwann, discípulo de Johannes Müller, destacado fisiólogo alemán. Theodor Schwann, (1810 – 1882), científico alemán nacido en el seno de una familia sumamente religiosa. Estudió medicina en Bonn, donde conoció a Müller, y en Berlín ocupó la cátedra de Anatomía. A partir de 1839 su carrera científica culmina debido, en principio, a la actitud crítica de científicos dedicados a la química frente a sus trabajos sobre la fermentación alcohólica. Ante tal circunstancia resurge fe religiosa, abandonada debido al racionalismo de los años anteriores, y se refugia en el Dios de su infancia. Schwann y Schleiden eran grandes amigos, y el mismo Schwann cuenta cómo una conversación con Schleiden, en Berlín, le sugirió la idea que daría origen a la teoría celular: “Un día que cenaba con el señor Schleiden, este ilustre botánico me indicó la importante función que desempeña el núcleo en el desarrollo de las células vegetales. Me acordé enseguida de haber visto un órgano semejante en las células de la cuerda dorsal del renacuajo, y en aquel momento comprendí la importancia que tendría mi descubrimiento si llegaba a demostrar que en las células de la cuerda dorsal este núcleo desempeñaba el mismo papel que el núcleo de las plantas en el desarrollo de los vegetales”. Esto ocurría en 1838, año en que Schleiden había publicado una breve memoria en la que se describía el desarrollo del bolso embrionario de diversas plantas y en la que se explicaba la independencia de las células que componen el organismo y la función directora del núcleo. A raíz de esta observación, Schwann se dedicó a descubrir la composición celular de los tejidos animales y a localizar los núcleos de las diferentes células. Al año siguiente, Schwann publicó una memoria en la que se exponían todas las bases de la teoría celular. Dibujos de células animales hechos por Schawnn (1839) - 56 - La teoría celular de Schwann exponía dos cosas: 1) El reconocimiento de que el organismo compuesto se desarrolla de células; 2) Una nueva filosofía inductiva, genética y mecánica. Tanto Schleiden como Schwann afirmaban que el organismo era un agregado (según ciertas leyes) de otros seres de orden inferior; contra la opinión vitalista de la unidad de la vida en el cuerpo orgánico y contra la fuerza vital unitaria. Schleiden aducía que la vida es el resultado de la colaboración de muchas células. Schleiden, botánico, y Schwann, zoólogo, estudiaron muchos tipos de tejidos en sus campos respectivos. Ambos llegaron a la conclusión de que la célula es la unidad estructural básica de todos los organismos. La célula constituye la unidad fundamental de los seres vivos. Todo organismo vivo está constituido por una o por multitud de células. Este es el enunciado básico de la teoría celular. La Teoría Celular, tal como se la considera hoy, puede resumirse en los siguientes postulados: 1. En principio, todos los organismos vivos están compuestos por una o más células. 2. En las células tienen lugar las reacciones químicas de un organismo vivo, incluidos los procesos que liberan energía y las reacciones biosintéticas. 3. Las células provienen tan solo de otras células preexistentes. 4. Las células contienen la información hereditaria de los organismos de los cuales son parte, y esta información pasa de célula progenitoras a células hijas. La segunda y tercera proposiciones fueron añadidas por el patólogo y también estadista Rudolf Virchow (1821 – 1902). En su trabajo “Patología celular” (1858), Virchow consideró la célula como la unidad básica metabólica y estructural. En ese mismo trabajo subrayó la continuidad de los organismos: “todas las células provienen de otras células (preexistentes)”. Por lo tanto, las células no surgen por generación espontánea a partir de materia inanimada. - 57 - Impacto que causó la Teoría Celular en la Biología del desarrollo Una de las numerosas contribuciones unificadoras de la Teoría Celular consistió en dar un nuevo significado a los términos óvulos y semen, que hasta entonces habían sido conceptos bastante confusos. Resulta curioso que hasta 1880, aproximadamente, siguiera existiendo mucha incertidumbre acerca del significado de la fecundación. Para los fisicismo (el fisicismo era la insistencia y creencia en ciertos principios dominantes en la física clásica), la fecundación no era más que el impulso o señal que iniciaba la segmentación de la célula huevo. Así interpretaba la fecundación Miescher, el descubridor del ADN en el año 1874. Con el tiempo, los citólogos como O. Hertwig y van Beneden demostraron que el espermatozoide aporta al huevo mucho más que una simple orden de iniciar la primera segmentación; también aporta el núcleo de la célula germinal (gameto) masculina. Este núcleo con su dotación haploide de cromosomas penetra en el óvulo y se suma a la dotación haploide del mismo formando el núcleo diploide del zigoto. Así, la fecundación restaura la diploidía y combina en la descendencia genes del padre y de la madre. (Mayr, 1998) LÍNEA DE TIEMPO: ALGUNOS EVENTOS EN BIOLOGÍA CELULAR 1639 1680 Robert Hooke observa “células” de corcho con un microscopio primitivo A.Leeuwenhoek (1632-1702) descubre espermatozoides en el semen 1688 Redi publica su trabajo sobre la generación espontánea 1839 Johannes Müller efectúa investigaciones microscópicas e histológicas 1839 Jacob Henle realiza una descripción general de la epidermis y el epitelio 1839 1839 Schleiden y Schwann proponen la Teoría Celular Robert Remak (1815-1865) descubrimiento de las células ganglionares del corazón humano - 58 - 1841 1852 1855 Albert Koelliker (1817-1905) descubre que cada espermatozoide es una célula, la célula germinal masculina Robert Remak demostró que el óvulo es una célula Rudolf Virchow afirma que todas las células provienen de células 1873 Camillo Golgi Da a conocer su procedimiento de tinción de las células nerviosas 1888 Santiago Ramón y Cajal Modifica el método de tinción de Golgi y logra esclarecer todas las estructuras del sistema nervioso 5. ESTRUCTURA CELULAR Todas las células están cubiertas por una membrana externa, llamada membrana plasmática, que las separa de otras células y del medio circundante con el cual intercambian materia y energía. Este intercambio esta altamente regulado y es selectivo. De esta forma la membrana plasmática debe actuar no sólo como limite celular sino también como barrera selectiva. Por lo tanto la célula, mantiene una composición química muy ordenada y diferente a la del entorno. (Como se verá en el capítulo de Transporte a través de la membrana.) Todas las células poseen un metabolismo o conjunto de reacciones químicas, que posibilitan el mantenimiento de la vida. Este metabolismo para sustentarse necesita de una o más fuentes de energía. Las células, necesitan de distintivos tipos de moléculas energéticas: Monedas energéticas, como el ATP Moléculas combustibles, como la glucosa o los ácidos grasos Moléculas de reserva de energía, como el glucógeno o el almidón Dentro de las reacciones para obtener y transformar diferentes forma de energía, son muy importantes las reacciones de oxido-reducción o reacciones REDOX. En este tipo de reacciones es esencial la participación de las coenzimas de oxido-reducción, como el NAD+ y el FAD. Todas las células, almacenan en forma de ADN, ácido desoxirribonucleico, a información necesaria para controlar sus actividades (reproducción, metabolismo), y para establecer su propia estructura. - 59 - Las células para perpetuarse necesitan reproducirse. Esto significa que la información almacenada en el ADN debe duplicarse para poder ser transmitida a las células hijas. El ADN tiene la excepcional característica de ser una molécula capaz de autorreplicarse, es decir de generar una copia de si misma. Este proceso es llamado duplicación o replicación. (Se verá en el Módulo Nº 3). Células Eucariotas y Procariotas Características Principales: Todas las células se parecen y responden a un patrón común por más diversas que sean. Las células de organismos pluricelulares son diferentes en su función, por ser distintas estructuralmente, pero todas concuerdan con un patrón común. Por ejemplo, aquellas especializadas en la síntesis de lípidos, tendrán mayor desarrollo del retículo endoplasmático liso y serán distintas de las neuronas especializadas en la transmisión del impulso nervioso, cuya especialización es tan grande que pierden su capacidad de reproducirse. A pesar de las semejanzas y diferencias entre las células y que todas cumplen con los postulados de la Teoría Celular, se distinguen dos grandes tipos de células: Procariotas (sin núcleo verdadero) y Eucariotas (con núcleo). Principales características comunes entre células eucariotas y procariotas 1- En ambos tipos celulares el ADN es el material genético. 2- Ambos tipos celulares poseen membranas plasmáticas como límite celular. 3- Poseen ribosomas para la síntesis proteica. 4- Poseen un metabolismo básico similar 5- Ambos tipos celulares son muy diversos en formas y estructuras. Los eucariontes son organismos cuyas células poseen un sistema de endomembranas (membranas internas) muy desarrollado. Estas membranas internas forman y delimitan organelas donde se llevan a cabo numerosos procesos celulares. De hecho él más sobresaliente de estos organelos es el núcleo, donde se localiza el ADN lineal. Justamente, el término eucarionte, significa núcleo verdadero (del griego eu significa “buen” y karyon, “núcleo” o “centro”). Las células eucariontes poseen diversos compartimentos internos, rodeados por membranas. De esta forma es más eficiente reunir a los sustratos y sus enzimas, en una pequeña parte del volumen celular total. Además de conseguirse una mayor velocidad, las membranas favorecen la aparición de estructuras reguladoras que orientan el flujo de moléculas y su posterior - 60 - conversión en otros productos. Ciertos procesos como la fotosíntesis y la cadena respiratoria están altamente organizados gracias a la localización de las enzimas en diferentes estructuras de membrana. Por otra parte, las membranas también impiden la aparición de sustratos en forma inespecífica en distintas regiones de la célula, ya que actúan como barrera selectiva. En cuanto al tamaño, podemos decir que en promedio una célula eucarionte es diez veces mayor que una célula procarionte. En cuanto al material genético, podemos decir que el ADN lineal eucariota posee una organización mucho más compleja que el ADN procarionte. Las células procariontes carecen de núcleo y generalmente son mucho menores que las células eucariontes. El ADN circular de las células procariontes no está rodeado por una membrana, pero puede estar limitado a determinadas regiones denominadas nucleoides. Las células procariontes, al igual que las células eucariontes, poseen una membrana plasmática, pero carecen de membranas internas, que formen organelas. Sin embargo, debemos precisar que en algunas células procariontes, la membrana plasmática forma laminillas fotosintéticas. Las células procariontes poseen una característica única, una pared de peptidoglicanos, un gran polímero de glúcidos y aminoácidos. Características Diferenciales entre el Modelo Celular Procariótico y Eucariótico Característica Núcleo Cromosomas ADN ADN extracromosómico Organelas citoplasmáticas Membrana plasmática Célula Procariótica No posee membrana nuclear Posee membrana nuclear Un único cromosoma Posee uno o más cromosomas circular y desnudo lineales unidos a proteínas (cromatina) Circular y grande. Proteínas Lineal y unido a proteínas asociadas débilmente Puede estar presente como plásmidos Presente en organelas No posee Mitocondrias y cloroplastos, (los cloroplastos presentes sólo en células vegetales) Contiene las enzimas de la Semipermeable, sin las cadena respiratoria, también funciones de la membrana puede poseer los pigmentos procariótica fotosintéticos Sistema de endomembranas No posee Pared celular Esteroles Célula Eucariótica Presenta REG, REL, Golgi, lisosomas, vacuolas y vesículas. Capa rígida de peptidoglucano (excepto micoplasmas) No poseen pared de peptidoglucano. Pueden poseer una pared de celulosa o quitina Ausentes (excepto micoplasmas) Generalmente presentes - 61 - Citoesqueleto Exocitosis y Endocitosis Ribosomas División Tamaño Ausente Presente. Formado por filamentos proteicos. Ausente 70 S en el citoplasma Presente 80 S en el retículo endoplásmico y en el citosol Fisión Binaria (amitosis) 0,2 a 10 mm Mitosis - Meiosis Siempre superior a 6 mm 1) Estructura de la célula procariota: Bacterias, Micoplasmas y Cianobacterias (algas azules-verdosas) Figura: Esquema de Escherichia coli2, un procarionte heterótrofo muy estudiado. 2 Escherichia coli: especie de bacteria habitante del colon humano y de otros animales. Utilizada como modelo biológico en investigaciones biomédicas. - 62 - En el siguiente cuadro, se muestran los componentes de celulares de los procariontes: Las bacterias son organismos unicelulares procariontes que se reproducen por fisión binaria. Contienen toda su información genética en un único cromosoma bacteriano circular. También poseen sistemas productores de energía y biosintéticos necesarios para el crecimiento y la reproducción. Poseen como característica particular una pared rígida de peptidoglicanos. Son generalmente de vida libre y poseen ADN extracromosómico en forma de plásmidos, estos codifican genes de resistencia a antibióticos o factores "sexuales" como los pili. Los micoplasmas son las bacterias más pequeñas de vida independiente. Son muy flexibles por lo que atraviesan los filtros de esterilización. Entre sus características principales se encuentran: a) carecen de pared celular, b) en su membrana plasmática poseen esteroles, que no son sintetizados por la bacteria sino que son absorbidos del medio de cultivo o del tejido donde se desarrolla. Los micoplasmas son resistentes a la penicilina (carecen de pared de peptidoglucano) y por la misma razón no toman la coloración de Gram. Las cianobacterias, son bacterias Gramnegativas. Se encuentran presentes en estanques, lagos, suelo húmedo, cortezas de árboles, océanos y algunas en fuentes termales. La mayor parte de las cianobacterias son autótrofos fotosintéticos. Contienen clorofila a, que también se encuentra en plantas y algas. La clorofila a y pigmentos accesorios se localizan en membranas fotosintéticas, llamadas laminas internas o laminillas fotosintéticas. Muchas especies de cianobacterias fijan nitrógeno, este proceso enriquece el suelo. Características de la pared celular en Bacterias Grampositivas y Gramnegativas: Por fuera de la membrana celular, se encuentra una pared celular rígida de peptidoglicano, que esta presente en todas las bacterias excepto los micoplasmas. La presencia de la pared protege a la bacteria de la diferencia de presión osmótica entre el medio interno de la bacteria y el medio exterior. De no existir la pared la bacteria estallaría. Además la pared cumple funciones de protección como por ejemplo contra sustancias tóxicas. - 63 - Existen dos tipos de pared bacteriana que pueden diferenciarse por la Tinción de Gram (siglo XIX). El primer grupo de bacterias son aquellas capaces de retener el colorante cristal violeta luego de la decoloración con alcohol-cetona. Estas bacterias son llamadas Grampositivas. El segundo grupo esta conformado por aquellas bacterias incapaces de retener el colorante luego del tratamiento decolorante, por lo tanto son llamadas Gramnegativas. 2) Estructura de las células eucarióticas: Figura: Esquema de un organismo eucarionte unicelular y fotosintético: Chlamydomonas - 64 - En el siguiente cuadro, se muestran los componentes celulares de los eucariontes: “Presentan este modelo celular, los organismos de los reinos Protista, Hongos, Plantas y Animales”. Si bien existe una gran diversidad entre estas células, el modelo básico es similar, presentando como estructura sobresaliente el núcleo celular. Figura: Esquema de la ultraestructura tridimencional de una célula animal y sus principales componentes - 65 - 3) Célula eucariótica animal y vegetal Figura: Esquemas de una célula vegetal (izquierda) y tridimencional de un cloroplasto con sus componentes (derecha) Cuadro: Modelos básicos de células Eucariotas Las células eucariontes poseen dos modelos estructurales básicos: Las células autótrofas fotosintéticas, son aquellas que sintetizan su propio alimento, es decir sus propias moléculas combustibles. En este caso las células eucariontes vegetales son células autótrofas fotosintéticas, por lo tanto utilizan la luz solar como fuente de energía. Transforman la energía solar en energía - 66 - química, por el proceso de fotosíntesis. La fotosíntesis en las células vegetales se lleva a cabo en una organela membranosa llamada cloroplasto. Dentro del cloroplasto se encuentran sacos membranosos apilados, denominados tilacoides, en cuyas membranas encontramos el pigmento llamado clorofila, esencial para la fotosíntesis. (Se verá en detalle en el capítulo de fotosíntesis) Las células heterótrofas son aquellas que no sintetizan su propio alimento sino que necesitan una fuente externa de energía tanto como de materiales de construcción de sus propias moléculas. Las células animales (y los hongos), son células eucariontes heterótrofas. Las células animales y las células vegetales poseen unas organelas membranosas llamadas mitocondrias, donde se lleva acabo la respiración celular. En este proceso son rotos los enlaces de alta energía de las moléculas combustibles orgánicas. Esta energía liberada es utilizada para la síntesis de las monedas energéticas como el ATP. El ATP es esencial para las diferentes funciones celulares. Para que este proceso se lleve a cabo dentro de las mitocondrias es necesaria la presencia de oxígeno. Por lo tanto en ambos tipos celulares son necesarias las mitocondrias, para obtener energía química en forma de ATP a partir de las moléculas combustibles. Pero es diferente el origen de las moléculas orgánicas utilizadas como combustibles. En el caso de las células vegetales (autótrofas), ellas sintetizan sus propias moléculas combustibles en los cloroplastos, en el proceso de fotosíntesis. En cambio las células animales (heterótrofas), necesitan una fuente externa de moléculas energéticas que sirvan como combustible celular. Principales diferencias entre células animales y células vegetales Estructura Pared celular Célula animal Ausente Célula vegetal Pared celular por celulosa. constituida Aparato mitótico Astral Anastral Centríolos Presente Ausente Vacuolas Vacuolas pequeñas (Huso acromático) Vacuolas grandes, puede ser una grande central Metabolismo Heterótrofo Autótrofo Mitocondrias Presentes Presentes Cloroplastos Ausentes Presentes - 67 - Figura: Esquema de la ultraestructura de una célula animal idealizada - 68 - Figura: Esquema de la ultraestructura de una célula vegetal idealizada que muestra sus principales características: la pared celular, los plasmodesmos, los cloroplastos y las vacuolas. 