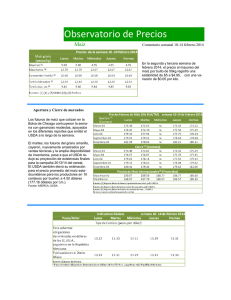
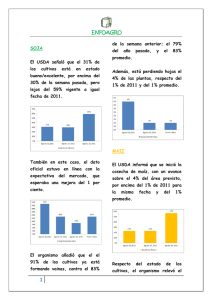
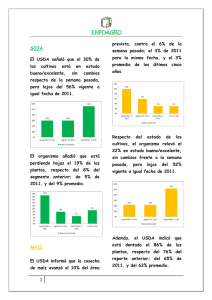
Revisión de la seguridad ambiental de la proteína Cry1Ac
Anuncio

Revisión de la seguridad ambiental de la proteína Cry1Ac Center for Environmental Risk Assessment, ILSI Research Foundation 1156 Fifteenth Street N.W., Washington D.C. 20005-1743 EE. UU. 26 de mayo de 2010 Introducción Este documento presenta una revisión completa de la información y los datos relevantes para la evaluación de riesgo ambiental del Cry1Ac y también presenta una declaración resumida acerca de la seguridad ambiental de esta proteína. Todas las fuentes de información aquí revisadas estaban disponibles públicamente y se incluyeron: expedientes presentados a las autoridades regulatorias; resúmenes de decisiones preparados por las autoridades regulatorias; bibliografía revisada por pares y resúmenes de productos preparados por los desarrolladores de productos. Las evaluaciones de riesgo ambiental relacionadas con la introducción de plantas genéticamente modificadas (GM) se realizan caso por caso y consideran la biología de la planta, la naturaleza del transgen y la proteína que produce, el fenotipo que concede el transgen y también el uso previsto de la planta y el medioambiente en el que se la introducirá (es decir, el ambiente receptor). Dichas evaluaciones son comparativas por necesidad y por lo general implican comparaciones con una línea progenitora sin transformar o con una isolínea muy relacionada (CBD 2000a, 2000b, Codex 2003a, 2003b, EFSA 2006, NRC 1989, OCDE 1992). El propósito de estas comparaciones es identificar los posibles riesgos que la planta GM podría presentar al ambiente, más allá de los ya aceptados de plantas similares. Cualquier riesgo identificado se puede evaluar en busca de probabilidades y posibles consecuencias. En once países, se han emitido las aprobaciones regulatorias para la liberación ambiental de plantas GM que expresan Cry1Ac e incluyen tres especies de plantas (CERA 2010, CFIA 1997, CTNBio 2005, BCH de Japón 1997, 1999, 2007, OGTR 2002b, 2003a, 2003c, 2006a, 2006c, USDA APHIS 1995, 1997a, 1997d, 2001, 2004) (Tabla 1). Un evento 1 del maíz (Zea mays) y otro del tomate (Lycopersicon 1 Evento significa un único evento de transformación: la incorporación de un transgen en el genoma de una planta. Un único evento de transformación se puede cruzar en múltiples líneas. esculentu) 2 han recibido aprobación mientras que 12 líneas de algodón 3 (Gossypium hirsutum) han recibido aprobación al menos en un país. Estas revisiones regulatorias, por lo general, han considerado el potencial del Cry1Ac para afectar negativamente los organismos no objetivo, el potencial de que la expresión de Cry1Ac afecte el potencial para convertirse en maleza de la planta modificada y el potencial para que el flujo de genes impacte en las malezas de parientes silvestres (CFIA 1997, CTNBio 2005, BCH de Japón 1997, 1999, 2007, OGTR 2002b, 2003a, 2003c, 2006a, 2006c, USDA APHIS 1995, 1997a, 1997d, 2001, 2004). Palabras clave Cry1Ac, Bacillus thuringiensis, resistencia a los insectos, genéticamente modificado, evaluación de riesgo ambiental Origen y función de Cry1Ac Bacillus thuringiensis y las δ-endotoxinas Cry Bacillus thuringiensis es una bacteria en forma de bastón, gram positiva, capaz de formar endosporas de larga vida. Por lo general, se la menciona como una bacteria del suelo aunque se encuentra en todo el medioambiente (Hofte y Whiteley 1989, Schnepf et al. 1998, OCDE 2007). Existe una enorme variación entre las especies con respecto a la producción de un rango de proteínas pesticidas que difieren en su modo de acción, la especificidad del objetivo y el mecanismo de expresión (Hofte y Whiteley 1989, Schnepf et al. 1998, OCDE 2007). Las proteínas pesticidas expresadas por las cepas de B. thuringiensis incluyen compuestos antihongos, β exotoxinas 4, proteínas insecticidas vegetativas (Vip) y las δ endotoxinas que incluyen las proteínas Cry (cristalina) y las proteínas Cyt (citolítica), estructuralmente no relacionadas (Hofte y Whiteley 1989, Schnepf et al. 1998, OCDE 2007). Se ha demostrado que la mayoría de ellas contribuye con la toxicidad frente 2 El Evento 5345 de tomate que expresa Cry1Ac fue desregulado por USDA APHIS pero nunca recibió registro como pesticida de la USEPA y no fue comercializado en ningún lugar del mundo. No será considerado en ninguna otra sección de este documento. 3 Esto incluye las aprobaciones de las líneas generadas a través de la reproducción y la transformación con transgenes adicionales. 4 también llamada thuringiensin Copyright © ILSI Research Foundation 2010 El presente trabajo está autorizado en virtud de la licencia Creative Commons AttributionNoncommercial-No Derivative Works 3.0 de Estados Unidos. Para ver una copia de esa licencia, visite http://creativecommons.org/ licenses/by-nc-nd/3.0/us/ o envíe una carta a Creative Commons, 171 Second Street, Suite 300, San Francisco, California, 94105, EE. UU. X X X X * * X Sudáfrica México Japón X India X Colombia X Canadá Burkina Faso MON-15985-7 MON 15985 DAS-21023-5 3006-210-23 31807/31808 DAS-21023-5 x DAS-24236-5 DAS-21023-5 x DAS-24236-5 x MON-01445-2 DAS-21023-5 x DAS-24236-5 x MON88913 Evento-1 ACS-GH001-3, MON-15985-7 LLCotton25 x MON15985 MON-15985-7 x MON-01445-2 MON-00531-6 x MON-01445-2 MON-15985-7, MON-88913-8 MON15985 x MON88913 MON-00531-6, MON-00757-7 MON531/757/1076 5345 DKB-89614-9 DBT418 Brasil Gossypium hirsutum (algodón) Lycopersicon esculentum (tomate) Zea mays (maíz) También conocido como Australia Nombre del evento Argentina Especie Estados Unidos Tabla 1. Aprobaciones regulatorias para la liberación al ambiente de plantas GM que contienen Cry1Ac. X X X X * * * X X * X X X X X X X X X X X X X X X X X X X X La X indica una aprobación regulatoria. *Acumulación de eventos que pueden considerarse aprobados para su liberación al medioambiente según las aprobaciones existentes de las líneas progenitoras GM de las cuales son derivados. Las aprobaciones dependen de los registros de los pesticidas, los cuales requieren una renovación periódica. a los insectos y algunas (se destacan las β exotoxinas y las proteínas Cyt) son muy tóxicas (Hofte y Whiteley 1989, Schnepf et al. 1998, OCDE 2007). Las preparaciones de aislados naturales de B. thuringiensis se usaron por primera vez como insecticida comercial en Francia en 1938 y se han registrado B. thuringiensis subespecie kurstaki (que, entre otras proteínas Cry, produce Cry1Ac) en US EPA desde 1961 (Kumar et al. 1996, Schnepf et al. 1998, USEPA 2001). Las preparaciones microbianas de B. thuringiensis actualmente están aprobadas para su uso en todo el mundo, incluido en Australia, Canadá, la Unión Europea y los Estados Unidos (AVPMA 2010, EU DG SANCO 2010, PMRA 2008, USEPA 2001). Dichas preparaciones contienen una mezcla de pesticidas microbianos, que incluyen proteínas Cry, que interactúan ampliamente entre sí para influir en la toxicidad y la especificidad de los insectos (Schnepf et al. 1998, OCDE 2007). Si bien se podría extrapolar cierta información sobre la seguridad ambiental de las proteínas Cry a partir de la experiencia con las preparaciones bacterianas, se debe tener presente que la actividad del rociado foliar bacteriano se debe a una combinación de varias δ endotoxinas como también de otras toxinas y calidades de la espora misma, que pueden tener un impacto sobre la selectividad y el rango de huésped (Schnepf et al. 1998, Tabashnik et al. 1992). Asimismo, el perfil de exposición para el rociado foliar de preparaciones bacterianas difiere de la expresión de proteínas Cry en una planta GM (OCDE 2007). Las δ endotoxinas de la proteína Cry reciben ese nombre porque son el componente principal de los cristales parasporales de la proteína, que son característicos en la formación de esporas en B. thuringiensis (Hofte y Whiteley 1989, Kumar et al. 1996, Schnepf et al. 1998, OCDE 2007). Se propuso una nomenclatura sistemática para identificar y diferenciar las proteínas Cry en 1989 y fue ampliamente adoptada (Hofte y Whiteley 1989, OCDE 2007). Este sistema se ha actualizado posteriormente para dar cuenta de las 2 proteínas Cry adicionales y ampliar el conocimiento de la función molecular y la familiaridad, lo cual conduce a algunas discrepancias menores en los nombres en comparación con la bibliografía anterior (Crickmore et al. 1998, Crickmore et al. 2005, OCDE 2007). Este documento utiliza la nomenclatura más reciente (Cry1Ac para la proteína, cry1Ac para el gen) pero la proteína en cuestión es análoga con la nomenclatura anterior CryIA(c). Todas las proteínas Cry1 están muy relacionadas entre sí por su secuencia y las proteínas designadas como Cry1A (incluidas Cy1Aa, Cry1Ab y Cry1Ac) son más del 85% idénticas en su secuencia de aminoácidos (Crickmore et al. 1998). Se ha determinado la estructura cristalina de Cry1Aa y presenta un alto nivel de similitud con otras estructuras conocidas de la proteína Cry (Cry3A, Cry2A, Cry4A y Cry4B) a pesar de las identidades de la secuencia, que pueden ser menores al 30% (Aronson y Shai 2001, Bravo et al. 2007, Crickmore et al. 1998, Kumar et al. 1996, OCDE 2007). En la nomenclatura original, las proteínas Cry se designaron según su actividad insecticida (las proteínas CryI fueron las activas contra los lepidópteros) y si bien ahora la nomenclatura depende de la secuencia, la especificidad del objetivo sigue mayormente intacta, de manera que las proteínas designadas como Cry1 presentan actividad específicamente contra los lepidópteros (Aronson y Shai 2001, Crickmore et al. 1998, Hofte y Whiteley 1989, Kumar et al. 1996, OCDE 2007). Mecanismo de la actividad insecticida de Cry1Ac Aunque existe una significativa variabilidad en la secuencia de aminoácidos y en el rango objetivo, se cree que el mecanismo general por el que las proteínas Cry (incluidas las Cry1Ac) logran la actividad insecticida es común en todo el grupo (Aronson y Shai 2001, Bravo et al. 2007, Crickmore et al. 1998, 2005, Hofte y Whiteley 1989, Kumar et al. 1996, OCDE 2007). Las proteínas Cry1 se producen en forma de protoxinas de 130-140 kDa en tamaño, con 1100-1200 residuos de aminoácidos (Aronson y Shai 2001, Bravo et al. 2007, Kumar et al. 1996, OCDE 2007). En el caso de la Cry1A, las protoxinas se dividen para generar toxinas activas que están compuestas por fragmentos de 60-70 kDa de la porción terminal N de la proteína (Knowles 1994, Kumar et al. 1996). Las toxinas supuestamente activas se unen a receptores específicos que se encuentran en la membrana plasmática de las células epiteliales del intestino medio de insectos susceptibles (Aronson y Shai 2001, Bravo et al. 2007, Kumar et al. 1996, OCDE 2007). Una vez unida a los receptores, la toxina puede insertarse en la membrana plasmática y formar poros oligoméricos transmembrana (Aronson y Shai 2001, Bravo et al. 2007, Kumar et al. 1996, OCDE 2007). Se cree que dichos poros forman canales iónicos que afectan el potencial trasmembrana, lo cual causa lisis osmótica (Aronson y Shai 2001, Hofte y Whiteley 1989, Kumar et al. 1996, OCDE 2007). Aún no se comprende totalmente el proceso bioquímico de la inserción en la membrana. Existen evidencias de que algunas proteínas Cry cuentan con receptores múltiples, o pueden estar unidas a múltiples sitios en un único receptor y se ha demostrado que la unión de receptores es necesaria pero no es suficiente para la toxicidad (Aronson y Shai 2001, Jenkins et al. 1999, OCDE 2007). Existe