5.5. Metabolismo. - Colegio Maravillas
Anuncio

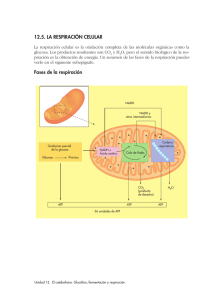
5.5. Metabolismo. 5.5.1. Concepto de metabolismo, catabolismo y anabolismo. 5.5.2. Aspectos generales del metabolismo: reacciones de oxidorreducción y ATP. 5.5.3. Estrategias de obtención de energía: energía química y energía solar. 5.5.4. Características generales del catabolismo celular: convergencia metabólica y obtención de energía. 5.5.4.1. Glucolisis. 5.5.4.2. Fermentación. 5.5.4.3. Respiración: ciclo de Krebs, cadena respiratoria y fosforilación oxidativa. 5.5.4.4. Balance energético del catabolismo de la glucosa 12. Explicar el concepto de metabolismo, catabolismo y anabolismo. Diferenciar entre catabolismo y anabolismo. Realizar un esquema de las fases de ambos procesos. 13. Reconocer y analizar las principales características de las reacciones que determinan el catabolismo y el anabolismo. 14. Destacar el papel de las reacciones de óxido-reducción como mecanismo general de transferencia de energía. 15. Destacar el papel del ATP como vehículo en la transferencia de energía. 16. Resaltar la existencia de diversas opciones metabólicas para obtener energía. 17. Definir y localizar la glucolisis, la β-oxidación, el ciclo de Krebs, la cadena de transporte electrónico y la fosforilación oxidativa indicando los sustratos iniciales y productos finales. 18. Comparar las vías anaerobias y aerobias en relación a la rentabilidad energética y los productos finales. Destacar el interés industrial de las fermentaciones. 19. Reconocer que la materia y la energía obtenidas en los procesos catabólicos se utilizan en los procesos biosintéticos y esquematizar sus fases generales. En la misma línea de lo indicado en la primera observación, no es necesario formular los intermediarios de las rutas metabólicas, aunque el alumno deberá conocer los nombres de los sustratos iniciales y de los productos finales. 5.5.1. Concepto de metabolismo, catabolismo y anabolismo. El metabolismo es el conjunto de reacciones y procesos físico-químicos que ocurren en una célula. Estos complejos procesos interrelacionados son la base de la vida a nivel molecular, y permiten todas las actividades de las células: crecer, reproducirse, mantener sus estructuras, responder a estímulos, etc. El metabolismo se divide en dos procesos conjugados: catabolismo y anabolismo. Las reacciones catabólicas liberan energía; un ejemplo es la glucólisis, un proceso de degradación de compuestos como la glucosa que liberan la energía retenida en sus enlaces químicos. Las reacciones anabólicas o de síntesis, en cambio, consumen la energía liberada en las reacciones catabólicas para recomponer enlaces químicos y construir componentes de las células como lo son las proteínas y los ácidos nucleicos. El anabolismo y el catabolismo son procesos acoplados ya que cada uno depende del otro. La economía que la actividad celular impone, obliga a organizar estrictamente las reacciones químicas del metabolismo en vías o rutas metabólicas, donde un compuesto químico (sustrato) es transformado en otro (producto), y este a su vez funciona como sustrato para generar otro producto, siguiendo una secuencia de reacciones bajo la intervención de diferentes enzimas (generalmente una para cada sustrato-reacción). Las enzimas son cruciales en el metabolismo porque agilizan las reacciones físico-químicas, pues hacen posibles reacciones termodinámicas "desfavorables". Las enzimas también se comportan como factores reguladores de las vías metabólicas, modificando su funcionalidad y por ende, la actividad completa de la vía metabólica en respuesta al ambiente y necesidades de la célula, o según señales de otras células. Una característica del metabolismo es la similitud de las rutas metabólicas básicas incluso entre especies muy diferentes. Por ejemplo: la secuencia de pasos químicos en una vía metabólica como el ciclo de Krebs es universal entre células vivientes tan diversas como un paramecio y organismos pluricelulares como el hombre. Esta estructura metabólica compartida es