6. COMPONENTES CELULARES Es este capítulo, veremos más detalladamente los componentes celulares que fueron presentados en los capítulos anteriores: membrana plasmática, pared celular, citoplasma, núcleo, sistemas de endomembranas, organelas, citoesqueleto. - 69 - Membrana plasmática: Como ya se ha visto, las células están separadas del medio que las rodea por una delgada lámina denominada membranas plasmática, que define los límites de las mismas. Como todas las membranas biológicas, es una estructura dinámica y fluida formada por fosfolípidos y proteínas. No solo define los límites de la célula sino, como se verá en el siguiente capítulo, permite que ésta exista como una entidad diferente de su entorno, regulando el tránsito de sustancias hacia fuera y hacia adentro de ella. La membrana generalmente está rodeada por un medio acuoso y en consecuencia, las moléculas de fosfolípidos se disponen en una bicapa, con sus colas hidrófobas apuntando hacia el interior y sus cabezas hidrófilas de fosfato apuntando hacia el exterior. Por su tamaño, no pueden visualizarse con el microscopio óptico pero si con el microscopio electrónico se identifica como una doble línea delgada y continua. Figura: Microfotografía electrónica de transmisión de una membrana plasmática. Se pueden ver tres líneas paralelas, dos líneas densas separadas por una capa intermedia clara. Este aspecto conocido como unidad de membrana no es reflejo de una estructura trilaminar a nivel molecular, sino es la expresión de como el osmio, usado como "colorante" se une a la membrana. - 70 - El modelo estructural más aceptado es el llamado modelo de mosaico fluido. Figura: Esquema del modelo de mosaico fluido de las membranas Aunque los lípidos y las proteínas, unidos a proteínas periféricas o a proteínas concentradas en el interior, parecen estar ancladas en una posición fija en la membrana, la estructura de la bicapa es fluída, de forma que la estructura en “mosaico” de fosfolípidos y proteínas cambia en el tiempo. Las proteínas incluidas en la bicapa, denominadas proteínas integrales de membranas, tienen una orientación espacial definida. Existe otro grupo de proteínas denominadas proteínas periféricas, que no atraviesan la bicapa lipídica sino que se unen a una u otra cara de la membrana mediante interacciones con otras proteínas. En las células eucariontes, todas las membranas, incluidas las organelas, tienen la misma estructura. Sin embargo la composición química de lípidos, proteínas y glúcidos difiere en las membranas de los distintos tipos celulares, de acuerdo a las funciones celulares y/o de las organelas. Esquema de un fosfolípido Corte esquemático de una vesícula de fosfolípido - 71 - Matriz extracelular: Los organismos multicelulares están compuestos no solo por células sino también por elementos intercelulares que se agrupan el la denominada matriz extracelular. La matriz celular es el ambiente especial que rodea a la célula. Está formada por un conjunto de materiales ubicados en el espacio extracelular, que puede a veces ser muy pequeño, y que le brinda a las células un ambiente particular. La matriz está formada por proteínas fibrosas y por la sustancia fundamental, constituida principalmente por proteínas y azúcares asociados en diferentes combinaciones, que cumplen funciones estructurales además de participar en la adhesión entre las células y la matriz. La matriz tiene un papel fundamental en el desarrollo de los tejidos y órganos controlando la diferenciación celular, la morfogénesis, la migración celular y su metabolismo. Otras funciones importantes de la matriz extracelular: - Rellenar los espacios no ocupados por las células - Conferir a los tejidos resistencia a la compresión y estiramiento - Constituir el medio por donde llegan los nutrientes y se eliminan los desechos celulares - Proveer a diversas clases de células de puntos fijo donde aferrarse - Ser el vehículo por donde migren las células cuando se desplazan de un punto a otro del organismo - Ser un medio por el arriban a las células sustancias inductoras (señales) provenientes de otras células. Los tejidos, y por extensión los órganos y sistemas, son el resultado de asociaciones de diferentes tipos de células y matrices extracelulares. Así, el tejido conectivo contiene las células que se encuentran dispersas en medio de una abundante matriz extracelular. Existen varios tipos de tejidos conectivos. En la lámina siguiente un ejemplo de tejido conectivo laxo: - 72 - Pared celular: Como se vio, una diferencia fundamental entre células animales y vegetales es que sólo las células vegetales están rodeadas por una pared celular. Se puede decir que es como un exoesqueleto de las células vegetales, que le confieren protección y sostén mecánico. Esta pared realiza muchas de las funciones que cumple la - 73 - matriz extracelular en las células animales, aunque la composición y la estructura son diferentes a la de la matriz extracelular. La pared celular si bien es rígida, no es estática. Así interviene en la diferenciación de las células vegetales durante el crecimiento. La pared celular mediante plasmodesmos3, conecta a las células vegetales entre sí formando tejidos, transmite señales que influyen en el crecimiento y la división celular y también en la morfogénesis. La pared celular es un filtro un poco más impermeable que la matriz extracelular de las células animales, sin embargo su estructura porosa deja pasar materiales solubles que toman contacto con la membrana plasmática. La pared celular está formada por las siguientes capas, desde el exterior hacia el interior: - Laminilla media: Sólo presente en plantas multicelulares, formada por compuestos pécticos. - Pared primaria: Formada por celulosa, hemicelulosa y compuestos pécticos. Se encuentra en todas las células vegetales. - Pared secundaria: Compuesta sólo por celulosa o celulosa y hemicelulosa. Es una pares suplementaria que puede estar ausente y su función es mecánica. ● Núcleo: El núcleo es la estructura típica de la célula eucarionte. En el núcleo de las células eucariontes se encuentran las moléculas que contienen la información hereditaria, moléculas de ADN. En su interior se también se sintetiza el ARN y se producen procesos claves relacionados con la regulación de la expresión genética. Es una estructura generalmente esférica y voluminosa comparada con el tamaño total de la célula. Está rodeada por una envoltura nuclear, formada por dos membranas, cada una de las cuales es una bicapa lipídica. A intervalos frecuentes, las membranas presentan poros nucleares por donde circulan materiales entre el núcleo y el citoplasma. En el interior del núcleo, el ADN se asocia a proteínas básicas llamadas histonas, formando una estructura fibrilar muy enrollada denominada cromatina y el nucleolo, sitio de ensamblaje de los ribosomas (estructuras esenciales para la síntesis de proteínas, formados por ARN ribosomal y proteína). El ARN ribosómico se sintetiza en el nucleolo, y las proteínas ribosómicas en el citoplasma, para pasar después al núcleo y de allí al nucleolo, donde se unen al ARN ribosomal para formar los ribosomas. El núcleo desempeña dos funciones fundamentales para la célula: En primer lugar, como vimos, lleva la información hereditaria que determina si un tipo particular de célula se desarrollará en un paramecio, un roble o un ser humano; y será semejante a sus progenitores. En segundo lugar, el núcleo ejerce una influencia continua sobre las actividades de las células, de esta forma asegura que las moléculas complejas que ellas requieren se sinteticen en la cantidad y tipo necesario. 3 Plasmodesmo: Puentes citoplasmáticos entre células vegetales adyacentes. - 74 - El siguiente recuadro, resume estructura y función de los componentes del núcleo: Características del Núcleo Celular y sus Componentes Estructura : Núcleo Función Descripción Celular Núcleo Envoltura Nuclear Estructura rodeada por una doble membrana con poros. Contiene cromatina/cromosomas y nucleolo. Estructura formada por dos unidades de membrana unidas a nivel de los poros nucleares. Regular la función celular. Control del metabolismo, reproducción (ciclo celular) y diferenciación celular. Continuación del REG. Posee poros que regulan el pasaje entre núcleo y citoplasma Nucleolo Cuerpo granular en el núcleo, Sitio de síntesis del RNA que consiste en ARN y ribosómico y de ensamble de los ribosomas. proteínas. Cromatina ADN asociado a proteínas, Empaquetamiento tanto estructurales (histonas) (plegamiento) de ADN. El como a proteínas regulatorias. ADN compone los genes. La cromatina es visible durante Funciones regulatorias de la transcripción genética. la interfase celular Cromosomas ADN asociado a proteínas, en estado superenrrollado. Visible en forma de estructuras cilíndricas cuando la célula se divide, ya sea en mitosis o meiosis. - 75 - Contienen los genes que son las unidades de información, que rigen las funciones y estructura celular. Citoplasma: Rodeando al núcleo encontramos el citoplasma, también llamado citosol o matriz citoplasmática. Es un coloide donde predominan como constituyentes agua, iones, enzimas y donde se encuentran incluidas las organelas celulares. El citoplasma se encuentra separado del ambiente exterior por la membrana plasmática y está altamente organizado. En hepatocitos y células musculares es común la presencia de gránulos de glucógeno. En las células denominadas adipositos, el citosol está ocupado por una gran gota de grasa. Se puede subdividir esquemáticamente al citoplasma en dos espacios: - Uno correspondiente al citosol, verdadero medio interno celular que se extiende desde la envoltura nuclear hasta la membrana plasmática y que llena el espacio no ocupado por el sistema de endomembranas, mitocondrias y peroxisomas. - Otro espacio encerrado en el interior de las organelas. En el citoplasma se encuentran unas estructuras, los ribosomas, que cumplen una función muy importante, puesto que en ellos se realiza la Síntesis de proteínas. Cuanto más proteínas esté sintetizando una célula más ribosomas tendrá. Si bien son organelas celulares, no poseen membranas (como las mitocondrias, peroxisomas y plásticos) y están constituidos por dos subunidades, cada una formada por ARN ribosómico y proteínas. Estas dos subunidades, denominadas mayor y menor, generalmente están separadas y se ensamblan en el nucleolo, en el momento de la síntesis de proteínas. Cuando hay varios ribosomas unidos a una molécula de ARN mensajero, se lo denomina polirribosoma. (Ver capítulo: Compuestos orgánicos) Sistemas de endomembranas: Como vimos, las células eucariontes poseen estructuras internas que las dividen en compartimentos especializados; estos compartimientos están limitados por membranas cerradas que tienen una permeabilidad selectiva. Cada compartimiento tiene una función diferente y contiene un grupo característico de enzimas que realizan las funciones propias de cada organela. Asimismo los distintos compartimientos si bien están separados físicamente, se encuentran interconectados funcionalmente. Dentro del sistema de endomembranas se distinguen los siguientes elementos: a) Retículo endoplasmático granular o rugoso (REG o RER). Es un grupo de cisternas aplanadas que se conectan entre sí mediante túbulos. Presente en todos los tipos celulares, se halla especialmente desarrollado en las células secretoras de proteínas. El REG ofrece una cara citosólica tachonada de ribosomas, a los que debe su aspecto rugoso. Los ribosomas se unen a las membranas del REG por su subunidad mayor, mediante receptores específicos, las proteínas integrales de las membranas cisternales conocidas como riboforinas. - 76 - b) Retículo endoplasmático agranular o liso (REA o REL). Su aspecto es más tubular y carece de ribosomas. Es poco conspicuo en la mayoría de las células, pero alcanza un notable desarrollo en las células secretoras de hormonas esteroides. c) Aparato o complejo de Golgi. Constituido por sacos discoidales apilados, como mínimo en número de tres, rodeados por pequeñas vesículas. Cada saco presenta una cara convexa y otra cóncava, esta última orientada hacia la superficie celular. En las células animales se ubica típicamente entre el núcleo y el polo secretor de la célula, en tanto en las células vegetales aparece fragmentado en varios complejos denominados dictiosomas o golgiosomas. d) Vacuolas y vesículas. Las vesículas son organelas con forma de bolsas formadas por membranas. Sus principales funciones son el almacenamiento y transporte de materiales, tanto dentro de la célula como hacia el interior y exterior. Las vacuolas son un tipo especial de vesículas presentes en las células vegetales y hongos, estas vacuolas son de gran tamaño y están llenas de fluído. e) Lisosomas: Son un tipo especial de vesículas formadas en el complejo de Golgi, presente en las células animales. Contienen enzimas hidrolíticas capaces de degradar macromoléculas como proteínas, polisacáridos, ácidos nucleicos y lípidos. Ejemplo: los glóbulos blancos con lisosomas encargados de capturar bacterias en el cuerpo humano. - 77 - La vía de tránsito intracelular implica un transporte desde el RE hasta el aparato de Golgi; a partir de éste hay dos caminos posibles: hacia las vesículas de secreción y desde allí a la membrana plasmática, o bien hacia los lisosomas: Otros tipos de organelas: Los peroxisomas son un tipo de vesícula grande presente en la mayoría de las células eucariotas. Poseen una sola membrana con enzimas que degradan los ácidos grasos, liberando energía en forma de calor y otras sustancias. Los peroxisomas son abundantes en las células hepáticas, donde participan de la degradación de etanol. Junto con las mitocondrias, constituyen los principales sitios de utilización del oxígeno dentro de la célula. - 78 - También en las plantas se encuentran los peroxisomas. Una forma interviene en la fotorrespiración y otra denominada glioxisoma se encuentra presente en las semillas en germinación. Las mitocondrias, son las organelas en las que se degradan moléculas orgánicas y se libera energía química en el proceso que se verá en el capítulo de respiración celular. En general cuanto mayor es el requerimiento de energía de una célula eucariota, más mitocondrias contiene esa célula. Las mitocondrias están rodeadas de dos membranas, una interna plegada hacia dentro formando crestas y otra externa lisa. Por su importante función en la respiración celular, se verá en mayor detalle en el capítulo de Respiración y Fotosíntesis. Los plástidos, son organelas que se encuentran sólo en las células de plantas y algas. Están rodeadas por dos membranas y hay diferentes tipos: Los leucoplastos, pueden almacenar almidón (o a veces proteínas y aceites) y son numerosos en por ejemplo raíces o tubérculos. Los cromoplastos, contienen pigmentos del tipo de los carotenoides. Los cloroplastos, que contienen clorofila y en ellos se producen los hidratos de carbono por el proceso de fotosíntesis. En el siguiente cuadro se hace un resumen de las características principales de las estructuras y organelas que forman los sistemas de endomembranas: Organización del Sistema de endomembranas Estructura Descripción Función Retículo endoplasmático Membranas internas en Síntesis de Proteínas rugoso (REG) forma de sacos aplanados y destinadas a secreción túbulos. Con ribosomas (exportación) o a la adheridos a su superficie incorporación de membranas. externa. La envoltura nuclear es parte del REG. Retículo endoplasmático Membranas internas donde Sitio de biosíntesis de lípidos liso (REL) predominan los túbulos. Sin y detoxificación de ribosomas adheridos. medicamentos. Aparato de Golgi Pilas de sacos membranosos Modificación de proteínas aplanados (dictiosomas). (glicosilación). Funcional y estructuralmente Empaquetamiento de polarizado. proteínas secretadas. Clasificación de las proteínas que se distribuyen a membrana plasmática, secreción o lisosomas. Lisosomas Vesículas (sacos) Contienen enzimas membranosas hidrolíticas, que desdoblan materiales ingeridos, secreciones y deshechos celulares. - 79 - Vacuolas Sacos membranosos principalmente, en plantas, hongos y algas. Transporte de materiales, desechos y agua. Principales organelas membranosas de la célula eucarionte Descripción Función Estructura Mitocondria Cloroplasto Peroxisomas Organelas semiautónomas. Metabolismo aeróbico. Sitio Poseen ADN y ribosomas tipo de muchas de las reacciones procarionte. Una doble de la respiración celular. membrana les sirve de envoltura. Allí se realizan el ciclo de La membrana interna forma las Krebs, la cadena respiratoria crestas mitocondriales. y la fosforilación oxidativa. Es decir la transformación de la energía de lípidos o glucosa (moléculas combustibles) en ATP (moneda energética). Organela semiautónoma. Posee La clorofila capta la energía ADN y ribosomas tipo luminosa para formar ATP y procarionte. Una doble otros compuestos con gran membrana envuelve a los cantidad de energía. Estos tilacoides. La clorofila, se compuestos altamente encuentra en las membranas energéticos sirven para tilacoidales. sintetizar, glucosa a partir de CO2. Vesículas membranosas que Sitio de muchas reacciones contienen diversas enzimas metabólicas. Enzimas que relacionadas con el metabolismo protegen de la toxicidad del del oxigeno y el peróxido de oxigeno, por ejemplo la hidrogeno. No poseen ADN ni catalasa. ribosomas Citoesqueleto: El citoesqueleto es un sistema de andamiaje interno. No sólo mantienen la organización celular y sus organelas sino que además le permite moverse, participar en las modificaciones de su morfología y dirige el tránsito intracelular. El citoesqueleto está constituido principalmente por tres integrantes fundamentales: microtúbulos, filamentos de actina (o microfilamentos) y filamentos intermedios. - 80 - También forman parte del citoesqueleto, los cilios y flagelos. Son estructuras largas y delgadas, presentes en muchos tipos de células eucariotas. Cuando son numerosos y cortos se llaman cilios y cuando son largos y escasos se llaman flagelos. Están asociados con el movimiento del organismo. La gran movilidad de los espermatozoides humanos se debe a su poderos flagelo único. Las células que se encuentran en el tracto respiratorio de nuestro cuerpo son ciliadas, ellas baten los cilios y crean una corriente ascendente de mucus que arrastra hacia la garganta los restos de polvo, polen, hollín alquitrán de tabaco y cualquier sustancia extraña inhalada. Muchos tipos de células eucariotas tienen en su citoplasma centríolos, generalmente se encuentran de a pares y en la región cercana a la envoltura nuclear, denominada centrosoma. El centrosoma es el principal centro organizador de microtúbulos y es importante en la formación del huso mitótico (Se verá en el capítulo de reproducción celular). Organización General del citoesqueleto Estructura Descripción Función Sostén estructural, participan en el Tubos huecos compuestos movimiento de organelas por la forma monomérica de Microtúbulos y la división celular la proteína tubulina. (aparato mitótico), (monómero globular) componentes de cilios, flagelos y centríolos. Sostén estructural, Estructura sólida en forma de participan en el Filamentos de actina huso consistente en la movimiento de la célula y (microfilamentos) proteína actina. (monómero sus organelos y en la globular) división celular. Sostén estructural. Forman Proteínas filamentosas, en redes que conectan la Filamentos intermedios forma de tubos. Compuestas membrana plasmática con por monómeros fibrosos. la envoltura nuclear. El huso mitótico se forma entre los centríolos Pares de cilindros huecos, durante la división de localizados cerca del centro Centríolos células animales, fija y de la célula, formados por organiza los microtúbulos. microtúbulos. Están ausentes en las plantas superiores. Proyecciones relativamente Movimiento de algunos cortas que se extienden desde organismos unicelulares. Cilios la superficie celular. Se utiliza para mover Compuestas por materiales en la superficie microtúbulos. de algunos tejidos. Proyecciones largas Locomoción celular de compuestas por microtúbulos. espermatozoides y Flagelos Cubiertos por membrana algunos organismos plasmática unicelulares. - 81 - Figura: Esquema de componentes del citoesqueleto 7. DIFUSIÓN Y ÓSMOSIS Como se vio en el ítem 6 “Componentes celulares”, las células están separadas del medio que las rodea por una delgada lámina denominada membrana plasmática, cuya función es crear una barrera selectiva para regular el intercambio de materiales entre la célula y el medio que la rodea. Sin esta membrana, la célula estaría expuesta a los rigores del mundo externo. ¿Por qué la glucosa disuelta en la sangre puede atravesar la membrana celular e ingresar en el citoplasma? ¿Cómo salen de la célula los productos de desecho, como urea? Para responder estas preguntas, a continuación se verán los diferentes mecanismos de transporte a través de la membrana celular. Transporte a través de la membrana celular A. Difusión: Es el movimiento de moléculas desde una zona de mayor concentración hacia una de menor concentración. La diferencia de concentración que existe entre una zona y otra se denomina gradiente. Ejemplo: Si colocamos un soluto en un solvente, las moléculas de soluto van a difundir desde la zona donde está más concentrada hacia la zona sonde se encuentra en menor concentración. Después de un tiempo toda la solución va a presentar la misma concentración de soluto. Esto ocurre cuando agregamos una gota de tinta dentro de un - 82 - vaso con agua, la tinta difunde a través del líquido y después todo el vaso presenta una coloración pareja. Difusión de una sustancia disuelta en un solvente. Este mecanismo no necesita aporte de energía externa, es suficiente con la energía cinética propia de las moléculas. a) Difusión Simple: Es el mecanismo por el cual la difusión se realiza entre compartimentos separados por una membrana permeable al soluto y sin requerir energía externa. Es un fenómeno espontáneo y ocurre con las moléculas no polares y de pequeño tamaño, las liposolubles y las polares pero sin carga eléctrica neta, cuando pasan a través de la membrana plasmática. El paso del agua a través de la membrana ocurre por difusión simple pero se lo denomina ósmosis. Siempre se da en forma espontánea y muy rápidamente. El agua (H2O) difunde desde el compartimiento de menor concentración de solutos o medio hipotónico, al de mayor concentración de solutos o medio hipertónico, de forma tal de igualar las concentraciones en ambos compartimientos. Después de un tiempo el resultado será dos medios isotónicos, es decir que la concentración a ambos lados de la membrana será la misma. Proceso osmótico sobre una célula viva. Ejemplo: Si colocamos un glóbulo rojo en una solución hipertónica (p ej. agua con sal), el H2O tenderá a salir por ósmosis hacia el medio extracelular, la célula (glóbulo rojo) - 83 - sufre el fenómeno de crenación4. Ahora si el medio extracelular es hipotónico (p ej. agua destilada), el H2O entrará a la célula, se hinchará y sufrirá el proceso de citólisis5(o lisado) deteniéndose todos sus procesos vitales. Se debe aclarar que un medio no es por sí solo hipotónico o hipertónico, cuando se usa esta terminología se está comparando un medio con respecto a otro. El proceso de ósmosis en una célula vegetal difiere del de una célula animal. Recordemos que en células vegetales existe por fuera de la membrana plasmática una pared celular que le confiere características distintas. Entonces: Si el medio es isotónico, habrá un equilibrio dinámico Si el medio es hipotónico, ingresará H2O a la célula vegetal y sus vacuolas se llenan incrementando la presión de turgencia, es decir aumenta el volumen de la célula y la pared va a dar contención impidiendo que la célula se destruya. Si el medio es hipertónico, la célula vegetal va a eliminar H2O y el volumen de la vacuola va a disminuir, produciendo que la membrana plasmática se despegue de la pared celular, ocurriendo la plasmólisis6. El H2O no solo entra y sale a la célula por ósmosis sino que también existen otros mecanismos por el cual se mueve el H2O como por ejemplo con acuaporinas (se verán más adelante) Osmosis. Efecto de los cambios de concentración de soluto en (a) células animales y (b) células vegetales 4 Crenación: destrucción de la célula cuando al estar en un medio hipertónico pierde gran cantidad de agua. La destrucción celular es por deshidratación. 