- Ninguna Categoria
INFOMUSA, Vol.8, N°1
Anuncio
INFOMUSA La Revista Internacional sobre Banano y Plátano Vol. 8 N° 1 Junio 1999 EN ESTE NUMERO Fase II del IMTP: sinopsis de los resultados Bananos y marchitamiento por Fusarium en Taiwan Estudio de bananos de Papua Nueva Guinea con respecto a nematodos Estudio de variedades de banano frente a Sigatoka amarilla en Australia GCV de las poblaciones de Fusarium oxysporum f. sp. cubense presentes en Cuba Crioconservación de meristemas en el banano: optimización de la regeneración Polinizaciones y cultivo de embriones en bananos Fertilidad polínica de algunos cultivares de Musa utilizados por el INIVIT Observaciones preliminares sobre los enemigos naturales asociados con el picudo negro del banano en Uganda Podredumbre de las frutas de los bananos en el Sudeste de Asia Libros etc. Anuncios Noticias de INIBAP Noticias de Musa Noticias de PROMUSA La misión de la Red Internacional para el Mejoramiento del Banano y el Plátano es aumentar de manera sostenible la productividad del banano y el plátano cultivados por pequeños productores para el consumo doméstico y mercados locales y de exportación. INFOMUSA La Revista Internacional sobre Banano y Plátano Vol. 8 N° 1 Junio 1999 EN ESTE NUMERO Fase II del IMTP: sinopsis de los resultados Bananos y marchitamiento por Fusarium en Taiwan Estudio de bananos de Papua Nueva Guinea con respecto a nematodos Estudio de variedades de banano frente a Sigatoka amarilla en Australia GCV de las poblaciones de Fusarium oxysporum f. sp. cubense presentes en Cuba Crioconservación de meristemas en el banano: optimización de la regeneración Polinizaciones y cultivo de embriones en bananos Fertilidad polínica de algunos cultivares de Musa utilizados por el INIVIT Observaciones preliminares sobre los enemigos naturales asociados con el picudo negro del banano en Uganda Podredumbre de las frutas de los bananos en el Sudeste de Asia Libros etc. Anuncios INIBAP tiene cuatro objetivos principales : • organizar y coordinar un esfuerzo global de investigación sobre banano y plátano para el desarrollo, la evaluación y la diseminación de cultivares mejorados y para la conservación y utilización de la diversidad de las Musaceas ; • promover y fortalecer colaboraciones en la investigación relacionada con banano y plátano a los niveles nacional, regional e internacional ; • fortalecer la capacidad de los SNIA para conducir actividades de investigación y desarrollo sobre banano y plátano ; • coordinar, facilitar y apoyar la producción, recopilación y el intercambio de información y de documentación sobre banano y plátano. Desde Mayo de 1994, INIBAP está dirigida y administrada por el Instituto Internacional de Recursos Fitogenéticos (IPGRI). Noticias de INIBAP Noticias de Musa Noticias de PROMUSA Vol. 8, N° 1 Publica : Red Internacional para el Mejoramiento del Banano y el Plátano (INIBAP) Jefe de redacción : Claudine Picq Comité editorial : Emile Frison, Jean-Vincent Escalant, Gisella Orjeda, Suzanne Sharrock Impreso en Francia Redacción : INFOMUSA, INIBAP Parc Scientifique Agropolis II, 34397 Montpellier Cedex 5, Francia. Teléfono +33-(0) 4 67 61 13 02 Telefax : +33(0)- 4 67 61 03 34 Correo electrónico : [email protected] La subscripción es gratuita para los países en vías de desarrollo. Se agradecen contribuciones en forma de artículos y cartas al editor. La redacción se reserva el derecho de editar los artículos. INFOMUSA no se responsabiliza por el material no solicitado. Sin embargo, trataremos de responder a cada una de las peticiones. Los artículos pueden ser citados o reproducidos sin cargos, con la mención de la fuente. También se publican ediciones de INFOMUSA en francés y en inglés. Cambio de dirección : Para evitar la perdida de sus ejemplares de INFOMUSA, notifique a INIBAP con seis semanas de antelación si cambia de dirección postal. Las opiniones expresadas en los artículos son responsabilidad de sus autores y no necesariamente reflejan los puntos de vista de INIBAP. 2 Vol. 8, N° 1 INFOMUSA Foto en la portada : Mercado en Guinea (E. Akyeampong, INIBAP) CONTENIDO Programa Internacional de Evaluación de Musa (IMTP) fase II sinopsis del informe final y resumen de los resultados . . . . . . . . . . . . . . . . . 3 Desempeño de los clones de banano frente al reto del marchitamiento por Fusarium en Taiwan . . . . . . . . . . . . . . . . . . . . . . . . 10 Estudio de los bananos procedentes de Papua Nueva Guinea con respecto a los nematodos noduladores y de lesiones de la raíz . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12 Estudio de 143 variedades de banano con respecto a su resistencia a la Sigatoka amarilla en el norte de Queensland . . . . . . . 15 Grupos de compatibilidad vegetativa de las poblaciones de Fusarium oxysporum Schlecht f. sp. cubense (E. F. Smith) Snyd. and Hans. presentes en Cuba . . . . . . . . . . . . . . . . . . . . . 22 Crioconservación de meristemas en el banano (Musa spp.): optimización de la regeneración . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23 Polinizaciones y cultivo de embriones en bananos . . . . . . . . . . . . . . . . . . . . . . 24 Estudio de la fertilidad polínica de algunos cultivares de Musa utilizados en el programa de mejoramiento genético del INIVIT . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 27 Observaciones preliminares sobre los enemigos naturales asociados con el picudo negro del banano Cosmopolites sordidus Germar en Uganda . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 28 Enfermedades de la podredumbre de las frutas de los bananos en el Sudeste de Asia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29 Ecología, distribución y estructura de las poblaciones de Fusarium oxysporum f.sp. cubense en Kenia . . . . . . . . . . . . . . . . . . . . . . 31 Libros etc. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31 Anuncios . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32 Noticias de INIBAP . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33 Noticias de Musa . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 35 Noticias de PROMUSA . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .de I a XVI INFOMUSA — Vol 8, N° 1 Ensayos multisitio Recursos geneticos Programa Internacional de Evaluación de Musa (IMTP) fase II sinopsis del informe final y resumen de los resultados Gisella Orjeda, Jean-Vincent Escalant y Natalie Moore l Programa Internacional de Evaluación de Musa (IMTP) se estableció en 1989 como una sociedad cooperativa entre los Sistemas Nacionales de Investigación Agrícola (SNIA), INIBAP, programas de mejoramiento y patólogos de varios institutos con el apoyo del PNUD. El propósito fue identificar en ensayos multisitio alrededor del mundo los híbridos resistentes de bananos y plátanos que cumplieran con requisitos locales y con los cuales los pequeños agricultores pudieran reemplazar los cultivares susceptibles existentes. Otro objetivo del IMTP fue estimular el mejoramiento suministrando información a los programas de mejoramiento sobre la respuesta patológica de sus variedades mejoradas. Como efecto secundario, el IMTP también ayudó a aumentar la capacidad de las organizaciones nacionales para llevar a cabo investigaciones en banano y plátano. El programa empezó evaluando el germoplasma proveniente del programa de mejoramiento de la FHIA con respecto a su resistencia a la Sigatoka negra (Mycosphaerella fijiensis). Siete híbridos tetraploides con amplias bases genéticas fueron evaluados junto con varios clones diploides de referencia (silvestres y comestibles) que representaban la serie completa de reacciones a la Sigatoka negra, desde los altamente resistentes hasta los altamente susceptibles. Los experimentos se establecieron en seis E países. A los administradores de los sitios se les brindó la capacitación, guías técnicas y financiamiento para llevar a cabo los experimentos. Cuatro años después, se publicó un documento final que presentó resultados detallados del IMTP. Se hicieron recomendaciones para la liberación de tres clones para distribución : los clones FHIA-01 y FHIA-02, ambas variedades de postre con desempeño excepcional y altamente resistentes a la Sigatoka negra, y el FHIA-03, un banano de cocción también con un desempeño excelente y resistente a la Sigatoka negra. Durante los últimos diez años estos tres clones han sido distribuidos en más de 50 países alrededor del mundo. El éxito de la fase I del IMTP fomentó el crecimiento del programa y se le solicitó a INIBAP desarrollar la iniciativa más adelante. En 1991, otra propuesta se sometió al PNUD que fue aprobada para un total de tres años. La propuesta también incluía apoyo a los programas de mejoramiento, indización del germoplasma donado por los programas de mejoramiento, apoyo a la investigación de los virus, publicación de los resultados y gastos de personal. Esta vez no se asignaron fondos para el establecimiento y mantenimiento de los sitios de ensayo. Sin embargo, la mayoría de los SNIA decidieron que estaban preparados para financiar ellos mismos los ensayos, debido a la relevancia del IMTP para sus programas. Para la fase II del IMTP, se expandieron todos los aspectos del programa. En vez de evaluar el germoplasma con respecto a la resistencia a un solo patógeno, en esta fase el germoplasma fue evaluado con respecto a su resistencia a tres enfermedades, Sigatoka negra : M. fijiensis, Sigatoka amarilla : M. musicola y marchitamiento por Fusarium : Fusarium oxysporum f. sp. cubense. Cuatro programas de mejoramiento proporcionaron el germoplasma, cuyo listado se presenta en la Tabla 1, y la cantidad de sitios de evaluación aumentó de seis a 37, a pesar del hecho de que los ensayos fueron financiados a expensas del instituto participante. En la mayoría de los ensayos en la fase II del IMTP las plantas fueron sembradas durante 1996 y 1997. El informe completo incluye los resultados de los sitios para el marchitamiento por Fusarium (Foc) en Australia, Brasil, Honduras, Indonesia, Malasia, Filipinas, Africa del Sur, España, Taiwan, y Uganda, sitios para la Sigatoka negra (BS) en Camerún, Costa Rica, Honduras, Nigeria, Filipinas, Tonga y Uganda un sitio para la Sigatoka amarilla (YS) en Colombia. Aunque muchos sitios proporcionaron sus datos, no fue posible realizar un análisis completo ya que faltó mucha información debido a las catástrofes naturales y a la recolección incompleta de datos en algunos de los sitios. Entre estos sitios está uno para el Foc en Australia, uno para la YS en Camerún, tres sitios en Cuba (uno para la BS, uno para la YS y uno para el Foc), cinco sitios en India (dos para la YS y tres para el Foc), dos sitios para el Foc en Indonesia, un sitio para el Foc en Malasia, un sitio para la YS en Sta. Lucia y dos sitios en Tailandia (uno para el Foc y uno para la YS). Tabla 1. Cultivares mejorados incluidos en los ensayos de resistencia a la Sigatoka y Fusarium en la fase II del IMTP. Nombre del cultivar Origen Genoma Pedigree Enfermedad Sogatoka PV-03.44 EMBRAPA1 AAAB PA-03.22 SH-3436-9 INIVIT1 AAAA Pacovan x Calcutta 4 ✓ Prata Aña x Calcutta 4 ✓ Variant somaclonal de SH-3436: ✓ Fusarium ✓ Highgate x SH-3142 FHIA1 AAAB Prata Aña x (Prata Aña x SH-3142) ✓ FHIA-03 AABB SH-3486 x SH-3320 ✓ FHIA-17 AAAA Gros Michel Highgate x SH-3362 FHIA-23 AAAA Highgate x SH-3362 AAA Variant de Giant Cavendish issu de culture in vitro ✓ AAA Variant de Giant Cavendish issu de culture in vitro ✓ FHIA-01 GCTCV-119 TBRI1 GCTCV-215 ✓ ✓ ✓ 1Programas de mejorameinto que proporcionaron el germoplasma : EMBRAPA Empresa Brasileira de Pesquisa Agropecuaria, Brésil FHIA Fundación Hondureña de Investigación Agrícola, Honduras INIVIT Instituto Nacional de Investigaciones en Viandas Tropicales, Cuba TBRI Taiwan Banana Research Institute, Taiwan INFOMUSA — Vol 8, N° 1 3 Este artículo es un resumen que brinda una sinopsis de los resultados del IMTP II. Por razones de espacio, presentamos solo los promedios para el peso del racimo y el puntaje para las enfermedades. El informe final completo incluye los promedios y coeficientes de varianza de los principales caracteres agronómicos y de la enfermedad, análisis de varianza y comparaciones de los promedios, así como todos sus gráficos más los gráficos de la evolución de la enfermedad a través de tiempo. Para recibir ejemplares del informe final, por favor dirígese al Coordinador del IMTP en INIBAP. Procedimiento utilizado para el análisis de los datos En los sitios para la Sigatoka, el diseño experimental consistía de un bloque completo aleatorio con doce genotipos, cinco réplicas o bloques y cinco plantas por parcela. La unidad experimental consistía de la parcela con cinco plantas y todos los análisis fueron realizados con sus promedios. En los sitios para el marchitamiento por Fusarium el diseño experimental consistía de un diseño aleatorio con 22 genotipos y 20 réplicas, la unidad experimental era cada réplica. En ambos casos, se incluyeron como controles de referencia los clones susceptibles y clones resistentes así, como el cultivar local del área. A los administradores de los sitios se les proporcionó protocolos de evaluación y ellos enviaron datos sin procesar al coordinador del IMTP. Los datos sin procesar procedentes de varios sitios fueron importados al programa Minitab v. 12.2 para su análisis. Sitios para la Sigatoka Características agronómicas Con respecto a las características agronómicas, se utilizó un análisis de varianza de dos vías con bloques y genotipos como fuente de varianza. Siguiendo el análisis de la varianza, los promedios de cada genotipo se compararon con el promedio del cultivar local utilizando el procedimiento de Dunnett. Se utilizó la comparación unilateral. Esta comparación múltiple de los promedios fue útil, por ejemplo, para determinar los mejores rendidores o clones más altos que el cultivar local. Sin embargo, en este artículo solo presentamos cifras con el promedio para cada genotipo del peso del racimo y de la cantidad de días hasta la cosecha, así como un gráfico que ilustra el peso del racimo promedio de los cultivares mejorados en cada sitio como una medición general de la calidad del sitio para cultivar los genotipos mejorados. después de la siembra, durante la emergencia de los racimos y durante la cosecha. En el informe completo, se realizó un análisis para cada período de evaluación. Luego, se compararon todos los tratamientos, primero con el promedio de referencia susceptible y luego con el promedio de referencia resistente. Se utilizaron dos pruebas consecutivas de Dunnett de dos vías con el fin de detectar, en primer lugar, aquellos genotipos que presentaron un fenotipo susceptible en mayor, igual o menor grado que el control susceptible, y, en segundo lugar, los genotipos que fueron más, igual o menos resistentes que el testigo resistente. En este resumen presentamos un gráfico para el índice de infección y la YLS promedios por genotipo en tres países representativos : Camerún, Costa Rica, y Tonga. En el informe completo se presenta un gráfico que ilustra el progreso del índice de infección durante el primer ciclo para comparar todos los genotipos en cada sitio. El informe completo también incluye tablas con promedios y coeficientes de variación de DDT y YLS. Clones de referencia utilizados para los análisis El Calcutta IR 124 fue sembrado en todos los ensayos como un clon altamente resistente. Sin embargo, este clon no fue utilizado para las comparaciones de los promedios ya que es un clon con una respuesta hipersensible y su índice de infección siempre es igual a cero. Evidentemente, todos los cultivares serían menos resistentes que el Calcutta. El Pisang Lilin, otra raza indígena resistente, también fue sembrado en todos los ensayos como clon de referencia altamente resistente. Desafortunadamente, debido a los problemas con los cultivos de tejidos, en todos los sitios se utilizaron variantes somaclonales enanas muy débiles. Por lo tanto, estas plantas no podrían ser utilizadas como un clon de referencia como se pretendía. El Pisang Ceylan, una raza indígena seleccionada para ser la referencia parcialmente resistente para la segunda ronda del IMTP, también fue sembrado en todos los ensayos en la fase II del IMTP. Este clon, en ausencia de otros clones resistentes adecuados, se utilizó como un control resistente para las comparaciones múltiples de los promedios de Dunnett. El Pisang Berlin fue utilizado como el clon de referencia susceptible. En uno o dos ensayos donde este clon no fue sembrado, el análisis se realizó utilizando el Niyarma Yik como clon de referencia altamente susceptible. vía con cada genotipo como fuente de varianza. Siguiendo los análisis de varianza, los promedios de cada genotipo se compararon con los del cultivar local. Una comparación unilateral de Dunnett de los promedios con un control se utilizó de la misma manera que en los análisis de la Sigatoka (ver la explicación presentada arriba). Como en los sitios para la Sigatoka, se presenta un gráfico dual que resume el peso promedio de los racimos por sitio y por genotipo mejorado a través de los sitios. El análisis completo se presenta en el informe. Síntomas internos Los registros de los síntomas internos también fueron analizados utilizando un ANOVA de una vía. La prueba de Dunnett unilateral fue realizada utilizando el clon de referencia susceptible al marchitamiento por Fusarium especificado por el protocolo. Un clon de referencia distinto fue utilizado dependiendo de la raza del Fusarium presente en el sitio. Sin embargo, en este artículo presentamos sólo una tabla que resume los promedios de los síntomas internos (rizoma) en los sitios infestados con la raza 1 del Foc (Tabla 2). También presentamos en este resumen dos gráficos duales (uno por cada raza del Foc) que muestran los promedios de los registros de los síntomas internos promediados por sitio y por cada genotipo a través del sitio. Síntomas externos El protocolo para la evaluación del marchitamiento por Fusarium también especificó Tabla 2. Promedios de los puntajes de decoloración del rizoma de los cultivares mejorados, clones de referencia y cultivares locales. Sitios de ensayos del IMTP II infestados con la raza 1 del Foc. Puntajes de decoloración del rizoma Genotipo Brasil FHIA-01 (AAAB) 1.47 Honduras Filippinas 1.00 4.14 FHIA-03 (AABB) 2.65 1.00 5.60 FHIA-17 (AAAA) 1.00 1.00 5.78 FHIA-23 (AAAA) 1.18 1.00 4.13 GCTCV 119 1.00 1.00 2.00 GCTCV 215 1.00 1.00 5.63 PA 03.22 1.15 1.07 5.00 PV 03.44 1.00 1.00 4.86 Gros Michel 2.40 1.20 6.00 Bluggoe 3.29 1.39 6.00 3.88 Pisang Ceylan 2.05 1.06 Williams 1.00 1.00 Local cultivars Indice de infección, tiempo de desarrollo de la enfermedad (DDT) y la hoja más joven manchada (YLS) El índice de infección se registró en tres etapas durante el primer ciclo, seis meses 4 Sitios para el marchitamiento por Fusarium Características agronómicas Con respecto a las características agronómicas, se utilizó un análisis de varianza de una FHIA-18 1.00 Latundan Prata Anã 5.70 2.78 Los datos completos son proporcionados en el informe final de la fase II del IMTP. INFOMUSA — Vol 8, N° 1 el registro de siete síntomas externos una vez al mes, empezando durante el tercer mes después de la siembra, con el propósito de visualizar si hay una clara evolución de los síntomas. Por lo tanto, los resultados finales presentan un gráfico de la evolución de los síntomas externos por genotipo a lo largo del experimento. Genotipo (todos los sitios) índice de infección 34 28 22 16 local Niy arma Yik P. lilin P. Berlin Calcutta Saba P. Ceylan Yang. km5 PA 03-22 SH 3436-9 FHIA 23 PV 03.44 Filipinas Honduras CR2 CR1 Cameroun 10 Figura 1. Promedios para el índice de infección durante la emergencia de los racimos – todos los genotipos. Camerun 12 40 P. Ceylan Var. Locale 35 FHIA 23 10 Indice de infección SH3436-9 30 P. Berlin PV 03.44 PA 03-22 Saba 25 8 Saba FHIA 23 20 Var. locale PV 03.44 PA 03-22 15 6 P. Berlin SH3436-9 4 P. Ceylan 10 5 YLS Diversidad del Foc a través de los sitios de ensayo del IMTP Las muestras del tejido vascular decolorado y seco de los seudotallos de las plantas infectadas fueron preparadas por los trabajadores en cada sitio y enviados a la Unidad de Fitopatología, DPI, Indooroopilly, Qld, Australia para su análisis. El Foc fue aislado del tejido afectado y se prepararon cultivos monoconidiales para cada aislado. Para caracterizar los aislados del Foc, se realizaron los análisis de los grupos de compatibilidad vegetativa (GCV) y de producción volátil en los laboratorios del DPI, y la impresión de huellas genéticas de ADN utilizando el análisis de amplificación de huellas (DAF) fue conducido por la Dra. Suzy Bentley en el CRC para los laboratorios de fitopatología tropical. Los resultados de los análisis se presentan en la Tabla 3. Estas técnicas se describen en el informe completo : Sitio (todos los genotipos) Yang.Km5 2 Calcutta 4 Calcutta 4 0 0 Yang.Km5 Indice de infección YLS Resultados Costa Rica 50 P. Ceylan PA 03.22 Yang.Km5 45 P. Berlin 9 PV 03.44 8 FHIA 23 Var. locale SH3436-9 40 Indice de infección Saba Calcutta 4 P. Berlin 7 PV 03.44 35 6 PA 03.22 Saba 30 5 P. Ceylan Var. Locale 25 SH3436-9 4 20 Yang.Km5 3 FHIA 23 15 2 Calcutta 4 10 1 5 0 0 Indice de infección YLS Figura 3. Tolerancia a la Sigatoka negra de diferentes genotipos en Costa Rica. YLS : Hoja más joven manchada. Tonga 40 Var. Locale Saba 30 PA 03-22 PA 03-22 8 Saba PV 03.44 FHIA 23 20 SH3436-9 6 P. Berlin Var. locale 4 P. Ceylan 10 5 10 PV 03.44 P. Berlin 25 12 SH3436-9 P. Ceylan 35 15 FHIA 23 Yang.Km5 YLS INFOMUSA — Vol 8, N° 1 Figura 2. Tolerancia a la Sigatoka negra de diferentes genotipos en Camerún.YLS : Hoja más joven manchada. YLS Cultivares mejorados Tolerancia a la Sigatoka negra (M. fijiensis) La respuesta de los clones que fueron examinados contra la Sigatoka negra, varió de acuerdo a diferentes factores bióticos y abióticos presentes en cada país. Un análisis de los resultados obtenidos indicó que el Tiempo de Desarrollo de la Enfermedad (DDT) no representó un parámetro confiable para la evaluación de los niveles de resistencia. Esto puede ser debido a las dificultades en la interpretación de los síntomas foliares en ciertas situaciones. Por ejemplo, cuando la presión de la enfermedad es alta, las numerosas lesiones que pueden ocurrir en la etapa 1, pueden unirse en superficie de la hoja y asemejar una lesión necrótica de la etapa 6. En contraste, el índice de infección parece proporcionar un parámetro más confiable permitiendo una mejor consistencia entre los países y una base más apropiada para la clasificación de los nuevos híbridos (Figura 1). En este resumen, los resultados obtenidos en Costa Rica, Camerún y Tonga se presentan como de países representativos de América Latina, Africa y Asia/Pacífico. Los híbridos de Honduras (FHIA-23 y SH-34369) mostraron un índice de infección promedio cercano al obtenido para el Pisang Ceylan (referencia resistente), permitiendo concluir que el FHIA-23 y el SH-3436-9 son tolerantes a la enfermedad de la Sigatoka negra (Figuras 2, 3, 4). Esta conclusión es Indice de infección Sigatoka negra Yang.Km5 2 Calcutta 4 Calcutta 4 0 Indice de infección 0 YLS Figura 4. Tolerancia a la Sigatoka negra de diferentes genotipos en Tonga. YLS : Hoja más joven manchada. 5 40 Peso (Kg) 35 30 25 20 15 10 6 ga nd a U H País FHIA23 PV-03.44 PA-03.22 SH-3436-9 Local Figura 5. Peso del racimo obtenido durante el IMTP II en diferentes genotipos. 700 600 500 400 300 200 100 nd ga U To ng a a s H Co Ca st on a du Ri er m sh O go ra ca un iro 0 Discusión País FHIA23 PV-03.44 PA-03.22 SH-3436-9 Local Figura 6. Días desde la siembra hasta la cosecha durante el IMTP II en diferentes genotipos. 250 200 Peso del dedo (g) La tolerancia a la Sigatoka negra de los mismos clones mostró algunas variaciones significativas de país a país. Ya que muchos factores, incluyendo el manejo, fertilidad de los suelos, presión del patógeno, presencia de otros patógenos y condiciones climáticas, influyen sobre la tolerancia de la planta, no es posible generalizar los resultados. Los efectos de estos factores y su papel en el rendimiento obtenido no son fáciles de demostrar o cuantificar. Sin embargo, en CRBP, Camerún, se realizó algún trabajo en esta área. Este trabajo permitió demostrar la influencia de la Sigatoka negra sobre el peso del racimo. Utilizando los resultados del IMTP con respecto al índice de infección durante la emergencia de los racimos y el peso de los dedos promedio a través de los sitios, se encontraron algunas buenas correlaciones con un coeficiente de correlación de – 0.71 (Figura 7). Esto indica que la cantidad de parámetros para los cuales se recolectan los datos, puede ser reducida, facilitando de este modo la recolección y el manejo de los datos. También reduciría la necesidad de interpretación visual de los síntomas en el campo. También fue notado que los clones altamente resistentes Calcutta 4 y Yangambi Km5 presentaron lesiones necróticas de la To ng a on du r a Co st Ba go O Ca m er Ri ca un sh iro 0 as 5 Ba Desempeño agronómico Los híbridos de la FHIA dieron muy buenos rendimientos. Los racimos del FHIA-23 y SH-3436-9 pesaron en promedio 30.6 y 22.3 kg respectivamente a través de los sitios, con un peso máximo de 39.37 kg en Camerún y de 28.8 kg en Tonga respectivamente. Los rendimientos de los híbridos de EMBRAPA fueron más bajos, con pesos de racimos promedio entre 10.1 y 9.27 kg a través de los sitios para el PV-03.44 y PA-03.22 respectivamente. Aunque los cultivares locales difieren entre los países, es interesante notar que el promedio a través de los sitios es de 16,5 kg, con un peso máximo de 22.78 kg en Tonga. Esto sirve para reforzar el buen desempeño de los híbridos de la FHIA (Figura 5). Sin embargo, los híbridos de la FHIA tienen un ciclo más largo en comparación con las referencias locales, con un promedio de 474 y 420.2 días para el FHIA-23 y el SH-3436-9 respectivamente (Figura 6). 45 Periodo siembra/cosecha (días) consistente con el registro de la Hoja Más Joven Manchada (YLS) obtenido en la mayoría de los países. Los híbridos de EMBRAPA, Brasil (PV 03.44 y PA 03.22) tienen índices de infección promedio que se encuentran entre aquellos para el Pisang Berlin y los cultivares locales. Además, los promedios de YLS en la mayoría de los países fueron más bajos que los de los cultivares locales susceptibles, permitiendo considerar a estos tan susceptibles, como los clones de referencia susceptibles. R2 = 0,5064 150 100 50 0 0 10 20 30 40 50 60 70 Indice de infección Figura 7. Correlación entre el índice de infección y peso del dedo en los bananos. El análisis se hizo utilizando todos los clones a través de los sitios en el marco del IMTP. etapa 6 en algunos de los sitios, un resultado no esperado para estos clones. Esto indica que se requiere realizar más estudios para determinar si están presentes algunas cepas de Mycosphaerella fijiensis más agresivas o realmente si una nueva especie de patógeno está involucrada en estos casos. INFOMUSA — Vol 8, N° 1 Tabla 3. Diversidad del Fusarium oxysporum f. sp. cubense en los sitios de ensayos de la fase II del IMTP. Localización y GCV Producción volátil Genotipos de Musa GCV 0124 inodoratum Bluggoe (ABB) inodoratum Calcutta IR 124 (AAw) inodoratum FHIA-02 (AAAB) inodoratum Gros Michel (AAA) inodoratum Ney Poovan (AB) inodoratum Pisang Ceylan (AAB) GCV 01213/16 Bluggoe (ABB) FHIA-03 (AABB) odoratum FHIA-23 (AAAA) odoratum Gros Michel (AAA) odoratum Igisahira Hazyview, Eastern Transvaal, Sudafrica odoratum Pisang Mas (AA) GCV 0120/15 GCV 0124 inodoratum Bluggoe (ABB) GCV 0125 inodoratum Bluggoe (ABB) inodoratum Pisang Awak (ABB) inodoratum Silk/Rasthali (AAB) odoratum Silk/Rasthali (AAB) GCV 0122 Cruz das Almas, Brazil Podavur, Tamil Nadu, India GCV 01219 Latundan (AAB) odoratum odoratum Cavendish (AAA) Pisang Lilin (AA) odoratum odoratum odoratum odoratum odoratum odoratum odoratum odoratum odoratum odoratum Bluggoe (ABB) Burro CEMSA (ABB) Chinese Cavendish (AAA) FHIA-03 (AABB) GCTCV 215 (AAA) FHIA-17 (AAAA) FHIA-23 (AAAA) Gros Michel (AAA) Williams (AAA) Yangambi Km5 (AAA) GCV 0120/15 odoratum odoratum odoratum odoratum odoratum odoratum odoratum FHIA-01 (AAAB) FHIA-03 (AABB) FHIA-23 (AAAA) Grande Naine (AAA) PV 03-44 (AAAB) Williams (AAA) Yangambi Km5 (AAA) Bluggoe (ABB) odoratum FHIA-23 (AAAA) odoratum GCTCV 215 (AAA) odoratum Pisang Ceylan (AAB) odoratum Pisang Mas (AA) odoratum Pisang Nangka (AAB) odoratum PV 03-44 (AAAB) Chiuju, Pingtung, Taiwan odoratum Williams (AAA) GCV 0121 odoratum Yangambi Km5 (AA) odoratum Kepok odoratum Bluggoe (ABB) odoratum Burro Cemsa (ABB) odoratum odoratum odoratum odoratum odoratum odoratum Cavendish (AAA) Gros Michel (AAA) PA 03.22 (AAAB) Pisang Ceylan (AAB) Pisang Mas (AA) Yangambi Km5 (AAA) odoratum FHIA-17 (AAAA) GCV 01213/16 odoratum FHIA-23 (AAAA) odoratum Pisang Berangan (AA) odoratum odoratum odoratum odoratum Bluggoe (ABB) FHIA-23 (AAAA) Gros Michel (AAA) PA 03.22 (AAAB) odoratum Pisang Ceylan (AAB) odoratum Pisang Mas (AA) Kichwamba, Bushenyi District, Uganda odoratum PV 03-44 (AAAB) GCV desconocido - No Foc odoratum Williams (AAA) odoratum Yangambi Km5 (AAA) ? ? ? Pisang Nangka (AAB) Williams (AAA) Yangambi Km5 (AAA) Ensayos sobre marchitamiento por Fusarium Consideraciones generales Los rendimientos fueron muy variables y dependían del sitio, tal como se esperaba. Ya que muchos factores, incluyendo el manejo, latitud, temperatura, presión del patógeno, etc., influyen sobre el desempeño agronómico, no es posible generalizar los resultados. La Figura 8 proporciona un sumario del peso promedio del racimo por genotipo y por sitio, considerando sólo los cultivares mejorados y locales, Yangambi Km5 y Gros Michel. El sitio más favorable desde el INFOMUSA — Vol 8, N° 1 inodoratum odoratum Selangor, Malasia GCV 01213/16 FHIA-17 (AAAA) Pisang Mas (AA) Pisang Awak (ABB) Burro CEMSA (ABB) Islas Canarias, España Solok, Indonesia GCV 01213/16 odoratum odoratum odoratum odoratum Sto Tomas, Davao del Norte, Mindanao, Filipinas odoratum GCV 01213/16 Genotipos de Musa Bago Oshiro, Davao City, Filipinas odoratum GCV 0124/5 Producción volátil GCV 0123 Wamuran, Australia GCV 0120 Localización y GCV Johor, Malasia Cudgen, Australie punto de vista de peso del racimo fue Taiwan donde las prácticas de manejo están muy desarrolladas. Los segundos mejores resultados fueron obtenidos en el sitio en Uganda. Es necesario notar que este sitio se encontraba en el campo de un agricultor, lo que indica el alto nivel de prácticas de manejo de los productores bananeros en esta área. La decoloración interna del rizoma se prorrateó entre 1 y 6, donde 1 indicaba la ausencia de decoloración ; 2 – puntos aislados de decoloración ; 3 – la decoloración de hasta un tercio del tejido vascular ; 4 – la decoloración entre 1/3 y 2/3 del tejido vascular ; 5 – la decoloración de más de 2/3 del tejido y 6 – la decoloración total. Los programas de mejoramiento han utilizado la decoloración interna del rizoma para estimar la resistencia del genotipo al Foc. Asimismo, este valor combinado con el desempeño agronómico es una medida de la tolerancia de la accesión. Sin embargo, en el caso de la fase II del IMTP era bastante difícil hacer una estimación total de la resistencia o tolerancia del genotipo de la planta a Foc debido a grandes diferencias de tasas a través de los sitios. Se 7 Sitio (todos los genotipos) Genotipo (todos los sitios) Peso del racimo (Kg) 29 23 17 11 FH IA FH -01 IA FH -03 IA FH -17 IA PV -2 03 3 P A .4 4 G CT 0 3 2 C 2 G V1 CT 1 CV 9 2 cv 15 Ya lo ng ca l G ro Km sM 5 ic he l A us tr al i Br a H on azi d l In ur do a s n Fi lip M esi a Fi (B ala lip a g s i a o (S to Os T o h) Su ma da s) fr i Es ca pa T a ña iw U an ga nd a 5 Figura 8. Promedios para el peso del racimo. Sitios con Foc – Cultivares mejorados. considera que las diferencias se debieron no sólo a las diferencias en la presión del patógeno, sino también a un elemento subjetivo de interpretación visual. El promedio de los síntomas internos para cada sitio puede ser considerado como una medida general de la presión del patógeno confundida con el efecto de la interpretación visual (Figura 9). Esto particularmente es verdad para los promedios de decoloración a través de los sitios infestados con la raza 1 del Foc, ya que hubo sólo tres sitios infestados con esta raza en todo el ensayo del IMTP y uno de ellos (Bago Oshiro en Filipinas) tenía promedios excepcionalmente altos (Figura 9). Finalmente, los resultados del análisis de los GCV se presentan en la Tabla 3. Cultivares mejorados Sitios infestados con la raza 1 del Foc Hubo sólo tres sitios en el ensayo infestados con la raza 1 del Foc : Brasil, Filipinas (Bago Oshiro) y Honduras. Como ya hemos mencionado, el sitio en Filipinos tenía promedios extremadamente altos. Sin embargo, de las numerosas muestras, enviadas para el análisis de los GCV desde este sitio, sólo cuatro rindieron Foc. De estos cuatro, tres aislados resultaron ser GCV sin determinar y sólo uno pertenecía a la raza 1. Por lo tanto, los promedios totales mencionados abajo en este resumen, deben ser considerados con prudencia. Para realizar las comparaciones con el clon de referencia y tasas de resistencia o tolerancia es preferible utilizar los puntajes promedio de cada genotipo dentro del sitio (Tabla 2). El puntaje promedio, considerando todos los genotipos a través de los sitios infestados con la raza 1 del Foc fue de 2.4. El clon de referencia susceptible Gros Michel tuvo un puntaje promedio de 3.2 a través de los sitios. El genotipo mejorado con el menor puntaje de decoloración a través de los sitios fue el GCTCV 119. Este clon tuvo un 8 puntaje promedio total de 1.3, lo que refleja su resistencia a la raza 1 del Foc. El genotipo con el segundo puntaje más bajo de decoloración fue el FHIA-23. Este híbrido tiene un puntaje de decoloración de 2.1 en los sitios infestados con la raza 1 del Foc. Aunque este puntaje fue inferior al promedio total y el más bajo para la raza 1 del Foc de todos los híbridos de la FHIA, el mismo ya indica una infección en el cormo. Debido a su excelente peso de racimo (Figura 8), este híbrido puede ser clasificado como muy tolerante. Aún falta realizar investigaciones para verificar si sus futuras generaciones retienen su tolerancia. El FHIA-01 mostró un puntaje promedio de 2.2. Aunque los puntajes del FHIA-01 ya indican más que sólo puntos aislados de decoloración en el tejido vascular, ellos permanecen aún más bajos que el puntaje promedio general. A pesar de que el peso de los racimos de los híbridos de la FHIA es el más bajo, el FHIA-01 mostró la varianza más baja a través de todos los sitios. Estos resultados permiten concluir que el FHIA-01 es tolerante a la raza 1 del Foc, que se desempeña bien bajo una variedad de ambientes y que responde positivamente a las buenas condiciones de manejo. El PV 03.44 tuvo un puntaje promedio de 2.3 en los sitios infestados con la raza 1, lo que estaba justo por debajo del promedio general para estos sitios. Sin embargo, el peso de su racimo fue usualmente bajo con promedio general de 8.4 kg y un promedio máximo de 11.3 kg en Súd Africa. El GCTCV 215 tuvo un puntaje promedio de 2.5 para los sitios con la raza 1, mayor que el del GCTCV 119 y ligeramente mayor que el promedio general. Sin embargo, es necesario notar que este promedio está substancialmente influenciado por los puntajes excepcionalmente altos (5.6) registrados en un sitio en Filipinas (Bago Oshiro). En los otros dos sitios infestados con la raza 1, este genotipo tuvo un puntaje promedio de 1 (ninguna decoloración). Ya que este genotipo siempre fue considerado como resistente a la raza 1, estos resultados contradictorios merecen una nueva evaluación. El FHIA-17 tuvo puntajes de decoloración por encima del promedio. Su puntaje promedio fue de 2.5 a través de los sitios. Sin embargo, como en el caso del GCTCV 119, es necesario notar que este promedio está substancialmente influenciado por los puntajes excepcionalmente altos (5.6) registrados en Filipinas (Bago Oshiro). En los otros dos sitios infestados con la raza 1, este genotipo tuvo un puntaje promedio de 1 (ninguna decoloración), lo que se ha de esperar de los clones Cavendish. El FHIA-17 fue el genotipo con el mejor rendimiento en todos los sitios. El mismo tenía un peso de racimo promedio de 25.2 kg. La FHIA ha informado que este genotipo es resistente a la raza 1 del Foc. Estos descubrimientos indican resultados contradictorios entre los sitios, lo que amerita realizar más investigaciones. El FHIA-03 tuvo un puntaje promedio de decoloración vascular de 3.08 a través de los sitios con la raza 1 de Foc, el puntaje más alto de todos los genotipos mejorados y muy similar al del Gros Michel, la referencia susceptible. Este puntaje está por encima del puntaje promedio total, cuando se toman en consideración todos los genotipos participantes, y ciertamente indica susceptibilidad. A pesar de su alto puntaje de decoloración, este híbrido dio algunos pesos del racimo muy buenos con un promedio de 20.4 kg a través de los sitios y un promedio máximo de 29.8 kg. Se puede decir que este genotipo muestra tolerancia en el primer ciclo de cultivo. Sería interesante verificar si la tolerancia se mantiene en las generaciones posteriores. Sitios infestados con la raza 4 del Foc Desafortunadamente, el cultivar Williams, la referencia susceptible para los sitios infestados con la raza 4, fue una variante somaclonal enana que no pudo ser utilizada para las comparaciones. El puntaje promedio de decoloración total para los sitios con la raza 4 fue de 2.4 considerando todos los genotipos. El GCTCV 119 tuvo un puntaje promedio de 1.3 a través de los sitios infestados con la raza 4. Como en los sitios infestados con la raza 1, este también fue el puntaje de decoloración más bajo de todos los cultivares mejorados a través de los sitios del ensayo. Sin embargo, existen indicaciones de que este genotipo puede ser bastante susceptible a la raza 4 del Foc. Aunque no se registraron infecciones para este genotipo en el sitio en Indonesia, las muestras de este genotipo enviadas anteriormente al QDPI confirmaron la infección. Su peso promedio del racimo fue de 10 kg, considerando su lado más bajo. Este genotipo fue muy variable con buenos pesos promedio del racimo entre 15 kg y 22 kg bajo las mismas condiciones y muy bajo (3 kg) INFOMUSA — Vol 8, N° 1 Genotipo (todos los sitios) Sitio (todos los genotipos) Síntomas internos 5 4 3 2 Raza 1 FHIA-01 FHIA-03 FHIA-17 FHIA-23 PV 03.44 PA 0322 GCTCV 119 GCTCV 215 Burro C emsa Saba P nangka CV rose PJB P lilin Bluggoe Wiliams P ceylan local cv P mas Yang Km 5 Calcutta Gros Michel iw ) an ña sh Ta iro pa O Es ag o Fi lip (B H o To on mas d ) Su ur da as fr ic a ia es on (S t In d Fi lip A us tr a Br lia az il 1 Raza 4 Figura 9. Promedios para los síntomas internos – razas 1 y 4 del Foc. en otras. Se cree que las prácticas de manejo tienen una gran influencia sobre el desempeño agronómico de este genotipo. El GCTCV 215 tuvo un puntaje promedio de 1.7 si no se consideran los datos de Indonesia, donde sólo una planta fue evaluada. Este promedio fue el segundo más bajo de todos los genotipos mejorados. Esta variante somaclonal tuvo un peso promedio del racimo de 11.9 kg a través de los sitios. Sin embargo, este promedio aumenta hasta 12.7 kg si los datos de Indonesia no se toman en consideración. Ya que este genotipo siempre fue considerado resistente a la raza 1 y susceptible a la raza 4, estos resultados contradictorios merecen una nueva evaluación. El FHIA-01 mostró una decoloración promedio de 2.04 a través de los sitios infestados con la raza 4 del Foc. El mismo tuvo un peso promedio del racimo de 17.6 kg a través de los sitios y un promedio máximo de 24 kg en condiciones óptimas de manejo. Dado el buen desempeño del FHIA-01 a través de los sitios, este genotipo puede ser clasificado tolerante a las razas 1 y 4 del marchitamiento por Fusarium. El PV 03.44 tuvo un puntaje de 2.1. Este puntaje estuvo por debajo del promedio total. Sin embargo, su peso del racimo usualmente fue bajo con un promedio general de 8.4 kg y un promedio máximo de 11.3 kg en Africa del Sur. El PA 03.22 también tuvo un puntaje bajo (2.3) en los sitios infestados con la raza 4. Similarmente al PV 03.44, este puntaje también fue por debajo del promedio general a través de los sitios y más bajo que el de las referencias susceptibles. Sin embargo, su peso del racimo también fue usualmente muy bajo con un promedio total de 6.4 kg y un promedio máximo de 11.6 kg en Africa del Sur. El FHIA-17 dio un puntaje promedio de 2.5 a través de los sitios, el mismo para los sitios con las razas 1 y 4 del Foc. Este puntaje está por encima del promedio general y INFOMUSA — Vol 8, N° 1 en Taiwan fue de 4.3. No obstante, el FHIA17 produjo los racimos más pesados de todos los genotipos en estos ensayos. El mismo tuvo un peso promedio del racimo de 25.2 kg a través de los sitios y un máximo de 43.8 kg en Taiwan. Nuestros resultados indican que este híbrido es muy tolerante a la raza 4 durante el primer ciclo de cultivo de la planta. Queda investigar si las generaciones posteriores retienen este nivel de tolerancia. El FHIA-03 tuvo un puntaje promedio de 3.09, lo que significa que alrededor de un tercio de tejido vascular mostró decoloración en ambos tipos de sitios. Estos puntajes están por encima del puntaje promedio total, cuando se consideran todos los genotipos participantes, y ciertamente indican susceptibilidad. Además, en algunos sitios, el FHIA-03 tuvo puntajes de decoloración muy altos (de 4.3 a 5.7) como en las Islas Canarias y Australia. A pesar de sus altos puntajes de decoloración, las plantas sobrevivientes de este híbrido dieron muy buenos pesos del racimo con un promedio de 20.4 kg a través de los sitios y un promedio máximo de 29.8 kg. La única excepción a esto se dio en las Islas Canarias. En este caso particular los datos se recolectaron sólo de dos plantas. El FHIA-23 tuvo un puntaje promedio de 3.1, muy por encima del promedio general, indicando susceptibilidad. Además, en Australia y en las Islas Canarias el mismo tuvo promedios por encima de 4 indicando una alta susceptibilidad a la raza 4 subtropical. A pesar de sus altos puntajes de decoloración, este híbrido fue el segundo mejor rendidor de todos los híbridos de la FHIA con un peso promedio del racimo de 22.3 kg y un promedio máximo de 46.8 kg en Taiwan. Cultivares indígenas El Bluggoe fue el genotipo con los mayores puntajes de decoloración en ambos tipos de sitios. El mismo tuvo un puntaje de 3.5 en los sitios con la raza 1 y 4.1 en los sitios con la raza 4, indicando una alta susceptibilidad a ambas razas del Foc. Otro genotipo que mostró susceptibilidad a ambas razas fue el Gros Michel con un puntaje de 3.2 para los sitios con la raza 1 y 3.5 para los sitios con la raza 4. Otros dos cultivares indígenas mostraron puntajes de decoloración superiores al puntaje general para ambos tipos de sitios, estos fueron el Burro CEMSA y Saba. En los sitios con la raza 1 del Foc, el Burro CEMSA y Saba tuvieron el mismo promedio general de 2.8. En los sitios infestados con la raza 4 del Foc, el Burro CEMSA tuvo un puntaje de 2.9 y el Saba, de 3. Algunos cultivares indígenas, por ejemplo, el Yangambi Km5 y el Pisang Mas tuvieron una reacción muy específica a la raza del Foc. El Yangambi tuvo un puntaje de 2.1 que fue más bajo que el promedio general para los sitios con la raza 1, pero tuvo un puntaje de 4.1, muy por encima del promedio para los sitios con la raza 4. La diferencia, aunque menos marcada, también fue evidente para el Pisang Mas. En los sitios con la raza 1, el Pisang Mas tuvo un puntaje de 2.06, mientras que para los sitios con la raza 4, su puntaje fue de 3.5. Los cultivares indígenas que consistentemente tuvieron puntajes más bajos que el promedio fueron el Cultivar Rose y el Pisang Nangka. Conclusiones Los híbridos de la FHIA consistentemente fueron los genotipos con mejor rendimiento en estos ensayos. Con pocas excepciones, sus racimos fueron más pesados que los racimos producidos por todos los otros cultivares, mejorados y locales. Los híbridos de la FHIA también respondieron muy bien al manejo cuidadoso y aplicación de fertilizantes, como en el caso de Taiwan. En resumen, los híbridos de la FHIA se desempeñaron bien bajo un rango de condiciones y respondieron aún mejor cuando las condiciones mejoraron. Un cultivar mejorado que merece una referencia especial es el GCTCV 119, que tuvo el puntaje de decoloración más bajo para ambas razas del Foc y buenos rendimientos bajo un buen manejo. Hemos tratado de combinar los datos de los pesos del racimo con los datos sobre la reacción al Fusarium y a la Sigatoka negra para dar una indicación de desempeño total y comparativo de las variedades en diferentes situaciones de enfermedad. Es importante subrayar que la resistencia sola no es útil. Se debe combinar con una buena producción, característica postcosecha y organolépticas aceptables. Las variedades mejoradas de banano contribuyen no solo a reducir la incidencia de la enfermedad, sino también a mejorar la producción de alimentos. Los datos completos y el análisis de los resultados se publicarán en breve. Para recibir ejemplares del informe final, por favor contacte el coordinador del IMTP en INIBAP. 