muy probablemente el resultado de la alta eficiencia de estas rutas, de su temprana aparición en la historia evolutiva y la selección molecular aleatoria. ESQUEMA GENERAL DEL METABOLISMO CELULAR El anfibolismo es el proceso metabólico en los que, gracias a unos sistemas enzimáticos y a algunas moléculas preexistentes, se oxidan metabolitos y se almacena gran cantidad de energía que posteriormente se usarán en rutas anabólicas. En los procesos anfibólicos (que son fuertemente endergónicos), la energía necesaria no puede proceder del propio sistema, sino que tiene que provenir del medio. Por las mismas razones, los procesos catabólicos y anfibólicos (que son exergónicos), no puede tener un número infinito de pasos intermedios, tiene que existir un aceptor último de electrones que no pertenezca al sistema. MOLÉCULAS QUE INTERVIENEN EN EL METABOLISMO Además de los enzimas son necesarias las siguientes moléculas: Metabolitos, (glucosa, ácidos grasos, acetil CoA, etc.) Nucleótidos, NAD+,NADP+, FAD, FMN. Permiten una reducción u oxidación de metabolitos, formando auténticos pares redox. Actúan como transportadores de electrones: Moléculas ricas en energía, generalmente vinculadas al grupo fosfato (típico de los ATP ADP AMP): Moléculas ambientales como el oxígeno, agua, CO2, alcohol etílico etc. RENDIMIENTO Y BALANCE ENERGÉTICO DEL METABOLISMO La célula obtiene su energía a partir de degradaciones oxidativas de moléculas como glucosa, grasas etc., esto implica la reducción de otras. En general, los procesos catabólicos son exergónicos para que las células puedan vivir y realizar procesos anabólicos o de síntesis. La cantidad de energía desprendida en un proceso exergónico depende del desnivel energético entre el estado inicial y final del sistema, es decir, depende de la diferencia existente entre el el potencial de reducción de la molécula que comienza la ruta y el de la última que se reduce y permite la oxidación de los anteriores. La energía química es la única que puede aprovechar el ser vivo, y este aprovechamiento puede realizarlo directamente mediante el acoplamiento energético o almacenando la energía en forma de enlaces ricos en energía enlaces fosfato del ATP). EL parámetro que mide el rendimiento o balance energético es el número de moléculas de ATP que surgen en el proceso de oxidación del metabolito oxigenado. Si la ruta es catabólica el balance es positivo, si es anabólica el balance es negativo. Balance energético positivo: Un ejemplo típico es la glucolisis (degradación de la glucosa a ácido pirúvico) Dos fosforilaciones del sustrato (glucosa y fructosa) ........... Dos reacciones de oxidación (gliceraldehido 3-fosfato)....... Dos reacciones de oxidación (gliceraldehído 1-3 difosfato).. -2 ATP +2ATP +2 ATP ______ +2ATP Balance energético negativo: Un ejemplo típico: (síntesis de un polipéptido de 10 aminoácidos) Unión de 10 aminoácidos a 10 ARNt (2 ATP cada uno).......... Acoplamiento de 9 aminoacil-ARNt en el ribosoma ............. Desplazamiento del ribosoma espués de 9 enlaces peptídicos .. -20 ATP -9 ATP -9 ATP ______ -38 ATP Toda la energía que se desprende en un proceso metabólico no se aprovecha, ya que parte de ella se traduce en forma de calor. Por esta razón se define como rendimiento energético como el porcentaje de energía almacenada respecto a la cantidad total desprendida en un proceso catabólico. Como en cada mol de ATP hay almacenada 7,3 Kcal, se puede calcular el rendimiento del proceso fácilmente. 