Vista - DEPARTIR
Anuncio

MEMORIAS
Primer Congreso Nacional del
Teocintle de Nicaragua
(Zea nicaraguensis Iltis & Benz)
Managua, Nicaragua
04 Noviembre, 2011
MEMORIAS
Primer Congreso Nacional del
Teocintle de Nicaragua
(Zea nicaraguensis Iltis & Benz)
Primer congreso Nacional del Teocintle de
Nicaragua (Zea nicaraguensis Iltis & Benz)
El teocintle (Zea nicaraguensis Iltis & Benz),
un recurso genético de importancia mundial,
ayudemos a conservarlo
Invitación al Primer congreso Nacional del Teocintle de Nicaragua
(Zea nicaraguensis Iltis & Benz) en la Universidad Nacional Agraria
COMITÉ ORGANIZADOR DEL CONGRESO
El Teocintle de Nicaragua, (Zea nicaraguensis, Iltis & Benz), un recurso
genético de importancia mundial, ayudemos a conservarlo
Dr. Carlos Henry Loáisiga Caballero
Responsable del Laboratorio de Biotecnología-UNA
Dr. Denis Salazar Centeno
Decano Facultad de Agronomía-UNA
Ing. MSc. Álvaro Benavides González-UNA
Coordinar del Proyecto FAO/APACUNCA
Ing. MSc. José Dolores Cisne-UNA
Coordinador del equipo DEPARTIR
Organizadores y expositores del Primer Congreso Nacional del teocintle de Nicaragua
Editores
Ing. Álvaro Benavides González
Ing. Juan Carlos Morán Centeno
INDICE DE CONTENIDO
Contenido
Página
Presentación
1
Descubriendo, caracterizando y utilizando germoplasma de los teosintes
centroamericanos. Dr. Robert McK. Bird.
2
Estado actual del teocintle (Zea nicaraguensis Iltis & Benz) en la Reserva
de Recursos Genéticos de Apacunca (RRGA). Ing. Álvaro Benavides
González.
17
Las áreas protegidas y los Recursos Genéticos. Ing. Edilberto Duarte.
32
La importancia de los recursos genéticos silvestres y el rol de la UNA en
su protección y aprovechamiento. Ing. Daniel Querol Lipcovich.
38
Diversidad genética y evolución del maíz silvestre de Nicaragua:
41
Teocintle (Zea nicaraguensis Iltis & Benz). Dr. Carlos Henry Loáisiga
Caballero.
Turismo sostenible y Áreas Protegidas. Dra. Matilde Somarriba Chang.
45
Poster
52
Elementos de discusión
54
Comentarios del teocintle entre expositores
55
Breve reseña bibliográfica de los expositores en el Congreso
56
Lista de participantes y temáticas desarrolladas
57
Invitados principales
58
Programa
60
Gráficas del Congreso
61
PRESENTACIÓN
La Reserva de Recursos Genéticos de Apacunca (RRGA) obtuvo este estatus mediante la
Ley 217 (Ley General del Medio Ambiente y los Recursos Naturales), el Artículo 154 y
publicada en La Gaceta (Diario Oficial de Nicaragua), No. 105 del 06 de junio de 1996.
Esta valiosa reserva genética está siendo mermada por las comunidades humanas aledañas,
que en su mayor parte se dedica a la ganadería y la agricultura. En este sentido la
Universidad Nacional Agraria se propuso, junto a otros actores incorporar a las
comunidades aledañas de Somotillo y Villanueva en el proceso de conservación y uso del
teocintle y especies asociadas, manteniendo los hábitats, y ampliar la investigación sobre el
conocimiento científico de la biología, ecología y genética del teocintle como pariente
silvestre del maíz.
La historia natural del teocintle es de vital importancia, considerando que el maíz (Zea
mays L.), tiene su origen en Mesoamérica, por tanto la variabilidad genética existente
simboliza un reservorio para el mejoramiento de plantas cultivadas, los estudios indican
que las especies silvestres que se encuentran emparentadas con el maíz, como es el caso del
teocintle (Zea spp.), esta amenazadas por el proceso de erosión genética (Miranda, 1997).
El grupo de Desarrollo Participativo Integral Rural (DEPARTIR) desarrolló el proyecto
denominado: Rescate, conservación y manejo sostenible del teocintle de Nicaragua
(Zea nicaraguensis Iltis & Benz) en la reserva de Recursos Genéticos de Apacunca, el
cual fue financiado por FAO durante el período 2010-2011. Esto con el objetivo de
coadyuvar a la conservación del teocintle y el mejoramiento de la calidad de vida de las
comunidades vinculadas a este importante recurso genético.
Las primeras investigaciones sobre el teocintle en Nicaragua se realizaron en 1996 por la
Universidad Nacional Agraria y fue reconocido tentativamente como Zea luxurians. En ese
mismo año, se reconocieron 7 poblaciones de teocintle en Somotillo y Villanueva,
municipios de Chinandega. Hoy en día, sólo existe una población de teocintle en El
Papalonal.
Los temas presentados y expuestos a discusión estuvieron enfocados a los recursos
fitogenéticos nacionales y los hábitats donde se encuentra el teocintle.
Ing. MSc. Álvaro Benavides González
Coordinador del Proyecto/FAO/APACUNCA
Descubriendo, caracterizando y utilizando germoplasma de los teosintes
centroamericanos
Dr. Robert McK. Bird
1. El Gran Teocintle por El Rodeo, Reserva de Apacunca, Chinandega
La importancia del teosinte nicaragüense se observa de varias maneras. Entre los teocintles se
distingue por muchas caracteres. Vamos a revisar los diversos teocintles. Estudios de unas de
las especies incluyendo visitas a Apacunca, y al final el proyecto japonés, seguido por
sugerencias para estudios y preguntas.
Hay varios teocintles los más comunes son los Teocintles Anuales de México (TAM) con
varias razas geográficas. También en México hay dos especies perennes (Z. perennis y Z.
diploperennis). En Centro América hay Z. nicaraguensis, Z. luxurians y Z. mays subespecie
huehuetenangensis.
El género Zea (los maíces y los teocintles) ahora se divide en dos secciones, uno con los
maíces y los TAM en una especies, el otro, distinguido por muchos caracteres, las especies Z.
nicaraguensis, Z. luxurians, Z. perennis y Z. diploperennis. Las poblaciones del
departamento guatemalteco de Huehuetenango en el oeste Guatemala son distintas, más o
menos, intermedio entre las dos secciones, unos caracteres de una sección, unos de la otra.
Para una historia del descubrimiento, la caracterización y la historia de cambios en
nomenclatura de los teocintles centroamericanos, ver la bibliografía, Z. luxurians en los años
1869/1872/1924/1967/1978, Z. nicaraguensis en 1989/2000, y lo de Huehuetenango en
1967/1980/1990.
El Gran Teocintle. Tomado de Alvaro Benavides (2002).
2
2.
Hugh Iltis y Alfredo Grijalva por Apacunca
En 1989 el ingeniero Allan Hruska encontró semillas del teocintle Nica al noreste de
Chinandega y les envió a un amigo en los EEUU, quien notificó al Dr. Hugh Iltis. El año
siguiente, Iltis organizó una búsqueda para el origen de las semillas con los Doctores: Alfredo
Grijalva y Bruce Benz, y lo encontraron cerca al Rancho Apacunca.
Semillas que ellos colectaron fueron depositadas en el banco de germoplasma de CIMMYT
en México.
En 1997, el Ing. Carlos Henry Loáisiga e Ing. Alvaro Benavides González colectaron semilla
de la misma población. Iltis y Benz dieron el apellido, Z. nicaraguensis, al Gran Teocintle en
2000. En 2002 Ing. Benavides terminó sus estudios del Gran Teosinte para su tesis de
maestría, y en 2011, Loaisiga finalizó su tesis doctorado tratando del mismo (los dos tesis
están a disposición en Internet).
El Dr. Grijalva y el Dr. Iltis sobre el río que bordeo al Rancho Apacunca.
Hábitat del teocintle. Tomado de Iltis & Benz (2000).
3. Vista de la zona de la Reserva de Apacunca, tomado por un satélite
Nota: Esta discusión está en un proceso de cambio porque, hace poco, tres poblaciones
fueron descubierto en México y descrito parcialmente en un artículo de hace dos meses. Dos
se relacionen con los perennes, y uno (del este del estado de Oaxaca) es curiosamente
semejante a Z. luxurians.
3
Vista de la zona de la Reserva de Apacunca, tomado por un satélite
4. Mis estudios de los teocintles en tres décadas
Empecé mis estudios de los teocintles en los años 70, y en 1978 cambié el apellido del
teosinte oriental de Guatemala a Zea luxurians. Hasta entonces, este teosinte tenía una
historia de cambios de nombre, últimamente a Zea mays subesp. luxurians, una tontería
porque hay muchas diferencias citoplásmicas, genéticas y morfológicas ya conocidas entre
estos, el teocintle y las poblaciones de maíz y los teocintles mexicanos anuales.
Usando un invernadero en Missouri, realice crucé Z. luxurians con un maíz pequeño. Los
híbridos crecieron a más que 4 m de altura, cada uno con cuatro tallos secundarios (hijos) de
4 m. Me sorprendió, y en los años 90 en México observé igual al cruzar Z. nicaraguensis con
un maíz convencional.
Mi interés en el Gran Teocintle siguió así – La semilla de Iltis, Grijalva y Benz en el
CIMMYT necesitaba un aumento para
disponer de suficiente semilla para los
investigadores. Esto fue en proceso en la
estación experimental de Tlaltizapán
cuando visité el lugar temprano en 1995. Y
las plantas casi floreciendo, pero todavía
muchos hijos basales crecieron, algo muy
inesperado. Pedí que me llevarán dos
plantas a la sede. El Batán, después de la
cosecha para ver si iban a seguir creciendo.
Sí, crecieron como plantas perenes, y corté
uno en tres partes. Sobrevivieron y devolví
un parte a Tlaltizapán adonde unos
científicos franceses lo cruzaron con un
maíz resultando en unas semillas híbridas
que crecieron a plantas de casi 5 m con
varios hijos tan altos. Una planta de
teocintle que quedó en El Batán floreció
dos veces más. Caramba, que especie, pero hay más.
4
Después, en Carolina del Norte, puse a crecer unas muestras del Gran Teocintle en un
invernadero y detrás de mi casa. Como fue observado en mis siembras del mismo en México,
empezó muy normalmente, como otros teocintles, hasta tener cinco hojas (tres semanas).
5. Planta tipo prostrada, a tres meses, sin inundación
A cuatro semanas, brotes o hijos empezaron a salir de los tallos, y de los hijos salieron otros
brotes; últimamente habían 22 a 31 en total. Una semana más tarde, los tallos estaban
inclinando hacía el suelo. Finalmente unos fueron completamente prostrado, otros inclinado
50°-70° grados de la vertical.
