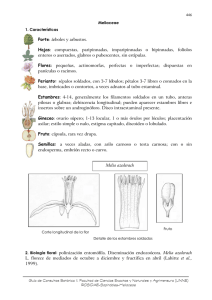
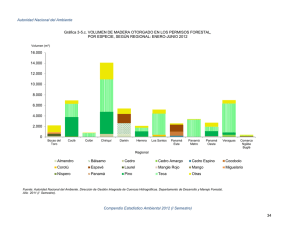
Descargar PDF - Fundación ProYungas
Anuncio

PROLOGO La actividad forestal, contrariamente a lo que se suele pregonar desde distintos ámbitos, se ha desarrollado en nuestro país y en Latinoamérica siguiendo una lógica que prioriza el beneficio de corto plazo sobre cualquier consideración ambiental o inclusive de sostenibilidad en la obtención de ingresos económicos. Los bosques nativos han sido sometidos a la práctica denominada curiosamente “floreo” que consistió en extraer los ejemplares presuntamente mejores, de fuste más recto y de mayor volumen, de las especies que presentaban mejores expectativas comerciales. Las extracciones se realizaron sin tener en cuenta condiciones de regeneración y de crecimiento de las especies en cuestión. De esta manera probablemente se perdió una porción importante de la variación genética de muchas de estas especies, a la vez que disminuyó el capital sujeto a aprovechamiento. Esta forma de explotación determinó posiblemente la imposibilidad de generar polos de desarrollo en torno a los bosques nativos, con la consecuencia señalada en este libro de que amplias poblaciones económicamente marginales se asientan en las cercanías de estas formaciones. La segunda consecuencia de esta forma de encarar la relación con los bosques nativos es que los imperativos económicos de corto plazo rápidamente pudieron demostrar la necesidad de reemplazar bosques “empobrecidos” o “degradados” por actividades de mayor rentabilidad como la agricultura o la ganadería. Al empobrecimiento no le sucedió pues el enriquecimiento reparador, sino la destrucción. Estos procesos han afectado de manera singular muchas especies nativas y los ecosistemas que las contienen, y entre ellas se destacan los cedros (Cedrela spp.) de nuestro continente. Como se menciona en la sección introductoria, los ecosistemas mas accesibles han sido sometidos a una extracción despiadada o directamente convertidos a sistemas agropecuarios. A su vez la escasa o nula generación de conocimientos acerca de aspectos fisiológicos, genéticos y ecológicos ha conspirado contra el desarrollo de tecnologías que pudieran salir al cruce de las mencionadas prácticas, ofreciendo alternativas de manejo sustentable que pudieran reemplazar la lógica prevaleciente en el tratamiento de las especies de Cedrela spp. Contrasta con esta realidad el potencial de desarrollo que presentan las especies de cedro, debido al valor de su madera y a sus tasas de crecimiento, que permitirían realizar forestaciones que no requieren de enormes superficies para garantizar ingresos económicos más que aceptables. Por ello mismo, serían aptas para pequeños y medianos productores que habitan en la cercanía de los ecosistemas de bosque nativo de las Yungas y de la Selva Misionera. Es por ello que muchos celebramos la aparición de este volumen dedicado a uno de los géneros productores de madera de mejor calidad del país. En el mismo se describen los últimos avances en la generación de conocimientos básicos acerca de la distribución, fenología, relaciones con disturbios de diferente tipo, producción de madera de diferentes características, productividad y existencias así como ejemplos aplicados de enriquecimiento y esfuerzos por generar cultivos basados en especies de Cedrela spp. Seguramente es mucho lo que queda por hacer, pero ningún partido se gana sin jugarlo y todos requieren del puntapié inicial que quizás sea uno de los destinos de este libro. En mi opinión la investigación forestal y ecológica debe ciertamente cambiar en nuestro país orientándose, tal como se muestra en los capítulos que siguen, a la generación de conocimientos y tecnologías que permitan un manejo sustentable de los recursos con más potencial para generar desarrollo económico, tales como las especies de cedros. El presente volumen representa en este sentido un ejemplo a seguir. Es de esperar que a los investigadores que presentan trabajos en este libro se les sumen otros, como parece estar sucediendo en otras regiones de la Argentina, para profundizar sobre los conocimientos aquí presentados, así como para generar los mismos o similares para otras especies de similar valor de las diversas regiones forestales de nuestro país. Tomás Schlichter Coordinador Programa Nacional Forestal, INTA [email protected] Ecología y producción de cedro (género Cedrela) en las Yungas australes Ediciones del Subtrópico Tucumán - Argentina Diciembre de 2006 Institución patrocinante: Gasoducto Nor Andino Argentino SA Agradecimientos: Marcelo Arturi Natacha Chacoff Ricardo Grau Juán Manuel Morales Darién Prado Pablo Yapura Colaboración en edición: Karina Buzza Fotografía de portada, diagramación y diseño editorial: Ignacio Paz Posse (www.cerrosyvalles.com) Esta publicación ha sido realizada en conjunto por el Laboratorio de Investigaciones Ecológicas de las Yungas (LIEY-UNT) y la Fundación ProYungas © 2006 Fundación ProYungas, C. C. 34 (4107) Yerba Buena, Tucumán, Argentina Prohibida la reproducción total o parcial por medios electrónicos o mecánicos, incluyendo fotocopia, grabación magnetófonica y cualquier sistema de almacenamiento de información, sin consentimiento expreso de los editores. ISBN: ISBN 978-987-23533-0-8 Impreso en Argentina - Artes gráficas Crivelli PROLOGO Tomás Schlichter introduccion 9- 18 Capítulo 1 Importancia del género Cedrela en la conservación y desarrollo sustentable de las Yungas australes. Alejandro Brown y Silvia Pacheco Seccion 1 Ecología, distribución geográfica y características de la madera 19- 30 Capítulo 2 Botánica y distribución del género Cedrela en el noroeste de Argentina. Alfredo Grau, M. Alicia Zapater y Roberto A. Neumann 31-40 Capítulo 3 Fenología de dos especies de cedro en un gradiente altitudinal del Parque Nacional Calilegua, Jujuy. Silvia Pacheco y Alicia Yapur 41-50 Capítulo 4 Biología reproductiva e importancia de la polinización en Cedrela lilloi. Valeria Aschero 51-58 Capítulo 5 Características y usos de las maderas de Cedrela de las Yungas de Argentina. Stella Maris Rivera 59-82 Capítulo 6 Variabilidad interanual de los caracteres anatómicos en el leño de Cedrela lilloi en el noroeste de Argentina. Ricardo Villalba, Silvia Delgado, Mónica De Membiela y Daniela Mendoza Seccion 2 Estructura, crecimiento y dinámica poblacional de bosques nativos 83-104 Capítulo 7 Diversidad, estructura y distribución de bosques con cedro en el noroeste de Argentina y sur de Bolivia. Lucio Malizia, Cecilia Blundo y Silvia Pacheco 105-116 Capítulo 8 Modelos de crecimiento de Cedrela lilloi en el sector norte de las Yungas argentinas. N. Ignacio Gasparri y Juan F. Goya 117-130 Capítulo 9 Incidencia de la herbívora en Cedrela balansae. Romina P. Scandalo, Gustavo A. Zuleta , M. Andrea Gainza y A. Silvia Haedo 131-142 Capítulo 10 Influencia de la ganadería trashumante y la apertura de claros en la supervivencia y el crecimiento de Cedrela lilloi en Tariquía, Bolivia. Marcela Zamora Petri Seccion 3 Producción y comercialización 143-154 Capítulo 11 Aprovechamiento forestal de cedro en las Yungas de Argentina. José M. Minetti 155-160 Capítulo 12 Producción de plantines de cedro en vivero. David Monteverde 161-170 Capítulo 13 Experiencias preliminares de propagación de Cedrela in vitro. Roxana J. Enrico y M. Eugenia Enrico 171-178 Capítulo 14 Crecimiento de Cedrela balansae en la plantación experimental de Valle Morado. Mariana Horlent y David Monteverde 179-192 Capítulo 15 Resultados comparativos de plantaciones experimentales de Cedrela balansae en INTA - Yuto, Jujuy. Elvio del Castillo, M. Alicia Zapater y M. Norma Gil 193-198 Capítulo 16 Enriquecimiento con cedro rosado y cedro misionero en Tucumán, Argentina. Jorge González Morenghi 199-208 Capítulo 17 Mercados para la madera de cedro. Ariel Zorrilla Seccion 4 209-224 Mapas y fotografías Importancia del género Cedrela en la conservación desarrollo sustentable Ecología y producciónyde cedro (género Cedrela)deenlas lasYungas Yungas australes australes. Pacheco, S. y A. Brown (Eds.). LIEY-ProYungas. Argentina. pp. 9-18. 2006. Importancia del género Cedrela en la conservación y desarrollo sustentable de las Yungas australes Brown A.1,2 y S. Pacheco1,2 1 Fundación ProYungas, CC 34 (4107), Yerba Buena, Tucumán, Argentina. 2 Laboratorio de Investigaciones Ecológicas de las Yungas (LIEY, UNT). [email protected] CONTEXTO ECOLOGICO Y SOCIOECONOMICO DE LAS YUNGAS AUSTRALES Las Yungas representan el ecosistema de selvas húmedas subtropicales de mayor extensión geográfica en la Argentina (casi 5.5 millones de hectáreas de bosques). Conjuntamente con la Selva Paranaense representan menos del 3% del territorio nacional, pero albergan más del 50 % de la biodiversidad del país. Esta importante biodiversidad debiera traducirse en una variedad de opciones para el desarrollo regional basadas en el uso sustentable de sus recursos naturales. Sin embargo, la realidad es diferente y estos sistemas se enfrentan a importantes procesos de degradación y transformación. De la Selva Paranaense en la Provincia de Misiones (hábitat de Cedrela fissilis), ha sido transformado casi el 60 % de su superficie y este remanente es casi todo lo que queda de una ecoregión que ocupaba importantes áreas de Brasil, Paraguay y Argentina (Placci y Di Bitetti 2006). Adi- cionalmente, estos bosques se encuentran intervenidos en porcentajes elevados quedando pocos espacios sin explotar fuera de las áreas protegidas e incluso dentro de las mismas. EL 89 % de la Selva Paranaense remanente se encuentra en niveles medianos a elevados de degradación y fragmentación y persisten menos de 40.000 ha de bosques prístinos (MacDonagh y Rivero 2006). Las Yungas australes son la expresión más meridional de un sistema que acompaña a los Andes desde Venezuela hasta el noroeste de Argentina. En nuestro país ocupa más de 5 millones de hectáreas, de las cuales un millón ya se encuentra transformado en tierras agrícolas (Brown et al. 2002). Este proceso avanza a una tasa de aproximadamente 10.000 ha anuales en las áreas pedemontanas (Gasparri y Grau 2006) poniendo en riesgo una parte importante de la superficie remanente de Selva Pedemontana (el piso inferior de las Yungas) por degradación y transformación (Brown y Malizia 2004). Brown A. y S. Pacheco En el noroeste de Argentina, las Yungas australes se extienden desde la frontera con Bolivia (22° S) hasta la Provincia de Catamarca (29° S) por más de 700 km de longitud Norte - Sur. Altitudinalmente, las Yungas ocupan desde el pedemonte (400 m s.n.m.) hasta los 2.500 - 3.000 m s.n.m. en las áreas cumbrales de la Sierras Subandinas y Precordillera oriental (Brown et al. 2001). En este importante gradiente altitudinal las características estructurales del bosque, las especies dominantes y la importancia forestal, varían marcadamente (Tabla 1). Los dos pisos altitudinales de mayor importancia forestal se caracterizan por la abundancia de distintas especies de cedro. En la Selva Pedemontana (SPM) es común el cedro orán (Cedrela balansae) y otras 9 especies de interés forestal y en el Bosque Montano (BM) se encuentran los rodales dominados por el cedro coya Tabla 1. Características estructurales de las áreas boscosas del Alto Bermejo (promedio y error estándar; entre paréntesis número de parcelas de 1 ha) (Malizia 2004). Altitud Area m basal s.n.m. (m2/ha) Densidad (ind./ha) Riqueza (especies/ha) Especies dominantes (en orden decreciente de abundancia) 600 25,7 ± 1,3 473,5 ± 27,5 39,0 ± 2,9 Anadenanthera colubrina, Phyllostilon rhamnoides, Trichilia hieronymi, Parapiptadenia excelsa, Calycophylum multiflorum Selva montana (8) 1.100 27,5 ± 2,1 458,3 ± 30,0 39,6 ± 2,4 Miconia molybdea, Croton densiflorus, Siphoneugenia occidentalis, Blepharocalix salicifolius, Inga marginata Ecotono selva-bosque (7) 1.600 33,1 ± 1,9 573,7 ± 34,3 28,2 ± 2,4 Allophylus edulis, Podocarpus parlatorei, Blepharocalix salicifolius, Parapiptadenia excelsa, Myrcianthes pseudomato Bosque montano (7) 2.100 36,0 ± 1,0 508,3 ± 70,5 14,2 ± 2,0 Podocarpus parlatorei, Prunus tucumanensis, Viburnum seemeni, Ilex argentina, Crinodendron tucumanum Selva pedemontana (8) 10 Importancia del género Cedrela en la conservación y desarrollo sustentable de las Yungas australes (Cedrela lilloi), además de nogal criollo (Juglans australis) y pino del cerro (Podocarpus parlatorei). En el espacio intermedio ocupado por la Selva Montana (SM) son escasas las especies de interés forestal. Los dos primeros pisos son utilizados en general por empresas de distinta magnitud económica. La Selva Pedemontana ha sido la más presionada por su cercanía a ciudades y rutas y ostenta los mayores niveles de degradación. Es explotada por pequeñas empresas de capitales limitados y en circuitos económicos que generalmente bordean la ilegalidad. El Bosque Montano, que requiere mayores inversiones en infraestructura y logística, es aprovechado por empresas de importantes capitales que concentran su actividad en la explotación del cedro, cuyo capital maderero se acumuló por espacio de varios siglos. Es decir la SPM sustenta una economía pauperizada de varias décadas de explotación, aunque en cierta medida con opciones de sustentabilidad, debido a la diversidad de especies maderables y las altas tasas de crecimiento. El BM, en cambio, es generador de riquezas, pero a costa del capital acumulado en centurias y que no presenta opciones claras de sustentabilidad. Esto se debe a la distribución agregada de las edades de los rodales y las dificultades de las poblaciones de C. lilloi de recuperarse de la explotación forestal, por el bajo reclutamiento en condiciones de bosque con cobertura cerrada. Si a estas características propias de las condiciones climáticas y de la fisiología de las dos especies de cedros, le sumamos su rol ecosistémico, es posible establecer diferencias importantes en cuanto a la funcionalidad que estos bosques debieran cumplir en la sustentabilidad del ecosistema Yungas al largo plazo y en la economía y sustentabilidad social de las poblaciones en la región. La Selva Pedemontana debiera considerarse como el “sistema forestal bajo manejo” por excelencia, por su importante diversidad forestal, sus altas tasas relativas de crecimiento, la cercanía a las rutas y centros poblados, su mayor estacionalidad climática que permite el acceso a las áreas boscosas durante prácticamente 10 meses al año, y por presentar menores costos de caminería. El Bosque Montano debiera considerarse como “bosque protector” dado el importante impacto ambiental que genera su explotación motivado por las abruptas pendientes, por lo costoso del desarrollo y mantenimiento de infraestructura, los pocos meses que se puede ingresar al bosque (menos de 6 meses al año), y por la ubicación topográfica de estos bosques en las cabeceras de cuenca y en el nivel de máxima frecuencia e intensidad de neblinas (“bosques nublados”). La Alta Cuenca del Río Bermejo (ACRB) en las Provincias de Salta y Jujuy, es el área de Yungas australes con mayor actividad forestal del noroeste de Argentina y el bloque forestal continuo de mayor superficie (más de 1,5 millones de ha). Apelando a la nomenclatura expuesta en el párrafo anterior, todo el sector de BM correspondiente a la Reserva de la Biósfera de las Yungas (RBYungas), debiera ser considerado como “bosque protector” sujeto a explotaciones forestales selectivas, con 11 Brown A. y S. Pacheco métodos de bajo impacto y restringidas en cuanto a su localización geográfica. En el pedemonte de esta área se desarrollan cerca de 100.000 ha de caña de azúcar bajo riego (Ingenios Ledesma y S. Martín del Tabacal) y una importante superficie de cultivos de hortalizas, maíz, cítricos y bananales que dependen del agua generada en las Yungas (Fig. 1). Por otra parte, el área de SPM ubicada mayoritariamente en el Departamento San Martín de la Provincia de Salta, debiera ser considerada como “sistema forestal bajo manejo” Esta área está bordeada a lo largo de su límite este por la ruta 34, asiento de una importante población de más de 150.000 personas que viven mayormente en condiciones económicas de marginalidad y que se reparten en seis municipios: Aguaray (8.000 hab.), Embarcación (18.000 hab.), General Mosconi (13.000 hab.), General Ballivián, Profesor Salvador Mazza (16.000 hab.) y Tartagal (56.000 hab.). En el Departamento San Martín en el 2001 había 10.854 hogares con Necesidades Básicas Insatisfechas (NBI), lo que significa el 35,3 % del total departamental. El 70,3 % de estos vivían bajo la Línea de Pobreza (LP) y el 42,5 % de estas personas estaban ubicadas bajo la Línea de Indigencia (LI). IMPORTANCIA ECOLOGICA DE LOS CEDROS EN LA REGION Desde los comienzos de la exploración y colonización de América Latina, los cedros han estado entre las especies forestales más importantes de la región noroeste de Argentina. Su madera blanda y fácil de 12 trabajar, pero a la vez resistente para su densidad, sumada a su perfume agradable y tolerancia al ataque de insectos una vez cortada, han sido los atributos que le han ganado un lugar entre las mejores maderas del mundo (Grau et al. este volumen). En el noroeste de Argentina habitan tres especies del género Cedrela (C. lilloi, C. balansae y C. saltensis), la primera del BM, la segunda típica de la SPM y la tercera habitante de las áreas altitudinalmente intermedias de Selva Montana (SM) (Grau et al. este volumen). En estos dos ambientes las condiciones climáticas son claramente contrastantes. La SPM se caracteriza por mayor temperatura media anual, de verano y de invierno y mayor precipitación media anual. En cambio en el BM, los valores de temperaturas anuales y estacionales son más bajos al igual que la precipitación media anual, pero son más comunes las neblinas. En la zona de ecotono los valores de temperatura y precipitación son intermedios entre los otros dos ambientes (Pacheco y Yapur este volumen). Las especies de Cedrela de las Yungas tienen un comportamiento fenológico similar a la comunidad vegetal del piso altitudinal al que pertenecen. C. balansae de la Selva Pedemontana, es una especie caducifolia con pérdida del follaje maduro y senescente durante tres meses en la estación seca del invierno y C. lilloi en el Bosque Montano es una especie semicaducifolia con pérdida del follaje maduro y senescente durante casi dos meses en la estación del invierno. Si bien ambas especies florecen abundantemente todos los años, C. lilloi presenta ciclos Importancia del género Cedrela en la conservación y desarrollo sustentable de las Yungas australes Figura 1. Pisos altitudinales de Yungas y áreas transformadas en la Alta Cuenca del Río Bermejo. 13 Brown A. y S. Pacheco muy largos de ausencia de producción de frutos, a diferencia de C. balansae que fructifica abundantemente todos los años (Pacheco y Yapur este volumen). Esto tiene implicancias sobre las opciones de regeneración vinculado con la generación de disturbios producto de la actividad forestal. Adicionalmente, se muestra que al menos C. lilloi es dependiente de una buena polinización para alcanzar niveles de producción de frutos abundantes (la fructificación aumenta cinco veces bajo polinización cruzada). En tal sentido, prácticas de manejo silvícola que aseguren la conservación de polinizadores efectivos de esta especie, son un requisito para asegurar la permanencia de las poblaciones de C. lilloi a largo plazo (Aschero este volumen). Las condiciones ambientales en que crecen las especies de cedro y posiblemente también particularidades genéticamente establecidas, les confieren características maderables diferenciables y por ende valores de mercado diferente. En C. lilloi por ejemplo, es común observar en el cepillado la aparición de pelusa, lo que podría atribuirse a la “madera de reacción” y que le ha valido el nombre vulgar de “cedro peludo”. Con respecto a C. balansae, si bien no presenta este defecto, su madera es considerada de calidad ligeramente inferior a C. lilloi y a C. fissilis (cedro misionero). Esta última es la madera considerada de referencia en cuanto a calidad para el género en nuestro país (Rivera este volumen). Las características anatómicas de la madera generan adicionalmente oportunidades 14 para estudios más complejos, como el desarrollo de reconstrucciones dendroclimáticas en áreas donde los bosques podrían ser la única fuente de información paleoclimática con amplia extensión temporal y resolución anual. Los factores ambientales actúan como fuerzas modeladoras del número y tamaño de los elementos constitutivos del xilema y en consecuencia, existe un gran potencial para derivar información climática a partir de otros caracteres anatómicos de la madera más allá del ancho de los anillos. La cronología final estudiada representa las variaciones en el crecimiento radial de los árboles muestreados durante los últimos 200 años donde el aumento en el tamaño del vaso medio es muy pronunciado durante los estados juveniles del árbol, disminuye en intensidad en los próximos 100 años y es muy reducido o nulo durante los últimos 50 años. Esto no está directamente relacionado con una disminución en crecimiento con la edad del árbol, sino que obedece a un principio geométrico de distribuir una cantidad dada de madera en círculos concéntricos cada vez de mayor diámetro (Villalba et al. este volumen). Potencial y limitaciones de la actividad forestal La información disponible sobre diversidad de árboles y estructura de las Yungas australes (o Subtropicales) es relativamente escasa. En ese sentido, el trabajo de Malizia et al. (este volumen) representa un aporte significativo a partir de la evaluación de 30 ha ubicadas en distintos pisos altitudinales en la ACRB. El estudio con- Importancia del género Cedrela en la conservación y desarrollo sustentable de las Yungas australes firma la presunción que los dos pisos de mayor concentración de especies de interés forestal son la SPM (30,4 % de los árboles son de interés forestal) y el BM (32 %), en tanto que la SM no registró árboles dominantes de interés forestal. Como reflejo de la disminución de la presión forestal con la altitud. se observó un incremento del área basal con la altitud y se registró una disminución en el número de tocones por ha. Adicionalmente, se generó un mapa de distribución de cedros a nivel regional que muestra que las dos especies de cedro estudiadas, se segregan espacialmente a lo largo del gradiente altitudinal (Malizia et al. este volumen). La degradación de los bosques tropicales y subtropicales es un proceso de pérdida gradual de cobertura que muchas veces es acompañado por una pérdida del valor comercial. En tal sentido, una de las premisas de la ordenación forestal es lograr la producción sostenida a perpetuidad, partiendo de la base que la extracción no supere el crecimiento del bosque. Por lo tanto, para determinar la capacidad de producción de un bosque y realizar la planificación de la explotación, son indispensables los inventarios forestales y el desarrollo de los modelos de crecimiento (Gasparri y Goya este volumen). En los bosques nativos del noroeste de Argentina, se realizan cortas selectivas respetando un diámetro mínimo de corta, legislado por cada provincia y para cada especie. Las intervenciones no son acompañadas por tratamientos silvícolas que garanticen la recuperación del bosque. En general, el aprovechamiento tradicional representa una reducción drástica de las existencias comerciales y si bien los bosques recuperan en parte la cobertura arbórea, la capacidad productiva queda comprometida. La distribución del crecimiento con la edad no es uniforme y los individuos al alcanzar los 30 a 40 cm de DAP presentan los mayores incrementos en diámetro con valores cercanos a 1cm/año. Árboles de este tamaño, cuando tienen condiciones de iluminación óptimas, pueden alcanzar hasta 2 cm/año. Esto permite prever que los turnos de corta para cedro podrían ser reducidos significativamente con medidas silvícolas que mejoren las condiciones de iluminación de los individuos de interés comercial (Gasparri y Goya este volumen). En las etapas de establecimiento, variables adicionales como la dinámica de claros, que favorecen el crecimiento de las plántulas establecidas, y la presión de pastoreo, son relevantes en asegurar el éxito de la renovación (Zamora este volumen). Sin embargo, estos estudios no son concluyentes y se requiere mayor esfuerzo de muestreo y mayor extensión geográfica del estudio, para asegurar una relación causal entre clareos y disminución de la presión de ganado con un mejor reclutamiento de ejemplares de cedro. Observaciones realizadas muestran que en áreas bajo intensa actividad forestal, el reclutamiento es deficiente a pesar de que los claros son abundantes. De la misma manera, en áreas de fuerte presión ganadera, con disturbios intensos como incendios, se observa una importante regeneración. También la herbivoría surgió como un factor relevante en la dinámica de reclutamiento del cedro. Se registró que 15 Brown A. y S. Pacheco un mayor crecimiento de los renovales se encuentra asociado al menor consumo por insectos en tanto que la supervivencia aumentó con el grado de protección a insectos y otros herbívoros (Scandalo et al. este volumen). La intensa tala selectiva que sufrieron los cedros en las Yungas argentinas llevó a sus especies a una situación poblacional crítica, al menos desde la óptica productiva. La intensidad de las explotaciones, los escasos conocimientos técnicos silvícolas, el ineficiente control de los organismos oficiales competentes y la escasa transparencia del mercado forestal en la región, son condicionantes del actual estado de las especies de cedro en los bosques nativos del noroeste de Argentina. Una medida importante para contribuir a salvaguardar el futuro de los cedros de las Yungas, es la reforestación a través de enriquecimientos o macizos con fines comerciales, aliviando la presión sobre las formaciones nativas (Minetti este volumen). Es fundamental que esta materia prima se genere en lugares accesibles que reduzcan los valores de flete y el impacto de abrir y mantener caminos en áreas de fuertes pendientes y lluvias torrenciales. La producción de plantines de Cedrela en cantidad y calidad, es fácilmente accesible y exitosa a través de la producción en viveros (Monteverde este volumen). En ese sentido una serie de experiencias se han desarrollando durante la última década, combinando distintas especies, densidades y tareas de mantenimiento (Horlent y Monteverde este volumen, del Castillo et al. este volumen). Estas experiencias generan la opción de que 16 las plantaciones forestales con especies nativas (como cedro) sean una alternativa productiva (a cielo descubierto o en fajas de enriquecimiento), frente a plantaciones con especies exóticas de rápido crecimiento (del Castillo et al. este volumen, Morenghi este volumen), e incluso frente a cultivos agrícolas como la soja. En ese sentido, técnicas de reproducción in vitro de cedro permitirán desarrollar plantaciones mas homogéneas genéticamente permitiendo seleccionar aquellos genotipos con más potencial para el desarrollo de plantaciones comerciales (Enrico y Enrico este volumen). Adicionalmente, estas plantaciones podrán cumplir la función de restaurar antiguas áreas de bosque en sitios ambientalmente estratégicos como márgenes de ríos o corredores para la fauna, que permitan reconectar áreas actualmente disyuntas. El desarrollo de Plantaciones MDL (Mecanismo de Desarrollo Limpio) que generan recursos adicionales a partir de la comercialización de bonos de carbono, podría ser una herramienta financiera importante para generar un cambio de paradigma productivo que permita utilizar la importante disponibilidad de mano de obra regional, retomando el rol protagónico que tuvieron los bosques y la madera en el desarrollo económico de la región. Los cedros por su alto valor de mercado, su productividad y la relativa facilidad para manejarlos en plantaciones o en el interior de bosques nativos, podrán ser un elemento clave en este proceso asociado a la certificación forestal que permitirá a los productores forestales de la región ser más competitivos tanto en el mercado interna- Importancia del género Cedrela en la conservación y desarrollo sustentable de las Yungas australes cional como posiblemente en el nacional en un futuro no muy lejano (Zorrilla este volumen). En cuanto a la preservación al largo plazo de las poblaciones silvestres de cedro, la situación es claramente contrastante entre las distintas especies. Por un lado, C. lilloi mantiene aún poblaciones prístinas en muchas quebradas protegidas por las dificultades de acceso. Además, presenta poblaciones importantes en el interior de reservas nacionales (PN El Rey, PN Calilegua, PN Baritú y RN El Nogalar) y algunas provinciales (por ejemplo La Florida y San Javier en Tucumán) que preservan poblaciones de distintos sectores geográficos (Norte, Centro y Sur) de las Yungas. Estas reservas preservan también importantes bosques viejos en distintos niveles altitudinales con lo que la diversidad genética de la especie estaría asegurada. La situación de C. balansae es diferente ya que su distribución geográfica es marginal a la ubicación de las reservas y en ninguna de ellas, hasta donde se conoce en la actualidad, se preservan bosques viejos de la especie. Se encuentran poblaciones de cedro orán en los alrededores del PP Pintascayo (Salta), en el PN Calilegua (Jujuy) y aledaños a este último, hay una cantidad importante en propiedad privada (Ingenio Ledesma). La superficie total de distribución de esta especie en la Alta Cuenca del Río Bermejo es de 8.500 km², de los cuales sólo 600 km² se encuentra dentro de áreas protegidas. En ese sentido, será relevante generar convenios de largo plazo para la preservación de estos ejemplares. En relación al cedro rosado (C. saltensis) dada su reciente identidad taxonómica, aún se desconoce su estatus regional, aunque poblaciones importantes podrían encontrarse en el interior del PN Baritú y del PN Calilegua dada su distribución altitudinal propuesta. Esperamos que los distintos artículos que conforman este libro generen la inquietud y el interés para desarrollar en forma sustentable esta opción productiva en la región y para generar en la sociedad y en las instituciones gubernamentales, la preocupación sobre el futuro de las poblaciones silvestres de las distintas especies de cedro y los sistemas naturales que los albergan. BIBLIOGRAFIA (sólo citadas las que no representan capítulos de este volúmen) Brown, A.D.; H.R. Grau; L. Malizia y A. Grau. 2001. Los Bosques Nublados de la Argentina. En: Bosques Nublados de Latinoamerica. M. Kappelle y A.D. Brown (Eds.). Editorial INBio, Costa Rica. pp: 623-659. Brown, A.D.; A.Grau; T. Lomáscolo y N.I. Gasparri. 2002. Una estrategia de conservación para las selvas subtropicales de montaña (Yungas) de Argentina. Ecotropicos 15: 147-159. 17 Brown A. y S. Pacheco Brown, A.D. y L.R. Malizia. 2004. Las Selvas Pedemontanas de las Yungas: en el umbral de la extinción. Ciencia Hoy 14: 52-63. Gasparri, N.I. y H.R. Grau. 2006. Patrones regionales de deforestación en el subtrópico argentino y su contexto ecológico y socioeconómico. En: La Situación Ambiental de la Argentina. Brown, A.D.; U. Martinez Ortiz; M. Acerbi y J. Corcuera (Eds.). Fundación Vida Silvestre Argentina. pp: 442-446. Mac Donagh, P. y L. Rivero 2006. ¿Es posible el uso sustentable de los bosques de la Selva Misionera? En: La Situación Ambiental de la Argentina. Brown, A.D.; U. Martinez Ortiz; M. Acerbi y J. Corcuera (Eds.). Fundación Vida Silvestre Argentina. pp: 210-217. Malizia, L. 2004. Diversity and distribution of tree species in subtropical andean forest. Tesis doctoral. Universidad de Missouri, St. Louis. Placci, G. y M. Di Bitetti. 2006. Situación Ambiental en la Ecoregión del Bosque Atlántico del Alto Paraná (Selva Paranaense). En: La Situación Ambiental de la Argentina. Brown, A.D.; U. Martinez Ortiz; M. Acerbi y J. Corcuera (Eds.). Fundación Vida Silvestre Argentina. pp: 197-225. 18 Ecología y producción de cedro (género Cedrela) en las Yungas australes. Pacheco, S. y A. Brown (Eds.). LIEY-ProYungas. Argentina. pp. 19-30. 2006. Botánica y distribución del género Cedrela en el noroeste de Argentina Grau, A.1; M. A. Zapater 2 y R. A. Neumann 2 1 Laboratorio de Investigaciones Ecológicas de las Yungas (LIEY, UNT). CC 34 (4107), Yerba Buena, Tucumán, Argentina. [email protected] 2 Facultad de Ciencias Naturales, UNSA. En el noroeste de Argentina el género Cedrela está representado por tres especies: C. lilloi, C. balansae y C. saltensis. En el pasado, los nombres C. angustifolia y en menor medida C. odorata y C. fissilis han sido usados en lugar de C. balansae. Esta especie es característica de la Selva Pedemontana o de Transición, sobre tierras planas o serranías bajas de las Provincias de Salta y Jujuy, extendiéndose desde el límite con Bolivia hasta aproximadamente 24º30’S, entre 300 y 800 m s.n.m. Cedrela lilloi es una especie característica de la Selva Montana y Bosque Montano en las Provincias de Salta, Jujuy, Tucumán y Catamarca, extendiéndose desde el límite con Bolivia hasta aproximadamente 28º 15’ S, entre 1.100 y 2.300 m s.n.m. en la Provincia de Salta, y entre 500 y 1.900 m s.n.m. en Tucumán y Catamarca. Cedrela saltensis es una especie descripta recientemente, que ocupa una franja montañosa en las Provincias de Salta y Jujuy, a una altura entre 700 y 1.100 m s.n.m., entre las áreas de distribución de C. balansae y C. lilloi, desde el límite con Bolivia al norte hasta los 24º 40’ S, aproximadamente. Se realiza una descripción de las características morfológicas más relevantes del género y se presenta una tabla comparativa que permite diferenciar las distintas especies. The genus Cedrela is represented by three species in northwestern Argentina: C. lilloi, C. balansae and C. saltensis. In the past, C. angustifolia and, to a lesser extent C. odorata and C. fissilis, have been used as names instead of C. balansae. This species is characteristic of the Piedmont or Transitional Forest, on lowlands or low ranges in Salta and Jujuy Provinces, extending from the Bolivian border to approximately 28º 15’ S, between 300 y 800 m asl. Cedrela lilloi is a characteristic species in the Montane Cloud Forest in the mountains of Salta, Jujuy, Tucumán and Catamarca, expanding from the Bolivian border to approximately 28º 15’ S, between 1.100 y 2.300 m asl in northern Salta Province, and between 500 and 1.900 m asl in Tucumán and Catamarca Provinces. Cedrela saltensis is a recently described species, present along a montane strip in Salta y Jujuy, from 700 to 1.100 m asl, between C. balansae and C. lilloi distribution areas, from the Bolivian border to 24º 40’ S approximately. A description of the most relevant morphological features of the genus and a comparative table to distinguish the different species is presented. 19 Grau, A; M. A. Zapater y R. A. Neumann IntroducciOn Desde los comienzos de la exploración y colonización de América Latina, los cedros han estado entre las especies forestales más importantes. Su madera blanda y fácil de trabajar, pero a la vez resistente para su densidad, sumada a su perfume agradable y tolerancia al ataque de insectos una vez cortada, han sido los atributos que le han ganado un lugar entre las mejores maderas del mundo. El nombre cedro ha sido tomado por el parecido con la madera de alta calidad del verdadero cedro, correspondiente al género de coníferas Cedrus (Pinaceae), conocido en el Mediterráneo desde tiempos bíblicos. En el lenguaje corriente el nombre cedro suele ir acompañado localmente de algún adjetivo que hace referencia a alguna característica destacable, como color de la madera o localidad de origen (por ejemplo cedro coya, cedro orán, cedro rosa). Cedro es también el término empleado para las distintas especies del género que aparecen en Brasil. Términos en idiomas nativos no se usan en el noroeste de Argentina ni aparecen en la literatura local, pues parecen haber sido mayormente desplazados en América Latina por el término español y portugués. En inglés se emplea “Spanish cedar”, que probablemente hace referencia a la madera de Cedrela originalmente comercializada por los españoles a partir del Caribe. En Centroamérica el cedro fue, y en parte todavía es, una madera de elección para estructuras, canoas y muebles. En los procesos de explotación forestal de esa región la primera madera aprovechada era la caoba (Swietenia macrophylla), y en segundo 20 lugar el cedro. Por su parte, en el noroeste de Argentina, el cedro ocupó siempre el primer lugar entre las maderas buscadas en cada sitio de aprovechamiento forestal. El Cedro y sus parientes Cedrela pertenece a las Meliaceae, una familia de árboles y arbustos predominantemente tropical. Este género se ubica dentro de la subfamilia Swietenioideae conjuntamente con Swietenia, Khaya y Toona, todos con especies productoras de maderas valiosas. Esta ubicación, propuesta en la revisión taxonómica clásica de Pennington y Styles (1975) ha sido recientemente confirmada usando secuencias de ADN nuclear y plastidial (Muellner et al. 2003). Khaya es originario de África, y fue recientemente introducido en el sur de Salta en el establecimiento La Moraleja en el Departamento de Anta. Toona es originario del sudeste de Asia y Australia. En el pasado Toona se consideraba incluido dentro de Cedrela, pero actualmente este género se restringe a las especies americanas. Toona ciliata var. australis (= T. australis), originario de Australia, se ha difundido ampliamente en plantaciones forestales en países tropicales y subtropicales, existiendo ensayos y plantaciones comerciales a cielo abierto y bajo cubierta en el noroeste de Argentina. Swietenia macrophylla, la caoba o mara boliviana tiene su extremo de distribución sur alrededor de los 17ºS en Bolivia (Mayhew y Newton 1998), y no crece silvestre en Argentina. Existen ejemplares cultivados en Libertador General San Martín y ensayos en La Moraleja (Salta) y en el INTA Yuto (Jujuy). Botánica y distribución del género Cedrela en el noroeste de Argentina El gEnero Cedrela El nombre botánico Cedrela fue empleado primeramente por Linneo, como deformación de Cedrus. Linneo (1759) creó el género en base a una lámina publicada por Browne (1756), en su descripción de la historia natural de Jamaica. La lámina mencionada representa claramente a una especie del género Cedrela y en base a ella Linneo describió a Cedrela odorata, aunque existen ciertas dudas sobre si el material representado corresponde plenamente a esta especie (Styles 1981). El género Cedrela está claramente definido y sus especies pueden distinguirse con relativa facilidad de los géneros afines, Swietenia y Toona. Por el contrario, la identificación de las distintas especies dentro de Cedrela es muchas veces dificultosa y ha dado lugar a confusiones. Esta dificultad en definir claramente las especies ha contribuido a la proliferación de especies creadas, que se acercan a unas 40, de las cuales sólo unas ocho se aceptan actualmente como válidas y cinco están presentes espontáneamente en Argentina (Styles 1981, Zapater et al. 2004). En el noroeste de Argentina tradicionalmente se aceptó la presencia de dos especies. Una de ellas, C. lilloi, es una especie característica de ambientes de montaña, claramente definida y descripta por C. de Candolle en base a material coleccionado por Miguel Lillo a comienzos del siglo XX. La segunda especie presente, característica de tierras planas y serranías bajas, fue originalmente identificada como C. balansae (por ejemplo T. Meyer en ma- terial de herbario LIL o Tortorelli 1956). Se trata de una especie descripta para el Paraguay, también por C. de Candolle. Sin embargo, más tarde Smith (1960), en una revisión centrada en el género, aunque sin visitar Argentina, clasificó el material argentino de esta especie como C. angustifolia, una especie descripta para México. Ese criterio fue adoptado por Legname (1982), en su trabajo sobre los árboles nativos del noroeste de Argentina. Por otro lado, en una revisión más completa y reciente del género y la subfamilia Swietenioideae para América, aunque también sin visitas al terreno en Argentina, Styles (1981) descartó el nombre C. angustifolia y puso en duda su validez taxonómica. Además, consideró que los materiales previamente identificados como C. balansae o C. angustifolia pertenecían en cambio a C. odorata o C. físsilis. Lamentablemente éste fue el criterio adoptado por Zuloaga y Morrone (1999) en el Catálogo de las Plantas Vasculares de la Argentina. Sin embargo, recientemente Zapater et al. (2004), en un análisis más completo que el de los autores previos, e incluyendo un amplio estudio de campo, reinstalaron a C. balansae como el nombre correcto para identificar esta especie. Por otro lado, descartaron la presencia de poblaciones silvestres de C. odorata y C. fissilis en el noroeste de Argentina, que sí están presentes en el noreste de Argentina. Además, estos autores determinaron la existencia de otra especie de Cedrela en la región, nueva para la ciencia, que describieron y denominaron C. saltensis Zapater y del Castillo. Cedrela saltensis es una especie muy afín a C. balansae, de 21 Grau, A; M. A. Zapater y R. A. Neumann la que se diferencia por el mayor tamaño y pubescencia de los folíolos, la reducida longitud de los peciólulos, y por sus frutos oscuros con lenticelas predominantemente alargadas, no anastomosadas. DescripciOn general del gEnero Cedrela en el noroeste de Argentina En este punto se realiza la descripción del patrón general al cual responden las especies nativas de Cedrela en el noroeste de Argentina y las diferencias más relevantes entre ellas. Para una descripción detallada de cada una de las especies consultar a Zapater et al. (2004). causa, individuos de C. balansae, situados en lugares en general más accesibles suelen presentar dimensiones menores que los de C. lilloi. El tronco es cilíndrico, aunque a veces, particularmente en C. lilloi, presenta la tendencia a aplanarse marcadamente en un sentido, cuando se encuentra creciendo en pendientes pronunciadas. La base suele presentar costillas marcadas, hasta 2 m de altura, en los ejemplares longevos. La copa de los ejemplares jóvenes es más bien angosta, pero se transforma en ancha, densa y ampliamente ramificada en ejemplares maduros (Foto 2). Se trata de árboles deciduos, que a la ma- Tanto ramas y ramitas como pecíolos y durez pueden alcanzar 20 - 30 m de altura raquis están densamente lenticelados. Las y 1,5 m de diámetro a la altura del pecho ramas y hojas presentan pubescencia. Esta (DAP), con una copa más o menos esfé- es muy abundante en C. saltensis, algo rica o en forma de paraguas, emergentes menos en C. balansae y muy poco en del nivel general del bosque. Arboles de estas dimensiones todavía son explotados con relativa frecuencia. Es posible encontrar árboles de dimensiones mayores (excepcionalmente de hasta 3 m), en lugares aislados y de difícil acceso (Foto 1). Seguramente ejemplares de estas dimensiones fueron frecuentes al inicio del proceso moderno de aprovechamiento sistemático de los bosques del noroeste de Foto 1. Ejemplar de cedro coya (Cedrela lilloi) en Osma, Salta, Argentina, a comienzos aproximadamente a 1.800 m s.n.m., con 3 m DAP y unos 25 m del siglo XX. Por esta de altura. 22 Botánica y distribución del género Cedrela en el noroeste de Argentina C. lilloi, donde suele restringirse a las nervaduras y peciólulos. Es poco lo que se conoce sobre las raíces de Cedrela. En el caso de C. balansae se ha observado que tienen la capacidad de regenerar tallos, una vez que el eje principal ha sido cortado (Fotos 3 y 4). Esta propiedad de rebrotar ha sido observada como respuesta a incendios, por lo cual éste fenómeno podría aumentar la densidad de individuos por hectárea. Las hojas son alternas, paripinadas, con (5-) 7 - 10 (-13) pares de folíolos opuestos o subopuestos, predominantemente oval lanceolados en C. balansae y C. lilloi, y generalmente elípticos en C. saltensis (Fig. 1). Sin embargo, la pubescencia y el tamaño de los peciólulos permite separar las especies mejor que la forma de los folíolos. Las cicatrices foliares son de forma semicircular y muy marcadas (Foto 5). Foto 2. Ejemplar de cedro coya (Cedrela lilloi) que ha crecido fuera del bosque, en la localidad de Horco Molle, Tucumán. Figura 1. Detalle de folíolos de Cedrela: A - cedro coya (C. lilloi), B - cedro rosado (C. saltensis) y C - cedro orán (C. balansae). La floración se produce durante los meses de noviembre y diciembre en C. lilloi y C. saltensis, en tanto que en C. balansae se extiende hasta el mes de enero. Las flores son pequeñas, de 7,0 - 8,5 mm de longitud y se encuentran en inflorescencias consistentes en tirsos multifloros, terminales o axilares, con varios cientos de flores. Las flores son pentámeras. Styles (1981) considera que en Cedrela todas las flores son imperfectas (es decir unisexuales), aunque con vestigios bien desarrollados del verticilo fértil opuesto. Este factor hace difícil la observación y definición clara de la presencia de flores imperfectas, particularmente en material de herbario. Aunque en C. balansae y C. saltensis no existen observaciones definitivas al respecto, en C. lilloi se ha determinado claramente la presencia de flores imperfectas y una marcada alternancia en la maduración y apertura de 23 Grau, A; M. A. Zapater y R. A. Neumann el androginóforo. En el caso de C. lilloi, las flores estaminadas poseen anteras desarrolladas, amarillas y dehiscentes. En las flores pistiladas en cambio éstas son pequeñas, arrugadas, marrones y carentes de polen. El ovario es pentalobado, pentalocular, con lóbulos pluriovulados, situado sobre el androginóforo. El estigma es discoide, papilado, situado sobre un estilo breve. Foto 3. Desarrollo de tallos a partir de un sis- tema radical de un ejemplar cortado de cedro orán (Cedrela balansae), en la localidad de Aguas Blancas, Salta. Puede verse el tronco original cortado tirado en el suelo y vigorosos retoños de raíz. El fruto es una cápsula ovoide a obovoide de dehiscencia septicida, que comienza a desarrollarse a partir del mes de diciembre y completa su maduración abriéndose en julio- agosto. El epicarpio de las cápsulas puede ser de color castaño claro a pardo oscuro y el pericarpio posee un espesor variable entre 1 - 3 mm según las especies. Posee exteriormente un diseño de lenticelas variable según las especies: prominentes, redondeadas y dispersas en C. lilloi; alargadas y aproximadas en C. saltensis, e inconspicuas y anastomosadas en C. balansae (Foto 6). Las semillas son de color castaño o pardo según la especie, con un ala bien desa- las flores pistiladas y estaminadas en cada árbol particular (Aschero este volumen). El cáliz, pubescente, está formado por cinco dientes o lóbulos irregulares con 1 - 2 hendiduras profundas hasta la base. Los pétalos son libres, pubescentes en el envés, blanquecino-amarillentos, imbricados y casi en contacto en su mayor parte con la columna de estambres y estilo lo que genera un aspecto de soldadura. Los cinco estambres por su parte, se originan y están en contacto en su mayor parte con 24 Foto 4. Detalle de un vigoroso retoño de raíz del grupo presente en la Foto 3. Botánica y distribución del género Cedrela en el noroeste de Argentina Distribucion El género Cedrela tiene una distribución predominantemente tropical, encontrándose en todos los países de América Latina a excepción de Chile. En su extremo septentrional llega en México hasta los 20º N. En Argentina C. odorata y C. fissilis están presentes en el nordeste (Misiones y Corrientes). Por su parte C. lilloi, C. balansae y C. saltensis están presentes en el noroeste (Fig. 2). Foto 5. Cicatrices foliares en el tallo de un ejemplar joven de cedro orán (Cedrela balansae). Las zonas de crecimiento longitudinal más lento durante el reposo invernal quedan marcadas por una mayor densidad de cicatrices foliares y pueden emplearse para estimar la edad de los ejemplares jóvenes. rrollada más grande que la porción de los cotiledones. La dispersión es por viento, y dado que la época de maduración de los frutos coincide con una mayor frecuencia de vientos del cuadrante N - NE, hay mayor densidad de plántulas hacia el sur de los árboles semilleros. Posteriormente, las lluvias del verano pueden dispersar las semillas por arrastre de agua en el sentido de la pendiente de cada sitio. Cedrela balansae se encuentra en los sectores de Selva Pedemontana de las Provincias de Salta y Jujuy, entre los 300 y 700 m s.n.m., al norte del paralelo 24º 30’ S y hasta el límite con Bolivia, con lluvias anuales de 700 a 1.500 mm. En la actualidad esta especie es más abundante en las zonas con serranías bajas. En las áreas de la Selva Pedemontana se ha visto muy reducida en su distribución sobre suelos planos y profundos, debido al aprovechamiento forestal intenso o a la transformación del bosque en campos agrícolas. Aunque esta especie se ha citado para las Provincias de Tucumán y Catamarca como En la Tabla 1 se presentan las características distintivas de cada una de las especies Foto 6. Cápsulas de cedro coya (Cedrela lilloi), cedro rosado (Cedrela saltensis) y cedro orán (Cedrela balansae). del género Cedrela. 25 Grau, A; M. A. Zapater y R. A. Neumann Tabla 1. Cuadro comparativo de las especies nativas del género Cedrela en el noroeste de Ar- gentina. *La denominacion cedro rosado suele usarse en las tres especies. C. lilloi C. saltensis C. balansae Ancho de folíolos y ápice 2,7 - 3,5 cm. 5,5 - 7 cm. 4,5 - 6,8 cm. ápice largamente agudo, subulado a filiforme. ápice agudo ápice agudo Pilosidad de los folíolos glabros, brillantes pubescentes en el haz y tomentosos en el envés glabros en el haz, subglabros a pubescentes en el envés Peciólulos 7 - 10 mm de largo, delgados, glabros hasta 2 mm de largo, gruesos, muy pubescentes 4 - 5 mm de largo, delgados, pubescentes Olor del follaje moderado suave, ligeramente desagradable muy fuerte y desagradable Cápsula obovoidea, a veces apiculada, parda oscura, con lenticelas blanquecinas redondeadas, no anastomosadas, prominentes ovoidea, parda oscura, con lenticelas ocráceas alargadas, no anastomosadas, algo prominentes ovoidea, castaño clara a castaña, lenticelas ocráceas alargadas no prominentes, anastomosadas, a menudo incospicuas. Color de las semillas castaño oscuro castaño oscuro castaño claro Nombres comunes en la región cedro coya, cedro tucumano, cedro rosado* cedro salteño, cedrillo, cedro rosado* cedro orán, cedro rosado* 26 Botánica y distribución del género Cedrela en el noroeste de Argentina Figura 2. Distribución del género Cedrela en el noroeste de Argentina. Cada punto repre- senta un ejemplar de herbario o una localidad donde la especie ha sido observada. 27 Grau, A; M. A. Zapater y R. A. Neumann escasa (Zapater et al. 2004), existen ahora dudas sobre su distribución austral. Los escasos ejemplares analizados ofrecen caracteres confusos e intermedios entre C. balansae y C. lilloi, por lo que deberían ser objeto de estudios más profundos. Por otra parte, resulta llamativa la falta de colecciones en los departamentos del sur de Salta, que conectan a dicha provincia con Tucumán y el extremo sur de la región. Cedrela lilloi es la especie con distribución más amplia en Argentina, extendiéndose desde el norte de la Provincia de Salta (22º S) hasta la Provincia de Catamarca (aproximadamente 28º 15’ S). En el norte de la Provincia de Salta se encuentra por encima de los 1.100 y hasta los 2.300 m s.n.m. En la Provincia de Tucumán ocupa un gradiente más amplio, entre los 500 y 1.900 m s.n.m. y un rango de precipitaciones entre 800 y 2.000 mm anuales. Cedrela saltensis está presente desde el norte de la Provincia de Salta hasta la Provincia de Jujuy 24º 40’ S, entre los 700 y 1.100 m s.n.m. EtnobotAnica y propiedades medicinales La familia Meliaceae tiene como característica la presencia de compuestos triterpénicos, usualmente células secretoras dispersas en los tejidos. En general, los compuestos almacenados en este tipo de estructuras tienen una acción relevante contra la herbivoría. En C. odorata se identificaron compuestos de naturaleza terpénica asociados al rechazo del follaje como alimento en coleópteros curculió28 nidos (Veitch et al. 1999). Sin embargo, todas las especies de Cedrela y Swietenia son atacadas por la mariposita del brote (Hypsipyla grandella), cuyas larvas atacan plántulas desde 0,5 a 3 m de altura en toda América Latina. Además de la actividad biológica contra insectos, en Cedrela se ha identificado actividad biológica contra microorganismos. Orientados por los usos tradicionales contra la malaria, distintos grupos han aislado la gedunina en C. odorata, un limonoide de naturaleza terpénica con acción in vitro contra Plasmodium falciparum, agente causante del paludismo (McKinnon et al.1997, Omar et al. 2003). Por otro lado, en C. lilloi se ha determinado la existencia de una importante actividad moduladora del sistema inmunitario, reduciendo las reacciones de hipersensibilidad (Nores et al. 1997). Esta actividad ha sido estudiada en otras especies de Cedrela y no está asociada a los compuestos de naturaleza terpénica sino a compuestos hidrosolubles (Benencia et al. 1995). Sin embargo, estudios etnobotánicos realizados en las Yungas del noroeste de Argentina no han detectado evidencias del uso de especies del género Cedrela como planta medicinal por parte de las comunidades locales (Levy-Hynes 1994, Hilgert 1998). Botánica y distribución del género Cedrela en el noroeste de Argentina BIBLIOGRAFIA Aschero, V. 2006. Biología reproductiva e importancia de la polinización en Cedrela lilloi. En Ecología y producción de cedro (género Cedrela) en las Yungas australes. Pacheco, S. y A. Brown (Eds.). LIEY- ProYungas. Argentina. pp 41-50. Benencia, F.; M.C. Courreges; M.M. Nores y F.C. Coulombie. 1995. Immunomodulatory activities of Cedrela tubiflora leaf aqueous extracts. Journal of Ethnopharmacology 49 (3):133-139. Browne, P. 1756. Civil and natural history of Jamaica, pl. 10. London (Osborn y Shipton). 158 pp. Hilgert, N. 1998. Las plantas vinculadas con el ámbito doméstico y la subsistencia de los campesinos de la cuenca del río Zenta, Dpto. Orán, Prov. Salta. Tesis Doctoral. Universidad Nacional de Córdoba. 342 pp. Legname, P.R. 1982. Árboles indígenas del Noroeste Argentino. Opera Lilloana 34. Tucumán, Argentina. 226 pp. Levy-Hynes, A. 1994. Uso de las plantas por las comunidades campesinas de las Yungas del extremo norte de Argentina. Tesis de Grado. Facultad de Ciencias NaturalesUniversidad Nacional de Tucumán. 42 pp. Linneo, C. 1759. Systema naturae (ed. 10). 2:940. MacKinnon, S.; T. Durst; J.T. Arnason; C. Angerhofer; J. Pezzuto; P.E. Sanchez-Vindas; L.J. Poveda y M. Gbeassor. 1997. Antimalarial activity of tropical Meliaceae extracts and gedunin derivatives. Jorunal of Natural Products 60 (4):336-341. Mayhew, J.E. y A.C. Newton. 1998. The silviculture of Mahogany (Swietenia macrophylla). CAB International, Wallingford, New York. 226 pp. Muellner, K.; R. Samuel; S.A. Johnson; M. Cheek; T.D. Pennington y M. Chase 2003. Molecular phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA sequences. American Journal of Botany 90 (3):471-480. Nores, M.M.; M.C. Courreges; F. Benencia y F.C. Coulombie. 1997. Immunomodulatory activities of Cedrela lilloi and Trichilia elegans aqueous leaf extracts. J. Ethnopharmacol. 55 (2):99-106. Omar, S.; J. Zhang; S. MacKinnon; D. Leaman; T. Durst; B.J. Philogene; J.T. Arnason; P.E. Sanchez-Vindas; L. Poveda; P.A. Tamez y J.M. Pezzuto. 