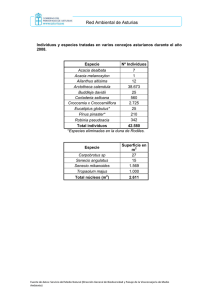
Unidad 3: BIOQUÍMICA METABÓLICA APLICADA
Anuncio

Unidad 3: BIOQUÍMICA METABÓLICA APLICADA Capítulo 7: Bioquímica de Forrajes Lección 31. Fotosíntesis, Redes alimentarias y flujo de energía Siguiendo la pista a la fuente última de energía utilizada por un organismo determinado en su hábitat natural, por ejemplo un zorro en un dominio dado de bosque, encontraríamos que existe una jerarquía de organismos, llamada cadena alimentaria, o trófica, que provee al zorro de la energía y de los materiales necesarios para sustentar su vida. Esta cadena alimentaria podría empezar con las células fotosintéticas de las plantas verdes, que convierten el Gas carbónico (CO2) en material celular nuevo. La planta puede, a su vez, ser consumida por larvas de insectos que, del mismo modo, pueden ser consumidas por sapos o pájaros que, a su vez pueden ser consumidos por el zorro. En una comunidad ecológica dada de organismos vivos, se interconectan entre sí muchas cadenas alimentarias individuales, formando lo que se llama una red alimentaria ó red trófica. Tal red alimentaria está formada por varias capas de organismos. En primer lugar, tenemos a los productores, aquellas células que pueden utilizar las formas más simples del carbono procedentes del medio ambiente, tales como el (CO2), Después, tenemos una capa primaria de consumidores, que se alimentan de los productores, seguida de ulteriores capas de consumidores. Finalmente, para completar el ciclo, están los desintegradores: bacterias y hongos, que provocan la descomposición y putrefacción de los consumidores muertos y que, de este modo, devuelven al suelo y a la atmosfera formas simples de carbono. Los organismos vivos se pueden clasificar en dos grandes grupos según su posición en la red alimentaria: autótrofos y heterótrofos. Organismos autótrofos (el término significa que se autoalimentan) son aquellos que pueden utilizar formas simples de carbono, a partir de los cuales elaboran todos sus componentes celulares. Los productores situados en la base de la red alimentaria son autótrofos; entre ellos están incluídas muchas bacterias y algas, así como pantas superiores. Los organismos heterótrofos (que se alimentan por otros ), por otra parte, no pueden utilizar formas simples de carbono, tales como (CO2), y que requieren formas más complejas: moléculas orgánicas como la glucosa(C 6H12O6). Los consumidores y desintegradores de la red alimentaria son heterótrofos y dependen, en última instancia, de los autótrofos para generar los complejos nutrientes que necesitan. 1 Examinemos ahora esta red alimentaria e investiguemos las fuentes de energía para cada capa de organismos. Al hacerlo, encontraremos que la gran mayoría de los organismos autótrofos productores obtienen su energía de la luz solar, la cual utilizan para convertir el Gas carbónico en materiales celulares más complejos mediante la fotosíntesis. Así, la mayor parte de los productores son autótrofos fotosintéticos. Pero cuando examinamos las capas sucesivas de consumidores, encontramos que ninguna de ellas tiene la capacidad de utilizar energía luminosa. Más bien, la mayor parte de ellos obtienen la energía que necesitan mediante la combustión de moléculas orgánicas complejas, tales como la glucosa, obtenidas a partir de los productores que ellos consumen. En este proceso, que requiere Oxígeno y que se llama respiración, la molécula sencilla y pequeña de anhídrido carbónico (CO2), es el producto final. Las células heterotróficas, por lo tanto obtienen energía mediante la degradación de moléculas nutrientes complejas a formas más simples En cada nivel de la red alimentaria se consume energía para realizar diversas clases de trabajo biológico, tal como la síntesis de material celular nuevo a partir de precursores simples, el movimiento de materiales en contra de gradientes y el trabajo de contracción o movimiento. Sin embargo, en cada nivel de la red alimentaria encontramos que hay pérdidas por degradación, de tal modo que cada vez que tiene lugar algún proceso físico o químico hay una conversión incompleta de energía de una clase en otra. Como resultado, parte de la energía hecha aprovechable para cada capa de organismos es disipada en el medio y, por lo tanto no es utilizable para realizar trabajo. Solamente una pequeña fracción de la energía solar absorbida por los productores en la capa básica de la red alimentaria alcanza siempre a la capa superior de los últimos consumidores a medida que estos consumidores últimos mueren y sus tejidos son degradados a productos orgánicos simples por los desintegradores, la energía de nuevo se pierde y se disipa en el medio. En definitiva, el flujo de energía que parte del sol y que corre a través de la red alimentaria se dispersa finalmente en el medio, generando la denominada entropía. ENERGIA SOLAR Y FOTOSÍNTESIS La luz sola visible, fuente de toda la energía biológica, es una forma de energía electromagnética o radiante que, en la última instancia, surge de la energía nuclear. A la temperatura inmensamente elevada del sol, la cual se cree que es de varios millones de grados centígrados, una parte de la enorme energía encerrada en el núcleo de los átomos 2 de hidrógeno es liberada a medida que estos últimos se convierten en átomos de helio (He) y positrones, mediante fusión termonuclear. En este proceso se libera un cuanto de energía en forma de radiación gamma. El cuanto se presenta por el termino (h), en el cual h es la constante de planck y es la frecuencia de la radiación gamma. Después de una compleja serie de reacciones en las cuales la radiación gamma es absorbida por los positrones, gran parte de la energía de la radiación gamma es emitida en forma de fotones o cuantos de energía luminosa. Las reacciones de fusión nuclear que tienen lugar en el sol son, en última instancia, la fuente de toda la energía biológica sobre la tierra. En general, tendemos a asociar el término (fotosíntesis) con el mundo visible de las plantas superiores: pastos, cultivos herbáceos y árboles. Pero esos organismos fotosintéticos macroscópicos realmente no constituyen sino una pequeña fracción de todos los organismos conocidos capaces de fotosintetizar. Se ha estimado que un 90 por 100 de las fotosíntesis que se lleva a cabo sobre la tierra es realizado en el mar por diversas clases de microorganismos entre los que se incluyen las bacterias, algas, diatomeas y dinoflagelados. Existe otro error común en torno a la fotosíntesis de las plantas superiores, no todas las células de las raíces, los tallos y los frutos de las plantas son capaces de fotosintetizar: son heterotróficas y, por lo tanto, se parecen a las células animales. Solamente las células que poseen el pigmento verde de la clorofila pueden llevar a cabo dicho proceso. Por otra parte, en la oscuridad, cuando no se dispone de energía solar, incluso estas células funcionas como las células heterotróficas: deben oxidar parte de su glucosa, a expensas del Oxígeno, para obtener energía en la oscuridad. La fotosíntesis consiste en la absorción de la energía radiante por la clorofila y otros pigmentos, seguida de la conversión de la energía luminosa absorbida en energía química, y la utilización de esa energía química para la reducción del anhídrido carbónico absorbido de la atmosfera para formar glucosa. En la mayor parte de los organismos fotosintéticos, en particular las plantas superiores, el Oxígeno molecular(O2) es el otro producto final importante, pero en otras, tales como las bacterias fotosintéticas, no se forma Oxígeno. La forma más simple de la ecuación global para la formación fotosintética de glucosa y Oxígeno a partir de anhídrido carbónico (Dióxido de Carbono) y agua en las plantas superior es: 3 Radiación UV 6CO2 + 6H2O + 686kcal E=h C6H12O6 + 6O2 Donde 686 Kcal, representa la cantidad mínima de energía útil que debe ser proporcionada por la luz solar absorbida para lograr la formación de un mol de glucosa a partir de un mol de CO2 y otro de H2O en condiciones estándar. En termodinámica química la unidad básica de energía es la caloría- gramo y se define como la cantidad de energía requerida, en forma de calor, para elevar la temperatura de 1g de agua a 15°C, exactamente en 1°C. La gran cantidad de energía necesaria para que tenga lugar la fotosíntesis es suministrada por la energía luminosa captada por la clorofila de las hojas. La ecuación fotosintética puede volver a escribirse, de modo que indique que la fuente de energía son los cuantos luminosos: nh , como sigue. 6CO2 + 6H2O + nh Radiación UV E=nh C6H12O6 + 6O2 Esta ecuación nos da solamente una visión global del proceso fotosintético; no dice nada acerca del mecanismo o del camino por el cual tiene lugar. Realmente, la fotosíntesis en las células de las plantas es un proceso mucho más complejo que lo que esta ecuación de apariencia sencilla puede sugerir. Existen más de un centenar de RBE consecutivas en la producción fotosintética de glucosa a partir de anhídrido carbónico y agua, cada uno de las cuales está catalizada por una molécula enzimática especifica, las cuales están agrupadas en dos etapas claves: Reacciones de Fase lumínica y Reacciones del Ciclo de Calvin La glucosa no es el único producto de la fotosíntesis. Durante dicho proceso se sintetizan también otros componentes carbonados de las células vegetales, tales como la celulosa, proteínas y lípidos. Todas estas sustancias, ricas en energía química, son utilizadas posteriormente como fuente de energía por los organismos heterotróficos, es decir, por los consumidores que se alimentan de plantas verdes. Lección 32. RESPIRACION EN CELULAS HETEROTRÓFICAS 4 La fase siguiente en el flujo de la energía biológica es la utilización de la energía de los carbohidratos, grasas y proteínas producidas en la fotosíntesis por los organismos heterótrofos, que oxidan estos materiales por medio del Oxígeno. Realmente, los organismos heterótrofos necesitan los complejos productos de la fotosíntesis por dos razones. En primer lugar, necesitan la energía química que pueden obtener por degradación de las estructuras complejas de alta energía de moléculas tales como la glucosa. Pero los heterótrofos necesitan también complejos compuestos de carbono, tales como la glucosa, como unidades estructurales para la síntesis de sus propios componentes celulares, ya que son incapaces de utilizar el anhídrido carbónico con este fin. Entre los heterótrofos están incluÍdos todos los organismos del reino animal, muchas bacterias y hongos, así como muchas células del reino vegetal. Se estima que más del 90 por 100 de todo el Oxígeno consumido por todos los heterótrofos es utilizado por microorganismos invisibles del suelo y del mar. La mayor parte de las células heterotróficas utilizan el Oxígeno que toman de la atmosfera para oxidar la glucosa y otros nutrientes, dando lugar a los productos finales estables, anhídrido carbónico y agua. Sin embargo, algunos heterótrofos son incapaces de utilizar el Oxígeno; degradan la glucosa en compuestos más simples, tales como el acido láctico: H3C-CH (OH)-COOH, en ausencia de Oxígeno, en el proceso llamado fermentación. Los productos de la fermentación, tales como el acido láctico, son posteriormente oxidados hasta CO2 y H2O por otros organismos heterotróficos, en particular, aquellos que utilizan Oxígeno. En definitiva, las células del mundo heterotrófico llevan a cabo la oxidación completa de los nutrientes orgánicos producidos por los autótrofos hasta convertirlos en el producto final, anhídrido carbónico. En el proceso global, mediante el cual las moléculas de alimentos son oxidadas por las células heterotróficas a expensas del Oxígeno, recibe el nombre de respiración. La ecuación química para la oxidación de la glucosa durante la respiración es : C6H12O6 + 6O2 RBE Mitocondria 6CO2 + 6H2O + 38ATP Vemos inmediatamente que esta ecuación es la inversa de la correspondiente a la fotosíntesis. Por otra parte, observamos que la combustión completa de un mol de glucosa puede producir un máximo de 38ATP de energía química útil. 5 Aunque la ecuación química de la respiración parece sencilla, no nos dice nada acerca de los mecanismos o del camino seguido por la respiración en las células heterotróficas. Realmente, hay más de 70 reacciones Bioquímicas enzimáticas (RBE) consecutivas en la oxidación de la glucosa en las células heterotróficas, las cuales se desarrollan en los procesos catabólicos celulares como: Glucólisis, Betaoxidación de ácidos grasos, desaminación y transaminación oxidativa, lipólisis, Ciclo de Krebs y Fosforilación Oxidativa (Adaptado de Lehninger.,1975, por el autor) Lección 33. Fijación Biológica del Nitrógeno (FBN) A pesar de que la atmósfera de la tierra posee un 78% del Nitrógeno gaseoso (N 2), siendo el principal reservorio de este elemento, este se encuentra en cantidades reducidas en los organismos , especialmente en las plantas. Sin embargo, como la mayoría de los seres vivos no pueden utilizar dicho nitrógeno atmosférico para biosintetizar aminoácidos, proteínas y otros compuestos nitrogenados, entonces, dependen del nitrógeno presente en los minerales del suelo. En consecuencia, a pesar de esta abundancia de nitrógeno en la atmósfera, la escasez de nitrógeno en el suelo constituye un factor limitante para el crecimiento y desarrollo de las plantas. Es importante anotar que tan sólo ciertos microorganismos son capaces de asimilar nitrógeno molecular transformándolo en biomoléculas utilizables para ellas (Bidwell, R., 1989). El proceso a través del cual circula nitrógeno a través del mundo orgánico y el mundo físico se denomina ciclo del nitrógeno. Este ciclo consta de las siguientes etapas: Fijación biológica del nitrógeno (FBN):Es un proceso simbiótico reductivo dado en bacterias fijadoras como las del género Rhizobium, que viven en nódulos de las raíces de leguminosas y de algunas plantas leñosas ó las cianobacterias que son propias