A las 15 semanas todos los tallos empezaron a crecer más verticalmente, a empezar la etapa
de floración. Se podían cortar los brotes y dejar crecerlos para tener varias clones de y
compararles en distintas condiciones. Por ejemplo, atrás de mi casa, metí unos en
contenedores de agua. Cubrí otros con una jaula de plástico negro para inducir floración
manteniendo 13 horas de oscuridad. Las mismas plantas fueron creciendo en el invernadero
sin florecer por culpa de luces problemáticas; finalmente tocaron al techo. Otras
características observadas en Carolina del Norte:
Con inundación, muchos raíces crecieron arriba del suelo, en el agua, hasta tocar al aire;
Unas raíces desarrollaron “cepillas” pequeñas, grupos de pequeños raíces;
Sin inundación, los entrenudos fueron muy cortos, con distancias entre el nudo 1 y el nudo
20 de 3.5 cm a 11.5 cm; con inundación, fueron mucho más largo, hasta 10 cm o más por
entrenudo.
Raíces adventicias salieron de muchos nudos del tallo principal.
La floración fue susceptible a largo de la noche, inducido por períodos oscuros de 12 horas
o más para 21 noches o menos, variando con la edad de la planta.
Escribí un resumen en el 2000, disponible en el Internet. En el año 2000 el Departamento
de Agricultura de los EE.UU otorgó fondos para observar y colectar el nuevo teocintle en
Nicaragua.
6. El teocintle en el bosque del estero
Acompañado del Dr. Loaisiga, Dr. Grijalva, y el Ing. Benavides viajé dos veces a Apacunca,
con el objetivo de para observar la situación y colectar muestras de semillas.
7. Una planta seca con ramas, muy impresionante
Allí hemos encontrado un “bosque” del teocintle, al lado de un bosque ribereño. La zona se
inunda varias veces al año y las plantas responden a la humedad, ya que crecen muy alto.
Para ese entonces, había dos metros de agua sobre las plantas de poblaciones de teocintle.
8. Raíces resultando de la inundación que mantenían la planta
Se colectaron 100 muestras de plantas individuales para la obtención de semillas, tratando de
tener una muestra representativa. Cinco muestras grandes que mezclaron la semilla de 15 a
26 plantas fueron para satisfacer pedidos de semilla.
5
Tallos secos de teocintle en El Papalonal, Apacunca.
9. Croquis de la área del teocintle en el año 2000 – una especie
El área en que se encuentra el Gran Teocintle extiende 200 m por 160 m, en manchas,
unas muy densas.
10. Zona del teocintle, tomado por satélite
En esta vista se nota la cerca que es el límite occidental de la población. Es una de las
especies más en peligro de extinción, por las vacas que le gusta, un finquero quien quiere
erradicarlo, un volcán que puede erupcionar, etc.
11. Tolerancia a inundaciones en tres teocintles centroamericanas: un proyecto japonés.
http://www.japanlandjourney.com.
6
12 Arrozales en terrazas en Japón
El gobierno japonés quiere cambiar unos arrozales regados del país a campos de producción
de maíz porque el consumo de carne ha aumentado. Para soportar más la producción de
carne, necesitan más grano y forraje. Piensan sustituir el maíz por arroz, pero el maíz no está
bien adaptado a inundaciones que son características de estos arrozales japoneses, que son
terrenos planos que no dejan que el suelo se seque.
En Japón las lluvias van de de mayo a julio y son fuertes (150 mm, mensual en promedio, en
Tokyo) y hay tormentas de agosto a octubre (148-216 mm, mensual).
13. Maíz dañado por la inundación. Mano y Omori 2007
Por este razón, unos científicos japoneses están investigando la adaptación a inundaciones de
los parientes del maíz, los teocintles, y de muchas líneas seleccionadas del maíz. El equipo:
Dr. Yoshiro Mano, Fumie Omori, Tadashi Takeda, y otros en el Instituto Nacional de la
Ciencia de Ganadería y Pradera, Nasushiobara, Tochigi, Japan.
7
14. Raíces respondiendo a la inundación, maíz Mi29 (izq.) y Z. nicaraguensis (der.)
Aunque los tallos del Gran Teosinte de Apacunca son impresionantes, las características de
mayor interés en estudios de tolerancia a inundaciones son las de raíces:
Raíces adventicios a la superficie del suelo (sin geotropismo) cuando están inundadas o
muy mojadas. Hay unos maíces con raíces superficiales en tales condiciones.
Aerénquima o canales internos al largo de la raíz.
Tolerancia a gases y sustancias químicas tóxicas en suelos ácidos o hipóxicos.
15. Aerénquima en raíces de un maíz, el teosinte Nica, un hibrido F1 y un segregante
F2, sin inundación
Hay dos genes ya conocido, Adh1 y Adh2, que ayudan en mantener unos maíces en
situaciones de inundaciones temporales; posiblemente están involucrados con la tercera
característica.
8
16. Aerénquima en un maíz, Z. luxurians y dos segregantes F2
El equipo japonés está comparando cómo varían los raíces superficiales y la aerénquima en
varios maíces y teosintes, y en híbridos de la primera generación y las más avanzadas.
Durante el proceso de mejoramiento de varios maíces para que soporten inundaciones, van a
seguir tomando en cuenta el aerénquima y el crecimiento de raíces sobre la superficie del
suelo.
9
Hasta ahora, el teosinte nicaragüense tiene más ventajas que los dos otros de Centroamérica.
¿ Cómo están los japoneses transmitiendo los caracteres de tolerancia a inundación a los
maíces ?
El proceso de seleccionar y avanzar las plantas mejoradas empieza en encontrar una serie de
marcadores que distinguen los padres, el maíz y el teocintle involucrado.
Hace décadas, Mangelsdorf desarrolló un “tester stock” que tiene un marcador fenotípico por
cada uno de los 10 pares de cromosomas. Con esto un mejorador puede ver si un carácter de
interés está asociado con uno o más de los 10 genes marcadores, sea o el ceroso, o el
amarillo, o el dulce. Pero las plantas de este material son débiles y no están adaptados a
muchos ambientes.
Mangelsdorf Tester Stock: bm2; lg1; a1; su1; pr1; y1; gl1; j1; wx1; g1 and r1 on
chromosomes 1 to 10 respectively; brown midrib2, liguleless1, anthocyaninless1, sugary1,
red aleurone1, yellow endosperm1, glossy1, japonica striping1, waxy1, golden plant1,
colored1
Usando esto, Mangelsdorf y Galinat calcularon que varias caracteres morfológicas que
distinguen los maíces de los teocintles se ubican en el cromosoma 4.
En realidad marcadores en gran cantidad y sin efecto fenotípico fueron necesarios para
mejorar al maíz en un gran escala. Ahora hay muchos que han resultado por los esfuerzos de
varios, especialmente la University of Missouri, el Brookhaven National Laboratory y la
compañía Pioneer Hi-Bred.
10
17. Los 10 cromosomas marcado por muchos SSRs (microsatélites) con carácteres
asociados
Entonces, se calcula un mapa de los genes de interés, en este caso cruzando el Z. luxurians
con una línea de maíz adaptado al centro de Japón. Con el mapa genético se puede hacer más
eficiente el proceso de transferencia. También, se puede identificar y eliminar los alelos que
controlan la forma de teocintle en vez del maíz.
Aquí se nota la posición de varios genes o más bien QTLs (loci de características
cuantitativas). Los marcadores son SSRs o microsatélites en las 10 cromosomas de Zea que
distinguen las especias en el experimento.
La transformación genética por retro-cruzamiento asistido por marcadores sigue así:
Crecer 150 o más plantas hibridas F2 o retrocruzadas po unas semanas,
tomar muestras de las hojas y extraer al ADN
por cada planta, amplificar los pedazos de ADN que contienen microsatélites de interés,
analizar cuales pedazos se correlacionen con el efecto de interés para tener un mapa
genético con los QTLs pertinentes,
seleccionar las plantas con los alelos de interés, y dejar crecerlos hasta que expresen el
fenotipo buscado,
hacer retrocruzas de los mejores materiales, y seguir el proceso.
18.
Sugerencias para estudios del Gran Teocintle y preguntas
Durante esta semana (primera de noviembre, 2011), hice algunos apuntes sobre estudios que
se podían realizar a lo inmediato en Nicaragua, pero al llegar a Managua encontré que ya
varios estudios se están desarrollando. Uno de ellos es el estudio del teocintle como forraje
para el ganado: su rendimiento por hectárea y su calidad.
11
El Ing. Benavides me explicaba que al momento él, y otros investigadores están en el proceso
de este estudio, incluyendo análisis del nivel de proteína durante el desarrollo de la planta.
Asimismo, se estaban buscando otros sitios ecológicos cercanos al hábitat del teocintle para
aumentar las áreas de teocintle. Esto ya está en proceso en Apacunca y en la Universidad
Nacional Agraria, además de que se están protegiendo el área actual de plantación de
teocintle.
Se debe continuar los estudios del Ing. Benavides en cuanto a fotoperiodismo usando luces
para interrumpir las noches y jaulas de plástico negro para extender las noches, también
variando la edad de las plantas cuando empieza el tratamiento.
Hacer cruces entre el Gran Teocintle y varios maíces, notando la variación encontrado en la
progenie. Hace décadas Drs. Mangelsdorf, Bianchi y Robertson reportaron que en unos
cruces entre maíz y Z. luxurians muchas mutaciones obvios aparecieron, algo importante para
mis teorías sobre la evolución del maíz y los teocintles.
¿ Qué es la variación genética en la población del Gran Teocintle ? Me parece que ya
tenemos una respuesta con el artículo de 2010 del Dr. Loaisiga y asociados.
¿ Cómo crece el teocintle en varias condiciones de inundación: notando duración, fechas,
profundidades de agua, etc.?
¿ Qué relación hay entre la postración de la planta y el crecimiento de los raíces arriba del
suelo?. Un problema aquí es que hasta ahora la postración no ha sido observada en
Nicaragua, probablemente por el carácter del suelo que mantiene mucha humedad o por unas
químicas especiales en suelos mojados.
19. Descubriendo, caracterizando y utilizando germoplasma de los teosintes
centroamericanos: Referencias a las investigaciones en Japón (en inglés).
Mano, Y., M. Muraki, T. Komatsu, M. Fujimori, F. Akiyama and T. Takamizo. 2002. Varietal
difference in pre-germination flooding tolerance and waterlogging tolerance at the seedling stage in
maize inbred accessions. Japanese Journal of Crop Science 71:361-367. [223 maize lines tested for
tolerance to soaking of seed for 8 days; subsets studied further comparing seedling to pre-germination
tolerance]
Mano, Y., M. Muraki, M. Fujimori and T. Takamizo and B. Kindiger. 2005. AFLP-SSR maps of
maize × teosinte and maize × maize: comparison of map length and segregation distortion. Plant
Breeding 124:432-439.
Mano, Y, M. Muraki, M. Fujimori, T. Takamizo and B. Kindiger. 2005. Identification of QTL
controlling adventitious root formation during flooding conditions in teosinte (Zea mays ssp.
huehuetenangensis) seedlings. Euphytica 142:33-42.
Mano, Y., F. Omori, M. Muraki and T. Takamizo. 2005. QTL mapping of adventitious root formation
under flooding conditions in tropical maize (Zea mays L.) seedlings. Breeding Science 55:343-347.
