File - Presentación del curso
Anuncio

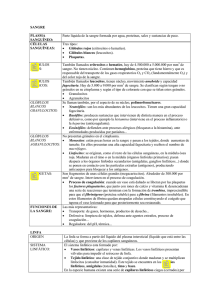
MORFOFISIOLOGÍA DEL SISTEMA CIRCULATORIO M.V.Z. J.Omar Hernández 1.- INTRODUCCIÓN El equilibrio del volumen y la composición de los líquidos corporales que constituyen el medio interno se mantiene por la homeostasis, que W.B.Cannon (1932) definió como “el conjunto de mecanismos reguladores de la estabilidad del medio interno”. Si falla la regulación el equilibrio se altera. Los temas a tratar en este contenido son: Sistema de distribución Sistema de estabilización 2.- SISTEMA DE DISTRIBUCIÓN 2.1 Corazón Desde el exterior el corazón está limitado por el saco pericárdico (pericardio), que protege el corazón y lo separa de otros órganos. El interior del pericardio está recubierto por tejido mesotelial. Hacia el interior se une al pericardio la capa más externa del corazón (epicardio), de textura fina. La fricción entre el pericardio y la capa exterior del corazón se reduce por la estructura en capas de ambas y un líquido lubricante denominado líquido pericárdico. Internamente al epicardio se halla una capa muscular en forma de tubo (miocardio). El diámetro del músculo cardíaco depende de la fuerza que se le exige y la cantidad de sangre que ha de albergar y bombear. Por lo tanto, el espesor del músculo varía según la talla del animal. En cada corazón se puede diferenciar una relación local de la fuerza muscular: La presión sobre la capa muscular es comparativamente baja en las aurículas del corazón. Por lo tanto, el músculo aquí sólo tiene entre 1 a 3 mm de grosor. Como la presión en el ventrículo derecho es superior a las aurículas, el grosor muscular es de 5 a 10 mm. El músculo que ha de soportar mayor presión es el del ventrículo izquierdo y, por lo tanto, el grosor de su pared es de entre 10 y 30 mm, ya que debe bombear la sangre hacia la circulación mayor o sistémica. Continuando en dirección a la cavidad del corazón se encuentra finalmente la capa más interna del corazón (endocardio). Su estructura es fina y está cubierta de tejido conectivo. La capa interior del corazón separa el músculo cardíaco de la cavidad, con lo que mediante una superficie fina lisa el torrente sanguíneo ofrece poca resistencia. La longitud de un corazón canino mediano adulto sano es de unos 12 a 13 cm, con anchura de 9,5 a 10,5 cm. El corazón pesa como 300 gramos y puede albergar alrededor de 0,6 a 1 litro de sangre. El volumen de eyección, es decir, la cantidad de sangre movilizada por contracción muscular en reposo es de unos 70 mililitros. 2.1.1 Ventrículos El corazón se compone de cuatro cavidades. Estas son las siguientes: 1. Dos ventrículos 2. Dos aurículas Desde los ventrículos del corazón se bombea sangre hacia una arteria de la circulación (ventrículo derecho = circulación pulmonar, arteria pulmonar, ventrículo izquierdo = circulación sistémica, arteria aorta). Sin embargo, la sangre que circula por las venas es la que regresa al corazón llegando a las aurículas (circulación pulmonar = venas pulmonares, circulación corporal = venas cava inferior y superior). De las aurículas pasa a los ventrículos para que desde allí, nuevamente pasan al circuito de la circulación correspondiente. El ventrículo izquierdo es el que ejerce mayor fuerza muscular, porque desde aquí la sangre se bombea a la circulación sistémica. Para evitar que la sangre fluya hacia atrás entre dos latidos, hay varios tipos de válvulas cardiacas: un tipo entre las aurículas y los ventrículos, y otro entre los ventrículos y las arterias. Las válvulas del corazón están formadas por pliegues de la capa interna cardíaca (endocardio). A la par que el latido del corazón, se abren y cierran de 35 a 40 millones de veces al año a través del torrente sanguíneo. Las válvulas atrioventriculares se disponen en la dirección del flujo sanguíneo y no le ofrecen ninguna resistencia, sino que son simplemente presionadas y desplazadas hacia la pared. Si inmediatamente tras el bombeo la sangre amenaza con refluir del ventrículo hacia atrás contra el flujo de la sangre, las válvulas atrioventriculares se cierran: se despliegan y presionan entre sí. Las válvulas auriculoventriculares cierran el paso entre las cavidades cardiacas, para que la sangre no fluya en dirección opuesta al flujo. En la pared interior del corazón están los orígenes del músculo (musculatura papilar), de donde salen cuerdas tendinosas, que al cerrarse una válvula evitan que se pliegue hacia la dirección opuesta. Las válvulas semilunares a su vez impiden que la sangre retorne de la arteria pulmonar y la aorta hacia los ventrículos, cuando tras el bombeo se dilatan de nuevo por la relajación muscular. VÁLVULA CARDIACA TIPO DE VÁLVULA FUNCIÓN Válvula tricúspide Válvula auriculoventricular Válvula de entrada entre la aurícula derecha (atrio) y el ventrículo derecho Válvula mitral Válvula auriculoventricular Válvula de entrada entre la aurícula y el ventrículo izquierdos Válvula pulmonar Válvula pulmonar Válvula de salida del ventrículo derecho a la circulación pulmonar Válvula aórtica Válvula aórtica Válvula de salida entre el ventrículo izquierdo y la circulación sistémica 2.2 Circulación de la sangre El corazón bombea la sangre a través del cuerpo. Los órganos, tejidos y células reciben aporte de oxígeno y vierten residuos como dióxido de carbono hacia la sangre para ser eliminados. Por ello la circulación de la sangre se divide en: Circulación menor (circulación pulmonar) Circulación mayor (circulación sistémica) 2.2.1.- Circulación menor La mitad derecha del corazón bombea sangre hacia la circulación menor o circulación pulmonar: el ventrículo derecho, bombea la sangre a través de la válvula pulmonar a la arteria pulmonar, desde allí, circula por las distintas ramificaciones de las arterias y arteriolas hasta los capilares de los pulmones, donde la sangre se enriquece con oxígeno y sigue fluyendo a través de los capilares hacia las vénulas y venas pulmonares hasta la aurícula izquierda. Desde aquí, la sangre entra a través de la válvula mitral en el ventrículo izquierdo. 2.2.2 Circulación mayor La circulación mayor o sistémica recibe impulso de la mitad izquierda del corazón: el ventrículo izquierdo bombea sangre enriquecida con oxígeno a través de la válvula aórtica hasta la arteria aorta. Desde allí continúa por las distintas divisiones arterias y posteriormente en las arteriolas más pequeñas. Finalmente, desemboca en los capilares, los medios de unión entre arterias y venas, que son responsables del intercambio de materiales entre la sangre y los tejidos. Aquí la sangre desprende oxígeno y nutrientes a las células y recoge los materiales de desecho, como el dióxido de carbono. La sangre pobre en oxígeno y rica en dióxido de carbono regresa al corazón: fluye desde los capilares a las venas muy pequeñas, las vénulas, que se unen para formar venas cada vez de mayor diámetro. La sangre fluye por las venas principales (cava inferior y superior) hacia la aurícula derecha y por la válvula tricúspide hacia el ventrículo derecho. 2.3 Fisiología Para mantener el flujo sanguíneo continuo, el corazón debe bombear con regularidad, lo que significa que el músculo cardíaco debe contraerse rítmicamente. Esto requiere que las células musculares sean estimuladas continuamente y que el estímulo se transmita a todas a la vez. 2.3.1 Potencial de acción En las células musculares del corazón, así como en las células musculares del esqueleto, existe una tensión eléctrica entre el interior y el exterior de la célula. 