2003. Traditionally-used antimalarials from the Meliaceae. Current Topics in Medicinal Chemistry 3 (2):133-9. 29 Grau, A; M. A. Zapater y R. A. Neumann Pennington, T.D. y B.T. Styles. 1975. A generic monograph of the Meliaceae. Blumea 22: 419-540. Smith, C.E. 1960. A revision of Cedrela (Meliaceae). Fieldiana: Botany 29:294-341. Styles, B.T. 1981. Swietenioideae. En Pennington, T.E. Meliaceae. Flora Neotropica 28:1-459. Tortorelli, L.A. 1956. Maderas y Bosques Argentinos. Acme Agency Press, Buenos Aires, Argentina. 646 pp. Veitch, N.C.; G.A. Wright y P.C. Stevenson. 1999. Four new tetranortriterpenoids from Cedrela odorata associated with leaf rejection by Exophthalmus jekelianus. Journal of Natural Products 62 (9): 1260-1263. Zapater, M.A.; E.M. del Castillo y T.D. Pennington. 2004. El género Cedrela (Meliaceae) en la Argentina. Darwiniana 42 (1-4): 347-356. Zuloaga, F. y O. Morrone (Eds.). 1999. Catálogo de las Plantas Vasculares de la República Argentina II. Fabaceae-Zigophyllaceae (Dicotyledoneae). Monographs in Systematic Botany from the Missouri Botanical Garden Volume 74. Missouri Botanical Garden Press. St. Louis. 1269 pp. 30 Ecología y producción de cedro (género Cedrela) en las Yungas australes. Pacheco, S. y A. Brown (Eds.). LIEY-ProYungas. Argentina. pp. 31-40. 2006. Fenología de cedro en el Parque Nacional Calilegua, Jujuy Fenología de dos especies de cedro en un gradiente altitudinal del Parque Nacional Calilegua, Jujuy Pacheco, S.1,2 y A. Yapur3 1 Fundación ProYungas, CC 34 (4107), Yerba Buena, Tucumán, Argentina. 2 Laboratorio de Investigaciones Ecológicas de las Yungas (LIEY, UNT) 3 Facultad de Ciencias Naturales - UNT. [email protected] Se estudió el patrón en la producción de hojas, flores y frutos de Cedrela balansae y Cedrela lilloi a lo largo de un gradiente altitudinal en el Parque Nacional Calilegua. Se trabajó en tres sitios ubicados a 600-900 (C. balansae), a 1.100 (C. balansae y C. lilloi) y a 1.400-1.600 m s.n.m. (C. lilloi). La expansión foliar, la floración y la senescencia tienen el mismo comportamiento fenológico para las dos especies a lo largo del gradiente altitudinal. Las diferencias se observan en la retención de hojas maduras y senescentes. En la parte baja, C. balansae se comporta como caducifolio ya que queda sin hojas maduras por siete meses, mientras que a 1.100 m s.n.m. conserva las hojas senescentes hasta la expansión. Los árboles de C. lilloi ubicados en el sitio más alto se comportan como semicaducifolios ya que pasan un mes sin hojas hasta el inicio de la brotación, mientras que en el sitio intermedio no tienen un período sin hojas. C. balansae produjo frutos los dos años de muestreo, mientras que C. lilloi produjo pocas flores en algunos individuos, que se perdieron antes de formar frutos. Se discute la ocurrencia de las fenofases y sus causas. This paper studies the pattern of leaf, flower and fruit production of Cedrela balansae and Cedrela lilloi through an altitudinal gradient in Calilegua National Park. There are three study sites, located at 600 - 900 (C. balansae), 1.100 (C. balansae and C. lilloi) and 1.400 - 1.600 (C. lilloi) m above sea level. Foliar expansion, flowering and senescence have the same phenological behaviour for both species through the gradient. There are differences in mature and senescent leaf retention. At lower altitude, the C. balansae behaves as deciduous, and has no mature leaves for seven months, while at 1.100 m a.s.l. it retains senescent leaves until leaf expansion. The C. lilloi trees located in the highest study site behave as deciduous, and have no leaves for a month until sprouting. In the intermediate site C. lilloi trees never loose their leaves at the same time. The C. balansae had fruit during the two years sample period, while the C. lilloi had only a few flowers in some individuals, which were lost before becoming fruit. This paper discusses phenology patterns observed and its causes. 31 Pacheco, S. y A. Yapur INTRODUCCION La fenología es el estudio de las fases del ciclo de vida de plantas o animales y su ocurrencia temporal a lo largo del año. Los datos básicos de los estudios fenológicos en árboles incluyen registros sobre la época, duración e intensidad de una o varias fenofases como fructificación, floración y foliación. Muchos de estos estudios están orientados a establecer las relaciones entre la ocurrencia de las fenofases y las características climáticas, o la disponibilidad de polinizadores o dispersores. En el caso de especies forestales, la fenología de floración y fructificación puede ser utilizada para determinar calendarios de colectas de semillas o de disponibilidad de frutos (Mariot et al. 2003). En cuanto a las estrategias fenológicas de las especies arbóreas, puede considerarse un gradiente de especies caducifolias o deciduas (pierden las hojas por más de tres meses en el año), especies semicaducifolias (pierden las hojas por uno o dos meses) y especies perennifolias o siempreverdes (conservan las hojas durante todo el año). En ambientes estacionales donde hay una estación de estrés por déficit hídrico o bajas temperaturas, los árboles en general tienen una estrategia caducifolia con pérdida parcial o total del follaje. En ambientes no estacionales, los árboles pueden retener el follaje maduro hasta la aparición de las hojas nuevas, de modo de conservar el tejido fotosintético a lo largo de todo el año. Sin embargo, la fenología puede entenderse como una respuesta plástica, y en un ambiente heterogéneo (espacial o temporalmente), algunos individuos de 32 la población pueden comportarse en forma distinta, con respuestas fenológicas dependientes de condiciones ambientales particulares. Las Selvas Subtropicales de Montaña del noroeste de Argentina o Yungas, se desarrollan a lo largo de un gradiente altitudinal desde los 400 a los 3.000 m s.n.m., donde se identifican pisos altitudinales con características ambientales contrastantes (Brown 1995a). Las especies de la Selva Pedemontana (400 - 700 m s.n.m.) están sometidas a temperaturas más altas y a un marcado déficit hídrico durante la estación del invierno (época seca), lo que lleva a una alta demanda evapotranspirativa que algunas especies evitan perdiendo el follaje durante esta estación. De un total de 115 especies de árboles de las Yungas analizadas, el 60 % de las especies de Selva Pedemontana son de régimen caducifolio (Brown y Malizia 2004). En la Selva Montana (700 - 1.500 m s.n.m.), sólo el 20 % de las especies son caducifolias durante el invierno (Brown 1995 b). En este ambiente la demanda evapotranspirativa durante el invierno es menor debido al aporte de las neblinas (Hunzinger 1995), al mayor almacenaje de agua en el suelo y a las temperaturas generales más bajas. El estrés del invierno no es debido al déficit hídrico, sino a las bajas temperaturas ya que son frecuentes las heladas y las nevadas. En el piso más alto de los Bosques Montanos (1.500 - 3.000 m s.n.m.) más del 80 % de las especies son caducifolias o semicaducifolias. Esto puede deberse a las bajas temperaturas del invierno y probablemente también a un mayor estrés hí- Fenología de cedro en el Parque Nacional Calilegua, Jujuy drico, dado que las lluvias disminuyen por encima de los 1.200 m s.n.m. Las especies con amplios rangos de distribución, que ocurren en más de un piso altitudinal, podrían tener comportamientos fenológicos distintos de acuerdo a su ubicación en el gradiente. Este trabajo se llevó a cabo con dos de las especies de Cedrela de las Yungas de Argentina: Cedrela balansae (cedro orán) en la Selva Pedemontana y Cedrela lilloi (cedro coya) en la Selva Montana y principalmente en el Bosque Montano. En el PN Calilegua, el rango de altura entre 900 y 1.100 m s.n.m. que corresponde a Selva Montana, puede considerarse como una zona de ecotono en la que se superponen las dos especies y representa la altura máxima en la que pueden encontrarse individuos de C. balansae y la altura mínima en la que puede distribuirse C. lilloi (Malizia et al. este volumen). Al encontrarse a lo largo del gradiente altitudinal, se espera que las dos especies tengan una respuesta fenológica acorde a las características de cada ambiente. Además, es esperable que dentro de cada especie, los individuos ubicados en la zona de ecotono, tengan un comportamiento fenológico distinto con respecto a la población presente en cada piso característico. El objetivo de este trabajo es presentar el patrón en la producción de hojas, flores y frutos de las dos especies de Cedrela a lo largo del gradiente altitudinal entre 600 y 1.600 m s.n.m. METODOS Area de estudio El trabajo se realizó en el Parque Nacional Calilegua (PN Calilegua) durante los años 2000 y 2001. El Parque tiene una superficie de 77.000 ha y representa principalmente el piso altitudinal de Selva Montana. También tiene una franja de Bosques Montanos y un parche de Pastizal de Neblina. La Selva Pedemontana ocupa sectores marginales del Parque en las áreas más bajas de ladera y pedemonte (Brown et al. 2002). Colección de datos climáticos Para obtener las características climáticas de los sitios de muestreo, se utilizaron datos climáticos obtenidos de la página WORLDCLIM (http://biogeo.berkeley. edu/worldclim/worldclim.html). Utilizando un Sistema de Información Geográfica se ubicaron puntos al azar dentro del PN Calilegua en las alturas de las estaciones de muestreo (600 a 900, 1.100 y 1.400 a 1.600 m s.n.m.). Luego se les asignaron los datos de precipitación media anual, estacionalidad en las precipitaciones, temperatura media anual, temperatura media de verano y temperatura media de invierno. Se calculó el promedio de cada variable climática para las tres alturas. Colección de datos de fenología Se trabajó con 70 árboles de las dos especies de Cedrela, distribuidos a lo largo de una transecta altitudinal (600 a 1.600 m s. n.m.), en seis estaciones de muestreo. Las estaciones de C. balansae estuvieron a 600, 700 - 900 y 960 - 1.100 m s.n.m. 33 Pacheco, S. y A. Yapur y las de C. lilloi a 960- 1.100, 1.400 y 1.600 m s.n.m. Las estaciones de 600 a 900 m s.n.m. se consideraron como típicas de la Selva Pedemontana, la estación de 1.400 a 1.600 m s.n.m. como típica de Bosque Montano y la estación de 960 1.100 m s.n.m. se consideró como Selva Montana y un área de ecotono donde se superponen las dos especies en el PN Calilegua. Mensualmente, se determinó la producción de hojas, flores y frutos (fenofases) mediante índices de abundancia: 0 (0 %), 1 (1 -5 %), 2 (6-25 %), 3 (26 - 50 %), 4 (51-75 %), 5 (76 -100 %). Los porcentajes se calcularon observando el total de la copa, por lo que un valor de 5 significa el 100 % de la copa con la fenofase observada. La categoría de hojas se dividió en hojas nuevas (hojas en desarrollo con el ápice aún enrollado, de color verde claro), hojas maduras (hojas totalmente expandidas, de color verde oscuro) y hojas senescentes (hojas amarillentas con pérdida de folíolos individuales). La categoría de flores se separó en botones florales y flores abiertas. La cantidad de frutos se determinó con una sola medida por año en el mes de mayo, antes de la apertura de los frutos y dispersión de las semillas. Debido a que no hubo diferencias entre las observaciones de los dos años, se analizó el comportamiento de cada especie en los tres sitios de muestreo usando el promedio de los dos años de observaciones para cada mes. RESULTADOS Características climáticas del Parque Nacional Calilegua La estación de muestreo de Selva Pedemontana en el PN Calilegua (600 a 900 m s.n.m.), se caracteriza por mayor temperatura media anual, de verano y de invierno y mayor precipitación media anual. En el sitio de Bosque Montano (1.400 a 1.600 m s.n.m.), los valores de temperaturas anuales y estacionales son más bajos al igual que la precipitación Figura 1. Características climáticas de las estaciones de muestreo en el Parque Nacional Ca- lilegua. A- Temperatura media anual, temperatura media de verano y temperatura media de invierno. B- Precipitación media anual. 34 Fenología de cedro en el Parque Nacional Calilegua, Jujuy media anual. En la zona de ecotono (960 a 1.100 m s.n.m.) los valores de temperatura y precipitación son intermedios (Fig. 1A y B). Fenología de Cedrela balansae La expansión foliar, la floración y la fructificación tienen el mismo comportamiento en cuanto a tiempo de ocurrencia, duración e intensidad, en la Selva Pedemontana y en el ecotono. La diferencia entre los dos ambientes se evidencia en las fases de hojas maduras y senescentes. Los árboles de la Selva Pedemontana se comportan claramente como caducifolios ya que quedan sin hojas maduras por siete meses (mayo a noviembre). Las hojas senescentes se conservan hasta el mes de junio y la mayoría de los árboles pierden el follaje maduro y senescente entre julio y octubre. Algunos individuos conservan hojas senescentes, pero con un porcentaje inferior al 1 % (Fig. 2A). En cambio, los árboles del ecotono no tienen un periodo sin hojas ya que pierden el follaje maduro en agosto y conservan las hojas senescentes hasta el momento de la expansión (Fig. 2B). La expansión foliar es sincrónica en los dos sitios (Selva Pedemontana y ecotono), comienza a fines de octubre y se extiende hasta mediados de diciembre (Fig. 2A y B). De diciembre a abril hay hojas maduras en la Selva Pedemontana. Los árboles del ecotono tienen hojas maduras de diciembre a agosto y tienen más cantidad de follaje. La senescencia empieza en mayo en los dos sitios. En la Selva Pedemontana casi todos los árboles pierden las hojas senescentes en junio, pero en el ecotono los árboles las conservan hasta noviembre cuando comienza la expansión (Fig. 2A y B). La floración es igual para los dos sitios, en diciembre aparecen los botones florales y las flores están abiertas hasta enero. Los árboles de los dos sitios produjeron frutos en los dos años de muestreo. La producción del año 2001 fue ligeramente mayor, coincidente con una mayor producción de flores. Los árboles Figura 2. Fenología de hojas de Cedrela balansae a lo largo del año. Cada punto es un pro- medio de los dos años de muestreo. A- individuos en Selva Pedemontana (600 - 900 m s.n.m.). B-individuos en ecotono (900 - 1.100 m s.n.m.). 35 Pacheco, S. y A. Yapur más chicos que produjeron frutos tenían 20 cm de diámetro a la altura del pecho. No hubo diferencias en la abundancia de frutos producidos por árbol (valor del índice), con relación al diámetro del árbol (rango de 20 a 92 cm de diámetro). Fenología de Cedrela lilloi La expansión foliar, la floración y el comienzo de la senescencia tienen el mismo patrón en el Bosque Montano y en el ecotono. La diferencia entre los dos sitios está dada por la retención de hojas maduras y senescentes. Los árboles del Bosque Montano se comportan como semicaducifolios ya que pasan un corto período sin hojas hasta el inicio de la brotación en septiembre. Pierden las hojas maduras al inicio de la estación seca y conservan las hojas senescentes hasta junio (Fig. 3A). Los árboles del ecotono prácticamente no tienen un período sin hojas porque con- servan hojas maduras hasta agosto y senescentes hasta la expansión de las hojas nuevas (Fig. 3B). La expansión foliar ocurre desde mediados de septiembre a mediados de octubre para los dos sitios. Los árboles del Bosque Montano tienen hojas maduras de noviembre a abril y los de ecotono, de diciembre a agosto. La senescencia comienza en mayo en los dos sitios y conservan hojas senescentes hasta agosto en el Bosque Montano y hasta noviembre en el ecotono (Fig. 3A y B). En el año 2000 la floración comenzó en noviembre en los tres sitios y fue abundante, pero en enero de 2001 se perdieron las flores. En el año 2001 sólo hubo flores en árboles de ecotono en el mes de noviembre, pero se perdieron en el mes de diciembre. En ninguno de los dos años hubo producción de frutos. Figura 3. Fenología de hojas de Cedrela lilloi a lo largo del año. Cada punto es un promedio de los dos años de muestreo. A- individuos en Bosque Montano (1.400 - 1.600 m s.n.m.). B- individuos en ecotono (900 - 1.100 m s.n.m.). 36 Fenología de cedro en el Parque Nacional Calilegua, Jujuy DISCUSION Las especies de Cedrela de las Yungas tienen un comportamiento fenológico similar a la comunidad vegetal del piso altitudinal al que pertenecen. C. balansae en la Selva Pedemontana es una especie caducifolia con pérdida del follaje maduro y senescente durante tres meses en la estación seca del invierno y C. lilloi en el Bosque Montano es una especie semicaducifolia con pérdida del follaje maduro y senescente durante casi dos meses en la estación del invierno. En el área del PN Calilegua donde se superponen las dos especies en el piso de la Selva Montana (1.100 m s.n.m.), Cedrela conserva el follaje maduro y senescente hasta la expansión de las hojas nuevas, por lo que no tienen un período caducifolio. El patrón general de producción de hojas, flores y frutos de Cedrela de las Yungas es similar al descrito para Cedrela mexicana en Costa Rica (Borcher 1983) y la época de floración y fructificación es similar a Cedrela fissilis en Brasil (Mantovani et al. 2003). El inicio de la expansión foliar en las dos especies de Cedrela es sincrónico en todo el gradiente altitudinal y en los dos años de muestreo. Para C. lilloi la expansión ocurre en el mes de octubre y para C. balansae, se inicia a fines de octubre, con un pico de expansión en noviembre. Esto es coincidente con el trabajo de Rivera et al. 2002 que definen a C. balansae como una especie de brotación primaveral, en las que la expansión es determinada por el aumento de la longitud del día cerca del equinoccio de primavera. En estas especies todos los árboles, independientemente del microam- biente en el que se encuentran, inician la expansión foliar en forma sincrónica, generalmente luego de dos y tres meses sin agua. El único factor constante entre años que podría controlar un patrón así, es el fotoperíodo (Thomas y Vince-Prue 1997). Dentro de la familia Meliacea, también se ha observado una sincronía en la expansión en Cedrela odorata en Manaus, Brasil (Lammi 2001 en Rivera et al. 2002). Es probable que la expansión antes del inicio de la época húmeda, permita optimizar la ganancia fotosintética ya que las hojas están expandidas al inicio de la estación de crecimiento. Esto puede ser una ventaja en ambientes estacionales con estación de crecimiento más corta. Las dos especies de Cedrela tienen hojas maduras durante la estación húmeda del verano y la senescencia comienza a fines de esta estación. Se ha propuesto que el comienzo de la senescencia a fines de la estación húmeda probablemente sea consecuencia de una combinación entre aumento en edad foliar y disminución del fotoperíodo, más que una respuesta al déficit hídrico (Borchert y Rivera 2001, Borchert et al. 2002). El inicio sincrónico de la senescencia en Cedrela a fines de la estación húmeda a lo largo del gradiente, podría deberse a esta combinación de factores. Sin embargo, la retención de las hojas maduras y senescentes varía entre las dos especies y entre sitios para la misma especie, lo cual sugiere una relación estrecha de la duración de esta fenofase con la humedad. C. balansae en la Selva Pedemontana es una especie caducifolia que pierde el follaje maduro durante los 37 Pacheco, S. y A. Yapur meses de la estación seca y C. lilloi en el Bosque Montano es una especie semicaducifolia que sólo está sin hojas un mes. En la zona de ecotono (con menor déficit hídrico y temperaturas más moderadas con relación a los pisos anteriores), las dos especies conservan sus hojas maduras hasta la expansión de las hojas nuevas. Es probable que en la Selva Pedemontana y en el Bosque Montano, el estrés por déficit hídrico y por bajas temperaturas respectivamente, limiten la retención del follaje. Al contrario de lo que ocurre en la zona de ecotono, donde tanto las precipitaciones como las temperaturas, son menos contrastantes entre el periodo de crecimiento y el de reposo. En este ambiente, puede ser una ventaja para el crecimiento retener las hojas maduras. Además, la retención de las hojas senescentes hasta el momento de la brotación, permite la translocación de nutrientes hacia las hojas nuevas. Las dos especies de Cedrela producen botones florales en noviembre y las flores están abiertas en diciembre y enero. Los frutos maduran durante la estación húmeda y la apertura ocurre a comienzos de la estación seca. La dispersión de las semillas es anemócora. Sin embargo, las dos especies tienen distinto comportamiento en cuanto a la edad de producción, a la cantidad y a la periodicidad de producción de frutos. C. balansae es una especie que produce flores y frutos todos los años. La producción puede comenzar en individuos de 20 cm de diámetro a la altura del pecho, que de acuerdo a la velocidad de crecimiento de esta especie, puede significar árboles de alrededor de 20 años. Esto es coincidente 38 con lo observado para Cedrela fissilis en Brasil (Mantovani et al. 2003). No hay una tendencia en la cantidad de producción y tamaño del árbol, aunque los árboles que produjeron más frutos fueron los de 50 a 60 cm de diámetro a la altura del pecho. C. lilloi produjo flores en los dos años de muestreo, pero se perdieron. La falta de producción de frutos registrada en Calilegua es un patrón regional ya que no se vieron frutos de esta especie en todo el gradiente de las Yungas desde hace por lo menos ocho años (Brown com. pers.). En el año 1997 hubo una producción abundante de frutos en Tucumán (obs. per.). No se sabe si esta falta de producción corresponde a un ciclo endógeno o a condiciones ambientales o a problemas con la polinización de las flores. La falta de semillas puede ser un problema en la colecta, por lo que sería importante conocer el ciclo de producción y desarrollar técnicas de conservación de semillas. Esta especie utiliza disturbios chicos dentro del bosque para establecerse. En consecuencia, la falta de producción anual de semillas es un factor limitante en la regeneración natural, ya que, su capacidad de colonización está condicionada a la coincidencia de años en los que haya producción de frutos y ocurrencia de disturbios adecuados en tamaño e intensidad. Agradecimientos Este trabajo fue financiado por el programa ProYungas y FOSDIC. Agradecemos a R. Grau y A. Brown por sus comentarios, y a los responsables del PN Calilegua. Fenología de cedro en el Parque Nacional Calilegua, Jujuy BIBLIOGRAFIA Borcher, R. 1983. Phenology and control of flowering in tropical trees. Biotropica 15 (2): 81-89. Borchert, R. y G. Rivera . 2001. Photoperiodic control of seasonal development and dormancy in tropical stem-succulent trees. Tree Physiology 21:13-221. Borcher, R.; Rivera G. y W. Hagnauer. 2002. Modification of vegetative phenology in a tropical semi-decidous forest by abnormal drought and rain. Biotropica 34 (1): 27-39. Brown, A.D. 1995a. Las selvas de montaña del noroeste de Argentina: problemas ambientales e importancia de su conservación. En: Investigación, Conservación y Desarrollo en Selvas Subtropicales de Montaña. Brown A.D. y H.R. Grau (Eds.). Proyecto de Desarrollo Agroforestal / LIEY. pp 9-18. Brown, A.D. 1995b. Fenología y caída de hojarasca en las Selvas Montanas del Parque Nacional El Rey, Argentina. En: Investigación, Conservación y Desarrollo en Selvas Subtropicales de Montaña. Brown A.D. y H.R. Grau (Eds.). Proyecto de Desarrollo Agroforestal / LIEY. pp 93-102. Brown, A.D.; A. Grau; T. Lomáscolo y N.I. Gasparri. 2002. Una estrategia de conservación para las selvas subtropicales de montaña (Yungas) de Argentina. Ecotrópico15: 147-159. Brown, A.D. y L.R. Malizia. 2004. Las Selvas Pedemontanas de las Yungas: en el umbral de la extinción. Ciencia Hoy 14 (83) 52–63. Hunzinger, H. 1995. La precipitación horizontal: su importancia para el bosque y a nivel de cuencas en la Sierra de San Javier, Tucumán, Argentina. En: Investigación, Conservación y Desarrollo en Selvas Subtropicales de Montaña. Brown A.D. y H.R. Grau (Eds.). Proyecto de Desarrollo Agroforestal / LIEY. pp 53 - 58. Malizia, L.; C. Blundo y S. Pacheco. 2006. Diversidad, estructura y distribución de bosques con cedro en el noroeste de Argentina y sur de Bolivia. En Ecología y Producción de cedro (género Cedrela) en las Yungas australes. Pacheco, S. y A. Brown (Eds.) LIEY-ProYungas. Argentina. pp. 83-104. Mantovani, M.; A.R. Ruschel; M. Sedrez dos Reis; A. Puchalski y R. O. Nodari. 2003. Fenologia reproductiva de espécies arbóreas em uma formação da floresta atlântica. R. Arvore, Viscosa-MG. 4: 451-458. 39 Pacheco, S. y A. Yapur Mariot, A.; M. Mantovani y J.C. Alencar. 2003. Uso e conservação de Piper cernuum Vell (Piperacea) na Mata Atlântica: I Fenologia reproductiva e dispersão de sementes. Revista Brasileira de Plantas Medicinais 5 (2): 1-10. Rivera, G.; S. Elliot; L.S. Caldas; G. Nicolossi; V.T.R. Coradin y R. Borchert. 2002. Increasing day-lenght induce spring flushing of tropical trees in the absence of rain. TREES 16 (7): 445-456. Thomas, B. y D. Vince-Prue. 1997. Photoperiodism in plants. Academic Press, San Diego. 40 Biología deCedrela) la polinización en Cedrela lilloi Ecologíareproductiva y producciónedeimportancia cedro (género en las Yungas australes. Pacheco, S. y A. Brown (Eds.). LIEY-ProYungas. Argentina. pp. 41-50. 2006. Biología reproductiva e importancia de la polinización en Cedrela lilloi Aschero, V. Laboratorio de Investigaciones Ecológicas de las Yungas (LIEY, UNT), CC 34 (4107), Yerba Buena, Tucumán, Argentina. Dirección Actual: CRICYT, CC 507, Av. Adrian Ruiz Leal s/n (5500), Mendoza, Argentina. [email protected] Los individuos de Cedrela lilloi portan flores unisexuales, es decir que sus flores son funcionalmente femeninas o masculinas aunque presentan vestigios no funcionales del otro sexo. A nivel inflorescencia, se observó separación temporal en la ántesis de las flores de cada sexo: la ántesis de las flores femeninas es intermedia a dos fases de antesis de flores masculinas. Esta asincronía de fases masculinas y femeninas entre las inflorescencias disminuye la probabilidad de autofecundación con polen del mismo individuo. Además, la producción de frutos y semillas fue mayor bajo polinización cruzada manual que bajo polinización autogama y natural. La separación temporal y espacial en los órganos sexuales funcionales y la influencia de la cantidad y calidad de polen en la producción de frutos y semillas, sugieren que la frecuencia de visitas y el comportamiento de los polinizadores serían factores importantes para la reproducción de la especie. The Cedrela lilloi trees have unisexual flowers, they function both as male or female. Nevertheless, each particular flower presents only vestiges of one of the sexes. In inflorescences, anthesis of flowers of different sexes is temporarily separated: anthesis of female flowers occurs within two events of male flowers anthesis. This flowering pattern diminishes the probability of self pollination. Furthermore, fruit and seed production was higher with manual crossed pollination, than in the self and natural pollination treatments. Temporal and spatial separation of sexual organs and influence of quantity and quality of polen in fruit and seed production suggest that frecuency of visits and foraging behaviour of pollinators would be important factors for reproduction in this species. 41 Aschero, V. IntroducciOn Las flores en las Angiospermas presentan una gran diversidad de tamaño, formas y colores y cumplen la función de promover la reproducción sexual. La mayoría de las plantas presentan flores hermafroditas, es decir, que portan órganos femeninos y masculinos (Richards 1997). Si bien la presencia del gineceo y estambres en una misma flor predomina, existen presiones selectivas que tienden a modificar la distribución de los órganos sexuales de tal patrón hermafrodita. Esta divergencia se explicaría principalmente como una estrategia para evitar la depresión por endogamia, al fomentar la fecundación de los óvulos con polen proveniente de otros individuos. De este modo, incluso en plantas con flores hermafroditas, la presentación del polen y del estigma puede presentar separación espacial (hercogamia) o temporal (dicogamia), disminuyendo la probabilidad de autofecundación y promoviendo la polinización cruzada. La calidad y la cantidad de polen depositado sobre el estigma para la fecundación de los óvulos, pueden limitar la producción de frutos y semillas. Para la mayoría de las plantas, los organismos responsables del proceso de polinización son los insectos (Proctor et al. 1996). El grado de dependencia de los polinizadores para la reproducción dependerá en parte de la distribución de sexos de la especie, siendo de mayor importancia en especies que presenten separación espacial o temporal en los órganos femeninos y masculinos. 42 Los estudios sobre el sistema reproductivo, la morfología y la fenología floral de las especies del género Cedrela son escasos y poco claros a pesar de la importancia económica de este género y del creciente interés en su producción y explotación sustentable. El objetivo de este trabajo es describir la expresión del sexo a nivel de flor e inflorescencia y la importancia de los polinizadores en la fructificación y producción de semillas en Cedrela lilloi. La sexualidad de las flores en el tiempo y en el espacio La sexualidad de las flores en Cedrela spp. ha sido interpretada de modos diferentes. En una de las primeras revisiones del género, Smith (1960) describió las flores como hermafroditas. Digilio y Legname (1966) también describen como hermafroditas las flores de C. lilloi. Sin embargo, Pennington (1981) en la última revisión de la familia Meliaceae describió las flores de Cedrela spp. como funcionalmente unisexuales, a pesar de presentar vestigios de órganos femeninos y masculinos. Dado que los estudios citados se basan principalmente en material vegetal herborizado y por lo tanto colectado en determinado momento de la floración, sería necesario realizar el seguimiento del desarrollo de las flores a campo para descartar la presencia de dicogamia o hercogamia en la especie. Por otra parte, a pesar de que Pennington (1981) describe las flores como unisexuales, no existen en su trabajo datos sobre la expresión del sexo a nivel de la inflorescencia. En Cedrela odorata las flores son unisexuales y a nivel de inflo- Biología reproductiva e importancia de la polinización en Cedrela lilloi rescencia, la antesis de flores pistiladas se da previamente a la antesis de flores masculinas (Cintron 1990). La importancia de los polinizadores en Cedrela Existe una gran variación temporal y espacial en la producción de frutos y semillas en C. lilloi (Pacheco y Brown com. pers.). Dicha variación, podría explicarse en parte por fallas en la polinización. La poca cantidad de polen depositado en el estigma por falta de polinizadores o la calidad del polen depositado, podrían afectar la producción de frutos y semillas en esta especie. La limitación en la producción de semillas por insuficientes visitas de polinizadores a las flores, ha sido registrada en Swietenia mahogany de la familia Meliacea (Mayhew y Newton 1998) y en otras especies (Parker 1997, Bierzychudek 1981, Galen 1985, Koptur 1984, Johnston 1991, Calvo 1993). metodos El trabajo se realizó en Horco Molle, Tucumán. El área se ubica a 650 m s. n. m. sobre la ladera este de la Sierra de San Javier, a unos 15 km de la ciudad de San Miguel de Tucumán. Para estudiar la sexualidad de las flores en el tiempo y el espacio a nivel de flor e inflorescencia, se escogieron cuatro individuos adultos de Cedrela lilloi. En ellos se marcaron dos inflorescencias por árbol y se observó cada dos días las flores abiertas durante todo el periodo de floración. Para describir la funcionalidad sexual de las flores, en el laboratorio se realizaron ex- perimentos de viabilidad del polen, receptividad del estigma y contenido de óvulos en el ovario. Para evaluar la viabilidad del polen, se realizaron tinciones de una antera por flor con solución de Alexander (Kearns e Inouye 1993) y se observó a microscopio la presencia de granos de polen viable (aquellos teñidos) en un campo de observación. La receptividad del estigma fue evaluada utilizando el método de la actividad de la enzima peroxidasa que consiste en embeber el estigma en una gota de solución de H2O2 sobre un portaobjetos y observar luego la presencia de burbujas sobre el estigma bajo lupa (la presencia de burbujas indica que el estigma está receptivo). Para observar la presencia de óvulos bajo lupa, se realizaron cortes transversales del ovario de cada una de las flores. En todas las flores recolectadas se analizó el contenido de óvulos y la presencia de polen en la antera bajo lupa. Algunas flores abiertas fueron colectadas y fijadas con la finalidad de analizar su morfología usando microscópica electrónica de barrido. Finalmente, para describir la sexualidad de la inflorescencia se registró la sexualidad de las flores en antesis a lo largo de la floración (la clasificación de las flores se realizó en base a las pruebas previamente descriptas). Para determinar la importancia de los polinizadores, se realizaron distintos tratamientos de polinización manual en cuatro individuos de Cedrela lilloi: 1) auto-polinización: se excluyeron los polinizadores embolsando inflorescencias con tela de tul, de modo que sólo puede depositarse el polen de la misma inflorescencia; 2) gei43 Aschero, V. tonogamia: se polinizaron manualmente las flores con polen del mismo individuo; 3) polinización cruzada: se polinizaron manualmente las flores con polen de los otros tres árboles; 4) polinización libre o control: las flores estuvieron disponibles a los polinizadores. Se marcaron inflorescencias con cables de color numerados para reconocerlas en el periodo de fructificación. Para cada uno de los tratamientos, se contó el número de flores presentes en cada una de las inflorescencias bajo estudio. En los tratamientos 2 y 3 las inflorescencias fueron embolsadas desde el comienzo de la floración. Una vez abiertas las flores, se retiraron las bolsas, se realizó el tratamiento y luego fueron embolsadas nuevamente hasta la cosecha de los frutos. En el laboratorio, se calculó para cada tratamiento el número de frutos en relación al número de flores funcionalmente femeninas que portaba cada inflorescencia y el número de semillas que contenía cada fruto. resultados En cuanto a la sexualidad de las flores en el tiempo y el espacio, se determinó que son funcionalmente unisexuales, a pesar de poseer vestigios no funcionales de órganos del otro sexo (Fig. 1). En las flores pistiladas (femeninas), el pistilo es robusto, las anteras no producen polen y se observan, a simple vista, atrofiadas y oscuras (Foto 1). En las flores estaminadas (masculinas), el pistilo es esbelto, las anteras producen abundante polen viable y los óvulos se encuentran atrofiados o ausentes (Foto 2). 44 figura 1. Esquema de flores estaminada y pistilada de Cedrela lilloi. En cada inflorescencia de C. lilloi ocurrió un único periodo de ántesis de flores femeninas intermedio a dos periodos de ántesis de flores masculinas. La proporción de flores masculinas producidas en la inflorescencia en relación a las femeninas es 4,03 ± 1,07. Entre las inflorescencias de un mismo individuo, se observó una marcada sincronía en la alternancia de las fases unisexuales, existiendo baja superposición temporal de los órganos femeninos y masculinos también a nivel individuo (Fig. 2). La sincronía de fases masculinas y femeninas entre las inflorescencias implica una disminución en la probabilidad de autofecundación con polen del mismo individuo. En cuanto a la importancia de los polinizadores en Cedrela lilloi, se observó que la producción de frutos por inflorescencia y de semillas por fruto, fue mayor en el tratamiento de polinización cruzada ma- Biología reproductiva e importancia de la polinización en Cedrela lilloi A B C D foto 1. Partes de la flor pistilada (femenina) de Cedrela lilloi. A - antera del estaminodio, B - superficie estigmática, C - gineceo, D - corte transversal del ovario. A B C D foto 2. Partes de la flor estaminada (masculina) de Cedrela lilloi. A- antera, B - polen, C - gineceo, D - corte transversal del ovario. 45 Aschero, V. Figura 2. Alternancia temporal en la antesis de flores masculinas y femeninas en los cuatro individuos de Cedrela lilloi estudiados. Sólo se representan los días con datos de muestreo. nual que en los tratamientos de autopolinización, geitonogamia y polinización libre (H = 13,83 para fructificación, F = 14,6 para contenido de semillas, P < 0,05). Al excluir los insectos en el tratamiento de autopolinización, sólo el 1 % de las flores en las inflorescencias produjeron frutos. En el tratamiento de polinización libre, el 2 % de las flores fructificaron. La fructifi46 cación aumentó cinco veces bajo polinización cruzada con relación a la polinización con polen del mismo individuo (geitonogamia), de tal modo que un 20 % de las flores en la inflorescencia se desarrollan como frutos (Tabla 1). El contenido de semillas por fruto alcanza los valores más bajos cuando la polinización se realiza exclusivamente con polen Biología reproductiva e importancia de la polinización en Cedrela lilloi Tabla 1. Producción de frutos y semillas bajo diferentes tratamientos de polinización en Cedrela lilloi. Entre paréntesis se indica el número de inflorescencias. Auto polinización Geitonogamia Polinización cruzada Polinización libre Frutos - flores en la inflorescencia 0,013 (72) 0,04 (4) 0,204 (12) 0,022 (86) Semillas por fruto 22,74 ± 9,81 13 ± 9,54 39,73 ± 11,31 24,29 ± 9,64 del mismo individuo (geitonogamia). Si bien C. lilloi produjo frutos bajo geitonogamia, el número de semillas por fruto fue 67 % menor con respecto al obtenido bajo polinización cruzada. Al excluir los insectos, el contenido de semillas por fruto no difiere del alcanzado bajo polinización libre. Esto sugiere que los polinizadores que visitaron las flores fueron escasos o poco eficientes en su servicio de polinización. En observaciones puntuales sobre las inflorescencias se pudieron identificar Coleópteros, Himenópteros, Lepidópteros, Dípteros y Tisanópteros. En total 25 especies de insectos visitaron las flores de C. lilloi, siendo las mariposas, polillas, abejas, abejorros y trips, los visitantes más comunes. DISCUSION La expresión del sexo en Cedrela lilloi a nivel individuo presenta separación espacial y temporal, existiendo una fase femenina intermedia a dos fases masculinas. La producción de frutos y semillas es dependiente de la acción de los polinizadores, especialmente de los insectos que tie- nen un patrón de forrajeo que promueve el cruzamiento entre individuos. El ciclo de floración donde la fase femenina se interpone entre dos fases masculinas se llama duodicogamia (Lloyd y Webb 1986) y ha sido registrado en algunas especies de Sapindaceae, Aceraceae, Hippocastraceae, Fagaceae, Annonaceae, Lauraceae y Rubiaceae (Stout 1928, de Jong 1976, Jaynes 1975, Rogstad 1994, Kubitzki y Kurz 1984, McDade 1986), pero nunca fue reportado para la familia Meliacea. Dentro de esta familia, C. lilloi podría ser la única con esta disposición y dinámica temporal de las flores. La especie Cedrela odorata presenta protoginia, es decir, la antesis de las flores femeninas ocurre primero que la de las flores masculinas (Cintron 1990). No existen estudios específicos acerca de la disposición y dinámica temporal de las flores de Cedrela balansae. La baja fructificación observada al excluir a los polinizadores y el efecto observado al agregar polen de otros individuos en el estigma, sugiere una importante influencia del comportamiento y abundancia de 47 Aschero, V. los polinizadores en la producción de frutos y semillas. Considerando que tanto la abundancia como el tipo de polen depositado en el estigma afectan la producción de frutos y semillas, una alta abundancia de polinizadores con comportamiento de forrajeo que promueva el cruzamiento entre individuos sería importante para alcanzar altos niveles reproductivos en Cedrela lilloi. BIBLIOGRAFIA Bierzychudek, P. 1981. Pollination limitation on plant reproductive effort. American Naturalist 117: 838-840. Calvo, R.N. 1993. Evolutionary demography of orchids: intensity and frecuency of pollination and the cost of fruiting. Ecology 74: 1033-1042. Cintron, B.B. 1990. Cedrela odorata L. Cedro hembra, spanish cedar. En: Burns, R.M. y B.H. Honkala (Eds.). Silvics of North America: 2. Hardwoods. Agric. Handb. 654. Washington, DC: U.S. Departament of Agriculture, Forest Service. pp. 250257. de Jong, P.C. 1976. Flowering and sex expression in Acer L A biosystematic study. Mededelingen Landbouwhogeschool Wageningen 76-2: 1-201. Digilio, A.P. y P.R. Legname. 1966. Los árboles indígenas de la Provincia de Tucumán. Opera Lilloana 15. Tucumán, Argentina. Galen, C. 1985. Regulation on seed set in Polemonium viscosum: floral scents, pollination and resources. Ecology 66: 792-797. Jaines, R.A. 1975. Chestnuts. En: Janick J. y J.N. Moore (Eds.). Advances in fruit breeding. West Lafayette, Purdue University Press. pp. 490-504. Johnston, M.O. 1991. Pollen limitation of female reproduction in Lobelia cardinalis and L. siphilitica. Ecology 72: 1500-1503. Kearns, C.A. y D.W. Inouye. 1993. Techniques for pollination biologists. University Press of Colorado. 583 pp. Koptur, S. 1984. Outcrossing and pollinator limitation of fruit set: breeding systems of neotropical Inga trees (Fabaceae: Mimosoideae). Evolution 38 (5): 1130-1143. Kubitzki, K. y D.H. Kurz. 1984. Synchronized dichogamy and dioecy in Neotropical Lauraceae. Plant Systematics and Evolution 147: 253-266. 48 Biología reproductiva e importancia de la polinización en Cedrela lilloi Lloyd, D.G. y C.J Webb. 1986. The avoidence of interference between the presentation of pollen and stigmas in angiosperms: Dicogamy. New Zealand Journal of Botany 22: 135-162. Mc Dade, L.A. 1986 Protandry, synchronized flowering and sequential phenotypic unisexuality in Neotropical Pentagonia macrophylla (Rubiaceae). Oecologia 68: 218-223. Mayhew, J.E. y A.C. Newton. 1998. The silviculture of Mahogany. CABI publishing, UK. 226 pp. Parker, I.M. 1997. Pollinator limitation of Cytisus scoparius (Scotch broom), an invasive exotic shrub. Ecology 78 (5): 1457-1470. Pennington, T.D. y B.T. Styles. 1975. A generic monograph of the Meliaceae. Blumea 22: 419-540. Proctor, M.; P. Yeo y A. Lack. 1996. The natural history of pollination. Timber Press, USA. 479 pp. Richards, A.J. 1997. Plant breeding systems. Chapman & Hall (Second edition). 529pp. Rogstad, S.H. 1994 The biosystematics and evolution of the Polyalthia hypoleuca species complex (Annonaceae) of Malesia. III. Floral ontogeny and breeding systems. American Journal of Botany. 81:145-154. Smith, C.E. 1960. A revision of Cedrela (Meliaceae). Fieldiana: Botany 29:294-341. Stout, A.B. 1928. Dichogamy in flowering plants. Bulletin of the Torrey Botanical Club 55:141-153. 49 Ecología y producción de cedro (género Cedrela) en las Yungas australes. Pacheco, S. y A. Brown (Eds.). LIEY-ProYungas. Argentina. pp. 51-58. 2006. Características y usos de las maderas de Cedrela de las Yungas de Argentina Rivera, S. M. Cátedra de Dendrología, Facultad. de Ciencias Agrarias y Forestales, Universidad Nacional de La Plata, Diag. 113 y 61 (1900), La Plata, Buenos Aires, Argentina. [email protected] En este trabajo se presenta un análisis a nivel genérico de las características anatómicas macro y microscópica de la madera de Cedrela. Se presentan los resultados de observaciones a simple vista y con microscopio, en secciones transversales y longitudinales. Se realiza una descripción de las propiedades tecnológicas y de uso y se presentan las características diagnósticas que permitirían diferenciar dos de las especies del noroeste de Argentina (C. balansae y C. lilloi). This work presents a generic analysis of macro and micro anatomical characteristics of Cedrela wood. It shows the results of nude eye and microscopic observations, both in transversal and longitudinal sections. It also describes technological and use properties, and diagnostic characteristics to differentiate C. balansae and C. lilloi, two species present in northwestern Argentina. Rivera, S. M. introduccion Los estudios que se han realizado hasta el momento sobre anatomía de la madera de Cedrela no han sido claros en la caracterización de las especies del género. De acuerdo a Record y Hess (1943) las muestras de madera de diferentes especies de Cedrela exhiben las mismas cualidades en los ensayos y tienen una estructura anatómica similar. Estos autores afirman que las variaciones en densidad y color probablemente son debidas a la influencia de los factores ambientales durante el crecimiento de los árboles. Browne en 1756, trabaja con el género Cedrela en conjunto y describe a la madera como de olor agradable, aromática, liviana, porosa y fácil de trabajar. Las referencias de trabajos actuales con estas maderas agrupan a las diferentes especies bajo el concepto de género, dada la dificultad para su caracterización a nivel específico (Chichignoud 1990, Viscarra y Lara 1992). Espósito y Urretavizcaya (1992) intentaron asociar los nombres vulgares tales como: cedro rosado, cedro salteño, cedro orán, cedro peludo, cedro coya, con las características xilológicas extraídas de citas bibliográficas y datos obtenidos de clasificaciones empíricas a campo realizadas por madereros locales; pero no pudieron establecer la correcta identidad específica de las muestras. El objetivo de este trabajo es describir las características anatómicas, macro y microscópicas, de las maderas del género Cedrela en el noroeste de Argentina. Se agregan además características de color, textura, diseño y algunas propiedades tecnológicas extraídas de la bibliografía. 52 Se trabajó con muestras de individuos de Cedrela balansae cercanos a la ciudad de Orán y con muestras de Cedrela lilloi de la localidad de Los Toldos, Salta. Las muestras fueron suministradas por el Dr. Ricardo Villalba. CARACTERÍSTICAS XILOLOGICAS DE Cedrela EN LAS YUNGAS ARGENTINAS Observables a simple vista Sección transversal. Se observa una amplia gama de tonalidades; el duramen varía entre rosados y castaños claros y la albura varía entre amarillos y rosados. Con el paso del tiempo la madera se oscurece por oxidación en su exposición al aire y la transición albura-duramen es más suave. Los anillos de crecimiento son demarcados. La distribución de los vasos o poros varía de difusa a semi-circular. El leño de primavera presenta vasos de mayor tamaño formando una hilera en correspondencia con la banda de tejido parenquimático. Estos vasos generalmente son solitarios, escasos y de forma circular. El resto del anillo de crecimiento presenta vasos o poros de menor tamaño, solitarios y múltiples de 2 a 3 hileras radiales. Algunas muestras presentan porosidad difusa, es decir los poros o vasos tienen una distribución homogénea en el período de crecimiento. Se ha observado también porosidad semi-circular a circular definida por 2 o 3 hileras de poros más grandes presentes en el leño de primavera que alternan con otros más pequeños, siempre rodeados por el tejido parenquimático. En cualquiera de estos casos, el anillo se define por bandas delgadas de tejido parenquimático. Este Características y usos de las maderas de Cedrela de las Yungas de Argentina también se presenta vasicéntrico, es decir rodeando al vaso. óptico. Las fotografías fueron tomadas con microscopio electrónico de barrido. Sección longitudinal. Se puede observar el diseño o las figuras que presentan los diferentes elementos anatómicos y las sustancias xilócromas. Las pequeñas marcas verticales presentes producen el “rayado” dado por los vasos; el diseño “veteado” está dado por las marcas que dejan los anillos de crecimiento y el “floreado” también por los anillos, pero en corte longitudinal tangencial. En las secciones longitudinales radiales donde los radios son cortados en su ancho máximo, producen un suave “jaspeado”. Las maderas de este género presentan una textura que es media y heterogénea o gruesa y heterogénea dependiendo de la relación entre el diámetro de los vasos, las fibras y la abundancia del tejido parenquimático. La densidad varía entre 0,4 y 0,7 g/cm³ y el “grano” o sea la alineación de los elementos verticales, puede presentarse recto u oblicuo. El conjunto de caracteres macroscópicos, la densidad y el aroma valoran notablemente sus cualidades estéticas. Las fibras de paredes muy delgadas producen un acabado liso que contrasta con el tejido vascular grueso otorgando un brillo dorado, acentuado en algunas secciones por el jaspeado de los radios. Sección transversal. Presenta anillos de crecimiento demarcados por el achatamiento radial de las fibras. La porosidad es semicircular a difusa. Los vasos son solitarios y múltiples, cortos, de contorno circular y angular y se presentan de 5 - 20 por mm². Se observa presencia de contenidos. Las paredes de las fibras son muy delgadas y de lumen amplio. El parénquima es paratraqueal vasicéntrico y en bandas marginales o terminales y axial difuso (Fotos 1 y 2). Observables con microscopio Para realizar estas descripciones se utilizaron secciones transversales, longitudinales tangenciales y radiales de 30 micrones de espesor. Se colorearon con safranina, se deshidrataron y se montaron en portaobjetos para ser observadas con microscopio Sección longitudinal tangencial. Presenta fibras septadas, radios uni, bi, tri y tetraseriados, puntuaciones intervasculares alternas de contorno poligonal y vasos con contenidos. Se observan cristales en las células marginales de los radios y en las células radiales. Las series parenquimáticas son de 3 a 8 células (Fotos 3 y 4). Sección longitudinal radial. Se observan elementos de vaso con placa de perforación simple, los radios homo y heterogéneos con presencia de células cuadradas y/o rectangulares y algunas con cristales (Foto 5). Propiedades tecnologicas y usos La madera del género Cedrela está recomendada para chapas decorativas, carpintería de exterior e interior, ebanistería e instrumentos musicales. Son apreciadas tanto a nivel local como en el mercado maderero mundial. Estas maderas reúnen una buena combinación de propiedades, tales como rápido secado y buena estabilidad. 53 Rivera, S. M. Foto 1. Corte transversal de cedro coya (Cedrela lilloi). Foto 2. Corte transversal de cedro orán (Cedrela balansae). 54 Características y usos de las maderas de Cedrela de las Yungas de Argentina Foto 3. Corte longitudinal tangencial de cedro coya (Cedrela lilloi). Foto 4. Corte longitudinal tangencial de cedro orán (Cedrela balansae). 55 Rivera, S. M. Foto 5. Corte longitudinal radial de cedro coya (Cedrela lilloi). Tortorelli (1956) describe a Cedrela lilloi como aquella madera que no presenta problemas en el maquinado. Sin embargo, en esta especie es común observar en el cepillado la aparición de pelusa, lo que podría atribuirse a la “madera de reacción”y le ha valido el nombre vulgar de “cedro peludo”. Esto podría deberse al comportamiento de la especie, que crece en diferentes condiciones ambientales, aunque también podría haber factores genéticos en juego. Con respecto a Cedrela balansae, si bien no presenta este defecto, su madera es considerada ligeramente inferior a C. lilloi y C. fissilis (cedro misionero). Esta última es la madera considerada de referencia en cuanto a calidad para el género en nuestro país. Tinto (1978) establece para C. balansae y C. fissilis las ventajas y limitaciones de su uso en la construcción de viviendas. Chichignoud (1990) describe 56 las características tecnológicas del género y con respecto a la densidad, afirma que su variabilidad radica en la procedencia de las maderas. En cuanto a la durabilidad, la establece entre media y buena y no describe métodos de impregnación. sintesis de caracteristicas diagnosticas Cedrela balansae es de madera liviana, su densidad es de aproximadamente 0,4 6 g/m3 y se clasifica como blanda. Se utiliza en trabajos de mueblería y carpintería, específicamente para molduras y persianas y marcos de puertas y ventanas (Tabla 1). Cedrela lilloi es de madera liviana, su densidad es de aproximadamente 0,430 g/cm³ presentando frecuentemente una abundante pubescencia. Esta característica determina su menor aptitud para el cepillado y lustrado (Tabla 1). Características y usos de las maderas de Cedrela de las Yungas de Argentina Tabla 1. Comparación de caracteres anatómicos entre cedro orán (Cedrela balansae) y cedro coya (Cedrela lilloi). Especie Porosidad Tamaño de poros Tipos de radios Altura Cedrela balansae Circular a semicircular 236 micrones Bi - triseriado heterocelular 192 micrones Cedrela lilloi Semicircular a difusa 172 micrones Mayormente biseriado heterocelular 247 micrones . BibliografIa Browne, P. 1756. The civil and Natural History of Jamaica. London (Osborn and Shipton). Chichignoud, G. 1990. Atlas de maderas tropicales de América latina. Association technique internationale des bois tropicane. Francia. 218 pp. Esposito, P. y M. F. Urretavizacarra. 1992. Dificultades para la identificación de las maderas del género Cedrela. 1er Encuentro de Anatomistas de la Madera. La Plata, Argentina. Record, S. J. y R. Hess. 1943. Timbers of the New World. Yale University Press, New Haven, CT. Tinto, J. 1978. Aporte del sector forestal a la construcción de viviendas. Folleto Técnico forestal N° 44 (2° edición). IFONA. República Argentina. Tortorelli, L. 1956. Maderas y Bosques Argentinos. Editorial Acme, Buenos Aires. Viscarra, S. y R. Lara. 1992. Maderas de Bolivia: Características y usos de 55 maderas tropicales. Cámara Nacional Forestal, Santa Cruz de la Sierra, Bolivia. 295 pp. 57 Variabilidad interanual de los caracteres anatómicos en el leño de Cedrella lilloi en el NOA Ecología y producción de cedro (género Cedrela) en las Yungas australes. Pacheco, S. y A. Brown (Eds.). LIEY-ProYungas. Argentina. pp. 59-82. 