Mano Y., F. Omori, T. Takamizo, B. Kindiger, R. McK. Bird and C. H. Loaisiga. 2006. Variation for
root aerenchyma formation in flooded and non-flooded maize and teosinte seedlings. Plant and Soil
281:269–279.
abstract
at
http://www.ars.usda.gov/research/publications/publications.htm?SEQ_NO_115=208167
Mano, Y, M. Muraki and T. Takamizo. 2006a. Identification of QTL controlling flooding tolerance
in reducing soil conditions in maize (Zea mays L.) seedlings. Plant Production Science 9:176-181.
Mano, Y. and F. Omori 2007. Breeding for flooding tolerant maize using “teosinte”as a germplasm
resource. Plant Root 1:17-21
Omori, F. and Y. Mano. 2007. QTL mapping of root angle in F2 populations from maize ‘B73’ x
teosinte ‘Zea luxurians.’ Plant Root 1:57-65.
12
Mano, Y, F. Omori, T. Takamizo, B. Kindiger, R. McK. Bird, C. H. Loaisiga, and H. Takahashi.
2007. QTL mapping of root aerenchyma formation in seedlings of a maize × rare teosinte “Zea
nicaraguensis” cross. Plant and Soil 295:103-113.
Mano, Y. and F. Omori 2008. Verification of QTL controlling root aerenchyma formation in a maize
x teosinte “Zea nicaraguensis” advanced backcross population. Breeding Science 58:217-223.
Mano, Y, F. Omori, B. Kindiger and H. Takahashi. 2008. A linkage map of maize × teosinte Zea
luxurians and identification of QTLs controlling root aerenchyma formation. MolecularBreeding
21:327-337.
Mano, Y., F. Omori, C. H. Loaisiga and R. McK. Bird. 2009. QTL mapping of above-ground
adventitious roots during flooding in maize x teosinte “Zea nicaraguensis” backcross population.
Plant Root 3:3-9.
Mano Y, F. Omori and K. Takeda. 2011. Construction of intraspecific linkage maps, detection of a
chromosome inversion, and mapping of QTL for constitutive root aerenchyma formation in the
teosinte Zea nicaraguensis. Molecular Breeding (online 19 November 2010, pp. 1-10.
20. Discubriendo, caracterizando y utilizando germplasma de los teosintes centroamericanos:
Una bibliografía limitada con anotaciones en orden cronológica
Rossignon, M. J. [Jules]. 1869. [letter read at meeting Extraits des? Procès Verbaux]. Bulletin de la
Société d'Acclimatation ser. 2, vol. 6:487-488. ["téozinté" on seed packet of luxurians from Sta. Rosa
sent to Durieu by MJR, Dir. Jardines Publicas . . . Guatemala]
Durieu de Maisonneuve, M.-C. [Michel-Charles] 1872. (untitled note in Extraits des ProcèsVerbaux). Bulletin de la Societe Nationale d'Acclimatation de France, Ser. 2, T. 9 (19):579-581.
[Series 2, Tome IX or vol. 19; definition of Reana luxurians Durieu as highly tillered plant with
maize-like, leafy stems reaching 3 meters, whose seed are enclosed in a thick, stony envelope (nearly
a nomen nudum but clearly the plant renamed as Euchlaena luxurians by Durieu and Ascherson
1876); maybe here gave source: Guatemala with name teozinte]
Collins, G. N. and J. H. Kempton. 1920. A teosinte-maize hybrid. Journal of Agricultural Research
19:1-37. [remarkable study; studied independence of 33 (mostly plant) key traits (6 of ear) in 127 F2
of Florida (Z. luxurians) x Tom Thumb grown at Chula Vista, CA; used punch cards and cardsorting; only paired rows segregated close to Mendelian; correlations in structured matrix! gave 8
groups: height, nodes above, tassel, male branch, alicole, nodes silking, prophyllary and number of
rows, plus 7 independents; 6 for earlets; 5-7 seed/earlet; only single-factor trait: paired ear spiklets –
maize dominant; at http://naldr.nal.usda.gov/NalWeb/HelpJAR.htm search authors/year; see Langham
1940]
Haines, H. H. 1924. The Botany of Bihar and Orissa (India), Part 6:1065. London. [placed
Euchlaena luxurians in Z. mexicana var. luxurians]
Collins, G. N. (with W. Popenoe). 1932. The rediscovery of teosinte in Guatemala. The Journal of
Heredity 23:260-265. http://jhered.oxfordjournals.org/content/23/7/261.full.pdf+html [mostly a letter
from Popenoe]
Kempton, J. H. and W. Popenoe. 1937. Teosinte in Guatemala. Contributions to American
Archaeology 23:200-217. Publication 483, Carnegie Institution of Washington, Washington, DC.
[collections of Z. luxurians & Huehue.; luxurians around Lake Retana - seed ripening in Oct.,
swampy; hybrids; MATs=lux x maize segr.]
Longley, A. E. 1937. Morphological characters of teosinte chromosomes. Journal of Agricultural
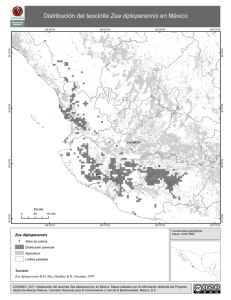
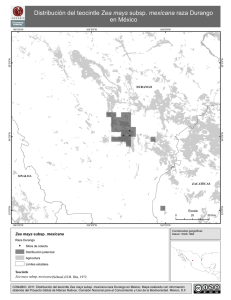
Research 54:835-862. [Z. lux & per, Huehue, Chalco & Durango, Tr. flo & dac - lengths of arms ;
confirms Florida is luxurians; internal (rare) and 7ST & 9ST knobs in Huehue; MAT shortest Kr 1 to
9, lux slightly longer Krs than maz ]
Longley, A. E. 1941b. Knob positions on teosinte chromosomes. Journal of Agricultural Research
62:401-413. [the 2 Guatemalan teosintes; Huehue has 4-5 knobs not in luxurians: (1L), 3L, 7S, 8S,
9S; maize Kr often slightly longer than lux]
13
O'Mara, J. G. 1942. A cytogenetic study of Zea and Euchlaena. University of Missouri Agricultural
Experiment Station, Research Bulletin 341:1-16. [maize x luxurians, maize x Huehue, Huehue x Flo
(luxurians); found 3 probable inversion differences: 4S, 8L, 9S; up to 6 univalents in Huehue x
luxurians; stickiness of luxurians knobs not in Huehue knobs - two types almost repel].
Rogers, J. S. 1950a. The inheritance of photoperiodic response and tillering in maize-teosinte
hybrids. Genetics 35:513-540. [multiple tester maize x (3 MATs, 2 luxurians, 1 Huehue); photoper
linkage to markers: Kr 8, 10 in most, Kr 1, 2, 6 in some; tiller no. linkage to markers: Kr1, 2, 7 in
most, Kr 4, 6, 8, 10 in some; relative dominance discussed].
Rogers, J. S. 1950b. The inheritance of inflorescence characters in maize-teosinte hybrids. Genetics
35:541-558. [multiple tester maize x (3 MATs, 2 luxurians, 1 Huehue), differing results, noted
spikelet pairing, rank number, ear grade and glume score; strong linkages: su1 (Kr 4) to EG & GS; a1,
bm2, lg1 (Kr 1-3) to 2-rank; none to paired; may confuse 1° & 2° earlets].
Reeves, R. G. 1953c. Comparative morphology of the American Maydeae. Texas Agricultural
Experiment Station Bulletin 761:1-26. College Station, Texas. [24 traits compared across tripsacum,
Z. luxurians, Mex. annual teosinte & maize]
Ting, Y.-C. 1966. Spontaneous chromosome inversions of Guatemalan teosintes (Zea mexicana).
Genetics 36 (1965):229-242. [only Z. luxurians., Jutiapa, Lake Retana & Florida; on 9S (all plants),
1L-Jut., 3L-Flo. In9 ~60% of short arm; In3 ~35%; In1 ~22%]
Wilkes, H. G. 1967. Teosinte: the Closest Relative of Maize. The Bussey Institution of Harvard
University, Cambridge, MA. 158 p. [Ph.D. dissertation, suggested model of introgression between
sections, extensive bibliography]
Bird, R. McK. 1978a. A name change for Central American teosinte. Taxon 27:361-363. [Z.
luxurians renamed]
Doebley, J. F. and H. H. Iltis. 1980. Taxonomy of Zea (Gramineae). I. A subgeneric classification
with key to taxa. American Journal of Botany 67:982-993. [define sections Luxuriantes and Zea].
Robertson, D. S. 1983. Genetic instability in maize-teosinte hybrids. Communicated in Maize
Genetics Cooperation News Letter 57:6. [the teosinte was Z. luxurians; seen in F1 and BC1 so
dominants]
Doebley, J. F., M. M. Goodman and C. Stuber. 1984. Isoenzymatic variation in Zea (Gramineae).
Systematic Botany 9:203-218. [12 enzymes, 21 loci, all teosinte populations, some sympatric maize –
4 peren. not same as 7 dip.; 6 lux; 5 maize more like 18 parv than 19 mex; 2 Huehue unlike others]
Smith, J. S. C., M. M. Goodman and C. W. Stuber. 1984. Variation within teosinte. III. Numerical
analysis of allozyme data. Economic Botany 38:97-113.
Laurie, D. A. and M. D. Bennett. 1985. Nuclear DNA content in the genera Zea and Sorghum.
Intergeneric, interspecific and intraspecific variation. Heredity 55:307-313. [maize: 9.8-13.5 pg,
MATs: 10.5-12.9 pg, Huehue: 12.2, Z. luxurians 18.3-18.5, Z. diploperennis: 10.6, Z. perennis: 21.1;
lowest maize: Seneca 60 sweet, highest maize: Zapalote Chico]
14
Doebley, J. F. 1989b. Molecular evidence for a missing wild relative of maize and the
introgression of its chloroplast genome into Zea perennis. Evolution 43:1555-1559.
[restriction of cpDNA w/ 21 endonucleases, 10 probes, 42 accessions 2 Trip spp, 3 lux, 3 dip,
3 per, 2 hue, 8 mex, 5 parv, 15 maz, but no passport data; 43 restriction-site polymorphic
sites; tree graph: T-Z 18 sites, Tp-Td 6 s, lux-p/d 3-4 s, L-Z 17-20 s, hue-{maz/mex 0-1s, par
1 s}, par-maz 1-2 s; Z. mays in 5 clades: mex, mex-parv, parv, mex-hue-mays, mays; Z per
from Piedra Ancha (Iltis et al. 1050) has 8 of 10 plants with cpDNA unlike per/dip. and
mostly like Z. mays but diff. – introgression].
Doebley, J. F. 1990c. Molecular systematics of Zea (Gramineae). Maydica 35:143-150.
[renames huehuetenangensis as a ssp of Z. mays; says it "may be of hybrid origin [or] . . .
may have experienced some form of genetic revolution. in a very short period of time" p.
147; cpDNA: 80 acc; Fig. 4 T-Z Δ 19 mut, sLux-sZea 8 mut in each of 2 “internodes”, luxper 2/internode, per-dip Δ 1; 5 sZea clades Δ 1-2 mut, maz in ABDE, prv in BCDE, mex in
ABD, hue in D; in D: 1 prv, 4 mex, 36 maz, 2 hue, in B: 22 mex, 15 prv, 8 maz; isoz: 99 Mex
& Guat acc; 3 pop of prv: Jalisco, Southern Guerrero, Central Balsas (Telo, Huet, M-M) see
Matsuoka et al. 2002].