cierta evidencia, basada parcialmente en experimentos que utilizan concentraciones inferiores a las mortales, de que podrían existir otras interacciones relevantes entre las proteínas Cry y los insectos que son su objetivo (Aronson y Shai 2001, Zhang et al. 2006). Expresión de Cry1Ac en plantas GM que resisten a los insectos El nivel de expresión de Cry1Ac en las plantas GM se determina mediante varios factores relacionados con los tipos de promotores y secuencias finales y los sitios de inserción de los genes. Por lo tanto, cada evento de transformación tiene como resultado un perfil de expresión distinto. Los datos del nivel de expresión de Cry1Ac en las plantas GM que han obtenido aprobaciones regulatorias están disponibles en los documentos de presentaciones y decisiones que son accesibles para el público (CFIA 1996, 1997, 2004, 2005, CTNBio 2005, 2009, OGTR 2003, 2006, USDA APHIS 1994, 1996, 1997a, 1997b, 2003, 2000, USEPA 2001, USFDA 1997). Los tipos de tejido y los métodos de recolección variaron en los estudios, pero todos utilizaron el método de ensayo por inmunoabsorción ligado a enzimas (ELISA) para cuantificar la cantidad de proteína Cry1Ac presente en una muestra dada. Por lo general, se tomaron una o más muestras de tejido de la planta en un centro de ensayo en el campo y se agruparon para su análisis. Por lo general, la cantidad de Cry1Ac se determinó en peso seco y luego se calculó para arrojar valores relevantes a nivel ambiental relacionados con el peso fresco total de la muestra y se representó en una proporción (por ejemplo, microgramos de la proteína Cry1Ac por gramo de peso fresco) (CFIA 1996, 1997, 2004, 2005, CTNBio 2005, 2009, OGTR 2003, 2006, USDA APHIS 1994, 1996, 1997a, 1997b, 2003, 2000, USEPA 2001, USFDA 1997). Las muestras se recolectaron de varios tipos de tejido y en distintas etapas de crecimiento, y se obtuvieron datos de plantas a través del tiempo y de distintas ubicaciones. En la mayoría de los casos, los datos se presentaron como un valor medio (por lo general, se promedió una media de medias como valores dentro de un estudio de campo y también entre distintos estudios) y un rango (por lo general, también un rango de medias que representaban la expresión promedio en un centro de estudio, aunque esto también varió, según cada caso en particular). En otros conjuntos de datos, se presentan las medias con la desviación típica o el error típico de las medias. (CFIA 1996, 1997, 2004, 2005, CTNBio 2005, 2009, OGTR 2003, 2006, USDA APHIS 1994, 1996, 1997a, 1997b, 2003, 2000, USEPA 2001, USFDA 1997). Las variaciones en la metodología empleada para recolectar las muestras hacen que sean inadecuadas las comparaciones cruzadas y estadísticas directas de los datos, pero el peso de la evidencia indica que las plantas GM expresaron la Cry1Ac a niveles muy bajos, en comparación con la proteína total disponible en la planta (consultar el anexo I y las referencias que allí se incluyen). La Tabla 2 incluye los valores más altos informados de la expresión de Cry1Ac en las plantas GM que presentan expresión, donde los datos estaban disponibles. El Anexo I contiene información adicional acerca de la expresión de Cry1Ac. Pruebas de Organismo no objetivo (ONO) e impactos de la exposición a la proteína Cry1Ac La proteína Cry1Ac tiene propiedades insecticidas sobre ciertos insectos lepidópteros cuando se alimentan con un sustrato que contiene la proteína Bt (Crickmore et al. 1998, 2005, Hofte y Whiteley 1989, OCDE 2007). El propósito de insertar el gen cry1Ac en un cultivo es el de ofrecer protección frente al daño causado por la alimentación de ese tipo de plagas. Otros organismos del sistema agrícola que no son plagas también pueden quedar expuestos a la proteína Cry1Ac y se los considera “organismos no objetivo” (ONO). Dicha exposición puede ser directa, a partir de la alimentación intencional o incidental con tejidos del cultivo como el polen u hojas en descomposición, o también puede ser indirecta, a partir de la alimentación con otros herbívoros que a su vez se alimentan del cultivo. Dado que la Cry1Ac Tabla 2. Información de concentraciones más altas de proteína Cry1Ac en el tejido vegetal GM. Especie Eventos Gossypium hirsutum MON-15985-7 Semilla 3,35±0,631 µg/g FW2 DAS-21023-5 Hoja joven 0,46-3,5 µg/g DW3 USDA 2003 DAS-21023-5 X DAS-24236-5 Flor 1,83 µg/g DW FSANZ 2004 31807/31808 Semilla4 2,5 µg/g FW FDA 1997 MON-00531-6 Hoja joven 5,00 ± 1,841 µg/g FW OGTR 2002 DKB-89614-9 Hoja recolectada 626,8 ± 141,62 ng/g FW USDA 1996 Zea mays Tejido con la expresión más alta Rango Cita 5 OGTR 2002 1 Desviación típica 2 FW = peso fresco 3 DW = peso seco. 4 Único tejido informado. 5 Error típico. 3 ha demostrado actividad pesticida, se ha considerado el posible daño a los ONO como parte de las evaluaciones de riesgo regulatorias de las plantas GM que expresan Cry1Ac, con una especial consideración para los ONO beneficiados que llevan a cabo funciones valiosas, del mismo modo que las especies amenazadas, en peligro de extinción o atractivas (CFIA 1996, 1997, 2004, 2005, CTNBio 2005, 2009, BCH de Japón 1997, 1999, 2007, OGTR 2002a, 2003b, 2003c, 2005, 2006b, USDA APHIS 1994, 1996, 1997a, 1997b, 1997c, 2000, 2003, USEPA 2001). Generalmente, se consideran las posibles exposiciones y se las utiliza para determinar qué organismos pueden verse afectados por el pesticida y luego se puede analizar a dichos organismos, o a especies sustitutas que los representen, en busca de efectos adversos. El impacto de los pesticidas sobre los ONO generalmente se determina mediante una serie secuencial de pruebas llamadas Nivel I, Nivel II, Nivel III y Nivel IV (USEPA 2007). La naturaleza exacta de cada nivel de pruebas depende de cada caso, pero habitualmente el nivel de realismo y complejidad de las pruebas excede los niveles (EFSA 2006, Romeis et al. 2008, Rose 2007, USEPA 2007, USEPA 2010). Los primeros estudios por niveles incluyen ambientes de laboratorio muy controlados, donde los ONO o la especie sustituta se exponen a altas concentraciones del pesticida en estudio con el propósito de determinar si existe algún efecto (Romeis et al. 2008, Rose 2007,USEPA 2010, USEPA 2007). Si no se observan efectos, por lo general, no es necesario realizar más pruebas en los niveles superiores (Romeis et al. 2008, Rose 2007, USEPA 2010, USEPA 2007). Si se observan efectos adversos en las pruebas de los primeros niveles o si existe una duda inaceptable, las pruebas adicionales continuarán, según sea necesario, hasta los últimos niveles para reducir la duda a un nivel aceptable para la toma de decisiones (EFSA 2006, Romeis et al. 2008, USEPA 2010, USEPA 2007). Vías de exposición ambiental Las decisiones regulatorias, por lo general, toman en cuenta tres vías principales de exposición además del contacto directo con la planta GM que expresa la proteína Cry1Ac, a saber: exposición al polen que contiene Cry1Ac y exposición a la Cry1Ac depositada en el suelo por la planta en descomposición y exposición tritrófica a través de la alimentación con herbívoros que consumen la planta GM (BCH de Japón 1999, OGTR 2003, USEPA 2001). La exposición a través del polen es limitada por los niveles de expresión generalmente bajos de Cry1Ac en el polen de las variedades que recibieron aprobaciones regulatorias (consultar el Anexo I para conocer los datos del nivel de expresión en polen de las variedades aprobadas) como también por la densidad de sedimentación del polen que disminuye rápidamente al aumentar la distancia de la planta original (CFIA 1997, FSANZ 2004, OGTR 2002a, 2003b, 2003c, 2005, 2006b, 2008, USDA APHIS 1994, 1996, 1997a, 1997b, 1997c, 2000, 2003, USEPA 2001). Aunque se puede presentar cierta exposición biológicamente significativa a poca distancia de los campos de cultivo, por lo general, las agencias regulatorias únicamente solicitan datos del impacto de Cry1Ac sobre las especies polinizadoras más representativas (por ejemplo, la abeja) (EU SCP 1998, BCH de Japón 1999, OGTR 2002b, 2003a, 2003c, 2006a, 2006c, USEPA 2001). Asimismo, la especificidad de la toxicidad de Cry1Ac sobre los lepidópteros y la evidencia que sugiere una exposición baja a través del suelo condujo a quienes emiten las regulaciones a solicitar pruebas únicamente de las especies de artrópodos que habitan el suelo (EU SCP 1998, OGTR 2002a, 2003b, 2003c, 2005, 2006b, USEPA 2001). En varios informes, se ha indicado que las proteínas Cry de las plantas GM pueden unirse a los sustratos de arcilla presentes en el suelo y que dichas proteínas unidas están protegidas de la digestión 4 de los microbios pero conservan su actividad insecticida (Crecchio y Stotsky 1997, Koskella y Stotzky 1997, OCDE 2007). Estos estudios utilizaron concentraciones muy altas de proteínas Cry en relación con la cantidad de sustrato unido, lo cual representa una exposición mucho más alta de la que posiblemente ocurra en un ambiente agrícola. Los estudios posteriores, bajo condiciones más relevantes para los campos agrícolas, han respaldado las conclusiones anteriores acerca de la degradación de Cry1Ac con una vida media de aproximadamente 9-40 días (Accinelli et al. 2008, Marchetti et al. 2007). En al menos un experimento de campo, no se detectó Cry1Ac mediante ELISA o bioanálisis en campos agrícolas en los que se había cultivado algodón que expresaba Cry1Ac (MON-00531-6) y cultivado en suelos durante tres a seis años consecutivos (Head et al. 2002). Las aprobaciones regulatorias de los eventos de Cry1Ac han considerado información sobre las tasas de degradación de la proteína Cry en un rango de tipos de suelo, pero no han solicitado pruebas adicionales de la toxicidad que implica la Cry1Ac para los organismos del suelo (CFIA 1997, CTNBio 2005, BCH de Japón 1997, 1999, 2007, OGTR 2002a, 2003b, 2003c, 2005, 2006b, USDA APHIS 1994, 1996, 1997a, 1997b, 1997c, 2000, 2003, USEPA 2001). Las posibles exposiciones bitróficas y tritróficas se tratan con pruebas ecotoxicológicas. Pruebas ecotoxicológicas de Cry1Ac sobre ONO no lepidópteros Las pruebas en ONO sobre la Cry1Ac purificada se han realizado en una gran variedad de especies de no lepidópteros para las presentaciones regulatorias relacionadas con las plantas GM que producen Cry1Ac (ANZFA 2002, OCDE 2007, USEPA 2001). Los organismos de prueba incluyeron Apis mellifera (abeja) adultas y larvas, coleópteros depredadores Hippodamia convergens (mariquita) y neurópteros Chrysoperla carnea (crisopa de alas verdes), Himenópteros parásitos Nasonia vitripennis, así como también especies que viven en el suelo, como los colémbolos Folsomia candida y Xanylla grisea. Ninguno de esos organismos presentó una respuesta significativa frente a la Cry1Ac con las concentraciones utilizadas en la prueba, lo cual condujo a observaciones de nivel de efectos no observados (NOEL). Además, se realizaron pruebas toxicológicas agudas en mamíferos como ratones (Mus musculus) (ANZFA 2002, USEPA 2001). Los resultados de todos estos estudios se sintetizan en la Tabla 3. Pruebas ecotoxicológicas de Cry1Ac sobre los lepidópteros no objetivo Danaus plexippus L. (mariposa monarca) Se sabe que las proteínas Cry1 tienen un efecto tóxico sobre ciertos insectos del orden de los lepidópteros (Crickmore et al. 1998, 2005, Hofte y Whiteley 1989, OCDE 2007). Puesto que los lepidópteros que se alimentan de las plantas modificadas para que expresen proteínas Cry1 son considerados una plaga, los estudios de los organismos no objetivo han considerado el impacto sobre los lepidópteros que incidentalmente puedan