2006. Variabilidad interanual de los caracteres anatómicos en el leño de Cedrela lilloi en el noroeste de Argentina Villalba, R.; S. Delgado; M. De Membiela y D. Mendoza. Departamento de Dendrocronología e Historia Ambiental Instituto Argentino de Nivología, Glaciología y Ciencias Ambientales (IANIGLA); CC 330 (5500), Mendoza, Argentina. [email protected] El ancho de los anillos de crecimiento es el parámetro dendrocronológico que tradicionalmente se emplea para inferir cambios en la historia de vida de los árboles y la relación con su entorno. Las distintas etapas de crecimiento y los factores ambientales, actúan modificando el número, tamaño y disposición de los elementos constitutivos del xilema. Por lo tanto, existe un gran potencial para derivar información silvícola y ambiental a partir de otros caracteres anatómicos de la madera. En este estudio presentamos el desarrollo de cuatro cronologías basadas en el ancho de los anillos, el área total de vasos de xilema, el número de vasos y el tamaño del vaso medio, derivadas de 10 ejemplares de Cedrela lilloi creciendo en las proximidades de Los Toldos (22º 60´S, 64 º61 ´W), Salta, Argentina. Las muestras provienen de un rodal coetáneo de esta especie y cubren el intervalo 1796-1999 (204 años). Las series de ancho de anillos se caracterizan por una reducción marcada en el espesor de los anillos con la edad. Una tendencia similar se observa en las series derivadas a partir del número de vasos por anillo de crecimiento. Por el contrario, las series de área total de vasos por anillos de crecimiento no muestran una tendencia marcada a la disminución, en tanto que el tamaño medio del vaso aumenta con la edad. La proporción del área total de vasos al área total del anillo, se incrementó durante los primeros 150 años de vida de los ejemplares estudiados, estabilizándose en los últimos 60 años. Estos cambios en el porcentaje de tejido conductor de agua en el tiempo, están asociados fundamentalmente al aumento y posterior estabilización en el tamaño de los vasos con la edad. Una combinación del tamaño medio del vaso, el número total de vasos por anillo y el porcentaje de tejido de conducción permitió identificar tres patrones bien definidos en el arreglo leñoso correspondientes al leño juvenil, adulto y maduro. Estos cambios en el leño reflejan distintas etapas en el crecimiento de los árboles estudiados. Así por ejemplo, el cambio de leño adulto a maduro coincide temporalmente con el turno biológico de corta (aproximadamente 150 años), hecho que permite inferir los cambios en las distintas etapas del crecimiento de los individuos de Cedrela lilloi a partir del arreglo de los elementos del plan leñoso. The ring width is the dendrochronological feature traditionally used to infer changes in tree live history and its environment. In addition to ring width, the number, size and arrangement of xylem elements are influenced by different stages in tree development and environmental changes. In consequence, the history on tree dynamics and the environment provided by tree rings can be complemented with 59 Villalba, R.; S. Delgado; M. De Membiela y D. Mendoza additional information inferred from changes in the spatial array of anatomical woody features over time. In this study we report the development of four chronologies based on ring width, total vessel area per ring, number of vessel per ring, and mean size of vessels per annual ring. Samples were collected from 10 individuals in an even-age stand of Cedrela lilloi near to Los Toldos (22º60´S, 64º61´W), Salta, Argentina. Dendrochronological series cover the interval 1796-1999 (204 years). Tree-ring series from Cedela lilloi show a conspicuous reduction in width with increasing age. A similar trend was recorded for changes in the number of vessel over time. In contrast, the total vessel area per ring shows almost none trend, whereas the mean size of the vessels increased over time. The ratio between annual vessel and ring areas shows a persistent increased during the early 150 years, follows by an interval of stabilization in the past 60 years. The changes in the proportions between water and no-water transport tissues in the wood are related to the sustained increase and later stabilization in vessel sizes over time. Three wood stages (juvenile, adult, mature) were identified from the examination of the temporal patterns of mean vessel size, number of vessel and percentage of water-transport elements. These patterns appear to be related to different stages in tree development. For example, the transition from adult to mature wood is coincident with the biological rotation age (150 years), suggesting the possibility of inferring changes in tree development of Cedrela lilloi from the examination of the arrangement of woody elements. Introduccion El empleo de los anillos de crecimiento de las plantas leñosas permite cuantificar en forma precisa las velocidades de crecimiento radial de los árboles en distintos ambientes, así como determinar diferencias en las estructuras poblacionales del bosque en relación a factores tales como el clima, el suelo, los disturbios naturales y la historia de uso. Todo estudio que requiera información sobre la estructura de edad y la dinámica de los bosques, la determinación de sus turnos de aprovechamiento, el establecimiento de las relaciones entre crecimiento leñoso, fructificación y/o variaciones climáticas o simplemente la reacción del bosque a un tratamiento silví60 cola determinado; necesita del fechado y la evaluación de los ritmos pasados y presentes del crecimiento. A esta lista se suma la posibilidad de desarrollar a partir de los anillos de crecimiento, reconstrucciones dendroclimáticas en áreas donde los bosques podrían ser la única fuente de información paleoclimática con una importante extensión temporal y resolución anual. Empleando las variaciones interanuales en el ancho de los anillos ha sido posible reconstruir para los últimos 3 o 4 siglos las fluctuaciones anuales o estacionales de la temperatura, la precipitación y otras variables de la circulación atmosférica en numerosas regiones del mundo (Villalba 2000). Variabilidad interanual de los caracteres anatómicos en el leño de Cedrella lilloi en el NOA El ancho de los anillos de crecimiento es el rasgo anatómico que tradicionalmente se usa en dendrocronología para estudios de dinámica forestal, para datar disturbios y procesos geodinámicos, o para reconstruir las variaciones climáticas pasadas (Fritts 1976, Schweingruber 1996). Su medición es relativamente sencilla y no destructiva. Debido a que los anillos tienen resolución anual (normalmente una banda de crecimiento es formada por año), es posible cuantificar muy precisamente la velocidad de crecimiento de un árbol, determinar la edad de una geoforma, datar la ocurrencia de disturbios (e.g. fuego, inundación), o establecer precisamente las relaciones entre el crecimiento y las variaciones anuales o estacionales del clima (Fritts 1976). Es importante hacer notar que las bandas anuales de crecimiento pueden proveer información adicional a aquella que se desprende de la simple medición del ancho de los anillos. Los factores ambientales actúan como fuerzas modeladoras del número y tamaño de los elementos constitutivos del xilema (Carlquist 1988). En consecuencia, existe un gran potencial para derivar información climática a partir de otros caracteres anatómicos de la madera más allá que el ancho de los anillos. Desde mediados de la década de 1970 fue posible determinar las variaciones interanuales en la densidad de la madera a través del empleo de rayos-X. Aún cuando los estudios densitométricos tienen en su mayoría aplicación tecnológica, se observó que las variaciones anuales de densidad, en particular la densidad del leño tardío, contenían también una señal climá- tica complementaria a la del ancho de los anillos (Schweingruber 1988, Briffa et al. 1992). De la misma forma, el desarrollo de sistemas de análisis de imágenes digitales desde comienzos de la década de 1990 ha permitido medir, en forma rutinaria, otros rasgos anatómicos de la madera. Estos registros nuevos pueden contener información xilológica, tecnológica o ambiental diferente o complementaria de la registrada en el ancho o la densidad de los anillos de crecimiento. En este trabajo se describe el desarrollo de cronologías en base a caracteres anatómicos de la madera de Cedrela lilloi C.DC. (cedro coya) en el noroeste de Argentina (NOA). Este estudio representa el primer desarrollo de cronologías en base al área total de vasos, el número de vasos y el tamaño del vaso medio por anillo de crecimiento para una especie en las regiones subtropicales de América del Sur. Estas cronologías son comparadas con la derivada a partir del ancho de los anillos procedentes de los mismos individuos. Nuestra hipótesis es que las variaciones climáticas producen importantes cambios en el crecimiento de los árboles. Estas variaciones se ven reflejadas en el patrón de sus anillos de crecimiento y en otras características anatómicas del leño tales como la densidad, el tamaño y distribución de los vasos. La lectura cronológica precisa de estas variables en el leño de los árboles proveerá información adicional al ancho de los anillos. Entre otras aplicaciones, puede ser usada para mejorar nuestro conocimiento de las variaciones temporales en las propiedades físico-mecánicas de la madera de 61 Villalba, R.; S. Delgado; M. De Membiela y D. Mendoza Cedrela lilloi, así como de las variaciones ambientales del pasado en el NOA. El hecho de contar con información sobre las variaciones interanuales de diversos componentes de la madera, nos permitirá valorar sus tendencias a lo largo del tiempo. Así por ejemplo, la información proveniente de nuestro análisis nos permitirá responder a preguntas tales como: 1) ¿Es estable en el tiempo la proporción en los volúmenes ocupadas por los vasos y los demás tejidos constituyentes del leño en Cedrela lilloi? Es de suponer que si las proporciones entre los distintos elementos constitutivos del leño cambian con el tiempo, también lo hagan las propiedades físico-mecánicas de la madera. 2) ¿Requieren los árboles de esta especie un número fijo de vasos para formar una cantidad determinada de madera, o la cantidad y el tamaño de vasos varía con el tiempo? Estos temas están íntimamente ligados a las distintas etapas de crecimiento de los árboles (juvenil, adulto, maduro) y la eficiencia de la especie en el uso del agua. 3) ¿Cambia la eficiencia en el uso del agua con el tiempo, o siempre se requiere transportar la misma cantidad de agua para formar una determinada cantidad de madera? ¿Cómo cambian estas relaciones entre el leño juvenil y el leño adulto en Cedrela lilloi? Por ello, la información presentada en este estudio resulta valiosa no sólo en el campo de la dendrocronología, sino también en estudios xilológicos, ecofisiológicos, silviculturales y de aplicación tecnológica de la madera. 62 ANTECEDENTES DE ESTUDIOS DENDROCRONOLOGICOS MEDIANTE ANALISIS DE IMAGENES Uno de los objetivos principales de la Dendrocronología durante los últimos años ha sido la búsqueda de variables anatómicas adicionales al ancho de los anillos que permitan fortalecer la aplicación de los métodos dendrocronológicos en la resolución de numerosos problemas ambientales. Las investigaciones en coníferas se han concentrado principalmente en los estudios de las variaciones intra-anuales de la densidad de la madera, usando radio-densitometría (Schweingruber 1988, Schweingruber y Briffa 1996). Recientemente, estudios preliminares han demostrado la potencialidad dendrocronológica resultante del análisis del tamaño de las fibrotraqueidas o de sus paredes celulares (Munro et. al. 1996). Los trabajos en angiospermas son menos numerosos. La división entre leño temprano y leño tardío en las angiospermas presenta algunas dificultades metodológicas (Blank 1997), y en contados casos se han empleado los rasgos anatómicos del leño en estudios dendrocronológicos. Sin embargo, la mayoría de los trabajos realizados coinciden en señalar la potencialidad de los caracteres anatómicos como proveedores de información adicional a la aportada por los anillos de los árboles. Es interesante destacar que gran parte de estos estudios se han realizado en especies con porosidad circular, donde los poros del leño temprano son más grandes que los del leño tardío tal como ocurre en Cedrela lilloi. Por ejemplo, Woodcock (1989) ob- Variabilidad interanual de los caracteres anatómicos en el leño de Cedrella lilloi en el NOA servó que las variaciones en el diámetro de los vasos del leño tardío de Quercus macrocarpa, Michx., especie con marcada porosidad circular que crece en los bosques nativos de Nebraska, están directamente relacionadas con las fluctuaciones anuales de la precipitación. Según los análisis de esta autora, el diámetro del vaso del leño tardío en Quercus macrocarpa es un indicador más preciso de los cambios en la precipitación anual que el ancho de sus anillos. En especies de mismo género, el diámetro y longitud de los vasos, así como su conductividad, han sido usados para evaluar las adaptaciones ambientales de los robles a lo largo de gradientes climáticos en el noreste de España. Villar-Salvador et al. (1997) en un estudio sobre tres especies, Quercus coccifera, Q. ilex y Q. faginea, observaron que el diámetro medio máximo del vaso de Q. coccifera y la Q. ilex aumenta con el incremento de precipitación, mientras que la densidad de los vasos disminuye. En la misma región, García Gonzalez y Eckstein (2003) encontraron que los cambios en el área de vasos del leño temprano de Quercus robur, L. responden a las variaciones de la precipitación durante el periodo febrero-abril (fines de invierno-comienzos de primavera), reflejando la importancia del agua disponible en el momento de formación de los vasos conductores tempranos. Trabajando con especies de porosidad difusa, donde los poros son de tamaño uniforme y se encuentran uniformemente distribuidos en todo el anillo, Sass y Eckstein (1992) notaron que el área media anual de vasos en Fagus sylvatica, L. se relaciona en forma positiva con la precipitación anual (medida desde julio del año anterior hasta agosto del año corriente). La temperatura tiene un efecto menor sobre el crecimiento y generalmente negativo debido probablemente a que la temperatura y la precipitación están negativamente correlacionadas durante el verano boreal (marzo-octubre). En América del Sur, sólo podemos mencionar algunos trabajos preliminares que han intentado establecer relaciones entre caracteres anatómicos del leño y parámetros ambientales. Villalba et al. (2002), analizaron las variaciones interanuales en el leño de ejemplares de Nothofagus pumilio (especie de porosidad difusa) provenientes del límite superior del bosque en el norte de la Patagonia. Estos autores observan que la información provista por características anatómicas tales como el área total o el número de vasos por anillo no difiere significativamente de la que surge del análisis del ancho de los anillos de crecimiento. El tamaño del vaso medio por el contrario, no se relaciona con el ancho de los anillos y su variabilidad pareciera estar más fuertemente controlada por las variaciones de la temperatura anual que el ancho de los anillos. Metodos Colección de las muestras Las muestras de Cedrela lilloi, empleadas en este estudio proceden de El Arranque, al este de la localidad Los Toldos (22º 60´ S, 64º 61´ W), Provincia de Salta, Argentina. 63 Villalba, R.; S. Delgado; M. De Membiela y D. Mendoza El muestreo se realizó a una altura media de 1.460 m s.n.m. Las muestras utilizadas para análisis de imágenes fueron colectadas durante una tala selectiva de individuos de Cedrela lilloi realizada por gente del lugar. Se muestreó un rodal coetáneo donde los individuos tenían un diámetro medio de 1,20 m y más de 20 m de altura. Aprovechando la corta de los árboles, pudimos obtener muestras desde el centro del fuste hasta la corteza con un ancho variable entre 7 y 10 cm. El hecho de contar con muestras de ese tamaño facilitó la toma posterior de las imágenes de video. Es muy difícil captar imágenes de buena calidad para el análisis de los elementos anatómicos de la madera a partir de muestras de barrenos de incrementos de 5 mm de diámetro, particularmente cuando las muestras están dañadas por ataques de hongos o presentan anillos irregulares. Con el fin de obtener imágenes de video de alta calidad, las muestras fueron pulidas mecánicamente usando lijas cuyas granulometrías varió entre 220 y 500. Posteriormente, las superficies fueron pulidas a mano con lijas de grano entre 800 y 1.200. En todos los casos, la superficie de pulido fue perpendicular a la dirección del grano en la madera. Para facilitar la visualización de los vasos en la madera, el lúmen de los vasos se rellenó con carbonato de calcio, sustancia de color blanco, incrementando de ésta forma el contraste entre los elementos conductores de la madera y los tejidos restantes. 64 Captura de imágenes y procesamiento de los datos La filmación se realizó con una cámara de video digital conectada a una lupa binocular y a una computadora personal. Las imágenes se capturaron, en la medida de lo posible, en el mismo sector de la muestra donde se realizó la medición del ancho de los anillos. Se generó una imagen por anillo a partir de un promedio de veinte tomas para reducir el ruido introducido por el sistema digital de captura. Las imágenes digitales fueron procesadas con el programa NIH Image 1.62 (Software de dominio público) del National Institute of Health (NIH). Este programa se desarrolló en el departamento de investigación (RSB) del NIH de los Estados Unidos (http://rsb.info.nih.gov/ij/download.html). Con el fin de reducir el tiempo de procesamiento de la imagen, se prepararon macros en lenguaje Pascal que automatizaron parcialmente el proceso de medición de los elementos leñosos. El primer macro incrementa la definición a través de un proceso de perfilado de la imagen original (Fig. 1). El operador delimita manualmente los límites izquierdo y derecho del sector de anillo de crecimiento. Los límites superior e inferior están dados por el tamaño de la imagen que se conserva en cada toma. Queda así enmascarada la imagen del sector del anillo donde se encuentran los elementos a medir y se calcula la superficie del anillo en cada toma. Se genera un archivo con las coordenadas del sector en estudio respecto de la imagen total, lo que permite posterior- Variabilidad interanual de los caracteres anatómicos en el leño de Cedrella lilloi en el NOA A B A C D Figura 1. Cuatro etapas en el procesamiento de una imagen de video de Cedrela lilloi desti- nado a la cuantificación del área individual de cada vaso, sus diámetros mayores y menores, y la posición x-y de cada vaso en la imagen: A - imagen original, B - separación de los vasos del resto del tejido y marcación del anillo de crecimiento, C - selección final, y D - numeración y medición de los vasos individuales. mente ubicar cada elemento en su posición relativa dentro del anillo. Posteriormente, la imagen en escala de grises se transforma en imagen binaria (blanco y negro) lo que permite realizar las mediciones de los elementos leñosos de interés. El macro habilita al operador para manejar el valor de gris que define el límite entre blanco y negro, de modo que resalte el elemento a estudiar, en nuestro caso el lúmen de los vasos. Frecuentemente, los vasos múltiples no son bien discriminados durante el proceso de digitalización binaria. Sin embargo, el programa incluye una función (watershed), incluida en el segundo macro, que separa automáticamente objetos conectados entre si. Este proceso que ayuda en la separación de vasos no es perfecto y necesita a menudo la edición manual de la imagen. El siguiente macro superpone la imagen editada a la original. Por comparación, el operador puede rectificar todos los errores del proceso semi-automático de digitalización y lograr una clara delimitación de los elementos a estudiar (Fig. 1). Una vez delimitados los elementos de interés, se procede a la medición de los mismos. En nuestro caso las variables cuantificadas fueron: área individual de cada vaso, distancia en el eje horizontal del borde de la imagen al centro de vaso, distancia en el eje vertical del borde de la imagen al centro del vaso, perímetro del vaso, eje mayor del vaso, eje menor del vaso, ángulo del eje mayor con la horizontal. Todas estas mediciones se realizan en forma automática aplicando el último macro desarrollado. Este macro genera una ima65 Villalba, R.; S. Delgado; M. De Membiela y D. Mendoza gen de los vasos delimitados a través del proceso de digitalización anteriormente descrito y otra imagen de los vasos numerados, la que permite asociar cada grupo de mediciones a su respectivo vaso. A partir de estas medidas básicas se derivaron un número importante de variables, destacándose entre ellas: Area total de vaso por anillo de crecimiento, resultante de la sumatoria de todos los vasos presentes por anillo de crecimiento. Número de vasos por anillo, resultante del conteo de todos los vasos presentes en cada anillo de crecimiento. Porcentaje del área de vasos en relación al área total del anillo, resultante de la división del área total de vasos por el área total del anillo de crecimiento. A su vez, teniendo en cuenta otras variables tales como la localización dentro del anillo o las dimensiones de los vasos, las variables se pueden reagrupar en vasos del leño temprano versus tardío, o vasos grandes versus pequeños y evaluar la relación de cada una de estas variables con el clima. Para facilitar el cálculo de estas variables, se desarrollaron una serie de programas en Lenguaje Fortran en el Departamento de Dendrocronología e Historia Ambiental del IANIGLA. Los macros del programa NIH y la serie de programas Fortran descriptos están a disposición de quien los necesite comunicándose con los autores. Desarrollo de las cronologías Previo a la toma de las imágenes de video, cada anillo de crecimiento fue fechado empleando técnicas dendrocronológicas 66 estándares (Stokes y Smiley 1968). Esta etapa inicial es fundamental para establecer con certeza la fecha de cada anillo analizado. Primero, las muestras fueron fechadas en forma visual asignando a cada anillo de crecimiento su año calendario, teniendo en cuenta los patrones comunes de anillos anchos y estrechos. Esta etapa se conoce como co-fechado visual (Holmes 1983). Una vez fechadas las muestras se procedió a medir el ancho de los anillos con un medidor de incrementos Bannister con una precisión de 0,01 mm. Las series de anchos de anillos procedentes de cada muestra se plotearon y se compararon entre sí, con la ayuda del programa COFECHA (Holmes 1983, Grissino-Mayer y Fritts 1997). Este programa realiza una verificación estadística del fechado señalando las muestras o sectores de las mismas que pudiesen tener problemas de fechado. De esta manera se logra una alta precisión en el fechado del material leñoso. Una vez logrado el fechado exacto de cada anillo de crecimiento se procedió a la toma de las imágenes de video. Todas las cronologías fueron desarrolladas utilizando los métodos de estandarización comúnmente empleados en el desarrollo de cronologías a partir de anillos de árboles (Fritts 1976, Cook y Kairiukstis 1990). Para eliminar la tendencia biológica en el ancho de los anillos de crecimiento (que implica una reducción en el espesor de los anillos a medida que se incrementa el diámetro del árbol), se utilizaron diferentes curvas de estandarización. Las curvas de estandarización empleadas incluyeron cur- Variabilidad interanual de los caracteres anatómicos en el leño de Cedrella lilloi en el NOA vas de tipo exponencial negativa, de funciones spline con ventanas de 256 y 128 años, y de Crecimiento Regional (RCS, Fritts 1976, Cook y Kairiukstis 1990, Briffa et al. 1996). El método que mejor reprodujo las oscilaciones a largo plazo es la función spline de 256 años, por lo que este método fue también empleado para estandarizar las series de anillos relativas a los rasgos anatómicos de la madera. La validez del ajuste de la función spline se comprobó muestra por muestra mediante criterio visual utilizando el programa TURBO-ARSTAN para Macintosh. Las series estandarizadas se promediaron con una media doble-robusta (Mosteller y Tukey 1977) que minimiza el efecto de los valores extremos al generar una cronología media (Cook y Holmes 1984, Cook 1985). Para desarrollar las cronologías de los caracteres anatómicos del leño, se emplearon para cada una de las 10 muestras procesadas los valores anuales de área total de vasos por anillo, número de vasos por anillo y tamaño del vaso medio en cada anillo. El tamaño del vaso medio en cada año de cada muestra se obtuvo dividiendo el área total de vasos por el número de vasos presente en cada anillo. RESULTADOS Caracteres anatómicos Ancho de anillos En la Figura 2 se han representado las variaciones interanuales en el ancho de los anillos de crecimiento de las 10 muestras que se utilizaron para el análisis de ima- gen. Si bien el número de muestras no es elevado, permite establecer una comparación valida entre las propiedades del ancho de los anillos y de otros caracteres anatómicos del leño de Cedrela lilloi. Se observa en cada una de las muestras individuales una clara tendencia a disminuir el ancho de los anillos con el tiempo, en respuesta al aumento del diámetro de los árboles con la edad (tendencia biológica o de crecimiento). El crecimiento radial medio por año entre las muestras, medido a través del ancho de los anillos, oscila entre 1,6 y 3,2 mm a-1 sobre un período de 204 años (1796-1999). La serie promedio de anchos de anillos de las 10 muestras analizadas muestra una tendencia negativa de 0,014 mm a-1 (Fig. 2). Para eliminar las tendencias de crecimiento, las series de anchos de anillos fueron estandarizadas (valores observados divididos por los valores estimados) usando funciones splines con una rigidez de 256 años. En general, las tendencias de crecimiento se asemejan a funciones exponenciales negativas y más raramente a líneas rectas de tendencias negativas (Fig. 2). La cronología final, que resulta de promediar las series estandarizadas, representa las variaciones en el crecimiento radial de los árboles muestreados durante los últimos 200 años. En la cronología se observa que a finales de la década de 1870 se produjeron los crecimientos más bajos de los dos últimos siglos. Otros períodos de crecimiento reducido se registraron en las décadas de 1940, 1950 y comienzos de la década de 1960. Por el contrario los períodos de mayor crecimiento se produjeron a 67 Villalba, R.; S. Delgado; M. De Membiela y D. Mendoza 80 40 80 40 Ancho de anillos (mm x 10) 80 40 80 40 80 40 80 40 80 40 80 40 80 40 80 40 80 Indice de ancho de anillos 40 2.0 1.5 1.0 0.5 0.0 1800 1850 1900 1950 2000 Año Figura 2. Variaciones interanuales en el ancho de los anillos de crecimiento de 10 ejemplares de Cedrela lilloi, en las cercanías de la localidad de Los Toldos, Salta, Argentina, durante el intervalo 1796-1999. Cada una de las series de ancho de anillos se muestra a su vez con la curva que representa la tendencia biológica de crecimiento radial calculada a partir de los datos originales con funciones spline de 256 años (Cook 1985). En la parte media de la figura se indica la serie media (Promedio) obtenida promediando las 10 curvas individuales. En la parte inferior se representa la cronología de Anchos de Anillos que resulta de promediar todas las series individuales previa estandarización, o sea la división de las series individuales de anchos de anillos por las curvas de tendencia calculadas a partir de las funciones spline. 68 Variabilidad interanual de los caracteres anatómicos en el leño de Cedrella lilloi en el NOA mediados de la década de 1880, alrededor de 1930 y a partir de 1971, con excepción de un corto intervalo de bajo crecimiento centrado en la década de 1980 (Fig. 2). sos durante los estados juveniles del leño de Cedrela (ver comentarios en sección Tamaño medio de vaso). Area total de vasos A diferencia de lo que ocurre con el ancho de los anillos, el exámen detallado de las series individuales indica claramente que si bien algunas muestras presentan una tendencia a la disminución del área total de vasos con la edad (LMC01, LMC04), otras series no tienen una tendencia definida (LMC02, LMC07) o inclusive muestran tendencias positivas (LMC06, LMC09, Fig. 3). Otra característica importante es que la variabilidad interanual en el área total de vasos no se ve seriamente afectada por la edad, factor muy marcado en las series de anchos de anillos (Fig. 2). La serie promedio de las 10 muestras analizadas presenta un aumento muy marcado en el área total de vasos durante los primeros 20 años, valores máximos entre los 30 y los 90 años y una disminución gradual a partir de los 100 años. En consecuencia, no podemos hablar para la variable área total de vasos de una tendencia sostenida a la disminución con la edad como ocurre con el ancho de los anillos de crecimiento. Número de vasos Las series individuales que representan los cambios en el número de vasos del leño de Cedrela lilloi con la edad se incluyen en la Figura 4. Es claro que el número de vasos es mayor durante los estados juveniles de crecimiento. Transcurrido los primeros 50-70 años de crecimiento, se registra una reducción importante en el número de vasos en combinación con una disminución en la variabilidad interanual de los mismos. En forma consistente, todas las muestras analizadas presentan esta tendencia negativa en el número de vasos con la edad (Fig. 4). En este aspecto, el comportamiento del número de vasos con la edad, medido a través de las series promedios, es muy similar al del ancho de los anillos (r = 0,90, n = 204 años, p< 0,001), pero algo diferente con el área total de vasos (r = 0,55, n = 204 años, p< 0,001). La cronología que resulta a partir de la estandarización de las series de números de vasos es remarcablemente similar a la cronología de anchos de anillos (r = 0,90, n = 204 años, p< 0,001). A pesar de estas diferencias observadas, entre las series de ancho de anillos y de área total de vasos, las cronologías resultantes son muy similares (r = 0,83, n = 204 años, p< 0,001). La mayor diferencia entre las cronologías está dada por índices menores en el área total de vasos durante los primeros 20 años de crecimiento en respuesta al notable tamaño menor de los va- Tamaño medio del vaso En marcado contraste con otros caracteres anatómicos del leño de Cedrela lilloi, el tamaño medio de los vasos aumenta en forma sostenida con la edad (Fig. 5). En las muestras empleadas que alcanzan 200 años de edad, el aumento en el tamaño del vaso medio es muy pronunciado durante los estados juveniles del árbol, disminuye 69 Villalba, R.; S. Delgado; M. De Membiela y D. Mendoza 300 200 100 300 200 100 Area total de vasos (mm2 x 100) 300 200 100 300 200 100 300 200 100 300 200 100 300 200 100 300 200 100 300 200 100 300 200 100 200 Indice de área total de vasos 100 1.5 1.0 0.5 1800 1850 1900 1950 2000 Año Figura 3. Variaciones interanuales en el área total de vasos por anillo de crecimiento de 10 ejemplares de Cedrela lilloi, en las cercanías de la localidad de Los Toldos, Salta, Argentina, durante el intervalo 1796-1999. Cada una de las series de área total de vasos se muestra a su vez con la curva que representa la tendencia del área total de vasos en el tiempo calculada a partir de los datos originales con funciones spline de 256 años (Cook 1985). En la parte media de la figura se indica la serie media (Promedio) obtenida promediando las 10 curvas individuales. En la parte inferior se representa la cronología de Area Total de Vasos que resulta de promediar todas las series individuales previa estandarización, o sea la división de las series individuales de área total de vasos por las curvas de tendencia calculadas a partir de las funciones spline. 70 Variabilidad interanual de los caracteres anatómicos en el leño de Cedrella lilloi en el NOA 150 100 50 150 100 50 Número de vasos 150 100 50 150 100 50 150 100 50 150 100 50 150 100 50 150 100 50 150 100 50 150 100 50 150 100 Indice de número de vasos 50 1.5 1.0 0.5 1800 1850 1900 1950 2000 Año Figura 4. Variaciones interanuales en el número de vasos por anillo de crecimiento de 10 ejem- plares de Cedrela lilloi, en las cercanías de la localidad de Los Toldos, Salta, Argentina, durante el intervalo 1796-1999. Cada una de las series de número de vasos se muestra a su vez con la curva que representa la tendencia del número de vasos en el tiempo calculada a partir de los datos originales con funciones spline de 256 años (Cook, 1985). En la parte media de la figura se indica la serie media (Promedio) obtenida promediando las 10 curvas individuales. Finalmente, en la parte inferior se representa la cronología de Número de Vasos que resulta de promediar todas las series individuales previa estandarización, o sea la división de las series individuales de número de vasos por las curvas de tendencia calculadas a partir de las funciones spline. 71 Villalba, R.; S. Delgado; M. De Membiela y D. Mendoza 4000 Vaso medio (µm2) 2500 4000 5500 4000 2500 2500 5500 4000 2500 5000 3000 4000 1000 2500 7000 4000 5500 4000 2500 1000 5500 4000 2500 5500 4000 2500 Indice de vaso medio 6000 1500 3000 1500 1.2 1.0 0.8 0.6 1800 1850 1900 1950 2000 Año Figura 5. Variaciones interanuales en el tamaño medio del vaso por anillo de crecimiento de 10 ejemplares de Cedrela lilloi, en las cercanías de la localidad de Los Toldos, Salta, Argentina, durante el intervalo 1796-1999. Cada una de las series de tamaño medio del vaso se muestra a su vez con la curva que representa la tendencia del tamaño medio del vaso en el tiempo calculada a partir de los datos originales con funciones spline de 256 años (Cook 1985). En la parte media de la figura se indica la serie media (Promedio) obtenida promediando las 10 curvas individuales. En la parte inferior se representa la cronología de Vaso Medio que resulta de promediar todas las series individuales previa estandarización, o sea la división de las series individuales de tamaño medio del vaso por las curvas de tendencia calculadas a partir de las funciones spline. 72 Variabilidad interanual de los caracteres anatómicos en el leño de Cedrella lilloi en el NOA en intensidad en los próximos 100 años y es muy reducido o nulo durante los últimos 50 años. Asimismo, en contraposición con el ancho de los anillos y el número de vasos, la variabilidad interanual es muy reducida durante los primeros años de crecimiento y aumenta paulatinamente con la edad (Fig. 5). La serie promedio, que representa las variaciones interanuales en el tamaño medio del vaso de todas las muestras analizadas, refleja claramente la tendencia positiva registrada en las muestras individuales. Esta tendencia positiva se opone marcadamente a la del ancho de los anillos (r = -0,67, n = 204 años, p < 0,001) y a la del número total de vasos (r = -0,81, n = 204 años, p < 0,001) y en menor grado a la del área total de vasos por anillo (r = -0,10, n = 204 años, p < 0,001). La cronología que resulta de estandarizar las series individuales presenta los valores más bajos de tamaño de vasos durante los primeros 30 años de crecimiento, probablemente en respuesta a la presencia de leño juvenil, caracterizado por vasos más pequeños. Otros períodos de vasos relativamente pequeños se registraron en la década de 1930, comienzos de los 40 y en particular durante el año 1998 (Fig. 5). Variancia común en los caracteres anatómicos del leño En Dendrocronología se emplean una serie de parámetros estadísticos para medir la variabilidad común a todas las series que conforman una cronología. Cuanto mayor es esta señal común, las series son más similares entre sí, o sea que es menor el ruido presente en cada una de las series temporales que integran el registro. En la Tabla 1 se listan algunos estadísticos que nos permiten comparar la señal común presente en cada una de las cronologías desarrolladas a partir de los caracteres anatómicos del leño de Cedrela lilloi. La evolución temporal de dos de estas variables, el Rbar y el EPS, se muestran en la Figura 6. El Rbar es la media de los coeficientes de correlación que resultan de comparar todos los pares posibles de segmentos de un largo determinado entre todas las series que forman la cronología (Briffa 1995). El EPS (Expressed Population Signal), es una medida de la proporción de la señal total existente en una cronología infinitamente replicada que está presente en las series que integran la cronología considerada. En base a los estudios de Briffa (1995), cuando los valores de EPS están próximos o son mayores a un valor de 0,85, el número de muestras que integra la cronología es adecuado o está captando un porcentaje adecuado de esa señal teórica que resulta de una replicación infinita de las muestras. Valores menores de 0,85 estarían indicando que la replicación en ese sector de la cronología es baja y que debería aumentarse el número de muestras para fortalecer la señal. Los valores de correlación mínima y máxima entre las series que conforman las cronologías, evaluados en períodos de 50 años de extensión, desplazados 25 años entre períodos, indican que la cronología de anchos de anillos posee los valores más elevados (0,179 y 0,728) sugiriendo mayor similitud entre las series de anchos de anillos que entre los otros caracteres anatómicos 73 Villalba, R.; S. Delgado; M. De Membiela y D. Mendoza Tabla 1. Estadísticos de las cronologías de caracteres anatómicos del leño de Cedrela lilloi. Sensib. media: sensibilidad media, estadístico comúnmente usado en dendrocronología, que se refiere al porcentaje medio de cambio entre valores anuales consecutivos de la variable considerada (Fritts 1976). Rbar: representa la media de los coeficientes de correlación entre todos los pares posibles de series de diferentes árboles. Auto correl (1): Autocorrelación de primer orden. Los valores de correlación mínima (Correl. min.) y máxima (Correl. max.) están basados en comparaciones de 50 años entre todas las series que integran las cronologías. Desvío estand. Skew Ancho 0.356 0.496 3.268 Area Total 0.232 0.005 Número Vasos 0.260 Vaso Medio 0.109 0.8 Auto correl (1) Rhar 0.316 0.375 3.779 0.205 0.168 3.427 -0.202 3.237 Ancho de anillos Area total Kurtosis Sensib. media Mín. Max 0.414 0.179 0.728 0.433 0.315 0.054 0.625 0.217 0.451 0.325 0.049 0.591 0.104 0.292 0.210 0.041 0.405 Número total Tamaño del vaso Rbar 0.6 0.4 0.2 EPS 0.9 0.8 0.7 0.6 1840 1860 1880 1900 1920 1940 1960 1980 2000 Figura 6. Evolución temporal de los estadísticos que mi- den la calidad de las cronologías. Los valores de Rbar y EPS fueron calculados para segmentos de 50 años de extensión, desplazados cada 25 años. Los coeficientes de variación para el Rbar se indican para cada segmento. Para el EPS, la línea punteada representa el valor de referencia de 0.85. 74 Correl (Tabla 1). En el otro extremo encontramos la cronología derivada a partir de las series de tamaño medio de los vasos, que muestra los valores más bajos tanto de correlaciones mínimas y máximas (0,041 y 0,405, respectivamente). Estos valores estarían indicando un porcentaje mayor de señal común presente en las series de anchos de anillos, seguidas por las de área total y número total de vasos y un porcentaje menor entre las series que componen la cronología de tamaño medio del vaso. Estos resultados son consistentes con los valores de Rbar que decrecen desde la cronología de anchos de anillos (0,414), a la de tamaño del vaso Variabilidad interanual de los caracteres anatómicos en el leño de Cedrella lilloi en el NOA medio (0,210), pasando por las de número total (0,325) y área total (0,315) de vasos (Tabla 1). La evolución temporal de los estadísticos Rbar y EPS, medidos en intervalos de 50 años, proveen similar información (Fig. 6). Siempre los valores de Rbar y EPS son más altos para la cronología de ancho de anillos, seguidos por las cronologías de número total y área total de vasos. Para estas tres cronologías los valores de EPS se encuentran muy próximos o son superiores al valor límite fijado de 0,85. Por el contrario, la cronología de tamaño medio de los vasos muestra valores menores de Rbar que las otras tres cronologías y valores de EPS fluctuando entre 0,6 y 0,7 durante los últimos 200 años. Interacciones entre caracteres anatómicos El contar con series temporales de diversos caracteres anatómicos del leño de Cedrela lilloi nos permite evaluar, con resolución anual, los cambios temporales en la distribución de los mismos. ¿Es la proporción entre elementos conductores del leño, representado por los vasos, y el resto de los tejidos en el plan leñoso constante, o varía con la edad y el tamaño de los árboles? La Figura 7 muestra las variaciones interanuales en el porcentaje del área total del anillo ocupada por tejido conductor, específicamente los vasos. Se observa que durante los primeros 40 años de vida de los árboles, durante la formación de leño juvenil, el porcentaje Porcentaje vasos (%) 25 20 15 10 Tamaño vaso 5 Número de vasos 1800 1820 1840 1860 1880 1900 1920 1940 1960 1980 2000 Figura 7. Variaciones interanuales en el porcentaje de área en el plan leñoso ocupada por el tejido conductor (línea llena), el tamaño medio del vaso (círculo gris claro) y el número de vasos (círculo gris oscuro) por anillo de crecimiento. Los valores de tamaño medio del vaso provienen de la serie media (Promedio) de la Figura 5, en tanto que los valores del número de vasos provienen de la serie media (Promedio) de la Figura 4. Para facilitar la comparación, la serie de número de vasos se muestra en forma invertida. 75 Villalba, R.; S. Delgado; M. De Membiela y D. Mendoza del leño ocupado por los elementos conductores raramente supera el 10 % del plan leñoso. Conforme aumenta la edad y el tamaño de los árboles, la superficie de elementos conductores por unidad de madera se incrementa, llegando a valores medios de un 18% durante los últimos 60 años. En la Figura 7 se muestran las variaciones interanuales en el porcentaje del leño ocupado por los vasos, junto con la evolución temporal del número de vasos y tamaño medio del vaso. A partir de estas observaciones se ve claramente que el aumento en el porcentaje del leño ocupado por los vasos está asociado con el aumento paulatino en el tamaño de los vasos y no con el número, que disminuye a medida que aumenta la edad del leño. DISCUSION En base al material leñoso colectado en las vecindades de la localidad de Los Toldos, Provincia de Salta, hemos desarrollado cuatro cronologías basadas en los siguientes caracteres anatómicos del leño de Cedrela lilloi: ancho de los anillos, área total de vasos por anillo de crecimiento, número total de vasos por anillo de crecimiento y tamaño medio de los vasos por anillo de crecimiento. Estas cronologías fueron desarrolladas a partir de la medición de 1.817 anillos de crecimientos y 88.354 vasos provenientes de 10 muestras de un rodal coetáneo con árboles de aproximadamente 200 años de edad. El período cubierto por las cuatro cronologías es 1796-1999 (204 años). Desde el punto de vista dendrocronológi76 co, las series que componen la cronología de anchos de anillos son las que poseen mayor porcentaje de señal común, seguidas de cerca por las series de área total y número de vasos (Tabla 1). Las series de tamaño medio del vaso, por el contrario, no presentan una señal común muy fuerte, lo que sugiere, comparativamente, la necesidad de contar con un número mayor de muestras para incrementar la calidad de las cronologías derivadas a partir de esta variable. Las series individuales de anchos de anillos presentan una clara tendencia a disminuir el espesor con la edad. Esta tendencia es común a todos los árboles estudiados y claramente se refleja en la curva de crecimiento medio (Fig. 2). Sin embargo, es importante señalar que esta reducción en el ancho de los anillos no está directamente relacionada con una disminución en crecimiento con la edad del árbol, sino que obedece a un principio geométrico de distribuir una cantidad dada de madera en círculos concéntricos cada vez de mayor diámetro. Esto queda claramente evidenciado cuando el crecimiento se expresa en área basal y no ancho de anillos. En la Figura 8, se han representado los incrementos corriente y medio en área basal para las 10 muestras estudiadas y se observa un aumento sostenido en la producción de madera hasta aproximadamente el año 1950, cuando los árboles muestreados con una edad aproximada de 150 años alcanzaban un diámetro medio de alrededor de 1 m (considerando una corteza de 4-5 cm de espesor a esa edad). Variabilidad interanual de los caracteres anatómicos en el leño de Cedrella lilloi en el NOA Dado que el área total de vasos por anillo de crecimiento no disminuye marcadamente con la edad (Fig. 3), en tanto que el número total de los vasos sigue una tendencia a disminuir similar al ancho de los anillos (Fig. 4), inferimos que el aumento del área ocupado por los vasos conductores respondía al incremento en el tamaño de los vasos con la edad (Fig . 7). Esta observación es consistente con la mayoría de los estudios en latifoliadas que indican un aumento en el diámetro de los vasos desde la médula hacia la corteza (Carlquist 1988). Esta tendencia responde a la necesidad de acomodar las demandas de agua a una copa más desarrollada con mayor masa foliar. Por otro lado, es importante señalar que el transporte de agua en los vasos, o conductividad, es proporcional a la cuarta potencia del radio del vaso (Zim- merman 1983), por lo que un vaso con un diámetro sólo dos veces mayor que otro transporta 16 veces más agua. En vasos de grandes dimensiones, la fricción capilar se reduce marcadamente. Por lo tanto, la posible reducción en el transporte de agua asociada a la disminución del número de vasos (Fig. 4) registrado en Cedrela lilloi, es ampliamente compensada por el aumento en el tamaño de los vasos con la edad (Fig. 5). Nuestro análisis de la evolución temporal de los caracteres anatómicos del leño de Cedrela lilloi manifiesta la existencia de una etapa de leño juvenil con un arreglo muy particular del plan leñoso. Esta etapa de leño juvenil está caracterizada por bandas de crecimiento con un mayor número de vasos (Fig. 4) de menor tamaño (Fig. 5) Crecimiento basal (m2) 0.008 0.006 0.004 0.002 0.000 1800 1825 1850 1875 1900 1925 1950 1975 2000 Figura 8. Crecimiento promedio corriente (ICA) y medio anual (IMA) calculados en función de los incrementos anuales en área basal a partir de las 10 series de anchos de anillos incluidas en la Figura 2. La edad del turno biológico de corta se alcanza aproximadamente a los 150 años de edad, donde la curva de IMA corta la curva de ICA. 77 Villalba, R.; S. Delgado; M. De Membiela y D. Mendoza y un menor porcentaje de elementos de conducción en relación a otros elementos constitutivos del leño (Fig. 7). Aún cuando no tan claramente definido como el leño juvenil, el análisis de estas variables anatómicas sugiere, durante los últimos 60 años de crecimiento, la presencia de un leño maduro caracterizado por un número reducido y estable de vasos, y la falta de tendencias de largo plazo en el tamaño del vaso medio y en el porcentaje del tejido de conducción. Entre estos leños de tipo “joven” y “maduro” distinguimos una etapa de transición prolongada de más de 100 años, caracterizada por una reducción paulatina en el número de vasos cada vez más grandes y representando porcentajes mayores del plan leñoso. Para facilitar la identificación de estas tres etapas en el leño de Cedrela lilloi, el número, tamaño y porcentaje de tejido conductivo fueron combinados en una única variable empleando análisis de componentes principales (Cooley y Lohnes 1971). Las amplitudes del primer componente (PC1), que explican más del 86 % de la variabilidad total en estas tres variables, se muestra en la Figura 9. Durante la etapa de leño juvenil que abarca aproximadamente los primeros 35 años de vida de los árboles, el incremento interanual en las amplitudes es el más marcado de toda la serie. Este período juvenil es seguido por un intervalo prolongado de “crecimiento sostenido” en las amplitudes. Finalmente, a partir de 1940, se observa Crecimiento basal (m2) 2 1 0 -1 -2 -3 1800 1820 1840 1860 1880 1900 1920 1940 1960 1980 2000 Figura 9. Amplitudes del Primer Componente (PC1) que resulta de un análisis de componentes principales de las tres variables incluidas en la Figura 7: porcentaje de vasos, tamaño medio del vaso y número total de vasos por anillo de crecimiento. En base a las tendencias dominantes en los distintos períodos, las amplitudes fueron divididas en tres intervalos correspondientes al leño juvenil (gris claro), leño adulto (gris oscuro) y leño maduro (negro). 78 Variabilidad interanual de los caracteres anatómicos en el leño de Cedrella lilloi en el NOA un período de “estabilidad”, donde no se registran cambios significativos en la tendencia de largo plazo de las amplitudes. En consecuencia, este análisis combinado de la evolución temporal de los caracteres anatómicos de Cedrela lilloi nos permite identificar a través del leño tres etapas en el desarrollo de los individuos de esta especie: juvenil, adulta y madura. Leños con alto número (cercanos a 100) de vasos pequeños (< 2000 μm2) y bajo porcentaje (< de 10%) de tejido conductor por anillo, corresponden a etapas juveniles en el crecimiento de los individuos de Cedrela lilloi. Por el contrario, leños con un número relativamente bajo (cercano a 40) de vasos de gran tamaño (> 3000 μm2) constituyendo más del 15% en promedio del plan leñoso transversal, provendrían de leños formados durante las etapas de madurez de los árboles. Las situaciones intermedias corresponderían al leño de tipo “adulto”. No es coincidente que la transición de las etapas adulta a madura en el leño ocurra aproximadamente a los 150 años de edad. En este momento se alcanza el turno biológico de corta, dado por la intersección de las curvas de crecimiento basal corriente y medio (Fig. 8). Estos cambios en la anatomía del leño de Cedrela lilloi con la edad, seguramente introducen modificaciones en las propiedades físico-mecánicas de la madera y por lo tanto tiene importantes implicaciones en los distintos usos y aplicaciones de esta madera de alto valor comercial. AGRADECIMIENTOS Este trabajo fue parcialmente financiado por el CONICET y la Agencia Nacional de Promoción Científica y Tecnológica (PICTR02-123). Ricardo Grau, Mariano Morales, Leonardo Paolini y Alberto Ripalta colaboraron en la colección del material leñoso. BIBLIOGRAFIA Blank, R. 1997. Ringporigkeit des Holzes und häufige Entlaubung durch Insekten als spezifische Risikofaktoren der Eichen. Forst und Holz 52(9): 235-242. Briffa, K.R. 1995. Interpreting High-Resolution Proxy Climate Data – The Example of Dendroclimatology. En von Storch, H. y A. Navarra (Eds.). Analysis of Climate Variability, Applications of Statistical Techniques. Springer, Berlin. pp: 77-94. Briffa, K.R.; P.D. Jones y F.H. Schweingruber. 1992. Tree-ring density reconstructions of summer temperature patterns across western North America since 1600. Journal of Climate 5(7): 735-754. Briffa, K.R.; P.D. Jones; T.S. Bartholin; F.H. Schweingruber; W. Karlen y S.G. Shiyatov: 1996. Tree-Ring Variables as Proxy-Climate Indicators: Problems with Low-Fre79 Villalba, R.; S. Delgado; M. De Membiela y D. Mendoza quency Signals. En Jones, P.D.; R.S. Bradley y J. Jouzel (Eds). Climate Variations and Forcing Mechanisms of the Last 2000 Years. NATO ASI Series. Vol. I41. Springer, Heidelberg. pp. 9-41. Carlquist, S. 1988. Comparative wood anatomy, systematic, ecological, and evolutionary aspects of dycotyledon wood. Springer-Verlag, New York. Cook, E.R. 1985. A time series analysis approach to tree ring standardization. Ph.D. dissertation, University of Arizona, Tucson. 171 pp. Cook, E.R. y R. Holmes. 1984. User’s manual for Program ARSTAN. Laboratory of Tree Ring Research,University of Arizona, Tucson. 15 pp. Cook, E. y L. Kairiukstis (Eds.). 1990. Methods of Dendrochronology, Kluwer Academic Publishers. Amsterdam, The Netherlands. pp. 104-132. Cooley, W.W. y P.R. Lohnes. 1971. Multivariate Data Analysis, Wiley, New York, U.S.A. Fritts, H.C. 1976. Tree Rings and Climate. Academic Press, New York. 567 pp. García Gonzalez, I. y D. Eckstein. 2003. Climatic signal of earlywood vessels of oak on a maritime site. Tree Physiology 23: 497-504. Grissino-Mayer, H.D. y H.C. Fritts. 1997. The International Tree-Ring Data Bank: an enhanced global database serving the global scientific community. The Holocene 7(2): 235-238. Holmes, R.L. 1983. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bulletin 43: 69-78. Mosteller, F. y J.W. Tukey. 1977. Data analysis and regression. Reading, MA, USA. Addison. Wesley. Munro, M.A.R.; P.M. Brown; M.K. Hughes y E.M.R. García. 1996. Image analysis of tracheid dimensions for dendrochronological use. En Dean, J.S.; D.M. Meko y T.W. Swetnam (Eds.). Tree Rings, Environment, and Humanity. Radiocarbon pp 843-852. Sass, U. y D. Eckstein. 1992. The annual vessel area of beech as an ecological indicator. En: Bartholin, T.S.; B.E. Berglund; D. Eckstein; F.H. Schweingruber y O. Eggertsson (Eds.). Tree Rings and Environment: Proceedings of the International Symposium, Ystad, South Sweden, 3-9 September, 1990. Lundqua Report (Department of Quaternary Geology, Lund University, Sweden). 34: 281-285. 80 Variabilidad interanual de los caracteres anatómicos en el leño de Cedrella lilloi en el NOA Schweingruber, F.H. 1988. Climatic information for the past hundred years in width and density of conifer growth rings. En: Wanner, H. y U. Siegenthaler (Eds.). Long and Short Term Variability. Lecture Notes in Earth Sciences 16: 35-55. Schweingruber, F.H. 1996. Tree Rings and Environment. Dendroecology. Berne: Paul Haupt Verlag. 609 pp. Schweingruber, F.H. y K.R. Briffa. 1996. Tree-ring density networks for climate reconstruction. En Jones, P.D.; R.S. Bradley y J. Jouzel (Eds.). Climatic Variations and Forcing Mechanisms of the Last 2000 Years. NATO ASI Series. 41: 43-66. Stokes, M.A. y T.L. Smiley. 1968. An Introduction to Tree-Ring Dating. University of Chicago Press, Chicago. 73 pp. Villalba, R. 2000. Dendroclimatology: a Southern Hemisphere Perspective. Paleo- and Neoclimates of the Southern Hemisphere: the state of the arts. Smolka, P. y W. Volkheimer (Eds.). Springer. pp. 105-143. Villalba, R.; S. Delgado; D. Mendoza. 2002 . Climatic influences on vessel size of Nothofagus pumilio in the Southern Andes. 6th International Conference on Dendrochronology. Quebec, Canada. Villar-Salvador, P.; P. Castro Díez; C. Perez-Rontomé y G. Montserrat Martí. 1997. Stem xylem features in three Quercus (Fagaceae) species along a climatic gradient in NE Spain. Trees 12: 90-96. Woodcock, D.W. 1989. Climate sensitivity of wood-anatomical features in a ring-porous oak (Quercus macrocarpa). Canadian Journal of Forest Research 19: 639-644. Zimmerman, MH. 1983. Xylem structure and ascent of sap. New York, USA: SpringerVerlag. 81 Ecología y producción de cedro (género Cedrela) en las Yungas australes. Pacheco, S. y A. Brown (Eds.). LIEY-ProYungas. Argentina. pp. 83-104. 2006. Diversidad, estructura y distribución de bosques con cedro en el noroeste de Argentina y sur de Bolivia Malizia, L.R. 1,2,3; C. Blundo1,2 y S. Pacheco1,2 Fundación ProYungas, CC 34 (4107), Yerba Buena, Tucumán, Argentina. 2 Laboratorio de Investigaciones Ecológicas de las Yungas (LIEY, UNT). 3 International Center for Tropical Ecology, University of Missouri - St. Louis. 1 Describimos la riqueza y composición de especies de árboles y la estructura del bosque en las Yungas de la Alta Cuenca del Río Bermejo y brindamos información sobre abundancia, parámetros estructurales y distribución de Cedrela balansae y C. lilloi. Los muestreos se realizaron en 30 parcelas de 1 ha cada una en 4 elevaciones aproximadas (600, 1.100, 1.600 y 2.100 m). La composición de especies varió relativamente poco entre parcelas en una misma elevación, pero hubo un recambio casi total entre parcelas ubicadas en los extremos del gradiente altitudinal. La riqueza de especies, géneros y familias disminuyó con la altitud, con un pico leve a los 1.100 m. La estructura del bosque también varió con la altitud, siendo mayor el área basal y menor la altura del dosel a los 2.100 m. Las especies de cedro presentaron abundancias intermedias en relación a las otras especies de árboles censadas. C. balansae fue registrada entre los 500 m y 1.100 m, con mayor densidad y área basal a los 600 m. C. lilloi se encontró entre los 1.000 m y 2.100 m, con mayor densidad a los 2.100 m y mayor área basal a los 1.600 m. Basados en modelos de distribución, estimamos que C. balansae ocupa unos 8.500 km2 y C. lilloi, 9.200 km2 en el área de estudio, y que ambos se presentan en simpatría sobre una superficie de aproximadamente 600 km2, entre los 1.000 y 1.100 m. Finalmente, analizamos los factores que podrían determinar los patrones estructurales y de diversidad encontrados, y analizamos posibles estrategias de conservación y uso sustentable del cedro en la región. We describe tree species richness and composition, and forest structure in the Yungas of the Upper Bermejo River Basin and provide information on abundance, distribution and structural parameters for Cedrela balansae and C. lilloi. We sampled 30 1-ha plots at four main elevations (600, 1100, 1600 and 2100 m). Species composition varied relatively little among plots at the same elevation, but showed high turnover among plots located at the extremes of the altitudinal gradient. Species, genera and family richness decreased with elevation, with a minor peak at 1100 m. Forest structure also varied with elevation, with highest basal area and lowest canopy height at 2100 m. Cedrela species showed intermediate abundances compared with other censused tree species. C. balansae was found between 500 and 1100 m, with highest density and basal area at 600 m. C. lilloi was found between 1000 and 2100 m, with highest density at 2100 m and highest basal area at 1600 m. Based on ecological niche models, we estimated that C. balansae occupied 8500 km2 and C. lilloi 9200 km2 in the study area, and that they occurred in sympatry over an area of about 600 km2, between 1000 and 1100 m. We finally discuss the factors that may determine the structural and diversity patterns found, and analyze possible conservation and sustainable-use strategies for Cedrela in the region. 