2.3.1.1 Potencial de acción del corazón. En los músculos esqueléticos esta tensión en reposo (potencial en reposo) es de unos -90 mV (mV = milivoltio), y en el corazón de unos -70 mV, con lo que el interior de la célula está cargado negativamente, al contrario que el exterior. Esta tensión se forma especialmente porque la membrana de la célula es permeable a los diferentes iones del cuerpo. Los estímulos mecánicos, químicos o eléctricos pueden cambiar esta permeabilidad. Sin esos estímulos, el voltaje se mantendría permanentemente. Esto es diferente en las células del músculo cardiaco. Para generar en el corazón una tensión de -70 mV, las diferencias de concentración de la membrana celular debe ser las siguientes: Iones de sodio (Na+) interior: exterior = 1 : 14 Iones de potasio (K+) interior: exterior = 35:1 También para los iones de cloro (Cl-) así como algunos otros iones hay diferencias de concentración. En contraste con las células de los músculos esqueléticos, el potencial en descanso de -70 mV en las células de músculo de corazón no es estable. Continuamente entran dentro de la célula pequeñas cantidades de iones, en particular iones de sodio. Esto cambia la tensión gradualmente desde -70 mV dentro de la célula hasta valores de -60 mV. Cuanto más positivo y menos negativo se vuelve el Interior de la célula, más permeable es la membrana celular al sodio (Na+) y por tanto hay más positivo en el interior de la célula. Desde el llamado umbral de unos -50 mV la permeabilidad de la membrana celular cambia muy rápidamente, y puede fluir más Na+. Esto conduce a una inversión de tensión de +30 mV en el interior de la célula. Durante este proceso la permeabilidad de Na+ disminuye de nuevo y aumentan el K+ y el Cl-. Sale K+ y entra Cl- en la célula. Esto ocurre hasta que el estado original está restaurado, es decir el interior de la célula de nuevo tiene una tensión de aproximadamente -70 mV. Entonces el proceso comienza de nuevo. El curso completo del voltaje de -70 mV a +30 mV hasta el anterior estado de -70 mV se denomina potencial de acción. La duración del potencial de acción de una célula muscular, dependiendo de la frecuencia cardíaca, es de hasta varios cientos de milisegundos. La duración de un potencial de acción de una célula del músculo esquelético es mucho más breve: uno o dos milisegundos. El potencial de acción (PA) puede dividirse en cuatro fases: Fase 1: el proceso de cambio de tensión de-70 mV a + 30 mV entre el interior y exterior de las células se denomina despolarización. La despolarización produce un breve valor pico (pico inicial) de alrededor de + 30 mV. Fase 2: tras el pico inicial el valor baja ligeramente a aproximadamente 0 mV, y permanece allí durante un tiempo (fase de meseta). Fase 3: la fase de meseta se une a la repolarización, en la que también se utiliza la bomba de sodio - potasio. Fase 4: la repolarización es seguida por el potencial de reposo. 2.3.2Bomba de sodio - potasio Durante la repolarización del músculo cardiaco el flujo de potasio (K+) del exterior es nuevamente transportado al interior de la célula y sale el sodio (Na+). Este proceso activo está controlado por la membrana celular a través de una proteína transportadora que se denomina bomba de sodio - potasio. En intercambio por la bomba de sodio - potasio es un proceso activo, es decir, un proceso que consume energía, pues los iones deben ser transportados contra su diferencia de concentración. Por cada tres iones de sodio, la bomba transporta dos iones de potasio. 2.3.3 Transmisión de estímulos Para funcionar, el músculo cardíaco cuenta con células musculares modificadas, que originan, coordinan y transmiten el estímulo en el corazón. Estos llamadas marcapasos (nodo sinusal y nodo atrioventricular o nodo AV) se ocupan de que las contracciones transcurran correctamente y en un corazón sano no aparezcan arritmias cardiacas. 2.3.3.1 Nodo sinusal El nodo sinusal (nodus sinuatrialis) está conformado por un conjunto de células despolarizadas, es decir células que pueden cambiar rápidamente su potencial de membrana al polo positivo, y son las que tienen mayor frecuencia propia de todas las células cardíacas. Se encuentra cerca del orificio de entrada desde la vena cava superior a la aurícula derecha. El nodo sinusal también es conocido como el marcapasos natural del corazón, porque impone su frecuencia a todo el músculo cardíaco (miocardio). La frecuencia propia del nodo sinusal suele ser de 70 a 80 “estímulos” por minuto, correspondiente al ritmo normal del corazón, el pulso. Para ajustar el latido a la carga física específica, la frecuencia del nodo sinusal puede ajustarse en respuesta a los estímulos de dos nervios (el vago y el simpático). Si aumenta la influencia del nervio vago más de lo normal ocurre la llamada bradicardia. Se trata de una alteración del ritmo cardíaco de 40 a 50 potenciales de acción y, por tanto, latidos por minuto. Si prevalece el sistema nervioso simpático, se provocará lo que se llama taquicardia sinusal; esta arritmia suele presentar de 100 a 150 latidos por minuto. La frecuencia del nodo sinusal puede ser afectada por medicamentos o por la hormona adrenalina, además de los estímulos facilitados por el sistema nervioso. 2.3.3.2 Nodo atrioventricular (nodo AV) Desde el nodo sinusal el estímulo eléctrico se extiende por tres conductos al músculo auricular hasta el llamado nodo atrioventricular (nodo AV). El nodo AV tiene menor ritmo propio que el nodo sinusal (aproximadamente 40 a 50 despolarizaciones o descargas por minuto). Normalmente los estímulos procedentes del nodo sinusal despolarizan los nodos AV y les imponen el ritmo sinusal. Si se perturba la transición del seno al nodo AV (llamado bloque AV), el corazón sigue el ritmo propio del nodo AV de 40 a 50 potenciales de acción por minuto. Esto es suficiente para el suministro de sangre al cuerpo en reposo. El ritmo intrínseco del nodo AV es de 40 a 50 potenciales de acción por minuto, lo que representa un suministro meramente suficiente de sangre al cuerpo en reposo. 2.3.3.3 Haz de His Un haz de His consiste en células musculares especializadas del sistema de conducción del estímulo inmediatamente debajo del nodo AV. También tienen un ritmo propio que es aún menor que el del nodo AV (aproximadamente 20 a 30 de polarizaciones por minuto). A unos cm de distancia del nodo AV hacia la punta del corazón (distal) el haz de His se divide en tres ramas: dos a la izquierda y una a la derecha. Estas, a su vez, se dividen en forma de red en las llamadas fibras de Purkinje. El haz de His emerge del nodo AV y se dirige desde aquí el estímulo hasta la capa interior de la musculatura cardiaca. Estas tres ramas van hasta los músculos papilares, que son estimulados y se contraen en primer lugar. Esto asegura que las válvulas auriculares se cierren herméticamente al comienzo de un latido del corazón y que no regrese sangre a las aurículas. 