encontrarse expuestos a las proteínas Cry. La mayoría de las investigaciones se han concentrado en la mariposa monarca (Danaus plexippus), una especie reconocida, apreciada y valorada de Norteamérica. Los primeros estudios con mariposas monarca (Jesse y Obrycki 2000, Losey et al. 1999) no evaluaron el material Cry1Ac de la planta; sin embargo, en investigaciones posteriores, se ha examinado la toxicidad de Cry1Ac sobre larvas de monarca tanto en los estudios de Nivel I con proteínas purificadas en una dieta artificial como en los estudios de Nivel II en los que se simuló la exposición mediante el polen del evento de maíz DKB-89416-9 (Hellmich et al. 2001). Estos Tabla 3. Resumen de pruebas ecotoxicológicas del Cry1Ac sobre organismos que no son objetivos ni pertenecen al orden de los lepidópteros. Especie Método de exposición Duración Resultados de la observación Larvas de Apis mellifera (abeja) Inyección única de solución de proteína purificada en las células que tienen larvas en desarrollo 1-3 días luego de la incubación hasta NOEL 20 ppm1 el brote del adulto Adultos de Apis mellifera (abeja) Alimentación con proteína purificada en una solución de agua y miel ND NOEL 20 ppm1 Nasonia vitripennis Proteína purificada en una dieta de agua y miel 23 días NOEL 20 ppm1 Hippodamia convergens (mariquita) Proteína purificada en una dieta de agua y miel 30 días NOEL 20 ppm1 Larvas de Chrysoperla carnea (crisopa de alas verdes) Proteína purificada mezclada en una pasta de huevos de sitotroga 11 días NOEL 20 ppm1 Folsomia candida (colémbolos) Proteína purificada en dieta artificial 21 días NOEL > 200 ppm1 Xenylla grisea (colémbolos) Proteína purificada en dieta artificial 21 días NOEL > 200 ppm1 Mus musculum (ratón) Dosis única por vía oral >3280 mg Cry1Ac/kg de peso 14 días No se observan efectos1 1 Datos informados en US EPA 2001 y en ANZFA 2002. estudios señalan que las larvas de monarca son sensibles a Cry1Ac y la exposición bajo condiciones de laboratorio puede causar un retraso en el desarrollo y la mortalidad a las larvas de monarca. Sin embargo, la exposición al polen del maíz que expresa Cry1Ac (evento DKB89416-9) en concentraciones >1600 granos de polen/cm2 de hojas de asclepias no afecta el crecimiento ni la supervivencia (Hellmich et al. 2001). Un estudio sobre la sedimentación del polen sobre las asclepias ubicadas en los campos de maíz y sus alrededores estableció que se espera que menos del 1% de las hojas de asclepia que se encuentran dentro del campo de maíz durante las dos semanas de antesis presenten concentraciones de polen superiores a 900 granos/cm2 (OCDE 2003, Tabla 4. Resumen de pruebas ecotoxicológicas de la mariposa monarca (D. plexippus).1 Especie Método de exposición Duración Resultado Danaus plexippus 1.º larva instar Proteína purificada incorporada a la dieta de prueba 7 días LC50 = 13,8 ng/ml dieta artificial2 EC50 = 0,9 ng/ml dieta artificial3 Danaus plexippus Granos de polen de DBT418 derramados en hojas de asclepias como sustrato alimentario (100 - >1600 granos de polen/ cm2) 4 días No se observaron efectos 1 Datos de Hellmich et al. 2001. 2 LC50 = Concentración en la que se espera el 50% de mortalidad de las larvas (concentración letal). 3 EC50 = Concentración que se espera que produzca el 50% de la inhibición de crecimiento por cálculo. los ONO (Romeis et al. 2006). Se ha creado una base de datos 5 que reúne dicha información, para facilitar el estudio constante (Duan et al. 2008, 2010, Marvier et al. 2007, Naranjo 2009, Wolfenbarger et al. 2008). Cuando se compararon las plantas GM que expresan proteínas Cry, incluido el algodón Cry1Ac, con las plantas de control que no habían sido tratadas con insecticidas químicos hubo una disminución en la abundancia de artrópodos, pero cuando las plantas de control sí habían sido tratadas con insecticidas, la abundancia de artrópodos fue significativamente mayor en las plantas GM que expresan proteínas Cry (Marvier et al. 2007, Naranjo, 2009 Wolfenbarger et al. 2008). En las comparaciones entre las plantas GM que expresan proteínas Cry y las plantas de control, ambas rociadas con insecticida, no se observaron diferencias significativas (Marvier et al. 2007). Los datos del metanálisis del algodón Cry1Ac indican que la disminución en la abundancia de artrópodos no objetivo comparada con las plantas de control que no habían sido rociadas se debe principalmente a una disminución de los lepidópteros, pero también se observaron disminuciones menores en la cantidad de coleópteros y hemípteros (Marvier et al. 2007). Cuando se agrupó a los artrópodos según su función (depredadores, parásitos, mezcla, herbívoros, omnívoros, detritívoros), se observaron disminuciones significativas de los depredadores en el algodón Cry1Ac en comparación con el control sin rociar (Naranjo 2009, Wolfenbarger et al. 2008). La disminución fue una consecuencia de las disminuciones en dos familias (nabidae y coccinélidos) y no una disminución uniforme (Naranjo 2009, Wolfenbarger et al. 2008). Se ha demostrado que la disminución es intrascendente para el control biológico de plagas no objetivo (Naranjo 2005a, 2005b). Pleasants et al. 2001). Este resultado confirma las evaluaciones de riesgo anteriores, en las cuales se predecían impactos insignificantes debido a la baja exposición de lepidópteros no objetivo al polen o a otros tejidos de la planta que contengan Cry1Ac (CFIA 1997, USDA APHIS 1994, 1996, 1997a, 1997b, 1997c, 2000, 2003, USEPA 2001). Los resultados de estos estudios se sintetizan en la Tabla 4. Estudios de campo de los organismos no objetivo de Cry1Ac Varias revisiones y metanálisis han estudiado los resultados netos de gran parte de la bibliografía que existe acerca de los estudios de campo sobre 5 La base de datos de los efectos no objetivo de los cultivos Bt la conserva el Centro Nacional de Análisis y Síntesis Ecológicos (National Center for Ecological Analysis and Synthesis, NCEAS) http://delphi.nceas.ucsb.edu/btcrops/. Los documentos deben cumplir con los siguientes criterios para ser incluidos en la base de datos: (i) incluir una especie de cultivo que haya sido transformada genéticamente para que exprese uno o más genes cry derivados de Bacillus thuringiensis; (ii) medir los efectos del cultivo transformado para uno o más grupos de invertebrados no objetivo; (iii) incluir una comparación con un control no transgénico o un rango de niveles de exposición a la planta transgénica o a sus productos (por ejemplo, polen); y (iv) estar escrito en inglés. 5 Establecimiento y persistencia en el medioambiente de las plantas que expresan Cry1Ac Biología de la especie de planta Por lo general, el punto de comienzo de las evaluaciones de riesgo ambiental de las plantas GM es la familiaridad con la biología de la especie de planta sin transformar o planta huésped dentro del ambiente que la recibe (OCDE 2006). La información acerca de la biología de la planta huésped se puede utilizar para identificar las características específicas de la especie que pueden verse afectadas por los rasgos nuevos, de manera de permitir que la planta transgénica se transforme en "maleza", que invada hábitats naturales o que sea perjudicial para el ambiente, de cualquier manera. También puede brindar detalles acerca de las interacciones importantes entre la planta y los demás organismos, y esos detalles pueden ser importantes para considerar los posibles perjuicios. Al considerar la biología de la planta huésped, el evaluador de riesgo puede identificar los peligros posibles que pueden estar asociados con la expresión de la nueva proteína (por ejemplo, Cry1Ac) y luego puede evaluar la probabilidad de que dichos peligros se hagan realidad. Por ejemplo, si la especie de la planta está muy aclimatada y necesita de gran intervención humana para crecer o reproducirse, el evaluador puede tener en cuenta lo anterior al considerar la probabilidad de que la planta GM se establezca fuera del cultivo. Datos del fenotipo La información acerca del fenotipo de las plantas GM que expresan Cry1Ac se recaba del laboratorio, el invernadero y los estudios de campo, y se presenta en las presentaciones regulatorias para: (1) identificar todo cambio intencional al fenotipo que pueda afectar la seguridad ambiental de la planta; y (2) para identificar todo cambio no intencional hecho sobre la biología de la planta que pueda afectar la seguridad ambiental. En las presentaciones regulatorias y en las publicaciones de pares, los datos del fenotipo se han concentrado en las características de la planta que podrían contribuir a su supervivencia o persistencia (por ejemplo, el potencial para convertirse en maleza), o que podrían afectar negativamente el rendimiento agrícola (por ejemplo, la susceptibilidad frente a enfermedades y los datos de rendimiento) (CFIA 1997, CTNBio 2005, BCH de Japón 1997, 1999, 2007, OGTR 2002a, 2003b, 2003c, 2005, 2006b, USDA APHIS 1994, 1996, 1997a, 1997b, 1997c, 2000, 2003). Puesto que la proteína Cry1Ac tiene el propósito de ofrecer resistencia a las plagas de insectos objetivo, este dato se considera al momento de realizar las observaciones de fenotipo. Algunos de los datos recolectados son cuantitativos (por ejemplo, altura de la planta o porcentaje de germinación de la semilla), mientras que otros datos son cualitativos y de observación (por ejemplo, no hay diferencia en la susceptibilidad frente a enfermedades) (USDA APHIS 1994, 1996, 1997a, 1997b, 1997c, 2000, 2003). En muchos casos, se observaron diferencias estadísticamente significativas entre las plantas GM que expresan Cry1Ac y los controles, pero fueron pequeñas y entraban dentro del rango informado sobre la especie cultivada (USDA APHIS 1994, 1996, 1997a, 1997b, 1997c, 2000, 2003). Conjuntamente, los datos de fenotipo no presentaron ningún patrón de cambio que respaldara la hipótesis de que la introducción de la proteína Cry1Ac tuviera un efecto no intencional sobre la morfología general o las características del fenotipo de las plantas, además de conceder resistencia a los insectos 6 en las plagas de lepidópteros. En el Anexo II, se sintetizan los datos de fenotipo de las plantas GM que expresan Cry1Ac. Potencial para convertirse en maleza en ambientes agrícolas Tanto el maíz como el algodón tienen cierto potencial para ser "voluntarias" a convertirse en maleza en las temporadas posteriores de cultivo (OCDE 2003, OCDE 2008, OGTR 2008). Las características que influyen en la capacidad de una planta para ser voluntaria son, en gran medida, las mismas que generalmente están presentes para convertirse en maleza, por ejemplo, el reposo vegetativo de la semilla, la dehiscencia y la competitividad (Baker 1974). No existen datos que indiquen una vinculación entre la expresión de la proteína Cry1Ac y el aumento de la capacidad de supervivencia o de pasar el invierno que pudieran alterar la prevalencia de maíz o algodón voluntarios en las temporadas posteriores de cultivo (CFIA 1997, CTNBio 2005, BCH de Japón 1997, 1999, 2007, OGTR 2002a, 2003b, 2003c, 2005, 2006b, USDA APHIS 1994, 1996, 1997a, 1997b, 1997c, 2000, 2003). No se espera que los voluntarios de la temporada siguiente que expresen Cry1Ac presenten ninguna dificultad de control y se los puede tratar del mismo modo que a los voluntarios convencionales de maíz y algodón. Potencial para convertirse en maleza en ambientes no agrícolas Los principales mecanismos mediante los cuales se puede introducir Cry1Ac en un medioambiente no agrícola son el movimiento y el establecimiento de la planta GM fuera