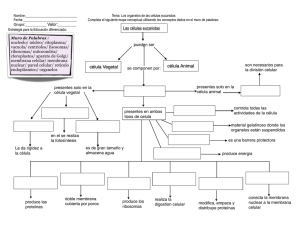
APUNTES DOCENTES
Anuncio

UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES ASIGNATURA: BIOLOGÍA PROFESOR: JUAN ANTONIO MANJARRÉS DUICA UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES TABLA DE CONTENIDO INTRODUCCIÓN ............................................................................................................................... 4 1. EL ORIGEN DE LA VIDA ............................................................................................................. 5 1.1. Teorías del Origen de la Vida ............................................................................................... 5 1.1.1. Generación espontánea ................................................................................................ 5 1.1.2. Pasteur ............................................................................................................................. 6 1.1.3. Teoría de Darwin ............................................................................................................. 6 1.1.4. Teoría de Oparin - Haldane ........................................................................................... 6 1.1.5. Spallanzani Y Needhad.................................................................................................. 7 1.1.6. La Panspermia ................................................................................................................ 7 1.1.7. Creacionismo................................................................................................................... 7 1.1.8. Fijismo .............................................................................................................................. 7 2. ORIGEN DEL UNIVERSO ........................................................................................................... 8 2.1. BIG BANG ............................................................................................................................... 8 2.2. Inflacionaria ............................................................................................................................. 8 2.3. De Creación Continua o Estado Estacionario ................................................................... 8 2.4. Universo Pulsante .................................................................................................................. 8 2.5. Agujero Negro ......................................................................................................................... 9 2.6. Galaxias ................................................................................................................................... 9 2.7. Los Púlsares ........................................................................................................................... 9 2.8. Los Cuásares .......................................................................................................................... 9 2.9. Las Estrellas ........................................................................................................................... 9 2.10. Vía Láctea ............................................................................................................................. 9 3. ÁCIDOS NUCLEICOS ................................................................................................................ 10 3.1. Ácido Desoxirribonucleico (ADN) ...................................................................................... 11 3.2. Ácido Ribonucleico (ARN) .................................................................................................. 13 4. FOTOSÍNTESIS .......................................................................................................................... 15 4.1. Etapas de la Fotosíntesis.................................................................................................... 16 4.1.1. Fase luminosa ............................................................................................................... 16 4.1.2. Fase oscura ................................................................................................................... 16 4.2. Estructura del Cloroplasto y de las Membranas Fotosintéticas ................................... 17 4.3. Fotosistemas......................................................................................................................... 17 4.4. Fotofosfoliración ................................................................................................................... 19 4.5. Fotorespiración ..................................................................................................................... 20 5. LA CÉLULA EN FUNCIONAMIENTO ...................................................................................... 22 5.1. Las Células Procarióticas ................................................................................................... 22 5.2. Las células eucarióticas ...................................................................................................... 22 5.3. Los Organelos del Sistema Endomembranoso............................................................... 25 5.3.1. El núcleo......................................................................................................................... 25 5.3.2. La envoltura nuclear ..................................................................................................... 25 5.3.3. El retículo endoplasmático rugoso ............................................................................. 26 5.3.4. El retículo endoplasmático liso ................................................................................... 28 5.3.5. El aparato de Golgi ....................................................................................................... 28 5.3.6. Los lisosomas ................................................................................................................ 29 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES 5.3.7. Las vacuolas .................................................................................................................. 30 5.3.8. Los cloroplastos ............................................................................................................ 31 5.3.9. Las mitocondrias ........................................................................................................... 32 6. MECANISMOS DE TRANSPORTE A TRAVÉS DE LA MEMBRANA CELULAR ............. 38 6.1. Los Fosfolípidos ................................................................................................................... 38 6.2. Transporte Pasivo ................................................................................................................ 40 6.3. Transporte Activo ................................................................................................................. 44 7. DIVISIÓN CELULAR................................................................................................................... 48 7.1. Mitosis .................................................................................................................................... 48 7.1.1. Fases de la Mitosis ....................................................................................................... 48 7.2. Meiosis ................................................................................................................................... 51 7.2.1. Meiosis I ......................................................................................................................... 51 7.2.2. Meiosis II ........................................................................................................................ 54 BIBLIOGRAFÍA ................................................................................................................................ 57 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES INTRODUCCIÓN La Biología es probablemente la más diversa de todas las ciencias, es la ciencia de la vida, rastrea las interacciones complejas dentro de los ecosistemas, relaciona los principales elementos constitutivos de los seres vivos, incluyendo al hombre y su relación con el entorno para contribuir a un ambiente sostenible en el tiempo. Esta relación le permite al hombre conocer su realidad ambiental, analizar los factores biológicos, físicos y químicos que rigen los procesos, la estructura y el comportamiento de los sistemas vivientes y elegir dar buen uso al planeta tierra, disposición adecuada de las basuras, el tamaño de la familia y muchos otros aspectos que afectan el medio que nos rodea. El presente material corresponde a un resumen de conceptos claves que pretende ser una herramienta accequible, amigable, fresca, esencial en el acompañamiento del trabajo independiente de los estudiantes de primer semestre de Tecnología en Recursos Ambientales de las Unidades Tecnológicas de Santander en su proceso de aprendizaje concebido éste como un acto cognitivo, voluntario, individual y actitudinal del estudiante que ocurre en un contexto colectivo y así tener un buen desempeño académico en la asignatura de Biología y adquirir las competencias básicas para su formación integral a través de los conceptos abordados en cada una de las unidades temáticas 4 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES 1. EL ORIGEN DE LA VIDA Desde que el hombre tuvo la capacidad de pensar y de razonar, se empezó a preguntar como surgió la vida, surgiendo así uno de los problemas más complejos y difíciles que se ha planteado el ser humano, en su afán de encontrar una respuesta, se intento solucionarlo mediante explicaciones religiosas, mitológicas y científicas, a partir de estas ultimas han surgido varias teorías y otras han sido descartadas. La cuestión del origen de la vida en la Tierra ha generado en las ciencias de la naturaleza un campo de estudio especializado cuyo objetivo es dilucidar cómo y cuándo surgió. La opinión más extendida en el ámbito científico establece la teoría de que la vida comenzó su existencia a partir de la materia inerte en algún momento entre hace 4.400 millones de años, cuando se dieron las condiciones para que el vapor de agua pudiera condensarse primera vez y 2.700 millones, cuando aparecen los primeros indicios de vida, que inducen a pensar en un origen biogénico de los minerales y sedimentos que se produjeron en esa época, así como los biomarcadores- moleculares que indican ya existía la fotosíntesis. Los progresos en esta área son generalmente lentos y esporádicos, aunque aún atraen la atención de muchos dada la importancia de la cuestión que se investiga. 1.1. Teorías del Origen de la Vida 1.1.1. Generación espontánea Sugiere que la vida se origina de la materia inerte según sus creencias y por observación suponían que del lodo se forman las lombrices, de la carne en descomposición, las moscas, de la ropa sucia y basura las ratas. La tesis de la generación espontánea fue defendida por Aristóteles, quien afirmaba, por ejemplo que era una verdad patente que los pulgones surgían del rocío que cae de las plantas, las pulgas de la materia en putrefacción, los ratones del heno sucio, los cocodrilos de los troncos en descomposición en el fondo de las masas acuáticas, y así sucesivamente. 5 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES 1.1.2. Pasteur En 1862, Louis Pasteur, medico francés, realizo una serie de experimentos encaminados a resolver el problema de la generación espontánea. Él pensaba que los causantes de la putrefacción de la materia orgánica eran los microorganismos que se encontraban en el aire. Para demostrar su hipótesis, diseño unos matraces cuello de cisne, en los cuales coloco líquidos nutritivos que después hirvió hasta esterilizarlos. Posteriormente, observo que en el cuello de los matraces quedaban detenidos los microorganismos del aire y aunque este entraba en contacto con la sustancia nutritiva, no había putrefacción de la misma. Para verificar sus observaciones, rompió el cuello de cisne de un matraz, y al entrar en contacto él liquido con el aire y los microorganismos que contenía él ultimo, se producía una descomposición de la sustancia nutritiva De esta manera quedo comprobada por él celebre científico la falsedad de la teoría de la generación espontánea. 1.1.3. Teoría de Darwin Charles Robert Darwin fue un naturalista inglés que postuló que todas las especies de seres vivos han evolucionado con el tiempo a partir de un antepasado común mediante un proceso denominado selección natural Las formas de vida no son estáticas sino que evolucionan; las especies cambian continuamente, unas se originan y otros se extinguen. El proceso de la evolución es gradual, lento y continuo, sin saltos discontinuos o cambios súbitos. Los organismos parecidos se hallan emparentados y descienden de un antepasado común. Todos los organismos vivientes pueden remontarse a un origen único de la vida. La selección natural es la llave, en dos fases, que explica todo el sistema. La primera fase es la producción de variabilidad: la generación de modificaciones espontáneas en los individuos. La segunda, la selección a través de la supervivencia en la lucha por la vida: los individuos mejor dotados, los que han nacido con modificaciones espontáneas favorables para hacer frente al medio ambiente van a tener más posibilidades de sobrevivir, de reproducirse y de dejar descendencia con estas ventajas. 1.1.4. Teoría de Oparin - Haldane Admiten que la atmósfera primitiva de la Tierra estaba constituida por una mezcla de gases muy rica en HIDRÓGENO y pobre en OXÍGENO. Los elementos biogenéticos, Carbono, Hidrógeno y Oxígeno, estaban combinados con el HIDRÓGENO formando METANO (CH4), AMONÍACO (NH3) y AGUA (H2O). Los compuestos mencionados, presentes en mezclas sometidas a la acción de las radiaciones ultravioletas, dieron origen a la formación de moléculas orgánicas tales como AZÚCARES y AMINOÁCIDOS. Características de la tierra: altas temperaturas (volcanes) producción constante de lluvias constantes relámpagos En esta atmósfera ocurrían reacciones químicas debido a la energía eléctrica de los rayos y a la energía térmica. Estas reacciones químicas formaron los primeros compuestos orgánicos. Estos compuestos orgánicos se mezclaron para formar monómeros (compuestos orgánicos + compuestos orgánicos). 