2.3.4 Transmisión defectuosa de un estímulo Cuanto más se aleja desde el nodo sinusal en la dirección de propagación del estímulo, más bajo es el ritmo propio de las células correspondientes. Cada célula individual de los músculos de trabajo del corazón (fibras miocárdicas) tiene un ritmo propio; este es tan lento que apenas tiene importancia para la función normal del corazón. Todo trastorno del músculo cardíaco debido a un estrechamiento de las arterias coronarias o incluso un cierre completo conduce a cambios más o menos fuertes en la propagación del estímulo eléctrico y, por lo tanto, a trastornos de contracción mecánica. En el caso más extremo, no tiene lugar la propagación de estímulos regulares. Entonces tienen lugar innumerables contracciones individuales del músculo cardíaco, se produce, entonces, lo que se denomina fibrilación auricular o fibrilación ventricular. En este último caso, el corazón no bombea sangre. Después de unos minutos aparecen daños irreversibles y pronto sigue la muerte. Entre otras cosas, la fibrilación ventricular puede causar un infarto de corazón. En este caso el corazón debe recuperar el ritmo y la despolarización adecuados utilizando una tensión creada desde el exterior por dos electrodos metálicos (> 1000 voltios). Los dispositivos que lo hacen posible se llaman desfibriladores. 2.4 VASOS SANGUINEOS Anatomía de vasos sanguíneos Los vasos sanguíneos funcionan como conductos por los cuales pasa la sangre bombeada a través de corazón. Los vasos sanguíneos pueden clasificarse en: 2.4.1 Arterias Por definición son aquellos vasos sanguíneos que salen del corazón y llevan la sangre a los distintos órganos del cuerpo. Todas las arterias excepto la pulmonar y sus ramificaciones llevan sangre oxigenada. Las arterias pequeñas se conocen como arteriolas que vuelven a ramificarse en capilares y estos al unirse nuevamente forman las venas. Las paredes de las arterias son muy elásticas y están formadas por tres capas. Sus paredes se expanden cuando el corazón bombea la sangre, de allí que se origine la medida de la presión arterial como medio de diagnóstico. Las arterias, contrario a las venas, se localizan profundamente a lo largo de los huesos o debajo de los músculos. Las arterias principales son la aorta y el tronco pulmonar. La aorta es un vaso sanguíneo grueso que sale del ventrículo izquierdo en forma de arco, del cual se originan las arterias que van al cuello, cabeza y miembros. La aorta desciende a lo largo de la columna vertebral por la cavidad torácica y abdomen, terminando en las dos arterias ilíacas que van a los miembros posteriores. Al pasar por cada cavidad del cuerpo se subdivide para suplir distintos órganos. El tronco pulmonar es la única arteria que lleva sangre venosa (con poco oxígeno), sale del ventrículo derecho hasta ingresar a los pulmones como arterias pulmonares para capilarizarse y sufrir el intercambio gaseoso. Durante la contracción ventricular, se genera una presión sanguínea relativamente alta; sin embargo, también es necesario que se conserve en el periodo que hay entre una contracción y otra. Por tal razón, las paredes de los grandes vasos que nacen en los ventrículos consisten principalmente en fibras elásticas. Estos vasos se conocen como arterias elásticas y la sangre que llega a ellos desde los ventrículos en contracción, estira la elastina de sus paredes. Después de la contracción ventricular, el rebote elástico de las paredes arteriales mantiene la presión sanguínea en la siguiente contracción ventricular. A diferencia de las arterias de mayor calibre, cuya función principal es conservar la presión sanguínea diastólica, la de las arterias que nacen de aquellas es distribuir la sangre a las diversas partes del cuerpo. Estas últimas requieren volúmenes desiguales de sangre, según su actividad, de modo que el diámetro de la luz de las arterias que se distribuyen en dichas partes debe regularse de manera continua para garantizar que llegue el volumen apropiado de sangre a ellas, en cualquier momento dado. Las paredes de estos vasos consisten principalmente en células de músculo liso dispuestas en las llamadas capas circulares, que en realidad tienen forma helicoidal. Dada la importancia de su componente de músculo liso, estos vasos también se conocen como arterias musculares. Las arteriolas distribuyen la sangre arterial en los lechos capilares, con una presión hidrostática relativamente baja. Como su nombre lo indica, en lo esencial son arterias de muy poco calibre, si bien tienen una luz relativamente angosta y paredes musculares gruesas. Las paredes de los vasos sanguíneos se ajustan a un plan generalizado, según el cual consiste en 3 capas o túnicas. Sin embargo estas capas no siempre presentan una definición tan clara como lo indica su descripción se denomina: a) Capa o túnica íntima, que es la más interna (endotelio vascular) b) Capa o túnica media, la intermedia. (musculo liso) c) Capa o túnica adventicia que es la externa. (tejido conectivo) La mayor parte de las arterias van acompañadas por un vaso linfático, nervio y una o dos venas formando una relación vasculonerviosa cubierta por tejido conectivo. En algunas animales y humanos las paredes arteriales se endurecen perdiendo y dando lugar a la condición de arteriosclerosis o ateroesclerosis. 2.4.2Venas Son vasos sanguíneos mayores que las arterias y que corren superficialmente a la fascia (Tejido conectivo que recubre a los músculos) como venas superficiales y acompañan a las arterias (dos por cada arteria) como venas profundas, Su circulación se debe a la presión de la sangre que efluye de los capilares, a la contracción de los músculos y del cierre de las válvulas. Forman dos sistemas de vasos, los de la circulación pulmonar y los de la circulación general. Las venas pulmonares llevan sangre oxigenada de los pulmones al atrio izquierdo. Comienza en los alvéolos hasta formarse en tres troncos venosos del pulmón derecho y dos para el izquierdo; uniéndose luego el lóbulo superior del pulmón derecho con el que sale del lóbulo medio para formar cuatro venas pulmonares: dos para cada pulmón. Las venas de la circulación general traen sangre de todas las regiones del cuerpo al atrio derecho del corazón. Incluyen las venas que se vacían en el corazón, las que van a la vena cava superior y a la vena cava inferior. Las venas están provistas de válvulas consistentes en valvas dispuestas a modo de permitir el flujo de la sangre hacia el corazón, pero no en la dirección opuesta. Tales valvas son pliegues de la túnica íntima con refuerzos centrales de tejido conectivo y también hay fibras elásticas en la cara de las valvas que miran hacia la luz del vaso. Las válvulas venosas tienen varias funciones: En primer término sirven para contrarrestar la fuerza de la gravedad al evitar el flujo retrógrado, pero también ejerce otros efectos por ejemplo las válvulas permiten el "ordeño" de las venas cuando los músculos que circundan a éstas se contraen y hacen las veces de dos bombas. Aún más, evitan que la fuerza de la contracción muscular origine una presión retrógrada en los lechos capilares drenados por las venas. La sangre venosa es de un color rojo oscuro. Contiene bióxido de carbono (CO2) y menos oxígeno que la arterial. 2.4.3 Capilares Son vasos sanguíneos que surgen como pequeñas ramificaciones de las arterias a lo largo de todo el cuerpo y cerca de la superficie de la piel. Llevan nutrientes y oxígeno a la célula y traen de ésta los productos de deshecho. Al reunirse nuevamente forman vasos más gruesos conocidos como vénulas que al unirse luego forman las venas. 