de ambientes acuáticos. La FBN , consiste en la conversión del nitrógeno gaseoso (N2) mediante la enzima nitrogenasa que cataliza la ruptura del triple enlace ,hasta producir amoníaco (NH3), forma utilizable para los organismos. La ecuación química es la siguiente: - N2 + 8e + 16 Mg ATP + 16 H2O Nitrogenasa Raices 6 2NH3 + H2 + 16 Mg ADP + 16 Pi + 8 Donde : e- = electrones; Mg= Magnesio ; ATP =Adenosín trifosfato ; ADP =Aenosín Difosfato ; Pi = Fosfato inorgánico Nitrificación: proceso de oxidación del amoníaco realizado por dos tipos de bacterias: Nitrosomonas y Nitrobacter (comunes en el suelo). Dicho proceso ocurre en dos etapas: A. Nitrosación: Producción de Nitrito (NO2-) Las bacterias nitrosomas oxidan el amoníaco produciendo nitrito (NO2-) , hidrógenos y agua; los H+ son utilizados en el metabolismo bacteriano para generar moléculas energéticas de ATP ; La ecuación química del proceso es como sigue: + 2 NH3 + 3 O2(g) Nitrosomas - 2 NO2 + 2 H + 2 H2O B. Nitratación : Producción de Nitrato (NO3-) Como el nitrito es un ión tóxico para las plantas limitando la mayoría de cultivos nitrobacter lo continúan oxidando , las , hasta generar el ión nitrato, el cual es fácilmente asimilado por las plantas; la ecuación es la siguiente: - 2 NO2 + O2 (g) Nitrobacter anteriormente se puede observar en la figura 4: 7 2 NO3 - Lo descrito Figura 4:Etapas dadas en el ciclo del Nitrógeno (Tomado de: http://quimica.laguia2000.com/conceptos-basicos/ciclo-del-nitrogeno ) Asimilación: las raíces de las plantas absorben el amoníaco (NH3) o el nitrato (NO3 -), e incorporan el nitrógeno en proteínas, ácidos nucleicos y clorofila. Cuando los animales se alimentan de vegetales consumen compuestos nitrogenados vegetales y los transforman en compuestos nitrogenados animales. Amonificación: consiste en la conversión de compuestos nitrogenados orgánicos en amoníaco, se inicia cuando los organismos producen desechos como urea (orina) y ácido úrico (excreta de las aves), sustancias que son degradadas para liberar como amoníaco el nitrógeno en el ambiente abiótico. El amoníaco queda disponible para los procesos de nitrificación y asimilación. El nitrógeno presente en el suelo es el resultado de la descomposición de materiales orgánicos y se encuentra en forma de compuestos orgánicos complejos, como proteínas, aminoácidos, ácidos nucleicos y nucleótidos, que son degradados a compuestos simples por microorganismos - bacterias y hongos - que se encuentran en el suelo. Estos microorganismos usan las proteínas y los aminoácidos para producir sus propias proteínas y liberan el exceso de nitrógeno en forma de amoníaco (NH3) o ion amonio (NH4+). Desnitrificación: es el proceso que realizan algunas bacterias ante la ausencia de oxígeno, degradan nitratos (NO3 -) liberando nitrógeno (N2) a la atmósfera a fin de utilizar el oxígeno para su propia respiración. Ocurre en suelos mal drenados. 8 A pesar de las pérdidas de nitrógeno, el ciclo se mantiene gracias a la actividad de las bacterias fijadoras de nitrógeno, capaces de incorporar el nitrógeno gaseoso del aire a compuestos orgánicos nitrogenados. En los sistemas naturales, el nitrógeno que se pierde por desnitrificación , lixiviación, erosión y procesos similares es reemplazado por el proceso de fijación y otras fuentes de nitrógeno. La interferencia antrópica (humana) en el ciclo del nitrógeno puede, no obstante, hacer que haya menos nitrógeno en el ciclo, o que se produzca una sobrecarga en el sistema. Por ejemplo, los cultivos intensivos, su recogida y la tala de bosques han causado un descenso del contenido de nitrógeno en el suelo (algunas de las pérdidas en los territorios agrícolas sólo pueden restituirse por medio de fertilizantes nitrogenados artificiales, que suponen un gran gasto energético). Por otra parte, la lixiviación del nitrógeno de las tierras de cultivo demasiado fertilizadas, la tala indiscriminada de bosques, los residuos animales y las aguas residuales han añadido demasiado nitrógeno a los ecosistemas acuáticos, produciendo un descenso en la calidad del agua y estimulando un crecimiento excesivo de las algas. Lo analizado anteriormente , se resume en la figura 5: Figura 5 : Resumen esquemático del ciclo del Nitrógeno (Tomado de : http://roble.pntic.mec.es/~mbedmar/iesao/quimica/ciclodel.htm) Lección 34. Forrajes : composición y análisis químico proximal EVALUACIÓN PROXIMAL DE TRES ESPECIES CON POTENCIAL FORRAJERO Puerto, Luz1 ; Granados, Jairo 2; Barreto, Leonor3 1,2,3 , Escuela de Ciencias Agrícolas, Pecuarias y del Medio Ambiente. ECAPMA-UNAD, 2010. 9 RESUMEN Se evaluó la composición fitoquímica de dos especies leguminosas y una herbácea, con el fin de conocer diferencias asociadas a la zona lumínica de muestreo (zona alta o fototrópica, zona media y zona baja o geotrópica). Se escogieron las leguminosas Acacia negra (Acacia decurrens), Matarratón (Gliricidia sepium) y la herbácea Botón de Oro (Tithonia diversifolia), las cuales fueron evaluadas en el laboratorio de nutrición animal de la Universidad Nacional Abierta y a Distancia (UNAD) Sede Bogotá D.C. Colombia, mediante un diseño totalmente aleatorizado y tres réplicas por zona lumínica de cada especie. Los contenidos que presentaron un nivel significativamente superior al resto fueron para los contenidos de materia seca para Acacia decurrens con 45,60%, cenizas para Tithonia diversifolia con 11,36%, materia orgánica para Acacia decurrens con 95,68%, nitrógeno total para Tithonia diversifolia con 2,58%, y proteína cruda fueron mayores en Tithonia diversifolia con (16,15%), seguido de Gliricidia sepium con (13,48%) y Acacia decurrens con (13,47%). Por lo tanto estas especies constituyen buenas alternativas como suplemento en los sistemas de producción ganadero en el trópico Colombiano. Palabras clave: Leguminosas, Herbácea, composición química, materia seca, cenizas, materia orgánica, nitrógeno total y proteína cruda. INTRODUCCIÓN Estas especies son conocidas por los ganaderos pero poco utilizadas, tal vez debido al desconocimiento acerca de sus propiedades proteicas, minerales y vitamínicas, adaptación, buen rendimiento de biomasa y rápida recuperación después del corte, resistencia a la sequia, asociación con otras especies arbóreas, mejoramiento del suelo, y lo mejor siendo fuentes generadoras de oxigeno y de agua; además del ahorro por concentrado que se tendría en el consumo en grandes y pequeñas producciones agropecuarias. Estas especies se encuentran usualmente en los caminos de las carreteras, actúan como barreras rompevientos y heladas, cercas vivas, plantas de adorno debido a su belleza, son muy fructíferas en época de verano; pueden ser suministradas en pastoreo o como forraje en hoja verde, en ensilaje, en hoja seca o molida. Es posible su almacenamiento por largos periodos; son un excelente alimento para los animales, dando como resultados aumento en la producción y calidad de la leche, aumento de fertilidad, calidad en cuanto a pigmentación de huevos y carne. Es un reto el uso de estas plantas arbóreas especialmente en la ganadería colombiana, las cuales al ser incluidas en la dieta animal se generaría una reducción en los costos además de ofrecer alternativas ecológicamente razonables y eficientes con el medio ambiente. El propósito del presente trabajo fue evaluar la composición del análisis químico proximal el cual incluye: materia seca cenizas, materia orgánica, nitrógeno total y proteína cruda 10 de tres especies potencialmente forrajeras como son de clima frío Acacia decurrens y de clima templado Tithonia diversifolia y Gliricidia sepium. MATERIALES Y MÉTODOS Características de la zona de muestreo La recolección del material vegetal se realizo así: Acacia decurrens (municipio Bojacá Cundinamarca, finca Acapulco, con 2598 msnm y temperatura media 14ªC), Tithonia diversifolia (municipio Silvania Cundinamarca, orilla de la carretera, con 1470 msnm y temperatura media 20ªC) y Gliricidia sepium (municipio La Mesa Cundinamarca, Finca Orquídeas, con 1198 msnm y temperatura media 20ªC). Recolección y preparación de las muestras Las muestras de las hojas fueron seleccionadas al azar por cada zona lumínica de la planta como son: alta o fototrópica, media y baja o geotrópica, fueron secadas a temperatura ambiente, colocadas en bolsas de papel y rotuladas de acuerdo a la zona lumínica de muestreo, para su posterior análisis en el laboratorio de la Universidad Nacional Abierta y a Distancia (UNAD). Las muestras fueron recolectadas en los meses de abril, mayo y junio de 2009, en esta época se observo un verano prolongado. La edad de las plantas no fue determinada, sin embargo se cree que las especies Acacia decurrens y Tithonia diversifolia eran plantas maduras y la especie Gliricidia sepium tenia aproximadamente un año según comentario del propietario de la finca donde se recolecto la muestra. RESULTADOS Y DISCUSIÓN Análisis Proximal En la tabla 1 se muestran los resultados obtenidos de los análisis en las especies evaluadas. Tabla 1. Composición bromatológica de análisis proximal Especie A. decurrens T. diversifoli a G. sepium EEM MS Cz MO NT PC -----------45,60a --------------4,32a (%) ---------------- 95,68a -----------2,15a 28,43a 11,36b 88,64b 2,58a 16,15a 30,37b 0,387 8,25c 0,387 91,75c 0,387 2,16b 0,052 13,48b 0,327 13,47a (MS)=Materia seca, (Cz)=Cenizas, (MO)=Materia orgánica, (NT)= Nitrógeno total, (PC)=Proteína cruda. Distintas letras en una columna indican diferencias significativas en las medias (P<0.05). EEM: Error estándar de la media. 11 Materia Seca (MS). El análisis de varianza muestra que se presentaron diferencias altamente significativas (P<0,01) entre los promedios de las medias de las especies trabajadas. Los promedios indican que la mayor especie que tuvo materia seca fue Acacia decurrens con 45,60%, seguido de Gliricidia sepium con 30,37% y Tithonia diversifolia con 28,43%, la especie Acacia decurrens tiene mayor porcentaje de materia seca que Gliricidia sepium y Tithonia diversifolia. Acacia decurrens supera a Gliricidia sepium en un 15.23% mientras que la diferencia con Tithonia diversifolia es de 17,17%. Los datos encontrados son similares a los reportados por (Londoño y Velasquez,1999) con 48,72% para Acacia decurrens, (Rosales, 1992) con 23% para Tithonia diversifolia, y (Laredo y Cuesta,1990) con 24,9% para Gliricidia sepium. (Gráfica 1). En cuanto a la prueba Duncan se analizó medidas en una columna con diferente letra, indicando diferencia estadística (P<0,05). 60% 50% %Materia Seca 48,72% 45,60% 40% 28,43% 23,00% 30% 30,37% 24,90% 20% 10% 0% -10% Fototropica Media Geotropica Gráfica 1. Promedio comparativo %Materia Seca Cenizas (Cz). El análisis de varianza muestra que se presentaron diferencias altamente significativas (P<0,01) entre los promedios de las medias de las especies trabajadas. Los promedios muestran que la mayor concentración de cenizas la tuvo la especie Tithonia diversifolia con 11,36% seguido de Gliricidia sepium con 8,25% y luego Acacia decurrens con 4,32% lo que demuestra que la especie Tithonia diversifolia, contiene un mayor contenido de minerales que Gliricidia sepium y Acacia decurrens. Tithonia diversifolia supera a Gliricidia sepium en un 3,11% mientras que la diferencia con Acacia decurrens es de 7,04%. Los datos encontrados son similares a los reportados por (Carvajal T,1999) con 4,29% para Acacia decurrens; mayores a los reportados (Navarro,1990) con 9,42% para Tithonia diversifolia y similares a los reportados por (Laredo y Cuesta,1990) con 8,90% para Gliricidia sepium. (Gráfica 2). 12 %Cenizas 15% 11,36% 9,42% 10% 5% 8,25% 8,90% 4,32% 4,29% 0% Fototropica Media Geotropica Gráfica 2. Promedio comparativo % Cenizas Materia Orgánica (MO). El análisis de varianza muestra que se presentaron diferencias altamente significativas (P<0,01) entre los promedios de las medias de las especies trabajadas. Los promedios indican que la mayor cantidad de materia orgánica se encontró en la especie Acacia decurrens con 95,68% seguido de Gliricidia sepium con 91,75% y por último de Tithonia diversifolia con 88,64%, lo que demuestra que la especie Acacia decurrens, contiene un mayor contenido de materia orgánica que Gliricidia sepium y Tithonia diversifolia. Acacia decurrens supera a Gliricidia sepium en un 3,93% mientras que la diferencia con Tithonia diversifolia es de 7,04%. Los datos encontrados son similares según los reportados por (Carvajal, T.) con 95,18% para Acacia decurrens. (Gráfica 3). 100% 90% 80% 70% 60% 50% 40% 30% 20% 10% 0% 95,68% 95,18% Fototropica %Materia Orgánica 88,64% 78,59% Media 91,75% Geotropica Gráfica 3. Promedio comparativo %Materia Orgánica 13 Nitrógeno total (NT). El análisis de varianza muestra que se presentaron diferencias significativas (P<0,05) entre los promedios de las medias de las especies trabajadas. Los promedios enseñan que la mayor concentración de nitrógeno total se presento en la especie Tithonia diversifolia con 2,58% seguido por Gliricidia sepium con 2,16% y por último Acacia decurrens con 2,15%, lo que demuestra que la especie Tithonia diversifolia, contiene un mayor contenido de nitrógeno total que Gliricidia sepium y Acacia decurrens. Tithonia diversifolia supera a Gliricidia sepium en un 0,42% mientras que la diferencia con Acacia decurrens es de 0,43%. (Gráfica 4). %Nitrógeno Total 3,00% 2,58% 2,50% 2,16% 2,15% 2,00% 1,50% 1,00% 0,50% 0,00% Adta Promedio Fototropica Tdtm Promedio Media Gstm Promedio Geotropica Gráfica 4. Promedio comparativo %Nitrógeno Total Proteína Cruda (PC). El análisis de varianza muestra que se presentaron diferencias significativas (P<0,05) entre los promedios de las medias de las especies trabajadas. Los promedios indican que la mayor concentración de proteína cruda se presento en la especie Tithonia diversifolia con 16,15% seguido de Gliricidia sepium con 13,48% y luego por Acacia decurrens con 13,47%, lo que demuestra que la especie Tithonia diversifolia, contiene un mayor contenido de proteína cruda que Gliricidia sepium y Acacia decurrens. Tithonia diversifolia supera a Gliricidia sepium en un 2,67% mientras que la diferencia con Acacia decurrens es de 2,68%. Los datos encontrados son similares según los reportados por (Londoño et al.,1999) con 14,86% para Acacia decurrens, (Navarro et al.,1990) con 14,84% para Tithonia diversifolia, y (Rosales et al., 1996) con 14,7% para Gliricidia sepium . (Gráfica 5). 