EXTENSIONES DEL ANÁLISIS MENDELIANO Sara Arce y Enric
Anuncio

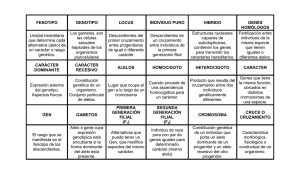
Sara Arce Gallego Enric Bertran Garcia de Olalla EXTENSIONES DEL ANÁLISIS MENDELIANO Interacciones entre genes. Epistasias: explicaciones bioquímicas La genética bioquímica es la disciplina que se encarga de estudiar las relaciones entre genes y enzimas. Diferentes genes cooperan para la obtención del producto final. Aquí se exponen algunos ejemplos que permiten justificar algunas epistasias estudiadas. Proporciones mendelianas 9:3:3:1 A modo de introducción, podemos hacer referencia a la segunda ley de Mendel, ya que a partir de las proporciones mendelianas 9:3:3:1, podemos entender el resto de proporciones expuestas a continuación. Nos fijaremos en el ejemplo de los pimientos Capsicum annuum. El color en el fruto de este vegetal viene determinado de la siguiente manera: Genotipo Y+_C+_ Y+_cc yyC+_ yycc Fenotipo rojo "red" naranja "peach" naranja "orange" blanco "cream" El color en estos pimientos resulta de su contenido en carotenoides de color rojo y amarillo, los cuales se sintetizan mediante rutas bioquímicas. El locus Y codifica para una enzima (primer paso de la ruta) y el C codifica para otra (último paso). Sara Arce Gallego Enric Bertran Garcia de Olalla Epistasia recesiva 9:3:4 Estas epistasias están presentes y se observan pues en genes que determinan el color del Labrador retriever. Estos suelen ser negros, marrones o amarillos, y la diferencia de estos vendrá determinada por las interacciones entre genes en dos locus. Un locus determina el tipo de pigmento producido por las células de la piel : un alelo dominante B codifica para el pigmento negro, mientras que el alelo recesivo correspondiente "b" lo hace para el marrón Por otro lado los alelos del segundo locus afectan a la deposición del pigmento en el eje del pelo : de esta manera el alelo E permite que el pigmento oscuro (bien negro o marrón) sea depositado, mientras que el recesivo "e" prevendrá esta deposición, causando pues que el pelaje sea amarillo. La presencia del genotipo ee en el segundo locus por tanto enmascara la expresión de los alelos negro y marrón en el primero locus. Los genotipos que determinan el color del "abrigo" del pelaje y sus fenotipos son : Genotipo Fenotipo B_E_ negro "black" bbE_ marrón "chocolate" B_ee amarillo bbee amarillo En el caso en el que cruzásemos un Labrador negro homocigoto para los alelos dominantes con uno amarillo homocigoto para los alelos recesivos, la progenie obtenida al cruzar la F1, sería una F2 en proporciones 9:3:4 . En este ejemplo de interacción de genes, el alelo e es epistatico de B y b, porque "e" enmascara la expresión de los alelos para los pigmentos marrón y negro, y los alelos B y b son hipostáticos para "e". En este caso, e es un alelo epistásico recesivo, ya que dos copias de "e" deben estar presentes para enmascarar a los pigmentos nuevamente negro y marrón. Sara Arce Gallego Enric Bertran Garcia de Olalla Epistasia dominante 12:3:1 Estas epistasias se ven en la interacción de dos locus determinantes del color de la calabaza de verano, comúnmente observable en amarillo, blanco o verde. Cuando una planta homocigota que produce estas calabazas blancas se cruza con otro homocigoto pero en este caso de color verde, se obtiene tras el cruce de la F1, los siguientes resultados: ¿Cómo puede la interacción de genes explicar estos resultados?En la F2 , como hemos visto, 12/16 de las plantas producen blacas y 3/16+1/16=4/16=1/4 de las plantas producen calabazas con color. Este resultado es el ya conocido 3:1 producido por el cruce entre dos heterocigotos, lo que sugiere que un alelo dominante en un locus inhibe la producción de pigmento, resultando así la progenie blanca. Si usamos el símbolo W para representar el alelo dominante que inhibe la producción de pigmento, entonces el genotipo W_ inhibe la producción de pigmento produciendo así el blanco, permitiendo en el caso de ww el pigmento, con los consiguientes resultados con color. Entre ambas plantas ww en F2 con fruto coloreado, observamos 3/16 amarillas y 1/16 verdes (relación 3:1). Este resultado se debe a que un segundo locus determina el tipo de pigmento producido, amarillo (Y_) domina sobre verde (yy). Este locus se expresa sólo en plantas ww, a las que les falta el al·lelo dominante inhibidor. Podemos asignar el genotipo wwY_ a plantas que producen calabazas amarillas y el genotipo wwyy a las que lo hayan verde. Los genotipos y sus fenotipos asociados serán: Genotipo Fenotipo W_Y_ Blancas "white squash" W_yy Blancas "white squash" wwY_ Amarillas "yellow squash" wwyy Verdes "green squash" El alelo W es epistasico de Y e "y" (suprime la expresión de este pigmento) produciendo genes. W es un