5.5.2. Aspectos generales del metabolismo: reacciones de oxidorreducción y ATP. LAS REACCIONES REDOX TRANSFIEREN ELECTRONES Y ENERGÍA Hemos visto anteriormente que el ADP es capaz de usar reacciones exergónica para fosforilarse y formar ATP, almacenando en sus enlaces la energía absorbida. Otra manera de transferir energía es transferir electrones. Las reacciones en las que hay transferencia de uno o dos electrones se denominan reacciones redox. La ganancia de electrones se denomina reducción. La pérdida de uno o dos electrones se denomina oxidación. A pesar de que la oxidación y la reducción siempre están asociadas al intercambio electrónico, también podemos pensar en estos términos cuando los átomos de hidrógeno (no iones de hidrógeno) se pierden o ganan. La razón es obvia, cuando una molécula pierde un átomo de hidrógeno se oxida y cuando gana un átomo de hidrógeno se reduce. OXIDACIÓN AH2 + B BH2 + A REDUCCIÓN La oxidación y reducción siempre ocurren juntas: a medida que un material se oxida, los electrones que pierden son transferidos a otro material, reduciéndolo. En una reacción redox denominamos al reactivo que se reduce agente oxidante y al que se oxida, agente reductor. Un agente oxidante acepta electrones: en el proceso de oxidar el agente reductor, el propio agente oxidante se reduce. Por el contrario, el agente reductor dona electrones; se oxida en la medida que reduce al agente oxidante. En el metabolismo de la glucosa, ésta es el agente reductor y el gas oxígeno, el agente oxidante. En una reacción redox, la energía se transfiere. EL G global de una reacción redox es negativo. EL ATP ACOPLA REACCIONES EXERGÓNICAS Y ENDERGÓNICAS E INTERVIENE EN LA TRNSFERENCIA DE ENERGÍA EN LAS CÉLULAS Todas las células vivas se basan en el ATP para capturar, transferir y almacenar la energía libre necesaria para realizar sus funciones vitales. Se puede considerar que el ATP es una moneda universal de intercambio energético en la célula. El ATP se produce en las células de diversas formas; cuando se hidroliza el ATP libera energía entregando ADP y un ión fosfato inorgánico además de energía libre: ATP + H2O ADP + Pi + energía libre En esta reacción es importante recordar que: Es exergónica, libera energía libre El equilibrio está desplazado hacia la derecha, es decir, hacia la producción de ADP. En el equilibrio de la célula, hay 10 millones de veces más ADP que ATP. Muchas reacciones exergónicas catalizadas por enzimas en la célula pueden proporcionar la energía para convertir ADP en ATP. En las células eucarióticas, la más común es la denominada “respiración celular”, en la cual la energía liberada por las moléculas de combustible (glucosa, ac. grasos, etc.), es atrapada y almacenada en el ATP. La síntesis e hidrólisis del ATP constituye un ciclo de acoplamiento de energía, en el que el ATP transporta la energía desde las reacciones exergónicas a las endergónicas. Cuando se forma ATP captura energía libre; posteriormente difunde a otros lugares de la célula, donde su hidrólisis libera energía libre para impulsar una reacción endergónica. Una célula activa requiere millones de moléculas de ATP por segundo para impulsar su maquinaria bioquímica. Una molécula de ATP se consume al minuto e haberse sintetizado. En reposo una persona promedio produce 40 kg. de ATP diarios lo que significa que cada moléculas de ATP sufre cerca de 10000 ciclos de síntesis e hidrólisis diarias. 5.5.3. Estrategias de obtención de energía: energía química y energía solar. Es importante conocer las formas de nutrición de los organismos según el tipo de materia que intercambian con su entorno y la fuente primaria de energía que les resulta útil para fabricar su propia biomasa. Clasificación de los organismos según su forma de nutrición. Para que un organismo pueda sobrevivir necesita: Una fuente ambiental de carbono para construir sus moléculas. Dependiendo de la fuente de carbono podemos clasificar a los seres vivos en: - Autótrofos, si asimilan el CO2 ambiental. - Heterótrofos, si usan moléculas orgánicas sencillas. Una fuente ambiental de hidrógeno (electrones) para reducir moléculas que, al aceptarlo, alcancen un elevado potencial de reducción, es decir, un potencial redox muy negativo. Según esta fuente, la clasificación sería: - Litótrofos, si el H procede de una fuente inorgánica - Organótrofos, si precisan de moléculas más complejas. Una fuente primaria de energía que haga posible esa reducción. Así, según el tipo de energía que aprovecha, se distinguen organismos: - Fotótrofos, si aprovechan la ENERGÍA SOLAR directamente. - Quimiótrofos, si solo se pueden servir de energía