14 20,00% 15,00% %Proteina Cruda 14,86% 13,47% 16,15% 14,84% 14,70% 13,48% 10,00% 5,00% 0,00% Fototropica Media Geotropica Gráfica 5. Promedio comparativo %Proteína Cruda Conclusiones y Recomendaciones Las especies estudiadas constituyen una importante fuente de forraje en la alimentación animal, especialmente de rumiantes, ya que los contenidos de proteína cruda fueron considerables, en donde se presento mayor porcentaje en la especie Tithonia diversifolia con (16,15%), seguido de Gliricidia sepium con (13,48%) y Acacia decurrens con (13,47%). Estadísticamente se observaron diferencias significativas (P<0,05) y altamente significativas (P<0,01) entre especies; con respecto a las zonas no se observaron diferencias estadística (P>0,05) para los análisis bromatológicos. Esto significa que se presentan diferencias entre las especies, pero no se presentan diferencias entre las zonas de cada especie. Se recomienda el uso de estas tres especies en sistemas silvopastoriles, ya que ayudan a favorecer la composición química de los pastos, hay sombra para el pasto y los animales, disminución de temperatura, disponibilidad de nutrientes y de agua, existiendo así mejoras en la fertilidad del suelo; además de incrementar significativamente las contribuciones económicas y sociales, que son fundamentales para el proceso de cambio. Se recomienda diseñar y evaluar dietas para rumiantes y monogástricos con estas especies arbóreas, con el fin de determinar el valor nutritivo, rendimiento y producción en la alimentación animal como: gallinas ponedoras, pollo engorde, cerdos, rumiantes y otros. Lección 35. Bioquímica del ensilaje Análisis químico de ensilajes de cogollo de caña, inoculados con dos clases de bacterias ácido lácticas Jairo E. Granados* José D. Neva**. Departamento de Química, Facultad de Ciencias Agrarias. UNAD, Bogota D.C. Colombia. Calle 14 S Nº 14-23. *Docente15 MSc. ECAPMA; [email protected]. ** Zootecnista UNAD PALABRAS CLAVES: Ensilaje, lactobacillus, ácido láctico, cogollo de caña Introducciòn: El ensilaje es una técnica de conservación de forrajes o productos agrícolas basado en los procesos bioquímicos de fermentación de carbohidratos solubles (CHOS), presentes en la microflora epifítica del forraje, que produce principalmente ácido láctico (Davies et al., 1998). Este método, permite mantener la calidad nutricional que tenía el forraje en el momento de corte, lo cual depende de factores como: concentración y disponibilidad de CHOS, tipo de bacterias ácido lácticas epifíticas y la cantidad de microorganismos indeseables, como la enterobacteria y el clostridium presentes en el material que se ensila (Davies et al., 1998). En este experimento se evaluò la inclusión de dos tipos de bacterias ácido lácticas en ensilajes de cogollo de caña y su efecto en la producción de ácidos láctico y acético, lo mismo que en la reducción del pH, cambios en la cantidad de proteína cruda, CHOS, materia seca y porcentaje de cenizas. Metodología: El experimento se desarrollò en dos etapas: 1. Preparación de 25 ensilados a partir de cogollo de caña tipo panelera fresca tipo Puerto Rico, que presentó un 8.10% de CHOS, los cuales se distribuyeron en un diseño al azar constituido por cinco tratamientos y cinco réplicas por cada uno. Los componentes utilizados en la preparación de los ensilajes experimentales que constituyeron cada tratamiento, se muestran en la Tabla 1. TABLA 1. Tratamientos utilizados en el experimento Component e T1 T2 T3 T4 T5 Cogollo (Kg) Melaza (Kg) Agua (Kg) Urea (Kg) Inóculo A*(g/dL) Inóculo B *(g/dL) 100.0 3.0 3.0 0.5 0.0 0.0 100 .0 3.0 3.0 0.5 0.5 0.0 100 .0 3.0 3.0 0.5 1.5 0.0 100. 0 3.0 3.0 0.5 0.0 0.5 100. 0 3.0 3.0 0.5 0.0 1.5 *IA: Lactobacillus plantarum, estreptococus faecium, pediococcus acidoláctico + complejo enzimático: Celulasa, amilasa y pentosana. *IB: Lactobacillus faecium; estreptococcus acidoláctico+complejo celulasa-amilasa Análisis químicos en el laboratorio: Al final del experimento (40 días), se tomaron porciones iguales de cada réplica, aproximadamente 500 g del ensilado. Para efectuar análisis de materia seca (MS); humedad (H); Cenizas (CZ); Nitrógeno (N); pH, carbohidratos solubles (CHOS); ácido láctico (AL); y ácido acético (AA). Esto se realizò según técnicas analíticas de la AOAC (1988). Resultados y Discusión 16 Los datos provenientes de las variables estudiadas, fueron analizadas por medio del paquete estadístico SPSS para Windows Versión 10.0. Se utilizò el análisis de varianza (ANAVA) en una sola vìa, para detectar las diferencias estadísticas entre los tratamientos. Además, se aplicaron pruebas de comparaciones múltiples de Tukey y 6 contrastes ortogonales. Los resultados, mostraron un incremento de la concentración de ácido láctico en los ensilados T2, T3, T4 y T5 a expensas del descenso en el nivel de carbohidratos solubles de cada tratamiento. Dicha reducción fue lineal (r 2 = 0.912) y significativa (p<0.05) con respecto a la producción de ácido láctico. Además, la disminución del pH fue causada por el incremento altamente significativo (p<0.01) del A.L. En los ensilajes tratados con el inòculo A. Este comportamiento se puede explicar por la probable acciòn conjunta de celulasas amilasas, pentosanas y bacterias ácido lácticas, sobre los procesos de hidrólisis y fermentación anaeróbica de los carbohidratos solubles presentes en el material ensilado, hasta producir gran cantidad de ácido láctico y en algunos casos ácido acético (Méndez, 1989). En general los inóculos bacterianos incrementaron la materia seca y el porcentaje de cenizas de los ensilados, lo cual causó una disminución apreciable de su humedad y un aumento significativo (p<0.05) de la proteína cruda. Se concluyó que el inòculo A, adicionado en el nivel 1.5 g/dL, generó una mayor preservación y calidad nutricional de los ensilados de cogollo de caña, demostrado en la recuperación de materia seca, aumento en la proteína cruda, notable degradación de los carbohidratos solubles e incremento significativo del ácido láctico y disminución del pH. Referencias Association of analytical chemists. 1980. oficial methods of análisis. 12th ed. AOAC. Washington, D.C. Alltech, Inc. 1999. Biotechnology Center. Bacterias y enzimas para conservación de ensilajes. Nicholasville, K.Y. 40356. Davies, D.R. J. merry, A.P. williams. E.L. Bakewell. D-K- Leemans and J.K.S. Tweed: 1998. Proteolysis during Ensilaje of Forages Varying in soluble sugar content. J. Dairy, SCi, 81: 444-453. Dubois, M.K.A. Guilles, J.K. Harnilton, P.A. Rebers and F. Smith. 1956. Colorimetric method for determination of sugars and related substances anal chem. 28:350-356. Méndez, G. 1989. curso de ensilajes I.C.A. Tibaitatá. Bogotá D.C. – Colombia. Capítulo 8. Metabolismo primario en animales y plantas (Tomado y adaptado de: Aurora Hilda Ramírez-Pérez y Silvia E. Buntinx Dios http://amaltea.fmvz.unam.mx/textos/alimenta/MET_CHO_LIP_PRO2.pdf) Introducción: El metabolismo es el ensamble de las transformaciones moleculares y de transferencia de energia que se desarrollan sin interrupciones dentro de la celula o del organismo. Los procesos son ordenados, interviniendo procesos de degradación 17 (catabolismo) y de síntesis orgánica (anabolismo). Se puede distinguir el metabolismo basal (durante el sueño) y el metabolismo en actividad (actividad cotidiana).Toda actividad celular y del organismo requiere de energía, pero también, de nutrimentos específicos (proteinas, ácidos nucleícos, lípidos, minerales, vitaminas), que deben moverse a través de membranas, con frecuencia contra un gradiente de concentración, lo que implica un gasto importante de energía. Los niveles de energía y las concentraciones de nutrimentos deben estar disponibles constantemente y deberán satisfacer la tasa de actividad y sus variaciones. Los organismos deben regular sus actividades metabólicas económicamente para evitar deficiencias o excesos de productos metabólicos. El organismo debe ser flexible para poder alterar su metabolismo ante cambios significativos en su medio (variaciones en las concentraciones o en el tipo de nutrientes). Lección 36. Metabolismo de carbohidratos: Glucólisis y Vía Pentosa Fosfato Los carbohidratos de la ración proporcionan mas del 50% de la energía necesaria para el trabajo metabólico, el crecimiento, la reparación, la secreción, la absorción, la excreción y el trabajo mecánico. El metabolismo de carbohidratos incluye las reacciones que experimentan los de orígen alimentario o los formados a partir de compuestos diferentes a estos La oxidación de este tipo de glúcidos proporciona energia, se almacenan como glucógeno, sirven para la síntesis de aminoácidos no esenciales. Glucólisis (Vía de Embden-Meyerhof) La glucólisis es un proceso común a todas las células, es la principal vía metabólica de utilización de hexosas, principalmente glucosa pero también directamente de la fructosa y de la galactosa. El conjunto de las reacciones permiten oxidar parcialmente la glucosa para formar piruvato con el objeto de liberar energía para sintetizar ATP. Esta vía se desarrolla totalmente en el citoplasma celular en condiciones anaeróbicas o aeróbicas. Pueden considerarse dos fases dentro de esta vía. 1. La primera parte o fase preparativa, la glucosa es activada y para ello se emplean dos ATP. Los enzimas hexocinasa y glucosinasa son responsables de la conversión de glucosa a glucosa 6-P. La hexocinasa se encuentra en todos los tejidos, tiene una gran afinidad por la glucosa y otras hexosas, puede llevar a cabo la reaccion aun a bajas concentraciones del enzima y es inhibido por la glucosa 6-P. La enzima glucocinasa se localiza en el hígado y en las celulas β del páncreas, tiene una baja afinidad por la glucosa, por ello es efectiva cuando la glucosa se encuentra a elevadas concentraciones, no es inhibido por el producto y está ausente o sus concentraciones son muy bajas en los rumiantes. La formación de fructosa 1, 6-bi fosfato se lleva a cabo por la fosfofructocinasa. Esta enzima está presente solo en la glucólisis, así, constituye un sitio de control. La adrenalina, el glucagón, aumento en los ácidos grasos libres, el citrato, y el ATP inhiben su actividad. 2. En la segunda parte de la glucólisis o fase productora de energía, se lleva a cabo la generación de ATP. 18 La vía colateral de las pentosas (ruta de la pentosa fosfato) Esta via metabólica ni requiere, ni produce ATP, se desarrolla en el citoplasma de las células de tejidos con elevada actividad lipogenética (hígado, tejido adiposo, glándula mamaria, cerebro en desarrollo). La molécula de glucosa 6-fosfato será transformada en y una pentosa fosfato. Los carbonos de la pentosa se transferirán en piezas de 2 a 3 carbonos entre moléculas. Los productos finales pueden contener de 3 a 7 átomos de carbono que serán utilizadas posteriormente en la glucólisis (triosas fosfato), en la síntesis de aminoácidos (eritrosa 4-fosfato), en la síntesis de ac: nucleicos, NAD, FAD, y CoA. En esta vía se genera también NADPH, esta coenzima se utilizará para la síntesis de ácidos grasos de cadena larga, de colesterol, la hidroxilación de ácidos grasos y esteroides, mantenimiento de la glutation reducido (GSSG) en los glóbulos rojos. Gluconeogénesis Es la producción de azúcares a partir de sustancias diferentes a los carbohidratos (lactato, aminoacidos, propionato y glicerol). Esta vía permite tener una fuente alterna de glucosa, remover el lactato (producido por los glóbulos rojos y el tejido muscular) de la sangre, remover el glicerol producido por el tejido adiposo. Esta vía metabólica se activa ante la disminución de la glucosa sanguínea, en el cerdo su activacion es el ayuno: cerdo, 24 h, hombre 8 y en el pollo 2 h. En el rumiante es una vía constantemente activa. La gluconeogénesis se encuentra bajo control hormonal (insulina, glucagón y adrenalina). Los rumiantes son eficientes para realizar la gluconeogénesis y su aparato digestivo se ha adaptado a una falta de azúcar y almidón, por lo que la capacidad para el manejo de estos carbohidratos es limitada. Asi, los rumiantes absorben la mayoría de su carbono dietario digerido (energía) en forma de ácidos grasos volátiles (AGV). Los AGV son producidos por la fermentación bacteriana de los carbohidratos en el rumen y en menor cantidad en el intestino grueso, pueden contribuir con 70-80% de la energía total que el animal necesita. Lección 37. Metabolismo de Aminoácidos y proteínas Las proteinas funcionan como enzimas, para formar estructuras, pero además los aminoácidos pueden utilizarse como fuente de energía o como sustratos para otras rutas biosintéticas. En los animales superiores, los aminoácidos provienen de la proteina de la dieta o por recambio metabólico de proteína endógena. El exceso de aminoácidos se degrada parcialmente para dejar esqueletos de carbono para biosíntesis o se degradan totalmente para producir energía. Los aminoácidos son catabolizados a través de la remoción del nitrógeno (N), a través de dos rutas principales: la transaminación y la desaminación oxidativa. En la transaminación, un aminoácido dona su grupo amino al α-cetoglutarato (ciclo de Krebs) se forma un α-cetoácido y glutamato, el coenzima utilizado es principalmente el piridoxal fosfato. Esta reacción es reversible y se encuentra ampliamente distribuída en los tejidos, especialmente: cerebro, corazón, riñón, hígado. Solo la lisina, treonina, prolina e hidroxiprolina no sufren transaminación. La regeneracion del α-cetoglutarato se consigue 19 mediante la desaminación