83 Malizia, L. R., C. Blundo y S. Pacheco INTRODUCCION La información disponible sobre diversidad de árboles (i.e. riqueza y composición de especies) y estructura (i.e., densidad y área basal) del bosque de las Yungas Subtropicales (sur de Bolivia y noroeste de Argentina) es relativamente escasa. Estudios realizados en otras regiones de bosque montano tropical muestran un empobrecimiento en la riqueza de especies con aumentos en la elevación, una alta tasa de recambio de especies entre pisos altitudinales, y marcados cambios en los parámetros estructurales del bosque, como por ejemplo una disminución en la altura de los árboles con la elevación, o un incremento en área basal hacia las partes altas (Richards 1952, Grubb 1974, Grubb y Stevens 1985, Proctor et al. 1988, Gentry 1995, Lieberman et al.1996, Vázquez y Givnish 1998). Existen estudios previos para las Yungas Subtropicales que analizan la riqueza y composición de especies arbóreas (Brown et al. 1985, Brown 1986, Brown y Ramadori 1989, Brown 1995, Grau y Brown 1995, Morales et al. 1995, Grau 1999) y unos pocos que consideran aspectos de estructura del bosque en el gradiente altitudinal (Brown 1986, Grau et al. 1997, Grau y Brown 1998). Sin embargo, estos estudios han sido basados en tamaños de muestreo relativamente pequeños y no han puesto énfasis en la Alta Cuenca del Río Bermejo, el área de mayor diversidad de las Yungas Subtropicales (Grau y Brown 2000). Las Yungas Subtropicales comprenden un rango altitudinal que se extiende aproximadamente desde los 400 a los 84 2.500 m s.n.m. Este gradiente altitudinal está asociado con cambios en las condiciones ambientales, principalmente en precipitación y temperatura, a los cuales la vegetación arbórea responde con cambios bien marcados en composición específica y probablemente en características estructurales a lo largo del gradiente. En base a la composición de especies se diferencian de modo aproximado tres pisos altitudinales de vegetación: el piso inferior de Selva Pedemontana, el piso intermedio de Selva Montana y el piso superior de Bosque Montano (Brown 1995, Brown et al. 2001). Las Yungas Subtropicales albergan dos especies de cedro, el cedro orán (Cedrela balansae) y el cedro coya (C. lilloi). En términos generales, C. balansae se distribuye principalmente en la Selva Pedemontana, mientras que C. lilloi está presente en la Selva Montana y el Bosque Montano. Recientemente se ha sugerido la presencia adicional de una tercera especie para la región, C. saltensis (Zapater et al. 2004), muy afín es su morfología con C. balansae (Grau et al. este volumen). Esta nueva especie no es considerada en este trabajo, ya que su descubrimiento fue posterior al trabajo de campo. Este hecho evidencia las limitaciones de la información disponible, que se profundizan al considerar parámetros más específicos como la distribución espacial, la abundancia, la distribución de tamaños y el área basal de estas especies. Los cedros se destacan por la calidad de su madera, que está entre las más cotizadas del mercado regional (aproximadamente US$ 170-300/m3 para C. lilloi y US$ 140- Diversidad, estructura y distribución de bosques con cedro en la Alta Cuenca del Río Bermejo 200/m3 para C. balansae como madera no aserrada), lo que los ha expuesto a un régimen de extracción intenso por parte de la industria forestal, sin una planificación que asegure la sustentabilidad económica y biológica de este recurso. Las dificultades de las autoridades competentes para ordenar el uso del recurso maderero y la escasez de información básica, dificultan la posibilidad de desarrollar programas de aprovechamiento sustentable a nivel regional para estas especies. En el mejor de los casos, la información que se genera sobre densidad y estructura es a nivel predial, lo que permite realizar planes de aprovechamiento a esta escala, pero sin un contexto regional que conduzca los planes locales. Todo esto resulta en que no existan estrategias de uso y conservación de las especies de cedro (ni de ninguna otra especie maderable) a escala regional. En este trabajo, presentamos aspectos de la diversidad (considerando número de especies, géneros y familias y composición y recambio de especies) y de la estructura del bosque (incluyendo densidad, área basal y altura máxima) a lo largo del gradiente altitudinal de Yungas del noroeste de Argentina y sur de Bolivia. Además, determinamos la importancia en términos estructurales (densidad, abundancia relativa y área basal) y modelamos el área de distribución de C. balansae y C. lilloi para este sector de Yungas. metodos Este estudio se enfoca principalmente en la Reserva de la Biosfera de las Yungas y zonas aledañas, abarcando una superficie aproximada de 2 millones de ha. Allí establecimos 30 parcelas de 1 ha cada una entre noviembre de 2002 y abril de 2004. Las parcelas se ubicaron desde el Parque Nacional Baritú, en el norte de la Provincia de Salta, y la Reserva de Flora y Fauna de Tariquía, en el sur de Bolivia, hasta la Serranía de Zapla, en el sur de la Provincia de Jujuy (Fig. 1). Estas parcelas fueron distribuidas en cuatro elevaciones aproximadas: 600, 1.100, 1.600 y 2.100 m s.n.m. (ocho parcelas en cada una de las dos elevaciones inferiores y siete parcelas en cada una de las dos superiores) (Apéndice). Todas las parcelas midieron 20 x 500 m, corregidas por pendiente para cubrir 1 ha (Condit 1998). En cada parcela identificamos a nivel de especie y medimos todos los árboles ≥ 10 cm de diámetro a 1,30 m del suelo (i.e., diámetro a la altura del pecho o DAP). Cada árbol fue marcado con una chapa de aluminio numerada y se le midió DAP y altura total. Además, registramos el número de tocones (árboles aserrados) por parcela para hacer una estimación de la intensidad de la extracción forestal ocurrida en los distintos bosques. Para cada parcela determinamos la riqueza (i.e., número) de especies, géneros y familias; la estructura del bosque en términos de densidad, área basal y altura máxima considerando todas las especies; y la intensidad de uso (número de tocones). Correlacionamos estos parámetros con la elevación de las parcelas. Debido a que algunas variables no cumplieron con los supuestos de normalidad y homocedasticidad, utilizamos correlaciones 85 Malizia, L. R., C. Blundo y S. Pacheco índice de similitud de Sørensen, utilizando el programa EstimateS (Colwell 2000). Para cada especie de cedro, determinamos densidad, abundancia relativa, área basal y distribución de tamaños (diámetros) y correlacionamos (Spearman) las tres primeras variables con la posición en el gradiente altitudinal de las parcelas. Además, comparamos las variables estructurales medidas para los cedros en parcelas ubicadas en áreas protegidas y en propiedades privadas, utilizando el test no paramétrico de comparación de medias (U de MannWhitney). Describimos la distribución de cedros a nivel regional (21°-25.5° S, ~ 65.000 km2) utilizando el modelo de nicho ecológico GARP (Genetic Algorithm for RuleFigura 1. Mapa de distribución de la vegetación en la Set Prediction; http://biodi.sdsc. Alta Cuenca del Río Bermejo, incluyendo la distribu- edu/), calculado con el programa ción de las 30 parcelas permanentes de muestreo y los Desktop GARP (disponible en 29 sitios de validación para los modelos de distribución http://beta.lifemapper.org/desktode GARP (ver Métodos). pgarp/). GARP busca asociaciones no aleatorias entre las característino paramétricas (Spearman). Analizamos cas ambientales de los sitios donde se enla composición florística en el gradiente cuentra presente la especie de interés y las altitudinal empleando datos de presencia- características de toda el área de estudio ausencia de especies en un ordenamiento (Stockwell y Noble 1992, Stockwell y Peno-métrico de escalamiento multidimen- ters 1999). Luego de generar 100 modelos sional (NMDS) basado en el índice de si- GARP para cada especie de cedro, obtumilitud de Sørensen, utilizando el progra- vimos un mapa hipotético de distribución ma PC-ORD (McCune y Mefford 1999). de cada especie resumiendo los 20 mejoFinalmente, calculamos el recambio de res modelos (Anderson et al. 2003, Leviespecies entre elevaciones mediante el ne et al. 2004). De este modo, cada punto 86 Diversidad, estructura y distribución de bosques con cedro en la Alta Cuenca del Río Bermejo (píxel) del mapa de distribución muestra un valor predictivo de presencia: 20 es el valor máximo donde todos los modelos seleccionados predicen la presencia de la especie y 0 es el valor mínimo donde ningún modelo predice la presencia de la especie. La obtención de los modelos se hizo en dos etapas. Primero generamos los modelos de distribución con GARP usando los datos de presencia de las 30 parcelas de muestreo (datos de “entrenamiento”). Luego verificamos la validez de estos modelos utilizando otras 29 parcelas de muestreo independientes (datos de “validación”). Los sitios de validación se ubicaron en lugares que no habían sido muestreados previamente (Fig. 1). Cada sitio cubrió aproximadamente 0,8 km2, que es el tamaño de píxel que se usó en la generación de los modelos (ver más abajo). En los sitios de validación sólo registramos presenciaausencia de especies. Evaluamos la significancia de los modelos de distribución de cada especie usando la prueba estadística no paramétrica de dos muestras de Mann-Whitney (Elith y Burgman 2002). Básicamente, evaluamos si los sitios con presencia confirmada (según los datos de validación) presentaron valores de GARP mayores que los sitios donde la especie no fue encontrada (según los datos de validación). Esta prueba es recomendable porque brinda una medida del desempeño del modelo que no depende de un umbral prefijado (Elith y Burgman 2002). Las bases de datos ambientales (coberturas de SIG) usadas para crear los modelos incluyeron variables climáticas, topográ- ficas y de cobertura de la vegetación. En total utilizamos 10 variables: temperatura media anual, isotermalidad (rango medio diario/rango anual), estacionalidad térmica (desvío estándar anual de las temperaturas mensuales medias), orientación de la ladera (utilizando transformaciones de seno y coseno), pendiente, elevación y tres variables estimadas de NDVI (Normalized Difference Vegetation Index, ver más abajo). Empleamos los datos de temperatura de WORLDCLIM (http://biogeo. berkeley.edu/worldclim/worldclim.html), disponibles en una grilla con resolución de 30 arco segundos (~1 km2 en el ecuador y ~0.72 km2 en el área de estudio) (Hijmans et al. 2004). Utilizamos datos digitales de elevación producidos por la NASA (ftp://edcsgs9.cr.usgs.gov/pub/data/srtm/), disponibles a escala global a una resolución de 3 arco-segundos (~ 90 m x 90 m en el ecuador). A partir de esta superficie de elevación calculamos pendiente media, orientación media y elevación media a una resolución de 30 arco segundos. Obtuvimos datos de NDVI del Land Processes Distributed Active Archive Center (http:// edcdaac.usgs.gov/1KM/1kmhomepage. html), disponibles a una resolución aproximada de 1,1 km2. NDVI es una medida de la reflectividad de la vegetación de la superficie de la tierra (Lillesand y Keifer 2000) y es representativa de la productividad de la vegetación (Chong et al. 1993). Basándonos en valores mensuales máximos, calculamos el valor máximo de NDVI, la estacionalidad anual y una medida intermedia de “verdor” (para más detalles consultar Parra et al. 2004). Todas 87 Malizia, L. R., C. Blundo y S. Pacheco las grillas de NDVI fueron re-calculadas a una resolución de 30 arco segundos para que coincidan con las grillas climáticas y de elevación. RESULTADOS Diversidad de especies Censamos un total de 15.028 árboles ≥10 cm de DAP pertenecientes a 138 especies, 115 géneros y 53 familias. El número promedio de especies a los 600 y 1.100 m s.n.m. fue de aproximadamente 39 especies/ha y luego disminuyó con la altitud (rS = -0,76, P < 0,001, n = 30) (Tabla 1, Fig. 2). El mismo patrón se repitió considerando número de géneros (rS = -0,76, P < 0,001, n = 30), lo cual refleja el hecho de que el 90 % de los géneros encontrados estuvieron representados por una única especie. El número promedio de familias presentó un pico a los 1.100 m s.n.m. y luego disminuyó hasta los 2.100 m s.n.m., mostrando una correlación negativamente menos marcada con la altitud (rS = -0,55, P < 0,01, n = 30) (Tabla 1, Fig. 2). El 49% de las familias estuvo representada por un sólo género, mientras que unas pocas familias estuvieron representadas por un número medio a alto de géneros: Anacardiáceas (4), Rubiáceas (4), Sapindáceas (4), Euforbiáceas (6), Mirtáceas (6) y Leguminosas (14). El número total de especies, géneros y familias por elevación alcanzó un valor máximo a los 1.100 m s.n.m. y luego disminuyó con la elevación (Tabla 1). Composición y recambio de especies La composición de especies varió claramente a lo largo del gradiente altitudinal, como se observa en el ordenamiento nométrico de escalamiento multidimensional (Fig. 3). El primer eje de este ordenamiento explicó el 77 % de la variación de los datos, mientras que el eje 2 sólo explicó el 5 % de la variación de los datos. Las parcelas de muestreo se dispusieron a lo largo del eje 1 aproximadamente en 4 grupos de puntos correspondientes con su elevación, desde los 600 a los 2.100 m s.n.m. Este agrupamiento de las parcelas sugiere que la composición de especies responde marcadamente a factores ambientales que cambian con la elevación. Tabla 1. Promedio (± error estándar) de especies, géneros y familias por hectárea de árboles ≥10 cm DAP para cada elevación de las Yungas de la Alta Cuenca del Río Bermejo, noroeste de Argentina y sur de Bolivia. En corchetes se indica el número total de especies, géneros o familias por elevación. Elevación aproximada (m) 600 (n = 8) 1.100 (n = 8) 1.600 (n = 7) 2.100 (n = 7) Número de especie 39,0 ± 2,9 [78] 39,6 ± 2,4 [95] 28,2 ± 2,4 [63] 14,2 ± 2,0 [38] Número de géneros 36,2 ± 2,9 [70] 36,7 ± 2,1 [81] 26,0 ± 2,2 [56] 13,2 ± 1,7 [35] Número de familias 22,1 ± 1,5 [34] 26,6 ± 1,2 [42] 21,1 ± 1,2 [36] 12,5 ± 1,3 [27] 88 Diversidad, estructura y distribución de bosques con cedro en la Alta Cuenca del Río Bermejo Figura 2. Número de especies, géneros y familias por elevación en las Yungas de la Alta Cuenca del Río Bermejo (noroeste de Argentina y sur de Bolivia). La Selva Pedemontana (ca. 600 m s.n.m.) estuvo caracterizada por Anadenanthera colubrina (13,8% de todos los individuos censados en este piso altitudinal), Phyllostylon rhamnoides (10,9 %), Trichilia hieronymi (7,2%), Parapiptadenia excelsa (5,9%), Calycophyllum multiflorum (5,5%), Astronium urundeuva (5,2 %) y Ruprechtia apetala (4,2 %). Con excepción de T. hieronymi, son todas especies deciduas que pasan la temporada seca sin follaje, respondiendo a la marcada estacionalidad climática del pedemonte. La Selva Montana (ca. 1.100 m s.n.m.) estuvo caracterizada por Miconia molybdea (16,3 %), Croton densiflorus (5,6 %), Siphoneugenia occidentalis (4,5%), Blepharocalyx salicifolius (4,3%), Inga marginata (3,9 %), Parapiptadenia excelsa (2,9 %) y Nectandra pichurin (2,7 %). La transición entre Selva Montana y Bosque Montano (i.e., 1.600 m s.n.m.) estuvo caracterizada por Allophylus edulis (13,8 %), Podocar- pus parlatorei (13,4 %), Blepharocalyx salicifolius (11,8 %), Parapiptadenia excelsa (6,1 %), Myrcianthes pseudomato (5,7 %), Juglans australis (4,2 %) e Ilex argentina (4,1 %). En fuerte contraste con la Selva Pedemontana, la mayoría de las especies dominantes en la Selva Montana son perennifolias (con excepción de P. excelsa y J. australis), lo que probablemente responde a una mayor disponibilidad de agua durante la época seca, en parte en forma de neblina. Finalmente, las especies típicas del Bosque Montano (ca. 2.100 m s.n.m.) fueron Podocarpus parlatorei (43,4 %), Prunus tucumanensis (13,3 %), Viburnum seemenii (11,2 %), Ilex argentina (7,7 %), Crinodendron tucumanum (5,2 %), Cedrela lilloi (3,1 %) y Alnus acuminata (2,7 %). Muchas de estas especies son de origen austral o boreal, lo que explicaría su adaptación a las bajas temperaturas invernales a las que están sometidas en este piso de vegetación. A esta 89 Malizia, L. R., C. Blundo y S. Pacheco Figura 3. Cambio en la composición de especies a lo largo del gradiente altitudinal de bosques en la Alta Cuenca del Río Bermejo (noroeste de Argentina y sur de Bolivia), evidenciado a partir de un ordenamiento no-métrico de escalamiento multidimensional (NMDS) con datos de presencia y ausencia de especies de árboles ≥10 cm de DAP. altitud, entre las más abundantes encontramos tanto especies perennifolias (las cuatro especies más abundantes) como caducifolias (C. tucumanun, C. lilloi y A. acuminata). en términos de composición de especies que los sitios ubicados en elevaciones adyacentes. La composición de especies varió relativamente poco al interior de un mismo piso altitudinal, particularmente dentro de la Selva Pedemontana (similitud de especies entre parcelas de 1 ha: 0,64 ± 0,03 EE), con un pico de variación de la composición en el bosque montano (0,46 ± 0,03 EE) (Tabla 2). El recambio de especies fue mayor entre elevaciones, con una disminución de los índices de similitud al aumentar la separación altitudinal entre parcelas (Tabla 2). De este modo, los sitios ubicados en elevaciones distantes tendieron a compartir menos especies y en consecuencia fueron menos similares La densidad de individuos no varió en función de la altitud (rS = 0,19, P = 0,31, n = 30), aunque fue mayor en promedio en el Bosque Montano (1.600 y 2.100 m s. n.m.), alcanzando un máximo de 573,7 ind/ha a los 1.600 m s.n.m. (Tabla 3). El área basal aumentó significativamente con la altitud (rS = 0,58, P < 0,001, n = 30), mientras que la altura máxima de los árboles disminuyó significativamente con la altitud (rS = -0,72, P < 0,0001, n = 28). Además, encontramos una tendencia a la disminución en el número de tocones por ha a medida que aumenta la altitud (rS = -0,36, P = 0,05, n = 30) (Tabla 3). 90 Estructura del bosque Diversidad, estructura y distribución de bosques con cedro en la Alta Cuenca del Río Bermejo Tabla 2. Variación en la composición de espe- cies dentro y entre elevaciones en las Yungas de la Alta Cuenca del Río Bermejo (noroeste de Argentina y sur de Bolivia), estimada con el índice de similitud de Sørensen promediado entre parcelas de 1 ha, considerando árboles ≥ 10 cm DAP. Comparaciones Similitud promedio ± EE Dentro de cada elevación 600 m 0,64 ± 0,03 1100 m 0,54 ± 0,02 1600 m 0,57 ± 0,02 2100 m 0,46 ± 0,03 Entre dos elevaciones consecutivas 600 m - 1100 m 0,40 ± 0,01 1100 m - 1600 m 0,31 ± 0,02 1600 m - 2100 m 0,39 ± 0,02 Una elevación de por medio 600 m - 1600 m 0,10 ± 0,01 1100 m - 2100 m 0,12 ± 0,01 Dos elevaciones de por medio 600 m – 2100 m 0,05 ± 0,002 Parámetros estructurales y distribución de las especies de cedro La densidad y la abundancia relativa por hectárea de los cedros (C. balansae y C. lilloi considerados juntos) aumentó significativamente con la elevación (densidad: rS = 0,47, P < 0,02, n = 26; abundancia relativa: rS = 0,47, P < 0,02, n = 26; Tabla 4). Analizando cada especie por separado, se observa que ocupan rangos altitudinales diferentes. Por un lado, C. balansae alcanza su densidad máxima en la Selva Pedemontana (600 m s.n.m.), llegando algunos individuos al sector basal de la Selva Montana (1.100 m s.n.m.). Por otro lado, C. lilloi se distribuye a partir de la Selva Montana (1.100 m s.n.m.), alcanzando densidades máximas en el bosque montano (2.100 m s.n.m.) (Tabla 4). Dentro de sus rangos de distribución característicos, C. lilloi presentó mayores densidades que C. balansae (Tabla 4). En relación a otras especies de árboles de las Yungas, C. lilloi fue relativamente común en las parcelas de muestreo en tres pisos de vegetación (247 individuos censados; posición número 15 en orden decreciente de abundancia considerando el total de individuos censados), mientras que C. balansae sólo se presentó en dos pisos de vegetación y a su vez fue menos común en cada uno (60 individuos censados, posición número 53 en orden decreciente de abundancia). La distribución de tamaños de los cedros nos muestra que la mayoría de los individuos midieron menos de 50 cm de DAP (Fig. 4). El 5 % de los individuos de C. balansae y el 9 % de los individuos de C. lilloi medidos en las parcelas de muestreo presentaron diámetros por arriba del tamaño mínimo de corta de 60 cm de DAP dispuesto para ambas especies por la Provincia de Jujuy. En el caso de la Provincia de Salta, donde el diámetro mínimo de corta es de 40 cm, el 30 % de los individuos de C. balansae y el 22 % de los individuos de C. lilloi presentaron DAP por arriba del diámetro mínimo legal. 91 Malizia, L. R., C. Blundo y S. Pacheco Tabla 3. Promedio (± error estándar) por hectárea de densidad, área basal y altura máxima de árboles ≥10 cm DAP, y número de tocones en el gradiente altitudinal de las Yungas de la Alta Cuenca del Río Bermejo (noroeste de Argentina y sur de Bolivia). (*) Correlaciones significativas (P<0,05) con la altitud. Parámetros por ha Elevación aproximada (m) 600 (n = 8) 1100 (n = 8) 1600 (n = 7) 2100 (n = 7) Densidad (ind/ha) 473,5 ± 27,5 458,3 ± 30,0 573,7 ± 34,3 508,3 ± 70,5 Area basal (m /ha)* 25,7 ± 1,3 27,5 ± 2,1 33,1 ± 1,9 36,0 ± 1,0 Altura máxima (m)* 24,8 ± 1,4 20,1 ± 1,1 19,7 ± 0,9 14,8 ± 0,6 2,9 ± 1,0 1,7 ± 0,5 0,1 ± 0,1 2 Num. de tocones (ind/ha) 3,8 ± 1,6 La densidad, la abundancia relativa y el área basal de los cedros no varió de modo significativo entre parcelas ubicadas en áreas protegidas y en propiedades privadas (no protegidas) (densidad: U = 48,0, P = 0,5; abundancia relativa: U = 42,5, P = 0,3; área basal: U = 47,0, P = 0,4). Los modelos de distribución desarrollados con GARP para cada una de las especies de cedro fueron significativos (pruebas de dos muestras de Mann-Whitney: C. balansae, U = 185,5, P = 0,0002; C. lilloi, U = 160,0, P = 0,0025), sugiriendo que reflejan de modo adecuado la distribución de estas especies en la región. Considerando sólo los píxel con una probabilidad de ocurrencia > 90 %, C. balansae abarcaría una superficie de aproximadamente 8.500 km2 en el área de estudio, mientras que C. lilloi abarcaría Tabla 4. Promedio (± error estándar) por hectárea de densidad, abundancia relativa y área basal para las especies de cedro, considerando árboles ≥10 cm DAP, en el gradiente altitudinal de las Yungas de la Alta Cuenca del Río Bermejo (noroeste de Argentina y sur de Bolivia). (*) Correlaciones significativas (P<0.05) con la altitud, considerando ambas especies en forma conjunta Parametros por especie Densidad (ind./ha)* C. balansae Abundancia (proporción) * C. balansae Area basal (m2/ha) C. balansae 92 Elevación aproximada (m) 600 1100 5 ± 1,3 3 ± 1,7 C. lilloi 9,7 ± 6,3 0,01 ± 0,002 C. lilloi C. lilloi 2100 15,7 ± 1,3 22,2 ± 11,5 0,005 ± 0,003 0,03 ± 0,02 0,5 ± 0,1 1600 0,03 ± 0,02 0,04 ± 0,02 0,3 ± 0,2 0,9 ± 0,4 2,6 ± 0,3 1,1 ± 0,7 Diversidad, estructura y distribución de bosques con cedro en la Alta Cuenca del Río Bermejo Figura 4. Distribución de diámetros (medidos a 1,3 m de altura) de Cedrela balansae (Cedbal) y C. lilloi (Cedlil) en las Yungas de la Alta Cuenca del Río Bermejo (noroeste de Argentina y sur de Bolivia). Figura 5. Area de distribución de C. balansae y de C. lilloi y área de superposición de ambas especies en las Yungas de la Alta Cuenca del Río Bermejo (noroeste de Argentina y sur de Bolivia), determinada a partir de modelos de nicho ecológico GARP. 93 Malizia, L. R., C. Blundo y S. Pacheco Figura 6. Area de distribución de Cedrela lilloi y C. balansae en el gradiente altitudinal de las Yungas de la Alta Cuenca del Río Bermejo (noroeste de Argentina y sur de Bolivia), determinado a partir de modelos de nicho ecológico GARP. una superficie aproximada de 9.200 km2 en el área de estudio (Fig. 5). En toda el área de estudio (aproximadamente 50.000 km2) sólo existirían, según los modelos, 600 km2 de superposición en la distribución de ambas especies, principalmente entre los 1.000 y los 1.100 m s. n.m. (Fig. 5). Según los modelos de distribución, C. balansae se distribuiría desde los 400 hasta los 1.400 m s.n.m., encontrándose el 90 % del área de distribución entre los 500 y los 1.100 m s.n.m. (Fig. 6). La distribución de C. lilloi sería más amplia y abarcaría desde los 700 hasta los 2.500 m s.n.m., con el 90 % del área de distribución entre los 1.000 y los 2.100 m s.n.m. (Fig. 6). 94 DISCUSION La diversidad de especies de árboles en las Yungas de la Alta Cuenca del Río Bermejo tendió a disminuir con la elevación. Existió además un marcado recambio de especies entre elevaciones, debido probablemente a las diferencias climáticas que se presentan a lo largo del gradiente altitudinal y a diferentes factores biogeográficos históricos relacionados con cada piso altitudinal de vegetación. La disminución de la diversidad de especies con la altitud ha sido reportada para otros bosques montanos del Neotrópico (Gentry 1988, Lieberman et al. 1996, Vásquez y Givnish 1998) y para otras regiones del mundo (Beaman y Beaman 1990, Kitayama 1992). Para las Yungas del noroeste de Argentina, tam- Diversidad, estructura y distribución de bosques con cedro en la Alta Cuenca del Río Bermejo bién se ha reportado previamente este patrón (Brown 1986, Grau y Brown 1995, Brown et al. 2001). Esta disminución de especies a lo largo de gradientes altitudinales no ocurre necesariamente de forma monotónica, sino que usualmente presenta un pico de riqueza a elevaciones intermedias (Rosenzweig y Abramsky 1993, Rosenzweig 1995, Rahbek 1997, 1995). Nuestros resultados apoyan la existencia de un pico de riqueza de especies a elevaciones intermedias (1.100 m s.n.m.), particularmente considerando el número total de especies censadas por piso altitudinal (Tabla 1). Considerando el número promedio (Tabla 1) o el número total de especies por hectárea, no hubo diferencias significativas entre la Selva Pedemontana (600 m s.n.m.) y la Selva Montana (1.100 m s.n.m.). De modo similar, Gentry (1992) encontró picos de riqueza de especies entre los 1.000 y los 1.500 m s.n.m. para los bosques andinos, y sugiere que este patrón está explicado por factores históricos que determinan la confluencia de especies de diversos orígenes biogeográficos, causando una superposición de especies en estas elevaciones intermedias. En el caso de las Selvas Montanas del noroeste de Argentina y sur de Bolivia (1.000 - 1.500 m s.n.m.), encontramos que allí confluyen especies más típicas de los bosques montanos (e.g., C. lilloi, J. australis, P. tucumanensis, Sambucus peruviana), que pertenecen a familias de origen Holártico (e.g., Meliaceae, Juglandaceae, Rosaceae y Caprifoliaceae, respectivamente), con especies de origen Gondwánico (e.g., Roupala cataractarum) y tropical (e.g., Clethra scabra, Cinnamomun porphyria, Styrax subargenteus, Miconia molybdea, Trichilia hieronymi). Esta diversidad de orígenes podría ser una respuesta a la variedad de situaciones climáticas que se presentan en el gradiente altitudinal de los Andes, y la confluencia de estas especies en la Selva Montana podría explicar el pico de riqueza encontrado a elevaciones intermedias. La estructura del bosque también varió con la altitud: el área basal y la densidad tendieron a aumentar, mientras que la altura máxima de los árboles tendió a disminuir con incrementos en la elevación. Estos patrones son similares a los encontrados en otros bosques montanos del Neotrópico (Lieberman et al. 1996, Vázquez y Givnish 1998). En relación a una menor área basal a elevaciones bajas, Vázquez y Givnish (1998) sugieren que esto es una consecuencia de la actividad forestal, que sería más intensa en las partes bajas y planas. De modo similar, Lieberman et al. (1996) explican los bajos valores de área basal encontrados en algunas parcelas con la ocurrencia de disturbios en el pasado, aunque no mencionan una asociación entre área basal y elevación. Según nuestro estudio, el mayor número de tocones encontrados en la Selva Pedemontana y la Selva Montana evidencia una mayor intervención forestal en las elevaciones bajas (Tabla 3). La combinación de un mayor número de especies de interés forestal y la topografía plana o con poca pendiente de la Selva Pedemontana facilitan en términos prácticos y económicos una mayor intensidad de la actividad forestal en el piso inferior de Yungas (Brown et al. 2001). 95 Malizia, L. R., C. Blundo y S. Pacheco Se han desarrollado varias hipótesis para explicar la disminución de la altura del dosel a medida que se asciende por el gradiente altitudinal (revisión de Stevens y Fox 1991). Las condiciones climáticas, las características edáficas y las interacciones bióticas son los principales factores que influyen en la captación de recursos (luz, agua y nutrientes) y en la inversión de estos recursos en crecimiento aéreo o subterráneo. Por un lado, las bajas temperaturas (heladas y nevadas), el viento y los suelos con bajo contenido de agua y/o nutrientes, propiciarían menor crecimiento aéreo para enfrentar las condiciones climáticas adversas, y mayor crecimiento subterráneo para facilitar la absorción de nutrientes en las raíces. Por otro lado, las altas temperaturas y la competencia por luz, propiciarían un mayor crecimiento aéreo para aumentar la captación de luz y dióxido de carbono, limitando el desarrollo del sistema radicular y el crecimiento subterráneo. En el área de estudio, las altas temperaturas y los suelos profundos de la parte baja del gradiente propiciarían el crecimiento en alto de los árboles, mientras que la disminución en temperatura y los vientos más intensos de la parte alta del gradiente serían responsables de la menor altura de los árboles. Las dos especies de cedro estudiadas se segregan espacialmente a lo largo del gradiente altitudinal de las selvas de montaña del noroeste de Argentina y sur de Bolivia. C. balansae ocupa la parte baja del gradiente, mientras que C. lilloi ocupa los sectores más elevados, y ambas especies se presentan en simpatría en el sector basal de las Selvas Montanas (principalmen96 te entre los 1.000 y 1.100 m s.n.m.). Ambas especies ocupan una superficie similar de aproximadamente 9.000 km2 en el área de estudio, calculada a partir de modelos de distribución de GARP que sintetizan el nicho ecológico observado de las especies. Es posible que en nuestros muestreos de campo, alguno de los ejemplares censados de C. balansae correspondan a C. saltensis, una nueva especie para la ciencia descripta recientemente (Zapater et al. 2004) y que es muy afín en su morfología foliar y aspecto general a C. balansae (Grau et al. este volumen). De modo interesante, la distribución de C. saltensis (Grau et al. este volumen) coincidiría en gran medida con el área de simpatría de C. balansae y C. lilloi modelada en este trabajo con GARP (Fig. 5). Este hecho sugiere un origen híbrido para C. saltensis, que debería ser evaluado con estudios filogenéticos para las especies de Cedrela en la región. Los modelos de nicho ecológico como GARP, si bien resultan extremadamente útiles para describir la distribución de las especies a escala regional, no brindan información sobre abundancia relativa de las especies, ni especifican cuales de las variables ambientales consideradas tienen un mayor peso relativo. Futuros estudios en esta dirección serán esenciales para entender mejor la relación entre distribución espacial, abundancia relativa e influencia de los factores ambientales. Estos datos, combinados con datos estructurales como densidad y área basal, permitirán mejorar las estrategias de conservación y uso sus- Diversidad, estructura y distribución de bosques con cedro en la Alta Cuenca del Río Bermejo tentable de las especies de cedro (y otras) a nivel regional. Asimismo, información sobre el efecto de las variables ambientales en la distribución y abundancia de las especies permitirá evaluar los impactos del cambio climático en la diversidad biológica de la región. 1.100 m s.n.m., con un pico en área basal en la parte baja de su distribución (Tabla 4). Además, las tasas de crecimiento de esta especie son probablemente mayores que las de C. lilloi, al encontrarse a menor elevación donde las temperaturas anuales son mayores. En nuestro estudio, no encontramos diferencias significativas en los parámetros estructurales medidos entre parcelas establecidas dentro de áreas protegidas y en fincas privadas. Esto probablemente se debió a que nuestras parcelas de muestreo estuvieron ubicadas en sitios de bosque relativamente bien conservados, tanto en áreas protegidas como en áreas privadas, por lo que esperábamos no encontrar diferencias marcadas entre estas áreas. Las actividades forestales pueden provocar cambios significativos en la dinámica y estructura de las poblaciones de árboles, especialmente para aquellas especies con alto valor forestal como los cedros. Tradicionalmente, estas especies han sido taladas indiscriminadamente sin planes de manejo que contemplen la sustentabilidad económica y ambiental de esta actividad, incluso en el corto plazo (Grau y Brown 2000). Las consecuencias directas de esto se relacionan con una disminución sistemática en los volúmenes aprovechables de madera, la pérdida de árboles semilleros y consecuente pérdida de variabilidad y calidad genética, la desaparición de árboles de gran porte que almacenan información climática de siglos (e.g., 300-500 años) en sus anillos de crecimiento y la degradación del bosque en general (incluyendo la fauna asociada) por uso de técnicas de aprovechamiento inadecuadas. Es de esperar que la generación de información de base sobre las especies arbóreas de las Yungas, junto con programas que incentiven el aprovechamiento sustentable de los recursos forestales y un rol más comprometido del Estado en su tarea de regulador del uso de los recursos naturales, permitan mejorar el uso y la conservación de las especies de cedro en el noroeste de Argentina y sur de Bolivia. En última instancia, esto debe- C. lilloi es probablemente la especie maderable más atractiva en términos económicos de las Yungas de la Alta Cuenca del Río Bermejo, ya que es relativamente común (Tabla 3 y Fig. 4) y tiene un alto valor de mercado (US$ 170 - 300/m3 en el mercado interno Argentina, Noviembre 2005). Como contraparte, el acceso a muchos de los sitios donde se encuentra puede resultar complicado, ya que se distribuye principalmente entre los 1.000 y 2.100 m s.n.m., con un pico en área basal aproximadamente a los 1.500 m s.n.m. (Tabla 4). C. balansae es también una especie maderable importante, aunque con un precio de mercado menor a C. lilloi. Aunque esta especie fue mucho menos abundante en el área de estudio (Tabla 3 y Fig. 4), sus sitios de distribución son generalmente más accesibles, ya que se distribuye principalmente entre los 500 y los 97 Malizia, L. R., C. Blundo y S. Pacheco ría traducirse en una mejora sustentable y equitativa de la calidad de vida de los habitantes de la región. AGRADECIMIENTOS Agradecemos a los que colaboraron en el establecimiento de las parcelas: Claudio Aguirre, Eneas Toranzo, María José Tulli, Erica Cuyckens, Gabriela Quintana, Uriel Colina y Rolando Guevara. El apoyo financiero para realizar este estudio estuvo provisto por la National Science Foundation (Investigador principal: John G. Blake), el International Center for Tropical Ecology (University of Missouri-St. Louis), la Fundación ProYungas e IdeaWild. BIBLIOGRAFIA Anderson, R.; D. Lew, y A. Peterson. 2003. Evaluating predictive models of species’ distributions: criteria for selecting optimal models. Ecological Modelling 162:211232. Beaman, J.H., y R.S. Beaman. 1990. Diversity and distribution patterns in the Flora of Mount Kinabatu. En: Baas, P.; K. Kalkman y R. Geesink (Eds.). The Plant Diversity of Malesia. Kluwer Academic Publishers, Dordrecht. pp: 147-160. Brown, A.D. 1986. Autoecología de bromeliaceas epifitas del noroeste argentino y su relación con Cebus apella. Tesis de doctorado, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, Buenos Aires. Brown, A.D. 1995. Fitogeografía y conservación de las selvas de montaña del noroeste de Argentina. En: Churchill, S.P.; H. Baslev; E. Forero y J.L. Luteyn (Eds.). Biodiversity and conservation of Neotropical Montane Forest. The New York Botanical Garden, New York. pp: 663-672. Brown, A.D. y E.D. Ramadori. 1989. Patrón de distribución, diversidad y características ecológicas de especies arbóreas de las selvas y bosques montanos del noroeste de la Argentina. Anales VI Congreso Forestal Argentino. pp: 177-181. Brown, A.D.; S.C. Chalukian y L.M. Malmierca. 1985. Estudio florístico-estructural de un sector de selva semidecidua del noroeste argentino. I. Composición florística, densidad y diversidad. Darwiniana 26: 27-41. Brown, A.D.; H.R. Grau; L.R. Malizia y A. Grau. 2001. Argentina. En: Kappelle, M. y A.D. Brown (Eds.). Bosques nublados del neotrópico. Instituto Nacional de Biodiversidad, Santo Domingo de Heredia, Costa Rica. pp: 623-659. 98 Diversidad, estructura y distribución de bosques con cedro en la Alta Cuenca del Río Bermejo Chong, D.; S. Mougin y J. Gastellu-Etchegorry. 1993. Relating the global vegetation index to net primary productivity and actual evapotranspiration over Africa. International Journal of Remote Sensing 14: 1517-1546. Colwell, R. K. 2000. EstimateS: Statistical estimation of species richness and shared species from samples. Versión 5. http://viceroy.eeb.uconn.edu/estimates. Condit, R. 1998. Field guide for tropical forest census plots: methods and results from Barro Colorado Island, Panama and a comparison with other plots. SpringerVerlag, Berlin. Elith, J. y M. Burgman. 2002. Predictions and their validation: rare plants in the Central Highlands, Victoria, Australia. En: Scott, J.; P. Heglund; M. Morrison; J. Haufler; M. Raphael; W. Wall y F. Samson (Eds.). Predicting species occurrences: issues of accuracy and scale. Island Press, Washington, DC. pp. 303-313. Gentry, A.H. 1988. Changes in plant community diversity and floristic composition on environmental and geographical gradients. Annals of Missouri Botanical Garden 75: 1-34. Gentry, A.H. 1995. Tropical Forest Biodiversity: distributional patterns and their conservational significance. Oikos 63: 19-28. Grau, A. y A.D. Brown. 2000. Development threats to biodiversity and opportunities for conservation in the mountain ranges of the Upper Bermejo River Basin, NW Argentina and SW Bolivia. Ambio 29: 445-450. Grau, A.; M.A. Zapater y R.A. Neumann. 2006. Botánica y distribución del género Cedrela en el Noroeste de Argentina. En: Pacheco, S. y A.D. Brown (Eds.). Ecología y producción de los cedros (género Cedrela) en las Yungas australes. pp.1930. Grau, H.R. 1999. Disturbances and tree species diversity along the elevational gradient of a subtropical montane forest of NW Argentina. Tesis de Doctorado, Departamento de Geografía, University of Colorado at Boulder, USA. Grau, H.R. y A.D. Brown. 1995. Patterns of tree species diversity along latitudinal and altitudinal gradients in the Argentinian Subtropical montane Forest. En: Churchill, S.P.; H. Baslev; E. Forero y J.L. Luteyn (Eds.). Biodiversity and conservation of Neotropical Montane Forest. The New York Botanical Garden, New York. pp: 295-300. 99 Malizia, L. R., C. Blundo y S. Pacheco Grau, H.R.; M.F. Arturi; A.D Brown y P.G. Aceñolaza. 1997. Floristic and structural patterns along a chronosequence of secondary forest succession in Argentinian subtropical montane forest. Forest Ecology and Management 95: 161-171. Grau, H.R. y A.D. Brown. 1998. Structure, composition and inferred dynamics of a subtropical Montane Forest of northwest Argentina. En: Dallmeier, F. y J.A. Comiskey (Eds.). Forest biodiversity in north, central and south America and the Caribbean. Man and the Biosphere Series, Vol. 21. UNESCO y The Parthenon Publishing Group. pp: 715-726. Grubb, P.J. 1974. Factors controlling the distribution of forest-types on tropical mountains: new facts and a new perspective. En: Flenley, J.R. (Ed.). Altitudinal Zonation in Malesia. Third Aberdeen-Hull Symposium on Malesia Ecology, University of Aberdeen. pp: 13-46. Grubb, P.J. y P.F. Stevens. 1985. The forest of the Fatima Basin and Mt. Kerigomna and a review of Montane and Sub-alpine Forest elsewhere in Papua New Guinea. Departamento de Biogeografía y Geomorfologia, Australian National University, Canberra, Australia. Hijmans R.J.; S. Cameron y J. Parra. 2004. WorldClim, versión 1.2. A square kilometer resolution database of global terrestrial surface climate. http://biogeo.berkeley. edu/ Kitayima, K. 1992. An altitudinal transect study of the vegetation of Mount Kinabatu, Borneo. Vegetatio 102: 149-171. Levine, R.; A. Peterson y M. Benedict. 2004. Geographic and ecological distributions of the Anopheles gambiae complex predicted using a genetic algorithm. Am. J. Trop. Med. Hyg. 70:105-109. Lieberman, D.; M. Lieberman; R. Peralta y G.S. Hartshorn. 1996. Tropical forest structure and composition on a large scale altitudinal gradient in Costa Rica. Journal of Ecology 84: 137-152. Lillesand T. y R. Keifer. 2000. Remote sensing and image interpretation, 4 edition. Wiley. McCune, B. y M. Mefford. 1999. PC-ORD: multivariate analysis of ecological data, versión 4.01. MjM Software Design. Morales, J.M.; J. Sirombra y A.D. Brown. 1995. Riqueza de árboles en las Yungas argentinas. En: Brown, A.D. y H.R. Grau (Eds.). Investigación, conservación y desarrollo en Selvas Subtropicales de Montaña. Laboratorio de Investigaciones 100 Diversidad, estructura y distribución de bosques con cedro en la Alta Cuenca del Río Bermejo Ecológicas de las Yungas (UNT) y Proyecto de Desarrollo Agroforestal. Tucumán. pp: 157-162. Parra, J.; C. Graham y J. Freile. 2004. Evaluating alternative data sets for ecological niche models of birds in the Andes. Ecography 27:350-360. Proctor, J.; Y.F. Lee; A.M. Langley; W.R.C. Munro y F.M. Robertson. 1988. Ecological studies on Gunung Silam, a small ultrabasic mountain in Sabah. I. Environment, forest structure and floristics. Journal of Ecology 76: 320-340. Rahbek, C. 1995. The elevational gradient of species richness: a uniform pattern? Journal of Biogeography 18: 200-205. Rahbek, C. 1997. The relationship among area, elevation and regional species richness in neotropical birds. American Naturalist 149: 876-902. Richards, P.W. 1952. The Tropical Rain Forest. Cambridge University Press, London. Rosenzwerg, M.L. y Z. Abransky. 1993. How are diversity and productivity related? En: Ricklefs, R. y D. Schlater (Eds.). Species diversity in ecological communities: historical and geographical perspectives. University of Chicago Press, Chicago. pp: 52-65. Rosenzweig, M. L. 1995. Species diversity in space and time. Cambridge University Press, New York. Stevens, G.C. y J.F. Fox. 1991. The causes of treeline. Annual Review of Ecology and Systematics 22: 177-191. Stockwell, D. y I. Noble. 1992. Induction of sets of rules from animal distribution data: a robust and informative method of data analysis. Math. Comput. Simul. 33: 385390. Stockwell, D. y D. Peters. 1999. The GARP modelling system: problems and solutions to automated spatial prediction. Int. J. Geogr. Inf. Sci. 13: 143-158. Vázquez, G. y T.J. Givnish. 1998. Altitudinal gradient in tropical forest composition, structure and diversity in the Sierra de Manantlán. Journal of Ecology 86: 9991020. Zapater, M.A.; E.M. del Castillo y T.D. Pennington. 2004. El género Cedrela (Meliaceae) en la Argentina. Darwiniana 42: 347-356. 101 102 533 420 434 468 439 460 638 396 462 469 637 402 416 355 426 Tabacal Yuchán Abra Grande Candado Chico San Martín Valle Morado 1 Aibal Valle Morado 2 Maroma Sidras Casabindo Fortuna Hondura Mesada Indiv. por ha Tecpetrol Sitio 36 31 37 36 46 40 38 36 50 28 34 48 30 43 43 Especies por ha 1200 1150 1100 1100 1100 1000 950 750 650 600 600 550 500 500 500 Elevación (m) 23° 42.0’ 24° 11.7’ 23° 59.6’ 23° 5.1’ 22° 14.8’ 23° 5.6’ 23° 26.2’ 24° 0.3’ 23° 26.5’ 23° 46.2’ 22° 37.6’ 23° 7.3’ 23° 56.4’ 23° 13.3 22° 55.9’ Lat. S (grados) 64° 52.1’ 65° 8.4’ 65° 3.9’ 64° 44.8’ 64° 33.4’ 64° 44.5’ 64° 40.0’ 64° 56.2’ 64° 31.2’ 64° 48.3’ 64° 26.7’ 64° 28.3’ 64° 54.3’ 64° 27.7’ 64° 27.0’ Long. O (grados) Parque Nacional Calilegua, Jujuy (AP) Finca Forestal Norteña, Palpalá, Jujuy (PP) Finca El Aibal, Fraile Pintado, Jujuy (PP) Finca San Andrés, Orán, Salta (PP) Reserva Nacional Tariquía, Tarija, Bolivia (AP) Finca San Andrés, Orán, Salta (PP) Finca Urundel, Urundel, Salta (PP) Finca El Aibal, Fraile Pintado, Jujuy (PP) Finca Urundel, Urundel, Salta (PP) Ingenio Ledesma, L. G. San Martín, Jujuy (PP) Finca Candado Chico, Orán, Salta (PP) Finca Abra Grande, Orán, Salta (PP) Ingenio Ledesma, L. G. San Martín, Jujuy (PP) Ingenio Tabacal, Orán, Salta (PP) Finca Abra Grande, Orán, Salta (PP) Localidad y sur de Bolivia), con datos del número de individuos y número de especies por hectárea, y ubicación de las parcelas, incluyendo elevación, coordenadas geográficas en su punto de inicio y localidad (propiedad privada [PP], área protegida [AP]). ApEndice: Lista de parcelas permanentes de muestreo en las Yungas de la Alta Cuenca del Río Bermejo (noroeste de Argentina Malizia, L. R., C. Blundo y S. Pacheco 426 499 655 448 559 496 699 629 530 727 629 511 675 460 208 348 Lipeo Loma Chata Toro Overo Zapla 1 Escaleras Nogalar 1 Baritú Monolito Aparicio Baritú Ocloyas Tablada Zapla 2 Pino Hachado Nogalar 2 Indiv. por ha Mesada Sitio Continuación ApEndice 12 14 15 4 16 19 20 27 32 19 31 20 33 36 53 36 Especies por ha 2200 2200 2050 2050 2000 2000 1950 1700 1700 1650 1650 1600 1550 1500 1200 1200 Elevación (m) 22° 16.3’ 23° 37.6’ 24° 14.1’ 23° 5.2’ 23° 58.2’ 22° 27.8’ 23° 3.6’ 23° 40.9’ 22° 27.7’ 22° 16.7’ 24° 7.4’ 24° 14.2’ 23° 4.2’ 23° 5.1’ 22° 26.1’ 23° 42.0’ Lat. S (grados) 64° 45.1’ 64° 55.8’ 65° 3.7’ 64° 50.9’ 65° 16.8’ 64° 44.3’ 64° 51.1’ 64° 54.2’ 64° 44.5’ 64° 43.5’ 65° 5.7’ 65° 4.5’ 64° 47.1’ 64° 48.0’ 64° 43.9’ 64° 52.1’ Long. O (grados) Reserva El Nogalar, Los Toldos, Salta (AP) San Francisco, Jujuy (PP) Serranía de Zapla, Palpalá, Jujuy (PP) Finca San Andrés, Orán, Salta (PP) Finca El Duraznal, Ocloyas, Jujuy (PP) Parque Nacional Baritú, Salta (AP) Finca San Andrés, Orán, Salta (PP) Parque Nacional Calilegua, Jujuy (AP) Parque Nacional Baritú, Salta (AP) Reserva El Nogalar, Los Toldos, Salta (AP) Las Escaleras, Jujuy (PP) Serranía de Zapla, Palpalá, Jujuy (PP) Finca San Andrés, Orán, Salta (PP) Finca San Andrés, Orán, Salta (PP) Parque Nacional Baritú, Salta (AP) Parque Nacional Calilegua, Jujuy (AP) Localidad Diversidad, estructura y distribución de bosques con cedro en la Alta Cuenca del Río Bermejo 103 Modelos de crecimiento de yCedrela lilloideen el sector de las Yungas argentinas Ecología producción cedro (géneronorte Cedrela) en las Yungas australes. Pacheco, S. y A. Brown (Eds.). LIEY-ProYungas. Argentina. pp. 105-116. 2006. Modelos de crecimiento de Cedrela lilloi en el sector norte de las Yungas argentinas Gasparri, N. I 1,2 y Goya, J. F. 2 Laboratorio de Investigaciones Ecológicas de las Yungas (LIEY, UNT), CC 34 (4107), Yerba Buena, Tucumán, Argentina. 2 Cátedra de Ordenación Forestal. Facultad de Ciencias Agrarias y Forestales (UNLP) 1 [email protected] La degradación de los bosques con presencia de cedros, es un problema ambiental y económico causado principalmente por la corta selectiva sin planificación. La sostenibilidad de la producción maderera se logra extrayendo volúmenes que no excedan la capacidad productiva del bosque. Para esto es necesario contar con herramientas de planificación como las funciones de crecimiento. Este trabajo tiene por objetivo explorar la relación de distintas variables sobre el crecimiento de individuos de Cedrela lilloi y ajustar una función de crecimiento individual. Para caracterizar la estructura del bosque, se instalaron 14 parcelas circulares concéntricas de 500 y 1.000 m2 en el norte de Salta, cerca de la localidad de San Andrés. De los individuos de cedro incluidos en las parcelas, se obtuvieron muestras de barreno para estudiar el crecimiento. Además, se tomaron medidas de tamaño (altura y diámetro a la altura del pecho), iluminación de copa y competencia. Se realizaron pruebas de correlación para evaluar la relación de las distintas variables sobre el crecimiento. El diámetro a la altura del pecho y el índice de iluminación de la copa resultaron ser las variables con mayor relación. Con estas dos variables se ajustó una función derivada del modelo de Chapmand y Richards para predecir el incremento en área basal. La función seleccionada utiliza como variable independiente mediciones rutinarias en inventarios forestales y puede ser aplicada de manera sencilla en la estimación del crecimiento de Cedrela lilloi. Selective logging of Cedrela spp. causes forest degradation and generates environmental and economical problems. Sustainable logging can be achieved by extracting volumes that do not exceed forest production capacity. Knowing growth variables can be an important tool for planning logging activities. The goal of this work is to explore relationships between different individual growth variables in Cedrela lilloi, adjust and individual growth function. To describe forest structure, 14 concentric circular plots (500 y 1.000 m2) were placed in forests located in northern Salta, nearby the locality of San Andrés. Increment core samples were extracted for each Cedrela individual to estimate growth. We also measured size (height and DBH), crown illumination and competition. Correlation tests were performed to evaluate the relationship between different variables and growth. DBH and crown illumination showed the highest correlation with growth. Using these variables, we adjusted a derived function from Chapmand and Richards model to predict basal area increase. The selected function uses as independent variables measurements that are routine in forest surveys and can be easily used for estimating growth in Cedrela lilloi. 105 Gasparri, N. I. y J. F. Goya introduccion La degradación de los bosques tropicales y subtropicales es un proceso de pérdida gradual de cobertura que muchas veces es acompañado por una pérdida del valor comercial. Comúnmente, en aquellos sitios con aptitud para la agricultura, la degradación es seguida por la deforestación. Una de las principales causas de la degradación es la corta selectiva de individuos comerciales sin planificación (Johons 1992). Esto produce una disminución de los volúmenes maderables y provoca daños en la estructura remanente que comprometen la recuperación del bosque. Una de las premisas de la ordenación forestal para lograr la producción sostenida a perpetuidad, es que la extracción no supere el crecimiento del bosque. Por lo tanto, para determinar la capacidad de producción de un bosque (posibilidad) y realizar la planificación, son indispensables los inventarios forestales y el desarrollo de los modelos de crecimiento. En los bosques tropicales, que presentan cierta complejidad estructural y donde generalmente se dispone de escasa información, modelos sencillos de crecimiento pueden representar un avance importante para la planificación y para demostrar la efectividad de las medidas aplicadas o la necesidad de modificar alguna de las prácticas de manejo (Vanclay 1995). Los modelos de crecimiento individual son fórmulas matemáticas que permiten estimar el incremento de un árbol durante un periodo de tiempo, o el tamaño de un individuo. Estos modelos se elaboran en función de la edad, del tamaño (diámetro a la altura del pecho y/o altura) o 106 de la competencia (área basal del rodal o iluminación de la copa). En plantaciones forestales donde se conoce la fecha de implantación con exactitud, es adecuado utilizar la edad del árbol como variable independiente. Sin embargo, en los bosques nativos resulta difícil conocer la edad de los árboles ya que muchas especies no marcan anillos anuales de crecimiento. En este caso, es preferible aplicar una fórmula que no utilice la edad, como por ejemplo la función de von Bertalanffy o de Chapman-Richards en su forma derivada, la cual tiene la virtud de poseer una base teórica y, por lo tanto, significado biológico (Richards 1959, Vanclay 1995). En los bosques nativos del noroeste de Argentina, se realizan cortas selectivas respetando un diámetro mínimo legislado por cada provincia y para cada especie. Las intervenciones no son acompañadas por tratamientos silvícolas que garanticen la recuperación de los bosques. El diámetro mínimo de corta en ocasiones tampoco es efectivo en su propósito de conservar recursos del bosque en el mediano y largo plazo (Gasparri et al. 2003). En general, el aprovechamiento tradicional representa una reducción drástica de las existencias comerciales y, si bien los bosques recuperan en parte la cobertura arbórea, la capacidad productiva queda comprometida varios años después (Pinazo y Gasparri 2003). Para avanzar en el uso planificado de los bosques nativos, es necesario contar con modelos de crecimiento individual de las principales especies comerciales como Cedrela lilloi y Juglans australis que además, presentan cualidades que las Modelos de crecimiento de Cedrela lilloi en el sector norte de las Yungas argentinas hacen aptas para los estudios dendrocronológicos (Villalba et al. 1985). El objetivo de este trabajo fue explorar variables que puedan explicar el crecimiento de los individuos de Cedrela lilloi y ajustar una fórmula de crecimiento individual para dicha especie. metodos El muestreo se realizó en el valle del Río Querusillar, en las proximidades de la localidad de San Andrés en el norte de la Provincia de Salta. Los sitios identificados a priori con Cedrela lilloi, fueron recorridos de manera sistemática respetando un rumbo fijo hasta encontrar un árbol adecuado para la muestra. Se procuró cubrir todo el rango de diámetros a la altura del pecho (DAP). En cada árbol seleccionado se establecieron dos parcelas circulares concéntricas donde se determinó la especie y se midió el DAP de todos los individuos. En la parcela mayor de 1.000 m² se relevaron los árboles mayores a 20 cm de DAP y en la subparcela de 500 m² incluida en la anterior, se relevaron los árboles con DAP entre 10 y 20 cm. Dentro de las parcelas se registró la pendiente máxima, la posición topográfica (filo, media loma alta, media loma, media loma baja y quebrada) y la exposición general y puntual. Para todos los individuos de cedro incluidos en las parcelas se tomaron muestras de barreno a la altura del pecho siguiendo la dirección de la pendiente. De cada individuo se cuantificaron las siguientes variables: diámetro a la altura del pecho, dos diámetros de copa en forma perpendicular (para lo cual se estimó la proyección de la copa en el plano), altura total y del fuste (con clinómetro), sanidad (buena, regular y mala) y el índice de iluminación de la copa (IIC) propuesto por Alder y Synott (1992). Este índice es una forma de estimar la disponibilidad de luz de los árboles y en su versión original presenta cinco valores: 1- Sin iluminación directa: la copa del árbol no recibe luz directa en ninguna de sus partes. 