2.5 VASOS LINFATICOS Anatomía de sistema linfático El sistema linfático se compone de un líquido circulante: la linfa, un sistema de vasos para recoger y transportar la linfa, y diversos órganos linfoides incluyendo los linfonodos, el bazo, bolsa cloacal, timo y las tonsilas. La función que este sistema desempeña se relaciona con el mantenimiento del equilibrio hídrico y proteico de los tejidos, y con la protección contra la invasión de substancias químicas y microorganismos extraños. 2.5.1 Linfa El líquido que circula por este sistema, la linfa o líquido tisular, se deriva del líquido intersticial. Una vez que penetra a los vasos linfáticos, recibe el nombre de linfa. Características y composición. La linfa tiene un peso específico que varía entre 1.018 y 1.024. Tiene una baja concentración de proteína, carece de eritrocitos y de trombocitos, y contiene poco oxígeno. En cuanto a substancias inorgánicas, su composición es similar a la del plasma, y en cuanto a las moléculas orgánicas que no sean proteínas ésta varía según la localización de la muestra de linfa que se analiza. La linfa que se extrae de los linfáticos del tubo digestivo tiene más glucosa y líquidos. En la linfa se encuentra también un número variable de leucocitos, que aumenta a medida que la linfa drena al sistema vascular sanguíneo. Formación y funciones. La linfa se forma a partir del plasma, por medio de un proceso de filtración. Ese proceso explica el bajo contenido proteico y la carencia de células del líquido. Como la presión de filtración en los capilares es mayor que la presión osmótica que regresa el líquido a los capilares, con una diferencia de aproximadamente 7 mm de Hg (32 y 25 mm de Hg.), los espacios tisulares tienen tendencia a acumular un exceso de líquido. El sistema linfático recoge este líquido y lo devuelve al sistema vascular, lo que ayuda a mantener concentraciones normales de líquidos en los tejidos. También hay que devolver al plasma las pequeñas cantidades de proteína que logran pasar a través de los capilares. Además, microorganismos, células muertas o substancias químicas extrañas pueden salir de los tejidos con el líquido y se eliminan o depuran en los linfonodos. 2.5.2. Vasos linfáticos El sistema de los vasos linfáticos empieza dentro de los tejidos en la forma de tubos cerrados en un extremo que reciben el nombre de capilares linfáticos (quilíferos en el intestino). Estos tienen un recubrimiento endotelial interno, similar al de los capilares sanguíneos. Dichos vasos se encuentran en mayor número en la piel, el tubo digestivo y el aparato respiratorio. El líquido tisular penetra por difusión a los capilares linfáticos, con ayuda de un pequeño gradiente de presión indirecto, que le confiere la presión arterial. Las paredes de los capilares linfáticos ofrecen poca resistencia a la entrada de substancias; son más permeables que los capilares del sistema sanguíneo. Los diminutos capilares forman vasos más grandes, similares a las vénulas y venas del sistema vascular sanguíneo. Las paredes son más delgadas que las de las venas del sistema sanguíneo y sus válvulas mucho más numerosas. Los vasos linfáticos más grandes convergen hasta terminar en los dos grandes conductos linfáticos. El conducto torácico corre a lo largo de la columna vertebral y se extiende desde la segunda vértebra lumbar hasta terminar en la vena subclavia izquierda. Su extremo inferior se ensancha para formar la cisterna del quilo. El conducto torácico recoge la linfa del lado izquierdo de la cabeza y del tórax, del miembro izquierdo y de toda la parte del cuerpo situada abajo de las costillas. El conducto linfático derecho drena la linfa de las partes restantes del cuerpo y vierte en la vena subclavia derecha. 2.5.3 Factores que intervienen en el flujo de la linfa El factor principal para hacer pasar la linfa por los vasos es un gradiente de presión que existe entre los tejidos y la vena subclavia. En los tejidos, la presión hidrostática es de aproximadamente 25 mm de Hg; en la vena subclavia, la presión puede ser nula o negativa. Además, las contracciones de los músculos esqueléticos ejercen acción de masaje en la linfa a través de los vasos, y las válvulas impiden el reflujo. Los movimientos respiratorios también aspiran linfa hacia la parte superior de los conductos 2.5.4 Órganos linfáticos Los órganos relacionados con la linfa son áreas de producción de leucocitos linfoides y de macrófagos de substancias determinadas que se encuentran en la linfa o en la sangre. Los linfonodos están situados a lo largo de los vasos linfáticos y por lo tanto, sirven para filtrar la linfa, cuando pasa por ellos. El bazo, situado en el trayecto de los vasos sanguíneos, desempeña funciones tanto linfáticas como sanguíneas. El timo, bolsa cloacal y las tonsilas no están situadas a lo largo de vaso alguno, y su función se limita a producir linfocitos linfoides y substancias químicas indispensables para el desarrollo de una capacidad inmunológica completa. Los nódulos son sencillamente conjuntos de células linfoides sin estructura organizada. 2.5.4.1 Linfonodos El tamaño de los linfonodos es variable, con un largo que va de 1 a 2 mm hasta 25 mm aprox. en perros de talla mediana. Están situados a lo largo del trayecto de los vasos linfáticos, y tienen tanto vasos linfáticos aferentes (que llegan a ellos) como eferentes (que salen de ellos). Las células del linfonodos incluyen linfocitos, monocitos, células plasmaticas y células macrófagos fijas. El linfonodo produce leucocitos linfoides, globulinas inmunes, y depura la linfa al eliminar las partículas de substancias. El linfonodo también capta células y se puede llegar a inflamar al desarrollarse una infección, o puede ser el sitio donde se alojen las células cancerosas. Los linfoncitos están concentrados en varias regiones del cuerpo. Las principales regiones son las que contienen los grupos axilares, inguinales cervicales, lumbares y sacros. 2.5.4.2 Bazo El bazo es un órgano de forma oval de unos 20 cm de largo en perros medianos, situado a la izquierda y ligeramente detrás del estómago Presenta una depresión, el hilio, que marca el punto de entrada y de salida de los vasos sanguíneos y de los linfáticos eferentes del bazo. Este órgano está rodeado por una cápsula elástica que contiene músculo liso y de la cual nacen las trabéculas que penetran en él. La parte interna del bazo consta de dos variedades de pulpa esplénica: l. La pulpa blanca, que rodea las arterias centrales del órgano y se compone de masas de linfocitos. 2. La pulpa roja, que ocupa el resto del órgano y consta principalmente de sangre que está de paso por el bazo o que se almacena dentro de éste. Desde el punto de vista funcional, el bazo sirve como sitio de producción de leucocitos linfoides, como área de fagocitosis de eritrocitos viejos, como reservorio de sangre y como lugar de formación de sangre fetal. 2.5.4.3 Timo El timo es una estructura formada por dos lóbulos y situada detrás del esternón. Este órgano está rodeado por una cápsula colágena que se extiende hacia el interior, formando tabiques que lo subdividen en lóbulos más pequeños. La corteza, rica en células linfoides, rodea la médula, en la que éstas son menos abundantes. Dentro de la médula se encuentran los corpúsculos tímicos, cuya función se desconoce. El timo produce células linfoides y substancias químicas que garantizan el desarrollo de la capacidad inmunológica en las células plasmáticas. El tamaño de este órgano aumenta hasta la pubertad, y luego sufre involución y substitución por grasa y tejido conectivo. 