6 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES Éstas se combinarían de una forma cada vez más compleja hasta quedar disueltas en una gotita de coacervado. Estas gotitas crecerían por fusión con otras y se reproducirían mediante fisión en gotitas hijas, y de ese modo podrían haber obtenido un metabolismo primitivo en el que estos factores asegurarían la supervivencia de la "integridad celular" de aquellas que no acabaran extinguiéndose. 1.1.5. Spallanzani Y Needhad En esos mismos tiempos, otro científico llamado Needhad, sostenía que había una fuerza vital que originaba la vida. Sus suposiciones se basan en sus experimentos: hervía caldo de res en una botella, misma que tapaba con un corcho, la dejaba reposar varios días y al observar al microscopio muestra de la sustancia, encontraba organismos vivos. Él afirmaba que el calor por el que había hecho pasar el caldo era suficiente para matar a cualquier organismo y que, entonces, la presencia de seres vivos era originada por la fuerza vital. Sin embargo Spallanzani no se dejó convencer como muchos científicos de su época, realizando los mismos experimentos de Needhad, pero sellada totalmente las botellas, las ponía a hervir, la dejaba reposar varios días y cuando hacia observaciones no encontraba organismos vivos. Esto lo llevo a concluir que los organismos encontrados por Needhad procedían del aire que penetraba a través del corcho. 1.1.6. La Panspermia Una propuesta más para resolver el problema del origen de la vida la presento Svante Arrhenius, en 1908 su teoría se conoce con el nombre de panspermia. Según ésta, la vida llego a la Tierra en forma de esporas y bacterias provenientes del espacio exterior que, a su vez, se desprendieron de un planeta en la que existían. A esta teoría se le puede oponer el siguiente argumento: Se tiene conocimiento de que las condiciones del medio interestelar son poco favorables para la supervivencia de cualquier forma de vida. Además, se sabe que cuando un meteorito entra en la atmósfera, se produce una fricción que causa calor y combustión destruyendo cualquier espora o bacteria que viaje en ellos. Este argumento tampoco soluciona el problema del origen de la vida, pues no explica cómo se formó ésta en el planeta hipotético del cual se habría desprendido la espora o bacteria. 1.1.7. Creacionismo Los creacionistas clásicos niegan la teoría de la evolución biológica y, especialmente, lo referido a la evolución humana, además de las explicaciones científicas sobre el origen de la vida. Por esto rechazan todas las pruebas científicas (fósiles, geológicas, genéticas, etc.). En el creacionismo clásico de origen cristiano se hace una interpretación literal de la Biblia y se sostiene la creación del mundo, los seres vivos y el cataclismo del Diluvio Universal tal como está descrito en el Génesis, sin pretender concretar de manera científica el origen de las especies. 1.1.8. Fijismo Es una creencia que sostiene que las especies actualmente existentes han permanecido básicamente invariables desde la Creación. Las especies serían, por tanto, inmutables, tal y como fueron creadas. 7 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES 2. ORIGEN DEL UNIVERSO 2.1. BIG BANG Su rasgo principal es que la expansión lleva asociada un enfriamiento y una transformación de la materia. Las etapas del origen del Universo según el modelo del Big – Bang se distinguen cuatro etapas iniciales y sucesivas: 1. 2. 3. 4. inflación confinamiento de quarks nucleosíntesis primordial recombinación 2.2. Inflacionaria Muestra los estudios de Alan Guth. Se basa en estudios sobre campos gravitatorios fortísimos, como los que hay cerca de un agujero negro. Supone que una fuerza única se dividió en las cuatro que ahora conocemos, produciendo el origen al Universo. 2.3. De Creación Continua o Estado Estacionario Muchos consideran que el universo es una entidad que no tiene principio ni fin. No tiene principio no comenzó con una gran explosión ni se colapsará, en un futuro, para volver a nacer. La teoría que se opone a la tesis de un universo evolucionario es conocida como "teoría del estado estacionario y nace en el siglo XX. 2.4. Universo Pulsante El momento en que el universo se desploma sobre sí mismo atraído por su propia gravedad es conocido 8 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES como Big Crunch" en el ambiente científico. El Big Crunch marcaría el fin de nuestro universo y el nacimiento de otro nuevo, tras el subsiguiente Big Bang que lo forme. 2.5. Agujero Negro Son cuerpos con un campo gravitatorio extraordinariamente grande, no puede escapar ninguna radiación electromagnética ni luminosa, por eso son negros. 2.6. Galaxias Son acumulaciones enormes de estrellas, gases y polvo. En el Universo hay centenares de miles de millones. Cada galaxia puede estar formada por centenares de miles de millones de estrellas y otros astros. 2.7. Los Púlsares Los estudios indican que un púlsar es una estrella de neutrones pequeña que gira a gran velocidad. El más conocido está en la nebulosa de Cangrejo. 2.8. Los Cuásares Se identificaron en la década de 1950. Más tarde se vio que mostraban un desplazamiento al rojo más grande que cualquier otro objeto conocido. La causa era el efecto Dopler, que mueve el espectro hacia el rojo cuando los objetos se alejan. 2.9. Las Estrellas Son masas de gases, principalmente hidrógeno y helio, que emiten luz. Vemos las estrellas, excepto el Sol, como puntos luminosos muy pequeños, y sólo de noche, porque están a enormes distancias de nosotros. 2.10. Vía Láctea La Vía Láctea es nuestra galaxia. Los romanos la llamaron "Camino de Leche". Es grande, espiral y puede tener unos 100.000 millones de estrellas, entre ellas, el Sol. 9 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES 3. ÁCIDOS NUCLEICOS Los ácidos nucleicos fueron descubiertos por Freidrich Miescher en 1869. En la naturaleza existen solo dos tipos de ácidos nucleicos: El ADN (ácido desoxirribonucleico) y el ARN (ácido ribonucleico) y están presentes en todas las células. Su función biológica no quedó plenamente confirmada hasta que Avery y sus colaboradores demostraron en 1944 que el ADN era la molécula portadora de la información genética. Los ácidos nucleicos tienen al menos dos funciones: trasmitir las características hereditarias de una generación a la siguiente y dirigir la síntesis de proteínas específicas. Tanto la molécula de ARN como la molécula de ADN tienen una estructura de forma helicoidal. Químicamente, estos ácidos están formados, como dijimos, por unidades llamadas nucleótidos: cada nucleótido a su vez, está formado por tres tipos de compuestos: 1. Una pentosa o azúcar de cinco carbonos: se conocen dos tipos de pentosas que forman parte de los nucleótidos, la ribosa y la desoxirribosa, esta última se diferencia de la primera porque le falta un oxígeno y de allí su nombre. El ADN sólo tiene desoxirribosa y el ARN tiene sólo ribosa, y de la pentosa que llevan se ha derivado su nombre, ácido desoxirribonucleico y ácido ribonucleico, respectivamente. Las dos pentosas 2. Una base nitrogenada: que son compuestos anillados que contienen nitrógeno. Se pueden identificar cinco de ellas: adenina, guanina, citosina, uracilo y timina. Las cinco bases nitrogenadas 10 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES 3. Un radical fosfato: es derivado del ácido fosfórico (H3PO4) Los ácidos nucleicos son polímeros lineales en los que la unidad repetitiva, llamada nucleótido (figura de la izquierda), está constituida por: (1) una pentosa (la ribosa o la desoxirribosa), (2) ácido fosfórico y (3) una base nitrogenada (purina o pirimidina). La unión de la pentosa con una base constituye un nucleósido (zona coloreada de la figura). La unión mediante un enlace éster entre el nucleósido y el ácido fosfórico da lugar al nucleótido. La secuencia de los nucleótidos determina el código de cada ácido nucleico particular. A su vez, este código indica a la célula cómo reproducir un duplicado de sí misma o las proteínas que necesita para su supervivencia. El ADN y el ARN se diferencian porque: - el peso molecular del ADN es generalmente mayor que el del ARN - el azúcar del ARN es ribosa, y el del ADN es desoxirribosa - el ARN contiene la base nitrogenada uracilo, mientras que el ADN presenta timina La configuración espacial del ADN es la de un doble helicoide, mientras que el ARN es un polinucleótido lineal, que ocasionalmente puede presentar apareamientos intracatenarios 3.1. Ácido Desoxirribonucleico (ADN) El Ácido Desoxirribonucleico o ADN (en inglés DNA) contiene la información genética de todos los seres vivos. 11 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES Cada especie viviente tiene su propio ADN y en los humanos es esta cadena la que determina las características individuales, desde el color de los ojos y el talento musical hasta la propensión a determinadas enfermedades. Es como el código de barra de todos los organismos vivos que existen en la tierra, que está formado por segmentos llamados genes. La combinación de genes es específica para cada organismo y permite individualizarnos. Estos genes provienen de la herencia de nuestros padres y por ello se utiliza los tests de ADN para determinar el parentesco de alguna persona. Además, se utiliza el ADN para identificar a sospechosos en crímenes (siempre y cuando se cuente con una muestra que los relacione). Actualmente se ha determinado la composición del genoma humano que permite identificar y hacer terapias para las enfermedades que se trasmiten genéticamente como: enanismo, albinismo, hemofilia, daltonismo, sordera, fibrosis quística, etc. Molécula de ADN con sus estructura helicoidal 12 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES Agentes mutagénicos y las diferentes alteraciones que pueden producir en el ADN Las mutaciones pueden surgir de forma espontánea (mutaciones naturales) o ser inducidas de manera artificial (mutaciones inducidas) mediante radiaciones y determinadas sustancias químicas a las que llamamos agentes mutágenos. Estos agentes aumentan significativamente la frecuencia normal de mutación. Así pues, distinguimos: 1) Radiaciones, que, según sus efectos, pueden ser: a) No ionizantes, como los rayos ultravioleta (UV) que son muy absorbidas por el ADN y favorecen la formación de enlaces covalentes entre pirimidinas contiguas (dímeros de timina, por ejemplo) y la aparición de formas tautómeras que originan mutaciones génicas. b) Ionizantes, como los rayos X y los rayos gamma, que son mucho más energéticos que los UV; pueden originar formas tautoméricas, romper los anillos de las bases nitrogenadas o los enlaces fosfodiéster con la correspondiente rotura del ADN y, por consiguiente, de los cromosomas. 2) Sustancias químicas que reaccionan con el ADN y que pueden Otra imagen para la molécula provocar las alteraciones siguientes: de ADN a) Modificación de bases nitrogenadas. Así, el HNO 2 las desamina, la hidroxilamina les adiciona grupos hidroxilo, el gas mostaza añade grupos metilo, etilo,... b) Sustitución de una base por otra análoga. Esto provoca emparejamientos entre bases distintas de las complementarias. c) Intercalación de moléculas. Se trata de moléculas parecidas a un par de bases enlazadas, capaces de alojarse entre los pares de bases del ADN. Cuando se produce la duplicación pueden surgir inserciones o deleciones de un par de bases con el correspondiente desplazamiento en la pauta de lectura. El “ayudante” del ADN La información genética está, de alguna manera, escrita en la molécula del ADN, por ello se le conoce como ―material genético‖. Por esto, junto con el ARN son indispensables para los seres vivos. El ARN hace de ayudante del ADN en la utilización de esta información. Por eso en una célula eucariótica (que contiene membrana nuclear) al ADN se lo encuentra sólo en el núcleo, ya sea formando a los genes, en cambio, al ARN se lo puede encontrar tanto en el núcleo como en el citoplasma. 3.2. Ácido Ribonucleico (ARN) Ácido nucleico formado por nucleótidos en los que el azúcar es ribosa, y las bases nitrogenadas son adenina, uracilo, citosina y guanina. Actúa como intermediario y complemento de las instrucciones genéticas codificadas en el ADN. 13 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES Tipos de ARN Existen varios tipos diferentes de ARN, relacionados con la síntesis de proteínas. Así, existe ARN mensajero (ARNm), ARN ribosómico (ARNr), ARN de transferencia (ARNt) y un ARN heterogéneo nuclear (ARN Hn). El ARN es normalmente el producto de la transcripción (reacción anabólica) de un molde de ADN, aunque en los retrovirus el ARN actúa de plantilla y el ADN de copia. ARNHn ARN heterogéneo nuclear = ARNm primario: localizado en el núcleo y de tamaño variable. Precursor del ARN mensajero, se transforma en él tras la eliminación de los intrones, las secuencias que no codifican genes. ARNm ARN mensajero: molécula de ARN que representa una copia en negativo de las secuencias de aminoácidos de un gen. Las secuencias no codificantes (intrones) han sido ya extraídas. Con pocas excepciones el ARNm posee una secuencia de cerca de 200 adeninas (cola de poli A), unida a su extremo 3' que no es codificada por el ADN. El ARN mensajero, es una cadena simple, muy similar a la del ADN, pero difiere en que el azúcar que la constituye es ligeramente diferente (se llama Ribosa, mientras que la que integra el ADN es Desoxi Ribosa). Una de las bases nitrogenadas difiere en el ARN y se llama Uracilo, sustituyendo a la Timina. La información para la síntesis de aminoácidos está codificada en forma de tripletes, cada tres bases constituyen un codón que determina un aminoácido. Las reglas de correspondencia entre codones y aminoácidos constituyen el código genético. La síntesis de proteínas o traducción tiene lugar en los ribosomas del citoplasma. Los aminoácidos son transportados por el ARN de transferencia, específico para cada uno de ellos, y son llevados hasta el ARN mensajero, dónde se aparean el codón de éste y el anticodón del ARN de transferencia, por complementariedad de bases, y de esta forma se sitúan en la posición que les corresponde. Una vez finalizada la síntesis de una proteína, el ARN mensajero queda libre y puede ser leído de nuevo. De hecho, es muy frecuente que antes de que finalice una proteína ya está comenzando otra, con lo cual, una misma molécula de ARN mensajero, está siendo utilizada por varios ribosomas simultáneamente. 