Newton, K. J. and K. M. Courtney. 1991. Molecular analyses of mitochondria from teosintecytoplasm-associated miniature plants. Maydica 36:153-159. [Z. diplo., lux., peren].
Goloubinoff, P., S. Pääbo and A. C. Wilson. 1993. Evolution of maize inferred from
sequence diversity of an Adh2 gene segment from archaeological specimens. Proceedings of
the National Academy of Sciences, U.S.A. 90:1997-2001. online [possible "that. the gene
pool of maize is millions of years old and that domestic races of maize stem from several
wild ancestral populations" p 1997; Adh2 of 5 modern, 3 ancient maizes (440, 1500,
(4500=1300) yBP), 2 MATs, lux, dip, Trip pilosum; all cloned so get both alleles per plant,
ca. 315 bp; 52 bp upstr, exons 1 & 2, intron A & 9 nt of B; counted synon+intron+upstr SNPs
for genetic dist calc; SNP differences: T-Z 7-14, L-Z 0-11, lux-maz 1-10, dip maz 0-11, mazmaz 1-12, maz-MAT 1-11; 2 haplotype blocks in maz (3-4 vs. 5-6) 8 SNP diff, in 1 lux plant
diff = 6 SNP, in one MAT plant diff = 10 SNP].
Benavides G., Alvaro and Carlos H. Loaisiga C. ms. Segunda Gira. Prospección y
recolección del teocintle anual (Zea luxurians D.) en la región de Villanueva, Apacunca,
Nicaragua. Programa de Recursos Genéticos Nicaragüenses, Universidad Nacional Agraria,
Managua. Marzo, 1997. 6 pg. Mimeographed.
Kato Y., T. A. 1998. Caracterización cromosómica. In: Sánchez G., J. J., T. A. Kato Y., M.
Aguilar S., J. M. Hernández C., A. López R. and J. A. Ruíz C. 1998. Distribución y
Caracterización del Teocintle. Instituto Nacional de Investigaciones Forestales, Agrícolas y
Pecuarias (INIFAP), Guadalajara, Mexico. [complex analyses & comparisons 64 Mexican
teosintes, 3 hue, 4 lux]
Provan, J., P. Lawrence, G. Young, F. Wright, R. Bird, G. Paglia, F. Cattonaro, M. Morgante
and W. Powell. 1999. Analysis of the genus Zea (Poaceae) using polymorphic chloroplast
simple sequence repeats. Plant Systematics and Evolution 218:245-256. [27 Latin American
maizes and 10 teosintes, 15 SSR loci 80-196 bp long by PCR, MATs close to one of two
maize groups (Canilla & Chulpi odd), Huehue very different; maize split = ca 50% of LuxZea split, lacks race labels, uses Euclidean distance]
Bird, R. McK. 2000. A remarkable new teosinte from Nicaragua: Growth and treatment of
progeny.
Communicated in Maize Genetics Cooperation Newsletter 74:58-59.
http://www.maizegdb.org/mnl/74/74bird.html [anyone may cite]
15
Iltis, H. H. and B. F. Benz. 2000. Zea nicaraguensis (Poaceae), a new teosinte from Pacific
coastal
Nicaragua.
Novon
10:382-390.
http://biostor.org/cache/pdf/13/23/b6/1323b6cc4c1a50bf838d4ad9e88e235c.pdf
[mainly
grows up to 5m tall, on floodplain at 9 masl, away from cultivated fields, flooded often and
deeply; closest to Z. luxurians; with Alfredo Grijalva]
Loáisiga, C. H. 2002. Caracterización agro-socio-económica del hábitat original del teocintle
(Zea nicaraguensis) en Nicaragua, Ministerio del Ambiente y los Recursos Naturales
(MARENA). Managua, Nicaragua.
Benavides G., A. 2002. Caracterización y evaluación ex situ de una población de teocintle
anual (Zea nicaraguensis Iltis & Benz) recolectada en el norte de Chinandega, Nicaragua. La
Calera,
2:6-13.
Universidad
Nacional
Agraria,
Managua.
http://www.una.edu.ni/diep/calera/dowload_pdf/Calera_Inv-SGI46_Num-_Ano-2002.pdf [29
variables; plantings Mar-Sep 1996, statistical analysis of variable to environment, 10-300
ears/plant, 4-5 seeds/ear, rachid leng. 7-10 mm; pl ht: Mar 340 cm, Apr 380, May 350, Aug
200, Sep 150 cm; SM: sin malezas, CM: con malezas]
Palaisa, K. A., M. Morgante, M. Williams and A. Rafalski. 2003. Contrasting effects of
selection on sequence diversity and linkage disequilibrium at two phytoene synthase loci.
The Plant Cell 15:1795-1806. http://www.plantcell.org/content/15/8/1795.full [ Y1 (and
PSY2); 75 white & yellow inbred lines + 5 teosinte taxa (PI 11083 – Nic mislabeled Z. lux);
85 non-coding SNPs, 21 coding, 32 indels; color figure of SNP linkage blocks with bipolar
pattern; yellow maize 1 group w/ low diversity; 34 white maizes in 4 groups, Gp 1: 36 maz,
Gp 2: 7 maz, 1 mex & 1 parv, Gp 3 variable 15 maz, Gp 4: 1 maz & 3 mex; 5 th white group 2 lux, 2 per, 2 dip & 1 parv; distances within 2 teos groups much less than between 4 white
maize and 1 yellow (n=31) groups].
Aragón Cuevas, F. 2006. Nueva poblacion de teocintle en Oaxaca. XXI Congreso Nacional
y Primero Internacional de Fitogenética, 3 al 8 de septiembre de 2006, Tuxtla Gutiérrez,
Chiapas, Mexico. [lux in Oax see Sanchez et al 2011; he coll’d it in 2004].
Ellneskog-Staam, P., C. H. Loáisiga and A. Merker. 2007. Chromosome C-banding of the
teosinte Zea nicaraguensis and comparison to other Zea species. Hereditas 144:96-101.
doi: 10.1111/j.2007.0018-0661.01989.x [C-banding; in Z. nicaraguensis 20 Kr; knobs: nic
term: S & L of Kr 1-3, L of Kr 4-9, sum: 12; lux term: S & L of Kr 1-3, L of Kr 4-10, sum:
13; dip term: S & L of Kr 1-2, L of Kr 4,6,9, S of Kr 5,8, sum: 9; maiz (Birko, Fr) term: S
& L of Kr 1, S of Kr 5, 7; int: L of Kr 2, 4, 8, 9, S of Kr 6, sum: 9; at variance with others].
*Loáisiga, C. H., A. K. Brantestama, O. Rocha, B. Salomon and A. F. Merker. 2010.
Genetic diversity and gene flow in the Meso-American teosintes. Genetic Resources and
Crop Evolution **
Loáisiga C., C. H. 2011. Cytogenetics, genetic diversity and phylogenetics of wild Zea
species, with emphasis on Zea nicaraguensis. PhD. Dissertation. Acta Universitatis
Agriculturae Sueciae, ISSN 1652-6880. Vol. 2011:41, 53 p. Sveriges Lantbruksuniversitet,
Alnarp, Skåne County, Sweden. http://pub.epsilon.slu.se/8148/1/loaisiga_C_110524.pdf
accessed 30 Sep 2011 [review of teos literature; # accessions: nic 1, lux 1, hue 1, per 2, dip 2,
parv 1, maz 1; Trip dac 2; all for seq of 8 cpDNA regions w/ Sorg bic; all teos for 33 morph
traits & 21 SSR markers; C-bands of nic, lux, dip & maz; C-bands: nic like lux with term
bands on short & long arms of Kr 1-9 unlike dip & maz (both only 9 detectable); nic avg’d
longest Kr, close to lux.
16
Loáisiga, C. H., A. K. Brantestama, O. Diaz, B. Salomon and A. F. Merker. 2011. Genetic
diversity in seven populations of Nicaraguan teosinte (Zea nicaraguensis Iltis et Benz) as
estimated by microsatellite variation. Genetic Resources and Crop Evolution 58:217-223.
Sánchez G., J. J., L. De La Cruz L., V. A. Vidal M., J. Ron P., S. Taba, F. SantacruzRuvalcaba, S. Sood , J. B. Holland, J. A. Ruíz C., S. Carvajal, F. Aragón C., V. H. Chávez T.,
M. M. Morales R., and R. Barba-González. 2011. Three new teosintes (Zea spp., Poaceae)
from México. American Journal of Botany 98:1537-1548. Online prepublication Sep 2011
http://xa.yimg.com/kq/groups/22176678/1323701150/name/SanchezEtAl2011_3newTeosinte
s.pdf
[A. perennial diploid population from Nayarit: early maturing plants and male
inflorescences with few tassel branches and long spikelets. B. perennial tetraploid population
from Michoacán: tall and late maturing plants and male inflorescences with many branches.
C. annual diploid population from Oaxaca (see Aragón-Cuevas 2006): male inflorescences
with fewer branches and longer spikelets than those found in the sister taxa Z. luxurians and
Z. nicaraguensis, plants with high thermal requirements, and very long seed dormancy, 17°
56′ 37 ” N, 96° 27′ 43” W, 80 masl, 8.8 km NE of San Felipe Usila, 76 km ESE of Coxcatlan.
Comparison using morphological, ecological, ploidy, and DNA markers of 20 teos, 15 pl/acc,
18 nrSSR loci ]. [email protected] in 2005 GoogleEarth.
17
Estado actual del teocintle (Zea nicaraguensis ILTIS & BENZ) en la
Reserva de Recursos Genéticos de Apacunca (RRGA)
Ing. MSc. Álvaro Benavides González
Objetivos del Proyecto:
1. Involucrar a las comunidades en el rescate, conservación, uso del teocintle y asegurar
que reciban algún beneficio.
2. Promover en las comunidades de Somotillo y Villanueva la preservación de los
hábitats para proteger la diversidad genética del teocintle, de las especies animales y
vegetales asociados.
3. Caracterización molecular y morfológica in situ de las poblaciones de teocintle.
4. Fortalecer mecanismos de coordinación entre el sector privado, instituciones no
gubernamentales, centros de investigación y la sociedad civil.
18
El DEPARTIR en Asamblea con familias en Apacunca.
El DEPARTIR desde finales del 2006 se dio a la tarea de buscar fondos adicionales a los
aportados por la UNA y con dichos fondos ha logrado tres procesos de intervención en
nueve comunidades rurales del país: El Castillito, El Pegador, Nueva Esperanza y Buena
Vista (municipio de Las Sabanas, departamento de Madriz); Los Ángeles, Malacatoya
(municipio de Granada, departamento de Granada); y comunidad de Apacunca, El Circuito,
El Jicote y Aquespalapa (municipios de Somotillo y Villanueva, departamento de
Chinandega).
La vivencia de estudiantes y profesores a lo largo de 5 años en las comunidades antes
mencionadas, ha permitido desarrollar y adaptar metodología y técnicas que facilitan
conocer el estado de la agricultura en las comunidades, el estado de la biodiversidad, y el
estado socioeconómico de las familias que conforman la comunidad. La unidad
fundamental de estudio es la Unidad Familiar Productiva (UFP), que integra a la familia, la
finca, la infraestructura de la comunidad, los recursos naturales de la comunidad.