9 Agradecimiento La fase II del IMTP representa el esfuerzo de muchos investigadores dedicados y programas de mejoramiento, programas nacionales de investigación agrícola y agencias de financiamiento alrededor del mundo. Gracias especiales a los administradores de los sitios por enviar sus datos, a los programas de mejoramiento que donaron germoplasma, al Dr. Mauricio Rivera de la FHIA por sus comentarios para este artículo y la Dra. Suzy Bentley del CRCTPP por Recursos genéticos su contribución a la impresión de huellas de ADN. ■ Nota : El Yangambi Km5 se incluye sólo como elemento de interés, dado que esta fue la accesión más solicitada en el Centro de Tránsito de INIBAP. Se incluye el cultivar local como referencia. G. Orjeda y J.V. Escalant trabajan en INIBAP, Parc Scientifique Agropolis, 34397 Montpellier, Cedex 5 Francia, y N. Moore, en el Queensland Department of Primary Industries, 80 Meiers Road, Indooroopilly, Qld 4068, Australia. Ensayos en Taiwan Desempeño de los clones de banano frente al reto del marchitamiento por Fusarium en Taiwan C. Y. Tang y S. C. Hwang l marchitamiento por Fusarium causado por Fusarium oxysporum f. sp. cubense es un problema serio para la producción bananera de Taiwan (Su et al. 1986). Se estimó que más de 1 500 ha., alrededor de un tercio del área de cultivo de banano en el sur de Taiwan, fue afectada por esta enfermedad. Tanto la raza 1 (GCV 0123) y la raza 4 (GCV 0121, 01213) del patógeno del Fusarium fueron reportadas en Taiwan (Pegg et al. 1992). Para la búsqueda de las diferentes fuentes de resistencia, un ensayo en el marco del IMTP organizado por INIBAP, fue llevado a cabo durante 1996-1997 en Pingtung, Taiwan. Veintiún clones más un cultivar local fueron incluidos en la evaluación frente al reto de esta enfermedad. Este trabajo es el informe de los resultados de este ensayo. E Materiales y métodos Veintiún clones experimentales (Tabla 1) fueron suministrados en forma de plántulas cultivadas a partir de tejidos por el ITC-INIBAP en 1995. Después de atravesar el proceso de cuarentena fitosanitario, las plántulas fueron transplantadas en potes colocados bajo una red por cinco meses. El ensayo en el campo fue establecido el 23 de mayo de 1996 en un vivero infectado, con suelo limoso con un pH 5-5.3 en el Taiwan Banana Research Institute (TBRI), Pingtung, Taiwan (N 22° 42’, E 120° 29’). Todos los clones se establecieron con éxito, con excepción del #16 (ITC0001, Pisang Lilin) que murió durante el proceso de aclimatación o en una etapa temprana después de trasladarlo al campo. Un cultivar Cavendish local, Tai-Chiao No. 2, fue utilizado como referencia. Las plantas fueron sembradas en una red de 2 m x 1.5 m. Catorce plantas de cada accesión fueron arregladas en un di10 seño aleatorio completo. Cada planta fue tratada como una unidad experimental. Los datos sobre los síntomas de la enfermedad y características agronómicas fueron tomados de acuerdo a las ‘Guías Técnicas para los sitios con Fusarium para la Fase II de IMTP’ (‘IMTP Phase II Technical Guidelines for Fusarium Sites’— INIBAP 1994). Los síntomas externos de la severidad de la enfermedad incluían los siguientes : amarillamiento del follaje, rajadura de la base del seudotallo, decoloración vascular en la base foliar, internudos cortos (prorrateados de 1 a 3) y cambios en las hojas nuevas (prorrateados de 1 a 2). Los datos de los síntomas externos fueron tomados una vez al mes empezando desde el cuarto al duodécimo mes después de la plantación. A la cosecha o cuando la planta resultó totalmente marchitada, los síntomas internos se determinaban examinando la decoloración en la porción más baja del cormo (prorrateados de 1 a 6). Los datos agronómicos, que comprenden la cantidad de días desde la plantación hasta la aparición del retoño y hasta la cosecha, altura de la planta al tiempo de la cosecha (cm), circunferencia del seudotallo a 30 cm del nivel del suelo (cm), altura del retoño seguidor al tiempo de la cosecha, cantidad de manos y dedos en el racimo, peso promedio de cada dedo (g), fueron registrados durante la floración o la cosecha. Resultados y discusión Síntomas externos de la enfermedad Cuatro meses después de la plantación, se registraron seis criterios de los síntomas externos del marchitamiento por Fusarium en el campo. En la Tabla 1 se presenta un resumen de los datos que muestran la severidad y la tasa del desarrollo de la enfermedad 12 meses después de la plantación. En general, los síntomas externos, como el amarilleamiento del follaje, rajadura del seudotallo, decoloración vascular y el marchitamiento representaron indicaciones claras del desarrollo de la enfermedad. Otros criterios como el cambio de nuevas hojas y el acortamiento de internudos no fueron distintivos. El síntoma de torcedura del pecíolo no fue obvio y por lo tanto, no fue registrado. De acuerdo a los síntomas externos, 21 cultivares pueden ser clasificados de la siguiente manera : 1. Resistente : SH-3481, SH-3565, SH-3444, GCTCV-119, GCTCV-215, Cultivar Rose, Pisang Jari Buaya, Calcutta IR 124, Williams y Tai-Chiao No. 2 (cultivar local) ; 2. Intermedio : SH-3649, Pisang Mas, Pisang Nangka, Pisang Ceylan ; 3. Susceptible : PV03.44, Burro Cemsa, Saba, Yangambi km5 ; 4. Altamente susceptible : PA03.22, Gros Michel, Bluggoe. Los clones altamente susceptibles, PA03.22, Gros Michel y Bluggoe mostraron los síntomas más severos así como el desarrollo más rápido de la enfermedad. Al quinto mes después de la plantación, alrededor de un tercio de las plantas mostraron el follaje amarillo y la rajadura del seudotallo. Más de la mitad de las plantas se marchitó 8 meses después de la plantación. Todos los clones investigados, como el SH-3481, SH-3565, SH3444, GCTCV-119, GCTCV-215, Cultivar Rose, Pisang Jari Buaya, Calcutta IR 124, Williams y Tai-Chiao No. 2 (cultivar local), mostraron ninguno o sólo ligeros síntomas externos. Síntomas internos Los síntomas internos, como los indicados por la decoloración vascular del rizoma, fueron examinados después de que la planta se marchitó debido a la enfermedad o a la cosecha. Las señales de los síntomas internos y las frecuencias de las plantas que muestran diferentes grados de severidad son mostradas en la Tabla 2. De acuerdo a esta observación, la clasificación de 21 cultivares se realizó como sigue : 1. Resistente : SH-3481, SH-3565, GCTCV-215, Cultivar Rose, Pisang Jari Buaya, Calcutta IR 124. 2. Intermedio : SH-3444, PV03.44, GCTCV-119, Tai-Chiao No. 2. 3. Susceptible : SH-3649, PA03.22, Burro CEMSA, Saba, Pisang Nangka, Yangambi Km5, Pisang Ceylan, Williams. 4. Altamente susceptible : Pisang Mas, Gros Michel, Bluggoe. Entre los cultivares resistentes, el SH3481, Calcutta IR 124 y el Cultivar Rose no mostraron ningunos síntomas de la enfermedad en todas las plantas investigadas. Estos cultivares pueden ser considerados altamente resistentes. Entre los cultivares GCTCV-215 y Pisang Jari Buaya, alrededor del 10 % de las plantas mostraron síntomas internos, por lo tanto estos cultivares fueron considerados resistentes. Las plantas enfermas de los clones susceptibles Gros Michel, Bluggoe y Burro CEMSA siempre mostraron la pudrición del rizoma. Aunque los síntomas INFOMUSA — Vol 8, N° 1 Tabla 1. Severidad del marchitamiento por Fusarium después de 12 meses de plantación en el ensayo desarrollado en el marco del IMTP y llevado a cabo en Taiwan. Código 01 02 03 04 05 06 07 08 09 10 11 12 13 14 15 16 17 18 19 20 21 Clon SH-3481 SH-3565 SH-3649 SH-3444 PV 03.44 PA 03.22 GCTCV 119 GCTCV 215 Burro CEMSA Pisang Mas Saba Pisang Nangka Cultivar Rose Yangambi km 5 Pisang Jari Buaya Calcutta IR 124 Gros Michel Bluggoe Williams Pisang Ceylan Tai-Chiao No. 2 Genoma Amarilleamiento del follaje (1) AAAB AABB AAAA AAAA AAAB AAAB AAA AAA ABB AA BBB AAB AA AAA AA AAw AAA ABB AAA AAB AAA 1.0 1.0 1.5 1.2 2.1 2.2 1.1 1.2 2.5 1.6 2.8 1.5 1.0 2.5 1.2 1.2 2.8 2.9 1.2 1.5 1.1 Rajaduras en la base del seudotallo 1.0 1.0 1.2 1.0 1.6 2.3 1.0 1.0 1.6 1.4 1.6 1.6 1.0 2.1 1.1 1.0 2.3 2.3 1.0 1.6 1.0 1.7 1.4 Promedio Decoloración del tejido vascular 1.5 1.0 1.5 1.2 2.0 2.3 1.0 1.1 2.5 1.6 2.7 1.7 1.1 2.4 1.2 1.0 2.6 2.9 1.3 1.5 1.4 Cambios en las nuevas hojas 1.7 Internudos cortos Marchitamiento 1.0 1.0 1.2 1.0 1.1 1.6 1.0 1.0 1.3 1.1 1.1 1.2 1.0 1.5 1.0 1.0 1.8 1.6 1.1 1.0 1.0 1.0 1.0 1.4 1.0 1.4 2.2 1.0 1.0 1.5 1.2 1.7 1.3 1.0 2.1 1.0 1.0 2.8 2.4 1.2 1.1 1.0 1.0 1.0 1.4 1.0 1.9 2.3 1.0 1.0 1.9 1.4 2.6 1.6 1.0 2.0 1.0 1.0 2.7 2.9 1.1 1.6 1.0 1.2 1.4 1.6 1) Afuera del cambio en las nuevas hojas, todos los parámetros fueron prorrateados de 1 a 3 donde 1 = síntomas ausentes y 3 = síntomas severos. El cambio en las nuevas hojas esta prorrateado 1 (= síntomas ausentes) y 2 (= sintomas presentes). Tabla 2. Ocurrencia del marchitamiento por Fusarium determinada por síntomas internos en el ensayo desarrollado en el marco del IMTP y llevado a cabo en Taiwan. Código 01 02 03 04 05 06 07 08 09 10 11 12 13 14 15 17 18 19 20 21 22 Clon SH-3481 SH-3565 SH-3649 SH-3444 PA 03.44 PV 03.22 GCTCV 119 GCTCV 215 Burro CEMSA Pisang Mas Saba Pisang Nangka Cultivar Rose Yangambi km 5 Pisang Jari Buaya Calcutta IR 124 Gros Michel Bluggoe Williams Pisang Ceylan Tai-Chiao No. 2 Genoma Nùmero de plantas evaluadas AAAB AABB AAAA AAAA AAAB AAAB AAA AAA ABB AA BBB AAB AA AAA AA AAw AAA ABB AAA AAB AAA 12 13 9 8 11 14 7 10 11 11 13 14 12 13 9 10 14 14 5 13 13 Promedio (1) Valor 1-6 (1) 1.0 1.7 4.3 2.3 2.8 4.3 2.3 1.2 4.9 5.7 4.8 3.5 1.0 4.8 1.1 1.0 5.9 5.9 3.2 3.4 2.3 DE 0.00 1.54 2.17 1.58 1.94 2.23 0.63 0.53 1.30 0.64 1.46 2.31 2.21 1.92 2.31 0.00 1.84 0.53 2.28 0.33 2.14 3.2 Min. Max. 1 1 1 1 1 1 1 1 3 4 2 1 1 1 1 1 4 4 1 1 1 1 6 6 5 6 6 6 3 6 6 6 6 1 6 2 1 6 6 6 6 6 1.6 4.9 % de plantas con síntomas Ausentes 100.0 76.9 22.2 50.0 36.4 28.6 71.4 90.0 0.0 0.0 0.0 50.0 100.0 15.4 88.9 100.0 0.0 0.0 40.0 30.8 53.8 45.4 ligeros 0.0 7.7 11.1 25.0 27.2 0.0 0.0 10.0 18.2 0.0 23.1 0.0 0.0 7.7 11.1 0.0 0.0 0.0 20.0 23.1 30.7 10.2 severos 0.0 15.4 66.7 25.0 36.4 71.4 28.6 0.0 81.8 100.0 76.9 50.0 0.0 76.9 0.0 0.0 100.0 100.0 40.0 46.2 15.3 44.3 Síntomas internos tienen una escala de 1 a 6 donde 1 = síntomas ausentes y 6 = síntomas severos. externos no fueron severos en el Pisang Mas en este ensayo, la ocurrencia de síntomas internos intensivos indicó que este cultivar es altamente susceptible. Datos agronómicos Debido a la escasez de tierra apropiada para conducir este experimento, la densidad de plantas en este ensayo fue más alta que la normal. En consecuencia, las plantas de este ensayo fueron altas y demoraron en su desarrollo, especialmente los clones como Cavendish y Pisang Mas que son menos vigorosos en desarrollo. Los datos agronómicos de este ensayo son mostrados en la Tabla 3. Entre los INFOMUSA — Vol 8, N° 1 clones resistentes, el Cultivar Rose fue el cultivar más precoz con un seudotallo delgado y racimo pequeño. Por otra lado, el Pisang Jari Buaya fue el cultivar más tardío con un seudotallo alto. Los híbridos de la FHIA (SH3481, SH-3565, SH-3649 y SH-3444) fueron los cultivares más robustos con seudotallos fuertes y racimos grandes. El GCTCV-215, un clon Cavendish, mostró un nivel moderado del peso de racimo en este ensayo. Conclusión En el transcurso del ensayo, las muestras del patógeno tomadas de los haces vasculares de las plantas enfermas, fueron enviadas a Aus- tralia para su identificación. Los GCV 0121, 01213 y 01216 fueron encontrados en estas muestras. Estos GCV tienen patrones de bandas de ADN similares y probablemente pertenecen al mismo grupo (N. Moore, comunicación personal). Por lo tanto, la observación en este ensayo debería ser interpretada como la reacción del patógeno ‘GCV 0121 a los genotipos de los bananos investigados. En este ensayo se observaron varios clones resistentes, incluyendo los híbridos mejorados, variantes somaclonales y cultivares nativos. Algunos de los híbridos mejorados, desarrollados por la FHIA, mostraron un alto nivel de resistencia y un racimo grande. El SH-3481 (FHIA-01) fue 11 Tabla 3. Características agronómicas de 21 clones en el ensayo desarrollado en el marco del IMTP y llevado a cabo en Taiwan. Código Clon Genoma Días desde la siembra hasta la parición Días desde la siembra hasta la cosecha Altura de Circumferenla planta cia del (cm) seudotallo (cm) Altura de vàstago (cm) Peso del racimo (kg) Nùmero Nùmero Peso de de promedio manos dedos del dedo (g) 01 SH-3481 AAAB 185 355 289 75.0 193 24.2 9.5 145 165 02 SH-3565 AABB 225 373 365 97.0 286 29.8 8.8 156 199 03 SH-3649 AAAA 318 401 346 94.0 204 43.9 12.1 205 209 04 SH-3444 AAAA 327 418 376 98.6 250 46.8 14.8 282 168 05 PV 03.44 AAAB 206 417 372 68.2 348 11.0 7.3 119 142 06 PV 03.22 AAAB 149 377 252 62.0 218 5.4 7.6 97 57 07 GCTCV 119 AAA 348 431 359 74.9 179 22.2 10.2 207 109 08 GCTCV 215 AAA 300 402 296 61.0 134 20.5 7.6 117 169 09 Burro CEMSA ABB 217 386 405 79.3 304 23.9 8.2 128 194 10 Pisang Mas AA 362 - 440 69.0 - 11 Saba BBB 215 375 409 80.6 317 23.5 - 7.7 - - 119 196 254 12 Pisang Nangka AAB 277 392 405 79.6 259 26.2 6.5 102 13 Cultivar Rose AA 167 273 234 38.6 209 3.2 9.2 134 24 14 Yangambi Km5 AAA 271 380 354 74.7 338 27.1 8.8 187 145 15 Pisang Jari Buaya AA 343 423 433 67.4 331 20.2 9.9 170 120 17 Calcutta IR 124 AAw 183 400 499 48.8 355 1.6 7.7 169 9 18 Gros Michel AAA 278 - - - - - 19 Bluggoe ABB 226 - 402 - - - 74.3 - 23 6.3 100 20 Williams AAA 355 393 158 53.7 105 24.0 6.0 139 21 Pisang Ceylan AAB 243 372 383 77.7 377 25.8 10.9 202 135 22 Tai-Chiao No. 2 AAA 293 408 299 74.7 160 26.8 10.4 190 139 260 388 343 72.4 254 156 145 Promedio 173 22.6 8.9 CV 16.3 8.8 8.9 9.1 26.8 24.9 10.8 21.7 21.7 DE 52.1 46.9 35.7 7.7 96.6 8.1 1.8 52.1 45.6 el mejor en términos de la altura de la planta, ciclo de desarrollo y nivel de resistencia. Sin embargo, la textura y el sabor de la fruta no son aceptables para los consumidores de Taiwan, quienes están acostumbrados al sabor del Cavendish o Latundan. Entre los clones Cavendish, el GCTCV-215 desarrollado por el TBRI a través de la selección a partir de las variantes somaclonales, continúa mostrando un buen nivel de resistencia al marchitamiento por Fusarium. El nivel de rendimiento y la calidad de consumo es similar a su clon materno ‘Pei-Chiao’, un Cavendish Gigante. Este clon fue registrado como ‘Tai-Chiao No. 1’y fue liberado a los productores con el fin de controlar el marchitamiento por Fusarium en Taiwan (Hwang et al. 1992). Para desarrollar nuevos cultivares con sabor especial distinto al del Cavendish, el Cultivar Rose puede ser el candidato para este propósito y será sujeto a más investigaciones. Agradecimiento Los autores agradecen a la Dra G. Orjeda por animarnos a publicar los resultados de este experimento. Este proyecto de investigación fue financiado por el Council of Agriculture (87AST-1.2-FAD-24). ■ INIBAP. 1994. IMTP Phase II Technical guidelines for Fusarium wilt sites. INIBAP. Montpellier, France. Pegg K.G., N.Y. Moore & S. Sorensen. 1992. Fusarium wilt in the Asian Pacific Region. Pp. 162-171 in Proceedings : International symposium on recent developments in banana cultivation technology (Valmayor R.V., S.C. Hwang, R. Ploetz & Recursos genéticos 12 Sun H.J., S.C. Hwang & W.H. Ko. 1986. Fusarial wilt of Cavendish banana in Taiwan. Plant Dis. 70 : 115-119. Los autores trabajan en el TBRI, P.O. Box 18, Chiuju, Pingtung, Taiwan 904. Resistencia a nematodos Estudio de los bananos procedentes de Papua Nueva Guinea con respecto a los nematodos noduladores y de lesiones de la raíz Ruth Stoffelen, Raf Verlinden, Ngo Thi Xuyen, Rony Swennen & Dirk De Waele Bibliografía Hwang S.C., W.H. Ko & C.P. Chao. 1992. Control of fusarial wilt of Cavendish banana by planting a resistant clone derived from breeding. Pp. 259280 in Proceedings of the Symposium on the non-chemical control of crop diseases and pests. Plant Protection Bulletin, special publication. The Plant Protection Society of the Republic of China. Taipei, Taiwan. S. W. Lee, eds). INIBAP/ASPNET, Los Baños, Philippines. apua Nueva Guinea posee una gran diversidad de Musa spp., incluyendo los bananos comestibles Fe’i y diploides AA. Esta es una de las últimas áreas que quedan donde los diploides AA todavía se cultivan en gran escala (Jarret et al. 1992). Actualmente, casi en todas las partes del Sureste de Asia, los triploides han reempla- P zado a los diploides AA comestibles debido a que tienen frutas y racimos más grandes (Simmonds 1962). Sin embargo, los programas de mejoramiento de banano y plátano dependen del desarrollo de los diploides agronómicamente superiores con resistencia a plagas y enfermedades (Stover y Buddenhagen 1986, Tenkouano et al. 1998). Durante los años 1988 y 1989, el International Board for Plant Genetic Resources (IBPGR, actualmente IPGRI) recolectó 264 accesiones de banano en las provincias continentales de Papua Nueva Guinea y en las INFOMUSA — Vol 8, N° 1 islas Nueva Bretaña, Nueva Irlanda y Manus. Las accesiones fueron caracterizadas taxonómica- y agronómicamente por el Queensland Department of Primary Industries en Australia y luego enviadas al Centro de Tránsito del Germoplasma de Musa de INIBAP. El objetivo de este estudio consistió en investigar la reacción de la planta huésped de una selección de las variedades diploides AA y Fe’i recolectadas en Papua Nueva Guinea a R. similis, Pratylenchus coffeae y Meloidogyne spp. El Radopholus similis y P. coffeae son las dos especies de nematodos más diseminadas e importantes asociadas con el banano y plátano (Gowen y Quénéhervé 1990). Estas dos especies de nematodos deben haber estado desarrollándose por mucho tiempo conjuntamente con las variedades de Musa ya que el género de los nematodos Radopholus y P. coffeae se consideran indígenas de los países del Pacífico (Sher 1968, Luc 1987, Bridge et al. 1997). Estos nematodos que inducen lesiones, son la causa de un sistema radical necrótico y reducido. Los nematodos nodulares de la raíz Meloidogyne incognita, M. arenaria, M. javanica y M. hapla también atacan seriamente a los bananos y plátanos. Estos endoparásitos sedentarios causan llagas en las raíces primarias y secundarias (De Waele y Davide, en imprenta). Las variedades seleccionadas representan todas las áreas de Papua Nueva Guinea donde las accesiones han sido recolectadas. La mayoría de las variedades mostraron en el campo un grado de sensibilidad bajo o intermedio a la Sigatoka amarilla, una enfermedad foliar causada por el hongo Mycosphaerella musicola (Arnaud y Horry 1997). Sin embargo, la tolerancia a este hongo no fue confirmada para todas las variedades bajo condiciones de invernadero (Daniells et al. 1996). A través del estudio, se siguieron la terminología de Bos y Parlevliet (1995) con respecto a la resistencia y susceptibilidad y la metodología descrita por Speijer y De Waele (1997). Materiales y métodos Preparación de las plantas Se seleccionaron veinticinco diploides (grupo AA) y siete variedades de banano Fe’i recolectados en Papua Nueva Guinea (Tablas 1-3). Los cultivares triploides (grupo AAA) de banano Cavendish 901 y Grande Naine fueron incluidos como cultivares de referencia, ya que son altamente susceptibles a R. similis, P. coffeae y Meloidogyne spp. Las plántulas propagadas in vitro fueron transplantadas en potes plásticos de 1 litro (diámetro 12 cm) llenados con arena limosa procesada en un autoclave y mantenidas en un invernadero a temperatura ambiente de 20-27°C y un fotoperíodo de 12 horas. Los potes fueron regados cuando era necesario y fertilizados con una solución hidropónica (Swennen et al. 1986) cada tres semanas después de la inoculación con nematodos. Preparación del inoculo de nematodos Las poblaciones de R. similis, P. coffeae y Meloidogyne spp. utilizadas en este estudio fueINFOMUSA — Vol 8, N° 1 Tabla 1. Reproducción de nematodos en 25 bananos diploides AA y dos cultivares de referencia, medida 8 semanas (R. similis ; Meloidogyne spp.) o 10 semanas (P. coffeae) después de la inoculación. Nombre de la accesión Número ITC Lote 1 Cantidades de R. similis por sistema radical Cantidades de P. coffeae por sistema radical Cantidades de ELF de Meloidogyne por sistema radical Pi=1,000 huevos y vermiformes Pi=1,009 huevos y vermiformes Pi=4,000 huevos y juveniles Tagomor 0989 41 872 c 796 a 110 a Jaruda 1017 35 209 bc 1 695 ab 83 a Kwaro 0943 27 247 bc 663 a 105 a To’o 1004 25 518 bc 1 979 ab 156 a Ulungan 0902 21 892 abc 781 a 87 a Yendisi 0942 21 621 abc 1 472 ab 144 a Ivi Ivi 0919 20 763 ab 1 970 ab 210 a Tamat 0899 19 412 ab 3 303 b 110 a Adina 0893 10 488 a n.e. 40 a Cavendish 901 0738 22 347 abc 853 a 185 a Grande Naine 1256 n.e. 718 a n.e. Lote 2 Pi=020 huevos y vermiformes Pi=1,154 huevos y vermiformes Pi=5,014 huevos y juveniles Mala 0869 1 327 b 8 828 a 288 c Meleng 0821 915 b 10 468 a 189 abc Tomolo 1187 850 b 14 069 a 211 bc Inori 0850 703 ab 10 638 a 301 c Odwa 0889 681 ab 8 852 a 129 ab Sepi i 0849 639 ab 18 479 a 266 c Lalalur 0771 476 ab 12 340 a 188 abc Igua 0891 188 a 9 469 a 180 abc Aivip 0593 n.e. 14 813 a 95 a Cavendish 901 0738 732 b n.e. Grande Naine 1256 483 ab 9 252 Lote 3 Pi=1,100 huevos y vermiformes n.e. a Pi=1,005 huevos y vermiformes 195 abc Pi=5,000 huevos y juveniles Spiral 1206 24 892 b 917 a 302 Bagul 0814 8 934 ab 918 a 254 a Kuspaka 0840 8 671 ab 1 075 a 317 abc Papat Wung i0904 8 075 ab n.e. 241 a Gonub 0995 5 622 a 958 a 286 ab Udiwasi 0930 5 610 a 816 a 424 c Yenai 0774 3 164 a 569 a 308 abc Cavendish 901 0738 27 201 b 859 a Grande Naine 1256 n.e. ITC = Centro de Tránsito de INIBAP ELF = hembras ponedoras de huevos Pi = población inicial n.e. = no examinado n.e. abc n.e. 384 bc Los datos para el análisis fueron transformados mediante log 10 (x + 1). Los promedios en la misma columna seguidos por la misma letra no difieren significativamente, de acuerdo al método de Tukey (P < 0.05) ron recolectadas originariamente en las raíces infectadas de banano en América Latina, Asia y Africa. Radopholus similis y P. coffeae cultivados monoxénicamente sobre los cultivos en los discos de zanahoria a 28°C en la oscuridad (Moody et al. 1973, Pinochet et al. 1995). El inoculo fue ajustado para que rindiera una suspensión de casi 1,000 huevos y nematodos juveniles vivos por planta en tres hoyos excavados en el suelo alrededor de las raíces. Las poblaciones de Meloidogyne spp. fueron cultivadas monoxénicamente sobre las raíces de tomate transformado genéticamente por Agrobacterium rhizogenes (Verdejo et al. 1988). El inoculo fue ajustado para rendir una suspensión de 3,300 – 5,000 huevos y especímenes juveniles por planta en tres hoyos hechos en el suelo alrededor de las raíces. Estimación de la resistencia de la planta huésped Las plantas fueron inoculadas con nematodos cuatro semanas después de la aclimatación. Las plantas inoculadas con R. similis y P. coffeae fueron cosechadas 8 y 10 semanas después de la inoculación, respectivamente. Todo el sistema radical fue pesado y cortado en pedazos de 2 cm. Una muestra adicional de 15 g de raíces frescas fue macerada en una licuadora durante dos períodos de 10 » separados por un intervalo de 5 ». Luego, los nematodos fueron concentrados utilizando tamices con poros de 250, 106 y 40 µm. La cantidad de nematodos fue calculada en 6 ml alícuotas en una suspensión de bajo volumen. Las plantas inoculadas con Meloidogyne spp. fueron cosechadas ocho semanas des13 Tabla 2. Reproducción de nematodos en 7 bananos Fe’i y dos cultivares de referencia medida, 8 semanas (R. similis, Meloidogyne spp.) o 10 semanas (P. coffeae) después de la inoculación. Nombre de la accesión Nùmero ITC Lote 4 Cantidades de R. similis por sistema radical Cantidades de P. coffeae por sistema radical Pi = 1,066 huevos y vermiformes Pi = 1,048 huevos y vermiformes Cantidades de ELF Meloidogyne por sitema radical Pi = 3,300 huevos y juveniles Asupina 1027 2 740 bc 3 411 b 1 323 e Wain 0813 1 904 bc 5 881 cd 802 cd Sar 0898 940 ab 4 648 bc 365 ab Utafan 0913 319 a 1 605 a 439 ab Rimina 1010 271 a 7 846 d 1 047 de Skai 0883 n.e. 6 889 d 290 a Menei 1021 n.e. 4 515 bc 490 bc Cavendish 901 0738 5 026 Grande Naine 1256 n.e. a 735 ITC = Centro de Tránsito de INIBAP ELF = hembras ponedoras de huevos Pi = población inicial n.e. = no examinado Nùmero ITC Lote 5 n.e. 2 265 n.e. cd Los datos para el análisis fueron transformados mediante log 10 (x + 1). Los promedios en la misma columna seguidos por la misma letra no difieren significativamente, de acuerdo al método de Tukey (P < 0.05) Tabla 3. Reproducción de nematodos en 5 bananos Fe’i un banano diploïde (‘Igua’) y un cultivar de referencia, medida 8 semanas después de la inoculación. Nombre de la accesión c Cantidades de R. similis por sistema radical Pi = 1,035 huevos y vermiformes El diseño experimental fue un bloque completo escogido al azar. Dentro de cada experimento cada genotipo fue replicado ocho veces. Previo al análisis estadístico, las densidades de la población de los nematodos y las cantidades de las ELF fueron transformadas mediante la fórmula log10 (x + 1). Todos los datos fueron sujetos al análisis de la varianza (ANOVA) y los promedios fueron separados por la prueba de Tukey a P<0.05. Wain 0813 2 555 c Skai 0883 1 005 bc Resultados Utafan 0913 613 bc Radopholus similis Menei 1021 69 ab Rimina 1010 43 a Igua 0891 1 208 bc Grande Naine 1256 2 032 cd Todas las variedades diploides estudiadas en los lotes 1 y 2, estadísticamente fueron tan susceptibles a R. similis como ‘Cavendish 901’o ‘Grande Naine’, los cultivares susceptibles de referencia (Tabla 1). Debido a que en el lote 2 la cantidad de nematodos por sistema radical de ‘Igua’fue muy bajo en comparación con otras variedades diploides y cultivares susceptibles de referencia, la respuesta de huésped de ‘Igua’ fue examinada nuevamente en el lote 5 (Tabla 3). Nuevamente la susceptibilidad de ‘Igua’con respecto a R. similis no difería significativamente (P < 0.05) de la del ‘Grande Naine’. En los lotes 3 y 4, se observaron diferencias significativas (P < 0.05) en la susceptibilidad con respecto a R. similis entre varios diploides y variedades Fe’i y ‘Cavendish 901’ (Tabla 2). De las variedades diploides, las cantidades de nematodos por sistema radical de ‘Gonub’, ‘Udiwasi’y ‘Yenai’ fueron significativamente más bajas (P < 0.05) en comparación con el cultivar susceptible de referencia. Sin embargo, debido a que el número total de nematodos recuperados fue más alto que la cantidad de nematodos inoculados, estas tres variedades no pueden ser consideradas resistentes a R. similis, sino sólo menos susceptible en comparación con el ‘Cavendish 901’. De las variedades Fe’i, las cantidades de nematodos por sistema radical de ‘Sar’, ‘Utafan’ y ‘Rimina’fueron significativamente más bajas (P < 0.05) en compara- ITC = Centro de Tránsito de INIBAP Pi = población inicial Los datos para el análisis fueron transformados mediante log 10 (x + 1). Los promedios en la misma columna seguidos por la misma letra no difieren significativamente, de acuerdo al método de Tukey (P < 0.05). pués de la inoculación. Todo el sistema radical fue pesado y cortado en pedazos de 4 cm. Para estimar la cantidad de hembras ponedoras de huevos (ELF), se hizo una muestra adicional de 5 g de raíces frescas. Para facilitar el proceso de conteo, las masas de huevos primeros fueron teñidas sumergiendo las raíces en una solución de floxina B (0.15 g/l) por 15’ (Hadisoeganda y Sasser 1982). Diseño experimental, observaciones y análisis de los datos Las variedades fueron divididas en cuatro lotes de unas nueve variedades cada uno. Cada lote incluyó al menos un cultivar susceptible de referencia. La reacción de la planta huésped a R. similis, P. coffeae y Meloidogyne spp. de las variedades en cada lote fue examinada en tres experimentos sucesivos. Las variedades, que mostraron resistencia o parecían ser menos susceptibles, fueron agrupadas en un quinto lote y su reacción de planta huésped fue examinada nuevamente. 14 ción con el cultivar susceptible de referencia (Tabla 2). El nuevo examen de ‘Utafan’ y ‘Rimina’en el lote 5 confirmó la resistencia de ‘Rimina’ a R. similis (Tabla 2). Debido a que el número total de nematodos recuperados fue también más bajo que el número de nematodos inoculados, el ‘Rimina’ puede ser considerado resistente a R. similis. En el lote 5 (Tabla 3) también se incluyeron el ‘Skai’y el ‘Menei’, otras dos variedades de Fe’i, no examinadas previamente en los lotes 1-4. El ‘Skai’ fue tan susceptible a R. similis como lo fue el ‘Grande Naine’, pero el ‘Menei’ parece ser resistente a R. similis : la cantidad de nematodos por sistema radical (en promedio de 69) fue significativamente más baja (P < 0.05) en comparación con el ‘Grande Naine’. Pratylenchus coffeae Todos los diploides y las variedades Fe’i cribadas en los lotes de 1 a 4 estadísticamente fueron al menos tan susceptibles a P. coffeae como el ‘Cavendish 901’ o ‘Grande Naine’ (Tablas 1-2) : las cantidades de nematodos por sistema radical no diferían significativamente de las cantidades de nematodos recuperados de los cultivares susceptibles o fueron aún significativamente más altas. Meloidogyne spp. De las variedades diploides, la cantidad de ELF por sistema radical del ‘Bagul’ y ‘Papat Wung’ fue significativamente más baja (P < 0.05) en comparación con el cultivar susceptible de referencia (Tabla 1). De las variedades Fe’i, la cantidad de ELF por sistema radical del ‘Sar’, ‘Utafan’ y ‘Skai’fue significativamente más baja (P < 0.05) en comparación con el cultivar susceptible de referencia (Tabla 2). Debido a que la cantidad total de ELF recuperada del ‘Bagul’, ‘Papat Wung’, ‘Sar’, ‘Utafan’ y ‘Skai’es todavía alta, estas variedades no pueden ser consideradas resistentes a Meloidogyne spp. Discusión Aunque sólo 25 de unas 120 variedades diploides recolectadas en Papua Nueva Guinea fueron incluidas en el presente estudio, el hecho de que las variedades seleccionadas provenían de lugares diferentes y de que no se descubrió resistencia a R. similis y P. coffeae, confirma las observaciones anteriores de que la resistencia a estas especies de nematodos es extremadamente rara en el germoplasma de Musa, aún en su centro de origen. Hasta la fecha, sólo se conocen dos fuentes de resistencia a R. similis confirmadas ampliamente : ‘Pisang Jari Buaya’ y ‘Yangambi Km5’ (Wehunt et al. 1978, Sarah et al. 1992, Price 1994). El grupo Pisang Jari Buaya (PJB) consiste de las variedades diploides AA, de las cuales algunas variedades muestran resistencia o son menos susceptibles a R. similis (Wehunt et al. 1978). Ninguna de las variedades diploides AA evaluadas en este estudio pertenecen al grupo Pisang Jari Buaya. La otra INFOMUSA — Vol 8, N° 1 fuente de resistencia a R. Similis es el ‘Yangambi Km5’, una variedad triploide AAA recolectada en Congo y posiblemente relacionada con algunas variedades en Malasia. Aunque siendo fértil por parte masculina y femenina, esta variedad no se está utilizando en los programas de mejoramiento de Musa debido a que todas las progenies producen hojas anormales o racimos erectos o semi-erectos. La observación de que una o posiblemente dos variedades Fe’i (‘Rimina’ y ‘Menei’) son resistentes a R. similis las hace particularmente interesantes. Aunque los bananos Fe’i tienen muchas semillas, polen estéril, un índice de cosecha bastante bajo y la erecta orientación de los racimos, su resistencia justifica la investigación combinando sus cualidades con las de otros bananos. La mayoría de las variedades de bananos cultivadas ampliamente son susceptibles a Meloidogyne spp. A menudo, se obtenían resultados inconclusos cuando las variedades de banano fueron cribadas con respecto a su resistencia a Meloidogyne spp. en una escala grande. En Brasil e India, muchas variedades de Musa fueron cribadas contra M. incognita y M. javanica pero ninguna de las dos fue encontrada resistente o aún moderadamente resistente (Zem y Lordello 1981, Patel et al. 1996). Sin embargo, en Filipinas, Davide y Marasigan (1985) cribaron 90 diferentes variedades de Musa con respecto a su reacción a M. incognita y encontraron nueve variedades que mostraban resistencia a M. incognita. Agradecimiento Los autores agradecen a J. Pinochet, M. Riveira, P. Speijer y J.L. Sarah por suministrar poblaciones de nematodos, a I. Van den Houwe (Centro de Tránsito de INIBAP) por suministrar el germoplasma de Musa y a J. Reynders por asistencia técnica. Se agradece la beca de la Universidad de Ghent que permitió a la Sra. N.T. Xuyen estudiar durante un año en Bélgica. Este trabajo fue realizado con el apoyo financiero del Fundo Común para los productos basícos, FAO y del Proyecto de Mejoramiento del Banano del Banco Mundial y la K.U.Leuven en el marco del grupo de trabajo en nematología del Programa Global para el Mejoramiento de Musa — PROMUSA. ■ Bibliografía Arnaud E. & J.P. Horry. 1997. Musalogue : a catalogue of Musa germplasm. Papua New Guinea collecting missions, 1988-1989. INIBAP, Montpellier, Francia. Bos L. & J. Parlevliet. 1995. Concepts and terminology on plant/pest relationships : toward consensus in plant pathology and crop protection. Annual review of Phytopathology 33 : 69-102. Bridge J., R. Fogain & P. Speijer. 1997. Nematodos lesionadores de los bananos. Pratylenchus coffeae (Zimmermann, 1898 Filip. & Shu Stek., 1941, Pratylenchus goodeyi Sher 1 Allen, 1953. Plagas de Musa. Hoja divulgativa No.2. INIBAP, Montpellier, Francia. INFOMUSA — Vol 8, N° 1 Daniells J. W, R.A. Peterson, D.J. Reid & N.J. Bryde. 1996. Selección de 165 accesiones de Papúa New Guinea para la detección de la resistencia a Sigatoka amarilla (Mycosphaerella musicola Leach) en el Norte de Queensland. INFOMUSA 5 (1) : 31-34. Davide R.G. & L.Q. Marasigan. 1985. Yield loss assessment and evaluation of resistance of banana cultivars to the nematodes Radopholus similis Thorne and Meloidogyne incognita Chitwood. Philippine Agriculturist 68 : 335-349. De Waele, D. & D.G. Davide. in press. The root-knot nematodes of banana. Musa Pest Fact Sheet. INIBAP, Montpellier, Francia. Gowen, S. & P. Quénéhervé. 1990. Nematode parasites of bananas, plantains and abaca. Pp. 431460 in Plant Parasitic Nematodes in Subtropical and Tropical Agriculture (M. Luc, R.A. Sikora & J. Bridge, eds.). CAB International, Wallingford, Reino Unido. Hadisoeganda, W.W. & J.N. Sasser. 1982. Resistance of tomato, bean, southern pea, and garden pea cultivars to root-knot nematodes based on host suitability. Plant Disease 66 : 145-150. Jarret, R.L., N. Gawel, A. Whittemore & S. Sharrock. 1992. RFLP-based phylogeny of Musa species in Papua New Guinea. Theoretical and Applied Genetics 84 : 579-584. Luc, M. 1987. A reappraisal of Tylenchina (Nemata). 7. The family Pratylenchidae Thorne, 1949. Revue Nématologie 10 : 203-218. Moody, E.H., B.F. Lownsberry & J.M. Ahmed. 1973. Culture of the root-lesion nematode Pratylenchus vulnus on carrot disks. Journal of Nematology 5 : 225-226. Patel, B.A., R.V. Vyas, D.J. Patel & R.S. Patel. 1996. Susceptibility of banana cultivars to root-knot nematodes (Meloidogyne spp.). INFOMUSA 5: 26-27. Pinochet, J., C. Fernandez & J.L. Sarah, 1995. Influence of temperature on in vitro reproduction of Pratylenchus coffeae, P. goodeyi, and Radopholus similis. Fundamental and applied Nematology 18 : 391-392. Price, N.S. 1994. Field trial evaluation of nematode susceptibility within Musa. Fundamental and Applied Nematology 17 : 391-396. Recursos genéticos Sarah, J.L., F. Blavignac, F. Sabatini & M. Boisseau. 1992. Une méthode de laboratoire pour le criblage variétal des bananiers vis-à-vis de la résistance aux nématodes. Fruits 47 : 559-564. Sher, S.A. 1968. Revision of the genus Radopholus Thorne, 1949 (Nematoda : Tylenchoidea). Proceedings of the Helminthological Society of Washington 35 : 219-237. Simmonds, N.W. 1962. The evolution of the bananas. Longmans, London, UK. Speijer, P. & D. De Waele. 1997. Screening of Musa germplasm for resistance and tolerance to nematodes. INIBAP Technical Guidelines 1. IPGRI, Rome, Italy ; INIBAP, Montpellier, Francia ; CTA, Wageningen, Países Bajos. Stover, R.H. & I.W. Buddenhagen. 1986. Banana breeding : polyploidy, disease resistance and productivity. Fruits 41 : 175-191. Swennen, R., E. De Langhe, J. Janssen & D. Decoene. 1986. Study of the root development of some Musa cultivars in hydroponics. Fruits 41 : 515-524. Tenkouano, A., J.H. Crouch, H.K. Crouch & R. Ortiz. 1998. Genetic diversity, hybrid performance, and combining ability for yield in Musa germplasm. Euphytica 102 : 281-288. Verdejo, S., B.A. Jaffee, & R. Mankau. 1988. Reproduction of Meloidogyne javanica on plant roots genetically transformed by Agrobacterium rhizogenes. Journal of Nematology 20 : 599-604. Wehunt, E. J., D.J. Hutchinson & D.I. Edwards. 1978. Reaction of banana cultivars to the burrowing nematode (Radopholus similis). Journal of Nematology 10 : 368-370. Zem, A.C. & L.G.E. Lordello. 