5 Citólisis: Destrucción celular debido al ingreso de agua a su interior. La consecuencia es pérdida de citoplasma y material genético hacia el exterior. 6 Plasmólisis: Cuando se libera agua de la célula vegetal, disminuye el agua de las vacuolas y disminuye el volumen celular. Finalmente se separa la membrana plasmática de la pared celular. - 84 - b) Difusión Facilitada: Hay moléculas que por su tamaño o polaridad no pueden atravesar fácilmente las membranas por difusión simple. Por ejemplo: glucosa, aminoácidos, iones, etc. Necesitan de transportadores, que son proteínas integrales de membranas. Este mecanismo ocurre siempre a favor del gradiente y por lo tanto no requiere de energía adicional. Este gradiente puede ser de concentración (como se vio difusión simple, recordemos que era cuando las moléculas pasan de un compartimiento de mayor concentración a uno de menor concentración, es decir que las moléculas) o bien por un gradiente de potencial eléctrico, es decir el soluto con carga eléctrica, independientemente de su signo, se desplazará de una zona donde la carga sea mayor hacia otra donde la carga sea menor. La difusión facilitada depende: Del gradiente de concentración de la sustancia a ambos lados de la membrana Del número de proteínas transportadoras existentes en la membrana De la rapidez con que estas proteínas hacen su trabajo Figura: Asociación de proteínas de membrana con la bicapa lipídica. Transmembrana (o intrínsecas o integrales), atraviesan la membrana como hélice o como láminas plegadas cerradas. Las proteínas transportadoras pueden ser agrupadas en: I. Canales Iónicos o Proteínas Canal: Son poros o túneles formados por una o varias proteínas transmembrana. Existen canales iónicos en todas las células y tanto en la membrana plasmática como en las membranas de los organoides. Estos canales son selectivos, es decir que sólo transportan un tipo de ión (p. ej. K+, Na+, etc.). El transporte de un ión es impulsado por un gradiente electroquímico. La mayoría de estos canales no están siempre abiertos, sino que - 85 - se abren ante estímulos o respuesta, ante por ejemplo de la presencia de una sustancia inductora. II. Proteínas “Carrier” o Permeasas: Las permeasas están formadas por proteínas transmembrana multipaso. Generalmente transportan una gran variedad de iones y otras moléculas polares sin carga como la glucosa. Vale recordar que este tipo de mecanismo es sin gasto de energía y a favor de gradiente electroquímico. La velocidad de este transporte es menor al realizado por los canales iónicos. Transporte facilitado por medio de una permeasa. Figura: Distintos mecanismos y estructuras utilizados por los solutos para atravesar las membranas de una célula. - 86 - Como se mencionó en Difusión y Ósmosis, el H2O puede movilizarse también por medio de proteínas transportadoras altamente específicas, denominadas acuaporinas. Son canales especiales con una estructura helicoidal (forma de hélice) que permite el paso selectivo de H2O. No son canales iónicos y este transporte es pasivo, sin gasto de energía. En algunas células por ejemplo en células renales, se necesita un mayor transporte de H2O que el logrado simplemente por difusión (ósmosis). Estas acuaporinas se encuentran tanto en célula animal como vegetal. Esquema de una acuaporina: B. Transporte Activo: Es un transporte que se realiza en contra de gradiente ya sea de concentración o eléctrico, y requiere gasto de energía en forma de ATP7. Este tipo de mecanismo ocurre cuando las células deben ingresar (o importar) moléculas que están en menor concentración en el medio extracelular que en el citoplasma y también porque necesitan mantener constante la composición iónica intracelular. Este mecanismo también es específico como la difusión facilitada pero con la diferencia que el transporte activo va en contra de gradiente. Para realizarse necesita de energía adicional externa (endergónico). El funcionamiento de estos sistemas de transporte activo es análogo al de las bombas hidráulicas que elevan agua hacia tanques ubicados a grandes alturas con el aporte de energía externa (motor). Por esta analogía se denomina bombas a las proteínas que transportan sustancias en forma activa. Estas proteínas tienen sitios específicos de unión con alta afinidad por el soluto que va a transportar. En un primer paso la proteína que actúa como bomba, se une al soluto formando un complejo proteína-soluto. Esto ocurre en el compartimiento donde el gradiente de concentración o eléctrico del soluto es menor. Una vez producida esta unión, tiene lugar un cambio de conformación en la proteína, que depende del suministro de energía. Como resultado, el sitio al que está unido el soluto queda expuesto al otro lado de la membrana, es decir al compartimiento donde el gradiente de concentración o eléctrico es mayor. La nueva conformación tiene una afinidad mucho menor por el soluto y por lo tanto lo libera al medio. Finalmente la proteína retoma su conformación inicial y queda en condiciones de iniciar un nuevo ciclo. La energía para este proceso proviene de diferentes fuentes según el tipo celular. Por ejemplo existen bombas que funcionan 7 ATP: Abreviatura de trifosfato adenosina, el principal compuesto celular portador de energía. - 87 - sobre la base de energía lumínica por medio de absorción de fotones como ocurre en las membranas de algunas bacterias, hongos y levaduras. Otras, como las bombas de H+ o Na+ presentes en bacterias, mitocondrias y cloroplastos, utilizan la energía eléctrica resultante de procesos de oxidorreducción. Sin embargo la mayoría de las proteínas transportadoras sólo son capaces de aprovechar la energía química contenida en los enlaces entre ciertos átomos de moléculas de ATP, debido a esto las bombas reciben el nombre de ATPasas transportadoras. Unas de las bombas más importante en células eucariotas animales son la Bomba Na+ K+ (bomba sodio/potasio) y la de K+- H+ (bomba potasio/hidrógeno). Bomba Na+ - K+ Su función es expulsar Na+ al espacio extracelular e introducir K+ al citosol8. Ambos son movilizados en contra de su gradiente electroquímico, estableciendo así diferencias de concentración y carga entre el espacio extra e intracelular para ambos iones. Debido a que se esta transportando simultáneamente dos solutos distintos en sentidos opuestos, estamos en presencia de un sistema de contratransporte. Es importante recordar que, si bien el Na+ sale y el K+ ingresa a la célula, ambos lo hacen en contra de su gradiente y, en consecuencia, hace falta hidrolizar ATP para movilizarlos (hace falta energía química). La Bomba Na+-K+ tiene simultáneamente funciones de proteína transportadora y de ATPasa (hidroliza ATP para obtener energía). Por lo menos un tercio de la energía que consume una célula animal se destina para impulsar esta bomba. En las células nerviosas, donde la actividad eléctrica es sumamente importante, este valor asciende al 60%. Cada ATPasa puede hidrolizar hasta 100 moléculas de ATP. Esto nos da idea de la importancia de este sistema de transporte. Figura: Esquema de la ATPasa. 8 Citosol: Contenido del citoplasma, excepto las organelas delimitadas por membranas como las mitocondrias y el retículo enplasmático. - 88 - Figura: Esquema de una Bomba Na+ - K+ Cotransporte Na+ - glucosa Cuando se vio Difusión facilitada, la glucosa era una de las moléculas que podía movilizarse por este tipo de transporte sin requerir energía adicional. Sin embargo, hay situaciones, como en las membranas apicales de las células del intestino delgado o en las membranas de células renales, donde se absorbe glucosa desde la luz del intestino o de los túmulos renales aunque las concentraciones extracelulares sean bajas. En estos casos se realiza mediante cotransporte Na+ - glucosa. En un primer paso una bomba Na+ - K+ expulsa iones Na+ al medio extracelular creando una menor concentración de este ión en el medio intracelular. En una segunda etapa, una permeasa (ver en Difusión facilitada, tipos de proteínas transportadoras) cotransporta el Na+ y glucosa hacia el citoplasma celular. De este modo el Na+ ingresa a favor de gradiente y arrastra con él a la glucosa que ingresa en contra de gradiente de concentración. Este tipo de transporte también se denomina transporte acoplado a gradiente iónico o transporte activo secundario (porque está indirectamente ligado a una bomba). Finalmente la glucosa atravesará la célula y saldrá por difusión facilitada a favor de su gradiente de concentración, hacia el torrente sanguíneo. - 89 - Mecanismo de cotransporte Na+/glucosa en epitelio intestinal Existen otros tipos de ATPasas transportadoras de iones como el Ca++, H+ y en el caso de organismos procariotes también transportan ión cobre Cu++ un metal pesado. Algunas de ellas se volverán a ver en los capítulos siguientes en Respiración y Fotosíntesis. C. Transporte en masa o intercambio a través de vesículas Los mecanismos de transportes vistos en los puntos A y B, ocurren para iones o moléculas relativamente pequeñas. Las moléculas de mayor tamaño (macromoléculas), ingresan o abandonan la célula por medio del transporte en masa. Este tipo de transporte se realiza siempre con gasto de energía (ATP), es diferente a los vistos. Estos mecanismos implican la fusión o escisión de membrana y son muy importantes para la célula. Hablamos de endocitosis y la exocitosis. I. Endocitosis: Es el proceso por el cual la membrana plasmática se repliega hacia dentro (se invagina) y se va haciendo más profunda al tiempo que va rodeando la sustancia que va a ingresar a la célula junto con una cantidad de material del medio extracelular. A continuación se produce un estrangulamiento de la membrana y se forma una vesícula intracelular llamada endosoma. - 90 - Existen tres mecanismos de endocitosis: Dentro de esta vesícula puede haber una macromolécula o incluso un microorganismo, en este caso el mecanismo se denomina fagocitosis y la vesícula de gran tamaño se llama fagosoma. Gran parte del material que ingresa por endocitosis es degradado en los lisosomas. En algunos organismos unicelulares (protistas) constituye un modo de alimentación, ellos engloban grandes partículas (p. ej. bacterias) por medio de prolongaciones de la membrana plasmática llamados pseudópodos y las ingresan formándose así un fagosoma. Posteriormente será degradada por las enzimas de los lisosomas. En los animales sólo se da en algunas células altamente especializadas, llamadas células fagocíticas, macrófagos de los tejidos y glóbulos blancos sanguíneos denominados neutrófilos. En estos casos la función no es nutricional, sino defensiva contra infecciones, ingiriendo microorganismos patógenos. Otra función es la de eliminar células muertas o dañadas, o restos celulares (p. ej. glóbulos rojos no funcionales). Puede ingresar a la célula, fluído o líquidos con sustancias disueltas, también mediante pequeñas vesículas. En este caso se denomina pinocitosis. La endocitosis puede estar mediada por un receptor o ligando. Este mecanismo captura macromoléculas específicas que necesitan acoplarse a moléculas receptoras específicas para ingresar a la célula. Cerca de estos receptores que se ubican en la cara interna de la membrana plasmática, se encuentran las proteínas periféricas de membrana que inducen la curvatura de la membrana y la formación de vesículas. Un ejemplo importante de este mecanismo de endocitosis mediada por un receptor, ocurre en las células animales cuando captura colesterol. El colesterol, debido a su carácter hidrofóbico, es transportado por la sangre unido a proteínas, formando complejos llamados lipoproteínas de baja densidad (LDL). Estas LDL se unen a receptores ubicados en la superficie celular y los complejos LDL-receptor son internalizados (o ingresados) en vesículas y luego transferidas a los endosomas. En el interior de los endosomas, el LDL se disocia del receptor y este es reciclado nuevamente a la membrana plasmática para captar nuevamente LDL. - 91 - Tipos de transporte en masa II. Exocitosis: Es el proceso inverso a la endocitosis. En la exocitosis, ciertas vesículas intracelulares se fusionan desde el lado interno de la membrana plasmática y el contenido de las vesículas se libera al exterior de la célula. Por medio de este mecanismo se exportan neurotransmisores, enzimas digestivas y hormonas como la insulina en respuesta a una señal o estímulo del medio externo. La fusión de membranas es mediada en general por proteínas llamadas de fusión. Este mecanismo ocurre en la mayoría de las células y además de permitir el intercambio de sustancias, también contribuyen al reciclado de componentes de la membrana plasmática. En la exocitosis hay ganancia de membrana plasmática mientras que en la endocitosis hay pérdida de membrana. - 92 - 8. RESPIRACIÓN Y FOTOSÍNTESIS La Respiración Celular es la degradación del alimento con la liberación paulatina de energía. Este control está ejercido por enzimas específicas. Haciendo un repaso de los contenidos de capítulos anteriores, recordemos que los azucares (hidratos de carbonos), ácidos grasos (lípidos), aminoácidos y nucleótidos son moléculas basadas en carbono. Los azúcares constituyen una fuente primaria de energía química para las células y son incorporados a polisacáridos para el almacenamiento de energía. Los ácidos grasos son también importantes para el almacenamiento de energía pero su función más significativa es la formación de las membranas celulares. Los polímeros formados por aminoácidos constituyen macromoléculas muy diversas denominadas proteínas. Los nucleótidos desempeñan un papel fundamental en la transferencia de energía y también son las subunidades a partir de las cuales se construyen las macromoléculas de información, ARN y ADN. Antes de desarrollar la respiración celular, se hará un breve resumen desde la incorporación de alimentos hasta la generación de energía en ATP: Las proteínas, los lípidos y los polisacáridos que constituyen la mayor parte de los alimentos que comemos, deben ser degradados a moléculas menores antes de que las células puedan utilizarlos. Así esta degradación enzimática o catabolismo, ocurre en tres etapas: Etapa I: Llamada digestión, ocurre principalmente en el intestino. Las macromoléculas se rompen (escinden) en monómeros (p. ej. proteínas en aminoácidos, las grasas en ácidos grasos y glicerol, etc.). Etapa II: Ocurre en el citoplasma celular cuando las pequeñas moléculas resultantes de la etapa I ingresan a la célula (por los mecanismos que vimos en el ítem anterior) y sufren una degradación posterior. La mayoría de los átomos de carbono e hidrógeno de los azúcares se transforman en piruvato que ingresa a las mitocondrias para formar acetil coenzima A (acetil CoA). Esta etapa corresponde a la glucólisis y se verá con más detalle por ser una de las vías centrales en el metabolismo de los organismos vivos. Etapa III: El acetil CoA se degrada en CO2 y H2 O en la mitocondria. En esta fase final se genera la mayor parte de ATP. Esta etapa corresponde al denominado ciclo de Krebs y transporte de electrones y se verán posteriormente en más detalles. - 93 - Figura: Esquema simplificado de las tres etapas del catabolismo que conducen desde el alimento hasta los productos de desechos. Esta serie de reacciones produce ATP que luego es utilizado para impulsar reacciones de biosíntesis y otros procesos celulares que requieren energía. - 94 - Así a través de la energía derivada originalmente de la combustión de los hidratos de carbonos y de las grasas se redistribuyen en forma de energía química, convenientemente empaquetada. El ATP como se vio, es el principal transportador de energía en los sistemas vivos, y es necesario para una gran variedad de procesos celulares vitales, desde la biosíntesis química hasta la contracción muscular durante el ejercicio físico. Aproximadamente el 40% de la energía libre emitida por la oxidación de la glucosa se conserva en forma de ATP. La respiración ocurre en distintas estructuras celulares. La primera de ellas, como ya se mencionó, es la glucólisis que ocurre en el citoplasma. La segunda etapa dependerá de la presencia o ausencia de O2 en el medio, determinando en el primer caso la respiración aeróbica (ocurre en las mitocondrias), y en el segundo caso la respiración anaeróbica o fermentación (ocurre en el citoplasma). Glucólisis La glucólisis, lisis o escisión de la glucosa, tiene lugar en una serie de nueve reacciones, cada una catalizada por una enzima específica, hasta formar dos moléculas de ácido pirúvico (o piruvato), con la producción de ATP. La ganancia neta es de dos moléculas de ATP y dos de NADH9 por cada molécula de glucosa. Las reacciones de la glucólisis se realizan en el citoplasma y pueden darse en condiciones anaerobias; es decir en ausencia de oxígeno. Los primeros cuatro pasos de la glucólisis sirven para fosforilar (incorporar fosfatos) a la glucosa y convertirla en dos moléculas del compuesto de 3 carbonos gliceraldehído fosfato (PGAL). En estas reacciones se invierten dos moléculas de ATP a fin de activar la molécula de glucosa y prepararla para su ruptura. Paso 1: El grupo fosfato Terminal se transfiere desde el ATP al carbono en la posición 6 de la glucosa y se forma glucosa 6-fosfato. Paso 2: La molécula se reorganiza. La glucosa se transforma en fructosa. Paso 3: La fructosa 6-fosfato gana un segundo fosfato que proviene de otro ATP y se produce fructosa 1,6 bifosfato. Paso 4: El azúcar de seis carbonos se escinde en dos moléculas de tres carbonos: la dihidroxiacetona fosfato y gliceraldehído fosfato. Paso 5: Las moléculas de gliceraldehído fosfato se oxidan, es decir que pierden los átomos de hidrógeno con sus electrones, y el NAD+ se reduce a NADH y H+. Un ión fosfato se une a la posición 1 del gliceraldehído fosfato. 