14 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES 4. FOTOSÍNTESIS La vida en la tierra depende fundamentalmente de la energía solar, la cual es atrapada mediante el proceso fotosintético, que es responsable de la producción de toda la materia orgánica que conocemos. La materia orgánica comprende los alimentos que consumimos diariamente tanto nosotros como los animales, los combustibles fósiles (petróleo, gas, gasolina, carbón); así como la leña, madera, pulpa para papel, inclusive la materia prima para la fabricación de fibras sintéticas, plásticos, poliéster, etc. La cantidad de carbono fijado por la fotosíntesis es espectacular, como lo demuestran las cifras de la 11 producción anual de materia orgánica seca, estimada en 1,55 x 10 toneladas, con aproximadamente 60% formada en la tierra, el resto en océanos y aguas continentales. Los organismos que en el curso de la evolución aprendieron a usar la energía solar y a transformarla en energía química son los llamados autótrofos, que están representados por bacterias y organismos del Reino Vegetal. La fotosíntesis, del griego antiguo υοτο (foto) "luz" y σύνθεσις (síntesis) "unión", es la base de la vida actual en la Tierra. Proceso mediante el cual las plantas, algas y algunas bacterias captan y utilizan la energía de la luz para transformar la materia inorgánica de su medio externo en materia orgánica que utilizarán para su crecimiento y desarrollo. Los organismos capaces de llevar a cabo este proceso se denominan foto autótrofos y además son capaces de fijar el CO2 atmosférico (lo que ocurre casi siempre) o simplemente autótrofos. Salvo en algunas bacterias, en el proceso de fotosíntesis se producen liberación de oxígeno molecular (proveniente de moléculas de H2O) hacia la atmósfera (fotosíntesis oxigénica). Es ampliamente admitido que el contenido actual de oxígeno en la atmósfera se ha generado a partir de la aparición y actividad de dichos organismos fotosintéticos. Esto ha permitido la aparición evolutiva y el desarrollo de organismos aerobios capaces de mantener una alta tasa metabólica (el metabolismo aerobio es muy eficaz desde el punto de vista energético). La otra modalidad de fotosíntesis, la fotosíntesis anoxigénica, en la cual no se libera oxígeno, es llevada a cabo por un número reducido de bacterias, como las bacterias púrpuras del azufre y las bacterias verdes del azufre; estas bacterias usan como donador de hidrógenos el H2S, con lo que liberan azufre. 15 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES 4.1. Etapas de la Fotosíntesis EL Objetivo de la fotosíntesis: es producir glucosa Para alimentarse. Se lleva a cabo en los cloroplastos, consiste en una serie de reacciones que requieren energía en forma de Luz. La glucosa está formada por carbono, hidrogeno, oxigeno. Este proceso ocurre en dos etapas diferentes: 4.1.1. Fase luminosa La fotosíntesis consta de dos fases, la fase luminosa necesita de la luz para llevarse a cabo, por lo tanto sólo se lleva a cabo durante el día. Primero, la clorofila de las plantas y de las algas captura la energía luminosa. Esta energía queda atrapada entre los enlaces de las moléculas de clorofila excitándola. Con esa energía, las células fragmentan las moléculas de agua que hay en su interior en sus dos componentes: hidrógeno (H), y oxígeno. Las moléculas de oxígeno se unen en pares, para formar el oxígeno que es liberado hacia la atmósfera (O2) y las de hidrógeno (H2) forman un gradiente el cual es aprovechado para formar energía química (ATP). Si te cuesta trabajo imaginar esto piensa en un represa, en ella formas un lago artificial y el agua al querer seguir su cause natural sale con bastante fuerza, nosotros los seres humanos usamos bobinas o ?molinos? que nos sirven para aprovechar esa fuerza y realizar un trabajo como el producir luz eléctrica, igual la *célula* al recibir energía luminosa y romper el agua (H2O) en hidrógenos y oxígenos desechando el oxígeno y guardando dentro del *cloroplasto* muchos hidrógenos, crea una especie de represa de hidrógenos y este al salir del interior del cloroplasto con una cierta energía esta se utiliza para formar energía química en forma de *(ATP)* la cual es utilizada por la planta y por todos los seres vivos en miles y millones de distintas funciones. 4.1.2. Fase oscura Esta fase es así llamada por no necesitar de la luz para efectuarse. Se lleva a cabo dentro de los *cloroplastos* tanto en el día como en la noche. En esta fase se utilizan los hidrógenos liberados y la energía química formada en la *fase luminosa* junto con el dióxido de carbono absorbido del medio ambiente para formar moléculas grandes de azúcar como la glucosa y el almidón. Esta fase es de construcción, en la que gracias a la energía obtenida piezas pequeñas como el carbono obtenido del dióxido de carbono y el hidrógeno se forman grandes moléculas. 16 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES 4.2. Estructura del Cloroplasto y de las Membranas Fotosintéticas La unidad estructural de la fotosíntesis es el cloroplasto. Los organismos fotosintéticos procariotas y eucariotas poseen sacos aplanados o vesículas llamadas tilacoides, que contienen los pigmentos fotosintéticos; pero solamente los cloroplastos de los eucariotas están rodeados por una doble membrana. Los tilacoides se disponen como una pila de panqueas, que recibe el nombre de grana. El interior del cloroplasto entre las granas es el estroma proteico, donde se encuentran las enzimas que catalizan la fijación del CO2. Las mitocondrias constituyen un sistema con dos membranas como los cloroplastos, pero los cloroplastos tienen tres compartimentos: el estroma, el espacio tilacoidal y el espacio entre las membranas. El cloroplasto en su interior tiene un ADN circular y ribosomas. 4.3. Fotosistemas En la fotosíntesis cooperan dos grupos separados de pigmentos o fotosistemas, que se encuentran localizados en los tilacoides. Muchos organismos procariotas solamente tienen el fotosistema I (es el más primitivo desde el punto de vista evolutivo). 17 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES Los organismos eucariotas poseen los fotosistemas I y II. El fotosistema I está asociado a las formas de clorofila a, que absorbe a longitudes de onda de 700 nm ( P 700 ), mientras que el foto sistema II tiene un centro de reacción que absorbe a una longitud de onda de 680 nm ( P 680 ). Cada uno de estos fotosistemas se encuentra asociado a polipéptidos en la membrana tilacoidal y absorben energía luminosa independientemente. En el fotosistema II, se produce la fotólisis del agua y la liberación de oxígeno; sin embargo ambos fotosistemas operan en serie, transportando electrones, a través de una cadena + transportadora de electrones. En el fotosistema I se transfieren dos electrones a la molécula de NADP y se forma NADPH, en el lado de la membrana tilacoidal que mira hacia el estroma. 18 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES 4.4. Fotofosfoliración Es la síntesis de ATP que se produce cuando se exponen cloroplastos aislados a la acción de la luz, en presencia de ADP y fosfato. La formación de ATP a partir de la reacción de ADP y fosfato, es el resultado del acoplamiento energético de la fosforilación al proceso de transporte de electrones inducido por la luz, de la misma forma que la fosforilación oxidativa está acoplada al transporte de electrones y al consumo de oxígeno en las mitocondrias. En el fotosistema I se realiza la síntesis cíclica de ATP, que es independiente de la fotólisis del agua y de la formación de NADPH; mientras que la fotofosforilación no cíclica, está acoplada al transporte de electrones desde el agua, en el foto sistema II a través de una cadena transportadora de electrones hacia el foto + sistema I, donde la ferrodoxina cede dos electrones al NADP para que se reduzca a NADPH. + H2O + NADP + Pi + ADP+ cloroplastos + luz + ½ O2 + NADPH + H + ATP + H2 O La molécula de H2 O del lado izquierdo de la ecuación, cede los dos electrones necesarios para la reducción + del NADP y el átomo de oxígeno que se libera en forma de ½ O 2. La molécula de H2O del lado derecho de la ecuación procede de la formación de ATP a partir de la reacción de ADP + Pi. 19 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES En la membrana tilacoidal como resultado de la fotólisis del agua y de la oxidación de la plastoquinona +) +) (PQH2) se generan protones (H ; que originan un fuerte gradiente de concentración de protones (H al ser transportados del lumen tilacoidal hacia el estroma. Este gradiente de pH a través de la membrana es responsable de la síntesis de ATP, catalizada por la ATP sintetasa o conocida también como factor de acoplamiento; ya que acopla la síntesis de ATP al transporte de electrones y protones a través de la membrana tilacoidal. La ATP sintetasa existe en los tilacoides del estroma y consta de dos partes principales: un tallo denominado CFo, que se extiende desde el lumen de la membrana tilacoidal hasta el estroma y una porción esférica (cabeza) que se conoce como CF1 y que descansa en el estroma. Esta ATPasa es similar a la de las mitocondrias donde sintetiza ATP. El flujo cíclico de electrones tiene lugar en algunos eucariotas y bacterias fotosintéticas primitivas. No se produce NADPH, sino ATP solamente. Esto puede ocurrir cuando las células pueden requerir un suministro + de ATP adicional, o cuando no se encuentre presente NADP para ser reducido a NADPH. En el fotosistema + II, el bombeo de iones H dentro del tilacoide crea un gradiente electroquímico que culmina con la síntesis de ATP a partir de ADP +Pi. 4.5. Fotorespiración Una de las propiedades más interesantes de la rubisco es que además de catalizar la carboxilación de la ribulosa 1,5 bifosfato, también produce su oxigenación; proceso conocido como foto respiración. La foto respiración da como resultado la liberación de CO 2, después de una serie de reacciones enzimáticas. ¡Es admirable que la rubisco de bacterias anaeróbicas autótrofas, cataliza la reacción de la oxigenada! La reacción de la carboxilación es favorecida a la oxigenación en una proporción de 3:1; lo que indica un 33% de ineficiencia en la carboxilación. El metabolismo del glicolato requiere la participación de las mitocondrias y de los peroxisomas. Sin embargo, es en las mitocondrias donde el aminoácido glicina, producido en los peroxisomas es descarboxilado liberando CO2. 20 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES El ritmo de la fotorrespiración de las plantas C-3 es bastante elevado, siendo 5 veces superior al de la respiración en la oscuridad; lo cual es perjudicial para estas plantas. Las plantas C-4, que muestran muy poca o ninguna foto respiración, son considerablemente más eficientes; ya que realizan la fotosíntesis a concentraciones más bajas de CO2 y a más elevadas tensiones de oxígeno. Las plantas C-4 son de origen principalmente tropical, habitan en condiciones de alta luminosidad y altas temperaturas. Esto les permite competir más eficientemente con las plantas C-3, al tener que cerrar los estomas para economizar agua y evitar la desecación; sin embargo pueden realizar la fotosíntesis a bajas tensiones de CO2, debido a que la enzima PEP-carboxilasa muestra una mayor afinidad por el CO 2 que la rubisco. La fotosíntesis aunque es un proceso bioquímico a nivel celular lo que podría resultar insignificante, tiene más repercusiones en nuestra vida de lo que podríamos imaginar. Tan solo de ella depende la calidad del aire que respiramos y gracias a ella la atmósfera primitiva se modificó permitiendo así la vida de animales, plantas y el hombre. La energía solar capturada por el proceso de la fotosíntesis es la fuente de cerca del 90% de toda la energía empleada por el hombre para satisfacer las demandas de calor, de luz y de potencia. En ella el impacto de los fotones de luz sobre la clorofila y la fotolisis del agua son el origen de un estado de desequilibrio molecular, que se reequilibra constantemente gracias al flujo de protones a través de la membrana de los tilacoides; esto se conoce como Fase Luminosa. La Fase Oscura consiste en la transformación de dióxido de carbono en glucosa y otros carbohidratos, utilizando para ello la energía química de los productos de la fosforilación. Esta energía almacenada en forma de ATP y NADPH2 se usa para reducir el dióxido de carbono a carbono orgánico. Esta función se lleva a cabo mediante una serie de reacciones llamada ciclo de Calvin, activadas por la energía de ATP y NADPH2. Dando como resultado el 02 liberado a la atmósfera y glucosa que sirve de alimento para la planta. Cada año, las plantas de nuestro planeta utilizan alrededor de 310,000 millones de toneladas de agua y 750,000 millones de toneladas de dióxido de carbono, para transformarlo en unos 510,000 millones de toneladas de materia y unos 550,000 millones de toneladas de oxígeno. Suena ilógico que a pesar de que el oxígeno de nuestra atmósfera no se acaba gracias al trabajo de todas las plantas, algas y bacterias, los humanos ―supuestamente seres racionales‖ seguimos talando árboles, contaminando el ambiente, con el smog de automóviles, basura en las calles, aerosoles, humo, etc. Y acabando con todo este oxígeno que es vital para nuestra vida. 21 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES 5. LA CÉLULA EN FUNCIONAMIENTO 5.1. Las Células Procarióticas Son pequeñas y estructuralmente simples. A lo largo del tiempo, han evolucionado dos clases de células estructuralmente diferentes. Las bacterias y las arqueobacterias consisten en células procarióticas, mientras que todas las demás formas de vida están compuestas por células eucarióticas. Se necesita un microscopio electrónico para ver claramente los detalles estructurales de cualquier célula, y esto es especialmente cierto para las células procarióticas debido a que son tan pequeñas. La mayoría de las células procarióticas miden entre 2 y 8 um de largo, lo cual en promedio es alrededor de un décimo del tamaño de una célula eucariótica típica. Una célula procariótica (del griego pro, antes, y karyon, núcleo) carece de núcleo; su ADN está enrollado en una región nucleoide (semejante a un núcleo), como se muestra en la figura 1, Debido a que ninguna membrana rodea a la región nucleoide, el ADN está en contacto directo con el contenido del resto de la célula. Nótense los ribosomas (puntos oscuros) en el interior de la célula. Por medio de instrucciones impartidas desde el ADN, los ribosomas unen aminoácidos y forman polipéptidos, los polímeros que constituyen las proteínas. El ADN ejerce control sobre las células por medio de los ARN mensajeros para especificar qué proteínas deben sintetizarse. La membrana plasmática rodea el citoplasma de la célula procariota. Alrededor de la membrana plasmática de la mayoría de las bacterias existe una pared celular bacteriana relativamente rígida y químicamente compleja. Esta pared protege a la célula y le ayuda a conservar una cierta forma. En algunos procariotas, existe una cubierta externa denominada cápsula, que rodea a la pared celular y protege aún más la superficie de la célula. Las cápsulas participan además en la adhesión de los procariotas a las superficies, por ejemplo a ramas y rocas en corrientes rápidas de agua, o a los tejidos en el cuerpo humano. Además de sus cubiertas externas, algunas bacterias poseen ciertas proyecciones en su superficie. Los pi11 (singular, pilus) son proyecciones cortas que también le ayudan a las bacterias a adherirse a las superficies. Los flagelos procarióticos son proyecciones largas que propulsan a la célula procariótica a través de su medio líquido. Figura 1. Una célula Procariota (35.000x) 5.2. Las células eucarióticas Poseen comportamientos funcionales Todas las células eucarióticas (del griego eu, verdadero, y karyon, núcleo), tanto animales, como vegetales, protistas y hongos, son fundamentalmente similares las unas a las otras y profundamente diferentes de las células procarióticas. Veamos una célula animal y una célula vegetal como representantes de las eucariotas. 