1981. Behaviour of banana cultivars exposed to infestation by M. incognita and M. javanica. Anais de Escola Superior de Agricultura « Luiz de Queiroz » 38 : 875-883. Ruth Stoffelen, Raf Verlinden, Rony Swennen y Dirk De Waele trabajan en el Laboratory of Tropical Crop Improvement, Catholic University of Leuven (K.U.Leuven), K. Mercierlaan 92, 3001 Heverlee, Bélgica ; Ngo Thi Xuyen en el Department of Plant Pathology, Hanoi Agricultural University (HAU), Gia Lam, Hanoi, Vietnam Evaluación en Australia Estudio de 143 variedades de banano con respecto a su resistencia a la Sigatoka amarilla en el norte de Queensland Jeff Daniells and Neil Bryde aniells et al. (1996) evaluaron la reacción a la Sigatoka amarilla de 165 accesiones de banano procedentes de Papua Nueva Guinea (PNG). Estas acce- D siones representaron dos tercios de las accesiones recolectadas por el International Board for Plant Genetic Resources en PNG durante 1988-89. Estamos informando aquí sobre las reacciones a la Sigatoka amarilla de las siguientes 65 accesiones procedentes de PNG, 15 Sin control, la Sigatoka amarilla puede dejar plantas sin hojas antes de la cosecha. así como la reacción de otras 78 accesiones procedentes de otras partes, que fueron escogidas para la caracterización agronómica como parte del proyecto de ACIAR “Fitomejoramiento de banano en el Pacífico Sur”. Jones (1995) examinó una cantidad de accesiones de diversas fuentes con respecto a la resistencia a la Sigatoka amarilla utilizando una técnica de selección en invernadero con plantas jóvenes micropropagadas. Daniells et al. (1996) compararon esta técnica rápida con su técnica de selección en el campo y encontraron que alrededor de 17 % de las variedades dieron un resultado erróneo al utilizar la técnica de selección precoz. Nuevamente comparamos las dos técnicas, esta vez con las variedades restantes comunes a los dos estudios. cada una. Sin embargo, ocasionalmente su cantidad fue reducida a 2-3 plantas debido a varias razones. Plantas de las variedades susceptibles a la Sigatoka amarilla fueron sembradas alrededor del perímetro de las parcelas de ensayo. Los bloques de ensayo recibieron prácticas de manejo de cultivo normales según Daniells (1984), excepto que no se aplicaron químicos para el control de la Sigatoka. A partir de marzo de 1994 y 1995, las hojas fueron prorrateadas con respecto a la presencia de la enfermedad en tres ocasiones con intervalo mensual. Contando desde la primera hoja no desenrollada completamente hacia abajo, se registró la hoja más joven con (i) 10 o más lesiones maduras [la hoja más joven manchada (HMJM), Stover y Dickson 1970] y (ii) con al menos 33 % de la lámina destruida por la enfermedad (HJ33, Jones 1993). Si las hojas no mostraban alguno o todos los síntomas arriba mencionados, se registraba el número total de hojas funcionales en la planta. Para promediar los resultados, si una planta no mostraba los síntomas de la enfermedad, se asumía que los mismos ocurrirían en la hoja que sigue a la hoja más vieja en la planta (número total de hojas más una, Jones 1993). También se determinó la tasa de emergencia foliar entre las ocasiones de prorrateo. Los promedios de la parcela para cada accesión fueron promediados en las tres ocasiones de muestreo. Los errores estándar de los promedios fueron calculados uniendo las varianzas de las tres ocasiones de muestreo. Resultados y discusión La severidad de la Sigatoka amarilla fue alta durante el período de preparación incluyendo los dos meses cuando ambos ensayos fueron evaluados con respecto a la presencia de la enfermedad. La HMJM para el cultivo susceptible Williams fue de 4.7 en el experimento 1 y de 5.4 en el experimento 2 y la HJ33 fue 7.3 en el experimento 1 y 8.3 en el experimento 2. Todos los valores fueron el promedio de 3 evaluaciones. La HMJM para las 143 accesiones en los 2 experimentos varió entre 4.6 y 16.0 y la HJ33, de 6.4 a 16.0 (Tablas 1-3). La tasa de emergencia foliar promedio de diferentes accesiones varió entre 0.4 y 0.9 hojas por semana pero no hubo relaciones entre la HMJM y la tasa de emergencia foliar (los datos no son presentados). Al colocar en orden, empezando por el valor más alto de la HMJM o más resistente en cada grupo genómico o especie, los resultados forman una gradación continua de respuestas lo que hace difícil de colocar las variedades en categorías, como resistentes o susceptibles, con algún grado de confiabilidad. El número pequeño de plantas por parcela sin réplicas también dificulta la interpretación de los datos y es criticable. Sin embargo, hubo un considerable nivel de reacciones a la Sigatoka amarilla en las variedades obviamente resistentes en la mayoría de los grupos genómicos. Daniells et al. (1996) detallaron los factores a considerar en la clasificación de la reacción a la enfermedad y arbitrariamente establecieron las siguientes 4 categorías : • Muy susceptible (MS) HMJM < 6.0, • Susceptible (S) HMJM 6.0 – 9.0 o 6.0 – 8.5 si HJ 33 < 11.5, • Resistente (R) HMJM 9.1 – 9.9 o 8.6 – 9.9 si HJ 33 > 11.5, • Altamente resistente (AR) HMJM ≥10.0 o si el total de hojas funcionales es < HMJM (sin manchas maduras) Hemos continuado con este sistema en el presente estudio. Todas las 8 accesiones de Australimusa que incluían cinco variedades Fe’i fueron altamente resistentes así como las dos accesiones de AAT (Tabla 4). Seis de las siete accesiones ABB fueron altamente resistentes y también cuatro de los siete híbridos AAAA. Materiales y métodos Los dos experimentos fueron realizados, uno durante 1994 y el otro en 1995, en South Johnstone, norte de Queensland (lat. 17°38’S). El experimento 1 abarcó plantas del primer ciclo de 39 accesiones de diferentes fuentes, mientras que el experimento 2 abarcó plantas del primer ciclo de las 65 accesiones de PNG y 43 accesiones de otros lugares. El arreglo de las plantas se estableció en hileras simples con una densidad de 1,333 plantas/ha con el fin de realizar la caracterización taxonómica y agronómica. Las plantas fueron cultivadas a partir de retoños en un vivero en diciembre de 1993 y diciembre de 1994 respectivamente y además, algunos retoños fueron seleccionados el mes siguiente para nivelar los ciclos de cultivo de todas las accesiones (Daniells y O’Farrell 1988). En la mayoría de los casos cada accesión fue representada por una parcela de cuatro plantas 16 Niyarma Yik (SF 248) de Papua Nueva Guinea. INFOMUSA — Vol 8, N° 1 Esta caracterización es sólo preliminar y la distribución en categorías puede cambiar. Se requiere realizar estudios adicionales en diversos ambientes. Las reacciones a la enfermedad obtenidas en el presente estudio concuerdan en general con la poca información publicada hasta ahora para una cantidad limitada de varie- dades (Cheesman y Wardlaw 1937, Wardlaw 1939, Simmonds 1966, Vakili 1968, Laville 1983). Las excepciones se aplican a las variedades como el Plátano Cuerno – altamente Tabla 1. Estimaciones de la enfermedad foliar Sigatoka amarilla en experimento 1. (Los valores mostrados representan un promedio de las tres ocasiones de muestreo). Germoplasma de Musaa Especies silvestres M. acuminata subsp. malaccensis (Selangor IR53) M. acuminata (non classée) 20,44 - Genoma AA Pisang Mas 20,18 Senorita Kluai Khai Inarnibal 20,16 20,09 20,46 Pisang Berlin 20,24 Kluai Hom Kluai Pa Siamee (II 249) SH-3142 20,10 20,11 20,47 Genoma AAA Subgrupo Cavendish Dwarf Cavendish Grande Naine Fast Cycle Grande Naine n( 1 Fast Cycle Grande Naine n( 2 Williams J.D.Dwarf HMJMb Código de la accesión Sinónimos HJ33c Reacciónd D2 13,5e (0,5)f 13,5 (0,5) AR MRS 12,3e (0,88) 12,3 (0,88) AR 6,5 (0,31) 9,3 (0,31) S 6,3 6,9 7,9 (0,24) (0,25) (0,55) 9,3 10,4 11,3 (0,42) (0,94) (1,33) S S S 7,9 (0,22) 11,4 (0,40) S 9,5 11,6 13,4e (0,72) (0,43) (0,60) 11,8g 13,9g 13,4 (0,49) (0,46) (0,60) R AR AR 5,4 (0,28) 8,2 (0,37) MS (0,29) (0,20) (0,20) 7,6 8,1 7,9 (0,38) (0,36) (0,34) MS MS MS Sucrier (KUL BS107) Sucrier Sucrier P.Lemak Manis (II 257) P.Lemak Manis (KUL BS611) - 20,05 - 30,42 30,45 30,46 Selección Novak 4,8 5,2 5,0 30,47 Selección Novak 4,9 (0,19) 8,0 (0,27) MS 4,7 4,8 (0,22) (0,35) 7,3 7,1 (0,29) (0,51) MS MS 5,1 5,0 (0,15) (0,26) 7,5 7,2 (0,33) (0,31) MS MS 4,7 5,8 5,3 5,7 10,8 6,3 (0,00) (0,36) (0,20) (0,33) (0,31) (0,43) 7,4 8,7 7,2 7,9 12,8g 9,3 (0,17) (0,57) (0,29) (0,32) (0,31) (0,47) MS MS MS MS AR S 30,31 30,50 Selección Daniells ex cv. Williams Mons Mari Selección Daniells ex cv. W&R1 W&R1 J.D.Special 30,48 30,43 Valery Red Dacca Nchumbahoka Inzirabustera Kluai Khai Bonng M.inediblus 30,36 30,49 30,59 30,58 30,39 - ex Papouasie-Nouvelle-Guinée Genoma AAAB Goldfinger 40,11 SH-3481/FHIA-01 6,1 (0,32) 12,0 (0,49) S King 7,5 8,9 9,1 (0,39) (0,62) (0,58) 10,8 12,5 11,3g (0,31) (0,42) (0,35) S R R Hua Moa 6,2 6,4 7,0 7,1 (0,26) (0,28) (0,41) (0,32) 9,7 8,8 9,5 10,4 (0,39) (0,36) (0,81) (0,66) S S S S 6,4 6,3 5,9 4,6 9,5 (0,49) (0,22) (0,15) (0,29) (0,99) 8,4 9,3 8,8 6,4 12,6 (0,53) (0,49) (0,19) (0,15) (0,62) S S MS MS R 13,2e (0,35) 13,2 (0,35) AR Genoma AAB Subgrupo plantain Horn Plantain Dwarf French Plantain Pisang Kelat Sous-groupe Maia Maoli/Popoulu Pacific Plantain Mangaro Torotea Ainu Puerto Rican Dwarf Plantain Subgrupo Pome Lady Finger (améliorée) Icecream Santa Catarina Prata Shaban Tomnam (NBH 10) Genome ABB Ducasse a. 21,38 21,24 21,42 21,13 21,52 21,25 21,21 Red Banano de altiplano de Africa Oriental Banano de altiplano de Africa Oriental 21,50 21,14 21,41 21,40 Prata Anã Prata Anã Rajapuri 12,03 Pisang Awak Clasificación está basada en la mejor información disponible – estudios adicionales lo harían más claro b. HMJM = hoja más joven manchada = posición numérica de la hoja con ≥10 lesiones maduras c. HJ33 = posición numérica de la hoja con más de un tercio de la hoja muerta debido a M. musicola d. AR = Altamente Resistente - HMJM ≥10.0 o TFL < HMJM (no se encontraron síntomas) R = Resistente - HMJM 9.1-9.9 o 8.6-9.9 si HJ33 > 11.5 S = Susceptible - HMJM 6.0-9.0 o 6.0-8.5 si HJ33 < 11.5 INFOMUSA — Vol 8, N° 1 MS = Muy Susceptible - HMJM < 6.0 e. La etapa HMJM no fue alcanzada en esta accesión f. Error Estándar g. La etapa HJ33 no fue alcanzada en esta accesión 17 Tabla 2. Estimaciones de la enfermedad foliar Sigatoka amarilla para las variedades de Papua Nueva Guinea en experimento 2. (Los valores mostrados representan un promedio de las tres ocasiones de muestreo). Germoplasma de Musaa Especies silvestres Eumusa M.acuminata subsp. banksii M.acuminata subsp. banksii x M.schizocarpa M.schizocarpa Australimusa M.maclayi subsp. maclayi var maclayi M.maclayi subsp. ailuluai M.jackeyi (nord Queensland) Banano Fe’ i Utafan Menei Asupina Skai Sar Genoma AAT Kabulupusa Umbubu Genoma AS Ungota Genoma AA NBB20 Niyarma Yik Kuyu Kwaro Wambo Wan Wiliman Maka Kwonta Gilasalasa Fu Des NBC4 NBA14 Buka Papat Wung Papat Meinje Sena Gonub Kombok Mah Vudu Beo Sira Suruh Ivi-Ivi Taoaya Loibwa Spiral Udiwasi Buladou Yendisi Te’engi Ulungan Maseru Hogolo Genoma AAA Manodop Kokadja Merik Genoma AAB Wandia Panteringan Duningi Bira Yagoa Waema Geagea Midi Kupulik Halaemoemofi Horul Mandape Boung Fu Samoa a. Códido de la accesión Sinónimos PNG059 PNG342 Hawain n°2 Sosi HJ33c Réacciónd 7,5 8,8 (0,26) (0,35) 9,7 11,8 (0,31) (0,58) S R PNG232 8,0e (0,00) 8,0 (0,00) AR PNG252 PNG340 BAN277 11,3e 12,9e 10,3e (0,52) (0,62) (0,64) 11,3 12,9 10,3 (0,52) (0,62) (0,64) AR AR AR PNG311 PNG293 PNG361 PNG274 PNG294 11,4e 11,2 11,2e 11,8e 9,5e (0,32) (0,66) (0,42) (0,32) (0,20) 11,4 11,2 11,2 11,8 9,5 (0,32) (0,66) (0,42) (0,32) (0,20) AR AR AR AR AR PNG332 PNG236 10,3 11,3e (0,31) (0,33) 12,6g 11,3 (0,36) (0,33) AR AR PNG362 9,8 (0,25) 13,0 (0,49) R 8,3 6,6 7,2 6,3 11,6 7,3 7,7 7,7 5,9 6,8 7,1 7,5 9,0 7,5 8,2 7,3 8,6 6,8 7,2 6,8 7,3 8,4 8,0 7,8 6,2 6,9 8,0 7,3 7,0 5,7 6,2 9,5 7,0 6,9 7,7 (0,47) (0,58) (0,41) (0,48) (0,53) (0,47) (0,39) (0,47) (0,43) (0,58) (0,22) (0,29) (0,71) (0,29) (0,19) (0,43) (0,40) (0,29) (0,27) (0,42) (0,27) (0,69) (0,43) (0,27) (0,31) (0,22) (0,27) (0,58) (0,33) (0,41) (0,86) (0,31) (0,79) (0,46) (0,27) 11,3 9,3 10,2 9,0 13,0 9,4 10,1 8,1 8,4 8,7 9,5 10,3 11,0 9,6 10,8 9,6 11,1 9,0 9,7 9,1 9,7 10,7 10,4 9,8 8,3 8,8 11,0 10,0 9,2 8,3 8,7 11,8 9,0 8,6 10,3 (0,98) (0,47) (0,56) (0,35) (0,33) (0,51) (0,48) (0,43) (0,54) (0,33) (0,39) (0,00) (0,58) (0,34) (0,51) (0,38) (0,36) (0,41) (0,27) (0,42) (0,64) (0,86) (0,38) (0,19) (0,39) (0,31) (0,43) (0,00) (0,58) (0,71) (0,64) (0,51) (0,86) (0,69) (0,43) S S S S AR S S S MS S S S S S S S S S S S S S S S S S S S S MS S R S S S 6,7 7,8 6,9 (0,27) (0,65) (0,34) 8,4 9,7 9,9 (0,33) (0,58) (0,57) S S S 8,6 9,0 6,8 8,4 9,2 8,8 8,9 7,1 11,0 8,8 7,7 8,5 8,2 8,3 (0,48) (0,47) (0,44) (0,38) (0,54) (0,27) (0,22) (0,41) (0,41) (0,29) (0,47) (0,39) (0,31) (0,48) 11,2 10,3 9,1 10,7 11,7 12,1 11,9 9,5 13,0 10,7 10,2 11,8 10,9 10,6 (0,57) (0,31) (0,32) (0,47) (0,46) (0,75) (0,49) (0,65) (0,41) (1,15) (0,38) (0,42) (0,74) (0,60) S S S S R R R S AR S S S S S BAN100 BAN21 PNG198 PNG350 PNG351 PNG355 PNG356 PNG243 PNG286 PNG338 PNG346 20,26 PNG345 PNG301 PNG297 PNG237 PNG231 PNG160 PNG156 PNG152 PNG140 PNG305 PNG306 PNG318 PNG319 PNG333 PNG010 PNG334 PNG337 PNG349 PNG358 PNG299 PNG309 PNG264 BAN101 20,45 PNG353 PNG348 PNG304 PNG354 PNG352 PNG328 PNG323 PNG322 PNG167 PNG307 PNG324 PNG258 PNG360 PNG347 PNG326 NBB19/SF248 (KUL BS269) SF215 (KUL BS267) OBb4 Amau (III 154) (tipo Iholena) Laknau (tipo Maia Maoli/Popoulu) La clasificación está basada en la mejor información disponible – estudios adicionales lo harían más claro b. HMJM = hoja más joven manchada = posición numérica de la hoja con ≥10 lesiones maduras c. HJ33 = posición numérica de la hoja con más de un tercio de la hoja muerta debido a M. musicola d. AR = Altamente Resistente - HMJM (10.0 o TFL < HMJM (no se encontraron síntomas) 18 HMJMb R = Resistente - HMJM 9.1-9.9 o 8.6-9.9 si HJ33 > 11.5 S = Susceptible - HMJM 6.0-9.0 o 6.0-8.5 si HJ33 < 11.5 MS = Muy Susceptible - HMJM < 6.0 e. La etapa HMJM no fue alcanzada en esta accesión f. Error Estándar g. La etapa HJ33 no fue alcanzada en esta accesión INFOMUSA — Vol 8, N° 1 resistente, de acuerdo con Simmonds (1966) y Laville (1983) y otras variedades de reacción ‘intermedia’ como Pome, Silk, Mysore y Red. Estas excepciones se deben a las diferencias en los criterios de clasificación de las reacciones a la enfermedad y ambiente, particularmente la presión del inóculo de la enfermedad. Ninguna de las accesiones de Papua Nueva Guinea tienen mucho potencial para la producción comercial en Australia debido a sus rendimientos mucho más bajos y porque ellos se adaptan mejor para los propósitos de cocción que de postre. Sin embargo, ellos pueden ser útiles como fuente de resistencia a la Sigatoka amarilla para los propósitos de mejoramiento. Entre otras accesiones evaluadas se encuentran algunas resistentes y altamente resistentes, que pueden tener un potencial comercial. Estas son las siguientes : Genoma Variedad Comentarios AA Kluai Hom Sabor dulce AAA Kluai Khai Bonng Pulpa de color rosado o naranja, fruta muy atractiva. AAAA AAB Yangambi Km5 Sabor algo ácido T6 De tipo Cavendish T12 De tipo Cavendish Dwarf French Excelente Plantain variedad de cocción de baja estatura ABB Mysore Sabor algo ácido Ducasse Producción australiana (30 ha, mercado principal mente étnico Kluai Namwa Khom Manejo de cultivo más fácil que de Ducasse que es más alto Cuatro de estas variedades (Yangambi Km5, Mysore, Ducasse y Kluai Namwa Khom) también tienen una buena resistencia a la Sigatoka negra y están siendo utilizadas por el QDPI en el Programa de Reemplazo de Bananos (Banana Replacement Programme) en el norte de Australia. Las variedades resistentes como las que se utilizan para reemplazar las variedades susceptibles creando de esta manera una zona de amortización con el fin de reducir el riesgo de la propagación de la Sigatoka negra desde Papua Nueva Guinea y Estrecho de Torres hacia las principales áreas productoras en el sur de Cairns. Las variedades arriba mencionadas pueden ser cultivadas a escala comercial sin la necesidad de realizar el control de la Sigatoka amarilla. Sin embargo, en algunas circunstancias otras enfermedades como las manchas de las hojas [Mycosphaerella musae, Veroneae (Chloridium) musae y Periconiella (Ramichloridium) musae] pueden requerir cierta atención mediante rociados, deshoje, etc. De acuerdo a Stover (1972) las manchas de las hojas representan un patógeno importante sólo en Australia. La resistencia a una enfermedad foliar no necesariamente confiere resistencia a otras enfermedades foliares. Esto requiere una atención constante por parte de los programas de mejoramiento. La reacción susceptible del Goldfinger (Tabla 1) es típica de los híbridos de la FHIA, los cuales a menudo tienen una mayor resistencia a la Sigatoka negra que a la Sigatoka amarilla. Esto se debe en parte a un legado de los progenitores masculinos SH-3142 y SH-3362 (Tablas 1 y 3) que son muy susceptibles a la Sigatoka amarilla. Probablemente, si los híbridos de la FHIA se cultivaran en una gran escala sin efectuar los programas de control de enfermedades foliares, ¡podríamos observar el resurgimiento de la importancia de la Sigatoka amarilla ! Todos los miembros del subgrupo Cavendish evaluados fueron uniformemente muy susceptibles a la Sigatoka amarilla. Se evaluaron siete híbridos tetraploides de Jamaica (Tabla 3). Los primeros híbridos del programa (IC2, Bodles Altafort y 2390-2) primeramente fueron desarrollados para la resistencia al marchitamiento por Fusarium. Ellos también fueron más resistentes a la Sigatoka amarilla que el Gros Michel susceptible al marchitamiento por Fusarium al cual se decidió reemplazar. Los híbridos desarrollados posteriormente (Calypso, T6, TU8, y T12) tenían una destacada resistencia a la Sigatoka amarilla (Tabla 3). Sesenta variedades de este estudio se encontraban entre las 147 examinadas por Jones (1995) para detectar resistencia utili- Paka fue altamente resistente a la Sigatoka amarilla. Kluai Khai Bonng es altamente resistente a la Sigatoka amarilla. Las variedades Fe’i como el Utafan son altamente resistentes a la Sigatoka amarilla. INFOMUSA — Vol 8, N° 1 19 zando plántulas jóvenes en una técnica rápida de cribado en invernadero. En la Tabla 5, se comparan los datos para cada una de las 60 variedades de los dos sistemas. Hubo diferencias en los resultados entre los dos sistemas : las más notables son dos variedades susceptibles en invernadero, pero resistentes en el campo y 7 variedades extremadamente resistentes en invernadero pero muy susceptibles o susceptibles en el campo. De esta manera la reacción en el invernadero de alrededor de 15 % de las variedades no fue confiable. Esto es casi lo mismo (17 %) que la comparación anterior de los dos sistemas efectuada por Daniells et al. (1996). Los sistemas de cribado precoz continúan teniendo sus limitaciones. ■ Bibliografía Cheesman E.E. & C.W. Wardlaw. 1937. Specific and varietal susceptibility of bananas to Cercospora Tabla 3. Estimaciones de la enfermedad foliar Sigatoka amarilla en experimento 2 (Los valores mostrados representan un promedio de las tres ocasiones de muestreo) Germoplasma de Musaa Especies silvestres Eumusa M.acuminata subsp. burmannicoides Rhodochlamys M.velutina Genoma AA Zarum Tereg Stephens Paka Tuu Gia Malaysian Blood Lakatan SH-3362 Genoma AAA Subgrupo Cavendish Williams Dwarf Parfitt Dwarf Cavendish Pubescent Williams Gros Michel Green Dacca Kru Inyamico HMJMb Código de la accesión Sinónimos BAN 19 Calcutta IR 124 ( KUL BS249) HJ33c 10,7e (0,26) 10,7 (0,26) AR BAN285 8,9e (0,58) 8,9 (0,58) AR BAN 49 BAN 51 BAN 8 20,23 BAN 53 BAN117 - 8,0 9,9 15,1e 13,8e 10,0 4,7 5,3 (0,42) (0,36) (0,43) (1,05) (0,43) (0,33) (0,28) 11,1 12,3 15,1 13,8 12,2 7,0 8,8 (0,53) (0,46) (0,43) (1,05) (0,33) (0,39) (0,31) S R AR AR AR MS MS 5,4 5,5 5,4 5,5 5,4 6,1 6,1 6,6 (0,34) (0,72) (0,36) (0,20) (0,28) (0,43) (0,32) (0,33) 8,3 7,9 8,3 7,8 7,8 8,4 9,0 9,1 (0,40) (0,78) (0,52) (0,40) (0,22) (0,38) (0,63) (0,33) MS MS MS MS MS S S S 13,1e (0,66) 13,1 (0,66) AR 12,9e 11,0e 13,0e 8,6 11,2 7,1 7,6 (0,74) (0,47) (0,39) (0,53) (0,29) (0,33) (0,27) 12,9 11,0 13,0 11,2 11,6g 9,0 10,2 (0,74) (0,47) (0,39) (0,29) (0,28) (0,27) (0,43) AR AR AR S AR S S 9,8 (0,70) 11,5 (0,98) R 7,1 8,3 8,8 9,5 7,2 7,9 7,2 6,6 9,1 8,6 (0,51) (0,43) (0,90) (0,35) (0,26) (0,53) (0,33) (0,42) (0,72) (0,36) 9,2 12,3 12,2 13,5 12,8 12,8 10,4 9,6 12,6 11,0 (0,61) (0,39) (0,43) (0,63) (0,49) (0,58) (0,51) (0,75) (0,58) (0,54) S S R R S S S S R S 15,1e 16,0e 12,3 15,2e 11,6e (0,64) (0,43) (0,71) (1,44) (0,58) 15,1 16,0 14,8 15,2 11,6 (0,64) (0,43) (0,65) (1,44) (0,58) AR AR AR AR AR 9,3 (0,00) 11,8 (0,29) R BAN118 BAN57 BAN91 BAN 81 BAN 166 BAN 165 BAN 297 BAN109 D1 (KUL BS610) Blood Banana AAA? Yangambi km5 Genoma AAAA T6 TU8 T12 2390 - 2 Calypso Bodles Altafort IC2 Genoma AB Ney Poovan Genoma AAB Pisang Raja French Reversion Bobby Tannap Mysore Pisang Raja Cere Sugar Lady Finger (non améliorée) Pacific Plantain Tereg Kud Genoma ABB Ducasse Kluai Namwa Khom Goly Goly Pot Pot Warwar Kluai Niu Mue Nang BAN 106 Green Red Dwarf Red/Figue Rose Naine Banano de altiplano de Africa Oriental Ibota (ITC 1123) BAN181 BAN180 BAN178 BAN 179 BAN 170 BAN 325 BAN176 61-86 T8;61-882 Buccaneer; 15-168-12 ex-Jamaïque T13? 65-3405-1? ex-Jamaïque ex-Jamaïque ex-Jamaïque Kluai Teparot BAN269 BAN186 BAN 229 BAN 290 BAN229 BAN211 BAN218 BAN241 BAN198 BAN50 BAN42 BAN256 BAN250 BAN264 BAN263 12,08 R5/5, tipo plantain Tipo plantain Silk Silk Tipo Pome Tipo Maia Maoli/Popoulu Laknau Pisang Awak Dwarf Pisang Awak Tipo Kalapua Tipo Kalapau Clasificación está basada en la mejor información disponible – estudios adicionales lo harían más claro S = Susceptible - HMJM 6.0-9.0 o 6.0-8.5 si HJ33 < 11.5 HMJM = hoja más joven manchada = posición numérica de la hoja con ≥10 lesiones maduras e.La a. b. c. HJ33 = posición numérica de la hoja con más de un tercio de la hoja muerta debido a M. musicola d. Réacciónd MS = Muy Susceptible - HMJM < 6.0 etapa HMJM no fue alcanzada en esta accesión f. Error Estándar g. La etapa HJ33 no fue alcanzada en esta accesión AR = Altamente Resistente - HMJM (10.0 o TFL < HMJM (no se encontraron síntomas) R = Resistente - HMJM 9.1-9.9 o 8.6-9.9 si HJ33 > 11.5 20 INFOMUSA — Vol 8, N° 1 leaf spot. Tropical Agriculture (Trinidad) 14 : 335-336. Daniells J.W. 1984. The banana industry in north Queensland. Queensland Agriculture Journal 110 : 282-290. Daniells J.W. & P.J. O’Farrell. 1988. The nurse sucker management system. Pp 67-71 in Memorias 1986 de la IV Reunión sobre Agrofisiología del Banano. (J.A.G. Cháves and R.R. Calderón, eds.). Asociación Bananera Nacional, San José, Costa Rica. Daniells J. W, R.A. Peterson, D.J. Reid & N.J. Bryde. 1996. Selección de 165 accesiones de Papúa New Guinea para la detección de la resistencia a Sigatoka amarilla (Mycosphaerella musicola Leach) en el Norte de Queensland. INFOMUSA 5(1): 3134. Jones D.R. 1993. Evaluating banana and plantain for reaction to black leaf streak disease in the South Pacific. Tropical Agriculture (Trinidad) 70 : 39-44. Jones D.R. 1995. Rapid assessment of Musa for reaction to Sigatoka disease. Fruits 50(1):11-22. Laville E. 1983. Les cercosporioses du bananier et leurs traitments. Comportement des variétés. Généralités. Fruits 38 : 147-151. Simmonds N.W. 1966. Bananas. Longman, London, UK. Stover R.H. 1972. Banana, Plantain and Abaca Diseases. CAB International, UK. Stover R.H. & J.-D. Dickson. 1970. Leaf spot of bananas caused by Mycosphaerella musicola : Me- thods of measuring spotting prevalence and severity. Tropical Agriculture (Trinidad) 47 : 289-302. Wardlaw C.W. 1939. Cercospora leaf spot disease of bananas. Nature 144 : 11-14. Vakili N.G. 1968. Responses of Musa acuminata species and edible cultivars to infection by My- cosphaerella musicola. Tropical Agriculture (Trinidad) 45 : 13-22. Los autores trabajan en Queensland Horticulture Institute QDPI, PO Box 20, South Johnstone, 4859 Australia. Tabla 4. La reacción de los grupos genómicos a la Sigatoka amarilla. Reacción a la enfermedad* Constitución genómica No de accesiones AR R S MS Especies silvestres Eumusa M.a. subsp.banksii M.a. subsp. malaccensis M.a.subsp. burmannicoides M.a. (non classée) M.a. subsp. banksii x M.schizocarpa M.schizocarpa 1 1 1 1 1 1 0 1(1)** 1(1) 1(1) 0 1(1) 0 0 0 0 1 0 1 0 0 0 0 0 0 0 0 0 0 0 Rhodochlamys M.velutina 1 1(1) 0 0 0 Australimusa M.maclayi et M.jackeyi 3 3(3) 0 0 0 Fe’I AAT AS AA AAA AAAA AAAB AB AAB ABB 5 2 1 51 24 7 1 1 35 6 5(5) 2(2) 0 6(4) 2(2) 4(4) 0 0 1 5(4) 0 0 1 3 0 0 0 1 9 1 0 0 0 37 7 3 1 0 23 0 0 0 0 5 15 0 0 0 2 0 Total 143 33(29) 16 72 22 Variedades comestibles * Ver tabla 1 ** ( ) indica el número de accesiones en las cuales la enfermedad no alcanzó la etapa de HMJM o HJ33 Tabla 5. Comparación de dos técnicas de evaluación del germoplasma de Musa con respecto a M. musicola. Prueba de plàntulas jóvenes en el invernadero Reacción en el campo Susceptible Parcialmente resistente Extremadamente resistente Muy susceptible SH-3142 SH-3362 Gros Michel Williams Dwarf Parfitt Grande Naine Valery Inzirabustera Santa Catarina Prata PNG286 Red Dacca Susceptible Pisang Mas Kokadja PNG 297 Lady Finger (mejorado) PNG243 Pisang Raja Niyarma Yik Pisang Berlin PNG140 PNG237 Sugar IC2 Inarnibal NBA 14 Green Dacca Plàtano Cuerto PNG 167 PNG258 PNG264 PNG347 Plàtano Pacific PNG358 PNG328 Plàtano French Enano Mysore Resistente Kluai Hom Tomnam Goldfinger Kluai Teparot Altamente resistente Total INFOMUSA — Vol 8, N° 1 Kluai Pa T12 Calypso 17 17 Calcutta IR 124 Musa jackeyi PNG340 PNG274 PNG293 PNG294 PNG311 PNG361 Paka Tuu Gia Kluai Khai Bonng Yangambi Km5 T6 TU8 Ducasse Kluai Namwa Khom Kluai Niu Mue Nang 26 21 Enfermedades Diversidad de Fusarium en Cuba Grupos de compatibilidad vegetativa de las poblaciones de Fusarium oxysporum Schlecht f. sp. cubense (E. F. Smith) Snyd. and Hans. presentes en Cuba Alicia Batlle y Luis Pérez os informes más antiguos de la marchitez o Mal de Panamá causado por Fusarium oxysporum f. sp. cubense (Foc) provienen de Cuba (Smith 1910), Puerto Rico (Fawcett 1911) y Jamaica (Ashby 1913). Según Stover (1962), la enfermedad era predominante en el clon Manzano (subgrupo Silk) y Gros Michel en Cuba. Desde entonces, los clones Gros Michel y Manzano han sido cada vez más atacados hasta su casi total desaparición de los mercados. El Mal de Panamá junto a la Sigatoka amarilla fueron los causantes del derrumbamiento de la industria bananera en Cuba en los años 40 (Calpouzos 1955). Han sido diferenciadas cuatro razas de Foc, tres de las cuales son patógenos primarios en bananos (Stover 1962, Stover y Buddenhagen 1986, Su et al. 1986, Ploetz 1990). La raza 1 está presente en la mayor parte de las regiones donde crecen los bananos y es patogénica por muchos clones incluyendo Manzano y Prata (subgrupo Silk AAB) y el Gros Michel (AAA). La raza 2 es patógena a los bananos de cocción ABB tales como el Bluggoe. La raza 3 es patógena de Heliconia spp. ; fue informada por Waite (1963), en Colombia, Costa Rica, Honduras y Panamá y no ataca los clones de bananos comestibles. La raza 4 ha sido descrita más recientemente por Su et al. (1977) y posteriormente identificada en diferentes países (Ploetz 1990, Pegg et al. 1996). El término raza cuando se aplica a Foc, no implica una relación genética definida con el hospedante como ocurre con otros patosistemas (Ploetz 1990, Pegg et al. 1996). Las razas no están definidas genéticamente sino son simples grupos de aislados virulentos a los mismos clones diferenciales, por lo tanto poblaciones diferentes de la misma raza pueden no tener los mismos genes de virulencia (Stover y Buddenhagen 1986). Aunque el mejoramiento genético y el control de la enfermedad pueden beneficiarse del conocimiento de la filogenia, hasta muy reciente poco era conocido de esta relación. Para el estudio de la variabilidad de las poblaciones, se han utilizado los caracteres convencionales como origen geográfico, caracteres fenotípicos y fisiológicos, la virulencia o razas específicas, la compatibilidad vegetativa y las comparaciones del ADN a L 22 través de los cariotipos electroforéticos y el polimorfismo de los fragmentos de restricción (RFLP). Los grupos de compatibilidad vegetativa (GCV) sirven como una vía natural para subdividir las poblaciones fungosas. En una población asexual, el intercambio de información genética se limita a individuos que pueden formar un heterocarión viable. Los loci que gobiernan la incompatibilidad del heterocarión se denominan het, tal, vc o vic (Leslie 1990). Los locus het actúan como parte de un sistema de reconocimiento que permiten a los individuos reconocerse entre sí y diferenciarse de otros. Estos locus het pueden delimitar patotipos de hongos fitopatógenos asexuales como ocurre en el género Fusarium (Correll et al.1987, Ploetz 1990). Han sido identificados 21 GCV para Foc (Correll et al. 1987, Ploetz y Correll 1988, Pegg et al. 1995) ; 15 de estos han sido encontrados en Asia donde se considera que Musa y Foc han co-evolucionado para generar líneas genéticamente diversas del patógeno. Grupos como el 0124/125 tienen una amplia distribución geográfica mientras otros como el 01210 pertenecen a zonas específicas. Así mismo, la raza 4 ha sido vinculada a los grupos 0120, 0121, 0122, 0126, 0129, 01211, 01213, 01215 entre otros. Materiales y métodos Con el objetivo de determinar los GCV a que pertenecen las poblaciones de Foc de Cuba, se realizó un muestreo en diferentes regiones del país de plantas de bananos y plátanos con síntomas de marchitez por Fusarium, de las cuales se obtuvieron aislamientos monoconidiales. Siguiendo la técnica de Puhalla modificada por Correll et al. (1987), se generaron mutantes nit auxotróficos en medio mínimo rico en clorato de potasio ; se identificaron posteriormente los fenotipos nit 1, nit 3 y nit M. deficientes de las enzimas nitrato reductasa, nitrito reductasa y el cofactor molibdeno respectivamente. Se probó la autocompatibilidad de los mutantes obtenidos eliminándose los autoincompatibles (fenotipos crin) y se probaron los nit 1 obtenidos de los aislados, contra una colección internacional de nit M. pertenecientes a todos los GCV de Foc, suministrada por los doctores Julio Hernández del ICIA (ex CITA), Tenerife, España y Ken Pegg de QDPI Queensland, Australia. Resultados y discusión Se determinó la presencia de los grupos 01210 a los que pertenecieron los aislados de la raza 1 hallados en el clon Manzano y 0124/125 sobre aislados de plantas del clon Burro criollo (tipo Bluggoe, ABB). La presencia del grupo 01210 ha sido reportado solo en el condado de Dade en Florida (Ploetz 1990), mientras que el grupo 0124/125 presenta una amplia distribución mundial (Ploetz 1990, Pegg et al. 1996). Todos los aislados de Foc obtenidos del clon Manzano (subgrupo Silk) reportados por Ploetz (1990) han correspondido al grupo 01210. Dado que el Mal de Panamá no había sido reportado hasta hace relativamente poco tiempo en Florida (Ploetz y Shepard 1989), podría considerarse que los aislados de Florida pudieran tener su origen en plantas del clon Manzano llevadas desde Cuba. Ploetz (información personal), determinó recientemente la presencia del grupo 0128 en aislados de Cuba enviados desde la Universidad Central de Las Villas. Los GCV 01210 y los 0124/0125 y 0128 de acuerdo a los estudios de la variabilidad genética de poblaciones de Foc en base a análisis RAPD realizados por Bentley y Bassam (1996), fueron ubicados en grupos diferentes, por lo que presentan poca relación genética y se asume que poseen orígenes independientes. Los resultados obtenidos hasta el presente permiten identificar estos tres GCV y sugieren la necesidad de continuar estudiando la variabilidad genética de Foc en Cuba utilizando un número mayor de aislados y otros marcadores moleculares. Al mismo tiempo, pueden ser tomados como referencia de los grupos presentes en Cuba con vista a los procedimientos cuarentenarios con el material de Musa que se introduzcan. Conclusiones • Las poblaciones de Foc presentes en el clon Manzano (subgrupo Silk), pertenecen al GCV 01210. • Las poblaciones encontradas sobre el clon Burro criollo pertenecen al GCV 0124/0125 • Se debe continuar un estudio exhaustivo de las poblaciones del patógeno en Cuba incorporando marcadores moleculares. • Los resultados obtenidos deben ser tomados en cuenta en los análisis de cuarentena del germoplasma que se introduce al INFOMUSA — Vol 8, N° 1 país y en las pruebas de resistencia de variedades al Foc que se realizan en los programas de Musáceas en diferentes instituciones del país. ■ Bibliografía Ashby. 1913. Banana disease in Jamaica. (Abstract). Science 31 : 754 -755. Bentley S. & B.J. Bassam. 1996. A robust DNA amplification fingerprinting system applied to analysis of genetic variation within Fusarium oxysporum f. sp. cubense. J. Phytopathology 144 : 207-213. Calpouzos L. 1955. Studies on the Sigatoka disease of bananas and its fungus pathogen. Atkins Garden and Research Laboratory Cienfuegos. 68pp. Correll J.C., C.J.R. Klittich & J.F. Leslie. 1987. Nitrate non-utilizing mutants of Fusarium oxysporum and their use in vegetative compatibility tests. Phytopathology 77 : 1640-1646. Fawcett. 1911. Report of the plant pathologist. Ann. Report. Puerto Rico Agric. Experim. Stat. 1910 : 35 – 36. Leslie J.F. 1990. Genetic exchange within sexual and asexual populations of the genus Fusarium. Técnicas de laboratorio P. 37-48 in Fusarium Wilt of Banana (R. Ploetz, ed.). APS. Minneapolis. Pegg K.G., R.G. Shivas, N.Y. Moore & S. Bentley. 1995. Characterization of a unique population of Fusarium oxysporum f. sp. cubense causing Fusarium wilt in Cavendish Bananas at Carnarvon, Western Australia. Aust. J. Agric. Research 46 : 167-178. Pegg K.G., N.Y.Moore & S. Bentley. 1996. Fusarium wilt of banana in Australia. Aust. J. Agric. Research 47 : 63-650. Ploetz R. 1990. Population biology of Fusarium oxysporum f. sp. cubense. Pp. 63-76 in Fusarium Wilt of Banana (R. Ploetz, ed.). APS. Minneapolis. Ploetz R. & J.C. Correll. 1988. Vegetative compatibility among races of Fusarium oxysporum f. sp. cubense. Plant Disease 72 : 325-328. Ploetz R.C. & E.S. Shepard. 1989. Fusarium Wilt of bananas in Florida. Mycol. Research 93 : 242-245. Smith. 1910. A Cuban banana disease. (Abstract). Science 31 : 754 – 755. Stover R.H. 1962. Fusarial wilt (Panama disease) of bananas and other Musa species. Phytopathological Paper 4. CMI. 117pp. Conservación de recursos genéticos Crioconservación de meristemas en el banano (Musa spp.) : optimización de la regeneración José G. Surga, Rony Swennen y Bart Panis isponemos actualmente de tres métodos para la crioconservación de tejidos meristemáticos de banano : (i) un método simple basado en el pre-cultivo de meristemas altamente proliferantes en presencia de una elevada concentración de sacarosa (Panis et al. 1996), (ii) un método basado en la vitrificación de meristemas altamente proliferantes (Panis et al. en prensa) y (iii) un método basado en la vitrificación de meristemas extraídos de plántulas in vitro (Thinh et al. en prensa). El objetivo de este trabajo es la optimización de la tasa de supervivencia de los meristemas utilizando el método de crioconservación simple para la optimización de la regeneración. D Materiales y métodos Se seleccionaron tres genotipos por su pertenencia a diferentes grupos genómicos o por la morfología del explante (meristema) que producen (Schoofs 1997). Los distintos cultivares empleados fueron suministrados por el Centro de Tránsito del INIBAP, Katholieke Universiteit Leuven (KUL), Bélgica. INFOMUSA — Vol 8, N° 1 Durante un mes, se procedió al cultivo de meristemas de los cultivares Bluggoe (ABB), Kisubi (AB) y Gran Enano (AAA) en un medio estéril (pH 5,8) de Murashige y Skoog, MS (1962), enriquecido con bencilaminopurina (BAP) con una concentración de 10-4 M, ácido indol acético (AIA) con una concentración de 10-6 M, sacarosa con una concentración de 0,1M y gelrite (2 gr/l) como soporte del medio. Seguidamente, los conjuntos meristemáticos (“scalps”) que medían entre 12 y 25 mm2 (Dhed’a et al.) fueron extraídos y cultivados durante tres semanas en un medio semejante al anterior pero con una concentración de BAP de 10-5 M y una concentración de sacarosa cuatro veces más alta (0,4 M). Al cabo de tres semanas, los pequeños “scalps” sobrevivientes se extrajeron utilizando una lupa binocular. Ocho explantes, con varios meristemas cada uno, fueron transferidos en criotubos (Biovial) de 2 ml que fueron introducidos en nitrógeno líquido (-196ºC) durante al menos 30 minutos y luego descongelados rápidamente en agua caliente (38 a 40ºC durante 90 segundos). Inmediatamente después, se procedió a la inoculación de los ocho explantes en una caja de Petri que contenía un medio de proliferación (Schoofs 1997) semi-sólido y sacarosa a 0,1 M Stover R.H. & Y. Buddenhagen. 1986. Banana breeding : Poliploidy, disease resistance and productivity. Fruits 41 : 175-191. Su H.J., T.Y. Chuang & W.S. Kong. 1977. Physiological race of Fusarial wilt fungus attacking Cavendish bananas of Taiwan. Taiwan Banana Research Institute. Spec. Public. 2. 21 pp. Su H.J., S.C. Hwang & W.H. Ko. 1986. Fusarial wilt of Cavendish bananas in Taiwan. Plant Disease 70 : 814 - 818. Waite B.H. 1963. Wilt of Heliconia spp. caused by Fusarium oxysporum f. sp. cubense race 3. Trop. Agric. 40 : 299 - 305. Los autores trabajan en el Instituto de Investigaciones de Sanidad Vegetal. Ministerio de Agricultura. Gaveta 634, 11300, Playa, Ciudad Habana, Cuba. Email: [email protected] ó 0,2 M. Seguidamente, se inocularon ocho explantes diferentes en un matraz Erlenmeyer con un medio de proliferación líquido con dos concentraciones de sacarosa (0,1 M y 0,2 M). Se colocaron los cultivos líquidos en agitadores rotativos a 70 r.p.m. Se utilizó como testigo un conjunto de cuatro explantes, que no habían sido sumergidos en nitrógeno líquido, por tratamiento. Los meristemas inoculados permanecieron una semana en la cámara de cultivo (25ºC) en la oscuridad. Al cabo de este periodo, los explantes fueron expuestos a la luz en condiciones térmicas idénticas. Las observaciones sobre el desarrollo de los meristemas comenzaron dos semanas después de la inoculación de los explantes en los distintos medios. Los meristemas que se desarrollaron tras la crioconservación fueron transferidos a un medio MS que contenía las sustancias de crecimiento BAP (10-6 M) y AIA (10-6 M) para su regeneración en plantas in vitro. La experiencia comprendía cinco repeticiones por cultivar. Resultados y discusión La elección de los cultivares empleados en este estudio es interesante debido a que pertenecen a diferentes grupos genómicos. Los resultados muestran que el porcentaje de supervivencia de los explantes tras congelación en nitrógeno líquido varía del 45,9% al 9,8% en un medio semi-sólido enriquecido con sacarosa 0,1 M (normal) (cuadro 1). Los mejores resultados se obtuvieron con el cv. Bluggoe (45,9%) y confirman los datos aportados por Panis et al. (1996) con material vegetal similar e idénticas condiciones. Los cvs. Kisubi (12,5%) y Gran Enano (9,8%) muestran índices de supervivencia más bajos que 23 los que encontró la KUL (INIBAP 1996) en los mismos clones (20,8 y 19,2% respectivamente). Se realizó un segundo ensayo con un medio idéntico al anterior pero con una concentración de sacarosa de 0,2 M. La tasa de supervi- (29,6% vs 12,5%). Gran Enano conserva en los dos casos una baja tasa de sobrevivencia. Se realizó un nuevo ensayo en el que se volvió a utilizar un medio líquido y una concentración de sacarosa de 0,2 M. El análisis de los resultados (cuadro 1) muestra que Tabla 1. Efecto del tipo de soporte del medio (líquido y semisólido) y de la concentración de sacarosa (0,1 m y 0,2 M) en el medio de regeneración sobre la tasa de sobrevivencia tras crioconservación. La experiencia incluye ocho explantes por repetición y cinco repeticiones por cultivar (n = 40). Cultivar medio semisólido 0.1 M 0.2 M tasa de sobrevivencia tras testigo crioconservación (%) (%) Bluggoe (ABB) Grande Naine (AAA) Kisubi (AB) tasa de sobrevivencia tras crioconservación (%) testigo (%) 45.9 ± 29.1 100.0 42.5 ± 22.7 9.8 ± 10.3 94.7 5.0 ± 6.8 60.0 100.0 12.2 ± 8.8 90.0 12.5 ± 0 100.0 medio liquido 0.1 M 0.2 M tasa de sobrevivencia tras testigo crioconservación (%) (%) Bluggoe (ABB) Grande Naine (AAA) Kisubi (AB) tasa de sobrevivencia tras crioconservación (%) testigo (%) 46.0 ± 21.9 100.0 67.8 ± 14.5 7.3 ± 11.2 100.0 23.7 ± 15.9 60.0 100.0 25.0 ± 24.8 90.0 29.6 ± 7.4 vencia de los explantes tras congelación en nitrógeno líquido figura en el cuadro 1. En general, en medio semi-sólido, el aumento de la concentración de sacarosa (0,1 a 0,2 M) no repercutió de forma significativa en la tasa de supervivencia de los explantes. Por ello, se utilizó otro tipo de soporte (líquido) conservando la misma concentración del medio, incluso en las concentraciones de sacarosa (0,1 y 0,2 M). El cuadro 1 muestra los resultados de la regeneración de explantes tras crioconservación en medio líquido con una concentración de sacarosa de 0,1 M en los tres cultivares estudiados. Cuando se comparan los resultados en medio líquido y semi-sólido con la misma concentración de sacarosa (0,1), se observa que la tasa de sobrevivencia en el cv. Bluggoe no ha experimentado cambios importantes (46% vs 45,9% respectivamente) mientras que se observa una clara mejoría en el cv. Kisubi Mejoramiento 100.0 el medio líquido con una concentración de sacarosa de 0,2 M es el más eficaz para los cvs Bluggoe y Gran Enano. Conclusión La tasa de sobrevivencia meristemática de los tres cultivares, evaluada en medio semi-sólido y líquido con 0,1 M y 0,2 M de sacarosa, es mayor en medio líquido con 0,2 M de sacarosa y varía entre 23,7 y 67,8%. El efecto positivo del medio líquido puede atribuirse a una dilución de los polifenoles desagregados. En un medio semi-sólido, estos polifenoles forman, tras la oxidación, una capa negra impenetrable alrededor de los meristemas. La ventaja de los medios que contienen 0,2 M de sacarosa reside en que el choque osmótico entre el medio de precultivo (0,4) y el medio de regeneración se ha vuelto más débil. La reacción de los explantes tras la crioconservación siguió en todos los tratamientos el mismo orden, con una mejor tasa de sobrevivencia para Bluggoe, seguido por Kisubi y Gran Enano. Esta reacción parece deberse a la calidad de los explantes que a su vez depende del cultivar como ya expuso Schoofs (1997). ■ Bibliografía Dhed’a D., F. Dumortier, B. Panis, D. Vuylsteke & E. De Langhe. 1991. Plant regeneration in cell suspension cultures of the cooking banana cv. Bluggoe (Musa spp. ABB group). Fruits 46 (2) : 125-135. INIBAP. 1996. Networking Banana and plantain : Annual Report 1996. INIBAP, Montpellier, France. Pp. 12-13. Murashige T. & F. Skoog. 1962. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 15 : 473-497. Panis B., N. Totté, K. Van Nimmen, L. Withers & R. Swennen. 1996. Cryopreservation of proliferating meristems of banana (Musa spp.). Plant Science 121 : 95-106. Panis B., H. Schoofs, N.T. Thinh & R. Swennen. (in press). Cryopreservation of proliferating meristem cultures of banana. Proceedings of a JIRCAS/IPGRI Joint International Workshop : Cryopreservation of Tropical Plant Germplasm, Current research progress and applications. Tsukuba, Japan, 20-23 Oct, 1998. Thinh N.T., H. Takagi and S. Yashima. (in press). Cryopreservation of in vitro-grown shoot tips of banana (Musa spp.) by vitrification method. Cryo-Letters. Schoofs H. 1997. The origin of embryogenic cells in Musa. PhD Tesis No. 300, Katholieke. Universiteit Leuven, Dissertationes de Agricultura. 