9 NADH: Forma reducida del dinucleótido de nicotinamida y adenina. Coenzima que funciona como aceptor de electrones. - 95 - Paso 6: El fosfato se libera de la molécula de bifosfoglicerato, reacciona con una molécula de ADP10 y se forma ATP. Paso 7: El grupo fosfato remanente se transfiere de la posición 3 a la posición 2. Paso 8: Se elimina una molécula de agua del compuesto de tres carbonos Paso 9: El fosfato se transfiere a una molécula de ADP y se forma otra molécula de ATP. Resumen de la Glucólisis ECUACIÓN DE LA GLUCÓLISIS Glucosa + 2 ADP + 2 Pi11 + 2 NAD+ 2 piruvato + 2 ATP + 2 NADH + 2 H+ + 2 H2O 10 ADP: Nucleótido compuesto por adenina, ribosa y un grupo fosfato; su fuente más común es la eliminación de un fosfato de una molécula de ATP. 11 Pi: Abreviatura de fósforo inorgánico. - 96 - Resumen de las dos etapas de la glucólisis. En la primera etapa se utilizan 2 ATP y la segunda produce 4 ATP y 2 NADH. Otros azúcares, además de la glucosa, como la manosa, galactosa y las pentosas, así como el glucógeno y el almidón, pueden ingresar en la glucólisis una vez convertidos en glucosa 6-fosfato. - 97 - Vías anaeróbicas El ácido pirúvico puede tomar por una de varias vías. Dos son anaeróbicas (sin oxígeno) y se denomina fermentación alcohólica y fermentación láctica. A la falta de oxígeno, el ácido pirúvico puede convertirse en etanol (alcohol etílico) o ácido láctico según el tipo de célula. Por ejemplo, las células de las levaduras pueden crecer con oxígeno o sin él. Al extraer jugos azucarados de las uvas y al almacenarlos en forma anaerobia, las células de las levaduras convierten el jugo de la fruta en vino al convertir la glucosa en etanol. Cuando el azúcar se agota las levaduras dejan de fermentar y en este punto la concentración de alcohol está entre un 12 y un 17 % según sea la variedad de la uva y la época en que fue cosechada. La formación de alcohol a partir del azúcar se llama fermentación. Fermentación alcohólica El ácido pirúvico formado en la glucólisis se convierte anaeróbicamente en etanol. En el primer caso se libera dióxido de carbono, y en el segundo se oxida el NADH y se reduce a acetaldehído. Otras células, como por ejemplo los glóbulos rojos, las células musculares y algunos microorganismos transforman el ácido pirúvico en ácido láctico. En el caso de las células musculares, la fermentación láctica, se produce como resultado de ejercicios extenuantes durante los cuales el aporte de oxígeno no alcanza a cubrir las necesidades del metabolismo celular. La acumulación del ácido láctico en estas células produce la sensación de cansancio muscular que muchas veces acompaña a esos ejercicios. Fermentación láctica En esta reacción el NADH se oxida y el ácido pirúvico se reduce transformándose en ácido láctico. - 98 - La fermentación sea alcohólica o láctica ocurre en el citoplasma. Esquema bioquímico del proceso de fermentación A) B) Alcohólica : 2 ácido pirúvico + 2 NADH === 2 etanol + 2 CO2 + 2 NAD+ Láctica : 2 ácido pirúvico + 2 NADH === 2 ácido láctico + 2 NAD+ La finalidad de la fermentación es regenerar el NAD+ permitiendo que la glucólisis continúe y produzca una provisión pequeña pero vital de ATP para el organismo. Respiración Aeróbica En presencia de oxígeno, la etapa siguiente de la degradación de la glucosa es la respiración, es decir la oxidación escalonada del ácido pirúvico a dióxido de carbono y agua. La respiración aeróbica se cumple en dos etapas: el Ciclo de Krebs y el transporte de electrones y la fosforilación oxidativa (estos dos últimos procesos transcurren acopladamente). En las células eucariotas estas reacciones tienen lugar dentro de las mitocondrias; en las procariotas se llevan acabo en estructuras respiratorias de la membrana plasmática. Estructura de las Mitocondrias: Las mitocondrias están rodeadas por dos membranas, una externa que es lisa y una interna que se pliega hacia adentro formando crestas. Dentro del espacio interno de la mitocondria en torno a las crestas, existe una solución densa (matriz o estroma) que contiene enzimas, coenzimas, agua, fosfatos y otras moléculas que intervienen en la respiración. La membrana externa es permeable para la mayoría de las moléculas pequeñas, pero la interna sólo permite el paso de ciertas moléculas como el ácido pirúvico y ATP y restringe el paso de otras. Esta permeabilidad selectiva de la membrana interna, tiene una importancia crítica porque capacita a las mitocondrias para destinar la energía de la respiración para la producción de ATP. La mayoría de las enzimas del ciclo de Krebs se encuentran en la matriz mitocondrial. Las enzimas que actúan en el transporte de electrones se encuentran en las membranas de las crestas. Las membranas internas de las crestas están formadas por un 80 % de proteínas y un 20 % de lípidos. En las mitocondrias, el ácido pirúvico proveniente de la glucólisis, se oxida a dióxido de carbono y agua, completándose así la degradación de la glucosa. El 95 % del ATP producido se genera, en la mitocondria. Las mitocondrias son consideradas organoides semiautónomos, porque presentan los dos ácidos nucleicos (del tipo procarionte) - 99 - Figura: Esquema de la ultraestructura de una mitocondria. (a) Esquema tridimensional, (b) Esquema de un corte al M.E.T. (c) Cresta mitocondrial (detalle). - 100 - Figura: Microfotografía electrónica de una mitocondria. Se observan las invaginaciones de la membrana interna que forman las características crestas, que identifican esta organela. Como puede apreciase en la figura c) de la página anterior, las crestas mitocondriales aparecen cubiertas por partículas en forma de hongo, que tienen un tallo más fino que las unen a la membrana. Estas estructuras son las llamadas partículas F1 y representan una porción de la ATPasa especial que interviene en el acoplamiento entre la oxidación y la fosforilación. Las partículas F1 se encuentran en la membrana interna, del lado relacionado con la matriz; le confieren una asimetría característica relacionada con la función de la ATPasa (este punto se verá más detalladamente al referirnos a la hipótesis quimiosmótica). Para concluir, es importante destacar que el ciclo de Krebs se lleva a cabo en la matriz mitocondrial; mientras que el transporte de electrones y la fosforilación oxidativa se producen a nivel de las crestas mitocondriales. Paso intermedio al Ciclo De Krebs El ácido pirúvico sale del citoplasma, donde se produce mediante glucólisis y atraviesa las membranas externa e interna de las mitocondrias. Antes de ingresar al Ciclo de Krebs, el ácido pirúvico, de 3 carbonos, se oxida. Los átomos de carbono y oxígeno del grupo carboxilo se eliminan como dióxido de carbono (descarboxilación oxidativa) y queda un grupo acetilo, de dos carbonos. En esta reacción exergónica, el hidrógeno del carboxilo reduce a una molécula de NAD+ a NADH. - 101 - Ahora la molécula original de glucosa se ha oxidado a dos moléculas de CO2, y dos grupos acetilos y, además se formaron 4 moléculas de NADH (2 en la glucólisis y 2 en la oxidación del ácido pirúvico). Cada grupo acetilo es aceptado por un compuesto llamado coenzima A dando un compuesto llamado acetilcoenzima A (acetil CoA). Esta reacción es el eslabón entre la glucólisis y el ciclo de Krebs. Ciclo De Krebs El ciclo de Krebs también conocido como ciclo del ácido cítrico es la vía común final de oxidación del ácido pirúvico, ácidos grasos y las cadenas de carbono de los aminoácidos. La primera reacción del ciclo ocurre cuando la coenzima A transfiere su grupo acetilo (de 2 carbonos) al compuesto de 4 carbonos (ácido oxalacético) para producir un compuesto de 6 carbonos (ácido cítrico). El ácido cítrico inicia una serie de pasos durante los cuales la molécula original se reordena y continúa oxidándose, en consecuencia se reducen otras moléculas: de NAD+ a NADH y de FAD+ a FADH2. Además ocurren dos carboxilaciones y como resultado de esta serie de reacciones vuelve a obtenerse una molécula inicial de 4 carbonos el ácido oxalacético. El proceso completo puede describirse como un ciclo de oxalacético a oxalacético, donde dos átomos de carbono se adicionan como acetilo y dos átomos de carbono (pero no los mismos) se pierden como CO2. - 102 - Figura: Esquema simplificado del Ciclo de Krebs Dado que por cada molécula de glucosa inicial se habían obtenido dos de ácido pirúvico y, por lo tanto dos de acetil CoA, deben cumplirse dos vueltas del ciclo de Krebs por cada molécula de glucosa. En consecuencia los productos obtenidos de este proceso son el doble del esquema que se detalla a continuación. - 103 - Balance parcial de la respiración Proceso Sustrato Glucólisis Glucosa Entrada al ciclo de Krebs 2 ácido pirúvico Ciclo de Krebs 2 Acetil CoA Glucosa Productos 2 ácido pirúvico 2 ATP 2 NADH 2 Acetil CoA 2 CO2 2 NADH 4 CO2 2 GTP (equivalentes a 2 ATP) 6 NADH 2 FADH2 6 CO2 2 ATP 2 GTP 10 NADH 2 FADH2 Observando el balance parcial del ciclo de Krebs, se comprueba que en este proceso no se obtiene energía directamente bajo la forma de ATP (sólo se obtiene 1 GTP que es equivalente a 1 ATP). En cambio se obtienen cantidades de coenzimas reducidas (NADH y FADH2), y es a través de la oxidación posterior que se obtendrá la energía para sintetizar ATP. Cada coenzima NADH equivale a 3 ATP y cada coenzima FADH2 equivale a 2 ATP. Transporte de Electrones o Cadena Respiratoria En esta etapa, que ocurre en las crestas mitocondriales, se oxidan las coenzimas reducidas, el NADH se convierte en NAD+ y el FADH2 en FAD+. Al producirse esta reacción, los átomos de hidrógeno (o electrones equivalentes), son conducidos a través de la cadena respiratoria por un grupo de transportadores de electrones, llamados citocromos. Los citocromos experimentan sucesivas oxidaciones y reducciones (reacciones en las cuales los electrones son transferidos de un dador de electrones a un aceptor). En consecuencia, en esta etapa final de la respiración, estos electrones de alto nivel energético descienden paso a paso hasta el bajo nivel energético del oxígeno (último aceptor de la cadena), formándose de esta manera agua. Cabe aclarar que los tres primeros aceptores reciben el H+ y el electrón conjuntamente. En cambio, a partir del cuarto aceptor, sólo se transportan electrones, y los H+ quedan en solución. - 104 - Fosforilación Oxidativa También este proceso ocurre en las crestas mitocondriales y el flujo de electrones está íntimamente acoplado al proceso de fosforilación, y no sucede a menos que también pueda verificarse este último. Esto, en un sentido, impide el desperdicio ya que los electrones no fluyen a menos que exista la posibilidad de formación de fosfatos ricos en energía. Si el flujo de electrones no estuviera acoplado a la fosforilación, no habría formación de ATP y la energía de los electrones se degradaría en forma de calor. Puesto que la fosforilación del ADP para formar ATP se encuentra acoplada a la oxidación de los componentes de la cadena de transporte de electrones, este proceso recibe el nombre de fosforilación oxidativa. En tres transiciones de la cadena de transporte de electrones se producen caídas importantes en la cantidad de energía potencial que retienen los electrones, de modo que se libera una cantidad relativamente grande de energía libre en cada uno de estos tres pasos, formándose ATP. Figura: Diagrama de la cadena respiratoria y de la fosforilación oxidativa asociada - 105 - Balance energético del proceso: Glucólisis = 2 ATP 2 NADH + H+ (x2) = 2 ATP = 4 ATP Piruvato ------ Acetil CoA 1 NAD + H+ (x3) = 3 ATP Krebs = 1 GTP 3 NADH + H+ 1 FADH2 (x3) (x2) ( = 1 ATP) = 9 ATP = 2 ATP X2 Porque es necesario dar dos vueltas al ciclo para oxidar la glucosa Vale aclarar que solo los NADH + H+ mitocondriales resultan equivalentes a 3 ATP. Aquellos NADH + H+ que fueron obtenidos en glucólisis y por lo tanto son citoplasmáticos, solo rinden 2 ATP (al igual que un FADH2) ya que la membrana interna mitocondrial resulta impermeable al NADH + H+ y estos deben transferir sus H+ a un intermediario de nivel energético equivalente al FADH2. Esto indica que por la oxidación total de una molécula de glucosa, se obtienen 36 ATP, en términos generales y puede variar según los distintos tipos celulares y según se parta de glucosa o glucógeno. Se resume en el siguiente diagrama, el metabolismo de los glúcidos o hidratos de carbono en una célula eucariota. Se puede observar que en el mismo, el piruvato luego de la glucólisis puede tomar dos vías diferentes dependiendo si está en presencia de oxígeno, entonces ingresará en el ciclo de Krebs en la matriz mitocondrial, o si en cambio no hay oxígeno presente, entonces ocurrirá fermentación en el citoplasma celular. La fermentación es común en procariontes, levaduras y en el ser humano. En este último, sólo dos tipos celulares degradan glucosa en forma fermentativa: los glóbulos rojos y las células musculares en condiciones de trabajo intenso. Vale aclarar que en el diagrama, ciclo ATC en la matriz mitocondrial, significa: ciclo del ácido tricarboxílico comúnmente llamado Ciclo de Krebs. - 106 - Otras Vías Catabólicas Sí la mayoría de los organismos no se alimentan directamente de glucosa, ¿cómo obtienen energía a partir de las grasas o proteínas? La respuesta está en que el ciclo de Krebs es un gran nudo del metabolismo energético. Otras sustancias alimenticias son degradadas y convertidas en moléculas capaces de ingresar al ciclo. Las grasas se desdoblan en sus componentes glicerol y ácidos grasos. Estos últimos son fraccionados en fragmentos de dos carbonos e introducidos en el ciclo de Krebs como acetil CoA. Las proteínas se degradan a aminoácidos, estos son desaminados (se les eliminan los grupos amino) y el esqueleto de carbonos se convierte en un grupo acetilo, ingresando al ciclo de Krebs. Los grupos amino si no se utilizan, se excretan como urea u otros desechos nitrogenados. - 107 - FOTOSINTESIS: En los primeros apuntes sobre El Origen de la Vida, se vio que aún hoy no se ha podido demostrar si las primeras células fueron heterótrofas o autótrofas, pero sí los científicos están de acuerdo en que la evolución sobre la Tierra no podría haber sido sino fuera por el surgimiento de organismos autótrofos fotosintéticos. Se nos puede plantear ¿qué relación existe entre los organismos autótrofos y la vida sobre la Tierra? Se puede decir que existe una conexión entre la ecología y la bioquímica a través del proceso de fotosíntesis. La vida sobre la Tierra depende de la fotosíntesis, porque por medio de esta, la energía lumínica del Sol que es captada por pigmentos particulares, forman hidratos de carbono y oxígeno libre (O2) necesario para el normal desarrollo del Hombre. Recordemos cómo era la atmósfera primitiva en el Origen de la Vida, no había oxígeno libre porque los primeros microorganismos que poblaron la superficie de la Tierra no realizaban fotosíntesis que produjera oxígeno como subproducto, todos los organismos eran anaerobios. Posteriormente la aparición de oxígeno atmosférico permitió la formación de la capa de ozono (O 3) y la gran diversidad biológica. El proceso de fotosíntesis es un claro ejemplo de cómo se transforma la energía: se transforma la energía lumínica en energía química. Volviendo a capítulos anteriores, se dijo que los plásticos son organelas que poseen dos membranas, hay diferentes tipos uno de ellos son los cloroplastos que tienen un rol fundamental en la fotosíntesis. Debemos aclarar que la fotosíntesis la realizan los organismos autótrofos, es decir aquellos que no necesitan incorporar del medio, sustancias orgánicas sintetizadas porque pueden fabricar su propio alimento a través de la fotosíntesis. Nos referimos a plantas, algas y algunas bacterias (tiobacterias púrpuras). Ecuación general de la fotosíntesis: CO 2 + H 2 O + Luz --------------- C 6 H 12 O 6 + H 2 O + O 2 La fotosíntesis se realiza en dos etapas: Etapa lumínica: como lo indica su nombre, esta etapa depende de la presencia de luz y consiste en transformar la energía lumínica en energía química (en forma de ATP). Como consecuencia de la hidrólisis del agua, esta se oxida, desprendiéndose O 2 a la atmósfera y reduciéndose la coenzima NADPH + H +. Se realiza en los tilacoides de las granas de los cloroplastos. - 108 - Etapa que fija el carbono: mal llamada etapa oscura porque no depende directamente de la luz pero indirectamente porque necesita de los productos derivados de la primera etapa lumínica, y consiste en la fijación de CO 2 para formar azúcares. Muchas de las enzimas que intervienen en esta etapa son reguladas o activadas de forma indirecta por la luz. Se realiza en el estroma de los cloroplastos. Etapa lumínica o fotoquímica Se inicia con la captación y absorción de la energía lumínica proveniente del Sol, por los pigmentos que se encuentran en las granas de los cloroplastos, la clorofila. La clorofila es sensible a determinadas longitudes de onda de la luz que coincide con casi todo el espectro de la luz visible: Figura: Se muestran las longitudes de ondas que corresponden a la luz visible. Hacia la izquierda del espectro (menor longitud de onda) se encuentran los rayos UV, rayos X y rayos gamma, que resultan nocivos para el material genético de las células. Hacia la derecha del espectro (mayor longitud de onda), se encuentran los rayos infrarrojos, microondas y ondas de radio, que carecen de energía suficiente para excitar electrones. La clorofila es el pigmento que le otorga el color verde a las plantas sin embargo también está presente en plantas y alga de distintos colores que realizan fotosíntesis. Existen varios tipos de clorofila pero la más importante es la clorofila a. Así por ejemplo en las algas verdes y plantas contienen un segundo tipo: clorofila b, en algas pardas y diatomeas están presentes la clorofila c. Todos los organismos fotosintéticos poseen otros pigmentos llamados carotenoides. Estos pigmentos accesorios funcionan como receptores suplementarios de luz, captando otras longitudes de onda y transfiriéndoselas hacia la clorofila a. - 109 - Figura: Ultraestructura del cloroplasto. (a) esquema tridimensional, (b) esquema de la ultraestructura al M.E.T, (c) grana de un cloroplasto - 110 - Figura: microfotografía electrónica de un corte de una parte del cloroplasto, donde se observan las granas. Las plantas, algas y cianobacterias poseen en los tilacoides complejos llamados fotosistemas I y II, formados por proteínas transmembrana. Estos fotosistema I y II difieren en el pico de absorción de la clorofila y trabajan juntos en las reacciones de oxidorreducción del transporte de electrones. Como se mencionó el proceso se inicia con la captación de luz lumínica por la clorofila. La luz incide sobre el Fotosistema II, excita los electrones aumentando así la energía “lanzando electrones cuesta arriba”. Estos electrones son reemplazados por electrones de moléculas de agua que, al escindirse, liberan O 2 y H+. Los electrones luego pasan “cuesta abajo”, a lo largo de una cadena de transporte de electrones, al Fotosistema I y de éste (nuevamente cuesta abajo) al NADP, que se reduce formando NADPH + H+ . Como resultado de este proceso se produce la fosforilación del ADP para obtener ATP, que serán utilizados en la siguiente etapa. Resumiendo: La energía luminosa de la fotosíntesis consiste en la transformación de: ENERGÍA LUMINOSA ENERGÍA QUÍMICA La energía química queda contenida en moléculas de dos tipos: [ [ ATP NADPH Como subproducto de esta etapa, se obtiene: [ O2 (oxígeno molecular) - 111 - Figura: Esquema simplificado de la etapa lumínica de la fotosíntesis Etapa que fija el carbono o bioquímica: Las enzimas presentes en el estroma de los cloroplastos, regulan una serie de reacciones bioquímicas que permiten la reducción del CO2 y su fijación por medio de la condensación a un compuesto intermedio el gliceraldehído 3P, que será el que permita la formación glucosa y otros compuestos orgánicos. Este proceso transcurre en forma de ciclo y se inicia a partir de una molécula de 5 átomos de carbono (la ribulosa difosfato) que acepta la adición de CO 2 y luego se escinde formando dos moléculas de fosfoglicerato o PGA. Cada molécula de PGA contiene tres átomos de carbono de ahí el nombre de Ciclo de Calvin o Ciclo del Carbono 3. Se puede hacer una analogía con el Ciclo de Krebs en las mitocondrias y al igual que éste, cada paso es catalizado por una enzima específica. - 112 - Haciendo un resumen de esta etapa, en el Ciclo de Calvin el NADPH y el ATP formados en las reacciones que capturan energía lumínica se utilizan para reducir el CO2. Figura: Diagrama simplificado del Ciclo de Calvin En cada vuelta al Ciclo de Calvin se incorpora una molécula de CO2. Tres vueltas al Ciclo introducen tres moléculas de CO2 (equivalente a un azúcar de tres carbonos) y produce una molécula de gliceraldehído fosfato. Por lo tanto se necesitan dar seis vueltas al Ciclo para introducir seis moléculas de CO2 y producir el equivalente a un azúcar de seis carbonos, como la glucosa. 6 CO2 + 12 H2O C6H12O6 + 6 O2 + 6 H2O - 113 - Fotorrespiración: Este proceso ocurre cuando la concentración de CO2 en la hoja de la planta es baja en relación con la concentración de O2 y también se ve favorecida a temperaturas ambientales altas, superiores a 28 ºC. Bajo estas condiciones, se inicia un proceso en los peroxisomas y mitocondrias denominado fotorrespiración en el cual se forman compuestos intermedios que consumiendo ATP, dan lugar a la producción de CO2 y H2O. En condiciones atmosféricas normales, hasta el 25% del carbono fijado durante la fotosíntesis se reoxida a CO2 durante la fotorrespiración, reduciendo así la eficiencia fotosintética de las plantas. 9. TIPOS CELULARES Hasta ahora se vio todo lo que ocurre dentro de una célula y sus componentes celulares. Esta visión tan microscópica y específica genera una incapacidad de trasladar estos conocimientos a un nivel de organización superior al celular. Sin embargo para poder avanzar en la comprensión de la biología, es necesario tener la información dada en los capítulos anteriores. Por tal motivo, en este capítulo se darán algunos ejemplos de la enorme especialización celular en organismos eucariontes, para de esta forma lograr una visión más amplia e integradora. La riqueza de especializaciones diferentes que se encuentran en las células de un animal superior, es incomparablemente mayor a la que puede presentar cualquier procariota. En un vertebrado se puede distinguir más de 200 tipos celulares que generan una gran diversidad de comportamientos dentro de un mismo organismo eucariota. Sin embargo estas diferentes células presentan aspectos básicos que son comunes a todas las células, como se vio en los capítulos anteriores. Para dar un ejemplo, se puede considerar la unión neuromuscular en la que interviene sólo tres tipos de células: la célula muscular, la célula nerviosa y la célula de Schwann. La célula muscular se ha especializado en la contracción y su citoplasma contiene un elevado número de filamentos de actina y numerosas mitocondrias que suministran ATP como combustible para el aparato contráctil. La célula nerviosa estimula el músculo que va a contraerse. Para ello transmite una señal excitadora procedente del cerebro o médula espinal. La célula nerviosa es muy larga y puede medir más de un metro desde su núcleo hasta su unión con el músculo. La célula de Schwann está especializada en producir membrana plasmática que va a rodear a la célula nerviosa para formar una vaina de mielina que actuará como aislante. - 114 - Ejemplos de algunos tipos celulares en vertebrados: - 115 - - 116 - - 117 - - 118 - BIBLIOGRAFIA Alberts, B; Watso, J; Bray, D; Lewis, J; Raff, M; Roberts, K. Biología Molecular de la Célula; 3ª Edición; Ediciones Omega S.A. Barcelona. 