2- Iluminación directa lateral: copa no expuesta a luz directa de forma vertical, pero sí lateral. 3- Iluminación directa vertical incompleta: copa con algo de iluminación directa vertical (entre el 10 y el 90%). 4- Iluminación directa vertical: copa con iluminación directa vertical total (más del 90 %), pero sin iluminación directa, al menos en uno de los laterales. 5- Emergentes: copa completamente expuesta a iluminación directa. En total fueron tomados como muestra 59 árboles de cedro distribuidos en 14 parcelas. Las muestras de barreno fueron acondicionadas mediante técnicas dendrocronológicas estándares (Stokes y Smiley 1968) y fueron medidas con lupa binocular utilizando platina de precisión. También se midió el grosor de la corteza para luego calcular los incrementos con corteza. En base a las muestras de barreno, se determinó el crecimiento de los 10 últimos años y con ellos se calculó el incremento acumulado (periodo 1989-1999) y el incremento periódico anual en diámetro 107 Gasparri, N. I. y J. F. Goya (IPA, promedio del periodo 1989 - 1999). Sólo se tomaron los últimos 10 años asumiendo que las condiciones ambientales se mantuvieron relativamente estables en este periodo. Se realizaron pruebas de correlación para identificar las variables que se asocian con el incremento. Las variables que intervinieron en las pruebas de correlación fueron las siguientes: DAP, índice de iluminación de copa, superficie de copa, altura total, competencia (calculada como área basal de individuos con DAP ≥ 20 cm y área basal de individuos con DAP mayor al del árbol considerado) y disponibilidad de humedad (fue codificada con valores ordinales que asignaban mayores valores a las situaciones definidas a priori como más favorables para el crecimiento. Las exposiciones más húmedas recibieron mayores valores que las secas y las posiciones de quebrada mayores que los filos). Finalmente, se ajustó una función como propuesta de un modelo matemático para predecir el crecimiento individual de cedro. La función seleccionada (Vanclay 1994, Monserud y Sterba 1996) en su forma general se presenta como la fórmula a) a) ln ΔY = a + b ln Y + cY2 + d (vc1; vc2...vci) + e (va1; va2...vai) + ε donde: ΔY : incremento de la variable de interés vc1; vc2...vc i : variables que reflejan la competencia va1; va2...vai : variables ambientales a; b; c; d y e : parámetros del modelo ε : error de la muestra 108 Esta función permite predecir el incremento de un individuo en función del tamaño y de otras variables que modifican la predicción como por ejemplo, la competencia o las condiciones ambientales. En este caso se decidió utilizar como variable dependiente el logaritmo natural del incremento en área basal del individuo y como variables independientes se forzó la inclusión de variables con mayor correlación y de fácil medición en los inventarios forestales de rutina (DAP e índice de iluminación de la copa). De esta manera, se pretende contar con un modelo de crecimiento que utiliza variables simples de medir en los inventarios forestales de rutina, evitando otras como por ejemplo la edad o el diámetro de la copa. resultados La situación más frecuente de iluminación de la copa fue la de valor 4 (61 %) que corresponde a iluminación directa vertical, pero con algún tipo de competencia lateral. Como segunda situación más frecuente (15 %) se encuentra la iluminación directa total que corresponde al valor 5. Las categorías correspondientes a situaciones de iluminación regular o mala (valores entre 1 y 3) reunieron en conjunto el 24 % de los árboles. Con el muestreo se cubrió un rango de tamaños entre los 10 y 120 cm de DAP. En la Tabla 1 se presentan los datos de tipo ambiental y estructural de las parcelas, que pueden ser tomados como referencias de bosques maduros de cedro en el área de trabajo. Estos datos son coincidentes con estructuras maduras de otros Bosques Modelos de crecimiento de Cedrela lilloi en el sector norte de las Yungas argentinas Tabla 1. Datos ambientales y estructurales de las parcelas donde se tomaron las muestras de cedro. Area basal total (ABT) y área basal de cedro (ABC) expresadas en m²/ha; densidad total (DT) y densidad de cedro (DC) expresada en individuos/ha; pendiente expresada en grados. Parcela ABT ABC DT DC Exposición general Exposición puntual Topografía 1 9,04 1,44 160 10 Sur Suroeste Quebrada 40 2 10,54 6,42 160 50 Sur Sur Quebrada 57 3 9,95 2,04 190 20 Sur Oeste Media loma alta 35 4 7,13 1,40 170 30 Sur Suroeste Media loma alta 35 5 24,85 2,65 210 30 Este Este Media loma baja 10 6 18,97 1,81 160 40 Este Este Media loma baja 38 7 20,08 3,16 140 30 Este Sur Media loma baja 25 8 34,70 8,18 240 20 Este Sureste Media loma 40 9 46,33 7,02 460 50 Este Sureste Filo 30 10 36,92 12,65 280 100 Sur Sureste Media loma alta 30 11 24,70 11,15 270 60 Este Sureste Media loma alta 35 12 32,76 11,12 170 10 Sur Este Media loma 30 13 18,44 6,36 250 80 Sur Sur Media loma alta 39 14 33,57 7,85 470 90 Este Este Media loma alta 44 Pendiente 109 Gasparri, N. I. y J. F. Goya Montanos maduros (Brown et al. 1985, Grau y Brown 1998, Pinazo et al. 2003). En la Tabla 2 se presentan los valores de incrementos periódicos anuales por clase diamétrica para Cedrela lilloi. Los mayores crecimientos se midieron para las categorías de tamaño intermedias, entre los 30 y 40 cm de DAP. Las pruebas de correlación (Sperman) mostraron que las variables relacionadas con el tamaño y la iluminación del individuo son las de mayor influencia al momento de explicar el crecimiento de los árboles. Las variables que presentaron una correlación estadísticamente significativa con el incremento periódico anual fueron el DAP (R2 = 0,311), el índice de iluminación de la copa (R2 = 0,329), la superficie de la copa (R2 = 0,320) y la altura total (R2 = 0,328). En cuanto a las variables que reflejan la competencia, no se encontraron correlaciones estadísticamente significativas. El valor de mayor correlación corresponde al área basal de los árboles cuyo DAP es mayor al del individuo considerado (R2 = - 0,129). En cuanto a las variables de tipo ambiental, con las muestras disponibles no se encontraron correlaciones estadísticamente significativas con el crecimiento. La función ajustada de crecimiento se presenta en la fórmula b). b) ln IAB = 10.722 + 1.1137 ln (DAP) 0.0001 (DAP2) + 0.3221(IIC) donde: ln IAB: logaritmo natural del incremento en área basal (m2) ln (DAP): logaritmo natural del diámetro a la altura del pecho (cm) IIC: índice de iluminación de la copa La fórmula presentó estadísticos acep- Tabla 2. Incremento periódico anual (IPA, 1989 - 1999) expresado en incremento diamétrico (cm/año) de árboles de cedro por clase diamétrica. Los árboles se asignaron a la clase diamétrica en función de su valor al inicio del periodo (1989); n: número de muestras de cada clase diamétrica. 110 Clase diamétrica IPA promedio IPA máximo IPA mínimo n < 10 0,707 1,535 0,311 5 10 a 20 0,605 1,245 0,161 20 20 a 30 0,646 0,905 0,415 11 30 a 40 0,996 1,999 0,340 11 40 a 50 0,830 1,660 0,351 4 50 a 60 0,794 1,335 0,461 3 60 a 70 0,577 0,699 0,454 2 > 70 0,472 0,737 0,258 3 Modelos de crecimiento de Cedrela lilloi en el sector norte de las Yungas argentinas tables y comparables con otros modelos similares desarrollados para especies de otros ecosistemas (Favrichon 1998). Los estadísticos pueden ser consultados de manera detallada en el Anexo I. Con esta fórmula se construyeron curvas tipo para individuos bajo distintas condiciones de iluminación. En la Figura 1 se observa el incremento en área basal que corresponde a la fórmula ajustada y en la Figura 2 se construyeron las curvas de incremento en DAP, a partir de los resultados obtenidos con la fórmula b). Las curvas muestran que cuanto mayor es la iluminación de la copa mayor es el crecimiento y que en el rango de 40 y 60 cm de DAP se producen los mayores incrementos en área basal. Figura 1. Incremento anual del área basal de árboles de Cedrela lilloi en función del DAP y la iluminación de copa (IIC). 1- Sin iluminación directa, 2- Iluminación directa lateral, 3- Iluminación directa vertical incompleta, 4- Iluminación directa vertical, 5- Emergentes. Figura 2. Incremento anual del diámetro a la altura del pecho de árboles de Cedrela lilloi en función del DAP y la iluminación de la copa (IIC). 1- Sin iluminación directa, 2- Iluminación directa lateral, 3- Iluminación directa vertical incompleta, 4- Iluminación directa vertical, 5- Emergentes. 111 Gasparri, N. I. y J. F. Goya Discusion La relación entre el tamaño y el crecimiento observada en el cedro es consistente con la base teórica del modelo de Chapmand y Richard (Richards 1959). El mismo expresa que el aumento de tamaño proporciona mayor capacidad de captar recursos (por lo tanto crecimiento rápido), pero una vez alcanzado un determinado tamaño la captación de recursos se vuelve cada vez más ineficiente, por lo que el ritmo de crecimiento disminuye. Se puede observar que el tamaño tiene influencia sobre el ritmo de crecimiento. Al agrupar los datos por clase diamétrica, los individuos que alcanzan los 30 a 40 cm de DAP, presentan los mayores incrementos en diámetro con valores cercanos a 1 cm/año. Arboles de este tamaño, cuando tienen condiciones de iluminación óptimas, pueden alcanzar hasta 2 cm/año, como lo indica el incremento máximo para esta categoría. Los datos anteriores permiten suponer que los turnos de corta para cedro podrían ser reducidos significativamente con medidas silvícolas que mejoren las condiciones de iluminación de los individuos de interés comercial (casi el 24% de los individuos incluidos en la muestra se encontraban con iluminaciones regulares a malas). Respecto al uso del Indice de Iluminación de Copa (IIC), se puede indicar que la condición identificada como IIC = 4 incluye distintas situaciones de iluminación lateral de la copa que pueden tener influencia sobre el crecimiento. Tal como está utilizado el índice en este trabajo, mejorar la situación de los individuos con IIC = 4 puede ser improcedente en la práctica, ya que 112 proporcionar una situación de iluminación total (IIC = 5) implica eliminar toda la competencia. Sin embargo, es probable que los individuos que tienen tres laterales bloqueados a la luz directa, mejoren su incremento luego de una intervención que libere dos de los laterales bloqueados. Por lo tanto, sería recomendable modificar el índice desglosando la categoría 4 y evaluar la real influencia sobre el incremento de las distintas situaciones de iluminación lateral incluidas en la categoría 4 del índice original. Si bien las variables relacionadas con el tamaño e iluminación de la copa parecen ser apropiadas para la predicción del crecimiento individual, sería importante avanzar en la evaluación de la influencia de las distintas situaciones ambientales y de la competencia. Aumentar la precisión del modelo incorporando índices de competencia y de calidad de sitio es especialmente importante cuando el valor de la madera es alto (como es el caso del cedro), y los márgenes de error en las estimaciones pueden significar importantes recursos económicos. La fórmula seleccionada se comporta bien con los datos de C. lilloi. Las variables que presentaron una mayor correlación y que posteriormente fueron incluidas en la función, son variables que pueden ser relevadas eficientemente en los inventarios forestales de rutina. El índice de iluminación de la copa debería ser incorporado como variable de medición en los inventarios de áreas donde se prevé aplicar esta fórmula. Por otra parte, este índice puede ser de utilidad para estimar las condicio- Modelos de crecimiento de Cedrela lilloi en el sector norte de las Yungas argentinas nes de iluminación de todos los individuos comerciales, lo cual puede ser relevante al momento de orientar la planificación silvícola. Finalmente, la fórmula aquí ajustada corresponde a una muestra limitada. Si se quisiera aplicarla en otras áreas de Bosques Montanos dentro de Argentina, debería hacerse primero una calibración del modelo, para lo cual se pueden seguir las recomendaciones de Vanclay (1994). AGRADECIMIENTOS Este trabajo fue financiado por el programa de subsidios de la Fundación ProYungas. Agradecemos por sus valiosos comentarios a Pablo Yapura, Patricio MacDonagh y Alfredo Grau. Agradecemos por su colaboración en las tareas de campo a Miguel Martín, Sebastián Lorenzati, El pampa y David Monteverde. BIBLIOGRAFIA Alder, D. y T.J. Synott. 1992. Permanent sample plot techniques for mixed tropical forest. Tropical Forestry Papers Nº25, Oxford Forestry Institute. 124 pp. Favrichon, V. 1998. Modeling the dynamics and species composition of tropical mixedspecies uneven-aged natural forest: Effects of alternative cutting regimes. Forest Science 44: 113-124. Brown, A.D.; S.C. Chalukian y L.M. Malmierca. 1985. Estudio florístico estructural de un sector de selva caducifolia del noroeste de Argentina, composición florística, densidad y diversidad. Darwiniana 26: 27-41. Gasparri, N.I.; M.A. Pinazo y J.F. Goya. 2003. Crecimiento individual y diámetro mínimo de corta de Juglans australis: simulación en un rodal maduro en el noroeste de Argentina. Yvyraretá 11: 27-32. Grau, H.R. y A.D. Brown. 1998. Structure, composition, and infrarred dynamics of a subtropical montane forest of northwestern Argentina, En: Dallmeier, F. y J.A. Comiskey (Eds.). Forest biodiversity in north, central and south America, and the Caribbean: research and monitoring. Man and the Biosphere Series. Vol. 21: 715-726. Johons, R.J. 1992. The influence of deforestation and selective logging operations on plant diversity in Papua New Guinea. En: Withmore, T.C. y J.A. Sayers (Eds.). Tropical deforestation and species extinction. Chapman and Hall and the World Conservation Union. 156 pp. Monserud, R.A. y H. Sterba. 1996. A basal area increment model for individual trees growing in uneven-aged forest stands in Austria. Forest Ecology and Management 80: 57-80. 113 Gasparri, N. I. y J. F. Goya Pinazo, M.A. y N.I. Gasparri. 2003. Cambios estructurales causados por el aprovechamiento selectivo en el bosque montano del norte de Salta, Argentina. Ecología Austral 13: 160-172. Pinazo, M.A.; N.I. Gasparri; J.F. Goya y M.F. Arturi. 2003. Caracterización estructural de un bosque de pino del cerro (Podocarpus parlatorei Pilg,) y nogal criollo (Juglans australis Griseb,) en un sector de la Alta Cuenca del Río Bermejo. Revista de Biología Tropical 51: 361-368. Richards, F.J. 1959. A flexible growth function for empirical use. Journal of Experimental. Botany 10: 290-300. Stokes, M.A. y T.L. Smiley. 1968. An introduction to tree-rings dating. University of Chicago Press, Chicago. 73 pp. Vanclay, J.K. 1994. Modeling forest growth and yield: applications to mixed tropical forest. CAB International, Wallingford UK. 312 pp. Vanclay, J.K. 1995. Growth models for tropical forests: a synthesis of models and methods. Forest Science 40: 7-42. Villalba, R.; J.A. Boninsegna y R.L. Holmes. 1985. Cedrela angustifolia and Juglans australis. Two tropical species useful in dendrochronology. Tree-ring Bulletin 45: 25-35. 114 Modelos de crecimiento de Cedrela lilloi en el sector norte de las Yungas argentinas ANEXO I: Estadísticos de la regresión Variable dependiente: Logaritmo Natural del Incremento anual en área basal (m²). Tamaño de la muestra: El número de pares de datos utilizados corresponde al número de años tomados para ajustar la función. Se tomaron los últimos cuatro años presentes en cada muestra. n: 236. Coeficientes de correlación: R= 0,768; R2 = 0,590; R2 ajustado = 0,585 Error estándar de la estimación: 0,592 Análisis de la varianza del modelo: Suma de cuadrados GL Cuadrado medio F P 111,202 0,000 Regresión 117,107 3 39,036 Residual 81,440 232 0,351 Residuales del Modelo ajustado: 115 Ecología y producción de cedro (género Cedrela) en las Yungas australes. Pacheco, S. y A. Brown (Eds.). LIEY-ProYungas. Argentina. pp. 117-130. 2006. Incidencia de la herbivoría en Cedrela balansae Scandalo1,2, R.P.; G.A. Zuleta1,2,3; M.A. Gainza1,2 y A.S. Haedo1 1 Facultad de Ciencias Exactas y Naturales - UBA, Intendente Güiraldes 2160, Ciudad Universitaria (1428 EGA), Cap. Fed., Argentina. 2 Departamento de Ecología y Ciencias Ambientales.CEBBAD. Universidad Maimónides. 3 CONICET. [email protected] La supervivencia y el crecimiento de los renovales de Cedrela balansae son comúnmente afectados por procesos de herbivoría. Los dos grupos que pueden representar una amenaza para la supervivencia son las larvas de lepidópteros (en particular, la mariposita del brote) y las hormigas cortadoras. A fin de determinar las consecuencias a mediano plazo de la herbivoría total y en particular la causada por insectos, se transplantaron 200 renovales de C. balansae, siguiendo un diseño de bloques al azar (DBA) con cuatro tratamientos: 1) exclusión de invertebrados (insecticida), 2) exclusión de vertebrados (clausura), 3) exclusión de ambos (mixto) y 4) sin exclusión (testigo). A los 360 días desde el transplante, se registraron parámetros asociados con el crecimiento, la herbivoría y la supervivencia. Se registró que un mayor crecimiento se encuentra asociado al menor consumo por insectos. La herbivoría por vertebrados fue baja (< 1 %), mientras que el consumo por invertebrados se registró en el 94 % de los individuos. El porcentaje de supervivencia aumentó con el grado de protección (testigo<clausura/insecticida<mixto), siendo la herbivoría por insectos la causa más importante de mortalidad. Survival and growth of Cedrela balansae are commonly affected by herbivory. The most dangerous taxa for sapling survival are lepidopteran larvae (especially Hypsipyla shoot borer) and leaf-cutting ants. In order to determine the middle term effects of herbivory on C. balansae, and specially the ones caused by insects, 200 saplings were transplanted to a native forest stand in the Yungas region. Saplings were subjected to one of four possible experimental treatments in a randomized block design: (1) protection against invertebrates (insecticide), (2) protection against vertebrates (enclosure), (3) protection against invertebrates and vertebrates, and (4) without protection (control). One year after transplantation, parameters related to survival, growth and herbivory were measured in all experimental units. Maximum growth was associated to minimum insect damage. Vertebrate herbivory was low (< 1 %), while invertebrate herbivory was recorded in 94 % of the individuals. Percent survival increased with increased protection (no protection<enclosure/insecticide<total protection), being insect herbivory the main cause of mortality. 117 Scandalo, R. P.; G. A. Zuleta; M. A. Gainza y A. S. Haedo INTRODUCCION La herbivoría es un factor clave en la regulación de la dinámica poblacional de las especies vegetales (Crawley 1997, Veblen et al. 1992, Relva y Veblen 1998). Los diversos grupos de herbívoros consumen distintas partes de las plantas, pudiendo incidir en su crecimiento y supervivencia. El impacto en el desempeño de la planta depende del tipo de tejido afectado, de la intensidad y frecuencia del ataque y de la escala de forrajeo (Honkanen y Haukioja 1994, Fonseca 1994, Newton et al. 1998, Roldán y Simonetti 2001, Wallin y Raffa 2001, Liang y Seagle 2002). En condiciones naturales los efectos de los herbívoros están generalmente acotados por factores regulatorios como la presencia de depredadores, parásitos y patógenos. Sin embargo, en ambientes perturbados como los bosques secundarios y las poblaciones vegetales artificialmente densas como las plantaciones forestales, la incidencia de la herbivoría puede incrementarse de manera considerable (Crawley 1997, Rao et al. 2001). Por ello, es necesario determinar la importancia que presenta la herbivoría en proyectos de reforestación. En este trabajo se analiza el impacto de la herbivoría sobre renovales del cedro orán (Cedrela balansae) en base a una revisión del estado de conocimiento sobre la ecología de los principales grupos de herbívoros de la especie y en base a los resultados de un experimento de reforestación con C. balansae en bosques perturbados de Yungas (Scandalo 2004). Se discuten 118 además las implicancias de la herbivoría para el manejo de plantaciones de esta especie. Riesgos de herbivoría en Cedrela Aunque la información sobre herbivoría en las Yungas es escasa, se sabe que existe una gran diversidad de herbívoros, tanto vertebrados como invertebrados, que pueden tener distintos impactos sobre las plantas de Cedrela (Mata 2001). Entre los invertebrados, aunque hay grupos que se alimentan de variadas estructuras vegetales (semillas, flores, frutos, raíces y tallos), los más importantes en cuanto a abundancia y riqueza de especies son los folívoros (que se alimentan de hojas), como caracoles, babosas y una gran cantidad de especies de insectos (Mata 2001). Se estima que los insectos consumen entre el 5 y el 10 % del área foliar por año (Dirzo 1987, Crawley 1997, Arturi et al. 1999), constituyendo la causa de mayor mortalidad en bosques tropicales (Dirzo y Rodriguez 1995, Crawley 1997, Mata 2001). En este tipo de ecosistemas, el mayor daño foliar en especies leñosas es producido por lepidópteros (Dirzo 1987, Coley 1987, Moubert-Quiros y Arguedas 1993), coleópteros, ortópteros (Coley 1987) y hormigas (Blanton y Ewel 1985, MoubertQuiros y Arguedas 1993). Las larvas de insectos y las hormigas cortadoras pueden representar una amenaza para la supervivencia de los renovales por los patrones de consumo que presentan. Incidencia de la herviboría en Cedrela balansae Riesgo de herbivoría por larvas de insectos Las larvas de lepidópteros y de coleópteros constituyen, por su abundancia y diversidad, uno de los grupos más importantes de folívoros en plantas jóvenes y adultas (Borror et al. 1989, Mata 2001). El consumo de hojas y brotes puede afectar el desempeño de las plantas al reducir el crecimiento, la morfología de la planta y la calidad de los tejidos vegetales (Rodgers et al. 1995). Entre los lepidópteros que atacan a especies de Cedrela se destaca la mariposita del brote, Hypsipyla grandella (Lepidoptera, Pyralidae) que es específico de la subfamilia Swietenioideae. Debido a su ataque sobre el follaje terminal y los meristemas apicales de los renovales, constituye una grave plaga forestal, ya que con apenas una larva por árbol, el daño resulta severo (Floyd y Hauxwell 2001). A pesar del conocimiento detallado sobre el ciclo de vida de H. grandella (Floyd y Hauxwell 2001), los factores que influyen en sus patrones de herbivoría han sido poco estudiados. En estudios realizados en la selva amazónica (Yamazaki et al. 1990), se encontró que la densidad poblacional de H. grandella y los daños causados por las larvas, se incrementan al comienzo de la estación lluviosa, momento que coincide con la aparición de nuevos brotes en los árboles. Patrones similares fueron encontrados en Venezuela (Tillmanns 1964) y Florida (Howard 1991). En contraste con esto, en plantaciones de Costa Rica, se encontró que H. grandella era activa todo el año, y los picos poblacionales coincidían con fluctuaciones térmicas (Taveras et al. 2004). Aparentemente, la interacción HypsipylaMeliáceas está regulada químicamente por la concentración de nutrientes y de compuestos metabólicos secundarios en la planta hospedador (Newton et al. 1998, Floyd y Hauxwell 2001). Esta regulación química tiene una base genética, que determina distinto grado de susceptibilidad al ataque por H. grandella dentro y entre las especies de Meliáceas (Newton et al. 1999). Por ejemplo, se ha encontrado que en variedades de Cedrela odorata con baja susceptibilidad al ataque, la concentración de nutrientes era baja y el contenido de taninos alto (Newton et al. 1999). Asimismo, se ha observado que ejemplares de C. odorata plantados en Australia sufren pocos ataques de H. robusta y que ejemplares de Toona ciliata, plantados en Costa Rica y en Brasil, no sufren el ataque de H. grandella (Bygrave y Bygrave 2001). Con respecto a las condiciones ambientales que favorecen el ataque, estudios realizados en Surinam con C. balansae (Vega 1976) indican que los factores que inciden en la intensidad del ataque son: a) la exposición a la luz (las plantas que fueron plantadas en sitios abiertos fueron atacadas antes que las plantadas en sombra) y b) el tipo de suelo (en los renovales que crecieron en suelos ricos en nutrientes la duración del ataque fue menor que en plantas que crecieron en suelos más pobres). 119 Scandalo, R. P.; G. A. Zuleta; M. A. Gainza y A. S. Haedo Riesgo de herbivoría por hormigas cortadoras Las hormigas cortadoras (Formicidae, Attini) colectan material vegetal que usan como sustrato para cultivar los hongos de los que se alimentan. Estas hormigas, representadas por los géneros Atta y Acromyrmex, son importantes herbívoros en toda la región Neotropical (Holldober y Wilson 1990) y suelen ser consideradas plagas agrícolas y silvícolas. En Argentina se registran tres especies de Atta y once de Acromyrmex, de las cuales al menos tres ocurren en las Yungas (Farji-Brener 1994). Algunas de estas hormigas forrajean sólo dicotiledóneas, otras sólo monocotiledóneas y un tercer grupo tiene una “dieta” mixta (Holldober y Wilson 1990, FarjiBrener 1994). Los estudios realizados sobre la actividad forrajera de las hormigas cortadoras indican que sus tasas de herbivoría son altas en todos los ecosistemas. En bosques tropicales consumen el 17 % de la producción de hojas (Cherrett 1989), y sus patrones de cosecha son selectivos, ya que suelen estar concentrados en algunas pocas especies vegetales (Rockwood 1976, Blanton y Ewel 1985, Vasconcelos 1997, Farji-Brener 2001, Rao et al. 2001, Cortés-Pérez y León-Sicard 2003). Sin embargo, existen grandes variaciones intra e interespecíficas en cuanto a las especies y tipo de material vegetal consumido. La selectividad del forraje depende de diversos factores físicos, químicos y biológicos de las plantas: a) palatabilidad:altos contenidos de agua y nutrientes en las 120 hojas favorecen el consumo; b) defensas anti-herbívoro: la presencia de metabolitos secundarios, taninos, terpenoides y alcaloides, disminuye la selectividad (Hubbel et al. 1984, Pelotto y del Pero de Martínez 2002); c) características físicas de las plantas: la resistencia mecánica o dureza de las hojas reduce el consumo (Stradling 1978, Camargo et al. 2004); d) presencia de hormigas agresivas en la planta: las especies vegetales visitadas con mayor frecuencia por hormigas nectívoras son menos abundantes en la dieta de las hormigas cortadoras (Folgarait et al. 1994, Pelotto y del Pero de Martínez 2002). Dado que las hormigas cortadoras suelen ser más abundantes en bosques secundarios que en bosques primarios (Farji-Brener 2001, Rao et al. 2001) y que se ha encontrado que sus tasas de herbivoría aumentan al reducirse la complejidad estructural y la diversidad de especies del ecosistema (Blanton y Ewel 1985, Folgarait et al. 1994, Vasconcelos 1997), son un grupo potencialmente importante de herbívoros en bosques naturales perturbados y en plantaciones forestales. Sin embargo, resulta difícil predecir el riesgo que representan ya que suelen cambiar su selectividad de acuerdo a la disponibilidad de recursos (Rao et al. 2001, Wetterer et al. 1998). METOdOS Area de estudio Este estudio se llevó a cabo en Valle Morado (64º 30´3,17” O 23º 26´40,12” S, Finca Urundel, Departamento de Orán, Salta, Argentina) en el marco de un proyecto de in- Incidencia de la herviboría en Cedrela balansae vestigación para una tesis de grado (Scandalo 2004). El área de estudio corresponde al sector norte de la Provincia Fitogeográfica de las Yungas (Cabrera 1976, Brown y Ramadori 1989, Brown 1995). La misma está sujeta a distintos disturbios antrópicos, como períodos repetidos de explotaciones forestales selectivas (uno finalizado en la década del 80 y otro que ocurre en la actualidad), reiteradas invasiones de fuego proveniente de áreas adyacentes (cultivos) y cacería de animales (pecaríes, tapires, corzuelas). Además, cuenta con un camino para acceso a un pozo de gas transitado regularmente. parcelas de 3 x 10 m, y en cada parcela se transplantaron en hilera diez renovales de C. balansae de un año de edad, distanciados 1 m entre si. A cada parcela se le asignó un tratamiento al azar: El sector bajo estudio corresponde a la Selva Pedemontana (500 - 550 m s.n.m.) y las especies dominantes son Calycophyllum multiflorum, Coccoloba tiliacea, Tipuana tipu y Anadenanthera macrocarpa. Las especies más abundantes son: Urera baccifera (19,5% del total de individuos) y Trichilia hieronymi (13,2 %) (Scandalo et al. 2001). 4- Testigo: sin exclusión de herbívoros. El clima es subtropical con precipitaciones anuales que superan los 1.200 mm. El 80 % de las precipitaciones ocurren entre noviembre y abril, con un máximo entre diciembre y marzo (Bianchi 1981). El efecto de la herbivoría sobre la supervivencia y el crecimiento de renovales fue evaluado según un diseño de bloques al azar (Steel y Torrie 1985). Para ello se eligieron al azar cinco picadas madereras abandonadas, debido a la posibilidad de diferencias entre micrositios (compactación del suelo, pendiente, intensidad lumínica). Cada bloque fue dividido en cuatro 1- Insecticida: se aplicó un insecticida carbofurado de amplio espectro para la exclusión de invertebrados, 2- Clausura: se instaló un alambrado para evitar el acceso de los ungulados presentes en el área (pecaríes, corzuelas y tapires), 3- Mixto: con exclusión de ambos tipos de herbívoros, A los 360 días desde del transplante (enero de 2003), se midieron parámetros asociados con el crecimiento (diámetro basal, altura, número de ramas, número de hojas), la herbivoría (número de hojas consumidas por vertebrados y por invertebrados, número de folíolos consumidos por invertebrados) y la supervivencia. La supervivencia se analizó por medio de un Análisis de Varianza (ANOVA) sin replicaciones, y previamente se realizó el Test de no aditividad de Tukey a fin de probar el supuesto de aditividad. Asimismo, se utilizó el Test de comparaciones Múltiples de Tukey-HSD (Zar 1984) para evaluar diferencias en la supervivencia entre los tratamientos. A fin de evaluar el efecto de los tratamientos de exclusión de herbívoros sobre el crecimiento y la herbivoría de los renovales, se realizó un Análisis de Correspondencia 121 Scandalo, R. P.; G. A. Zuleta; M. A. Gainza y A. S. Haedo Múltiple (MCA) con cuatro parámetros de crecimiento y tres parámetros de herbivoría, para la variable suplementaria Tratamientos: Testigo (TR1), Clausura (TR2), Insecticida (TR3) y Mixto (TR4). resultados Se registraron diferencias significativas en el porcentaje de supervivencia entre los tratamientos (ANOVA: F = 8,65, P = 0,002, comparaciones múltiples de Tukey-HSD: mixto vs. testigo P< 0,03), y entre los bloques (F=5,61, P= 0,009). Asimismo, se observó que el porcentaje de supervivencia promedio fue mayor en los tratamientos con insecticida (entre 62 y 80 %), respecto a sin insecticida (40 - 46 %), y que éste aumentó con el grado de protección (Tabla 1). En el MCA utilizado para relacionar los parámetros de crecimiento y herbivoría registrados a 360 días desde el transplante, los primeros dos ejes explicaron el 38,58 % 1. Porcentaje de supervivencia promedio y error estándar por tratamiento. Resultados del Test de comparaciones múltiples de Tukey-HDS. Letras distintas indican diferencias significativas, P<0,03. Tabla Tratamiento Media ± EE Testigo 40,00 ± 10,49 a Clausura 46,00 ± 7,48 ab Insecticida 62,00 ± 11,14 ab Mixto 122 80,00 ± 5,48 b de la variabilidad total (Fig. 1). El eje 1 se encontró influenciado positivamente por los parámetros asociados a menor herbivoría: mayor número de ramas (RA2), bajo porcentaje de hojas consumidas por invertebrados (% HI1), y el tratamiento que combina exclusión de vertebrados e invertebrados (mixto) (TR4). Se encontró influenciado negativamente por aquellos parámetros asociados a mayor herbivoría: presencia de hojas consumidas por vertebrados (HCV2), alto porcentaje y número de hojas consumidas por invertebrados (% HI3, HCI2), intermedio y alto porcentaje de foliolos consumidos por invertebrados (% FI2, % FI3), y tratamientos sin insecticida (es decir, testigo y clausura) (TR1 y TR2, respectivamente). El crecimiento vegetativo no contribuyó al primer eje. El eje 2 se encontró influenciado positivamente por parámetros asociados a menor crecimiento y mayor herbivoría: elevado porcentaje de hojas y foliolos consumidos por invertebrados (% HI3, % FI3), reducido número de hojas totales (HO1), reducido diámetro (DI1), y tratamientos sin insecticida (TR1, TR2); y negativamente por parámetros asociados a mayor crecimiento y menor herbivoría: mayor altura (AL2), valores intermedios de consumo de hojas por invertebrados (% HI2, HC2), y tratamiento de exclusión de invertebrados (TR3). Asimismo, en la Figura 1 se observa que los individuos que presentan mayor daño por invertebrados (% FI2, % FI3, % HI3, HI2) se encontraron asociados a los tratamientos sin insecticida (testigo y clau- Incidencia de la herviboría en Cedrela balansae Figura 1. MCA de parámetros asociados con el crecimiento vegetativo y la herbivoría, en relación a cuatro tratamientos de exclusión de herbívoros. sura) (TR1, TR2); mientras que aquellos que presentan daño leve o moderado por invertebrados (% FI1, % HI1, % HI2, HI1) se encontraron asociados a los tratamientos con insecticida (insecticida y mixto) (TR3, TR4). La contribución relativa de los bloques a los dos primeros ejes del MCA fue baja. Al analizar el porcentaje de individuos consumidos por herbívoros, se registró mayor incidencia de herbivoría por larvas (principalmente de lepidópteros, exceptuando larvas de la mariposita, 84 %) respecto a otros grupos de herbívoros como barrenadores (12%), babosas (2 %), hor- migas (0 %) y vertebrados (2 %) (Fig. 2). Es importante resaltar que, en un experimento realizado simultáneamente con una especie exótica (Toona ciliata o cedro australiano), se registró también mayor incidencia de larvas (85 %) y alta incidencia de hormigas cortadoras que eliminaron todas las hojas de los renovales atacados (15 %), mientras que otros herbívoros no registraron incidencia (babosas, larvas y vertebrados 0%) (Fig. 2). Asimismo, se determinaron los invertebrados hallados sobre los individuos transplantados, a partir de la colecta manual de los mismos y se registró la presencia de in123 Scandalo, R. P.; G. A. Zuleta; M. A. Gainza y A. S. Haedo Figura 2. Incidencia de herbívoros en cedro orán (Cedrela balansae) y tona (Toona ciliata) a 360 días desde el transplante. Se registra el porcentaje de renovales consumidos en mayor grado por cada grupo de herbívoros. vertebrados consumiendo hojas-foliolos. Se determinaron 50 invertebrados, siendo los grupos más abundantes: larvas no determinadas (26%), arácnidos (22 %), himenópteros (18%), y lepidópteros (14 %). Los invertebrados hallados consumiendo follaje corresponden al grupo de los folívoros masticadores: larvas no determinadas (26%), hormigas de la especie Atta saltensis (10%, particularmente consumiendo follaje de T. ciliata), y larvas de Hypsipyla grandella (2 %, consumiendo follaje de C. balansae) (Tabla 2). Tabla 2. Porcentaje de invertebrados regis- Es necesario resaltar que el insecticida no fue efectivo contra el ataque de la mariposita del brote, por lo que no se registró diferencias en la intensidad del ataque en renovales de C. balansae bajo distintos tratamientos. Lepidópteros 14 (2*) Himenópteros 18 (10*) Heterópteros 4 Homópteros 2 124 trados en colecta manual sobre renovales de cedro orán (Cedrela balansae) y tona (Toona ciliata). (*) Porcentaje de invertebrados registrados consumiendo hojas/foliolos de individuos transplantados. Los números entre paréntesis indican: para lepidópteros (porcentaje de larvas de H. grandella), y para hormigas (porcentaje de adultos de Atta saltensis). Grupo Porcentaje Arácnidos 22 Coleópteros 12 Dípteros 2 Larvas no determinadas 26* Incidencia de la herviboría en Cedrela balansae Conclusiones En individuos creciendo bajo exclusión de invertebrados se registró menor daño, lo que indicaría la efectividad del insecticida aplicado y más importante aún, que la herbivoría fue un factor significativo en la supervivencia. En el período bajo estudio, los parámetros de crecimiento no manifestaron una clara diferencia entre los tratamientos aplicados. Sin embargo, se observó cierta asociación entre menor consumo por insectos y mayor crecimiento. El registro de herbivoría por vertebrados fue bajo (< 1%), mientras que el consumo por invertebrados se registró en el 94 % de los individuos. A los 360 días del inicio del experimento, se observó que el porcentaje de supervivencia aumentó con el grado de protección (testigo < clausura/insecticida < mixto), siendo la herbivoría por insectos la causa más importante de mortalidad. Implicancias para el manejo de plantaciones Los cedros son especies de gran importancia, tanto económica como ecológica. Son las especies maderables más codiciadas y de mayor valor en el mercado (Minetti este volumen), a la vez que cumplen una función esencial en la dinámica de regeneración natural de las Yungas dadas sus características de especies claro-dependientes (Grau 2000). A partir de la década del 60, comienza a verificarse una creciente y preocupante disminución de la abundancia y distribución de cedros (Minetti este vo- lumen). Esto se debió a una extracción no sustentable de madera (causa directa), talas rasas para agricultura, ganadería extensiva en remanentes de selva (causas indirectas, sinérgicas) y a una falta de política efectiva de reforestación y conservación (causa primaria), pese a las tempranas alertas sobre la necesidad de ejecutar planes de manejo forestal (Overgaard 1975). En este contexto, las medidas prioritarias para futuros planes de acción son la comercialización de cedros provenientes de plantaciones (Brown y Pacheco este volumen) y la rehabilitación de ecosistemas degradados mediante reforestaciones (Scandalo 2004) como soluciones a los problemas económico y ecológico, respectivamente. Ambos tipos de medidas enfrentarán los riesgos de la herbivoría. La incidencia de Hypsipyla grandella en plantaciones y ensayos de Cedrela spp. por ejemplo, es considerada una seria amenaza a la viabilidad de los emprendimientos forestales. En ensayos realizados en nuestra área de estudio (Valle Morado), el porcentaje de plantas atacadas varió entre el 35 y el 80 % (Horlent y Monteverde este volumen) provocando la muerte de numerosos individuos, independientemente de la procedencia de la semilla. Es decir que la susceptibilidad no dependería de factores genéticos y el patrón de herbivoría no sería selectivo. En condiciones naturales, en cambio, la incidencia de este lepidóptero parece tener menor impacto si bien se registraron ataques en los brotes terminales de las plántulas, incluso en individuos de hasta 20 cm de altura con tallos muy delgados no lignificados (Zamora Petri este volumen). 125 Scandalo, R. P.; G. A. Zuleta; M. A. Gainza y A. S. Haedo En algunos casos, los ataques parecen haber influenciado sobre la supervivencia de plántulas debido su debilitamiento (Grau y Pacheco 1996). En síntesis, estos antecedentes y nuestros resultados ponen en evidencia la necesidad de aplicar medidas de control de la herbivoría, al menos en plantaciones con fines comerciales o para transplantes en proyectos de rehabilitación ecológica. AGRADECIMIENTOS La investigación fue financiada por la Fundación ProYungas (Tucumán) y UBACyT (Bs. As.). Agradecemos la colaboración brindada por: Shell-CAPSA, Sr. Fernando Nocetti, Ing. Carlos Suárez, UNSa, Universidad Maimonides, Madenor, y especialmente a la Lic. Alejandra Christen. BIBLIOGRAFIA Arturi, M.; M. Relva y A.D. Brown. 1999. Consumo foliar de las especies arbóreas en bosques templados xeromórficos del Este de la Provincia de Buenos Aires, Argentina. Revista Chilena de Historia Natural 72: 63-71. Blanton, C.M. y J.J. Ewel. 1985. Leaf-cutting ant herbivory in successional and agricultural tropical ecosystems. Ecology 66: 861-869. Bianchi, A. 1981. Las precipitaciones del Noroeste Argentino. INTA. Salta. 388 pp. Borror, D.J.; C.A. Triplehorn y N. Johnson. 1989. An Introduction to the Study of Insects. Sixth Edition. Saunders College Publishing, Philadelphia. 800 pp. Brown, A.D. 1995. Fitogeografía y Conservación de las Selvas de Montaña del Noroeste de Argentina. En: Churchill et al. (Eds.). Biodiversity and conservation of Neotropical Montane Forests. The New York Botanical Garden. pp 663-672. Brown, A.D. y E. Ramadori. 1989. Patrón de distribución, diversidad y características ecológicas de especies arbóreas de las selvas y bosques montañosos del noroeste de Argentina. Anales VI Congreso Forestal Argentino. Pp 177-181. Brown, A. y S. Pacheco 2006. Importancia del género Cedrela en la conservación y desarrollo sustentable de las Yungas australes. En: S. Pacheco y A. Brown (Eds.). Ecología y producción de cedro (género Cedrela) en las Yungas australes. pp. 9-18. Bygrave, F. L. y Bygrave P. L. 2001. Host preference of the Meliaceae shootborer Hypsipyla: Further information from grafting Cedrela odorata and Cedrela fissilis on Toona ciliata (Australian Red Cedar). Australian Forestry 64: 216-219. 126 Incidencia de la herviboría en Cedrela balansae Cabrera, A. 1976. Regiones Fitogeográficas Argentinas. Enciclopedia de Agricultura y Jardinería. Acme. Camargo, R.S.; L.C. Forti; C.A.O. de Matos; J.F. Lopes y A.P.P. de Andrade. 2004. Physical resistance as a criterion in the selection of foraging material by Acromyrmex subterraneus brunneus Forel, 1911 (Hym., Formicidae). Journal of Applied Entomology 128 (5): 329-331. Cherrett, J.M. 1989. Leaf-cutting ants: biogeographical and ecological studies. En Lieth, H. y M.J. Verger (Eds.). Ecosystems of the world. Tropical rain forest ecosystems. Elsevier, New York. pp 473-488. Coley, P. 1987. Patrones de defensa de las plantas: por qué los herbívoros prefieren ciertas especies? Revista Biología Tropical 35 (1): 151-164. Cortés-Pérez, F. y T.E. León-Sicard. 2003. Modelo conceptual del papel ecológico de la hormiga arriera (Atta laevigata) en los ecosistemas de sabana estacional (Vichada, Colombia). Caldasia 25: 403-417. Crawley, M. 1997. Plant Ecology. M. Crawley ed. Blackwell Science KK. Japan. Dirzo, R. 1987. Estudios sobre las interacciones planta-herbívoro en “Los Tuxtlas”, Veracruz. Revista Biología Tropical 35 (1): 119-132. Dirzo, R. y C. Rodriguez. 1995. Plant – herbivore interactions in Mesoamerican tropical dry forests. En: Bullock, S.; S. Mooney y E. Medina (Eds.). Seasonally dry tropical forests. Cambridge University, Massachusetts. pp 304-345. Grau, H.R. 2000. Regeneration patterns of Cedrela lilloi (Meliaceae) in northwestern Argentina subtropical montane forest. Journal of Tropical Ecology 16: 1-16. Grau, H.R. y S.E. Pacheco. 1996. Demografía y crecimiento de renovales de Cedrela lilloi durante dos años, en un bosque subtropical de montaña de Tucumán Argentina. Yvyraretá 7: 7-11. Farji-Brener, A.G. 1994. Leaf-cutting ants (Atta and Acromyrmex) inhabiting Argentina: patterns in species richness and geographical range sizes. Journal of Biogeography 21: 391-399. Farji-Brener, A.G. 2001. Why are leaf-cutting ants more common in early secondary forests than in old growth tropical forests? An evaluation of the palatable forage hypothesis. Oikos 92: 169-177. 127 Scandalo, R. P.; G. A. Zuleta; M. A. Gainza y A. S. Haedo Floyd, R.B. y C. Hauxwell (Eds.). 2001. Hypsipyla shoot borers in Meliaceae. Proceedings of an International Workshop, Kandy, Sri Lanka 20–23 August 1996. ACIAR Proceedings No. 97, 189 pp. Folgarait, P.J.; A.G. Farji-Brener y J.J. Protomastro.1994. Influence of biotic, chemical and mechanical plant defences on the foraging pattern of the leaf-cutter ant (Acromyrmex striatus) in a subtropical forest. Ecologia Austral 4: 11-17. Fonseca, R. 1994. Herbivory and the long-lived leaves of an Amazonian ant-tree. Journal of Ecology 82: 833-842. Honkanen, T. y E. Haukioja. 1994. Why does a branch suffer more after branch-wide than after tree-wide defoliation?. Oikos 71: 441-450. Holldober, B. y E.O. Wilson. 1990. The ants. The Belknap Press of Harvard University Press, Cambridge. Horlent, M. y D. Monteverde. 2006. Crecimiento de renovales de Cedrela balansae en la plantación experimental de Valle Morado. En: S. Pacheco y A. Brown (Eds.). Ecología y producción de cedro (género Cedrela) en las Yungas australes. pp. 171-178. Howard, F.W. 1991. Seasonal incidence of shoot infestation by mahogany shoot borer (Lepidoptera: Phyticidae) in Florida. Florida Entomologist 74: 150–151. Hubbel, S.P.; J.J. Howard y D.F. Wiemer. 1984. Chemical leaf repellency to an Attini ant: seasonal distribution among potential host plant species. Ecology 65: 10671076. Liang, S. y S. Seagle. 2002. Browsing and microhabitat effects on riparian forest woody seedling demography. Ecology 83: 212-227. Mata, A. 2001. Diagnóstico de los insectos herbívoros presentes en las plantaciones de monocultivos de pilón, cedro y laurel, ubicadas en las parcelas experimentales del Proyecto Huertos, Estación Biológica La Selva, Puerto Viejo de Sarapiquí, Heredia Costa Rica. Informe de Práctica de Especialidad. Cartago, CR. Instituto Tecnológico de Costa Rica. 106 pp. Minetti, J.M. 2006. Aprovechamiento forestal de cedro en las Yungas de Argentina. En: S. Pacheco y A. Brown (Eds.). Ecología y producción de cedro (género Cedrela) en lasYungas australes. pp. 143-154. Moulaert-Quiros, A. y M. Arguedas. 1993. Insectos herbívoros asociados con diez especies forestales en la Región Huetar Norte de Costa Rica. Revista Forestal Centroamericana 2 (2): 24-26. 128 Incidencia de la herviboría en Cedrela balansae Newton, A.C.; J.P. Cornelius; J.F. Mesén; E.A. Corea y A.D. Watt. 1998. Variation in attack by the mahogany shoot borer, Hypsipyla grandella (Lepidoptera: Pyralidae), in relation to host growth and phenology. Bulletin of Entomological Research 88: 319–326. Newton, A.C.; A.D. Watt; F. López; J. Cornelius; J.F. Mesén y E.A. Corea. 1999. Genetic variation in host susceptibility to attack by the mahogany shoot borer, Hypsipyla grandella (Zeller). Agricultural and Forest Entomology 1: 11–18. Overgaard, J. 1975. Aprovechamiento y transporte forestal en el Noroeste Argentino. NOA II Forestal. IFONA, UNDP y FAO. Salta Pelotto, J.P. y M.A. del Pero de Martínez. 2002. Chemical defenses in the tree Ziziphus mistol against the leaf-cutting ant Acromyrmex striatus. Ecología Austral 12: 1118. Rao, M.; J. Terborgh y P. Nuñez. 2001. Increased herbivory in forest isolates: implications for plant community structure and composition. Conservation Biology 15: 624-633. Relva, M. y T. Veblen. 1998. Impacts of introduced large herbivores on Austrocedrus chilensis forests in northern Patagonia, Argentina. Forest Ecology and Management 108: 27-40. Rockwood, L.L. 1976. Plant selection and foraging patterns in two species of leaf-cutting ants (Atta). Ecology 57: 48–61. Rodgers, H.L.; M.P. Brakke y J.J. Ewel. 1995. Shoot damage effects on starch reserves of Cedrela odorata. Biotropica 27: 71-77. Roldán, A. y J. Simonetti. 2001. Plant - mammal interactions in Tropical Bolivian Forests with different hunting pressures. Conservation Biology 15 (3): 617-623. Scandalo, R. 2004. Efecto de la herbivoría sobre el establecimiento de renovales de Cedrela balansae Sessé y Moriño y Toona ciliata M. Roem en un sector de Yungas perturbado por tala selectiva. Tesis de grado. Universidad de Buenos Aires. Scandalo, R.; M. Cagnoni y G. Zuleta. 2001. Análisis estructural de la vegetación en selvas de Yungas sometidas a perturbación antrópica (Salta, Argentina). Libro de resúmenes de la I Reunión Binacional de Ecología. Bariloche, Argentina. Steel, R. y J.Torrie. 1985. Bioestadística: principios procedimientos. McGraw-Hill. 613 pp. 129 Scandalo, R. P.; G. A. Zuleta; M. A. Gainza y A. S. Haedo Stradling, D.J. 1978. The influence of size on foraging in the ant Atta cephalotes, and the effect of some plant defense mechanisms. Journal of Animal Ecology 47: 173-188. Taveras, R.; L. Hilje; P. Hanson; R. Mexzón; M. Carballo y C. Navarro. 2004. Population trends and damage patterns of Hypsipyla grandella (Lepidoptera: Pyralidae) in a mahogany stand, in Turrialba, Costa Rica. Agricultural and Forest Entomology 6: 89-98. Tillmanns, H. 1964. Apuntes bibliográficos sobre Hypsipyla grandella Zeller. Boletin del Instituto Forestal Latinoamericano. Mérida, Venezuela. 14: 82-92. Vasconcelos, H.L. 1997. Foraging activity of an Amazonian leaf-cutting ant: responses to changes in the availability of woody plants and to previous plant damage. Oecologia 112: 370-378. Veblen, T.; M. Mermoz; C. Martin y T. Kitzberger. 1992. Ecological impacts of introduced animals in Nahuel Huapi National Park, Argentina. Conservation Biology 6: 71-83. Vega, C.L. 1976. Influencia de la silvicultura en el comportamiento de Cedrela in Surinam. En: Whitmore, J.L. ed. Studies on the shoot borer Hypsipyla grandella (Zeller) Lep. Pyralidae Volume III. IICA Miscellaneous publication no. 101, CATIE, Turrialba, Costa Rica, 26–49. Wallin, K. y K. Raffa. 2001. Effects of folivory on subcortical plant defenses: can defense theories predict inter guild processes?. Ecology 82: 1387-1400. Wetterer, J.K.; D.S. Gruner y J.E. Lopez. 1998. Foraging and nesting ecology of Acromyrmex octospinosus (Hymenoptera: Formicidae) in a Costa Rican tropical dry forest. Florida Entomologist 81: 61-67. Yamazaki, S.; A. Taketani.; K. Fujita, C. Vasques y T. Ikeda. 1990. Ecology of Hypsipyla grandella and its seasonal changes in population density in Peruvian Amazon forest. Japan Agricultural Research Quarterly 24, 149–155. Zamora Petri, M. 2006. Influencia de la ganadería trashumante y la apertura de claros en la supervivencia y crecimiento de Cedrela lilloi en Tariquía, Bolivia. En: S. Pacheco y A. Brown (Eds.). Ecología y producción de cedro (género Cedrela) en las Yungas australes. pp. 131-142. Zar, J. 1984. Biostatistical Analysis. Prentice-Hall. New Jersey. 718 pp. 130 Influencia Ecología de la ganaderia trashumante la apertura de claros en Cedrella lilloi y producción de cedro y(género Cedrela) en las Yungas australes. Pacheco, S. y A. Brown (Eds.). LIEY-ProYungas. Argentina. pp. 131-142. 2006. Influencia de la ganadería trashumante y la apertura de claros en la supervivencia y el crecimiento de Cedrela lilloi en Tariquía, Bolivia Zamora Petri, M. Naturaleza Tierra y Vida (NATIVA), Ingavi La Paz 1064 Tarija, Bolivia. [email protected] El objetivo de este trabajo fue estudiar la influencia de la apertura de claros en el bosque y de la ganadería de trashumancia, sobre el crecimiento y la mortalidad de plántulas de Cedrela lilloi. Se trabajó en condiciones naturales, con plantas ubicadas en claros y sotobosque. Se realizaron medidas de crecimiento y mortalidad en cuatro momentos a lo largo de un año. Las aperturas del dosel del bosque favorecen significativamente el crecimiento de las plántulas debido al mayor ingreso de radiación solar directa. Por el contrario, la presencia de ganado aumenta significativamente la mortalidad de las plántulas debido principalmente al efecto del daño físico causado por el pisoteo. The study evaluated the influence of canopy gaps and cattle in seedling growth and mortality of Cedrela lilloi. Growth and mortality rates were measured in saplings located both in gaps and in the understory four times during one year. Canopy gaps significantly influenced saplings growth, mainly due to the increased solar radiation, essential for photosynthesis, as well as to environmental changes that favored the development of the saplings. Conversely, livestock presence in the area increases sapling mortality mainly due to the effect of physical damage caused by trample. 