de las áreas cultivadas y del flujo de genes de la planta GM a una población naturalizada o a un grupo de familiares compatibles sexualmente (Mallory-Smith y Zapiola 2008). Las evaluaciones de riesgo de las plantas GM que expresan Cry1Ac han considerado los posibles impactos asociados con ambos tipos de movimiento (CFIA 1997, CTNBio 2005, BCH de Japón 1997, 1999, 2007, OGTR 2002a, 2003b, 2003c, 2005, 2006b, USDA APHIS 1994, 1996, 1997a, 1997b, 1997c, 2000, 2003). Si bien en ciertos contextos todas las plantas pueden considerarse malezas, no se considera que el maíz ni el algodón sean una maleza invasiva o agresiva fuera de los sistemas agrícolas. El maíz tiene una capacidad muy restringida para establecerse sin la intervención humana pero el algodón puede persistir bajo condiciones favorables y a veces puede necesitar control (OCDE 2003, OCDE 2008, OGTR 2008). Los datos agronómicos indican que la Cry1Ac no tiene un impacto significativo sobre los rasgos asociados con el potencial para convertirse en maleza (CFIA 1997, CTNBio 2005, BCH de Japón 1997, 1999, 2007, OGTR 2002a, 2003b, 2003c, 2005, 2006b, USDA APHIS 1994, 1996, 1997a, 1997b, 1997c, 2000, 2003). Aunque el no estar sometidas a los factores naturales de control (incluidos los insectos herbívoros) se ha presentado como explicación parcial del éxito de las especies invasivas (Blumenthal 2005, Keane y Crawley 2002, Mack 1996, Mason et al. 2004), la mayoría de las decisiones regulatorias han acordado que es poco probable que el agregado de resistencia a las plagas de lepidópteros haya conducido a que el algodón que expresa Cry1Ac se haya transformado en invasivo en los ambientes que no son agrícolas (CFIA 1997, CTNBio 2005, BCH de Japón 1997, 1999, 2007 USDA APHIS 1994, 1996, 1997a, 1997b, 1997c, 2000, 2003). Las decisiones regulatorias en Australia antes de 2006 restringían la liberación del algodón Cry1Ac en el norte de Australia debido a la duda acerca del impacto que la resistencia a los insectos tendría sobre la capacidad del algodón para persistir. Sin embargo, estudios posteriores señalaron que los lepidópteros herbívoros no representaban un factor significativo como para limitar la expansión del algodón en el norte de Australia y la restricción se eliminó (OGTR 2002a, 2003b, 2003c, 2005, 2006b, 2006c). Movimiento del transgen a familiares sexualmente compatibles El movimiento de transgenes de una planta GM a sus familiares silvestres se realiza mediante el polen, y la producción de híbridos reproductivamente viables depende de la proximidad física y temporal de las plantas GM con las especies compatibles sexualmente. Ni el maíz ni el algodón tienen familiares a los que se considere invasivos en el ecosistema ni malezas de importancia agrícola de amplia distribución para los que la hibridación es una preocupación (OCDE 2003, OCDE 2008, OGTR 2008). El maíz se hibrida libremente con teosintes silvestres, pero se cree que la introgresión de genes es limitada (Baltazar et al. 2005, OCDE 2003, Serratos et al. 1995). Las poblaciones de teosintes silvestres se limitan a la zona de México, Guatemala y una única población en Nicaragua; y si bien algunos agricultores mexicanos la consideran una maleza grave, otros la tratan como beneficiosa (Serratos et al. 1995). El algodón tiene varios familiares silvestres con los cuales potencialmente podría hibridarse (OCDE 2008, OGTR 2008). La EPA de los Estados Unidos ha restringido la liberación del algodón que expresa Cry1Ac en Hawaii debido a la incertidumbre acerca de los efectos que podría producir sobre las poblaciones de G. tomentosum (USEPA 2001). La EPA de los Estados Unidos también ha restringido su liberación en el sur de Florida porque se desconoce el impacto del flujo de genes sobre las G. hirsutum naturalizadas en cuanto al desarrollo de resistencia de los insectos (USEPA 2001). En Australia, el desconocimiento del impacto del flujo de genes sobre poblaciones naturalizadas de G. hirsutum y G. barbadense llevó a la restricción para plantar algodón Cry1Ac en el norte del país hasta 2006, cuando los estudios establecieron que la depredación de lepidópteros no era significativa en el control de dichas poblaciones (OGTR 2002a, 2003b, 2003c, 2005, 2006b, 2006c). Brasil también ha establecido una zona de exclusión para el cultivo de algodón Cry1Ac, con el propósito de evitar el posible flujo de genes hacia especies silvestres en el noroeste del país (CTNBio 2005). Análisis de composición de las plantas Cry1Ac El análisis detallado de la composición es un componente estrictamente científico de la caracterización de las plantas GM y es un requisito de las autoridades regulatorias para obtener las aprobaciones de seguridad de alimentos y pienso GM (Codex 2003a, 2003b, EFSA 2006A, FAO/ OMS 1996, OCDE 1992, OMS 1995). La selección de análisis que se realizan depende de la naturaleza del producto y su intención de uso. Los cultivos GM que expresan Cry1Ac y que presentan resistencia a los insectos, por lo general, se someten a un análisis proximal (proteína cruda, grasa cruda, fibra, humedad y cenizas) (ANZFA 2002, Berberich et al. 1996, CFIA 1996, 1997, 2002, 2003, 2005, CTNBio 2005, Hamilton et al. 2004, BCH de Japón 1997, 1999, 2007, OGTR 2002a, 2003b, 2003c, 2005, 2006b, USDA APHIS 1994, 1996, 1997a, 1997b, 1997c, 2000, 2003). También se han realizado los análisis detallados de la composición de ácidos grasos y aminoácidos, así como también el análisis de metabolitos secundarios importantes que tiene propiedades tóxicas o antinutricionales (por ejemplo, el gosipol en algodón) (ANZFA 2002, Berberich et al. 1996, CFIA 1996, 1997, 2002, 2003, 2005, CTNBio 2005, Hamilton et al. 2004, BCH de Japón 1997, 1999, 2007, OGTR 2002a, 2003b, 2003c, 2005, 2006b, USDA APHIS 1994, 1996, 1997a, 1997b, 1997c, 2000, 2003). Los datos recabados pueden ser útiles como indicadores de cambios no intencionales en la planta transformada (Codex 2003a, 2003b, Nickson y McKee 2002). En el Anexo III, se sintetizan los datos de los análisis de composición disponibles al público. Si bien se observaron algunas diferencias estadísticamente significativas en la composición, las plantas GM que expresan Cry1Ac estuvieron dentro del rango normal que se observa en la especie de cultivo (ANZFA 2002, Berberich et al. 1996, Hamilton et al. 2004, USDA APHIS 1996, 1997c, 2000, 2003). Los análisis regulatorios subsiguientes no consideraron que dichas diferencias fueran importantes en el contexto de seguridad ambiental (CFIA 1997, CTNBio 2005, BCH de Japón 1997, 1999, 2007, OGTR 2002b, 2003a, 2003c, 2006a, 2006c, USDA APHIS 1995, 1997d, 2001, 2004). Considerando los datos de los eventos aprobados, no hay patrones de cambios sistemáticos o confiables en la composición proximal de plantas que expresan Cry1Ac. Esto indica que la expresión de Cry1Ac no tiene ningún efecto biológicamente significativo sobre el metabolismo general de las plantas transformadas. Conclusión La proteína Cry1Ac expresada en plantas GM con resistencia a los insectos es derivada de las bacterias comunes del suelo Bacillus thuringiensis y es específicamente tóxica para los lepidópteros. Las pruebas de toxicidad con un rango representativo de organismos no objetivo (ONO) arrojó valores de NOEL en concentraciones que representan diez veces o más las concentraciones ambientales esperadas de Cry1Ac. Los metanálisis de los estudios de campo sugieren que el cultivo de plantas de algodón GM que expresan Cry1Ac redujo levemente la abundancia de artrópodos no objetivo en comparación con el algodón sin rociar, aumentó la abundancia de artrópodos en comparación con el algodón que había sido rociado con insecticida y no tuvo un efecto visible cuando se trató a las plantas GM y a los controles con insecticida, lo cual coincide con las prácticas convencionales de control de insectos. La Cry1Ac en plantas puede ser tóxica para los lepidópteros no objetivo, pero las evaluaciones regulatorias de riesgo para los productos aprobados han concluido que la baja probabilidad de exposición tiene como resultado un riesgo adicional insignificante en comparación con otras prácticas agrícolas. El peso de las pruebas de los análisis de fenotipo y de composición demuestra que la expresión de Cry1Ac en los eventos aprobados de algodón y maíz no alteró la fisiología general de la planta y dichas plantas no tienen más probabilidades de convertirse en malezas ni en invasoras que sus equivalentes convencionales. 7 Referencias Artículos de publicaciones y libros Accinelli C. Koskinen W. C., Becker J. M. y Sadowsky M. J. (2008). Mineralization of the Bacillus thuringiensis Cry1Ac endotoxins in soil. Journal of Agricultural and Food Chemistry. 56:1025-1028. Aronson A. I. y Shai Y. (2001). Why Bacillus thuringiensis insecticidal toxins are so effective: unique features of their mode of action. FEMS Microbiology Letters 195: 1-8. Baker H.G. (1974). The evolution of weeds. Annual Review of Ecology and Systematics 5: 1-24. Baltazar B.M., Sanchez-Gonzales J. J., Cruz-Larios, L. y Schoper, J.B. (2005). Pollination between maize and teosinte: an important determinant of gene flow in Mexico. Theoretical Applied Genetics 110:519-526. Berberich S. A., Ream J. E., Jackson T. L., Wood R., Stipanovic R., Harvey P., Patzer S. y Fuchs R. L. (1996). The composition of insect-protected cottonseed is equivalent to that of conventional cottonseed. Journal of Agricultural and Food Chemistry 44(1):365-371. Jenkins J. L., Lee M. K., Sangadala S., Adang M. J. y Dean D. H. (1999). Binding of Bacillus thuringiensis Cry1Ac toxin to Manduca sexta aminopeptidase-N receptor is not directly related to toxicity. Cartas de la Federación de las Sociedades Bioquímicas Europeas (FEBS) 462: 373-376. Jesse L. C. H. y Obrycki J. J. (2000). Field deposition of Bt transgenic corn pollen: lethal effects on the monarch butterfly. Oecologia 125:241-248. Keane R. M. y Crawley M. J. (2002). Exotic plant invasions and the enemy release hypothesis. Trends in Ecology and Evolution 17(4):164-170. Koskella J. y Stotzky G. (1997). Microbial utilization of free and clay-bound insecticidal toxins from Bacillus thuringiensis and their retention of insecticidal activity after incubation with microbes. Applied and Environmental Microbiology 63(9): 3561-3568. Kumar P. A., Sharma R. P. y Malik V.S. (1996). The insecticidal proteins of Bacillus thuringiensis. Advances in Applied Microbiology 42: 1-43. Losey J. E., Rayor L. S., Carter M. E. (1999). Transgenic pollen harms monarch larvae. Nature 399: 214. Mack R.N. (1996) Predicting the identity and fate of plant invaders: emergent and emerging approaches. Biological Conservation 78:107-121. Blumenthal D. (2005). Interrelated causes of plant invasion. Science 310: 243-244 Mallory-Smith C. y Zapiola M. (2008). Gene flow from glyphosate-resistant crops. Pest Management Science 64: 428-440. Bravo A. Gill S. S. y Soberon M. (2007). Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control. Toxicon. 