1996. Alberts, B et al. Molecur Biology of the cell. 4th Edition. Garland Publishing. 2002. Asimov, I. Breve Historia de la Biología. 3ª Edición. Editorial Universitaria de Buenos Aires. 1975. Avers, Ch. Biología celular. Grupo Editorial Iberoamérica. 1991. Broks, T. Biology of the Microorganisms. 8th Edition. Prentice Hall. N. Y. 1997 Campbell, N. Biology. 4th Edition. The Benjamin Cummings Publishing Company Inc. California. 1997. Castro, R; Handel, M; Rivolta, G. Actualizaciones en Biología. 8ª Edición. Editorial Universitaria de Buenos Aires. 1990. Castro, R. et al. Investigación y Ciencia. Nº 39. Diciembre de 1979. CONICET-SENOC. Módulo 3 y 4. 1984. Curtis, H; Barnes, N. Biología. 7ª Edición. Editorial Médica Panamericana. Buenos Aires. 2008. Curtis, H; Barnes, N. Invitación a la Biología. Editorial Médica Panamericana. Buenos Aires. 1987. De Robertis, E; Hib, J. Fundamentos de Biología Celular y Molecular. 4ª Edición. Editorial El Ateneo. Buenos Aires. 2004. De Robertis, E; Hib, J. Biología Celular y Molecular. 12ª Edición. Editorial El Ateneo. Buenos Aires. 1996. Dickerson, R. La Evolución Química y el origen de la Vida. Investigación y Ciencia Nº 26. Noviembre 1978. Harper, H. Manual de Bioquímica. Ediciones El Manual Moderno. México. 1995. Holtzman, E; Novikoff, A. Estructura y Dinámica Celular. 3ª Edición. México. Editorial Interamericana. 1986. Karp, G. Biología Celular y Molecular. Editorial Mc Graw Hill Interamericana. México. 1998. - 119 - Kuchel, P; Ralston, G. Bioquímica General. Serie Schaum. Editorial Mc Graw Hill. México. 1994. Lehninger, A. Bioquímica. 2ª Edición. Editorial Omega S.A. México. 1985. Lehninger, A; Nelson, D; Cox, M. Principios de Bioquímica. 4ª Edición. Editorial Omega S.A. España. 2005. Moreno Azorero, R; Schwartzman, G. Principios de Biología Celular. Editorial El Ateneo. Buenos Aires. 1986. Smith and Wood. Biología Celular. Editorial Addison-Wesley. Iberoamericana S.A. 1997. Smith and Wood. Moléculas Biológicas. Editorial Addison-Wesley. Iberoamericana S.A. 1998. Solomon, E; Ville, C; Davis, P. Biología. 4ª Edición. Editorial Mc Graw Hill Interamericana. México. 1998. Stryer, L. Bioquímica. Editorial Reverté. España. 1995. - 120 - MÓDULO Nº 3: GENÉTICA 1. REPRODUCCIÓN CELULAR La división celular por mitosis es un proceso complejo por el cual los materiales celulares se reparten en partes iguales entre las dos células hijas, resultando virtualmente idénticas entre sí e idénticas a su progenitora, es decir que cada nueva célula derivada del proceso mitótico, hereda una copia exacta de la información genética de la célula progenitora. Esto de debe en parte a que cada célula hija recibe alrededor de la mitad del contenido del citoplasma de la célula madre o progenitora. Esta división implica cambios previos ocurridos a nivel bioquímico, fundamentalmente la duplicación del ADN. Figura: Ciclo de División Celular Una célula típica, a lo largo de su vida, alterna entre fases de crecimiento y división denominado ciclo celular. En general consta de un período donde ocurre un importante crecimiento y aumento de la cantidad de organoides (interfase), un período de división celular (mitosis o meiosis) y la citocinesis. La interfase, a su vez, involucra períodos donde la célula realiza los procesos vitales propios de su función. Se la subdivide en tres períodos o etapas: G1, S y G2: Etapa G1 es un período muy variable en su duración donde ocurre un crecimiento general y duplicación de las organelas del citoplasma, excepto mitocondrias y cloroplastos los cuales se originan por división de estas estructuras preexistentes (recordar que ambos contienen ADN y ribosomas que les permite dividirse de forma más o menos independiente del núcleo celular). - 121 - Este período puede durar desde horas hasta años como ocurre por ejemplo en las neuronas (que no se dividen), en éstas el ciclo se detiene en G1 por lo que se dice que están en G0. En cambio en el otro extremo tenemos a las células que van a dar origen a los glóbulos rojos y en ellas el período G1 ocurre muy rápido puesto que la vida media de un glóbulo rojo es muy corta (120 días en el Hombre y 55 días en el ratón). En este período la célula posee una actividad bioquímica intensa, el ADN está en forma de cromatina, la célula aumenta de tamaño, sintetiza ribosomas y una gran diversidad de proteínas como: enzimas, proteínas del citoesqueleto, de membrana, etc., algunas estructuras como Aparato de Golgi, lisosomas y vacuolas derivan del retículo endoplasmático que aumenta de tamaño y se renueva constantemente, también en esta etapa se comienzan a duplicar los centríolos (recordar el capítulo de Componentes Celulares). Etapa S: es el proceso clave de replicación del ADN. Durante esta etapa se sintetiza ADN, histonas y otras proteínas asociadas al ADN. Etapa G2: esta fase precede a la etapa de división celular. Durante este período la célula se prepara para la división celular, se terminan de duplicar los centríolos y el ADN, que estaba en estado de cromatina, comienza a pasar al estado de cromosomas. Figura: Fases del Ciclo Celular - 122 - En el siguiente esquema se representa las etapas del Ciclo Celular de una célula hipotética que contiene dos cromosomas: Mitosis (o división del núcleo o cariocinesis) Del esquema de arriba vemos que el proceso de mitosis comienza al finalizar el período G2 y termina originando dos células hijas que inician un nuevo ciclo (interface- G1). Ambas células hijas contienen un conjunto completo de cromosomas y más o menos la mitad del citoplasma y de las organelas de la célula madre. - 123 - En organismos unicelulares, la mitosis es una forma de reproducción asexual. En organismos pluricelulares es el medio por el cual a partir de una sola célula (huevo o cigota) pueden crecer y reparar los tejidos dañados. La mitosis es semejante en todas las células eucariotas aunque existen algunas diferencias entre las células animales y vegetales. Las fases de la mitosis son: Profase, Metafase, Anafase y Telofase. Cuando una célula está en interfase, el material cromosómico está disperso y se observan como finos cordones. Al iniciarse la mitosis, la cromatina se arrolla lentamente y se condensa en forma compacta. Esta condensación sería necesaria para los complejos movimientos y separación de los cromosomas durante la mitosis. Cuando los cromosomas condensados se tornan visibles, cada uno consiste en dos réplicas llamadas cromátides unidas entre sí por el centrómero. Dentro de éste hay estructuras proteicas, los cinetocoros. Figura: Esquema de un cromosoma replicado Etapas de la Mitosis Profase: el comienzo de la profase está indicado por la visualización de los cromosomas. Cada uno de esos cromosomas está constituido por dos cromátidas (recordar que la duplicación del ADN ocurre en la interfase previa). Durante la profase los cromosomas tienden a una máxima espiralización, haciéndose más gruesos y cortos. Hacia el final de la profase, el nucléolo tiende a desintegrarse para finalmente - 124 - desaparecer y se desintegra la membrana nuclear, en tanto que comienza a constituirse las fibras del huso mitótico (en el citoplasma) a partir de los centríolos previamente duplicados. El huso acromático (o mitótico) está constituido por microtúbulos de igual naturaleza que aquellos que forman los centríolos. (Repasar el capítulo de Componentes Celulares). Algunas veces se denomina prometafase a la transición entre la profase y la metafase. Se trata de un período muy corto durante el cual se termina de desintegrar la envoltura nuclear y se acaba de armar el aparato mitótico. Metafase: los cromosomas, que poseen su máxima espiralización (o condensación), se unen a determinadas fibras del huso a la altura de los centrómeros y se disponen en el plano ecuatorial de la célula. Anafase: en esta etapa, los microtúbulos del huso van acortándose de modo que cada una de las cromátides hermanas se mueven hacia los polos de la célula. De este modo se opera la repartición del material genético duplicado en la interfase. - 125 - Telofase: en la última etapa de la mitosis, los cromosomas hijos separados en dos grupos, se descondensan (desespiralizan); la envoltura nuclear se forma nuevamente a partir del retículo endoplasmático y se sintetizan nuevos nucléolos en cada uno de los núcleos hijos. Citocinesis: consiste en la separación del citoplasma para limitar dos nuevas células. En las células animales se produce típicamente una constricción ecuatorial en la célula, que avanza de afuera hacia adentro. En las células vegetales, luego o simultáneamente a la telofase y a partir de vesículas del dictiosomas, se forma una placa en el ecuador de la célula (el fragmoplasto) que avanza desde el centro hacia la periferia hasta alcanzar las paredes y finalmente fundirse con ellas. Recordar cuando vimos las diferencias entre - 126 - células vegetal y animal, que las células vegetales no posee centríolos pero igual se forma un huso mitótico denominado huso anastral Mitosis en célula vegetal: - 127 - Meiosis o división reduccional La inmensa mayoría de los organismos eucariontes (ej.: moscas, erizos de mar, peces, Hombre) se reproducen sexualmente, es decir producen descendencia que poseen características genéticas de los dos progenitores. La reproducción sexual requiere en general de dos progenitores y siempre involucra dos procesos: meiosis y fecundación. Por el medio de la fecundación las dotaciones de genéticas de ambos progenitores se reúnen y forman un nuevo individuo. Por medio de la meiosis, se mantiene un número constante de cromosomas de generación en generación, que es característico de cada especie y también es un mecanismo que origina variabilidad al combinarse el material genético dentro de los mismos cromosomas. En los organismos eucariontes más complejos los cromosomas siempre existen en pares, hay invariablemente dos de cada clase formando parejas, cada uno de ellos se llama homólogo. Así los 46 cromosomas humanos, constituyen 23 pares. Figura: Ciclo vital sexual. Se observa alternancia de generaciones haploides y diploides En las gametas la cantidad de cromosomas es exactamente la mitad, existiendo sólo uno de cada clase. Esto ocurre porque son células destinadas a unirse, así cuando un espermatozoide fecunda a un óvulo se reconstituye el número normal de cromosomas de la especie. - 128 - Como en las células somáticas tenemos dos cromosomas de cada clase decimos que son diploides, en cambio a las gametas las llamamos haploides. Habitualmente designamos el número haploide como “n” y al diploide como “2 n”. Por ejemplo para la especie humana: n = 23 y 2n = 46. La célula diploide resultante de la fusión de dos gametas se la denomina huevo o cigoto. Cuando una célula u organismo tiene más de dos dotaciones completas de cromosomas por núcleo, lo denominamos poliploides. Etapas de la meiosis La meiosis consiste en dos divisiones nucleares sucesivas, denominadas meiosis I y meiosis II. En la meiosis I se aparean y luego se separan los cromosomas homólogos y en la meiosis II se separan las cromátidas de cada homólogo. Igual que en la mitosis, es durante la interfase cuando se duplican los cromosomas. El resultado son cuatro células, cada una con un miembro del par de homólogos presentes en la célula progenitora y con la mitad del número de cromosomas que tenía la progenitora. Es decir pasa de ser una célula diploide a una haploide. La meiosis I a su vez se divide en: Profase I, Metafase I, Anafase I y Telofase I: Durante la profase I los cromosomas homólogos (contienen dos cromátidas cada uno) se aparean o se juntan longitudinalmente y se condensan por espiralización. Hacia el final de la profase I los cromosomas están completamente espiralizados y unidos íntimamente, conformando un bivalente o tétrada (por tener cuatro cromátidas). En este paso es cuando se puede producir el intercambio de porciones de cromátidas, es decir ocurre la recombinación cromosómica. Hacia el final de la profase I los cromosomas homólogos comienzan a separarse. No obstante la separación no es completa puesto que los cromosomas homólogos permanecen unidos por los puntos de intercambio o quiasmas. Los quiasmas son considerados la expresión del fenómeno genético llamado crossing over o recombinación genética, por medio del cual determinadas porciones de cromosomas con determinada información genética, se intercambian entre los miembros homólogos de los pares. En la metafase I los bivalentes o tétradas se disponen en la placa ecuatorial de la célula, se asocian a los microtúbulos del huso, ya desorganizada la envoltura nuclear. En la anafase I, migran a los polos de la célula los homólogos constituidos cada uno por dos cromátidas ya recombinadas; de modo que en la telofase I se tendrán dos núcleos, cada uno con un solo miembro del par de homólogos pero con dos cromátidas. Entre la telofase I y la segunda división, se puede operar un breve período o intercinesis. En el comienzo de la profase II, la situación es similar a lo visto en mitosis, esto es, cada cromosoma está duplicado pero sólo existe un número “n” de cromosomas. - 129 - Las etapas de esta meiosis II son idénticas a la mitosis. En la metafase II los cromosomas, cada uno formado por dos cromátidas, se ubican en el plano ecuatorial. En la anafase II, se separarán (en cada uno de los núcleos) las cromátidas hermanas, de modo que, en la telofase II, se tendrán cuatro núcleos, cada uno con un solo ejemplar del par de homólogos y constituido por una sola cromátida recombinada. En los siguientes esquemas se visualizan las etapas descriptas: Meiosis I: Profase I - 130 - Anafase I Telofase I Meiosis II: Anafase II Telofase II - 131 - Gametogénesis El proceso antes descripto es el que ocurre en aquellas células destinadas a formar células sexuales. Según el tipo de organismo del que se trate, podemos hablar de una gametogénesis (es decir que la meiosis produce gametas) o esporogénesis (cuando los productos son esporas ). En el caso de las gametas, se originan por meiosis los óvulos femeninos y los espermatozoides masculinos. En ambos casos, se trata de células especiales, las gametogonias, las que en los órganos reproductivos (ovarios y testículos) van a experimentar la meiosis y así originar las gametas. - 132 - Figuras: Esquema de las etapas de esperrmatogénesis y ovogénesis - 133 - Comparación entre Mitosis y Meiosis - 134 - Características conceptuales del proceso de Muerte Celular Como vimos, para que un organismo pluricelular se desarrolle y se mantenga con eficiencia no sólo es importante la división celular para aumentar el número de células, sino también un proceso de muerte celular programada genéticamente denominado apoptosis. La división y muerte celular ocurren en forma coordinada. En vertebrados, la apoptosis controla el número de neuronas durante el desarrollo del sistema nervioso, elimina linfocitos defectuosos y da forma a los órganos en desarrollo. Por ejemplo, las células de la cola de los renacuajos se eliminan por apoptosis durante la metamorfosis de estos anfibios. En los embriones humanos, las células que forman membranas interdigitales se eliminan también por apoptosis durante el desarrollo embrionario. Las células también pueden morir a través de un proceso no controlado denominado necrosis. En este caso, la célula se hincha y explota, derramando su contenido hacia el entorno. En los vertebrados esto produce una inflamación que recluta glóbulos blancos y puede lesionar el tejido normal que lo rodea. Senescencia o envejecimiento celular es un estado en el cual ingresa la célula y no tiene retorno, es decir no nunca salen de este estado. Como vimos, las células no se dividen eternamente, sin embargo este envejecimiento celular no ocurre en todos los tipos celulares. Existe una relación entre el acortamiento progresivo que ocurre en los extremos de los cromosomas y la limitación en el número de divisiones celulares. 2. EXPERIMENTOS DE MENDEL Antes de introducirnos en los experimentos de Mendel, veremos algunos conceptos necesarios para comprender los fenómenos genéticos. La genética es una rama de la Biología que estudia la herencia y la variación de las causas que determinan las diferencias y similitudes entre individuos. Sabemos que los cromosomas están constituidos por ADN y que este ADN es el material genético. Cada secuencia de bases de ADN que codifica para un ARN se lo denomina Gen Por lo tanto, al ser un cromosoma una larga doble hélice de ADN, los cromosomas están constituidos por numerosos genes. Cada gen ocupa en la secuencia de nucleótidos del cromosoma, siempre un mismo y definido lugar que se denomina Locus1 Un individuo presentará entre todos sus cromosomas, un conjunto de genes que van a constituir el Genoma de ese individuo. Ese genoma lleva información genética para distintos caracteres. 1 El plural de locus es Loci. - 135 - El material genético responsable de una característica dada es el Genotipo. Sin embargo el material genético no podemos observarlo a simple vista; para estudiar un individuo necesitamos caracteres que sean visibles. Cuando la expresión del genotipo puede ser observable o medible, lo llamamos Fenotipo El fenotipo si bien depende del genotipo, también se encuentra influenciado por condiciones ambientales en los cuales se desarrolla el individuo. Así por ejemplo una planta puede heredar la capacidad de florecer y dar frutos pero si se mantiene en condiciones de oscuridad, temperatura y humedad desfavorables, no podrá florecer ni dar frutos. Por tal motivo, se dice que el fenotipo es la síntesis del genotipo y el ambiente. La variante de un gen que ocupa el mismo locus en un cromosoma se denomina Alelos. Supongamos una característica dada para un organismo, por ejemplo para una planta el color de sus semillas. Esta característica color de semillas, estará codificada por un par de genes que están en un par de cromosomas homólogos. Supongamos también que existen dos posibilidades para la característica: semillas amarillas y semillas verdes; existirá entonces codificación distinta para cada caso pero siempre en el mismo par de genes. A cada una de esas posibilidades se las llama alelos o genes alelos. Entonces un par de homólogos presentan dos alelos para un mismo gen. Estos alelos pueden ser iguales o distintos. Siguiendo el ejemplo anterior, una planta de arvejilla puede heredar un gen para semilla amarilla y el otro alelo para semilla verde, sin embargo las semillas que produce la planta son amarillas. En este caso uno de los alelos tapa los efectos del otro, ese alelo que se pone de manifiesto (gen para color amarillo) se llama Dominante. El alelo que queda oculto no puede expresarse (gen para color verde) se denomina Recesivo. Al simbolizarlos se le asigna una letra mayúscula al gen dominante, y la correspondiente minúscula al alelo recesivo: A= gen para el alelo dominante a= gen para el alelo recesivo - 136 - Si los dos genes que gobiernan un rasgo son iguales (ambos dominantes o ambos recesivos), el individuo que los porta se denomina Homogigota para ese rasgo. Si los dos genes que gobiernan un rasgo son distintos (uno dominante y otro recesivo), el individuo que los porta se denomina Heterocigota para ese rasgo. Entonces los tres posibles genotipos para un rasgo son: Homocigota dominante Homocigota recesivo Heterocigota2 Ej.: AA Ej.: aa Ej.: Aa Los fenotipos posibles para el rasgo "color de semilla" de arvejillas son: semilla verde semilla amarilla 2 Los individuos heterocigotos se nombran siempre anteponiendo el gen dominante. - 137 - Entonces: GENOTIPO FENOTIPO AA Semilla amarilla Aa Semilla amarilla aa Semilla verde Leyes de Mendel Gregor Mendel nació en 1822. Luego de recibir formación universitaria en física, matemática y ciencias naturales, inició en 1857 sus experimentos genéticos con plantas de arvejas de jardín o arvejilla (Pisum sativum) en los jardines del Monasterio de Agustinos de Brünn (Austria) donde era sacerdote. En 1866, Gregor Mendel publica “Experimentos sobre híbridos en plantas” en una revista de la Sociedad de Naturistas de Brünn. Las conclusiones a las que arriba y que son la piedra fundamental de la genética actual, fueron ignoradas por más de 30 años. Mendel no utilizó esta planta de forma azarosa sino porque presentaban características apropiadas para sus experimentos: El tiempo entre una generación y otra es corto (una estación de cultivo) Ocupan poco espacio y se puede trabajar con gran número de muestras (individuos) para poder hacer estudios estadísticos Normalmente la arvejilla se autofecunda y esto permite la formación de líneas puras. Presentan características observables (fenotípicas) bien definidas y diferenciales, entre las cuales Mendel eligió siete de ellas que eran caracteres unitarios y aparecían en forma constante. Por ejemplo: Color de flor (blanca / violeta). Forma de semilla (lisa / rugosa). Tallo (corto / alto). Vaina (amarilla / verde). Primera Ley de Mendel Cruza monohíbrida La primera ley de Mendel se refiere a la herencia de un carácter o rasgo tomado independientemente para su estudio (cruza monohíbrida) y predice el comportamiento del par