22 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES La figura 2, ilustra una célula animal ideal, y muestra los detalles vistos con el microscopio electrónico de transmisión. Una mirada rápida a la figura confirma que las células eucarióticas son mucho más complejas que las células procarióticas. La diferencia más obvia es la variedad de estructuras en el citoplasma. Nótese que la mayoría de las estructuras están compuestas por membranas (en el dibujo se muestran como líneas blancas en la sección transversal). En las eucarióticas, las membranas dividen al citoplasma en compartimientos, que los biólogos denominan organelos membranosos (sus nombres aparecen subrayados en la figura, así como también el término membrana plasmática). Los organelos membranosos que se muestran en la figura 2 son el núcleo, retículo endoplasmático, aparato de Golgi, mitocondria, lisosomas y peroxisomas. Aunque la mayoría de los organelos no tienen color, les hemos asignado aquí y en muchos otros sitios en el libro, un código de color para su fácil identificación. Por ejemplo, utilizamos púrpura consistentemente para el núcleo, el centro de control de la célula eucariótíca, cada vez que se lo presenta en una figura. Muchas de las actividades químicas de las células, conocidas en conjunto como metabolismo celular, tienen lugar en los espacios llenos de fluido en el interior de los organelos membranosos. Estos espacios son importantes como sitios donde se mantienen condiciones químicas específicas, que incluso varían de organelo en organelo. Los procesos metabólicos que requieren condiciones diferentes, pueden tener lugar simultáneamente en una única célula porque se desarrollan en organelos separados. Por ejemplo, mientras el retículo endoplasmático está dedicado a la producción de una hormona esteroidea, los peroxisomas vecinos pueden estar fabricando peróxido de hidrógeno (H2O2) que es un producto secundario tóxico de ciertas reacciones metabólicas. Sólo gracias a que el H 2O2 se encuentra confinado en los peroxisomas, donde se le convierte rápidamente en H2O, es que los esteroides están protegidos para no ser destruidos. Otro beneficio de las membranas internas es que aumentan enormemente el área total de la membrana de una célula eucariótica. Una célula eucariótica típica, con un diámetro diez veces mayor que una célula procariótica típica, tiene un volumen cito- plasmático mil veces mayor, pero el área de la membrana plasmática es sólo cien veces mayor que la de la célula procariótica. En las células eucarióticas, las membranas internas (de los organelos) proveen la superficie donde tienen lugar muchos procesos metabólicos importantes; en efecto, muchas proteínas enzimáticas esenciales para los procesos metabólicos son partes componentes de las membranas de los organelos. Sin el sistema de membranas internas, las células eucarióticas probablemente no tendrían suficiente superficie de membrana para cubrir las necesidades metabólicas. Figura 2. Una célula Animal (8000x) 23 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES Casi todos los organelos mencionados hasta ahora están presentes tanto en las células de animales como de plantas. Como puede verse al comparar las figuras 2 y 3, la excepción es el lisosoma, que no está presente en las células vegetales. Pero hay además otras diferencias entre las células vegetales y animales. Una de ellas es el par de centríolos presentes en la célula animal, y ausentes en las células vegetales. Una célula animal puede además tener un flagelo, y algunas células animales pueden tener más de uno. Por el contrario, en las plantas sólo las células espermáticas de unas pocas especies llevan flagelos. (El flagelo eucariótico es diferente del flagelo procariótico tanto en estructura como en función.) Una célula vegetal posee ciertas estructuras que no posee la célula animal. Por ejemplo, la célula vegetal tiene una pared celular rígida, un tanto gruesa (como las células de los hongos y de muchos protistas). Las paredes celulares protegen a las células vegetales y les ayudan a conservar su forma. Las paredes celulares vegetales, que difieren químicamente de las paredes celulares procarióticas, contienen el polisacárido celulosa. A diferencia de las células animales típicas, muchas células vegetales maduras poseen una forma poligonal, tal como se muestra en la figura 3. Otro de los organelos presentes en las células vegetales pero no en las animales es el cloroplasto, donde se lleva a cabo la fotosíntesis. (Las membranas fotosintéticas también pueden encontrarse en algunos protistas.) La gran vacuola central es exclusiva de las células vegetales, y es un gran depósito de agua y de una importante variedad de otras sustancias químicas. En la mayoría de las células vegetales maduras, la vacuola central contiene enzimas que llevan a cabo la digestión celular, con lo que cumple la misma función que el lisosoma en la célula animal. Además, al incorporar agua adicional y expanderse, la vacuola central colabora con el alargamiento de la célula. Aunque hemos enfatizado en los organelos membranosos, las células eucarióticas contienen también otras estructuras no membranosas (aquellas cuyos nombres no están subrayados). Entre ellas se encuentran el centríolo, flagelo y citoesqueleto, todas las cuales contienen tubos de proteínas denominados microtúbulos. Además, puede verse que los ribosomas, que son los sitios de síntesis de las proteínas y están representados por numerosos puntos oscuros en ambas figuras, se distribuyen por todo el citoplasma, como también lo hacen en las células procarióticas. Además de tener ribosomas en el fluido citoplasmático, las células eucarióticas poseen muchos ribosomas adosados a ciertas partes del retículo endoplasmático (haciéndolo ―rugoso‖) y al exterior del núcleo. 24 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES Figura 3. Una célula Vegetal (8.000x) ¿Cuál de los siguientes organelos no pertenece a la lista: mitocondria, cloroplasto, ribosoma, lisosoma, peroxisoma? ¿Por qué? 5.3. Los Organelos del Sistema Endomembranoso 5.3.1. El núcleo Es el centro de control genético de la célula. El núcleo es el centro de control genético de una célula eucariótica. Su ADN, al igual que el ADN de un procariota, es el código genético de la célula. El ADN nuclear está adherido a proteínas, formando fibras muy largas denominadas cromatina (los hilos morados en la figura 4). Cada fibra constituye un cromosoma. Durante la reproducción celular, la cromatina se superenrolla, y forma estructuras mucho más gruesas que pueden ser observadas con un microscopio óptico como cromosomas individuales. 5.3.2. La envoltura nuclear Rodea al núcleo, y consiste en una doble membrana perforada con poros que controlan el flujo de materiales hacia adentro y hacia afuera del núcleo. Adjunto a la cromatina dentro del núcleo existe una masa de fibras y gránulos llamado nucléolo. El nucléolo, que consiste en partes del ADN de la cromatina combinado con ARN y proteínas, es el lugar donde se forman los ribosomas. 25 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES Figura 4. El Núcleo Diversos organelos celulares están relacionados a través del sistema endomembranoso. Ahora nos entraremos en los organelos eucarióticos que están formados por membranas interrelacionadas. Algunas de estas membranas están físicamente conectadas entre sí y algunas no, pero en conjunto constituyen una red citoplasmática que los biólogos denominan sistema endomembranoso. Muchos de los organelos de este sistema trabajan juntos en la síntesis, almacenamiento y exportación de moléculas importantes. Uno de los organelos, el retículo endoplasmático (RE), es el ejemplo típico de la directa interrelación entre las partes que constituyen el sistema de endomembranas. (El término retículo endoplasmático proviene de las palabras en griego que significan ―red dentro de la célula‖.) Como explicaremos enseguida, existen dos tipos de RE, el RE rugoso y el RE liso. Estos organelos difieren en estructura y función, pero las membranas que los forman son continuas. Las membranas del RE rugoso son también continuas con la membrana externa de la envoltura nuclear. El espacio dentro del RE está separado del fluido citoplasmático por la membrana del RE. De esta manera, las membranas interconectadas del RE y la envoltura nuclear dividen el interior de la célula en dos compartimientos separados. La división de la célula en compartimientos es una función primordial del sistema de endomembranas. 5.3.3. El retículo endoplasmático rugoso Fabrica membranas y proteínas. El término ―rugoso‖ en el retículo endoplasmático rugoso, o RE rugoso, se refiere a la apariencia de este organelo en las micrografías electrónicas. Como muestra la figura 4, la rugosidad resulta de la presencia de los ribosomas, los cuales tachonan las membranas de este organelo. El RE rugoso es una trama de sacos planos interconectados, con dos funciones principales. Una de ellas es fabricar más membrana. Algunas de las proteínas hechas por los ribosomas del RE son insertadas en la membrana del RE, así como también lo son los fosfolípidos hechos por las enzimas del RE. Como resultado, la membrana del RE se agranda, y parte de ella termina formando parte más tarde de otros organelos. La otra función principal del RE rugoso es fabricar proteínas que son secretadas por la célula. Un ejemplo de una proteína de secreción es un anticuerpo, el cual es una molécula de defensa elaborada y secretada por los glóbulos blancos. 26 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES Figura 4. Retículo Endoplasmático Rugoso Los ribosomas del RE rugoso sintetizan los polipéptidos del anticuerpo, los cuales se ensamblan para formar proteínas funcionales en el interior del RE. La figura 4 muestra la síntesis y el empaquetamiento de una proteína de secreción formada por un único polipéptido. (1) A medida que el polipéptido es sintetizado, pasa al interior del RE. (2) Pequeñas cadenas de azúcares se adicionan entonces al polipéptido, con lo que la molécula se transforma en una glucoproteína (gluco- significa azúcar). Cuando la molécula está lista para ser exportada fuera del RE, (3) el RE la empaqueta dentro de un pequeñísimo saco denominado vesícula transportadora. Esta vesícula se (4) separa a partir de la membrana del RE. La proteína de secreción viajará ahora hasta el aparato de Golgi para su posterior procesamiento. A partir de allí, una vesícula transportadora que contiene a la molécula definitiva, se abrirá paso hasta la membrana plasmática y liberará su contenido fuera de la célula. ¿Qué hace que el RE rugoso sea rugoso? Figura 5. Retículo Endoplasmático liso 27 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES 5.3.4. El retículo endoplasmático liso Posee una variedad de funciones. Como indica la figura 5, el retículo endoplasmático liso, o retículo endoplasmático liso, se continúa a partir del retículo endoplasmático rugoso. El retículo endoplasmático liso es una trama de túbulos interconectados desprovistos de ribosomas. La mayor parte de su actividad es llevada a cabo por enzimas embebidas en su membrana. Una de las funciones más importantes del retículo endoplasmático liso es la síntesis de lípidos, incluidos los ácidos grasos, fosfolípidos y esteroides. Cada uno de estos productos es fabricado por un tipo particular de células. En los mamíferos, por ejemplo, el retículo endoplasmático liso de las células de los ovarios y los testículos sintetiza las hormonas sexuales esteroidales. Las células de nuestro hígado también poseen grandes cantidades de retículo endoplasmático liso, con otros tipos de funciones. Ciertas enzimas en el retículo endoplasmático liso del hígado ayudan a regular la cantidad de azúcar liberada al torrente sanguíneo. Otras enzimas hepáticas ayudan a degradar las moléculas de las drogas y de otras sustancias potencialmente perjudiciales. Las drogas detoxificadas por estas enzimas incluyen, entre otras, sedativos tales como los barbitúricos, estimulantes tales como las anfetaminas y ciertos antibióticos. Cuando las células del hígado responden a las drogas, pueden ocurrir ciertas complicaciones no deseadas. A medida que las células se exponen a dichas sustancias químicas, la cantidad de retículo endoplasmático liso y sus enzimas detoxificadoras aumenta, incrementándose entonces la tolerancia del cuerpo a las drogas. Esto implica que se necesitan dosis cada vez más altas de una droga para conseguir un efecto particular, tal como la sedación. Otra complicación es que las enzimas detoxificadoras a menudo no pueden distinguir entre sustancias químicas relacionadas y por tanto responden a muchas de ellas de modo virtualmente similar, rompiendo una gran variedad de sustancias extrañas en la sangre. Como resultado, el crecimiento del retículo endoplasmático liso en respuesta a una droga puede aumentar la tolerancia a otras drogas, incluidas importantes medicinas. El uso de barbitúricos, por ejemplo, puede disminuir la eficacia de ciertos antibióticos. El retículo endoplasmático liso tiene además otra función, que es el almacenamiento de iones calcio. En el tejido muscular, éstos son importantes para la contracción. Cuando una señal nerviosa estimula a una célula muscular, los iones calcio fluyen desde el retículo endoplasmático liso al fluido citoplasmático, donde se dispara la contracción de la célula. 