49(4): 423-435. http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=1857359 Marchetti E., Accinelli C., Talame V. y Epifani R. (2007). Persistence of Cry toxins and cry genes from genetically modified plants in two agricultural soils. Agronomy for Sustainable Development 27(3): 231-236. CERA. (2010). Base de datos de cultivos GM. Center for Environmental Risk Assessment (CERA), ILSI Research Foundation, Washington D.C. http://ceragmc.org/index.php?action=gm_crop_database Marvier M., McCreedy C., Regetz J., Kareiva P. (2007). A meta-analysis of effects of Bt cotton and maize on nontarget invertebrates. Science 316:1475–1477. Crecchio C. y Stotsky G. (1998). Insecticidal activity and biodegradation of the toxin from Bacillus thuringiensis subsp. Kurstaki bound to humic acids from soil. Soil Biology and Biochemistry 30(4):463-470. Crickmore N., Zeigler D. R., Feitelson J., Schnepf E., Van Rie J., Lereclus D., Baum J. y Dean D. H. (1998). Revision of the nomenclature for the Bacillues thuringiensis pesticidal crystal proteins. Microbiology and Molecular Biology Reviews 62(3): 807-813. Crickmore N., Zeigler D.R., Schnepf E., Van Rie J., Lereclus D., Baum J., Bravo A. y Dean D.H. (2005). Bacillus thuringiensis Nomenclatura de toxinas (Página de inicio). [citado en enero de 2010]. http://www.lifesci.sussex.ac.uk/Home/Neil_ Crickmore/Bt/. Duan JJ, Marvier M, Huesing J, Dively G, Huang ZY (2008) A meta-analysis of effects of Bt crops on honey bees (Hymenoptera: Apidae). PLoS ONE 3(1):e1415 Duan JJ, Lundgren JG, Naranjo S, Marvier M (2010) Extrapolating non-target risk of Bt crops from laboratory to field. Biol Lett 6:74–77 Ellstrand N.C. (2003). Current knowledge of gene flow in plants: implications for transgene flow. Philosophical Transactions of the Royal Society, Series B 358:1163-1170. Hamilton K. A., Pyla P.D., Breeze M., Olson T., Li M., Robinson E., Gallagher S. P., Sorbet R. y Chen Y. (2004). Bollgard II cotton: compositional analysis and feeding studies of cottonseed from insect-protected cotton (Gossypium hirsutum L.) producing the Cry1Ac and Cry2Ab2 proteins. Journal of Agricultural and Food Chemistry 52: 6969-6976. Head G., Surber J. B., Watson J. A., Martin J. W. y Duan J. J. (2002). No detection of Cry1Ac protein in soil after multiple years of transgenic Bt cotton (Bollgard) use. Environmental Entomology 43(1): 30-36. Hellmich R. L. Siegfried B. D., Sears M. K., Stanley-Horn D. E., Daniels M. J., Mattila H. R., Spencer T., Bidne K. G. y Lewis L. C. (2001). Monarch larvae sensitivity to Bacillus thuringiensis-purified proteins and pollen. Actas de las Academias Nacionales de Ciencias de los EE. UU. 98(21):11925-11930. Hofte H. y Whiteley H. R. (1989). Insecticidal crystal proteins of Bacillus thuringiensis. Microbiological Reviews 53(2): 242-255. 8 Mason P., Braun L., Warwick S.I., Zhu B., Stewart Jr. C.N. (2003) Transgenic Bt-producing Brassica napus: Plutella xylostella selection pressure and fitness of weedy relatives. Environmental Biosafety Research 2:263-276. Naranjo S.E. (2005a) Long-term assessment of the effects of transgenic Bt cotton on the abundance of non-target arthropod natural enemies. Environmental Entomology 34: 1193–1210. Naranjo, S.E. (2005b) Long-term assessment of the effects of transgenic Bt cotton on the function of the natural enemy community. Environmental Entomology 34: 1211-1223. Naranjo S. E. (2009). Impacts of Bt crops on non-target invertebrates and insecticide use patterns. CAB Reviews: Perspectives in Agriculture, Veterinary Science, Nutrition and Natural Resources 4(11): http://fbae.org/2009/FBAE/ website/images/pdf/imporatant-publication/impacts-of-bt-crops-on-non-targetinvertebrates-and-insecticide-use-patterns.pdf Nickson T. E. y McKee M. J. (2002). Ecological assessment of crops derived through biotechnology. En Thomas J. A. y Fuchs R. L. (eds.) Biotechnology and safety assessment (tercera edición) (pp233-252). Academic Press, San Diego, CA NRC (1989). Field testing genetically modified organisms: framework for decisions. Academia Nacional de Ciencias, Comisión de la National Research Council (NRC) sobre la evaluación científica de la introducción al medioambiente de plantas y microorganismos genéticamente modificados. National Academy Press, Washington, D.C. NRC (2000). Genetically modified pest-protected plants: science and regulation. Academia Nacional de Ciencias, National Research Council (NRC). National Academy Press, Washington D.C. NRC (2002). Environmental effects of transgenic plants: the scope and adequacy of regulation. Academia Nacional de Ciencias, National Research Council (NRC). National Academy Press, Washington D.C. Pleasants J. M., Hellmich R. L., Dively G. P., Sears M. K., Stanley-Horn D. E., Mattila H. R., Foster J. E., Clark P. y Jones G. D. (2001). Corn pollen deposition on milkweeds in and near cornfields. Actas de las Academias Nacionales de Ciencias de los EE. UU. 98(21): 11919-11924. http://www.pnas.org/content/98/21/11919. full Rose R.I. (ed.) (2007) Informe oficial sobre las pruebas en niveles de los efectos de protectores insecticidas proteínicos incorporados a las plantas sobre invertebrados no objetivo para las evaluaciones regulatorias de riesgo. USDA-APHIS y Agencia de Protección Ambiental de los Estados Unidos, Washington, DC, EE. UU. http:// www.epa.gov/pesticides/biopesticides/pips/non-target-arthropods.pdf Romeis J., Meissle M . y Bigler F. (2006). Transgenic crops expressing Bacillus thuringiensis toxins and biological control. Nature Biotechnology 24(1): 63-71. Romeis J., Barsch D., Bigler F., Candolfi M. P., Gielkens M. M. C., Hartley S.E., Hellmich R. I., Huesing J. E., Jepson P. C., Layton R., Quemada H., Raybould A., Rose R. I., Schiemann J., Sears M. K., Shelton A. M., Sweet J., Vaituzis Z. y Wolt J. D. (2008). Assessment of risk of insect-resistant transgenic crops to nontarget arthropods Serratos J.A., Wilcox M.C., Castillo F. (Eds.) (1995). Actas de un foro: flujo de genes entre variedades autóctonas de maíz, variedades de maíz mejoradas y teosintes: consecuencias para el maíz transgénico. Insituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias de México (INIFAP), Centro Internacional de Mejoramiento de Maíz y Trigo (CIMMYT) y Comité Nacional de Bioseguridad Agrícola de México (CNBA). Schnepf E., Crickmore N., van Rie J., Lereclus D., Baum J., Fetelson J., Ziegler D. R. y Dean D. H. (1998). Bacillus thuringiensis and its pesticidal crystal proteins. Microbiology and Molecular Biology Reviews 62(3): 775-806. Tabashink B.E. (1992). Evaluation of synergism among Bacillus thuringiensis toxins. Applied and Environmental Microbiology 58(10): 3343-3346. Wolfenbarger L. L., Naranjo S. E., Lundgren J. G., Bitzer R. J. y Watrud L. S. (2008). Bt crops effects on functional guilds of non-target arthropods: a meta-analysis. PloS One 3(5): e2118 http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2346550/pdf/ pone.0002118.pdf Zhang X., Candas M., Griko N. B., Taussig R., Bulla L. A. Jr. (2006). A mechanism of cell death involving an adenylyl cyclase/PKA signaling pathway is induced by the Cry1Ab toxin of Bacillus thuringiensis. Actas de las Academias Nacionales de Ciencias de los EE. UU. 103(26):9897-9902. CFIA (2005). Documento de decisión DD2005-51: Determination of the safety of Dow AgroSciences Inc. insect resistant cotton (Gossypium hirsutum) event 3006210-23. Agencia de Inspección Alimentaria de Canadá (AIAC) Ottawa, Canadá. http://www.inspection.gc.ca/english/plaveg/bio/dd/dd0551e.shtml CFIA (2009). Introducción a las políticas de la CFIA sobre el manejo de casos de incumplimiento. Agencia de Inspección Alimentaria de Canadá (AIAC) Ottawa, Canadá. http://www.inspection.gc.ca/english/plaveg/bio/nonapp/nonappe.shtml. Codex (2003a). Principles for the risk analysis of foods derived through modern biotechnology. Comisión del Codex Alimentarius (Codex), Roma http://www. codexalimentarius.net/download/standards/10007/CXG_044e.pdf. Codex (2003b). Guideline for the conduct of food safety assessment of foods derived from recombinant DNA plants. Comisión del Codex Alimentarius (Codex), Roma http://www.codexalimentarius.net/download/standards/10021/CXG_045e. pdf. CTNBio (2005). Commercial release of genetically modified cotton, Bollgard Cotton (531). Comisión Técnica Nacional de Bioseguridad de Brasil (CTNBio), São Paulo http://www.ctnbio.gov.br/index.php/content/view/3663.html CTNBio (2009). Commercial release of genetically modified cotton, Bollgard Cotton (MON 15985): Technical Opinion no. 1832/2009. Comisión Técnica Nacional de Bioseguridad de Brasil (CTNBio), São Paulo. EC (2001). Directiva 2001/18/EC del Parlamento Europeo y del Consejo. Comisión Europea, Bruselas, Bélgica. http://ec.europa.eu/environment/biotechnology/pdf/ dir2001_18.pdf. EFSA (2006a). Guidance document of the scientific panel on genetically modified organisms for the risk assessment of genetically modified plants and derived food and feed. Autoridad Europea de Seguridad Alimentaria (EFSA), Bruselas, Bélgica. http://www.efsa.europa.eu/en/scdocs/doc/gmo_guidance_derived_feed_food.pdf. Publicaciones regulatorias EU DG ARD (2007). Economic impact of unapproved GMOs on EU feed imports and livestock production. Dirección General Europea de Agricultura y Desarrollo Rural (EU DG ARD), Bruselas, Bélgica. ANZFA (2002) Informe de evaluación preliminar de la solicitud A380: Food from insect-protected and glufosinate ammonium-tolerant DBT418 corn. Australia New Zealand Food Authority (ANZFA), Canberra, Australia y Wellington, Nueva Zelanda. EU DG SANCO (2010). Base de datos de pesticidas de la UE [búsqueda de Bacillus thuringiensis]. Dirección General de la Unión Europea, Salud y Política de Consumidores (EU DG SANCO) Bruselas, Bélgica http://ec.europa.eu/sanco_ pesticides/public/index.cfm?event=activesubstance.selection APVMA (2010). Sistema público de información sobre productos químicos (Public Chemical Information System, PUBCRIS) [búsqueda de Bacillus thuringiensis]. Australian Pesticides and Veterinary Management Authority (AVPMA), Symonston, Australia. http://services.apvma.gov.au/PubcrisWebClient/welcome.do EU SCP (1998). Opinión del comité científico sobre las plantas respecto del algodón modificado genéticamente y tolerante a los insectos notificado por la compañía Monsanto (notificación C/ES/96/02). Comisión Europea. Comité científico de las plantas de la Dirección General de Protección de la Salud y del Consumidor, Bruselas. CBD (2000a). Protocolo de Cartagena sobre Seguridad de la Biotecnología. Secretaría del Convenio sobre la Diversidad Biológica (CBD), Montreal. http:// www.cbd.int/biosafety/protocol.shtml. CBD (2000b). Protocolo de Cartagena sobre Seguridad de la Biotecnología Anexo III: Evaluación de riesgos. Secretaría del Convenio sobre la Diversidad Biológica (CBD), Montreal. http://www.cbd.int/biosafety/articles.shtml?a=cpb-43 CFIA (1996). Documento de decisión 96-14: Determination of environmental safety of Bollgard insect resistant cotton (Gossypium hirsutum L.). Agencia de Inspección Alimentaria de Canadá (AIAC) Ottawa, Canadá. http://www.inspection. gc.ca/english/plaveg/bio/dd/dd9614e.shtml CFIA (1997). Documento de decisión 98-23: Determination of Dekalb Genetics Corporation’s European Corn Borer Resistant Corn (Zea mays L.) Line DBT418. Agencia de Inspección Alimentaria de Canadá (AIAC) Ottawa, Canadá. http:// www.inspection.gc.ca/english/plaveg/bio/dd/dd9823e.shtml CFIA (2002). Acuerdo bilateral entre Canadá y Estados Unidos sobre biotecnología agrícola, Apéndice II: Environmental characterization data for transgenic plants intended for unconfined release. http://www.inspection.gc.ca/english/plaveg/bio/ usda/appenannex2e.shtml CFIA (2003). Documento de decisión DD2003-45: Determination of the safety of Monsanto’s insect resistant Bollgard II cotton (Gossypium hirsutum L.). Agencia de Inspección Alimentaria de Canadá (AIAC) Ottawa, Canadá. http://www.inspection. gc.ca/english/plaveg/bio/dd/dd0345e.shtml FAO/OMS (1996). Biotecnología y seguridad alimentaria. Informe de una conferencia conjunta de FAO/OMS. Organización de las Naciones Unidas para la Agricultura y la Alimentación (FAO)/Organización Mundial de la Salud (OMS), Documento 61 sobre alimentos y nutrición, Roma, Italia. http://www.fao.org/ag/ agn/food/pdf/biotechnology.pdf. FAO/OMS (2006). Food safety risk analysis: a guide for national food safety authorities. Organización de las Naciones Unidas para la Agricultura y la Alimentación (FAO) y Organización Mundial de la Salud (OMS), Roma. http:// www.who.int/entity/foodsafety/publications/micro/riskanalysis06.pdf. FSANZ (2004). Informe de evaluación final, formulario A518: Food derived from insect-protected, herbicide-tolerant cotton line MXB-13. Food Safety Australia New Zealand (FSANZ) Canberra, Australia y Wellington, Nueva Zelanda. http://www.foodstandards.gov.au/standardsdevelopment/applications/ applicationa518foodd2314.cfm BCH de Japón (1997). Outline of the biological diversity and risk assessment report (Mon-00531-6). Biosafety Clearing House (BCH) de Japón. Tokio, Japón. BCH de Japón (1999). Biological diversity risk assessment report: DBT418. Biosafety Clearing House (BCH) de Japón. Tokio, Japón. BCH de Japón (2007). Outline of the biological diversity risk assessment report: Type 1 use approval for cotton LLCotton25 x MON15985 (ACS-GHØØ1-3 x MON-15985-7). Biosafety Clearing House (BCH) de Japón. Tokio, Japón. OCDE (1992). Recombinant DNA safety considerations. Organización para la Cooperación y el Desarrollo Económico (OCDE), París, Francia. 