oxidativa del glutamato catalizada por la glutamato deshidrogenasa unida al NAD. El amoníaco resultante de la desaminación de, se transforma en urea en el hígado para detoxificarlo. En muchos órganos (cerebro, intestino, músculo esquelético), la glutamina es el transportador del exceso de N. En el músculo esquelético existe el ciclo glucosa-alanina (Ciclo de Cori) para transportar el amoníaco al hígado bajo la forma de alanina. La formación de urea involucra una serie de pasos de la ornitina en arginina. La urea se forma a partir de la arginina. El ciclo de la urea utiliza cinco enzimas: argininosuccinato sintasa, arginasa, arginosuccinato liasa (los tres se encuentran en el citosol), ornitina transcarbamoilasa y carbamoil fosfato sintasa (presentes en la mitocondria). El amonio libre formado en la desaminación oxidativa del glutamato se convierte en carbamoil fosfato, reacción catalizada por la carbamoil fosfato sintetasa I y que requiere dos ATP. El carbamoil fosfato transfiere su grupo amino a la ornitina y forma citrulina. Esta debe transportarse a través de la membrana mitocondrial al citosol, donde se formará la urea. En cada vuelta del ciclo de la urea se eliminan dos N, uno que se origina de la desaminación oxidativa del glutamato y el otro del aspartato. El fumarato es el vinculo entre el ciclo de la urea y el de Krebs. Después de la desaminación, el esqueleto de carbono de los aminoácidos puede ser utilizado para la producción de energía. El catabolismo de los aminoácidos involucra su conversión a intermediarios en el ciclo de Krebs, su conversion a piruvato o a acetil-CoA. Este último puede oxidarse en el ciclo de Krebs o puede convertirse en acetoacetato y lípidos. Los aminoácidos que forman acetoacetato son cetogénicos, ya que no pueden convertirse en glucosa. Los aminoácidos que forman α-cetoglutarato o acidos dicarboxílicos de cuatro carbonos estimulan el funcionamiento del ciclo de Krebs y son considerados glucogénicos. Lección 38. Betaoxidación de ácidos Grasos Los Ácidos grasos (AG) son los componentes principales de los lÍpidos complejos (triacilgliceroles, fosfolÍpidos). Los triacilgliceroles son la forma más importante de almacenamiento de energía en los animales. Este tipo de almacenamiento presenta sus ventajas, al oxidarse el C de los AG producen más ATP que cualquier otra forma de C, además, los lípidos están menos hidratados que los polisacáridos, por lo que ocupan menos espacio. Los AG se incorporan a las membranas celulares. El principal órgano de interconversión y metabolismo de lípidos es el hígado. Oxidación de los ácidos grasos Cuando el aporte de energía de la dieta es insuficiente, el animal responde con la señal hormonal, que se transmite al tejido adiposo por medio de la liberación de adrenalina, glucagón u otras hormonas. Estas se unen a la membrana de la célula adiposa y estimulan la síntesis. Los trigliceridos se hidrolizan a diglicéridos, liberando un ácido graso del carbono 1 o 3 del glicerol. Los diglicéridos y los monoglicéridos son hidrolizados rápidamente para producir ácidos grasos y glicerol. El ácido graso no esterificado sale a la sangre y se une a la albúmina para ser transportado a otros tejidos, y el glicerol 20 será utilizado por el hígado para la producción de glucosa. Los AG se oxidan en el carbono β, de ahí el nombre de β-oxidación y se degradan a ácido acético y un ácido graso con dos carbonos menos. La β-oxidación inicia con una reacción de deshidrogenación (acil-CoA deshidrogenasa), utilizando a FAD como coenzima. El producto de esta reacción es un enoil-CoA y . El enoil-Coa es hidratado por la enoil-CoA hidrasa, se produce un hidroxiacil-CoA. El grupo hidroxilo de este compuesto es oxidado por y la hidroxiacil-CoA deshidrogenasa, se produce β-cetoacil-CoA y NADH. El último paso es catalizado por una tiolasa, produciendo acetil-CoA y un acil-CoA, con dos carbonos menos que el sustrato inicial. Estos pasos se repiten hasta que en la última secuencia de reacciones el butiril-CoA es degradado a dos acetil-CoA. En los rumiantes, la oxidación de AG de cadena impar puede representar tanto como el 25% de sus requerimientos de energía. La oxidacion de un AG de 17 carbonos da por resultado 7 acetil-CoA y un propionil-CoA. El propionil-CoA es también un producto de la degradacion de valina e isoleucina. El propionil-CoA es convertido en succinil-CoA y será utilizado en el ciclo de Krebs. Lección 39. El ciclo de Krebs (CK) También denominado: ciclo del ácido tricarboxílico- CATC o del ácido cítrico-CAC La glucólisis y el ciclo de Krebs son consideradas las vías metabólicas centrales, participan en la degradación de casi todos los componentes que la célula es capaz de degradar y proveen el poder reductor y los materiales de construcción. El proceso general es el de metabolismo respiratorio aeróbico. En estas condiciones, el es el último aceptor de energía. En este ciclo se pueden mencionar dos procesos separados pero relacionados: 1. El metabolismo oxidativo, hay remoción y flujo de electrones de sustancias orgánicas y transferencia a coenzimas. 2. Hay reoxidación de las coenzimas a través de la transferencia de electrones acompañada directamente de la generacion de ATP. En anaerobiosis, la glucólisis es la fase inicial del catabolismo de la glucosa. Los otros componentes del metabolismo de respiración son el ciclo de Krebs (continuacion de la oxidación del piruvato), la cadena de transporte de electrones y la fosforilación oxidativa de ADP a ATP a través de un gradiente de protones generado en el transporte de electrones. El proceso completo genera de 36 a 38 moléculas de ATP/mol de glucosa, en cada vuelta del ciclo de Krebs entran dos moles de acetil CoA y se liberan 2 carbonos, lo que regenera la molécula de oxaloacetato (OAA). Lección 40. Foto-respiración 21 ACTIVIDAD FOTORRESPIRATORIA EN EL FRUTO DE MORA(Rubus glaucus) Alexandra Ortiz *; Jenny Rincón*Jairo Granados** *Ingenieras Agrónomas; **Docente, MSc; ECAPMA RESUMEN El fruto de mora de castilla (Rubus glaucus) es un fruto blando por lo que se considera altamente perecedero teniendo una vida útil de 3-5 días, en consecuencia se obtienen grandes pérdidas en la poscosecha. El principal objetivo de este trabajo fue la caracterización fisicoquímica y la evaluación de las enzimas Peroxidasa (POD) y polifenoloxidasa (PFO) en este fruto las que intervienen en el pardeamiento enzimático durante la maduración de los frutos; así como evaluar la efectividad de tres inhibidores el Acido ascórbico, Cloruro de Sodio y Cloruro de Calcio en tres concentraciones 1, 2 y 4%. Los frutos de mora de castilla (Rubus glaucus) fueron muestreados de dos fincas ubicadas en los municipios de San Bernardo y Silvania, los cuales fueron recolectados en dos índices de madurez 2 y 5 de acuerdo a la tabla de color diseñada por ICONTEC conocida como NTC 4106. Se encontró que en la mayoría de las variables no hubo diferencias significativas entre las fincas, pero si entre los índices lo cual es coherente ya que son frutos de dos índices con madurez fisiológica diferentes. Como mejor tratamiento en la inhibición de las estas enzimas evaluadas se encontró la aplicación del Cloruro de Calcio en las tres concentraciones al 1, 2 y 4% siendo la última concentración la más efectiva. Palabras claves: Fruto de Mora (Rubus Glaucus), Peroxidasa, inhibidores enzimáticos. INTRODUCCION La mora de Castilla (Rubus glaucus) es originaria de las zonas tropicales de América latina (Castellanos, 2003), en Colombia los principales departamentos productores son, Cundinamarca, Santander, Huila, Antioquia y Caldas donde se concentra el 95% de la producción (Rojas et al., 2004). El fruto de la mora (Rubus glaucus) está incluido en el grupo de lo que algunos autores denominan “frutos blandos” ya que sufre un rápido ablandamiento, igualmente, el pardeamiento de la pulpa se debe a la acción de enzimas como: la polifenoloxidasa (PFO), la cual en presencia de oxígeno cataliza la oxidación de numerosos compuestos como los fenoles y la peroxidasa (POD) la cual a niveles altos de peróxido de hidrogeno H2O2 oxida fenoles a quinonas generando pardeamiento por la reacción denominada polimerización (Candan, 2004). El fenómeno de pardeamiento de frutos y vegetales durante el crecimiento, colección, almacenamiento y procesado, es un problema de primera magnitud en la industria alimentaria y se reconoce como una de las principales causas de pérdida de calidad y valor comercial. Este pardeamiento produce cambios importantes tanto en la apariencia (colores oscuros) como en las propiedades organolépticas (sabor, textura) de alimentos comestibles (Mayer, 1987, citado por Sellés, 2007). La peroxidasa es una hemoproteina monomérica que cataliza la oxidacion de un amplio número de sustratos (fenoles, aminas, aromaticas e hidroquinonas) utilizando peroxido de 22 hidrogeno como cofactor a quinonas. Se encuentra en los peroxisomas, cloroplastos, vacuolas y en la pared celular (Narváez, 2002, citado por Hernandez, 2006). Es miembro de un grupo de enzimas llamadas oxido reductoras, la encontrada en plantas superiores, es la ferriprotoporfirina (hematina) en general reportada como EC. 1.11.1.7 (Daoudi, 2004). Hernández (2006) evaluó la actividad de peroxidasa según la técnica descrita por Dalisay y Kúc (1995), la cual se fundamenta en la determinación del cambio de Absorbancia por minuto a 436 nm, producido por la oxidación de un sustrato de la enzima como la o-dianisidina en presencia de peróxido de hidrógeno. La PFO cataliza la reacción dependiente de oxigeno que transforma o-difenoles en o-quinonas, esta transformación tiene lugar en dos pasos: hidroxilación de monofenoles en o-difenoles, y oxidación de estos o-difenoles a o-quinonas, que corresponden a las dos actividades consecutivas realizadas por la polifenoloxidasa (Pérez, 2003), esta actividad causa cambios en el color, sabor, en la calidad de los productos procesados y daños nutricionales (García, 2004). Esta es la razón fundamental por la que el contenido en fenoles y la actividad PFO se consideran determinantes en la calidad de los frutos y vegetales (Casado, 2004). Se sabe que las enzimas PFO y POD extraídas de diferentes tejidos vegetales pueden ser inactivadas por tratamientos térmicos y por ciertos compuestos químicos usados en la industria de los alimentos, como cloruro de sodio, sulfitos, ácido ascórbico, sacarosa, EDTA y ácido benzoico (Rivera et al., 2004). El objetivo de este trabajo fue el de evaluar y caracterizar física y químicamente los frutos de mora variedad Castilla (Rubus glaucus) y la actividad enzimática de peroxidasa (POD) y polifenoloxidasa (PFO), principales enzimas involucradas en el pardeamiento enzimático del fruto durante la maduración y senescencia y a su vez aplicar tratamientos permitidos por la industria alimentaria que inhiban su actividad, a fin de alargar la vida útil del fruto reduciendo las pérdidas en el manejo de poscosecha manteniendo la calidad comercial de estos frutos. MATERIALES Y METODOS MATERIAL VEGETAL Las muestras se recolectaron de dos fincas, la primera se encuentra localizada en el municipio de San Bernardo (F1) ubicado en el departamento de Cundinamarca (Colombia) con una temperatura promedio de 20 ºC, situada a una altura de 2300 m.s.n.m, y la segunda finca está ubicada en la vereda Noruega alta del municipio de Silvania (F2), Cundinamarca, a 1470 m.s.n.m. con temperatura media de 20ºC, la recolección de estas se hizo en dos índices de madurez 2 y 5 de acuerdo a la escala de color según la NTC 4106, se seleccionaron los frutos de forma homogénea por sus características físicas, sin daños mecánicos u ocasionados por plagas o enfermedades, para luego aplicar los respectivos análisis fisicoquímicos y enzimáticos. ANÁLISIS FÍSICO-QUÍMICO Para determinar el pH se realizó por medio de un pHmetro calibrado a temperatura 20°C donde se pesó 1 gr de fruto picado se le adicionó 10 ml de agua destilada, se agitó 5 min 23 y luego se procedió a medir el pH, se adicionó 40 ml de agua destilada, se agito 5 min y luego se procedió a titular con NaOH 0,1 N hasta alcanzar un pH de 8,5 en la solución, para la determinación de índice de acidez. Los grados Brix se midió por medio de un refractómetro digital, la firmeza se determinó mediante un Penetrómetro BERTUZZI midiendo la fuerza (Newtons) necesaria para penetrar la pulpa y las medida de longitud y diámetro se tomaron utilizando un Calibrador Vernier digital. Las anteriores variables se evaluaron en 20 frutos enteros para cada índice. ACTIVIDAD ENZIMATICA Extracción de la enzima. En la técnica que se implementó se utilizó los polvos de acetona que permiten que el extracto presente una alta actividad con una mayor estabilidad, porque con la acetona se retiran los fenoles, β-carotenos y ácidos orgánicos del extracto disminuyendo así la reacción de las enzimas (Baquero, 2005). Se utilizó 5 g de fruta picada homogenizados con 25 ml de acetona 4°C se filtró al vacío, y el retenido (polvos de acetona) fue lavado dos veces con 25 ml de acetona. Los polvos de acetona fueron suspendidos en 25 ml buffer de fosfato pH 7, luego, la suspensión fue centrifugada a 10000 rpm durante 30 min, el sólido se desechó, el sobrenadante fue mantenido refrigerado. Las medidas de actividad enzimática fueron desarrolladas a las condiciones de temperatura, en las que se obtuvieron las máximas respuestas, evaluadas en reportes anteriores y en este trabajo. Medida de Actividad de Peroxidasa. La actividad de esta enzima se estimó por espectrofotométria del incremento de la absorbancia a 470 nm cada 15 s durante 300 s, utilizando como sustrato el guayacol. La mezcla con un volumen final de 4 ml, contenía 1 ml de Buffer Fosfato pH 7, 2 ml de H 2O2 0.03 M en Buffer Fosfato pH 7, 1 ml de Guayacol y 2 ml de extracto diluido, manteniendo la mezcla a 22°C. Para el blanco se utilizó Buffer Fosfato pH 7. Una unidad de actividad (UPOD) se definió como el cambio en una