5.3.5. El aparato de Golgi Completa, clasifica y despacha los productos celulares. El aparato de Golgi recibió el nombre del biólogo y médico italiano Camilo Golgi, cuya carrera se prolongó hasta inicios del siglo xx. Utilizando el microscopio óptico, Golgi y sus contemporáneos descubrieron este organelo membranoso y varios otros más en células animales y vegetales. El microscopio electrónico ha revelado que el aparato de Golgi consiste en una pila de sacos aplanados. Como puede verse en la ilustración que sigue, estos sacos no están interconectados, como ocurre con los sacos del retículo endoplasmático. Una célula puede contener sólo unos pocos aparatos Golgi o cientos de ellos. La cantidad de aparatos de Golgi de una célula se correlaciona con el grado de actividad de dicha célula en la secreción de proteínas, un proceso que consiste en múltiples etapas y que se inicia, como hemos visto recientemente, en el retículo endoplasmático. El aparato de Golgi desempeña diversas funciones en estrecha colaboración con el retículo endoplasmático. En su función de almacén molecular y fábrica de refinamiento, el aparato de Golgi recibe y modifica las sustancias manufacturadas por el retículo endoplasmático. Uno de los lados del aparato de 28 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES Golgi funciona como muelle donde se reciben las vesículas de transporte producidas por el retículo endoplasmático. Cuando un aparato de Golgi recibe vesículas de transporte que contienen moléculas de glicoproteínas, por ejemplo, incorpora ese material y lo modifica químicamente. Una de las funciones de esta modificación química parece ser marcar y clasificar las moléculas en grupos diferentes para darles destinos diferentes. De acuerdo con una hipótesis popular, las moléculas se mueven de un saco a otro en el Golgi dentro de vesículas transportadoras. El sitio de ―despacho‖ del aparato de Golgi funciona como una terminal desde donde los productos de secreción ya terminados, empaquetados también en vesículas de transporte, se mueven hacia la membrana plasmática para su exportación desde la célula. Como alternativa, los productos terminados pueden volverse parte de la membrana plasmática en sí o de otro organelo, tal como un lisosoma. Figura 6. El Aparato de Golgi 5.3.6. Los lisosomas Digieren los alimentos y desechos de las células. El cuarto componente en el sistema de endomembranas, el liso- soma, es producido por el retículo endoplasmático rugoso y el aparato de Golgi. El nombre lisosoma deriva de dos palabras griegas que significan ―cuerpo de ruptura’ puesto que los lisosomas consisten en enzimas digestivas (hidrolíticas) incluidas en un saco membranoso. La formación y las funciones de los lisosomas están indicadas en la figura 6. El retículo endoplasmático y el aparato de Golgi cooperan en la fabricación de los lisosomas. Primero (figura 7), el retículo endoplasmático rugoso reúne a las enzimas y las membranas; luego, el aparato de Golgi refina químicamente las enzimas y libera lisosomas maduros. Los lisosomas ejemplifican lo que es el tema central en la estructura de la célula eucariótica, la formación de compartimientos. La membrana de los lisosomas encierra un compartimento donde se almacenan las enzimas digestivas y se las aísla sin peligro del resto del citoplasma. Sin lisosomas, una célula no podría tener enzimas hidrolíticas activas pues se digeriría a sí misma. Los lisosomas poseen varios tipos diferentes de funciones digestivas, como muestra la figura 7. Muchas células encierran a los nutrientes dentro de pequeños sacos citoplasmáticos denominados vacuolas 29 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES digestivas. Los lisosomas se fusionan con las vacuolas digestivas, exponiendo los nutrientes a las enzimas hidrolíticas que los digieren. Los productos moleculares más pequeños de la digestión, tales como los aminoácidos, abandonan el lisosoma para ser reusados por la célula. Los lisosomas ayudan también a destruir las bacterias perjudiciales. Nuestros glóbulos blancos ingieren a las bacterias dentro de vacuolas, y las enzimas lisosomales que se vacían dentro de esas vacuolas rompen las paredes celulares bacterianas. Además, los lisosomas funcionan como centros de reciclaje de los organelos dañados. Sin perjudicar a la célula, un lisosoma puede englobar y digerir partes de otro organelo, poniendo disponibles sus moléculas para la construcción de nuevos organelos. Los lisosomas también juegan un papel vital en el desarrollo embrionario. Por ejemplo, las enzimas lisosomales destruyen las células de las membranas que unen los dedos en los embriones humanos en los estadíos tempranos del desarrollo. Figura 7. Formación y funciones del lisosoma Los lisosomas anormales pueden causar enfermedades graves. La importancia de los lisosomas en la función celular y en la salud humana, es sorprendentemente evidente cuando se piensa en ciertos desórdenes moleculares graves denominados enfermedades de almacenamiento lisosomal. Una persona afectada con una enfermedad de almacenamiento lisosomal no posee ninguna de las enzimas hidrolíticas del lisosoma. Los lisosomas anormales se llenan con sustancias no digeridas, lo cual interfiere eventualmente con otras funciones celulares. La mayor parte de estas enfermedades son graves en la primera infancia. En la enfermedad de Pompe, se acumulan cantidades perjudiciales del polisacárido glucógeno en las células del hígado, debido a que los lisosomas carecen de una de las enzimas que digieren el glucógeno. La enfermedad de Tay-Sachs causa estragos en el sistema nervioso. En este desorden, los lisosomas carecen de una enzima digestiva de lípidos, y las células nerviosas en el cerebro se dañan a medida que se acumula el exceso de lípidos. Afortunadamente, las enfermedades de almacenamiento son raras en la población general. Con respecto a la enfermedad de Tay-Sachs, los portadores del gen anormal pueden ser identificados aun antes de que decidan tener hijos. 5.3.7. Las vacuolas Participan en el mantenimiento general de la célula. Al igual que los lisosomas, las vacuolas son sacos membranosos que pertenecen al sistema endomembranoso. Las vacuolas presentan formas y tamaños diferentes y desempeñan una amplia variedad de funciones. La vacuola alimenticia trabaja en colaboración con un lisosoma. Aquí, en la figura 8, se ve la vacuola central de una célula vegetal, la cual puede funcionar como un gran lisosoma. La vacuola central también ayuda a la célula vegetal a crecer en tamaño al absorber agua, y pude almacenar sustancias químicas vitales o productos de desecho del metabolismo celular. Las 30 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES vacuolas centrales en los pétalos de las flores pueden contener pigmentos que atraen a los insectos polinizadores. Otras contienen venenos que las protegen de los animales que se alimentan de plantas. La figura 9 muestra un tipo muy diferente de vacuola, en el protista por ejemplo el Paramecium. se alimenta de bacterias y se desplaza por medio de cilios. Nótense las dos vacuolas contráctiles, que semejan el eje de una rueda con radios centrales. Los ―radios‖ colectan el exceso de agua de la célula, y el eje central la expele hacia el exterior. Esta función es necesaria para los protistas de agua dulce, porque están constantemente incorporando agua del ambiente. Si no poseyeran alguna manera de liberarse del exceso de agua, el fluido celular se volvería demasiado diluido como para mantener la vida, y eventualmente la célula se hincharía y explotaría. Por esta razón, la vacuola contráctil es vital en el mantenimiento del medio interno celular. Figura 8. Vacuola Central en la célula de una planta Figura 9. Vacuolas contráctiles Revisión del Sistema de Endomembranas. Las relaciones que existen entre los organelos principales en el sistema endomembranoso (Figura 6). pueden verse las conexiones estructurales directas entre la envoltura nuclear, el retículo endoplasmático rugoso y el retículo endoplasmático liso. Las flechas rojas muestran las conexiones funcionales dentro del sistema de endomembranas, y cómo las vesículas de transporte viajan desde el retículo endoplasmático al aparato de Golgi y desde allí hasta otros destinos. Algunas de las vesículas se desarrollan para convertirse en lisosomas y vacuolas. Otras vesículas de transporte se fusionan con la membrana plasmática y contribuyen a ésta con sus propias membranas. El color azul dentro de los organelos del sistema y por fuera de la célula resalta el hecho de que un producto del retículo endoplasmático puede salir de la célula sin tener que cruzar en realidad una membrana. La vesícula de transporte que contiene ese producto se fusiona con la membrana plasmática y libera su producto al exterior. A continuación, estudiaremos dos organelos membranosos que no forman parte del sistema endomembranoso, el cloroplasto y la mitocondria. Ambos contienen ADN y ribosomas y fabrican algunas de sus propias proteínas; el resto de éstas son hechas por los ribosomas libres del citoplasma. Los cloroplastos y las mitocondrias son los procesadores de la energía, convierten la energía en aquellas formas que las células vivas pueden utilizar. 5.3.8. Los cloroplastos Transforman la energía solar en energía química. La mayor parte del mundo vivo funciona con la energía provista por la fotosíntesis, que consiste en la conversión de la energía lumínica del sol en energía química 31 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES de las moléculas de azúcar. Los cloroplastos son los organelos fotosintetizadores de las plantas y los protistas. El sistema de captación de energía solar del cloroplasto es mucho más eficiente que cualquier otro que el ingenio humano haya podido producir. Al igual que un organelo lleva a cabo procesos complejos en etapas múltiples, existen membranas internas que dividen al cloroplasto en tres compartimentos principales. Uno de los compartimentos es el delgado espacio intermembranoso, comprendido entre las membranas exteriores e interior del cloroplasto. El segundo, el espacio delimitado por la membrana interna, contiene un fluido espeso denominado estroma y una red de túbulos y discos huecos formados por membranas. El espacio en el interior de los túbulos y de los discos constituye el tercer compartimento. Los discos se disponen en pilas, cada una denominada granum (en plural, grana). Los grana son los paquetes de energía solar del cloroplasto, los sitios donde la clorofila realmente atrapa la energía del sol. Cada una de las partes del cloroplasto desempeña una función especial en la conversión de la energía solar en energía química. Figura 10. El Cloroplasto 5.3.9. Las mitocondrias Obtienen la energía química de los alimentos. Las mitocondrias son organelos que transforman la energía química de una forma en otra. Las mitocondrias llevan a cabo la respiración celular, en la cual la energía química de los alimentos tales como los azúcares es convertida en la energía química de una molécula denominada ATP (adenosin trifosfato). El ATP es la fuente principal de energía para el trabajo celular. La estructura de la mitocondria se ajusta a su función. En la figura puede verse que la mitocondria, al igual que el cloroplasto, está rodeada por dos membranas. Sin embargo, la mitocondria tiene sólo dos compartimentos. El espacio entre las membranas forma un compartimento lleno de fluido. La membrana interna rodea al segundo compartimento, el cual contiene un fluido denominado matriz mitocondrial. Muchas de las reacciones químicas de la respiración celular se llevan a cabo en la matriz. La membrana interna está muy plegada, y las moléculas de enzimas que producen el ATP se encuentran embebidas allí. Los pliegues, denominados crestas, aumentan el área superficial de la membrana, con lo que favorecen la capacidad de la mitocondria para producir ATP. 32 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES Figura 11. La Mitocondria El esqueleto interno de la célula ayuda a organizar su estructura y actividades. Muchos de los organelos que ya hemos descrito proporcionan algún tipo de soporte estructural a las células. Pero además, las células eucarióticas contienen una red de finas fibras de soporte, que en conjunto se denomina citoesqueleto. La esfera cerca del centro, teñida de anaranjado, es el núcleo. Las fibras del citoesqueleto se extienden a través de todo el citoplasma. Además de proveer un soporte estructural, las fibras del citoesqueleto están involucradas en el movimiento celular. Ciertos estudios recientes sugieren que regulan las actividades celulares al transmitir en forma mecánica ciertas señales desde la superficie hasta el interior celular. Tres tipos principales de fibras constituyen el citoesqueleto: microfilamentos, el tipo de fibras más fino; microtúbulos, el más grueso; y los filamentos intermedios, con un grosor medio. Los microfilamentos son bastones helicoidales sólidos compuestos principalmente por una proteína globular denominada actina. Nótese en el dibujo inferior de la izquierda en la figura 12, que un microfilamento es una doble cadena enroscada de moléculas de actina. Los microfilamentos de actina participan en el cambio de forma y el movimiento de las células, al ensamblarse (agregar subunidades) en un extremo mientras se desensamblan (pierden subunidades) en el otro. El movimiento ameboideo (de arrastre) del protista Amoeba y, por cierto, de nuestros glóbulos blancos involucran este tipo de proceso. Además, los microfilamentos de actina a menudo interaccionan con otros tipos de filamentos proteicos para producir la contracción de las células. Esta función de los microfilamentos se conoce muy bien a partir de estudios realizados sobre células musculares. Los filamentos intermedios son un grupo muy variado. Están constituidos por proteínas fibrosas en vez de proteínas globulares y tienen una estructura de cordón. Los filamentos intermedios funcionan principalmente como bastones de refuerzo para resistir la tensión, pero también ayudan a fijar ciertos organelos. Por ejemplo, el núcleo suele conservar su posición gracias a una jaula entretejida de filamentos intermedios. Los microtúbulos son tubos rectos y huecos compuestos por proteínas globulares denominadas tubulinas. Los microtúbulos se alargan añadiendo subunidades que consisten en pares de tubulina. Se desensamblan fácilmente de la manera opuesta, y las subunidades de tubulina pueden ser entonces reusadas en otro mícrotúbulo. Los microtúbulos