9 OCDE (1993). Safety considerations for biotechnology: scale-up of crop plants. . Organización para la Cooperación y el Desarrollo Económico (OCDE), París, Francia. OGTR (2009b). Informe anual de las operaciones del regulador de la tecnología genética (08-09). Office of the gene technology regulatory (OGTR) Department of Health and Ageing, Canberra, Australia. OCDE (2003a). Consensus document on the biology of Zea mays subsp. Mays. Organización para la Cooperación y el Desarrollo Económico (OCDE), París, Francia. PMRA (2008). Re-evaluation decision document: Bacillus thuringiensis. Health Canada, Pest Management Regulatory Agency (PMRA), Ottawa. [consultado el 18 de febrero de 2010] http://www.hc-sc.gc.ca/cps-spc/pubs/pest/_decisions/rvd200818/index-eng.php OCDE (2003b). Description of selected key generic terms used in chemical hazard/risk assessment. Organización para la Cooperación y el Desarrollo Económico (OCDE), París. http://www.olis.oecd.org/olis/2003doc.nsf/LinkTo/ NT00004772/$FILE/JT00152557.PDF OCDE (2006). Points to consider for consensus documents on the biology of cultivated plants. Organización para la Cooperación y el Desarrollo Económico (OCDE), París, Francia. OCDE (2007) Consensus Document on Safety Information on Transgenic Plants Expressing Bacillus thuringiensis - Derived Insect Control Protein. Serie sobre la armonización de la supervisión regulatoria en Biotechnology, n.º 42. Organización para la Cooperación y el Desarrollo Económico, París. OCDE (2008). Consensus document on the biology of cotton (Gossypium spp.). Organización para la Cooperación y el Desarrollo Económico (OCDE), París, Francia. OCDE (2009). OECD schemes for the varietal certification or the control of seed. Organización para la Cooperación y el Desarrollo Económico (OCDE), París, Francia. OGTR (2002a).DIR 012/2002 Risk Assessment and Risk Management Plan: Commercial Release of Bollgard II cotton. Office of the Gene Technology Regulator (OGTR), Canberra, Australia. http://www.ogtr.gov.au/internet/ogtr/publishing.nsf/ Content/dir012-2002 OGTR (2003a) Notificación de la decisión de emitir una licencia en la solicitud de algodón DIR023/2002. Office of the Gene Technology Regulator (OGTR), Canberra, Australia. http://www.ogtr.gov.au/internet/ogtr/publishing.nsf/Content/ dir023-4/$FILE/dir023notific.rtf OGTR (2003b) Risk assessment and risk management plan for DIR023/2002. Office of the Gene Technology Regulator (OGTR), Canberra, Australia. http://www.ogtr. gov.au/internet/ogtr/publishing.nsf/Content/dir023-3/$FILE/dir023finalrarmp.pdf OGTR (2003c). DIR 022/2002 - Application and license for dealings involving an intentional release into the environment: Commercial release of insecticidal (INGARD event 531) cotton. Office of the Gene Technology Regulator (OGTR), Canberra, Australia. http://www.ogtr.gov.au/internet/ogtr/publishing.nsf/Content/ dir022-4/ OGTR (2005). Full risk assessment and risk management plan for commercial release of genetically modified cotton lines (MON-15985-7, MON-88913-8). Office of the Gene Technology Regulator (OGTR), Canberra, Australia. http://www.ogtr. gov.au/internet/ogtr/publishing.nsf/Content/dir059-3/$FILE/dir059finalrarmp1.pdf OGTR (2006a). Dir 059/2005 Notificación de la decisión de emitir una licencia. Office of the Gene Technology Regulator (OGTR), Canberra, Australia. http://www. ogtr.gov.au/internet/ogtr/publishing.nsf/Content/dir059-3/$FILE/dir059notific.pdf OGTR (2006b). Full risk assessment and risk management plan (RARMP) for commercial release of genetically modified (GM) herbicide tolerant and/or insect resistant cotton lines north of latitude 22o south. Office of the Gene Technology Regulator (OGTR), Canberra, Australia. http://www.ogtr.gov.au/internet/ogtr/ publishing.nsf/Content/dir066-3/$FILE/dir066rarmp2.pdf OGTR (2006c). Decisión de emitir una licencia para la solicitud DIR 066/2006: Commercial release of five herbicide tolerant and/or insect resistant GM cotton lines in Northern Australia. Office of the Gene Technology Regulator (OGTR), Canberra, Australia. http://www.ogtr.gov.au/internet/ogtr/publishing.nsf/Content/ dir066-3/$FILE/dir066notifc.pdf OGTR (2008). Biología de Gossypium hirsutum L. y Gossypium barbadense L. Office of the gene technology regulatory (OGTR) Department of Health and Ageing, Canberra, Australia. OGTR (2009a). Marco para el análisis de riesgo. Office of the gene technology regulatory (OGTR) Department of Health and Ageing, Canberra, Australia. 10 USDA APHIS (1994). Petición para la determinación del estado no regulado: línea de algodón Bollgard 531 (Gossypium hirsutum L.) con el gen de Bacillus thuringiensis subespecie kurstaki. http://www.aphis.usda.gov/brs/aphisdocs/94_30801p.pdf. USDA APHIS (1995). Determinación de USDA/APHIS sobre una petición 94-30801 de Monsanto Agricultural Company en busca del estado no regulado de líneas de algodón que resisten lepidópteros 531, 757, 1076. Departamento de Agricultura de los Estados Unidos, Servicio de Inspección de Sanidad Animal y Vegetal, Washington D.C. http://www.aphis.usda.gov/brs/aphisdocs2/94_30801p_com.pdf USDA APHIS (1996). Petición para que se determine el estado no regulado: maíz con protección contra insectos (Zea mays L.) con el gen cryIA(c) de Bacillus thuringiensis subespecie Kurstaki. Departamento de Agricultura de los Estados Unidos, Servicio de Inspección de Sanidad Animal y Vegetal, Washington D.C. http://www.aphis.usda.gov/brs/aphisdocs/96_29101p.pdf. USDA APHIS (1997a). Petición para que se determine el estado no regulado: línea de tomate 5345 que resiste los insectos (Lycopersicon esculentum) y que produce la proteína Cry1Ac que controla a los insectos de Bacillus thuringiensis subespecie kurstaki. Departamento de Agricultura de los Estados Unidos, Servicio de Inspección de Sanidad Animal y Vegetal, Washington D.C. http://www.aphis. usda.gov/brs/aphisdocs/97_28701p.pdf USDA APHIS (1997b). Petición para que se determine el estado no regulado: algodón BXN con Bt. Departamento de Agricultura de los Estados Unidos, Servicio de Inspección de Sanidad Animal y Vegetal, Washington D.C. http://www.aphis. usda.gov/brs/aphisdocs/97_01301p.pdf USDA APHIS (1997c) USDA/APHIS petición 97-013-01 p para la determinación de estado no regulado para los eventos 31807 y 31808. Departamento de Agricultura de los Estados Unidos, Servicio de Inspección de Sanidad Animal y Vegetal, Washington D.C. http://www.aphis.usda.gov/brs/aphisdocs2/97_01301p_com.pdf USDA APHIS (1997d). USDA/APHIS petición 96-291-01p para determinar el estado no regulado para la línea de maíz DBT418 protegida contra insectos Evaluación ambiental y hallazgo de impacto no significativo. Departamento de Agricultura de los Estados Unidos, Servicio de Inspección de Sanidad Animal y Vegetal, Washington D.C. http://www.aphis.usda.gov/brs/aphisdocs2/96_29101p_ com.pdf. USDA APHIS (2000). Solicitud para la determinación del estado no regulado del artículo regulado: evento de algodón Bollgard II n.º 15985 (Gossypium hirsutum L.) que produce la proteína Cry2Ab que controla los insectos derivada del Bacillus thuringiensis subespecie kurstaki. Departamento de Agricultura de los Estados Unidos, Servicio de Inspección de Sanidad Animal y Vegetal, Washington D.C. http://www.aphis.usda.gov/brs/aphisdocs/00_34201p.pdf USDA APHIS (2001). Aprobación de la petición de la Compañía Monsanto (00342-01p) en busca de la determinación de estado no regulado para el evento de algodón Bollgard II n.º 15985 que produce la proteína Cry2Ab que controla insectos, derivada del Bacillus thuringiensis subespecie kurstaki. Departamento de Agricultura de los Estados Unidos, Servicio de Inspección de Sanidad Animal y Vegetal, Washington D.C. http://www.aphis.usda.gov/brs/aphisdocs2/00_34201p_ com.pdf. USDA APHIS (2003). Petición para la determinación del estado no regulado: B.t. Cry1Ac evento de algodón resistente a los insectos n.º 3006-210-23. Departamento de Agricultura de los Estados Unidos, Servicio de Inspección de Sanidad Animal y Vegetal, Washington D.C. http://www.aphis.usda.gov/brs/aphisdocs/03_03602p. pdf. USDA APHIS (2004). Aprobación de peticiones de Mycogen/Dow 03-036-01p y 03-036-02p en busca de la determinación del estado no regulado para los eventos de algodón resistente a los insectos n.º 281-24-236 y 3006-210-23 genéticamente modificados para que expresen B.t. Cry1F y Cry1Ac sintéticas, respectivamente: evaluación ambiental y hallazgo de impacto no significativo. Departamento de Agricultura de los Estados Unidos, Servicio de Inspección de Sanidad Animal y Vegetal, Washington D.C. http://www.aphis.usda.gov/brs/aphisdocs2/03_03601p_ com.pdf. USDA APHIS (2007b). Política de APHIS en respuesta a la presencia en bajo nivel de plantas genéticamente modificadas reguladas. Departamento de Agricultura de los Estados Unidos, Servicio de Inspección de Sanidad Animal y Vegetal, Washington D.C. USEPA (1992). Framework for ecological risk assessment. Foro de evaluación de riesgos, Agencia de Protección Ambiental de los Estados Unidos (USEPA), Washington D. C. http://oaspub.epa.gov/eims/eimscomm. getfile?p_download_id=36361http:// cfpub.epa.gov/ncea/cfm/recordisplay. cfm?deid=30759. USEPA (1998). Guidelines for ecological risk assessment. Foro de Evaluación de Riesgos, Agencia de Protección Ambiental de los Estados Unidos (USEPA), Washington D. C. http://oaspub.epa.gov/eims/eimscomm. getfile?p_download_id=36512 USEPA (2001). Bt Plant-Incorporated Protectants October 15, 2001 Biopesticides Registration Action Document. Agencia de Protección Ambiental de los Estados Unidos (USEPA), Washington D. C. http:// www.epa.gov/pesticides/biopesticides/pips/ bt_brad.htm USEPA (2007). White paper on tier-based testing for the effects of proteinaceous insecticidal plant-incorporated protectants on non-target arthropods for regulatory risk assessments. Agencia de Protección Ambiental de los Estados Unidos (USEPA), Washington D. C. http://www.epa.gov/ pesticides/biopesticides/pips/non-targetarthropods.pdf USEPA (2010). Oficina de