La metodología permite analizar la UFP y la comunidad en su conjunto y permite a la
Universidad promocionar valores, actitudes y el desarrollo de técnicas y actividades
investigativas que fortalezcan a la comunidad y las familias campesinas. Por tanto, la
metodología desemboca en la diversificación y el establecimiento de modelos de
producción sostenibles, que a la vez signifiquen una mejora de calidad de vida de la
comunidad y de las familias.
19
El proceso de intervención de la UNA en la comunidad rural es un proceso participativo, de
abajo hacia arriba, en el cual la familia en su finca es el punto de partida, para la participación
activa de mujeres, niños y hombres de la familia campesina en la gestión de su propio
desarrollo. Son el productor y la productora quienes dinamizan el proceso de identificación de
problemas y soluciones e implementan las alternativas seleccionadas.
Para ello, se imparte un curso de capacitación a estudiantes y profesores que garantice
relaciones horizontales Campesino-Estudiantes-Docentes.
Propuestas conformadas a partir de la canasta de problemas encontradas en Apacunca:
1. Talleres de capacitación sobre Buenas Prácticas Agrícolas
2. Indicadores de calidad del suelo para mejorar las propiedades físicas y de fertilidad
3. Tácticas de manejo de ratas mediante el fomento de depredadores, uso de trampas,
eliminación de madrigueras y uso de raticidas naturales
4. Estudio etnobotánico y de prospección de especies originarias de la Reserva de
Recursos Genético de Apacunca
5. Práctica de campo de ensilaje de pastos y teocintle
6. Sistema de perforación de pozo
7. Manejo alternativo de plagas y enfermedades mediante la crianza depredadores,
naturales, prácticas culturales, etc.
8. Tácticas y alternativas para el manejo de gallina ciega.
9. Tacticas y alternativas para el manejo de mosca blanca
10. Crianza de patos
11. Establecimiento de parcela de reprodución de teocintle
12. Identificación de factores que ocasionan alto porcentaje de aborto en el ganado
Trabajos de Investigación que se desarrollan en Apacunca:
1. Caracterización in situ y ex situ del teocintle
2. Inventario básico de especies asociadas al teocintle
3. Estudio básico de etnobotánica y etnozoológico de especies
4. Diagnóstico de plagas y enfermedades asociadas al teocintle
5. Caracterización molecular básica del teocintle
6. Caracterización morfológica básica de especies silvestres vegetales
Biodiversidad
La Reserva de Recursos Genéticos de Apacunca (RRGA) en Chinandega, Nicaragua, fue
creada en 1996. Representa un reservorio de genes y fuente de variabilidad genética
interesante para la mejora genética de algunas especies cultivadas. Asimismo, se
encuentran especies animales de alto potencial, y un alto potencial ecoturístico.
20
Biodiversidad de Apacunca
El tonkuá (Benincasa hispida (thunb)) es una cucúrbita que los pobladores la utilizan para
hacer un dulce de excelente sabor que tiene una gran demanda en el mercado. A través del
proyecto FAO/Apacunca, se realizó día de campo y que un especialista capacitó a 10 mujeres
integrantes del proyecto sobre la preparación de este dulce.
Frutos de Tonkuá en manos del Sr. Cristobal Aguilera, municipio de Aquespalapa
21
Características del hábitat y suelo del teocintle de Nicaragua
22
El teocintle de Nicaragua
La variabilidad genética en los parientes silvestres de las plantas cultivadas representa el
reservorio para la mejora genética. Muchas de estas especies silvestres a partir de las cuales
evolucionaron las actuales plantas cultivadas, aún sobreviven en condiciones naturales, y
otras están en peligro de extinción. El maíz como especie mesoamericana está emparentado
con especies silvestres y/o malezas conocidas como teocintle.
Se conocen cuatro razas de teocintles en México y dos en Guatemala, una de las razas de
Guatemala fue clasificada como Zea luxurians [(Durie & Ascherson) Bird] y reportada en
Honduras y Nicaragua. Estudios realizados por Iltis & Benz concluyen que el teocintle anual
de Nicaragua tiene algunas características a los otros teocintles, por lo que lo clasificaron
como Zea nicaraguensis.
En la Mesa Central de México y Honduras el teocintle desapareció, y las poblaciones del sur
de Guatemala están en peligro. Asimismo, en Nicaragua las poblaciones reportadas en
Apacunca, Somotillo, son vulnerables. Lo anterior lleva a plantear la necesidad de estudiar y
establecer un plan de manejo del teocintle en Nicaragua para conservar el reducto que aún se
encuentra en la zona de Apacunca.
El teocintle:
del Nahualt, Teoxintli
teotl divinidad o cosa sagrada
centli espiga de maíz
Nombres comunes en Nicaragua:
Teosinto, tiosinto, teosinte, teocintle, maíz de playa, maíz de costa, maíz de laguna, maíz
forrajero, maíz de pájaro, maíz silvestre, maicillo.
Usos comunes:
Pasto natural, cazar animales, enramadas (secar maíz), viviendas temporales.
Los primeros estudios de caracterización del teocintle en Nicaragua se realizaron en 1996
en la UNA, para este año la especie fue clasificada de manera preliminar como Zea
luxurians, ya que se creía era la continuación de las poblaciones de Honduras (hoy extinta)
y procedente de Guatemala. Otros estudios revelaron que el teocintle de Nicaragua difería
de los otros teocintles.
Z. nicaraguensis, difiere de teocintles del área mesoamericana por los siguientes aspectos:
Se encuentra a una altitud entre 0 y 10 msnm en la depresión de Apacunca.
La mayoría de las plantan superan los 5 m, más de 5 cm de diámetro en tallo, longitud
de hoja superior a 100 cm y raíces prominentes.
Longitud de panojas de más de 50 cm, esto indica que los descriptores que conforman
la panoja son mayores que en otros teocintles mesoamericanos.
Tiene de 5 a 8 rugosidades transversales bien pronunciadas en las glumas,
especialmente cuando están jóvenes.
La forma de la semilla es trapezoidal (casi romboide) y la parte central presenta forma
circular (transversalmente).
Mayor longitud de semilla, y frecuentemente tiene 5 ó 6 cariopsis por “mazorca”.
23
El teosinto de Nicaragua en su hábitat natural (El Papalona, Apacunca)
La distribución del teocintle en Nicaragua está limitada al occidente del país, departamento
de Chinandega. En la actualidad esta especie se encuentra en la comunidad Cayanlipe (12°
52' 25" N, 86° 54' 50" O, 15 msnm) y Apacunca (12° 53' 45" N, 86° 59' 00" , 9 msnm) en
los municipios de Villanueva y Somotillo, respectivamente.
La población de Apacunca se encuentra a 3.5 km al noreste de la hacienda que lleva el
mismo nombre, específicamente en El Papalonal (Apacunca) a la orilla del río Bocana que
es un afluente del estero Palo Grande. El área de El Papalonal se encuentra dentro de un
área de 42 hectáreas propiedad del Señor Oscar Martínez. Actualmente es la población más
grande y abarca una hectárea. Dicha población puede variar en área cada año y está
condicionada por el pastoreo del ganado, quema del pasto y condiciones ambientales,
principalmente.
24
Innundaciones en el hábitat del Teocintle de Nicaragua, en épocas de lluvias
Nomenclatura usada en la clasificación de teocintle y maíz (Kato et al., 2009)
Wilkes (1967)
Iltis & Doebley (1980)
Doebley (1990a)
Doebley (2003)
Sección ZEA
Zea mays L.
Sección ZEA
Zea mays L.
Sección ZEA
Zea mays L.
Sección ZEA
Zea mays L.
Sección EUCHLAENA spp. mexicana
Euchlaena mexicana
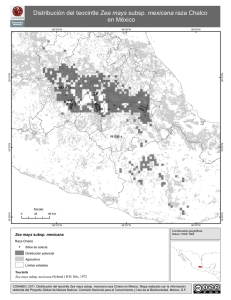
Raza Chalco
spp. mexicana
spp. mexicana
Sin. Zea mexicana
Raza Chalco
Raza Mesa Central
Raza Nobogame
Raza Mesa Central
Raza Nobogame
Raza Nobogame
Raza Mesa Central
Raza Nobogame
ssp. parviglumis
var. huehuetenangensis
Raza Balsas
ssp. huehuetenangensis
ssp. parviglumis
ssp. huehuetenangensis
Raza Balsas
Raza Huehuetenango
Raza Guatemala
ssp. mays
Sección
LUXURIANTES
Zea luxurians
ssp. mays
Sección
Sección LUXURIANTES LUXURIANTES
Zea luxurians
Zea luxurians
Zea perennis
Zea perennis
Zea perennis
Zea perennis
Zea diploperennis
Zea diploperennis
Zea diploperennis
Raza Chalco
Zea nicaraguensis
25
Investigaciones sobre teocintle en Nicaragua
In situ
Ex situ
Relación de teocintles mesoamericanos (Benavides, 2002)
6.00
Z. perennis_a
Num_Hijo
Vol_100S
Nud_APRL
X23
Num_SeMz
X21 X22
X26
X1
Anc_Glu
3.00
Long_RPP
Z. huehuetenangensis
X25
CP 2 (25.2%)
X16
Z. perennis_b
Pes_100S
X32
Nud_MzHij
X5
0.00
Long_Hoja
Z. nicaraguensis
Z. luxurians
Alt_Pla
Z. diploperennis
X19
X30
Nud_MzRL
X12
X20
X9
X31
Anc_Hoja
-3.00
X15
Z. parviglumis
X6
X7
X14
X17
-6.00
-5.00
-2.50
0.00
2.50
CP 1 (41.3%)
Relación de teocintles mesoamericanos (Benavides y Loáisiga, 2010)
26
5.00
Los resultados obtenidos del presente estudio, permiten hacer las consideraciones siguientes:
Es posible agrupar y relacionar teocintles utilizando técnicas de taxonomía numérica.
Algunas variables de tallo, hojas, inflorescencia y semillas aportaron la mayor
variación para diferenciar teocintles de Mesoamérica.
En base a características morfológicas, los teocintles de México difieren de los
teocintles de Guatemala y Nicaragua. Z. luxurian de Guatemala y Z. nicaraguensis
presentaron características morfológicas semejantes, y a su vez estos comparten
semejanzas con Z. huehuetenagensis de Guatemala.
Z. huehuetenangensis
Z. nicaraguensis
Z. luxurians
Z. perennis_b
Z. parviglumis
Z. diploperennis
Z. perennis_a
0.00
19.57
39.13
58.70
Relación de teocintles mesoamericanos (Benavides y Loaísiga, 2010)
27
78.26
PROBLEMÁTICA
Desastres naturales
1
Daños ocasionados a la infraestructura vial que conduce a la Reserva de Recursos Genéticos
de Apacunca, durante la temporada de lluvias del año 2010 y 2011.
28
Reserva en áreas de productores
2
Georeferenciación de parcelas en la RRGA (Querol, 2010)
29
3
Avances del área de pastizales para la crianza de Ganado Bovino en el área de la Reserva de
Recursos Genéticos de Apacunca
4
Pérdida del bosque primario en el área de la Reserva de Recursos Genéticos de Apacunca, para
la siembra de caña de azúcar y pastos mejorados.