de genes que gobierna dicho rasgo. Planteo del cruzamiento: - 138 - Especie: plantas de arvejillas Carácter a analizar: color de la semilla Se parte de dos individuos progenitores o generación parental, a los que supondremos provenientes de líneas puras u homocigóticas para este carácter. Se les asigna arbitrariamente los símbolos para identificarlos: Gametas: Una vez planteada la cruza, cabe preguntarse qué genes portarán las gametas de los respectivos progenitores. Para responder a esto recordemos que: Ambos alelos del par ocupan loci equivalentes en cromosomas homólogos La gametogénesis tiene lugar por división meiótica. En el siguiente esquema, recordamos la meiosis: - 139 - - 140 - Así, cada progenitor produce sólo un tipo de gametas. Lo representamos del siguiente modo: - 141 - Unión de gametas y obtención de los descendientes: Filial 1 (F1): Se procede a la unión de los genes que llevan las gametas, tal como ocurre en la fecundación. Así se obtiene la constitución genética de la F1. Resumiendo lo hasta aquí expuesto: Resultados del cruzamiento Se deben explicar el Genotipo y el Fenotipo de los descendientes. De esta cruza sólo es posible obtener individuos heterocigotas, por lo tanto amarillos. Entonces: Si ahora se cruzan dos individuos de la F1, el procedimiento es el mismo. Planteo de cruzamiento: Gametas: Dado que la constitución genética de ambos progenitores es la misma, analizaremos la gametogénesis una vez: - 142 - Cada padre origina dos tipos de gametas: Unión de gametas y obtención de los descendientes: Filial 2 (F2): Cualquier gameta de un padre tiene la misma probabilidad de fecundar a cualquier gameta del otro padre. Para expresar todos los cruzamientos potenciales, en este caso, es aconsejable distribuir las gametas en un tablero de Punnett: - 143 - a A A AA Aa a Aa aa Las intersecciones nos indicarán las distintas fecundaciones posibles, y, por lo tanto, los potenciales descendientes de esta cruza. Resultados del cruzamiento: Pueden expresarse en cualquiera de las siguientes formas: 25% AA; 50% Aa; 25% aa Proporción genotípica: ¼ AA; ½ Aa; ¼ aa 1 AA; 2Aa; 1 aa 75% amarillo; 25% verdes Proporción fenotípica ¾ amarillos; ¼ verdes 3 amarillos; 1 verde Éstas son las proporciones típicas de una F2 en cualquier cruza monohíbrida, y representan una probabilidad estadística a la cual se acercan los resultados reales cuando el número de cruzas es muy grande. Notar que en la F2 reaparece el fenotipo recesivo de la G.P. que había quedado enmascarado en la F1. Estos resultados fueron expresados por Mendel textualmente así: “… resulta claro que los híbridos forman semillas que tienen uno u otro de los dos caracteres diferenciales; y de estos, la mitad vuelve a desarrollar híbridos, mientras que la otra mitad produce plantas que permanecen constantes, ya sea porque reciben el carácter dominante o el recesivo en igual número”. Esta conclusión constituye la Primera Ley de Mendel o Principio de Segregación: “Los alelos recesivos enmascarados en la F1 heterocigota de un cruzamiento entre dos líneas puras (homocigotos), reaparecen en la F2 con una proporción específica (3/4: 1/4) debido a que los miembros de un pareja alélica, se segregan (separan) uno de otro, sin sufrir ninguna modificación, cuando un individuo heterocigoto forma gametas”. - 144 - Cruzamiento prueba: ¿Cómo podía saber Mendel, entre sus plantas, cuáles eran líneas puras y cuáles eran híbridos? Mendel resolvió este problema cruzando a la planta de genotipo desconocido con una planta homocigota recesiva para el color de la semilla. ¿Qué resultados se obtienen de esta cruza de prueba? Examinemos los dos casos posibles: 1) Si el individuo de genotipo desconocido es homocigota dominante, el 100% de la descendencia presentará el fenotipo correspondiente al gen dominante: 2) Si el individuo incógnita es heterocigota, el 50% de la descendencia tiene el fenotipo dominante (igual al progenitor incógnita) y el 50% tiene el fenotipo recesivo (igual al progenitor de prueba). Por lo tanto sólo aparecen individuos con el rasgo recesivo (aa) si el progenitor es heterocigota (Aa) y nunca si es homocigota dominante (AA). Cuando el homocigoto recesivo es uno de los progenitores, entonces se habla de Retrocruza, es decir un tipo particular de cruzamiento prueba. Segunda Ley de Mendel Cruza polihíbrida: En los cruzamientos anteriores hemos considerado la herencia de un solo par de genes. Sin embargo, los individuos no heredan los genes de uno en uno, sino que los heredan - 145 - todos al mismo tiempo. Se hace necesario entonces el estudio de cruzamientos en los que se tome en cuenta la transmisión simultánea de dos o varios pares de genes. A estas cruzas se las denomina dihíbridas, trihíbridas, etc. La segunda Ley también llamada Principio de Segregación Independiente, puede enunciarse en términos actuales como: “Los miembros de parejas alélicas diferentes se distribuyen o combinan independientemente unos de otros cuando se forman las gametas de un individuo híbrido para los caracteres correspondientes.” Siguiendo con el ejemplo de las arvejillas, ahora se analizan la herencia simultánea de los caracteres color y textura de la semilla, para comprender la Segunda Ley: Planteo del cruzamiento: G.P. fenotipos semillas amarillas lisas X semillas verdes rugosas G.P genotipos AA.BB X aa.bb Gametas: - 146 - Unión de gametas: F1: Resultado de la cruza : F1 Genotípicamente: 100% Aa.Bb (heterocigota para ambos caracteres). Fenotípicamente: 100% semillas amarillas y lisas. Cruzamiento entre dos individuos de la F1: obtención de la F2 Planteo del cruzamiento: F1 Aa. Bb X Aa.Bb Unión de gametas: AB Ab aB ab AB AA.BB* AA.Bb Aa.BB Aa.Bb* Ab AA.Bb AA.bb* Aa.Bb* Aa.bb aB AaBB Aa.Bb* aa.BB* aa.Bb ab Aa.Bb* Aa.bb aa.Bb aa.bb* *dobles homocigotas / *dobles heterocigotas - 147 - Resultado de la cruza Proporción genotípica de F2 Proporción fenotípica de F2 1/16 AA.BB 9/16 amarillas lisas 1/16 AA.bb 3/16 amarillas rugosas 1/16 aa.BB 3/16 verdes lisas 1/16 aa.bb 1/16 verdes rugosas 2/16 AA.Bb 2/16 Aa.BB 2/16 Aa.bb 2/16 aa.Bb 4/16 Aa.Bb A pesar de que los caracteres color y textura se heredan simultáneamente y producen una razón fenotípica 9:3:3:1, considerándolos separadamente la razón es 3:1. Los caracteres se heredan independientemente. Genética humana Las enfermedades genéticas son aquellas en las que una o varias alteraciones en el material genético contribuyen de manera significativa a las manifestaciones clínicas. En las últimas décadas el conocimiento de la genética humana creció enormemente. Tanto la citogenética como la genética molecular permitieron perfeccionar métodos de diagnóstico que hoy hacen posible confirmar la presencia de una determinada enfermedad en personas con síntomas clínicos e incluso detectar alelos afectados capaces de desencadenar una enfermedad en una persona sana pero portadora de ese alelo, quienes a su vez pueden transmitirlo o no a su descendencia. Se pueden realizar detecciones de anomalías en fetos en desarrollo e identificar en recién nacidos, patologías que pueden ser tratadas en edad temprana. En los últimos años, se hizo que ciertas enfermedades hereditarias son más frecuentes en algunos grupos étnicos que en otros. El campo de la genética es muy amplio. También se realizan estudios para la identificación y filiación de personas con gran certeza. - 148 - Las enfermedades originadas por alteraciones en el material genético, pueden clasificarse en tres categorías principales: Enfermedades cromosómicas. Cuando ocurre un exceso o una deficiencia de material cromosómico. Ejemplos: Síndrome de Down (o trisomía 21), Síndrome de Turner, Síndrome de Edwards. Enfermedades monogénicas. Cuando ocurre una mutación3 en un único gen. Ejemplos: Talasemia, Fibrosis quística, Anemia falciforme, Enfermedad de Huntington, Albinismo. Enfermedades multifactoriales. Ocurren mutaciones en varios genes, los cuales interactúan entre sí y también con factores ambientales. Ejemplos: Cáncer de mama, Hipertensión, Hipercolesterolemia. (Más adelante se verán en forma más detenida, algunas alteraciones cromosómicas.) Ingeniería genética Es una rama dentro de Biología que permite manipular la información de un organismo y en consecuencia, los productos de su expresión génica. En la actualidad se realizan muchas investigaciones desde este campo para encontrar una solución a los problemas de un mundo globalizado: enfermedades como el cáncer y el SIDA, el incremento de la población junto a la disminución de los espacios destinados a la agricultura o el aumento de los desechos tóxicos por parte de las industrias. Diferentes técnicas y herramientas moleculares son utilizadas para beneficio del Hombre. Unas de las más difundidas seguramente son las de clonación por un lado y la obtención de animales y plantas transgénicas por otro. Los beneficios que se obtienen son muchos. Lograr animales transgénicos como modelo para investigaciones en medicina y el estudio de enfermedades. También como biorreactores, es decir como sistemas capaces de ofrecer algún producto biológico, generalmente se utilizan vacas, ovejas, cerdos y aves. Muchos animales de granja transgénicos poseen mejores características nutricionales como por ejemplo la leche con mayor aporte de proteínas. En el caso de las plantas transgénicas tiene una importante aplicación en cuanto a nivel productivo. Así se obtienen plantas que son resistentes a ciertos herbicidas, el ejemplo más conocido es la soja transgénica y su resistencia al herbicida glifosato. Otras especies de importancia agrícola son resistentes al ataque de insectos, de virus, o a condiciones ambientales extremas como sequía, alta salinidad, bajas temperaturas. 3 Mutación: cambio en la información genética; cambio heredable en la secuencia del ADN de un cromosoma. - 149 - 3. HERENCIA NO MENDELIANA No todas las características se heredan de una manera tan simple como el color de las semillas de arvejilla. Hasta ahora hemos analizado que la herencia simple, en la que solamente interviene un par de genes, es la excepción y no la regla. En las páginas anteriores dijimos que un cromosoma contiene a muchos genes. En realidad, los cromosomas son grupos de genes “ligados” (unidos) que se comportan como unidades hereditarias; por lo tanto, todos los genes de un mismo cromosoma tienden a heredarse juntos. Si dos pares de genes pertenecen al mismo par de cromosomas homólogos: ¿segregarán en forma independiente uno de otro? No, no siempre es así. Al pertenecer al mismo par de cromosomas homólogos tienden a transmitirse juntos, en otras palabras, la segregación de dos o más pares de genes que forman parte del mismo par de cromosomas homólogos no es independiente. Cuando ocurre esto la proporción observada es diferente de la clásica 9:3:3:1 de la Segunda Ley de Mendel, que vimos y analizamos. Se dice entonces que los genes están ligados y se heredan en bloque. Lo mismo ocurre cuando hay crossing over, recordar las etapas de la Meiosis, el cual es un mecanismo que introduce variabilidad y genera un resultado diferente a los trabajos de Mendel. Entonces ¿Mendel se equivocó? No fue así. El material que Mendel seleccionó para trabajar arrojó resultados que concordaban con lo que sus hipótesis le hacían esperar. Pero con otros materiales no se puede llegar a los mismos resultados. El crossing over anula los efectos del ligamiento. Cuanto mayor es la distancia entre dos genes de un cromosoma, mayor es la frecuencia de crossing over entre ellos. El entrecruzamiento, como se vio, es una fuente de variabilidad genética. - 150 - Hoy, entonces, se considera que existen genes que se comportan respondiendo a la herencia mendeliana, mientras que otros quedan incluidos en la herencia NO Mendeliana, tal es el caso del ligamiento y crossing over, como así también los casos que se describirán a continuación: Dominancia incompleta Alelos múltiples Codominancia Herencia ligada al sexo Dominancia incompleta No se necesita que una característica sea completamente dominante sobre la otra. Muy a menudo la combinación de genes diferentes tiende a producir grados variables de dominancia parcial o incompleta; en este caso, el fenotipo del heterocigota resulta diferente del de ambos homocigotas. En el cruzamiento de cierto tipo de ganado se ve un ejemplo de dominancia incompleta. Si un animal rojo se cruza con una blanco, se produce un animal de un color intermedio, un ruano; pero cuando dos miembros de esta generación se cruzan, los rasgos empiezan a segregarse de nuevo, mostrando que los alelos mismos, como opinaba Mendel, permanecen inalterados, dando una razón de uno rojo a dos ruanos a uno blanco. La dominancia parcial o incompleta es aquella condición en la cual un gen dominante no logra imponer su expresión en forma total en el heterocigota. Nótese, en el siguiente esquema, que si la piel roja fuese dominante sobre la blanca (o viceversa), se obtendría una razón de 3 rojos a 1 blanco. - 151 - Alelismo múltiple y Codominancia Hasta ahora vimos ejemplos de genes que se presentan bajo dos formas alternativas distintas, en la mayoría de los casos un alelo dominante y otro recesivo. Sin embargo muchos genes presentan tres o más formas alternativas que pueden ocupar el mismo locus, cada uno de los cuales determina un fenotipo distinto. Se dice que estos genes presentan alelos múltiples. Un ejemplo sencillo de alelos múltiples es el de los grupos sanguíneos en el Hombre, que además presenta codominancia, es decir que el fenotipo del heterocigota está determinado por la expresión individual de ambos genes. Ej.: grupo sanguíneo AB. Este sistema consiste de tres posibles alelos para el par de genes que controla el grupo sanguíneo: Alelo SA determina el grupo sanguíneo A y es dominante frente al alelo s que, en homocigosis, determina el grupo sanguíneo O. El alelo SB determina el grupo sanguíneo B y también es dominante frente al alelo s. Entre los alelos SA y SB, se manifiesta una relación de codominancia, como se dijo, ambos alelos se expresan fenotípicamente por igual y el grupo sanguíneo resultante, de un individuo de genotipo SA SB, se denomina AB. - 152 - Entonces queda determinado el Sistema ABO: Genotipos Homocigota Dominante SA SA Heterocigota SA s Homocigota Dominante SB SB Heterocigota SB s Codominancia SA SB Homocigoto Recesivo ss Fenotipos (grupos sanguíneos) Grupo A Grupo A Grupo B Grupo B Grupo AB Grupo O Herencia ligada al sexo En la especie humana, los genes "diferenciadores" del sexo se encuentran en cromosomas particulares: los cromosomas sexuales, gonosomas o alosomas. El par sexual puede estar constituido por: Cromosomas del par sexual Sexo Dos cromosomas X = XX Femenino Un cromosoma X y un cromosoma Y = XY Masculino El sexo de un individuo queda determinado en el momento de la fecundación, dependiendo del cromosoma sexual que aporta el espermatozoide (X ó Y), ya que el óvulo siempre aporta un X. Los cromosomas X e Y son morfológicamente diferentes y esto crea un mecanismo hereditario particular, para ciertos caracteres, como se verá a continuación. El cromosoma Y es bastante más pequeño que el cromosoma X, lo que indica que el cromosoma X contiene varios genes que no están presentes en el cromosoma Y. - 153 - Uno de los brazos del cromosoma X es homólogo del brazo mayor del cromosoma Y, es decir ambos brazos portan genes para las mismas características, y son los que permiten el apareamiento de los dos cromosomas durante la meiosis. Los brazos restantes de ambos cromosomas son de tamaño diferente y se sabe que parten genes particulares y exclusivos de cada uno, de ahí el nombre de brazos heterólogos. La herencia de los genes ubicados en los brazos homólogos de los cromosomas X e Y es independiente del sexo; en cambio, la herencia de los genes ubicados en los brazos heterólogos está ligada al sexo. Es conocido que el daltonismo (ceguera al color) y la hemofilia (trastorno en la coagulación de la sangre) son enfermedades genéticas hereditarias. Su transmisión hereditaria está ligada al sexo, es decir está ligada al cromosoma X debido a que los genes responsables de ambas enfermedades están ubicados en el brazo heterólogo del cromosoma X, y por lo tanto, no se encuentra en el cromosoma Y. Los alelos responsables de ambas enfermedades son recesivos frente a los alelos normales. 4. ALTERACIONES CROMOSÓMICAS Vimos que la constancia en el número de cromosomas es fundamental para que un organismo se desarrolle normalmente. Las alteraciones cromosómicas consisten en el exceso o en la deficiencia en la cantidad de material genético, ya sea por cambios en la estructura del cromosoma o en su número. Estos cambios son visibles en los estudios cromosómicos que se realizan a partir del análisis de los cromosomas observados con un microscopio óptico. Estos estudios se denominan cariotipo. El cariotipo es la identificación y ordenamiento sistemático de los pares cromosómicos. - 154 - Alteraciones estructurales humanas Translocaciones: Es el intercambio de material genético entre cromosomas no homólogos. Translocaciones recíprocas: Cuando las roturas ocurren en dos cromosomas diferentes y se produce un intercambio mutuo de material genético. Los portadores de este tipo de translocación suelen tener fenotipos normales sin embargo su descendencia puede presentar una trisomía parcial o una monosomía parcial y un fenotipo anormal. Translocaciones Robertsonianas: Se producen cuando los brazos largos de dos cromosomas acrocéntricos (cromosomas 13, 14, 15, 21 y 22) no homólogos se unen por el centrómero y los brazos cortos se pierden. - 155 - Deleciones: Son roturas cromosómicas con la consiguiente pérdida de material genético. Son clasifican en terminales e intersticiales. Ejemplos: Síndrome del maullido del gato (Cri du Chat) Descrito por el característico llanto del niño, A medida que el niño crece, el llanto suele hacerse menos obvio, lo que dificulta el diagnóstico. Se debe a una deleción del brazo corto del cromosoma 5. Síndrome de Wolf – Hirschhorn: Se debe a una deleción distal del brazo corto del cromosoma 4. - 156 - Duplicaciones: Son excesos de material genético y suelen tener consecuencias menos graves que las deleciones. Inversiones: Es el resultado de dos roturas en un cromosoma, seguidas por la reinserción del fragmento perdido en su lugar de origen, pero en sentido inverso. Pueden ser Pericéntricas (si incluye el centrómero) o Paracéntricas (si no incluye el centrómero). - 157 - Alteraciones numéricas humanas Estas alteraciones numéricas se producen durante la meiosis, cuando los miembros de los pares de cromosomas homólogos se separan, de manera que las células hijas reciben la mitad de los cromosomas que presenta la célula madre. Si en lugar de separarse, los miembros del par se desplazan juntos hacia el mismo polo celular (falta de disyunción) las células resultantes presentarán un desbalance cromosómico; la mitad tendrá cromosomas de más y la otra mitad cromosomas de menos. Entonces al fusionarse una gameta normal con una desbalanceada (con cromosomas de más o de menos), la cigota resultante presentará una alteración cromosómica numérica Algunos conceptos para una mejor comprensión: Euploidías: Cuando una célula contiene en su núcleo un número de cromosomas múltiplo de 23 (n). Haploidías: Cuando n = 23 Diploides: Cuando 2n = 46 (número normal de cromosomas en la especie humana) Poliploides: Cuando 3n = 69, 4n = 92, etc. Aneuploidías: cuando las células no contienen en su núcleo un número de cromosomas múltiplo de 23 y presenta ganancia o pérdida de cromosomas. Las principales son las monosomías y trisomías, que generalmente están causadas por ausencia de disyunción durante la Meiosis. - 158 - Monosomías: Síndrome de Turner: (XO) Estos individuos carecen del cromosoma sexual Y; es decir que son XO y por lo tanto tienen 45 cromosomas. Su apariencia es de mujer; son estériles (no tienen ovarios o los tienen poco desarrollados). Presentan pechos pocos desarrollados, crecimiento general inferior a lo normal y usualmente retraso mental. Una de cada 5000 mujeres sufre este síndrome. Trisomías: Síndrome de Down (cromosoma autonómico 21) Estos individuos presentan un cromosoma autonómico de más, perteneciente al par 21 (trsomía del cromosoma 21) y tienen por lo tanto 47 cromosomas. Probablemente es la anormalidad congénita más común, con una frecuencia de uno cada 600 nacimientos. Este síndrome produce retraso mental, cuerpo corto con dedos gordos, lengua más grande de los normal, pliegue del párpado parecido al de las razas mongólicas. - 159 - SÍNDROME DE EDWARDS (Cromosoma 18) - 160 - SÍNDROME DE PATAU (Cromosoma 13) SÍNDROME XXX SÍNDROME DE KLINEFELTER: (XXY) Estos individuos poseen un cromosoma sexual X de más, es decir que son XXY y por lo tanto tienen 47 cromosomas. - 161 - Tienen apariencia de varones, con órganos sexuales poco desarrollados, son estériles (no producen esperma); pechos más desarrollados de lo común en el hombre, frecuentemente restrasados mentales. Uno de cada 500 hombres sufre este síndrome. - 162 - BIBLIOGRAFIA Alberts, B; Watso, J; Bray, D; Lewis, J; Raff, M; Roberts, K. Biología Molecular de la Célula; 3ª Edición; Ediciones Omega S.A. Barcelona. 1996. Alberts, B et al. Molecur Biology of the cell. 4th Edition. Garland Publishing. 