2.5.4.4 Tonsilas En el cuerpo se encuentran tres pares de tonsilas. Las tonsilas palatinas están situadas a ambos lados de la cavidad oral, a la altura del paladar blando; las tonsilas faríngeas (adenoides) están situadas en la parte superior de la garganta (nasofaringe), y las tonsilas linguales, en la base de la lengua. La estructura de todas ellas es similar . Abajo del órgano se encuentra una cápsula parcial de tejido colágeno. La única función que corresponde a las tonsilas es la elaboración de linfocitos. 2.5.4.5. Tejido linfoide asociado a mucosas En las paredes de los sistemas digestivo y respiratorio se encuentran masas de linfocitos sin cápsula, que se denominan nódulos linfáticos. Los nódulos del íleon reciben el nombre especial de placas de Peyer. Los folículos proporcionan una fuente adicional de células linfoides y constituyen una barrera de defensa contra los microorganismos que pueden entrar al cuerpo a través de las paredes de estos sistemas. 2.5.4.6. Bolsa cloacal La bolsa de Fabricio es un órgano pequeño redondeado, que se encuentra situado dorsalmente a la parte caudal de la cloaca de las aves, incluida en el tejido conjuntivo que rodea a la misma. Los linfocitos de la bolsa provienen de las células pluripotenciales: Linfoblasto de la médula ósea roja, llegan a la bolsa en el caso de las aves y es aquí donde adquieren la capacidad inmunológica de convertirse bajo el estímulo antigénico en células plasmáticas, que son las que sintetizan los anticuerpos específicos para neutralizar al antígeno que provocó su formación (inmunidad de tipo humoral), estas células plasmáticas se van a localizar en los tejidos conjuntivos, y permanece activa por 72 horas, después muere por apoptosis muerte celular programada). Los linfocitos B, tienen tres destinos: activarse bajo la acción de un antígeno, muerte celular programada o quedarse como célula de memoria. En los mamíferos no existe un órgano especializado para la producción o diferenciación de los linfocitos B, a tal efecto los inmunólogos le confieren ese rol a los nódulos linfáticos de las vías digestivas, en el bazo y en los nódulos linfáticos periféricos. Función: Los linfocitos B en las aves, se maduran y capacitan en la bolsa de Fabricio y tienen la propiedad de convertirse en células plasmáticas y producir anticuerpo específico contra el antígeno que provocó su activación. 3 SISTEMA DE ESTABILIZACIÓN Anatomía de sistema vascular Los organismos unicelulares o muy simples que viven en un medio líquido, utilizan éste como fuente de nutrientes y como vía de excreción de los desechos. Al aumentar el número de células es inevitable que las células que se ubican más profundan se alejen cada vez más del contacto con el medio ambiente. Proporcionar nutrientes a estas células y eliminar sus deshechos se vuelve una tarea cada vez más difícil. En animales primitivos estos problemas se solucionaron parcialmente por medio de conductos que pasan entre las células y que comunican con el medio ambiente. El pulso de la medusa hace que el líquido se retire de los canales y fluya. Al aumentar una demanda de nutrientes un sistema tipo abierto de flujo y reflujo se vuelve insuficiente. El líquido debe circular con más rapidez y seguir la dirección de su flujo. Pueden llenarse las necesidades al aislar los vasos o conductos del contacto directo del ambiente y proporcionar una bomba para hacer circular el líquido que contiene. Los tres componentes que han evolucionado son: Un líquido para circular, una bomba para provocar su circulación y vasos para contenerlo. El sistema circulatorio humano es del tipo cerrado y tiene dos subdivisiones: el sistema vascular sanguíneo, incluyendo sangre, corazón, vasos sanguíneos y el sistema vascular linfático, que comprende la linfa, los vasos linfáticos y los órganos linfáticos. 3.1 Sangre La sangre se considera como un tejido conectivo porque se origina de células similares. La sangre representa cerca del 8% del peso corporal total peso de un individuo, y tiene un volumen de cinco a seis litros en un canino de raza grande. Sus principales funciones se centran en su capacidad de disolver sustancias o tenerlas en suspensión y, por lo tanto, transportarlas por todo el cuerpo. Comprende los elementos figurados: glóbulos rojos, glóbulos blancos, plaquetas y una parte líquida sin células, el plasma. El plasma es el líquido, tiene una coloración amarilla clara, puede variar; se forma de agua, sales minerales, glucosa, proteínas (como albúminas y globulinas), algunos lípidos como el colesterol, algunas hormonas principalmente. Las funciones principales de la sangre son: 1.- Transporta a las células, elementos nutritivos, oxígeno, y extrae de los mismos productos de desecho. 2.- Transporta hormonas y se encarga del equilibro hídrico. 3.- Interviene en el equilibrio de ácidos, bases, sales (Equilibrio acido-base) 4.- Toma parte importante en la regulación de la temperatura del cuerpo, al enfriar los órganos como el hígado y músculos, donde se produce exceso de calor, cuya pérdida del mismo es considerable, y calentar la piel. 5.- Transporte de células cuando existe proceso activado de tipo inflamatorio, además de ser imprescindible desde el punto de vista inmunológico. 6.- Importante en la hemostasisque es el proceso en donde se utiliza la cascada de coagulación para evitar la pérdida de sangre del lecho vascular. 3.2. Plasma Aunque la sangre aparece como un líquido rojo, homogéneo, al fluir de una herida, se compone en realidad de un líquido amarillento llamado plasma en el cual flotan los elementos: glóbulos rojos, los cuales dan su color a la sangre, glóbulos blancos y plaquetas. Estas últimas son pequeños fragmentos celulares, convenientes para desencadenar el proceso de coagulación, los cuales derivan las células de mayor tamaño de la médula ósea. El plasma es una mezcla compleja de proteínas, aminoácidos, hidratos de carbono, lípidos, sales, hormonas, enzimas, anticuerpos y gases en disolución. Es ligeramente alcalino, con un Ph de 7.4. Los principales componentes son el agua (del 90 al 92 por ciento) y las proteínas (7 al 8 por ciento).El plasma contiene varias clases de proteínas, cada una con sus funciones y propiedades específicas: fibrinógeno, globulinas alfa, beta y gama, albúminas y lipoproteínas. El fibrinógeno es una de las proteínas destiladas al proceso de coagulación; la albúmina y las globulinas regulan el contenido de agua dentro de la célula y en los líquidos intercelulares. La fracción gamma globulina es rica en anticuerpos, base de la comunidad contra determinadas enfermedades infecciosas. La presencia de dichas proteínas hace que la sangre sea unas seis veces más viscosa que el agua. Las moléculas de las proteínas plasmáticas ejercen presión osmótica, con lo que son parte importante en la distribución del agua entre el plasma y los líquidos tisulares. Las proteínas del plasma y la hemoglobina de los glóbulos rojos son importantes amortiguadores acidobásicos que mantienen el ph de la sangre y de las células corporales dentro de una pequeña variación. 3.3. Elementos figurados Las células hemáticas representan una categoría de células libres del tejido conectivo, son producidas por los tejidos hemopoyéticos y al entrar al torrente sanguíneo quedan suspendidas en el plasma sanguíneo. 