prevención, pesticidas y sustancias tóxicas (Office of Prevention, Pesticides and Toxic Substances, OPPTS), Pautas armonizadas de prueba. Agencia de Protección Ambiental de los Estados Unidos (USEPA), Washington D. C. http://epa.gov/oppts/pubs/frs/home/guidelin. htm USFDA (1997). Biotechnology Consultation Note to the File BNF No. 000047. Administración de Alimentos y Medicamentos de Estados Unidos (FDA), Silver Spring MD. http://www.fda.gov/Food/ Biotechnology/Submissions/ucm161166. htm OMS (1995). Application of the Principles of Substantial Equivalence to the Safety Evaluation of Foods or Food Components from Plants Derived by Modern Biotechnology. Informe de un taller de la OMS. Organización Mundial de la Salud (OMS), Ginebra. Anexo I: Resumen de datos de expresión de la proteína Cry1Ac Las tablas que aparecen a continuación presentan un resumen de datos de publicaciones revisadas por pares y de presentaciones regulatorias. Los datos se presentan en el formato en el que están disponibles en el documento citado, con el propósito de facilitar las referencias cruzadas. En las fuentes citadas, se puede encontrar información adicional acerca de las metodologías de recolección y de toma de muestras. Tabla I.1. Expresión de Cry1Ac en el evento DBT418 con Zea mays (USDA APHIS 1996).1 Tejido Genotipo Hoja Autógamo Etapa de crecimiento V6-V7 Media SE5 2 SE5 Media SE5 Media SE5 7,12 88,1 19,7 ND ND 240,4 52,22 ND ND 24,6 1,94 ND ND 324,6 44,14 ND ND 45,8 9,65 ND ND 626,8 141,62 ND ND Autógamo ND ND 5,7 0,84 ND ND 36,7 13,88 ND ND Het.3 ND ND BLD7 BLD ND ND 12,1 2,18 ND ND ND ND BLD BLD ND ND 34,2 7,48 ND ND Autógamo 7,0 1,75 11,1 1,78 ND ND 10,8 1,87 ND ND Het.3 5,1 0,97 8,2 2,31 ND ND 10,7 1,51 ND ND 2 2 Híbrido 11,9 1,72 10,2 2,92 ND ND 23,1 3,12 ND ND Autógamo2 ND ND ND ND ND ND 42,86 16,60 ND ND Het.3 ND ND ND ND ND ND 37,16 3,976 ND ND Híbrido4 ND ND ND ND ND ND 36,06 8,146 ND ND Autógamo ND ND BLD BLD ND ND ND ND ND ND Het. ND ND 14,1 1,37 ND ND ND ND ND ND Híbrido4 ND ND BLD BLD ND ND ND ND ND ND Autógamo ND ND BLD BLD ND ND ND ND ND ND Het.3 ND ND BLD BLD ND ND ND ND ND ND ND ND BLD BLD ND ND ND ND ND Autógamo ND ND 27,6 9 ND ND ND ND 124,2 16,476 Het. ND ND 6,7 1,02 ND ND ND ND 41,2 6,426 Híbrido4 ND ND 14,1 2,82 ND ND ND ND 69,96 17,766 Autógamo2 ND ND ND ND 97,2 10,66 ND ND ND ND Het. ND ND ND ND 19,4 3,25 ND ND ND ND Híbrido4 ND ND ND ND 59,5 18,18 ND ND ND ND 2 2 Híbrido Planta completa sin cepellón Media 6,5 4 Planta completa SE5 5,66 3 Polen Media 44,6 4 Seda Senescencia 27,4 Híbrido Grano6 Recolección Híbrido4 4 Cepellón Pasta Het. 3 Tallo 33,6 Polen derramado 2 3 3 ND 6 6 1 Los datos se presentan en ng/g en peso fresco, a menos que se indique lo contrario. 2 Autógamos (2/3 alelos de Cry1Ac). 3 Híbrido comercial (Heterocigótico de Cry1Ac). 4 Híbrido (Homocigótico de Cry1Ac). 5 Error típico. 6 ng/g peso seco. 7 BLD = Por debajo del límite de detección (51,6 ng/g peso seco). 8 ND = No disponible. 11 Tabla I.2. Expresión de Cry1Ac en Gossypium hirsutum evento MON-00531-6 (USDA APHIS 1994).1 Tejido 1992 1993 Tabla I.6. Expresión de Cry1Ac en Gossypium hirsutum evento DAS 21023 (USDA APHIS 2003).1 1999 Tejido Media Rango Media Rango Media Rango Media SD Hoja joven (3-6 semanas) 1,92 0,46-3,5 Hoja joven 1,56 1,10-2,04 2,59 0,41-5,91 5,00 1,84 Hoja terminal 1,44 0,24-2,4 Hoja junio Hoja julio Hoja agosto 3,55 ND 0,13 ND Hoja septiembre 1,3 ND 0,23 ND Semilla 2 1,40 ND 3 5,12 ND ND ND Flor 1,92 1,3-2,4 1,49 ND 3,21 ND ND ND Cuadrado 1,84 1,0-3,1 ND ND Cápsula (inicial) 0,77 0,46-1,1 ND ND Planta completa (almácigo) 1,59 0,8-2,2 0,86 0,49-1,62 2,18 1,13-3,41 4,30 0,86 Planta completa (polinización) 1,15 0,57-2,1 Polen 11,5 ng/g ND ND ND ND ND Planta completa (defoliación) 0,81 0,31-1,3 Néctar BLD 4 ND ND ND ND ND Raíz (almácigo) 0,20 0,092-0,44 Planta completa 0,044 ND ND ND ND ND 1 Los datos se presentan en µg/g en peso fresco, a menos que se indique lo contrario. 2 Desviación típica. 3 ND = No disponible. 4 BLD= Por debajo del límite de detección (1,6 ng/g). Media Hojas -- 20 días después de la plantación 2,93 Hojas -- 130 días después de la plantación 3,02 Semillas 6,88 Tabla I.4. Expresión de Cry1Ac en Gossypium hirsutum evento MON-00531-6 (OGTR 2002).1 Hoja 1998 ND-0,23 ND-0,11 Polen3 1,44 0,9-2,4 Semilla3 0,57 0,332-0,78 Tabla I.7. Expresión de Cry1Ac en la Expresión de Cry1Ac en Gossypium hirsutum evento DAS-21023-5 X DAS-24236-5 (FSANZ 2004).1 1 Los datos y los valores de los estudios de campo en Brasil se expresan en µg/g en peso fresco. Tejido 0,10 0,05 1 Los valores están expresados en µg/g en peso seco, a menos que se indique lo contrario. 2 Por debajo del límite de cuantificación del método (0,001 a 0,375 ng/mg según la matriz). 3 Los valores se expresan en µg/g en peso fresco. Tabla I.3. Expresión de Cry1Ac en Gossypium hirsutum evento MON-00531-6 (CTNBio 2005).1 Tejido Raíz (polinización) Raíz (defoliación) 1999 Tejido Media Hoja joven (3-6 semanas) 1,82 Hoja terminal 1,31 Flor 1,83 Cuadrado 1,82 Cápsula (inicial) 0,64 Media STD Media STD Planta completa (almácigo) 1,37 1,95 1,21 2,05 0,71 Planta completa (polinización) 1,05 Semilla 3,22 0,77 2,64 0,63 Planta completa (defoliación) 0,6 Planta completa 0,13 0,04 <0,07 ND2 Raíz (almácigo) 0,17 Polen 0,04 0,01 0,01 <0,01 Raíz (polinización) 0,072 Raíz (defoliación) ND Polen 1,45 1 Los datos informados están expresados en µg/g en peso fresco (estos datos provienen de la presentación regulatoria del Evento 15985). 2 ND = No disponible. Tabla I.5. Expresión de Cry1Ac en Gossypium hirsutum evento MON15985-7 (OGTR 2002).1 Tejido 1998 Media 1999 SD2 Media 0,55 Semilla 3 1 Los datos se presentan en µg/g en peso seco, a menos que se indique lo contrario. 2 Los datos están calculados con algunos valores que se encuentran por debajo del límite de cuantificación del método (0,001 a 0,375 ng/mg según la matriz). 3 Los valores se expresan en µg/g en peso fresco. SD Hoja 2,75 1,32 2,07 0,61 Semilla 3,35 0,63 2,6 0,66 Planta completa 0,17 0,08 0,08 0,01 Polen 0,02 0,01 0,05 0,07 1 MON-15985-7 es una retransformación de MON-00531-6 con una proteína Cry adicional. Estos datos se pueden considerar como información adicional del evento MON-00531-6. Los datos se informan en µg/g en peso fresco. 2 SD= Desviación típica. 12 3 Tabla I.8. Expresión de Cry1Ac en Gossypium hirsutum evento 31807/31808 (FDA 2007). Tejido Expresión máxima Semilla 2,5 ppm1 1 Equivalente a 2,5 µg/g peso fresco. Anexo II: Resumen de análisis de fenotipo de las plantas GM que expresan Cry1Ac Las tablas que aparecen a continuación presentan un resumen de datos de publicaciones revisadas por pares y de presentaciones regulatorias. Los datos se presentan en el formato en el que están disponibles en el documento citado, con el propósito de facilitar las referencias cruzadas. En las fuentes citadas, se puede encontrar información adicional acerca de las metodologías de recolección y de toma de muestras. Tabla II.1. Resumen del análisis de fenotipo del evento MON-00531-6 (USDA APHIS 1996).1 Característica del fenotipo Resultado informado Observaciones de diferencias Potencial para convertirse en maleza Sin diferencias significativas2 Brote Sin diferencias significativas2 Aumento del brote en un lugar Vigor del almácigo Sin diferencias significativas2 Aumento del vigor en un lugar Reposo vegetativo Sin diferencias significativas Cierto reposo vegetativo en un lugar aparentemente debido a semillas producidas en invernadero Germinación Sin diferencias significativas2 Morfología Sin diferencias significativas2 Tiempo hasta la floración Sin diferencias significativas2 Fructificación Sin diferencias significativas2 Formación de la cápsula Sin diferencias significativas2 Desarrollo de la cápsula Sin diferencias significativas2 Producción Sin diferencias significativas2 Susceptibilidad a la enfermedad Sin diferencias significativas2 Voluntariado Sin diferencias significativas2 2 1 Resumido a partir del texto descriptivo de USDA APHIS 1996. 2 Aquí "significativo" no hace referencia a la significancia estadística. La información sobre el análisis estadístico no está incluida. Tabla II.2. Resumen de brote medio, floración y fechas de recolección de los Eventos MON-15985-7 y MON-00531-6 (USDA APHIS 2000). 1 Centro Evento o n.º de línea Porcentaje de Porcentaje de Primera flor blanca Fecha del primer Fecha de recolección almácigos brotados almácigos brotados observada recuento de cápsulas (7 días) (14 días) partidas 71 82 21/7/1998 8/9/1998 16/10/1998 Arizona 1 15985 80 85 21/7/1998 8/9/1998 16/10/1998 DP50B2 DP503 70 73 21/7/1998 8/9/1998 16/10/1998 Arizona 2 15985 51 72 27/7/1998 8/9/1998 18/11/1998 DP50B 70 76 23/7/1998 8/9/1998 18/11/1998 DP50 49 62 23/7/1998 8/9/1998 18/11/1998 68 2/8/1998 21/9/1998 27/10/1998 Louisiana 1 15985 52 DP50B 48 56 31/7/1998 21/9/1998 27/10/1998 31/7/1998 21/9/1998 27/10/1998 DP50 48 67 31/8/1998 14/10/1998 Louisiana 2 15985 73 82 20/7/1998 DP50B 68 71 20/7/1998 31/8/1998 14/10/1998 14/10/1998 DP50 54 56 20/7/1998 31/8/1998 Mississippi 1 15985 70 75 30/7/1998 4/9/1998 19/10/1998 DP50B 75 76 30/7/1998 4/9/1998 19/10/1998 DP50 78 62 30/7/1998 4/9/1998 19/10/1998 Mississippi 2 15985 63 78 20/7/1998 8/9/1998 5/10/1998 DP50B 76 73 20/7/1998 8/9/1998 5/10/1998 DP50 74 69 20/7/1998 8/9/1998 5/10/1998 Carolina del Sur 15985 88 94 23/7/1998 9/9/1998 27/10/1998 DP50B 77 87 23/7/1998 9/9/1998 27/10/1998 DP50 60 78 23/7/1998 9/9/1998 27/10/1998 15985 73 93 23/7/1998 2/9/1998 28/9/1998 Texas4 DP50B 83 83 20/7/1998 2/9/1998 28/9/1998 DP50 64 69 23/7/1998 2/9/1998 9/10/1998 1 USDA APHIS 2000 es una presentación regulatoria de MON-15985-7 que contiene información acerca de MON-00531-6 como control parental GM. 2 DP50B = progenitor transgénico de 15985 (evento 531). 3 DP50 = progenitor no transgénico de DP50. 4 La fechas de recolección del centro de Texas son distintas debido a una humedad excesiva que hubiese aumentado la putrefacción de la cápsula. 13 Tabla II.3. Resumen de altura media: proporción de nudos, días hasta la floración máxima y recuento total de cápsulas partidas del Evento MON15985-7 y MON-00531-6 (USDA APHIS 2000). Evento o n.º de línea Altura: Proporción de nudos Evento o n.º de línea 159852 % 4.º día de germinación 76 % 9.º día de germinación 77 % de germinación fría a 18 ºC Día 7 72 15985 1,70 Promedio de días hasta la floración máxima 15,29 407 DP504 DP50B2 1,77 15,03 431 DP50 1,72 15,77 284 1 USDA APHIS 2000 es una presentación regulatoria de MON-15985-7 que contiene información acerca de MON-00531-6 como control parental GM. 2 15985 = MON-15985-7. 3 DP50B = progenitor transgénico de 15985, MON-00531-6. 4 DP50 = progenitor no transgénico de DP50. 3 Promedio de cantidad total de cápsulas partidas por parcela Tabla II.4. Pruebas de germinación y de vigor en el almácigo sobre las semillas de dos ubicaciones del MON-15985-7 (USDA APHIS 2000).1 1 USDA APHIS 2000 es una presentación regulatoria de MON-15985-7 que contiene información acerca de MON-00531-6 como control parental GM. 2 DP50B = progenitor transgénico de 15985. 