que proporcionan rigidez y forma en una cierta área pueden desensamblarse y luego reensamblarse en algún otro lugar en la célula. Otras de las funciones importantes de los microtúbulos son proveer anclaje a los organelos y actuar como rieles para el movimiento de éstas dentro del citoplasma. Por ejemplo, un liso- soma puede moverse a lo largo de un mícrotúbulo para alcanzar una vacuola alimenticia. Los microtúbulos también guían el 33 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES movimiento de los cromosomas cuando las células se dividen, y, como veremos después, son la base del movimiento ciliar y flagelar. Figura 12. Fibras del Citoesqueleto. Los cilios y flagelos se mueven cuando se doblan los microtúbulos. El papel del citoesqueleto puede verse claramente en los flagelos y cilios de las células eucarióticas, éstos son apéndices locomotores que salen de ciertas células. Los flagelos y los cilios de las células eucarióticas poseen una estructura y un mecanismo de movimiento común. Los numerosos apéndices cortos que impulsan a protistas tales como el Trichodina a través del agua se llaman cilios. Los apéndices más largos, en general menos numerosos, presentes en otros protistas se denominan flagelos. Algunas células de organismos multicelulares también poseen cilios o flagelos. Por ejemplo, los cilios de las células que tapizan la tráquea humana barren el mucus con partículas hacia afuera de los conductos respiratorios. La mayoría de los animales y algunas plantas poseen espermatozoide flagelado. Como puede verse en la figura 13, un cilio o flagelo está compuesto por un centro de microtúbulos envuelto en una extensión de la membrana plasmática. Un par central de microtúbulos está rodeado por un anillo de nueve dobletes de microtúbulos. A este diseño, presente en casi todos los flagelos y cilios de las células eucarióticas, se le conoce como arreglo 9 + 2. Nótese que este diseño se extiende todo a lo largo del organelo, pero es diferente en la base. Aquí, los nueve dobletes se prolongan dentro de una estructura de anclaje denominada cuerpo basal, el cual posee un diseño de nueve tripletes de microtúbulos. El par central de microtúbulos termina por encima del cuerpo basal. Cuando un cilio o flagelo comienza a crecer, el cuerpo basal parece funcionar como una base para el ensamblaje de microtúbulos a partir de subunidades de tubulina. Los cuerpos basales tienen una estructura idéntica a los centriolos. En los cilios y flagelos, los microtúbulos proporcionan tanto el soporte como el mecanismo locomotor que impulsa la acción de batido de estos organelos. La figura 13 muestra la posición de dos dobletes de microtúbulos en un flagelo que se encuentra quieto (a la izquierda) y en proceso de doblarse (a la derecha). La curvatura de estos organelos involucra la participación de ―botones‖ de proteínas adosados a cada doblete de microtúbulos, los brazos de dineína, en rojo en las ilustraciones. Utilizando energía del ATP, los brazos de dineína se toman del doblete adyacente y ejercen una fuerza de deslizamiento a medida que tratan de ―caminar‖ a lo largo de él. Los dobletes se mantienen unidos entre sí por medio de entrecruzamientos (que no se muestran en la ilustración); si no se los mantuviera en su lugar, esta acción de caminar haría que un doblete resbalara a lo largo del otro. En su lugar, los microtúbulos (y en consecuencia, todo el flagelo o el cilio) se doblan. 34 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES Figura 13. Estructura de un flagelo o cilio de una célula eucariota Las superficies celulares protegen, sostienen y unen a las células. Con toda seguridad, se imaginará fácilmente que la delicada membrana plasmática no puede hacer frente por sí sola a todos los desafíos que ofrece el ambiente exterior de una célula. En efecto, la mayoría de las células poseen superficies de cubierta adicionales que rodean a la membrana plasmática. Debido a que la mayoría de los procariotas existen como células individuales o como agregados de células, sus cubiertas superficiales interactúan principalmente con entornos no celulares. Por el contrario, la mayoría de los organismos eucarióticos están compuestos por muchas células, las cuales se organizan en un organismo funcional único. En las plantas, las paredes celulares rígidas no sólo protegen a las células, sino que también proporcionan el soporte esquelético que mantiene a las plantas erguidas sobre el suelo. Las paredes celulares vegetales, típicamente unas 10-100 veces más gruesas que la membrana plasmática, consisten en fibras del polisacárido celulosa embebidas en una matriz de otros polisacáridos y proteínas. Esta fuerte construcción de fibras en una matriz recuerda a la fibra de vidrio, lo cual se observa también en su resistencia. La figura 12 muestra cómo están dispuestas las paredes de las células vegetales. Nótese que las paredes celulares poseen múltiples capas. Entre las paredes de células adyacentes existe una capa de polisacáridos adhesivos (en marrón oscuro) que mantiene a las células unidas entre sí. Las paredes de las células vegetales maduras pueden ser muy fuertes; son el componente principal de la madera, por ejemplo. A pesar de su grosor, las paredes de las células vegetales no aíslan totalmente a las células unas de otras. Para funcionar de manera coordinada como parte de un tejido, las células deben poseer uniones celulares, es decir estructuras que las conecten entre sí. Como muestra la figura 12A, existen numerosos plasmodesmos, canales entre células vegetales adyacentes, que forman un sistema circulatorio y de comunicación que conecta las células en los tejidos vegetales. Nótese que la membrana plasmática y el fluido citoplasmático en el dibujo se extienden a través de los plasmodesmos, de manera tal que el agua y otras moléculas pequeñas pueden pasar fácilmente de una célula a otra. Las células de un tejido vegetal comparten el agua, alimento y mensajes químicos a través de los plasmodesmos. Las células animales carecen de paredes celulares rígidas, pero la mayoría de ellas segregan y están embebidas en una capa adherente de glicoproteínas, denominada matriz extracelular (figura 12B). Esta capa ayuda a mantener juntas a las células de los tejidos y puede tener también funciones de protección y de sostén. Los científicos han descubierto que la matriz extracelular ayuda a regular el comportamiento celular, probablemente al contactar proteínas en la membrana plasmática, las cuales a su vez entran en contacto con fibras del citoesqueleto. 35 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES Las células adyacentes en muchos tejidos animales también se conectan entre sí por medio de las uniones celulares; existen tres tipos generales. Las uniones estrechas mantienen a las células unidas, formando una capa a prueba de filtraciones. Una capa de tejido de este tipo es la que tapiza el tracto digestivo, previniendo que los contenidos se filtren hacia los tejidos vecinos. Los desmosomas sujetan a células adyacentes entre sí o con la matriz extracelular. Las uniones de anclaje tachonan a las células con fibras del citoesqueleto pero a su vez permiten el paso de materiales por los espacios que quedan entre ellos. Los desmosomas son canales con una función similar a los plasmodesmos en las plantas; permiten el flujo de agua y de otras moléculas pequeñas entre células vecinas. Estas uniones son especialmente comunes en los embriones animales, donde la comunicación química entre las células es esencial para el desarrollo. Figura 12A Paredes Celulares Vegetales y uniones celulares Animal y uniones celulares. Figura 12B. Superficie de la Célula Los organelos de las células eucarióticas comprenden cuatro categorías funcionales. En este capítulo, hemos introducido varias estructuras celulares importantes. Para proporcionar un marco a toda esta información y reforzar la idea de que la estructura se correlaciona con la función, en la tabla de la derecha se agrupan los organelos de la célula eucariótica en cuatro categorías de acuerdo con su función general. La primera categoría es de la manufactura. Aquí no sólo incluimos la síntesis de moléculas, sino también su transporte dentro deja célula. La segunda categoría incluye tres organelos que degradan y reciclan materiales perjudiciales o que ya no se necesitan. (Se incluyen aquí las vacuolas, las que, por ser multifuncionales, no encajan claramente dentro de ninguna de nuestras categorías) La tercera categoría contiene los dos organelos que procesan la energía. La cuarta categoría es soporte, movimiento y comunicación intercelular. Estas tres funciones están relacionadas porque, para que el movimiento se desarrolle, debe existir algún tipo de soporte rígido contra el cual se pueda aplicar cierta fuerza. Y cuando una estructura de sostén forma el límite exterior de una célula, está necesariamente involucrada en la comunicación de esa célula con sus vecinas. Dentro de cada una de estas cuatro categorías existe una similitud estructural que subyace en la función general de los organelos. En la primera categoría, la manufactura depende fuertemente de una red de membranas metabólicamente activas. En la segunda categoría, todos los organelos de la lista están compuestos por sacos formados por una membrana simple, dentro de los cuales los materiales pueden ser químicamente degradados. En la tercera categoría, las expansiones de membranas metabólicamente activas dentro de los organelos, permite a los cloroplastos y a las mitocondrias llevar a cabo transformaciones de energía complejas que promueven el funcionamiento de la célula. Aun en la más diversificada de las categorías, y en la cuarta categoría, existe un tema estructural común dado por los distintos tipos de fibras involucradas en el funcionamiento de la mayoría de los organelos que se mencionan. Podemos resumir aún más esta información enfatizando en que estas cuatro categorías de Matriz extracelular organelos forman un equipo integrado, y que las (en animales) propiedades de la vida a nivel celular emergen Uniones celulares de las funciones coordinadas de los miembros de este equipo. El movimiento celular es una de estas propiedades básicas. En ese caso, las 36 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES mitocondrias proporcionan la energía que hace que los microtúbulos se doblen y que los cilios y flagelos se agiten. Como veremos las propiedades distintivas de la vida dependen de las acciones coordinadas de los organelos celulares. LOS ORGANELOS EUCARIÓTICOS Y SUS FUNCIONES Función general: Manufactura NÚCLEO Síntesis de ADN; síntesis de ARN; ensamblado de subunidades ribosomales (en el nucléolo) RIBOSOMAS Síntesis de polipéptidos (proteínas) RE rugoso Síntesis de proteínas de membrana, proteínas de secreción y enzimas hidrolíticas; formación de vesículas transportadoras RE liso Síntesis de lípidos, metabolismo de carbohidratos en las células de hígado; detoxificación en las células del hígado; almacenamiento de iones calcio APARATO DE GOLGI Modificación, almacenamiento temporal y transporte de macromoléculas; formación de lisosomas y de vesícula de transporte Función general: Degradación LISOSOMAS Digestión de nutrientes, bacterias y organelos dañados; destrucción de ciertas células durante el desarrollo embrionario PEROXISOMAS Diversos procesos metabólicos, incluyendo la degradación del H2 O2, un producto secundario del metabolismo VACUOLAS Digestión (similar a los lisosomas); almacenamiento de sustancias químicas; alargamiento celular; equilibrio del agua Función general: Procesamiento de la energía CLOROPLASTOS (en Transformación de la energía lumínica en energía química contenida en plantas) los azúcares MITOCONDRIAS Transformación de la energía química del alimento en energía química contenida en el ATP Funciones generales: Sostén, movimiento y comunicación entre células CITOESQUELETO Mantenimiento de la forma celular; anclaje de organelos; movimiento de (incluidos cilios, flagelos y organelos dentro de la célula; movimiento celular; transmisión mecánica de centriolos en las células señales desde el exterior al interior de la célula animales) PAREDES CELULARES Mantenimiento de la forma celular y sostén esquelético; protección (en plantas, hongos y superficial; unión de células en tejidos algunos protistas) MATRIZ EXTRACELULAR Unión de células en tejidos; protección superficial; regulación de las (en animales) actividades celulares UNIONES CELULARES Comunicación entre células; unión de células en tejidos Todas las formas de vida comparten características fundamentales. Las células son complejas. Sin embargo, podríamos argumentar que la estructura celular es más simple que lo que podríamos esperar, si consideramos todas las complejas funciones que una célula lleva a cabo y la diversidad de formas de vida que existe sobre la Tierra. Aunque es prácticamente casi cierto que la Tierra es el único planeta con vida en nuestro sistema solar, también es aceptable la idea de que las condiciones en algunas de las lunas de los planetas exteriores o en planetas de algún otro lugar hayan posibilitado la evolución de la vida. Supongamos por un momento que acabamos de saber que en realidad existen organismos extraterrestres. Tales organismos podrían ser muy diferentes de los que conocemos. ¿Qué características podríamos predecir si utilizamos nuestro conocimiento sobre los procariotas y eucariotas que viven en la Tierra? Podríamos, al menos, imaginar que los organismos extraterrestres tendrían estructuras complejas y que las características de su estructurafunción serían comunes a la de los organismos de la Tierra. Podríamos, además, predecir que estarían formados por algún tipo de unidades fundamentales, como las células, separadas de su ambiente por algún 37 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES tipo de membrana. Para poder reproducirse y adaptarse al ambiente, estos organismos tendrían que tener algún tipo de maquinaria genética en cada ―célula‖, tal vez contenida en un centro de control similar al núcleo. La base molecular de la herencia en otro planeta podría no ser el ADN; las moléculas biológicas allí podrían incluso estar basadas en un elemento distinto del carbono. Pero cualquiera que sean los detalles de su química, estas formas extrañas de vida tendrían que llevar a cabo algún tipo de metabolismo. Nuestra especulación sobre la vida extraterrestre nos ayuda a resumir lo que hemos aprendido en este capítulo acerca de la vida celular tal como la conocemos: todas las formas de vida en nuestro planeta comparten las características fundamentales de (1) estar formadas por células, cada una de ellas encerrada por una membrana que mantiene condiciones internas muy diferentes a las de sus alrededores; (2) poseer ADN como material genético; y (3) llevar a cabo metabolismo, el cual involucra la interconversión de formas diferentes de energía y de—materiales químicos. 