257pp. J. G. Surga trabaja en el FONAIAP- Centro Nacional de Investigaciones Agropecuarias, Departamento de cultivos regionales, Recinto Universitario el Limón, Apdo. 4653, Maracay, Venezuela ; R. Swennen y B. Panis en la Katholieke Universiteit Leuven, Laboratory of Tropical Crop Improvement, Kardinaal Mercierlaan 92, B-3001 Heverlee, Bélgica. Desarrollo de híbridos Polinizaciones y cultivo de embriones en bananos S. de O. e Silva, K.M. da Silva, M. de F. Borges y R. P de Oliveira as primeras investigaciones en el área del mejoramiento genético de banano fueron realizadas en los años veinte en Honduras, Trinidad y Tobago y Jamaica, buscando el control de la enfermedad del marchitamiento por Fusarium o Mal de Panamá (Fusarium oxysporum f. sp. cubense). En la L 24 década siguiente fue obtenido el primer híbrido tetraploide de banano, cruzando el cultivar ‘Gros Michel’ (AAA) con un diploide silvestre (AA). Este esquema de hibridación (3n x 2n) viene siendo utilizado en los principales programas de mejoramiento genético de banano (FHIA 1992, Shepherd 1992, IITA 1993). El programa de mejoramiento genético de banano actualmente desarrollado en el EMBRAPA-CNPMF esta basado en cruzamientos entre triploides comerciales (Maçã, Yan- gambi No. 2, Figue Pomme Naine) y diploides mejorados (Híbrido AB : Maçã x Musa balbisiana) y tiene como objetivo el desarrollo de bananos mas productivos, resistentes a las principales enfermedades (Sigatoka amarilla, Sigatoka negra, Mal de Panamá) y plagas (nemátodos y picudos). El proceso de polinización puede ser afectado por varios factores. La eficiencia del polen puede estar relacionada con la presencia de translocaciones en los cromosomas, INFOMUSA — Vol 8, N° 1 afectando su fertilidad y/o el crecimiento del tubo polínico en los ovarios (Shepherd et al. 1987a, 1987b). Factores como humedad y temperatura, también pueden tener gran influencia en la producción de semillas tetraploides (Shepherd et al. 1994). Así mismo, se observa una gran diferencia en la producción de semilla (calidad y cantidad) dependiendo de la variedades utilizadas como padres. El cultivo de embriones ha proporcionado un incremento en el numero y en la uniformidad de germinación de las semillas híbridas obtenidas. El objetivo de este trabajo consiste en la evaluación de los resultados obtenidos después de ser realizadas polinizaciones de diploides (AA) y triploides (AAB), así como en el cultivo de embriones diploides (AA) y tetraploides (AAAB) de banano. ploides SH-3263 y 2803-01 como progenitores masculinos. madre que el sentido del cruzamiento en la obtención de semillas. Los trabajos realizados en plantas triploides del sub-grupo ‘Prata’ permitieron la obtención de semillas en cantidades muchos mas pequeñas que usando plantas diploides. Lo mismo se obtuvo al utilizar plantas madre del cultivar Yangambi No. 2, aunque comparándose los triploides entre ellos, se noto la superioridad de este último cultivar. (Tabla 3). En ambos casos, se pudo observar un mejoramiento en el numero de semillas obtenidas usando los di- Cultivo de embriones Los resultados obtenidos del cultivo in vitro de los embriones rescatados, variaron mucho de un cruzamiento al otro (Tabla 4). Sin embargo, se puede notar que los embriones procedentes de cruzamientos entre diploides presentan en general un porcentaje de germinación mas alto que en caso de hibridación entre diploide y triploide. Además, se pudo verificar que ninguno de los embriones Tabla 1. Polinizaciones realizadas en bananos diploides dentro del periodo 1993/95. EMBRAPA-CNPMF, 1996. Año Polinizacions buenas malas 1993 13 741 2 914 2 406 718 Materiales y métodos 1994 71 4 785 17 929 14 431 676 Polinizaciones 1995 70 5 164 49 573 18 491 1 318 Las inflorescencias fueron protegidas con bolsas de plástico. El polen extraído de anteras colectadas en la mañana, fue colocado sobre los estigmas de las flores femeninas en cuanto se abrían. Total 154 10 690 70 416 35 328 2 712 Semillas Semillas/100 frutos Tabla 2. Polinizaciones usando el diploide SH-3263 durante 1995. EMBRAPA-CNPMF, 1996. Padre1 x Madre2 Cultivo de embriones Las semillas fueron extraídas de frutos maduros. Después de ser lavadas en agua corriente, y una vez quitada la pulpa, la semillas fueron embebidas en agua destilada. Dependiendo de la siembra inmediata de los embriones o de su conservación, la embebición fue respectivamente de 24 horas y 4 días. La desinfección de las semillas se realizo en condiciones estériles en cámara de flujo laminar. Después de 10 minutos en nitrato de plata al 0.5% y de 5 minutos en cloruro de sodio al 5%, la semillas se lavaron tres veces en agua destilada esterilizada. Después de ser extraídos en cámara de flujo laminar, bajo estereoscopio, los embriones fueron inoculados en placas de Petri sobre un medio de cultivo MS agregándose de 30g. l-1 de sacarosa y 7g. l-1 Agar. Después de dos semanas de cultivo en oscuridad, las plántulas obtenidas fueron trasladadas a la luz en el mismo medio de cultivo (temperatura 26°C + 2°C ; foto periodo de 16 horas, intensidad luminosa 1 800 lux). Las plantas enraizadas fueron transferidas en un invernadero, donde permanecerán 90 días antes de meterse al campo. No de flores polinizadas Polinizaciones No de flores polinizadas Semillas buenas malas Semillas /100 frutos SH-3263 x 1318-01 4 464 21 648 3 379 5 394 SH-3263 x 0337-02 2 96 542 786 1 383 SH-3263 x TH03-01 1 60 2 4 10 SH-3263 x 0323-03 1 91 524 1 167 1 858 SH-3263 x 2803-01 5 404 2 68 17 SH-3263 x 0116-01 1 72 1 516 629 2 979 SH-3263 x 1 304-01 1 198 11 347 4 830 82 2803-01 x SH-3263 1 205 16 1 590 35 581 10 863 11 723 Total 1 SH-3263 : hybride diploïde introduit du Honduras 2 1318-01 13: ‘Malaccensis’; 18: ‘Sinwobogi)’; 0337-01 (03: ‘Calcutta’*; 37: ‘Galeo’); TH03-01 (TH: ‘Terrinha’; 03: ‘Calcutta’); 0323-03 (03: ‘Calcutta’; 23: cultivar with no name); 2803-01 (28: ‘Tuggia’; 03: ‘Calcutta’); 0116-01 (01:’Borneo’*; 16: Guyot); 1304-01 (13: ‘Malaccensis’; 04: ‘Madang’*). * ‘Calcutta’ = (M. acuminata spp. burmannicoides); ‘Borneo’ = (Musa acuminata spp. microcarpa); ‘Madang’ = (M. acuminata spp. banksii). Resultados y discusión Polinizaciones Durante el periodo de 1993/1995, numerosas polinizaciones fueron realizadas, en bananos diploides y triploides. En el programa de mejoramiento de los diploides, se logro aumentar el numero de semillas obtenidas, utilizando el diploide híbrido SH-3263 procedente de la Fundación Hondureña de Investigación Agrícola (FHIA) Honduras (Tablas 1 y 2). También se pudo verificar la importancia que tiene tanto la especificidad de la planta INFOMUSA — Vol 8, N° 1 Embrión zigótico en germinación in vitro 25 Tabla 3. Polinizaciones en cultivares tipo ‘Prata’ y Yangambi No. 2 dentro del periodo 1992/95. EMBRAPA-CNPMF, 1996. Año Cruzamiento1 Polinizaciones No de flores polinizadas Semillas Buenas 1994 1503-01 x PC 1503-01 x PP 3 123 49 2 283 10 02 0,53 1503-01 x SM 4 188 01 01 1,06 58 2 716 11 03 1,59 2803-01x PC 16 1 202 17 01 1,50 2803-01x PP 18 1 302 11 02 0,99 2803-01x PV 185 10 297 120 30 1,46 2803-01x SM 04 191 223 12 992 SH-3263 x PC 20 1 040 SH-3263 x PP 9 506 SH-3263 x PV 104 SH-3263 x SM 15 Sous-total 1995 Malas 129 1503-01 x PV Sous-total 1995 2 Semillas /100 frutos Total 02 1,03 148 35 4,89 8 1 0,86 5 034 101 10 2,20 507 11 2 2,56 148 7 087 120 13 5,62 1992 M53 x Yang. No. 2 97 5 583 194 25 3,92 1993 M53 x Yang. No. 2 120 5 184 51 16 1,29 1994 M53 x Yang. No. 2 116 5 674 164 37 2,89 1 1 503-01 : híbrido diploide (Madu x Calcutta) ; 2803-01 : híbrido diploide (Tuugia x Calcutta) ; SH3263 : híbrido diploide obtenido en Honduras ; PC : Prata común ; PP : Prata Ponta Aparada ; PV : Pacovan ; SM : Prata Santa María ; M53 : híbrido diploide obtenido en Jamaica ; Yang. No. 2 : Yangambi No. 2. Tabla 4. Cultivo de embriones en diferentes cruzamientos usando diploides y triploides de bananos. EMBRAPA-CNPMF, 1996. Endosperma Normal Embrión Normal Genotipo1 Semillas Germinación 86B94 03 4087A 13 9287A 03 03 86B79 16 03 17 07 4087A 13 13 9287A 01 86B79 03 Porcentaje de germinación (%) 10 9 279 8 979 Anormal 86B94 02 9 279 8 979 Normal y anormal 86B94 03 4087A 26 23 88 0 9287A 04 03 75 86B79 19 05 26 8 979 17 07 41 PC 27 08 30 PP 13 05 38,5 PV 278 122 44 SM 24 07 30 Yang. x M53 144 23 16 23,5 9279 Réducido Normal y Anormal 86B94 34 08 4087A 12 07 9287A 03 03 100 9 279 02 01 50 8 979 16 15 93,75 PC 09 02 22 aislados de semilla sin endosperma lograron germinar, aunque se pudo obtener plantas a partir de semilla con endosperma reducido. También se obtuvo una mejor taza de germinación de las semillas procedentes de cruzamientos con triploide de tipo ‘Prata’. Así, los porcentajes de germinación obtenidos a partir de embriones aislados de semillas de Yangambi No. 2 fueron en general mas bajos y en cuanto a los embriones del banano ‘Maçã’ no permitieron obtener plantas tetraploides. Las polinizaciones en banano no siempre permiten la obtención de semillas, sobre todo cuando se trata de cruzamientos entre triploides y diploides. Estos trabajos permitieron evidenciar la especificidad existente en los cruzamientos. También se verificó la gran utilidad del cultivo in vitro en el rescate de embriones de bananos procedentes de semillas con un endospermo reducido. ■ Referencias FHIA. 1992. Programa internacional de mejoramiento genético de banano y plátano. Informe anual 1990. FHIA, La Lima, Honduras. 25pp. IITA. 1993. Plantain and banana improvement program. Annual report 1991. IITA, Ibadan, Nigeria. 208pp. Shepherd K. 1992. History and methods of banana breeding. Pp. 108-110 in Planning program and management review of the International Network for the Improvement of Banana and Plantain. Report to TAC. CGIAR, Washington DC, USA. Shepherd K., J.L.L Dantas & E.J. Alves. 1987a. Aspects of banana breeding at the Centro Nacional de Pesquisa de Mandioca e Fruticultura, Brasil. Pp. 78-86 in Proceedings of ACORBAT VII meeting (J.J. Galindo & R. Jaramillo, eds). CATIE, Turrialba, Costa Rica. Shepherd K., J.L.L Dantas & E.J. Alves. 1987b. Banana breeding in Brazil. Pp. 78-83 in Banana and plantain breeding strategies (G. Persley & E. De Langhe, eds). ACIAR proceedings No. 21. ACIAR, Canberra, Australia. Shepherd K., J.L.L Dantas & S. de O. Silva. 1994. Breeding Prata and Maçã for Brazil. Pp. 157-168 in The improvement and testing of Musa : a global partnership (D.R. Jones, ed.). INIBAP, Montpellier, Francia. 58,3 86B79 PP 02 01 50 PV 41 09 22 SM 03 01 33,3 Yang. x M53 99 26 26,3 1 Diploides : TH03-01 (TH : Terrinha ; 03 : Calcutta) ; 79 : 2803-01 (28 : Tuugia ; 03 : Calcutta) ; 86 : 0337-01 (03 : Calcutta ; 37 : Galeo) ; 87 : 0338-02 (03 : Calcutta ; 38 : Heva) ; 89 : 1318-01 (13 : Malaccensis ; 18 : Sinwobogi) ; 92 : 0323-03 (03 : Calcutta ; 23 : Cultivar sin nombre) ; 94 : SH-3263 (híbrido diploide introducido de Honduras). Triploides : PC : Prata Común ; PP : Prata Ponta Aparada ; PV : Pacovan ; SM : Prata Santa Maria, cruzados con varios diploides ; Yang. : Yangambi No. 2 x M53 : diploide introducido de Jamaica. 26 Los autores trabajan en EMBRAPA-CNPMF, Cx. P. OO7, 44380-000, Cruz das Almas-BA, Brazil. INFOMUSA — Vol 8, N° 1 Mejoramiento Estudios de fertilidad Estudio de la fertilidad polínica de algunos cultivares de Musa utilizados en el programa de mejoramiento genético del INIVIT René Landa, Aymé Rayas, Teresa Ramírez, José Ventura, Julia Albert y Omayra Roca l mejoramiento genético de los bananos y plátanos resulta difícil debido a diferentes parámetros como la partenocarpia, la esterilidad y la poliploidía. Los bananos y plátanos aptos para el consumo son por la mayoría estériles, volviéndose muy complicado y largo su mejoramiento (Kulasekaran 1986). La diseminación rápida de la enfermedad de la Sigatoka negra, así como su virulencia, orientaron rápidamente los esfuerzos de los programas de mejoramiento hacia la búsqueda de clones elites para la producción de plantas resistentes. Una de las vías mas utilizadas consiste en el uso de métodos convencionales de mejoramiento basados en la hibridación de clones triploides con fertilidad femenina residual con diploides mejorados como progenitor masculino (Roman 1988). La producción de polen en los cultivares de Musa sp. depende en primer lugar de causas genéticas. Sin embargo, se conoce también la influencia de los parámetros ambientales (Horry 1995). Por esto puede ser de mucha utilidad estudiar el comportamiento de los genitores en el sitio mismo donde se desarrolla el programa de mejoramiento. E Materiales y métodos Este trabajo se desarrollo en el laboratorio de citogenética del Instituto de Investigaciones de Viandas Tropicales (INIVIT). Todos los cultivares utilizados dentro del programa de mejoramiento fueron evaluados, utilizando el método de Alexander “Diferencial de granos de polen” (Roman et al. 1988). Se aislaron las anteras de botones florales maduros, tomados al azar 10 flores clon. Las anteras fueron colocadas sobre un portaobjeto usando una o dos gotas del colorante de Alexander. Después de un ligero aplastamiento de las anteras, se observaron 200 granos de polen por cada flor mediante un microscopio. Los granos de polen fértiles aparecen teñidos de rojo intenso, al contrario de los granos verdes que no tienen fertilidad. Resultados y discusión Los resultados obtenidos con los clones diploides revelan en forma general una gran INFOMUSA — Vol 8, N° 1 mejoramiento genético de los bananos y plátanos. Se pudo corroborar la importancia de las condiciones ambientales en el desarrollo de un polen fértil, así como eliminar el clon “AA de Zanzíbar’ por tener una producción de polen limitada. ■ cantidad de polen fértil (Tabla 1). Sin embargo, se pudo notar algunas diferencias entre ellos. Así el clon Zanzíbar apareció poco interesante debido al porcentaje muy bajo de polen fértil que presento en nuestras condiciones experimentales. Cabe re- Tabla 1. Cultivares diploides fértiles. Valores encontrados para el número total de granos de polen (fértiles e infértiles) y los % que representan del total. Cultivar No total de granos Nùmero de granos % de granos Fértiles Infértiles Fértiles Infértiles 0.9 BB de Vietnam 200 198.0 1.8 99.1a Pisang Jari Buaya 200 196.2 3.8 98.1a 1.9 SH-3362 200 188.2 11.2 94.4b 5.6 SH-3142 200 190.0 9.8 95.1b 4.9 Mundo 200 184.5 15.5 92.5c 7.7 200 183.7 17.0 91.8c 8.5 181.6 45.1 135.8 27.6d 74.5 Paka AA de Zanzibar ES + 0.19 – CV = 2% Tabla 2. Cultivares triploides. Valores encontrados para el número total de granos de polen (fértiles e infértiles) y los % que representan del óptimo establecido. Cultivar No total de granos Nùmero de granos Fértiles Infértiles % de granos Fértiles Infértiles Pelipita 141.9 56.4 86.3 39.7a 54.5 Somaclon Saba 138.4 63.6 78.1 43.3a 61.6 Saba 131.1 56.4 74.7 42.9a 57.2 Hembra 3/4 129.3 53.8 73.5 41.6a 56.8 Highgate 118.2 46.7 71.5 39.6a 60.4 Burro CEMSA 133.4 54.8 79.1 41.2a 54.2 Burro CEMSA enano 137.2 56.6 80.3 42.6a 58.4 ES + 0.101 – CV = 2% saltar la fertilidad excepcional del clon “Pisang Jari Buaya” dado en la literatura consultada por tener muy poca cantidad de polen (Rowe 1981). Aquí se puede corroborar la influencia de los parámetros ambientales en la producción de polen. También se obtuvieron altos porcentajes de polen fértiles en los clones ‘Mundo’, ‘Paka’, ‘SH-3362’ y ‘SH-3142’. Los buenos resultados obtenidos con los dos diploides SH coinciden con lo reportado por Pinochet y Rowe (1979). El análisis de los resultados obtenidos con los clones triploides revelan bajas cantidades de polen fértiles, sin diferencia entre ellos (Tabla 2). Los clones tetraploides resultaron mejores productores de polen que los triploides, con porcentajes de polen fértiles intermedios del 57,8 a 61,4%. Esto concuerda con lo reportado por Menéndez y Shepherd (1975). A través de este trabajo, se pudo recoger una muy valiosa información que debería permitir una mejor orientación del programa de Bibliografía Kulasekaran M. 1986. Banana breeding. Faculty of Agriculture, Tamil Nadu University, Coimbatore, India. Menéndez T & K. Shepherd. 1975. Breeding new bananas. World Crops 27:104-112. Pinochet J. & P. Rowe. 1979. Progress in breeding for resistance to Radopholus similis on bananas. Nematropica 9:76-78. Roman & M. Isabel. 1988. Estudio de la fertilidad del polen en cultivares de plátano (Musa spp.). Ciencia y Técnica de la Agricultura. Viandas Tropicales 11(1). Rowe P. 1981. Breeding an ‘intractable’ crop: bananas. Pp. 66-83 in Genetic Engineering for Crop Improvement (K.O. Rochie and J.M. Lyman, eds). The Rockefeller Foundation, New York. Los autores trabajan en el Instituto de Investigaciones en Viandas Tropicales (INIVIT), Apdo. 6, CP 53000, Santo Domingo, Villa Clara, Cuba. 27 Plagas Control biológico del picudo negro Observaciones preliminares sobre los enemigos naturales asociados con el picudo negro del banano Cosmopolites sordidus Germar en Uganda W. Tinzaara, E. Karamura y W. Tushemereirwe tos) como agentes de control biológico para controlar C. sordidus en Uganda. Materiales y métodos l picudo negro del banano, Cosmopolites sordidus Germar, es la principal plaga en las regiones productoras de banano (Stover y Simmonds 1987), y una de las principales limitaciones de la producción bananera, especialmente en los sistemas de producción a pequeña escala (Bujulu et al. 1983, Sikora et al. 1989). El control del picudo negro del banano siempre ha dependido del uso de insecticidas químicos que son altamente tóxicos, muy costosos y pueden causar serios problemas residuales en los bananos cosechados. Actualmente se investigan medidas alternativas de control. El enfoque del manejo integrado de plagas (IPM) con un énfasis sobre el control biológico puede proporcionar una opción plausible para el control de esta plaga. Los estudios de los enemigos naturales de esta plaga, provenientes del Sureste de Asia (Neuenschwander 1988) y Kenia (Koppenhoefer 1993), han generado un inventario de depredadores, hongos entomopatogénicos y nematodos. En Uganda, sin embargo, sólo los patógenos fungosos han sido reportados como agentes promisorios del control biológico de los picudos (Allard et al. 1993, Nankinga 1994). El papel de los enemigos naturales indígenas de C. sordidus no ha sido explorado en Uganda. Este estudio está dirigido a evaluar el potencial de los enemigos naturales endémicos (depredadores y parási- E El estudio fue realizado en mayo de 1997 en los campos de Kawanda, sembrados con el banano de cocción Nakyetengu (AAA-EA), con el fin de determinar la composición de los enemigos naturales y su abundancia utilizando la técnica de búsqueda manual (Koppenhoeffer 1993). Se investigaron residuos de seudotallos y sus fragmentos. La búsqueda se realizaba semanalmente durante cuatro semanas. Los depredadores recuperados se mantuvieron en frascos con formalina al 5% para realizar posteriormente su identificación y registro. Durante la búsqueda también se recolectaron picudos en diferentes etapas de desarrollo. La identificación de los depredadores fue realizada en el Museo de Insectos del Kawanda Agricultural Research Institute. Los picudos en diferentes etapas de desarrollo recolectados en el campo, fueron incubados en platos petri con el fin de determinar la emergencia parasitoide. El tamaño de las muestras dependió de las diferentes etapas de desarrollo de los picudos recolectados durante la búsqueda de depredadores. Exámenes de bioensayo La determinación en laboratorio de los depredadores que tienen potencial para controlar el picudo del banano, se realizó tomando en cuenta diferentes etapas de desarrollo de los picudos recolectados en los cultivos establecidos y depredadores recolectados en las plantaciones bananeras de Kawanda. Cada uno de los depredadores sospechosos fue alimentado con huevos, larvas y pupas de los picudos. Antes de realizar cada ensayo de alimentación, se privó a los depredadores de alimentación durante 24 horas, después de lo cual se les suministraron picudos en diferentes eta- pas de desarrollo. En cada prueba de alimentación, los picudos en diferentes etapas de desarrollo fueron colocados abiertamente en los platos petri sobre los pedazos delgados de los tejidos de cormos, donde también se introdujeron los depredadores. Las tasas de los depredadores, alimentados durante las pruebas, fueron las siguientes : Depredador : huevos = 1:2, Depredador : Larvas = 1:2 y Depredador : Pupas 1 :1. La prueba de alimentación con los huevos escondidos en los tejidos del cormo fue realizada con el fin de obtener información preliminar sobre la capacidad de búsqueda de los depredadores. Las cantidades de huevos, larvas y pupas de picudos, consumidos o exterminados por los depredadores, se registró durante cada período de alimentación de 24 horas. Las pruebas de alimentación se repitieron seis veces. Resultados y discusión Los depredadores, pertenecientes a siete géneros diferentes y de abundancia variable, se encontraban en los ambientes de los picudos como lo son los túneles en cormos y seudotallos en descomposición (Tabla 1). Los depredadores más comunes fueron Odontomachus sp. (Hormigas) y Dactylosternum sp. (Familia Hydrophilidae). Se informó que la mayoría de estos animales son depredadores potenciales de C. sordidus en otros agro-ecosistemas (Koppenhoeffer 1993). Se espera, sin embargo, que los futuros estudios detallados en Uganda proporcionarán especies con un mayor potencial para controlar C. Sordidus, en comparación con las especies detalladas por Koppenhoeffer (1993). Igual que en el caso de Koppenhoefer (1993), también emergieron los no parasitoides en diferentes etapas de desarrollo de los picudos incubados. Esto podría estar relacionado con el tamaño de la muestra en la observación de un sólo sitio o Tabla 1. Composición y abundancia de los depredadores asociados con C. sordidus. Depredador Abundancia por búsqueda Euborellia sp. (Dermaptera) ** Labia sp. (Dermaptera) ** Dactylosternum sp. (Hydrophilidae) *** Thyreocephalus sp. (Staphilinidae) * Paederus sp. (Staphilinidae) * Fourmis (Odontomachus sp.) Eulissus sp. (Staphilinidae) *** * *= <10, **= >10, ***= <50 28 INFOMUSA — Vol 8, N° 1 puede ser atribuido al fallo del parasitoide local de adaptarse a una plaga introducida. Los datos preliminares sobre el potencial de los depredadores de consumir los picudos en diferentes etapas de desarrollo se presentan en la Tabla 2. Los datos muestran que Euborellia sp., Labia sp. y Thyreceophalus sp. son depredadores de un potencial de depredación promisorio. Las tasas de picudos consumidos o exterminados diariamente pa- estar complementadas con los experimentos en cajas en las condiciones de campo. Los resultados presentados en este trabajo son preliminares, pero proporcionan información vital que forma la base para futuras investigaciones. Las futuras encuestas deben incluir mayor cantidad de muestras y sitios en diferentes condiciones agro-ecológicas para poder aclarar el papel de los enemigos naturales indígenas con respecto a la Tabla 2. Potencial de los depredadores de apresar a los picudos en diferentes etapas de desarrollo. Depredador Proporción promedio de los picudos consumidos o eliminados por día (± S.E.) Huevos al descubierto Huevos escondidos Larvas al descubierto Pupas al descubierto Euborellia sp. 0,97 ± 0,02 0,09 ± 0,04 0,83 ± 0,07 Labia sp. 0,50 ± 0,02 0,74 ± 0,05 0,82 ± 0,03 0,12 ± 0,02 Dactolysternum sp. 0,13 ± 0,02 0,30 ± 0,04 0,13 ± 0,02 0,05 ± 0,01 0,52 ± 0,04 Thyreocephalus sp. 0,47 ± 0,05 0,22 ± 0,04 0,48 ± 0,04 0,97 ± 0,02 Eulissus sp. 0,07 ± 0,01 0,40 ± 0,03 - - recen disminuir de huevos a pupas para todos los depredadores, con excepción de Thyreocephalus sp., que consumió una alta tasa de pupas del picudo por día. Aunque Dactylosternum sp. fue abundante en los ambientes de los picudos, su potencial para controlar los picudos fue insignificante en los estudios del laboratorio. La capacidad de búsqueda de los depredadores, estimada utilizando la tasa de consumo de los huevos escondidos, fue alta para Labia sp. Por otro lado, Euborellia sp., que fue muy activa en los ensayos (alta velocidad de movimiento), mostró una capacidad de búsqueda muy baja. La capacidad de búsqueda de otros depredadores fue generalmente baja. Los resultados de las pruebas de alimentación sin opciones en las condiciones de laboratorio tiene la debilidad de sobrestimar el papel de los depredadores, que normalmente no se alimentan de una presa dada y pueden ser forzados a hacerlo debido a falta de alternativas (Waage y Millis 1990). Las futuras pruebas en el laboratorio deben Enfermedades regulación de la población de los picudos del banano. Agradecimiento Los autores agradecen al personal del Museo de Insectos de Kawanda quienes ayudaron en la identificación de los depredadores, y a la Fundación Rockefeller y CIID por financiar el estudio. ■ Bibliografía Allard G.B., C. Nankinga & M. Ogenga-Latigo. 1993. The potential of the indigenous fungal pathogens as a component of integrated management of the banana weevil C. sordidus in Uganda : A new research project. Pp. 118-123 in Biological and integrated control of highland banana and plantain pests and diseases. Proceedings of a research coordination meeting. (C.S. Gold and B. Gemmill, eds). IITA, Ibadan, Nigeria. Bujuru J., B. Uronu & C.N.E Cumming. 1983. The control of the banana weevil and plant parasitic nematodes in Tanzania. East Africa Forest Journal 49 : 1-3 Podredumbre bacterial Enfermedades de la podredumbre de las frutas de los bananos en el Sudeste de Asia Agustín Molina E n la sección de las noticias de Musa de INFOMUSA Vol. 7 (2), se informó que la enfermedad sanguínea está cau- INFOMUSA — Vol 8, N° 1 sando pérdidas de los bananos de cocción Kepok (Saba) en ciertas áreas de Indonesia. También se informó que se está verificando la eficacia de los métodos, que resultaron exitosos en el control de la enfermedad Bugtok en Filipinas, para controlar la enfermedad Figueroa W. 1990. Biocontrol of the banana weevil, Cosmopolites sordidus (Germar) with Steinermatid nematodes. Journal of the Agricultural University of Puerto Rico 74 : 15-19 Koppenhoeffer A.M. 1993. Search and Evaluation of Natural enemies of the banana weevil. Pp. 87-96 in Biological and integrated control of highland banana and plantain pests and diseases. Proceedings of a research coordination meeting. (C.S. Gold and B. Gemmill, eds). IITA, Ibadan, Nigeria. Nankinga C. 1994. Potential of Indigenous fungal pathogens for the biological control of the banana weevil Cosmopolites sordidus Germar in Uganda. M. Sc. Thesis. Makerere University, Uganda Neunschwander P. 1988. Prospects and proposals for biological control of Cosmopolites sordidus Germar (Coleoptera : Curculionidae) in Africa. Pp. 54-57 in Nematodes and the borer weevil in bananas. Present status of research and outlook. INIBAP, Montpellier, France. Sikora R.A., N.D. Bafokuzara, A.S.S. Mbwana, G.W. Oloo, B. Uronu & K.V. Seshu Reddy. 1989. The interrelationship between banana weevil, root lesion nematode and agronomic practices and their importance for banana decline in the United Republic of Tanzania. Plant Protection Bulletin 37 : 151-157 Stover R.H. & N.W. Simmonds. 1987. Bananas. Tropical Agricultural series, Scientific and Technical, U.K. 467 pp. Treverrow N.R.A., E. Bedding, B. Dettmann & C. Maddox. 1991. Evaluation of entomopathogenic nematodes for the control of Cosmopolites sordidus, a pest of bananas in Australia. Annals of Applied Biology 119 :139-145 Waage J.K. & N.J. Millis. 1990. Understanding and measuring the impact of natural enemies on pest populations. in Biological Control Training Manual (D.J. Girling, ed.). CAB, International Institute of Biological Control, UK. W. Tinzaara y W. Tushemereirwe trabajan en el Kawanda Agricultural Research Institute, P.O. Box 7065, Kampala, Uganda. E. Karamura es el Coordinador Regional de INIBAP para Africa Oriental y del Sur, P.O. Box 24384, Kampala, Uganda. sanguínea, asumiendo que esta enfermedad sanguínea es la misma enfermedad de Bugtok. Se debe señalar que este no es cierto. Los términos “Bugtok”, “Tibaglon” y “Tapurok” son términos locales utilizados en Filipinas para describir la fruta infectada, que tiene apariencia decolorada y es dura, aún madura. Una planta enferma puede parecer normal, pero la pulpa de la fruta está podrida (seca), de color negro y no sirve para el consumo humano. La enfermedad es causada por la bacteria Ralstonia solanacearum raza 2 (anteriormente conocida como Pseudomonas solanacearum), sobre la existencia de la cual se informó aún en el año 1965. La enfermedad afecta los bananos de cocción Saba y Cardaba y en los años recientes se observaron daños de proporciones epidémicas. Se co29 noce que la bacteria se transmite por insectos como resultado de su alimentación en flores infectadas. Aunque también se informó sobre la transmisión mecánica y por el suelo, la transmisión por insectos sigue siendo el principal método de la propagación de “Bugtok”/”Tibaglon” en los bananos de cocción. Se conoce también que Ralstonia solanacearum causa la enfermedad del Moko, que afecta comúnmente las plantaciones comerciales de Cavendish en América Latina y Filipinas. Los síntomas externos son el amarilleo y colapso de las 3 hojas más jóvenes. Estos síntomas progresan hacia las hojas más viejas. Internamente, los tejidos vasculares se descoloran y se tornan necróticos, especialmente cerca del centro del seudotallo. Esto causa el marchitamiento y el colapso de las hojas jóvenes. La bacteria es mas sistémica en la infección con Moko, moviéndose en dirección hacia arriba, en contraste con el movimiento lento hacia abajo de la bacteria en la infección con Bugtok. La infección avanzada en las plantas con frutas también puede causar la podredumbre de las frutas. Esto ocurre raramente en las plantaciones comerciales, ya que la detección temprana lleva a la eliminación también temprana de las plantas infectadas. Sin embargo, si esto ocurre de todos modos, la podredumbre de la fruta en la infección con Moko es similar a los síntomas de Bugtok. La propagación del Moko en una plantación comercial de Cavendish ocurre principalmente con cuchillos contaminados utilizados para el deshije regular y otras actividades de poda. De aquí que la infección comienza desde las partes básales de la planta. La inoculación de la fruta es prácticamente inexistente en las plantaciones comerciales de Cavendish, ya que las frutas se embolsan lo que las protege de los insectos. Además, también se remueven los brotes masculinos reduciendo la posibilidad de inoculación por insectos a través de las flores masculinas. El análisis reciente del ADN realizado mediante las técnicas RFLP y PCR, ha confirmado que la misma cepa de la bacteria causa ambas enfermedades, el Moko y el Bugtok. Las diferencias en la sintomatología se deben a las diferencias en los sistemas de manejo bajo los cuales se producen diferentes variedades. En el caso de la producción de los bananos de cocción por pequeños agricultores, raramente se practica el deshije, el embolse de la fruta y la remoción del brote masculino. Además, los brotes masculinos de las variedades de los bananos de cocción son más dulces y se utilizan como alimento. Ellos parecen ser más atractivos para los insectos que los brotes masculinos de Cavendish, que son más amargos y no pueden ser consumidos. La enfermedad sanguínea, causada por Pseudomonas celebensis (la identidad taxonómica todavía necesita ser comprobada), es una enfermedad devastadora de los bananos en Indonesia. Se reportó por primera vez en Sulawesi en los años 30, y luego se propagó a Java Occidental en los años 80. Recientemente, está causando daños en la parte sur de la isla de Sumatra. Como Bugtok en Filipinas, la enfermedad causa daños considerables al banano de cocción Pisang Kepok, que también es un cultivar con la constitución genómica de balbisiana. Esta enfermedad prácticamente ha eliminado el Pisang Kepok en la isla de Sulawesi. La enfermedad sanguínea es sistémica como el Moko y puede propagarse por las herramientas de poda contaminadas, así como por el suelo. Sin embargo, las observaciones en el campo indican que los síntomas de la podredumbre de la fruta son los más comunes en el Pisang Kepok. Esto indica que como en el Bugtok, la inoculación de la enfermedad se produce a través de las flores. Esta conclusión es apoyada por las observaciones de los fitopatólogos de Indonesia, que consisten en que un cultivar de tipo Pisang Kepok, Frutos mostrando una decoloración de la pulpa debida a la enfermedad de Bugtok. que no produce un brote masculino desarrollado, es significativamente menos afectado por la enfermedad sanguínea. Teniendo en cuenta esta información, actualmente se llevan a cabo ensayos de campo en la isla de Sumatra con el fin de desarrollar una estrategia de manejo integrado de plagas para la enfermedad sanguínea. Se examinará la eficacia de la estrategia de manejo de la enfermedad desarrollada para el Bugtok en Filipinas, basada en la eliminación temprana de los brotes masculinos y saneamiento, con respecto a la enfermedad sanguínea. Se espera que esta técnica sencilla, sola o en combinación con otras técnicas, como es el uso del material de plantación sano, ayudará a resolver el problema de la enfermedad sanguínea en Indonesia. ■ Para obtener más información, por favor diríjase al Dr Agustín Molina, Coordinador Regional de INIBAP para Asia y el Pacífico, C/o PCARRD, Los Baños, Laguna 4030, Filipinas. La piedra y el banana : un mito indonesio La gente nativa de Poso, un distrito de Celebes Centrales, cuenta que en el principio el cielo estaba muy cerca de la tierra y que el Creador, quien vivía en el cielo, acostumbraba a bajar sus regalos a los hombres con una soga. Un día, bajó de este modo una piedra ; pero nuestros primeros padre y madre no sabían que hacer con ella y preguntaron a su Creador : « ¿Qué hemos de hacer con esta piedra ? Dé nos algo más. » El Creador obedeció y tiró de la soga ; la piedra subió hasta desaparecer de la vista. Dentro de poco vieron la soga bajar del cielo otra vez, y esta vez en su extremo estaba un banano en vez de una piedra. Nuestros primeros padres corrieron hacia el banano y lo tomaron. Luego vino una voz del cielo diciendo : « Ya que ustedes escogieron el banano, su vida será como la de este. Cuando el árbol del banano tiene descendencia, el tallo progenitor muere ; así ustedes morirán y sus hijos ocuparán su lugar. Si hubieran escogido la piedra, su vida sería como la de la piedra, sin cambios e inmortal. » El hombre y su mujer lamentaron su fatal elección, pero ya era demasiado tarde ; es así que por comer un banano, la muerte llegó al mundo. J.G. Frazer, The Belief in Immortality, 1 (Londres, 1913), op. 74-75, citant A.C. Kruijt. 30 INFOMUSA — Vol 8, N° 1 Tesis Ecología, distribución y estructura de las poblaciones de Fusarium oxysporum f. sp. cubense en Kenia Tesis Ph D. (en inglés). University of Kent at Canterbury, Reino Unido por Jackson N Kung’u usarium oxysporum f. sp. cubense (Foc), un hongo del suelo, está causando significativas pérdidas de cultivos a los productores bananeros en Kenia. A través de una encuesta intensiva sobre la enfermedad, se descubrió que el marchitamiento por Fusarium fue un problema serio en todas las principales áreas de cultivo de banano del país. Los cultivares afectados fueron Bluggoe (ABB), Gros Michel (AAA), Muraru (AA ?) y Wang’ae (AB). La enfermedad nunca fue encontrada por encima de la zona de humedad 3. Como un prerequisito para el desarrollo de las medidas sostenibles de control, se estudió la variabilidad genética de Foc en Kenia determinando las agrupaciones de incompatibilidad vegetativa de más de 200 aislados de Foc. La compatibilidad fue registrada con los probadores entre los seis existentes GCV (0124, 0125, 0128, 01220, 01222 y 01212), pero varios aislados fueron compatibles con dos o más probadores del complejo 0124/5/8/20/22. Los aislados pertenecientes al GCV 01212, sin embargo, fueron distintos y no formaron conexiones cruzadas con cualesquiera otros probadores de GCV. Ya que la incompatibilidad vegetativa es una característica controlada genéticamente, se consideró que los aislados pertenecientes a los GCV 0124, 0125, 0128, 01220 y 01222 forman un linaje clonal (CL1) y aquellos del GCV 01212 pertenecen a un linaje clonal diferente (CL2). Este resultado luego fue confirmado por diferentes reacciones de estos aislados en un medio rectificado con el sulfato de cobre. En este medio, las colonias CL2 variaron de un color gris pálido a un gris oscuro o negro y formaron complejos con cobre que se precipitaron en el medio por debajo de los micelios. En contraste, las colonias CL1 fueron de un color gris púrpura pálido y no formaron complejos con el cobre en el medio. Mientras que el linaje clonal CL1 fue muy diseminado en el país y fue aislado de un gran rango de cultivares, el linaje CL2 fue encontrado principalmente en un área contigua a Tanzania (donde fue detectado por primera vez) y sólo fue encontrado en el cultivar Wang’ae (= Ney Poovan). Las pruebas de resistencia de los aislados del Foc al fungicida benomil indicaron que ninguno de los aislados examinados ha sido resistente al fungicida a una dosis de 2.86 g por litro. F INFOMUSA — Vol 8, N° 1 Las pruebas del poder patógeno confirmaron la existencia de las razas 1 y 2 en Kenia. Se encontró que ellos pertenecen al linaje CL1. No se pudo asignar los aislados del linaje CL2 junto con el algunos del linaje CL1 a ninguna raza, lo que sugiere la posibilidad de la existencia de al menos dos razas más en Africa Oriental. La confirmación de una nueva raza o razas fue impedida por una falta de cultivares diferenciales adecuados. Ya que el cultivar Wang’ae fue encontrado susceptible a los aislados de ambos linajes clonales, recomendamos su uso como un cultivar de referencia para las futuras pruebas de poder patógeno que implica los aislados de Africa oriental. El análisis en racimos sobre la presencia o ausencia de los metabolitos secundarios no volátiles producidos por los aislados de Foc in vitro, sugirió una correlación compleja entre los quimiotipos del Foc y otros factores como la raza, cultivar huésped y localización geográfica. Se determinaron los perfiles de los metabolitos hongo-huésped de los cultivares Bluggoe, Gros Michel, Muraru, y Wang’ae. La actividad antifungosa de alguno de estos metabolitos contra una cepa de Cladosporium cucumerinum y Aspergillus niger sugirió que estos compuestos fueron fitoalexinas. La selección o desarrollo de los cultivares de banano resistentes a los linajes clonales CL1, CL2 y otras variantes de Foc podrían representar un mejor enfoque hacia el control del marchitamiento por Fusarium en Kenia. ■ continua de varios aspectos de micropropagación de Musa han llevado a la modificación de los protocolos establecidos y a la adopción de nuevos. El manejo de nuevo germoplasma, que proviene de los programas de mejoramiento, también tuvo alguna influencia sobre los procedimientos establecidos. Por estas razones, esta segunda edición está muy a tiempo y ciertamente ayudará a los prácticos a mejorar y ampliar sus aptitudes en este campo. Para ordenar su ejemplar, dirígase a : International Institute for Tropical Agriculture, C/o Lambourn & Co, 26, Dingwall road, Croydon CR9 3EE, RU. Numerical taxonomic studies of the East African highland bananas (Musa AAA-East Africa) in Uganda Deborah A. Karamura ISBN: 2-910-810-31-3 Los trabajos de investigación desarrollados en el marco de esta tesis fueron financiados por el Department for International Development (DFID, Reino Unido). Libros, etc. Shoot-tip culture for the propagation, conservation and distribution of Musa germplasm D.R. Vuylsteke ISBN: 978 131 140 1 Este manual de 82 páginas es una actualización comprensiva de la primera edición escrita por el mismo autor y publicada en 1989 por el International Plant Genetic Resources Institute (IBPGR, actualmente IPGRI). La micropropagación desempeña un papel importante en la conservación, multiplicación y distribución del germoplasma de Musa a nivel nacional, regional e internacional. Durante la última década, la experimentación El propósito de este estudió consistió en determinar el patrón de variación que existe en los bananos de los altiplanos de Africa Oriental (Musa AAA) cultivados en Uganda, estimar los niveles de diversidad causada por diferentes condiciones de crecimiento y establecer un sistema provisional de clasificación e identificación. Se emplearon las técnicas de taxonomía numérica para determinar el patrón de la variación. Se emplearon sesenta y un caracteres morfológicos para determinar las diferencias entre 238 accesiones disponibles para el estudio. Los bananos de los altiplanos de Africa Oriental han sido mantenidos como un subgrupo dentro de un Grupo (Musa AAA). Las unidades más pequeñas que se distinguen dentro del subgrupo son clones y éstos han sido agrupados en series de clones. Se delinearon cinco series de clones basándose principalmente en los caracteres cualitativos. Las series de clones fueron suficientemente 31 Banana streak disease – an illustrated field guide J.W. Daniells, J.E. Thomas & A.D.W. Geering distintas para que las nuevas accesiones pudieran integrarse. Actualmente, estos datos se utilizan para desarrollar la hipótesis sobre la base evolutiva de los bananos de los altiplanos de Africa Oriental y explotar su vigor, resistencia a plagas y enfermedades en los clones representativos de las diferentes series de clones propuestas. Los ejemplares de esta tesis publicada por INIBAP, están disponibles a pedido en la sede de INIBAP, Unidad de Información y Comunicaciones. Cytogenetics of the genus Musa Ken Shepherd ISBN: 2-910-810-30-5 Este libro de 160 páginas sobre la « Citogenética del género Musa » proporciona información importante que es resultado de toda una vida dedicada a la investigación bana- Information Series QI99018 ISSN 0727-6273 Para más información, dirígase a : INIFAP, Campo experimental Tecomán, Km. 35 Carretera Colima-Manzanillo, Apartado Postal No. 88, Tecomán, Colima, México 28100. Tel./Fax : (332) 4 01 33 Banana offtypes – an illustrated guide J.W Daniells, M.K. Smith & S.D. Hamill Information Series QI99019 ISSN 0727-6273 Cytogenetics of the genus Musa Ken Shepherd IPGRI nera. Ken Shepherd no solo dedicó toda su vida al estudio de los bananos, sino que se ha concentrado sobre un área desatendida por otros investigadores, la citología de Musa. La publicación de este libro está especialmente a tiempo, dado que recientemente surgió un interés en la cariología de Musa y en los nuevos métodos que actualmente están disponibles para el estudio de los cromosomas de Musa. Por lo tanto, INIBAP está particularmente honrada en estar asociada con el esfuerzo del Dr Shepherd y publicamos este libro con el conocimiento de que la información que contiene será de gran valor para los investigadores de Musa en todo el mundo durante los años venideros. Para solicitar un ejemplar del libro, dirígase a la Unidad de Información y Comunicaciones en la sede de INIBAP. Manejo integrado de la Sigatoka negra del plátano Mario Orozco-Santos ISSN: 1405-7050 Este manual técnico de 96 páginas, muy bien ilustrado, presenta los descubrimientos prácticos de los 10 años de investigación de la Sigatoka negra realizada por el Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP) de México, con el propósito de proporcionar a los técnicos y productores bananeros opciones para el manejo de la enfermedad. 