30
6
Cacería no controlada de reptiles y aves dentro del en el área de la Reserva de Recursos
Genéticos de Apacunca.
7
Contaminación ambiental con desechos sólidos proveniente de la parte alta de la cuenca del rio
Villa Nueva.
31
8
?
Teocintle
90
mz
500
mz
Avance de la frontera agrícola (cultivo de la caña de azúcar), dentro de la Reserva de Recursos
Genéticos de Apacunca. Círculo redondo en rojo es un área próxima a sembrar el cultivo de
caña de azúcar.
Necesidades
Fortalecer mecanismos de coordinación entre el sector privado, instituciones no
gubernamentales, centros de investigación y la sociedad civil.
Elaboración de convenios de colaboración con propietarios privados en cuyos
terrenos se encuentren poblaciones de teocintle
32
Conservación del teocintle
(Zea nicaraguensis
ILTIS & BENZ)
En parcelas de pequeños productores
En la Universidad Nacional Agraria
Conservación y establecimiento de áreas teocintle en la Universidad Nacional Agraria, y en
fincas de productores ubicado en el al área de amortiguamiento. Productor Antonio García.
33
Las áreas protegidas y los Recursos Genéticos
Ing.MSc. Edilberto Duarte
El Gobierno de Reconciliación y Unidad Nacional priorizando los temas y acciones de
conservación de la Madre Tierra y el Bienestar de la Humanidad y como parte integrante
del Plan Nacional de Desarrollo Humano (PNDH), la “Declaratoria universal del bien
común y los derechos de la Madre Tierra y la Humanidad.” y las directrices de la
Convención de Diversidad Biológica (CDB). Por lo cual el ministerio esta subdividido de
la siguiente manera:
De igual manera se está trabajando en diversas estrategias y planes de acción entre
los cuales tenemos:
1.
2.
3.
4.
5.
6.
7.
Conservación y uso sostenible de la biodiversidad
Viabilidad económica del uso sostenible de la biodiversidad
Fortalecimiento de la información y monitoreo de la biodiversidad
Gestión institucional y coordinación interinstitucional
Armonización de políticas y marco jurídico-normativo
Educación y participación ciudadana
Uso y valoración de los recursos genéticos
34
El SINAP fue creado a través de la Ley General del Ambiente (Ley 217) mediante el Arto.
17. En la actualidad existen 72 áreas protegidas en diferentes categorías de manejo:
Entendiendo por diversidad biológica toda aquella, variabilidad de organismos vivos de
cualquier fuente, incluidos, entre otras cosas, los ecosistemas terrestres y marinos y otros
ecosistemas acuáticos y los complejos ecológicos de los que forman parte; comprende, la
diversidad dentro de cada especie, entre las especies y de los ecosistemas.”
35
La biodiversidad en Nicaragua es un tema de relevancia estratégica para el país, en este
sentido, el gobierno, desarrolla acciones en todos los frentes con el propósito de restaurar
zonas que han sido degradadas, la conservación de zonas que se mantienen en excelentes
condiciones y proteger importantes recursos naturales, la biodiversidad así mismo fomenta
y facilita la investigación científica por su connotación cimera en el aporte de los criterios
para la formulación de políticas públicas, estrategias y regulaciones que conllevan a la
conservación y protección de la biodiversidad, además, se realizan intercambios del
conocimiento a nivel nacional e internacional, lo que indica el amplio alcance de una
voluntad política del gobierno hacia la madre tierra
Por ejemplo en el Mapa actualizado de Ecosistemas y Formaciones vegetales de Nicaragua
se identifican un total de 68 clases de ecosistemas, con la diferencia que ahora se ha
realizado un aglomerado de 44 ecosistemas. Además de incluir 5 ecosistemas acuáticos
(Embalse, Estuario, Laguna costera).
36
Taxas
Mamiferos
Aves
Reptiles
Anfibios
Peces
Plantas superiores
Corales
Moluscos
Insectos
Crustaceos
Rotiferos
Nicaragua
225
706
163
76
698
58
58
1908
8514
88
57
Mundo
4.3
9.7
6.6
4.0
22
240
N:D
60
1,200,000
4.0
N:D
%
5.8
6.9
2.62
1.55
2.92
2.71
N:D
3.33
0.71
2.2
N:D
En el pacifico de Nicaragua encontramos los siguientes ecosistemas:
Bancos de moluscos como concha negra (Anadara utberculosa y Anadara similis) y Barba
de hacha (Mytella guayanensis), Zonas de fondos lodosos asociados a manglares en áreas
estuarinas donde la abundancia de especies de bivalvos comerciales es notable. áreas de
anidación de tortugas carey (Eretmochelys imbricata), áreas de anidación de tortugas verde
o torita (Chelonia mydas agassizy), áreas de anidación de tortugas paslama (Lepidochelys
olivacea), Bancos de moluscos como concha negra (Anadara utberculosa y Anadara
similis) y Barba de hacha (Mytella guayanensis), Zonas de fondos lodosos asociados a
manglares en áreas estuarinas donde la abundancia de especies de bivalvos comerciales es
notable, áreas de anidación de tortugas carey (Eretmochelys imbricata), áreas de anidación
de tortugas verde o torita (Chelonia mydas agassizy), áreas de anidación de tortugas
paslama (Lepidochelys olivacea), Áreas de anidación de tortugas tora (Dermochelys
coriácea), áreas de congregación y/o reproducción de aves marinas y playeras, áreas de
congregación de pargos y meros (Lutjanus sp. Y Epinephelus sp.), áreas de crecimiento de
camarones y otras especies de la familia Penaeidae. Áreas de congregación de tiburones y
otras especies pelágicas, áreas de agregación de cocodrilos y lagartos (Crocodrylus acutus
y Caiman rocodilus).
37
38
39
La importancia de los recursos genéticos silvestres y el rol de la UNA en
su protección y aprovechamiento
Ing. MSc. Daniel Querol
Introducción
Algunas ideas alrededor de 30 años cambiantes
Cuál es la importancia de estos recursos en Nicaragua y
Cuál es la función de la Universidad y del programa recursos genéticos
Antecedentes
Los 2 millones de años como recolectores, nos dejan marcados. Colectando y
colectando, Ardillas y museos
10,000 como sedentarios y agricultores y criadores de animales
Cuanto rato dando vueltas recolectando?
Algunos términos y conceptos sobre recursos genéticos y su evolución:
el germoplasma
los recursos genéticos 70s, tesoro olvidado
las especies útiles 80s, pueblos indígenas recolectores, extractivistas
la biodiversidad 90s
Carbon Sink !!!! la banalización
(Demasiado cuantificado en dinero y cifras)
40
Donde estaban la UNA y el REGEN
Para los agrónomos no ha cambiado el concepto de utilización en la línea
Paisaje
domesticación
cultivo
de especie
(variedad)
Recolección
Clasificación
Almacenamiento
En ciertos casos caracterización
Post-REGEN, Álvaro Benavides y DEPARTIR
Usos de los recursos genéticos silvestres en Nicaragua
en su configuración paisaje: servicios ambientales y turismo
en su configuración en proceso de domesticación: novedades como guillas, guillitas,
camarones y bejucos y teocintle
en su configuración cultivo (variedad): Raicilla y en Teocintle escucharemos al
siguiente expositor
Usos de los recursos genéticos silvestres
en su configuración paisaje: servicios ambientales y turismo
Usos de los recursos genéticos silvestres
En proceso de domesticación: novedades como guillas y camarones
Usos de los recursos genéticos silvestres
41
en su configuración en proceso de domesticación: novedades como guillitas, bejucos
y teocintle
Usos de los recursos genéticos silvestres
en su configuración cultivo(variedad) escucharemos al siguiente expositor en el caso
Teocintle
Raicilla
Tomate silvestre
Ideas para el final de la tarde
No contamos con un estado con visión científica a largo plazo (ejemplos con
meteorito y con REGEN)
No debiera existir una contradicción operativa entre conservación y desarrollo ya que
son acciones complementarias. Los modelos de conservación aplicados son los que
tienen intrínseca una deformación al incorporar un proceso de exclusión humana si no
es empresarial.
Debemos analizar que nuevos mecanismos (incluyendo los monetizados como el
comercio de carbono y REDD) pudieran servir como alternativas a los procesos
estatales y no gubernamentales que se han demostrado bastante poco eficientes
El REGEN o el equipo DEPARTIR podría conformar un observatorio que alerte ante
peligros de desaparición o merma de la biodiversidad silvestre o cultivada,
movilizando apoyo científico, político y económico.
42
Diversidad genética y evolución del maíz silvestre de Nicaragua:
Teocintle (Zea nicaraguensis Iltis & Benz)
Dr. Carlos Henry Loáisiga Caballero
Introducción
Los centros de diversidad de especies silvestres emparentadas con especies cultivadas son
reservorios de importante germoplasma para el mejoramiento genético. Estos pueden contener
importantes fuentes de resistencia o tolerancia en contra de plagas, enfermedades y factores
abióticos (Frankel 1984).
La región Mesoamericana, del cual Nicaragua forma parte, es uno de esos centros con alta
diversidad genética en muchas especies tales como; maíz (Zea mays L.), frijol común (Phaseolus
vulgaris L.) Pitahaya (Hylocereus undatus (Haworth) Britton et al Rose), Cucúrbitas (Cucurbita
spp) Cacao (Theobroma cacao L.) Aguacate (Persea americana L.) y Achiote (Bixa orellana L.).
La Figura1, muestra la región mesoamericana así como la distribución de los teocintles.
Figure 1. Distribución de los teocintles en la región mesoamericana.
Zea mays L. ssp. parviglumis (Teocintle o teosinte) es el más cercano pariente y probablemente el
progenitor del maíz moderno (Zea mays L. ssp. mays (Doebley et al. 1984), ambos pertenecen al
género Zea, el cual está compuesto por cuatro especies y cuatro subespecies, la siguiente tabla
describe la clasificación según Doebley 1990.
Accession
9476
8837
9475
9479
9477
9478
4290
Origen
México
México
México
Guatemala
México
Guatemala
Nicaragua
Nombre de accession
Las Joyas
Zea perennis
Piedra ancha
Teosinte, huehuetenagensis
Teosinte, Las Balsas
Teosinte
Teocintle
43
Especies
Zea diploperennis1
Zea perennis1
Zea perennis2
Zea mays ssp. huehuetenangensis
Zea mays ssp. parviglumis
Zea luxurians
Zea nicaraguensis
Altitud
1950
940
1600
1300
850
800
10
Recientemente (1990), en Nicaragua se descubrieron varias poblaciones de teocintle que crece casi
al nivel del mar, siendo el único germoplasma entre los maíces silvestres en la región que crece a
nivel del mar, suelos completamente inundados de agua y alcanza hasta seis metros de altura
(Loáisiga, 2011).
Diversidad genética
Estudios posteriores utilizando marcadores moleculares (microsatelites) se compararon los
teocintles de la región y los resultados mostraron que todos los marcadores fueron polimórficos y
detectaron 109 alelos.