3.4. Flujo laminar En condiciones fisiológicas el tipo de flujo mayoritario es el denominado flujo en capas o laminar. El fluido se desplaza en láminas coaxiales o cilíndricas en las que todas las partículas se mueven sin excepción paralelamente al eje vascular. Se origina un perfil parabólico de velocidades con un valor máximo en el eje o centro geométrico del tubo 3.5 Eritrocitos Los eritrocitos, glóbulos rojos o hematíes constituyen el tipo más común de células hemáticas, existen unos 5 millones de eritrocitos por cm3 de sangre. El diámetro de los eritrocitos tiene un valor promedio de 7.2 a 8 µm, tienen tiene forma de un disco bicóncavo, excepto a los de ave que son ovales y con núcleo. Los constituyentes moleculares particulares de su membrana celular y su contenido coloidal, son los que determinan y conservan la forma característica del eritrocito. Alrededor del 38% del eritrocito es una proteína, la hemoglobina, y un 66% de agua. Sintetizan la proteína de intercambio de gases, hemoglobina. Los eritrocitos son fagocitados en hígado, bazo y médula ósea, con el reciclamiento de sus productos. La hemoglobina es el pigmento rojo que da el color en la sangre, cuya misión exclusiva es transportar casi todo el oxígeno y la mayor parte del bióxido de carbono. La hemoglobina tiene la notable propiedad de formar una unión química poco estrecha con el oxígeno; los átomos de oxígeno están unidos a los átomos de hierro en la molécula de la hemoglobina. En el órgano respiratorio, pulmón, el oxígeno se difunde hacía en interior de los glóbulos rojos desde el plasma, y se combina con la hemoglobina (Hb) para formar oxihemoglobina (HbO2): Hb + O2 = HbO2. La reacción es reversible y la hemoglobina libera el oxígeno cuando llega a una región donde la tensión oxígeno es baja, en los capilares de los tejidos. La combinación de oxígeno con la hemoglobina y su liberación de oxihemoglobina están controladas por la concentración de oxígeno y en menor grado por la concentración de bióxido de carbono. La hemoglobina se compone de un pigmento, el hem, combinado de una proteína la globina. Entre las anomalías en la formación de eritrocitos se encuentran los procesos que tienen por resultado una forma anormal de las células o poiquilocitosis o un tamaño de las células fuera de lo común o anisocitosis. La deficiencia de hemoglobina por pérdida de eritrocitos se denomina anemia. 3.6 Plaquetas Las plaquetas sanguíneas o trombocitos, son fragmentos de citoplasma granulado relativamente pequeñas, tienen forma de disco y un diámetro de 2 a 3 µm. Se desprenden de unas células grandes llamadas megacariocitos. Las plaquetas no poseen núcleo, mantienen un período de vida corto debido a restos de RNA mensajero derivados del magacariocito. El número de plaquetas en la sangre varía entre 150,000 a 400,000 por milímetro cúbico. La causa principal para que cese el sangrado es la adhesión de las plaquetas a la superficie interna de la pared del vaso en la vecindad del corte. Se las observa como discos aislados biconvexos y ovales. La parte externa de las plaquetas se tiñe de un color azul pálido. La forma se mantiene por la presencia de microtubulos ordenados en una circunferencia. Si no participan en la hemostasis, las plaquetas viven entre ocho a diez días, luego son fagocitadas por los macrófagos. 3.7 Leucocitos Son células con gran movilidad que realizan sus funciones más importantes fuera del torrente sanguíneo. Su número es de 5,000 a 9,000 por milímetro cúbico de sangre. Se los clasifica en cinco tipos distintos según sus características de tinción específicas y su morfología celular y funciones específicas. Al microscopio de luz pueden dividirse en: Leucocitos granulares Leucocitos no granulares 3.7.1. Leucocitos granulares + -14 m A.- Neutrófilos Su cantidad es del 50% al 70% de los leucocitos. Su número absoluto se considera entre 3,000 a 6,000 por milímetro cúbico de sangre. Su función más importante es actuar en las inflamaciones primordialmente agudas. B.- Eosinófilos Se encuentran entre el 1% y el 4% de las células de sangre periférica. Su número absoluto es de 120 a 350 por milímetro cúbico de sangre. Tiene una función reguladora en las alergias. Un número elevado de eosinófilos en la sangre periférica puede ser un indicador de cuadros de parasitosis. C.- Basófilos Constituyen solo el 0.5% de los leucocitos de la sangre periférica. Su número llega a 40 por milímetro cúbico de sangre. Pueden acumularse en zonas donde se producen reacciones alérgicas. 3.7.2. Leucocitos no granulares A.- Linfocitos+- 8 -12 m Los linfocitos comprenden entre el 20% y el 50% de los leucocitos sanguíneos. El número total es de 1,500 a 4,000 por milímetro cúbico. Los linfocitos pequeños se clasifican en dos grupos: los linfocitos T y los linfocitos B B.- Monocitos+- 20 m Los monocitos comprenden de 2% al 8% de los leucocitos sanguíneos. El número absoluto son de 200 a 30 por milímetro cúbico de sangre. Los monocitos sirven como precursores de los macrófagos. Tiene una vida media de tres días, para luego migrar fuera del torrente sanguíneo. 3.8. VOLUMEN Y CORPORALES DISTRIBUCIÓN NORMAL DE LOS LÍQUIDOS En el individuo adulto, el agua corporal total (ACT) se estima en un 60 % del peso corporal magro, Estos valores varían en función de la especie, edad, sexo y hábito corporal. Así, éste valor puede ser mucho menor en un individuo obeso, alrededor del 50% del peso corporal, ya que el tejido adiposo contiene poca agua. LÍQUIDOS CORPORALES BASE 100 KILOS DE PESO VIVO Volumen extracelular ( 15 litros ) Volumen plasmático ( 3 litros ) Volumen de hematíes ( 2 litros ) Volumen intracelular Liquido intersticial ( 25 litros ) (15 litros) El ACT se distribuye en 2 compartimentos principales: 1-El Agua Intracelular (AIC) que corresponde a dos tercios del ACT, unos 25 litros aproximadamente. 2-El Agua Extracelular (AEC) que representa el tercio restante y que se distribuye entre los compartimentos intersticial, plasmático y transcelular, constituyendo los 15 litros de agua restante. Este volumen de líquido transcelular, estimado en un 2,5 % del ACT, incluye los fluidos formados por glándulas (glándulas salivares, páncreas ) así como los líquidos del líquido cefalorraquídeo, árbol traqueo bronquial, tracto gastrointestinal, sistema genitourinario y ojos (humor acuoso) . Además, hemos de asumir que 1/4 del AEC se encuentra en el espacio vascular, mientras que los 3/4 restantes ocupan el espacio intersticial. 3.8.1. Composición ionica de los líquidos del organismo La composición de los dos compartimentos principales, extracelulares e intracelulares, difieren en forma significativa. Además, ningún compartimento es completamente homogéneo, y también varían los diversos tipos celulares que los componen. Por supuesto, la amplia diferencia en la composición de los compartimentos intracelular y extracelular es el resultado de barreras de permeabilidad y mecanismos de transporte, tanto activos como pasivos, que existen en las membranas celulares. Dentro de los factores que determinan el movimiento entre los distintos compartimentos, la ósmosis es el principal factor que determina la distribución de los líquidos en el organismo. La osmolaridad de todos los fluidos orgánicos es el resultado de la suma de electrolitos y no electrolitos presentes en un compartimento. Un organismo fisiológicamente estable mantiene una presión osmótica casi constante y uniforme en todos los compartimentos. Cuando se producen cambios de concentración de solutos confinados preferentemente en un compartimento, se trata de restablecer el equilibrio osmótico mediante la redistribución del disolvente, el agua. Por lo tanto, un cambio en un compartimento como el vascular tiene repercusión en el intracelular. En la práctica médica diaria, el compartimento vascular es el más fácilmente accesible a la exploración y modificación según las necesidades. 