química. Un aceptor último de hidrógenos (electrones), que permita la oxidación del aceptor anterior, liberando la energía necesaria para la síntesis de biomoléculas. Así podemos hacer la siguiente clasificación: - Aerobios, si el oxígeno es el último aceptor. - Anaerobios, si es otra sustancia la que finalmente recibe los electrones. Un suministro ambiental de agua, sales minerales y nitrógeno para construir proteínas. Puesto que los organismos autótrofos son generalmente litótrofos y los heterótrofos organótrofos, podemos simplificar esta clasificación de la siguiente manera: CLASIFICACION DE LOS ORGANISMOS DEPENDIENDOS DE SU TIPO DE NUTRICIÓN LITÓTROFOS (Autótrofos) ORGANÓTROFOS (Heterótrofos) FOTÓTROFOS Fotolitotrofos. Bacterias fotosintéticas del azufre y metafitas. Fotoorganótrofos Bacterias purpúreas no Metazoos y los Fungi QUIMIÓTROFOS Quimiolitótrofos Bacterias quimiosintéticas Quimiorganótrofos Muchas bacterias, sulfurosas 5.5.4. Características generales del catabolismo convergencia metabólica y obtención de energía. celular: El catabolismo comprende el metabolismo de degradación oxidativa de moléculas orgánicas, cuya finalidad es la obtención de energía necesaria para que la célula realice sus funciones vitales. La célula tiene que disponer de un último aceptor de electrones de los hidrógenos desprendidos en las rutas de oxidación. Según la naturaleza de este aceptor clasificamos a los seres vivos en: aeróbicos o aerobios, si el aceptor es oxígeno molecular (O2), o anaeróbicos o anaerobios, si es otra molécula (NO2-, SO42-). Desde una perspectiva evolutiva, los seres anaerobios son mucho más antiguos que los aerobios ya que la atmósfera primitiva era reductora en vez de oxidante. Todas las transformaciones moleculares que desprenden energía en los procesos catabólicos son reacciones de oxidación. Las reacciones de este tipo son aquellas en las que se transfieren electrones de un átomo o molécula a otro. Toda oxidación requiere una reducción, por lo que estos procesos se denominan redox (ver epígrafes anteriores). La transferencia de electrones en un proceso catabólico se realiza en un orden preciso que viene determinado por el potencial re reducción de cada par redox, comenzando por el que tenga potencial más negativo. Un par redox está compuesto por las dos especies que intervienen en la reacción de oxido-reducción. Cuanto mayor sea la diferencia entre el potencial de reducción del estado inicial y del estado final de la ruta catabólica, tanto mayor será la energía desprendida en el proceso. Los átomos de H liberados en las reacciones de oxidación van acompañados de gran cantidad de energía que estaba almacenada en los enlaces de los que formaban parte. Los transportadores de hidrógeno son nucleótidos no nucleicos como el NAD+, el NADP+ o el FAD, que captan los H liberados por las moléculas oxidadas y los transfieren a las moléculas aceptoras, que finalmente se reducirán. 5.5.4.1. Glucolisis. También denominada ruta de Emdben- Meyerhof, ocurre en le citosol; no necesita oxígeno y es un proceso en el que una secuencia de reacciones catalizada por encimas específicos, degrada una molécula de glucosa hasta convertirla en dos de ácido pirúvico o piruvato. Las etapas se podrían resumir así: Etapa 1: Fosforilación de la glucosa con consumo de una molécula de ATP Etapa 2: Isomerización de la glucosa 6-P en fructosa 6-P Etapa 3: Fosforilación de la fructosa 6-P con gasto de una molécula de ATP, formándose fructosa 1-6 bifosfato. Etapa 4: Rotura de la fructosa 1-6 bifosfato en dos triosas en equilibrio; la 3fosfogliceraldehído(G3P) y la 3-dihidroxiacetona(DAP). A partir de este punto seguimos la ruta de una de estas moléculas y posteriormente multiplicamos por dos los resultados energéticos conseguidos. Etapa 5: El DAP se reorganiza para formar su isómero. Etapa 6: Oxidación y fosforilación del 3 fosfo-gliceraldehído (G3P), empleando un Pi y reduciendo dos moléculas de NAD+ Etapa 7: Desfosforilación del ácido 1,3-bifosfoglicerladehído, formándose una molécula de ATP por cada una