3 DP50 = progenitor no transgénico de DP50. DP50B3 83 83 80 88 89 82 Tabla II.5. Resultados de germinación y reposo vegetativo de las semillas recolectadas en las tres ubicaciones de 1999 del Evento MON-15985-7 y MON-00531-6 (USDA APHIS 2000).1 Media de psdv3 Media de pgerm4 (%) Media de psvfh5 (%) Media de pdegen6 (%) (en reposo vegetativo) (%) 1,2 0,0 95,1 4,1 5 1598 0,0 0,0 95,2 5,4 DP50B2 Rango de ref. (0-41) (0-1) (53-99) (1-20) 26,46 0,0 1,2 73,96 10 1598 0,0 1,3 78,5 21,7 DP50B1 Rango de ref. (0-28) (0-3) (38-91) (9-62) 0,0 95,4 0,0 5,46 20 1598 0,0 97,4 0,0 3,1 DP50B1 Rango de ref. (0-6) (74-100) (0-13) (0-26) 0,0 93,96 0,0 6,66 30 1598 0,0 98,6 0,0 2,2 DP50B1 Rango de ref. (0-0) (83-100) (0-0) (0-17) 0,0 85,9 0.0 14,9 40 1598 0,0 89,3 0,0 11,1 DP50B1 Rango de ref. (0-0) (70-96) (0-0) (4-30) 0,0 NC7 NC7 NC7 5/20 1598 0,1 NC7 NC7 NC7 DP50B1 Rango de ref. (0-29) NC7 NC7 NC7 7 10/20 1598 1,9 7,5 0,0 NC DP50B1 0,0 NC7 1,2 5,8 (0-79) (1-31) Rango de ref. (0-18) NC7 0,0 NC7 0,0 5,1 20/30 1598 0,0 NC7 0,0 3,7 DP50B1 Rango de ref. (0-2) NC7 (0-1) (0-17) 1 USDA APHIS 2000 es una presentación regulatoria de MON-15985-7 que contiene información acerca de MON-00531-6 como control parental GM. 2 DP50B = progenitor transgénico de 15985, MON-00531-6. 3 psdv = porcentaje de semillas duras viables. 4 pgerm = porcentaje de semillas germinadas. 5 psvfh = porcentaje de semillas viables firmes e hinchadas. 6 pdegen = porcentaje de semillas degeneradas. 7 Indica una diferencia significativa en comparación con DP50B por cuanto P ≤0,05. 8 NC = no comparado. No se comparan las medias combinadas posibles debido a una variedad significativa por interacción del centro por cuanto P ≤0,05. Informes anecdóticos adicionales: se informa que la susceptibilidad frente a la enfermedad en MON-15985-7 es similar al control. Temperatura (ºC) 14 Variedad Tabla II.6. Características agronómicas del Evento DAS-21023-5 Líneas que expresan la proteína Cry1Ac en comparación con la variedad progenitora PSC355 (USDA APHIS 2003). Variable Unidades 3006-210-3 (Cry1Ac) PSC355 (Nulo) Cantidad de ubicaciones Hábito de crecimiento Altura de la planta Pulgadas 39,9 41,5 17 Total de nudos Cantidad por planta 17,4 17,6 16 Altura: proporción de nudos Pulgadas por planta 2,29 2,35 17 Nudos de la primera rama con fructificación Nudo 6,7 6,6 17 Ramas con fructificación Cantidad por planta 11,7 12,1 16 Posición de fructificación total Cantidad por planta 25,6 26,6 17 Germinación y floración vegetal prematura Cantidad por planta 2,3 1,6 16 Brote en el campo % 63,6 82,3 19 Vigor frío % 32 38 20 Germinación y brote 4 días con calor % 64 65 20 7 días con calor % 80 82 20 Germinación total % 85 87 20 Semilla en reposo vegetativo % 0,6 0,3 20 Cantidad por planta 2,9 2,6 16 Vigor vegetativo Ramas vegetativas Período de floración Días hasta la primera flor Días 61,9 60,6 18 Nudo de flor blanca – 15 días Nudo 12,9 12,9 17 Nudo de flor blanca – 30 días Nudo 17,0 16,8 15 Potencial reproductivo Porcentaje de retención – total % 49,0 44,4 16 Porcentaje de retención – 1.a posición % 58,5 54,3 16 Porcentaje de cápsulas abiertas % por planta 73,5 75,4 17 Peso de semilla de algodón por cápsula Gramos por cápsula 5,5 5,1 19 Porcentaje de pelusa % 37,9 37,3 19 Índice de semilla (con pelusa) Gramos por 100 semillas 11,0 10,7 17 Pelusas por acre Libras por acre 1005 993 17 Largo Pulgadas 1,160 1,147 19 Fuerza Gramos por tex 31,9 32,6 19 Calidad de la fibra Micronaire Unidades de micronaire 4,51 4,96 19 Uniformidad del largo % 85,8 85,7 19 Reflectancia % 76,0 74,6 19 Amarilleamiento Escala de Hunter +b 8,3 8,4 19 Informes anecdóticos adicionales: susceptibilidad frente a la enfermedad (sin diferencia). 15 Tabla II.7. Ejemplos de características de rendimiento en el campo de los eventos 31807 y 31808 en comparación con la variedad comercial utilizada como control (USDA APHIS 1997c). Parámetro de evaluación % de frutas dañadas por Helicoverpa zea (% transformado por la raíz cuadrada de su arcoseno) Cuadrados dañados por Heliothis zea Control (por ejemplo, Coker 130) 12,87 Evento 31807 Evento 31808 1,88 0,61 15,4 2,92 2,92 0 0 0 Incidencia de tumores del cuello Infección con virus del mosaico de la coliflor Susceptibilidad frente a Phomopsis, Verticillium y demás patógenos de hongos normales del algodón Niveles de plagas con insectos que no son objetivo, por ejemplo, el pulgón del algodón, chinches manchadas, ácaros tetraníquidos y gorgojos de la cápsula Tolerancia al bromoxinil 0 0 0 Dentro del rango esperado Dentro del rango esperado Dentro del rango esperado Dentro del rango esperado Dentro del rango esperado Dentro del rango esperado No Sí Sí Germinación de la semilla normal normal normal Morfología de la planta normal normal normal Período de floración normal normal normal Producción normal normal normal Calidad de la fibra normal normal normal 0 0 0 Incidencia de plantas de algodón voluntarias y fuera de estación Tabla II.8. Comparaciones medias de los porcentajes de germinación del Evento 31807 (USDA APHIS 1997c). Evento o cepa N Porcentaje de germinación cálida Porcentaje de germinación fría 317071 4 99 92 31803 4 98 86 3 97 87 4 97 89 4 97 92 4 95 88 1 31807 A 31807 C Coker 130 2 ST4742 LSD (0,05) CV (%) 2 5 1,6 3,5 1 Plantas GM que expresan Cry1Ac. 2 Variedades sin transformación. Tabla II.9. Rendimiento agronómico de DBT418-híbrido convertido (evento DK-B89614-9) en comparación con la versión convencional del mismo híbrido (USDA APHIS 1996).1 Rasgo Equivalente del híbrido sin convertir DBT418 Rendimiento (busheles/acre) 130,4 129,5 Humedad de los granos (%) 13,9 14,32 Peso de prueba (lb) 55,0 55,0 Recuento del espacio final 61,2 61,1 Vigor del almácigo (escala 1 a 9) 6,5 6,21 Altura de la planta (in) 89,2 88,5 Altura de la espiga (in) 42,5 41,0 Suma térmica del polen 1339 1342 Suma térmica de la seda 1335 13421 Capacidad de permanecer verde (escala de 1 a 9) 4,2 4,31 Sin daños (escala de 1 a 9) 4,1 4,91 Espigas caídas (%) 0,04 0,04 Tallos alojados (%) 3,1 2,9 Raíces alojadas (%) 2,9 5,0 Plantas estériles (%) 3,4 5,1 1 Informes anecdóticos adicionales: susceptibilidad frente a la enfermedad (sin diferencia). 2 Estadísticamente distintos del control en el nivel P = 0,5. 16 Anexo III: Resumen de análisis de composición de las plantas GM que expresan Cry1Ac Las tablas que aparecen a continuación presentan un resumen de datos de publicaciones revisadas por pares y de presentaciones regulatorias. Los datos se presentan en el formato en el que están disponibles en el documento citado, con el propósito de facilitar las referencias cruzadas. En las fuentes citadas, se puede encontrar información adicional acerca de las metodologías de recolección y de toma de muestras. Tabla III.1. Análisis proximal de los granos de Zea mays evento DBT418 (ANZFA 2002).1 Componente analizado DBT4182 Media Control2 Proteína 9,02 Desviación típica 0,22 Aceite 4,05 0,05 Media Desviación típica 8,56 0,16 3,92 Tabla III.2. Análisis proximal del forraje de Zea mays evento DBT418 (ANZFA 2002).1 Rango de bibliografía Componente analizado DBT4182 6,0-12,0 Proteína 6,81 Desviación típica 0,23 3,1-5,7 Aceite 2,77 0,07 2,82 0,04 Media Control3 Media Desviación típica 7,12 0,29 0,06 Rango de bibliografía 3,5-15,9 0,7-6,7 Fibra 1,96 0,03 2,02 0,03 2,0-5,5 Fibra 20,56 0,03 20,57 0,38 2,0-5,5 Ceniza 1,32 0,01 1,30 0,02 1,1-3,9 Ceniza 4,33 0,15 4,28 0,13 1,3-10,5 Humedad 8,14 0,04 8,22 0,04 7-23 Humedad 66,68 0,04 66,96 0,04 ND 1 Los valores son en porcentaje de peso seco. 2 Tamaño de la muestra = 30. 1 Los valores son en porcentaje de peso seco. 2 Tamaño de la muestra = 24. 3 Tamaño de la muestra = 30. Tabla III.3. Análisis proximal de la semilla de Gossypium hirsutum evento MON-00531-6 (Berberich et al.1996).1 Característica Coker 312 Media Rango MON-00531-6 Media Rango Rango de bibliografía Proteína 27,00 23,3-28,4 27,56 22,8-31,0 12-32 Grasa 22,95 19,6-25,1 23,23 22,2 16,1-26,7 Ceniza 4,63 4,3-5,0 4,53 3,9-4,7 4,1-4,9 Carbohidratos 45,40 42,8-47,6 44,68 42,0-46,7 ND2 Calorías/100 g 496,32 479-508 498,11 495-511 ND Humedad 12,36 9,6-15,9 13,43 11,2-14,7 5,4-10,1 1 Todos los valores son en % de peso seco, excepto la humedad (porcentaje de peso fresco) y las calorías/100 g. 2 ND= No disponible. Tabla III.4. Análisis proximal de la semilla de Gossypium hirsutum evento MON-15985-7 (USDA APHIS 2000).1 Componente Evento 15985 % de proteína 26,13 21,45-28,82 26,06 21,93-28,15 25,96 21,76-27,79 Rango de referencia comercial 21,76-28,15 % de grasa 20,52 17,54-27,42 20,37 16,04-23,48 19,74 15,44-23,64 15,44-23,83 Media Rango DP50B (evento 531) Media Rango DP50 (no transgénico) Media Rango % de ceniza 4,36 3,93-4,81 4,38 4,06-4,67 4,34 3,76-4,85 3,76-4,85 % de fibra, cruda 16,83 14,93-17,95 17,17 15,42-19,69 17,19 15,38-19,31 15,38-20,89 % de carbohidratos 49,09 42,97-52,69 49,23 46,85-51,93 49,94 45,64-52,44 45,64-53,62 Calorías/100 g PS 485,33 468,50-520,01 484,45 463,09-498,71 481,57 457,77-499,84 457,77-500,49 5,99 4,34-7,59 6,05 4,22-7,28 6,03 3,97-7,26 3,97-8,47 % de humedad 1 MON-15985-7 es MON-00531-6 retransformado para que exprese una proteína Cry adicional. Estos datos pueden considerarse información adicional sobre el evento de Cry1Ac MON-00531-6. No se informan diferencias estadísticamente significativas entre 15985 y la línea progenitora de DP50B. 17 Tabla III.5. Análisis proximal de la semilla de Gossypium hirsutum evento MON-15985-7 (Hamilton et al. 2004).1 Componente MON 15985 DP50 Rango de bibliografía Media Rango Media Rango 4,28 3,85-4,92 4,32 3,76-5,23 3,87-5,29 Calorías (Kcal/100 g) 489,65 468,50-520,01 487,11 457,77-501,84 471,39-506,95 Carbohidratos 47,95 42,97-52,69 48,55 43,69-52,44 45,28-53,62 Grasa total 21,33 17,54-27,42 20,85 15,44-24,29 17,37-25,16 Ceniza Fibra cruda 16,07 13,81-17,95 16,22 13,45-19,31 13,85-17,94 FDA2 25,68 21,40-31,95 25,26 21,10-34,80 21,10-34,80 FDN3 38,75 34,90-46,20 38,97 34,75-43,13 32,92-45,83 Humedad 4,86 2,32-7,59 4,88 2,91-7,26 2,25-7,49 Proteína 26,26 21,45-28,82 26,12 21,76-28,24 24,54-30,83 1 MON-15985-7 es MON-00531-6 retransformado para que exprese una proteína Cry adicional. Estos datos se pueden considerar información adicional sobre el evento de Cry1Ac MON-00531-6. Todos los valores se expresan en % de peso seco excepto la humedad, que se expresa en % de peso fresco. No se han informado diferencias estadísticamente significativas (P ≤0,05). 2 FDA = Fibra detergente ácido. 3 FDN = Fibra detergente neutro. Tabla III.6. Análisis proximal de la semilla de Gossypium hirsutum evento DAS-21023-5 (USDA APHIS 2003). Proximal (%) Ceniza Semilla Cry1Ac Semilla de control 4,0 3,9 Tabla III.7. Análisis de la fibra nutricional de la semilla de Gossypium hirsutum evento 31807/31808 (USDA APHIS 1997c).1 Valores de bibliografía 3,76-4,851 Grasa total 23,3 21,9 15,4-23,81 Humedad 2,8 3,2 3,97-8,471 Proteína 27,3 26,7 21,8-28,21 Carbohidratos 42,8 44,3 45,6-53,61 Calorías (Kcal/100 g) 490 481 ND Fibra cruda 15,7 17,0 15,4-20,91 Fibra detergente ácido 22,6 24,4 262, 37,51 Fibra detergente neutro 34,1 34,7 372, 52,61 Evento Fibra cruda FDA2 FDN3 31707 30,2 39,7 49,4 31803 31,8 36,8 45,7 31807 32,1 42,1 48,8 31808 31,4 40,4 47,5 42317 31,4 41,8 48,5 Coker 130 31,8 38,1 46,3 Stv. 474 30,4 40,4 47,6 St. LA887 32,5 42,4 49,0 DPL 51 33,8 41,4 48,6 1 Expresado como porcentaje de semilla con pelusa por peso. 2 Fibra detergente ácido. 3 Fibra detergente neutro. 1 Informado a partir del documento borrador de consenso de la OCDE, 2002. 2 Informado de la NCPA, Guía de productos de pienso de la semilla de algodón. Tabla III.8. Análisis proximal de la harina de semilla de algodón de Gossypium hirsutum evento 31807/31808 (USDA APHIS 1997c).1 Evento Humedad Proteína Ceniza 2,98 Grasa/aceite crudo 2,53 31707 31803 49,52 6,82 2,00 2,42 49,41 6,36 31807 1,69 2,14 53,31 7,16 31808 2,38 2,27 51,02 6,55 42317 1,69 1,73 49,17 6,53 Coker 130 3,14 2,39 53,10 7,14 Stv. 474 2,34 2,13 44,92 6,18 St. LA887 3,05 2,45 46,12 6,44 DPL 51 2,74 3,01 45,54 6,92 1 Los valores son en porcentaje de peso. 18