3.8.2. Composición del líquido extracelular Como ya hemos comentado, la composición del líquido extracelular es muy distinta a la del líquido intracelular. En cambio, la composición de los diferentes espacios en que se divide el líquido extracelular es muy parecida. En el suero, el sodio (Na+) es el catión predominante y alcanza una concentración media de 142 mEq/L (normal:136-145 mEq/L). Las concentraciones de otros cationes como el potasio (K+) , el calcio (Ca++) y el magnesio (Mg++) son mucho menores. El K+ tiene una concentración media de 4 mEq/L (normal: 3,5-5,0 mEq/L), el Ca++ de 5 mEq/L (normal: 3,5-5,5 mEq/L) y el Mg++ de 2 mEq/L (normal: 1,5-2,5 mEq/L). Los iones del hidrógeno (H+) se hallan a una concentración muy baja (4 x 10-5 mEq/L), pero ésta es crítica, ya que de ella depende el pH del medio (pH de 7.4). El anión predominante en el suero es el cloro (Cl-) cuya concentración es de alrededor de 103 mEq/L (normal: 96-106 mEq/L), seguido del ión bicarbonato (COH3) de 26 mEq/L (normal: 24-27 mEq/L) y de las proteínas de aproximadamente 6-8 gr/dl. En cantidades menores, se hallan los iones sulfato (SO-), fosfatos (HPO4 y H2PO4-) y diversos ácidos orgánicos. Entre estos últimos, figuran los ácidos lácticos, pirúvico, cítrico y otros procedentes del metabolismo de los hidratos de carbono, de los lípidos, así como de diferentes aminoácidos. En condiciones normales, la concentración de los ácidos orgánicos es muy baja, inferior a 1 mEq/L, excepto para el ácido láctico. La composición iónica del líquido intersticial es muy parecida a la del suero, pero no idéntica. Las proteínas, debido a su elevado peso molecular, apenas difunden al líquido intersticial y su concentración en este medio es inferior a 2 gr/dL. 3.8.3. Composición del líquido intracelular A diferencia del medio extracelular, en el interior de la célula el catión principal es el potasio (156 mEq/L), seguido del magnesio (26 mEq/L), mientras que la concentración de sodio es muy baja (10 mEq/L). En relación con los aniones intracelulares, las mayores concentraciones corresponden a los iones del fosfato (95 mEq/L), seguidos de las proteínas (16 gr/dl) y los sulfatos (20 mEq/L). Las concentraciones de cloro y bicarbonato son muy pequeñas. DISTRIBUCIÓN DE LÍQUIDOS EN EL ORGANISMO Líquido intravascular 1º espacio Líquido intersticial (extracelular) 1º espacio Líquido intracelular 2º espacio Líquido en cavidades naturales pleura peritoneo luz intestinal 3º espacio (cavidades) Líquido que ha salido al exterior del organismo 4º espacio (exterior) 3.8.4 Movimiento del agua entre los compartimentos El agua y los solutos disueltos fluyen entre los compartimentos corporales por difusión, convección o por mecanismos de transporte específicos. Las fuerzas que gobiernan estos intercambios son, principalmente, las presiones hidrostática y osmótica y para, algunos solutos que atraviesan las membranas celulares, las bombas transportadoras. Además, el organismo intercambia a diario con el medio exterior una cantidad de agua y solutos. La fuerza capaz de provocar el paso de agua por una membrana semipermeable debido a las diferencias en la concentración de los solutos a ambos lados de ésta, constituye la presión osmótica. La presión osmótica depende exclusivamente del número de partículas disueltas (moles) por unidad de volumen, con independencia de su carga eléctrica, peso o fórmula química. El número total de partículas disueltas constituye la osmolaridad, si su concentración se expresa por unidad de volumen total de la solución (moles/L de suero), o en términos de osmolalidad, si se expresa por unidad de volumen sólo el disolvente (moles/kg H2O). De hecho, la actividad osmótica depende de la osmolalidad, pero en la práctica, y debido a que las soluciones biológicas son muy poco concentradas, la diferencia entre ambos valores es pequeña y ambos términos se utilizan a menudo de forma indistinta. En ausencia de osmómetro, la concentración total de solutos del suero puede calcularse con una fórmula sencilla a partir de las concentraciones de sodio, cloro, glucosa y urea (los principales solutos del líquido extracelular). El siguiente cálculo emplea un sodio plasmático de 140 mEq/L, una glucemia de 90 mg/dL y un BUN* de 14 mg/dL. [ glucosa ] [ BUN ] [ urea ] Osmolalidad Plasmática = 2 x [Na+] +---------- + --------- + -------18 2,8 6 90 14 = 2 x ( 140 ) + -------- + ------18 2,8 = 290 mOsm/kg H2O La concentración de sodio se duplica para incluir la contribución osmótica del cloruro. La glucemia y el BUN se miden en mg/dL, los factores 18 y 2,8, los pesos atómicos divididos por 10, se emplean para pasar mg/dL a mOsm/kg H2O. Los valores obtenidos por esta fórmula difieren sólo de 1 a un 2 % de los valores obtenidos por osmometría y pueden utilizarse con fines clínicos. * Si se emplea la urea hay que dividir por 6 (el peso de la urea es 60). 3.8.5 Intercambios entre los espacios intracelular e intersticial. El movimiento del agua es pasivo y depende de las diferencias (gradientes) de presión hidrostática y de presión osmótica transmembrana. Las diferencias de presión hidrostática pueden omitirse y, por consiguiente, son los gradientes de presión osmótica los que determinan los movimientos de agua a su través. El espacio extracelular es el más expuesto a variaciones primarias de su osmolalidad, dada su relación más directa con el medio ambiente. Por lo tanto, el grado de hidratación celular depende fundamentalmente de las variaciones de la osmolalidad extracelular. Un aumento de la osmolalidad extracelular por pérdida de agua (deshidratación ) causa un flujo de agua desde la célula hasta el espacio extracelular, y ambos espacios experimentan una depleción de volumen. Un descenso de osmolalidad por hiperhidratación, causa un flujo de agua hacia el interior de la célula y ambos espacios experimentan una expansión de volumen. Cuando la osmolalidad extracelular se altera por ganancia o pérdida de solutos, los volúmenes extracelular e intracelular varían en direcciones opuestas. 