de las moléculas implicadas. Etapa 8: Isomerización del ácido 3-fosfoglicérico, cambiando su grupo fosfato del C3 al C2. Etapa 9: Formación de un doble enclace como consecuencia de la pérdida de un átomo de hidrógeno y un grupo –OH en el ácido 2-fosfoglicérico. Etapa 10: Desfosforilación del ácido fosfoenol pirúvico (PEP), dando ácido pirúvico y ATP BALANCE ENERGÉTICO Se necesitan dos moléculas de ATP para comenzar la ruta, una vez comenzado se producen dos moléculas de NADH y cuatro de ATP. Por tanto, el balance es de dos moléculas de NADH y dos moléculas de ATP por cada glucosa. La ecuación global es: Glucosa + 2 ADP + 2Pi + 2NAD+ 2 Ácido pirúvico + 2ATP + 2 NADH + 2H+ + 2 H2O Etapas clave de la glucolísis: En la etapa 5, si el NADH producido no se vuelve a oxidar, la ruta se detendrá. El modo de oxidarse dependerá de la disponibilidad de oxígeno: En condiciones aerobias, las moléculas de NADH ceden sus electrones a la cadena de transporte electrónico, que los conducirá hasta el oxígeno, produciendo agua y regenerando el NAD+, que se reutilizará en la glucolisis. En estas condiciones, el ácido pirúvico entra en la mitocondria y se transforma en grupos acetilo, que formarán el acetil coenzima A (acetil CoA), que se incorpora en la respiración celular. En condiciones anaerobias, ya sea en bacterias o en células eucarióticas sometidas en condiciones de anoxia, el NADH se oxida a NAD + mediante la reducción del ácido pirúvico. Estas etapas hacen posible que se produzca energía de forma anaeróbica (FERMENTACIONES) y ocurre en el citosol. Otras rutas catabólicas: Β-oxidación de los ácidos grasos. Los ácidos grasos son moléculas que el organismo usa como reserva de energía metabólica. En el citoplasma celular se hidrolizan las grasas (triacilglicéridos) por acción de las lipasas originándose glicerina más ácidos grasos. Los fosfolípidos también se hidrolizan en ácido fosfórico y ácidos grasos. Antes de ser oxidados, los ácidos grasos se activan en la membrana mitocondrial externa uniéndose al acetil-CoA. El catabolismo de los ácidos grasos ocurre en la matriz mitocondrial y en los peroxisomas, y consiste en la oxidación del carbono β, eliminándose de forma secuencial unidades de dos átomos de carbono. El carbono C3 (carbono β) es el que sufre esta oxidación; también se denomina hélice de Lynen en honor a uno de sus descubridores y también porque la cadena se va acortando progresivamente. La carnitina actúa como lanzadera de ácidos grasos a través de un enzima translocasa , para pasar la membrana mitocondrial. El resultado final es la obtención de acetil-CoA que se incorpora al ciclo de Krebs. 5.5.4.2. Fermentación Cuando el catabolismo de la glucosa ocurre en condiciones anaerobias y el último aceptor de hidrógenos (o de electrones) no es el oxígeno, sino que es una molécula orgánica sencilla, la ruta se denomina fermentación. Posiblemente sea una de las rutas más antiguas ya que la atmósfera primitiva era reductora y no había oxígeno. Algunas células de metazoos (músculo estriado) y metafitas (oxidando NADH extramitocondrial), ocasionalmente pueden trabajar en condiciones de anaerobiosis, y realizar fermentaciones. En el mundo de los moneras (bacterias), las fermentaciones es la forma habitual de oxidar la glucosa ya que sus necesidades energéticas son menores que la de los seres pluricelulares eucariotas. Fermentación alcohólica o etílica: Los vegetales, hongos y levaduras (Saccharomyces cerevisiae) poseen la enzima piruvato deshidrogenasa responsable de este fenómeno. Desde las primeras civilizaciones se han descrito fermentaciones alcohólicas usando levaduras para obtener bebidas con más o menos graduación alcohólica. El efecto Pasteur es el que se usa para promocionar o frenar la fermentación de los mostos, aumentando o disminuyendo la concentración deO2. Si se elimina el oxígeno, se fermenta el azúcar de fruta y si aumentamos la concentración de oxígeno se degradaría el ácido pirúvico hasta CO 2 liberando energía. Fermentación láctica: En esta fermentación se obtiene ácido láctico, a