131 Zamora Petri, M. Introduccion La regeneración natural de las especies arbóreas se define como el proceso por el cual las plantas se reemplazan unas a otras. El éxito de esta etapa depende de factores ambientales (condiciones microclimáticas y edáficas adecuadas para la germinación de las semillas y el desarrollo de las plántulas) y de las interacciones entre las especies (dispersores, competidores y depredadores). El objetivo de este trabajo fue estudiar la regeneración natural del cedro coya (Cedrela lilloi) en el sector sur de las Yungas de Bolivia, en relación a la estructura de la vegetación y la mortalidad causada por depredadores. En particular se determinó la influencia que ejerce la apertura de claros en el dosel del bosque y la ganadería de trashumancia, sobre la supervivencia y crecimiento de plántulas de Cedrela lilloi. Importancia de los claros en la regeneración natural de especies arbóreas Uno de los principales efectos de la apertura de claros en el dosel por la caída de un árbol, es la exposición del suelo del bosque a la luz solar directa. Los claros son utilizados por las especies “pioneras o heliófitas” para el establecimiento y reclutamiento, que requieren condiciones ambientales de mayor exposición para germinar y establecerse. Por otro lado, las especies que permanecen en el sotobosque en un estado retardado del crecimiento (Uhl y Nepstad 1990), también aprovechan el claro para alcanzar el dosel, aumentando sus tasas de 132 crecimiento en respuesta al aumento en la radiación. En el bosque húmedo de Costa Rica, el 63 % de las especies de dosel son clarodependientes, entre las que se destacan maderas valiosas como Cedrela spp. y Swietenia macrophylla (Hartshorn 1989). La especie Cedrela odorata también es clasificada como una especie pionera (Lamprecht 1990). En la Reserva Nacional de Flora y Fauna Tariquía (de aquí en más llamada Reserva Tariquía), el género Cedrela se halla principalmente representado por Cedrela lilloi. Observaciones de campo sugieren que aunque es posible encontrar plántulas bajo el dosel cerrado del bosque, su crecimiento se encuentra influenciado por la apertura de claros. Grau (2000) determinó que Cedrela lilloi se caracteriza por un rápido crecimiento, en comparación con otros géneros neotropicales como Jacaranda, Cecropia y Solanum. Las tasas de crecimiento elevadas y el patrón espacial agregado, permiten, caracterizar a la especie como claro dependiente en el modo de regeneración (Grau 2000). Influencia de la ganadería en la regeneración natural de especies arbóreas El aumento de las perturbaciones antrópicas en los ambientes naturales ha llevado a que en muchas áreas, sean más comunes que las perturbaciones naturales (Uhl y Nepstad 1990). La recuperación que sigue a una perturbación antrópica de gran magnitud (por ejemplo pastizales ganaderos abandonados), es en general lenta y en Influencia de la ganaderia trashumante y la apertura de claros en Cedrella lilloi muchos casos se desvía hacia una formación estabilizada no boscosa (Uhl y Nepstad 1990). La actividad ganadera, en su mayoría, se caracteriza por la habilitación de pasturas en tierras con bosque, que generalmente implica la sobreexplotación del suelo debido a la extracción de nutrientes de manera tan rápida que estos no pueden ser reemplazados. A esto se suma la excesiva concentración de ganado, lo que lleva a una compactación y erosión del suelo, con daños irreversibles y el consecuente abandono de las tierras (Serrao y Toledo en Anderson 1990, Faminow 1998). La regeneración del bosque luego de este tipo de perturbaciones es muy lenta. Esto se debe a que las plántulas de las especies arbóreas deben competir con especies herbáceas y a la elevada mortalidad de las plántulas causada por el pisoteo y la depredación por parte del ganado (Serrao y Toledo en Anderson 1990, Uhl y Nepstad 1990). La actividad ganadera practicada en la Reserva Tariquía no implica la habilitación de tierras boscosas para este fin, sino que aprovecha los recursos del bosque para el pastoreo del ganado en la época seca, lo que se conoce como ganadería de trashumancia. La misma consiste en el pastoreo en diferentes pisos altitudinales, de manera cíclica estacional y de acuerdo a la disponibilidad de pastos. El ciclo comienza con un período de pastoreo en zonas del valle en la época húmeda (diciembre a abril) y continúa con el traslado de ganado a áreas de bosque durante la época seca (mayo a noviembre). El ganado trashumante que pastorea dentro de la Reserva Tariquía, procede en su mayoría de comunidades ubicadas fuera de la misma y en menor proporción de las comunidades ubicadas en su interior. La Reserva soporta casi toda la carga ganadera del Valle Central de Tarija y zonas aledañas a la Reserva. Según el censo ganadero realizado en el 2000, la cantidad de ganado que ingresa a la Reserva es de 19.065 cabezas. La práctica de la trashumancia resulta en una distribución más homogénea de la intensidad de pastoreo y se la identifica como una forma de ampliar el uso de la tierra y la presión de pastoreo (Molinillo 1993). Esta práctica quizás reduzca los efectos en la compactación del suelo, pero la ganadería aún representa un factor importante en la determinación de la composición y estructura del bosque (Brown y Grau 1990). metodos Area de estudio El estudio se realizó en la comunidad de Chiquiacá ubicada al noreste de la Reserva Nacional de Flora y Fauna Tariquía, Departamento de Tarija, Bolivia. La Reserva cuenta con una superficie de 246.870 ha, abarcando las Provincias de Arce, O’Connor, Gran Chaco y Cercado (21° 45’ - 22° 20’ S y 64° 05’ - 64° 36’ O). El área de la Reserva pertenece a la vertiente oriental andina, la mayor parte del área corresponde a la provincia biogeográfica de Yungas (dominio amazónico) y en menor proporción al Chaco serrano (dominio chaqueño) (Cabrera y Willink 1973). Las temperaturas medias anuales varían 133 Zamora Petri, M. entre18 y 22°C. La precipitación promedio va desde los 1.000 a los 2.000 mm anuales, concentrándose el 85 % en los meses de noviembre a marzo (Gonzalez et al. 1996). En la comunidad de Chiquiacá, se encuentra un bosque alto húmedo submontano semi siempreverde distribuido entre los 600 y 1.200 m s.n.m. con un relieve quebrado. Domina la vegetación arbórea de porte alto de 20 a 25 m en el dosel, con emergentes que pueden sobrepasar los 30 m. Entre las especies dominantes se encuentran: Cinammomum porphyria y Ocotea puberula (Lauráceas), Cedrela lilloi (Meliaceas), Blepharocalyx salicifolius y Eugenia spp. (Myrtaceas) y Juglans australis (Junglandaceas). La zona presenta un alto epifitismo donde dominan bromeliáceas y orquidáceas. Para evaluar la influencia de los claros en el crecimiento de Cedrela lilloi, se establecieron 10 parcelas de 400 m² cada una (20 x 20 m). Las parcelas fueron distribuidas en un rango altitudinal entre 850 y 1.050 m s. n.m. y la distancia entre cada una fue de aproximadamente 1.000 m. La selección de las parcelas no fue al azar, debido a que el patrón de distribución espacial de las plántulas de Cedrela lilloi tiende a ser agregado. Por esto, previo a la selección de las parcelas, se realizó un diagnóstico general de la zona para identificar las agregaciones de plántulas. De las diez parcelas establecidas, sólo dos presentaban una clara apertura de dosel, una de origen natural ocasionada por la caída de un árbol y la otra de origen an134 trópico debido a la actividad forestal. Las restantes ocho parcelas fueron consideradas como de dosel cerrado. Dado que la presencia de agregados de plántulas no es común, no fue posible establecer otras parcelas bajo claros. Se consideró que las parcelas estaban ubicadas bajo un claro cuando el 80 % del dosel se encontraba libre de follaje y que estaban bajo el dosel cerrado, cuando el 80 % del dosel se encontraba cubierto de follaje. A pesar de que las parcelas no fueron instaladas a la misma altura, ya que se cubrió un rango altitudinal de 200 m, las características del bosque fueron similares en las diez parcelas. El estrato arbóreo presentaba una clara dominancia de Myrtaceas como B. salicifolius y Eugenia spp. y un sotobosque poco poblado, en el que se podían observar helechos y plántulas principalmente de Myrtaceas. La pendiente de las parcelas fue menor a 20º. Dentro de cada una de las diez parcelas, se marcaron 18 plántulas de Cedrela lilloi (aproximadamente el 30 % del total de plántulas de cada parcela) con una ficha indicando el número de parcela y el número de plántula. Durante un año se midió el crecimiento en altura en cuatro fechas: t0) en julio de 1998 cuando se marcaron y tomaron las medidas iniciales de las plántulas, t1) en octubre de 1998, t2) en enero de 1999 , y t3) en julio de 1999. La diferencia del crecimiento de las plántulas establecidas en claros y en sotobosques, en función de los controles realizados (t0 - t3) y las diferencias intrínsecas de una parcela a otra, se analizaron Influencia de la ganaderia trashumante y la apertura de claros en Cedrella lilloi mediante un Análisis de Varianza de tres vías para medidas repetitivas (ANDEVA) (Zar 1996). Todo el procesamiento de datos fue elaborado utilizando el programa estadístico SPSS 8.0 para Windows. Para el análisis estadístico fueron descartadas aquellas plántulas marcadas que murieron a lo largo del tiempo de muestreo. Para evaluar la influencia de la ganadería de trashumanancia en la supervivencia de plántulas de Cedrela lilloi, se trabajó con las diez parcelas seleccionadas para el caso anterior. Dentro de cada una se establecieron dos sub-parcelas de 36 m² (6 x 6 m). Una de las dos sub-parcelas fue cercada con malla, con el propósito de aislar el impacto del ganado (control experimental). La otra sub-parcela permaneció sin clausura, pero delimitada con estacas en sus extremos. Dentro de cada una, se marcaron 9 plántulas con fichas plastificadas en las que se indicó el número de plántula y parcela. Los controles fueron simultáneos a los realizados para evaluar la influencia de la apertura de dosel en el crecimiento de las plántulas (t0-t3). Durante cada control se determinó el número de plántulas muertas y se identificó cuantitativamente la cau- sa de la mortalidad. Se consideró muerte por efecto del ganado cuando la plántula presentaba evidentes signos de daño físico como ruptura o aplanamiento de sus partes y muerte por sequía cuando la plántula presentaba una absoluta deshidratación de la raíz, tallo y hojas. La diferencia de la mortalidad de plántulas observadas en las sub-parcelas con ganado, con relación a las sub-parcelas sin ganado, en función a los tres controles realizados, se analizó mediante un Análisis de Varianza de dos vías para medidas repetitivas (ANDEVA). Resultados En cuanto a la influencia de los claros en el crecimiento de cedro, se observó que las plántulas que se desarrollaron bajo el dosel abierto tuvieron un crecimiento mayor con relación a las del sotobosque. Las plántulas de claros superaron en más de 20 cm a las plántulas de dosel cerrado, que sólo alcanzaron un crecimiento anual total de 5 cm (Tabla 1). El análisis de varianza de tres vías mostró que el desarrollo de las plántulas muestra diferencias significativas entre los tiempos Tabla 1. Crecimiento (media ± error estándar) de plántulas de cedro (Cedrela lilloi) desarrolla- das en claros del bosque (dosel abierto) y bajo el sotobosque (dosel cerrado). Cobertura del dosel Tamaño inicial Tamaño final Crecimiento Claros (dosel abierto) 13,41cm ± 1,01 40,16 cm ± 2,70 ≈ 26,75 cm Sotobosque (dosel cerrado) 9,33 cm ± 0,37 14,04 cm ± 0,74 ≈ 4,71 cm 135 Zamora Petri, M. de muestreo (t0 – t3). Esto indica que el crecimiento no es constante a lo largo del tiempo, sino que durante el periodo de lluvias el crecimiento es mayor. Estas diferencias en el crecimiento fueron más evidentes en plántulas que se desarrollaron bajo claros. El crecimiento de las plántulas del sotobosque bajo el dosel cerrado (regeneración preexistente) fue casi homogéneo a lo largo del tiempo, frente a un incremento significativo en el crecimiento de plántulas bajo condiciones de iluminación. Este aspecto caracteriza a Cedrela lilloi como una especie oportunista de claros (Tabla 2 y Fig. 1). FIGURA 1. Tamaño a lo largo del año de muestreo, de las plántulas de Cedrela lilloi establecidas en claros (dosel abierto) y en el sotobosque (dosel cerrado) (promedio ± error estándar). Tabla 2. Resultados de ANDEVA de 3 vías para medidas repetitivas para crecimiento de plán- tulas de Cedrela lilloi desarrolladas en claros del bosque (dosel abierto) y bajo el sotobosque (dosel cerrado). Se presenta el estadístico F para las tres fuentes de variación (tiempo, apertura del dosel y parcelas) y para las interacciones * = Efecto significativo P<0,01. Fuentes de variación Grados de libertad Estadístico F Apertura de dosel: abierto y cerrado 1 182,820 * Parcelas (2, 8) 7 3,087 * Interacción: tiempo x apertura de dosel 3 27,821 * Interacción: tiempo x parcelas 21 0,275 * Interacción: apertura de dosel x parcelas 1 26,460 * Interacción: apertura de dosel x controles x parcelas 3 2,058 136 Influencia de la ganaderia trashumante y la apertura de claros en Cedrella lilloi Figura 2. Mortalidad durante el año de muestreo de plántulas de C. lilloi en lugares con presencia de ganado y sin presencia de ganado (promedio ± error estándar). En cuanto a la influencia de la ganadería de trashumancia en la supervivencia de Cedrela lilloi se observó una mayor mortalidad de plántulas en áreas con presencia de ganado vacuno con relación a áreas con exclusión de ganado. El porcentaje máximo de mortalidad registrado fue del 34 % en áreas sin exclusión de ganado y del 22 % en áreas con exclusión de ganado (Tabla 3 y Fig. 2). El daño causado por el ganado sobre las plántulas de Cedrela lilloi, se evidencia como daño físico cau- Tabla 3. Porcentajes de mortalidad de plántulas de cedro (Cedrela lilloi) creciendo en parcelas con exclusión (sin efecto del ganado) y en parcelas sin exclusión (expuestas al ganado) (promedio ± error estándar). Mortalidad máxima Mortalidad mínima Sin ganado 22,22 % ± 7,94 3,33 % ±1,69 Con ganado 34,44 % ± 5,35 16,67 % ± 4,45 137 Zamora Petri, M. Tabla 4. Resultados de ANDEVA de 2 vías para medidas repetitivas para la mortalidad de plántulas de Cedrela lilloi desarrolladas en parcelas con y sin exclusión del ganado. Se presenta el estadístico F para las dos fuentes de variación (tiempo y ganado) y para la interacción. * = efecto significativo P< 0,01. Fuentes de variación Grados de libertad Estadístico F Tiempo (t1- t3) 2 13,220 * Ganado: con exclusión y sin exclusión 1 7,571 * Interacción: tiempo x ganado 2 0,415 sado por el pisoteo. Se encontró una variabilidad significativa en los porcentajes de mortalidad de plántulas entre los tiempos de muestreo. Esto se debe a que por la estacionalidad hídrica hay mayor mortalidad de las plántulas en la época seca (Tabla 4). Esto se manifestó a pesar de haber trabajado con una muestra desbalanceada (2 parcelas en claros y 8 parcelas en el sotobosque). Estudios previos reportan un incremento de 30 a 60 cm en altura/año en individuos de Cedrela lilloi mayores a 2 m de altura bajo condiciones de iluminación (Grau y Pacheco 1996, Grau 2000). Discusion La disponibilidad de luz es un factor determinante en el crecimiento de C. lilloi y no así en la germinación de las semillas. Por esto se caracteriza a la especie como oportunista de claros, por cuanto depende de ellos sólo para el crecimiento (Grau 2000) y no para la germinación, como las especies pioneras (Kyereh et al. 1999). Influencia de los claros en el crecimiento de cedro Uno de los efectos más notables de la apertura de claros en el dosel del bosque, es la mayor disponibilidad de luz solar directa por parte de las plantas. Debido a las condiciones naturales en las cuales se desarrolló el estudio, no se puede afirmar que las condiciones de luz fueron iguales en cada apertura del dosel. Sin embargo, las diferencias encontradas en el crecimiento de las plántulas, muestran una clara influencia de la luz en el crecimiento y desarrollo de esta especie. El crecimiento promedio de las plantas en claros superó en más de 20 cm a las plántulas bajo el sotobosque. 138 El mayor crecimiento de las plántulas de Cedrela lilloi ocurrió principalmente en la época lluviosa (noviembre a marzo), durante la cual se registra el 85 % de la precipitación anual. Esta disponibilidad de humedad permite un incremento en la superficie foliar, favoreciendo el desarrollo de la fotosíntesis que incide en el crecimiento de las plántulas. Este crecimiento Influencia de la ganaderia trashumante y la apertura de claros en Cedrella lilloi es aún mayor en plántulas bajo claros. El trabajo se llevó a cabo con individuos pequeños (post-germinación) en los que el sistema radicular se encuentra poco desarrollado. Esto constituyó un factor determinante en la falta de crecimiento de las plántulas durante la época seca. La ausencia de crecimiento refleja la disminución en la tasa fotosintética (principalmente en plántulas bajo sombra) como respuesta a la pérdida de hojas. La falta de humedad del suelo también parece influir en la mortalidad de plántulas (Poorter 1998). Otro aspecto que podría haber influido en el crecimiento y desarrollo de las plántulas de Cedrela lilloi, es su distribución espacial, ya que las plántulas de esta especie tienden a ser espacialmente agregadas a una escala de pocos cientos de metros cuadrados (Grau 2000). Este hecho también se observó en el estudio, en el que las agrupaciones tienden a ocupar un área aproximada a los 400 m². La distribución de las plántulas se encuentra relacionada con la dispersión anemócora de sus semillas, en la que la dirección del viento y el tipo de follaje juegan un papel importante en la distribución espacial que presentan. Este aspecto podría afectar el crecimiento y desarrollo de las plántulas creando competencia intraespecifica, tanto por luz como por nutrientes (Baldiviezo 1993). Influencia de la ganadería de trashumancia en la supervivencia de Cedrela lilloi Las plántulas están formadas por tejidos jóvenes frágiles y con alta susceptibilidad al ataque de plagas, depredadores, condi- ciones climáticas adversas y actividades antrópicas como la ganadería. Estos factores constituyen procesos que condicionan la supervivencia de las plántulas (Baldiviezo 1993). Por lo tanto, se debe evaluar el efecto de la ganadería practicada en la Reserva de Tariquía sobre la regeneración de especies como Cedrela lilloi, en conjunto con las causas naturales de mortalidad. En las plantas analizadas se encontraron evidencias de ataque de Hypsipyla grandella en los brotes terminales de las plántulas. En plántulas de hasta 20 cm de altura con un tallo muy delgado y no lignificado, la larva destruye únicamente el ápice. Sin embargo, en plántulas más grandes se observó el desarrollo de la larva dentro de la plántula. Por otro lado, la marcada estacionalidad de las lluvias es otra causa natural, que influyó significativamente en la mortalidad de las plántulas, lo cual se refleja en los distintos muestreos realizados a lo largo del año. En cuanto al efecto de la ganadería en particular, los resultados de este trabajo muestran una mayor mortalidad de plántulas en zonas expuestas a la influencia del ganado. El ganado disminuye la probabilidad de reclutamiento de las plántulas, debido al daño físico producido por el pisoteo. El ramoneo también es un efecto importante debido a que ocasiona, en la mayoría de los casos, la muerte de los individuos afectados. Esto es más evidente en la época seca, que es cuando el ganado ingresa al monte y las plántulas se encuentran poco vigorosas y por consiguiente más vulnerables al efecto del pisoteo. El efecto del ganado también fue documentado como 139 Zamora Petri, M. una de las probables causas de bajas tasas de regeneración natural en Cedrela (Baldiviezo 1993). Si bien el efecto de la ganadería aumenta la mortalidad de las plántulas, también se podría considerar la incidencia que tiene sobre la cobertura del sotobosque. El pastoreo disminuye la cobertura lo que podría aumentar la llegada de la luz, lo cual resultaría beneficioso para el desarrollo de las plántulas. Por esto, es posible que algunas densidades de pastoreo favorezcan el crecimiento a pesar de aumentar también la mortalidad. Se crea así una situación delicada en la que las condiciones óptimas para la regeneración de la especie estarían dadas por un balance de disturbio que maximice el compromiso crecimiento-supervivencia. De todos modos el efecto de un mayor crecimiento demostrado en este trabajo por mayor llegada de radiación directa, se evaluó en aperturas de dosel y no en coberturas de sotobosque. Sin embargo, este posible efecto doble de la ganadería puede ser de gran importancia para el cedro y podría considerarse al momento de evaluar el manejo de la ganadería de trashumancia. BibliografIa Anderson, A. 1990. Alternativas a la deforestación (Ed.). Abya-Yala, Quito. 415 pp. Baldivieso, J.P. 1993. Estudio de la regeneración natural de Cedrela balansae. Tesis de grado, Universidad Juan Misael Saracho, Tarija. Brown, A.D y H.R. Grau. 1990. Reserva Natural de Tariquía - Dpto. Tarija Informe Inédito PROMETA. Cabrera, A. y A. Willink. 1973. Biogeografía de América Latina. Serie Biología. Monog. 13. OEA. Faminow, M. 1998. Cattle, deforestation and development in the Amazon. An Economic, agronomic and environmental perspective, New York. CAB International. Gonzalez, J.; G.J. Scrocchi y E.O. Lavilla. 1996. Relevamiento de la biodiversidad de la Reserva Nacional de Flora y Fauna Tariquía. Fundación Miguel Lillo. Tucumán. 130 pp. Grau, H.R. y S.E. Pacheco. 1996. Demografía y crecimiento de renovales de Cedrela lilloi durante dos años, en un bosque subtropical de montaña de Tucumán, Argentina. Yvyraretá 7: 7-11. Grau, H.R. 2000. Regeneration patterns of Cedrela lilloi (Meliaceae) in northwestern Argentina subtropical montane forest. Journal of Tropical Ecology 16: 1-16. 140 Influencia de la ganaderia trashumante y la apertura de claros en Cedrella lilloi Hartshorn, G.S. 1989. Gap-phase dynamics and tropical tree species richness. En: HolmNielsen, L.B.; I.C. Nielsen y H. Balsev (Eds.). Tropical Forests: Botanical Dynamics, Speciation and Diversity. London, UK Academic. pp. 65-73. Kyereh, B.; M.D. Swaine y J. Thompson. 1999. Effect of light on the germination of forest trees in Ghana. Journal of Ecology 87: 772-783. Lamprecht, H. 1990. Silvicultura en los trópicos. Eschborn. GTZ. 335 pp. Molinillo, M. 1993. Is traditional pastoralism the cause of erosive processes in mountain enviroments? The case of the cumbres calchaquies in Argentina. Mountain Research and Development 13 (2): 189-202. Poorter, L. 1998. Seedling growth of Bolivian rain forest tree species in relation to light and water availability. PROMAB, Beni. 169 pp. Uhl, C. y D. Nepstad. 1990. Perturbaciones naturales y antropogénicas en la amazonia. En: Anderson, A. (Ed.). Alternativas a la deforestación. Eschborn. pp 45-79. Zar J.H. 1996. Biostatical analisys. (III edición.) Prentice-Hall, Nueva Jersey. 523 pp. 141 Aprovechamiento de cedro en lasenYungas de Argentina Ecología y producción de forestal cedro (género Cedrela) las Yungas australes. Pacheco, S. y A. Brown (Eds.). LIEY-ProYungas. Argentina. pp. 143-154. 2006. Aprovechamiento forestal de cedro en las Yungas de Argentina Minetti, J. M. I.N.T.A.- Mosconi, R.N. 34, Km 1425 (4562), Gral. E. Mosconi, Salta, Argentina. [email protected] El cedro es la especie de mayor valor en el mercado de productos maderables y se lo utiliza para carpintería general, construcciones y otros usos menores. Se describe el sistema de aprovechamiento típico y las pautas operatorias establecidas entre los dueños de las tierras y los propietarios de aserraderos. Además, se plantea que la tala selectiva que sufrieron los cedros en las Yungas argentinas llevó a sus especies a una situación crítica. Debido a esto, es imprescindible realizar planes de manejo forestal que tengan como fin la sustentabilidad del recurso y de la actividad. Estas acciones de manejo que incluyen controlar los aprovechamientos ilegales, respetar los rodales protectores, seleccionar y preservar rodales e individuos como semilleros, deben encararse en forma urgente a través de alianzas públicas y privadas. Una medida para contribuir a salvaguardar el futuro de los cedros de las Yungas es la reforestación a través de enriquecimientos o macizos, aliviando la presión sobre las formaciones nativas. At present, Cedrela is the most valuable timber species in wood market. It is used for carpentry, woodwork and other smaller uses. This work describes the most typical exploitation system and the operating guidelines between landowners and sawimill owners. It is stated that selective logging of Cedrela in Argentinian Yungas caused a crtitical situation for this species. Now, it is essential to develop a forestry management strategy to assure resource persistence and economical sustainability. These actions include efective control to avoid illegal operations, preservation of protective stands and individuals as seed sources, and should be urgently implemented trough public and private alliances. A way to contribute to preserve Cedrela in Yungas is reforestation throw seedling production and enrichment activities, reducing as a consequence pressure on native forests. 143 Minetti, J. M. aprovechamiento de cedro en la región La utilización de madera de cedro con fines comerciales en el noroeste de Argentina data de la época colonial, aunque probablemente, la población prehispánica también haya aprovechado esta madera para la elaboración de utensilios tales como las bateas. Ya en el siglo XIX se conocían sus excelentes propiedades para la construcción de muebles y carpintería rústica. El cedro, junto a otras especies de maderas duras o semiduras, se utilizaba en la fabricación de casas, muebles, carretas, envases y herramientas. Debido a la ausencia de estructuras de transporte, esta madera se comercializaba fundamentalmente en el mercado local. El avance del ferrocarril en la Provincia de Tucumán en 1876 y su continuación hacia las otras provincias del norte, impulsó el desarrollo de la actividad forestal a un ritmo comercial importante y permitió la explotación forestal a lo largo de la red ferroviaria (Saravia Toledo 1994). La introducción de maquinaria de aserrraje movida por motores a vapor y la facilidad de transporte dada por los trenes hacia los mercados del centro del país, hicieron de la actividad forestal un sector socioeconómico importante en la región. En un informe del Departamento Nacional de Agricultura del año 1876, el inspector de Agricultura de la Provincia de Salta cita: “...Los bosques que encierra esta vasta provincia, son suficientes para proveer a medio mundo de las mejores maderas de construcción, para obras de todas clases y 144 muebles exquisitos como ser: el cedro...”. En el mismo documento se describe el potencial forestal de los distintos departamentos provinciales, donde el cedro encabeza la lista de especies más valiosas. Ya en esa época, existían en el Departamento La Caldera dos aserraderos que proveían de tablas a la capital salteña. Durante la primera mitad del siglo XX, la explotación forestal fue una actividad importante en las Yungas argentinas y fue el motor de las economías de los pueblos que iban surgiendo hacia el interior de las provincias. El cedro era y es en la actualidad, la especie más codiciada y de mayor valor en el mercado de productos maderables. En esa época el aprovechamiento se hacía en forma rudimentaria y manual, usando el hacha para el apeo y trozado, bueyes para el rodeo de la madera hasta los sitios de acopio y camiones para el transporte hasta los aserraderos (Foto 1). Sin embargo, el transporte más importante era el ferrocarril, que llevaba la producción a la Capital Federal desde donde se redistribuía al interior de la región pampeana y se exportaba al exterior. Se comercializaban rollos y tablas que se industrializaban en las principales ciudades del centro del país (Buenos Aires, Rosario, Córdoba y Santa Fé). En los primeros años de la segunda mitad del siglo XX empiezan a mecanizarse muchas tareas relacionadas con la actividad forestal, tanto en la selva como en la industria. La aparición de las motosierras, los tractores y topadoras, las máquinas de aserrado con nuevas tecnologías y la obtención de láminas para terciados, dieron un fuerte impulso al sector forestal deter- Aprovechamiento forestal de cedro en las Yungas de Argentina minando un período de auge económico. Luego, durante las dos últimas décadas del siglo XX, la actividad sufre una gran recesión, fundamentalmente, por cuestiones macroeconómicas y disminución de los volúmenes maderables, donde el cedro presenta la situación más crítica. En todo el período histórico mencionado, el cedro fue la primera especie con la cual se realizó un aprovechamiento, que llegó hasta prácticamente el agotamiento de los individuos con excelentes aptitudes forestales (fustes rectos, cilíndricos y largos, diámetros entre 50 y 80 cm). Desde mediados del siglo XX a la fecha, se escucharon opiniones o referencias al agotamiento o devastación de la selva y bosque con cedro. Sin embargo, existen todavía algunos rodales y ejemplares de excelente porte forestal en sitios privados de difícil acceso o en Reservas y Parques Nacionales. Otros factores, como los desmontes para actividades agropecuarias, los incendios y la ganadería vacuna extensiva a monte, también contribuyeron a la drástica disminución de cedros en la región. Pautas operatorias entre dueños de tierras y propietarios de aserraderos Los productores forestales, que iniciaron el desarrollo del aprovechamiento de maderas nativas en el noroeste de Argentina, eran dueños de las tierras con bosques. Esta situación se mantuvo hasta que la escasez de madera los obligó a buscar materia prima en otras propiedades. Esto último dio lugar a los contratos de arriendo, operatoria que tomó mayor relevancia en las tres últimas décadas del siglo XX. En este tipo de operatoria a veces se establecen pautas de aprovechamiento (determinados lotes, ciertos cuidados en la apertura de picadas y apeo de árboles), entre los madereros y los dueños de la tierra con bosque. Normalmente el dueño del aserradero es quien se encarga de la construcción de vías de saca, de la corta, rodeo y transporte de la madera. Existe también la figura de contratistas de la industria forestal que realizan todas o algunas de las actividades del aprovechamiento (apeo, rodeo y transporte hasta los aserraderos). Foto 1. Viga de cedro labrada con hacha en la ciudad de Orán, Salta. Julio 1948. Foto Humberto Chagra. Entre las pautas operatorias más frecuentes entre los dueños de los aserraderos y los propietarios del bosque, pueden citarse: 145 Minetti, J. M. Derecho de monte o aforo: se fija entre las partes un precio por m3 de cedro y se establece un porcentaje en carácter de retribución al dueño del bosque, que varía entre un 25 y un 35% del valor presente en el canchón del aserradero. Este mecanismo es muy común en las áreas selváticas de las Provincias de Salta y Jujuy, donde ese porcentaje puede aumentar hasta un 40 % en los casos de rodales de cedro rosado. Derecho de monte o aforo al barrer: es una variante del modo anterior que consiste en fijar un único porcentaje independiente de las especies. Normalmente oscila entre un 15 y un 30 %, dependiendo de los tipos de madera o calidad del bosque. Este modo ocurre principalmente cuando los rodales de cedro son escasos. Alquiler de monte: se fija un monto mensual a modo de alquiler del bosque, tratándose de extraer el máximo de madera en el menor tiempo posible. Por ejemplo, en los alrededores de la ciudad de Orán, Salta, en un bosque de condiciones relativamente buenas, durante el año 2000 los valores de alquiler oscilaban entre $ 1.500 y $ 2.000 mensuales para toda la finca (las cuales pueden tener entre 1.000 a 5.000 ha). Estos datos corresponden a estadísticas del año 2000 cuando un peso argentino equivalía a un dólar estadounidense. Esta forma es frecuente en la zona de ecotono entre Yungas y Bosque chaqueño o en áreas a desmontar con fines agrícolas. Por ejemplo, en Tartagal y hacia el este de la Ruta Nacional Nº 34, hasta el año 2000, el valor del alquiler podía oscilar entre los $ 400 y $ 600 por mes para toda una finca, dependiendo de 146 las especies y calidad del monte. Esta operatoria irracional puede estar relacionada a explotaciones ilegales. Por unidades: los pilotes o trocillos y despuntes se comercializan por unidades de piezas con medidas estándares. Excepcionalmente se realizan operatorias en forma de cargas, lo cual es frecuente en especies de maderas duras como quebrachos o algarrobos. Para el cedro, normalmente se cubican estas piezas igual que los rollos. Sin embargo, el valor comercial suele ser menor por tratarse de pequeñas dimensiones que corresponden a los despuntes o tal vez alguna rama recta. Sistemas de aprovechamiento El aprovechamiento forestal en las Yungas se realiza en forma estacional en la época seca, desde mayo hasta noviembre. De esta forma, los aserraderos deben acopiar madera para tener stock durante los meses lluviosos del verano. Como sistema de aprovechamiento entendemos a las operaciones que se ejecutan (idealmente en el marco de un plan de manejo forestal) para lograr la extracción de los rollos o productos maderables del bosque y el transporte hasta los centros de acopio o industrialización. La sumatoria de actividades del aprovechamiento forestal que se desarrollan dentro del bosque y las interrelaciones socio-productivas entre los distintos actores que intervienen, se denomina obraje. Para los cedros, desde mediados del siglo XX hasta nuestros días, el sistema de aprovechamiento típico Aprovechamiento forestal de cedro en las Yungas de Argentina consiste en el marcado de los árboles que superan los diámetros mínimos de corta, el apeo, desrame, trozado y despunte. Estas operaciones se realizan con motosierra. El diámetro a la altura del pecho (DAP) mínimo de corta para cedro en las Provincias de Salta, Jujuy y Catamarca es de 40 cm, mientras que en Tucumán puede ser de 35 o 40 cm (Overgaard 1975). Posteriormente, se realiza el rodeo de la madera, operación que consiste en el arrastre de los rollos o trozas con moto arrastradoras o tractores hasta los canchones de carga. Aquí normalmente se mide la madera y se carga en forma manual o con tractores provistos de elevadores o grúas a camiones o acoplados. Con los cedros, lo más frecuente es llevar los rollos enteros en camiones denominados localmente vigueros (Foto 2). madera. Este sistema se basa en la existencia de árboles individualmente valiosos que, por estar en áreas inaccesibles o no formar parte de rodales, quedaron fuera de la acción de los obrajes tradicionales. Esta modalidad de aprovechamiento no es estacional como la tradicional. Existen antecedentes de esta operatoria en otras regiones tropicales de América y el mundo, no sólo con cedro, sino también con otras especies de maderas valiosas (Lamprecht 1990). El rallado de la madera consiste en la obtención de tablas o tablones al pie del tocón de los árboles apeados, utilizando para tal fin una motosierra normalmente de espada larga y una guía que puede ser de madera o estar incorporada a la máquina como accesorio. Se obtienen tablones de 2 o 3 pulgadas de espesor, por 10 a 15 pulgadas de En la Alta Cuenca del Río Bermejo un ancho (según dimensiones de los rollos) y sistema rudimentario, o de pequeña es- por 2 a 3 m de largo, que son transportados cala de aprovechamiento, es el conocido a lomo de burro o por fuerza humana, a como obraje individual o rallado de la través de las picadas abiertas para llegar a los árboles. Esta operación es peligrosa, ya que normalmente exige el desplazamiento en condiciones de mucha pendiente, hasta llegar al borde de algún camino o sitio de acopio. Desde el punto de vista del rendimiento, esta modalidad es extremadamente ineficiente, ya que sólo se aprovecha una tercera parte del volumen total de los Foto 2. Camión viguero con tres rollos de cedro en Orán. Agosto fustes. El rendimiento 2003. Foto Madenort SRL. 147 Minetti, J. M. en volumen sólido de tablones con relación al de rollos es de sólo un 30 a 36 %, debido al espesor del corte de la motosierra, las condiciones incómodas de trabajo y la técnica en sí misma. En un principio, el sistema de rallado de la madera permitía el aprovechamiento de cedros en sitios de dificil acceso. Esto generó una fuerte presión sobre los rodales remanentes, que cumplían con la función de protección de laderas y cabeceras de cuencas. En muchos casos, las fuertes pendientes en las que se encontraban estos cedros hacían deslizar los árboles recién apeados, perdiéndose la madera en las quebradas o precipicios. CubicaciOn y clasificaciOn de la calidad de madera A partir del Proyecto “NOA II Inventario y Desarrollo Forestal del Noroeste Argentino 1975”, se obtuvieron datos significativos para el manejo de los bosques nativos de la región. Para el caso del cedro se elaboró una función de volumen, la cual sigue vigente y es utilizada por técnicos, investigadores forestales y grandes empresas que cuentan con inventarios forestales realizados con metodología estadística. La cubicación por lo común se realiza una vez apeados los árboles, y sólo excepcionalmente con árboles en pie. En las Yungas argentinas los rollos se cubican tomando en cuenta el diámetro en punta fina y el largo total. Esta operación suele hacerse normalmente en el bosque una vez rodeada la madera o, menos frecuentemente, en el canchón del aserradero. El diámetro citado es medido en la sección más corta de la punta fina del rollo, pasando por la médula del árbol y hasta el inicio de la corteza, descontándose el valor del ancho de la albura o blanco en uno o ambos extremos. En el caso de la madera muerta o seca naturalmente en el bosque (llamados palos campana), no se hace descuento. Este descuento por la albura es conocido como “blanco” y puede explicarse mejor con la siguiente figura (Fig. 1). Figura 1. Diagrama de cubicación tradicional (sistema de “un blanco”) en el norte de Salta. 148 Aprovechamiento forestal de cedro en las Yungas de Argentina Según se descuente en uno o en los dos extremos del diámetro, la forma de cálculo se denomina “un blanco” o “dos blancos” respectivamente, aunque lo común para cedro es lo primero. Si se observa algún defecto o problema sanitario sobre la sección en la punta fina del rollo, al diámetro medido con “un blanco” se le descuenta lo afectado por el problema de sanidad. El sistema de “un blanco” es una práctica errónea en el cálculo del volumen que subestima el valor real en un 30 a 35 % en el caso de los rollos de cedro. El nombre de “blanco” corresponde a la albura y su valor no sólo depende de la especie, sino también de la edad o dimensiones del árbol y del sitio. Considerando dimensiones, uniformidad del grano visible de la madera, agujeros, rajaduras y edad de corte, los rollos o rollizos, se pueden clasificar en cuatro clases (Tabla 1). La clase especial está en desuso y sólo hacía notar una diferencia de calidad maderable a favor de la especie cedro rosado respecto al cedro orán. La diferenciación entre las dos especies es un tema reciente, ya que durante mucho tiempo se las consideraba iguales, a pesar que los madereros distinguían las maderas fundamentalmente por el intenso color rojizo de la primera. Tabla 1. Clasificación de calidad de rollos o rollizos de cedro. Clase Características Especial Hace referencia a rollos de cedro rosado con las características de la clase “primera”. Representa la calidad de máxima excelencia muy apreciada para mueblería fina. Actualmente en desuso ya que son escasas las existencias maderables en las selvas y bosques del noroeste de Argentina. Primera Rollo de aspecto cilíndrico: recto y con poca diferencia entre los diámetros extremos. Diámetro en punta fina mayor de 40 cm y largo mayor de 6 m. No presenta rajaduras ni signos de enfermedad. Grano visible muy uniforme. Segunda Los diámetros y largos pueden ser menores a las clases anteriores y la forma no es tan recta o presenta una curvatura. Tercera Con más de una torcedura, de dimensiones menores o presenta otros defectos como perforaciones por galerías de insectos. 149 Minetti, J. M. EstadIsticas provinciales y comercio Las Provincias de Salta, Jujuy y Tucumán, llevan registros de extracción de rollos de cedro en m³ por año (Tabla 2). Estas estadísticas subregistran los valores de extracción, pues el circuito ilegal (informal) de esta especie hace que las cifras reales se ubiquen por encima de las oficiales en cada año considerado. Los equipos técnicos provinciales encargados de la regulación en la extracción forestal, no cuentan con medios para efectivizar el control de los planes de extracción ya aprobados y la mayoría de estos planes se realizan sin un inventario forestal exhaustivo o con rigor estadístico (Suárez 1998). A esto último se suma la fuerte influencia de la extracción de cedros en forma ilegal. El aprovechamiento ilegal de cedro se realiza por personas que extraen madera de cedro de fincas privadas o fiscales y la comercializan en forma de tablones directamente en las carpinterías. Esta operatoria suele denominarse en el norte de Salta y Jujuy como “robo hormiga de la madera” y por lo general sólo utiliza una motosierra y un carro tirado por tracción animal o un vehículo viejo. Para citar un ejemplo, en la ciudad de Tartagal gran parte de las carpinterías que trabajan con cedro no compran la madera en los aserraderos, sino que son provistos de materia prima por el circuito informal. Es frecuente ver por las rutas de Tartagal, Gral. E. Mosconi, Aguaray (Departamento San Martín, Salta), Palma Sola y El Fuerte (Departamento Santa Bárbara, Jujuy), carros tirados por caballos o vehículos viejos con 2 a 3 rollos de cedro. 150 Tabla 2. Extracciones de cedro en m³/año en las Provincias de Tucumán, Salta y Jujuy. Las celdas en blanco no significan ausencia de extracción, sino falta de datos. Año Tuc. Salta Jujuy 1981 - 1982 - 8.131 8.184 - 1983 - 10.487 - 1984 - 11.435 - 1985 - 7.495 - 1986 - 7.190 - 1987 - 8.714 - 1988 - 8.925 - 1989 - 1990 - 1991 - 1992 - 1993 - 1994 - 1995 - 1996 450 1997 - 1998 - 1999 336 2000 266 2001 179 2002 441 2003 - 2004 - 5.570 - 2.488 - 3.686 - 4.366 - 6.163 - 13.208 - 4.501 1.809 10.174 - 5.887 1.019 2.969 1.356 3.495 748 9.059 2.990 - - - - Aprovechamiento forestal de cedro en las Yungas de Argentina Productos y medidas usuales de comercializaciOn Zona de producciOn, centros fabriles y sitios demandantes La madera de cedro se usa principalmente para carpintería general, para la fabricación de muebles finos, aberturas, puertas, ventanas, chapas y terciados (a través de debobinado y faqueado), para molduras, revestimientos, construcciones y otros usos menores. Las zonas de producción de cedro corresponden a: Los productos usuales del cedro son (tomado y adaptado del trabajo de Suárez 1998): Tablas: suelen ser de dimensiones variables. Por ejemplo de más de 220 cm de largo y más de 6 pulgadas de ancho, de 1, 1½, 2, 2½ y 3 pulgadas de espesor. Batientes: pueden ser de 2 x 3; 2 x 4; 1 ½ x 3 y 1½ x 4 x 60 a 22 cm. Se las usa en aberturas. Alfajías: de ½ x 1½ x 40 a 90 cm. Se las usa para remanufactura de pequeñas piezas, para postigones y persianas. Rollizos aserrados en sándwich: son los rollos aserrados en tablas de 1 o 2 pulgadas de espesor pero con corteza, lo cual determina que los rollos conserven la forma a pesar de estar divididos en varias tablas. Normalmente las tablas se separan con finos listones de madera, para permitir el aireado y secado de toda la madera. Provincia de Salta. Departamento Orán: en las sierras de Santa María, Abra Grande y áreas de influencia de las ciudades de San Ramón de la Nueva Orán, Pichanal y Urundel, que a la vez actúan como centros de industrialización y/o aserrado (Foto 3). Departamento Iruya: en Finca Santiago y Finca La Candelaria del Municipio de Isla de Cañas. Departamento Santa Victoria: Municipio de Los Toldos. Esta localidad se encuentra a más de 120 km de distancia de la ciudad de Orán y tiene acceso en vehículos sólo por territorio boliviano. Departamento San Martín: en las sierras de San Antonio y Aguarague, en los seis Municipios que integran el citado Foto 3. Rollo de cedro rosado en aserradero y fábrica de aberturas en la ciudad de Orán. Julio 2002. 151 Minetti, J. M. Departamento (Embarcación, Gral. Ballivián, Gral. Mosconi, Tartagal, Aguaray y Profesor Salvador Mazza). La extracción en los Departamentos Metán, Rosario de la Frontera, Anta y Capital es de menor cuantía volumétrica que en el norte de la provincia. Provincia de Jujuy. Departamentos Ledesma, Santa Bárbara, Capital, San Pedro y El Carmen. Los centros con mayor concentración de aserraderos y carpinterías son las localidades de San Pedro, Caimancito, y en menor medida Libertador General San Martín y San Salvador de Jujuy. Algunos de los aserraderos de la Provincia de Jujuy también trabajan con maderas provenientes de la Provincia de Salta (por ejemplo en las ciudades de Libertador Gral. San Martín y San Pedro). Provincia de Tucumán. Departamentos Burruyacu, Tafí Viejo, Famaillá, Monteros, Chicligasta, Alberdi y La Cocha. Existen centros de transformación primaria en San Miguel de Tucumán y localidades cercanas a las áreas de extracción que en la mayoría de los casos son aserraderos rudimentarios. Los principales centros de consumo son las capitales provinciales del centro y norte del país y otras ciudades grandes. Las provincias que más madera de cedro demandan son Santa Fé, Córdoba y en menor medida Buenos Aires y Mendoza. Los consumidores son fábricas de muebles y aberturas, carpinterías o corralones mayoristas. Generalmente el cedro se asierra casi totalmente en la zona. Como excepciones pueden citarse los ejemplares selectos 152 para faqueo, operación no realizada por ninguna industria en Salta o en Jujuy, que demandan rollos enteros. El faqueo consiste en la obtención de chapas de madera por cortes transversales lo que genera un valor del producto superior al de las chapas obtenidas a través del debobinado de los rollos. Prácticamente en la actualidad, no existe oferta de rollos para faqueado. El transporte se realiza exclusivamente en camiones que utilizan las Rutas Nacionales Nº 34 y 9. Conclusiones La tala selectiva que sufrieron los cedros en las Yungas argentinas llevó a sus especies a una situación crítica, que difícilmente pueda revertirse. La intensidad de las explotaciones, los escasos conocimientos técnicos silvícolas, el ineficiente control de los organismos oficiales competentes y la escasa transparencia del mercado forestal en la región, son condicionantes del estado actual de las especies de cedro en el noroeste de Argentina. Debido a esto, es imprescindible realizar planes de manejo forestal que además de cumplir con las normas vigentes, tengan como fin lograr la sustentabilidad del recurso y de la actividad. Existen acciones que deben encararse en forma urgente, a través de alianzas públicas y privadas, tales como frenar los aprovechamientos ilegales, respetar los rodales que protegen las cabeceras de cuenca o márgenes de cursos de agua, seleccionar y preservar rodales e individuos como semilleros o para futuros planes de mejoramiento forestal. Aprovechamiento forestal de cedro en las Yungas de Argentina Por último, una medida importante para contribuir a salvaguardar el futuro de los cedros de las Yungas es la reforestación a través de enriquecimientos o macizos con fines comerciales, aliviando la presión sobre las formaciones nativas. En este sentido, se están realizando esfuerzos y existen productores forestales que, están incursionando en la reforestación con cedros en las Provincias de Salta, Jujuy y Tucumán, ante la imposibilidad de obtener materia prima de los bosques nativos. Además, debe generarse mejor material de divulgación, más información científica sobre el manejo de cedro y difundirse con mayor ímpetu las experiencias y conocimientos existentes en organismos como el INTA, las Universidades, las Secretarías de Medio Ambiente provinciales (con oficio de Direcciones de bosques) y la Secretaría competente del Gobierno Nacional. BibliografIa Lamprech, H. 1990. Silvicultura en los Trópicos: los ecosistemas forestales en los bosques tropicales y sus especies arbóreas; posibilidades y métodos para un aprovechamiento sostenido. GTZ. Alemania. Pp. 335. Overgaard, J. 1975. Aprovechamiento y Transporte Forestal en el Noroeste Argentino. NOA II Forestal. IFONA, UNDP y FAO. Salta. Saravia Toledo, C. 1994. Limitantes de la producción forestal de la región NOA. Mimeo. Fundación para el Desarrollo del Chaco, Salta. Suarez, C. 1998. Estudio del Mercado Argentino de Maderas en Rollos y Aserradas de algunas Especies Seleccionadas. Mimeo. Orán, Salta. 153 Ecología y producción de cedro (género Cedrela) en las Yungas australes. Pacheco, S. y A. Brown (Eds.). LIEY-ProYungas. Argentina. pp. 155-160. 2006. Producción de plantines de cedro en vivero Monteverde, D. Fundación ProYungas CC 34 (4107), Yerba Buena, Tucumán, Argentina. Dirección actual: Piedrabuena 1952 (1846), José Mármol, Bs.As., Argentina. [email protected] En este capitulo se expone la experiencia práctica desarrollada por la Fundación ProYungas en la producción de plantines de cedro durante los años 1999 a 2004. Se describen los pasos a seguir desde la selección de árboles semilleros hasta la implementación de la plantación. Primero se detallan los caracteres a tener en cuenta en la selección de individuos para la cosecha de acuerdo a los objetivos de la plantación que se desea realizar y la técnica de colecta de semillas. Luego se presentan los métodos para el tratamiento y acondicionamiento de las semillas para su siembra, la técnica de producción del plantín y las pautas para realizar plantaciones con estas especies. This chapter exposes the experience developed by ProYungas Foundation in Cedrela seedlings production during the 1999-2000 period. I present the methodology to implement a successful plantation. First, I describe important tree variables for mother plant selection depending on the specific goals of the plantation, and the appropriate technique for seed collection. Second, I present the methods for seeds treatment and storing before planting them, seedlings production techniques, and guidelines to create and maintain plantations. 155 Monteverde, D. INTRODUCCION Este trabajo ofrece una guía práctica para la producción de plantines de cedro basada en la experiencia adquirida en el marco de diversos proyectos realizados por la Fundación ProYungas entre 1999 y 2004. Entre ellos, se encuentran los Planes de Revegetación de Valle Morado (Shell CAPSA) y Gasoducto Nor Andino Argentina, la implementación de la plantación Experimental de Valle Morado (Shell CAPSA) y los viveros comunitarios de San Andrés, Los Naranjos (“Proyecto de diversificación productiva con comunidades indígenas”) y Los Toldos. La producción de plantines de Cedrela lilloi o cedro coya se hizo en Los Toldos y la producción de Cedrela balansae o cedro orán se hizo en Valle Morado. Las técnicas de producción aplicadas fueron iguales, por lo que se utilizará el nombre cedro para hacer referencia a ambas especies. SelecciOn de Arboles semilleros Para producir plantines de cedro para plantaciones a campo, es importante realizar una selección lo más intensa posible de los árboles semilleros. En la mayoría de los casos, sólo puede hacerse una selección fenotípica de los individuos, ya que se realiza sobre rodales naturales. Por lo tanto, se desconoce qué proporción de la expresión del carácter es de origen genético. Además, en la selección en rodales naturales no se pueden elegir individuos en base a su crecimiento (que es uno de los factores más importantes en la silvicultura), ya que no se sabe con precisión la edad 156 de cada individuo ni qué efecto pudo haber tenido el sitio sobre el mismo. En la selección fenotípica los caracteres que deben considerarse son los siguientes: Forma del individuo: se seleccionan individuos que presenten pocas ramificaciones y características adecuadas de rectitud, altura de fuste y desarrollo de la copa. Sanidad del individuo: se seleccionan individuos que no presenten evidencias de quemaduras de sol, áreas podridas en el fuste, signos de ataque de insectos o enfermedades en la copa, que afectan tanto a la calidad del fuste como al normal desarrollo del individuo. Calidad y cantidad de frutos: se seleccionan individuos que tengan producción abundante y continua de frutos a lo largo de varios años y que presenten una buena sanidad. La selección de los individuos puede hacerse considerando la valorización de los caracteres en forma cualitativa o cuantitativa, para lo cual habría que desarrollar un sistema de clasificación de cada uno de los caracteres mencionados. De acuerdo al objetivo de la plantación deben priorizarse los caracteres para la selección de los individuos. Por ejemplo, para la producción de madera de calidad, deben elegirse individuos con buena forma, mientras que para plantaciones destinadas a la producción de semillas, debe priorizarse la calidad y la cantidad de frutos como carácter para la selección. Producción de plantines de cedro en vivero Epoca y forma de cosechar los frutos Entre la ultima semana de julio y la primera de agosto debe hacerse un monitoreo casi diario de los individuos de los cuales se cosecharán las semillas. Al observarse la dehiscencia de las primeras cápsulas debe realizarse la cosecha, ya que es probable que en menos de una semana las cápsulas se abran totalmente y diseminen las semillas perdiendo la posibilidad de colectarlas. No es conveniente cosechar temprano para “asegurar la cosecha” ya que pueden obtenerse semillas que no estén completamente desarrolladas. La cosecha de las cápsulas puede hacerse con una tijera de poda larga, aunque en la mayoría de los casos estarán fuera del alcance y habrá que subir al árbol para obtenerlas. Uno de los métodos más prácticos es utilizar una escalera de soga con peldaños de madera, que sea fácil de transportar (es probable que haya que internarse en el interior de la selva para llegar al individuo seleccionado). Para “colgar” la escalera al árbol, se debe arrojar la soga con una piedra atada a uno de los extremos por la horqueta más accesible del árbol. Una vez en el árbol, el operario deberá moverse por ella asegurado con un arnés y cosechar los frutos con la tijera de podar (Foto 1). Acondicionamiento de las semillas Una vez obtenidas las cápsulas, deben extenderse al sol para que se produzca la apertura total de las mismas, teniendo la precaución de no someterlas a fuerte insolación (Foto 2). Lo más conveniente es Foto 1. Cosecha manual de semillas de cedro orán (Cedrela balansae) utilizando una escalera de soga. exponerlas al sol a la mañana temprano y durante la tarde colocarlas bajo media sombra. Una vez que se han abierto totalmente las cápsulas se terminan de extraer manualmente las semillas que hayan quedado pegadas a las mismas. El resultado es una mezcla de semillas vanas (sin embrión en desarrollo) fácilmente removibles, con semillas buenas de mayor peso. Las mismas pueden ser separadas tomando puña157 Monteverde, D. Siembra y cuidados en el almAcigo Foto 2. Cápsulas de cedro orán (Cedrela balansae) extendidas al sol para que se produzca la apertura luego de la cosecha. dos de semillas y soplando de cerca para que “vuelen” las vanas. Por cada cápsula hay entre 15 y 20 semillas viables. PreparaciOn del almAcigo Para favorecer y homogeneizar el proceso de germinación se pueden sumergir las semillas en agua por 12 horas y luego dejarlas orear por 6 horas antes de sembrarlas. En este momento, si se considera conveniente, se puede aplicar algún fungicida para prevenir el ataque de hongos en el almácigo. La siembra se realiza al voleo en agosto - septiembre, a razón de 50 g/ m². Una vez distribuidas sobre la cama de siembra, se las tapa con una fina capa de mantillo y se las compacta presionando suavemente con una tabla para mejorar el contacto de la semilla con el sustrato. El almácigo debe estar ubicado bajo media sombra de 70 - 80 % con un túnel de plástico para evitar los excesos de humedad que puedan provocar las precipitaciones (Foto 3). Se debe regar diariamente con poca cantidad de agua, de manera que el sustrato permanezca húmedo sin que se produzca acumulación de agua. Pueden La cama de siembra del almácigo se prepara con mantillo que se “solariza” previamente para disminuir la cantidad de hongos y bacterias patógenas que puedan afectar la semilla. Para esto, se debe preparar un cordón de mantillo de 1 metro de ancho, de hasta 30 cm de profundidad, por el largo que se desee. Se tapa con un plástico transparente para que en el interior se alcancen altas temperaturas durante 45 a 60 días en épocas de alta insolación (primavera). Una vez solarizado, se procede a su tamizado para obtener un material fino y homogéneo que se distribuye en el Foto 3. Almácigo de cedro orán (Cedrela balansae) en almácigo. Valle Morado, Salta. 158 Producción de plantines de cedro en vivero realizarse aplicaciones de Almacigol para prevenir el ataque de dumping-off, pero en general solarizando el sustrato y teniendo un buen control de la humedad del almácigo es suficiente. La germinación puede comenzar entre los 10 y los 25 días de realizada la siembra. Repique la altura del cuello de la planta para lograr que las raíces queden en contacto con el sustrato (Foto 4). Cuidados en canchas de crIa y rustificaciOn Las canchas de cría deben estar ubicadas bajo media sombra del 70 - 80 % durante los primeros 45 días de haber repicado los plantines. A medida que éstos se van desarrollando y el tallo comienza a lignificarse, se puede correr la media sombra en días frescos y nublados o durante las mañanas para comenzar a rustificarlos. Durante esta etapa la incidencia de la mariposita del brote de las Meliáceas (Hypsipyla grandella) no es seria, sólo algunos plantines pueden ser atacados. Para asegurar la rustificación completa, durante los últimos 20 o 30 días, es conveniente exponer a los plantines a cielo abierto para someterlos a condiciones similares a las del campo. El repique se debe realizar cuando las plántulas desarrollan su primer par de hojas verdaderas o tienen alrededor de 5cm de altura. Generalmente, esto ocurre entre 30 y 45 días después de la siembra. Debe realizarse en días frescos y nublados si es posible, o bien temprano en la mañana. Si los días son calurosos es recomendable trabajar a última hora de la tarde. Los bolsines utilizados deben ser de 20-25 cm de alto y de 6-7 cm de diámetro de boca y deben estar llenos con una mezcla 2:1 de tierra y mantillo. Previo al repique de plantines, es conveniente realizar un riego a los bolsines y hacerles perforaciones en la base. El transplante se debe hacer con cuidado, evitando producir daños en las frágiles raíces de las plántulas. Para ello es conveniente remover desde abajo el almácigo a fin de aflojar las raicillas y mantener las plántulas en un recipiente con agua mientras se realiza el repique. Una vez introducidos en el bolsín se aplica una Foto 4. Repique de plántulas de cedro orán (Cedrela balansae) presión suave de la tierra a en Valle Morado, Salta. 