3.8.6 Intercambios plasmático. entre los espacios intersticial y Los mismos principios básicos se aplican a la distribución entre estos dos componentes del espacio extracelular. Con la diferencia de que la pared de los capilares no constituye una barrera que se oponga a la difusión simple de la mayoría de solutos que contribuyen a la osmolalidad del medio extracelular. Sin embargo, es relativamente impermeable a las especies moleculares más grandes, como las proteínas. La agregación de estas moléculas dentro del componente vascular aumenta la osmolalidad y si no existiese una fuerza opuesta, todo el líquido extracelular pasaría al plasma. La presión osmótica ejercida por las proteínas séricas y, en particular, por la albúmina se denomina presión oncótica. Dado que las proteínas permanecen confinadas en el interior de los capilares, ellas ejercen la única fuerza osmótica efectiva que se opone a la salida de agua fuera del árbol vascular. El aumento de la presión hidrostática y/o la disminución de la presión oncótica de las proteínas séricas constituyen la causa más frecuente de acumulación de líquido en el espacio intersticial (edema). El equilibrio de estas fuerzas, fuerzas de Starling, es el determinante de la distribución estable del volumen entre ambos compartimentos. En general, estas fuerzas están ajustadas de modo que alrededor de un cuarto del líquido extracelular se encuentra dentro del sistema vascular y el resto corresponde al espacio intersticial. La ley de Starling de los capilares puede expresarse por la ecuación: Qf = Kf [( Pc - Pi ) s ( pc - py)] Qf es el flujo total de líquido a través de la membrana capilar; Kf, el coeficiente de filtración de líquido; Pc, la presión hidrostática capilar; Pi, la presión hidrostática intersticial; s, el coeficiente de reflexión; pc, la presión oncótica capilar (plasmática) y py, la presión oncótica intersticial. El estudio de esta ecuación revela la presencia de cuatro fuerzas de Starling coloidales e hidrostáticas que actúan a cada lado de la pared capilar. La presión hidrostática dentro del capilar (Pc), es la fuerza dominante que filtra líquido fuera del espacio vascular. La presión hidrostática intersticial (Pi) es generalmente negativa, pero se acerca a cero con acumulación de líquido de edema, y puede hacerse positiva si se acumula en grandes cantidades. La presión oncótica plasmática (pc) es la única fuerza de Starling que retiene líquido dentro del espacio vascular. La presión oncótica intersticial (py), en cambio, favorece la retención de líquido en el espacio intersticial. La concentración de proteína intersticial puede estar diluída por líquido de edema pobre en proteínas que cruza la membrana vascular. El aumento de Pi y la reducción de py sirven como asas de retroalimentación negativa que limitan la formación de edema. Según esto, el gradiente neto de presión hidrostática (Pc- Pi), que desplaza líquido a través de la membrana, y el gradiente neto de presión oncótica, que retiene líquido dentro del espacio vascular ( pc - pi ), determina el flujo de líquidos a través de las membranas capilares. Por último, el sistema linfático sirve de drenaje, demorando la acumulación del exceso de líquido filtrado. De este modo el aumento de flujo linfático compensa el aumento de desplazamiento de líquido transvascular. El coeficiente de filtración de líquido (Kf) representa la cantidad neta de líquido que cruza el lecho capilar para un desequilibrio dado de las fuerzas de Starling. Además de la propia membrana capilar, que puede ser el sitio principal de ingreso de proteínas, el movimiento de líquidos y solutos del espacio vascular hacia los linfáticos es afectado por la conductividad hidráulica de la membrana basal vascular, el gel intersticial y el linfático terminal. El coeficiente de reflexión (s) es una medida de la capacidad de la membrana capilar para servir como barrera contra el movimiento de proteínas. Para que una membrana capilar sea totalmente impermeable a las proteínas, s debe ser igual a uno y las proteínas deben ejercer toda su fuerza osmótica a través de esta perfecta membrana-barrera. Para una membrana capilar, que las proteínas pueden atravesar con tanta facilidad como el agua, sería igual a cero y las proteínas no ejercerían ninguna fuerza osmótica. Se ha calculado, que el s promedio es de 0.9 para los lechos capilares sistémicos, y 0.7 para los capilares pulmonares. En estados de permeabilidad capilar aumentada este valor puede disminuir a 0.4. Estos datos implican que pc - pies máxima en el tejido sistémico intacto, un poco menor en el pulmón intacto y mínima en capilares muy permeables. En caso de membranas capilares muy permeables, el edema se forma por excesivo egreso de proteínas y líquido hacia el espacio intersticial, con reducción de la presión oncótica efectiva a través de la membrana. Las alteraciones de las presiones físicas o de la integridad de la membrana capilar pueden explicar la formación de edema. Una de las consecuencias terapéuticas más importantes, es que el volumen plasmático no puede ser aumentado específicamente a menos que el líquido administrado contenga un coloide. La administración de solución salina a un individuo que ha perdido sangre reexpanderá el volumen del líquido extracelular, pero la mayor parte de la expansión se producirá en el compartimento intersticial. 3.8.7 Intercambios con el exterior El organismo intercambia agua y electrolitos con el exterior a través de las vías pulmonar, cutánea, digestiva y renal. En condiciones normales, las entradas y salidas se equilibran y el balance corporal permanece inalterado. Se acepta generalmente que si un paciente gana o pierde una sustancia, tiene un equilibrio positivo o negativo. Si no se producen cambios significativos, el equilibrio es neutro. Esto a menudo se denomina “estar en equilibrio”. El manejo adecuado de muchos pacientes incluye un registro diario cuidadoso de ingresos / pérdidas y del peso corporal. Los ingresos, incluyen la ingestión por vía oral, infusiones, transfusiones, etc. Las pérdidas incluyen orina, vómitos, heces y otras pérdidas intestinales. Las pérdidas insensibles por pulmón y piel no se miden, se utilizan valores promedio. Finalmente, destacar que el efecto de una perturbación en el equilibrio externo de la composición de los líquidos orgánicos es independiente del mecanismo fisiológico individual que está implicado. Agua: El balance diario en un adulto de 70 kg, las pérdidas inevitables diarias que tienen que ser sustituidas son las pérdidas insensibles ( 800 mL ), las pérdidas fecales ( 200 mL ) y el sudor (100 mL ). Además, deberá producirse orina suficiente para excretar una carga de solutos de 600 mOsm producidos diariamente por el cuerpo. La capacidad de concentración de la orina en los enfermos quirúrgicos, lesionados y muy graves está disminuída. En estos enfermos hay una cantidad mínima obligatoria de aproximadamente 900 mL. El paciente de 70 kg necesita en total 2.000-2.500 mL/día de H2O para mantener su equilibrio líquido. Sodio: Las pérdidas de sodio alcanzan unos 30 mEq/día por las heces y el sudor, con una pérdida variable por la orina. En casos de gran privación de agua, los riñones pueden absorber casi todo el sodio filtrado como respuesta al aumento de secreción de aldosterona. La administración aproximada de 1-2 mEq/kg/día de sodio a los adultos o de 1 mEq/kg/día a los niños, no sólo sustituirá las pérdidas obligatorias, sino que suprimirá también la secreción de aldosterona en proporción suficiente para ayudar a las pérdidas de potasio. Si se utiliza ClNa, también se cubren los requerimientos orgánicos diarios de cloruros. Potasio: Las pérdidas diarias de potasio por la orina y el sudor alcanzan los 40-60 mEq. Por lo general, su sustitución con 0,5-1,0 mEq/kg/día basta para mantener el equilibrio de este ión en el enfermo con riñones normales. 4 REFERENCIAS Guyton y Hall. 2009. Tratado de fisiología médica. 10 edición México Sánchez., A . 2006. Sistema nervioso., Revisión bibliográfica, Facultad de Ciencias Veterinarias -Universidad de Buenos Aires., Argentina. Murray. R. 2009. Harper bioquímica ilustrada. 28ª edición. México. Comack, H. D., 2001. Histología de Ham., Editorial Harla., 9 ed. México Ganong, W. F., 2010., Fisiología Medica, Editorial Manual moderno - MC Graw Hill, 23 edición., México Cunningham J. G., 2006., Fisiología Veterinaria., Elsevier., 3ra edición., España Welsch U., 2009., Histología Sbotta Welsch, Editorial Médica Panamericana. Madrid España, 2ª reimpresión.