alelo epistásico dominante porque , en contraste con el "e" en Labrador retriever explicado anteriormente, una sola copia de este alelo es suficiente para inhibir la producción de pigmento. Las calabazas de verano nos ofrecen una buena oportunidad o ejemplo para considerar cómo las epistasias a menudo surgen cuando los genes afectan a una serie de pasos de una vía bioquímica. El pigmento amarillo en la calabaza es más probable que se produzca en dos pasos de la vía bioquímica. El compuesto incoloro (blanco) es convertido por la enzima I (todo lo mencionado representado en la Sara Arce Gallego Enric Bertran Garcia de Olalla siguiente y última foto), en el compuesto verde B, el cual es convertido en el C por la acción del enzima II. Este último compuesto es el del color amarillo. Plantas con el genotipo ww producen enzima I y en ocasiones la verde o amarilla dependiendo de si la enzima II está presente. Cuando el alelo Y está presente en el segundo locus, la enzima II es producida y el compuesto B convertido en C, produciendo un fruto amarillo. Cuando dos copias de "y" ,las cuales no codifican una enzima II funcional, está presente, la calabaza se vuelve verde. La presencia de W en el primer locus inhibe la conversión del compuesto A en B; plantas con genotipo W_ no hace que el compuesto B y su fruto se vuelva blanco, independientemente de qué alelos estén presentes en el segundo locus. Muchos casos de epistasia surge de esta manera. El gen A (como el W) el cual tiene un efecto en una temprana fase de la ruta bioquímica, será epistatico para genes (como Y e "y") que afecta a la subsecuencia de pasos de la ruta, porque el efecto de la enzima en la tardía depende del producto de esta temprana o de las primeras fases. Epistasia recesiva doble 9:7 Ahora consideraremos otro ejemplo detallado de epistasia. El albinismo es la ausencia de pigmento y es un rasgo genético común en muchas plantas y animales. El pigmento es casi siempre producido a través de una ruta bioquímica de muchos pasos; por lo tanto, el albinismo puede implicar interacción entre genes. Robert T.Dillon y Amy R.Wethington encontraron que el albinismo en el caracol de agua dulce común Physa heterostroha puede resultar de la presencia de cualquiera de los dos alelos recesivos en dos locus diferentes. Se obtuvieron caracoles inseminados de una población natural y fueron colocados en recipientes con agua, donde estos pusieron sus huevos. Algunos de los huevos eclosionaron en caracoles albinos. Cuando se cruzaron dos caracoles albinos, toda la F1 eran pigmentados. El cruzamiento de esta, produjo una F2 que consistía en caracoles tanto pigmentados como albinos. ¿Cómo se dio esta relación 9:7? La relación 9:7 observada en esta F2 como hemos mencionado, puede ser entendida como una modificación de la relación 9:3:3:1 obtenida cuando dos individuos heterocigotos para dos locus se cruzan. La proporción 9:7 surge cuando alelos dominantes para ambos locis (A_B_) producen caracoles pigmentados; cualquier otro genotipo produciría caracoles albinos. La relación 9:7 en estos caracoles está producida probablemente por dos pasos en la vía de producción de pigmento. El pigmento (compuesto C) es producido solamente después de que el compuesto A se haya convertido a B por el enzima I y después de que este B se haya convertido en C por el enzima II. Sara Arce Gallego Enric Bertran Garcia de Olalla Al menos un alelo dominante A en el primer locus es requerido para producir enzima I, y igualmente al menos un alelo dominante B en el segundo locus es requerido para producir el enzima II. El albinismo surge en la ausencia del compuesto C, lo cual puede suceder de 3 maneras. La primera de ellas: dos alelos recesivos en el primer locus (genotipo aaB_) puede prevenir la producción del enzima I, y por tanto el compuesto B nunca será producido. Por segunda parte, dos alelos recesivos en el segundo locus (genotipo A_bb) pueden prevenir la producción del enzima II. En este caso el compuesto B nunca será convertido en compuesto C. Por último, el tercer caso, dos alelos recesivos pueden estar presentes en ambos locus (aabb), causando pues la ausencia de ambos enzimas I y II. En este ejemplo de interacción entre genes, "a" es un epistàsico de B, y "b" lo es de A; ambos son alelos epistáticos recesivos porque la presencia de dos copias de cualquier alelo a o b, es necesario para suprimir la producción de pigmento. Este ejemplo difiere de la supresión del color del abrigo de pelo del Labrador retriever en que estos alelos recesivos en cualquiera de los dos locus son capaces de suprimir la producción de pigmentos en los caracoles, mientras que alelos recesivos en un solo locus suprime la expresión de pigmento en Labradores. Tabla resumen de las epistasias