32 Son numerosas las enfermedades y las plagas del banano que pueden desarrollarse en nuevas plantaciones a través de material de plantación infectado. Solo las plantas de cultivo de tejidos pueden ser garantizadas libres de estas enfermedades y plagas. A pesar de todo, la mayoría de los agricultores siguen utilizando material de plantación tradicional (pedazos de cormo y hijuelos). Según los agricultores del Queensland, Australia, la mayo razón de no utilizar plantas de cultivos de tejidos es, después del costo (mas bajo), el riesgo de obtener plantas fuera de tipo (offtype). Esta guía ilustrada, publicada por el Queensland Department of Primary Industries, tiene como meta ayudar a las personas encargadas de viveros, el personal de los sectores de agroindustría, investigación, extensión así como los productores, a identificar las plantas fuera de tipo y manejarlas de manera adecuada. Se espera que este manual favorecerá la utilización del cultivo de tejidos y ayudara a disminuir el riesgo de enfermedades y plagas en la industria bananera. Esta guía presenta información sobre la manera de reconocer los bananos fuera de tipo en los viveros y en el campo y incluye 57 ilustraciones de color mostrando las anormalidades asociadas a esto tipo de plantas. La enfermedad del rayado del banano fue observada por primera vez en Australia en 1992, y encontrada después en varias plantaciones comerciales y huertas individuales. En 1996, el Queensland Horticulture Institute inició un proyecto con el propósito de comprender mejor esta enfermedad y desarrollar estrategias para su control. Esta publicación presenta informaciones sobre la causa, la transmisión, los síntomas así como medidas de control para la enfermedad del rayado del banano en Queensland. El amplio rango de síntomas observados durante el periodo del proyecto se ilustra a través de 41 fotos de color y un mapa proporciona detalles sobre la distribución actual de la enfermedad en Australia. Se espera que esta guía ayudará en la identificación de la enfermedad en el campo. Para más información sobre estas dos guías ilustradas, dirígase a : Department of Primary Industries, GPO Box 46, Brisbane, Qld 4001, Australia. ■ ✡nnonce Anuncio Taller internacional sobre la producción y comercialización de bananos orgánicos producidos por pequeños agricultores 1-4 de Noviembre 1999, República Dominicana Muchos agricultores a pequeña escala se enfrentan con la creciente dificultad de competir directamente con los productores a gran escala en una economía de libre mercado. Debido a este hecho, se hizo clara la necesidad de buscar alternativas para la producción y diversificación para estos agricultores. El creciente interés en la producción orgánica y en su mercado por parte de los consumidores en los países importadores ofrecen una posible solución. En respuesta a esta demanda, tres organismos internacionales, CAB International, INIBAP y CTA, están organizando conjuntamente un taller para examinar estos tópicos, los retos y oportunidades, que enfrentan los pequeños productores de bananos orgánicos. El taller tendrá un enfoque específico sobre los requerimientos y limitaciones a la producción y comercialización de bananos orgánicos INFOMUSA — Vol 8, N° 1 producidos por pequeños productores en la región de América Latina y el Caribe. Se planea que sean representados los principales participantes en la cadena entera, desde el campo hasta la mesa (productores, servicios de investigación y extensión, miembros de las asociaciones de bananeros, ONG, certificadores, exportadores e importadores, agentes, comerciantes y minoristas). El objetivo de la reunión consistirá en brindar un foro imparcial que catalice las discusiones entre los principales participantes con el fin de crear un entendimiento común sobre los tópicos clave, implicados en una iniciativa de producción orgánica, y determinar cómo esto puede ser orientado e impulsado hacia delante de manera conjunta. El taller consistirá principalmente de las sesiones de discusión y planeamiento, aunque se presentarán algunas charlas por parte de los principales oradores y se incluirá un viaje para visitar las zonas de producción de bananos orgánicos en la República Dominicana. Los organizadores del taller también ofrecen la posibilidad de un viaje posterior a Cuba para un taller en el campo y para ver la producción orgánica de los híbridos mejorados de Musa con resistencia a las principales enfermedades. La producción de estas nuevas variedades, que mantiene una gran promesa para la producción orgánica a una escala más amplia, es muy avanzada en Cuba. Le invitamos a participar en este taller (indicando si le interesa o no visitar Cuba, que será por cuenta de los participantes). Es necesario destacar que con el fin de asegurar un diálogo eficaz, el taller será limitado a una cantidad restringida de participantes. Se harán esfuerzos para asegurar una representación balanceada de cada una de las principales categorías de participantes. Una cantidad limitada de participantes de los países en vías de desarrollo será patrocinada por los organizadores. Para obtener más información sobre el taller, dirígase a : INIBAP (E. Frison, S. Sharrock) Tel : +33 467 61 13 02 ; Fax : +33 467 61 03 34 ; correo electrónico : inibap@cgiar. org CAB International (M. Holderness) Tel : +44 1 491 829 043 ; Fax : +44 1 491 829 100 ; correo electrónico : m. holderness@cabi. org CTA (I. Boto) Tel : +31 317 467 157 ; Fax : +31 317 460 067 ; correo electrónico : boto@cta. nl. Noticias de INIBAP Contrataciones Jean-Vincent Escalant se ha unido recientemente a INIBAP como Científico encargado de Recursos Genéticos de Musa, después de la partida de Jean-Pierre Horry. Jean-Vincent es un ciudadano francés, obtuvo su Doctorado en Fitofisiología en la Universidad de Montpellier, Francia en 1987. Jean-Vincent tiene más de 10 años de experiencia en la investigación de Musa, especialmente en las áreas de cultivo in vitro, embriogénesis somática y biotecnología INFOMUSA — Vol 8, N° 1 de Musa en general. Trabajó en CATIE, Costa Rica, de 1988 a 1996, donde se convirtió en el Líder del Grupo de la Unidad de Biotecnología. En este cargo, realizó investigaciones de una gran variedad de cultivos tropicales, frutas y árboles, incluyendo el banano y el plátano. Estando en Costa Rica, Jean-Vincent desarrolló estrechas relaciones con la Oficina Regional de INIBAP para América Latina y el Caribe. Brindó asistencia técnica a INIBAP-LACNET en el área de biotecnología y manejó el grupo de trabajo temático sobre ‘Biotecnología para los bananos y plátanos domésticos en América Latina y el Caribe’. De 1997 a 1999, Jean-Vincent trabajó como Coordinador Científico en el CRBP, Camerún, donde desempeñó un papel principal en el manejo, organización y orientación de la investigación de banano y plátano. También trabajó estrechamente con la Oficina Regional de INIBAP para Africa Occidental y Central y brindó apoyo a la red regional de investigación de Musa, MUSACO. Hélène Doco, una ciudadana francesa, fue contratada recientemente para la posición de Editora de Web. Hélène tiene experiencia en Agronomía y Ciencias de la Información. Habla con fluidez en inglés y alemán. Hélène tiene siete años de experiencia en el manejo de información y documentación agronómica y trabajó principalmente para el CIRAD, Montpellier, donde estuvo a cargo de la edición del boletín bibliográfico Sésame y del diseño y mantenimiento de un servidor de Internet. Su principal tarea en INIBAP es el desarrollo y mantenimiento de un sitio Web de INIBAP que incluirá el acceso a las bases de datos de INIBAP, publicaciones electrónicas, noticias, etc. Guido Ponsioen, un ciudadano holandés, ha sido seleccionado para la posición de Especialista en Información y Comunicaciones y asumió sus labores el 12 de abril. Guido obtuvo su calificación en el manejo de la información en 1993 y empezó a trabajar como documentalista a medio tiempo en la Facultad de Medicina de la Universidad de Leiden, Países Bajos. Desde 1996 hasta principios de 1999, Guido también se desempeñó como bibliotecario asistente a medio tiempo en ISNAR, uno de los centros de CGIAR basado en La Haya. En su nueva posición, Guido estará a cargo principalmente del manejo de Musalit, la base de datos trilingüe de INIBAP y del mejoramiento de su acceso a los usuarios. INIBAP aprovecha esta oportunidad para dar la bienvenida a Jean-Vincent, Hélène y Guido y desearles éxitos en sus nuevas posiciones. del mejoramiento de las plantas en el marco del programa de investigación de hortalizas. Durante los cinco años que trabajo en INIBAP, Jean-Pierre Horry tuvo como papel dirigir y animar el programa de conservación e intercambio de recursos genéticos de la Red. En el marco de este programa, mantuvo la colección de germoplasma del Centro de Tránsito de INIBAP (ITC) y aseguro la disponibilidad de las accesiones para el intercambio. Participó a la elaboración de la normas y acuerdos que rigen ahora la introducción y el intercambio de germoplasma en el ITC. Así, averiguó que el material conservado por INIBAP respetaba los acuerdos firmados con la FAO. Desarrollo también en colaboración con el CIRAD el proyecto de caracterización por marcadores moleculares de la colección del ITC. Invierto también muchos de sus esfuerzos en el desarrollo del sistema de información sobre el germoplasma de Musa (MGIS — Musa Germplasm Information System), aportando sus conocimientos científicos y el apoyo indispensable para llevar a buen término este proyecto. Participo de manera activa a las reuniones de desarrollo del software y condujo tres talleres regionales de capacitación en la metodología del MGIS. A partir de 1994, dedico parte de su tiempo a la actualización de la lista de descriptores para el banano publicada en 1991 por el IBPGR. Esta nueva lista, que incluye 121 descriptores, fue publicada por el IPGRI en colaboración con el CIRAD en 1997. Procuró después que esta lista fuera utilizada por los investigadores como una lista de referencia para todos sus trabajos de identificación varietal en colecciones establecidas o durante expediciones de colección. Supervisó la publicación del primer Musalogue sobre variedades colectadas en Papúa-Nueva Guinea. Durante una visita a la FHIA, Honduras en 1992, Jean-Pierre había resaltado el interés de conservar el trabajo de caracterización y evaluación desarrollado en los años 50 y 60 por el Dr Allen sobre la diversidad de la colección de germoplasma de la United Fruit Co. en Honduras. Hoy en día, un libro que refleja el trabajo del Dr Allen esta en preparación en colaboración estrecha con la FHIA. Jean-Vincent Escalant Hélène Doco Guido Ponsioen Jean-Pierre Horry Partida de Jean-Pierre Horry Jean-Pierre Horry, Doctor en Fitomejoramiento, fue encargado del germoplasma en INIBAP de 1994 a Abril 1999, compartiendo con la Red su amplia experiencia en taxonomía y en mejoramiento de las variedades de banano. Ahora ha vuelto en el CIRAD, su instituto de origen, para tomar la responsabilidad 33 Acuerdos de Transferencia de Material (ATM) La Red Internacional para el Mejoramiento del Banano y el Plátano suministra el material que se adjunta con las siguientes condiciones : Acuerdo de transferencia de germoplasma designado La Red Internacional para el Mejoramiento del Banano y el Plátano como parte del Instituto Internacional de Recursos Fitogenéticos (referido debajo como INIBAP) pone a disposición el material descrito en la lista adjunta como parte de su política de aprovechamiento máximo del material genético con fines de investigación. El material o se adquirió antes de la entrada en vigor del Convenio sobre la Diversidad Biológica ; o bien, si se adquirió después de la entrada en vigor del Convenio sobre la Diversidad Biológica, se obtuvo quedando entendido que podría ponerse libremente a disposición con fines de investigación agrícola o mejoramiento. El material se mantiene en depósito con arreglo a las condiciones de un acuerdo entre INIBAP y la FAO, y el receptor no tiene derecho a obtener derechos de propiedad intelectual (DPI) sobre el germoplasma o la información conexa. El receptor puede reproducir y utilizar el material con fines de investigación agrícola y mejoramiento y lo puede distribuir a otras partes, siempre que el receptor esté también dispuesto a aceptar las condiciones del presente acuerdo1. Por consiguiente, el receptor acuerda por la presente no reclamar la propiedad sobre el germoplasma que reciba ni solicitar DPI sobre ese germoplasma o la información conexa. Acuerda asimismo garantizar que cualquier persona o institución a disposición de la cual pueda poner posteriormente muestras de germoplasma esté vinculada por la misma disposición y se comprometa a trasmitir las mismas obligaciones a los receptores futuros de germoplasma. INIBAP no ofrece garantías en cuanto a la seguridad o el título del material, ni en cuanto a la exactitud o corrección de cualquier dato de pasaporte o de otro tipo suministrado con el material. Tampoco ofrece ninguna garantía en cuanto a la calidad, la disponibilidad o la pureza (genética o mecánica) del material que suministra. La situación fitosanitaria del material se garantiza sólo con arreglo a (1) Esto no impide que el receptor pueda distribuir o reproducir las semillas con el fin de ponerlas directamente a disposición de los agricultores o consumidores para su cultivo, siempre que se cumplan las demás condiciones establecidas en el ATM. 34 lo descrito en el certificado fitosanitario adjunto. El receptor asume la plena responsabilidad del cumplimiento de la reglamentación de cuarentena/bioseguridad del país receptor en cuanto a la importación o distribución de material genético. Previa solicitud, INIBAP facilitará la información que pueda estar disponible, además de la que se proporciona con el material. Los receptores deberán suministrar a INIBAP los datos de rendimiento obtenidos durante las evaluaciones. El material se suministra con la condición expresa de que se acepten las condiciones del presente acuerdo. La aceptación del material por parte del receptor constituye la aceptación de las condiciones del presente acuerdo. Acuerdo de transferencia de variedades mejoradas de Musa 1. La Red Internacional para el Mejoramiento de Bananos y Plátanos (la cual en adelante se denominará INIBAP) como parte del Instituto Internacional de Recursos Fitogenéticos traspasa el germoplasma enumerado abajo y la información relacionada al receptor bajo los términos y condiciones de este acuerdo. 2. Este germoplasma ha sido adquirido por INIBAP del (los) programa(s) de mejoramiento, el (los) cual (es) en adelante se denomina (n) ‘el (los) Proveedor (es)’) bajo los términos y condiciones establecidos en el (los) ‘Acuerdo(s) para la adquisición de variedades mejoradas de Musa’. 3. El receptor accede a no reclamar los derechos de propiedad sobre este germoplasma, ni solicitar protección de la propiedad intelectual sobre el germoplasma recibido. El receptor también accede a asegurar que cualquier persona o institución, a la cual el mismo pueda entregar muestras de este germoplasma, estará sujeto a las mismas disposiciones. 4. El receptor puede utilizar este germoplasma con fines de investigación, evaluación y producción de acuerdo con los términos de este acuerdo. 5. El receptor, al distribuir este germoplasma, establecerá claramente que el mismo ha sido suministrado por el (los) Proveedor (es), como se especifica en el catálogo de germoplasma. El receptor requiere la obtención del mismo compromiso por parte de cualquier otro receptor subsiguiente al cual se le puede distribuir este germoplasma en el futuro. 6. Del receptor se requiere : a) concertar un acuerdo con el (los) Proveedor (es) antes de : • realizar la propagación in vitro del material de plantación para su venta, • producción de la fruta para exportar, • producción comercial en un país en vías de desarrollo, • reclamar propiedad o protección de los derechos sobre la propiedad intelectual del material derivado esencialmente del germoplasma suministrado bajo este acuerdo. • Renombrar el germoplasma recibido bajo este acuerdo b)Proporcionar a INIBAP cualesquiera datos de evaluación recolectados sobre el germoplasma traspasado bajo este acuerdo ; c) informar a INIBAP los nombres de personas o instituciones a las cuales se le suministraron materiales recibidos bajo este acuerdo ; d)informar al Proveedor del germoplasma sobre todas las publicaciones o documentos relacionados con este material ; e)obtener el mismo compromiso de cualquier receptor subsiguiente al cual pueda distribuir este germoplasma en el futuro. 7. INIBAP comparará los datos de evaluación y pondrá gratuitamente esta información a disposición del (los) Proveedor (es) y otras partes interesadas. Catálogo de germoplasma Nùmero ITC Nombre de la accesión Para el Receptor Para INIBAP Firma : Firma: Nombre : Nombre : Título : Título : Fecha : Fecha : Proveedor INFOMUSA — Vol 8, N° 1 Jean-Pierre efectuó numerosas misiones científicas en el mundo para INIBAP con el fin de aportar a los institutos de investigación, sus conocimientos científicos en recursos genéticos y mejoramiento varietal. Tenemos también que resaltar aquí que durante cinco años, la sede de INIBAP en Montpellier disfruto de la ‘fuerza tranquila’ de Jean-Pierre así como de su sentido de humor. Les deseamos a Jean-Pierre muchos éxitos en sus nuevas funciones ! República de Filipinas – Acuerdo con INIBAP El Gobierno de Filipinas, a través del Departamento de Relaciones Extranjeras, firmó recientemente un Memorándum de Entendimiento con INIBAP. Filipinas desempeña un papel dirigente en la investigación de bananos y plátanos en la región de Asia y fuera de ella, y es un socio importante en ASPNET, la red regional de investigación en Musa. El Acuerdo formaliza el establecimiento y operación de la Oficina Regional de INIBAP para Asia y el Pacífico en Filipinas y brindará oportunidades para el fortalecimiento de los ya fuertes enlaces entre el Gobierno de Filipinas e INIBAP. Nuevos acuerdos para la distribución de germoplasma En octubre de 1994, IPGRI firmó un acuerdo con la FAO referente a la colocación de las accesiones designadas en la colección de germoplasma de Musa de INIBAP, bajo los auspicios de la FAO como parte de la Red Internacional de Colecciones ex situ. El germoplasma designado en este acuerdo es mantenido por INIBAP « en depósito para el beneficio de la comunidad internacional, en particular de los países en vías de desarrollo, de acuerdo con el Compromiso Internacional sobre los Recursos Fitogenéticos. » En octubre de 1998, un nuevo Acuerdo sobre la Transferencia de Material de Germoplasma, designado bajo los acuerdos con la FAO, fue confirmado y aprobado para su uso a través del sistema de CGIAR. Este Acuerdo sobre la Transferencia de Material no requiere la firma del receptor de germoplasma. En vez de esto, utiliza un enfoque, que obliga al receptor a cumplir con los términos y condiciones expresados en el Acuerdo sobre la Transferencia de Material con tal de que el receptor acepte y retenga el material. El texto del nuevo Acuerdo sobre la Transferencia de Material fue adoptado por INIBAP y su texto completo se presenta en p. 34. Firma del acuerdo entre INIBAP y el Gobierno de Filipinas - (la izquierda, el Secretariado de Asuntos extranjeros, P.L. Siazon Jr, a la derecha, E. Frison, Director de INIBAP). INFOMUSA — Vol 8, N° 1 En adición al germoplasma ‘designado por la FAO’, INIBAP también mantiene material que no está cubierto por los acuerdos con la FAO. Este material lo representan variedades mejoradas adquiridas por INIBAP de los programas de mejoramiento bajo los términos y condiciones de un acuerdo especial sobre la adquisición de germoplasma. Este acuerdo permite a INIBAP distribuir estas variedades para la investigación, evaluación y producción sólo bajo los términos de un ‘Acuerdo para la transferencia de las variedades mejoradas de Musa’. Este acuerdo requiere que los receptores establezcan un acuerdo específico con el proveedor originario del germoplasma (el programa de mejoramiento) en ciertas circunstancias. De los receptores se requiere firmar este acuerdo, cuyo texto completo se presenta abajo, antes de que el material sea distribuido por el Centro de Tránsito de INIBAP. INIBAP ha desarrollado un nuevo formato para ordenar el germoplasma, tomando en cuenta el hecho de que actualmente se requieren diferentes acuerdos sobre la transferencia para diferentes tipos de germoplasma. Para facilitar el pedido de material, pronto se confeccionará un listado del germoplasma disponible en INIBAP en su sitio Web, junto con las versiones electrónicas del formulario para el pedido de germoplasma y acuerdos de transferencia. Para obtener más información con respecto a la distribución de germoplasma por INIBAP, dirígase a S. Sharrock, Sede de INIBAP (Fax : +33 4 67 61 03 34, correo electrónico : s. sharrock@cgiar. org) o I. Van den Houwe, Centro de Tránsito de INIBAP (Fax : +32 16 32 19 93, correo electrónico : ines.vandenhouwe@agr. kuleuven. ac. be. ■ Noticias de Musa Asia y el Pacífico Podredumbre del pedúnculo en India Se notó que la podredumbre del pedúnculo está causando una seria amenaza a la producción bananera en algunas áreas de Tamil Nadu, el segundo estado productor de importancia en India. El problema es particularmente pronunciado en las regiones con altas temperaturas (mayores de 35°C) y donde predominan las enfermedades de la mancha de la hoja. Por lo tanto, se realizaron estudios para determinar la causa de la enfermedad, patogenia de los hon- Figura 1. Banano mostrando síntomas de podredumbre del pedúnculo. Figura 2. Podredumbre del pedúnculo : síntomas típicos gos implicados, susceptibilidad de varias variedades y prácticas de manejo para prevenir la ocurrencia de la enfermedad. Las observaciones revelaron que la emergencia de los racimos se produce principalmente durante los meses de verano y que debido al número reducido de hojas funcionales por enfermedades de la mancha de la hoja, la porción superior del pedúnculo queda expuesta al sol caliente. El pedúnculo pierde el color y por consiguiente empieza a pudrirse (Figura 1). La pudrición continúa propagándose hacia arriba y hacia abajo por el pedúnculo, lo que da como resultado dedos de tamaño reducido y dedos sin desarrollarse o parcialmente llenos. Las observaciones también mostraron que cuando la infección ocurre en una etapa temprana, la enfermedad puede adquirir suficiente severidad como para propagarse en los dedos y producir una completa destrucción del racimo (Figura 2). Se recolectaron muestras de los cultivares Karpuravalli (Pisang Awak — ABB) y Rasthali (Silk – AAB), se aislaron dos hongos, Colletotrichum gloeosporioides y Botryodiplodia theobromae, y se confirmó su patogeneidad. Se descubrió que tanto el Pisang Awak y Silk, como otros cultivares de importancia comercial incluyendo el Panchanadan (Pome – AAB) y Robusta (Cavendish – AAA), resultaron susceptibles a la podredumbre del pedúnculo. La incidencia de la enfermedades en las fincas bananeras encuestadas varió de un 5 a un 25 % de plantas infectadas. Se descubrió que se puede prevenir la enfermedad aplicando al pedúnculo un rociado profiláctico con carbendazin a una concentración de 0.1 % inmediatamente después de la emergencia del retoño y cubriendo el pedúnculo con hojas para asegurar su protección contra el sol. Este trabajo fue realizado por R. Thangavelu, B. Padmanaban y P. Sundararaju en el National Research Centre on Banana, Tiruchirapalli, India. Estudios de nutrientes en el banano utilizando 32P Los investigadores de la Kerala Agricultural University han estado estudiando el movimiento de nutrientes en las plantas de banano con la ayuda de 32P radioactivo. En el primer estudio, basándose en el conocimiento de que el látex suministra una guía útil para determinar el potencial hídrico de la planta (Milburn et al. 1990), los investigadores decidieron examinar al látex como una posible alternativa para el estudio del estado y el reciclaje de los nutrientes en bananos. Para el estudio se utilizó el cultivar Mysore (AAB). Una dosis estándar de 1mCi 32P fue inyectada en el seudotallo de la planta madre en ocho lugares definidos. Luego se midió la recuperación de la radioactividad en la savia recolectada del seudotallo a 10 cm 35 aproximadamente sobre el nivel del suelo y también de las muestras tomadas de la vena media de la segunda hoja totalmente abierta. El muestreo fue realizado dos veces, 15 y 25 días después de la aplicación. La recuperación de la actividad fue bien observada en todos los tratamientos en la savia de las muestras tanto del seudotallo, como de las hojas. El análisis químico del látex de banano mostró que el mismo es rico en iones, especialmente K +, Mg2 +, Cl- y NO3-, por lo tanto se considera que el látex brinda una alternativa para el muestreo destructivo, secado, digestión y análisis químico del tejido de la planta en los estudios de los nutrientes. El 32P radioactivo también fue utilizado para estudiar el patrón de distribución de los nutrientes desde la madre hacia el hijo en las plantas de banano con cantidad variable de retoños. Debido a la escasez de material de plantación, en muchos países se practica la producción comercial de retoños. Por lo tanto, este estudio fue realizado utilizando el cultivar Mysore para investigar el nivel de competición entre los retoños en la misma mata, utilizando uno, dos o tres retoños por mata. Los resultados mostraron que cuando sólo se retiene un retoño, la absorción de nutrientes por este de la planta madre continúa por un período de tiempo prolongado. Sin embargo, cuando se retienen dos o tres retoños, el primer retoño rápidamente completa la absorción de nutrientes y se vuelve independiente, permitiendo que los nutrientes se dirijan a los retoños subsiguientes. Bibliografía : Milburn J.A., J. Kallarackal y D.A. Baker. 1990. Water relations of the banana I. Using solute potential of the latex for predicting the potential of field-grown banana. Aust. J. Plant Physiol. 17: 57-68. Más información sobre este trabajo se puede obtener de los autores : S. Kurien, B.K. Anil, P. Suresh Kumar, P.A. Wahid y N.V. Kamalam, College of Horticulture, Kerala Agriculture University, India. Embriogénesis somática en los cultivares indonesios de banano En el marco del proyecto titulado « Identificación de germoplasma y uso de las técnicas in vitro para apoyar el mejoramiento genético de los bananos en Indonesia », se examinaron varias técnicas para la producción de embriones somáticos en las variedades indonesias de banano. La técnica con la utilización de flores masculinas Escalant et al, (1994) fue aplicada a 5 cultivares indonesios populares : Baranang (AAA), Ambon Kuning (AAA), Raja (AAB), Raja Sereh (AAB) y Kepok Kuning (ABB). Los resultados mostraron que la mayoría de los explantes (flores masculinas inmaduras) fueron capaces de producir callos. El callo tenía una textura compacta o friable, su color era blanco o amarillo y algunas veces estaba acompañado por nódulos. A la edad de 5-6 meses, los callos se tornaron de color marrón y aparecieron algunos nódulos amarillos suaves. Estos nódulos tenían entre 1 y 4 mm de largo y era posible arrancarlos fácilmente de los callos. La cantidad de nódulos varió entre los cultivares y 36 las posiciones de las flores. En general, las flores de las filas de la 4 a la 9 produjeron una gran cantidad de nódulos. La transferencia de los nódulos a un medio libre de hormonas dio como resultado la multiplicación de los mismos. También se produjo una pequeña cantidad de raíces y brotes en este medio, aparentemente a través de la organogénesis. La adición de citoquinina al medio de regeneración causó que los nódulos se multiplicaran y, en el caso de Baranang y Ambon Kuning, en la superficie de los nódulos se produjo un callo amarillo friable. El estudio histológico de este callo reveló la presencia de células embriogénicas, produciendo embriones y proembriones somáticos. Estos resultados muestran por primera vez que los embriones somáticos pueden ser obtenidos a partir de los cultivares indonesios, abriendo así una puerta para un futuro trabajo sobre los sistemas de propagación en masa y transformación para estos cultivares. Bibliografía : Escalant J.V., C. Teisson y F. Côte. 1994. Amplified somatic embryogenesis from male flower of triploid banana and plantain cultivars (Musa spp.) in vitro. Cell Dev. Biol. 30 : 181-186. Más información sobre este trabajo está disponible con Rita Megia, Department of Biology, FMIPA, Bogor Agricultural University, Indonesia. Investigación sobre el cultivar Saba en Filipinas La Universidad de Filipinas en Los Baños (UPLB) ha empezado un proyecto, financiado con fondos de DOST-PCARRD y apoyo técnico de INIBAP-ASPNET, con el objetivo de mejorar la productividad del cultivar Saba. Los métodos de producción, que están evaluados para pequeños productores, incluyen el estudio de la densidad de población, el uso de fertilizantes y el manejo de la enfermedad de Bugtok. El proyecto esta también evaluando varios lineajes de Saba para procesamiento. El objetivo principal es identificar el mejor lineaje de Saba para su procesamiento en chips, que representa un producto de exportación de los mas importantes en Filipinas. Otro producto en evaluación se llama localmente « Pinoy Fries » y consiste en tajadas de Saba preparadas como « papas a la francesa ». Este producto podría remplazar la papas fritas en los restaurantes de comida rápida. Otro aspecto del proyecto se dedica a la utilización de hojas secas de banano como substrato para la producción de una especie de hongo tropical llamado « Volvariela vulvacea ». Este tipo de hongo es muy popular en Asia y las hojas de banano resultan como un substrato muy eficiente. La producción y utilización mejorada e integrada de Saba podría mejorar la vida de numerosos pequeños productores de banano en el país. Más información sobre el proyecto se puede obtener de Gus Molina, Coordinador regional de INIBAP-ASPNET, C/o zando un molino manual en morteros. Esto requiere una mano y un mortero esterilizados para cada muestra. Para minimizar la contaminación cruzada y hacer el manejo de las muestras más fácil, se exploró la posibilidad de utilizar bolsas plásticas para la recolección y molido de las muestras. Muestras que consistían de aproximadamente 3 g de tejido foliar fueron recolectadas por duplicado en bolsas plásticas (polietileno) de calibre 150 de tamaño 24 x 13 cm. A una serie de muestras en bolsas plásticas se le añadió un búfer de carbonato a un pH 9.8 y a una tasa de 1 ml/g, luego la muestra fue molida con una mano de mortero. La misma mano fue utilizada nuevamente para moler todas las muestras, ya que la misma no había sido contaminada por la savia debido a su confinamiento en la bolsa. Las alicuotas de 100 ml de la savia expresada fueron colocadas con la ayuda de una pipeta en los pozos de un plato de microtitulado de 96 pozos. La segunda serie de muestras fue procesada de la misma manera, pero para extraer el antígeno se utilizaron manos y morteros individuales. Para realizar la prueba ELISA se liberó una cantidad suficiente de viriones mediante el molido en las bolsas plásticas. Como la misma bolsa fue utilizada para recoger y moler las muestras, se eliminaron las posibilidades de mezclar contaminar las muestras. Por lo tanto, esta técnica proporciona un método útil para el cribado masivo del germoplasma de Musa para determinar enfermedades virales. Más información sobre esta técnica se puede obtener de Duleep Kumar Samuel, Fruit Crop Virology Lab., Indian Institute of Horticultural Research, Hessaraghatta Lake Post, Bangalore – 560 089, India. Investigaciones en curso en Maroochy, Australia El cultivo de tejidos desempeña un papel importante en la industria bananera de Australia especialmente en relación con el suministro de material de plantación libre de plagas y enfermedades. Para apoyar la industria bananera en Australia, la Maroochy Horticulture Research Station realiza investigaciones relacionadas con el cultivo de tejidos y produce material vegetal examinado en relación a la presencia de patógenos para los proyectos de investigación y desarrollo en la región. A continuación se describen las investigaciones actuales y los resultados obtenidos recientemente. Para obtener más información, dirígese a M. Smith o S. Hamill, MHRS, QDPI, PO Box 5083, SCMC, Nambour, Queensland 4560, Australia. Ver también noticias de PROMUSA para obtener información respecto a la investigación sobre la transformación genética de Musa en Queensland. PCARRD, Laguna 4030, Los Baños, Filipinas. El uso de bolsas plásticas para la recolección de muestras de virus de banano y extracción de antígenos La extracción de antígenos a partir de las muestras de banano normalmente se realiza utili- Uso de un marcador basado en SCAR para la detección precoz de los tipos enanos en los bananos Cavendish micropropagados Con el fin de obtener un método más robusto y confiable para detectar tipos enanos en los bananos Cavendish micropropagados, se deINFOMUSA — Vol 8, N° 1 sarrolló un marcador basado en PCR, que define la región amplificada caracterizada de la secuencia (sequence characterized amplified region — SCAR), en comparación con un marcador RAPD (Plant Cell Reports 16 : 118123). Este marcador basado en SCAR permite la detección precoz in vitro de los tipos enanos. También se utilizó como una guía precoz para detectar los factores que promueven la inestabilidad genética durante la micropropagación. Los tipos enanos pueden surgir temprano durante el proceso de multiplicación y fueron detectados en el cuarto subcultivo después de la iniciación de los brotes apicales. Los resultados también indican fuertemente que la multiplicación de los brotes adventicios es el principal factor que contribuye a la formación de los tipos enanos. Los brotes adventicios son promovidos por las mayores concentraciones de la fitohormona benzilaminopurina (BAP), seleccionando con preferencia los racimos muy pequeños, retoños o estructuras parecidas a los bulbos como los propágulos para la multiplicación subsiguiente. También se descubrió que los tipos enanos tienen una mayor tasa de multiplicación que las plántulas de control y su cantidad puede aumentar a una tasa desproporcionada durante la micropropagación. Se descubrió que la inestabilidad inherente del cultivar a micropropagar es otro factor principal que influye sobre la producción de los tipos enanos. Se determinó que el ‘New Guinea Cavendish’producía enanos con una mayor frecuencia que el ‘Williams’. Más, los tipos enanos fueron más estables in vitro, y las condiciones que favorecen la inducción del enanismo en las plantas de control no favorece la reversión de los enanos en plantas normales. Los bananos micropropagados son más susceptibles al marchitamiento por Fusarium en subtrópicos, que los bananos cultivados de manera convencional Se comparó la reacción de los bananos micropropagados cultivados en el campo, Musa cv. Williams (AAA, subgrupo Cavendish) y cv. Goldfinger (AAAB, FHIA-01), a la raza 4 subtropical de Fusarium oxysporum f. sp. cubense (Foc), con la reacción de las plantas cultivadas a partir de material de plantación convencional (secciones del rizoma llamados pedazos). Se determinó el intercambio de gas en las hojas de las plantas y se midieron el crecimiento y la acumulación de la materia seca. Se realizaron comparaciones entre estos parámetros poco después de la plantación y durante el invierno y la primavera cuando un alto porcentaje de plantas empezó a mostrar síntomas externos del marchitamiento por Fusarium. Los bananos micropropagados fueron significativamente más susceptibles a la raza 4 del Foc, que las plantas derivadas de los pedazos. Este hecho no dependía de las épocas de siembra, cultivares utilizados o de si los pedazos fueron establecidos primero en el invernadero (como las plantas micropropagadas) o se plantaron directamente en el campo. Esta INFOMUSA — Vol 8, N° 1 mayor susceptibilidad no parece ser una consecuencia de las diferencias en las tasas máximas de fotoasimilación, de una mayor demanda de fotoasimilación o de una falta de reservas de carbohidratos después de que la planta se haya establecido. En el QDPI-Indooroopilly y el CRC-Tropical Plant Protection en Brisbane, se realizan estudios para determinar si los aislados no patógenos de Fusarium oxysporum y rizobacterias, recuperados a partir de los bananos cultivados en los suelos que suprimen el marchitamiento, tienen potencial para el control biológico. Estos aislados, que tienen la habilidad de colonizar los tejidos de los bananos, pueden inducir una resistencia sistémica en el huésped o proporcionar una competencia saprofítica con el patógeno. Es posible que el ambiente aséptico en el cual crecen los bananos micropropagados contribuye a una mayor susceptibilidad, cuando éstos se siembran en los suelos infectados con el Foc, debido a la ausencia de los microorganismos beneficiosos en el material de plantación micropropagado. La caracterización y detección precoz de un tipo anormal de los bananos micropropagados Lady Finger Un tipo anormal ha sido identificado en los bananos micropropagados Lady Finger (Musa, AAB, Subgrupo Pome) caracterizado por un crecimiento lento y un racimo pobre. El peso de los racimos fue aproximadamente el 25 % del peso de los racimos de las plantas normales de Lady Finger y toda la fruta producida no servía para la comercialización. Este tipo anormal particular es el más común que se presenta en las plantas micropropagadas de Lady Finger y, en dos casos, bloques enteros de 3 000 y 1 500 plantas contenían enteramente este único tipo anormal. La detección de las plantas anormales fue posible durante el establecimiento y cultivo de las plantas en el invernadero. Los tipos anormales fueron detectados por la presencia de estrías cloróticas en las hojas. En los casos más severos, las estrías se fundieron en manchas cloróticas que desarrollaron áreas necróticas delgadas las cuales eventualmente producían huecos o desgarramiento en las hojas. La expresión de los síntomas no fue mejorada al añadir el fertilizante y aunque los síntomas fueron similares a los de la deficiencia severa de Ca y B, las plantas tanto normales como anormales tenían niveles similares de estos elementos en las hojas. Las plantas anormales también mostraron un crecimiento lento en el invernadero y produjeron seudotallos y hojas significativamente más pequeños (P < 0.05) que las plantas normales. Las plantas anormales pueden ser detectadas fácilmente después de cuatro semanas de liberarlas de las probetas utilizando la presencia de las estrías cloróticas en las hojas como criterio principal. Fue posible realizar la discriminación máxima entre la quinta y séptima semanas y en la etapa de la hoja 6 cuando todos los tipos anormales pueden ser detectados. Las plantas de banano micropropagadas tienen tasas de crecimiento similares, pero retoños con morfología diferente que las plantas cultivadas en contenedores y propagadas a partir de los pedazos del cormo Las plantas de banano (Musa sp. AAA, subgrupo Cavendish) micropropagadas produjeron significativamente más retoños que las plantas cultivadas a partir del material de plantación convencional (secciones del rizoma con un punto de crecimiento, llamadas pedazos). Sin embargo, cuando los pedazos fueron establecidos en los contenedores en un invernadero y cultivadas bajo las mismas condiciones que las plantas micropropagadas, las diferencias se hicieron menores. Realmente, la altura, la acumulación de materia seca y el área foliar de las plantas madres de las plantas micropropagadas fueron similares a las de las plantas cultivadas a partir de los pedazos en contenedores. El crecimiento de las plantas derivadas a partir de pedazos que han sido establecidas directamente en el campo, siempre se demoraba más que el de las plantas procedentes de las otras dos fuentes. Las diferencias más significativas entre las plantas micropropagadas y aquellas cultivadas a partir de los pedazos en contenedores se mostraron en una mayor área foliar y el peso seco de los retoños de las plantas micropropagadas. Estos retoños con hojas más grandes fueron catalogadas como hijos de agua. Estas diferencias pueden estar relacionadas con la densidad de las raíces y el punto de origen del retoño en el rizoma y tienen implicaciones comerciales para el manejo de los bananos micropropagados. Africa Efecto del sistema de cultivo sobre la severidad de la Sigatoka negra en Nigeria La mayoría de los estudios sobre la Sigatoka en Nigeria han sido realizados en regiones de bosques húmedos, y se dispone de mucho menos información sobre la severidad de esta enfermedad en la Sabana de Guinea nigeriana. En esta región la precipitación no se distribuye uniformemente a través del año, sino que ocurre durante la estación lluviosa desde el mes de abril hasta el mes de octubre. En esta región se reconocen tres sistemas de cultivo de bananos, en patios traseros (suelos enriquecidos con desperdicios orgánicos caseros), Fadamas (suelo aluvial con un alto contenido de materia orgánica) y campos distantes (suelos arenosos arcillosos). Las principales variedades que se cultivan son plátanos (cvs. Agbagba y Ogakwu, AAB), bananos de postre (cvs. Valery y Ogede, AAA) y bananos de cocción (cv. Ogwokwu-Ige, ABB). Para el estudio, que duró más de cuatro años, se seleccionaron seis sitios que representan cada uno de los tres sistemas de cultivo. La severidad de la Sigatoka fue evaluada durante la floración siguiendo los procedimientos recomendados. También se analizaron muestras de suelo de los sitios con respecto al contenido de materia orgánica. Los resultados mostraron que la severidad de la enfermedad fue significativamente más alta 37 en los cultivares que crecían en los campos distantes, donde el contenido de materia orgánica fue bajo (0.95 %), en comparación con la severidad de la enfermedad en las plantas cultivadas en los patios traseros (2.9 % de materia orgánica) y en Fadamas (2.7 % de materia orgánica). La colocación de los desechos caseros alrededor de las matas de plátanos como una fuente constante de materia orgánica en Africa Occidental es considerada como una de las razones de la productividad sostenible de estos cultivos. Similarmente, los nutrientes orgánicos son altos en el sistema basado en Fadamas como resultado de los depósitos fluviales. Estos resultados indican el papel importante que desempeña la fertilidad de los suelos en el manejo de la Sigatoka negra. Este trabajo fue realizado por H.O.A. Oluma y F.N. Onyezili, Department of Biological Sciences, University of Agriculture, PMB 2373, Makurdi, Benue State, Nigeria. Investigación del marchitamiento por Fusarium en Africa del Sur Recientemente se lanzó en Africa del Sur un proyecto con el propósito de obtener una estrategia práctica sostenible para el manejo de Fusarium oxysporum f. sp. cubense (Foc). Los objetivos del proyecto son los siguientes : • Entender la estructura de la población de Foc en Africa del Sur • Estudiar la epidemiología (distribución, diseminación) • Desarrollar una identificación rápida de Foc en Africa del Sur. • Estudiar el papel de los factores edáficos, poblaciones microbianas, clima, nutrición y riego sobre el desarrollo del marchitamiento por Fusarium. • Desarrollar un programa de manejo integrado para disminuir el impacto de la enfermedad sobre la producción bananera. Más información sobre el proyecto se puede obtener de A. Severn-Ellis, ITSC, Private Bag X11208, Nelpruit 1200, Africa del Sur. América Latina y el Caribe Aislamiento de las toxinas a partir del hongo de la Sigatoka amarilla, Mycosphaerella musicola Los investigadores cubanos están trabajando en el aislamiento de las toxinas fungosas con el propósito de utilizarlas en la selección in vitro del germoplasma para detectar la resistencia a las enfermedades de la Sigatoka y del Fusarium. En el caso de la Sigatoka amarilla, los extractos crudos de una fracción tóxica fueron aislados a partir del hongo Mycosphaerella musicola. La fitotoxicidad de estos extractos fue examinada mediante la inoculación directa de una hoja, seguida por una incubación durante 72 horas en una cámara de crecimiento húmeda así como por la inmersión total de la hoja. Las fracciones que presentan la mayor actividad biológica fueron concentrados 10 veces y luego examinados nuevamente. Las toxinas fueron purificadas, utilizando un proceso químico que implica solventes orgánicos, y la actividad biológica fue evaluada inoculando una hoja con la fracción 38 pura y sumergiendo la punta de la hoja en una solución acuosa de la fracción. La actividad máxima se obtuvo con los extractos tomados del hongo durante la fase de su crecimiento óptimo. La actividad se hizo evidente de 24 a 48 horas después de la inoculación y dio como resultado síntomas foliares típicos de la infección con la Sigatoka amarilla. Se considera que las fracciones de toxinas obtenidas de esta manera pueden ser utilizadas en selección precoz del germoplasma para detectar la resistencia a la Sigatoka amarilla. 