6. MECANISMOS DE TRANSPORTE A TRAVÉS DE LA MEMBRANA CELULAR 6.1. Los Fosfolípidos Los Fosfolípidos de las membranas forman una Bicapa. Los lípidos, en especial los fosfolípidos, son el componente estructural principal de las membranas. La figura 1 ilustra la estructura de un fosfolípido. La fórmula estructural de la resalta las dos partes que constituyen la molécula, las cuales poseen interacciones opuestas con el agua. La cabeza es polar y, por tanto, hidrofílica. La doble cola es no polar e hidrofóbica. En la parte superior se muestra el símbolo para fosfolípidos que se usa comúnmente al ilustrar las membranas. Figura 1. Molécula de un fosfolípido La estructura de las moléculas de los fosfolípidos se ajusta muy bien a su papel en las membranas. En el agua, los fosfolípidos forman espontáneamente una lámina doble estable denominada bicapa fosfolipídica (figura 2). Sus cabezas hidrofilicas se ubican hacia afuera, exponiéndose al agua, y sus colas hidrofóbicas apuntan hacia adentro, protegidas del agua. Ésta es la disposición que adoptan las membranas de fosfolípidos en el ambiente acuoso dentro de los organismos vivos. Figura 2. Bicapa Fosfolipídica 38 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES El interior hidrofóbico de la bicapa es una de las razones por las que las membranas son selectivamente permeables. Las moléculas no polares, hidrofóbicas, son solubles en lípidos y pueden pasar fácilmente a través de las membranas, Por el contrario, las moléculas polares, hidrofilicas, no son solubles en lípidos. El hecho de que las moléculas polares puedan pasar a través de la membrana depende de ciertas moléculas de proteína presentes en la bicapa fosfolipídica. En efecto, gran parte de la permeabilidad selectiva de una membrana se debe a sus proteínas de membrana. ¿Por qué los fosfolípidos tienden a organizarse en una bicapa en un ambiente acuoso? La membrana es un mosaico fluido de fosfolípidos y proteínas El dibujo siguiente muestra la estructura de la membrana plasmática. Al igual que otras membranas celulares, comúnmente se la describe como mosaico fluido. La palabra ―mosaico‖ denota una superficie hecha de pequeños fragmentos, como piezas de un mosaico colorido cementadas en un piso o un cuadro. Una membrana es un mosaico en el sentido de que posee diversas moléculas de proteínas embebidas en un marco de fosfolípidos. El mosaico de membrana es fluido debido a que la mayor parte de las proteínas individuales y de las moléculas de fosfolípidos pueden moverse lateralmente dentro de la membrana. Para ayudar a mantener la delicada membrana en su lugar, algunas de las proteínas están ligadas tanto al citoesqueleto como al material extracelular adyacente. Nótense las colas retorcidas de muchos de los fosfolípidos. Como resultado de dobles enlaces en las colas lipídicas, estos dobleces hacen más fluida a la membrana al evitar que los fosfolípidos adyacentes se empaqueten fuertemente entre sí. El esteroide colesterol inserto en la membrana, ayuda a estabilizar los fosfolípidos a la temperatura corporal (alrededor de 37°C para los humanos) y mantener a la membrana fluida a temperaturas menores. En una célula viva, la bicapa de fosfolípidos se mantiene casi tan fluida como el aceite de una ensalada a temperatura ambiente. Cada una de las dos superficies de una membrana celular es diferente. Para la membrana plasmática, la superficie externa posee carbohidratos (cadenas de azucares, en verde) unidas covalentemente a proteínas y lípidos de las membrana. Una proteína con azúcares unidos se llama glucoproteína, mientras que un lípido con azúcares es un glucolípido. La porción de carbohidrato en una glucoproteína y un glucolípido varía de una especia a otra, así como también entre individuos de una misma especie, y aun entre distintos tipos celulares en un mismo individuo. Muchos de ellos funcionan como etiquetas de identificación de las células que sirven como reconocimiento para otras células. La capacidad para distinguir entre diferentes células es crucial para la vida, porque permite a las células en un embrión separarse entre sí para formar tejidos y órganos. También posibilita a las células del sistema inmune reconocer y rechazar las células extrañas, tales como bacterias infecciosas. La porción proteica, así como también los carbohidratos en las glicoproteínas, juegan un papel importante en el reconocimiento de superficies celulares. A continuación, veremos algunas otras funciones de las proteínas de membrana. Figura 3. La membrana Plasmática de una célula animal 39 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES Las proteínas hacen de la membrana un mosaico de funciones. El término ―mosaico‖ que se aplica a una membrana hace referencia no sólo a la distribución de las proteínas en la bicapa fosfolipídica, sino también a las diversas actividades que desarrollan dichas proteínas. En efecto, las proteínas desempeñan la mayor parte de las funciones de las membranas. Se han encontrado más de cincuenta tipos diferentes de proteínas en la membrana plasmática de los glóbulos rojos humanos, y probablemente hay muchas más aunque en número demasiado escaso como para que se las haya detectado hasta ahora. Hemos mencionado hasta aquí varias funciones de las proteínas de membrana, incluyendo el anclaje de la membrana al citoesqueleto y fibras externas, proveer con etiquetas de identificación, y formar uniones entre células adyacentes. ¿Qué otros papeles desempeñan las proteínas de membrana? Muchas proteínas de membrana son enzimas, que pueden funcionar como equipos catalíticos en las líneas de ensamblado de moléculas. Otras proteínas funcionan como receptores para mensajeros químicos de otras células. Una proteína receptora posee una forma tal que encaja con la de un mensajero específico, como una hormona, de la misma manera como una enzima encaja con su sustrato. A menudo, la unión del mensajero dispara una reacción en cadena que involucra a otras proteínas, las cuales retransmiten el mensaje a una molécula que desarrolla cierta actividad en el interior de la célula. Esto se denomina transducción de señales. Por último, algunas proteínas de membrana ayudan a mover sustancias a través de la membrana. Aunque las moléculas pequeñas como el O2 pasan libremente a través de la bicapa fosfolipídica, muchas de las moléculas esenciales necesitan ayuda de las proteínas para entrar o salir de la célula. La hormona epinefrina puede estimular una célula hepática para hidrolizar el glucógeno almacenado y liberar azúcar sin haber entrado a la célula. Explíquelo. Figura 4. Proteínas de Membrana 6.2. Transporte Pasivo El transporte pasivo consiste en la difusión a través de una membrana. La naturaleza de los fosfolípidos y el tipo de proteínas en una membrana determinan si una sustancia en particular puede o no cruzar la membrana. Ciertas moléculas pueden cruzar sin que la célula realice ningún trabajo, pero aun en estos casos se necesitan las proteínas de transporte, y los principios de la física determinan cuándo las moléculas pueden cruzar y en qué dirección lo harán. Difusión es la tendencia de partículas de cualquier tipo a separarse espontáneamente unas de otras hacia regiones donde están menos concentradas. La difusión no requiere ningún trabajo; es el resultado del movimiento al azar (energía cinética) de los átomos y las moléculas, y es promovida por la tendencia universal del orden a deteriorarse y convertirse en desorden (entropía). Debido a que la célula no realiza ningún trabajo cuando las moléculas difunden a través de su membrana, la difusión de cierta sustancia a través de una membrana biológica se denimina transporte pasivo. 40 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES La figura ilustra el transporte pasivo, y muestra cómo la concentración afecta la dirección en la que una sustancia cruza una membrana. En la figura 5, las moléculas de un colorante en solución tienden a difundir desde el lado de la membrana donde se encuentran más concentradas hacia el lado donde están menos concentradas, hasta que las soluciones a ambos lados posean iguales concentraciones del colorante. Dicho de otro modo, el colorante difunde a favor de su gradiente de concentración hasta que alcanza el equilibrio. En el equilibrio, las moléculas continúan moviéndose hacia adelante y hacia atrás, pero sin que haya ningún cambio neto en la concentración a ambos lados de la membrana. . Figura 5. Transporte pasivo El transporte pasivo es extremadamente importante para todas las células. En los pulmones, por ejemplo, el transporte pasivo a favor de gradientes de concentración es el único medio por el cual el oxígeno (O 2, esencial para el metabolismo) penetra en las células rojas, y el dióxido de carbono (CO 2, un desecho metabólico) sale de ellas. El agua es otra de las sustancias que cruzan las membranas por transporte pasivo. Ósmosis es el transporte pasivo de agua Por el hecho de que la célula contiene una solución acuosa en su interior y que además está rodeada por otra, y debido a que la membrana plasmática es permeable al agua, las moléculas de ésta pueden pasar fácilmente hacia adentro y hacia afuera de las células. La difusión de las moléculas de agua a través de una membrana selectivamente permeable es un caso especial de transporte pasivo conocido como ósmosis. La figura 6, muestra lo que ocurre si una membrana permeable al agua, pero no a los solutos, separa dos soluciones con diferentes concentraciones del soluto, por ejemplo una molécula como la glucosa. La solución con la mayor concentración de soluto se dice que es hipertónica (hyper, por sobre; tonos, tensión). La solución con la menor concentración de soluto es hipotónica (hypo, por debajo). El agua cruza la membrana hasta que las concentraciones del soluto (moléculas por mililitro de solución) sean iguales a ambos lados. En la representación amplificada de la figura 6, se puede ver lo que ocurre a nivel molecular. Las moléculas polares de soluto atraen grupos de moléculas de agua, de manera tal que son menos las moléculas de agua que se difunden libremente a través de la membrana. Con menor cantidad de moléculas de soluto, la solución hipotónica posee mayor cantidad de moléculas libres de agua, por lo que hay un movimiento neto de agua desde la solución hipotónica hacia la hipertónica. El resultado es la diferencia en los niveles de agua que se muestra en la figura 6. Acabamos de mostrar lo que ocurre con un único tipo de molécula de soluto; pero el mismo movimiento neto de agua ocurrirá sin importar la cantidad de solutos diferentes presentes. La dirección de la ósmosis está determinada solamente por la diferencia en la concentración total de soluto, y no por la naturaleza de los 41 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES solutos. Por ejemplo, el agua de mar contiene una gran variedad de solutos, pero de todas maneras perderá agua hacia una solución que contenga una concentración suficientemente elevada de un único soluto. Sólo si las concentraciones totales de solutos son las mismas a ambos lados de la membrana, las moléculas de agua se moverán a igual velocidad en ambas direcciones. Las soluciones que poseen igual concentración de soluto se dice que son isotónicas (isos, igual). Figura 6. El efecto general de la ósmosis Figura 7. Comportamiento de las células en diferentes soluciones. El equilibrio del agua entre las células y su entorno es crucial para los organismos. La figura 7 se ilustra cómo se aplica el principio de la ósmosis a las células vivas. Cuando una célula animal, tal como el glóbulo rojo que se muestra en la figura, es inmerso en una solución isotónica, el volumen de la célula permanece constante porque la célula incorpora agua a la misma velocidad con que la pierde. En esta situación, la célula y su entorno están en equilibrio porque ambas soluciones poseen la misma concentración total de solutos. Decimos que un organismo o célula en esta situación es isotónico respecto de la solución que le rodea. Muchos animales marinos, como las estrellas de mar y los cangrejos, son isotónicos respecto del agua de mar. Cuando una célula animal se encuentra en una solución hipotónica, la cual posee una concentración de soluto menor que la célula. La célula gana agua, se hincha y puede estallar (lisarse) como un globo demasiado inflado. Cuando una célula animal en una solución hipertónica se plasmoliza y puede morir por la pérdida de agua. Para que un animal pueda sobrevivir en el caso de que sus células estén expuestas a un ambiente hipertónico o hipotónico, el animal debe poseer un mecanismo para evitar la entrada excesiva o la pérdida de agua. El control del balance del agua se denomina osmorregulación. Por ejemplo, un pez de agua dulce, que vive en un ambiente hipotónico, posee riñones y agallas que trabajan constantemente para evitar una acumulación excesiva de agua en el organismo. 42 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES Los problemas del balance de agua son algo diferentes en las células vegetales debido a sus paredes celulares rígidas. Una célula vegetal inmersa en una solución isotónica es fláccida, y la planta se marchita en esta situación. Por el contrario, una célula vegetal está turgente, y las plantas están saludables, en un ambiente hipotónico, como se ve en la figura 8. Figura 8. Comportamiento de la célula vegetal y animal en diferentes soluciones Para estar turgente, una planta necesita un flujo neto de agua. Aunque la pared celular algo elástica se expande un poco, la presión que ejerce previene que la célula absorba demasiada agua y estalle, como le ocurriría a una célula animal en ese ambiente. La figura 8, parte c, muestra que en un ambiente hipertónico una célula vegetal no se encuentra nada mejor que una célula animal. A medida que la célula vegetal pierde agua, se plasmoliza y su membrana plasmática se separa de la pared celular. Por lo general, esto termina con la vida de la célula. Ciertas proteínas específicas facilitan la difusión a través de las membranas. Al discutir el tráfico de sustancias a través de membranas, nos hemos centrado hasta ahora en las moléculas pequeñas, incluida el agua, que difunden libremente a través de las membranas biológicas. Numerosas sustancias que no difunden libremente a través de las membranas, debido a su tamaño o carga, pueden cruzar por medio de mecanismos especiales que promueven y regulan su movimiento. Muchas moléculas se mueven a través de una membrana con la ayuda de proteínas de transporte presentes en la membrana. Cuando una de estas proteínas hace posible que una sustancia se mueva a favor de su gradiente de concentración, el proceso se denomina difusión facilitada. Sin la proteína, la sustancia no puede cruzar la membrana, o bien difunde a través de ella muy lentamente como para ser de utilidad a la célula. Al igual que la ósmosis, la difusión facilitada es un tipo de transporte pasivo porque no requiere ningún gasto de energía. Como ocurre con todo transporte pasivo, la fuerza propulsora es el gradiente de concentración. 