159 Monteverde, D. PlantaciOn La época recomendable para realizar la plantación es a fines del verano o principio de otoño (los plantines tienen de 6 a 7 meses de edad si se comenzó el almácigo en septiembre) cuando las precipitaciones son abundantes, el cielo se mantiene generalmente cubierto y las temperaturas ya no son tan elevadas. Si es posible, la plantación debe efectuarse durante días nublados y/o con llovizna. Los días muy calurosos y con fuerte insolación pueden producir quemaduras en el cuello de la planta, pro- 160 vocando en algunos casos la muerte del ejemplar. El suelo debe estar libre de malezas y con dos rastreadas para facilitar la operación de poceado y la instalación del plantín. En el caso de que heladas severas afecten a los plantines, se puede hacer un manejo de los rebrotes en la temporada siguiente, eligiendo el más vigoroso y de mejor forma para su conducción. Durante los primeros meses es importante realizar un buen control de malezas para evitar que compitan con el plantín. de propagación deYungas Cedrela in-vitro EcologíaExperiencias y producción preliminares de cedro (género Cedrela) en las australes. Pacheco, S. y A. Brown (Eds.). LIEY-ProYungas. Argentina. pp. 161-170. 2006. Experiencias preliminares de propagación de Cedrela in vitro Enrico R. J.1 y E. E. Enrico 2 Facultad de Ciencias Naturales e Instituto Miguel Lillo-UNT, Miguel Lillo 205 (4000), S. M. de Tucumán, Argentina. 2 Laboratorio de Investigaciones Ecológicas de las Yungas (LIEY, UNT). 1 [email protected] Usualmente la propagación de las especies nativas de Cedrela es a través de semillas. Esto limita la producción de plantines, aumenta la variabilidad genética y dificulta el establecimiento de plantaciones en base a material seleccionado. El objetivo de este trabajo fue desarrollar un sistema de propagación in vitro de Cedrela balansae. Para esto se trabajó con estacas uninodales, que produjeron múltiples brotes en medio MS adicionado con 0,3 mg/L de BAP y 0,01 mg/L de ANA. La combinación con la auxina indujo el enraizamiento de los explantes. También se logró la proliferación a partir de meristemas, usando MS al 50 %, con 0,5 mg/L de BAP. Cedrela native species are usually propagated by seed. This limits seedling production, increases genetic variability and makes the establishment of selected stands more difficult. The aim of this project was to develop an in vitro propagation system for Cedrela balansae. We used soft wood cuttings, which produced the highest number of shoots on MS medium added with 0,3 mg/L of BAP and 0,01 mg/L of ANA. Auxin addition induced the rooting of soft wood cuttings. The proliferation was also achieved starting from meristems, using MS medium at 50 % strength, with BAP 0,5 mg/L. 161 Enrico, R. J. y E. E. Enrico introduccion En la historia de la domesticación de los vegetales, las especies forestales no han recibido tanta atención como los cultivos agrícolas, debido probablemente a su gran tamaño y sus generaciones relativamente largas. Su reproducción se realiza casi exclusivamente a partir de semillas, lo que ocasiona una gran variabilidad genética que hace difícil su mejoramiento. La propagación vegetativa es un método de reproducción usado por los productores forestales y ha tenido un papel importante en la multiplicación de árboles de “elite” (Ivanova 1981). En el caso de las estacas, se ha observado que las mismas pierden su capacidad de enraizar a medida que el árbol de origen es más viejo. Por otro lado, es necesario que los árboles usados para su reproducción sean lo suficientemente adultos para expresar todo su potencial genético. Sweet (1973) observó que al utilizar ramas maduras para enraizar hay problemas plagiotrópicos. Esto, sumado al hecho de que una estaca produce únicamente una planta y que los volúmenes necesarios de materiales de siembra para establecer plantaciones son altos, indica que esta técnica no es la más eficiente para satisfacer a corto plazo, las demandas de plantines seleccionados. Mediante el cultivo in vitro es posible obtener un número elevado de plantas por genotipo, reducir el tiempo de multiplicación, aumentar la sanidad del material propagado, multiplicar rápidamente una variedad de la cual existen pocos individuos y facilitar el transporte de material de 162 un país a otro debido a las menores restricciones fitosanitarias. Los primeros trabajos de cultivo de tejidos en Cedrela fueron iniciados por Maruyama et al. en 1989, quienes a partir de ápices lograron obtener plántulas completas de cedro. En 1997, Maruyama et al. obtuvieron semillas artificiales encapsulando los ápices obtenidos por micropropagación. En la actualidad no existen protocolos desarrollados específicamente para las especies de Cedrela nativas del noroeste de Argentina (C. lilloi, C. balansae y C. saltensis) y la propagación comercial se realiza en base a semillas. El objetivo de este trabajo es establecer los protocolos de micropropagación para Cedrela balansae. metodos Se trabajó con material vegetal procedente de plantines y ejemplares adultos de Cedrela balansae cultivados en la Localidad de Horco Molle, Provincia de Tucumán, Argentina. Las experiencias se realizaron entre los meses de octubre y enero. Preparación de la planta madre y elección del tipo de explante Para reducir el número de microorganismos sobre la superficie vegetal, se empleó un régimen de irrigación mínimo necesario, humedeciendo el sustrato sin salpicar. Además, se aplicó sobre el material vegetal un fungida sistémico (Benomyl al 1,5 %) y un antibiótico (Agrimicina al 1 %), cada 15 días. Para el establecimiento de estos protocolos se utilizaron diferentes tipos de explantes: Experiencias preliminares de propagación de Cedrela in-vitro 1- Microestacas o segmentos de tallos uninodales, conteniendo una yema axilar, provenientes de: - plantines de vivero de 3 - 4 meses de hasta 25 cm de altura, - plantines de vivero de 1 - 2 años de edad, - árboles de 8 años de edad. Las estacas leñosas de 1 a 3 cm de diámetro se colocaron en cámaras húmedas con 80 a 100% de humedad relativa y baja intensidad lumínica (DDFF de 74 µmol. m-2.seg-1). Fueron cubiertas parcialmente por sustrato formado por tierra de crecimiento del árbol donante. Este sistema se emplea para inducir la producción de brotes con características juveniles. 2-Meristemas (0,3 - 0,4 mm). Establecimiento del cultivo e inducción a la brotación En esta etapa los principales procesos a controlar fueron la desinfección superficial de los explantes y la selección de los medios de cultivo, para que los explantes pudieran responder bajo las condiciones in vitro. Para lograr el establecimiento in vitro de los diferentes explantes se probó el desinfectante hipoclorito de sodio al 0,6; 0,9; 1,2 y 1,8%, con diferentes tiempos de exposición (10, 15 y 25 minutos). Posteriormente, se enjuagaron seis veces con agua destilada estéril. En el caso de las microestacas provenientes de plantines de 1-2 años de edad, pre- vio a los ensayos, se realizó un lavado de toda la rama con jabón en polvo, sin contenido de enzimas, en agitación durante 10 minutos y luego, un enjuague con agua corriente, durante 20 minutos, con el fin de eliminar agentes contaminantes del lugar de origen. El ensayo de desinfección consistió en la aplicación de hipoclorito de sodio al 1,2 % combinado con etanol 70 %, en dos tiempos de acción (30 o 60 segundos para el alcohol y 15 o 20 minutos para la lavandina) (Tabla 1). Todas las soluciones estaban adicionadas con tween 80, 1 gota cada 100 ml, para disminuir la tensión superficial y aumentar el contacto del agente desinfectante con los explantes, permitiendo un mayor efecto. En cuanto a los medios de cultivo, se trabajó con MS (Murashige y Skoog 1962) como formulación salina básica, con distintas diluciones y adicionado de sacarosa al 30 g.l-1 como fuente de carbono. En todos los casos, el pH de los medios fue ajustado a 5,8 empleando soluciones de KOH y HCl, antes de la adición del agar Sigma A - 1296 (7 g.l-1). Los medios de cultivo fueron distribuidos en tubos de vidrio 15 x 90 mm, colocándose 5 ml de medio en cada uno. Los recipientes fueron esterilizados en autoclave a 1,5 Kg.cm-2 de presión y 120º C, durante 20 minutos. Para inducir a la brotación se adicionó al medio de cultivo una citocinina sola o en combinación con una auxina. Se realizaron cuatro ensayos: Ensayo 1: se establecieron meristemas y 163 Enrico, R. J. y E. E. Enrico estacas uninodales en medio MS al 50 %, con 30 g.l-1 de sacarosa, 100 mg.l-1 de ácido ascórbico, como antioxidante y 0,5 mg.l-1 de BAP (6-bencilaminopurina). explante cultivado. Se realizaron dos repeticiones de 10 recipientes por tratamiento. Ensayo 2: se evaluó el establecimiento de segmentos nodales conteniendo una yema axilar, provenientes de plantines de vivero de 1 a 2 años de edad. Los medios utilizados corresponden a la formulación MS a la mitad de su concentración estándar, con 30 g.l-1 de sacarosa, 100 mg.l-1 de ácido ascórbico y concentraciones variables de BAP, combinadas o no con 0,5 mg.l-1 de ANA (ácido naftalen-acético) (Tabla 2). Preparación de la planta madre y elección del tipo de explante RESULTADOS El cultivo en cámara húmeda de estacas leñosas provenientes de árboles adultos mostró resultados positivos en la producción de brotes para iniciar el cultivo in vitro (Foto 1). Ensayo 3: se utilizó el mismo medio básico que en el ensayo 2, con diferentes combinaciones de citocinina-auxina (Tabla 3). Ensayo 4: se probaron diferentes concentraciones de la formulación salina básica de MS (12,5; 25; 50 y 100 %), con 30 g.l-1 de sacarosa, 0,3 mg.l-1 de BAP, 0,01 mg.l-1 de ANA. En todos los casos, los recipientes cultivados fueron tapados con Resinite AF 50 (Casco S.A.C Company Bs. As.). Los explantes fueron incubados a temperatura constante (27 ± 2ºC) y a una densidad de flujo fotónico (DDFF) de 116 µmol.m-2.seg-1 y 14 hs de fotoperiodo, empleando lámparas fluorescentes de tipo “luz día”. Los meristemas se incubaron previamente en oscuridad, durante 15 días, para evitar la oxidación en la fase de establecimiento, y posteriormente en luz, en igual condiciones a las antes citadas. Para el análisis de los datos, se tuvieron en cuenta el número de hojas y la longitud de los vástagos desarrollados a partir de cada 164 Foto 1. Producción de brotes con caracterís- ticas juveniles a partir de estacas leñosas de ejemplares adultos de campo, mediante el empleo de cámara húmeda. Establecimiento del cultivo e inducción a la brotación Desinfección. Dado que se trata de una especie leñosa, la desinfección superficial de los tejidos para ser cultivados resultó particularmente difícil. En los primeros ensayos in vitro, la contaminación fue cercana al 100 %, impidiendo el desarrollo del explante. Experiencias preliminares de propagación de Cedrela in-vitro Cuando se utilizó sólo hipoclorito de sodio como agente desinfectante, se observó que, independientemente del tiempo de acción, en concentraciones por debajo del 1,2 %, la infección superaba el 50 %. Al elevar la concentración al 1,8 %, se observó fitotoxicidad. El tratamiento que presentó el menor porcentaje de infección y que ocasionó el menor daño a los tejidos fue el de hipoclorito de sodio al 1,2 % de concentración. Al probarlo en diferentes tiempos de acción, se obtuvieron los siguientes porcentajes de infección: con 10 minutos, 37,5 %; con 15 minutos, 30 % y con 25 minutos, 25 %. A pesar de ser este último el porcentaje más bajo de infección, la exposición más prolongada provocó en algunos explantes síntomas de fitotoxicidad. En las microestacas provenientes de brotación en cámara húmeda, los porcentajes de contaminación estuvieron cercanos al 90 %. Cuando se desinfectaron microestacas provenientes de plantines de 1-2 años de edad mantenidos en vivero, se observó que al aumentar el tiempo de exposición al alcohol, disminuía notablemente el porcentaje de infección. El mejor tratamiento resultó la combinación de etanol 70 % durante 60 segundos, seguido de hipoclorito de sodio al 1,2 % por 15 minutos (Tabla 1). Inducción a la brotación. Cuando se trabajó con segmentos uninodales de plantas jóvenes (Ensayo 1), se observó brotación superior al 50%. Mientras que, en el cultivo de meristemas (Ensayo 1), se logró el establecimiento en un 33 % de los tubos Tabla 1. Porcentajes de infección de las mi- croestacas provenientes de plantines de vivero, empleando como desinfectantes alcohol 70 % e hipoclorito de sodio con 1 gota de Tween 80 para 100 mL. Hipoclorito % Trat. Etanol 70 % de sodio 1,2 % infectados (Seg.) (Minutos) 1 60 15 15,00 2 30 15 79,07 3 60 20 25,74 4 30 20 85,00 cultivados. En ambos casos, inicialmente se observó crecimiento satisfactorio, con despliegue de hojas color verde intenso, pero al transcurrir el tiempo, comenzó a evidenciarse vitrificación hasta la muerte posterior del explante. El antioxidante empleado mostró ser efectivo, registrándose bajos porcentajes de oxidación. Cuando se trabajó con segmentos nodales conteniendo una yema axilar provenientes de plantines de vivero de 1 a 2 años de edad (Ensayo 2), se obtuvieron altos porcentajes de respuestas del explante, en los medios que contenían solamente BAP (Tabla 2). Buscando la relación óptima citocinina/ auxina se decidió ampliar el rango de dosis, tanto de BAP como de ANA, como se muestra en la Tabla 3 (Ensayo 3). Se observó un alto porcentaje de brotación en el tratamiento 6, con 0,3-0,01 mg.l-1 de BAPANA, presentando además, producción de 165 Enrico, R. J. y E. E. Enrico Tabla 3. Efecto de diferentes concentraciones de BAP combinadas con ANA, adicionados a MS al 50 %, con 30 g.l-1 de sacarosa, sobre la inducción a brotación de segmentos nodales, con una yema axilar, provenientes de plantines de 1 – 2 años de edad de C. balansae. Trat. BAP (mg.l-1) ANA (mg.l-1) % Brotados 1 0 0 10 2 0,1 0 20 3 0,1 0,01 10 4 0,1 0,1 0 5 0,3 0 40 Entre nuestros resultados existe un 20 % de explantes cultivados en medios con combinación de citocinina/auxina, que enraizaron espontáneamente (Foto 3) sin cambiar de medio de cultivo. Esto indica- 6 0,3 0,01 70 7 0,3 0,1 10 8 0,5 0 40 9 0,5 0,01 20 Tabla 2. Inducción de brotación de segmen- 10 0,5 0,1 0 Foto 2. Múltiples brotes producidos en la eta- pa de iniciación del cultivo in vitro de Cedrela balansae, a las 4 semanas de cultivo. múltiples vástagos de aspecto más vigoroso (Foto 2). tos uninodales empleando diferentes concentraciones de BAP, combinadas o no con 0,5 mg.l-1 ANA, adicionados a MS al 50%, con 30 g.l-1 de sacarosa. Trat. BAP (mg.l-1) ANA (mg.l-1) % Brotados 1 0,1 0 70 2 0,5 0 40 3 1 0 50 4 0,1 0,5 20 5 0,5 0,5 0 6 1 0,5 10 166 ría la importancia del uso de auxinas en este tipo de organogénesis. A pesar de los bajos niveles de reguladores de crecimiento, los explantes mostraron bases ensanchadas (hiperplasia), razón por la cual se probaron diferentes concentraciones de MS (Ensayo 4). Transcurridos 35 días de cultivo, se observó que, empleando las diluciones más bajas de la formulación salina básica, aumentaba la oxidación y la deformación del explante (hiperplasia). La concentración estándar (MS completo) presentó el mejor aspecto de los vástagos, con el mayor número de hojas (Tabla 4). Experiencias preliminares de propagación de Cedrela in-vitro este tipo de material es a menudo imposible o muy difícil de desinfectar (Enjalric et al. 1988, Barghchi 1988, Badilla Mora et al. 1992). Por este motivo, para controlar las posibles infecciones, se buscó proteger un conjunto de plantas bajo condiciones de invernáculo o cámaras de crecimiento, pulverizándolas con fungicidas y bactericidas de amplio espectro. Foto 3. Vástago de C. balansae enraizado, después de los 60 días de iniciado el cultivo, en medios que contenían auxinas. Tabla 4. Efecto de la dilución de la formula- ción del medio MS sobre la brotación, la longitud de los vástagos y la producción de hojas durante la iniciación del cultivo in vitro de C. balansae. Todos los medios contienen 0,3mg. l-1 de BAP y 0,01mg.l-1 de ANA. Trat. Dilución Long. de MS (%) brote 1 100 2 50 3 25 4 12,5 Núm. % hojas brotados 0,66 5,3 37,5 1,4 2 20 0,54 3 40 0,86 4 30 DISCUSION Las contaminaciones microbiológicas más severas reportadas, se han originado a partir de explantes tomados directamente del campo de plantas sin protección, plantas infectadas o bien de plantas creciendo cerca del suelo. Existen antecedentes de que La adición de BAP, como único regulador de crecimiento en el medio de cultivo, ha provocado inducción de brotación en otras especies de Cedrela. Guevara Berger et al. (1992) obtuvieron brotación de yemas de C. tonduzii con la adición de 0,5 mg.l-1 de BAP en medio MS. Rodríguez et al. (2003) logran mayor calidad de explante y buenos niveles de multiplicación con la adición de 0,5 mg.l-1 de BAP. Perez et al. (2002) probaron en C. odorata con diferentes citocininas y obtuvieron el mayor número de brotes con la misma dosis de BAP y medio MS. En cambio para cotiledones de C. odorata, Andrade Torres et al. (2004), encuentran el mayor número de brotes con 0,25 mg.l-1 de BAP, en medio WPM (Loyd y Mc Cown 1980). Empleando MS como solución salina básica, a diferencia de C. tonduzii y C. odorata, en nuestros ensayos con C. balansae, los mayores porcentajes de brotación se obtuvieron con dosis menores de BAP. En contraposición con Valverde Cerdas et al. (1998) que no lograron organogénesis en C. odorata usando sólo citocinina, y sí cuando la combinan con auxinas, en el presente trabajo se obtuvo el mismo porcentaje de brotación tanto usando citocinina solamente como combinada con 167 Enrico, R. J. y E. E. Enrico auxina. Es conocido que la combinación de una citocinina con una auxina estimula la producción de nuevas yemas, activa la división celular y el desarrollo de tallos laterales respectivamente. En este trabajo el empleo de estos reguladores de crecimiento en el medio de cultivo, muestra que relación entre citocinina y auxina es más importante que su concentración en sí misma. La elección del uso o no de auxinas en la etapa de inducción se determinará con posterioridad, de acuerdo con los resultados que se obtengan para las siguientes etapas de la micropropagación (multiplicación y enraizamiento). En referencia a la solución salina, el MS fue el medio empleado para inducir brotación de plantines de semillas de C. odorata, Perez et al. (2002). En cambio para cotiledones de la misma especie el mayor número de brotes se encontraron en medio WPM (Andrade Torres et al. 2004). En otros trabajos no se han encontrado diferencias significativas entre el uso de MS y WPM, para el desarrollo de callo y raíces a partir de segmentos de hojas de C. fissilis, con combinaciones de BAP y ANA (Niella et al. 1996). Nuestros ensayos muestran que el BAP es una citocinina eficiente para inducir la producción y brotación de yemas in vitro en C. balansae, y que adicionada al medio MS produce vástagos de aspecto vigoroso. Los resultados obtenidos en nuestro trabajo indican la posibilidad de concretar un sistema de reproducción masiva y eficiente para esta especie. Se estudia el desarrollo de las siguientes etapas de la micropropagación a fin de establecer un protocolo completo. BIBLIOGRAFIA Andrade-Torres, A.; A. Alonso-López; A. Villanueva-Jiménez; M. Mata-Rosas; J. Murguía González y L. Solís-Ramos. 2004. Micropropagación de cedro rojo (Cedrela odorata l.) como herramienta para su manejo. http://www.colpos.mx/cveracruz/ SubMenu_Publi/Avances2004/Micropropagaci%F3n_de_cedro_rojo.html Badilla Mora, M.V.; D.N. Hidalgo; B.E. Guevara y G.O. Murill. 1992. Cultivo in vitro de plántulas de jaúl (Alnus acuminata). Tecnología en Marcha. 11: 3-9. Barghchi, M. 1988. Micropropagation of Alnus cordata (Loisel.) Loisel. Plant Cell, Tissue and Organ Culture 15: 233-244. Enjalric, F.; M.P. Carron y L. Lardet. 1988. Contamination of primary cultures in tropical areas: the case of Hevea brasiliensis. Acta Hort. 225: 57-65 Guevara-Berger, E.; N. Hidalgo-Dittel y O. Murillo-Gamboa. 1992. In vitro culture of Cedrela tonduzii. Tecnología en Marcha. 11: 10-16. 168 Experiencias preliminares de propagación de Cedrela in-vitro Ivanova, Z. 1981. Rapid vegetative propagation of conifers. Scientia Horticulturae 14: 347 - 355. Lloyd, G. y B. Mc Cown. 1980. Commercially-feasible micropropagation of moutain laurel, Kalmia latifolia, by use of shoot-tip culture. Comb. Proc. Int. Plant Propagators Soc. 30: 421-427. Maruyama, E.; K. Ishii; A. Saito y K. Migita. 1989. Screening of suitable sterilization of explants and proper media for tissue culture of eleven tree species of Peru-Amazon forest. J. Agric. Sci. 33: 252-261. Maruyama, E.; I. Kinoshita; K. Ishii y K. Ohba. 1997. Germoplasm conservation of the tropical forest trees Cedrela odorata L., Guazuma crinita Mart and Jacaranda mimosaefolia D. Don., by shoot tip encapsulation in calcium-alginate and storage at 12-25ºC. Plant Cell Reports. 16: 393-396. Murashige, T. y F. Skoog. 1962. A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiologia Plantarum 15: 53-58. Niella, P.; A.M. Noguera; J.L. Vera y F.O. Niella. 1996. Cultivo in vitro de guatambu blanco (Balfourodendron riedelianum) y cedro misionero (Cedrela fissilis). Yvyraretá 7 (7) Pérez, J.; J.F. Mesén-Sequeira; L. Hilje-Quirós y M.E. Aguilar. 2002. Development of a method for the micropropagation of selected genotypes of Cedrela odorata L. Opitimization of the multiplication phase. Revista Forestal Centroamericana 38. 67-71. Rodriguez, R; M. Daquinta; I. Capote; D. Pina; Y. Lezcano y J.L. González-Olmedo. 2003. Nuevos aportes a la micropropagación de Swietenia macrophylla X Swietenia mahogani (caoba híbrida) y Cedrela odorata (cedro) http://www.inca.edu. cu/otras_web/revista/CT24(3),%202003-INTERNET.htm Sweet, G.B. 1973. Effects of maduration on growth and form of vegetative propagules of radiata pine. N.Z. for Sci. 3: 191 - 210. Valverde-Cerdas, L. M. Dufour; V.M. Villalobos-Arambula. 1998. Organogénesis in vitro en Albizia guachapele, Cedrela odorata y Swietenia macrophylla (Fabaceae, Meliaceae). Revista de Biología Tropical 46: 225-228. 169 Ecología y producción de cedro (género Cedrela) en las Yungas australes. Pacheco, S. y A. Brown (Eds.). LIEY-ProYungas. Argentina. pp. 171-178. 2006. Crecimiento de Cedrela balansae en la plantación experimental de Valle Morado Horlent, M. y D. Monteverde Fundación ProYungas, CC 34 (4107), Yerba Buena, Tucumán, Argentina. Dirección actual: Piedrabuena 1952 (1846), José Mármol, Bs.As., Argentina. [email protected] La plantación experimental de Valle Morado cuenta con ensayos para evaluar el crecimiento y desarrollo de especies exóticas y nativas en el sector norte de las Yungas argentinas. Uno de los más relevantes es el ensayo de progenies de Cedrela balansae. El mismo cuenta con 42 familias provenientes de semillas obtenidas de individuos seleccionados en un área comprendida entre las localidades de Orán y Ledesma. La plantación se llevó a cabo en el año 2001 y los resultados que aquí se presentan corresponden a las mediciones de los años 2002 y 2003. Las características que se evaluaron hasta el momento fueron el crecimiento en diámetro y en altura, la forma general del individuo, el ataque de la mariposita del brote (Hypsipyla grandella), el quemado de la corteza y la mortalidad de los individuos. Los análisis realizados hasta el momento son preliminares, pero permiten observar la existencia de variabilidad entre las familias. Se destaca la importancia de continuar con este ensayo en el futuro y ampliarlo a otras localidades para contribuir al desarrollo forestal de la región. The experimental plantation of Valle Morado has dedicated several pieces of land to evaluate growth and development of exotic and native species from the north of the argentinian Yungas. One of the most important units concerns the Cedrela balansae progenies trials. It summons up 42 families coming from seeds collected from selected trees inside an area located between Orán and Ledesma towns. The plantation was established in 2001 and first data obtained in 2002 and 2003. Up to now the only variables that have been observed are the diameter and height growth, the tree shape, the attack of the Hypsipyla grandella, the burnt of the bark and the mortality rate. The analyses are preliminary but they show that a strong variability it exists among families, which is an important fact in improvement programs. It shows how important is to continue with these trials and extend them to other areas to promote the forestry development of the region. 171 Horlent, M. y D. Monteverde INTRODUCCION El conocimiento de los hábitos y el potencial de crecimiento de una especie, así como el desarrollo de técnicas silviculturales y la obtención de material genético superior, son aspectos fundamentales para difundir su cultivo y que resulte en una actividad económicamente rentable. Existen especies nativas con excelentes condiciones como recurso forestal. Sin embargo, no se cultivan a escala comercial por la incertidumbre que implica realizar una inversión de riesgo, con una especie de la que se desconoce su desarrollo en plantaciones. La plantación experimental de Valle Morado está ubicada en el Departamento de Orán, Provincia de Salta y fue establecida por Shell CAPSA y la Fundación ProYungas. El objetivo con el que se creó la plantación, fue el de aportar información sobre el desarrollo de especies nativas y así generar alternativas de producción de maderas valiosas en plantaciones comerciales. El grupo de especies nativas utilizado en los ensayos incluye alrededor de 20 especies, entre las que se encuentran: tipa blanca (Tipuana tipu), afata (Cordia trichotoma), pacará (Enterolobium contortisiliquum), tipa colorada (Pterogyne nitens) y cedro orán (Cedrela balansae). La plantación cuenta además, con ensayos de especies exóticas que se cultivan con éxito en otras regiones del país y del mundo y que presentan un alto potencial de producción en la región. En este grupo se trabajó con especies de eucaliptos (Eucalyptus y Corymbia), grevilea o roble 172 sedoso (Grevillea robusta) y con tona o cedro australiano (Toona ciliata). El cedro orán, por ser una especie nativa de madera valiosa de la que se desconoce su desarrollo en plantaciones, recibió especial atención al diseñar los ensayos. Se realizaron ensayos de progenies, con el objetivo de conocer las características genéticas de la especie y la capacidad de transmitir sus cualidades madereras de una generación a otra; ensayos de densidad donde se evaluaron prácticas silviculturales que permiten optimizar el desarrollo de la plantación, y ensayos donde se evaluó el crecimiento del cedro orán plantado como especie única y combinado con especies exóticas o nativas. En este capítulo se describen los resultados del ensayo de progenies. ensayos de progenies Los ensayos de progenies se establecen para evaluar la calidad genética de determinados individuos a través del cultivo de su descendencia. Para entender los ensayos de progenies hay dos conceptos importantes para definir, el fenotipo y el genotipo. Por fenotipo se entiende la apariencia externa de un individuo, que depende tanto de su constitución genética, como de las características ambientales en las cuales se desarrolla. Mientras que el genotipo se refiere al conjunto de genes que residen en los cromosomas del núcleo de cada célula y no puede observarse directamente, sino sólo a través de pruebas bien elaboradas (Barrett 1980, Zobel y Talbert 1994). Mediante los ensayos de progenie, pueden diferenciarse a los progenitores con una Crecimiento de Cedrela balansae en la plantación experimental de Valle Morado superioridad fenotípica consecuencia de haber crecido en un buen ambiente, de aquellos que presenten una superioridad genotípica. Conocer el genotipo es importante, ya que es lo que realmente se transmite de generación en generación. A partir de los ensayos de progenies es posible seleccionar individuos con una constitución genética superior, que aseguren una provisión de semillas de mayor calidad para la realización de plantaciones comerciales. Todas las especies exóticas que se cultivan en nuestro país, como los pinos y eucaliptos, han pasado por un proceso intenso de selección genética. Por esta razón presentan generalmente gran uniformidad, buena forma y rendimientos superiores a los que se obtienen con especies que no han sido mejoradas. Para realizar el mejoramiento de una especie, se deben identificar las características que se quieren mejorar y el destino del producto (producción de pulpa para papel o producción de madera para muebles). Dentro de las características a mejorar, la tasa de crecimiento es casi siempre la “característica fundamental” en los programas de selección (Zobel y Talbert 1994). Otras características de importancia son la forma del fuste, las bifurcaciones y ramificaciones, la resistencia a enfermedades, la proporción de albura y duramen y la anatomía de la madera. Para establecer un ensayo de progenies, se seleccionan individuos con buenas características fenotípicas de acuerdo a los objetivos productivos de la especie. Como estas características (crecimiento y forma del árbol), son el resultado de la interacción entre el potencial genético y el ambiente, se hace una selección poco exigente de los individuos. De esta forma se garantiza una variabilidad amplia a partir de la cual se realiza el mejoramiento. De estos árboles seleccionados, se obtienen las semillas para la producción de plantines. El sitio elegido para el ensayo debe ser homogéneo para minimizar el efecto ambiental, y poder identificar aquellos individuos que presentan superioridad genotípica. Este tipo de ensayo debe evaluarse hasta el cumplimiento del turno de corta y de forma continua durante todo el tiempo de crecimiento, ya que existe la posibilidad de que ciertas características se manifiesten recién al final del ciclo. Por ello, no se deben tomar decisiones importantes de inversión hasta no contar con resultados definitivos derivados de muchos años de análisis. Por otro lado, es necesario registrar los datos desde el comienzo del ensayo, dado que los ciclos de cultivo de los árboles son de varios años hasta décadas y las necesidades de información son en general, más inmediatas. Además, esto permite descartar progenitores con poco potencial en forma temprana, lo que se denomina aclareo genético. De acuerdo a este criterio se ralea el ensayo con el objetivo de dejar en pie sólo los mejores individuos y brindarle a cada árbol seleccionado el espacio suficiente para desarrollar su copa y que se convierta en un buen productor de semillas. Alcanzado este punto estamos en presencia de un huerto semillero, el cual proveerá semillas que permitirán establecer plantaciones genéticamente mejoradas. 173 Horlent, M. y D. Monteverde Ensayo de progenies de Cedrela balansae en Valle Morado El objetivo de este ensayo fue evaluar la variabilidad genética existente entre 42 árboles seleccionados, analizando características como crecimiento, forma, resistencia al ataque de la mariposita del brote (Hypsipyla grandella) y quemado de la corteza. En base a esta información se pretende seleccionar los progenitores genéticamente superiores para la provisión de semillas mejoradas. En una etapa posterior, se realizará un raleo para dejar los mejores individuos de las mejores familias, y así poder constituir un huerto semillero. El ensayo de progenies de C. balansae de Valle Morado se estableció en febrero de 2001 y cuenta con descendientes de 42 progenitores distintos (42 famiFigura 1. Localidades de donde provienen las semillas lias) los cuales fueron selecciona- de cedro orán (Cedrela balansae) para el ensayo de prodos en las localidades de Campo genies de la plantación experimental de Valle Morado, Chico, Los Naranjos, Valle Mo- Provincia de Salta. rado (Provincia de Salta), Yuto y Ledesma (Provincia de Jujuy) (Fig. 1). En cada localidad, se ubicaron Las técnicas de recolección de semillas y grupos de cedros dentro de los cuales se la producción de plantines se describe en seleccionó el mejor individuo a través de el capitulo 12 (Monteverde). Los plantiuna evaluación cualitativa de la forma nes obtenidos en el vivero se llevaron a (rectitud del fuste y ramificaciones) y de plantación definitiva en febrero de 2001, la sanidad. Los individuos seleccionados, a un distanciamiento de 4 m entre filas y distanciados entre sí 200 m como mínimo, 2,5 m entre plantas (1.000 individuos por fueron identificados y georeferenciados hectárea). Cada plantín, identificado en con el fin de poder reubicarlos para futuras el vivero en función de su progenitor, fue cosechas (Tabla 1). ubicado en 20 bloques (20 repeticiones) y 174 Crecimiento de Cedrela balansae en la plantación experimental de Valle Morado Tabla 1. Datos de procedencia y características de los individuos de cedro orán (Cedrela balansae) seleccionados para el ensayo de progenies de la plantación experimental de Valle Morado, Provincia de Salta. Se indica el número de individuos (n), la altura promedio sobre el nivel del mar, y los valores mínimos y máximos de DAP y altura de los individuos cosechados por localidad. Sitio n m s.n.m. DAP (cm) Altura (m) Campo Chico 3 400 40 20 Los Naranjos 8 800 - - Valle morado 4 500 43 - Yuto 6 400 35 - 46 20 - 22 Ingenio Ledesma 21 400 33.5 - 58 16 - 27 42 tratamientos (42 familias). Se aplicó el diseño estadístico bloques incompletos al azar, ya que cada bloque contenía sólo 40 familias. Cada tratamiento o familia se encuentra representado por una parcela que contiene 2 individuos (Fig. 2). En el año 2002 y 2003 se realizaron las mediciones de crecimiento en diámetro a la altura del pecho (DAP), diámetro a la base (DAB), altura total y se determinó el número de individuos afectados por ataque de la mariposita del bro- Figura 2. Diseño esquemático del ensayo. te y la forma general del árbol. En esta última variable se evaluó en conjunto la rectitud del fuste, las ramificaciones y bifurcaciones y el desarrollo de la copa. Se la clasificó en cinco categorías: 1) excelente: individuos de fuste recto sin bifurcaciones, copa con poca profusión de ramas; 2) muy buena: árboles de fuste recto que no presentan bifurcaciones, pero tienen un gran desarrollo de copa; 3) buena: árboles de buen fuste, pero que presentan alguna curvatura y ramificaciones; 4) regular: árboles que presentan fuste irregular, ramificaciones y bifurcaciones, pero que se podrían recuperar u obtener algún producto a través de la realización de podas y 5) mala: individuos con mala forma y presencia de rebrotes y ramificaciones laterales. 175 Horlent, M. y D. Monteverde RESULTADOS Crecimiento. El crecimiento total promedio para todas las familias fue de 5,4 cm de DAP (crecimiento anual de 2,7 cm, con una variabilidad entre 3,1 y 7 cm). Dentro de este rango, el 70 % de las familias presentaron un crecimiento total mayor al promedio. En relación al crecimiento en altura, se obtuvo un valor total promedio de 3,3 m a razón de 1,65 m/año. El valor más bajo de crecimiento en altura fue de 1,7 m y el mayor de 4,4 m (Tabla 2). Ambas medidas de crecimiento están asociadas positivamente. Muerte del brote apical. Todas las familias plantadas en el ensayo de progenies, presentaron ataque de Hypsipyla grandella. Teniendo en cuenta los individuos sobrevivientes al momento de la última medición, el porcentaje de plantas atacadas por familia, varió entre el 34,8 y el 79,5%. De las 42 familias del ensayo, el 65 % presentaron más de la mitad de sus individuos con algún grado de ataque. Quemado de la corteza. Este daño se produce por acción directa del sol. Genera desde un manchado en la corteza hasta heridas abiertas de tipo crónicas que pueden afectar 176 Tabla 2. Crecimiento en diámetro a la altura del pecho (DAP) y en altura para cedro orán (C. balansae), en el ensayo de progenies en Valle Morado, Provincia de Salta Crec. en DAP fam. prom. ds max. 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 5,9 5,4 5,9 3,9 6,0 6,4 6,5 4,5 4,9 3,6 3,3 3,5 3,2 3,2 3,1 3,3 3,4 5,4 6,2 5,6 5,7 5,9 6,0 6,4 5,8 5,8 6,7 6,2 6,0 6,1 4,6 3,9 5,7 6,2 6,2 6,6 6,3 6,0 5,6 6,9 6,7 7,0 1,6 2,0 1,6 1,2 2,5 1,7 2,6 2,1 1,8 1,0 0,8 1,5 1,7 1,0 1,0 1,0 1,6 1,9 1,8 1,8 2,4 1,3 1,7 2,3 1,4 2,1 1,7 2,3 1,8 2,0 1,4 1,3 1,8 2,1 1,9 2,7 1,7 2,4 1,7 1,9 2,0 1,8 9,7 9,0 8,7 6,0 10,7 10,4 15,7 8,6 10,0 5,4 5,0 6,5 7,3 4,8 5,3 5,1 6,6 8,0 9,6 10,8 10,0 8,3 9,2 9,1 8,7 10,1 9,8 9,5 9,1 9,9 8,0 5,9 8,5 9,7 9,9 10,6 8,7 10,4 8,8 10,6 11,1 10,2 Crec. en altura prom. ds max. 3,6 3,5 3,5 2,8 3,6 3,9 3,8 2,8 3,4 2,7 2,1 2,4 1,7 2,5 2,2 2,5 2,1 3,3 3,5 3,4 3,4 3,2 3,8 3,8 3,7 3,5 4,0 3,6 3,6 3,8 2,9 2,4 3,3 3,6 3,6 3,8 3,6 3,5 3,6 4,0 4,1 4,4 0,6 0,7 0,6 0,8 0,7 0,5 0,7 1,1 0,6 0,7 0,7 0,7 0,7 0,5 0,6 0,7 0,7 0,9 0,9 0,7 1,0 0,8 0,6 0,9 0,9 0,8 0,7 0,9 0,7 0,6 0,8 0,8 1,1 0,7 0,7 0,9 0,6 1,0 0,6 0,5 0,6 0,7 4,7 4,9 4,8 4,6 4,8 4,7 5,8 4,7 4 4,1 3,4 3,9 3,2 4,3 3,3 4,2 3,6 4,8 4,5 4,5 5,3 4,3 4,8 4,8 4,6 4,5 5,3 5 4,8 5,2 4 4,1 4,6 4,9 4,7 5,2 4,6 5,3 4,8 4,7 5,2 5,7 Crecimiento de Cedrela balansae en la plantación experimental de Valle Morado la calidad de la madera. También se ve afectada la sanidad de todo el árbol, ya que estas heridas son una vía de entrada directa de hongos, bacterias e insectos. Al igual que en la variable anterior, se registró la presencia del quemado de la corteza en cada individuo, sin determinar la intensidad. Del total del ensayo, el 30 % de las plantas presentaron signos de quemado en la corteza. Analizando los datos por familias, se encontró que el porcentaje de plantas con esta afección varió entre el 10 y el 50 %. Forma del árbol. La proporción de individuos clasificados en las mejores categorías de forma (excelente y muy buena), no es uniforme en cada familia. Puede haber entre un 39 y un 90% de individuos con estas categorías dentro de una familia. La forma del árbol está fuertemente ligada al ataque de la mariposita. La muerte del brote apical estimula la bifurcación y el crecimiento de brotes laterales, dando como resultado un árbol cuya forma no es deseable para la actividad forestal. Mortalidad. A diferencia del ataque de la mariposita y el quemado de la corteza, en este caso se encontraron familias en las que la mortalidad fue nula hasta la fecha de la última medición (abril de 2003). Del total de las 42 familias evaluadas, ocho no presentaron individuos muertos. En las familias restantes, la proporción de individuos muertos varió entre 2,5 y 47,5%. DISCUSION Los resultados presentados corresponden a los primeros dos años de desarrollo de Cedrela balansae en el ensayo de progenies de la plantación experimental de Valle Morado. Si bien estos ensayos requieren un análisis continuo hasta completar el turno de crecimiento, en este trabajo se encontraron tendencias de mayores tasas de crecimiento en algunas familias, al poco tiempo de establecida la plantación. Esto demuestra que sería posible detectar diferencias entre genotipos en forma temprana, lo cual podría tener consecuencias comerciales importantes. Las dos medidas de crecimiento analizadas (diámetro y altura), presentan una fuerte asociación. Esto significa que las familias que más crecieron en diámetro también lo hicieron en altura. Al traducir esto en el rendimiento en metros cúbicos que se obtendrían de las distintas familias, se revelan diferencias contrastantes entre unas y otras. Futuras plantaciones realizadas con material proveniente de las familias que presentaron los mayores crecimientos en altura y diámetro, presentarían rendimientos superiores en comparación con otras cuyas plantas no han tenido ningún proceso de selección y mejoramiento. Si además estas familias que crecen más, son las más sanas, tienen mejor forma y resisten el ataque de plagas, la ganancia genética que es posible obtener con el mejoramiento es enorme y se traduce directamente en beneficios económicos para los productores. Sin embargo, por el momento el resto de las variables analizadas, como mortalidad, quemado de la corteza y ataque de insectos, no presentaron ninguna asociación ni 177 Horlent, M. y D. Monteverde entre ellas, ni con el crecimiento. Hasta ahora sabemos que hay familias que crecen más, algunas que tienen mejor forma, otras que sufren menos ataques de insectos, pero no encontramos familias que claramente reúnan buenas características del conjunto de variables analizadas. Por ello es importante seguir adelante en el análisis de este ensayo, ya que es probable que con el tiempo se vayan definiendo más claramente algunas tendencias y se determine la real incidencia de algunos aspectos, como el ataque de mariposita o el quemado de la corteza en la calidad y en el rendimiento final de los árboles. Esto permitirá conocer de manera más concluyente la calidad genética de las distintas familias. BIBLIOGRAFIA Barrett, W.H.G. 1980. Elementos y principios de la genética. En: FAO 20/1. Mejora genética de árboles forestales: 18-26. Zobel, B. y Talbert, J. 1994. Técnicas de mejoramiento genético de árboles forestales. Editorial Limusa: 545 pp. 178 Resultados comparativos de plantaciones Cedrela balansae en Yungas INTA -Yuto, Jujuy Ecología y producción de cedrode(género Cedrela) en las australes. Pacheco, S. y A. Brown (Eds.). LIEY-ProYungas. Argentina. pp. 179-192. 2006. Resultados comparativos de plantaciones experimentales de Cedrela balansae en INTA - Yuto, Jujuy del Castillo, E. M.1; M. A. Zapater 2 y M. N. Gil 2 . 1 INTA - Yuto, R.N. 34 Km 1286 (4518), El Bananal, Jujuy, Argentina. 2. Universidad Nacional de Salta [email protected] Este trabajo resume información sobre ensayos de distanciamientos y tratamientos de riego, realizados en plantaciones experimentales bajo cubierta y a cielo abierto, con la especie Cedrela balansae (cedro orán). Se compara el desarrollo del cedro con especies nativas y exóticas. En plantaciones bajo cubierta se encontró que el cedro orán tiene mayor crecimiento que otras especies nativas, pero no alcanza al de Toona australis. El espaciamiento en el que el cedro alcanzó mayor crecimiento fue el de 5x5 m. El ataque de la mariposita del brote (Hypsipyla grandella) en plantaciones bajo cubierta, es reducido. En plantaciones a cielo abierto, no se encontró diferencias en el crecimiento a distintos espaciamientos y el crecimiento es similar al de Toona australis. En comparación a otras especies del género Cedrela, el cedro orán y el cedro mexicano, alcanzan los mayores crecimientos, y el primero supera a las otras especies en tratamientos sin riego. En la plantación comercial analizada se demuestra que la especie implantada tiene un gran potencial productivo si se efectúan los tratamientos sanitarios necesarios y si se maneja la plantación con escamondos de corrección. This paper resumes information that resulted from experiments with distance and water supply treatments, implemented in Cedrela balansae (cedro orán) understory and open experimental plantations. Cedro growth is compared with other native and non native species. In understory plantations cedro orán had higher growth compared to other native species, but it did not grow as much as Toona australis. Distance treatment in which cedro had higher growth was 5 x 5 m. Hypsipyla grandella did not significantly attack plants in understory plantations. In open plantations, there were no differences in growth in distance treatments, and growth is similar to Toona australis. Comparing to other Cedrela species, cedro orán and Mexican cedro have higher growth, and cedro orán grows more than all other species in no artificially watered treatments . The studied commercial plantation show that the species have high reproductive potential if receives adequate health treatment and if pruning is correctly managed. 179 del Castillo, E.M.; M.A. Zapater y M.N. Gil Introduccion Para la producción de maderas nativas de alto valor forestal, es imprescindible conocer sus características de desarrollo bajo distintos sistemas de manejo y en comparación a otras especies nativas y exóticas que se comercializan actualmente. La Estación Experimental de Cultivos Tropicales del INTA-Yuto, en la Provincia de Jujuy, posee plantaciones con ensayos de este tipo para la especie Cedrela balansae (cedro orán). Los estudios que se presentan a continuación corresponden a ensayos de evaluación del desarrollo en plantaciones experimentales bajo cubierta y a cielo abierto. Los ensayos se llevaron a cabo en ambientes de Selva Pedemontana y su umbral al Chaco semiárido, en las Provincias de Salta (Anta Muerta y La Moraleja) y Jujuy (INTA - Yuto ). plantaciones experimentales bajo cubierta Las plantaciones bajo cubierta o enriquecimiento constituyen uno de los métodos silvícolas más apropiados para la recuperación o restauración de la Selva Subtropical. Se pueden realizar en rodales que fueron explotados intensamente, de donde se extrajeron selectivamente los mejores individuos y las especies de mayor valor económico. La forestación bajo cubierta es una alternativa productiva importante por las siguientes razones: a) los incrementos en diámetro y altura han mostrado resultados satisfactorios; b) el precio de las maderas valiosas utilizadas en forestaciones bajo cubierta es 180 diez veces superior a las especies exóticas usadas tradicionalmente en forestaciones a cielo abierto, como Eucalyptus y Pinus. La madera de cedro de primera calidad se paga entre $400 y $600 por m3; mientras que especies como Eucalyptus, de madera blanca y blanda, valen entre $30 y $40 por m3; c) la forestación bajo cubierta incrementa la productividad a una cifra superior a los 10 m3 / ha / año y la renta anual es cuatro a cinco veces superior a la renta de las especies exóticas; d) se mantiene la biodiversidad de la selva ya que únicamente se modifica la franja que se abre para la plantación, manteniéndose sin alteración el área entre fajas. Características técnicas del sistema La forestación bajo cubierta consiste en la apertura de fajas en un bosque que ha sido intensamente explotado y tiene una baja densidad de especies arbóreas con importancia comercial. El diseño de fajas se realiza con un rumbo determinado. La distancia entre rumbos y entre plantines en las fajas, depende de la densidad de árboles que se quiere obtener. La densidad de plantación más aconsejable para especies nativas valiosas varía entre 200 - 250 individuos por hectárea, lo que se logra realizando la apertura de rumbos cada 10 o 20 m. El ancho de faja debe ser de 4 a 6 m, con una o dos filas por faja, con distanciamientos de 4 o 5 m entre plantas. El tamaño óptimo de los plantines al momento de realizar la plantación es de 0,80 - 1,50 m de altura, el cual se alcanza a los 12 o 15 meses de iniciada la siembra. Resultados comparativos de plantaciones de Cedrela balansae en INTA -Yuto, Jujuy El desmonte puede ser realizado en forma manual o mecánica dependiendo de la topografía y de la superficie a implantar. En los espacios entre fajas se puede favorecer la regeneración natural de las especies valiosas a través de cortas de limpieza y liberación. ser profundos, franco a franco arenosos y levemente ácidos con pH de 6,5 a 6,7. Las precipitaciones medias anuales deben variar entre 900 a 1.400 mm, y las temperaturas mínimas deben ser superiores a -4 º C. La evapotranspiración potencial no debe ser negativa. En la elección de las especies a utilizar en la plantación se debe: a) seleccionar especies nativas o exóticas de buena calidad maderable y con buen crecimiento; b) favorecer las especies nativas de mejor adaptación al método, teniendo en cuenta aspectos ecológicos, como ser competencia y tolerancia; c) seleccionar especies adaptadas a las características edáficas y climáticas del sitio y d) seleccionar especies no susceptibles a enfermedades. La plantación debe realizarse durante la época de lluvias (noviembre a marzo), en pozos de 20 cm de profundidad, sin enterrar el cuello de la planta. Al tapar se debe mezclar la tierra profunda con el humus superficial, apisonando el suelo contra las raíces. Los cuidados culturales deben realizarse en forma periódica para controlar la invasión del estrato herbáceo y enredaderas, que incrementan en forma notable cuando se realiza la apertura del dosel arbóreo. Otro aspecto importante a considerar son las cortas de liberación y limpieza, que se deben realizar dos o tres veces al año. En cuanto a las características del sitio, se deben seleccionar aquellos que se adapten a los requerimientos de la especie a cultivar, especialmente desde el punto de vista edáfico y climático. En lo posible, se debe realizar en terrenos planos o con ligeras pendientes (no mayores a un 15 %) para no producir grandes disturbios y no tener inconvenientes en el trazado de caminos de acceso (tanto para realizar la plantación como para la extracción de madera en el futuro). Las terrazas aluviales y los pies de serranías constituyen sitios de buena aptitud. Los suelos deben contener abundante materia orgánica en superficie (característica común de los suelos de los ecosistemas de Selvas Subtropicales), Ensayos bajo cubierta A continuación se presentan los resultados de dos ensayos llevados a cabo en el INTA-Yuto, con especies nativas y exóticas en plantaciones bajo cubierta (Foto 1). Las plantaciones se ubicaron a 349 m s.n.m. en una bajada aluvional subelevada, con un relieve suavemente ondulado. El suelo está fuertemente desarrollado con perfil A1, A2, B2, B3, C, de textura mediana en superficie a medianamente fina en profundidad, bien a imperfectamente drenados, moderado a 181 del Castillo, E.M.; M.A. Zapater y M.N. Gil fuertemente ácido, bajo contenido de materia orgánica, capacidad de intercambio catiónico medio a moderadamente alto, y porcentaje de saturación de bases bajo. El clima es subhúmedo- seco, con nulo a pequeño exceso de agua y con concentración estival de las precipitaciones. La precipitación media anual es de 862 mm, con una máxima de 1.419 mm y una mínima de 332 mm, demostrando gran variabilidad interanual. Desarrollo de cedro orán en relación a tona y grevilea o roble sedoso. El diseño experimental consistió en la apertura de fajas de 4 m de ancho, orientadas de norte a sur, perpendiculares a la dirección de la pendiente y distanciadas 20 m. Los plantines de las tres especies, con una edad de 6 meses, fueron implantados de manera intercalada a una distancia de 5 m en abril de 1994 (edad: 10 años). Durante los tres primeros, se realizaron cortas de limpieza, de liberación y escamondos (eliminación al ras de las ramas del fuste). Los resultados muestran que la tona (Toona australis) fue la especie que logró los mayores incrementos anuales en diámetro a la altura del pecho (DAP) y altura total. Superó en 11,2 cm en DAP y en 1,4 m en altura al cedro orán (Cedrela balansae). Además superó en 16,1 cm en DAP y 2,19 m en altura a la grevilea (Grevillea robusta). Un incendio afectó parcialmente el ensayo un año después de realizada la implantación. T. australis fue particularmente sensible al fuego y no pudo recuperarse de este efecto, presen182 Foto 1. Ensayo bajo cubierta de cedro orán (Cedrela balansae) en INTA-Yuto, Jujuy. tando los mayores porcentajes de fallas (58,3 %). En cambio, C. balansae, que posee la facultad de una pronta recuperación, tuvo una falla de 27,2 %. G. robusta fue alcanzada por el fuego en menor medida (fallas de un 8,3 %) y por ello, registra el mayor valor de área basal (5 m²/ha) a pesar de las menores dimensiones alcanzadas con respecto a las otras especies. Esto se refleja también en el volumen maderable y su respectivo incremento anual (Tabla 1). Resultados comparativos de plantaciones de Cedrela balansae en INTA -Yuto, Jujuy tabla 1. Valores promedios dasométricos y epidométricos para cedro orán (Cedrela balansae), tona (Toona australis) y grevilea (Grevillea robusta) en una plantación bajo cubierta, a los 10 años de edad. C. balansae T. australis G. robusta DAP (cm) 19,2 30,4 14,3 Altura total (m) 9,0 10,4 8,21 Altura fuste (m) 6,0 5,1 5,56 Area basal (m²/ha) 1,54 4,73 5,00 Volumen madera (m³/ha) 5,54 14,47 16,68 Volumen ramas (m³/ha) 2,77 15,04 7,95 Incremento DAP (cm/año) 1,92 3,04 1,43 Incremento altura total (m/año) 0,9 1,04 0,55 Incremento altura fuste (m/año) 0,6 0,51 0,82 Incremento área basal (m²/ha/año) 0,15 0,47 0,50 Incremento volumen madera (m³/ha/año) 0,55 1,44 1,66 Incremento volumen ramas (m³/ha/año) 0,27 1,50 0,79 Desarrollo de cedro orán, tipa colorada y lapacho rosado usando tres espaciamientos de plantación. El diseño experimental consistió en fajas de 2,5 m de ancho distanciadas 10 m entre sí. Tres bloques al azar, con tres tratamientos de 4, 5 y 6 m de distanciamiento de plantación, con 4 repeticiones y tres especies por parcela. La fecha de implantación fue abril de 1996 (edad: 8 años). Se realizaron cortas de limpieza, cortas de liberación y escamondos durante los tres primeros años (Foto 2). El cedro orán (C. balansae) registró los mayores incrementos en las variables estudiadas para los tres distanciamientos analizados. Esta especie presentó diferencias significativas en el crecimiento, Foto 2. Ensayo de forestación bajo cubier- ta de cedro orán (C. balansae), tipa colorada (Pterogyne nitens) y lapacho rosado (Tabebuia impetiginosa) en INTA-Yuto, Jujuy. 