En general, el número de alelos en las especies silvestres tiende a ser constante, a menos que
sucedan factores evolucionarios adversos (Fontdevila et al. 1999). Los resultados mostraron
diferentes frecuencias de alelos, por ejemplo; las poblaciones de Z. diploperennis y Z. perennis
fueron muy similares. Pero en general solo tres especies presentaron siete o más alelos raros, siendo
estos Z. diploperenis, Z. perennis y Z. nicaraguensis, en relación a los alelos únicos, casi todas las
especies presentaron igual cantidad excepto Z. luxurians quien solo obtuvo uno (Figura 2).
Finalmente, la heterocigosidad observada fue mayor Z. luxurians que en Z. nicaraguensis.
10
Raro
Único
Numero alelos
8
6
4
2
0
Z. perennis
Z. parviglumis
Z. nicaraguensis
Z. diploperennis
Z. huehuetenangensis Z.luxurians
Especies
Figura 2. Numero de alelos raros y únicos en los teocintles de la región mesoamericana.
Estas poblaciones fueron estadísticamente diferentes, la variación genética entre accesiones fue de
solamente 19.8 % en cambio el resto o sea 80.1 se debe a variación dentro de accesiones, lo que
indica que cada población presenta su propia diversidad genética.
La recombinación y el flujo génico juegan un importante rol en la dinámica de la diversidad
genética en los sistemas de cultivos tradicionales (Elias et al. 2000a). De igual manera otros
factores presentan influencia en los niveles de flujo génico tales como; baja producción de semilla,
esterilidad y dormancia en la semillas, manejo del cultivo, ecología del lugar, distancia física,
sincronía en la floración y selección humana (Baltazar et al. 2002).
En la región de Centro América, los datos revelan que la diferenciación genética y el flujo génico es
más similar entre Z. luxurians y Z. nicaraguensis (Fst = 0.095 y Nm = 2.38) que entre Z. luxurians y
Z. huehuetenangensis (Fst = 0.1304 y Nm = 1.66) a pesar que los dos últimos se encuentran más
cerca, ambos en Guatemala. Finalmente, la diversidad genética resulto ser un poco mayor entre los
teocintles de la región centro americana que los oriundos de la región mexicana.
44
Evolución
La domesticación del maíz (Zea mays L.) esta inexorablemente ligada al desarrollo de las culturas
del nuevo mundo y el maíz sigue siendo una fuente dominante de alimento. El maíz es la
preeminencia de la agricultura y ha conducido a su uso como un sistema modelo para la genética y
la biología molecular (Doebley, 1990a).
Durante muchos años las relaciones entre las especies Zea fueron objeto de mucha controversia. La
mayor dificultad en la taxonomía del maíz y la identificación de sus parientes más cercanos fue la
ausencia de una inflorescencia pistilada parecida a una mazorca en cualquiera de las otras especies.
Por lo tanto, los estudios comparativos de los cultivos y sus antepasados salvajes son altamente
relevantes para el fitomejoramiento. Los cultivos se diferencian de sus antepasados salvajes en su
expresión de los rasgos de importancia agronómica. El conocimiento del mecanismo que controla la
expresión de las características agronómicas proporcionan la base para las nuevas mejoras en los
cultivos (Matsuoka 2005).
Para determinar el parentesco genético entre los teocintles, se utilizaron ocho cebadores (primers)
para ampliar y secuenciar regiones del cloroplasto de las diferentes especies estudiadas. Asi mismo
se utilizo material de Sorgo (Sorghum bicolor L.) y Tripsacum (Tripsacum dactyloides) como
especies comparadores o fuera de grupo.
Los resultados indicaron que de 2347 inserciones y dilecciones (indels), 2214 fueron constantes y
133 variables, de las cuales solamente 57 presentaron información parsinómica (variación). De
igual manera las ocho regiones o cebadores utilizados, no todas amplificaron exitosamente sus
regiones en los materiales evaluados. El realizar el árbol de consenso estricto (Figura 3), se observo
que todos los materiales pertenecientes a la sección Luxuriantes, aparecen al mismo nivel, tal y
como lo describen otros investigadores (Bukcler and Holtsford 1996; Bukcler et al. 2006) utilizando
los mismos materiales pero usando otras regiones del genoma. La sección Mays presento una
pequeña variación dentro de la especie Z. parviglumis.
En conclusión, el teocintle nicaragüense debe ser considerado como otra especie diferente de los
otros teocintles de Mesoamérica, sin embargo, se sugiere utilizar nuevas regiones de cloroplastos
para una mayor determinación de la ubicación del Z. nicaraguensis dentro del género Zea.
Figura 3. Relación de teocintes de la región mesoamericana.
45
Bibliografia
Baltazar B, Schoper J (2002) Crop-to-crop gee flows: dispersal of transgenes in maize, during field
test and commercialization. In: Proc 7th Int Symp Biosafety Genet Modified Organisms. Beijing
Chinahttp://www.bba.de/gentech/isbgmo.pdf
Buckler E, Holtsford H 1996 Zea systematic: ribosomal ITS evidence. Mol. Biol. Evol. 13: 612622.
Buckler E, Goodman M, Holtsford T, Doebley J and Sanchez J 2006. Phylogeography of the wild
subspecies of Zea mays. Maydica 51: 123-134.
Doebley, J.F., Goodman, M, & Stuber, C. 1984. Isozyme variation in Zea (Graminaceae) Syst. Bot.
9:203-218.
Doebley, J.F. 1990. Molecular evidence for gene flow among Zea species. BioScience 40: 443-48.
Elias M, Panaud O, Robert T (2000a) Assessment of genetic variability in traditional cassava.
(Manihot esculenta Crantz). RAPD markers. Gen Mol Biol 21:105.
Fontdevila A, Moya A (1999) Introduccion a la genética de poblaciones. Editorial Sintesis. Madrid
España. 350 pp.
Frankel O. 1984. Genetics perspective of germoplasm conservation. Cambrige UK. Pp 161-180.
Loáisiga, C., Brantestam, A., Rocha, O., Salomon, B., & Merker A. 2011. Genetic diversity and
gene flow in Meso-American teosintes. Genetic Resources and Crop Evolution DOI:
10.1007/s10722-010-9637-6.
46
Turismo sostenible y Áreas Protegidas
Dra. Matilde Somarriba Chang
Cuando hablamos de turismo sostenible debemos tomar en cuenta los siguientes principios
de la sostenibilidad entre los que tenemos: satisfacer los usos y demandas actuales sin
deteriorar los sistemas naturales y culturales ni las futuras oportunidades para disfrutarlos.
Turismo rural ó Agro turismo: Esta actividad se realiza en un espacio rural, habitualmente
en pequeñas localidades, con una participación activa del visitante en actividades del
mundo agrario y rural, tomando en cuenta la alimentación con productos de cosecha propia,
montar caballo.
Cuando se habla de ecoturismo, nos estamos refiriendo al viaje que corresponden a Zonas
naturales que conserva el ambiente y mejora el bienestar de las poblaciones locales, para
ello se pretende entender la historia natural y la cultura local, con apoyo de la interpretación
y de la educación ambiental.
¿Existe potencial para estos tipos de turismo en Centroamérica y Nicaragua?
Principales indicadores económicos del turismo: Comparativo con valores promedios 20032008
Países
Aporte al PIB
Aporte
al Aporte a las Crecimiento
empleo
exportaciones
real del PIB
turístico
Promedio anual
Promedio anual
Promedio anual
Promedio anual
2003-2008
2003-2008
2003-2008
2003-2008
Costa Rica
14.3
13.9 %
18.8 %
6.2 %
El Salvador
8.5
7.5 %
22.3 %
7.7 %
Guatemala
6.9
6.0 %
20.5 %
4.2 %
Honduras
9.8
8.1 %
15.6 %
8.3 %
Nicaragua
6.4
5.2 %
18.2 %
5.4 %
Panamá
11.1
10.6 %
14.0 %
14.7 %
Latinoamérica
6.8 %
6.5 %
9.2 %
5.3 %
Nota: Los valores correspondientes a 2008 son proyecciones y los correspondientes a 2007 son estimaciones.
Fuente: CST por país del world travel& Tourism council (WTTC y OE, 2008)
El turismo de naturaleza, agroturismo y ecoturismo en Nicaragua
Los principales tipos de turismo basado en la naturaleza en Nicaragua son: ecoturismo, agro turismo
(rural), turismo de aventura, turismo científico (Somarriba et. al. 2001)
La afluencia del sector turístico está creciendo en Nicaragua al igual que en varios países de
Latinoamérica con bellezas por descubrir.
47
250
231
184
Millones de Dólares
200
152
167
150
100
50
0
2003
2004
2005
2006
Aproximadamente 100,000 visitantes/ año visita las áreas protegidas (14% de turistas en
.
El Papel del ecoturismo en el manejo de Áreas Protegidas: se deben tomar en cuenta los
siguientes aspectos.
El turismo no necesita ser masivo y destructor; Beneficios económicos para las áreas
protegidas; Implementado del modo adecuado, mejora relaciones entre las comunidades
locales y la administración de las A. P.
Principios del Ecoturismo: se concentra en los aspectos siguientes.
Minimizar impactos ecológicos y contribuir a la conservación de la biodiversidad; educar a
los visitantes y a los anfitriones; y proveer beneficios financieros y empoderamiento para
las comunidades locales (TIES, 1990).
Agro y Eco turismo en la Canta Gallo, Condega, Estelí
Zona de Producción Sostenible: Se permite la investigación científica, senderos, educación
ambiental y ecoturismo. Humedales de Apacunca.
48
Agro y Eco turismo en la Reserva de Recursos Genéticos de Apacunca, municipios de
Somotillo y Villanueva
Estudios vinculados al ecoturismo y las áreas protegidas
49
Estudio para el manejo y regulación de la Visitación Turística, Área Protegida Volcán
Cosigüina, Nicaragua: Senderos Guacamaya, El Humedal y San Luis
Identificación del Potencial Eco turístico en 7 fincas de la Reserva Natural Cerro Apante (Rosales,
L. 2006) y en 3 fincas de la Reserva Natural Cerro Arenal, Matagalpa, Nicaragua (Hidalgo, S.
2007)
Ecoturismo comunitario en diez fincas del parque ecológico municipal Canta Gallo, Condega,
Estelí
50
51
Conclusiones sobre los impactos ecológicos
• Reducción significativa de la cobertura vegetal en una banda adyacente a los senderos.
Ancho promedio ≤ 3 m, no es considerable para la superficie de la reserva.
• Riqueza de especies e Índice de Shannon de especies arbóreas en los senderos no difiere
significativamente de las parcelas testigo.
• En el caso de especies herbáceas y arbustivas, el análisis estadístico se da una diferencia
altamente significativa. Indicando un efecto en el soto bosque debido al uso.
•
Conclusiones sobre impactos socioeconómicos para las comunidades
Los beneficios más relevantes del turismo en Mombacho son de tipo económico y
ecológico; para los productores en Datanlí son de tipo económico y social.
Limitaciones de tipo social son de mayor peso en Datanlí y en cambio la conservación de la
reserva se percibe como un peligro inminente en Mombacho.