2002. Baker y Allen. Biología e Investigación Científica. Fondo Educativo Interamericano. Castro, R; Handel, M; Rivolta, G. Actualizaciones en Biología. 8ª Edición. Editorial Universitaria de Buenos Aires. 1990. Curtis, H; Barnes, N. Biología. 7ª Edición. Editorial Médica Panamericana. Buenos Aires. 2008. Curtis, H; Barnes, N. Invitación a la Biología. Editorial Médica Panamericana. Buenos Aires. 1987. Lacadena, J. R. Genética. Editorial A.G.E.S.A. 1976. Rieger, R; Michaelis, A; Green, M. M. Glossary of Genetics and Cytogenetics. Editorial Springer, Verlag, Berlin Heildelber. New York. 1976. Saez, F. A; Cardoso, H. Citogenética básica y biología de los cromosomas. Serie de monografías de O.E.A. 1978. Satansfield, W. Genética. 3º Edición. Editorial Mc Graw Hill Interamericana. México. 1992. Senoc – Promec. Biología. Módulo 3. Strickberger, M. Genética. Editorial Omega. 1974. Solomon, E; Ville, C; Davis, P. Biología. Editorial Mc Graw Hill Interamericana. México. 1987. Solomon, E; Ville, C; Davis, P. Biología. 4º Edición. Editorial Mc Graw Hill Interamericana. México. 1998. Weisz, P. La Ciencia de la Biología. Editorial Omega. Welch, C et al. Ciencias Biológicas (de las moléculas al hombre). Cia. Editorial Continental S.A. de C.V. México. - 163 - MÓDULO 4 EVOLUCIÓN Y DIVERSIDAD BIOLÓGICA 1. TEORÍA DE CHARLES DARWIN La enorme diversidad de seres vivos existentes sobre la Tierra llamó la atención del Hombre desde épocas tempranas. De las múltiples teorías formuladas para explicar esa diversidad, hoy en día quedan solamente aquellas que se ubican dentro del término general de la evolución, es decir, las teorías evolutivas. Evolución significa cambio gradual, continuo y ordenado en el tiempo. Puede definirse biológicamente como un cambio en el tiempo de la diversidad y adaptación de los seres vivos. Originalmente, la idea de evolución se oponía a las hipótesis de cambios catastróficos, como sostenía Cuvier, con creaciones especiales de nuevas formas de vida cada cierto tiempo. La mayor disponibilidad de información respecto de los organismos que nos han precedido en la Tierra permitió mostrar que los cambios habían sido relativamente continuos y graduales Las teorías evolucionistas se distinguieron por los diferentes mecanismos propuestos para explicar los cambios en el tiempo. El primer intento científico y coherente de explicar cómo opera la evolución, se debe al naturalista francés Jean B. De Lamarck: La Teoría de Lamarck La teoría de Lamarck, propuesta en 1809, se basa en las siguientes premisas: Los organismos son guiados a través de su existencia por una fuerza innata y misteriosa que les permite sobreponerse a la adversidad del medio. El medio ambiente actúa como una "fuerza modificadora" sobre los organismos, imponiendo necesidades que hacen surgir nuevos órganos y funciones. El "uso y desuso de las partes": el desarrollo de los órganos y su actividad están en relación constante con el uso que de ellos se haga. Si un órgano es utilizado, entonces crece y se hace más eficiente. De lo contrario, puede degenerar y atrofiarse. La herencia de los caracteres adquiridos: lo que se ha adquirido, impreso o modificado en la organización de los individuos durante el curso de su vida, es conservado y trasmitido a sus descendientes. - 164 - Un ejemplo hipotético para comprender lo propuesto por Lamarck: Supóngase que un determinado animal ancestral con el cuello corto se alimentaba con hojas de árbol. A medida que consumía las hojas inferiores del árbol estiraba el cuello para alcanzar las de más arriba. Debido a este estiramiento continuado a lo largo de toda la vida, el cuello se volvió algo más largo y entonces los descendientes heredaban un cuello algo más largo. Éstos a su vez se alimentaban de las hojas de los árboles y seguían estirando sus cuellos; y esto seguiría durante muchas generaciones. Con el tiempo llegó a formarse un animal con el cuello muy largo, semejante a una jirafa actual. Esta teoría (en algunos aspectos incontrastable) tuvo gran éxito y contribuyó a la expansión de la idea de la evolución. Pero con el tiempo resultó insostenible. Es correcto que el uso y desuso conduce a la adquisición de caracteres. Por ejemplo, se sabe que el ejercicio desarrolla músculos poderosos. Sin embargo Lamarck se equivocó al suponer que estas variaciones adquiridas (no genéticas) son heredables. Los caracteres adquiridos no son heredables ya que son efectos producidos por el ambiente y el desarrollo, no por los genes. Sólo son heredables los caracteres genéticos y sólo si estos caracteres están regulados por genes presentes en células reproductoras. Lo que ocurra a otras células que no sean las gametas, a causa del uso y desuso, o por otras causas, no afecta a los genes de las gametas. Por consiguiente, a pesar de que Lamarck observó los efectos del uso y desuso correctamente en algunos casos, estos efectos no pueden tener un papel en la evolución. Darwin y Wallace, en 1859, propusieron que la producción de nuevas formas se debía al fenómeno de variación al azar (también sostuvieron, como Lamarck, la herencia de los caracteres adquiridos); reconocieron la acción fundamental del proceso de selección natural, y la importancia del aislamiento reproductivo entre poblaciones como mecanismo de diversificación. La carencia de una teoría de la herencia hizo que la idea básica de Darwin quedara expuesta a fuertes críticas. Finalmente, el redescubrimiento de las leyes de Mendel (recordar que vimos que fueron ignoradas por más de 30 años) y más tarde el reconocimiento de los fenómenos citológicos y moleculares en la formación de las gametas, contribuyeron a la formulación de la Teoría Moderna o Teoría Sintética de la Evolución. La Teoría de Darwin Como adelantáramos, Darwin y Wallace juntos enunciaron una nueva teoría que sustituyó a la de Lamarck. Darwin también elaboró la nueva teoría en forma de libro. Este famoso trabajo, titulado "Sobre el origen de las especies mediante selección natural o la conservación de las razas favorecidas en la lucha por la vida" fue publicado en 1859. En esencia, la teoría de Darwin se basa en algunas observaciones, y en conclusiones extraídas a partir de ellas. Darwin sabía que la superproducción de descendientes es la regla general en la naturaleza y que sólo unos pocos suelen sobrevivir. Entonces se pregunta: ¿En qué se diferencian los supervivientes de los que perecen? ¿Tienen características distintas, o es el azar el que determina los que sobreviven y los que mueren precozmente? ¿Es puro - 165 - azar o se trata de un proceso dirigido cuyas causas pueden ser determinadas? Muchos son presa de sus enemigos, otros no consiguen suficiente alimento, no encuentran refugio o, sencillamente, son desbancados por sus propios congéneres. Pero ¿y los que sobreviven? ¿Acaso son más veloces, están más camuflados que los demás, son menos vulnerables a las enfermedades, aprovechan mejor su alimento, o tienen mejor olfato y pueden detectar al enemigo, o a sus presas, antes que otros miembros de su misma especie? Cuando la numerosa descendencia se enfrenta a las condiciones del medio ambiente, generalmente serán los "mejores", los "más aptos" los que logren sobrevivir. En esto consiste precisamente la " lucha por la existencia" -como había sido denominada ya por Malthus- denominación que fue adoptada por Darwin y que siempre se prestó a malentendidos. La situación queda mejor descrita por la expresión "supervivencia de los más aptos", de los mejor adaptados al entorno. Así pues, Darwin observa que en la naturaleza se produce una selección análoga al proceso de la selección artificial que efectúa el criador de animales; la selección de los más aptos, de los más capacitados, que está determinada por las condiciones ambientales: es decir que existe una selección natural. Pero para que pueda tener lugar tal selección es necesario que se dé una superproducción de seres vivos. Naturalmente, la selección sólo podrá darse si existen diferencias en las características de los descendientes de una misma especie; pero esto es algo que se da frecuentemente en todas las especies. Unos individuos resultan ser ligeramente más grandes, otros más pequeños; unos presentan una pigmentación más clara y otros más oscura; algunos pueden correr más rápidamente que el promedio de sus congéneres; algunas plantas presentan más ramificaciones que otras; este animal es más miedoso, aquél más agresivo... Estas diferencias, que Darwin denomina variaciones, pueden poner al ser vivo en situación ventajosa o desfavorable. En el último caso no estará en condiciones de superar las dificultades que le presente su entorno o de afrontar la competencia de otros seres vivos, por lo que pasará a formar parte de aquellos que sucumban antes de tiempo. En cambio, las variantes favorables que mejoran la adaptación al medio de un ser vivo, permiten su supervivencia, su elección en el proceso de selección natural. Los individuos que sobreviven dan origen a la próxima generación y de este modo las variaciones útiles se trasmiten a dicha generación y a las subsiguientes. Con el transcurso de muchos años de selección natural, los descendientes posteriores pueden resultar bien distintos de sus antecesores; esta diferencia puede ser suficientemente pronunciada como para que se los considere especies separadas. Así, la selección natural se constituye en el principal agente del cambio evolutivo. Aunque han pasado casi 150 años desde la publicación de El Origen de las Especies el concepto original de Darwin constituye todavía el marco global de comprensión del proceso y se puede resumir en un marco conceptual basado en cinco premisas: Los organismos provienen de organismos similares a ellos. En otras palabras, hay cierta estabilidad en el proceso de reproducción. En la mayoría de las especies, el número de descendientes que sobreviven y se reproducen en cada generación es menor que el número inicial de descendientes. En cualquier población existen variaciones entre individuos y algunas de estas variaciones son heredables. El número de individuos que sobreviva y se reproduzca dependerá de la interacción entre las variaciones heredables individuales y el ambiente. Algunas - 166 - variaciones capacitan a ciertos individuos para vivir más tiempo y dejar mayor descendencia que otros, en un ambiente determinado. Darwin llamó a estas variaciones favorables y sostuvo que éstas tendían a ser cada vez más frecuentes de una generación a la otra. Dado un tiempo suficiente, la selección natural, actuando sobre dos poblaciones de organismos de una misma especie, puede producir una acumulación de cambios tal que esas poblaciones terminen constituyendo dos especies diferentes. Uno de los ejemplos mejor estudiados de selección natural es el de una polilla (Biston betularia) que habita en bosques de abedules. Estas polillas eran bien conocidas por los naturalistas británicos del siglo XIX. Estas polillas de color claro se posaban sobre los troncos de abedules que a su vez estaban recubiertos de líquenes, lo que favorecía a las polillas porque parecían invisibles y así escapaban de los pájaros predadores de estas polillas. En 1845, todas las polillas eran de color claro pero en ese año fue capturada una especie de color negro en un centro industrial de Manchester. Por esta época se inicia la revolución industrial en Inglaterra y las partículas de humo provenientes de la quema de carbón para hacer funcionar las máquinas industriales, contaminaban el ambiente. Comienzan a morirse los líquenes que recubrían los troncos de abedules. En los pueblos cercanos a estas industrias, todo se cubría de hollín, los árboles, las rocas y hasta el suelo se teñían de negro. Durante este tiempo, se encontraban más polillas negras que claras. ¿Por qué? Las polillas claras se mimetizaban en los troncos con líquenes y estaban más seguras. Cuando desaparecen los líquenes y los troncos se cubren de hollín, estas mismas polillas claras quedaban más expuestas y eran fácilmente visibles a los pájaros que las predaban. Así las polillas negras ahora eran invisibles, podían escapar de las aves, reproducirse y dejar mayor descendencia. Surge ahora la pregunta ¿De dónde provienen las polillas negras? El color negro era el resultado de una mutación rara y recurrente, de modo que las polillas negras siempre habían estado ahí pero en cantidades pequeñas. Cuando los troncos estaban cubiertos de líquenes, las polillas negras eran fácilmente visibles y en consecuencia predadas por las aves. Cuando desaparecen los líquenes y los troncos se cubren de hollín, se revierte la situación. Así en la década de 1950 sólo se podían observar algunos individuos de color claro en zonas que estaban muy lejos de los centros industriales. Es importante, con este ejemplo, entender que la polilla negra no es superior en sí misma a la clara o viceversa. La adaptación entonces, como comprendió Darwin, es una cuestión de tiempo y de lugar. - 167 - Origen y Evolución de las jirafas según las ideas de Lamarck (izquierda) y Darwin (derecha) Las jirafas primitivas de cuello corto y patas de Entre las antiguas jirafas se daban algunas longitud normal, intentaban constantemente -sobre diferencias en cuanto a la longitud de sus cuellos y todo en épocas de sequía- ramonear las verdes hojas patas; estas variaciones se transmiten de los árboles. A causa del continuo estiramiento del hereditariamente. cuello y las patas estos miembros comenzaron a alargarse. Sus descendientes empezaron a nacer con el cuello y Las jirafas más altas de cada generación disfrutaban las patas más largos, y el ejercicio y "afán" de de la ventaja de alcanzar más alimentos que las estirarse hizo que sus miembros continuaran pequeñas, por lo que se reproducían mucho más estirándose cada vez más. intensamente. - 168 - Debido a este continuo estiramiento, llegaron con el tiempo a convertirse en las jirafas que conocemos hoy día con sus cuellos y patas largos. Con el tiempo la selección natural favorece a los ejemplares de cuellos y patas largos. 2. ENFOQUE EVOLUTIVO La articulación de la Teoría de la Evolución de Darwin con los principios de la genética mendeliana se conoce como la Síntesis neodarwiniana o la Teoría Sintética de la Evolución. Durante los últimos 60 años, la teoría sintética ha dominado el pensamiento científico y ha sido fuente de nuevas ideas y líneas experimentales de investigación. Los principales artífices de la teoría Sintética fueron, junto con Huxley, Theodosius Dobzhansky, Ernest Mayr, Sewell Wright, Ledyard Stebbins, George Gaylord Simpson y Bernard Rensch. El Mecanismo Evolutivo según La Teoría Sintética Los cambios en el curso de la evolución ocurren sobre los organismos individuales. Sin embargo, sólo son observables en el seno de los grupos y en el transcurso de sucesivas generaciones. Por esto, los cambios que importan a la evolución son aquéllos heredables y la unidad de evolución no es el individuo sino la población. Se entiende por tal a una asociación de individuos de la misma especie que coexisten en un hábitat y, lo más importante, que pueden cruzarse entre sí, son potencialmente interfértiles. Dada esta potencial interfertilidad o panmixia (del griego pan: todo y mixis: mezcla) los genes de - 169 - los individuos que conforman una población fluyen libremente entre sus integrantes mediante el proceso de reproducción sexual. La sumatoria de los genes de los individuos que componen una población es el patrimonio génico (llamado asimismo pool, complejo, reservorio o acervo génico) de la misma. El pool génico es una abstracción: obviamente, los genes de un pool se hallan en diferentes combinaciones de alelos, constituyendo genotipos de individuos particulares. Las Leyes de Mendel nos explican la forma en que estos alelos se transmiten en la reproducción sexual, determinando el genotipo de cada individuo. Siendo que el fenotipo –es decir las características morfofisiológicas y etológicas de un organismo- está determinado en gran medida por su genotipo, la herencia se comporta hasta cierto punto como un factor conservador, que tiende a mantener las generaciones sucesivas dentro de un esquema común. Pero en poblaciones naturales, el pool génico se ve enriquecido en cada generación por nuevas variantes génicas. Esta variación genética es el producto de la mutación, que introduce nuevos alelos y de la reproducción sexual, que favorece la recombinación de los genes existentes, creando diversos genotipos. Así, la existencia de variación genotípica (sumada a factores exógenos) acarrea la variación fenotípica. Como señalara Darwin, entre las “variaciones individuales” algunos fenotipos resultan más ventajosos que otros frente a determinadas condiciones ambientales. Éstos serán los “elegidos”, los favorecidos por la selección natural. En efecto, es el medio quien da la medida del grado de adaptación de cada fenotipo. Pero, ¿qué se entiende por aptitud en términos evolutivos? La aptitud no es sólo la capacidad de confrontar exitosamente con las dificultades del ambiente, sino gracias a ello y además de ello, alcanzar la posibilidad de reproducirse. Esto significa que la generación siguiente heredará los alelos determinantes del fenotipo seleccionado. En consecuencia, dichos alelos –en tanto las condiciones del medio no varíen- irán aumentando su frecuencia en el acervo génico de la población. (Recordar el ejemplo de la polilla en Inglaterra para comprender cómo actúa la selección natural sobre una población.) A continuación, se analizan los mecanismos propuestos por la Teoría Sintética de la Evolución: 1) Variación Hereditaria: Todos los organismos dejan descendientes que son ligeramente distintos de ellos. Esta diversidad en los descendientes que provienen de reproducción sexual, se origina por diversos mecanismos: a) Recombinación cromosómica: al formarse las gametas en la meiosis, cada par cromosómico se separa independientemente de los otros, es decir, cada gameta lleva una proporción impredecible de cromosomas “maternos” y “paternos” y es muy poco probable que se formen gametas idénticas. b) Recombinación génica: también durante la meiosis, el proceso de entrecruzamiento o crossing over permite que los cromosomas homólogos intercambien información. Como consecuencia, no hay cromosomas netamente “maternos” o “paternos”, sino que prácticamente todos serán recombinantes. - 170 - Estos dos mecanismos llevan a la formación de gametas distintas entre sí y distintas de las producidas por la generación anterior. Luego, cuando las gametas se unen por fecundación, actúa un tercer mecanismo que introduce variación: c) Recombinación gamética o recombinación sexual: cada individuo se origina por la recombinación de dos juegos de información, uno paterno y otro materno; de acuerdo a la forma en que se comporta dicha información, el organismo no se parecerá estrictamente a ninguno de los dos progenitores. Así la reproducción sexual da origen a descendientes casi siempre distintos entre sí y distintos de los que le dieron origen. Los mecanismos vistos hasta acá, recombinan genes ya existentes en la población. Si se analiza la disponibilidad de genes (para cierta característica) en la población entera, se comprueba que hay procesos por los cuales aparecen nuevos genes: d) Mutación: es la alteración al azar de la información que llevan los genes. La mutación puede afectar los genes de diversas maneras y en una extensión variable. Es un mecanismo que genera información heredable nueva. La frecuencia de mutación puede ser modificada por diversas influencias, como la presencia de ciertos otros genes, la aplicación de radiaciones o de sustancias químicas. Pero la naturaleza de esta nueva información es aleatoria, no está determinada por estas influencias. La información nueva no es necesariamente positiva para un individuo, también puede resultar perjudicial o neutra. e) Inmigración: entrada a la población de individuos de la misma especie que aportan genes que antes no se encontraban. Los genes existentes también pueden desaparecer de la población mediante: f) Emigración: salida de individuos que son portadores únicos de un gen particular. g) Deriva Génica: cuando un grupo reducido de individuos queda separado de una población grande, la parte del pool genético que pasa a la nueva población puede o no ser representativa de la original (tal vez algunos alelos no estén presentes y otros se hallen representados en exceso).Por otro lado el reducido tamaño de la población conduce al incesto, por lo que los alelos - que llegaron a ella por azar – tienden a recombinarse en forma homocigótica. Debe destacarse que es importante no solo que aparezcan genes nuevos, circunstancia poco frecuente, sino que se forman nuevas combinaciones de genes. 