183 del Castillo, E.M.; M.A. Zapater y M.N. Gil Tabla 2. Valores promedios dasométricos y epidométricos para cedro orán (Cedrela balansae), tipa colorada (Pterogyne nitens) y lapacho rosado (Tabebuia impetiginosa) en una plantación bajo cubierta con diferentes espaciamientos de plantación, a los 8 años de edad. C. balansae P. nitens T. impetiginosa Espaciamiento (m) Espaciamiento (m) Espaciamiento (m) 4 4 5 6 4 5 6 5 6 DAP (cm) 12,5 16,5 11,0 6,1 5,4 5,5 5,9 4,0 5,9 Altura total (m) 6,7 7,6 4,7 4,6 5,2 4,9 4,4 5,4 Altura fuste (m) 3,8 4,6 2,7 2,9 3,1 3,2 2,9 3,7 Incremento DAP (cm/año) 1,56 2,04 1,37 8,3 4,8 0,76 0,67 0,69 0,74 0,50 0,74 Incremento altura total (m/año) 0,84 1,04 0,95 0,59 0,57 0,65 0,61 0,55 0,67 Incremento altura fuste (m/año) 0,47 0,60 0,57 0,34 0,36 0,39 0,40 0,36 0,46 cuando el espaciamiento de plantación fue de 5 m. Tanto la tipa colorada (Pterogyne nitens) como el lapacho rosado (Tabebuia impetiginosa) no presentaron diferencias significativas entre ellas, y tampoco se verificó correlación entre sus crecimientos y las distancias de implantación (Tabla 2). plantaciones experimentales a cielo abierto Este sistema silvícola es el que mejor responde si consideramos evolución y crecimiento. Para su implementación se deben tener en cuenta los siguientes aspectos: Selección de Sitios. Es el factor de mayor relevancia. Se deben elegir aquellos sitios o lugares cuyas características ambientales (topografía, suelo y clima) sean com184 patibles con las exigencias de la especie a utilizar. En el caso de especies nobles, con maderas de alta calidad, se requieren suelos ricos en nutrientes y bien drenados, con pendientes inferiores al 15 %, registros pluviométricos mínimos de 900 mm/año y temperaturas mínimas superiores a los -2 ºC. Preparación del Sitio. Este sistema se caracteriza por la eliminación total de la vegetación natural existente. Se realiza en sitios que prácticamente han sido desvastados en su estructura y densidad de especies valiosas, e invadidos por especies secundarias sin valor comercial. La limpieza total del sitio, según la topografía del terreno, puede hacerse mecánicamente con topadora, desraizando para favorecer las Resultados comparativos de plantaciones de Cedrela balansae en INTA -Yuto, Jujuy limpiezas posteriores; o con motosierra, realizando tala rasa a toco bajo, cuando las pendientes son superiores a un 10 %. En ambos casos, se debe recuperar o rescatar la madera remanente de las especies de valor comercial. Densidad-espaciamiento. El tipo de enraizamiento y la necesidad de la especie en cuanto a los requerimientos de agua y sustancias nutritivas del suelo, son determinantes para fijar el espaciamiento o densidad. Los espaciamientos más usuales son 4 x 4, 5 x 4, y 5 x 5 m; con densidades de 625, 500 y 400 plantas por hectárea respectivamente. Epoca de plantación. Cuando se utilizan especies exigentes en cuanto a disponibilidad de agua, es conveniente realizar la plantación durante los meses de lluvia (diciembre a febrero), cuando el suelo está saturado de humedad. Poda de yemas o desbrote. La aplicación de la técnica de desbrote o poda de yemas es imprescindible a partir del primer año de implantación, durante la primavera-verano cuando la planta alcanza 1,50 - 2 m de altura. Se deben evitar lesiones y procurar acelerar el proceso de cicatrización. Cerramiento. En los sitios o lugares que puedan ser afectados por el ganado vacuno, es necesario clausurar con alambrado. Tratamientos silviculturales. Las plantas jóvenes son sensibles a la competencia con herbáceas. Por ello, las plantaciones deben mantenerse limpias durante los primeros años hasta que se produzca el cierre del dosel. La práctica del escamondo, hasta alcanzar una altura de 6 m, es importan- te para obtener una producción de madera libre de nudos, a partir de un diámetro de 3 - 4 cm. Tratamientos silvícolas intermedios. Estos tratamientos tienen como objetivo principal eliminar la competencia entre especies para que el crecimiento no disminuya. Es importante realizar un espaciamiento adecuado para evitar competencia por la disponibilidad de radiación solar, agua y nutrientes del suelo. Esto se logra disminuyendo la densidad inicial de plantación, operación que se denomina raleo. Ensayos a cielo abierto A continuación se presentan los resultados de tres ensayos en plantaciones a cielo abierto. Dos ensayos se realizaron en el INTA - Yuto, donde se comparó el desarrollo del cedro orán con especies exóticas y el crecimiento del cedro orán a tres distancias de espaciamiento. Las características edáficas y climáticas de este sitio fueron descriptas en los ensayos de plantaciones bajo cubierta. El tercer ensayo que compara el desarrollo de cuatro especies de Cedrela, se realizó en Anta Muerta, Departamento de Orán en la Provincia de Salta. La plantación está ubicada en un piedemonte a 405 m s.n.m., con pendientes del 2 al 5 %. El suelo es profundo, franco a franco arcilloso. El clima es subhúmedo - seco, con nulo a pequeño exceso de agua. La temperatura media anual es de 21,6 °C y la precipitación media anual es de 1.274 mm. 185 del Castillo, E.M.; M.A. Zapater y M.N. Gil Desarrollo de cedro orán, tona y paraíso gigante. Se trabajó con cinco parcelas de 10 plantas y un espaciamiento común de plantación de 4 x 4 m. La fecha de implantación fue abril de 1990 (edad: 14 años). Se realizaron cortas de limpieza y escamondos durante los tres primeros años, y se produjo un raleo natural por fallas y caída de árboles por acción del viento (Foto 3). El paraiso gigante (Melia azedarach) superó a las otras especies en su crecimiento en diámetro. Sin embargo, registró un crecimiento inferior en altura total, lo que determinó que su incremento en volumen maderable resultara menor. Comparando el cedro orán (C. balansae) y la tona (T. australis), ambas con igual DAP, se observó que la última registró un crecimiento en altura total ligeramente superior, pero que no fue significativo. Analizando las alturas de fuste, y por lo tanto el volumen maderable, las diferencias resultan significativas en favor de T. australis, debido a los severos ataques de la mariposita del brote (Hypsipyla grandella) en C. balansae (Tabla 3). Desarrollo de cedro orán usando cuatro distanciamientos de plantación. Se usaron parcelas de 7 filas con 7 plantas cada una con cuatro espaciamientos (3,5 x 3,5; 3 x 4; 4 x 4; 5 x 5 m). La fecha de implantación fue abril de 1994 (edad: 10 años). Se realizaron cortas de limpias y escamondos durante los tres primeros años. Se produjo un raleo natural por fallas y caída de árboles por acción del viento. Los mayores incrementos en DAP se verificaron a espaciamientos de 3 x 4, 4 x 4 y 5 x 5 m, sin detectarse diferencias significativas entre ellos. En cuanto a altura total, no se encontraron diferencias entre los tratamientos. A esta edad de plantación, el espaciamiento no pareció influir en el desarrollo de los inviduos (Tabla 4). Desarrollo de cedro orán, cedro misionero, cedro rosado y cedro mexicano. El diseño experimental consistió en bloques completos al azar con cinco repeticiones. Se trabajó con un espaciamiento de plantas de 5 x 4 m en parcelas de cuatro filas. La fecha de implantación fue diciembre de 2000 (edad: 4 años). Foto 3. Plantación experimental a cielo abier- to de cedro orán (Cedrela balansae) en INTAYuto, Jujuy. 186 Como resultado se observó que de las cuatro especies, el cedro orán (C. balansae) y el cedro mexicano (C. odorata) se destacaron netamente por su crecimiento en diámetro y altura (Tabla 5). Resultados comparativos de plantaciones de Cedrela balansae en INTA -Yuto, Jujuy tabla 3. Valores promedios dasométricos y epidométricos para cedro orán (Cedrela balansae), tona (Toona australis) y paraiso gigante (Melia azedarach) en una plantación a cielo abierto, a los 14 años de edad. C. balansae T. australis M. azedarach DAP (cm) 26,9 26,9 29,1 Altura total (m) 11,5 11,9 9,0 Altura fuste (m) 6,6 7,2 4,1 Area basal (m²/ha) 23,79 31,69 32,03 Volumen madera (m³/ha) 94,20 136,90 78,79 Volumen ramas (m³/ha) 69,94 89,36 94,16 Incremento DAP (cm/año) 1,9 1,9 2,08 Incremento altura total (m/año) 0,82 0,8 0,64 Incremento altura fuste (m/año) 0,47 0,5 0,29 Incremento área basal (m²/ha/año) 1,70 2,26 2,29 Incremento volumen madera (m³/ha/año) 6,72 9,77 5,62 Incremento volumen ramas (m³/ha/año) 4,99 6,38 6,72 Desarrollo de cedro rosado, cedro orán y cedro mexicano con y sin riego localizado. Este ensayo se realizó en La Moraleja, Departamento de Anta en la Provincia de Salta. La plantación está ubicada en llanuras y piedemonte a 360 m s.n.m, con pendientes del 1 al 6 %. Los suelos son profundos, bien drenados, de texturas moderadamente finas a finas. La temperatura media anual es de 20,5° C y la precipitación media anual es de 817 mm. El diseño experimental consistió en bloques completos al azar con tres especies y cinco repeticiones. Se trabajó con un espaciamiento de 5 x 4 m en parcelas de 4 filas con 5 plantas cada una y dos tratamientos de riego. La fecha de implan- tación fue septiembre de 2001 (edad: 3 años). Se realizaron cortas de limpieza y escamondos. En el tratamiento sin riego el cedro orán (C. balansae) superó significativamente a las otras especies en diámetro y altura. No se encontraron diferencias en el tratamiento con riego. En las otras dos especies se produjo un importante incremento en diámetro con la aplicación de riego localizado. Para el cedro rosado (C. saltensis) el incremento en DAP fue del orden de 0,7 cm, y para el cedro mexicano (C. odorata), de 1,4 cm. Sólo C. saltensis tuvo un incremento en altura en el tratamiento con riego (Tabla 6). 187 del Castillo, E.M.; M.A. Zapater y M.N. Gil tabla 4. Valores promedios dasométricos y epidométricos para Cedrela balansae, en una plan- tación a cielo abierto con cuatro espaciamientos de implantación, a los 10 años de edad. 3,5 x 3,5 m 3x4m 4x4m 5x5m DAP (cm) 18,0 20,2 20,4 20,66 Altura total (m) 9,1 10,4 10,6 9,40 Altura fuste (m) 5,1 6,8 5,3 3,90 Area basal (m²/ha) 18,19 23,80 20,51 12,61 Volumen madera (m³/ha) 55,66 97,10 65,22 29,50 Volumen ramas (m³/ha) 43,65 51,40 65,22 41,61 Incremento DAP (cm/año) 1,8 2,02 2,04 2,06 Incremento altura total (m/año) 0,90 1,04 1,06 0,94 Incremento altura fuste (m/año) 0,50 0,68 0,53 0,39 Incremento área basal (m²/ha/año) 1,82 2,38 2,05 1,26 Incremento volumen madera (m³/ha/año) 5,56 9,71 6,52 2,95 Incremento volumen ramas (m³/ha/año) 4,36 5,14 6,52 4,16 PLANTACION COMERCIAL DE cedro oran En esta plantación, ubicada en Anta Muerta, en el Departamento Orán de la Provincia de Salta, se trabajó con un espaciamiento de 4 x 4 m sobre una superficie de 45 hectáreas. La fecha de implantación fue diciembre de 2000 (edad: 4 años) y se realizaron cortas de limpieza y escamondos (Foto 4). En comparación con los ensayos experimentales realizados con esta especie, se observó un bajo crecimiento diamétrico y excesivo desarrollo en altura. Esto se explica por el mal manejo de la plantación, sin empleo de agroquímicos y un escamondo severo realizado hasta los 2/3 de la 188 Foto 4. Plantación comercial de cedro orán (Cedrela balansae) en Anta Muerta , Salta. altura total. El 24 % de las plantas presentaron de uno a tres ataques de Hypsipyla grandella (Tabla 7). Resultados comparativos de plantaciones de Cedrela balansae en INTA -Yuto, Jujuy Tabla 5. Valores promedios dasométricos y epidométricos para cuatro especies de Cedrela en una plantación a cielo abierto, a los cuatro años de edad. DAB (diámetro a la base). C. saltensis C. fissilis C. balansae C. odorata DAB (cm) 4,8 3,1 6,0 5,4 Altura total (m) 1,25 1,0 1,76 1,7 Altura fuste (m) 0,87 0,8 1,31 1,30 Incremento DAB (cm/año) 1,6 1,03 2,00 1,80 Incremento altura total (m/año) 0,42 0,33 0,59 0,57 Incremento altura fuste (m/año) 0,29 0,27 0,44 0,43 Tabla 6. Valores promedios dasométricos y epidométricos para tres especies de Cedrela en una plantación a cielo abierto con riego y sin riego, a los tres años de edad. C. saltensis C. balansae C. odorata S/riego C/riego S/riego C/riego S/riego C/riego DAP (cm) 5,09 5,79 5,49 5,46 4,44 5,84 Altura total (m) 2,65 3,48 3,75 2,83 3,13 2,93 Altura fuste (m) 1,90 2,38 2,63 4,10 2,03 2,18 Tabla 7. Valores promedios dasométricos y epidométricos para Cedrela balansae en una plan- tación comercial, a los cuatro años de edad. C. balansae DAP (cm) 4,30 Altura total (m) 2,90 Altura fuste (m) 2,20 Area basal (m²/ha) 1,20 Volumen madera (m³/ha) 1,58 Volumen ramas (m³/ha) 0,50 (Continua en pag sgte.) 189 del Castillo, E.M.; M.A. Zapater y M.N. Gil continuacion Tabla 7. Valores promedios dasométricos y epidométricos para Cedrela balansae en una plantación comercial, a los cuatro años de edad. C. balansae Incremento DAP (cm/año) 1,07 Incremento altura total (m/año) 0,72 Incremento altura fuste (m/año) 0,55 Incremento área basal (m²/ha/año) 0,30 Incremento volumen madera (m³/ha/año) 0,39 Incremento volumen ramas (m³/ha/año) 0,12 anAlisis de resultados y discusiOn Los ensayos se llevaron a cabo en ambientes de Selva Pedemontana de las Provincias de Salta (Anta Muerta y La Moraleja) y Jujuy (INTA - Yuto). Este último se emplaza en el ecotono entre la Selva Pedemontana y el Chaco semiárido. Se verifican importantes diferencias climáticas entre los sitios, especialmente con respecto a los registros pluviométricos y frecuencia de heladas. El Sitio Anta Muerta, con más de 1.000 mm anuales de precipitación, es el más húmedo. La Moraleja por otra parte, se encuentra en una zona con mayor riesgo de heladas. Los estudios realizados corresponden a sistemas de plantaciones experimentales bajo cubierta y a cielo abierto. En este último caso, también se analiza la respuesta de algunas especies de Cedrela a la aplicación de riego localizado. Las Plantaciones Bajo Cubierta se realizaron en INTA-Yuto y las principales conclusiones son: 190 1- El cedro orán (C. balansae) tiene un crecimiento significativamente superior, en comparación con el lapacho rosado (T. impetiginosa) y la tipa colorada (P. nitens). 2- A diferentes distancias de implantación en las fajas de enriquecimiento, a la edad de 8 años, C. balansae muestra un importante incremento en las magnitudes dasométricas a espaciamientos de 5 x 5 m. 3- Cuando se compara a C. balansae con especies exóticas de rápido crecimiento como el paraiso gigante (Grevillea robusta) y la tona (Toona australis), se observa que ésta última supera al cedro orán en incrementos diamétricos anuales de 1,12 cm. 4- En este sistema de implantación el ataque de Hypsipyla grandella sobre C. balansae es reducido, en comparación a lo que ocurre en plantaciones a cielo abierto. Resultados comparativos de plantaciones de Cedrela balansae en INTA -Yuto, Jujuy Las Plantaciones a Cielo Abierto, se realizaron en los tres sitios y las principales conclusiones son: 1- En ensayos de 10 años de edad, no se verifican diferencias significativas en el crecimiento de C. balansae, como respuesta a los diferentes espaciamientos de plantación. 2- A los 14 años de edad, C. balansae y Toona australis, alcanzan un crecimiento similar. 3- En plantaciones de tres años, el cedro orán (C. balansae) y el cedro mexicano (C. odorata) superan en desarrollo al cedro rosado (C. saltensis) y al cedro misionero (C. fissilis); en ambos casos con valores similares de crecimiento. Esto se modifica a los 18 meses de edad, cuando C. fissilis supera a los otros cedros en altura. 4- C. balansae a los tres años supera en crecimiento a C. saltensis y C. odorata en el tratamiento sin riego localizado. Con la aplicación de riego, C. saltensis y C. odorata manifiestan incrementos en diámetro que superan a C. balansae. 5- El tratamiento con riego localizado a los 14 meses produce un incremento significativo de las magnitudes dasométricas en todas las especies, especialmente C. saltensis. Respecto a la forestación comercial de C. balansae se demostró que la especie tiene un gran potencial productivo, en tanto se efectúen los tratamientos sanitarios necesarios (para combatir la mariposita del brote) y se manejen adecuadamente los escamondos de corrección. 191 Enriquecimiento cedro rosado y cedro misionero Ecología ycon producción de cedro (género Cedrela)en enTucumán, las YungasArgentina australes. Pacheco, S. y A. Brown (Eds.). LIEY-ProYungas. Argentina. pp. 193-198. 2006. Enriquecimiento con cedro rosado y cedro misionero en Tucumán, Argentina González Morenghi, J. M. Darwin 140, (4107), Yerba Buena, Tucumán, Argentina. [email protected] En este capítulo se presenta la experiencia acumulada en tres años de desarrollo de plantaciones de cedro bajo cubierta, en Tucumán. Se detallan los pasos de instalación y mantenimiento de la plantación desde la producción de los plantines. La elección del sitio de plantación involucra el mantenimiento de alambrado, la planificación de caminos de acceso y saca y el diseño de las fajas en la plantación. La preparación del sitio de plantación, implica el diseño de las fajas. Finalmente, se describen los cuidados de mantenimiento que deben realizarse hasta los primeros tres años de establecida la plantación. This chapter presents the tree years experience of the development of cedar plantation in Tucumán. Installation procedures and plantation maintenance are detailed since the production of the saplings. The selection of the planting site involves the preservation of the enclosure system, the planning of the access paths and the design of rows in plantation. The preparation of the planting site, implies the row design. Finally, the maintenance care of the first tree years of plantation are described. 193 González Morenghi, J. M. introduccion La Empresa Forestadora Las Tablas S.R.L. implementó sus primeras plantaciones en el año 2000 en la localidad de Potrero de las Tablas, Provincia de Tucumán. En un primer momento, se forestaron 36 ha con especies exóticas. En el año 2002 se comenzaron a plantar 40 ha con cedro rosado y cedro misionero (Cedrela fissilis), a un ritmo de 10 ha por año. En el año 2003 se implantaron 3 ha de Toona ciliata var. australis y durante el año 2005, se plantaron bajo cubierta 1 ha de teca (Tectona grandis) y 1 ha de tipa colorada (Pterogyne nitens). El objetivo de este trabajo es transmitir la experiencia acumulada en estos años, explicando las pautas de manejo de las plantaciones de cedro bajo cubierta en el pedemonte tucumano. La plantación está ubicada a 18 km de las ciudades de Lules y Raco, sobre la ruta provincial N° 341, Km 25 (26 °49 ´7” S y 65° 26´ 32” W). La altura aproximada es de 800 m s.n.m., la precipitación media anual varía entre 1.100 y 1.400 mm y la vegetación corresponde a Selva Montana. El bosque ha sufrido extracción forestal, por lo que sólo se encuentran cedros aislados de poca edad. PRODUCCION DE PLANTINES Los plantines se preparan el año anterior al de la plantación (Foto 1). Es importante iniciar la plantación con la seguridad de contar con calidad genética en semillas y plantas. Actualmente las semillas de cedro misionero son provistas por un Banco de 194 Foto 1. Plantín de cedro. Semillas de Misiones y los plantines de cedro rosado, por un vivero forestal de Salta. El vivero para producir y mantener las plantas de cedro hasta el momento de la plantación, debe estar en un terreno nivelado y debe tener techos altos que proporcionen sombra, pero permitan la entrada de luz. La producción de plantas a partir de semillas, puede hacerse directamente en macetas plásticas de 10 cm de base x 16 cm de alto, rellenas con tierra sarandeada mezclada con mantillo del lugar. Las semillas deben conservarse en frio, pero la noche anterior a la siembra deben ponerse en remojo a temperatura ambiente. Generalmente, la siembra se realiza en los meses de octubre y noviembre. El riego deber ser constante, pero no intensivo. Las plantas se mantienen bajo una cubierta de media sombra y deben cuidarse del ataque Enriquecimiento con cedro rosado y cedro misionero en Tucumán, Argentina de babosas e insectos. Al año siguiente se seleccionan las mejores plantas y en los meses de septiembre-octubre comienza el transporte al lugar de plantación. ELECCION DEL SITIO DE PLANTACION Para elegir el sitio en el cual se va a instalar la plantación, se deben considerar las siguientes características: Estado de alambres perimetrales y/o construcción de alambrado nuevo. Es importante controlar que la plantación se mantenga con un alambrado en buen estado y que las puertas de acceso estén bien cerradas con cadena y candado, de modo de evitar el ingreso de personas o animales que puedan dañarla. En el verano los vientos y tormentas tiran árboles y ramas que dañan el alambrado, por lo que es necesario realizar un mantenimiento periódico del mismo. el grado de insolación, humedad, y vientos presentes en la misma. Nuestra plantación se encuentra en cerros bajos junto a un valle que tiene como característica el ingreso constante de brisa y viento desde el sur, con heladas cortas y fuertes en invierno. Debido a estas condiciones, la plantación está orientada de este a oeste, aprovechando la cobertura del bosque circundante que protege de vientos helados a las plantas. A la vez, se aprovechan plenamente las horas de sol, disminuyendo la superficie de sombra que ocasiona el monte entre fajas, cuando la plantación está orientada de sur a norte. PREPARACION DEL SITIO DE PLANTACION Caminos de acceso. Es importante proyectar el camino tratando de reducir distancias y tareas de limpieza y planificar el ancho de las curvas de modo que pueda girar un vehículo cargado. Lo ideal es que se pueda entrar y salir de la plantación durante todo el año. Los caminos que en el futuro serán necesarios para sacar los árboles, deben planificarse con anticipación. Estos pueden ser los mismos utilizados para el traslado de plantines y elementos de trabajo (palas, piolas de medición, canastos, mochilas de pulverización, agua y agroquímicos), o bien puede habilitarse una alternativa diferente. En esta etapa, es importante tener planificado el trabajo, estableciendo las actividades que se harán cada día. Una vez decidido el espacio a plantar, se inicia la construcción de las picadas o fajas. Se marca la línea de cabecera, se numeran las cabeceras de cada picada y se comienzan a desmontar las líneas de plantación. El ancho de las picadas debe tener como mínimo 3 m y el largo será de acuerdo a lo oportunamente proyectado. La distancia entre fajas es de 10 m. Se debe decidir si los árboles que quedan en la línea de plantación y entre las fajas, serán cortados, anillados o quedarán en pie para un futuro aprovechamiento. En ciertos casos, debido al gran tamaño de algunos individuos, es conveniente espaciar la plantación de modo que no queden plantas bajo la dominancia de la copa de estos árboles (Foto 2). Orientación de las picadas o fajas. La orientación de la plantación repercute en Una vez limpia la picada y partiendo de la cabecera, se estira una piola de 50 m 195 González Morenghi, J. M. anudada cada 5 m y se marca con la pala el lugar donde se realizará el pozo. Plantando con este espaciamiento y dejando interfajas de 10 m, se plantan a razón de 200 plantas por ha. En caso de plantar cada 4 m, la densidad aumenta a 250 plantas por ha. Cuando se marca la ubicación de cada plantín, se aplica glifosato en un círculo de 1 m alrededor de la marca. De esta manera, se da una mayor protección a la planta recién puesta, del ataque de enredaderas y malezas circundantes. Cuando la humedad de la tierra es poca, se realiza un riego de asiento de 0,5 litros por pozo al momento de plantar. Se quita parcialmente el plástico inferior de la maceta, dejando la parte superior de la bolsa junto con la planta, de modo de evitar pérdida de humedad y reducir el stress hídrico. El plantín se coloca con el tallo recto, se tapa con el mantillo superficial que se sacó del pozo y se presiona con los puños para que las raíces queden apretadas con la tierra. Una vez concluida la plantación de las primeras picadas, se aplican insecticidas preventivos contra hormigas usando pulverizadoras. A los 30 días, se controla la plantación y se refallan los cedros secos o dañados. La cantidad de personal, encargado de la limpieza y de realizar la plantación, dependerá de los recursos de mano de obra calificada de la zona, al igual que de los recursos económicos disponibles. Las actividades de limpieza de las picadas se pueden alternar con las de plantación. El mismo personal interviene para ambas tareas y se requieren como mínimo tres personas, uno 196 Foto 2. Individuo de cedro a un año de ser plantado bajo cubierta en Potrero de Las Tablas, Tucumán. que cava, uno que planta y uno que acarrea las plantas y ayuda a los otros. TAREAS Y CUIDADOS POSTERIORES A LA PLANTACION Una vez concluida la plantación, se realizan tareas de mantenimiento y control que se repiten periódicamente durante los primeros dos años y/o hasta que las copas de los cedros superen el dosel del monte contiguo (Foto 3). A continuación se detallan dichas actividades: Limpieza y desmalezado. Durante el primer y segundo año, la limpieza se realiza con machetes y con las manos. La actividad principal es desenredar, arrancar o cortar las enredaderas que crecen prendidas de los tallos y hojas. Los cedros son fácilmente torcidos por estas plantas debido a su rápido crecimiento y por lo general deben ser raleados. Las tareas se realizan cada 30 días a partir del plantado y se Enriquecimiento con cedro rosado y cedro misionero en Tucumán, Argentina en los hormigueros. Los cebos tóxicos no son recomendables, ya que la humedad del ambiente hace que pierdan efectividad. Control de la mariposita del brote (Hypsipyla grandella). El ataque de esta mariposa produce la muerte del brote apical de la planta de cedro. Una vez ocurrido el ataque, se debe podar la planta lo que retrasa su crecimiento. En nuestro caso, se realizan pulverizaciones preventivas cada 20 - 30 días de noviembre a marzo, con una mezcla de insecticidas de alto valor residual, gran capacidad de volteo y fuerte olor. Foto 3. Plantas de cedro en su segundo año de crecimiento en una plantación bajo cubierta en Potrero de Las Tablas, Tucumán. repiten periódicamente hasta abril-mayo, reiniciándose en los meses de agostoseptiembre. A partir del segundo año y cuando la planta ha crecido al menos 2 m de altura, la limpieza de enredaderas y malezas se hace con herbicida, sin macheteo. Control de hormigas. Desde el inicio de la plantación y a modo de prevención, se pulverizan las plantas con insecticidas de amplio poder residual. El control de las hormigas debe ser permanente ya que consumen las hojas de plantas jóvenes y adultas. Debido a que no comen el brote apical, la planta se recupera rápidamente sin problemas en su crecimiento. El mejor mecanismo de control es aplicar veneno en polvo o con pulverizador, directamente Control de corzuelas. Es común que estos animales utilicen las picadas en busca de alimento. Si bien no comen las plantas, pueden dañar la corteza de las plantas jóvenes. En general, se observa que las heridas generadas se curan naturalmente dejando una cicatriz claramente visible al principio, que luego se pierde con el crecimiento del árbol. No se ha observado disminución en el crecimiento de los cedros por esta razón. TAREAS Y CUIDADOS A PARTIR DEL TERCER AÑO A partir del tercer año, las plantas tienen alturas promedio superiores a los 3 m, inclusive hay individuos de 4 o 5 m, por lo que las tareas de mantenimiento de la plantación disminuyen considerablemente (Foto 4). Eventualmente, es necesario podar alguna rama lateral o anillar algún árbol ubicado entre fajas que creció hacia la plantación. La actividad silvícola disminuye notablemente. Ya no se trabaja con 197 González Morenghi, J. M. continuarse con el control de alambrados y con un mínimo mantenimiento de los futuros caminos de saca. El riesgo de incendio es mínimo ya que el bosque se mantiene verde todo el año. CONCLUSIONES Foto 4. Plantas de cedro en su tercer año de crecimiento en una plantación bajo cubierta en Potrero de Las Tablas, Tucumán. insecticidas, aunque se continúa de manera más espaciada aplicando herbicidas en la base y en la senda de recorrido. Debe 198 La plantación y el manejo silvícola de una forestación bajo cubierta, requiere de un proyecto que incorpore técnicas específicas de manejo y cuidados intensivos hasta el segundo o tercer año de implantado el bosque. Esto permitirá lograr el éxito del objetivo buscado, lo que se traduce en volúmenes y calidad de madera. El cedro tiene un excelente crecimiento tanto en diámetro como en altura. Una buena carga genética de las semillas usadas y una ordenada planificación de las tareas, permiten en sólo dos años tener una plantación con la cual se pueden hacer proyecciones reales de lo que la forestación ofrece como alternativa económica. Mercados para de cedro Ecología y producción de cedro (género Cedrela) en la lasmadera Yungas australes. Pacheco, S. y A. Brown (Eds.). LIEY-ProYungas. Argentina. pp. 199-208. 2006 . Mercados para la madera de cedro Zorrilla, A. Saavedra 706 (3500), Resistencia, Chaco, Argentina. [email protected] En este trabajo se describen los tipos de productores locales que extraen madera del bosque, se detallan los mecanismos que utilizan y los centros donde comercializan su productos. Se presentan estadísticas de la Provincia de Salta para la extracción de cedro en los últimos 20 años. Se describen las características de funcionamiento, precios y destino de los productos, y los atractivos y desventajas del mercado doméstico (actividades de orden local, regional y hasta nacional) y del mercado internacional. Se presenta además, una reseña histórica de la creación de la certificación forestal, sus objetivos de creación, los requisitos que se deben cumplir para lograr la certificación, los países de Latinoamérica con mayor superficie certificada y la situación en Argentina. In this work we describe the type of local producers that extract wood from the forest, providing details about the mechanisms they used, and about the urban areas where they sell their products. In addition, we present data about Cedrella extraction from Salta Province in the last 20 years. We also describe some general aspects of Cedrella comercialization, such as price and destination of the products, and advantages and disadvantages of the domestic (local, regional and even national activities) and international market. We present an historical review of the creation of the system of forest certification, mentioning its objectives and the requirements the products have to fulfil. We mentioned the Latin American countries that have the largest certified area and the status of Argentina, as well. 199 Zorrilla, A. Principales usos de la madera de cedro En el noroeste de Argentina, se encuentran presentes tres especies de cedro, que se distribuyen en las Provincias de Salta, Jujuy, Tucumán y Catamarca (C. lilloi, C. balansae y C. saltensis). En la selva paranaense de la Provincia de Misiones, están citadas dos especies más, C. odorata y C. fissilis (Grau et al. este volumen). Los múltiples usos que tiene la madera de cedro se deben a que es muy vistosa, perfumada y fácil de trabajar, comparada con otras especies forestales de las Yungas de Argentina. En el pasado, el género era abundante en la zona (Minetti este volumen) y su madera era muy utilizada en el mercado de productos maderables. Esta situación llevó a una concepción generalizada entre las poblaciones rurales de que la presencia del cedro determinaba la aptitud maderable de un bosque. La madera del cedro es utilizada para la fabricación de una amplia gama de productos tales como muebles rústicos (mesas, sillas, mesadas, bancos, banquetas), muebles finos (juegos de dormitorio, mesas, sillas, bodegas), aberturas (ventanas, puertas), pisos (tarugado, parquet, entablonado), tirantería, machimbres y zócalos, y utensillos domésticos (fuentes, platos, bateas). MARCO DE EXPLOTACIÓN Y COMERCIO La madera de cedro se extrae del bosque por distintos tipos de productores: Grupos familiares. Generalmente habitan en zonas boscosas, alejadas de los grandes 200 centros urbanos. El grupo familiar se encarga de la extracción, trozado y comercialización. Para el trabajo utilizan hachas como elemento de corte habitual, a veces motosierras para hacer el “rayado” y carros tirados por caballos o bueyes para el arrastre y comercialización de la madera. El sistema de salarios no obedece a una escala legalmente establecida, sino que son pactos domésticos del momento y no hay aportes previsionales de ningún tipo. La comercialización es local, generalmente destinada a pequeñas carpinterías o aserraderos. Los productos consisten en trozas (troncos) y madera aserrada cruda (sin secar ni cepillar). Pequeños productores. Esta categoría difiere de la anterior en que ocasionalmente ocupan mano de obra externa. Estas personas generalmente ya desarrollan otros trabajos en las fincas, como ser el cuidado del ganado y actividades agrícolas. El personal contratado cobra por jornales, con precios informales y generalmente no hay aportes previsionales para los empleados. El uso de motosierras es frecuente, también puede haber uso de algún tipo de maquinarias para el arrastre y comercialización y la venta se efectúa en camiones. La comercialización y los productos obtenidos son iguales que en la categoría anterior. Medianos productores. Las actividades son desempeñadas principalmente por mano de obra contratada, sin embargo los niveles de gestión son ocupados por los titulares de la empresa. La mano de obra contratada cobra jornales oficiales, aunque se considera que una importante proporción de estos empleados están oficialmen- Mercados para la madera de cedro te registrados. Algunos empleados reciben aportes patronales y a su vez están cubiertos por seguros contra accidentes de trabajo. Es común la utilización de cartografía básica para la exploración forestal basada principalmente en personal familiarizado con el lugar que buscan los sitios donde se ubican los árboles (monteros). Se utiliza maquinaria propia o contratada, como ser motosierras, arrastradores mecánicos de las trozas hasta los caminos forestales y camiones para el traslado a los aserraderos. La comercialización es local, provincial, nacional y en pocos casos hay ventas al exterior. Se realiza con un importante grado de organización, aunque los precios obedecen a una escala muy variable de acuerdo a la necesidad del que oferta y del que demanda. Los productos consisten en madera aserrada seca y cepillada, pisos, tirantería, zócalos, machimbres, muebles y componentes. Grandes productores. Se trata de productores madereros que aprovechan el bosque en grandes extensiones, aunque no necesariamente de propietarios de grandes extensiones forestales. Las tareas son desempeñadas por contratistas, que a su vez subcontratan mano de obra. Este es el grupo más profesional con planes de negocios, planificación de las actividades, utilización de cartografía digital, presencia de empleados profesionales con capacitación frecuente, mecanización de la cosecha y con integración empresarial vertical, o en búsqueda de lograrlo. La mayoría de las veces, son propietarios también de la industria asociada al aprovechamiento forestal. La comercialización es local, provincial, nacional e internacional. Los productos consisten en madera de primera categoría, madera aserrada seca, cepillada y clasificada, pisos, muebles y componentes. Previo a la explotación forestal de un bosque, la Autoridad de Aplicación (Secretarías de Medio Ambiente o Dirección de Recursos Naturales, según sea el caso) debe aprobar el Plan de Aprovechamiento Forestal respectivo. Los productores que van a extraer la madera son los titulares del mismo y delegan la elaboración del plan a un profesional habilitado (en general ingeniero agrónomo o forestal). Además deben abonar el “Derecho de Inspección” que permite la inspección de la autoridad provincial y la Guía Forestal con la que el maderero transporta la madera a los centros de industrialización. Según las estadísticas oficiales, en el año 2002 se extrajeron unos 9.000 m3 de rollos de cedro en la Provincia de Salta, lo cual representa el 2 % del total de madera extraída en forma de rollos en la provincia. El mayor volumen se extrajo de la jurisdicción Tartagal (Departamento San Martín), donde alcanzó la cifra de 5.600 m3, seguido por el volumen extraído de la jurisdicción Orán (Departamento Orán), con 3.000 m3. Sin embargo, existe un porcentaje de la madera que se comercializa que no es de fuentes legales, por lo que no está registrada en las estadísticas. Conocer cuál es el porcentaje del mercado informal es difícil y se sugieren cifras que varían entre el 20 % y el 60 % del total. En cuanto al volumen aprovechado de 201 Zorrilla, A. cedros en relación a otras especies, en las estadísticas de los últimos 20 años de la Secretaría de Medio Ambiente y Desarrollo Sustentable de la Provincia de Salta, se observa un punto máximo de extracción de madera de bosques nativos (incluye Yungas y Chaco) en el año 1989, con 396.391 m³ de rollos. A partir de esta fecha y más fuertemente a partir del año 1995, el volumen fue decreciendo hasta llegar a los valores actuales de 35.128 m³. La Dirección General de Estadísticas de la Provincia de Salta registra para el año 2001, que la comercialización del sector maderas (que incluye madera aserrada, puertas, ventanas, muebles y demás productos) alcanzó un valor de US$ 3,6 millones. En este caso también debería considerarse que el valor es más alto, aunque en este segmento la producción y comercialización ilegal, es sensiblemente más baja. MERCADO DOMÉSTICO En este trabajo se define como mercado doméstico a todas las actividades de orden local, regional y hasta nacional vinculadas a la comercialización de cedro. La provisión de madera a las industrias se realiza en forma de rollos y principalmente durante la “época seca” del año (mediados de mayo hasta diciembre e inclusive principios de enero). Esto se debe a que las lluvias estivales dificultan las operaciones de extracción de madera fuera de estos meses. Con relación a la tenencia de la tierra, hay industriales que tienen bosques propios de 202 donde extraen cedro. Hay otros que arriendan o alquilan campos para proveerse de madera y otro grupo compra madera a terceros. Los industriales que aprovechan varias especies y tienen campos propios con cedro, prefieren cortar de sus campos bajos volúmenes de madera y comprar el resto. Esto se debe principalmente a que no desean ser dependientes del precio fluctuante de esta especie, ni tampoco de la inestabilidad de la oferta de cedro, percibiendo la ventaja que representa para el futuro ser poseedores de cedro en sus fincas. Los precios de los rollos de cedro dependen de la calidad de la madera y la época del año, la cual marca la cantidad disponible en el mercado. El precio de los rollos ubicados cerca de un camino accesible para el transporte, fluctúa entre un mínimo de US$ 100 / m3 y un máximo que puede llegar hasta US$ 300 / m3 alcanzando la madera aserrada valores entre US$ 900 y 1.100 / m3. Esto corresponde a madera sana, sin deformaciones, troncos rectos y de largos considerables y sin manchas. Los pagos se realizan desde operaciones al contado hasta cancelaciones a 60 días. Dentro de las Yungas, las explotaciones de cedro se concentran actualmente en la Alta Cuenca del Río Bermejo (ACRB) y el volumen de extracción supera ampliamente al volumen de crecimiento de la especie en los rodales sujetos a explotación. Esta explotación irracional tiende a disminuir el volumen de oferta a futuro. Para algunos industriales, otro factor de incertidumbre que se suma al mercado de cedros, es el efecto que producirá la extracción y comercialización de la madera de tona Mercados para la madera de cedro (Toona ciliata), que actualmente constituye la especie más desarrollada en plantaciones en la región. Sin embargo, los volúmenes disponibles que entrarán en turno de corta en pocos años, no son importantes. Las ventas de productos de cedro se hacen a consumidores finales o distribuidores en grandes centros urbanos. Si bien muchos venden a Buenos Aires, Córdoba, Rosario y Mendoza, hay otros que se limitan al mercado regional como Salta, Jujuy y Tucumán. Los productos que se venden son machimbres, zócalos, tablones entre 1 y 2 pulgadas, aberturas, tirantería, muebles y utensillos. Los precios en el mercado nacional en mayo de 2006 eran para madera rolliza de $ 650-700 el m2 (US$ 200-250 el m2) y de $ 3,5-4,3 el pie2 de tabla o tablón. A su vez, el precio de batienes para aberturas era de $ 3,9-4 el pie2 . A partir de consultas realizadas a industriales forestales, se destaca la elaboración de productos alternativos a partir de la madera de cedro, por ejemplo madera para obras y molduras, que se podrían elaborar y vender fácilmente en el mercado nacional. Sin embargo, consideran que el mayor desafío a enfrentar es la dificultad de reconvertirse tecnológicamente después de muchos años de dificultades económicas. Los atractivos del mercado nacional se basan en un retorno rápido del dinero y la presencia de reglas conocidas por todos y de fácil acceso. Es un nicho seguro que no requiere de óptima calidad de aserrado, ni impone grandes restricciones en cuanto al secado de la madera y porcentajes de hume- dad con el que se entregan los productos. Las desventajas más marcadas son la falta de transparencia y la presencia de madera ilegal que produce permanentes caídas de precios. Los volúmenes y la incorporación de valor agregado que podrían valorizar en gran medida a los productos de madera de cedro, podrían incrementarse si la industria de la construcción tuviera el hábito de una mayor utilización de productos de madera. Por otra parte, el mercado doméstico está atado a los avatares de la economía y fuertemente influenciado por la demanda del sector de la construcción, por lo que las perspectivas son casi siempre inciertas. De todas formas, para los industriales de la madera, este mercado resulta un “refugio” para momentos de incertidumbre en las ventas internacionales. MERCADO INTERNACIONAL En el mercado internacional la madera de cedro que se encuentra en las Yungas no es suficientemente conocida. Sin embargo, su comercialización no tiene mayores dificultades, ya que forman parte de una familia botánica muy difundida por su madera valiosa. La mayoría de los productores que han tenido experiencias de exportación de productos de cedro, lo hicieron a través de intermediarios. Esto se debe principalmente a dos factores: 1) los intermediarios conocen perfectamente como se maneja el mercado y 2) los requisitos legales de exportación no son temas que el productor quiera manejar. Las exportaciones son de bajos volúmenes y esporádicas y los mercados de destino actuales son Inglaterra, España, 203 Zorrilla, A. Líbano y Estados Unidos. En la década de los noventa hubo exportaciones de algunas industrias que se interrumpieron debido a la política económica de paridad cambiaria. Al salir de la convertibilidad y con precios más competitivos, las exportaciones volvieron paulatinamente a crecer. Los productos que están siendo comercializados en este mercado son tablones de 1 y 2 pulgadas, con 15 % de humedad y de anchos y largos máximos; zócalos y machimbres con 8 % de humedad; batientes para puertas y ventanas; y pisos. Uno de los objetivos actuales es el mercado chino, presentándose diferencias según sea para el mercado doméstico chino o para reexportación. En el primer caso, los principales productos demandados son pisos y partes de muebles. Mientras que en el segundo, se suma madera dimensionada para muebles de exterior y es necesario una certificación FSC, ya que se re-exporta a Inglaterra y Alemania. El consumidor externo busca medidas largas, anchas y de primera calidad. Con estos elevados requerimientos, de 1 m3 se obtienen 50 pies madereros FAS (Sistema de Clasificación Americano FAS = First and Second). Para comparar los rendimientos, si el comprador no tuviera estos requerimientos de calidad, se obtendrían de 1 m3 de madera, 150 pies madereros de medidas cortas y angostas. En general, los precios de los productos comercializados no varían con respecto al mercado nacional. El cedro rosado tiene un sobreprecio con respecto al cedro orán del 5 al 10 %. Según la empresa Forestal 204 Santa Bárbara, algunos de los valores que se manejan por pié cuadrado sin considerar IVA son: - Primera calidad: US$ 1,10 (madera de 2,10 m y 2x4 pulgadas), - Segunda calidad: US$ 0,93, - Tercera calidad: US$ 0,76. Las fortalezas del mercado de exportación son: 1) la demanda está asegurada si se cuenta con un buen producto y precios competitivos; 2) se considera que los precios van en alza y 3) existe la posibilidad de agregar nuevos productos a la cadena industrial. Algunas características del mercado de exportación son vistas como debilidades del sistema de producción por parte de los industriales. El mercado internacional tiene requerimientos de calidad de aserrado, “packaging” (embalaje), volúmenes y porcentajes de humedad que hacen difícil conformar al comprador, debido a las condiciones actuales de producción en el noroeste de Argentina. Además de esto, la zona de producción está alejada de los puertos de exportación (más de 1.000 km) y el costo del flete incide de manera importante en la comercialización. Las perspectivas del mercado internacional son favorables siempre y cuando los costos locales no suban de manera significativa (fletes, impuestos, etc). Otro punto importante es que la relación cambiaria entre el peso y el dólar se mantenga. Finalmente, la mayoría de los industriales consultados creen que es indispensable una reconversión industrial para ser más Mercados para la madera de cedro competitivos en este mercado tan exigente, con incorporación de tecnología, capacitación del personal, mejoras en la gestión empresarial y también en los canales de comercialización. LA CERTIFICACIÓN FORESTAL VOLUNTARIA Y EL NEGOCIO DEL CEDRO En la década de los ochenta se impusieron fuertes boicots principalmente en Europa, orientados a no comprar productos de madera proveniente de bosques nativos tropicales y subtropicales. El objetivo de estas campañas, emprendidas por grandes organizaciones ambientalistas con el apoyo de los consumidores, era detener la destrucción de las selvas tropicales lluviosas, como la Selva lluviosa del Amazonas, los bosques de Java, de la Isla de Borneo, de Congo y otros. Estas campañas no dieron resultado, porque el propietario de las selvas concluyó que si nadie le compraba su madera era mejor desmontar y producir granos (soja, poroto, etc) que se venden facilmente en el mercado mundial. Como respuesta a esta iniciativa y con el objeto de revertir la respuesta negativa de los productores forestales de los países tropicales, comenzó una proliferación de sellos “ecológicos” o “verdes”. Estos los habilitaba básicamente para entrar en los mercados europeos, mediante la certificación de productos forestales que provenían de un manejo ambientalmente adecuado. En 1993 una nueva corriente instaló el FSC (Forest Stewardship Council) una organización sin fines de lucro con sede actual en Bonn, Alemania (en sus comienzos había estado en Oaxaca, México). Está conformada por miembros que se reúnen en Cámaras para discutir los estándares de manejo forestal certificado y actualizarlos. El FSC acredita a empresas para que efectúen las certificaciones y las audita permanentemente. Este Consejo de Manejo Forestal cuenta con el apoyo de grupos ambientalistas como Greenpeace, WWF (World Wildlife Fund) y Amigos de la Tierra (Friends of the Earth); además de recibir financiación de varias entidades privadas. Pronto se fueron sumando organizaciones de empresas, empresas individuales, organizaciones comunitarias, comunidades aborígenes, otras ONGs y centros de estudio. A partir de los ’90 el FSC comenzó a certificar en forma independiente bosques nativos de diferentes partes del mundo. Si bien, el enfoque primero hizo énfasis en los bosques tropicales, hasta el momento, son los bosques templados y boreales los que más han crecido en superficie certificada. Los requisitos que una operación forestal debe cumplir para lograr la certificación forestal del FSC están descriptos en un documento llamado “Principios y Criterios del FSC”. Dicho documento establece que la operación forestal (que no es más que un bosque bajo aprovechamiento) debe cumplir con todas las leyes que regulan al sector (las laborales, las ambientales y otras que puedan existir localmente) y debe demostrar clara tenencia de la tierra (puede ser propietario o no, pero debe tener derechos para hacer el aprovechamiento). Todos sus empleados y los de sus contratistas deben desarrollar sus activida205 Zorrilla, A. des bajo buenas condiciones laborales, en cuanto a salarios, seguros, aportes, capacitación, edades, campamentos, transporte, equipos, etc. Otro punto fundamental es la interacción que se tiene con comunidades aborígenes, comunidades rurales y vecinos en general, en caso de que se vean afectadas por el aprovechamiento forestal. El proceso de certificación tiene tres componentes: la parte interesada que es el propietario (también puede ser un grupo de propietarios, cooperativa o administrador de recursos forestales), un comprador de productos certificados y un certificador que se apoya en estándares para asegurar que el productor transmita un mensaje cierto al consumidor. Las operaciones forestales que pasan exitosamente el proceso de evaluación de campo para certificación, utilizan un logotipo en sus productos. Con esta certificación se amplían las posibilidades de comercializar en mercados muy exigentes, con buenos precios y demanda constante. Grandes cadenas de compradores a nivel mundial (mueblerías, fábricas de aberturas, etc.) y distribuidores, buscan proveerse de maderas certificadas. El consumidor final es el que moviliza toda la cadena y lleva a un industrial o a un intermediario a comprar maderas certificadas. Tal es el caso de los países con una cultura ambientalmente fuerte que apunta a no ser co-responsable de la destrucción de los bosques y la explotación de su gente. Los mercados más exigentes en estas condiciones son países de Europa como Inglaterra, Alemania, Holanda, Dinamarca, Francia; pero no dejan de ser importantes determi206 nadas zonas de Estados Unidos y Canadá, así como Japón y China. El FSC hace hincapié en que las operaciones forestales deben conocer primero y utilizar después, todos los bienes y servicios que brinda el bosque para maximizar el valor económico y social de éstos, pero siempre en su medida justa sin sobre-explotar los recursos. Las operaciones forestales deben tener un Plan de Manejo donde se describen todas las operaciones que se llevan a cabo en el campo, la forma de monitorearlas, evaluarlas y ajustarlas. El monitoreo de las actividades y la interpretación de sus resultados es fundamental para tener registros que permitan mejorar las prácticas forestales. Los bosques y sitios con alto valor de conservación son celosamente considerados por la certificación forestal. Se restringen las actividades en lugares con endemismos o con características únicas de biodiversidad y en áreas con valor social y cultural. Si las operaciones forestales se desarrollan en áreas con alto valor de conservación deben ser proactivas (tomar medidas favorables) para minimizar el impacto y resguardar estos ecosistemas. La certificación forestal avala buenas prácticas forestales, sin importar si corresponden a bosques nativos o plantaciones. Todos los Principios y Criterios son aplicados por igual, con la diferencia que en plantaciones forestales se consideran además el uso de agroquímicos, de organismos con alteraciones genéticas o riesgos de monocultivo en grandes extensiones, entre otras. Mercados para la madera de cedro El FSC es uno de los sistemas de certificación forestal voluntaria que están siendo aplicados en el mundo, aunque hay otros más como el SFI (Estados Unidos), CSA (Canadá), PEFC (Europa), CerFor (Chile) y CertFlor (Brasil). Sin embargo, el FSC es el único que cuenta con el apoyo de una gran red de organismos y organizaciones que le dan mayor credibilidad. El sistema de certificación forestal es voluntario, es decir, que no constituye una exigencia de algún país para permitir el ingreso de madera y otros productos provenientes de bosques nativos o implantados. En América Latina los países con mayor cantidad de hectáreas de bosques nativos certificados son Bolivia y Brasil. Argentina cuenta hasta el momento con una sola operación forestal de bosques nativos certificada (Forestal Santa Bárbara). Esta operación hace aprovechamiento forestal sustentable en Yungas en las Provincias de Salta y Jujuy (ACRB). Las maderas que trabaja son cedro, nogal, quina, palo barroso, palo amarillo, palo blanco, cebil, lapacho, mora amarilla y otras. Otras operaciones en la misma región están actualmente orientando su actividad hacia la certificación forestal y la producción de “madera controlada” que es una categoría menos exigente de garantizar que la operación está en proceso de mejora continua de su desempeño empresarial orientada a la sustentabilidad social y ambiental de su actividad. CONCLUSIONES Varias empresas del noroeste de Argentina venden exitosamente productos de cedro en el mercado externo. Las principales dificultades de este mercado están dadas por la alta calidad de aserrado que exigen los compradores y los requerimientos de secado. Las escuadrías que se venden son grandes y todas de primera calidad, generando volúmenes importantes de madera de segunda y tercera calidad que sólo se pueden vender hasta el momento, en el mercado doméstico. Cuando se conocen y se aplican exitosamente los requerimientos del mercado externo, la madera de cedro es fácil de colocar en el exterior. Algunas de las empresas exportadoras son: Forestal Santa Bárbara, Maderera del Norte, Madenort y Maderera Las Moras. En la actualidad, están desarrollándose nuevos proyectos en la región con vistas a industrializar este tipo de maderas. En ese sentido, esta alternativa creciente de operaciones forestales dependerá de armar esquemas de manejo sustentables del bosque nativo, que valoricen el recurso y que generen una demanda laboral adecuada y fundamentalmente respetuosa de los derechos y de la calidad de vida de quienes participan en la actividad forestal. Esto implica un cambio profundo, tanto en el manejo del recurso, como también de la cultura de la relación laboral en el negocio forestal en la región noroeste de Argentina. 207 Mapas y Fotografías Localidades mencionadas en el texto y áreas protegidas en el noroeste de Argentina y sur de Bolivia en el área de distribución de Cedrela. Distribución de las tres especies de Cedrela en relación a la distribución de Yungas en Argentina y sur de Bolivia. Cedro de gran tamaño en la antigua estancia El Rey. Fotografía del archivo de la Administración de Parques Nacionales. Fecha aproximada 1947. Tocón de cedro que dio origen al actual P. N. El Rey. Fotografía del archivo de la Administración de Parques Nacionales. Fecha aproximada 1947. Aserradero en la zona de Orán en la década del treinta. Fotografía gentileza Ingenio San. Martin de Tabacal. Viga labrada con hacha en la ciudad de Orán. Julio 1948. Fotografía de H. Chagra. Ambiente de Selva Montana en la Sierra de San Javier, Tucumán. Entre las nubes se destacan grandes ejemplares de cedro. Fotografía de I. Paz Posse. Bosque Montano en la Reserva Nacional El Nogalar de Los Toldos, Salta. Fotografia de M. Molinillo. Cedrela lilloi en un ambiente de Selva Montana, en PN. Calilegua, Jujuy. Fotografía de A. Grau. Ambiente de Bosque Montano en la Finca San Andres, Salta. Fotografía de Tramontini/ProYungas. Ejemplar de cedro coya en la quebrada de Osma en la Provincia de Salta. Fotografía de R. Neumann Cedro coya en la Reserva Natural El Nogalar de los Toldos, Salta. Fotografía de L. Malizia. Cedrela lilloi en la Quebrada de las Cañas, Horco Molle, Tucumán. Fotografía de A. Grau. Ejemplar de cedro coya en los alrededores de Tiraxi, Jujuy. Fotografía de J. Llanes. Copa de C. lilloi en Horco Molle, Tucumán. Fotografía de A. Grau. Flor masculina y femenina de cedro. Fotografía de A. Grau. Inflorecencia de cedro. Fotografía de A. Grau. Inflorescencia de C. lilloi y detalle de las flores estaminada y pistilada. Fotografía de A. Grau. Frutos de cedro coya de los alrededores de Tiraxi, Jujuy. Fotografía de J. Llanes. Renoval de cedro coya. Fotografía de L. Malizia. Acarreo de troncos de cedro en áreas de fuerte pendiente en Salta. Fotografia de R. Villalba. Camión viguero con rollos de cedro en la Provincia de Salta. Fotografía de R. Villalba. Plantación de Cedrela balansae de cinco años de edad (plantación experimental en Valle Morado, Salta). Fotografía de Tramontini/ Proyungas. Tablas obtenidas por el método de “rallado”, en los alrededores de Los Toldos, Salta. Fotografía de R. Villalba. Rollos de cedro en un aserradero de la ciudad de Orán, Salta. Fotografía de A. Brown. Tablas de cedro y productos en preparación en una carpintería de la ciudad de Orán, Salta. Fotografía de E. Balducci. Puerta de cedro en la iglesia del Ingenio Tabacal, Salta. Fotografía de Tramontini/ ProYungas. Bateas de cedro realizadas por artesanos de Los Toldos. Fotografía de D. Rode. Mango de guitarra elaborado con madera de cedro. Fotografía de F. Jayat.