unidad de absorbancia por min. La actividad específica se expresó en unidades de actividad por mg de proteína (UPOD/min/mg proteína). Medida de Actividad de Polifenoloxidasa. Para medir la actividad de esta enzima se empleo como sustrato el catecol efectuando la lectura de absorbancia a 420 nm cada 15 s durante 300 s. La mezcla de reacción consistió en 2 ml de Catecol 20 mM en Buffer Fosfato pH 7, 0.8 ml de Buffer Fosfato pH 7 y 0.2 ml de extracto enzimático. Se incubo la mezcla durante 3 minutos a una temperatura de 20°C. El blanco se utilizó el mismo que en POD. La unidad de actividad se manejó igual que en la Peroxidasa. INHIBIDORES Se evaluó el efecto en los frutos recolectados de dos estados de madurez el Cloruro de calcio (CaCl2), Cloruro de sodio (NaCl) y el Ácido Ascórbico en tres concentraciones al 1%, 2% y 4% en cada tratamiento sometido a tres repeticiones. La descripción se presenta en el cuadro 1. El tiempo de inmersión se determinó con ensayos previos utilizando 2 tiempos a 10 y 20 min en donde se tomó el pH, IA y ºBrix de los frutos para 24 observar los efectos del tiempo en estas características, al no arrojar diferencias se determinó sumergir el fruto por 10 min para cada tratamiento. Cuadro 1. Descripción de tratamientos. TRATAMIENTO DESCRIPCION T1 Ácido Ascórbico 1% T2 Ácido Ascórbico 2% T3 Ácido Ascórbico 4% T4 Cloruro de sodio 1% T5 Cloruro de sodio 2% T6 Cloruro de sodio 4% T7 Cloruro de calcio 1% T8 Cloruro de calcio 2% T9 Cloruro de calcio 4% T10 Testigo ANÁLISIS ESTADÍSTICO Para el análisis de los resultados se utilizaron los elementos de la estadística descriptiva como promedios, desviación típica estándar y coeficiente de variación. A partir de las medidas de actividad de las diferentes variables se calcularon los valores promedio de las tres repeticiones, se efectuó los ANAVA y se evaluaron las diferencias entre tratamientos por la prueba Duncan. Todo este tipo de análisis estadístico se desarrolló con base en el programa SPSS, versión 10 para Windows. DISCUSION Y RESULTADOS PROPIEDADES FISICAS Y QUIMICAS EN FRUTOS DE MORA (Rubus glaucus) Se encontraron en las variables físicas y químicas diferencias estadísticas entre los índices de los 40 frutos evaluados de cada finca, lo cual es coherente ya que un índice tiene un mayor grado de madurez y adquiere ciertas características de un fruto más maduro, como el obtener mayores dimensiones y peso en los frutos del índice 5, como se puede observar en la tabla 1 en la que se presenta todos los datos obtenidos para los dos índices; se presenta una menor firmeza en el índice 5 (Grafico 1) ya que el fruto al estar más maduro es más blando a consecuencia de procesos químicos como la perdida de turgencia, la degradación de la pectina de la pared celular en las que interviene enzimas como la PME y PGA; el contenido de azúcar también es mayor en este índice (Grafico 2) debido a que en las frutas ocurre un comportamiento general en donde el aumento del contenido de azúcar es a medida que avanza su madurez en donde el almidón o ácidos orgánicos se han hidrolizado en azucares como fructosa, sacarosa y glucosa (Rojas et al., 2004), por tanto se va a tener una mayor concentración de azucares en los últimos índices de madurez lo que se puede deber a una mayor translocación de fotosintetizados 25 hacia el fruto o a que ciertos polisacáridos de reserva que forman parte de las substancias cementantes de las células se hidrolicen (Hernández, 2006). Tabla 1. Propiedades físicas y químicas encontradas en el índice 2 y 5 en el fruto de mora (Rubus glaucus). INDICE 2 VARIABLE MIN-MAX Longitud (mm) Diámetro (mm) Peso fresco (g) Firmeza (N) pH Grados Brix Índice de Acidez Índice de Madurez MEDIA INDICE 5 21 - 33 16 - 22 2,84 6,26 DESV ESTAN 2,37 1,15 2,86 - 6,55 3,97 0,61 5,11 - 11 1,56 10 - 82 2,46 - 2,27 4,4 - 8,6 3,26 - 4,9 26,55 18,45 4,8 49,42 15,16 0,06 0,49 0,22 7 - 27,2 1,86 - 3,29 5 - 10,7 0,77 - 2,33 29,88 21,43 7,48 17,36 4,8 0,34 1,18 0,24 1-2.19 1.58 0.22 0.77-6.52 5.09 0.24 60 49,34 INDICE 2 INDICE 5 40 30 20 15,71 10 0 F1 19,02 GRADOS BRIX FIRMEZA (N) 50 49,5 9 8 7 6 5 4 3 2 1 0 MIN-MAX MEDIA 21 - 44 15 - 24,24 2,57 7,41 DESV ESTAN 4,04 1,26 1,36 8,06 INDICE 2 6,76 5,93 6,6 INDICE 5 F2 F1 F2 FINCAS FINCAS Gráfica 1 y 2. Media de Firmeza y Grados Brix por fincas e índices del fruto de mora (Rubus glaucus). ACTIVIDADE ENZIMÁTICA Peroxidasa (POD). La actividad de Peroxidasa se encontró desde 0,16 a 0,83 UA/min/mg de proteína este resultado es acorde a lo encontrado por Baquero (2009), quien encontró una actividad máxima de 0,9 UA/min/mg en tomate de árbol y por Baquero (2005) en pitaya valores de 0,03 hasta 3,1UA/min/mg de proteína, a diferencia de lo reportado por estos autores el fruto de mora presenta una baja actividad. Como se aprecia en la gráfica 3 existe una mayor actividad de esta enzima en los frutos con una maduración más avanzada lo cual es acorde ya que en los últimos índices de 26 UA/min/mg de proteína madurez se empieza a evidenciar el pardeamiento enzimático por la aparición de daños en las membranas debido al procesos de senescencia, daños por insectos, aparición de patógenos (principalmente Botrytis) labores del cultivo (manejo cosecha, poscosecha) lo que hace más fácil el contacto sustrato-enzima. 0,5 0,44 0,42 0,4 0,29 0,3 INDICE 2 0,23 INDICE 5 0,2 0,1 0 F1 FINCAS F2 Gráfica 3. Actividad de Peroxidasa en el furo de mora (Rubus glaucus) sin la aplicación de tratamientos UA/min/mg de proteina Polifenoloxidasa (PFO). La actividad de esta enzima se encontró entre los rangos de 0,03 y 0,24UA/min/mg de proteína los cuales son más bajos que lo reportado por Gasull (2006), encontrando para la pera valores de 1 UA/min/mg de proteína y para manzana 0.9 UA/min/mg de proteína, lo cual es lógico ya que estas frutas son reportadas como las más sensibles al pardeamiento enzimático. 0,1 0,08 0,06 0,079 0,074 0,062 0,044 INDICE 2 INDICE 5 0,04 0,02 0 F1 FINCAS F2 Gráfica 4. Actividad de Polifenoloxidasa en el furo de mora (Rubus glaucus) sin la aplicación de tratamientos. En cuanto a la actividad de esta enzima no tuvo un comportamiento similar en los índices de las dos fincas (Grafico 4), existe gran heterogeneidad en los resultados de las publicaciones referentes a parámetros que afectan a la actividad enzimática de la polifenoloxidasa (pH óptimo, latencia, especificidad por el sustrato, etc.) entre especies y dentro de la misma especie, entre variedades y diferentes estados de desarrollo (la actividad enzimática es más elevada en frutos jóvenes que en los maduros) (Robards et al., 1999, citado por Pérez, 2003), como también se reportó que la actividad de la PFO es alta en los tejidos en desarrollo y en los tejidos meristemáticos, pero disminuye al madurar 27 las células (Cary et al., 1992; Shahar et al., 1992 citado por Gooding, 2001) como se observa comparando los dos índices de la F1.En otra investigación con frutos de guanábana, determinaron que existe una correlación positiva entre la actividad de la enzima y los compuestos fenólicos, en la medida que avanza la maduración de frutos verdes a maduros e igualmente señalan que la concentración de la enzima en el punto de maduración óptima del fruto es mínima (García, 2004). Capítulo 9. Metabolismo especiales aplicados Lección 41. Metabolismo secundario de las plantas Metabolitos secundarios en las especies Gliricidia sepium y Tithonia diversifolia Luz Elena Santacoloma Varón 1 y Jairo Enrique Granados2 ; 1,2 Docentes investigadores, ECAPMA, UNAD Resumen Se recolectaron muestras de 2 especies forrajeras (hoja peciolo) de las especies Gliricidia sepium, y Tithonia diversifolia en diferentes condiciones edáficas (departamentos del Valle del Cauca y Cesar) y se evaluó su contenido de polifenoles totales, taninos totales, taninos condensados y saponinas. El propósito fué detectar el efecto de las condiciones de conductividad eléctrica sobre la concentración de estas fitobiomoléculas secundarias metabolizadas por la célula vegetal. Introducción El uso de leguminosas y asteráceas en sistemas de alimentación animal es una práctica que se ha venido implementando desde hace varios años en muchas regiones del trópico; por el alto contenido de proteína y minerales que contienen. Entre las leguminosas se destaca la arbórea Gliricidia sepium, especie poco valorada como fuente suplementaria de proteína, especialmente en la época seca, cuando los forrajes son escasos y de mala calidad (Palma et al 1999). Entre las asteráceas sobresale la Tithonia diversifoli, planta de gran adaptabilidad a diversos pisos térmicos y con alto contenido de nutrientes para rumiantes y no rumiantes. No obstante a lo anterior, es importante tener en cuenta un factor que en cierta medida puede constituir una limitante en el uso de estas plantas como material forrajero y es la presencia de metabolitos secundarios que pueden afectar la 28 digestibilidad, los cuales no solamente pueden interferir con los procesos de digestión y absorción de los nutrientes, sino que, previo paso al torrente sanguíneo, pueden desencadenar variados efectos sistémicos en los animales (Jackson et al., 1996). El contenido de varios tipos de estos metabolitos secundarios, está influenciado por el genotipo de la planta (la especie y la variedad), las características ambientales (radiación solar y disponibilidad de agua), la velocidad de crecimiento, la madurez, la condición nutricional del suelo, la depredación y las enfermedades (Waterman y Mole 1995). Así mismo, la aparición de los compuestos secundarios está relacionada con los mecanismos de defensa de la planta y los efectos del suelo y el clima (Harborne, 1993). Kumar 1997 plantea. que los distintos compuestos que puede producir una especie presentan una determinada distribución dentro de los órganos, tejidos y células de una planta, y ello responde frecuentemente a las influencias ambientales. Entre los metabolitos secundarios, se destacan los taninos, por su abundancia en la naturaleza, y particularmente en los forrajes; Posada (2005) los define como un grupo de polifenoles solubles en agua, de relativo alto peso molecular, con presencia en casi todas las plantas, particularmente en las dicotiledóneas, de las cuales hacen parte las leguminosas. Leinmúller 1991 explica que su distribución al interior de ellas varía entre especies y a nivel de célula vegetal se encuentran en las vacuolas citoplasmáticas o en la pared celular. La síntesis de taninos, por su parte, se presenta a partir de esqueletos de carbono del metabolismo primario, como ocurre con los rebrotes más jóvenes de la planta, que también necesitan las reservas de carbono para su crecimiento (Haukioja et al 1985). Así mismo, los taninos tienen la capacidad para formar enlaces químicos más o menos estables al interactuar con proteínas, carbohidratos y otras macromoléculas de los alimentos, o con las enzimas digestivas; además se ha demostrado que los taninos pueden ser tóxicos para bacterias; en los rumiantes se refleja en cambios de la digestión ruminal de proteínas y carbohidratos, y en la producción de ácidos grasos volátiles ( Makkar et al 1988, Reed 1995). Para Norton (1997), los aspectos que inciden en el contenido de taninos en los árboles y que tienen relación con la planta son; la composición genética, la especie, el grado de madurez, el estado de crecimiento y la defoliación por herbívoros. Según este mismo autor los factores ambientales que mayor incidencia tienen en el contenido de taninos son; la estación climática, la humedad ambiental y luminosidad. Otros aspectos inherentes a la planta que inciden en el contenido de taninos es la parte de ésta a la cual pertenece, siendo diferente el contenido de estos metabolitos en el peciolo, que en la hoja. Por su parte, Oncina et al 2000 y Fedoreyev et al., 2000 han demostrado que las partes de la planta, presentan productividades diferentes de metabolitos secundarios y que se acumulan cantidades de metabolitos diferentes de acuerdo a la parte de la planta a la cual corresponde. Los taninos pueden dividirse, según su estructura química, en taninos condensados (proantocianidinas) , y taninos hidrolizables (derivados de los ácidos gálico y elágico). Al 29 respecto, Romero y Palma 2001 encontraron que factores como el pastoreo provocan una disminución en el contenido de taninos condensados y de los fenoles totales en G sepium, probablemente, porque se presenta una mayor competición por reservas para la generación de hojas, más que para la producción de taninos condensados. El efecto de la época, también varió según la naturaleza del tanino, sin mostrar tendencias consistentes. Los mismos autores reportan que los taninos adheridos a proteína representaron alrededor de 70-75% de los taninos condensados totales, siendo más alto los valores en época de secas que en la de lluvias. Los taninos condensados pueden llegar a producir efectos depresivos sobre el consumo y la digestibilidad de la materia seca y el nitrógeno, ya que provoca saciedad y limita por lo tanto el consumo de materia seca (Flores et al 1999; Gutiérrez et al 2003). Gershenzon 1984 reporta que la disponibilidad de agua contribuye a la producción de taninos condensados; las plantas al encontrarse en periodo de escasez de agua, cierran sus estomas y restringen el proceso de fotosíntesis. De este modo, se esperaría una relación negativa entre el estrés hídrico y la producción de compuestos fenólicos. Por su parte, la leguminosa G sepium puede ser considerada como una de las leguminosas tropicales con menor contenido de taninos condensados, en comparación con otras como Flemingia macrophylla con 388 g/kg (Cano et al 1994), Desmodium ovalifolium 238 g/kg, Acacia mangium 100 g/kg, Callyandra sp 194 g/kg (Jackson y Barry 1996). Esta concentración pudiera explicar el comportamiento de los animales de desarrollar un grado de gustocidad medio para G. sepium (Kaito et al 1996). Por tanto, cconsiderando todos los análisis químicos realizados, la hoja de Matarratón es un excelente forraje para dietas tropicales, por que presenta los compuestos nutricionales más altos, una buena tasa de degradabilidad y los principios tóxicos más bajos que los otros forrajes estudiados Respecto a la especie forrajera Tithonia diversifolia, en un análisis de metabolitos secundarios realizado por Rosales (1992), no se encontraron ni fenoles ni taninos, mientras que Vargas (1994) encontró un bajo contenido de fenoles y ausencia de saponinas. Murgaruline et al (1993) encontraron en esta especie un compuesto citotóxico Tagitinin. Dutta et al (1993), citado por Rosales encontraron Hispidulin, además del compuesto Tagitinin, a los cuales le atribuyen el efecto repelente contra los insectos. Las saponinas son otro tipo de metabolitos secundarios conformados por compuestos que en forma glucosídica (como esteroides reguladores del crecimiento) pueden generar en las plantas alto crecimiento, desarrollo, rápida recuperación ante la poda y brotes abundantes (Ashok., K.J Vincent R.M y Nessler 2000. También Kumar 1997, en dependencia de las concentraciones y las estructuras químicas específicas los compuestos saponínicos pueden constituir factores antinutricionales en rumiantes y monogástricos por conferirle a los forrajes un sabor amargo, provocar espumas consistentes, e interferir en la absorción de los alimentos No obstante también puede tener un efecto positivo en el metabolismo digestivo, al generar complejos con otros metabolitos secundarios con características tóxicas (Makkar., 1997). 