En Mombacho se emplea guarda parques y trabajadores locales, es un beneficio económico
para las comunidades vecinas
Las comunidades y productores en la zona de amortiguamiento no perciben que reciben
beneficios del turismo desarrollado en la zona núcleo. Desean participar activamente de las
actividades ecoturisticas
Conclusiones sobre Capacidad de carga
La visitación al Sendero El Cráter no excede la capacidad de carga efectiva calculada aún
pero está cerca de ese límite (estudiantes de colegio)
Si la tendencia de la visitación continúa, requerirá que la administración de la RNVM
restringa el número de visitas en los períodos picos. La CM se puede incrementar
sustancialmente si se mejora la infraestructura y las capacidades del personal, esto
aumentaría la CCE. Para esto se recomienda una estrategia de mejoras de acuerdo a las
debilidades o carencias encontradas en el análisis de la CM.
52
Dar a conocer y apreciar aún más las bellezas naturales de nuestro país
Conocer y compartir con las familias rurales de Nicaragua
53
POSTER
54
55
ELEMENTOS DE DISCUSIÓN
Compra-permuta de una finca en Apacunca para garantizar la finca donde hay
remanente de teocintle.
Mantener en el proyecto los componentes de desarrollo e investigación.
Crear una comisión técnica con visión integral (plan de manejo, convenio colaborativo,
POA, etc.).
Nombrar un director (a) de áreas protegidas. Y mantener un técnico en la zona.
Considerar el concepto de cuenca, integral. Buscar sinergias con otros proyectos
(proyecto del golfo).
Proponer a las alcaldías el cobro de impuestos por la alimentación de semovientes para
destinar dinero al manejo de la reserva.
UNAN-León: incluir en el proyecto el ordenamiento de fincas, evitar el despale,
gestión general de la parte turística. No se aterriza con los estudios actuales.
Posiblemente combinar visiones de corto, mediano y largo plazo. Hay problemas con
la inundación. Debería estar en territorio gubernamental.
La UNA tiene liderazgo, importante involucrarse decididamente en el Comité ya
existente. Reactivar el Comité local de manejo. Reasumir las experiencias de los
trabajos de investigaciones realizadas por los investigadores UNA y UNAN-León
Nombrar quien convocará al comité de manejo de la reserva y definir el papel de
MARENA a nivel central. Llevar el mensaje a la dirección del ministerio, cuyo
respaldo es de mucha importancia. El Ministerio debe involucrarse, en lo referente a
los planes que surjan deberá buscarse financiamiento.
El MARENA debe convocar y mejorar la estabilidad laboral de los delegados en la
zona.
La delegación departamental del MARENA en Chinandega, debe reconocer la
importancia del teocintle en la región. Se debe Proponer una visita a Apacunca para ver
in situ el estado de la zona después de las lluvias.
Invitar al director (a) de áreas protegidas para que asista a la reunión del comité, de
manejo de la reserva.
Se propone la participación de SINAPRED para evaluar la situación de los habitantes
de la zona.
Se debe dar prioridad a la situación de Don Oscar Martínez.
La delegación del MARENA enviará el POA 2011 para su evaluación y planificación.
Se puede negociar y cumplir los acuerdos sostenidos por intendencia de la propiedad,
con el señor Martínez y evitar que se siga afectando los remanentes de teocintle por
parte del señor Martínez.
56
Breve reseña bibliográfica de los expositores en el Congreso
Robert Mckelvy Bird
Biólogo de profesión, ha trabajado en origen y evolución de especies del genero Zea, desde
el punto de vista arqueológico y morfológico en el continente americano durante los
últimos 30 años. Igualmente ha estudiado los teocintles mesoamericanos. Actualmente es
profesor asociado de la Universidad de Carolina del Norte, USA.
Edilberto Duarte
Ingeniero Forestal, graduado en la Universidad Nacional Agraria. Ha trabajado en el
estudio de la diversidad florística de especies arbóreas principalmente del trópico seco.
Obtuvo su Maestría en la Universidad de Chapingo, México. Actualmente es Director de
Biodiversidad del Ministerio de Recursos Naturales y del Ambiente (MARENA).
Matilde Somarriba
Ingeniera agrónoma graduada en la Universidad Nacional Agraria. Ha trabajado por
muchos años el manejo de cuencas hidrográficas y recientemente finalizo sus estudios de
doctorado en el manejo de áreas protegidas y recursos naturales.
Álvaro Benavides González
Ingeniero Agrónomo graduado en la Universidad Nacional Agraria. Ha trabajado por
muchos años en la caracterización morfológica de recursos genéticos vegetales, obtuvo su
Maestría en el estudio del teocintle nicaragüense. Actualmente miembro del programa
Desarrollo Rural Participativo (DEPARTIR).
Juan Betancourt
Productor de la zona y dirigente campesino por muchos años, conocedor de los problemas
que ha tenido la Reserva de Recursos Genéticos Apacunca (RRGA) a través del tiempo e
importante defensor de la flora y fauna.
Carlos Henry Loáisiga
Ingeniero Agrónomo graduado en la Universidad Nacional Agraria. Ha dedicado muchos
años a la caracterización de recursos genéticos vegetales, recientemente obtuvo su
doctorado en el estudio del teosintle nicaragüense.
Daniel Querol Lipcovich
Genetista graduado en la Universidad Autónoma de Chapingo, México. Ha trabajado por
muchos años es el estudio de los recursos genéticos vegetales de Nicaragua, fue fundador
del Programa Recursos Genéticos Nicaragüenses (REGEN) y consultor-asesor del grupo de
investigación Desarrollo Rural Participativo (DEPARTIR).
57
Anexos
LISTA DE PARTICIPANTES Y TEMÁTICAS A ABORDAR
Expositores:
Dr. Robert Bird: El gran teocintle de Apacunca: Estudios del teocintle tras tres décadas,
tolerancia a inundaciones.
Ing. MSc. Álvaro Benavides González: Situación actual del teocintle en la Reserva
genética de Apacunca.
Ing. MSc. Edilberto Duarte: Las áreas protegidas y los Recursos Genéticos
Ing. MSc. Daniel Querol: La importancia de los recursos genéticos silvestres y el rol de la
UNA en su protección y aprovechamiento
Dr. Carlos Henry Loáisiga Caballero: Citogenética, diversidad genética y filogenia en
especies silvestres del género Zea, con énfasis en Zea nicaraguensis.
Dra. Matilde Somarriba Chang: Turismo sostenible y Áreas Protegidas
58
Invitados Participantes:
1.
Dr. Gero Vaagt, Representante FAO Nicaragua, Managua.
8.
Ing Edilberto Duarte, Responsable Biodiversidad (MARENA)
9.
Lic. Ana Reyes Zavala (Investigadora UNAN LEON)
21.
Lic. Mauricio Álvarez (Investigador UNAN-LEON)
10.
Lic. Santos Méndez Periodista del RECACH (Chinandega)
11.
Sr. Juan Betancourt (Dirigente comunitario y Pte. de Cooperativas de Apacunca)
12
Sr. Antonio García (Guardaparque Estación Biológica Apacunca)
13.
Sr. Julio Martínez (Guardaparque Estación Biológica Apacunca)
14.
Ing. Miguel Matus FACA UNA
21.
Ing. Yelba Cañada (Tesista)
Otros invitados que no aparecen en lista oficial
1.
Ing. José Dolores Cisne (DEPARTIR – UNA)
2.
Ing. Daniel Querol (DEPARTIR – UNA)
3.
Ing. Álvaro Benavides (DEPARTIR – UNA)
4.
Ing. Juan Carlos Morán (DEPARTIR – UNA)
5.
Dr. Robert Bird (USA)
6.
Dra. Matilde Somarriba (FARENA-UNA)
7.
Dr. Dennis Salazar (Decano FAGRO)
8.
Dr Oscar Gómez (DEPARTIR)
9.
Dr. Guillermo Reyes (DEPARTIR)
10.
Dr. Carlos Loaísiga (DEPARTIR )
11.
Dr. Bryan Mendieta (FACA UNA)
12.
Ing. Vidal Marín (FAGRO UNA)
13.
Ing. Nelson Artola
14.
Personal de Logística y Periodismo
59
PRIMER CONGRESO NACIONAL:
EL TEOCINTLE NICARAGUENSE, SU ESTADO ACTUAL Y SU IMPORTANCIA
EN MESOAMERICA
PROGRAMA (Noviembre 4)
08:00
08:30
08:45
09:00
09:30
08:30 AM
08:45 AM
09:00 AM
09:30 AM
10:00 AM
10:00 10:30 AM
10:30 11:00 AM
Genética
Inscripción de participantes
Apertura del Congreso Ing. Telémaco Talavera (Rector UNA)
Metodología del Congreso (PhD. Carlos Loaisiga)
Representante FAO Nicaragua (Dr. Gero Vaagt)
Áreas Protegidas y Recursos Genéticos (Caso teocintle)
MSc. Edilberto Duarte (MARENA)
Refrigerio
Incorporación de especies silvestres en Programas de Mejora
Ing. María Isabel Martínez (INTA)
11:00 11:30 AM
Áreas Protegidas y Agroturismo
Dra. Matilde Somarriba (Vice decana FARENA)
11:30 11:45 AM
Estado actual del teocintle en la Reserva
MSc. Álvaro Benavides González (UNA)
11:45 12:00 M
Situación del Teocintle en Apacunca
Sr Juan Betancourt, Poblador de la Reserva
12:00 01:00 PM
Almuerzo
01:00 01:30 AM
Caracterización Molecular del Teocintle nicaragüense
PhD. Carlos Henry Loáisiga Caballero (UNA)
01:30 02:00 AM
Importancia de los recursos genéticos silvestres y el rol de la UNA en
su Protección y conservación.
MSc. Daniel Querol L.
02:00 02:45 AM
Descubriendo, caracterizando y utilizando germoplasma de los
teocintles Centroamericanos.
Dr. Robert Bird, North Caroline University. USA
02:45 03:00 PM
Refrigerio
03:00 04:00 PM
Discusión técnica del teocintle y la Reserva
03:00 03:30 PM
Alianzas nacionales e internacionales (proyectos – fondos)
03:30 04:00 PM
Clausura (Dr. Denis Salazar, Decano FAGRO)
60
Gráficas del Congreso
Autoridades de la Universidad Nacional Agraria, Invitados Especiales y Pobladores
de las comunidades de Apacunca y Aquespalapa.
61
Ing. Telémaco Talavera, Rector de la Universidad Nacional Agraria.
Dr. Gero Vaagt, representante de la FAO en Nicaragua.
62
Dr. Robert McK. Bird, Profesor asociado a la Universidad Carolina del Norte, USA.
Ing. Edilberto Duarte, representante del Ministerio de Recursos Naturales y del Ambiente
(MARENA) de Nicaragua.
63
Ing. Miguel Obando, representante del Instituto Nicaragüense de Tecnología Agropecuaria
(INTA) de Nicaragua.
64
Dr. Dennis Salazar, Decana de la Facultad de Agronomía (FAGRO), Universidad
Nacional Agraria.
65
Dra. Matilde Somarriba Chang, Vicedecana de la Facultad de Recursos Naturales y del
Ambiente (FARENA), Universidad Nacional Agraria.
66
Lic. Santos Méndez, Periodista del RECACH, Chinandega.
Sr. Juan Betancourt, Dirigente comunitario y Presidente de Cooperativas de Apacunca.
67