2 6 1 5 4 3 7 Más información sobre el proyecto se puede obtener de M. Folgueras Montiel y M. Ramos Leal, INIVIT, 5 300 Santo Domingo, Villa Clara, Cuba. Nuevo dispositivo para evaluar la resistencia a la caída de los dedos Los investigadores de Brasil han desarrollado un nuevo dispositivo para medir la resistencia de banano a la caída de los dedos. El dispositivo, llamado ‘Detatcher’, consiste de un penetrómetro montado sobre un chasis de madera (Figura 3) y ha sido utilizado para evaluar la resistencia a la caída de los dedos de las accesiones en el Banco de Germoplasma Activo de Banano en Cruz das Almas, Brasil. Más información sobre el proyecto se puede obtener de V.M. Medina, R.C. Cerqueira y S. de Oliveira e Silva, CNPMF, Caixa Postal 007, CEP 44380-000, Cruz das Almas, Bahia, Brasil. Encuesta sobre el daño causado por el huracán Mitch en América Central Los plátanos son los cultivos alimentarios más importantes en América Central. Después de la devastación del huracán Mitch en octubre de 1998, se efectuaron misiones para evaluar el daño al sector platanero en Honduras, Guatemala y Nicaragua a principios de 1999. Estas misiones descubrieron que la mayor parte de las áreas de producción platanera de Honduras y Guatemala fue destruida y que los plátanos ya no estaban disponibles en los mercados de estos países. La escasez de plátanos en los mercados dio como resultado un aumento de precios sin precedentes, y esto, aunado con la pérdida de ingresos por empleos, ha puesto al plátano fuera del alcance de una gran cantidad de consumidores, rurales y urbanos. Las pérdidas para las economías de Honduras, Guatemala y Nicaragua fueron estimadas en alrededor de 30,000,000. Se desarrolló un proyecto para rehabilitar el sector platanero en Honduras, Nicaragua y Guatemala y se realizan los esfuerzos para identificar fuentes de financiamiento para este importante trabajo. Dirección del movimiento del banano Figura 3. Esquema del dispositivo de evaluación de la resistencia a la caída de los dedos. Cruz Das Almas, BA, Brazil. 1. Penetrometro 2. Tren de tracción 3. Chasis de madera 4. Taqué 5. Cursor del taqué 6. Penetrometro/traba al tren de tracción 7. Banano biológico fueron evaluados con respecto a su habilidad de reducir la enfermedad de la podredumbre de la corona del banano. En total, se evaluaron 12 cepas. La ESC11, una cepa de Pseudomonas syringae registrada por EPA y el BW7 se desempeñaron de la mejor manera. A una concentración de 1 X 108 de las unidades que forman colonias (ufc)/ml, la ESC11 redujo la severidad de la enfermedad en un 73 % aproximadamente. El BW7 a 1 X 108 de ufc/ml resultó ser más variable y redujo la severidad de la enfermedad en 33 % y 56 % en dos experimentos. El tratamiento comercial estándar con tiabendazol (TBZ) e imazalil redujo la severidad de la enfermedad en 70 %. El sulfato de potasio de aluminio (alumbre) se utiliza comúnmente para reducir la secreción del látex en la corona cortada del banano. Se realizaron dos estudios para investigar la compatibilidad de la ESC11 y BW7 con este químico. La habilidad de la ESC11 y el BW7 de proporcionar el control de la podredumbre de la corona disminuyó con la presencia de alumbre en los pedícelos. Con respecto a ESC11, 5 X 107 ufs/ml proporcionaron el mejor control en presencia de 1 % de alumbre, mientras que para el BW7, el mejor control se obtuvo a 3 X 10 8 ufc/ml. Estudios posteriores que incluirán ensayos en el campo a pequeña escala y escala comercial serán conducidos en Costa Rica en 1999 para demostrar la eficacia y confiabilidad de la ESC11 bajo condiciones de campo. También se realizarán estudios en el laboratorio para investigar el posible modo de acción. ■ Control biológico de la podredumbre de la corona del banano La podredumbre de la corona en el banano se considera la enfermedad postcosecha más seria y es causada principalmente por las especies de Fusarium, incluyendo F. moniliforme. Se está llevando a cabo una investigación por la EcoScience Corporation, EEUU, sobre los posibles métodos de control biológico de este patógeno. Los agentes potenciales de control Esta investigación se lleva a cabo por S.M. Williamson, O. Anas y T.B. Sutton, North Carolina State University, Raleigh, NC, EEUU ; M. Guzmán CORBANA, San José, Costa Rica ; D.H. Marin BANDECO, San José, Costa Rica ; Xixuan Jin, EcoScience Corporation, East Brunswick, NJ, EEUU. INFOMUSA — Vol 8, N° 1 Direcciones de INIBAP • Sede Parc Scientifique Agropolis II 34397 Montpellier Cedex 5 FRANCIA e-mail : [email protected] http ://www.cgiar.org/ipgri/inibap/ Director : Dr Emile FRISON e-mail : [email protected] Coordinadora de Mejoramiento Genético : Dra Gisella ORJEDA e-mail : [email protected] Encargado de Recursos Fitogenéticos Dr Jean-Vincent ESCALANT e-mail: [email protected] Encargada de la Conservación de Germoplasma : Sra Suzanne SHARROCK e-mail: [email protected] Jefe Información y Comunicación : Sra Claudine PICQ e-mail: [email protected] Coordinadora MGIS: Sra Elizabeth ARNAUD e-mail: [email protected] Administrador Financiero: Sr Thomas THORNTON e-mail: [email protected] • Red Regional para América Latina y el Caribe Coordinator Regional : Dr Franklin ROSALES C/o CATIE, Apdo 60-7071 Turrialba COSTA RICA Fax : (506) 556 24 31 e-mail : [email protected] • Red Regional para Asia y el Pacífico Coordinator Regional : Dr Agustín MOLINA C/o PCARRD Los Baños, Laguna 4030 - FILIPINAS Fax : (63-49) 536 05 78 e-mail : [email protected] • Red Regional para Africa Occidental y Central Coordinator Regional : Dr Ekow AKYEAMPONG Experto Asociado, Entomología Stjin MESSIAEN C/o CRBP, B.P. 12438 Douala, CAMERUN Fax : (+237) 42 91 56 e-mail : [email protected] • Red Regional para Africa Oriental y del Sur Coordinator Regional : Dr Eldad KARAMURA PO Box 24394 - Kampala, UGANDA Fax : (256-41) 22 35 03 e-mail : [email protected] • Centro de Tránsito INIBAP (ITC) Oficial Encargado : Ing. Ines VAN DEN HOUWE Katholieke Universiteit Leuven Laboratory of Tropical Crop Improvement Kardinaal Mercierlaan 92 B-3001 Heverlee, BELGICA Fax : (32-16) 32 19 93 e-mail : [email protected] • Expertos Asociados, Nematología Inge VAN DEN BERGH VASI Van Dien, Than Tri Hanoi, VIETNAM Fax : (84) 4 861 39 37 e-mail: [email protected] Thomas MOENS - C/o CORBANA Estación de investigaciones La Rita Apdo 390-7210 Guápiles, COSTA RICA Fax : (506) 763 30 55 e-mail: [email protected] Recomendaciones para los autores Los textos mecanografiados deberán ser preparados en inglés, francés o español y enviados al Jefe de Redacción. Todas las páginas (incluyendo las tablas, figuras, leyendas y referencias) deberán estar enumerados consecutivamente. El título deber ser lo más corto posible. Incluya el nombre completo de todos los autores y sus direcciones completas al momento de realizar el estudio. También indique a la persona quien recibirá la correspondencia respecto al trabajo. Si el manuscrito ha sido preparado en una computadora, por favor, envié una copia en disquete (o por correo electrónico) junto con el ejemplar impreso, indicando el nombre y la versión del procesador de palabras utilizado. • Resúmenes : Un resumen que no exceda 200-250 palabras deberá ser enviado en el mismo idioma del manuscrito, así como las traducciones (incluyendo el título) en los otros dos idiomas, si es posible. • Siglas : Las siglas deberán ser transcritas completamente la primera vez que éstas aparecen en el texto, seguidas por las siglas entre paréntesis. INFOMUSA — Vol 7, N°2 • Bibliografía : Todas las referencias bibliográficas deberán ser presentadas en orden alfabético de autores. La referencia en el texto debe indicar el nombre del autor y el año de publicación (por ejemplo : Sarah et al. 1992). Por favor, siga los siguientes ejemplos : Artículos de edicciones periódicas : Sarah J.L., C. Blavignac & M. Boisseau. 1992. Une méthode de laboratoire pour le criblage variétal des bananiers vis-à-vis de la résistance aux nématodes. Fruits 47 (5) : 559564. Libros : Stover R.H. & N.W. Simmonds. 1987. Bananas (3 ra edition). Longman, London, United Kingdom. Artículos (o capítulos) de publicaciones no periódicas : Bakry F. & J.P. Horry. 1994. Musa breeding at CIRAD-FLHOR. Pp. 168175 in The Improvement and Testing of Musa : a Global Partnership (D.R. Jones, ed.). INIBAP, Montpellier, Francia. • Tablas : Las tablas deberán estar enumeradas consecutivamente y las referencias en el texto se harán de acuerdo a esta numeración. Cada tabla debe incluir un título. • Illustraciones : Las illustraciones deberán estar numeradas consecutivamente y las referencias en el texto se harán de acuerdo a esta numeración. Cada ilustración debe incluir un título sencillo y claro. Gráficos : suministrar los datos correspondientes junto con los gráficos. Dibujos : si es posible, suministrar dibujos originales. Fotografias a colores : suministrar pruebas de buena calidad y películas o diapositivas originales. Notas : Si el material de plantación utilizado para los experimentos descritos proviene o está registrado en el banco de germoplasma de INIBAP, debe indicarse su número de accesión (código ITC) dentro del texto o en forma tabular. Gracias por seguir nuestras recomendaciones. Esto facilitará y acelerará el trabajo de edición. 39 Publicaciones de INIBAP Disponibles en la sede central en Montpellier INIBAP 1999. Annual Report 1998. INIBAP 1999. K. Shepherd. Cytogenetics of the genus Musa. INIBAP 1999. D.A. Karamura. Numerical taxonomic studies of the East African highland bananas (Musa AAA-East Africa) in Uganda. INIBAP 1998. E. Akyeampong (ed.) Musa Network for West and Central Africa. Report of the first Steering Committee meeting held at Douala, Cameroon, 8-10 December 1998. INIBAP 1998. E.A. Frison y S.L. Sharrock (eds). Banana streak virus : a unique virus-Musa interaction ? Proceedings of a workshop of the PROMUSA virology working group held in Montpellier, France, 19-21 January 1998. INIBAP 1998. C. Picq (ed.). Segundo seminario/taller de la Red regional de información sobre banano y plátano de America Latina y el Caribe. San José, Costa Rica, 10-11 de Julio 1997. INIBAP 1998. B.K. Dadzie. Post-harvest characteristics of black Sigatoka resistant banana, cooking banana and plantain hybrids. INIBAP Technical Guidelines 4. INIBAP 1998. G. Orjeda en colaboración con los grupos de trabajo de PROMUSA sobre Sigatoka y Fusarium. Evaluación de los bananos para las enfermedades de Sigatoka y marchitamiento por Fusarium. Guías Técnicas INIBAP 3. CIRAD/INIBAP 1998. Les bananes. INIBAP/ACIAR 1997. E. Arnaud y J.-P. Horry (eds). Musalogue, a catalogue of Musa germplasm : Papua New Guinea collecting missions 1988-1989. INIBAP/FHIA/NRI/ODA 1997. B.K. Dadzie y J.E. Orchard. Evaluación rutinaria postcosecha de híbridos de bananos y plátanos : Criterios y métodos. Guías Técnicas INIBAP 2. INIBAP/CTA 1997. P.R. Speijer y D. De Waele. Tamizado de germoplasma de Musa para resistencia y tolerancia a los nematodos. Guías Técnicas INIBAP 1. (in press) INIBAP/The World Bank 1997. E.A. Frison, G. Orjeda y S. Sharrock (eds). PROMUSA : A Global Programme for Musa Improvement. Proceedings of a meeting held in Gosier, Guadeloupe, March 5 and 9 1997. INIBAP 1997. E. Arnaud (ed.). Bananos y Plátanos : Directorio de Investigadores. INIBAP-IPGRI/CIRAD 1996. Descriptores para el banano (Musa spp.). INIBAP 1996. C. Picq y R. Raymond (eds). Valorizar los bananos y los plátanos : el rol de la Red Internacional para el Mejoramiento del Banano y el Plátano. INIBAP/CIRAD/MARDI 1996. E. Frison, J-P Horry y D. De Waele (eds). New Frontiers in Resistance Breeding for Nematode, Fusarium and Sigatoka. Proceedings of a workshop held in Kuala Lumpur, Malaysia, 2-5 October 1995. Disponibles en la oficina regional de INIBAP-ASPNET INIBAP/ASPNET 1998. Minutes : Sixth meeting of INIBAP/ASPNET Regional Advisory Committee (RAC) hosted by the Vietnam Agricultural Science Institute (VASI) in Hanoi, Vietnam, 21-23 October 1997. INIBAP/ASPNET 1997. V. N. Roa y R. V. Valmayor (eds). Minutes : Sixth meeting of INIBAP/ASPNET Regional Advisory Committee (RAC) hosted by National Research Center on Banana (ICAR) in Tiruchirapalli, India, 26-28 September 1996. INIBAP/ASPNET 1996. R. V. Valmayor, V. N. Roa y V. F. Cabangbang (eds). Regional Information System for Banana and Plantain - Asia and the Pacific (RISBAP) : Proceedings of a consultation/workshop held at Los Baños, Philippines, 1-3 April 1996. (ASPNET Book Series N° 6). INIBAP/ASPNET 1996. V. N. Roa y R. V. Valmayor (eds). Minutes : Fifth meeting of INIBAP/ASPNET Regional Advisory Committee (RAC) hosted by MARDI in Serdang and Langkawi, Malaysia, 6-9 October 1995. INIBAP/ASPNET 1994. V. N. Roa y R. V. Valmayor (eds). Proceedings, Fourth Meeting of the Regional Advisory Committee, Asia and Pacific Network hosted by Taiwan Banana Research Institute in Chiuju, Pingtung, Taiwan, 21-25 November, 1994. INIBAP/ASPNET 1994. R. Valmayor et al. (eds). Banana Nematodes and Weevil Borers in Asia and the Pacific : Proceedings of a conference held in Sergang, Selangor, Malaysia, 18-22 April 1994. (ASPNET Book Series N° 5). INIBAP/ASPNET-TBRI 1993. R.V. Valmayor et al. (eds). Proceedings : International Symposium on Recent Development in Banana Cultivation Technology. Chiuju, Pingtung, Taiwan, 14-18 December 1992. (ASPNET Book Series N° 4). PROMUSA N° 3 Contenido Symposio Internacional sobre la biología molecular y celular del banano • Confección de mapas y genómica . . . . . . . . . . . . . . . .p. I • Bioquímica y maduración de la fruta . . . . . . . . . . . . . . .p. VIII PROMUSA Un programa global para el mejoramiento de Musa • Virología . . . . . . . . . . . . . . . . .p. XI • Cultivo de células y transformación del banano . .p. XII • Patología fungosa e interacciones huésped/ planta . . . . . . . . . . . . . . . . . .p. XIV • Algunas noticias de PROMUSA . . . . . . . . . . . .p. II à IV ¿ Qué es PROMUSA? El Programa Global para el Mejoramiento de Musa (PROMUSA) es un amplio programa que tiene el propósito de involucrar a todos los principales actores del mejoramiento de Musa. Este programa fue desarrollado como un medio de enlazar el trabajo realizado para resolver los problemas de los productores de bananos para la exportación, además de las iniciativas dirigidas al mejoramiento de la producción de bananos y plátanos a nivel de subsistencia y pequeña escala para los mercados locales. El programa global se construye sobre los logros existentes en la investigación y está basado en las iniciativas de las investigaciones en curso. Por lo tanto, PROMUSA es un mecanismo para maximizar los logros y acelerar el impacto de todo el esfuerzo mundial del mejoramiento en el área de Musa. El programa representa un mecanismo innovador que reúne investigaciones que se realizan tanto dentro, como fuera del CGIAR, creando nuevas asociaciones entre los Sistemas Nacionales de Investigación Agrícola (SNIA) e instituciones de investigación en países desarrollados y en vías de desarrollo. La formación de tales asociaciones también contribuirá a fortalecer la capacidad de los SNIA con respecto a la conducción de las investigaciones relacionadas con Musa. La principal meta de PROMUSA es desarrollar un amplio rango de variedades mejoradas de banano, a partir de las cuales los productores de todo el mundo puedan seleccionar aquellas que más responden a sus necesidades. El programa reúne el mejoramiento convencional basado en las técnicas de hibridación, con el mejoramiento mediante la ingeniería genética y la biotecnología. Este amplio esfuerzo de mejoramiento genético es apoyado por las investigaciones que se realizan sobre plagas y enfermedades específicas dentro de varios grupos de trabajo de PROMUSA. Un mecanismo eficaz para la evaluación de nuevas variedades, desarrollado en el marco de PROMUSA, también representa un componente esencial del programa. INFOMUSA — Vol 8, N° 1 Simposio internacional sobre la biología molecular y celular del banano Resúmenes de las comunicaciones Este simposio, organizado del 22 al 25 de marzo en Ithaca, NY, EEUU por el Boyce Thompson Institute for Plant Research, brindó una importante oportunidad para la presentación de resultados e intercambio de información sobre el estado de la biología molecular y celular del banano. En el marco de su contribución al simposio, INIBAP publica, debajo los resúmenes de las presentaciones en este simposio. Los organizadores del simposio agradecen a los patrocinadores : Agritope Inc., Boyce Thompson Institute for Plant Research Inc., DNA Plant Technology Corporation, Monsanto y Zeneca Plant Science. Confección de mapas y genómica El estudio de las Musáceas : confección de mapas y genómica Pierre Lagoda, F.C. Baurens, L.M. Raboin y J.L. Noyer CIRAD-AMIS Biotrop, B.P. 5035, F-34032 Montpellier Cedex 1, Francia El mejoramiento moderno de cultivos está basado en la selección con la ayuda de marcadores moleculares e introgresión de caracteres agronómicos de interés como la resistencia a plagas o la calidad. Se investigan muchos cultivos en el nivel molecular utilizando una mayor cantidad de sistemas de marcadores mejorados. Los cultivos tropicales como Musa son raramente incluidos, y si se incluyen en las iniciativas internacionales de análisis genómico para estudiar sus estructuras genéticas a veces muy complejas. Sin embargo, estos cultivos son de importancia para el mundo por razones socioeconómicas y ecológicas. Los bananos y plátanos pertenecen a la familia de las Musáceas y se encuentran entre las monocotiledóneas más altas. Los bananos de postre se cultivan extensivamente para su exportación a Europa (Occidental, Central y Oriental), China y América del Norte, mientras que los bananos de cocción y algunos bananos de postre muy a menudo se cultivan en los patios para el consumo local en los países tropicales (aunque estos cultivos también empiezan a exportarse). Los principales cultivares son triploides, altamente estériles, partenocárpicos y propagados por clones. Ellos son susceptibles a varias enfermedades que amenazan seriamente a las plantaciones. El estudio de las Musáceas, emocionantemente complejo a finales de 1800, se benefició en gran manera del sistema taxomorfológico de inventario desarrollado por los científicos británicos en un esfuerzo de 20 años con el fin de clasificar con coherencia especies silvestres y cultivares (Cheesman, Shepherd, Simmonds 1940-1960). Este sistema aún se mantiene bajo el fuego analítico de los marcadores moleculares modernos. Un fenómeno común en las accesiones de bananos es la variabilidad de la estructura de los cromosomas : • las translocaciones a nivel diploide acuminata, posiblemente el problema más importante en los proyectos de confección de mapas de los bananos, son responsables de las irregularidades durante la meiosis, • la actividad de (retro)transposones o metilationes puede ser responsable de la creación PROMUSA I de las plantas fuera de tipo (variantes) durante la propagación in vitro. Para entender mejor la genética del banano, se necesita obtener más conocimientos sobre la compleja estructura genómica y relaciónes filogenicas de los híbridos, cultivares y tipos silvestres. De este modo, se puede identificar a los ancestros diploides fértiles de los actuales cultivares poliploides. También es necesario obtener más conocimientos sobre los genes de interés que se encuentran en la biodiversidad de los bananos y la modulación de su expresión (transcriptoma). Esto ayudaría a desarrollar nuevas estrategias de mejoramiento basadas en hibridación con la ayuda de marcadores y transformación genética. Lo obligatorio para el manejo y monitoreo de estas estrategias es el cribado rápido y confiable de grandes cantidades de accesiones, lo que implica una estrecha cooperación entre los « centros de biotecnología » y « centros de biodiversidad » a nivel mundial. Bibliografía Cheesman E.E. 1947. Classification of the bananas. Kew Bull 2 : 97-117. Simmonds N.W. 1962. The Evolution of the Bananas. Longman Group UK Limited, London, 268 pp. Simmonds N.W. & K. Shepherd. 1955. The Taxonomy and origins of the cultivated bananas. Linnean Soc London 55 : 302-312. Baurens F.C., J.L. Noyer, C. Lanaud & P.J.L. Lagoda. 1996. Use of competitive PCR to assay copy number of repetitive elements in banana. MGG 253 : 57-64 Baurens F.C., J.L. Noyer, C. Lanaud & P.J.L. Lagoda. 1997. Assessment of a repetitive DNA family Brep 1 in Musa acuminata. Theo. Appl. Gene. 95 : 922931. Baurens F.C., J.L. Noyer, C. Lanaud & P.J.L. Lagoda. 1998. Inter-Alu like PCR profiling of banana. Euphytica 99 : 137-142. Baurens F.C., J.L. Noyer, C. Lanaud & P.J.L. Lagoda. 1997. Copia-like elements in banana. J. Genet. Breed. 51 : 135-142. Baurens F.C., J.L. Noyer, C. Lanaud & P.J.L. Lagoda. 1997. STS markers to draft the banana chloroplast genome. Fruits 52 : 247-259. Carreel F., S. Fauré, D. González de León, P.J.L. Lagoda, X. Perrier, F. Bakry, H. Tezenas du Montcel, C. Lanaud & J.P. Horry. 1994. Evaluation de la diversité génétique chez les bananiers diploïdes (Musa sp.). Génétique sélection évolution 26(S1) : 125s-136s. Escalant J.V., F.X. Côte, A. Grapin, E. Auboiron, T. Legavre, P.J.L. Lagoda, F. Carreel, F. Bakry, J.P. Horry, F. Kerbellec, C. Teisson & H. Tezenas du Montcel. 1994. Biotechnologies for Musa improvement : strategies and results. Pp. 22-26 in Banana and Plantain Breeding : Priorities and Strategies. INIBAP, Montpellier, France. Fauré S., J.L. Noyer, J.P. Horry, F. Bakry, C. Lanaud & D. Gonzales de León. 1993. A molecular-based II PROMUSA linkage map of diploid bananas (Musa acuminata). Theor. Appl. Genet. 87 : 517-626. Ford-Lloyd B.V., D. Kaemmer, G. Kahl & P.J.L. Lagoda. 1997. Molecular marker assisted analysis of the Musa genome complex. Pp. 24-28 in Networking Banana and Plantain : INIBAP Annual Report 1996. INIBAP, Montpellier, France. Grapin A., J.L. Noyer, F. Carreel, D. Dambier, F.C. Baurens, C. Lanaud & P.J.L. Lagoda. 1998. Diploid Musa acuminata genetic diversity assayed with Sequence Tagged Microsatellite Sites. Electrophoresis 19 :1374-1380 Kaemmer D., D. Fischer, R.L. Jarret, F.C. Baurens, A. Grapin, D. Dambier, J.L. Noyer, C. Lanaud, G. Kahl & P.J.L. Lagoda. 1997. Molecular breeding in the genus Musa : a strong case for STMS marker technology. Euphytica 96 : 49-63. Lagoda P.J.L. & J.L. Noyer.1994. Update on Musa genome mapping at CIRAD-AGETROP. INFOMUSA 3(2): 4 Lagoda P.J.L., D. Dambier, A. Grapin, F.C. Baurens, C. Lanaud & J.L. Noyer. 1998. Non-radioactive sequence tagged microsatellite site analyses : a method transferable to the tropics. Electrophoresis 19 : 152-157. Confección de mapas del genoma y análisis genético de la resistencia a la Sigatoka negra en el banano F. Carreel1, C. Abadie2, J. Carlier3, K. Tomekpe2, P.J.L. Lagoda3 y Frédéric Bakry3 1 CIRAD, Neufchâteau, 97130 Capesterre BE, Guadalupe, Francia 2 CRBP, BP 832, Douala, Camerún 3 CIRAD, BP 5035, 34032 Montpellier, Francia La producción bananera está seriamente amenazada por la enfermedad de la Sigatoka negra causada por Mycosphaerella fijiensis. Se necesitan numerosas aplicaciones de químicos para controlar el impacto del hongo sobre las hojas lo que reduce los rendimientos y causa maduración prematura de la fruta. El mejoramiento genético para desarrollar la resistencia a la Sigatoka negra parece ser el método de control más duradero, eficaz desde el punto de vista de los costos e impacto ambiental. El cribado para detectar la resistencia llevó a la identificación de dos formas de resistencia reportadas como resistencia alta (RA, respuesta de hipersensibilidad) y resistencia parcial (RP). En conexión con el programa de mejoramiento, se desarrolló una guía para la confección del mapa del genoma con el fin de realizar análisis genéticos y de QTL para caracterizar la forma RA. Se obtuvo una población segregada F2 de 153 plantas mediante un cruzamiento entre dos accesiones de banano silvestre seminífero, Musa acuminata burmannicoides tipo Calcutta 4 y M. a. banksii tipo Madang. Se construyó un mapa de enlace me- Algunas noticias de PROMUSA Una reunión del subgrupo de Cartografía genética del grupo de trabajo sobre el Mejoramiento genético es el principal evento que tuvo lugar en el marco de PROMUSA desde la última reunión global. Esta reunión fue celebrada en ocasión del Simposio internacional sobre la biología molecular y celular del banano en la Universidad Cornell, EEUU. Subgrupo de Cartografía genética A la reunión del subgrupo de Cartografía genética asistieron científicos de varias compañías del sector privado. Esta fue la primera vez que el sector privado ha sido representado en una reunión de PROMUSA y por lo tanto, las discusiones se enfocaron sobre el papel que PROMUSA podría desempeñar en el desarrollo de las asociaciones entre los sectores público y privado. Se reconoció que PROMUSA podría proporcionar un foro adecuado para este tipo de colaboración, particularmente, en el área de la investigación precompetitiva. Se acordó que INIBAP dirigirá el seguimiento de los contactos con las compañías del sector privado. Inicialmente, el enfoque se concentrará en determinar cómo el sector privado podría contribuir a los propósitos y objetivos de PROMUSA e identificar los mecanismos para facilitar estas interacciones. Taller de trabajo sobre la genómica de Musa Después del simposio en Cornell y de las discusiones del grupo de trabajo en Mejoramiento genético de PROMUSA, se hizo claro que en la investigación genética de Musa se están haciendo avances rápidos. Varios proyectos de investigación en curso han permitido construir mapas genéticos, clonar genes, realizar ensayos de expresión, examinar los promotores y transferir construcciones de genes en los cultivares. Sin embargo, se acordó que habría un mayor beneficio en unir INFOMUSA — Vol 8, N° 1 varios equipos de investigación para poder obtener un mejor entendimiento del genoma de Musa. La creación de herramientas para estudiar el genoma, la propia transcripción, harán una contribución mayor al rápido progreso en la investigación de mejoramiento de Musa en el siguiente milenio. Se decidió que se podría lograr mejor progreso a través del desarrollo de una iniciativa sobre la genómica de Musa. Esta iniciativa podría empezar con un enfoque sobre la ‘cartografía’, incluyendo la genética, citogenética, mapas físicos y de expresión del transcriptoma. Sin embargo, se necesitarán los aportes de todas las áreas de la biología y agronomía de banano, así como nuevas habilidades científicas (bioinformática) para crear y afinar las herramientas (bibliotecas BAC, EST, bibliotecas cADN, juegos de diagnóstico de alto rendimiento, aparatos micro/macro, chips de ADN). Las herramientas desarrolladas de este modo permitirán resolver problemas específicos, como la localización de los puntos de translocación, análisis de los genes objetivos, clonación de genes basada en la cartografía, silenciamiento transgénico, etc. Como un primer paso en esta iniciativa de la genómica, INIBAP organizará un taller a en el marco de PROMUSA, en Montpellier en enero de 2000. Este taller estará abierto a cualquier persona que participe activamente en la investigación de la genómica de Musa, de los sectores público y privado. Para obtener más información sobre este taller, por favor dirígase a Jean-Vincent Escalant, INIBAP, Montpellier, Francia. Email: [email protected] Grupo de trabajo en Sigatoka La información sobre la evaluación de una cantidad de variedades de Musa con respecto a su resistencia a la Sigatoka amarilla por parte de los investigadores del National Research Centre on Bananas (NRCB) fue publicada en la sección MusaNews del último número de INFOMUSA (Vol. 7(2), p. 43). Posteriormente, se observó que los resultados de los ensayos de cribado presentados parecían estar en conflicto con los de otros científicos que han trabajado con INFOMUSA — Vol 8, N° 1 diante 110 marcadores AFLP en asociación con 39 marcadores RFLP co-dominantes y marcadores SSR para anclar los grupos de enlace al mapa genético principal de banano. Los resultados mostraron segregaciones alélicas distorsionadas significativas en la progenie F2 para 58 marcadores que destaca el uso de los estudios moleculares para los análisis de herencia. El primer mapa concluyó clasificando los marcadores en 11 grupos de enlace. Las correlaciones realizadas con la ayuda de las observaciones en el campo, que determinaron 6 clases de la severidad de la infección, llevaron a la identificación de un marcador RFLP asociado fuertemente con la resistencia y un segundo QTL colocado en un grupo de enlace diferente con un nivel significativo más bajo. Bibliografía De Langhe E.A.L. 1969. Bananas, Musa spp. Pp. 5378 in Outlines of perennial crops breeding in the tropics (F.P. Ferwerda and F. Wit, eds). Landbouwhogeschool Wageningen, The Netherlands. Fauré S., J.L. Noyer, J.P. Horry, F. Bakry, C. Lanaud & D. Gonzalez de León. 1993. A molecular-based linkage map of diploid bananas (Musa acuminata). Theor. Appl. Genet. 87 : 517-626. Fauré S., J.L. Noyer, F. Carreel, J.P. Horry, F. Bakry & C. Lanaud. 1994. Maternal inheritance of chloroplast genome and paternal inheritance of mitochondrial genome in bananas (Musa acuminata). Curr. Genet. 25 : 265-269. Lagoda P.J.L. & J.L. Noyer. 1994. Update on Musa genome mapping at CIRAD-AGETROP. INFOMUSA 3(2): 4 Lagoda P.J.L., C. Amson, G. Aubert, E. Auboiron, F. Bakry, F.C. Baurens, F. Carreel, L. Costet, D. Dambier, S. Fauré, E. Fouré, J. Ganry, D. Gonzalez de León, A. Grapin, J.P. Horry, C. Jenny, J.L. Noyer, P. Ollitrault, L.M. Raboin, F. Reschoeur, H. Tézenas du Montcel, K. Tomekpe & C. Lanaud. 1998. Acronyms to study the Musaceae. Acta Horticulturae 460/461 : 113-121 Lagoda P.J.L., J.L. Noyer, D. Dambier, F.C. Baurens, A. Grapin & C. Lanaud. 1998. Sequence tagged microsatellite site (STMS) markers in the Musaceae. Molecular Ecology 7 : 657-666 Análisis del genoma de Musa utilizando la citometría de flujo y citogenética molecular v Jaroslav Dolezel, M.A. Lysak, M. Dolezelova y M. Valarik Institute of Experimental Botany, Laboratory of Molecular Cytogenetics and Cytometry, Sokolovska 6, CZ-77200 Olomouc, República Checa - E-mail : [email protected] La citogenética de Musa ha sido poco desarrollada debido principalmente a las dificultades con el análisis de pequeños cromosomas. Hasta la fecha, no se conoce la ploidia de muchas accesiones, la identificación de cromosomas individuales dentro del cariótipo no es posible. Esta situación contrasta con un rápido progreso hecho en el análisis del genoma de Musa a nivel molecular. Sin embargo, la aplicación de la citometría de flujo y citogenética molecular está cambiando el mundo de la cariología de Musa. Este trabajo evalúa los progresos alcanzado en esta área en nuestro laboratorio. Se demostró que la citometría de flujo es adecuada para el análisis del contenido del ADN nuclear en Musa (Dolezel et al. 1994, 1997). El método está basado en la preparación de suspensiones de núcleos intactos de pequeñas cantidades de tejidos frescos. Luego el ADN de los núcleos en suspensión se tiñe por un fluorocromo adecuado y la fluorescencia de los núcleos individuales se analiza utilizando un citómetro de flujo. El análisis puede ser realizado a alta velocidad y miles de núcleos pueden ser medidos dentro de pocos minutos. Debido a que los núcleos son analizados individualmente, se puede detectar sub-poblaciones que se diferencian por el contenido de ADN (por ejemplo, mixoploidia). El método es rápido, no destructivo y requiere sóloamente pequeñas cantidades de tejido foliar, y de este modo encuentra un gran número de aplicaciones tanto en taxonomía, como en mejoramiento. La citometría de flujo ha sido utilizada para clasificar nuevamente la ploidia de varios clones bien conocidos los cuales anteriormente se consideraban tetraploides. Resultó que tres de los cuatro clones fueron triploides (Horry et al. 1998). Este resultado inesperado indica que los cultivares tetraploides naturales acuminata x balbisiana son aún más raros de lo que generalmente se creía. Más, demuestra que no se puede confiar en la morfología de la planta solamente para determinar su constitución genómica. Actualmente se dispone de métodos nuevos o mejorados, que incluyen el conteo de cromosomas y la citometría de flujo, y de este modo la ploidia de cualquier accesión puede ser determinada rápidamente y con suficiente confianza. cada vez más el método también está siendo utilizado en los programas de mejoramiento para cribar la ploidia en progenies obtenidas en diferentes cruzamientos. Además, se ha desarrollado un procedimiento integrado que implica una poliploidización in vitro y el cribado de la ploidia mediante citometría de flujo para la producción en masa de los poliploides no quiméricos (Van Duren et al. 1996). También se demostró que la citometría de flujo es adecuada para la estimación del tamaño del genoma. Los resultados del análisis del PROMUSA III contenido de ADN demostraron que los genomas de Musa son pequeños (~ 537 - 615 Mbp) y que los genomas A y B de Musa difieren en tamaño, siendo el genoma B más pequeño en un 12 % promedio (Dolezel et al. 1994, Lysak et al. 1999). No se encontraron variaciones en el tamaño del genoma entre las accesiones de M. balbisiana. Una pequeña pero significativa variación fue encontrada entre las subespecies y clones de M. acuminata. Esta diferencia puede estar relacionada con el origen geográfico de accesiones individuales. La variación más grande en el tamaño del genoma fue encontrada entre los clones triploides de Musa. La variación puede ocurrir debido tanto a la diferente constitución genómica, como a las diferencias en el tamaño de sus genomas A. El análisis estadístico sobre el tamaño del genoma dio los datos agrupadoslo que está muyrelevante con la clasificación taxonómica de Musa aceptada generalmente (Lysak et al. 1999). Basándose en estos resultados, se propuso que un análisis comparativo del tamaño del genoma en diploides y triploides podría ayudar a identificar progenitores diploides putativos de los clones triploides de Musa cultivados. En especies de algunas plantas, la citometría de flujo demostró ser útil para diferenciar tipos de cromosomas individuales que se utilizaron como fuente especifica de ADN cromosómicos (Dolezel et al. 1995). Una vez desarrollada para Musa, la técnica simplificaría el aislamiento de los genes y la confección de su mapa. Aunque la citometría de flujo demostró ser extremadamente útil, es imposible confeccionar mapas genómicos detallados sin el análisis de la estructura y el comportamiento de los cromosomas durante la mitosis y meiosis. Hasta hace poco esto era difícil debido al pequeño tamaño de los cromosomas de Musa. Se desarrolló un nuevo método para la preparación de metafase en placas adecuadas para el análisis de cromosomas de alta resolución (Dolezel et al. 1998). El método permite un conteo confiable de cromosomas y el análisis de la morfología básica de los cromosomas. Además, las metafases en placas resultaron adecuadas para la localización de las secuencias de ADN en los cromosomas mitóticos utilizando métodos de citogenética molecular. Mientras que la hibridación genómica in situ (GISH) ha sido utilizada para determinar el origen parental de los cromosomas en los híbridos, la hibridación fluorescente in situ (FISH) permite confeccionar mapas físicos de las secuencias de ADN hacia los cromosomas de Musa. La hibridación FISH con sondas para los genes rARN en las especies diploides seminíferas silvestres M. acuminata y M. balbisiana mostró que las secuencias de rADN 18S-25S están localizadas sólo en un par de cromosomas y que los sitios de rADN 5S están localizados en dos o tres pares IV PROMUSA de cromosomas. Es interesante notar que sólo se observaron cinco sitios de rADNA 5S en las variedades comestibles diploides propagadas vegetativamente, lo que indica su origen híbrido (Dolezelova et al. 1998). La identificación de cromosomas individuales utilizando secuencias de ADN en los mapas físicos permitirá realizar el análisis de su comportamiento y segregación durante los programas de evolución y mejoramiento. Además, las secuencias ADN de una sola copia o de pocas copias cartografiadas físicamente proporcionarán sitios de anclaje para integrar los mapas físicos y genéticos. Para concluir, la aplicación de la citometría de flujo y citogenética molecular estimularon un resurgimiento del interés en la cariología de Musa y expandieron significativamente el conocimiento del genoma de Musa. Se puede esperar que estos métodos se utilizarán con mayor frecuencia en la genética y mejoramiento de Musa. Agradecimiento Este estudio fue realizado como parte del Programa Global para el Mejoramiento de Musa (PROMUSA) y apoyado por el contrato de investigación No. 8145/RB de la Agencia Internacional para la Energía Atómica (International Atomic Energy Agency), Viena, Austria. Bibliografía la Sigatoka amarilla. Por ejemplo, algunas accesiones de los cultivares Pisang Awak, Bluggoe, Mysore y Ney Poovan, a los cuales normalmente se refiere como cultivares resistentes a la Sigatoka amarilla, fueron considerados susceptibles. En vista de la identificación de una ‘nueva’ enfermedad de la mancha foliar causada por Mycosphaerella/Septoria en India, tal como informó el grupo de trabajo sobre la Sigatoka en PROMUSA No. 2 (INFOMUSA Vol. 7(2), p. X), actualmente se realizan esfuerzos para confirmar la identidad de los patógenos presentes en este estudio. Está claro que se necesita más información sobre los hongos que causan las enfermedades de la mancha foliar en Asia, y especialmente sobre las reacciones a estas de los nuevos híbridos resistentes a la Sigatoka negra que se desarrollan por los programas de mejoramiento. Por lo tanto se está desarrollando una propuesta para un proyecto colaborativo sobre el hongo Mycosphaerella/Septoria en el marco del grupo de trabajo sobre la Sigatoka. v Dolezel J., M. Dolezelova & F.J. Novak. 1994. Flow cytometric estimation of nuclear DNA amount in diploid bananas (Musa acuminata and M. balbisiana). Biol. Plant. 36 : 351-357. v Dolezel J., M. Dolezelova, N. Roux & I. Van den Houwe. 1998. A novel method to prepare slides for high resolution chromosome studies in Musa spp. INFOMUSA 7: 3-4. v Dolezel J., S. Lucretti & J. Macas. 1995. Flow cytometric analysis and sorting of plant chromosomes. Pp. 185-200 in Kew Chromosome Conference IV (P. Brandham and M.D. Bennett, eds.). Royal Botanic Gardens, Kew, 1995. v Dolezel J., M.A. Lysak, I. Van den Houwe, M. Dolezelova & N. Roux. 1997. Use of flow cytometry for rapid ploidy determination in Musa species. INFOMUSA 6: 6-9. Dolezelova M., M. Valarik, R. Swennen, J.P. Horry & J. Dolezel. 1998. Physical mapping of the 18S-25S and 5S ribosomal RNA genes in diploid bananas. Biol. Plant. 41 : 497-505. Horry J.P., J. Dolezel, J., M. Dolezelova & M.A. Lysak. 1998. ¿ Existen tetraploides AxB naturales ? INFOMUSA 7: 5-6. Lysak M.A., M. Dolezelova, J.P. Horry, R. Swennen & J. Dolezel. 1999. Flow cytometric analysis of nuclear DNA content in Musa. Theor. Appl. Genet. (in press). Van Duren M., R. Morpurgo, J. Dolezel & R. Afza. 1996. Induction and verification of autotetraploids in diploid banana (Musa acuminata) by in vitro techniques. Euphytica 88 : 25-34. Grupo de trabajo sobre Fusarium El grupo de trabajo sobre Fusarium está organizando su siguiente reunión que se celebrará en Malasia, los días 21-22 de octubre, en la ocasión del seminario y taller de trabajo sobre “El banano y el manejo del marchitamiento por Fusarium: hacia un cultivo sostenible” que organizan MARDI e INIBAP (Ver el panfleto incluido en este número). Los miembros del grupo de trabajo presentarán resultados de sus investigaciones durante el seminario y el taller, permitiendo de este modo a otros participantes, que no son miembros del grupo de trabajo, también beneficiarse con esta información. La reunión del grupo de trabajo se enfocará sobre la identificación de las prioridades para las futuras investigaciones y desarrollo de proyectos colaborativos. Para obtener más información, dirigirse a la Dra Gisella Orjeda, INIBAP, Montpellier, Francia. Email: [email protected] INFOMUSA — Vol 8, N° 1 Investigación de la estructura genómica compleja del banano cultivado (Musa spp.) mediante citometría de flujo, hibridación de ADN genómico in situ y análisis de secuencias repetidas. Angélique D’Hont, A. Paget-Goy, C. Jenny, J.L. Noyer, F.-C. Baurens, P. Lagoda y F. Carreel. CIRAD, BP 5035, 34032 Montpellier Cedex 1, Francia y CIRAD, Neufchâteau, Sainte Marie, 97130 Capesterre B/E, Francia. Los genomas de cuatro diferentes especies silvestres de Musa se encuentran en los bananos cultivados : los genomas A, B, S y T relacionados con M. acuminata, M. balbisiana, M. schizocarpa y especies de Australimusa, respectivamente. La mayoría de los clones cultivados son triploides o diploides. Ellos han sido clasificados en grupos genómicos de acuerdo a la cantidad de cromosomas y caracteres morfológicos. Los principales grupos son AA, AAA, AAB y ABB. La citometría de flujo revela que los tamaños de los genomas A, B, S y T difieren significativamente. M. balbisiana tiene el genoma más pequeño (1.03 pg/2C, 2n = 2x = 22), seguida por M. acuminata (1.11 pg/2C, 2n = 2x = 22) y M. schizocarpa (1.18 pg/2C, 2n = 2x = 22). El valor más alto se obtuvo para las especies M. textilis de Australimusa (1.27 pg/2C, 2n = 2x = 20). De este modo, el tamaño del genoma puede ser utilizado para determinar las especies eimplicadas en los clones diploides interespecíficos así como la ploidia de los clones de banano. La hibridación genómica in situ (GISH) a menudo permite visualizar la porción del genoma aportada por cada especie progenitora en los híbridos interespecíficos (D’Hont et al. 1995, 1996). De esta manera, se desarrolló esta técnica para caracterizar el origen específico de los cromosomas en los clones interespecíficos de banano (D’Hont et al. 1999). Los cromosomas de los cuatro genomas A, B, S y T fueron diferenciados mediante la GISH. Sin embargo, una porción distal de los cromosomas permaneció sin etiquetar, tal como fue reportado acerca de las especies con genomas pequeños (Barre et al. 1997). La GISH fue utilizada para determinar la estructura exacta del genoma de varios clones cultivados interespecíficos. Los resultados en su mayoría fueron consistentes con la constitución cromosómica estimada a través de descriptores fenotípicos con una excepción notable, el clon ‘Pelipita’, que abarca cromosomas 8A y 25 B en vez de la 11A y 22 B pronosticados. La técnica también permitió determinar los cromosomas adicionales de unos pocos clones sin clasificar sobre la única base de descriptores fenotípicos y conteos de INFOMUSA — Vol 8, N° 1 cromosomas. Se supone que la GISH se apoya esencialmente en la detección de las secuencias repetidas, cuyas cantidades constituyen la principal fuente de variación en el tamaño básico del genoma en las plantas. Hemos mostrado que la complejidad del genoma de Musa spp. en un 25 % es altamente repetitiva y en un 50 % es medio repetitiva y hemos aislado varias secuencias repetidas (tipo Copia [Baurens et al. 1997a], tipo Alu [Baurens et al. 1998], Brep1 [Baurens et al. 1997b]). Como un ejemplo, la cantidad de copias repetitivas especificas de los genomas A y S de la familia Brep1 varía de 0 copias en los genomas B y T a más de 6 000 en M.a. burmannicoides (1 % del genoma) y varía en un orden de magnitud aproximadamente entre M.a. banksii y M.a. malaccensis (Baurens et al. 1996). Hemos mostrado la presencia de un locus de rADN 18S-25S por cada genoma básico en las especies de Eumusa estudiadas. Este locus podría ser cartografiado sobre el mapa genético disponible (Fauré et al. 1993) para tener un primer puente entre los mapas físicos y genéticos. El desarrollo de puentes similares (secuencias repetidas, BAC a.o.) para varios grupos de enlace permitirá asignar los diferentes cromosomas a los grupos de enlace respectivos y complementará con eficacia los esfuerzos en la cartografía genética. Bibliografía D’Hont A, P. Rao, P. Feldmann, L. Grivet, N. IslamFaridi, P. Taylor & J.C. Glaszmann. 