30 En consecuencia, es necesario realizar investigaciones sobre los efectos de los factores ambientales, como son los edáficos, en la producción de metabolitos secundarios, en las interacciones suelo- planta- así como en la determinación de los factores que inciden en su concentración. Para ello se parte de una hipótesis fundamental y es que el contenido de metabolitos secundarios de las plantas varía en función de las características del suelo, expresadas en variables como la conductividad eléctrica. Metodología Zona de muestreo La recolección del material vegetal y de suelos se realizó en fincas ubicadas en la zona central del Valle, en el municipio de la Paz y de Pueblo Bello en el Cesar con las condiciones climáticas que se presentan en la Tabla 1. Tabla 1. Condiciones climáticas de las regiones donde fueron tomadas las muestras de forraje Condiciones climáticas Zonas de muestreo Temperatura Precipitación promedio (Grados promedio centígrados) lluvia) Tuluá (Valle del 27 Cauca) San Diego (César) 29 Pueblo Bello 21 (César) (mm Altura (m.s.n.n) 1200 950 1311 500 - 1200 Fuente: Plan de desarrollo de los municipios (2011) Recolección de las muestras: La fracción vegetal (hojas y tallos tiernos) de Gliricidia sepium y Tithonia diversifolia fue colectada a partir de plantas adultas en los tres sitios mencionados durante el mes de agosto de 2011 en el cual predomina tiempo seco en los tres lugares. El material fue llevado al laboratorio de Nutrición Animal de la sede Nacional José Celestino Mutis de la UNAD y fue sometido a proceso de secado a temperatura ambiente, en un lugar ventilado durante 72 horas. Posteriormente, las muestras fueron molidas, tamizadas y se extrajeron los extractos para realizarse el análisis de los metabolitos secundarios. Preparación y extracción del material para análisis fitoquímico Las muestras se secaron a la sombra durante 30 días, posteriormente se molieron hasta menos de 1 mm de tamaño de partícula en un molino de martillo para laboratorio. Se prepararon 3 fracciones (etanol, eter dietílico y agua) con 10 g de cada muestra y finalmente se efectuó un tamizaje fitoquímico de acuerdo a Cuéllar (1991). Por último, se cuantificaron las cantidades de polifenoles totales, taninos totales y taninos condensados 31 presentes en las muestras, utilizando etanol como solvente y tomando como base las técnicas descritas por AOAC (1995). Métodos analíticos para los fitometabolitos estudiados Los métodos utilizados se describen en el cuadro 1 Cuadro 1. Identificación de variables analizadas y la técnica analítica utilizada Variable Técnica analítica Fuente Polifenoles Totales (PFT) Espectrofotometría con Dewanto reactivo de Folin Ciocalteu al. (2002), et Terrill et al. Taninos condensados (TC) Taninos Totales Espectrofotometría con n –butanol -HCl Folin-Denis Ciocalteu y Folin Posada, Et al 2006 Fuente: Laboratorio de Nutrición animal UNAD., (2011) Para la prueba de las saponinas se tomó 1 mL de metanol y se añadieron 9 mL de agua, para luego filtrarse esta solución. Se tomó 1 mL de este filtrado en un tubo de prueba pequeño, se agitó vigorosamente por 30 segundos y se dejó en reposo durante 15 minutos. La espuma sobrenadante indica la presencia de saponinas en el forraje. La proporción de saponinas se midió de acuerdo a la altura (h) de la espuma sobrenadante: Tabla 1.Determinación de saponinas según la altura de la espuma Rango h (mm) 0.0 0.1-5.0 5.1-9.0 9.1-14 Mayor de 14 Indicador Prueba negativa Contenido muy bajo Contenido bajo Contenido moderado Contenido alto Fuente: Cuéllar (1991) Con respecto a los análisis de pH y conductividad eléctrica de las muestras de suelo se utilizó la técnica descrita en IGAG (2009) Los resultados obtenidos para cada variable se sometieron a un análisis de varianza en doble vía, con el fin de determinar diferencias estadísticas entre zonas muestreadas y especies evaluadas. Adicionalmente, se realizó un análisis de correlación múltiple, utilizando el coeficiente de Pearson, con el propósito de detectar las posibles correlaciones entre conductividad eléctrica, pH y el contenido de los metabolitos 32 secundarios cuantifcados en las especies muestreadas. Para lo anterior, se utilizó el paquete estadístico SPSS, versión 11.5. Lección 42: Bioquímica de la Madera La madera es una mezcla de tres polímeros naturales: celulosa, lignina y hemicelulosas, que guardan una relación aproximada de 50:25:25, según la especie, las variaciones biológicas, tales como las diferencias genéticas que pueda haber dentro de una especie, y las condiciones de crecimiento. La celulosa y las hemicelulosas son polímeras de hidratos de carbono formados a partir de moléculas de monosacáridos, y la lignina es un polímero de unidades fenilpropánicas (Browning, 1963). La celulosa es un polímero de la glucosa de cadena larga que únicamente se diferencia del almidón, que es también un polímero glucósico, por la configuración de las moléculas glucósicas. El carácter fibroso de las células leñosas es el resultado de la disposición lineal, orientada y cristalina de su elemento más abundante, la celulosa. Las hemicelulosas son polímeras de cadena más corta o ramificada de sacáridos con cinco átomos de carbono (pentosas), tales como la xilosa, o de seis átomos de carbono (hexosas) aparte de la glucosa. Son de naturaleza amorfa y sirven, con la lignina, para formar la matriz en la cual se incrustan las fibrillas de celulosa. Aunque la estructura de la celulosa es la misma en las diferentes especies las hemicelulosas varían considerablemente entre especies, especialmente entre las frondosas y las coníferas. Las hemicelulosas de las frondosas son, en general, más ricas en pentosas mientras que las hemicelulosas de las coníferas suelen contener más hexosas. La lignina, que es el tercer componente de la membrana celular, es un polímero tridimensional que se forma a partir de las unidades fenilpropánicas que se han desarrollado aleatoriamente formando una molécula grande y compleja con muchas diferentes clases de ligamentos entre los bloques estructurales. La estructura de la lignina también varía según se trate de una especie frondosa o conífera. Los grupos fenílicos de la lignina de frondosas se sustituyen más con grupos metóxilos que los de la lignina de coníferas. Una de las consecuencias de esta diferencia es que las ligninas de frondosas están menos reticuladas y son más fáciles de disolver durante la conversión a pasta. La lignina actúa de cemento entre las fibras leñosas y como agente endurecedor entre las fibras en la producción de pasta de madera química. Se disuelve por varios tratamientos químicos, quedando la celulosa y las hemicelulosas en forma fibrosa. Algunas hemicelulosas se pierden durante este proceso a causa de su menor peso molecular, mayor solubilidad y más fácil hidrolización. Además de los componentes poliméricos del tabique celular que constituyen la fracción principal de la madera, hay diferentes especies que contienen diversas cantidades y clases de materiales extraños que se llaman, en conjunto, extractivos. En las coníferas, se encuentran con frecuencia cantidades considerables de resinas que consisten tanto en 33 ácidos grasos como en los llamados ácidos resinosos que se derivan de terpenos simples tales como la trementina y el aceite de pino. Los taninos son fenoles polihidroxílicos que se encuentran en el duramen y corteza de muchas especies. El caucho se obtiene de la corteza interior de ciertos árboles, en forma de látex. También pueden obtenerse de diversas especies aceites aromáticos y azúcares solubles. El aceite de cedro y el azúcar de arce son ejemplos bien conocidos. 43. Productos químicos derivados de la madera Tomado de: IRVING S. GOLDSTEIN, profesor del Departament of Wood and Paper Science, de la Universidad del Estado de Carolina del Norte, Estados Unidos. Además de la celulosa, que es el polímero más empleado hoy día y que se utiliza principalmente en su estado natural fibroso después de extraído, se siguen empleando todavía cantidades considerables de los llamados productos «silviquímicos» (Goheen, 1972) a pesar de la preponderancia de las sustancias químicas derivadas del petróleo, o productos «petroquímicos». Licores de pulpación. Los fragmentos químicos de los polímeras del tabique celular que quedan en solución después de la fabricación de la pasta pueden, con frecuencia, aislarse de los licores de la fabricación de pasta y utilizarse. En general, los licores resultantes de la fabricación de pasta por medios alcalinos se queman durante la recuperación de los productos químicos empleados, pero los licores resultantes de la fabricación de pasta al sulfito suelen tratarse para obtener subproductos útiles. La lignina sulfonada puede precipitarse en forma de sulfonatos de lignina y utilizarse como productos tánicos, adhesivos, aglutinantes, dispersantes, etc. Los sacáridos contenidos en el licor de sulfito agotado se fermentan con levadura para producir alcohol etílico y suplementos de forrajes y piensos. Mediante la oxidación alcalina suave de los sulfonatos de lignina, se obtiene vainillina para aromatizantes y odorantes. La lignina de álcali obtenido del sulfato o del licor negro kraft se precipita y utiliza como diluente de resinas, para refuerzo del cancho y la estabilización de emulsiones. Entre los productos volátiles obtenidos del licor negro kraft figuran el dimetil sulfuro, dimetil sulfóxido y dimetil sulfona, que son útiles como disolventes y reactivos químicos. Talol. La industria de calafateantes navales ha quedado en gran parte desplazada por la recuperación de los componentes oleorresinosos de la madera resultantes del procedimiento de fabricación de pasta kraft (Uprichard, 1978; Zinkel, 1975). Algunas esencias volátiles como la trementina se recuperan de los gases de alivio expulsados de los digestores. El licor resultante de la fabricación de pasta con álcali convierte a los ácidos grasos y a los ácidos resinosos en sales sódicas que se despuman del licor negro concentrado y se acidifican para producir talol crudo. 34 Hidrólisis de la madera. La hidrólisis de la madera, o sea la conversión de los polímeras de carbohidratos que contiene la madera en monosacáridos mediante reacción química con agua en presencia de catalizadores ácidos, es un procedimiento que se conoce desde hace 150 años y que se ha empleado en escala comercial en los Estados Unidos durante la primera guerra mundial, en Alemania durante la segunda guerra mundial, y se utiliza actualmente en la U.R.S.S. El producto principal es la glucosa, que puede convertirse después en etanol o levadura. Polímeros celulósicos. La celulosa química de gran pureza, o pasta soluble, es el material de partida de derivados poliméricos de la celulosa tales como el rayón y el celofán (que son ambos celulosas regeneradas); ésteres celulósicas tales como el acetato y el butirato para la producción de fibras; películas y aplicaciones de moldeo, y éteres celulósicas tales como la carboximetilcelulosa, la etilcelulosa, y la hidroxietilcelulosa, que se utilizan como gomas. Extractivos. Todavía se sigue obteniendo de los tocones de pino, por destilación al vapor o por extracción, cierta cantidad de trementina y colofonia. La arabinogalactana, que es una goma hemicelulósica extraída del alerce, se utiliza como sucedáneo de la goma arábica. Los ácidos fenólicos extraídos de la corteza de varias coníferas se emplean como diluentes para adhesivos resinosos sintéticos y como aglutinantes y dispersantes. Las ceras que se extraen de la corteza del abeto Douglas pueden utilizarse en las aplicaciones generales de la cera, y el caucho natural sigue siendo un material importante. Productos químicos que se obtendrán de la madera En el futuro, la utilización de la madera para la obtención de productos químicos revestirá dos categorías principales: por una parte, la extensión y ampliación de los procedimientos actuales, y por otra, la transformación de los polímeras de la membrana celular en materias primas químicas de poco peso molecular. Además de la ampliación material de las operaciones actuales, se persigue también su extensión a otros aprovechamientos afines en la utilización de los productos y extractivos. Los ejemplos que aquí se citan son ilustrativos y no Imitadores. Los extractivos obtenidos de la corteza y el leño tienen un potencial mucho mayor que el que indica su aprovechamiento actual (Hillis, 1978; Laver, 1978). Los polímeras celulósicas podrían tener una mayor importancia si se pudieran reducir los costos de energía y mejorar sus propiedades (Allan, 1978; Goldstein, 1977). Se puede estimular la producción de oleorresina de los pinos aplicando herbicidas (Roberts, 1973). Se pueden obtener polímeros hidrocarbúricos con el cultivo comercial de nuevas plantas (Calvin, 1978), y producir fenoles de poco peso molecular a partir de la lignina, subproducto de los licores de la fabricación de pasta (Goheen, 1971; Benigni y Goldstein, 1971). Es posible recuperar ácidos sacarínicos del licor negro kraft (Sarkanen, 1976). El follaje puede producir aceites esenciales, clorofila y proteína (Barton, 1978). 