2) Selección Natural: Como vimos anteriormente, debido a los procesos de variación hereditaria, se observa que los individuos presentarán una gran gama de fenotipos diferentes. - 171 - Algunos de estos rasgos serán adaptativos y otros no. Recordemos que los caracteres adaptativos son aquellos que les permiten a los individuos sobrevivir y dejar mayor descendencia. Las condiciones del ambiente se llaman presiones de selección. Las ventajas adaptativas pueden ser, entre otras: Morfológicas: por ejemplo un color o una forma particular que permitan que un animal se oculte más fácilmente de sus predadores, como vimos en el ejemplo de las polillas claras y negras. Fisiológicas: por ejemplo, la adquisición de una enzima que permita mejorar la eficiencia metabólica a bajas temperaturas. De comportamientos: por ejemplo, la adquisición de hábitos nocturnos (o diurnos) que mejoren la situación de competencia por el alimento. Esta “lucha por la existencia” no se desarrolla de una forma brutal y cruel, se trata más bien de un proceso estadístico en el que algunos individuos dejan más descendencia que otros. Este proceso se denomina reproducción diferencial y en ello consiste la concepción actual de la selección natural. Reformulando el concepto de aptitud decimos que está determinada por la capacidad de propagar los alelos propios en las generaciones siguientes, a expensas de otros alelos diferentes. Entonces, según este punto de vista, el organismo mayor y más poderoso de una población podría ser muy “apto” en cuanto a comportamiento, pero si fuera estéril, carecería de importancia en el aspecto reproductor y, por lo tanto, en el evolutivo. Recíprocamente, un organismo débil, enfermo, podría ser “inepto” en comportamiento, pero tener una descendencia numerosa. Claro que, generalmente (aunque no necesariamente) los que dejan más descendientes son los organismos mejor preparados para sobrevivir en su ambiente. Por lo tanto, la reproducción diferencial tiende a imponer en la población la información genética de los fenotipos más ventajosos. Para ilustrar la forma en que interactúan los distintos agentes que participan del proceso evolutivo vistos hasta ahora, describiremos un caso de evolución reciente que fue observado por el hombre: El DDT y las moscas En 1948, el químico suizo Paul A. Muller recibió el premio Nobel por el descubrimiento del DDT. Este insecticida provocó euforia por su efectividad, aunque ésta fue efímera, pues pronto los insectos se hicieron inmunes a su acción. ¿Qué había ocurrido? Se descubrió que entre los insectos libres se daban casos de mayor o menor resistencia al insecticida (variación fenotípica). Es decir que frente a la presión selectiva ejercida por el DDT, algunos individuos resultaban más aptos. - 172 - Por otro lado, experimentos efectuados con la mosca de la fruta demostraron que los rasgos que conferían resistencia eran heredables; estaban inscriptos en el genotipo y existía variación genética para ellos. Dichos rasgos se transmitían, en principio, en forma independiente. Ellos son: Capa de quitina tres veces menos permeable al DDT que en otros insectos. Presencia de enzimas que les permiten transformar al DDT, muy rápidamente, en un compuesto inocuo. Mayor cantidad de grasa corporal donde se acumula el DDT sin que cause daño. Mayor tamaño, por ende soportan dosis mayores. Comportamientos tales como abandonar las casas a la noche (cuando habitualmente se fumiga) y evitar las superficies verticales (que son las más rociadas). Los individuos que no presentaban ninguno de estos rasgos perecieron en las primeras fumigaciones: fueron eliminados por selección natural. Los que tenían alguno de ellos, en cambio, sobrevivieron y se cruzaron entre sí para dar origen a la siguiente generación (reproducción diferencial), que heredó así los alelos para la resistencia. Éstos, que habían aparecido en el pool génico por mutación y que hasta el descubrimiento del DDT carecieron de importancia adaptativa, aumentaron su frecuencia progresivamente al ser favorecidos por la selección. Esto es un ejemplo de preadaptación. Con el correr de las generaciones no sólo aumentaba el número de individuos resistentes, sino también el grado de resistencia. Dado que los sobrevivientes portaban al menos uno de los alelos ventajosos, al entrecruzarse generaban descendientes que reunían en su genotipo los genes heredados por ambas líneas (recombinación gamética). Las gametas de estos insectos podían llevar a la vez genes “buenos” paternos y maternos (recombinación cromosómica). Por último, sus nietos heredaban los alelos de ambos orígenes ligados en el mismo cromosoma, gracias al proceso de crossing over (recombinación génica). En este caso se puede leer claramente cómo a partir de un pool con alta variabilidad genética, la presión selectiva del medio y la reproducción sexual y diferencial generan cambios en las frecuencias de alelos y por consiguiente en las características fenotípicas de una población. En el marco de la Teoría Sintética esto es evolución. - 173 - El Origen de las Especies A partir del Origen de la Vida, hace unos 3.800 millones de años, se ha originado la diversidad de organismos que han poblado la Tierra desde entonces y hasta la actualidad, mediante un proceso de especiación. Así mediante pequeños cambios que ocurren dentro de las poblaciones, la microevolución, se pueden originar nuevas especies. Las especies son grupos de poblaciones naturales que se cruzan o pueden entrecruzarse y se encuentran reproductivamente aisladas de otros grupos similares. El detalle esencial de esta definición es el aislamiento reproductivo. Sabemos que existen dos especies aparte cuando ambas pueden ocupar el mismo espacio sin entrecruzarse. En términos de genética de poblaciones, los miembros de una especie comparten un reservorio génico netamente separado de los reservorios génicos de otras especies. En la especiación, entonces, se presentan dos cuestiones importantes: Una cuestión central es cómo un reservorio de genes se separa de otro para iniciar una trayectoria evolutiva aparte. Otra cuestión es cómo dos especies, a menudo muy similares entre sí, habitan el mismo lugar al mismo tiempo, pero se mantienen reproductivamente aisladas. Mecanismos de especiación y algunos modelos: Se han protestos muchos modelos para explicar cómo ocurre la especiación. Si se considera el papel de la selección natural en la divergencia, el tiempo involucrado en la especiación y las barreras al flujo génico, los procesos de especiación pueden clasificarse en dos grandes categorías: Especiación por divergencia: ocurre un establecimiento gradual del aislamiento reproductivo como resultado de la divergencia entre dos poblaciones entre las que el flujo génico se ve interrumpido por alguna barrera espacial o ecológica. En todo momento las poblaciones evolucionan en forma adaptativa. Especiación instantánea o cuántica: ocurre cuando el establecimiento del aislamiento reproductivo es súbito y se origina en poblaciones en las que prevalecen fuerzas evolutivas como la deriva génica. Especiación por divergencia: Se reconocen tres modelos principales. El modelo clásico de especiación por divergencia se denomina especiación alopátrica. La especiación ocurre en forma gradual e involucra varias etapas. Se inicia con la separación de los individuos de una población a causa de la aparición de una barrera geográfica. - 174 - Las barreras geográficas son de muy distintos tipos. A menudo las islas se prestan para la aparición de especies nuevas; tal fue el caso de Australia cuando se fraccionó de Pangea. La aparición de áreas pantanosas entre dos hábitats puede ser una barrera efectiva; unos pocos metros de tierra seca pueden aislar a dos poblaciones de caracoles, etc. Un aislamiento de este tipo tiene como consecuencia que el flujo genético se produzca separadamente en cada una de las poblaciones, dando lugar a un desarrollo independiente. Incluso el punto de partida para esta evolución será distinto: habrá genes del patrimonio genético original que estén más representados en un grupo que en el otro y viceversa. Además las mutaciones y combinaciones que ocurran estarán sometidas a diferentes presiones de selección. Primero surgirán dos razas distintas y tras un largo período de aislamiento llegarán a establecerse dos subespecies. Los cambios en el medio ambiente impulsarán a ambas subespecies a emigrar y colonizar nuevas áreas, e irán experimentando cada vez mayores divergencias genéticas, hasta llegar a constituir dos especies distintas que habitan en diferentes regiones, es decir dos especies alopátricas (del griego allos: diferente y del latín patria: hogar). Si posteriormente se llegara a producir el encuentro de ambas especies en un territorio común, ya no sería posible el entrecruzamiento. Una vez que el aislamiento geográfico ha favorecido la divergencia y la especiación, empiezan a actuar un sinnúmero de otros mecanismos aisladores: son los mecanismos de aislamiento reproductivo. No todos los mecanismos de aislamiento actúan en todos los casos, pero sirven para fortalecer los límites entre las especies. Cualquier combinación favorable alcanzada en una de las especies se “diluiría” si el flujo genético se restableciera entre ellas. En el siguiente esquema, se entenderá mejor el proceso de especiación alopátrica. Cabe aclarar que el concepto de panmixia se refiere a un patrón particular de intercambio reproductivo en el cual cada individuo de un sexo se aparea con el del sexo opuesto al azar, con exclusión de toda preferencia fenotípica (y por lo tanto genotípica), es decir con independencia de las características de sus genotipos. Una población panmíctica es cuando todos los individuos que conforman esa población se aparean al azar. - 175 - Como vimos, es importante que existan mecanismos de aislamiento reproductivo para asegurar la integridad del reservorio génico y de la especie como tal. A continuación, los mecanismos de aislamiento reproductivo precigóticos, es decir aquellos mecanismos que impiden la formación de una cigota híbrida: Aislamiento ecológico. Si dos poblaciones son capaces de aparearse, pero no lo hacen porque viven en nichos ecológicos distintos, se dice que están aisladas ecológicamente. Por ejemplo: dos poblaciones de ciervos son capaces de entrecruzarse en el laboratorio. En la naturaleza una población vive del bosque y otra de un campo cercano. Cada - 176 - población permanece casi completamente dentro de su propio nicho. En esta situación son prácticamente nulas las posibilidades de entrecruzamiento. Aislamiento estacional. Aunque dos poblaciones habiten una misma región, sus períodos reproductores pueden ocurrir en distintas estaciones del año. Por ejemplo: en el norte de Europa, el saúco rojo florece dos meses antes que el saúco negro; sin embargo es posible cruzar las dos especies artificialmente. Aislamiento etológico. Se debe a diferencias de comportamiento. Por ejemplo: en ciertos peces, aves e insectos, el macho y la hembra llevan a cabo rituales de cortejo muy elaborados, en los que cada movimiento de uno desencadena una respuesta específica del otro. Estos comportamientos vienen determinados genéticamente, y cualquier cambio en el ritual haría que el otro miembro de la pareja perdiera interés. Aislamiento mecánico. Una pequeña modificación anatómica, aparentemente mínima, puede ser suficiente para evitar la cópula. Aislamiento gamético. En animales se manifiesta mediante mecanismos modificadores de la acidez de las vías genitales, que ciertos tipos de gametas no pueden soportar. Por ejemplo: algunos animales marinos como las ostras o los erizos de mar, liberan sus gametas al medio donde podrían fertilizarse con las gametas de otras especies. Sin embargo, esto no ocurre debido a la existencia de barreras químicas muy específicas. Pero también pueden ocurrir mecanismos de aislamiento reproductivo poscigótico , esto implica la formación de la cigota a partir de dos especies diferentes pero se ven afectados el desarrollo, la viabilidad o la fertilidad de la primera o la segunda generación de híbridos: Aislamiento genético. El hecho de que se produzca la fusión de las gametas no asegura que el cigoto pueda desarrollarse. Un cigoto producto de especies diferentes, en la mayoría de los casos no pasa de las primeras divisiones. Por ejemplo: los embriones híbridos que provienen de la cruza de cabra y oveja mueren en las primeras divisiones. Esterilidad del híbrido. Los híbridos completan su desarrollo pero son estériles. Por ejemplo: El asno y la yegua pueden cruzarse dando un híbrido, la mula. Si bien es fuerte y sobrevive, no puede reproducirse porque sus gónadas no se desarrollan. Vimos que la especiación por divergencia, involucraba tres modelos diferentes. Especiación alopátrica, es uno de lo más clásicos en cuanto a ocurrencia. Otro modelo es la especiación simpátrica (del griego sym: junto y del latín patria: hogar). Ocurre entre poblaciones que conviven en un mismo territorio y la barrera que origina esta especiación no es geográfica, sino ecológica o etológica (de comportamiento). Los individuos portadores de distintas variantes se adaptan a distintos compartimientos ambientales y acumulan diferencias genéticas que conducen a la aparición de mecanismos de aislamiento reproductivo. Un ejemplo de este proceso lo encontramos en una mosca de la fruta (Rhagoletis pomonella) de América del Norte. Existen dos grupos de moscas que se alimentan de los frutos de dos tipos diferentes de árboles: - 177 - espinos y manzanos. Las moscas que viven en los manzanos se aparean entre sí y las que viven en los espinos también se aparean entre sí. A pesar de que son potencialmente fértiles, la probabilidad de que dejen descendencia híbrida es muy baja porque las moscas muestran preferencias de hábitat. Además las moscas del manzano y del espino se reproducen en diferentes épocas del año. Originalmente ambos grupos de moscas se alimentaban del espino pero ocurrió lo que se denomina un salto de nicho cuando comenzó a explotar el manzano. En consecuencia, estas dos subpoblaciones de moscas adaptadas a cada tipo de fruto, fueron acumulando diferencias genéticas. Son un ejemplo de lo que se denomina razas ecológicas que están en camino de convertirse en nuevas especies. 3. EVOLUCIÓN DE LOS HOMÍNIDOS Los humanos (Homo sapiens), junto con nuestros antecesores más cercanos ya extintos, pertenecemos al grupo de los homínidos. Pero también somos mamíferos placentarios y primates, al igual que los traseros, lo lemúres, los monos y los antropomorfos, entre otros. Las principales tendencias en la evolución de los primates han sido los cambios en la estructura anatómica de las manos y los brazos, el incremento en la agudeza y estereoscopía visual, la prolongación del cuidado de las crías y la postura vertical de la espalda al colgarse o sentarse. Todas estas características están relacionadas con adaptaciones a la vida arbórea. Entre los primates se pueden reconocer dos grupos principales: Prosimios: aparecieron hace unos 55 millones de años; en la actualidad son arborícolas, de pequeño tamaño y de hábitos nocturnos. Dentro de este grupo se encuentran los lemures, tarseros, loris y galagos. Antropoides: se habrían originado a partir del tronco de los prosimios hace unos 40 millones de años y están constituidos por dos grandes linajes: los monos del Nuevo Mundo o platirrinos y los monos del Viejo Mundo o catarrinos. Los humanos y los antropomorfos (gibones, orangutanes, gorilas y chimpancés) son catarrinos y conforman el grupo de los hominoides. La pregunta que se puede formular es: ¿cuál de estas especies está más estrechamente emparentada con nuestra especie? ¿Cuánto hace que el linaje al que pertenecemos los humanos divergió del resto de los simios? ¿A qué época se remonta el último antepasado común compartido con ello? Darwin consideraba que los parientes más próximos a los humanos eran los simios africanos (chimpancés y gorilas) y que el gran simio asiático, el orangután, tenía un vínculo más distante. - 178 - En 1963, Morris Goodman demostró que los chimpancés, los gorilas y los humanos forman un grupo natural, como había anticipado Darwin. Por otra parte, “el reloj molecular”, un método que permite estimar el tiempo de divergencia entre dos taxones, permitió estimar que la divergencia entre el linaje de los humanos y el de chimpancé – gorila habría ocurrido muy recientemente, hace aproximadamente de 5 a 8 millones de años. Las principales características del linaje de los homínidos son: Andar erecto Cerebro mayor en proporción con el tamaño corporal Patrón morfológico del primer molar con dos cúspides en lugar de la única que poseen los simios africanos. Existen dos hipótesis sobre la evolución del hombre moderno: 1. Modelo del Candelabro o Multirregional: este modelo propone que hubo múltiples migraciones tempranas desde África hacia Europa y Asia, que comenzaron tal vez hace más de un millón y medio de años y por las que se establecieron diferentes poblaciones de Homo erectus. Estas poblaciones que evolucionaron en formas graduales y separadas unas de otras, habrían originado las distintas variantes actuales de Homo sapiens. - 179 - 2. Modelo del Arca de Noé o Fuera de África: propone que es probable que un pequeño grupo de humanos modernos, previamente diferenciados como Homo sapiens, migrara desde algún sitio de África, en épocas más recientes, hace unos 100 a 150 mil años, y colonizara el planeta, reemplazando a las poblaciones previamente existentes que se extinguieron. Se han encontrado restos de humanos modernos en algunas localidades antes que en otras, con una distribución consistente con las rutas de migración de los hombres modernos a partir de su centro de dispersión hipotético, lo cual da sustento a este modelo. También este modelo del Arca de Noé, ha recibido apoyo de datos que provienen de la biología molecular, a través del análisis de ADN mitocondrial. Este método molecular, denominado reloj mitocondrial, permite analizar hechos recientes de la evolución (medidos en miles de años en lugar de millones de años). La ventaja del ADN mitocondrial es que por un lado no tiene enzimas de reparación del ADN, por lo tanto este ADN evoluciona a un atas más rápida que el ADN nuclear (que posee enzimas reparadoras de ADN). La otra gran ventaja del ADN mitocondrial es que, las mitocondrias se transmiten sólo a través de la línea materna, los cambios en las secuencia de nucleótidos que se estudian nunca son el resultado de recombinación sexual, sino sólo de acumulación de mutaciones. - 180 - ¿Es posible que los huesos fosilizados de un simio más antropoide (semejante al hombre) o de un hombre más pitecoide (semejante al simio) estén esperando, perdidos en algunos estratos antiquísimos, las investigaciones de un paleontólogo que no ha nacido todavía? THOMAS H. HUXLEY Esta frase de Huxley, expresa claramente lo difícil de fundamentar y explicar un suceso histórico tan largo en el tiempo y a su vez complejo, cuando todavía se siguen encontrando nuevos registros fósiles que dan lugar a la formulación de nuevas hipótesis sobre la evolución del Hombre. La nomenclatura de las especies que participaron en la evolución humana aún es muy controversial y sus relaciones no se han aclarado por completo. Sin embargo, cuatro hipótesis ampliamente consensuadas orientan actualmente la interpretación del patrón evolutivo de los homínidos: La evolución de nuestro linaje, parece estar caracterizada por una serie de eventos cladogenéticos1 (cuando un linaje evolutivo se desdobla en dos o más linajes separados). Nuestra supervivencia fue simplemente casual y nuestro éxito no es el resultado de un plan preconcebido de progreso lineal. 1 Cladogénesis: es uno de los principales patrones de cambio macroevolutivo, conocido también como evolución por ramificación, partición o división. - 181 - Los patrones del cambio cultural, como los del cambio biológico en el nivel macroevolutivo, se caracterizan por largos períodos de estasis (sin cambios o modificaciones) y breves momentos de cambios profundos. En general, los cambios culturales no ocurren en forma gradual. La cultura se transmite de manera vertical y horizontal, y esta última es mucho más rápida. Cuando se establece una tecnología, se difunde con rapidez y se mantiene durante un período sin mayores cambios, hasta que se inventa una nueva tecnología que reemplazará a la primera. La capacidad de caminar en dos pies es un rasgo que caracteriza a todo el linaje. Este rasgo, y no nuestra inteligencia, sería el que nos puso en el camino hacia la humanidad. - 182 - BIBLIOGRAFIA Ayala, F. La Naturaleza inacabada. Ensayos en torno a la evolución. Biblioteca Científica Salvat. 1994. British Museum Natural History. El Origen de las Especies. Editorial Akal. 1981. Curtis, H; Barnes, N. Biología. 7ª Edición. Editorial Médica Panamericana. Buenos Aires. 2008. Dawkins, R. El gen egoísta. Biblioteca Científica Salvat. Nº 9. 1985. El Correo de la UNESCO. De ratones y mariposas nocturnas. Mayo 1982. Gould, S. J. El pulgar del panda. Biblioteca de Divulgación Científica Muy Interesante. Nº 16. 1986. Lewin, R. Evolución humana. Biblioteca Científica Salvat. Nº 64. 1987. Miller J; Van Loon, B. Darwin para principiantes. Editorial Era Naciente SRL. 1995. PROMEC. Biología. Estructura Modular 5. Módulo 2. 1985 Schwoerbel, Wolgfgang. Evolución. Biblioteca Científica Salvat Nº 43. 1984. Simpsom, G. G. El sentido de la evolución. Editorial EUDEBA. 1984. Stansfield, W. Genética. 3º Edición. Editorial Mc Graw Hill. 1992. Weisz, P. B. La ciencia de la Biología. Editorial Omega. 1975. - 183 -