partir del ácido pirúvico procedente de la glucólisis. De esta manera regenera el NAD+, necesario para continuar la ruta de la glucólisis. Los lactobacilos son bacterias G+ y anaerobias que necesitan moléculas orgánicas para ceder H +. Existen dos grandes grupos de bacterias que realizan fermentación láctica: las homofermentativas (solo producen ácido láctico) y de las que se obtienen leche fermentada, yogur y queso; y las heterofermentativas (que producen otro tipo de sustancias). Las “agujetas” son el resultado de la fermentación de la glucosa en ácido pirúvico y éste en ácido láctico debido a la falta de O2 en el sobreesfuerzo físico. Fermentación butírica Consiste en la degradación de sustancias glucosídicas (almidón y celulosa) en determinados productos como el ácido butírico, el hidrógeno, el dióxido de carbono y otras sustancias malolientes. La realizan Bacillus amilobacer y Clostridium butiricum (descomponenen los restos vegetales del suelo) Fermentación pútrida Se denomina putrefacción, degrada restos animales y vegetales de origen proteínico. Se obitenen, indol, cadaverina, escatol (malolientes) 5.5.4.3. Respiración: ciclo de Krebs, cadena respiratoria y fosforilación oxidativa. En la respiración aerobia de la glucosa hemos llegado a obtener dos moléculas de ácido pirúvico (GLUCOLISIS), pero la oxidación total consume O2 y libera CO2, este proceso EN CONJUNTO se denomina respiración aerobia y consta de estas etapas: 1. GLUCOLISIS (ya vista) 2. DESCARBOXILACIÓN OXIDATIVA DEL ÁCIDO PIRÚVICO 3. CICLO DE KREBS 4. TRANSPORTE DE ELECTRONES DESCARBOXILACIÓN OXIDATIVA DEL ÁCIDO PIRÚVICO Antes de comenzar el ciclo de Krebs, el ácido pirúvico (piruvato) sufre una oxidación. El carbono y los oxígenos del grupo funcional se separan de la molécula, rinden un CO 2 y se forma un grupo acilo CH3-CO. Esta reacción está catalizada por el enzima piruvatodeshidrogenasa. Acoplada a esta reacción se forma un NADH a partir de la reducción de un NAD+; puesto que en la glucólisis se forman dos de ácido pirúvico, en total obtenemos dos de NADH por cada una de glucosa. Cada grupo acilo se une a un nucleótido llamado coenzima A; así se forma el acetilcoenzima A que se incorpora al ciclo de Krebs en la mitocondria. 3. CICLO DE KREBS Se denomina también ciclo de los ácidos tricarboxílicos o ciclo del ácido cítrico. Consiste en una cadena cíclica de reacciones catalizadas por enzimas específicos. Estas reacciones se dan en la matriz mitocondrial. El malato se oxida a oxalacelato con formación de NADH+ 8 Fumarato y agua raccionan dando malato 7 El succinato se oxida a fumarato con la formación de un FADH2 6 El succinil CoA libera la coenzima A y se convierte en succinato; la energía liberada convierte el GDP en GTP, que a su vez, convierte el ADP en ATP 5 BALANCE ENERGÉTICO En cada vuelta del ciclo se genera una molécula de GTP, tres de NADH y una de FADH 2 . Se necesitan dos vueltas del ciclo para oxidar al máximo una de glucosa (se refiere a los carbonos), ya que de cada glucosa se obtienen dos de ácido pirúvico; por lo tanto el balance final será: 2 GTP + 6 NADH + 2 FADH2. 4. TRANSPORTE DE ELECTRONES (RESPIRACIÓN) La molécula de glucosa que inició la glucólisis está completamente oxidada. Parte de la energía se ha usado en la síntesis de ATP; sin embargo, la mayor parte de la energía se encuentra en los electrones que aceptaron la NAD+ y el FAD. Estos electrones están en un estado electrónico más alto del que tenían antes de comenzar la glucólisis. En el transporte electrónico, los electrones son conducidos a través de una cadena formada por aceptores de electrones que los captan a un nivel electrónico ligeramente inferior al que lo precede. El destino de los protones y electrones es el siguiente: Los electrones pasan por una serie de transportadores asociados a la membrana de la mitocondria denominados cadena respiratoria. El flujo de electrones provoca el transporte activo de protones a través de la membrana interna de la mitocondria hacia fuera de la matriz, generando un gradiente de concentración. Los