35 Conversión de los polímeros de la membrana celular. La porción principal de la madera consiste en los componentes del tabique celular. En cuanto a volumen, esta fuente de materia prima supera con mucho a los elementos extractivos o subproductos químicos y representa un recurso potencial para satisfacer todas las necesidades químicas en sustitución de los productos petroquímicos. Se puede lograr una producción en gran escala de sustancias químicas, importantes desde el punto de vista industrial, derivadas de la lignocelulosa, siguiendo diversos procedimientos (Goldstein, 1976a). Estas cantidades industriales pueden proveer los bloques estructurales químicos fundamentales para la conversión en polímeros sintéticos (Goldstein, 1975a). Madera total. Los polímeros del tabique celular en su estado natural mixto pueden descomponerse en preparados más simples mediante tratamientos rigurosos que implican temperaturas altas y también, en algunos casos, presiones elevadas. Estos procedimientos no son de por si selectivos y son idénticos a los que se emplean con buenos resultados para la transformación en productos químicos de los desperdicios sólidos orgánicos o del carbón. En la gasificación, la madera se calienta a temperaturas de hasta 1000°C para formar una mezcla de monóxido de carbono y de hidrógeno como principales productos (Prahacs et al., 1971). Como subproductos se obtienen pequeñas cantidades de etileno, acetileno, propileno, benceno y tolueno. El monóxido de carbono y el hidrógeno que se forman de igual manera que en la gasificación del carbón se pueden: (a) tratar más aún para obtener hidrógeno con vistas a la producción de amoniaco; (b) convertir por catálisis en metanol; (c) enriquecer con hidrógeno y someter a reacción para formar metano, o (d) convertir por catalización en una mezcla de hidrocarburos alifáticos por el procedimiento FisherTropsch. La madera se licúa por reacción con monóxido de carbono y agua a una temperatura de 350-400°C y a una presión de 4000 libras por pulgada cuadrada (280 kg/cm 2), en presencia de varios catalizadores (Appell, 1971). Se produce un aceite viscoso (rendimiento: 40-50%) que puede tratarse ulteriormente para convertirlo en productos químicos de la misma forma en que los productos petroquímicos se derivan del petróleo. La pirolisis, o degradación térmica de la madera en ausencia de aire u oxigeno, transforma la madera en carbón de leña, gas y aceite (Saltes, 1978; Wender, 1974). El rendimiento relativo de cada producto dependerá de las condiciones de la pirolisis y de la composición del substrato, pero a una temperatura de 900°C las cifras típicas serían 2535% de carbón de leña, 3045% de gas y hasta 8% de alquitrán y aceite. El gas consiste principalmente en hidrógeno, monóxido de carbono y metano, mientras que el alquitrán y el aceite contienen elementos aceitosos ligeros como el benceno y el tolueno, así como preparados mixtos de temperatura de ebullición más elevada. Celulosa. La transformación selectiva de la celulosa de polímero glucósico en glucosa monomérica puede lograrse por diversos medios. La hidrólisis, para obtener glucosa, puede catalizarse bien sea por ácidos o enzimas, pero ningún procedimiento resulta tan 36 fácil como la hidrólisis del almidón debido al carácter cristalino de la celulosa. El contenido de lignina no impide la hidrólisis ácida, pero una celulosa que contenga mucha lignina será resistente a la hidrólisis enzimática, por lo cual habrá que delignificar en parte la madera o molerla fina para someterla a la hidrólisis por enzimas. La hidrólisis ácida con ácido diluido a elevadas temperaturas produce la descomposición de parte de la glucosa formada, que se convierte en hidroximetilfurfurol (Harris, 1975), lo que limita al 50% aproximadamente el rendimiento neto de azúcar. La hidrólisis ácida fuerte a temperaturas más bajas puede dar rendimientos casi cuantitativos de glucosa (Kusama, 1960). Shafizadeh (1978) ha demostrado que la destilación en seco de la celulosa a 400-500°C rinde casi 80% de un alquitrán que contiene principalmente levoglucosana que puede convertirse en glucosa con un rendimiento del 50% basado en celulosa. Para evitar toda contaminación y reacción con otros productos de descomposición, se necesitaría celulosa limpia de otros elementos del tabique celular. La transformación de la celulosa en glucosa es el primer paso en la utilización química en gran escala de la celulosa. La fermentación de la glucosa para obtener etanol mediante técnicas industriales acreditadas de alto rendimiento ofrece grandes perspectivas. El etanol es un importante producto químico industrial que se produce actualmente por hidratación del etileno. Puede también tener una amplia aplicación como combustible para motores de combustión interna. En la deshidratación del etanol para obtener etileno, o sea la reacción inversa a la actual formación de etanol a partir de etileno del petróleo, también el rendimiento es elevado. De igual forma, del etanol se obtiene fácilmente butadieno por procedimientos industrialmente acreditados, pero que, debido a la baratura del petróleo, han quedado anticuados. La transformación de la glucosa vía etanol en etileno y butadieno representa la principal utilización potencial de la celulosa para obtener productos químicos, debido a la importancia del etileno, tanto como producto químico orgánico de mayor volumen que como bloque estructural para la obtención de productos petroquímicos y plásticos, y el butadieno como agente para la producción de caucho sintético. La glucosa se transforma asimismo en productos químicos que actualmente carecen de importancia desde el punto de vista industrial, pero que pueden tenerla en condiciones económicas apropiadas. Uno de estos procedimientos es la producción de hidroximetilfurfurol y su ulterior conversión en ácido levulínico mediante la acción de ácidos minerales calientes sobre la glucosa (Harris, 1975). El empleo de la glucosa como substrato de fermentación general permitiría la producción, a partir de la madera, de un amplio surtido de antibióticos, productos químicos, vitaminas y enzimas (Seeley, 1976). Por ejemplo, el ácido láctico podría transformarse en ácido acrílico y en acrilatos. 44. Ciclo del Carbono en sistemas agroforestales 37 El carbono es un bioelemento que da estructura y soporte a los organismos vivos.. Biomoléculas como: proteínas, ácidos nucleicos, carbohidratos, lípidos, enzimas , hormonas y aminoácidos , tan esenciales para la vida ,contienen carbono. Se encuentra como dióxido de carbono (CO2) ó gas carbónico, en atmósfera, océanos y en los combustibles fósiles almacenados bajo la superficie de la Tierra. El movimiento global del carbono entre el ambiente abiótico y los organismos se denomina ciclo del carbono. El ciclo básico comienza cuando las plantas, a través de la fotosíntesis, hacen uso del dióxido de carbono (CO2) presente en la atmósfera o disuelto en el agua. El carbono (del CO2) pasa a formar parte de los tejidos vegetales en forma de carbohidratos , grasas y proteínas, y el oxígeno es devuelto a la atmósfera o al agua mediante la respiración. Así, el carbono pasa a los herbívoros que consumen las plantas y de ese modo utilizan, reorganizan y degradan los compuestos carbonados mediante las RBE de la respiración celular. Estas reacciones metabólicas producen CO2, H2O , ATP ; también se libera gas carbónico como producto secundario del metabolismo, pero parte se almacena en los tejidos animales y pasa a los carnívoros, que se alimentan de los herbívoros. En última instancia, todos los compuestos del carbono se degradan por descomposición, y el carbono que es liberado en forma de CO 2 a la atmósfera, es utilizado de nuevo por las plantas en su proceso fotosintético,la figura 2, integra la secuencia de las etapas del ciclo. Figura 2.Secuencia de reacciones en el ciclo del carbono (Tomado de: http://www.windows2universe.org/earth/Water/co2_cycle.html&lang=sp ) 38 En resumen, las reacciones más importantes que ocurren en el ciclo del carbono son las siguientes: 1. Fotosíntesis : El dióxido de carbono de la atmósfera es fijado por las plantas en el ciclo de Calvin y convertido en glucosa (azúcar aldohexosa , C6) según la ecuación química: Radiación UV 6CO2 + 6H2O + 686kcal E=h C6H12O6 + 6O2 2. Respiración celular :Los animales consumen los vegetales y a través de sus procesos digestivos , absortivos y metabólicos , oxidan carbohidratos como la glucosa a través de las RBE dadas en : glucólisis ,Ciclo de Krebs y Fosforilación Oxidativa ,hasta producir gas carbónico , agua y energía , según la ecuación química : C6H12O6 + 6O2 RBE Mitocondria 6CO2 + 6H2O + 38ATP 3. Descomposición : Bacterias y hongos descomponen plantas muertas y la materia animal, devolviendo carbono al medio ambiente. 4. Fosilización: En algunos casos el carbono presente en las moléculas biológicas no regresa inmediatamente al ambiente abiótico, por ejemplo el carbono presente en la madera de los árboles ó el que formó parte de los depósitos de hulla a partir de restos de árboles antiguos que quedaron sepultados en condiciones anaerobias antes de descomponerse. Hulla, petróleo y gas natural son llamados combustibles fósiles porque se formaron a partir de restos de organismos antiguos y contienen grandes cantidades de compuestos carbonados como resultado de la fotosíntesis ocurrida hace millones de años. 39 45. Captura de Carbono en sistemas Agroforestales CUANTIFICACIÓN DEL ALMACENAMIENTO DE CARBONO EN SISTEMAS SILVOPASTORILES EN ZONAS PRODUCTORAS DE LECHE EN CUNDINAMARCA Ángel Alberto Terreros Riveros3, Sandra Castiblanco Guzmán4, Jenny Bustos García5, Jairo 6 7 8 Enrique Granados Moreno , Leonor Barreto de Escovar , Luz Mery Bernal . RESUMEN Las prácticas productivas ganaderas deben encaminarse en acciones conservacionistas, mitigando el impacto que ejercen sobre los ecosistemas y el calentamiento global. Se cuantifico el almacenamiento de carbono en ganadería tradicional y sistemas silvopastoriles conformados por Alnus acuminata, Sambucus nigra, Acacia decurrens, Acacia melanoxylon, en asocio con pasturas compuestas por Pennisetum clandestinum, Holcus lanatus, Lolium spp, Trifolium pratense, Trifolium repens, en tres fincas ganaderas, determinando la línea base para la cuantificación de carbono. Se utilizo método alométrico para determinar la biomasa arriba del suelo del componente forestal, el valor para la relación de raíz es de 0,27 y el factor de proporción de carbono equivalente a 0,5 por unidad de materia seca para el material vegetal, según IPCC 2002. El porcentaje de carbono orgánico en suelos se determino por el método Walkley Black, y la lectura se realizo por espectrofotometría; El carbono contenido en el suelo (t C/ha) se calculo a partir de los valores de porcentaje de carbono y densidad aparente. En la evaluación se propuso un diseño en bloques al azar con arreglo factorial 3 x 4, donde el factor A = Finca, el factor B = Sistema (Un sistema de ganadería tradicional y tres tipos de arreglos de sistemas silvopastoriles). El análisis de varianza demostró diferencias altamente significativas (p < 0,01) en el total de carbono almacenado entre fincas, entre sistemas y en la interacción de finca*sistema, demostrando la gran capacidad de almacenamiento de carbono de los sistemas silvopastoriles frente a los sistemas de ganadería tradicional. Palabras Claves: Captura de Carbono, Componente Forestal, Biomasa Arriba del Suelo, Modelos Alométricos, Carbono Orgánico en Suelo. INTRODUCCIÓN El impacto de las actividades agropecuarias en los diferentes ecosistemas es considerado de suma importancia por las agencias de desarrollo e institutos de investigación; cuyos efectos en definitiva se traducen en baja fertilidad, baja capacidad de retención de humedad, alta susceptibilidad a la erosión, perdida de la cobertura vegetal, problemas de compactación, deforestación, sistemas de manejo de praderas extractivos que 3 Zootecnista. [email protected]. UNAD. Bogotá. Estudiante Tesista de Zootecnia. [email protected]. UNAD. Bogotá. 5 Estudiante Tesista de Zootecnia. [email protected]. UNAD. Bogotá. 6 Químico. Ph.D. (c). M.Sc. Investigador. [email protected]. UNAD. Bogotá. 7 Zootecnista. M.Sc. (c). Espec. Nutrición Animal Sostenible. Investigador. [email protected]. UNAD. Bogotá. 8 Bióloga. Ph.D. Investigadora. [email protected]. UNAD. Bogotá. 