1995. Identification and characterisation of intergeneric hybrids, S. officinarum X Erianthus arundinaceus, with molecular markers and in situ hybridization. Theor. Appl. Genet. 91 : 320-326. D’Hont A., L. Grivet, P. Feldmann, P.Rao, N. Berding & J.C. Glaszmann. 1996. Characterisation of the double genome structure of modern sugarcane cultivars (Saccharum spp.) by molecular cytogenetics. Mol. Gen. Genet. 250 : 405-413. D’Hont A., A. Paget-Goy, J. Escoute & F. Carreel. 1999. The interspecific genome structure of cultivated banana, Musa spp. revealed by genomic DNA in situ hybridization. Theor. Appl. Genet. (in press). Barre P., M. Layssac, A. D’Hont, J. Louarn, A. Charrier, S. Hamon & M. Noirot. 1997. Relationship between parental chromosomic contribution and nuclear DNA content in the coffe interspecific hybrid : C. pseudozanguebariae x C. liberica var. dewevrei. Theor. Appl. Genet. 96 : 301-305. Baurens F.C., J.L. Noyer, C. Lanaud & P. Lagoda. 1997a. Copia-like elements in banana. J Genet Breed 51 : 135-142. Baurens F.C., J.L. Noyer, C. Lanaud & P. Lagoda. 1998. Inter-Alu like PCR profiling of banana. Euphytica 99 : 137-142. Baurens F.C., J.L. Noyer, C. Lanaud & P. Lagoda. 1997b. Assessment of a repetitive DNA family Brep 1 in Musa acuminata. Theor. Appl. Genet. 95 : 922931. Baurens F.C., J.L. Noyer, C. Lanaud & P. Lagoda. 1996. Use of competitive PCR to assay copy number of repetitive elements in banana. . Mol. Gen. Genet. 253 : 57-64. Faure S., J.L. Noyer, J.P. Horry, F. Bakry, C. Lanaud & D. Gonzalez de León. 1993. A molecular markerbased linkage map of diploid bananas (Musa acuminata). Theor. Appl. Genet. 87(4): 517-526. Utilización del AFLP fluorescente para detectar enanismo. Caso del banano ‘Curare enano’ (Musa spp.). Iris Engelborghs, S. Remy y R. Swennen Laboratory of Tropical Crop Improvement, Catholic University of Leuven, Kardinaal Mercierlaan 92, 3001 Heverlee, Bélgica Email : [email protected] El ‘polimorfismo de la longitud de fragmentos amplificados’ (amplified fragment length polymorphism) o la técnica AFLP fue optimizada para el banano (Musa spp.). Esta técnica, basada en la amplificación selectiva de los fragmentos de restricción sin un conocimiento previo de secuencias permite la detección de muchos polimorfismos de una manera bastante rápida. Actualmente, las plantas de banano se propagan in vitro aunque la ocurrencia frecuente de variantes somaclonales permanece siendo un serio ‘cuello de botella’. La detección temprana de estos tipos anormales con una técnica de impresión de huellas moleculares como lo es el AFLP, es necesaria antes de transplantar en el campo las plántulas obtenidas mediante la técnica in vitro. Las técnicas de impresión de huellas moleculares se utilizan comúnmente para caracterizar genomas (Becker et al. 1995, Foisset et al. 1996), definir relaciones taxonómicas (Caetano-Anollés et al. 1995, Ghareyazie et al. 1995) y evaluar la diversidad genética (Virk et al. 1995, Hongtrakul et al. 1997, Hart y Seefelder 1998). Lagoda et al. (1998) revisaron los estudios de diversidad genética en Musa basándose en isoenzimas, RFLP, RAPD, ADN microsatélite, marcadores SST (‘sitio específico’ o ‘sequence tagged site’). La técnica de ‘polimorfismo de la longitud de fragmentos amplificados’ (AFLP) (Zabeau y Vos 1993) recientemente desarrollada es una nueva técnica poderosa que todavía no se aplica de manera general a Musa. Con este método, los juegos de fragmentos de restricción son visualizados mediante PCR sin conocimiento previo de la secuencia de nucleotidos. El procedimiento de AFLP (Zabeau y Vos 1993) fue adaptado para Musa spp. basándose en el protocolo de Vos y colaboradores (1995). PROMUSA V Las principales modificaciones se refieren a los siguientes pasos : omisión de la selección con magnetos, sin un real “touchdown” PCR, uso de un secuenciador automatizado para la separación de los fragmentos y la detección fluorescente de los fragmentos en vez del marcado radioactivo. El uso del marcado fluorescente no solo lo hace más fácil de manejar en comparación con una detección basada en la radioactividad, sino también brinda la posibilidad de detección y análisis computarizados de los fragmentos separados lo que reduce el tiempo y los esfuerzos al manejar muchas muestras con perfiles AFLP complejos. Este estudio está enfocado a los tipos enanos de banano, los cuales son los variantes somaclonales más importantes para Musa (Reuveni 1986, Sandoval et al. 1989, Smith y Drew 1990, Côte et al. 1992). Estas plantas de estatura más baja, con un tallo más grueso y con las hojas más anchas y cortas, en contraste con los tipos enanos que ocurren en la naturaleza tienen un racimo más pequeño. A menudo la variación sólo es visible después de que las plántulas cultivadas in vitro se siembran en un invernadero o en el campo, algunas veces después de la floración (Vuylsteke et al. 1996). Se ahorrarían tiempo y recursos al detectar muy temprano los variantes somaclonales. Los perfiles de AFLP de los tipos enanos de ‘Curare enano’ que ocurren en la naturaleza (verdaderos) fueron comparados con los tipos anormales de tamaño normal. El ADN pre-amplificado fue utilizado como medida para un paso de amplificación selectiva con 4 a 5 nucleotidos selectivos. Dependiendo de la combinación de los iniciadores (28 examinados) se detectó hasta un 10 % de polimorfismos entre los tipos enanos y normales de banano. Bibliografía Becker J., P. Vos, M. Kuiper, F. Salamini & M. Heun. 1995. Combined mapping of AFLP and RFLP markers in barley. Mol. Gen. Genet. 249 : 65-73. Caetano-Anollés G., L.M. Callahan & P.E. Williams. 1995. DNA amplification fingerprinting analysis of bermudagrass (Cynodon) : genetic relationships between species and interspecific crosses. Theor. Appl. Genet. 91 : 228-235. Côte F.X., X. Perrier & C. Teisson. 1992. Somaclonal variation in Musa spp. : theoretical risks, risk management, future research prospects. Pp. 192199 in Biotechnology applications for banana and plantain improvement. INIBAP, Montpellier, France. Engelborghs I., R. Swennen & S. Van Campenhout. 1998. Capacidad del AFLP para detectar diferencias genéticas y variantes somaclonales en Musa spp. INFOMUSA 7(2): 3-6. Foisset N., R. Delourme, P. Barret, N. Hubert, B. Landry & M. Renard. 1996. Molecular-mapping analysis in Brassica napus using isozyme, RAPD VI PROMUSA and RFLP analysis on a double-haploid progeny. Theor. Appl. Genet. 93 : 1017-1025. Ghareyazie B., N. Huang, G. Second, J. Bennett & G.S. Khush. 1995. Classification of rice germplasm. I. Analysis using AFLP and PCR-based RFLP. Theor. Appl. Genet. 91 : 218-227. Hart L. & S. Seefelder. 1998. Diversity of selected hop cultivars detected by fluorescent AFLPs. Theor. Appl. Genet. 96(1): 112-116. Hongtrakul V., G. Heustis & S. Knapp. 1997. Amplified fragment length polymorphisms as a tool for DNA fingerprinting sunflower germplasm : genetic diversity among oilseed inbred lines. Theor. Appl. Genet. 95 : 400-407. Lagoda P.J.L., C. Amson, G. Aubert, E. Auboiron, F. Bakry, F.C. Baurens, F. Carreel, L. Costet, D. Dambier, S. Fauré, E. Fouré, J. Ganry, D. Gonzales de Léon, A. Grapin, J.P. Horry, C. Jenny, J.L. Noyer, P. Ollitrault, L.M. Raboin, F. Reschoeur, H. Tézenas du Montcel, K. Tomepke & C. Lanaud. 1998. Acronyms to study the Musaceae. Pp. 113121 in Proceedings of the International Symposium on Biotechnology of Tropical and Subtropical Species. Part 2. (R.A. Drew, ed.). Brisbane, Queensland, Australia, ISHS 29. Virk P.S., B.V. Ford-Lloyd, M.T. Jackson & H.J. Newbury. 1995. Use of RAPD for the study of diversity within plant germplasm collections. Heredity 2 :170-179. Sandoval J., A. Tapia, L. Muller & E. Villalobos. 1989. Observations about the variability found in micropropagated plants of Musa cv. ‘Curare’ (AAB). in Abstracts of the IX ACORBAT Conference, Merida, Venezuela, 25-29 September, 1989. Smith M.K. & R.A. Drew. 1990. Growth and yield characteristics of dwarf off-types recovered from tissue-cultured bananas. Austr. J. Exp. Agr. 30 : 575-578. Vuylsteke D., R. Swennen & E.A.L. De Langhe. 1996. Field performance of somaclonal variants of plantain (Musa spp., AAB group). J. Amer. Soc. Hort. Sci. 121(1): 42-46. Vos P., R. Hogers, M. Bleeker, M. Reijans & T. Van der Lee. 1995. AFLP : a new technique for DNA fingerprinting. Nucleic Acids Research 23 : 44074414. Zabeau M. & P. Vos. 1993. European Patent Application, publication No. EP0534858. Relación de los elementos retrotransposables con la mutación ‘hojas largas y angostas’ de Musa Eli Khayat1, S. Regev2, A. Duvdevano1 y M. Amira1 1 2 Rahan Meristem, Kibbutz Rosh Hanikra, 22825 Israel Technion, Dept of Biology, Technion City, Haifa 3200 Israel Hemos identificado y aislado a partir de Musa un retrotransposón activo de tipo Ty-Copia utilizando iniciadores PCR degenerados to- mados del retrotransposón Tos-17’ que fue aislado a partir de arroz (Hiroshika et al. 1996). La activación transcripcional del retroelemento fue inducida por una prolongada permanencia de las plántulas de ‘Grande Naine’ en cultivo de tejidos. Para determinar la relación de estos resultados con variantes somaclonales, hemos analizado la amplificación del retroelemento en el genoma de varios mutantes, insensibles y muy sensible a las giberelinas. Se detectaron señales de amplificación y fuerte hibridación en el genoma del mutante ‘Hojas largas y angostas’ (HLA). El mutante HLA que puede ser detectado durante el cultivo in vitro, tiene hojas distintivas, largas y angostas, y los internodos más largos que la línea parental. Nuestros datos de hibridación revelan que el homólogo Tos-17’ del banano fue amplificado en las regiones cromáticas específicas en la HLA. Debido a la gran cantidad de regiones codificadoras y la naturaleza triploide del genoma de Musa, es probable que los retroelementos desciendan en una secuencia no funcional. Por otro lado, si los variantes somaclonales encontrados con frecuencia derivan de la activación transcripcional de los retroelementos, este hecho apoyará la hipótesis de inserciones no aleatorias en el genoma. Perspectivas de los mejoradores con respecto a la biotecnología para el mejoramiento de Musa Dirk Vuylsteke, J. Hartman y A. Tenkouano IITA, Ibadan, Nigeria, c/o Lambourn & Co, 26 Dingwall Road, Croydon CR9 3EE, Reino Unido El banano y el plátano (Musa spp L.) representan el principal cultivo y producto alimentario tropical a nivel mundial. Siendo un cultivo poliploide de propagación vegetativa, su mejoramiento genético mediante hibridación convencional es difícil, si no imposible. El mejoramiento genético es un proceso complejo que envuelve la creación de poblaciones heterogéneas, caracterización cualitativa y cuantitativa de estas poblaciones, el análisis genético de la variación cualitativa y cuantitativa y la selección y el estudio de los genotipos mejorados durante varios años y de los sitios para liberar los mejores cultivares. La biotecnología raramente permitirá atajos significativos en el proceso, pero puede mejorar la utilización de la variabilidad genética y vencer ciertas barreras o impedimentos biológicos. Las simples técnicas de cultivo de tejidos han facilitado considerablemente el manejo y mejoramiento del germoplasma de Musa. Las técnicas de genética molecular están siendo aplicadas para mejorar la eficaINFOMUSA — Vol 8, N° 1 cia en el mejoramiento de Musa a través del análisis de los genomas y el desarrollo de los sistemas de mejoramiento asistidos por marcadores. En el IITA se estudian marcadores candidatos SSRLP con respecto a caracteres importantes como la partenocarpia, la precocidad y la regularidad en la formación de brotes que podrían aumentar en gran medida la eficacia del mejoramiento de Musa. También se desarrollaron iniciadores PCR para el diagnóstico de los virus, que ya están brindando un método específico y sensible para detectar el virus del rayado del banano entre las plantas in vitro. Se necesita desarrollar diagnósticos confiables para el movimiento internacional seguro del germoplasma. Las plantas transgénicas con resistencia a enfermedades fungosas y virales serán útiles, si se distribuyen correctamente, lo que permitiría su uso subsiguiente en el mejoramiento convencional. Un enfoque holístico hacia el mejoramiento de Musa producirá cultivares superiores. Por lo tanto, los programas de mejoramiento de Musa multidisciplinarios deben prevalecer en los esfuerzos de mejoramiento. Estos programas se beneficiarán de las biotecnologías que han sido claramente dirigidas para complementar el mejoramiento convencional de Musa. Nos interesan las interacciones huésped/patógeno de la enfermedad de la Sigatoka negra. Por lo tanto, se utilizaron las mediciones de fitoalexinas y el despliegue diferencial RT-PCR para investigar las etapas tempranas en la estimulación de las defensas de la planta en las hojas de banano por M. fijiensis. Existen evidencias de que los filtrados de cultivos fungosos no sólo contienen extractores potenciales, sino también supresores de las reacciones de defensa de Musa. Las fitoalexinas de tipo fenilfenalenona pueden ser parte de estas reacciones, al menos en el cultivar de banano Yangambi Km5. Desarrollo de los marcadores de microsatélites específicos de los loci de Mycosphaerella fijiensis y reacciones de defensa inducida en las hojas de Musa Oligonucleótidos químericos autocomplementarios (Co) compuestos de residuos de ADN y ARN modificados fueron evaluados como medios para 1) crear sustituciones básicas estables, específicas de los sitios en un gen nuclear y 2) introducir un marco de lectura en un transgen nuclear en las células de las plantas. Para demostrar la creación de mutaciones alélicas específicas en un miembro de una familia de genes, se diseñaron CO para señalar el codón para la prolina 196 de SuRA, un gen de acetolactato sintasa (ALS) de tabaco. Una sustitución del aminoácido en el Pro196 de la ALS confiere al herbicida el fenotipo de resistencia que puede ser utilizado como un marcador seleccionable en las células de las plantas. Los CO se diseñaron para que contengan un dominio de homología de 25 nucleótidos compuesto por una región de cinco desoxiribonucleótidos (que aloja una base simple que desbalancea la secuencia ALS nativa) rodeada por regiones, cada una de las cuales está compuesta por diez ribonucleótidos. Después de la subsiguiente recuperación de las células de tabaco resistentes al herbicida en un medio selectivo, los análisis de las secuencias de ADN identificaron conversiones de bases en el gen ALS al nivel del codón para la prolina 196. Para demostrar una insersión local específica de una base simple en un gen señalado, se utilizaron los CO para restaurar la expresión de un transgen de pro- Dieter Kaemmer1, C. Neu1, O. Harprecht1, K. Weising1, K.L. Acosta2, J.G. Luis2, F. Gauhl3, A. James4, F. Khan1, C. Navarro5, C. Gimenez6, A. Gutierrez-Rojas7 y G. Kahl1 1 Plant Molecular Biology, Biocentre, Johan Wolfgang Goethe University, D-60439 Frankfurt am Main, Alemania 2 Instituto Universitario de Bio-Orgánica « Antonio González », Universidad de la Laguna, Avda Astrofísico Sánchez 2, 38206 - La Laguna, Tenerife, España 3 Chiquita Int. Ltd., San José, Costa Rica 4 CICY, Dpto de Biotecnología, Km 7 Antigua Carretera a Progreso, 9731 Merida, México 5 Micropropagación – Agromod, S.A. de C. V. Central Ote. S/N, Placa Com. K.L.-4 30700 Tapachula, México 6 Universidad Central de Venezuela, Laboratorio de Biotecnología Vegetal, Cal. S.C. de Bello Monte, 1040 Caracas, Venezuela 7 Programa National Biotecnología Agrícola, CORPOICA, Santafé de Bogotá, A.A. 240142 - Las Palmas, Colombia Se desarrollaron marcadores de microsatélite específicos de los loci para la cartografía genética del patógeno de banano M. fijiensis y el aislamiento de sus genes no virulentos,. El desempeño de estos marcadores se demuestra comparando dos pequeñas poblaciones procedentes de América y Africa, respectivamente. INFOMUSA — Vol 8, N° 1 Una herramienta para la genómica funcional de las plantas : los oligonucleótidos quiméricos de RNA/DNA causan mutaciones in vitro específicas de los genes P.R. Beetham, P.B. Kipp, X.L. Sawycky y Gregory D. May Boyce Thompson Institute, Tower Road, Ithaca NY 14853, EEUU teína fluorescente inactiva (GFP) que ha sido diseñado para que contenga una deleción de una base simple. La recuperación de las células fluorescentes confirmó la corrección de la deleción. Nuestros resultados demuestran la aplicación de una nueva tecnología para modificar loci genéticos individuales catalizando una sustitución de la base o una adición de la base a los genes nucleares específicos ; este enfoque debería tener una mayor utilidad en el área de la genómica funcional de las plantas. Análisis de diferencia representativas en el desarrollo de los marcadores de banano Christopher A. Cullis1 y K. Kunert2 1 Department of Biology, Case Western Reserve University, Cleveland OH 44106-7080, EEUU 2 Forestry and Agricultural Biotechnology Institute, University of Pretoria, Pretoria, Africa del Sur Entre las técnicas moleculares para desarrollar marcadores confiables con el fin de monitorear o identificar la variación se encuentran polimorfismos por restricción de longitud de fragmentos (RFLP), ADN polimorfico amplificado al azar (RAPDs), polimorfismos de fragmentos de longitud amplificada (AFLPs). Todas estas técnicas han sido utilizadas para generar polimorfismos entre varios genotipos de banano, y se identificó un gran número de polimorfismos. Sin embargo, ya que la variación entre los aislados es grande, fue difícil identificar el diagnóstico de variantes de un carácter particular o de un genoma particular (por ejemplo, secuencias específicas del genoma B). El análisis de las diferencias representativas (RDA) es un método mediante el cual se puede identificar la variación de ADN entre los individuos relacionados estrechamente. Esencialmente, el método consiste en una substracción de todas aquellas secuencias que se mantienen en común entre los dos individuos, y luego permite la amplificación de las secuencias que resultan diferentes. El resultado es una serie de fragmentos amplificados que representan algunas de las diferencias entre los ADN de salida. Se aplicó El RDA con el fin de identificar las cuatro líneas de secuencias de las variantes, que representan los genotipos AAA, AA y dos AAAB del banano. Se obtuvieron clones con diferencia entre los tipos AA y AAAB substraídos con el ADN a partir del tipo AAA. De las líneas caracterizadas, una fue compartida entre los dos tipos. La comparación entre la distribución de los clones de diferencia y los polimorfismos identificados en los genes de ARN ribosomal fue impresionante. Estos experimentos sólo identificaron fragmentos po- PROMUSA VII limórficos que resultaron diferentes entre dos individuos. Sin embargo, al reunir las muestras, se podría identificar el diagnóstico de los fragmentos de un genoma particular. El RDA también ha sido utilizado par aislar los marcadores asociados con la variación inducida por el cultivo de tejidos. Este trabajo fue apoyado en parte por NovoMark Technologies LLC, Cleveland, Ohio. Identificación de los marcadores RAPD ligados a las secuencias de los genomas A y B en Musa Michael Pillay, D. C. Nwakanma y A. Tenkouano Plantain and Banana Improvement Program, IITA, High Rainfall Station, Onne, PMB 008, Nchia Eleme, Rivers State, Nigeria Los bananos y los plátanos (Musa spp.) se originaron a partir de la hibridación intra e interespecífica entre dos especies diploides silvestres, M. acuminata Colla. y M. balbisiana Colla, que contribuyeron con los genomas A y B, respectivamente (Simmonds 1995). La poliploidia y la hibridación dieron origen a una cantidad de clones diploides, triploides y tetraploides con diferentes intercambios de los genomas A y B. De esta manera, los bananos de postre y de altiplano son principalmente AAA, los plátanos son AAB y los bananos de cocción son ABB. También ocurren otras combinaciones incluyendo AB, AAAB, AABB y ABBB. Simmonds y Shepherd (1955) clasificaron Musa en grupos genómicos señalando categorías de características morfológicas. La morfología puede ser influenciada por el ambiente y este sistema de clasificación puede resultar inconsistente. Se necesitan técnicas más estables y confiables para determinar los grupos genómicos en Musa. Este estudio es dirigido a identificar marcadores RAPD (ADN polimorfo amplificado al azar) para los genomas A y B. Se utilizaron ochenta iniciadores de operones 10-mer para amplificar el ADN del clon ‘Calcutta 4’ de M. acuminata spp. burmannicoides (genomas AA) y el clon Honduras de M. balbisiana clone (genomas BB). Se identificaron tres iniciadores (A17, A18 y D10) que produjeron fragmentos únicos en las dos especies. Estos iniciadores fueron examinados en una muestra de 48 genotipos nativos e híbridos que representaban varias combinaciones genómicas. Los patrones de las bandas obtenidas con estos marcadores RAPD hicieron posible aclarar la composición genómica de todas las plantas. Este método representa un sistema rápido y confiable para la identificación de los genomas en Musa. En VIII PROMUSA este estudio se identificaron marcadores PCR que son específicos a los genomas A y B en Musa, lo que podría facilitar la caracterización y manipulación genómica en las líneas de mejoramiento. La omposición genómica de los híbridos complejos puede ser determinada en la etapa de vivero utilizando unos pocos gramos de tejido foliar, venciendo problemas en el mejoramiento de Musa que incluyen propagación lenta, un ciclo de vida muy largo y grandes espacios requeridos para los ensayos en el campo. Bibliografía Simmonds N.W. 1995. Bananas Musa (Musaceae). Pp. 370-375 in Evolution of crop plants, 2nd Edition (J. Smartt J, N.W. Simmonds, eds). Longmans Scientific and Technical, Essex, UK Simmonds N.W. & K. Shepherd. 1955. The taxonomy and origins of the cultivated bananas. J. Linn Soc (London) Bot. 55 :302-312. Bioquímica y maduración de la fruta Expresión de genes en bananos en proceso de maduración Graham B. Seymour Horticulture Research International, Wellesbourne, Warwick CV35 9EF, Reino Unido La fisiología y la bioquímica del proceso de maduración de los bananos han sido estudiadas extensamente, y los informes en la literatura sobre la modificación de la intensidad en la respiración de la fruta se remontan a 1931. Desde entonces se conocieron los principales eventos bioquímicos asociados con el proceso de maduración en bananos que incluyen un marcado ascenso en la biosíntesis de etileno, la conversión de almidón en azúcares, la producción de compuestos volátiles, desarreglo de las paredes celulares y desintegración de la clorofila en la cáscara. Sin embargo, las vías bioquímicas precisas responsables por estos cambios, no son bien comprendidas y sus estudios han sido impedidos por los altos niveles de almidón y compuestos fenólicos en las frutas. Recientemente, aparecieron nuevas herramientas moleculares que han permitido estudiar el patrón de la expresión de genes en estas frutas. Este trabajo describe la clonación de los genes relacionados con la maduración contenidos en los tejidos de la cáscara y la pulpa y cómo estas secuencias de genes pueden mejorar nuestro entendimiento de los principales eventos moleculares y bioquímicos responsables por el control del proceso de ma- duración en los bananos (ver Medina-Suarez et al. 1997, Clendennen y May 1997). Bibliografía Medina-Suarez R., K. Manning, J. Fletcher, J Aked, C.R. Bird & G.B. Seymour. 1997. Gene expression in the pulp of ripening bananas. Plant Physiology 115 : 453-461 Clendennen S.K. & G.D. May. 1997. Differential gene expression in ripening banana fruit. Plant Physiology 115 : 463-469 Control de la maduración en banano Colin Bird Zeneca Plant Science, Jealott’s Hill Research Station, Bracknell, Berkshire, RG42 6ET, Reino Unido Muchas de las prácticas de producción en la industria bananera son determinadas por la necesidad de controlar el comienzo y la tasa de maduración para asegurar que la fruta de calidad consistente llegue al consumidor. La sincronización de la cosecha determina la vida verde postcosecha que se requiere para que la fruta llegue a los mercados distantes. La cosecha temprana es dictada por una variedad de razones y resultará en la pérdida del peso potencial del tallo. Una habilidad mejorada para controlar el inicio de la maduración podría tener un impacto substancial sobre las prácticas empleadas en la producción de bananos. Como en otras frutas climatéricas, el etileno desempeña un papel clave en la regulación de la iniciación y la subsiguiente tasa de maduración del banano. Varios mecanismos que se utilizan para disminuir la producción de etileno demoran la maduración en tomates y otras frutas. Esta investigación examina el papel del etileno en la maduración del banano y describe el trabajo dirigido a controlar la producción de etileno en las plantas de banano modificadas genéticamente. Caracterización de las transcripciones específicas de las frutas y sus promotores asociados en los bananos Cavendish Stephanie K. Clendennen, J.A. Kellogg, C.B. Phan, N.M. Webb, V.R. Bommineni, H. Mathews y D. Ry Wagner Agritope, Inc. 16160 SW Upper Boones Ferry Rd, Portland OR 97224, EE UU La estrategia para controlar el etileno de Agritope implica la expresión de la S-adenosil metionina hidrolasa (SAMase) de una manera que es específica de los tejidos y que es regulada a medida que se desarrolla. Para aislar un promoINFOMUSA — Vol 8, N° 1 tor específico de la fruta para controlar la expresión de SAMase en el banano, primero fue necesario identificar una transcripción propia, específica de la fruta y expresada de manera abundante. Anteriormente se identificó la pectasa liasa (PEL) como específica de la fruta y asociada con la maduración en bananos. Existen al menos dos diferentes pectate liasas de la secuencia cADN que indican que la PEL es representada por una pequeña familia de genes en el banano. La presencia de dos genes PEL similares fue confirmada mediante la amplificación PCR del ADN genómico con iniciadores de oligonucleótidos complementarios a las regiones conservadas de la secuencia codificadora de la PEL que mide un intrón. Se clonaron y secuenciaron dos productos de diferentes tamaños (PEL1 y PEL2) y se diseñaron los iniciadores de oligonucleótidos específicos de los genes. En adición al análisis de pectate liasa, se identificaron varios fragmentos anónimos no relacionados del cADN como específicos de la fruta y expresados abundantemente en el banano mediante un despliegue diferencial. El análisis de 36 fragmentos de despliegue diferencial específicos de la fruta condujo a la selección eventual de dos para realizar el aislamiento y caracterización de los promotores. Los fragmentos deseados de cADN de despliegue diferencial de banano (G1A y G8A) fueron clonados y secuenciados y se descubrió que los mismos muestran una homología limitada de secuencias nucleótidas con genes de otras plantas que codifican enzimas biosintéticas. Igual que la PEL, el G1A y G8A son miembros de pequeñas familias de genes representadas por dos miembros similares cada una. Las transcripciones de PEL, G1A y G8A fueron confirmadas como específicas de las frutas y de la maduración asociados mediante la hibridación con la técnica de Northern. Teniendo la información sobre las secuencias específicas de los genes, se diseñaron los iniciadores oligonucleótidos para penetrar en el genoma del banano y eventualmente aislar las regiones reguladoras asociadas con cada uno de los genes específicos de la fruta. Se discutirán los ensayos funcionales de los promotores putativos específicos de la fruta en las construcciones de los genes reporteros. Conversión de carbohidratos durante la maduración del banano – un estudio del metabolismo de carbono Randolph M. Beaudry Michigan State University, East Lansing, MI 48824-1325, U.S.A. La maduración en el banano está marcada por el aumento de las tasas de producción de etileno y INFOMUSA — Vol 8, N° 1 CO2 y por la conversión de cantidades relativamente grandes de carbono en forma de almidón en substancias que alteran la percepción del sabor de la fruta. Durante la maduración, el almidón, que comprende aproximadamente el 80 % del peso seco de la fruta, se convierte en sacarosa, glucosa, fructosa. Una fracción mucho más pequeña se convierte en compuestos aromáticos, principalmente los esteres, que representan una diminuta fracción de la reserva de carbono. Incongruentemente, en términos de la calidad del sabor de la fruta fresca, las dos reservas de carbono son de importancia relativamente igual. Los cambios en el metabolismo de carbohidratos están acompañados por marcados desplazamientos en actividades enzimáticas y reservas de metabolitos (Beaudry et al. 1987, 1989). El comienzo del aumento de la respiración coincide con un descenso del fosfo-enol-piruvato, la acumulación de la glucosa-6-fosfato y piruvato y por acumulación de la fructosa 1,6-bifosfato relativo a la fructosa 6-fosfato. Los datos indican un fomento de la glicolisis vía fosfofructoquinasa dependiente de ATP y quinasa de piruvato. El pico en el flujo respiratorio coincide con la tasa máxima de conversión de carbohidratos gluconeogénicos de almidón en azúcares, un proceso que requiere de energía. La tasa del flujo de carbono hacia los azúcares es aproximadamente de 40 a 50 veces más alta que el flujo de carbono a través de la vía respiratoria. El control simultáneo de la partición de carbono es en parte ayudado por un declive en la sacarosa que degrada la enzima sacarosa sintasa y una promoción alostérica de la sacarosa que forma la enzima sacarosa fosfato sintasa vía aumentos marcados en glucosa-6-fosfato (Cordenunsi and Lajolo 1995). La glucosa-6-fosfato también mejora la actividad del fosfoenolpiruvato carboxilasa, un proceso que puede contribuir al incremento de malato y citrato durante la maduración (Law and Plaxton 1995). La producción de esteres, responsables por el aroma del banano, empezó inmediatamente después de ocurrir el pico en la actividad respiratoria y aumentó a medida que la demanda de energía requerida para convertir el almidón en azúcares se aproximó a cero (Song y Beaudry, datos no publicados). La capacidad del tejido para producir esteres estuvo presente al menos 1 día antes de este período, sin embargo, la limitación sugerida de substrato controla la producción de esteres y la disponibilidad de este substrato aumenta marcadamente después de ocurrir el pico respiratorio. Bibliografía Beaudry R.M., N. Paz, C.C. Black & S.J. Kays. 1987. Banana ripening : Implications of changes in internal CO2 and ethylene concentrations, pulp fructose 2,6bisphosphate and activity of some glycolytic enzymes. Plant Physiol. 85 : 277-282. Beaudry R.M., R. Severson, C.C. Black & S.J. Kays. 1989. Banana Ripening : Implications of changes in the concentration of critical glycolytic intermediates and fructose 2,6-bisphosphate. Plant Physiol. 91 : 1436-1444. Cordenuns, B.R. & F.M. Lajolo 1995 Starch breakdown during banana ripening : Sucrose synthase and sucrose phosphate synthase. J. Agric. Food Chem. 43 :347-351. Law D.R. & W.C. Plaxton. 1995. Purification and characterization of a novel phosphoenolpyruvate carboxylase from banana fruit. Biochem. J. 307 :807-816. Proceso de metabolismo de almidón en sacarosa en bananos Joao Nascimento, B.R. Cordenunsi y F.M. Lajolo Departamento de Alimentos e Nutrio Experimental (FCF-USP), Av. Lineu Prestes 580, Bloco 14, CEP 05508-900, Saó Paulo/SP, Brasil El mecanismo de la degradación rápida del almidón y la acumulación de azúcar soluble en bananos todavía no se ha establecido con exactitud. A pesar de la importancia de este cambio en la fisiología de la fruta o de su calidad para el consumo, poco se sabe de los mecanismos implicados. El contenido de almidón se reduce de unos 14-22 % a casi cero en unos pocos días, mientras que el contenido de sacarosa aumenta de menos de 1 % a casi 15 %. Este proceso parece ser el mismo para varios cultivares de banano durante la maduración, sin embargo, el nivel máximo de almidón y de azúcares en las frutas inmaduras y maduras puede ser muy diferente. La cantidad de almidón puede variar de 14 % en un cultivar Mysore inmaduro a 23 % en un cultivar Nanico inmaduro. El contenido de sacarosa también puede variar, desde 16 % en los cultivares Prata-Comum y Nanica maduros, hasta sólo 5 % en un Mysore maduro. La degradación del almidón en bananos parece ser un proceso exo-corrosivo, tal como se observa al revisar los resultados obtenidos mediante un microscopio electrónico y la distribución del tamaño de los gránulos. A medida que continuó el proceso de maduración, apareció una acumulación de pequeños gránulos y, a diferencia de lo que sucede en algunos cereales, no se observaron perforaciones en la superficie de los gránulos, sólo estrías, indicativas de la degradación del almidón. También se estudió el papel de muchas enzimas que probablemente están implicadas en la degradación del almidón durante la maduración de banano. Se observó un incremento marginal de la fosforilasa de almidón y, después que se inició el proceso de degradación del almidón, se observó una dis- PROMUSA IX minución de la fosforilasa hasta alcanzar su valor inicial. En las rodajas de banano infiltradas con la glucosa-1-P marcada se observó una acumulación de sacarosa marcada, lo que muestra que la fosforilasa de almidón podría influir sobre la degradación de almidón, debido a que el producto de su actividad podría ser utilizado para la síntesis de sacarosa. La actividad de las enzimas implicadas en la hidrólisis de almidón, la ß-amilasa y la α-amilasa, también aumentaron durante el proceso de maduración. El aumento en la actividad de la α-amilasa coincidió con la actividad de degradación del almidón, pero su pico ocurrió exactamente después de que la mayor parte del almidón ya se había degradado. Es posible que hasta la mínima actividad de la αamilasa fue suficiente para promover el acceso inicial hacia los gránulos de almidón. La actividad de la ß-amilasa también aumentó después de la cosecha, alcanzando su máximo durante el período de degradación del almidón. La detección de niveles de maltosa superiores a los de los oligosacáridos producidos por la α-amilasa en suspensiones de almidón incubadas con extractos crudos con EDTA, podría indicar la importancia de la ß-amilasa para el proceso de degradación del almidón. Se observó que durante el proceso de maduración el contenido total del almidón bajó, pero no se detectaron cambios en la relación amilosa/amilopectina. Este resultado indica la existencia de un sistema de enzimas capaz de hidrolizar las bandas glucosídicas α1,4 y α-1,6 y que las enzimas implicadas podrían tener una acción coordinada, ya que no hubo acumulación de cualesquiera de los constituyentes polímeros de los gránulos de almidón. Este sistema parece actuar, ya que la actividad de las glucosidasas α-1,4 y α 1,6 aumentó al proceder la degradación del almidón. En relación a las enzimas implicadas en la síntesis de la sacarosa en plantas, la sacarosa sintasa (SS) y la sacarosa-fosfato sintasa (SFS), se descubrió que su actividad siguió tendencias opuestas durante la maduración. La actividad de la sacarosa-fosfato sintasa es baja durante el desarrollo de la fruta y aumenta al alcanzar la madurez fisiológica. Después de la cosecha, este aumento se acelera alcanzando su máximo al producirse la acumulación de la sacarosa. El aumento de la actividad fue seguida estrechamente por un aumento en SFS de mARN y proteína, mostrando que la expresión de genes puede contribuir al aumento en la actividad de SFS durante la maduración del banano. La actividad de SS fue alta durante el desarrollo de la fruta, reduciéndose durante la maduración. Similar a lo observado para la SFS, el mARN y la proteína de la SS también X PROMUSA experimentaron una disminución. Estos resultados sugieren que es la SFS, y no la SS, la que está implicada en la síntesis de sacarosa durante la maduración de bananos y similar a lo observado para otras enzimas, la expresión de genes desempeña un papel importante en la regulación del nivel de actividad de estas enzimas en bananos. La actividad de SS y SFS fue afectada por la cosecha y la relación de la actividad SS/ SFS fue diferente para los bananos en maduración en la planta y en los cortados. Cuando la fruta se deja madurar en la planta, todo su comportamiento es similar al de la fruta cosechada, pero su velocidad es diferente. En las frutas sin cosechar estos cambios en la actividad y en el contenido de almidón y azúcar son más lentos. El aumento en la actividad de SFS fue bajo, alcanzando sólo el 50 % del valor obtenido para las frutas cosechadas. Es posible la existencia de metabolitos u hormonas transportados a las frutas controlando el nivel de ambas enzimas. En realidad, las rodajas de banano infiltradas con el ácido indol-acético (IAA) mostraron una demora en la maduración. El efecto más destacado del tratamiento con IAA fue la degradación del almidón, que se demoró casi siete días más en relación con las rodajas de control. La demora en la degradación del almidón fue estrechamente seguida por una demora en la actividad total de amilasa. Los borrones de Northern del ARN total del IAA infiltrado y de la rodaja de control, provista con una sonda de ß-amilasa de banano, mostró que la transcripción de ß-amilasa también fue afectada por el tratamiento con IAA, lo que es consistente con la demora en la actividad total de amilasa y degradación de almidón. Esto sugiere que esta hormona puede afectar la maduración, y la cosecha puede representar un cambio dramático en el suministro de la hormona afectando la actividad y expresión de enzimas implicadas en la degradación de almidón y síntesis de sacarosa. En conclusión, durante la maduración de bananos, el almidón se transforma principalmente en sacarosa mediante una acción concertada de varias enzimas, controladas a nivel de transcripción (amilasas, SPS, SS) o no (fosforilasa). Es necesario realizar más trabajos sobre el control de la expresión de estas enzimas. Agradecimiento Deseamos agradecer al Dr G. B. Seymour por la clonación de ß-amilasa y el apoyo financiero de FAPESP y CNPq. Bibliografía Areas J.A.G. & F.M. 1981. Starch transformation during banana ripening : I. the phosphorylase and phosphatase behavior in Musa acuminata. J. Food Biochem. 5: 19-37. Cordenunsi B.R. & F.M. Lajolo. 1995. Starch breakdown during banana ripening : sucrose synthase and sucrose-phosphate synthase. J. Agric. Food Chem. 43 (2) : 347-351. Garcia E. & F.M. Lajolo. 1988. Starch transformation during banana ripening : the amylase and glucosidase behavior. J Food Science 53 (4) : 11811186. Mota R.V., F.M. Lajolo & B.R. Cordenunsi. 1997. Carbohydrate composition of some banana cultivars (Musa spp.) during ripening. Ciênc. Tecnol. Aliment, 17 (2) : 94-97. Nascimento J.R.O., B.R. Cordenunsi & F.M. Lajolo. 1999. Sucrose synthase activity and expression during development and ripening in bananas. Submitted to publication. Nascimento J.R.O., B.R. Cordenunsi & F.M. Lajolo. 1997. Partial purification and characterization of sucrosephosphate synthase from pre-climacteric and climacteric bananas. J. Agric. Food Chem. 45 : 11031107. Nascimento J.R.O., B.R. Cordenunsi, F.M. Lajolo & M.J.C. Alcocer. 1997. Banana sucrose-phosphate synthase gene expression during fruit ripening. Planta 203 : 283-288. Terra N.N., E. Garcia & F.M. Lajolo. 1983. Starch-sugar transformation during banana ripening : the behavior of UDP-glucose pyrophosphorylase, sucrose synthetase and invertase. J Food Science 48 : 10971100. Caracterización molecular de la señalización de etileno y la vía de control de desarrollo que regula la maduración de la fruta de tomate J. Giovannoni Department of Horticultural Sciences and Crop Biotechnology Centre, Texas A & M University, College station, TX 778432133, EEUU La maduración de las frutas carnosas es regulada por una combinación de las características de desarrollo, hormonales y ambientales. En el tomate se identificaron mutaciones de genes sencillas que influyen sobre las vías que representan las tres clases generales de los sistemas de señalización. Los ejemplos incluyen el mutante receptor de etileno Neverripe (Nr), los mutantes de respuesta a la luz de alta pigmentación (hp-1, hp-2), el inhibidor de la maduración (rin) y el regulador de desarrollo de maduración (nor). Los esfuerzos de nuestro grupo se han centrado en las tecnologías basadas en genomas para la clonación de genes para todas las clases e incluyen la clonación basada en la confección de mapas y las estrategias de la confección de mapas de los genes candidatos. La clonación basada en confección de mapas dio como resultado el aislaINFOMUSA — Vol 8, N° 1 miento de los genes candidatos para los loci rin y nor. Los cADN candidatos para rin y nor muestran la segregación conjunta con el locus objetivo, los patrones de expresión específica de maduración, y las alteraciones de las secuencias de ADN en los alelos derivados de los mutantes versus las isolíneas cercanas normales. Un enfoque similar es una vía que se explora actualmente para el aislamiento del gene hp-1. Un esfuerzo de colaboración para la confección de mapas de genes candidatos dio como resultado el aislamiento del receptor de etileno Nr parecido a ERS y los esfuerzos actuales se enfocan sobre la caracterización de los pasos de señalización de etileno hacia abajo. El aislamiento de los genes arriba mencionados representa un fundamento para el desarrollo de un modelo molecular para el control de la maduración. Virología Papel de los satélites de ADN en la réplica del virus bunchy top del banano y la infección causada por el mismo C. Horser, S. Hermann, R. Harding y James Dale Centre for Molecular Biotechnology, School of Life Sciences, Queensland University of Technology, Brisbane, Queensland, Australia A partir de las infecciones con el virus bunch top del banano (BBTV) se aislaron componentes del cssADN, que potencialmente codifican proteínas asociadas con su replicación (Reps), y una estructura en forma de tallo en bucle que podría actuar como origen de replicación. Sin embargo, estos posibles satélites de ADN, a los que hemos denominado ADNS1 y ADN-S2, no comparten las regiones conservadas dentro de la región intergénica característica de los seis cssADN originales. Más, solo hemos sido capaces de identificar estos componentes satélites en los aislados asiáticos del BBTV, lo que sugiere que estos componentes satélites no son partes integrantes del virus. Actualmente tenemos una fuerte evidencia de que estos componentes satélites no están implicados en la replicación del BBTV. Cuando 1.1 mers clonados de ADN-S1 fueron bombardeados solos dentro de las suspensiones celulares del banano, los mismos fueron capaces de ser extraídos y replicados, lo que indica que el ORF en el ADN-S1 fue una Rep. Sin embargo, cuando 1.1 mers de ADN-1 fueron bombardeados dentro de las suspensiones celulares, los mismos no fueron capaces de ser extraídos o replicados ; este hecho solo ocurrió en presencia de 1.1 mers INFOMUSA — Vol 8, N° 1 de ADN-5, los cuales parecen codificar una proteína retinoblastoma responsable de introducir las células anticipadamente infectadas en la fase S. Más, cuando 1.1 mers de ADN-3, componente que codifica la proteína de revestimiento, fueron bombardeados dentro de las células junto con el ADN-1 y ADN-5, los tres fueron extraídos y replicados, lo que indica que la Rep, codificada por ADN-1, fue capaz de iniciar su propia replicación, el ADN-3 y el ADN-5. De manera inversa, el ADN-S1 no requirió el ADN5 para su replicación y, lo que es importante, fue capaz de iniciar su propia replicación, pero no la replicación de ADN-5 o ADN-3. Esto proporciona un fuerte evidencia de que para la replicación del BBTV solo se necesita la Rep, codificada por ADN-1, y que el ADN-S1 y el S2 son ADN satélites o virus satélites. Un promotor del virus baciliforme de la caña de azúcar induce la expresión transgénica en banano y otras plantas monocotiledóneas y dicotiledóneas P.M. Schenk1,2, T. Remans1,3, R.G. Dietzgen2, M.J. Bernard2, R. Swennen3, M.W. Graham2, J.M. Manners1 y László Sági3 1 Cooperative Research Centre for Tropical Plant Pathology Level 5, John Hines Bldg, St. Lucia, Qld 4072, Australia 2 QDPI, Queensland Agricultural Biotechnology Centre, the University of Queensland, St. Lucia, Qld 4072, Australia 3 Laboratory of Tropical Crop Improvement, Katholieke Universiteit Leuven, Kardinaal Mercierlaan 92, 3001 Heverlee, Bélgica - Email : [email protected] Un fragmento de ADN de 1369 bp (Sc) fue aislado a partir de un clon completo del badnavirus baciliforme de la caña de azúcar (ScBV) que mostró tener una actividad promotora en los ensayos con la expresión transitiva utilizando plantas monocotiledóneas (banano, maíz, mijo y sorgo) y dicotiledóneas (tabaco, canola, girasol y Nicotiana benthamiana). También se analizó la expresión estable del promotor de las plantas transgénicas de banano y tabaco. Estos experimentos mostraron que este promotor podría inducir un alto nivel de expresión del gen informador b-glucuronidase (GUS) en la mayoría de las células de estas plantas. Por ejemplo, el nivel de expresión fue comparable con el promotor de ubiquitina de maíz en los ensayos estandarizados con la expresión de genes transitivos con el maíz. En las plantas transgénicas de banano, los niveles de la expresión GUS fueron variables para diferentes líneas transgénicas y generalmente fueron comparables con las actividades del promotor de ubiquitina de maíz y del promotor mejorado CaMV 35S. En adición, el promotor Sc parece expresarse de una manera casi constituyente en las plantas transgénicas de banano y tabaco. De esta manera, el promotor del virus baciliforme de la caña de azúcar representa una herramienta útil para conseguir una expresión de alto nivel de genes foráneos en las plantas transgénicas mono y dicotiledóneas que podría ser utilizada de manera similar a la de los promotores CaMV 35S o ubiquitina de maíz. Bibliografía Schenk P.M., L. Sági, T. Remans, R.G. Dietzgen, M.J. Bernard, N.W. Graham, R. Swennen & J.M. Manners. 1999. A promoter from sugarcane bacilliform badnavirus drives transgene expression in banana and other monocot and dicot plants. Plant Molecular Biology (in press). Expresión episomal inducida por el estrés de las secuencias del badnavirus del rayado del banano (BSV) integradas en el genoma de Musa Tsitsi Ndowora1, Ganesh Dahal1, Daryle LaFleur1, Glyn Harper2, Roger Hull2, Neil E. Olszcwski1 y Ben Lockhart1 1 Department of Plant Pathology, University of Minnesota, St Paul, MN 55108, EEUU 2 John Innes Centre, Norwich Research Park, Colney, Norwich NR4 7UH, Reino Unido Las secuencias genómicas del badnavirus del rayado del banano (BSV), un pararetrovirus de las plantas, están integradas en el genoma de Musa spp. Cuando ciertos genotipos de Musa, libres de virus y enfermedades, son propagados in vitro, la infección con el BSV se desarrolla en los propágulos, pero no en el hijuelo producido por la propagación vegetativa normal. Un integrante viral consiste de un genoma parcial de BSV y es considerado incapaz de dar origen a un genoma viral capaz de replicación. Un segundo integrante contiene más que un genoma viral completo. La secuencia nucleótida de esta forma integrada es casi idéntica a la del BSV que ocurre en el tejido de las plantas derivadas de los cultivos de tejidos, y el arreglo de las secuencias virales dentro de este locus sugiere que el mismo puede dar origen a un genoma viral episomal capaz de replicación vía recombinación homóloga. Se propone que los estrés asociados con la propagación in vitro dispara la activación episomal del mismo o de las secuencias virales integradas. Se discutirán los resultados de la investigación en curso cuyo objetivo consiste en identificar y caracterizar integrantes adicionales del BSV presentes en Musa y el posible papel de otros factores de estrés en iniciar la expresión episomal de las secuencias virales integradas. PROMUSA XI Análisis funcional de los genes del virus bunchy top del banano R. Wanitchakorn, A. James, G. Hafner, C. Horser, R. Harding y James Dale Centre for Molecular Biotechnology, School of Life Sciences, Queensland University of Technology, Brisbane, Queensland, Australia El virus bunchy top del banano (BBTV) es un miembro del grupo de nanovirus y como tal, contiene viriones isométricos y un genoma del componente múltiple de cssADN. Anteriormente hemos descrito seis diferentes componentes de cssADN que parecen estar presentes en todas las infecciones con el BBTV (Burns et al. 1995). Más, hemos demostrado que siete mARN diferentes se transcriben a partir de estos seis componentes (Beetham et al. 1997, 1998), cada uno del ADN-2 al 6 y dos del ADN-1. También hemos demostrado que el principal gen de ADN-1 codifica una proteína Rep que tiene actividades de corte y ligamiento y pueden iniciar la replicación propia y de otros componentes del ADN del BBTV (Hafner et al. 1997, Horser et al. Sin publicar) y que el gen del ADN-3 es el gen de la proteína de revestimiento (Wanitchakorn et al. 1997). Actualmente, tenemos evidencias de que (i) el ADN-5 codifica una proteína de retinoblastoma de tipo fijación que está implicada en el desvío de las células prematuramente infectadas en las células del componente de replicación de ADN (fase S), (ii) el ADN-4 codifica una proteína de movimiento que se dirige a la periferia de la célula, (iii) el ADN-6 codifica una proteína que se dirige al núcleo, pero también interactúa con la proteína de movimiento que se dirige al núcleo codificada por el ADN-4 y por lo tanto es una nucleoproteína transportadora potencial y (iv) el pequeño gen codificado por el ADN-1 es esencial para la iniciación de la replicación in vivo pero no se requiere para el corte o ligamiento in vitro y por lo tanto es posible que esta proteína es equivalente a la proteína mejoradora de replicación de los geminivirus. Estos resultados sugieren que el BBTV es muy similar en muchas maneras a los begomovirus, y requiere una proteína de movimiento y una nucleoproteína transportadora para el movimiento de célula a célula y que la nucleoproteína transportadora no solo es una proteína de revestimiento como la que se encuentra en los mastrivirus. Bibliografía Beetham P.R., G.J. Hafner & J.L. Dale. 1997. Two mRNAs are transcribed from banana bunchy top DNA-1. J. Gen. Virology 78 : 229-236. Burns TM, R.M. Harding & J.L. Dale. 1995. The genome organization of banana bunchy top virus : XII PROMUSA analysis of six ssDNA components. J. Gen. Virology 76 : 1471-1482. Hafner G.J., M.R. Stafford, L.C. Wolter, R.M. Harding & J.L. Dale. 1997. Nicking and joining activity of banana bunchy top virus replication protein in vitro. J. Gen. Virology 78 : 1795-1799. Wanitcharkorn R., R.M. Harding & J. Dale. 1997. Banana bunchy top virus DNA-3 encodes the viral coat protein. Arch. Virology 142 : 1673-1680. Cultivo de células y transformación del banano Desempeño en campo de las plantas de banano derivadas de las suspensiones de células embriogénicas (Musa AAA, cv. Grande Naine) F.X. Côte1, M. Folliot2, R. Domergue1 & C. Dubois1 1 CIRAD-FLHOR & CIRAD-BIOTROP, B.P 5035, 34032 Montpellier Cedex 1, Francia 2 CIRAD-FLHOR, BP 153, 97202 Fort de France, Martinica Se necesita un método eficaz de regeneración mediante embriogénesis somática para apoyar el mejoramiento genético y mejorar la propagación masiva de las especies de Musa. Plagas y numerosas enfermedades virales y fungosas amenazan seriamente a los bananos y plátanos. La transformación genética ofrece una posibilidad para mejorar cultivares estériles. La hibridación somática también puede contribuir al mejoramiento genético de Musa. Ambas tecnologías requieren de un método eficaz a través de la embriogénesis somática. El uso de los bananos, derivados a partir de los brotes cultivados in vitro como material de plantación, es una de las técnicas de mejoramiento más recientes diseñadas para este cultivo, que proporciona plantas de una calidad sanitaria muy alta, las cuales, al combinar las técnicas de prácticas agronómicas adecuadas, producirán altos rendimientos y reducirán el uso de plaguicidas (nematicidas). Sin embargo, el costo de las vitroplantas producidas a través de cultivo de brotes in vitro permanece alto en comparación con los métodos convencionales. Debido a su alta eficacia de multiplicación, la técnica de embriogénesis somática probablemente reducirá estos costos de manera significativa. Se han desarrollado varios métodos de embriogénesis somática con la utilización de diferentes explantes para bananos y plátanos (Cronauer y Krikorian 1988, Novak et al. 1989, Escalant y Teisson 1989, Dhed’a et al. 1991, Ma 1991, Marroquin et al. 1993, Escalant et al. 1994, Grapin et al. 1996). Sin embargo, hasta donde sabemos, no se han publicado informes sobre la variación somaclonal, crecimiento y rendimiento de las plantas de banano derivadas a partir de estos cultivos in vitro. Recientemente se informó sobre un método de regeneración a través de suspensiones de células embriogénicas para el cv. Grande Naine, el banano de postre más común (Côte et al. 1996). En este trabajo presentamos las características de crecimiento de estas plantas y las comparamos con las plantas producidas mediante el método convencional de propagación de brotes in vitro. Durante la fase de aclimatación se observaron dos tipos de variantes entre 500 plantas derivadas de suspensiones de células embriogénicas. El primer tipo relacionado con las plantas de banano con “hojas variegadas o deformadas” también fue observado en las plantas derivadas a partir de los brotes in vitro. El segundo tipo se refería a las plantas con “hojas dobles” (es decir, una de las primeras hojas que se desarrolló durante la fase de aclimatación consistía de dos partes que se fundían en la vena central). Sin embargo, durante el crecimiento en el campo, las variantes produjeron plantas que no diferían morfológicamente de otras plantas. De este modo, ninguna de las plantas derivadas de las suspensiones de células mostraron caracteres anormales en el campo. El análisis estadístico mostró que no hubo diferencias significativas entre las plantas producidas mediante la técnica de micropropagación con respecto a su altura y circunferencia, largo de la hoja de referencia, cantidad de inflorescencias y frutas, peso del racimo, período entre la plantación y la cosecha. Este estudio mostró que es posible regenerar plantas de banano con un comportamiento agronómico similar al de las plantas producidas mediante el método convencional de cultivo de brotes in vitro, a partir de las suspensiones de células embriogénicas bajo las condiciones de cultivo utilizadas. Bibliografía Côte F.X., R. Domergue, S. Monmarson, J. Schwendiman, C. Teisson & J.V. Escalant. 1996. Embryogenic cell suspensions from the male flower of Musa AAA cv. Grand nain. Physiol. Plant 97 : 285-290. Cronauer S.S. & A.D. Krikorian. 1988. Plant regeneration via somatic embryogenesis in the seeded diploid Musa ornata Roxb. Plant Cell Rep 7: 23-25. Dhed’a D., F. Dumortier, B. Panis, D. Vuylsteke & E. De Langhe. 1991. Plant regeneration in cell suspension cultures of cooking banana cv. « Bluggoe » (Musa spp. ABB group). Fruits 46 : 125-135. Escalant J.V. & C. Teisson. 1989. Somatic embryogenesis from immature zygotic embryos of INFOMUSA — Vol 8, N° 1 the species Musa acuminata and Musa balbisiana. Plant Cell Rep.7 : 665-668. Escalant J.V., C. Teisson & F.X. Côte. 1994. Amplified somatic embryogenesis from male flowers of triploid banana and plantain cultivars (Musa spp.). In Vitro Cell. Dev. Biol.30 : 181-186. Grapin A., J. Schwendiman & C. Teisson. 1996. Somatic embryogenesis in plantain banana. In Vitro Cell. Dev. Biol. 32 :66-71. Ma S.S. 1991. Somatic embryogenesis and plant regeneration from cell suspension culture of banana. Pp.181-188 in Proceedings of symposium on tissue culture of horticultural crops. Taipei, Taiwan 8-9 March 1988. Marroquin C.G., C. Paduscheck, J.V. Escalant & C. Teisson. 1993. Somatic embryogenesis and plant regeneration through cell suspensions in Musa acuminata. In Vitro Cell. Dev. Biol.29 : 43-46. Novak F.J., R. Afza, M. Van Duren, M. Perea-Dallos, B.V. Conger & T. Xiolang. 1989. Somatic embryogenesis and plant regeneration in suspension cultures of dessert (AA, AAA) and cooking (AAB) bananas. Bio/Tech. 46 : 125- 135. Transformación genética de los bananos Cavendish (Musa spp. grupo AAA) cv. ‘Grande Naine’ vía bombardeo con microproyectiles Douglas K. Becker1, B. Dugdale1*, M.K. Smith2, R.M. Harding1, J. L. Dale1 1 Centre for Molecular Biotechnology, School of Life Sciences, Queensland University of Technology, GPO Box 2434 Brisbane, Queensland 4001, Australia. 2 Centre for Subtropical Fruit, Maroochy Department of Primary Industries, Queensland Horticulture Institute, PO Box 5083, Sunshine Coast Mail Centre, Nambour, Queensland 4560, Australia. *Dirección actual, Boyce Thompson Institute for Plant Research, Tower Road, Ithaca, New York 14853, EEUU. Hemos desarrollado un método para la transformación y regeneración estables de los bananos Cavendish banana (Musa spp. grupo AAA) cv. ‘Grande Naine’, utilizando el bombardeo con microproyectiles. Las suspensiones de células embriogénicas fueron iniciadas utilizando flores masculinas inmaduras como explantes (Côte et al. 1996). Las células fueron bombardeadas con un plásmido que contenía el gen reportero de proteína (gfp) verde fluorescente bajo el control del promotor del virus del mosaico de la coliflor 35S (CaMV 35S) y el gen marcador seleccionable de neomicina fosfotransferasa (npt II) bajo el control de un nuevo promotor derivado del virus bunchy top del banano (BBTV) (Dugdale et al. 1998). La regeneración bajo la selección con kanamicina fue monitoreada desde las células transformadas únicas hasta las plantas enteras utilizando el gfp. La eficacia de la transformación fue alta con un promedio de 24 plantas transgénicas generadas INFOMUSA — Vol 8, N° 1 por cada bombardeo. La caracterización molecular de los transformantes demostró que los transgenes se han integrado de manera estable en el genoma del banano. Bibliografía Côte F.X., R. Domergue, S. Monmarson, J. Schwendiman, C. Teisson & J.V. Escalant. 1996. Embryogenic cell suspensions from the male flower of Musa AAA cv. Grand nain. Physiol. Plant 97 : 285-290. Dugdale B., P.R. Beetham, D.K. Becker, R.M. Harding & J.L. Dale. 1998. Promoter activity associated with the intergenic regions of banana bunchy top virus DNA-1 to -6 in transgenic tobacco and banana cells. Journal of General Virology 79 :2301-2311 Transformación mediante Agrobacterium de los cultivos de suspensiones de células embriogénicas de banano Juan B. Pérez Hernández1, R. Swennen2, V. Galán Saúco1 y L. Sági2 1 Departamento de Frutas Tropicales, Instituto Canario de Investigaciones Agrarias, Apartado 60, 38200 La Laguna, España - Email : [email protected] 2 Laboratory of Tropical Crop Improvement, Katholieke Universiteit Leuven, Kardinaal Mercierlaan 92, 3001 Heverlee, Bélgica Para elaborar una transformación de bananos eficaz mediante Agrobacterium, se investigaron los pasos consecutivos en la interacción planta-bacteria. La chemotaxia de agrobacterias hacia los tejidos se estudió utilizando un sistema múltiple de platos de agar. Se descubrió que los tejidos extraídos de las hojas, cormo y raíces, así como los tejidos de las plantas cultivadas in vitro de las razas indígenas de banano son capaces de producir una reacción quemotáctica positiva de A. Tumefaciens, que podría ser mejorada si se produce una herida extensa. La suspensión de Agrobacterium fue examinada mediante la microscopía de fluorescencia con rayos ultravioleta y microscopía electrónica, las cuales demostraron la presencia de las bacterias ligadas individualmente o fijadas masivamente a la superficie de las células y tejidos individuales de banano. La inducción de los genes vir fue detectada utilizando una fusión de genes lacZ :virE. La transferencia de T-ADN, monitoreada con una construcción del gene informador gusAintron en ensayos histoquímicos transitivos, se observó en cultivos de células embriogénicas de los cultivares Grande Naine, Williams, Three Hand Planty, y FHIA-18. Las colonias transgénicas putativas se seleccionaron en geneticin o Basta a una frecuencia que es diez veces mayor lo que resulta después de un bombardeo con partículas. Finalmente, un total de más de 250 plantas trans- génicas han sido regeneradas y confirmadas mediante el análisis PCR, manchado histoquímico y análisis de Southern. Bibliografía Pérez Hernández J.B., S. Remy, V. Galán Saúco, R. Swennen & L. Sági. 1999. Chemotactic movement to wound exudates and attachment of Agrobacterium tumefaciens to single cells and tissues of banana. Journal of Plant Physiology (in press). Pérez Hernández J.B., S. Remy, L. Sagi, R. Swennen & V. Galán Saúco. 1998. Chemotactic movement to wound exudates and attachment of Agrobacterium tumefaciens to single cells and tissues from banana plants. Acta Horticulturae 490 : 463-468. Análisis de la expresión del gen uidA de los transgenes en las plantas de banano Grande Naine (AAA) cultivadas en el campo Y. Moy, B. Hanson, D. Engler y Neal Gutterson DNA Plant Technology Corporation, 6701 San Pablo Avenue, Oakland California 94 608, EE UU Hemos desarrollado un método eficaz para la producción de plantas de banano transgénicas. El método para la entrega del ADN emplea el cultivo conjunto de las flores masculinas derivadas a partir de los callos embriogénicos con la cepa EHA101 de Agrogacterium tumefaciens, y es altamente eficaz. Utilizando este método, hemos producido poblaciones de plántulas de banano transformadas independientemente que contenían genes quiméricos uidA para evaluar la actividad de tres promotores diferentes : el 35S mejorado, el Ubi-1 de maíz, y el SpMAS. Estas plantas han sido evaluadas con respecto a la expresión de beta-glucuronidasa primero en el laboratorio, y luego maduradas y sembradas en los ensayos en el campo, establecidos en Costa Rica y en el sur de México. Se presentarán los resultados iniciales de la expresión del promotor en los tejidos vegetativos. Transformación y regeneración del cultivar de banano Rasthali (AAB) T.R. Ganapathi1, N. Higgs2, Joyce Van Eck2, P.J Balint-Kurti2 y G.D. May2 1 Nuclear Agricultural and Biotechnology Division, Bhabha Atomic Research Center, Trombay, Mumbai 400 085, India 2 Boyce Thompson Institute for Plant Research, Cornell University, Tower Road, Ithaca, New York 14853-1801, EEUU Hemos desarrollado un sistema de transformación y regeneración para el cultivar local de banano Rasthali (AAB). Las suspensiones de cé- PROMUSA XIII lulas embriogénicas fueron generadas a partir de los callos iniciados de los tejidos de las plantas del banano Rasthali propagadas in vitro. Estas suspensiones celulares fueron utilizadas en los experimentos de transformación con Agrobacterium. Se generaron aproximadamente 200 transformantes independientes. Se presentarán informes sobre la generación de las suspensiones celulares de Rasthali, y la transformación, regeneración y el análisis de las plantas transgénicas. Patología fungosa e interacciones huésped/planta Las enfermedades más importantes del banano : trabajo actual y futuro en Sigatoka negra y marchitamiento por Fusarium Randy C. Ploetz University of Florida, Tropical Research & Education Center, 18905 SW 280 th Street, Homestead, FL 33031-3314 EEUU La Sigatoka negra, causada por Mycosphaerella fijiensis, y el marchitamiento por Fusarium, causado por Fusarium oxysporum f. sp. cubense (Foc), son las enfermedades del banano más significativas en todo el mundo. Las cepas de M. fijiensis procedentes de Africa, América, Papua Nueva Guinea, Filipinas y Pacífico Sur se examinaron con los análisis RFLP. El desequilibrio gamético entre 19 loci nucleares principalmente mostró asociaciones no significativas que son indicativas de una reproducción sexual al azar. En contraste, el desequilibrio gamético entre los pares de 36 loci de RFLP del Foc no fue significativamente al azar. De este modo, la dispersión global de los aislados con haplotipos idénticos y una fuerte correlación entre los marcadores genéticos independientes (por ejemplo, los GCV, mitocondrias y los haplotipos RFLP de loci múltiples) indican que este patógeno tiene una estructura clonal de poblaciones. Adicionalmente, los análisis de los loci RFLP y las secuencias de los genes nucleares (factor de elongación de la traducción 1—αa y ARN ribosomico de la pequeña subunidad mitocondrial) demostraron independientemente que el patógeno no es monofilético ; en las colecciones mundiales se detectaron al menos tres poblaciones independientes evolutivas del patógeno. Esta es la primera vez que se demostró que este hecho ocurre en las formas especiales (formae speciales) de F. oxysporum. XIV PROMUSA Los orígenes y el movimiento de ambos patógenos parecen ser bastante similar. La diversidad genética de las poblaciones de M. fijiensis fue la más alta en Papua Nueva Guinea y en Filipinas, y el 88 % de los alelos encontrados en otras regiones también fueron encontrados en las poblaciones asiáticas. Con excepción de una única población de Foc que se encuentra en Malawi, todas las poblaciones de este patógeno se encuentran o están relacionadas con las poblaciones encontradas en el sur y sureste de Asia. Estos datos son corroborados por los datos de impresión de huellas de ADN. En resumen, los datos disponibles apoyan la hipótesis de que M. fijiensis y Foc se desarrollaron conjuntamente con su banano huésped en Asia. En ambos hongos se reconoce una diversidad patogénica significativa. Las cepas de M. fijiensis en el Pacífico Sur afectan a los clones que se utilizan como progenitores “resistentes” en los actuales programas de mejoramiento, y existe el potencial para que se desarrollen nuevos patotipos de este patógeno vía recombinación meiótica. Aunque el último mecanismo no está disponible en el Foc, las poblaciones procedentes del Sureste asiático afectan una gran extensión de germoplasma, incluyendo el importante subgrupo Cavendish. Hasta la fecha, los laboriosos esquemas de mejoramiento convencionales han producido los únicos ejemplos de los nuevos híbridos que resisten a estas enfermedades. Sin embargo, se anticipa que un mapa saturado de enlace del genoma de Musa acuminata mejorará los esfuerzos de mejoramiento y que el trabajo con las poblaciones huéspedes segregadas eventualmente permitirán la selección con la ayuda de los marcadores para desarrollar la resistencia en el futuro [en esta reunión se informó sobre un marcador RFLP que está enlazado con la resistencia a la Sigatoka negra (Careel et al.)]. La transformación genética del banano con respecto a su resistencia a estas enfermedades representa el reto más difícil y aún no ha sido exitoso. Sin embargo, la corta historia de estos esfuerzos y la diversidad de enfoques y genes que están disponibles para estos trabajos sugieren que un banano desarrollado con la ayuda de la ingeniería genética y resistente a las enfermedades, puede ser producido en el futuro. Bibliografía Bentley S., K.G. Pegg, N.Y. Moore, R.D. Davis & I.W. Buddenhagen. 1998. Genetic variation among vegetative compatibility groups of Fusarium oxysporum f. sp. cubense analyzed by DNA fingerprinting. Phytopathology 88 :1283-1293. Carlier J., M.H. Lebrun, M.F. Zapater, C. Dubois, & X. Mourichon. 1996. Genetic structure of the global population of banana black leaf streak fungus, Mycosphaerella fijiensis. Molecular Ecology 5 :499510. Carlier J., X. Mourichon, D. Gonzalez de León, M.F. Zapater & M.H. Lebrun. 1994. DNA restriction fragment length polymorphisms in Mycosphaerella species that cause banana leaf spot diseases. Phytopathology 84 :751-756. Fullerton R.A. & T.L. Olsen. 1995. Pathogenic variability in Mycosphaerella fijiensis Morelet, cause of black Sigatoka in banana and plantain. New Zealand Journal of Horticultural Science 23:39-48. Koenig R.L., R.C. Ploetz & H.C. Kistler. 1997. Fusarium oxysporum f. sp. cubense consists of a small number of divergent and globally distributed clonal lineages. Phytopathology 87 :915-923. O’Donnell K., H.C. Kistler, E. Cigelnik & R.C. Ploetz. 1998. Multiple origins of the fungus causing Panama disease of banana : concordant evidence from nuclear and mitochondrial gene genealogies. Proceedings of the National Academy of Science USA 95 :2044-2049. Ploetz R.C. & K.G. Pegg. 1997. Fusarium wilt of banana and Wallace’s line : Was the disease originally restricted to his Indo-Malayan region ? Australasian Plant Pathology 26 :239-249. Caracterización de un gen de resistencia análogo y adaptación de la técnica cADN-AFLP en banano Ilse Wiame, I. Engelborghs, R. Swennen y L. Sági Laboratory of Tropical Crop Improvement, Katholieke Universiteit Leuven, Kardinaal Mercierlaan 92, 3001 Heverlee, Bélgica - Email : [email protected] Hasta la fecha, se han identificado cinco clases de genes de resistencia a las enfermedades de las plantas (genes R) que están involucrados en la transducción de señales (Baker et al. 1997). Para la clase que contiene un sitio de unión de nucleótidos (NBS) y una repetición rica en leucinas (LRR), se diseñaron los iniciadores basados en los dominios conservados (Leister et al. 1996) y se utilizó la clonación basada en PCR para aislar genes de resistencia putativos análogos en algunas variedades de banano. Todos los clones de banano del tamaño esperado de la variedad Zebrina GF (ITC0966), que es resistente a la Sigatoka negra, resultaron de la amplificación del iniciador derivado de una región rica en leucina solamente, para la temperatura de hibridación tanto alta como baja. Sin embargo, hemos descubierto un clon con homología significativa a los genes pertenecientes a otra clase de los genes R. Esta clase incluye los genes Cf de tomate que confieren resistencia al patógeno fungoso Cladosporium fulvum y son caracterizados por una región transmembrana y una LRR (Jones et al. 1994, Dixon et al. 1996,ThoINFOMUSA — Vol 8, N° 1 mas et al. 1997). La secuencia traducida de aminoácidos fue homóloga a la región terminal carboxila de dos productos de genes alélicos (Cf-4 y Cf-9), que comprende la parte transmembrana. Una vuelta de PCR inverso estiró el clon en la LRR. En este punto, la secuencia traducida también fue homóloga a Cf-2: 33 % de aminoácidos idénticos y 48 % de aminoácidos similares sobre una longitud de 313 aa. En la LRR del clon de banano la secuencia coherente para las LRR de la planta podría ser reconocida claramente (Kajava 1998). La secuencia correspondiente a la región terminal amina ha sido clonada por otra vuelta de PCR inverso y su secuencia nucleotídica está siendo determinada. También hemos adaptado al banano la técnica cADN-AFLP más confiable (Bachem et al. 1996) como una alternativa al despliegue diferencial para el aislamiento de los genes expresados diferencialmente (Jones et al. 1998). Brevemente, el ARN se transcribe a la inversa en el cADN que subsecuentemente es digerida por las enzimas de restricción y ligada a los adaptadores. Debido a que la amplificación se realiza con iniciadores específicos de adaptadores, en contraste con el despliegue diferencial, se generan productos de amplificación no específicos y la mayoría de los productos contendrán la secuencia codificadora. Para demostrar la eficacia de este método se analizó la expresión de los genes específicos de la hoja y de la base foliar de dos variedades de banano con dos pares de iniciadores anclados. En promedio, se identificaron 11 bandas diferenciales por cada combinación de iniciadores después de realizar una novedosa electroforesis ultrarápida en gel (QuickPoint, Novex). Estos geles pequeños, en contraste con los geles secuenciadores utilizados comúnmente, sólo requieren muy poco tiempo de funcionamiento (15-20 min) y pueden ser convenientemente teñidos con plata. Esta técnica será utilizada para identificar y caracterizar genes inducidos por patógeno fungoso en banano. Bibliografía Bachem C.W.B., R.S van der Hoeven, S.M. de Bruijn, D. Vreugdenhil, M. Zabeau & R.G.F. Visser. 1996. Visualization of differential gene expression using a novel method of RNA fingerprinting based on AFLP : analysis of gene expression during potato tuber development. Plant J. 9 : 745-753. Baker B., P. Zambryski, B. Staskawicz & S.P. DineshKumar. 1997. Signalling in plant-microbe interactions. Science 276 : 726-733. Dixon M.S., D.A Jones, J.S. Keddie, C.M. Thomas, K. Harrison & J.D.G. Jones. 1996. The tomato Cf-2 disease resistance locus comprises two functional genes encoding leucine-rich repeat proteins. Cell 84 : 451-459. Jones D.A., C.M. Thomas, CK.E. Hammond-Kosack, P.J. Balint-Kurti & J.D.G. Jones 1994. Isolation of INFOMUSA — Vol 8, N° 1 the tomato Cf-9 gene for resistance to Cladosporium fulvum by transposon tagging. Science 266 : 789-793. Jones J.T. & B.E. Harrower. 1998. A comparison of the efficiency of differential display and cDNA-AFLPs as tools for the isolation of differentially expressed parasite genes. Fundam. Appl. Nematol. 21 : 81-88. Kajava A.V. 1998. Structural diversity of leucine-rich repeat proteins. J. Mol. Biol. 277 : 519-527. Leister D., A. Ballvora, F. Salamini & C. Gebhardt. 1996. A PCR-based approach for isolating pathogen resistance genes from potato with potential for wide application in plants. Nature Genet. 14 : 421-429. Thomas C.M., D.A. Jones, M. Parniske, K. Harrison, P.J. Balint-Kurti, K. Hatzixanthis & J.D.G. Jones. 1997. Characterization of the tomato Cf-4 gene for resistance to Cladosporium fulvum identifies sequences that determine recognitional specificity in Cf-4 and Cf-9. Plant Cell 9 : 2209-2224. plantas transgénicas independientes. Para medir con precisión el área foliar necrosada (AFN) y para la clasificación de transformantes independientes de acuerdo con su tolerancia al hongo se utilizaron la captura de imagen por computadora y el cálculo del área con un software utilizando un índice necrótico (IN). El cálculo de este índice se realiza dividiendo el AFN del control, una línea no transformada, por el AFN de la línea transformada. A partir del cribado de 48 líneas transgénicas independientes, se identificaron 2 líneas que mostraron una alta tolerancia al hongo (IN mayor que 4) y una alta expresión de PAM (más de 4 veces mayor que la de base) de acuerdo a las mediciones con ELISA. Las pruebas en el campo revelarán si estos transformantes se comportan igual bajo la presión natural de la enfermedad. Bibliografía Desarrollo de un ensayo con muestra foliar para evaluar la tolerancia del banano a los hongos Serge Remy, I. Deconinck, R. Swennen y L. Sági Laboratory of Tropical Crop Improvement, Katholieke Universiteit Leuven, Kardinaal Mercierlaan 92, 3001 Heverlee, Bélgica - Email : [email protected] Varios genes que codifican proteínas antimicrobianas (PAM) han sido introducidos en los cultivos de suspensiones de células embriogénicas de banano vía bombardeo con partículas (Sagi et al. 1995) con el fin de generar plantas transgénicas con resistencia a Mycosphaerella fijiensis, el patógeno fungoso más importante del banano. En una gran cantidad de líneas transgénicas independientes, la integración, transcripción y expresión estables de transgenes fueron demostradas mediante las técnicas PCR y Southern (Remy et al. 1998a, 1998b), RT-PCR y Northern y ELISA, respectivamente. Para evaluar la tolerancia a los hongos en líneas transgénicas, se ha desarrollado un bioensayo sencillo, sensible y reproducible utilizando discos foliares. Un disco foliar de 5x5 cm fue extraído de las plantas transgénicas cultivadas en el invernadero y utilizado para la inoculación con un hongo de prueba. Cuatro días después de la infección, se observó una clara respuesta diferencial entre los transformantes independientes, mientras que un hongo no patogénico no fue capaz de inducir síntomas de enfermedad indicando que el ensayo es específico para las interacciones específicas de huésped-patógeno. Las transformaciones con diferentes construcciones de genes promotores dieron como resultado un amplio rango de tolerancia al hongo entre las Sági L., B. Panis, S. Remy, H. Schoofs, K. De Smet, R. Swennen & B.P.A. Cammue. 1995. Genetic transformation of banana and plantain (Musa spp.) via particle bombardment. Biotechnology 13, 481485. Remy S., I. François, B.P.A. Cammue, R. Swennen & L. Sági. 1998a. Co-transformation as a potential tool to create multiple and durable disease resistance in banana (Musa spp.). Acta Horticulturae 461 : 361365. Remy S., A. Buyens, B.P.A. Cammue, R. Swennen & L. Sági. 1998b. Production of transgenic banana plants expressing antifungal proteins. Acta Horticulturae 490 : 433-436. Estudio de las interacciones Mycosphaerella/banano utilizando patógenos transgénicos expresando una proteína verde fluorescente Peter J. Balint-Kurti1, Mi Jin2, J. Clardy2, S.S. Mungekar1, A.C.L. Churchill1 y G.D. May1 1 Boyce Thompson Institute for Plant Research, Tower Road, Ithaca, NY 14853, USA 2 Department of Chemistry and Chemical Biology, Cornell University, Ithaca, NY USA - Email : [email protected] Un sistema de transformación genética fue desarrollado para tres especies estrechamente relacionadas de Mycosphaerella, todas patógenas de los bananos y plátanos (Musa spp.). M. fijiensis y M. musicola, agentes causales de la Sigatoka negra y amarilla respectivamente, y una especie de Mycosphaerella recientemente caracterizada con un Spetoria anamorfo (J. Carlier, personal communication) fueron transformados con una construcción llevando el gen sintético sGFP (S65T) codificando para una proteína verde fluorescente (GFP). Transformantes de cada especie expresaron el GFP constitutivamente a través todas las hiphas y PROMUSA XV las conidiosporas. Usando estos transformantes, se observó la interacción banano/Mycosphaerella en mas detalles y con mas facilidad que con los métodos anteriores. Transformantes individuales fueron caracterizados y su crecimiento en los huéspedes susceptibles fueron examinados en detalle. La expresión de GFP permitió observar el crecimiento del hongo bajo la superficie de la hoja. El crecimiento a la superficie de la hoja fue globalmente similar en cada una de las tres especies observadas. La penetración de la hoja se hizo exclusivamente por los estomas. Las hifas crecieron mas entre que dentro la células y no se evidencio estructuras de tipo hostoria. Estructuras de infección conocidas como stomapodia se formaron sobre las aperturas de los estomas en ambos huésped y non - huésped especies, sugiriendo que no se necesita un señal banano - específico para tal evento.. Sin embargo, la penetración de los estomas solo se noto en las hojas del banano. Se observo un crecimiento abundante de las especies Mycosphaerella/Septoria sobre el non – huésped tabaco. Muchas veces, la muerte de la célula foliar ocurre bien delante de la hifa creciente, sugiriendo la producción por Mycosphaerella de una fitotoxina difundible. Varias fracciones de un extracto de M. musicola no mostraron una especificidad ‘luz - activada’ en la fitotoxicidad contra células de banano en suspensión.. La existencia de toxinas activadas por la luz podría explicar la reducción muchas veces observada en los síntomas de la enfermedad de Sigatoka, cuando los bananos se cultivan bajo sombra. Experimentos in vitro así como observaciones en plantación, sugieren que las tres especies examinadas producen un auto–inhibición de la germinación de las conidiosporas. Un extracto con una actividad inhibitoria fue aislado a partir de esporas de Mycosphaerella. Estudio de poblaciones de Mycosphaerella fijiensis y mejoramiento genético de bananos para resistencia a la enfermedad de las rayas negras Jean Carlier1, A. El Hadrami1, H. Hayden2, M.F. Zapater1 and F. Lapeyre1 1 Laboratoire Phytrop, CIRAD-AMIS, BP 5035, 34032 Montpellier, France 2 Botany Department, The University of Queensland, Brisbane 4072, QLD, Australia Conocimientos sobre la amplitud y distribución de la variabilidad dentro de Mycosphaerella fijiensis son necesarios para un programa de mejoramiento genético eficiente y para una resistencia durable a la enfermedad XVI PROMUSA de la rayas negras. Los estudios de poblaciones pueden contribuir a la definición de estrategias para ambas hibridación y evolución de la resistencia en tiempo y espacio (Leung et al. 1993, Milgroom and Fry 1997). La estructura genética de la población de M. fijiensis fue estudiada en forma global, a nivel de país y campo, utilizando los marcadores moleculares (Carlier et al. 1994, 1996, Hayden et al. 1998, Carlier et al. resultados no publicados). Estos estudios mostraron que las poblaciones de M. fijiensis pueden mantener un alto nivel de diversidad genética en una pequeña escala, y que la recombinación juega un papel importante en este patógeno. Por consecuencia, se podría también que la poblaciones de M. fijiensis mantengan un alto nivel de variabilidad patogénica, permitiendo una adaptación a genotipos huéspedes resistentes introducidos recientemente, lo que fue observado con ‘Paka’ y ‘T8’ en Rarotonga en las Islas de Cook (Fullerton and Olsen 1995). El papel importante de la recombinación genética debería también estar considerado en los programas de hibridación del banano. Combinar genes de resistencia especifica en cultivares individuales (piramidal) tiene el riesgo de no estar durable y el uso de una mezcla de variedades o de resistencia parcial ofrece quizás una solución mas durable. La región Australasia-Asia del Sudeste es el centro de diversidad de M. fijiensis. Los efectos fondadores acompañando las introducciones de M. fijiensis en América Latina, en las Islas del Pacifico y en Africa han llevado a una reducción de la diversidad genética en estas regiones. Estos resultados son consistentes con la hipótesis en que M. fijiensis tiene su origen en Australasia-Asia del Sudeste y fue introducido recientemente en otras regiones productoras de bananos (Mourichon and Fullerton 1990). Los análisis de población de M. fijiensis deberían incluir estudios de su variabilidad patogénica. La caracterización de la resistencia del germoplasma a la enfermedad es también esencial para la hibridación en bananos. Es con esta finalidad que estamos desarrollando una técnica de inoculación de M. fijiensis, utilizando fragmentos de hojas de banano (El Hadrami et al. 1998). Se obtiene así modelos de infección y una evolución de los síntomas similares a los que se observaron en condiciones de campo. Todos los estadios diferentes del ciclo de vida de M. fijiensis fueron reproducidos en los ensayos con fragmentos de hoja y se distinguieron cultivares con fenotipo resistente. Diferencias significativas en la eficiencia de la infección y en el tiempo de incubación fueron destacadas en seis accesiones parcialmente resistentes de bananos con 12 aislamientos de M. fijiensis. Otros parámetros del ciclo de vida del patógeno se están investigando, y la fiabilidad de los ensayos con fragmentos de hojas se está evaluando en el campo utilizando las mismas accesiones de banano. El método de inoculación sobre fragmentos de hojas podría utilizarse para la caracterización de la resistencia parcial de cultivares y la evaluación de la variación de la agresividad del patógeno. Tal estudio, incluyendo ensayos epidemiológicos, ayudaría en identificar cuales son los componentes de la resistencia parcial que reducen significativamente la taza de desarrollo de la enfermedad al campo. Este método con marcadores moleculares podría también estar utilizado en la evaluación de la evolución de las poblaciones del patógeno en repuesta a la presión de selección ejercida por los cultivares resistentes y además, para ajustar una estrategia de resistencia durable. Bibliografía Carlier J., X. Mourichon, D. Gonzales de León, M.F. Zapater & M.H. Lebrun. 1994. DNA restriction fragment length polymorphisms in Mycosphaerella species causing Banana leaf spot diseases. Phytopathology 84 :751-756 Carlier J., M.H. Lebrun, M.F. Zapater, C. Dubois & X. Mourichon. 1996. Genetic structure of the global population of Bananas black leaf streak fungus Mycosphaerella fijiensis. Molecular Ecology 5 : 499510 El Hadrami A., M.F. Zapater, F. Lapeyre, C. Abadie & J. Carlier. 1998. A leaf disk assay to assess partial resistance of banana germplasm and aggressiveness of Mycophaerella fijiensis, the causal agent of black leaf streak disease. 7 th International Congress of Plant Pathology, Edinburgh, Scotland. Hayden H.L., E. A. Aitken & K.G. Pegg. 1998. Population analysis of Mycosphaerella fijiensis in the Torres Strait, Australia and nearby regions. 7 th International Congress of Plant Pathology, Edinburgh, Scotland. Leung H., R.J. Nelson & J.E. Leach. 1993. Population structure of plant pathogenic fungi and bacteria. Advances in Plant Pathology 10 :157-205. Milgroom M. G. & W.E. Fry. 1997. Contributions of population genetics to plant disease epidemiology and management. Advance in Botanical Research 24:1-30. INFOMUSA — Vol 8, N° 1
 0
0
Anuncio
Documentos relacionados
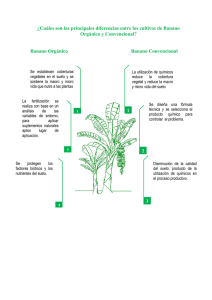







Descargar
Anuncio
Añadir este documento a la recogida (s)
Puede agregar este documento a su colección de estudio (s)
Iniciar sesión Disponible sólo para usuarios autorizadosAñadir a este documento guardado
Puede agregar este documento a su lista guardada
Iniciar sesión Disponible sólo para usuarios autorizados