43 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES Figura 9. Proteína transportadora que provee un conducto para el pasaje de un soluto. La figura 9, muestra el mecanismo más común por el cual las proteínas de membrana facilitan la difusión. La proteína transportadora (en morado) atraviesa la membrana y forma un conducto para el pasaje de un soluto en particular (anaranjado). La velocidad de la difusión facilita depende de cuantas moléculas de proteínas transportadora están disponibles en la membrana y de cuán rápido viaje el soluto específico a través de ella. 6.3. Transporte Activo Las células utilizan energía durante el transporte activo. En contraposición al transporte pasivo, el transporte activo requiere el uso de energía por parte de la célula para mover moléculas a través de la membrana. En dicha situación, la proteína transportadora bombea activamente un soluto determinado a través de una membrana en contra del gradiente de concentración del soluto, esto es, lejos del sitio donde se encuentra en menor concentración. Las proteínas de membrana por lo general utilizan ATP como fuente de energía para el transporte activo. La figura 10, muestra un sistema de transporte activo que involucra el pasaje de dos solutos diferentes a través de una membrana, en direcciones opuestas. La proteína transportadora posee un sitio de unión diferente para cada soluto. (1) El transporte activo comienza cuando uno de los solutos se une a la proteína transportadora. (2) Luego de la unión, un grupo fosfato se transfiere desde el ATP a la proteína. (3) La sobrecarga de energía que proviene de dicha transferencia (fosforilación) hace que la proteína modifique su forma y libera la molécula de soluto al otro lado de la membrana. (4) Al liberar la proteína al primer soluto, su forma y posición actual le permite al segundo soluto unirse a ella. (5) El grupo fosfato es entonces liberado. Esto hace que la proteína retorne a su forma original, (6) liberando al segundo soluto sobre el lado opuesto de la membrana. 44 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES Figura 10. Transporte activo de dos solutos a través de una membrana. Sistemas de transporte activo como éste, que acoplan el pasaje de dos solutos, son muy comunes en las membranas celulares, y transportan tanto iones como moléculas sin carga. Un ejemplo muy importante es la + + bomba de sodio-potasio (Na —K ) la cual participa en la generación de las señales nerviosas en las células neuronales. La exocitosis y la endocitosis transportan moléculas grandes. Hasta ahora nos hemos centrado en cómo el agua y los solutos pequeños entran y salen de las células moviéndose a través de la membrana plasmática. La historia es diferente para las moléculas grandes, como las proteínas. Como se muestra en la figura 11, la célula se vale de un proceso denominado exocitosis (del griego exo, hacia afuera, y kytos, célula) para exportar materiales voluminosos. Como primer paso en este proceso, una vesícula membranosa llena de macromoléculas se desplaza hacia la membrana plasmática. Una vez allí, la vesícula se fusiona con la membrana plasmática, y el contenido se vacía fuera de la célula. Cuando lloramos, por ejemplo, las células de las glándulas lacrimales se valen de la exocitosis para exportar una solución salina que contiene proteínas. En otro ejemplo, ciertas células del páncreas producen la hormona insulina y la secretan al torrente sanguíneo por medio de la exocitosis. Figura 11. Endocitosis y Exocitosis 45 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES La figura 11 ilustra el proceso opuesto a la exocitosis. En la endocitosis (endo, dentro de), la célula incorpora macromoléculas u otras partículas mediante la formación de vesículas o vacuolas a partir de la membrana plasmática. La figura 12 muestra tres tipos de endocitosis. La micrografía en color muestra a una ameba incorporando una partícula de alimento, lo cual es un ejemplo de fagocitosis, o ―alimentación celular’ La ameba engloba a su presa por medio de extensiones del citoplasma, llamadas pseudópodos, con las que formará una vacuola. La vacuola se fusiona entonces con un lisosoma, y las enzimas hidrolíticas del lisosoma digieren el contenido. La micrografía del centro muestra la pinocitosis, o ―toma de líquidos‖. Esta célula se encuentra en el proceso de tomar gotas de fluido de su alrededor por medio de pequeñas vesículas. La pinocitosis no es específica, ya que incorpora cualquiera y todos los solutos disueltos en las gotas de líquido. Contrariamente a la pinocitosis, el tercer tipo de endocitosis, denominada endocitosis mediada por receptor, es altamente específica. La micrografía de la derecha muestra uno de los pasos en el proceso. La membrana plasmática se ha hundido para formar una cavidad, la cual está tapizada con proteínas receptoras a las que se han adherido moléculas específicas de los alrededores. La cavidad se cerrará para formar una vesícula que conducirá a dichas moléculas al interior del citoplasma. Figura 12. Tres tipos de Endocitosis. Las membranas con fallas pueden sobrecargar la sangre de colesterol. Pocas moléculas aparecen en las noticias tan frecuentemente como el colesterol. A éste se le conoce como posible factor de las enfermedades del corazón, a pesar de que es esencial para el funcionamiento normal de nuestras células. Como hemos mencionado anteriormente, el colesterol es un componente de las membranas, así como 46 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES también la materia prima para sintetizar otros esteroides, como las hormonas sexuales. Los problemas tienden a aparecer sólo cuando se tiene demasiado colesterol en la sangre. En la mayoría de nosotros, las células del hígado remueven el exceso de colesterol de la sangre a través de la endocitosis mediada por receptores. En consecuencia, tenemos sólo pequeñas cantidades de esta sustancia en la sangre. El colesterol circula en la sangre principalmente en partículas denominadas lipoproteínas de baja densidad o LDL. Como muestra la figura 13, un LDL es un glóbulo de colesterol (y de otros lípidos) rodeados de una membrana simple de fosfolípidos en las que se embeben ciertas proteínas. Una de las proteínas del LDL (en rosa) se une a un tipo específico de proteína receptora (en morado) de las membranas celulares. Normalmente, las células del hígado toman LDL de la sangre por endocitosis mediada por receptor en la que los receptores son estas proteínas receptoras de LDL. Figura 13. Una célula efectuando la endocitosis mediada por receptores para incorporar una LDL En todo el mundo, alrededor de uno de cada 500 bebés humanos, heredan una enfermedad llamada hipercolesterolemia, que se caracteriza por un nivel excesivamente elevado de colesterol en la sangre. En los casos severos (alrededor de uno en un millón de personas), no hay receptores de LDL funcionales. Las personas con una forma muy leve de enfermedad poseen receptores funcionales, pero en cantidades más bajas. En cualquiera de estos casos, los LDL tienden a acumularse en la sangre. Los niveles altos de LDL en la sangre ponen en peligro la vida debido a que los LDL pueden depositar el colesterol en el interior de los vasos sanguíneos, obstruyéndolos eventualmente. El bloqueo de los vasos sanguíneos que alimentan el corazón produce enfermedades coronarias. Los individuos con una hipercolesterolemia severa pueden morir de enfermedades del corazón durante la infancia temprana. Los casos más leves también son peligrosos, pero pueden ser tratados con medicación y una dieta baja en grasas. 47 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES 7. DIVISIÓN CELULAR 7.1. Mitosis Mitosis es la división nuclear más citocinesis, y produce dos células hijas idénticas durante la profase, profase, metafase, anafase y telofase. La interface frecuentemente se incluye en discusiones sobre mitosis, pero la interfase, técnicamente, no es parte de la mitosis, más bien incluye las etapas G1, S y G2 del ciclo celular. 7.1.1. Fases de la Mitosis Profase. Los filamentos de cromatina se condensan para formar cromosomas, cada uno de los cuales está compuesto por dos fibras separadas denominadas clemátides, que están unidas en un punto mediante una región especializada, el centrómero. A medida que avanza la profase, un centriolo se desplaza a cada uno de los lados de la célula. Cerca de los centriolos se forman micro túbulos, que se proyectan en todas las direcciones. Al final de la profase, la cubierta nuclear degenera y los nucléolos han desaparecido. Metafase. Los cromosomas se alinean en el ecuador con las fibras fúsales procedentes de cada para de centriolos sujetas a sus centrómeros. 48 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES Anafase. Los centrómeros se separan, y cada clemátide es ahora un cromosoma. Cuando los centrómeros se dividen, el número de cromosomas se duplica, produciendo dos copias idénticas de 46. Las dos copias de cromosomas son atraídas por las fibras del huso hacia los polos de la célula. El citoplasma se estrecha a lo largo de las células del ecuador a medida que las membranas celulares se mueven unas hacia otras. Telofase. A partir del retículo endoplasmático se forma una nueva cápsula nuclear y reaparecen los nucléolos como organelos diferenciados. Los núcleos de las dos células hijas adquieren el aspecto de los núcleos en la interfase. Se ha completado el proceso de la mitosis. 49 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES Interfase. El período existente entre las divisiones celulares activas se denomina interfase. Durante este período el DNA y sus proteínas asociadas tienen el aspecto de hilos de cromatina dispersos dentro del núcleo. Cuando comienza la replicación, las dos cadenas de cada molécula de DNA se separan y cada una de ellas sirve de modelo para la producción de una nueva cadena de DNA. Durante la interfase, también se duplican los centriolos situados dentro del centrosoma. 50 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES 7.2. Meiosis 7.2.1. Meiosis I La Meiosis I consiste en la primera etapa de la llamada Meiosis completa, compuesta por dos fases. A su vez, la Meiosis I también contiene distintas fases o procesos muy similares a los de la mitosis. Se trata de un proceso también llamado mitosis mitocondrial ya que origina células con la mitad de cromosomas. Este proceso sólo tiene lugar en las gónadas, células diploides encargadas de la reproducción de las células sexuales. 7.2.1.1. Fases La replicación del ADN sucede antes del comienzo de la meiosis I. 51 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES PROFASE I: Los eventos de la Profase I son similares a los de la Profase de la mitosis: la cromatina se condensa en los cromosomas, el nucléolo se disuelve, desaparece la membrana nuclear, y se forma el huso mitótico. Durante la profase I, justo después de que se condense la cromatina, los cromosomas homólogos se sobre cruzan. Esto sólo ocurre en la meiosis. Los cromosomas sobre cruzados se llaman bivalentes. Este proceso es clave en la Meiosis, ya que permite que las células nuevas que se creen sean distintas entre ellas y con la célula original. La condensación de los cromosomas permite que estos sean vistos en el microscopio. METAFASE I: En esta fase intermedia, los cromosomas, de dos en dos (por grupo de homólogos sobre cruzados), se alinean en la placa ecuatorial (zona central de la célula), agarrados a las fibras del huso cromático por sus centrómeros. Es una fase que sucede muy rápida. ANAFASE I: Las fibras del huso acromático se rompen, tirando de cada uno de los cromosomas de cada par de homólogos hacia un polo celular. Las fibrillas acaban contrayéndose también en los distintos polos. 52 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES TELOFASE I: En la Telofase I los cromosomas forman dos núcleos hijos. Es la etapa final de la Meiosis I, y se caracteriza por ser inversa a la Profase I. En él desaparecen los restos del huso acromático, aparece una membrana nuclear a partir de los restos de la ya destruida en cada uno de los polos, se desespirilizan las cromátidas. 53 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES Grafica meiosis l: 7.2.2. Meiosis II Es muy similar a la mitosis sin embargo ella no prescindía por la duplicación del ADN, este hecho es de gran importancia puesto que determina que las células resultantes sean haploide. Se distinguen las siguientes etapas: 7.2.2.1. Fases PROFASE II: En esta etapa el ADN vuelve a empaquetarse reconstituyendo los cromosomas duplicados. Desaparece la membrana nuclear y se reinicia la formación de las fibras del huso. METAFASE II: Los cromosomas duplicados se disponen en la placa ecuatorial de igual manera que en una metafase mitótica. 54 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES ANAFASE II: En esta etapa las cromatidas hermanas de cada cromosoma se separan obteniéndose cromosomas simples los que se desplazan hacia los polos opuestos. TELOFASE II: En esta última etapa desaparecen las fibras del huso acromático, la membrana nuclear se reorganiza y los cromosomas desaparecen por descondensación del ADN. 55 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES Grafica MEIOSIS ll: 56 UNIDADES TECNOLÓGICAS DE SANTANDER APUNTES DOCENTES BIBLIOGRAFÍA CURTIS, Barnes. Biología. Sexta edición. Panamericana. Buenos Aires. 2000 BIGGS, Alton. Biología la dinámica de la vida. Mc Graw-Hill. BERNESTEIN, Ruth. Biología. Séptima edición. Mc Graw-Hill. FRIED, George. Biología Shawm. Sexta edición. Mc Graw-Hill. KIMBALL, John. Biología. Addison Wesley. 1986. MADER, Sylvia. Biología. Décima edición. Mc Graw-Hill OPARIN. A. Ivanovich. El Origen de la vida. Panamericana editorial Ltda. Bogotá. 2000 VILLE. A, Claude. Biología. Mc Graw-Hill. Mexico.1988. 57