4 40 acrecientan la perdida de la capacidad productiva de estos y en fuentes de emisión de Gases de Efecto Invernadero (GEI). Así mismo, es común en los últimos tiempos hablar de cambio climático y de sus efectos negativos en el medio, tanto que las investigaciones científicas sobre las emisiones de gases de efecto invernadero durante los últimos 10 años predicen que el cambio climático tendrá impactos negativos ambientales, sociales y económicos a nivel global. Los impactos pueden incluir aumento del nivel de los mares, erosión costera, cambios dramáticos en patrones climáticos, aumento de enfermedades tropicales, la pérdida acelerada de biodiversidad, y la desertificación (Stuart y Moura Costa, 1998). Numerosas modelaciones del cambio climático global sugieren que se puede contribuir sustancialmente al control de los niveles de CO 2 en la atmósfera mediante la calidad del manejo forestal, la conservación del bosque en peligro de deforestación, la rehabilitación de bosques, forestación, reforestación9 o promoción de la agroforestería. Debido a que el suelo almacena cantidades considerables de carbono, las prácticas que promueven un aumento en el carbono orgánico del suelo también pueden tener un efecto positivo en la fijación (Stuart, M.D. y Moura Costa, P. 1998). De hecho la Decisión 19/COP-9 10 ha definido como “reservorios de carbono” a la biomasa superficial, la biomasa subterránea, los detritos, la madera muerta y el carbono orgánico del suelo. Los Sistemas Agroforestales y entre ellos los Silvopastoriles 11 pueden mantener y hasta aumentar las reservas de carbono en la vegetación y los suelos; fomentando a su vez prácticas sostenibles con bajos insumos, debido al ciclaje de nutrientes y la estratificación de la vegetación perenne con lo que se disminuye la presión sobre los recursos naturales.Para efectos de homogenizar los conceptos y de acuerdo a la Decisión 17/COP–7 12 de la Convención Marco Sobre el Cambio Climático (CMCC) del Panel Intergubernamental Sobre Cambio Climático (IPCC); se acordó que el almacenamiento o stock de carbono hace referencia a la capacidad de un ecosistema de mantener una determinada cantidad promedio de carbono por ha, expresándose en toneladas de carbono por ha (t C ha-1). El carbono fijado se refiere a la capacidad de una unidad de área cubierta por vegetación para fijar carbono en un período determinado, adicional a la cantidad almacenada en el momento actual (Segura, 1997), expresándose en t C ha-1 año-1. El carácter de adicional es considerado, según la Decisión 19/COP–9 de la CMCC, si la absorción neta efectiva de GEI por los sumideros supera la suma de las variaciones del carbono almacenado que se habrían producido de no realizarse la actividad del proyecto de forestación o 9 Forestación: Conversión, por actividad humana directa de tierras que carecieron de bosque durante un periodo mínimo de 50 años, en tierras forestales mediante plantación, siembra o fomento antropógeno de semilleros naturales. Reforestación: Conversión por actividad humana directa de tierras no boscosas en tierras forestales mediante plantación, siembra o fomento antropógeno de semilleros naturales en terrenos donde antiguamente hubo bosques, pero que actualmente están deforestados. 10 Novena Reunión de la Conferencia de las partes en el Convenio sobre la Diversidad Biológica. 11 Sistema de producción del suelo que implica el asocio espacial y simultáneo de praderas (que sustentan la alimentación animal) y leñosas perennes multipropósito. 12 Séptima Reunión de la Conferencia de las partes en el Convenio sobre la Diversidad Biológica. 41 reforestación del Mecanismo de Desarrollo Limpio registrado, en el marco del Protocolo de Kyoto. Se está reconociendo el papel de las pasturas en el ciclo del carbono, por cuanto las gramíneas seleccionadas con altos rendimientos de biomasa y bien adaptadas cumplen un papel importante en la reducción de la emisión de carbono a la atmósfera debido a la producción de biomasa aérea y raíces y deposición de materia orgánica en el suelo. Se han reportado tasas de fijación de carbono del orden de 0,1 a 4,3 t C ha-1 año-1 para Sistemas Agroforestales y Silvopastoriles en Centroamérica (Kursten. 1993, Pomareda. 1999). Un estudio realizado en la Zona Norte de Costa Rica reporto niveles de almacenamiento de hasta 233 t C ha-1 mientras que el suelo de un sistema silvopastoril con regeneración natural de laurel (Cordia alliodora) almacenó de 180 – 200 t C ha-1. En Guápiles, Costa Rica, Andrade (1999) estimó el almacenamiento de carbono sobre el suelo en sistemas silvopastoriles con Acacia mangium y Eucalyptus deglupta en combinación con pasturas B. brizantha, B. decumbens, y P. maximum obteniéndose valores que oscilan de 3,7 t C ha-1 a 4,7 t C ha-1 donde el componente arbóreo aporta un promedio de 76 a 94% del carbono total. Arias (2001) reporta para un sistema silvopastoril en Venezuela con la especie Gliricidia sepium, un almacenamiento de 309 Kg. C ha -1 y una tasa de fijación de 124 Kg C ha -1 año-1; mientras que en el sistema de cultivos en callejones el almacenamiento fue de 654 Kg. C ha-1 y la tasa de fijación de 327 Kg C ha-1 año-1. MATERIALES Y MÉTODOS Localización El presente estudio se realizo en la finca San Pedro, localizada en el municipio de Mosquera, Cundinamarca, Colombia. El municipio presenta una temperatura promedio de 12,8 °C, una humedad del 82% y una precipitación anual de 600 – 700 mm.; En la finca Villa Nataly y El Ciprés, localizadas en Usme (Localidad Quinta de Bogotá), Colombia. La región presenta una temperatura promedio de 8 °C, y una precipitación anual de 1000 a 1500 mm. En el trabajo de investigación se analizaron los siguientes sistemas y arreglos silvopastoriles dentro de las fincas seleccionadas de la siguiente forma: 1) Finca San Pedro: Sistema de ganadería tradicional (SGT) conformado por Kikuyo (P. clandestinum) 90%, Trébol Rojo (T. pratense) 5% y Raigrás (Lolium spp) 5%; Cerca Viva compuesta por Aliso (A. acuminata), Acacia negra (A. decurrens) y Acacia japónica (A. melanoxylon); Banco de Proteína compuesto por Sauco (S. nigra), y como componente forrajero Kikuyo (P. clandestinum) en un 90%. Estos arreglos silvopastoriles llevan establecidos hace 4 años aproximadamente. 2) Finca Villa Nataly: SGT conformado por Kikuyo (P. clandestinum) 40,5%, Raigrás (Lolium spp) 10,8%, Falsa poa (H. lanatus) 21%, Trébol rojo (T. pratense) 7,8% y Trébol blanco (T. repens) 1,1% y el 18% restante está representado por arvenses y malezas; Cerca Viva compuestos por Aliso (A. acuminata); Arreglo de Sombra y Ramoneo, compuesto por Aliso (A. acuminata); Banco de Proteína compuesto principalmente por Acacia negra (A. decurrens), y otras especies presentes en 42 más baja densidad como Acacia japónica (A. melanoxylon) y Aliso (A. acuminata). Estos arreglos silvopastoriles tiene 5 años de establecidos aproximadamente. 3) Finca El Ciprés: SGT conformado por Kikuyo (P. clandestinum) 80%, Falsa poa (H. lanatus) 10%, y Trébol rojo (T. pratense) 10%; Viva compuesto por Aliso (A. acuminata). Este arreglo está establecido aproximadamente hace 2 años. Para el análisis de los datos se empleo un diseño experimental en bloques al azar, con tres repeticiones y arreglo factorial 3 x 4, donde el factor A = Finca (Tres fincas evaluadas) y el factor B = Cuatro sistemas (Un sistema de ganadería tradicional y tres tipos de arreglos de sistemas silvopastoriles), evaluando el efecto de la finca, el efecto del sistema y la interacción de finca*sistema. Se utilizo también la prueba de Duncan para la determinación de diferencias estadísticas de las medias entre las tres fincas y entre los cuatro sistemas evaluados. Unidades de Muestreo En cada sistema se estableció una unidad de muestreo denominada parcela permanente de muestreo (PPM), donde la forma y el tamaño fueron determinados por el tipo de arreglo teniendo en cuenta la densidad de siembra y el tiempo de establecimiento (Tabla 1). Tabla 1. Parcelas permanentes de Muestreo (PPM) Establecidas Finca San Pedro Arreglo Silvopastoril Cerca Viva de Aliso Acacias Banco de Proteína de Sauco Cerca Viva de Aliso Villa Nataly El Ciprés Banco de Proteína de Aliso y Acacias Siglas Forma PPM Tamaño PPM 2 CV-AA Lineal 40 m BP-S Circular 1.257 m2 Lineal 46 m BP-AA Circular 314 m2 Sombra y Ramoneo de Aliso SR-A Circular 1.257 m Cerca Viva de Aliso CV-A Rectangular 293 m2 Latitud: 4°41'17.14" N Longitud: 74°13'12.41" O Latitud: 4°41'38.17" N Longitud: 74°12'55.75" O Latitud: 4°25'47.25" N Longitud: 74° 7'58.66" O 2 CV-A Coordenadas* Latitud: 4°25'47.97" N Longitud: 74° 7'59.33" O 2 Latitud: 4°25'49.78" N Longitud: 74° 8'1.79" O Latitud: 4°23'37.27" N Longitud: 74°10'20.15" O * Datos obtenidos con GPS Garmin Colorado 300 (Autores., 2009). 43 Se realizó un mapeo de los sistemas evaluados y de las PPM, con Geoposicionador (Garmin Colorado 300), la medición del Diámetro a la Altura del Pecho (DAP) a 1,30 m de la base del árbol, la determinación de la altura comercial de cada árbol por medio de clinómetro marca Suunto, el muestreo de forraje y hojarasca, y el muestreo de suelo a 30 cm de profundidad para hallar densidad aparente y porcentaje de carbono orgánico en el Laboratorio de Nutrición Animal de la Universidad Nacional Abierta y a Distancia (UNAD), sede José Celestino Mutis (Calle 14 Sur No. 14 – 23). Depósitos de Carbono en los Sistemas Evaluados Se definieron como depósitos de carbono para la cuantificación del almacenamiento de carbono los siguientes estratos: Biomasa Arriba del Suelo: Comprende árboles en pie de Aliso (A. acuminata), Sauco (S. nigra), Acacia Negra (A. decurrens), Acacia Japónica (A. melanoxylon); y pastos Kikuyo (P. clandestinum), Falsa Poa (H. lanatus), Raigrás (Lolium spp) Trébol Rojo (T. pratense) y Trébol Blanco (T. repens). Para la determinación de la biomasa arriba del suelo del componente forestal se utilizo algunos modelos y ecuaciones alométricas seleccionados teniendo en cuenta la zona climática (Tabla 2), basándose en las medidas tomadas en las PPM (DAP y altura total). Tabla 2. Ecuaciones Alométricas para Determinar la Biomasa de Arboles Zona Climática Rango DAP Ecuación R2 <900 mm 3-30 Y = Log^ {-0,535 + log10 (BA)} 0,94 <900 mm 3-30 Y = 0,299 * (DAP)2 0,94 <1500 mm 5-40 Y = exp {-1,996 + 2,32*ln(DAP)} 0,89 Autor Martínez - Yrizar et al. (1992). Martínez - Yrizar et al. (1992). Brown et al. (1989). N° (1) (2) (3) Fuente: Martínez – Yrizar et al. (1992). Brown et al. (1989). Fundación Solar. (2000). Donde: Y = Biomasa arriba del suelo en kilogramos; DAP = Diámetro a la altura del pecho en centímetros; BA 2 2 = Área basal en cm (BA = PI * r ); r = radio. El valor obtenido de biomasa expresada en kg, se dividió en 1000 para pasar a toneladas; este valor se multiplicó por 0.5 (Proporción de carbono por unidad de materia seca según IPCC 2002) lo que da toneladas de carbono almacenado, luego se dividió en el área de la PPM donde se obtuvo el dato de cada árbol expresado en m2, obteniendo toneladas de carbono por m2 (t C/m2); este valor se multiplico por 10000 para convertirlo a toneladas de carbono por hectárea (t C/ha). Por último se realizo la sumatoria de los valores obtenidos de cada árbol medido en la PPM y así se obtuvo el total de t C/ha de este componente del sistema evaluado. La biomasa de la hojarasca y los forrajes se incluyeron en los cálculos para biomasa arriba del suelo, ya que se muestrearon y pesaron conjuntamente en un marco de 50 x 50 44 cm (3 muestras por sistema silvopastoril y 5 muestras para sistema de ganadería tradicional). Para el cálculo de la biomasa en estas fuentes se obtuvo el valor del contenido de humedad, determinando la materia seca de sub-muestras recolectadas de 200 g extraída de la mezcla de las muestras pesadas, por medio del secado en mufla a 100 °C durante 4 horas. Este valor se cálculo de la siguiente manera (Fundación Solar., 2000): CH = (Phs - Pss) / Phs; Donde: CH = Contenido de humedad; Phs = Peso húmedo sub-muestra (g); y Pss = Peso seco sub-muestra (g). Con el valor de contenido de humedad se procedió a calcular la proporción del peso húmedo que corresponde a biomasa: Y = Pht - (Pht * CH); Donde: Y = Biomasa en gramos; Pht = Peso húmedo total de la muestra (g); y CH = Contenido de humedad. El valor obtenido se dividió en 1000000 para obtener toneladas, luego se multiplico por 0.5 (Proporción de carbono por unidad de materia seca según IPCC 2002) quedando toneladas de carbono almacenado, por último se dividió en el total de metros muestreados, dando como resultado t C/m2 y al multiplicarlo por 10000 m 2 se obtuvo t C/ha. Biomasa Abajo del Suelo: Hace referencia a las raíces de la vegetación del ecosistema estudiado. El cálculo de este componente se realizo con el valor para R (Relación de raíz) para el bosque seco tropical que es de 0.27 según el cuadro 3A.1.8. de IPCC (2002), de la biomasa calculada arriba del suelo, expresada en t MS/ha. Para hallar el contenido de carbono en este componente se multiplico por 0.5 (Proporción de carbono por unidad de materia seca según IPCC 2002) obteniendo el valor en t C/ha. Para las especies forrajeras y herbáceas, se descontó el 5% que representa la proporción de hojarasca del total de biomasa calculada arriba del suelo. Hojarasca: Comprende residuos orgánicos de hojas, ramas, frutos y semillas que se encuentran en la superficie del suelo. El cálculo de este componente fue incluido en los cálculos de biomasa y contenido de carbono arriba del suelo conjuntamente con los forrajes. Suelo: Se cuantifico el contenido de carbono en los primeros 30 cm de profundidad del suelo; utilizando una muestra obtenida a 20 cm de profundidad para determinar la densidad aparente por el método del “cilindro de volumen conocido” descrito en MacDicken (1997) y otra de aproximadamente 200 g obtenida a 30 cm de profundidad, para determinar el porcentaje de carbono orgánico en laboratorio por el método WalkleyBlack, en el cual el suelo se oxida con una solución de Dicromato de potasio estandarizada, utilizando el calor producido por la dilución de ácido sulfúrico concentrado, en la solución crómica. El porcentaje de carbono orgánico se determinó por colorimetría (espectrofotometría), cuantificando el color verde del ácido crómico reducido a una Absorbancia máxima (λmax) de 590 nanómetros (nm), la cual es proporcional a la materia orgánica que reacciona. Además, se realizó la respectiva curva de calibración con patrones de Sacarosa (Adaptado de García, G. Ballesteros, G). Para hallar el contenido de carbono en el suelo, se utilizo el siguiente cálculo: COS = CO * DA * P; Donde: COS = C almacenado (t C ha-1), P = Profundidad de muestreo (cm); DA = Densidad aparente del 45 suelo (g/cm-3), y CO = Contenido de carbono (%) determinado por método de WalkleyBlack. 46