protones difunden nuevamente hacia la matriz mitocondrial a través de un canal de protones que acopla esta difusión con la síntesis de ATP. La cadena respiratoria tiene tres componentes principales: 1. Tres grandes complejos proteicos y sus enzimas asociados 2. Una proteína denominada citocromo c 3. Un componente no proteico denominado ubiquinona (Q) Estos complejos proteicos están asociados a las crestas de la membrana interna de las mitocondrias Los electrones captados por el NADH entran en la cadena cuando son transferidos al FMN (flavin mononucleotido que se reduce) El FMN reducido cede los electrones al CoQ volviéndose a oxidar. La CoQ se reduce. El NADH2 (NADH + H+) pasa los hidrógenos a Q , gracias al primer gran complejo proteico denominado NADH-q reductasa , formando el QH2 Posteriormente el QH2 pasa los hidrógenos al citocromo c, éste los pasa a la citocromo c oxidasa que se los cede en último lugar al oxígeno. El oxígeno reducido toma dos iones hidrógeno y forma agua. La NADH+H+ y el CoQ transportan protones y electrones, el resto de la cadena transporta solo electrones Si no existiese el oxígeno no habría ninguna molécula capaz de captar los electrones, en este caso el proceso se detendría. Complejos de la cadena respiratoria: COMPLEJO I: NADH deshidrogenasa. Recoge un par de electrones del NADH y los cede al CoQ por medio de nucleótido FMN que contiene un núcleo Fe-S. COMPLEJO II: Succinato deshidrogenasa. Incluye el enzima que cataliza la oxidación del ácido succínico. Cede los electrones al CoQ. COMPLEJO III : (Citocromo b-c1). Cataliza el paso de los electrones del CoQ al citocromo b COMPLEJO IV: (Citocromo oxidasa).Está formado por citocromos a y a3 que tienen iones de cobre. Recoge los electrones procedentes del citocromo c y los lleva hasta el oxígeno. La unión de este oxígeno con protones de la matriz da lugar a agua metabólica. FOSFORILACIÓN OXIDATIVA El movimiento de los electrones en la cadena respiratoria hace posible una liberación de energía que se usa en la fosforilación del ADP en el proceso conocido como fosforilación oxidativa. Al proceso global de síntesis de ATP, acoplado al transporte de electrones por la cadena respiratoria, se denomina fosoforilación oxidativa Por cada dos electrones que pasan desde el NADH al oxígeno se fosforilan 3 ADP formando 3 de ATP. Por cada dos electrones que pasan desde el FADH2 se forman dos ATP. Para explicar este fenómeno, Peter Mitchell formuló una hipótesis basada en fenómenos quimioosmóticos, (generación de un gradiente de protones a través de la membrana interna de las mitocondrias) Los tres complejos diferenciados que describimos anteriormente en la cadena de transportadores, actúan como bombas de protones. Cuando los electrones son transportados, los protones son bombeados desde la matriz mitocondrial al espacio intermembranoso. Por cada par de electrones que recorre la cadena, se bombean diez protones hacia el espacio intermembranoso. De esta forma se consigue un gradiente electroquímico entre la matriz y el espacio intermembrana, capaza de generar una fuerza prontomotriz de 230 mV. Existen unos canales denominados partículas F, por donde pueden circular los protones. En realidad cada partícula F es un sistema ATP-sintasa con una porción F0 anclada a la membrana de la cresta y una porción F1 que sobresale a la matriz. F0 y F1 están formadas por subunidades proteicas diferentes. Cuando la diferencia de potencial entre la matriz y el espacio intermembrana es lo suficientemente grande, los protones fluyen por este canal haciendo rotar F0 y provocando la síntesis de una molécula de ATO por cada 3 protones que pasan. 5.5.4.4. Balance energético global del catabolismo de la glucosa Los procesos metabólicos celulares deben ser exergónicos en su conjunto ya que hace falta energía para todos los procesos vitales. Precisamente el balance energético permite conocer la relación entre lo que se obtiene y lo que se consume en cada una de las etapas que hemos estudiado. En el caso concreto de la glucosa el balance total sería: