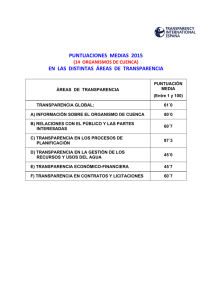
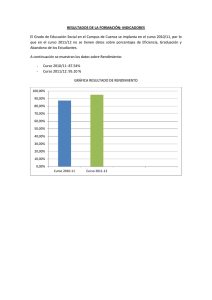
XXIX Jornadas de Paleontología
Anuncio

EN MEMORIA La Sociedad Española de Paleontología tiene el placer de dedicar este Libro de Resúmenes al Profesor Don Jaime Truyols Santonja, primer Presidente de la Sociedad y maestro de muchas generaciones de paleontólogos XXIX Jornadas de Paleontología Simposio del Proyecto PICG 596 LIBRO DE RESÚMENES C. Álvarez-Vázquez e I. López Rodríguez (eds.) Real Jardín Botánico de Córdoba Córdoba, 2013 © De los textos y figuras: los autores © De la edición: Real Jardín Botánico de Córdoba Imprime: I.S.B.N.: 978-84-938181-2-8 Depósito legal: Diseño de portada: Elena Moreno Barroso Fotos de portada: Carmen Álvarez-Vázquez, Mª Luisa Martínez Chacón, Isabel Rábano, Pedro Cózar y Juan Carlos Gutiérrez Marco. Composición y diseño del libro: C. Álvarez-Vázquez e I. López Rodríguez XXIX Jornadas de Paleontología “La Paleontología del Paleozoico” y Simposio del Proyecto PICG 596 Córdoba, 2 a 5 de octubre de 2013 Organizado por: Centro Paleobotánico, Real Jardín Botánico de Córdoba Departamento de Paleontología, Universidad Complutense de Madrid Sociedad Española de Paleontología Con la colaboración de: Ayuntamiento de Córdoba Delegación de Agricultura y Medio Ambiente, Diputación de Córdoba Programa Internacional de Correlación Geológica (IUGS, UNESCO) COMITÉ ORGANIZADOR Presidente: Roberto Wagner (Real Jardín Botánico Córdoba) Secretaria: Carmen Álvarez Vázquez (Real Jardín Botánico Córdoba) Vocales: Sergio Rodríguez (Universidad Complutense Madrid) John Knight (Derbyshire, Inglaterra) Inmaculada López Rodríguez (Real Jardín Botánico Córdoba) Elena Moreno Barroso (Real Jardín Botánico Córdoba) Juan Antonio Madueño Trujillo (Real Jardín Botánico de Córdoba) Elena Moreno Eiris (Universidad Complutense Madrid) Antonio Perejón (Universidad Complutense Madrid) Coordinadores Excursión Pre-Congreso: Elena Moreno Eiris (Universidad Complutense Madrid) Silvia Menéndez (IGME) Antonio Perejón (Universidad Complutense Madrid) Coordinadores Excursión Post-Congreso: Sergio Rodríguez (Universidad Complutense Madrid) Roberto Wagner (Real Jardín Botánico Córdoba) Carmen Álvarez Vázquez (Real Jardín Botánico Córdoba) Pedro Cózar (Universidad Complutense Madrid) Ismael Coronado (Universidad Complutense Madrid) COMITÉ CIENTÍFICO Julio Aguirre Rodríguez Departamento de Estratigrafía y Paleontología, Universidad de Granada María José Comas Rengifo Departamento de Paleontología, Universidad Complutense de Madrid Ángela Delgado Buscalioni Departamento de Biología, Universidad Autónoma de Madrid Esperanza Fernández Martínez Facultad de Ciencias Biológicas y Ambientales, Universidad de León Eladio Liñán Guijarro Departamento de Ciencias de la Tierra, Universidad de Zaragoza Jordi Martinell Callicó Departament d’Estratigrafía, Paleontología i Geociències Marines, Universidad de Barcelona Eduardo Mayoral Alfaro Departamento de Geodinámica y Paleontología, Universidad de Huelva Eustoquio Molina Martínez Departamento de Ciencias de la Tierra, Universidad de Zaragoza Jorge Morales Romero Museo Nacional de Ciencias Naturales, Madrid Isabel Rábano Gutiérrez del Arroyo Museo Geominero, Madrid Luis Carlos Sánchez de Posada Departamento de Geología, Universidad de Oviedo José Ignacio Valenzuela Ríos Departamento de Geología, Universidad de Valencia Presentación El pasado año 2012, el Real Jardín Botánico de Córdoba celebró el XXV aniversario de su inauguración. La actividad desarrollada por esta Institución, hoy día como parte esencial del Instituto Municipal de Gestión Medio Ambiental (IMGEMA), ha permitido que goce de un merecido prestigio que le ha convertido en uno de los más conocidos y con mayor prestigio en el ámbito de la comunidad conservacionista y científica. El Centro Paleobotánico, creado en 1983 en base a la donación de la colección del Profesor Roberto Wagner, forma parte del mismo, con su Colección y Museo, desde los primeros años de su gestación. Que la Sociedad Española de Paleontología nos haya designado como sede de sus XXIX Jornadas nos honra y llena de orgullo. Nuestras instalaciones, el gran valor patrimonial de la colección de Paleobotánica de la que somos custodios, y el ilusionado trabajo que se desarrolla en el Centro Paleobotánico hacen que nuestra Institución sea un lugar ideal para celebrar la reunión anual de esta Sociedad. A esto habría que añadir el marco incomparable de la ciudad de Córdoba. La celebración de un Congreso de estas características es siempre el resultado del trabajo y la colaboración de muchas personas e Instituciones. Desde aquí quiero agradecer la colaboración desinteresada de los miembros del Comité Organizador, Comité Científico y Junta Directiva de la Sociedad Española de Paleontología, así como la de los conferenciantes invitados, la de los coordinadores de las Sesiones Científicas y del Simposio PICG 596, y la de todos los trabajadores del Real Jardín Botánico de Córdoba. Mencionar también que la edición de este volumen ha sido posible gracias a la colaboración de la Delegación de Agricultura y Medio Ambiente de la Diputación de Córdoba. El éxito de la convocatoria es evidente por la calidad de las 86 contribuciones científicas que se reúnen en este libro y que resumen una parte de las investigaciones paleontológicas que se están realizando actualmente en España. Hay que agradecer a todos los ponentes su participación y aportación a las Jornadas. Pero, las Jornadas no se celebran sólo para presentar nuevos trabajos y discutir cuestiones científicas. Son el lugar para reencontrarse con los colegas, charlar, conocer a los miembros más jóvenes de la Sociedad, … para ver y ser visto y, por supuesto, también para disfrutar de la ciudad que las acoge. La ciudad de Córdoba que lo alberga, se siente orgullosa como anfitriona y sobre todo por tener la oportunidad de enseñar su rico Patrimonio cargado de un legado de distintos pueblos y culturas que le han dado forma a lo largo de los siglos. Romanos, Visigodos, Árabes, Judíos y Cristianos nos ofrecen una Ciudad Patrimonio de la Humanidad y del Mundo. El enclave de Córdoba nos da una mezcla bien estructurada de Patrimonio edificable y Patrimonio Natural que es su Río Guadalquivir y su Sierra, que es un ejemplo del Bosque Mediterráneo. Cultura y Gastronomía, Costumbres y Fiestas son sin duda, una buena carta de identidad absolutamente envidiables y exquisitas. Les ánimo a trabajar para que estas Jornadas sobre Paleontología sean todo un éxito y para que disfruten de esta nuestra ciudad que les acoge y que ya es la suya también. Cuando regresen a sus destinos de origen, deseo que guarden el mejor recuerdo de Córdoba y se lleven los mejores recuerdos de ella. Por todo ello, les doy la bienvenida. Córdoba, septiembre de 2013 Rafael Jaén Concejal de Patrimonio, Casco Histórico y Naturaleza Ayuntamiento de Córdoba Presidente del Real Jardín Botánico de Córdoba Jaime Truyols en el recuerdo Como todos los miembros de la Sociedad Española de Paleontología conocen, el pasado día 28 de agosto falleció en Oviedo Jaime Truyols Santonja (para muchos de nosotros Don Jaime), maestro de paleontólogos y de geólogos que cultivan variadas disciplinas de las Ciencias de la Tierra. A todos los miembros de la comunidad paleontológica y del conjunto de la comunidad geológica se nos ha ido una persona que es, a la vez, amigo y persona crucial en el desarrollo de la Paleontología moderna en España. Afortunadamente, nos queda su obra, plasmada en escritos, enseñanzas y en su ejemplo. Como atinadamente señala Leandro Sequeiros en mensaje dirigido a los paleontólogos españoles, el desenlace era esperado, pero no por ello menos sentido. Con toda propiedad puede aplicarse al profesor Truyols el término de “maestro” en el más noble sentido de la palabra. Su vida académica fue una magistral lección de vida para todos los que tuvimos el privilegio de recibir sus enseñanzas y de crecer científicamente bajo su tutela. Dotado de amena oratoria no exenta de cierta solemnidad, dotado, también, de un estilo literario en el que se conjugaban amenidad y rigor, provisto de encomiable capacidad de síntesis, cultura enciclopédica y espíritu humanista, el profesor Truyols ejerció su magisterio en el aula, en el laboratorio, en el campo, en reuniones y congresos científicos, en su propio domicilio, e incluso en placenteras charlas en bares o restaurantes, en torno a un buen plato o alrededor de una buena botella de vino que tanto le deleitaban. Magisterio académico, sí, pero también magisterio vital; al fin, para producir buenos frutos ambos deben combinarse. Autoridad sostenida en un prestigio bien ganado, capacidad para escuchar, meditar y asimilar ideas ajenas, espíritu abierto a la comprensión de problemas científicos o de situaciones difíciles, y un profundo sentido de la amistad eran otras tantas características de su personalidad. De ello pueden dar testimonio muchos paleontólogos y muchos geólogos, desde Finisterre hasta el Cabo de Gata o desde los confines nororientales de su Cataluña natal hasta el Cabo San Vicente, pero también numerosos colegas diseminados por los cinco continentes. Como sucede con los auténticos maestros, su obra ha trascendido su propia vida. Su hija, Montse es la primera conocedora y heredera de su obra. Desde estas líneas compartimos su dolor. No es éste el lugar adecuado para dar detalles concretos de la vida de Jaime Truyols, que, por otra parte, se encuentran reflejados en diversos artículos, como, por ejemplo, “Bio-bibliografía del Prof. Jaime Truyols Santonja” publicado en el “Liber amicorum”, que como homenaje le dedicaron el Departamento y la Facultad de Geología de la Universidad de Oviedo (“Trabajos de Geología”, 21) o la semblanza “Jaime Truyols. Paleontólogo, Maestro y primer Presidente de la Sociedad Española de Paleontología”, que vio la luz en el número extraordinario que le dedicó esta sociedad también en 1999. En el contexto de su fructífera vida poco importan en estos momentos algunos artículos más que añadir a su extensa bibliografía, pero no podemos resistirnos a señalar dos hitos que marcarían la vida del profesor Truyols, que discurrió en un periodo crucial para el desarrollo de la Paleontología, marcado por el nacimiento y desarrollo de la Teoría Sintética de la Evolución y por la influencia de G.G. Simpson. Uno de tales hitos es su incorporación a la sección de Paleontología del Museo de Sabadell, tras finalizar la licenciatura en Ciencias Naturales; el otro, su traslado Oviedo y, especialmente, la obtención de la cátedra de Paleontología de dicha universidad en 1964. El Museo de Sabadell era ya entonces una institución importante en la innovación de la paleontología española. Allí Jaime Truyols realizó una intensa y fructífera labor en torno a los materiales neógenos de la Península Ibérica, bajo la dirección del profesor Miquel Crusafont y entró en contacto con reputados especialistas en el campo de la Paleomastología (Piveteau, Viret, Von Koeningswald, entre otros). Especial resonancia tuvieron los artículos morfométricos y evolutivos, como “A biometric study of the evolution of Fissipid carnivores” (Evolution X) o “Masterometry and evolution again” (Evolution 20). Su traslado a Oviedo (año 1961) y, especialmente, la obtención de la cátedra de Paleontología (año 1964), representó un profundo cambio en la vida del profesor Truyols: la renuncia a la línea de investigación que había desarrollado hasta entonces. Sin duda no fue ésta una decisión fácil, pero sí (al menos desde la óptica del Paleozoico cantábrico) acertada y generosa. En él prevalecieron las ideas de que había contraído el compromiso de contribuir al conocimiento de la geología del Paleozoico y que la Sección de Ciencias Geológicas, a la que se incorporaba, debía acoger equipos de investigación aglutinados en torno a la geología de la región. Así creó especialistas en los grupos más relevantes del Paleozoico (especialmente del Devónico y Carbonífero), procurando que estancias en diversos países abrieran nuevos horizontes a aquellos jóvenes investigadores. La renuncia al estudio del Cenozoico y el cambio de los mamíferos por los invertebrados sin duda no fueron fáciles, pero, todos los que formamos parte de su equipo damos testimonio de lo atinado de su dirección, la valía de sus consejos y la ayuda que nos proporcionó su comprensión. Todos, sin excepción, nos consideramos afortunados. Un equipo como aquel no habría podido consolidarse sin su sabia dirección. Hoy, recortes presupuestarios y sesgadas visiones de la ciencia y la universidad amenazan seriamente su continuidad. Pero su vida científica y sus vivencias no quedaron limitadas a la zona cantábrica. Desde Oviedo, mirando al horizonte y con su vista superando las nieblas y la orografía cantábrica, Jaime Truyols mantuvo relaciones científicas y humanas con investigadores de muy distintas procedencias que cultivaban disciplinas muy diversas. Subrayar su papel en la constitución de nuestra sociedad y en el alumbramiento de nuestra revista resulta ocioso. La Sociedad Española de Paleontología tampoco hubiera sido la misma sin su fundacional presidencia y su prudente y decidido impulso. Aún a riesgo de resultar repetitivos queremos reiterar las palabras expresadas ya hace catorce años en el número extraordinario de la Revista Española de Paleontología: Nosotros, sus prójimos, somos conscientes de habernos encontrado en el curso de nuestras vidas con una persona prudente, de ánimo pacificador y recto criterio científico y humano en quien siempre se puede confiar. La Paleontología y la Sociedad Española de Paleontología han tenido la misma fortuna. No podría ser de otro modo. La personalidad del profesor Truyols tenía que proyectarse obligatoriamente con fuerza pareja hacia las personas, hacia la disciplina paleontológica y hacia los instrumentos creados para el desarrollo de esta rama del saber. En nombre de todos, gracias, Don Jaime. Gracias por sus lecciones, por sus consejos (que nunca fueron órdenes), y por su amistad. En definitiva, gracias por su magisterio. Como ocurre con los auténticos maestros siempre quedará algo del Profesor Truyols entre nosotros Luis C. Sánchez de Posada y Jenaro García-Alcalde Departamento de Geología, Universidad de Oviedo Índice de contenidos El Carbonífero de la Zona Cantábrica: sucesión estratigráfica y registro paleontológico (Conferencia inaugural) L.C. Sánchez de Posada …………………………………………………………...………………….. 23 El significado geológico de las cuencas carboníferas con flora fósil en Sierra Morena (Conferencia de clausura) R.H. Wagner ……………………………………………………………………….………………….. 41 COMUNICACIONES ORALES Bioestratigrafía de alta resolución con foraminíferos planctónicos del límite Cretácico/ Paleógeno de Moncada, Cuba I. Arenillas, J.A. Arz y R. Rojas-Consuegra …………………………………………….…………… 59 New plesiosaur specimens from the Guadalajara Province and a review of the Iberian Peninsula plesiosaur fossil record N. Bardet, A. Pérez-García, L.M. Castillo de la Fuente, X. Pereda Suberbiola and P. Vincent …....... 61 The plant fossil record just reveal two great extinction events B. Cascales-Miñana and C.J. Cleal ………………………………………………….......….................. 63 Composition and Dynamics of the Great Phanerozoic Evolutionary Floras C.J. Cleal and B. Cascales-Miñana ………………………………………........…….……................… 64 “Dinoichnofacies” barremienses de Teruel (España) A. Cobos, F. Gascó, R. Royo-Torres y L. Alcalá …………………………………..….……………… 65 Estimando la masa corporal de los hiénidos fósiles (Mammalia, Carnivora, Hyaenidae) a partir del esqueleto dental y postcraneal. Implicaciones paleoecológicas y evolutivas C. Coca Ortega y J.A. Pérez Claros …………………………...……………………………….…...… 67 Geología y Paleontología del yacimiento finicretácico de La Solana (Valencia) J. Company, X. Pereda Suberbiola y J.I. Ruiz-Omeñaca …………………………………………...… 69 Escenario paleoambiental de la Crisis de Salinidad del Messiniense en la Cuenca del Bajo Segura (sureste de España) H. Corbí ………………………………………………………………………………………………. 71 Bailiella cf. tenuicincta (Linnarsson, 1879), una fauna escandinava en el Cámbrico medio de las Cadenas Ibéricas (NE de España) J.B. Chirivella Martorell, R. Gozalo, M.E. Dies Álvarez y E. Liñán ……………….………………… 73 Evolución de la Complejidad en el cráneo de tetrápodos: una aproximación a partir de la teoría de redes B. Esteve-Altava, J. Marugán-Lobón, H. Botella y D. Rasskin-Gutman ………………………….….. 75 Cambio ambiental y evolución del comportamiento depredador en cánidos extintos B. Figueirido, A. Martín-Serra, C. Casares-Hidalgo, P. Palmqvist, J.A. Pérez-Claros y C.M. Janis … 77 Análisis crítico de la “icnita” de Arroyo Cerezo, primera huella de dinosaurio descrita en España J.A. Gámez Vintaned y M. Belinchón ………………………………...………...……………….…… 79 Experiencias de divulgación del Patrimonio Paleontológico de la Sierra de Aracena y Picos de Aroche I. Garzón González ………………………………………………….………………………………… 81 Asociaciones de macrofósiles en rocas metamórficas del Complejo Nevado-Filábride (Zonas Internas de la Cordillera Bética) en Águilas, Murcia (SE España). Tafonomía y biocronoestratigrafía C. Laborda-López, J. Aguirre y S. Donovan ………………………………………..………………… 83 El GSSP de la base del Piso Santoniense en la “Cantera de Margas”, Olazagutia, Navarra M.A. Lamolda, C.R.C. Paul y J. Mª Pons ……………………………………………………..……… 85 Microvertebrados del nuevo yacimiento “Sondeo Galería” del Calvero de la Higuera (Pleistoceno Medio final, Pinilla del Valle, Madrid) C. Laplana, P. Sevilla, H.-A. Blain, C. Araujo, J.L. Arsuaga, E. Baquedano y A. Pérez-González …. 87 Inferencias sobre las habilidades locomotrices de la subfamilia Borophaginae Canidae, Carnivora, Mammalia) mediante el estudio morfométrico de su esqueleto apendicular A. Martín-Serra, B. Figueirido, F.J. Serrano y P. Palmqvist …...………….……………………….… 89 El registro de Paroniceratinae en España y su distribución paleogeográfica G. Martínez y M.J. Comas-Rengifo ……………………………………………………...…………… 91 Icnología de medios marinos someros en el Neógeno Superior del Archipiélago de Cabo Verde E. Mayoral, J. Ledesma-Vazquez, B.G. Baarli, A. Santos, R. Ramalho, M. Cachão, C.M. da Silva y M.E. Johnson ……………………………………………………………………………….…………. 93 Oxfordian ammonites from Gerecse and Pilis Mountains (Hungary): Some taphonomic and biostratigraphic remarks G. Meléndez and I. Fözy ………………………………………………………………..........……….. 95 Microbioerosión en esqueletos de invertebrados marinos del Plioceno temprano de la región de Murcia A.R. Molinu, R. Domènech, D. García-Ramos y J. Martinell ………………………...……………… 97 Acritarchs from the Ordovician of the Ossa-Morena Zone (SW Spain); new biostratigraphic data T. Palacios and S. Jensen ……………………………………………………………..………………. 99 Actividad hidrotermal en los ambientes lacustres de edad Mioceno final a Pleistoceno medio de la cuenca de Baza y sus implicaciones paleoecológicas P. Palmqvist, J.M. García-Aguilar, A. Guerra-Merchán, F. Serrano, A. Flores-Moya y B. Martínez-Navarro ………………………………………………………………………….………. 101 Evidence of gastropod-like predation on Lower Jurassic bivalves from the Iberian Peninsula and the Mesozoic quiescence phase R. Paredes ……………………………………………………………………………….…………….. 103 Tortugas europeas continentales tras la crisis finicretácica: Sistemática y paleobiogeografía A. Pérez-García …………………………………………………………………………..…………… 105 Revisión del estatus del podocnemídido europeo Papoulemys (Chelonii, Pleurodira) A. Pérez-García y F. de Lapparent de Broin ……………………………………………..…………… 106 Las tortugas gigantes del Mioceno medio de la Meseta Castellana: Revisión del (malparado) material clásico A. Pérez-García y E. Vlachos ………………………………………………………………………… 107 Compartiendo escenario en el Jurásico Superior de Asturias. Huellas y rastros de pterosaurios, cocodrilos y dinosaurios L. Piñuela, J.C García-Ramos y J.I. Ruiz-Omeñaca ……………………………………..…………… 109 Dientes de condrictios del Triásico Medio de las cordilleras Costero-Catalanas C. Pla, H. Ferrón, A. Márquez-Aliaga y H. Botella …………………………………………...……… 111 Tendencias y patrones de diversidad de los conodontos durante el Triásico P. Plasencia, C. Martínez-Pérez, B. Cascales-Miñana, M. Mazza y H. Botella ……………………… 112 Asociaciones de foraminíferos del Toarciense inferior en la Sección de San Andrés (Cuenca Vasco-Cantábrica) R.W. Salazar Ramírez y C. Herrero ………………………………………………………...………… 113 Presencia de Podichnus en braquiópodos órtidos del Ordovícico Inferior (Tremadociense) del NW de Argentina A. Santos, E. Mayoral, E. Villas, Z. Herrera y G. Ortega ……………………..……………………… 115 Primer registro fotográfico de una excavación paleontológica de cetáceos: el fósil de Cuevas del Almanzora (Almería) de 1878 J.R. Sendra Sáez ………………………………………………………...…………………………….. 117 Icnofósiles (coprolitos) del Konservat Fossil-Lagerstätte del Plioceno de Cuevas del Almanzora (Almería) J.R. Sendra Sáez ………………...…………………………………………………………………….. 119 Tafonomía de la producción de cubiertas cementadas en mamíferos marinos J.R. Sendra Sáez ………………………………………………………………………………………. 121 Nuevos datos sobre la presencia de manglares (Familias Avicenniaceae y Rhizophoraceae) en el Plioceno superior de Cuevas del Almanzora, Cuenca de Vera (Almería) J.R. Sendra Sáez ……………………………………………………...……………………………….. 123 Evidencias tafonómicas de carroñeo o predación sobre cetáceos: el neurocráneo del Tortoniense de Burguillos (Sevilla) J.R. Sendra Sáez e I. Bajo Campos …………………………………….……………………………… 125 Estimación de la masa corporal y tendencias evolutivas en aves primitivas F.J. Serrano, P. Palmqvist, A. Martín-Serra y J.L. Sanz …………………………….………………... 127 Paleoecología de las plantas vasculares en el Maastrichtiense inferior de las Cuencas Surpirenaicas S. Villalba-Breva, J. Marmi, B. Gomez, C. Martín-Closas y V. Daviero-Gomez …………….……… 129 PÓSTERS Asociaciones fósiles del Eoceno medio-superior de la Cuenca de Pamplona y áreas colindantes (Navarra) H. Astibia, J. Elorza, S. Ortiz y A. Payros ……………………………………….…………………… 133 Los braquiópodos del Bajociense superior-Bathoniense inferior del Subbético Oriental. Datos preliminares J.F. Baeza-Carratalá, F. García Joral y J. Sandoval …………………………………...……………… 135 La “fauna de koninckínidos”. Precursores de la extinción del Toarciense Inferior (ETOAE) en el Subbético Oriental J.F. Baeza-Carratalá, F. García Joral, J.E. Tent-Manclús y A. Giannetti ……………...……………… 137 Bioestratigrafía comparada y bioeventos (ammonoideos y foraminíferos) del tránsito Aaleniense/Bajociense (Jurásico Medio) en Hontoria del Pinar (Noroeste de la Cordillera Ibérica) M.L. Canales, M.S. Ureta, L. Hernández y A. García-Frank ………………….……………………… 139 Estudio preliminar de los microvertebrados del yacimiento del Pleistoceno Medio de La Salema (Facheca, Alicante) E. Colomina, F.J. Ruíz-Sánchez, V.D. Crespo, S. Mansino, R. Marquina y P. Montoya ……..……… 141 Ammonites del Valanginiense inferior (Cretácico inferior) de la Cordillera Bética. Nuevos datos bioestratigráficos M. Company Sempere y J.M. Tavera Benítez ………………...……………………………………… 143 Primer registro del género Eumyarion (Mammalia, Cricetidae) en la cuenca de Ribesalbes-Alcora (Castellón, España) V.D. Crespo, F.J. Ruiz-Sánchez, M. Freudenthal, S. Mansino, R. Marquina, E. Colomina y P. Montoya ……………………………………………………………………………………………. 145 Unionoids (Bivalvia) and their associated microbialites from the Late Jurassic of Asturias (Spain) G. Delvene, R. Lozano, M. Munt, L. Piñuela and J.C. García-Ramos …………………………..…… 147 Los ornitópodos de Barrihonda-El Humero en la Formación Villar del Arzobispo de Riodeva (Teruel, España) F. Gascó, J. Verdú, A. Cobos, R. Royo-Torres y L. Alcalá ………………………………...………… 149 Sedimentología, tafonomía e icnología de secuencias sedimentarias en una plataforma abierta mixta del Cretácico inferior (Provincia de Alicante, Cordillera Bética) A. Giannetti, P. Monaco, H. Corbí y J.F. Baeza-Carratalá …………………………………………… 151 Ordovician and Devonian trace fossils from the Ossa-Morena Zone of south-west Spain, with a first report of Lophoctenium comosum Richter from the Ossa-Morena Zone S. Jensen and T. Palacios ………………………………………..……………………………………. 153 Una nueva forma del género Eliomys (Gliridae, Mammalia) del tránsito MiocenoPlioceno en el Este de la Península Ibérica S. Mansino, A. García-Alix, F.J. Ruiz-Sánchez, V.D. Crespo, E. Colomina, R. Marquina y P. Montoya …………………………………………………………………………………………….. 155 Valoración patrimonial del complejo de yacimientos de La Muntanyeta Blanca d´Almenara (Almenara, Castellón) R. Marquina, F.J. Ruiz-Sánchez, V.D. Crespo, S. Mansino y E. Colomina ………………………….. 157 Interés didáctico de las rutas geológico-paleontológicas en la Cordillera Ibérica y el Pirineo (Castilla La Mancha-Aragón, NE España) G. Meléndez, A. Calonge, G. Fermeli y R. Barella …………………………………………...……… 159 Registro de los foraminíferos durante el Óptimo Climático del Eoceno Inferior en el paleomargen continental noribérico S. Ortiz, A. Payros, I. Millán, E. Apellaniz, J. Arostegi, N. Martínez-Braceras y X. Orue-Etxebarria ……………………………………………………………………………………. 161 La colección de arqueociatos del Cerro de las Ermitas de Córdoba del Museo Geominero (IGME): nuevos datos sobre su origen A. Perejón, S. Menéndez, I. Rábano y E. Moreno-Eiris ……………………………………………… 163 Una nueva especie de Neochelys (Chelonii, Pleurodira) en el Eoceno inferior de Francia A. Pérez-García y F. de Lapparent de Broin ………………………………………………………….. 165 Reconocimiento de Paleotestudo en España e identificación como un taxón muy abundante en niveles aragonienses y vallesienses A. Pérez-García y X. Murelaga …………………………………………….…………………………. 166 Diversidad y sistemática de las tortugas del Mioceno medio de Mara-II (Zaragoza) A. Pérez-García, X. Murelaga y J.I. Canudo …………………………..……………………………… 167 Interés cronoestratigráfico de los bioeventos de foraminíferos planctónicos registrados en torno al GSSP de la base del Santoniense (Olazagutía, Navarra) I. Pérez-Rodríguez, J.A. Arz e I. Arenillas …………………………………………………………… 169 Icnofósiles del Ordovícico inferior de Aldeaquemada (Sierra Morena, Jaén): consideraciones taxonómicas, paleoecológicas y paleogeográficas F.J. Rodríguez-Tovar, M. Stachacz, A. Uchman y M. Reolid ………………………...……………… 171 Sobre el registro de Megacricetodon (Mammalia, Cricetidae) en el tránsito Mioceno inferior/Mioceno medio en la cuenca del Río Magro (Valencia, España) F.J. Ruiz-Sánchez, M. Freudenthal, V.D. Crespo, S. Mansino, R. Marquina y E. Colomina …….….. 173 Revisión taxonómica de Frogdenites y Labyrinthoceras (Sphaeroceratidae, Ammonitina) del Tethys Occidental J. Sandoval ……………………...…………………………………………………………………….. 175 Un Metaxytherium (Mammalia, Sirenia) del Tortoniense de Paterna (Valencia) J.R. Sendra Sáez …………………………………………………….………………………………… 177 Ctenophora fósil del Konservat Fossil-Lagerstätte del Plioceno de Cuevas del Almanzora, Cuenca de Vera (Almería) J.R. Sendra Sáez ………………………………………………………………………………………. 179 Un escaldicétido (Mammalia, Cetacea, Physeteridae) del Tortoniense de la Cuenca de Alcoy (Alicante) J.R. Sendra Sáez ……………………………………………………………….……………………… 181 Scaldicetus degiorgii Varola, Landini & Pilleri 1988 (Mammalia, Cetacea, Scaldicetidae) del Mioceno (Tortoniense) de la Cuenca del Guadalquivir J.R. Sendra Sáez e I. Bajo Campos ………………………………………………………..………….. 183 Un Metaxytherium del Plioceno inferior de Cuevas del Almanzora (Almería) J.R. Sendra Sáez y F.A. Fortea Soler ……………………………………………………...………….. 185 Bats in islands: new data from the Pliocene-Early Pleistocene fossils from Menorca (Spain) P. Sevilla, J. Quintana and M. Furió ……………………………………………………..…………… 187 Análisis bioestratigráfico basado en los foraminíferos del tránsito AalenienseBajociense (Jurásico Medio) en la sección Serra da Boa Viagem II (Cuenca Lusitánica, Portugal) S. Silva, M.L. Canales y M.H. Henriques ……………………………………………..……………… 189 Tapetes microbianos como elemento clave en la conservación de tejidos blandos y huesos I. Villalba, M. Iniesto, M.C. Guerrero, A.D. Buscalioni y A.I. López-Archilla ……….……………... 191 The Carboniferous forest at Verdeña (Palencia): A case study for the conservation of a forest preserved as imprints R.H. Wagner, L.J. Sardina and J.B. Diez ……………………………………………...……………… 193 SIMPOSIO PICG 596 Evolución temprana de las especies de Idiognathodontidae (conodontos) en torno al límite Misisípico/Pensilvánico S. Blanco Ferrera y J. Sanz López ……………………………………………...…………………….. 197 Primera aproximación al estudio de los montículos de fango del Devónico Inferior (Lochkoviense-Praguiense) de la Formación Lebanza (Zona Cantábrica) E. Fernández-Martínez, L.P. Fernández, C. Vera de la Puente y A. Ernst …………………………… 199 Análisis de los patrones de escamación en telodontos e inferencias sobre su modo de vida H. Ferrón y H. Botella ………………………………………………………………………………… 201 Estudio de los patrones de escamación y el modo de vida de tiburones actuales para inferencias paleoecológicas en taxones fósiles H. Ferrón, C. Martínez-Pérez y H. Botella ……………………………………………………………. 203 Conodontos, biofacies y microfacies del Givetiense de Renanué (Devónico Medio, Pirineos Aragoneses) J-C. Liao y J.I. Valenzuela-Ríos ……………………………………………………………………… 205 Kozlowskia (Productidina, Brachiopoda) del Bashkiriense superior/Moscoviense inferior (Carbonífero), Cordillera Cantábrica (N de España) M.L. Martínez Chacón y C.F. Winkler Prins ………………………………………………….……… 207 Aplicación del Synchrotron X-Ray tomographic microscopy al estudio funcional de los elementos conodontales C. Martínez-Pérez y P.C.J. Donoghue ………………………………………………………...……… 209 Patrones de integración morfológica en el género Polygnathus (Conodonta) C. Martínez-Pérez y S. de Esteban Trivigno ………………………………………………………….. 211 Polygnáthidos del Emsiense en los Pirineos Centrales Españoles y su importancia para la comprensión de la evolución temprana del grupo C. Martínez-Pérez y J.I. Valenzuela-Ríos …………………………………………………………….. 213 Cirtospiriféridos del Devónico de la Zona Centroibérica meridional M.V. Pardo Alonso ……………………………………………………………………………………. 215 Corales rugosos del Misisípico de Alabama (USA) S. Rodríguez y D.C. Kopaska-Merkel ………………………………………………………...……… 217 Revisión del criterio de correlación mediante conodontos para el límite Misisípico/Pensilvánico J. Sanz López y S. Blanco Ferrera ……………………………………………………………………. 219 Correlación de las secuencias de conodontos del Lochkoviense (Devónico Inferior) de Nevada (USA) y de los Pirineos Centrales Españoles J.I. Valenzuela-Ríos y M.A. Murphy …………………………………………………………….…… 221 Conodontos del Lochkoviense (Devónico Inferior) en los Pirineos Centrales Españoles y su correlación con las secuencias de Bohemia (República Checa) J.I. Valenzuela-Ríos, L. Slavík, H. Calvo, A. Hušková, J-C. Liao y L. Koptíková …………………... 222 Formaciones Escalada y Fito (Unidad del Ponga, Zona Cantábrica): Contenido en fusulinas y correlación C. González Parra y E. Villa Otero …………………………………………………………...………. 223 C onferencia inaugural 22 El Carbonífero de la Zona Cantábrica: sucesión estratigráfica y registro paleontológico L.C. Sánchez de Posada Departamento de Geología, Universidad de Oviedo, c/ Jesús Arias de Velasco s/n, 33005 Oviedo, España. [email protected]. Introducción El Carbonífero de la Zona Cantábrica ocupa una extensión superficial de unos 9000 km2 de las provincias de Asturias, León, Cantabria y Palencia, y se sitúa dentro de lo que Lotze (1945) ha denominado Zona Cantábrica (en adelante ZC), la parte externa del hercínico español, que ha experimentado deformación en condiciones superficiales, sin desarrollo del metamorfismo que caracteriza a las áreas más internas. Los detallados estudios sobre el índice de alteración de los conodontos (llevados a cabo sobre todo en las unidades del Ponga y Picos de Europa, ver Fig. 1) muestran valores correpondientes a la diacaizona, aunque en algunos enclaves puedan alcanzarse valores próximos al límite ancaizona/epicaizona. Una revisión de las regiones tectonosedimentarias en que se ha dividido la ZC (Fig. 1) da cuenta de que los materiales carboníferos afloran en todas ellas, aunque, como consecuencia en gran parte de su carácter sinorogénico, tienen características muy distintas en unas áreas y en otras. Baste por el momento decir que, si se exceptúa el Estefaniense continental discordante, Figura 1. Mapa geológico sintético de la ZC, mostrando las principales unidades de ésta. (Basado esencialmente en Julivert, 1971 y Pérez Estaún et al., 1988). 23 tres vectores básicos señalan tales diferencias: 1) La edad de los afloramientos más altos del Carbonífero aumenta de oeste a este (Bashkiriense en la Unidad de Somiedo-Correcillas, parte baja del Moscoviense en la Unidad de la Sobia-Bodón, Moscoviense terminal en la Cuenca Carbonífera Central de Asturias y Unidad del Ponga, y Gzheliense en la Unidad de Picos de Europa); 2) Los sedimentos marinos alcanzan edades más altas cuando más al este nos situemos, y 3) En conjunto, las calizas llegan a tener edades también más altas en las unidades más orientales. Varios autores dividieron la sucesión carbonífera Cantábrica en tres tramos. El inferior, que se extiende desde la base del sistema hasta el Serpukhoviense (Namuriense, Arnsbergiense), tiene naturaleza condensada, característica bastante constante a lo largo de la región, y según numerosos autores representa la transición entre las sucesiones pre- y sinorogénica. El tramo medio (que se extiende hasta el Asturiense, ex Westfaliense D) está constituido por sucesiones que muestran considerables diferencias litológicas entre distintas regiones y que son normalmente consideradas como sucesiones sinorogénicas, depositadas en una cuenca de antepaís. Finalmente, el tramo superior, de edad Asturiense tardío y Estefaniense, muestra sucesiones parálicas en unas regiones e intramontañosas en otras. El contenido paleontológico del primero de los intervalos es notablemente distinto del de los dos intervalos superiores. En efecto, en la parte inferior de la sucesión, los elementos macropaleontológicos son los cefalópodos, que aparecen acompañados por algunos trilobites, braquiópodos, bivalvos y restos de crinoideos. Los microfósiles más estudiados son, sin duda, los conodontos (que junto a los cefalópodos constituyen herramientas clave en la datación). Igualmente se han descrito ostrácodos, radiolarios y microforaminíferos. En contraste, en la parte alta de la sucesión, los macrofósiles mayoritarios son braquiópodos, corales y, en algunos tramos bivalvos y gasterópodos. Los hallazgos de cefalópodos son muy esporádicos. El registro micropaleontológico está constituido esencialmente por foraminíferos y ostrácodos, y, en menor proporción, por conodontos. Algunos tramos han proprorcionado abundante macro y microflora. Una muestra de los fósiles de origen marino y continental que aparecen en estos intervalos se muestra en las Figs 2 y 3. En las líneas siguientes destacamos los rasgos fundamentales de estos tres tramos (Fig. 4), junto con algunas características relevantes de sus asociaciones fósiles. Dada su continuidad geográfica con las sucesiones del tramo medio, una parte de las sucesiones del tramo superior se describen conjuntamente con aquellas. En cambio, se trata aparte el Estefaniense continental, Figura 2. Fósiles marinos del Carbonífero de la Cordillera Cantábrica. 1-2. Discosiphonella mammilosa (King, 1943) (porífero esfinctozoo), Demués (Asturias), Formación Demués, Kasimoviense, barra de escala = 10 mm (García-Bellido et al., 2004). 3-4. Secciones transversales de corales rugosos, Pensilvánico, Cordillera Cantábrica: 3. Petalaxis sp.; 4. Asturiphyllum sp. (Braga et al., 2009). 5. Foraminíferos fusulinoideos, ejemplares de los géneros Fusulina y Fusulinella, Formación Fito, Moscoviense superior, aproximadamente x 14 (fotografía E. Villa). 6-7. Braquiópodos: 6. Kutorginella cf. stepanovi (Lapina, 1957), vista anterior de la valva ventral, Calizas del Cuera, tránsito Bashkiriense/Moscoviense, Sierra del Cuera (Asturias), x 1,5 (Martínez Chacón & Bahamonde, 2008); 7. Tegulispirifer sp., vista ventral, Calizas del Cuera, Moscoviense superior, Hontoria (Llanes, Asturias), x 1,75 (Martínez Chacón, 1991). 8-9. Prolecanites primitivus (Kullmann, 1963) (cefalópodo ammonoideo), Formación Alba, Viseense, Matallana (León), x 1,5 (Olóriz & Rodríguez-Tovar, 2009). 10. Hollinella (Praehollinella) aff. camoni Bless, 1968 (crustáceo ostrácodo), Mina La Camocha (Gijón, Asturias), transición Bashkiriense/Moscoviense, aproximadamente x 45. 11. Declinognathodus bernesgae SanzLópez et al., 2006 (conodonto), Formación Alba, Serpukhoviense, Millaró (León), aproximadamente x 70 (Sanz-López et al., 2006). 24 25 discordante, que hoy aparece como afloramientos dispersos, en enclaves que muchas veces representan importantes cuencas carboníferas (en el sentido minero del término). Figura 3. Algunos ejemplos característicos de la macroflora de la Cordillera Cantábrica. a. Neuropteris ovata Hoffman (pteridosperma), Cuenca Carbonífera de Guardo (Palencia), Asturiense (ex Westfaliense D) superior, x 1. b. Pecopteris camertonensis Kidston (helecho; detalle de unas pínulas fértiles), Peñacorba, Barruelo (Palencia), Cantabriense superior, x 3. c. Diplazites emarginatus Göppert (helecho), Cuenca Carbonífera de Villablino (León), Estefaniense B, x 3. d. Linopteris palentina Wagner (pteridosperma), Tejerina (León), Cantabriense inferior, x 3. Todos los ejemplares pertenecen a la colección de Paleobotánica del Centro Paleobotánico del Real Jardín Botánico de Córdoba. La parte inferior de la sucesión (Tournaisiense-Kinderscoutiense) Incluye las formaciones Baleas, Vegamián, Alba, Cuevas (u Olleros) y Barcaliente, con características y asociaciones fósiles muy distintas. Las dos formaciones basales del Carbonífero (Baleas y Vegamián) son, al menos en parte, sincrónicas, pero tienen características litológicas y paleontológicas muy distintas: la primera está formada por unos 10 m de calizas claras bioclásticas, y la segunda por lutitas negras que, en 26 conjunto, encajan bien en la denominación de “black shales”. La formación Baleas ha proporcionado pocos restos fósiles reconocibles; los niveles más bajos de la formación corresponden al Fameniense terminal (Zona de praesulcata) y los más altos al Tournaisiense superior (Zona de anchoralis latus). La formación Vegamián es un intervalo estratigráfico depositado en condiciones disaeróbicas o anaeróbicas, de baja tasa de sedimentación y de aguas calmas. Conodontos, ostrácodos, braquiópodos, trilobites, bivalvos y algún cefalópodo son los fósiles más estudiados en la formación Vegamián. Distintos autores han subrayado la semejanza faunística de la formación Vegamián con diversas localidades de la “facies Kulm” de Europa occidental. La formación Alba (Formación Genicera o “caliza griotte”), que sigue a las anteriores, es la unidad litoestratigráfica más ampliamente extendida en la ZC. Está constituida mayoritariamente por calizas rojas o rosadas, con algunas intercalaciones lutíticas y, en ocasiones, por una banda de chert intermedia. Cefalópodos y conodontos son sus fósiles más conocidos, pero a ellos deben añadirse otros (aunque en general no muy abundantes), como microforaminíferos, ostrácodos, corales, trilobites, bivalvos, braquiópodos y crinoideos. Si bien la litología de la Formación Alba es muy diferente a la de la Formación Vegamián, el contenido faunístico, al menos en lo que se refiere a trilobites, braquiópodos, bivalvos y, en parte ostrácodos, sigue mostrando afinidades con la facies Kulm de Europa occidental. Cefalópodos y conodontos son los elementos básicos para su datación. La formación se extiende entre el Tournaisiense superior y el Serpukhoviense. En los últimos años, las secciones de Millaró (provincia de León), Vegas de Sotres (Parque Nacional de Picos de Europa, Asturias) y Triollo (provincia de Palencia) han alcanzado notable relevancia en el estudio global del límite Viseense/Serpukhoviense. Las tres muestran sucesiones continuas, bien expuestas y con abundante fauna de conodontos, entre ellos representantes de la línea filética Lochriea nodosa-L. ziegleri; la última de estas especies probablemente será designada como indicador biostratigráfico del inicio del piso Serpukhoviense. La sección de Vegas de Sotres, al contrario de lo que ocurre en otras secciones de la ZC, contiene, además, abundantes foraminíferos calcáreos retrabajados que se encuentran en estudio. En relación con los niveles de tránsito entre el Viseense y el Serpukhoviense, debemos hacer mención, finalmente, a la sucesión de Triollo (encuadrada en la Región del Pisuerga-Carrión y, a veces, considerada como una formación independiente), que, además de conodontos, ha proporcionado una abundante fauna de ostrácodos, con elementos característicos de la denominada “Megasociación Turíngica”, típica de ambientes de aguas calmas y, en general, distales. En muchas secciones de la Unidad de Somiedo-Correcillas, sobre la Formación Alba se sitúa una sucesión esencialmente detrítica, con tramos turbidíticos y algunas intercalaciones calcáreas (Formación Cuevas o Formación Olleros), que ha proporcionado conodontos y cefalópodos que alcanzan como máximo el Bashkiriense inferior (Kinderscutiense en la escala de Europa occidental). Se admite normalmente que representa un surco situado entre el orógeno y los ambientes en los que se depositó la formación Barcaliente. Tanto la parte superior de la formación Alba (Miembro Santa Olaja) como la parte baja de la Formación Cuevas, proporcionaron cefalópodos y ostrácodos pertenecientes a la “Megasociación Turíngica” y a la “Asociación de Entomozoidos”. Estas asociaciones de entomozoidos están constituidas esencialmente por una especie y representan uno de los registros más jóvenes de este tipo de asociaciones. En el resto de la región astur-leonesa, la Formación Alba está cubierta por 250 a 450 m de calizas micríticas, muy pobres en fósiles (Formación Barcaliente, parte baja de lo que se conocía como “Caliza de Montaña”). Su edad se ha establecido sobre la base de los cefalópodos contenidos en los niveles subyacentes (Serpukhoviense, Arnsbergiense en la escala estratigráfica de Europa occidental) y suprayacentes (Bashkiriense, Kinderscutiense, en términos de la escala 27 europea occidental), así como de algunos hallazgos de conodontos, encontrados a lo largo de la sucesión en varias secciones. No puede descartarse, sin embargo, que en el sector septentrional de la región del Ponga se alcancen edades más altas. En el contexto que nos ocupa, resulta necesario dedicar unas palabras a la problemática del denominado “Mid-Carboniferous Boundary” y su estudio en la ZC. La decisión de situar este limitotipo (límite entre los subsistemas Misisípico y Pensilvánico y entre los pisos Serpukhoviese y Bashkiriense) en la capa 61B de la sección de Arrow Canyon (Nevada) y de utilizar como indicador biostratigráfico la primera aparición de “Declinognathodus noduliferus s.l.” espoleó las investigaciones en niveles próximos a dicho límite en varias zonas del mundo, entre ellas España y, particularmente, la ZC. Los trabajos llevados a cabo en la Formación Barcaliente (ZC) y en la Formación Iraty (Pirineos) mostraron (Sanz-López et al., 2006) que en las secciones de Millaró y de Azcárate, cerca de la base de ambas formaciones, aparecía una nueva subespecie de D. noduliferus (D. noduliferus bernesgae) cuya primera aparición tenía lugar bastante por debajo de las formas de “Declinognathodus noduliferus s.l.” entonces conocidas. Ante este hecho, los autores mencionados analizaron la compleja problemática en torno a la definción del “Mid-Carboniferous Boundary” y de su caracterización bioestratigráfica. Parte de los problemas analizados derivan de lo que podría calificarse de difuso concepto de “D. noduliferus s.l.”. En el momento en que se estableció el limitotipo, D. noduliferus incluía 4 subespecies: D. noduliferus noduliferus, D. noduliferus lateralis, D. noduliferus inaequalis y D. noduliferus japonicus, hoy consideradas como especies independientes, y cuya aparición no era simultánea. En la práctica, la aparición del taxon índice nominal correspondía a la primera aparición del género. Sanz-López et al. (2006) consideran que la primera aparición de D. inaequalis es el mejor criterio de correlación con el limitotipo del límite Misisípico/Pensilvánico y que esta especie debería establecerse formalmente como indicador bioestratigráfico. La aparición de las sucesivas especies proporcionaría un marco interesante de correlación entre diferentes áreas, que debería ser refinado y complementado con otros criterios. Las partes media y alta de la sucesión (salvo el Estefaniense intramontano) El techo de la Formación Barcaliente (o de la Formación Cuevas en la Unidad de SomiedoCorrecillas) marca un incremento en la diversidad de la sucesión carbonífera en la Zona Cantábrica, aunque, en propiedad, la diferenciación comienza antes, durante el intervalo en el que en la Unidad de Somiedo-Correcillas se está depositando la secuencia terrígena de la Formación Cuevas y en amplias áreas de la región cantábrica se depositan las calizas de la formación Barcaliente. En la Unidad de La Sobia-Bodón, esta parte media de la sucesión está integrada por las formaciones Valdeteja y San Emiliano; en la Cuenca Carbonífera Central, por los grupos Lena y Sama (la Formación Valdeteja aflora sólo en la prolongación septentrional de la Cuenca Carbonífera Central por debajo de la cobertera postpaleozoica); en el sector central y meridonal de la Unidad de Ponga, por las formaciones Ricacabiello, Beleño, Escalada y Fito; en el sector septentrional de la misma unidad, por las denominadas “Calizas del Cuera”; en Picos de Europa, por las formaciones Valdeteja, Picos de Europa y algunos niveles situados por encima de ésta; y en la región del Pisuerga Carrión, por una amplia gama de materiales, muy diferentes en las distintas áreas (ver Figs 1, 4). La Formación Valdeteja (constituida por calizas más o menos masivas, frecuentemente bioclásticas, cuyo espesor puede alcanzar los 700 m) es rica en fósiles, y muchas veces incluye bioconstrucciones algales y microbiales. 28 Su parte baja tiene edad Bashkiriense inferior. La parte alta, notablemente diacrónica, puede alcanzar el Moscoviense inferior. Según diversos autores, este intervalo estratigráfico representa plataformas carbonatadas de márgenes inclinados, elevadas respecto a los ambientes circundantes. El paso de la Formación Barcaliente a la Formación Valdeteja supone un fuerte aumento en la variedad y abundancia de fósiles pertenecientes a grupos de naturaleza bentónica: (cianobacterias, algas calcáreas, foraminíferos, braquiópodos, corales, briozoos, y algunos trilobites, ostrácodos y bivalvos, como formas más representativas) y, también, un incremento en la oxigenación del fondo. Muchas de las formas de trilobites y braquiópodos son reminiscentes del Misisípico. Figura 4. Representación simplificada de la sucesión en varias unidades de la ZC (no se incluye la unidad del Pisuerga Carrión). 1. Unidad de Somiedo-Correcillas. 2. Unidad de La Sobia-Bodón. 3. Cuenca Carbonífera Central. 4. Unidad del Ponga. 5. Unidad de Picos de Europa. Unidades litoestratigráficas: B, Formación Baleas; Vg, Formación Vegamián; A. Formación Alba; Br, Formación Barcaliente; Cu, Formación Cuevas; Vl, Formación Valdeteja; SE, Formación San Emiliano; L, Grupo Lena; S, Grupo Sama; Rc, Formación Ricacabiello; Bñ, Formación Beleño; E, Formación Escalada; F, Formación Fito; PE Formación Picos de Europa; D, Formación Demués; LL, Formación Las Llacerias; P, Formación Puentellés; C, Formación Cavandi (modificado de Sánchez de Posada et al., 2001). En los últimos años, la paleoecología de los braquiópodos de la formación Valdeteja, junto con los presentes en las “Calizas del Cuera” y, de modo especial, las adaptaciones a la vida en sustratos duros ha sido analizada de modo detallado. Unidades de La Sobia-Bodón y del Aramo. Dentro de ellas, la sucesión continúa con la Formación San Emiliano, parcialmente sincrónica con la Formación Valdeteja. En su sección 29 tipo (valle del río Curueño, provincia de León) esta última llega a alcanzar el Moscoviense inferior, mientras que en la región de San Emiliano la parte media de la Formación San Emiliano es todavía Bashkiriense. En el área tipo (N de León), la Formación San Emiliano alcanza un espesor máximo de 1800 a 2000 m, con edades comprendidas entre un Bashkiriense relativamente bajo y el Moscoviense inferior (Vereyense). Por otra parte, los hallazgos de macro y microflora indican una edad Yeadoniense o Langsettiense temprano para la parte baja, y Duckmantiense para los niveles terminales. Estos datos señalan una correlación aproximada del límite Bashkiriense inferior/Bashkiriense superior con el límite Namuriense/Westfaliense y una equivalencia del límite Bashkirinese/Moscoviense con niveles correpondientes a la parte alta del Langsettiense o a la parte baja del Duckmantiense. Pero la Formación San Emiliano ha proporcionado muchos otros restos fósiles, tanto de organismos marinos, especialmente en el miembro medio, como continentales. Entre ellos pueden mencionarse, algas, foraminíferos, poríferos, corales, ostrácodos y, sobre todo, braquiópodos. Las algas y Donezella (que a veces forma montículos de considerables dimensiones) jugaron un papel importante en la constitución de estos materiales. Los braquiópdos, por su parte, son los macrofósiles mayoritarios. El grupo incluye un considerable número de formas que aparecen en terrenos más antiguos en otras áreas; al alcanzarse los niveles más someros de la parte superior, los braquiópodos cambian considerablemente y, aunque persisten algunas formas, aparecen otras características del Bolsoviense de Europa Occidental y del Moscoviese y aún de niveles superiores de la antigua Unión Soviética. Las características de los braquiópdos encajan bien con una posición geográfica de la ZC dentro del Paleotetis, quizás formando parte de un golfo en la región noroccidental. En la prolongación norte de la unidad de La Sobia-Bodón, la explotación minera de La Camocha ha permitido estudiar una sucesión, cubierta hoy por la cobertera postpaleozoica. La serie estratigráfica consta de unos 2300 m de alternancias de calizas, areniscas y capas de carbón. Los restos fósiles extraídos de La Camocha son abundantes: megasporas, microsporas, macroflora, braquiópodos, ostrácodos y cefalópodos. Los datos conjuntos de los hallazgos aislados de cefalópodos y de la micro y macroflora indican una edad Langsettiense y quizás Yeadoniense en los tramos más bajos, pero la base de la sucesión no ha sido cortada en el curso de las explotaciones mineras. Cuenca Carbonífera Central de Asturias. Constituye la unidad tectonoestratigráfica que hasta el declive de la minería proporcionó la mayoría del carbón de la región asturiana. Sus características estratigráficas más evidentes son la ausencia de depósitos pre-carboníferos (salvo un pequeño espesor de areniscas del Devónico Superior), el enorme espesor de la sucesión carbonífera (unos 6000 metros), y la profusión de capas de carbón explotables, especialmente en los niveles superiores. La sucesión sedimentaria presenta alguna característica en común con la de las unidades de La Sobia-Bodón y del Aramo (depósito sobre la “caliza de montaña” de una potente serie esencialmente detrítica con frecuentes capas de caliza en la parte baja y carbones en la parte alta). No obstante, las diferencias son también importantes: en la Cuenca Carbonífera Central la sucesión Westfaliense es mucho más potente, alcanza edades más altas, contiene mayor cantidad de conglomerados y las capas de carbón son mucho más numerosas. La división estratigráfica más utilizada actualmente en la Cuenca Carbonífera Central de Asturias parte de la ya antigua diferenciación de dos conjuntos (“Assise de Lena” y “Assise de Sama”; Barrois, 1882), a los cuales se da normalmente consideración de grupos (a veces formaciones), y que suelen subdividirse en paquetes mineros (a veces considerados también 30 como formaciones). Los materiales alcanzan una edad Myachkoviense, en términos de la escala estratigráfica global, Asturiense en términos de la escala de Europa occidental. Un punto controvertido es la posible existencia de materiales estefanienses en la parte alta de la sucesión, hecho planteado esencialmente a partir del estudio de la microflora. La sucesión de la Cuenca Carbonífera Central ha proporcionado, como cabe suponer, numerosos restos fósiles, predominantemente marinos en la parte inferior, y continentales en la superior. Un aspecto que debe ser destacado es la propuesta preliminar (Wagner et al., 2002) del Asturiense como un piso de la escala estratigráfica regional de Europa Occidental, en sustitución del antiguo Westfaliense D. El limitotipo propuesto (que debe ser estudiado con mayor profundidad) se sitúa en el denominado sector de Riosa-Olloniego (parte N de la Cuenca Carbonífera Central de Asturias), dentro de la parte alta del Paquete Canales, en el que algunos autores han mencionado la presencia de Neuropteris ovata, especie que históricamente se vinculó a la base del Westfaliense D. La presencia en el conjunto de la ZC de numerosas sucesiones del Westfaliense D en facies terrestres y marinas, con abundante contenido fósil, hace de la región un punto gran de interés en la caracterización paleontológica de las sucesiones de este tramo. Unidad del Ponga. Muestra dos tipos de sucesiones muy diferenciadas. En los sectores central y meridional, sobre la Formación Barcaliente se dispone un conjunto constituido por alternancia de tramos detríticos y carbonáticos (el más potente de estos últimos constituye la Formación Escalada). En cambio, en el sector septentrional, gracias a una progresiva sustitución de los materiales detríticos por materiales carbonáticos, la sucesión llega a ser enteramente calcárea, más semejante a la de la Unidad de Picos de Europa que a la del sector meridional de la unidad del Ponga. Salvo en la región septentrional, sobre la Formación Barcaliente aparece una sucesión condensada de lutitas, de edad comprendida entre el Serpukhoviense y el Bashkiriense superior (Formación Ricacabiello). La formación, que sustituye a la formación Valdeteja (y quizás incluso a tramos superiores) de otras áreas, ha proporcionado asociaciones variadas, claramente distintas de asociaciones coetáneas, tanto a escala regional como global, con un aspecto más antiguo que las edades a que pertenecen. Probablemente representan faunas de aguas calmas, situadas en la parte distal de la plataforma y desarrolladas sobre fondos blandos en un área que recibía escasos aportes. La Formación Beleño (alternancia de lutitas, limolitas, areniscas y algunas calizas en la parte superior, con espesor medio de unos 400 metros), que sucede a la Formación Ricacabiello, es muy pobre en fósiles. En la parte baja o media se ha encontrado una asociación con esporas atribuída al Langsettiense. La edad de los tramos más altos debe establecerse sobre la base de los foraminíferos contenidos en la parte baja de la formación siguiente, Formación Escalada (calizas masivas, claras, de unos 300 m de espesor), de carácter claramente diacrónico, lo mismo que la Formación Fito (alternancia de lutitas, areniscas y calizas). La edad de la Formación Escalada oscila entre el Moscoviense inferior (Kashiriense) y el superior (Podolskiense); la edad de la Formación Fito es Moscoviense superior, entre el Podolskiense y el Myachkoviense. Las edades son más jóvenes cuánto más orientales son las sucesiones estudiadas. Como se ha indicado, un importante cambio lateral de facies determina que en la Unidad del Ponga, la naturaleza de los materiales se haga progresivamente más calcárea hacia el noreste, de modo que en el sector norte, la sucesión es casi enteramente calcárea. La dificultad de diferenciar formaciones distintas ha llevado a denominar “Calizas del Cuera” a los tramos situados por encima de la formación Barcaliente. 31 En los último años, el sector septentrional de la Unidad del Ponga ha alcanzado considerable resonancia gracias a la existencia de una magnífica plataforma carbonatada (plataforma carbonatada del Cuera), de edad Bashkiriense a Moscoviense inferior, que ha sido minuciosamente estudiada por diversos autores. La estratificación vertical permite el estudio de una sección transversal de la plataforma, que representa una de las secciones transversales mejor conservadas de las plataformas pensilvánicas. Los detallados estudios llevados a cabo han posibilitado conocer la geometría de la plataforma, la contribución de distintos organismos en su construcción, los diferentes ámbitos de depósito, y la relación entre algunos de estos organismos y los ambientes. Una descripción de las facies puede verse en Della Porta et al. (2005), que estudiaron la distribución de distintos organismos en los ambientes de la plataforma. Contrariamente a las fusulinas, en la plataforma del Cuera los estudios sobre braquiópodos se refieren a faunas localizadas en la parte alta del talud de la misma. La mayoría de los braquiópodos estudiados proceden de los niveles de tránsito entre el Bashkiriense y el Moscoviense. Se trata de asociaciones variadas en las que muchos productidos muestran adaptaciones bastante peculiares que les permitieron vivir anclados al sustrato. Pero los braquiópodos de la Sierra del Cuera no son los únicos existentes en las denominadas “Calizas del Cuera”. En la costa, unos kilómetros al norte, aflora una sucesión, la denominada sección de Hontoria, Playa de La Huelga o San Antolín, que muestra, en facies casi exclusivamente calcárea, un conjunto de materiales que abarcan desde el Tournaisiense hasta el Moscoviense superior. Las fusulinas se distribuyen a lo largo de toda la serie, los conodontos son más abundantes en la parte baja (en facies atribuídas a ambientes de talud y pie de talud), y corales, braquiópodos y ostrácodos son más abundantes en la parte alta. Dos hechos destacables son la buena representación de los límites Serpukhoviense/Bashkiriense y Bashkiriense/ Moscoviense, y la existencia de niveles con variados palinomorfos en la parte alta. Estos restos, abundantes y bien conservados, han permitido establecer una edad Bolsoviense superior o Asturiense inferior para estos tramos, y correlacionarlos con los paquetes Caleras o Generalas de la Cuenca Carbonífera Central, y con la parte alta de la Zona de Forbach y parte baja de la Zona de St. Avold de la Cuenca de Lorena. En algunas escamas de la Unidad del Ponga, la sucesión carbonífera termina con un conjunto discordante. Está integrado por un espesor variable (máximo 250 m) de conglomerados, areniscas, lutitas, alguna capa de carbón y de calizas, depositados en abanicos aluviales costeros, con facies propias de abanicos subacuáticos, de playas, de cordones litorales e incluso de plataformas marinas carbonatadas o lutíticas. Los foraminíferos y braquiópodos indican un Myachkoviense terminal o un Kasimoviense basal. Unidad de Picos de Europa. El carbonífero de la región de Picos de Europa está constituido por una sucesión de más de 1000 m compuesta casi exclusivamente por calizas que abarcan desde el Tournaisiense hasta el Myaschkoviense, que comprende las formaciones Las Portillas (o Baleas), Alba, Barcaliente, Valdeteja, Picos de Europa y un conjunto de niveles superiores de características muy variadas. La sucesión recuerda a la expuesta en la parte N de la Unidad del Ponga, si bien en ésta no están representados los tramos más altos existentes en Picos de Europa. La Formación Picos de Europa, que se apoya sobre la Formación Valdeteja, es enteramente calcárea, y tiene un espesor aproximado de 600 metros. Los tramos basales están constituidos normalmente por calizas tableadas, con algunos niveles de chert y lutitas calcáreas (“Miembro Tableado”), mientras la parte superior está formada por calizas masivas, en su mayoría de tonos grises o rosados (“Miembro Masivo”). La formación tiene edades comprendidas entre el Moscoviense inferior (quizás Bashkiriense superior en algunos puntos) para su base, y Moscoviense superior (Myachkoviense) para el techo. 32 Como en la Unidad del Ponga, las formaciones Valdeteja y Picos de Europa representan una plataforma marina con dos partes diferenciadas: un área interna compuesta por packestones, wackestones y greinstones con restos esqueléticos variados y acumulaciones de riodofíceas y cianobacterias, y un área externa con disposición cliniforme. Como queda dicho, las rocas situadas inmediatamente por encima tienen litología muy variada, consecuencia de su naturaleza sinorogénica: sucesiones parálicas, especialmente visibles en el área noroccidental (sector de Gamonedo-Cabrales), calcáreas (Formación Las Llacerias y Formación Puentellés) o turbidíticas (Formación Cavandi). Algunos de estos tramos han proporcionado fusulínidos de edad comprendida entre el Moscoviense superior y el Gzheliense y floras del Cantabriense (en el área de Gamonedo/Inguanzo) y Estefaniense B (en el área de Cabrales). En los ultimos años, las secciones de Las Llacerias y Demués han llamado especialmente la atención. La primera abarca desde la base del Carbonífero hasta el Kasimoviense; algunos tramos (esencialmente los situados en la proximidades del límite Moscoviense/Kasimoviense, correpondientes a la parte alta de la Formación Picos de Europa y a la que le sucede, Formación Las Llacerias) han sido minuciosamente investigados dentro del marco del proyecto para el establecimiento del límite Moscoviense/Kasimoviense, proporcionando una serie de datos bioestratigráficos relevantes para la caracterización de dicho límite. La sección de Demués es, desde el punto de vista macrofaunístico, una de las sucesiones más fosilíferas y continuas del Kasimoviense de la ZC. Ha proporcionado asociaciones relativamente ricas de microflora, foraminíferos, poríferos, corales, ostrácodos y braquiópodos, así como de otros grupos que aún no han sido estudiados Los foraminíferos y palinomorfos dan datos bastante precisos de la edad de los distintos tramos en términos de la escala estratigráfica global y de la escala estratigráfica regional de Europa occidental. La sucesión abarca desde el Moscoviense superior (Myachkoviense) hasta la parte media del Kasimoviense (Khamonichiense) y, en términos continentales, probablemente desde el Cantabriense inferior hasta un nivel próximo al límite Barrueliense/Saberiense. Unidad del Pisuerga-Carrión. Tiene características bastante distintas del ámbito astur-leonés. Entre las más relevantes pueden mencionarse las siguientes: 1) Presencia de unidades alóctonas con facies devónicas más distales que las del ámbito astur-leonés. 2) Grado de metamorfismo algo mayor que el del Dominio Astur-Leonés. 3) Abundancia de discontinuidades estratigráficas a lo largo del Carbonífero. La sucesión carbonífera de la región del Pisuerga-Carrión presenta enormes diferencias entre unas áreas y otras, hasta el punto de que la geología de la región merecería una charla independiente. Las disparidades en los espesores, en la naturaleza de las rocas y en las edades representadas entre áreas próximas determinan que la región se haya subdividido en un conjunto de dominios con estratigrafía muy diferente. Como han subrayado numerosos autores, la presencia de condiciones sinorogénicas contínuas desde el Namuriense hasta el Estefaniense y la presencia de disconformidades mayores constituyen dos de las características más relevantes de la región. Ya en 1962, Koopmans estableció la existencia de tres grupos litoestratigráficos separados por discordancias: Ruesga (desde la base del Carbonífero hasta la discordancia de Curavacas), Yuso (desde la discordancia mencionada hasta la discordancia Astúrica), y Cea (desde la última a la discordancia Sálica, de edad Pérmico inferior), divididos en numerosas formaciones de validez en los distintos sectores que constituyen la Unidad del Pisuerga-Carrión. Wagner (1970) ha señalado la existencia de tres disconformidades mayores, correspondientes a tres fases de deformación: Palentina, de edad Westfaliense A inferior (sobre ella se deposita la 33 formación conglomerática de Curavacas), Leónica, de edad Westfaliense D, y Astúrica de edad Estefaniense A (reconocible sólo en la parte occidental). La Fig. 5 muestra las formaciones diferenciadas en distintas áreas de la unidad. Figura 5. Esquema de correlación de las unidades estratigráficas de la Unidad del Pisuerga-Carrión (reproducido de Fernández et al., 2004). Especial relevancia en el ámbito general de la estratigrafía del Carbonífero tiene la propuesta sucesiva de dos pisos (Cantabriense y Barrueliense) que, en la situación actual de los trabajos encaminados a la construcción de una escala estratigráfica global del Carbonífero, deben considerarse como parte de la escala estratigráfica regional del Carbonífero de Europa occidental. El Cantabriense fue propuesto por Wagner en 1965 (en un informe interno de la Subcomisión de Estratigrafía del Carbonífero, después traducido al español y publicado en Wagner, 1966) para cubrir el vacío existente entre el techo del Westfaliense D, representado en el estratotipo de Sarre-Lorena (Francia) por la disconformidad asociada al conglomerado de Holz, y la base del Estefaniense A, ligado tradicionalmente al “Assise de Rive-de-Gier” en la cuenca carbonífera de St. Étienne. La Cordillera Cantábrica reúne excelentes condiciones para esta propuesta, habida cuenta de la existencia en la región palentina de floras con elementos intermedios entre los propios del Westfaliense D (Asturiense) y del Estefaniense y la presencia en la región de restos fósiles de origen marino. Finalmente, el Cantabriense fue aceptado como piso basal del Estefaniense. Por lo que respecta al Barrueliense, cabe mencionar que el análisis de los afloramientos típicos del Estefaniense A (esencialmente el “Assise de Rive-de-Gier”, a las que el Estefaniense A estuvo ligado tradicionalmente, y los importantes afloramientos en las regiones de Carmeaux y Cévennes) reveló la existencia de algunos problemas de índole estratigráfica y algunas complicaciones tectónicas. Estos hechos, junto a la naturaleza restringida del registro fósil (consistente casi exclusivamente en restos de flora continental) hacen de las localidades 34 mencionadas un punto basante distante del ideal para el limitotipo del Estefaniense A. Por ello, Bouroz et al. en el curso del Septième Congrès International de Stratigraphie et de Géologie du Carbonifère (Bouroz et al., 1972) hicieron la propuesta de sustituir el Estefaniense A por el piso Barrueliense, situando su límite inferior en la base de la Formación Barruelo. El Estefaniense continental intramontañoso Diversos lugares de la ZC muestran materiales carboníferos con características muy distintas a las descritas. Se trata de sucesiones de carácter predominantemente continental, que a veces alcanzan potencias de varios miles de metros y que están constiuidas esencialmente por conglomerados, areniscas, lutitas y capas de carbón, con predominio de origen fluvial y lacustre. Su edad está comprendida entre el Barrueliense superior y el Estefaniense B o C. El registro fósil está constituido fundamentalmente por macroflora, bivalvos no marinos y ostrácodos continentales. Estos materiales aparecen en afloramientos asislados, frecuentemente en relación con fallas importantes y muchas de ellas (Tineo, Cangas de Narcea, La Magdalena, Ciñera-Matallana, Sabero…) representan (o han representado) importantes yacimientos de carbón, por lo han sido intensamente investigadas. Conclusión • El Carbonífero de la ZC ofrece la única sucesión de Europa occidental con abundancia de materiales y asociaciones fósiles marinos desde el Tournaisiense hasta el Gzheliense. Condicionado en gran parte por el desarrollo de la orogenia Varisca, el Carbonífero cantábrico muestra en el ámbito astur-leonés una clara diferencación tanto espacial (edades más jóvenes, sedimentación marina más persistente en el tiempo, mayor desarrollo de plataformas carbonatadas y plataformas carbonatadas más jóvenes en sentido oriental), como vertical (parte inferior de la sucesión con características relativamente uniformes y menor variedad de fósiles de grupos bentónicos, partes intermedia y alta con importante registro fósil de organismos bentónicos, fuerte compartimentación de la cuenca y materiales de con abundante flora fósil. • Como consecuencia de su deformación en condiciones superficiales y de la ausencia de metamorfismo significativo, el registro fósil del Carbonífero cantábrico es rico, variado y bien conservado. Las asociaciones bentónicas marinas que poblaron los mares pensilvánicos estaban integradas fundamentalmente por foraminíferos, poríferos, corales, braquiópodos y ostrácodos. • La variedad de ambientes en que se depositaron los materiales y las asociaciones fósiles que contienen constituyen un reto a investigaciones de carácter paleoecológico. • La continuidad del registro sedimentario, la alternancia de restos de origen contiental y marino, y la presencia de grupos de reconocido interés en correlación y datación ofrecen un marco propicio para estudios evolutivos y estratigráficos, así como para la investigación de la correlación entre escalas estratigráficas basadas en elementos continentales y marinos. En resumen: el registro fósil de Carbonífero cantábrico es un elemento crucial en el conocimiento de la estratigrafía, paleoecología, paleobiogeografía y biodiversidad de las asociaciones del Carbonífero a escala regional y global. Agradecimientos. El presente trabajo se ha beneficiado de la Subvención del Gobierno del Principado de Asturias SV-PA13-ECOEMP-46 al proyecto “Estudio paleontológico del Carbonífero cantábrico en intervalos estratigráficos considerados de interés prioritario por la Comisión Internacional de Estratigrafía”. Agradezco sinceramente a mis compañeros del área de Paleontología de la Universidad de Oviedo y a Carmen Álvarez Vázquez la ayuda prestada. 35 Bibliografía Barrois, Ch. 1882. Recherches sur les terrains anciens des Asturies et de la Galice. Mémoires de la Société Géologique du Nord, 2 (1), 1-630. Bouroz, A., Knight, J.A., Wagner, R.H., Winkler Prins, C.F. 1972. Sur la limite Westphalien-Stéphanien et sur les subdivisions du Stéphanien inférieur sensu lato. Compte Rendu 7e Congrès International de Stratigraphie et Géologie du Carbonifère, Krefeld 1971, I, 241-261. Braga, J.C. (coordinador), Fernández-Martínez, E., García-Bellido, D., Méndez-Bedia, I., Moreno-Eiris, E., Perejón, A., Rodríguez, S. & Soto, F. 2009. Poríferos y Cnidarios. En: Martínez Chacón, M.L. & Rivas, P. (eds.), Paleontología de invertebrados. Sociedad Española de Paleontología, Instituto Geológico y Minero de España, Universidad de Oviedo, Universidad de Granada, 33-110. Della Porta, G., Villa, E. & Kenter, J.A.M. 2005. Facies Distribution of Fusulinida in a BashkirianMoscovian (Pennsylvanian) carbonate platforms top (Cantabrian Mountains, NW Spain). Journal of Foraminiferal Research, 35 (4), 344-367. Fernández, L.P., Bahamonde, J.R., Barba, P., Colmenero, J.R., Heredia, N., Rodríguez-Fernández, L.R., Salvador, C., Sánchez de Posada, L.C., Villa, E., Merino Tomé, O. & Motis, K. 2004. Secuencia sinorogénica. En: Vera, J.A. (ed.), Geología de España. Sociedad Geológica de España, Instituto Geológico y Minero de España, 34-42. García-Bellido, D., Snowbari-Daryan, B. & Rigby, J.K. 2004. Cystothalamia vandegraffi new species and other Sphinctozoa from the Upper Carboniferous of Spain. Journal of Paleontology, 78 (6), 10371055. Julivert, M. 1971. Décollement tectonics in the Hercynian Cordillera of Northwest Spain. American Journal of Science, 270, 1-29. Koopmans, B.N. 1962. The sedimentary and structural history of the Valsurvio Dome, Cantabrian Mountains, Spain. Leidse Geologische Mededelingen, 26, 121-232. Lotze, F. 1945. Zur Gliederung der Varisziden der Iberischen Meseta. Geotektonische Forschungen, 6, 78-92 (Traducido al español en Publicaciones Extranjeras sobre Geología de España, 1950, 5, 149166). Martínez Chacón, M.L. 1991. Braquiópodos carboníferos de la costa E de Asturias (España). II: Spiriferida y Terebratulida. Revista Española de Paleontología, 6 (1), 59-88. Martínez Chacón, M.L. & Bahamonde, J.R. 2008. Brachiopods from a steep-slope facies of a Pennsylvanian carbonate platform (Sierra del Cuera, Asturias, North Spain). Fossils and Strata, 54, 133-143. Olóriz, F. & Rodríguez-Tovar, F.J. 2009. Clase Cephalopoda. En: Martínez Chacón, M.L. & Rivas, P. (eds.), Paleontología de invertebrados. Sociedad Española de Paleontología, Instituto Geológico y Minero de España, Universidad de Oviedo, Universidad de Granada, 300-361. Pérez Estaún, A., Bastida, F., Alonso, J.L., Marquínez, J., Aller, J., Álvarez-Marrón, J., Marcos, A. & Pulgar, J.A. 1988. A thin-skinned tectonics model for an arcuate fold and thrust Belt: The Cantabrian Zone (Variscan Ibero-Armorican Arc). Tectonics, 7, 517-537. Sánchez de Posada, L.C., Martínez Chacón, M.L., Méndez, C.A. & Villa, E. 2001. Rasgos paleontológicos del Carbonífero marino cantábrico. En: Gámez Vintaned, J.A. & Liñán, E. (eds.), La Era Paleozoica. El desarrollo de la vida marina. Homenaje al Prof. Jaime Truyols. Institución Fernando el Católico, 191-222. Sanz-López, J., Blanco-Ferrera, S., García-López, S. & Sánchez de Posada, L.C. 2006. The MidCarboniferous boundary in northern Spain: difficulties for correlation of the Global Stratotype Section and Point. Revista Italiana di Paleontologia e Stratigrafia, 112 (1), 3-22. Wagner, R.H. 1966. El significado de la flora en la Estratigrafía del Carbonífero. Boletín de la Real Sociedad Española de Historia Natural (Geología), 64, 203-208. 36 Wagner, R.H. 1970. An outline of the Carboniferous Stratigraphy of Northwest Spain. In: Streel, M. & Wagner, R.H. (eds.), Colloque sur la Stratigraphie du Carbonifère. Congrès et Colloques de l´Université de Liège, 55, 429-463. Wagner, R.H., Sánchez de Posada, L.C., Martínez Chacón, M.L., Fernández, L.P., Villa, E. & Winkler Prins, C.F. 2002. The Asturian Stage: a preliminary proposal for the definition of a substitute for Westphalian D. In: Hills, L.V., Henderson, C.M. & Bamber, E.W. (eds.), Carboniferous and Permian of the World. Canadian Society of Petroleum Geologists, Memoir 19, 832-850. C onferencia de clausura 40 El significado geológico de las cuencas carboníferas con flora fósil en Sierra Morena R.H. Wagner Centro Paleobotánico, IMGEMA-Real Jardín Botánico de Córdoba, Avenida de Linneo, s/n, 14004 (Córdoba). [email protected]. Introducción Sierra Morena incluye una parte de tres de las zonas tectono-estratigráficas de Lotze (1945): Surportuguesa, Ossa-Morena y Lusitano oriental-Alcúdica. Consiste en franjas de rumbo NWSE que se juntaron durante la segunda parte del Carbonífero, el Subperiodo Pensilvánico. Se trata de terrenos venidos de una situación próxima a las Islas Británicas mediante fallas intracontinentales de tipo strike-slip (Fig. 1). Estos movimientos laterales de gran envergadura son post-Misisípicos y pre-Autuniense, quedando bien acotados en el tiempo por la datación paleobotánica de varias cuencas en la parte occidental de la actual Península Ibérica. Figura 1. Principales fallas strike-slip continentales del Pensilvánico en la parte occidental de la Península Ibérica. 41 Las grandes fallas strike-slip La presencia de grandes fallas de tipo strike-slip en las zonas de Ossa-Morena y Lusitano occidental-Alcúdica de Lotze (1945), delimitando distintos terrenos, fue señalada, en primer lugar, por Burg et al. (1981) que determinaron este tipo de deformación tectónica para una franja de 20 km de anchura a la que llamaron “Coimbra-Córdoba shear zone” (autores posteriores modificaron esta nomenclatura a “zona de cizalla Badajoz-Córdoba) (Fig. 2). Mencionaron que fue una estructura “varisca”, dejando entrever que la orogénesis varisca sería anterior a la cuenca westfaliense de Peñarroya. Ya se sabía que la Zona Ossa-Morena se descomponía en una serie de dominios (Delgado et al., 1977) de composición tectono-estratigráfica diferente. Posteriormente, Martínez-García (1986, 2006) subdividió el Macizo Iberico, tal como lo conocemos actualmente, en una gran cantidad de terrenos, acudiendo a fallas strike-slip para la delimitación de algunos de ellos. Figura 2. Mapa de la zona de cizalla Badajoz-Córdoba mostrando la situación de las cuencas carboníferas implicadas. Observar la diferencia fundamental entre la cuenca westfaliense de Peñarroya, condicionada por una gran falla strike-slip intracontinental, y la cuenca tournaisiense de Valdeinfierno, Utilizando criterios estructurales, la envergadura de estas fallas se estableció en 72 km de traslado sinistral (Burg et al., 1981). No lograron datar estos movimientos, aunque parecía haber consenso entre los diferentes autores en que afectaban a los estratos anteriores al Pensilvánico. Efectivamente, el Carbonífero más moderno (Subperíodo Pensilvánico) estaba considerado como una cobertera, posterior a la deformación tectónica. Esta apreciación se modificó notablemente cuando se pudo estudiar a fondo la cuenca minera de Peñarroya-Belmez-Espiel en la provincia de Córdoba. La gran cantidad de datos mineros, y, sobre todo, los muchos miles de metros de sondeos testificados con detalle por el Servicio Geológico de la empresa minera ENCASUR en los años 80 y 90 del siglo pasado han permitido determinar que la cuenca de Peñarroya se estableció como consecuencia de una gran falla strike-slip intracontinental (Wagner, 1999). Esta falla bordeaba la zona de cizalla Badajoz-Córdoba por su lado NE y se reveló como un elemento tectónico de primer orden. La datación de las subcuencas sucesivas de la cuenca de Peñarroya y el análisis tectono-estratigráfico de las mismas permitieron entender su historia geológica, así 42 como el tiempo implicado. Posteriormente (Wagner, 2004) se vió que la cuenca de Peñarroya no era la única que se formó como consecuencia de una gran falla strike-slip. En efecto, resulta evidente que varias de las grandes fallas strike-slip en la parte occidental de la Península podían datarse por sus cuencas carboníferas adosadas y provocadas por los movimientos sinsedimentarios sobre la falla correspondiente. La datación de estas cuencas probó que las grandes fallas strike-slip continentales eran de edad pensilvánica, un intervalo de 20 a 25 Ma. Aunque en ningún caso se podía contar con la cantidad de datos obtenidos en Peñarroya, los indicios eran suficientes para establecer que las grandes fallas strike-slip eran de este Subperiodo en todos los casos, y que tenían una importancia capital para juntar los terrenos constituyentes de las zonas de Ossa-Morena y Lusitano Occidental-Alcúdica. La cuenca westfaliense de Peñarroya Es emblemática la cuenca hullera westfaliense de Peñarroya (Córdoba). Muestra todas las características de una cuenca resultante de los movimientos continuados sobre una gran falla strike-slip intracontinental comparable en su envergadura y comportamiento a la falla terciaria y actual de San Andrés en California. Es más, por tener una historia acabada, permite entender mejor lo que está pasando con la falla actual en California, aún en activo. La gran falla intracontinental strike-slip que provocó la cuenca hullera de Peñarroya (Wagner, 1999) movió un terreno metamórfico del Dominio de Sierra Albarrana (Delgado et al., 1977), con Precámbrico y Ordovícico, en dirección SE. Actuó como placa activa, y provocó el hundimiento de una placa pasiva adyacente con rocas del Dominio Obejo-Valsequillo (op. cit.), donde se acumularon los sedimentos westfalienses de Peñarroya (Fig. 3). Figura 3. Cuenca westfaliense de Peñarroya (con subcuencas sucesivas) adosada a la zona de cizalla BadajozCórdoba, que actuó como placa activa. Tomado de Wagner (1999). 43 Los términos empleados reflejan un comportamiento diferente. La placa pasiva quedaría en posición estática, aunque sufriendo una fracturación provocada por el movimiento lateral de la placa activa. Aunque esta tuvo, sobre todo, un traslado lateral, hubo también un efecto vertical importante, con subida de la placa activa. Es evidente que la gran falla tuvo pequeñas adláteres provocando duplicaciones (detectadas por la minería). La fracturación de la placa pasiva, provocada por los esfuerzos laterales, dió como resultado una falla sintética subparalela a la falla grande, que determinó el otro borde de la cuenca subsidente (westfaliense), con un escarpe donde se instaló una sucesión de abanicos aluviales. La placa pasiva experimentó también un movimiento ligeramente basculante hacia la gran falla, generando un área lacustre en épocas de pluviosidad importante (como lo eran los tiempos westfalienses de la franja paleoecuatorial). El resultado fue una cuenca con un borde activo elevado, con arrastre de los sedimentos consolidados, marcando etapas transtensivas y transpresivas, respectivamente, y abanicos aluviales mal preservados por efectos tectónicos en el contacto con la placa activa. El otro borde fracturado por la falla sintética también experimentó una elevación provocando un escarpe con sucesivos abanicos aluviales (Andreis & Wagner, 1983). El paleorelieve correspondiente se estimó en varios centenares de metros (op. cit.) (Fig 4). Figura 4. Distribución de facies en la cuenca strike-slip de Peñarroya. Tomado de Wagner (1999). Siendo así que las grandes fallas strike-slip intracontinentales nunca son totalmente rectas, se presentan irregularidades en su trazado que se conocen, por sus efectos tectónicos como releasing bend y restraining bend (Christie-Blick & Biddle, 1985). Puesto que la placa activa experimentó también un levantamiento paulatino a la vez que se trasladó lateralmente, se entiende que el movimiento vertical produciría un cabalgamiento sobre la placa pasiva en las etapas transpresivas de restraining bend. La placa pasiva cedió en su contacto con la falla intracontinental en etapas transtensivas (de releasing bend) para acomodar una cuenca de sedimentación continental, con varias subcuencas sucesivas que se trasladaron en el sentido del movimiento lateral de la placa activa (Fig. 5). Dentro de cada subcuenca había un traslado del 44 depocentro en el mismo sentido. Las subcuencas fueron deformadas tectónicamente en las sucesivas etapas transpresivas, por lo que la deformación tectónica de las distintas subcuencas es diferente (Fig. 6). Es una imagen complicada, aunque comprensible. Se entiende que tanto la distribución de facies como los efectos tectónicos fueron determinantes para el desarrollo de la historia de la minería en Peñarroya. Figura 5. Esquema del comportamiento del releasing bend en una falla strike-slip intracontinental con las fracturas inducidas en la placa pasiva subsidente y traslado del depocentro en la dirección del movimiento de la placa activa. Tomado de Wagner (1999). La datación de las subcuencas sucesivas procede del estudio de unos 20.000 ejemplares de megaflora obtenida, en su mayor parte, de sondeos en la cuenca minera de PeñarroyaBelmez-Espiel (Álvarez-Vázquez, 1995). La placa activa, que consta de un Precámbrico metamórfico y Ordovícico, muestra también sedimentos misisípicos (Viseense y Serpujoviense) depositados en discordancia total. Es una configuración que recuerda a la situación existente en la Europa caledoniana de las Islas Británicas. En efecto, el origen de la placa activa debe buscarse a una distancia de muchos centenares de kilómetros hacia el NNW, en dirección del Atlántico actual (Fig. 1) y cercano a las Islas Británicas. La placa pasiva consta de rocas menos metamórficas (principalmente filitas) del Paleozoico Inferior a las que siguen sedimentos devónicos. El contraste es total. El inicio de la gran falla strike-slip que provocó la generación de la cuenca westfaliense de Peñarroya no se ha podido datar con precisión porque esta cuenca está cubierta por un Terciario discordante en su extremo noroccidental, donde se encuentra su parte más antigua. Los sedimentos hulleros que se explotaron en la parte más NW conocida son de edad Langsettiense superior (Fig. 6, parte superior). En dirección SE, a lo largo de los aproximadamente 50 km de afloramientos correspondientes a la cuenca minera, se aprecian edades westfalienses sucesivas hasta llegar al Bolsoviense inferior (?) en su parte más oriental. Ahí se pierde el registro por un cabalgamiento de las rocas de la placa activa. Esto no quiere decir que la cuenca no continuara más hacia el SE. Con estas dataciones ya se demuestra que las etapas de transtensión, provocando la generación de subcuencas en etapas sucesivas y correspondientes al paso de un releasing bend en la gran falla strike-slip, duró unos 4 Ma para un traslado de unos 50 km. Significa un movimiento lateral contínuo de algo más de 1 m por siglo. La cuenca minera de Peñarroya es como una ventana a la historia de esta gran falla. Siendo así que las etapas de restraining bend producían un cabalgamiento de la placa activa sobre la cuenca hullera instalada en la placa pasiva adyacente, es lícito pensar que esta cuenca tuviera más longitud de la que aparenta por los afloramientos. Hay indicios para suponer que la gran falla strike-slip en el borde de la zona de cizalla de Badajoz-Córdoba haya actuado durante buena parte del Pensilvánico. En este caso llegaríamos a unos 16 Ma y un traslado lateral de la placa activa de unos 150 km. 45 I. Langsettiense superior II. Duckmantiense superior Figura 6. Cortes a través de dos subcuencas sucesivas de Peñarroya. Tomado de Wagner (2004). Otras fallas strike-slip Aunque esta falla debe ser de las mayores en la parte occidental de la Península Ibérica, no es la única. Algunos autores (p.ej. Ábalos & Eguiluz, 1991) se dieron cuenta de que la zona de cizalla Badajoz-Córdoba consta de una serie de terrenos orientados NW-SE, separados por fallas strike-slip. Aunque estos autores supusieron una edad misisípica (“Carbonífero Inferior”) para los movimientos en dirección, podrían ser posteriores. Los retazos de cuencas misisípicas marinas con intervalos continentales se encuentran en varias franjas diferentes separadas por falla, siendo probable que los contactos mecánicos por strike-slip sean posteriores a la formación de la(s) cuenca(s) misisípica(s), cuyo(s) borde(s) son desconocidos. La única evidencia directa de movimientos strike-slip de edad misisípica se encuentra en la pequeña cuenca pull-apart de Valdeinfierno (Roldán & Rodríguez, 1986), de edad Tournaisiense superior-Viseense inferior (Fig. 2). Está claro que un pull-apart no es lo mismo que una cuenca controlada por una falla strike-slip intracontinental. Sin embargo, el caso de Valdeinfierno indica que ya hubo movimientos strike-slip en el Misisípico, aunque a más pequeña escala que los correspondientes a las grandes fallas strike-slip intracontinentales de edad pensilvánica. 46 Otro caso de fallas strike-slip intracontinentales de edad pensilvánica lo encontramos en la zona fallada de Valongo, al NE de Oporto. Esta zona arrumbada también NW-SE (Fig. 7) muestra una serie de cuencas carboníferas sucesivas con edades de Bolsoviense al Estefaniense C en una longitud de unos 80 a 90 km. La zona fallada de Valongo debe entenderse como el límite de la Zona Lusitano oriental-Alcúdica con la Zona GalaicoCastellana de Lotze (1945). Es un límite de primer orden. Este dato confirma que la gran falla de Badajoz-Córdoba no es la única falla de strike-slip intracontinental que actuó durante el Pensilvánico en la parte occidental de la Península Ibérica. Hay que añadir que la configuración de terrenos en Ossa-Morena sugiere que hay más fallas de este tipo, aunque no sean siempre datables (por no tener cuencas carboníferas adosadas). El factor acumulativo hace que una parte de estos terrenos podría haber sufrido un traslado total del orden de mil kilómetros, con lo cual nos situaríamos en un área al WSW de las Islas Británicas (Fig. 1). Las relaciones exactas están aún por investigar, ya que exige recomponer el puzzle de terrenos precámbricos y del Paleozoico Inferior. Figura 7. Situación de las cuencas carboníferas en la zona de Valongo: 1. Serra de Rates (Bolsoviense); 2. Casais-Alvarelhos (Bolsoviense); 3. Ervedosa (Asturiense); 4. Douro (Estefaniense C). Además, 5. Buçaco (Autuniense), adosada a una falla strike-slip posterior (y menos importante). El cese de los movimientos strike-slip en dirección SE es post-Estefaniense C, tal como se ha podido establecer en Valongo, pero anterior a las cuencas autunienses (Estefaniense final) de Guadalcanal (Sevilla)-Fuente del Arco (Badajoz) y Valdeviar (Sevilla), en la parte más suroccidental de Ossa-Morena (y sin relación directa con la Zona Surportuguesa). Sobre todo la cuenca de Valdeviar tiene un historial que demuestra su desarrollo con independencia de las estructuras tectónicas plegadas y falladas de Ossa-Morena (Wagner & Mayoral, 2007). Pertenece a una etapa posterior, caracterizada por un volcanismo importante fini-carbonífero, que se extendió en el tiempo a lo largo del Pérmico. Los pitones de los volcanes de esta época se encuentran de pie, sin deformación tectónica, en varios lugares de la Zona de Ossa-Morena (p.ej. el que sostiene el castillo medieval de Almodóvar del Río). El ensamblaje de terrenos mediante varias grandes fallas strike-slip intracontinentales tuvo lugar, pues, durante casi todo el Pensilvánico, con la excepción de su parte terminal, el Autuniense. Es a partir del Autuniense o de la parte final del Estefaniense C que se terminó la constitución del Macizo Ibérico. Significa que el Macizo Ibérico no tuvo entidad geológica antes de finales del Carbonífero. Zona fallada de Valongo Esta zona fallada, al nordeste de Oporto, que está orientada NW-SE y que tiene una longitud aproximada de 80-90 km, contiene varias cuencas carboníferas que representan, en su conjunto, buena parte del Pensilvánico (Fig. 7) (Es preferible “Zona fallada de Valongo” a la nomenclatura portuguesa de “surco carbonífero dúrico-beirão”). En esta franja fallada, de menos de 1 km de anchura, se encuentran sucesivamente, del NW al SE, estratos continentales del Bolsoviense 47 (probable), Asturiense y Estefaniense C. Los carbones de este último fueron explotados en la cuenca minera del Douro. Muestran el efecto de strike-slip, con una duplicación estructural de la sucesión estratigráfica en una parte de la zona minera que delata claramente el movimiento lateral (Fig. 8). Movimientos laterales siniestros fueron observados en estratos pre-carboníferos, así como un cabalgamiento desde el NE (Domingos et al., 1983), reflejando una situación tectónica complicada. A pesar de faltar datos detallados, la alineación de cuencas carboníferas de edades sucesivas muestra el traslado clásico del área de sedimentación en el sentido de movimiento de la placa activa, hacia el SE (Wagner, 2004). Figura 8. Cuenca Estefaniense C de Douro con duplicación por falla strike-slip. Tomado de un informe inédito de Wagner, Ribeiro & Lemos de Sousa. Dataciones de las cuencas hulleras provocadas por strike-slip Estas dataciones se basan en los estudios paleobotánicos realizados en las últimas décadas. No se podrían haber hecho sin recolecciones importantes, sumando decenas de miles de muestras recogidas con un control geológico riguroso favorecido por el acceso a las minas de carbón y una gran cantidad de sondeos testificados con detalle. Se ha reconstruído la composición de las floras sucesivas, teniendo en cuenta el factor paleoecológico y con el máximo rigor taxonómico, garante de las comparaciones dentro de Europa y más allá. Son los elementos florísticos de las facies aluvial/pantanosas correspondien- 48 tes a un clima lluvioso en la franja paleoecuatorial de la época pensilvánica. El enfoque es geológico/paleoambiental más bien que botánico, que haría más hincapié en el sentido evolutivo de los elementos paleobotánicos. En el transcurso de estos estudios se han podido encajar los diferentes registros de flora de las cuencas carboníferas en Sierra Morena en una sucesión de biozonas erigidas en base de una información más amplia (Wagner & Álvarez-Vázquez, 2010) (Fig. 9). Figura 9. Cuadro estratigráfico del Sistema Carbonífero mostrando los pisos y subpisos y las biozonas florísticas correspondientes (Parte del cuadro publicado por Wagner & Álvarez-Vázquez, 2010). Valdeinfierno (Córdoba-Sevilla) Los datos paleoflorísticos más antiguos en Sierra Morena proceden, sobre todo, de la pequeña cuenca pull-apart de Valdeinfierno (Córdoba-Sevilla) (Roldán & Rodríguez, 1986). Cabe precisar que una cuenca pull-apart resulta de dos fallas strike-slip, paralelas, y actuando en 49 sentidos contrarios, produciendo así el hundimiento de un área generalmente pequeña, de forma rómbica. Constituyen auténticos agujeros de hundimiento muy rápido. En el caso de Valdeinfierno, la cuenca medía solamente 3 por 4 km, pero acumuló unos 1.000 m de sedimentos en los que destacan gruesas brechas de coluvión y abanico aluvial. La flora de Valdeinfierno es la clásica de la parte alta del Tournaisiense y del Viseense inferior en Europa (Wagner, 2001). Consta de elementos florísticos del ambiente pantanoso en el fondo de un valle flanqueado por el paleorelieve generado por las dos fallas strike-slip, además de las plantas que vivían sobre el paleorelieve existente. La mezcla de estos elementos paleoflorísticos producía una composición algo diferente de la que se conoce de las floras sucesivas del Pensilvánico, donde predominaban los elementos higrófilos característicos de una llanura aluvial. Una flora de la misma edad Tournaisiense superior-Viseense inferior se recogió de una franja en las inmediaciones de Puebla de los Infantes (Sevilla). Incluye el yacimiento de Cerrón del Hornillo (Laveine in Robardet et al., 1986) y otro yacimiento cercano (Wagner & ÁlvarezVázquez, 2010). Varios retazos de estratos generalmente marinos con intervalos terrestres de edades viseenses se han registrado en los dominios (terrenos) de Valencia de las Torres-Cerro Muriano, Sierra Albarrana y Córdoba-Alanís de Ossa-Morena. Incluyen los afloramientos de Matachel-Benajarafe, donde hay un importante elemento volcánico (Pascual & Pérez-Lorente, 1975; Wagner et al., 1983), así como la alineación subparalela formada por los Santos de Maimona, Berlanga y Casas de Reina. Aunque los distintos retazos fueron atribuídos a una sola cuenca misisípica, despedazada por fallas strike-slip (Wagner et al., 1983; Wagner, 2001), esto parece ahora una simplificación inadmisible. Es posible que se trate de restos de varias cuencas diferentes de edades parecidas. Se nota que las edades registradas abarcan tanto el Viseense como el Serpujoviense, ocupando un amplio espacio en el tiempo (hasta unos 23 Ma). Las floras corresponden a una llanura costera, y se parecen a las que se conocen de otras áreas europeas. Revisten un interés especial las floras serpujovienses recogidas en distintas localidades a lo largo del borde suroeste de la cuenca minera westfaliense de Peñarroya-Belmez-Espiel. No se ha realizado todavía el estudio pormenorizado del material florístico de los intervalos continentales en los estratos generalmente marinos del Viseense y Serpujoviense, pero existe una documentación parcial (Wagner, 2001). Las sucesivas floras de la cuenca de Peñarroya Son las mejor estudiadas en cuanto a su composición y su contexto geológico (ÁlvarezVázquez, 1995). La flora más antigua corresponde a la parte noroccidental de la cuenca. Es la llamada “zona de Antracitas”, donde un foco térmico posterior (¿de edad pérmica?) aumentó el rango de los carbones por una pérdida de volátiles. Coincide con una subcuenca (la más antigua) que ocupa la parte noroccidental de la cuenca minera y que se extiende hacia el sureste al área de carbones coquizables de mayor productividad industrial en Peñarroya-Pueblonuevo. La distribución de facies muestra una zona lacustre que se extendió al lado de la gran falla, lo cual se explica por el basculamiento del fondo de la cuenca hacia la gran falla strike-slip que provocó el hundimiento de la placa pasiva donde se instaló la cuenca de sedimentación. Las floras fósiles se han recogido, sobre todo, en los estratos sucesivos a la facies lacustre y que corresponden a las facies pantanosas que se instalaron posteriormente. La composición de la flora refleja estas facies pantanosas, y es parecida a la de las cuencas parálicas clásicas del Norte de Europa y de Norteamérica (Apalaches). Por tanto, cabe pensar que la cuenca de Peñarroya se encontraba no solamente en una paleolatitud parecida sino prácticamente a nivel del mar (aunque no hay prueba directa de facies costeras). Sorprende la poca representación de elementos florísticos “extracuenca” (Wagner & ÁlvarezVázquez, 2010, Table 1), correspondientes a la vegetación de ladera que debería encontrarse en el caso de una cuenca de sólo 1-2 km de anchura. De hecho, solamente dos especies de 50 Rhacopteris, correspondientes a dos hallazgos excepcionales, podrían ser “extracuenca”. La inmensa mayoría de las especies (72) son de facies “hullera”, y corresponden a una llanura aluvial con clima lluvioso. Los pocos elementos “extracuenca” se recuperaron de sedimentos lacustres con restos flotados “alóctonos”. Sin embargo, hay que reconocer una tendencia a la desintegración de los restos vegetales por motivos tafonómicos que haría menos frecuente el registro florístico de ladera. La segunda subcuenca de Peñarroya, situada hacia el sureste, contiene una flora del Duckmantiense superior (Álvarez-Vázquez, 1995). Esta segunda subcuenca corresponde a una etapa posterior a la deformación tectónica de la primera (de “Antracitas”) y su flora fósil es netamente más moderna que la anterior. Confirma que la etapa de deformación tectónica que corresponde al paso de un restraining bend de la gran falla strike-slip ha llevado un tiempo palpable (equivalente al Duckmantiense inferior). La composición de esta flora se parece también a la de las cuencas parálicas del Norte de Europa, con muy pocas diferencias que corresponden a las pteridospermas (“helechos con semillas”) con una diseminación más complicada que la de los helechos. Por otra parte, se nota un aumento en el número de especies “extracuenca” (7). Se podría especular sobre la posibilidad de un cierto aislamiento, pero el resto de la flora no lo avala. La deformación tectónica de la segunda subcuenca es claramente diferente de la que sufrió la primera (Fig. 6), lo cual concuerda con la distinta edad deducida por las floras fósiles. Parece que se haya generado una tercera subcuenca (establecida después del paso del restraining bend) en la parte más suroriental de la cuenca, pero los datos no son totalmente concluyentes. Por otra parte, cabe señalar un traslado de depocentro hacia el SE, con la correspondiente diferencia de edad que parece haber alcanzado al Bolsoviense inferior. No se puede determinar con precisión, ya que las floras de esta edad se distinguen mal de las del Duckmantiense superior. Los datos geológicos tampoco muestran precisión, aunque una campaña de sondeos sugirió una deformación tectónica distinta a la de la segunda subcuenca. Cuencas posteriores a las grandes fallas intracontinentales Dentro del ámbito de Sierra Morena se observa la ausencia de sedimentos estefanienses con la excepción de su parte final, el Autuniense. Sin embargo, el Estefaniense C está representado en la cuenca del Douro, zona fallada de Valongo, cerca de Oporto y, posiblemente, en la cuenca de Puertollano (Ciudad Real) (que ahora se piensa que podría ser posterior, del Autuniense). Hay que señalar que Ábalos et al. (2002, p. 175) mencionaron la presencia de cuencas intramontañosas estefanienses en Ossa-Morena, pero esta afirmación no está avalada por ningún dato real. De hecho, en el mismo libro (“The Geology of Spain”), Colmenero et al. (2002, fig. 7.13) indican que los tiempos estefanienses en la parte sur del Macizo Ibérico corresponden a “erosión y/o ausencia de sedimentación”. Las cuencas autunienses (post-Estefaniense C) de Guadalcanal y Valdeviar (Sevilla) son posteriores al ensamblaje final de los terrenos aportados por las grandes fallas strike-slip intracontinentales. El Macizo Ibérico se constituyó por primera vez a finales del Carbonífero (pre-Autuniense). Sin embargo, los movimientos strike-slip no cesaron totalmente, ya que hay fallas de este tipo, aunque de mucha menor envergadura, de orientación N-S aproximada, cuya edad se deduce por la cuenca autuniense de Buçaco, cerca de Coimbra, adosada a la falla. Estas fallas N-S tuvieron un desplazamiento muy limitado. Cuenca de Valdeviar (Sevilla) 51 Los 320 m de sedimentos autunienses que se conocen de la cuenca de Valdeviar (Fig. 10) comienzan por el relleno de un paleovalle escarpado en el extremo NW de la cuenca. Después hubo una expansión paulatina de la cuenca en dirección sureste, con un solapado progresivo sobre un basamento de rocas ígneas y del Paleozoico Inferior plegado y fallado (ver Fig. 11). Interpretada antiguamente como un semigraben adosado a una falla identificada como el límite entre las zonas de OssaMorena y Surportuguesa (Simancas, 1983, 1985; Wagner, 2004), un estudio más moderno de la cuenca de Valdeviar (Wagner & Mayoral, 2007) mostró que la falla al NE de esta cuenca es posterior y sin relación aparente con la sedimentación carbonífera. De hecho, parece que el límite con la zona Surportuguesa se sitúe a una cierta distancia al WSW y que la cuenca de Valdeviar se haya asentado sobre la antigua zona de Ossa-Morena (Mencionar que la distinción entre las zonas del esquema de Lotze, 1945, parece carecer de sentido a partir del Autuniense). Figura 10. Mapa de la cuenca autuniense de Valdeviar (Sevilla) mostrando el solapamiento de las distintas unidades estratigráficas en dirección SE. Al relleno de valle en el extremo NW de la cuenca de Valdeviar siguió una etapa de sedimentación fluvial (local), seguido por un episodio de coladas de basalto (Simancas & Rodriguez Gordillo, 1980). Los basaltos se acuñan hacia el SE adaptándose a un paleorelieve residual y muestran también un espesor menor en dirección NE. A los basaltos siguen lutitas rojas de facies lacustre que sufrieron los efectos de una rubefacción debida a unas condiciones climáticas secas, con lluvias ocasionales. La rubefacción logró homogeneizar los sedimentos, borrando en gran parte su laminación. Sobre este ambiente sedimentario se vertieron cenizas volcánicas con un espesor del orden de 40 m, correspondientes a un volcanismo local. El microclima asociado a un volcanismo explosivo generó facies lacustres con una flora higrófila y mesófila (Wagner & Mayoral, 2007). También se registraron lavas, que han quedado sin 52 estudiar. Chimeneas correspondientes a este volcanismo se han detectado a unos 5-7 km hacia el ENE de Valdeviar (Wagner & Mayoral, 2007). Se trata de un volcanismo explosivo de composición riodacítica (Sierra et al., 2009), bastante común en tiempos finicarboníferos y pérmicos de España y de Europa Occidental en general. Los efectos de este volcanismo se prolongaron al encontrarse troncos silicificados en los sedimentos siguientes que consisten en unos 150 m de capas rojas con caliches y coladas de barro provocadas por lluvias fuertes de forma ocasional en un régimen semi-desértico. Aunque las muestras de flora de esta etapa son más esporádicas, se nota la presencia de elementos más xerófilos. Figura 11. Esquema mostrando el solapamiento progresivo en dirección SE de las distintas unidades estratigráficas de Valdeviar. Tomado de Wagner & Mayoral (2007). La composición de la flora fósil refleja las distintas etapas. La importante colección de fósiles vegetales obtenida de las tobas volcánicas incluye helechos, pteridospermas y esfenofitas, además de la licópsida Sigillaria brardii, que refleja un régimen húmedo relacionado, tal vez, con las frecuentes erupciones volcánicas. Los elementos “extracuenca”, como Plagiozamites rochei, sólo reflejan una diferencia de paleoambiente. La composición de esta flora sugiere un régimen pantanoso-lacustre. Esta misma impresión da la flora (con escasos ejemplares) encontrada en las lutitas lacustres que sufrieron la rubefacción en las frecuentes etapas de evaporación del agua de los lagos convertidos en charcos. Las lluvias estivales movilizaron los suelos de las colinas en el entorno de la cuenca de sedimentación, formando coladas de barro e incorporando troncos y ramas de coníferas de la vegetación de ladera. La flora recogida por Gavala (1927) en las capas rojas superiores consta, sobre todo, de elementos correspondientes a lugares con suelos mejor drenados. Ya no son las facies pantanosas. Estos elementos se identificaron con la asociación mesófilo-xerófila de Gothan & Gimm (1930) del Rotliegend inferior de Europa Central, aunque llama la atención la ausencia de peltaspermáceas en Valdeviar, que son frecuentes en el Rotliegend. Sin embargo, hay que notar que el registro es aún bastante incompleto. Consideraciones florísticas generales 53 Las condiciones climáticas juegan siempre un papel importante en la distribución de la flora. Influyen directamente en las facies. Hay que reconocer que las floras carboníferas se han registrado casi exclusivamente de las facies pantanosas asociadas a llanuras aluviales en ambientes propicios a la generación de capas de carbón. Esto significa un registro muy parcial y sesgado, algo que los libros de texto paleobotánicos no siempre recogen adecuadamente. Hay indicios esporádicos de que algunos grupos de plantas, tales como las cicadópsidas, ya existían en el Carbonífero, aunque sin dejar apenas huella en el registro geológico. Las coníferas son también de gran antigüedad, pero corresponden a facies (hábitat) poco comunes antes de finales del Pensilvánico. Por tanto, conviene examinar el registro de la flora carbonífera siempre en relación con el clima y el ambiente sedimentario que, a su vez, debe ser considerado en su contexto geotectónico, con cuencas sedimentarias de distinto tipo. Este enfoque geológico es fundamental para poder juzgar el registro paleobotánico. Aparte de estos parámetros generales, conviene reconocer también que el estado fragmentario de los fósiles vegetales impone la necesidad de reunir un gran número de ejemplares, una necesidad que se hace imperativa cuando se trata de determinar un nivel cronoestratigráfico. De hecho, se necesitan miles de ejemplares. Las grandes colecciones, con muchos miles de ejemplares, que se han reunido de varios niveles del Carbonífero de Sierra Morena, permiten trabajar con más precisión de lo que suele ser posible en otros casos. Estas colecciones se conservan, en su mayor parte, en el Real Jardín Botánico de Córdoba. Conclusiones La geología de Sierra Morena, conocida por sus dominios variados, bastante diferentes en rocas precámbricas y del Paleozoico Inferior sobre todo, y con registros más esporádicos del Devónico y del Misisípico, se entiende solamente en el contexto de terrenos tectónicos aportados por varias grandes fallas strike-slip intracontinentales. El comportamiento de estas fallas de gran envergadura demuestra una procedencia del área atlántica, cercana a las Islas Británicas. La datación por las floras fósiles ha sido crucial para fijar un intervalo en el tiempo de unos 20-25 Ma (Pensilvánico) para la actividad de las grandes fallas strike-slip. La composición de estas floras es parecida a la que se conoce de toda la franja paleoecuatorial. Sorprende la poca cantidad de elementos “extracuenca”, ya que la poca anchura de las cuencas de sedimentación y la evidencia de un paleorelieve (aunque moderado) darían las circunstancias para encontrar ambientes distintos a la facies “hullera”. La actividad de las grandes fallas strike-slip cesó hacia finales del Carbonífero. El Macizo Ibérico quedó constituído en el Autuniense (el subpiso más alto del Estefaniense). Bibliografía Ábalos, B. & Eguiluz, L. 1991. Deformación transpresiva carbonífera en la Zona de Cizalla BadajozCórdoba (Macizo Ibérico Meridional). Revista Sociedad Geológica de España, 4, 229-249. Ábalos, B., Carreras, J., Druguet, E., Escuder Viruete, J., Gómez Pugnaire, M.T., Lorenzo Álvarez, S., Quesada, C., Rodríguez Fernández, L.R. & Gil-Ibarguchi, J.I. 2002. Variscan and Pre-Variscan Tectonics. In: Gibbons, W. & Moreno, T. (eds), The Geology of Spain. Geological Society, London, 155-183. Álvarez-Vázquez, C. 1995. Macroflora del Westfaliense inferior de la cuenca de Peñarroya-Belmez-Espiel (Córdoba). I (Texto): 1-393, text-figs 1-21; II (Láminas): Láms 1-100; III (Anexos): Anexo 1: 1-7; Anexo 2: 1-111; Anexo 3: Cuadros 1-9; Anexo 4: Planos 1-4. Tesis Doctoral, Departamento de Geología, Universidad de Oviedo (No publicada). Andreis, R.R. & Wagner, R.H. 1983. Estudio de abanicos aluviales en el borde norte de la cuenca Westfaliense B de Peñarroya-Belmez (Córdoba). En: Lemos de Sousa, M.J. (ed.), Contributions to the Carboniferous Geology and Palaeontology of the Iberian Peninsula. Universidade do Porto, Faculdade de Ciências, Mineralogia e Geologia, 171-227. 54 Burg, J.P., Iglesias, M., Laurent, Ph., Matte, Ph. & Ribeiro, A. 1981. Variscan intracontinental deformation: The Coimbra-Córdoba shear zone (SW Iberian Peninsula). In: Lister, G.S., Behr, H.-J., Weber, K. & Zwart, H.J. (eds.), The Effect of Deformation on Rocks. Tectonophysics, 78, 161-177. Christie Blick, N. & Biddle, K.T. 1985. Deformation and basin formation along strike-slip faults. In: Biddle, K.T. & Christie-Blick, N. (eds.), Strike-slip deformation, basin deformation, and sedimentation. Society of Economic Paleontologists and Mineralogists, Special Publication 37, 1-34. Colmenero, J.R., Fernández, L.P., Moreno, C., Bahamonde, J.R., Barba, P., Heredia, N. & González, F. 2002. Carboniferous. In: Gibbons, W. & Moreno, T. (eds.), The Geology of Spain. Geological Society, London, 93-116. Delgado-Quesada, M., Liñán, E, Pascual, E. & Pérez-Lorente, F. 1977. Criterios para la diferenciación de dominios en Sierra Morena Central. Stvdia Geologica, XII, 75-90. Domingos, L.C.G., Freire, J.L.S., Gomes da Silva, F., Gonçalves, F., Pereira, E. & Ribeiro, A. 1983. The structure of the Intramontane Upper Carboniferous Basins in Portugal. Memórias dos Serviços Geológicos de Portugal, 29, 187-194. Gavala, J. 1927. Mapa Geológico de España a escala 1:50.000. Memoria correspondiente a la hoja de Cantillana. Boletín Instituto Geológico y Minero de España, XLIX, 1-79, mapa geológico y cortes. Gothan, W. & Gimm, O. 1930. Neuere Beobachtungen und Betrachtungen über die Flora des Rotliegenden von Thüringen. Arbeiten aus dem Institut für Paläontologie und Petrographie der Brennsteine, 2 (1), 3974. Lotze, F. 1945. Zur Gliederung der Varisziden der Iberischen Meseta. Geotektonische Forschungen, 6, 1-12 [Traducción, 1950: Observaciones respecto a la división de los varíscides de la Meseta Ibérica. Publicaciones extranjeras sobre Geología de España, 5, 149-166]. Martínez-García, E. 1986. The Iberian Massif. In: Martínez-García, E. & Gil Ibarguchi, J.I. (eds.), International Conference on Iberian Terranes and their regional correlation, Oviedo 1986, 1-42. Martínez-García, E. 2006. Proterozoic-Lower Paleozoic terrane accretion and Variscan domains in the Iberian Massif (Spain and Portugal). Zeitschrift der deutschen Gesellschaft für Geowissenschaften, 154 (4), 559-574. Pascual, E. & Pérez Lorente, F. 1975. El magmatismo ácido superficial al sur de Villanueva del ReyVillaviciosa de Córdoba (Sierra Morena, Córdoba). Cuadernos de Geología, Universidad de Granada, 6, 15-30. Robardet, M., Weyant, M., Laveine, J.-P. & Racheboeuf, P. 1986. Le Carbonifère inférieur du Synclinal du Cerrón del Hornillo (Province de Seville, Espagne). Révue de Paléobiologie, 5 (1), 71-90. Roldán, F.J. & Rodríguez, J. 1986. La cuenca carbonífera de Vadeinfierno (dominio de Sierra Albarrana, zona de Ossa Morena). Un ejemplo de sedimentación relacionada con accidentes de desgarre. Acta Geológica Hispánica, 21-22 (1986-87), 321-327. Sierra, S., Moreno, C. & Pascual, E. 2009. Stratigraphy, petrography and dispersion of the lower Permian syn-eruptive deposits in the Viar Basin, Spain. Sedimentary Geology, 217, 1-29. Simancas, J.F. 1983. Geología de la extremidad oriental de la Zona Sudportuguesa. Texto: 1-439. Tesis Doctoral, Universidad de Granada (No publicada). Simancas, J.F. 1985. Estudio estratigráfico de la cuenca del Viar. 5ª Reunión del Grupo de Ossa-Morena, Belmez (Córdoba), 7-17. Temas Geológico Mineros, Instituto Geológico y Minero de España. Simancas, J.F. & Rodríguez Gordillo, J.F. 1980. Magmatismo basáltico hercínico tardío en el NW de Sevilla. Cuadernos de Geología, Universidad de Granada, 11, 49-60. Wagner, R.H. 1999. Peñarroya, a strike-slip controlled basin of early Westphalian age in Southwest Spain. Vĕstník Českého geologického ústavu, 74, 2, 87-108. Wagner, R.H. 2001. Fósiles vegetales. Serie Recursos Naturales, 6. Departamento de Medio Ambiente y 55 Protección Civil, Diputación de Córdoba, 1-105. Wagner, R.H. 2004. The Iberian Massif: a Carboniferous assembly. Journal of Iberian Geology, 30, 93-118. Wagner, R.H. & Álvarez-Vázquez, C. 2010. The Carboniferous floras of the Iberian Peninsula: A synthesis with geological connotations. Review of Palaeobotany and Palynology, 162 (3), 238-324. Wagner, R.H. & Mayoral, E. 2007. The Early Permian of Valdeviar in Sevilla province, SW Spain: basin history and climatic/palaeogeographic implications. Journal of Iberian Geology, 33 (1), 93-124. Wagner, R.H., Coquel, R. & J. Broutin 1983. Mississippian floras of the Sierra Morena. S.W. Spain: a progress report. In: Lemos de Sousa, M.J. (ed.), Contributions to the Carboniferous Geology and Palaeontology of the Iberian Peninsula. Universidade do Porto, Faculdade de Ciências, Mineralogia e Geologia, 101-126. S imposio PICG 596 58 Evolución temprana de las especies de Idiognathodontidae (conodontos) en torno al límite Misisípico/Pensilvánico S. Blanco Ferrera1 y J. Sanz López1 1 Departamento de Geología, Facultad de Geología, Universidad de Oviedo, c/ Arias de Velasco s/n, 33005 Oviedo. [email protected]; [email protected]. La crisis de biodiversidad del Serpukhoviense o del final del Misisípico (ver referencias en Walliser, 1996) es considerada un acontecimiento de extinción de segundo orden en la historia de la vida. Sin embargo, algunos autores (McGhee et al., 2012) lo sitúan en quinto lugar en cuanto a su impacto ecológico. El acontecimiento del Serpukhoviense supuso el final de la “Edad de los Crinoideos”, o la reorganización de las faunas de corales rugosos en las comunidades bentónicas marinas, mientras que tuvo lugar un reemplazamiento de las superfamilias y familias en las comunidades nectónicas y planctónicas, tanto de ammonoideos como de conodontos. En general, la reorganización de los ecosistemas marinos estuvo asociada a una pérdida de diversidad y la presencia de géneros de baja velocidad evolutiva que persistirán durante la edad de hielo del Paleozoico tardío. Entre los conodontos del Serpukhoviense, los representantes de la Familia Gnathodontidae y Mestognathidae fueron sustituidos por los géneros de la Familia Idiognathodontidae en aguas profundas, mientras que tuvo lugar un relevo a nivel genérico dentro de la Familia Cavugnathidae en aguas someras. El intervalo de tiempo estimado para el reemplazo de faunas fue corto en muchas cuencas, pero amplio en otras con registro de aguas profundas, como ocurre en las montañas Tián (Tian Shan, Asia Central) y en el norte de la península Ibérica. En el estratotipo de la Formación Barcaliente, la aparición de los primeros Idiognathodontidae del género Declinognathodus se produce unos 145 metros por debajo de la extinción local de los géneros atribuidos al Misisípico. Esta primera aparición está próxima a un cambio del medio sedimentario generalmente descrito en la zona Cantábrica, como una profundización del surco de antepaís y el desarrollo de una rampa carbonatada con un fondo pobremente oxigenado. Además, y antes de la desaparición de las especies misisípicas, ya se observa una diversidad de los primeros declinognátidos: Declinognathodus bernesgae, D. praenoduliferous, D. cf. noduliferus y D. cf. inaequalis. La aparición de D. inaequalis indica la base del Pensilvánico y es seguida por los hallazgos más bajos de D. lateralis, Idiognathoides y Neognathodus. A partir de los cambios en las morfologías de los elementos P1 de los primeros representantes de los géneros de la Familia Idiognathodontidae y su posición en diferentes secciones estratigráficas se propone una hipótesis filogenética (Fig. 1). Los primeros Declinognathodus evolucionaron probablemente desde Gnathodus postbilineatus, a través de la reducción de la plataforma rostral y la pérdida de su ornamentación, pero sobre todo del desarrollo de crestas transversales entre los dientes de la carena y del parapeto caudal. De esta manera, el surco adcarenal es corto o está ausente en los elementos de tamaño mayor de estos Declinognathodus. Sin embargo, el desarrollo de crestas transversales en la plataforma es un carácter que tenderá a ir desapareciendo durante la evolución posterior del grupo, dando lugar a taxones con surcos de distinta longitud y profundidad. Además, un carácter de primer orden en la evolución a nivel específico es la ornamentación y las características del parapeto rostral. Así, algunas especies de Neognathodus presentan largos parapetos, mientras que estos están ausentes en Idiognathoides; salvo en algunos elementos raros con nódulo rostral dentro de asociaciones dominadas por elementos sin nódulo. 59 Los primeros hallazgos de Declinognathodus en el Serpukhoviense son excepcionales, en coincidencia con las características de un taxón "progenitor durante crisis". Estos hallazgos preceden a la diversificación y distribución amplia de la Familia Idiognathodontidae durante el Bashkiriense más temprano, en un intervalo de tiempo de menos de un millón de años, en el que las especies misisípicas de conodontos fueron reduciendo su distribución hasta desaparecer. Figura 1. Hipótesis evolutiva de Declinognathodus, Idiognathoides y Neognathodus entre el final del Serpukhoviense y el Bashkiriense temprano a partir de dibujos de holotipos y ejemplares significativos. Agradecimientos. Por su soporte económico, a los proyectos CGL-2009-09109 (Ministerio de Economía y Competitividad, Fondos FEDER) y SV-PA-13-ECOEMP-46 (Principado de Asturias). Este trabajo representa una contribución al IGCP-596. Bibliografía McGhee, G.R.Jr., Sheehanm P.M., Bottjer, D.J. & Droser, M.L. 2012. Ecological ranking of Phanerozoic biodiveristy crises: The Serpukhovian (early Carboniferous) crisis had a greater ecologic impact than the end Ordovician. Geology, 40, 147-150. Walliser, O.H. 1996. Global events in the Devonian and Carboniferous. In: Walliser, O.H. (ed.), Global events and event stratigraphy in the Phanerozoic. Springer-Verlag, Berlin, 225-250. 60 Primera aproximación al estudio de los montículos de fango del Devónico Inferior (Lochkoviense-Praguiense) de la Formación Lebanza (Zona Cantábrica) E. Fernández-Martínez1, L.P. Fernández2, C. Vera de la Puente2 y A. Ernst3 1 Facultad de Ciencias Biológicas y Ambientales, Campus de Vegazana, 24071 León. [email protected]. Facultad de Geología, c/ Jesús Arias de Velasco s/n, 33005 Oviedo. [email protected]; [email protected]. 3 Institut für Geowissenschaften der Christian-Albrechts-Universität zu Kiel, Ludewig-Meyn-Str. 10, D-24118 Kiel, Germany. [email protected]. 2 Introducción La sucesión devónica de la Zona Cantábrica registra siete episodios arrecifales caracterizados por el desarrollo de biohermos y biostromos construidos por metazoos coloniales con esqueleto carbonatado, preferentemente corales tabulados, corales rugosos y esponjas del grado estromatoporoideos (Fernández-Martínez et al., 2010 y referencias). Junto a ellos se han reconocido, durante el Devónico Inferior, dos episodios singulares por el desarrollo de montículos de fango. Esta denominación alude básicamente a depósitos dominados por barros carbonatados, habitualmente generados o inducidos por comunidades microbianas y que, en el caso concreto de los ejemplos cantábricos, contienen también diversos metazoos esqueletizados. Este trabajo es una primera aproximación al estudio del más antiguo de estos episodios, que tiene lugar en la Formación Lebanza (Lochkoviense medio-Praguiense basal). Es destacable que estos montículos constituyen el primer registro de bioconstrucciones del Devónico cantábrico y, por tanto, de todo el Paleozoico de la Zona Cantábrica. Contexto geológico y estratigráfico La Formación Lebanza (Krans et al., 1980) es una unidad carbonatada que aflora en varias bandas limitadas por fallas y localizadas en la Unidad del Pisuerga Carrión (norte de Palencia). En su área de referencia, el valle alto del río Arauz, la formación consta de 150-160 m de calizas con margas y lutitas subordinadas. Ha sido dividida en cinco miembros de límites poco definidos y representativos de diferentes facies desarrolladas en una plataforma carbonatada somera. Las capas estudiadas se sitúan en la parte baja del miembro C. El análisis de las microfacies presentes en la parte inferior de este miembro ha permitido interpretar estos materiales como depositados en áreas posiblemente situadas en la parte trasera y protegida de las barras (de tipo shoals y lóbulos de washover) que han quedado registradas en el miembro B. Los montículos de fango En la sección del Arroyo de Pineda, donde han sido muestreados con mayor precisión, afloran 3 montículos, uno al lado de otro dentro del mismo intervalo estratigráfico. La longitud de base máxima es de 3,75 m, por una altura máxima de 2,5 m. Sus taludes son simétricos y tienen una inclinación próxima a los 30-35º. Los montículos están formados por caliza gris oscuro de aspecto masivo y homogéneo, afectada por una intensa bioturbación y que no presenta indicios de diferentes etapas de desarrollo. Microscópicamente son calizas floatstone de briozoos y corales tabulados con una matriz wackestone bioclástica. Se han reconocido hasta tres generaciones de fango: M1) micrita oscura posiblemente autóctona; M2) microesparita clara que rellena diversos tipos de poros; y 61 M3) micrita oscura grumosa que aparece como costras rodeando a granos y esqueletos y se interpreta como de origen orgánico. También se han observado varios tipos de porosidad y cuatro generaciones de cementos. El contenido fósil Tanto a simple vista como en lámina delgada, la fauna de metazoos es muy abundante. El grupo mejor representado es el de los briozoos (Ernst et al., 2012), seguido por los braquiópodos, y corales tabulados. Junto a ellos aparecen fragmentos de crinoideos, espinas de equínidos y, más ocasionalmente, gasterópodos y ostrácodos. Briozoos, corales y braquiópodos aparecen frecuentemente completos y, en especial el último grupo, en posición de vida. Es muy habitual la presencia de organismos con hábitos de vida reptantes e incrustantes (Fig. 1). Estas incrustaciones y la abundancia de bioperforaciones son dos de los rasgos más llamativos de los bioclastos presentes en estos montículos. Por lo que respecta a los corales tabulados, una altísima proporción de los mismos corresponde a representantes de la superfamilia Auloporicae y concretamente a los géneros Aulopora Goldfuss 1829, Mastopora Sokolov 1952 y Remesia Kettner 1934. Los dos primeros géneros son formas de hábito incrustante, que inician su crecimiento sobre un bioclasto o sobre un tapiz microbiano, y continúan elevándose ligeramente sobre su base. Por otra parte, Remesia se encuentra siempre como una coralita única, separada de su colonia, la cual se describe habitualmente como reptante. En ambos casos se trata de colonias con escaso número de coralitas y un desarrollo muy limitado. Sin embargo, el coral tabulado más frecuente es Hillaepora Mironova 1960 de la familia Pachyporidae, cuyas colonias suelen encontrarse total o parcialmente rodeadas por briozoos incrustantes (Fig. 1). Agradecimientos. A Paco Soto e Isabel Méndez Bedia, por su ayuda y buen humor en el campo. Este trabajo es una contribución a los proyectos PICG 596 y al proyecto CGL2011-24775 del Ministerio de Economía y Competitividad. Bibliografía Figura 1. Pequeña colonia de Hillaepora completamente rodeada por un briozoo incrustante. La disposición del briozoo indica que esta incrustación se produjo en vida del coral. Ernst, A., Fernández, L.P., Fernández-Martínez, E. & Vera, C. 2012. Bryozoan fauna from mud mounds of the Lebanza Formation (Lower Devonian) in the Arauz area (Pisuerga-Carrión Province, Cantabrian Zone, NW Spain). Geodiversitas, 34 (4), 693-738. Fernández-Martínez, E., Fernández, L. P., Méndez-Bedia, I., Soto, F. & Mistiaen, B. 2010. Earliest Pragian (Early Devonian) corals and stromatoporoids from reefal settings in the Cantabrian Zone (N Spain). Geologica Acta, 8, 301-323. Krans, T. J., Guit, T. A., & van Ofwegen, L. P. 1980. Facies-patterns in the Lower Devonian carbonates of the Lebanza Formation (Cantabrian Mountains, Province of Palencia, NW Spain). Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 163, 192-230. 62 Análisis de los patrones de escamación en telodontos e inferencias sobre su modo de vida H. Ferrón1 y H. Botella1 1 Departamento de Geología, Universidad de Valencia, Avda. Dr. Moliner, 50, 46100 Burjassot (Valencia). [email protected]; [email protected]. Los telodontos son un grupo fósil de vertebrados agnatos cuyo rango estratigráfico abarca desde el Ordovícico Superior hasta el Devónico Superior. Aunque sus relaciones y posición filogenética respecto a los otros grupos de agnatos paleozoicos está aún poco clara, la monofilia del grupo es ampliamente aceptada. La superficie corporal de los telodontos está cubierta por escamas aisladas cuya morfología, histología y proceso de desarrollo presentan una gran similitud con las encontradas en neoseláceos (Karätajûte-Talimaa, 1998). Aunque los restos articulados conocidos son de gran importancia para estudiar algunos aspectos de su morfología externa y anatomía interna, la mayoría de especies se conocen únicamente a partir de escamas aisladas. La abundancia y ubicuidad de estos restos desarticulados han hecho de los telodontos un grupo importante para el establecimiento de correlaciones en el Paleozoico medio, siendo muy estudiados desde un punto de vista bioestratigráfico y paleogeográfico. No obstante, sólo unos pocos trabajos los han tratado desde una perspectiva paleobiológica teniéndose hasta la fecha escasos conocimientos sobre su ecología (ver Turner, 1992, 1999). En este sentido, siguiendo la metodología descrita en Ferrón et al. (2013, en este mismo volumen) se han analizado los patrones de escamación de las 139 especies de telodonto descritas hasta la fecha con el fin de conocer algunos aspectos de su modo de vida. Para ello, se determinó la disposición corporal y superficie cubierta por cada tipo funcional de escamas descritas en tiburones actuales. Mediante los datos obtenidos ha sido posible la comparación tanto cualitativa como cuantitativa con los grupos ecológicos o modos de vida definidos para tiburones en Reif (1985). Los patrones de escamación observados en los ejemplares de telodonto que preservaban escamaciones completas o parciales fueron muy similares a los que encontramos hoy en día en tiburones actuales. Cinco escamaciones concretas fueron descritas para telodontos: (1) Escamación con una cobertura casi total de escamas con función de defensa ante ectoparásitos y epibiontes, que se asoció a telodontos de velocidad baja/moderada que muy probablemente formaban cardúmenes; (2) Escamación con una cobertura casi total de escamas con función de protección ante la abrasión con el sustrato, que se asoció a telodontos demersales de fondos duros; (3) Escamación con una cobertura casi total de escamas con funciones generalizadas, que se asoció a telodontos lentos de aguas abiertas; y finalmente dos patrones de escamación (4 y 5) que no pudieron ser relacionadas con ninguno de los observados en tiburones actuales. El conjunto de los resultados, incluyendo también los obtenidos mediante el estudio de las especies conocidas únicamente por restos desarticulados, mostró una diversidad notable en cuanto a los modos de vida presentes en telodontos. Especies estrictamente pelágicas están representadas en cuatro órdenes distintos (Shieliiformes, Phlebolepidiformes, Thelodontiformes y Furcacaudiformes), lo que indica que la adaptación al medio pelágico apareció independientemente al menos cuatro veces en telodontos. Un porcentaje similar de especies han sido relacionadas con un modo de vida bentopelágico asociado a plataforma externa y talud continental, estando bien representadas en el orden de los Loganelliiformes y habiéndose encontrado algunas de ellas en facies de origen turbidítico. Un mayor número de especies serían 63 nadadoras de velocidad lenta/moderada que muy probablemente formarían cardúmenes, estando bien representadas a partir del Silúrico en Shieliiformes y Furcacaudiformes. Finalmente, el modo de vida mayoritario parece haber sido el de los demersales de fondos duros, lo que contrasta con las interpretaciones realizadas hasta el momento que asociaban a los telodontos a fondos blandos, arenosos o fangosos. Una posible interpretación podría encontrarse en el hecho de que los telodontos son los únicos agnatos paleozoicos con un esqueleto micromérico, mientras que la gran mayoría del resto de agnatos paleozoicos (con excepción de los minoritarios ciclóstomos y los anáspidos) poseían un gran escudo cefálico rígido aplanado dorsoventralmente, con grandes prolongaciones también rígidas, no articuladas. Los hábitats de sustratos duros con relieve abrupto suelen ofrecer a estas especies protección ante depredadores, pero requiere que las mismas posean cuerpos relativamente flexibles que les permita buscar refugio entre cuevas, grietas o incluso en ambientes arrecifales. Esto parece improbable para el resto de ostracodermos con sus rígidas y pesadas corazas, pero no para los telodontos de cuerpo flexible cubierto con pequeñas escamas. Agradecimientos. Este trabajo ha sido financiado por el proyecto CGL2011-24775 del MINECO y es una contribución al PIGC-596-IUGS. Bibliografía Ferrón, H., Martínez-Pérez, C. & Botella, H. 2013. Estudio de los patrones de escamación y el modo de vida de tiburones actuales para inferencias paleoecológicas en taxones fósiles. XXIX Jornadas Paleontología, Córdoba. Libro de Resúmenes. Karätajûte-Talimaa, V. 1998. Determination methods for the exoskeletal remains of early vertebrates. Mitteilungen aus dem Museum für Naturkunde in Berlin, Geowissenschaftliche Reihe, 1, 21-52. Reif, W.E. 1985. Squamation and ecology of sharks. Courier Forschungsinstitut Senckenberg, 78, 255 pp. Turner, S. 1992. Thelodont lifestyles. In: Mark-Kurik, E. (ed.), Fossil Fishes as Living Animals, 1, 21-40. Turner, S. 1999. Early Silurian to Early Devonian thelodont assemblages and their possible ecological significance. In: Boucot, A.J. & Lawson, J. (eds.), Palaeocommunities: A Case Study From the Silurian and Lower Devonian, 42-78. 64 Estudio de los patrones de escamación y el modo de vida de tiburones actuales para inferencias paleoecológicas en taxones fósiles H. Ferrón1, C. Martínez-Pérez 1, 2 y H. Botella1 1 Departamento de Geología, Universidad de Valencia, Avda. Dr. Moliner, 50, 46100 Burjassot (Valencia). [email protected]; [email protected]. 2 School of Earth Sciences, University of Bristol, Wills Memorial Building, Queen’s Road, Bristol BS8 1RJ, United Kingdom. [email protected]. Las escamas desarticuladas son uno de los microictiolitos más abundantes del registro fósil, encontrándose la evidencia más temprana de éstas, y a la vez de vertebrados con exoesqueleto, en el Ordovícico Medio (Samson et al., 2012). Un gran número de grupos extintos de vertebrados presentan escamaciones microméricas (e.g. Astráspidos, algunos heterostráceos, telodontos, elegestolépidos, mongolépidos, acantodios y la mayoría de grupos de elasmobranquios), habiendo sido algunos de ellos muy estudiados taxonómicamente dada su importancia en bioestratigrafía pero raramente analizados desde un punto de vista paleobiológico, conociéndose muy poco acerca de su ecología y modo de vida. Sin embargo, el estudio de las escamaciones de tiburones actuales, constituidas también por elementos microméricos, podría ser útil para aproximarse a la ecología de estos grupos fósiles. En este sentido, existen algunos trabajos clásicos (Raschi & Tabit, 1992; Reif, 1982, 1985) donde se ha relacionado la morfología y la función de escamas de tiburones actuales y se ha puesto de manifiesto la estrecha relación existente entre sus patrones de escamación y su modo de vida. Reif (1982, 1985) estableció cuatro tipos funcionales de escamas placoideas de tiburones (escamas con una función de protección ante la abrasión con el sustrato, de defensa ante ectoparásitos y epibiontes, de reducción de la fricción con el agua y de acomodación de fotóforos en tiburones bioluminiscentes) describiendo una o varias morfologías características para cada uno de ellos. Adicionalmente, estableció siete grupos ecológicos de tiburones y describió para cada uno el tipo funcional de escamas predominante. Sin embargo, dado que varios tipos funcionales de escamas pueden coexistir en un mismo individuo es conveniente describir los patrones de escamación tanto cualitativamente, teniendo en cuenta la distribución corporal de los tipos funcionales, como cuantitativamente, en función de los porcentajes de cobertura de cada tipo funcional. Esto se hace especialmente necesario si el objetivo es comparar dichos patrones de escamación con los encontrados en grupos extintos que generalmente sólo pueden estudiarse a partir de escamaciones parciales. En el presente trabajo, y a partir del estudio de la relación entre los patrones de escamación y la ecología de tiburones actuales, se ha establecido una metodología útil para inferir algunos aspectos sobre el modo de vida de vertebrados acuáticos con exoesqueletos similares a partir del análisis de sus escamaciones. Un total de 50 ejemplares de tiburones actuales, pertenecientes a 43 especies distintas, fueron estudiados en las colecciones del Museu del Cau del Tauró de l’Arboç (Tarragona, España) y el Museu de Zoologia de Barcelona (Barcelona, España). En cada uno de ellos se describieron y fotografiaron las distintas variantes morfológicas de escamas encontradas, determinándose su distribución y calculando posteriormente el área de cobertura corporal ocupada por cada una de ellas. En paralelo, las morfologías de cada tipo funcional propuesto por Reif (1982, 1985) fueron caracterizadas por morfometría clásica y un posterior análisis discriminante, el cual permitió la asignación de las variantes morfológicas observadas en los 50 ejemplares estudiados a alguno de los tipos funcionales. Los patrones de escamación quedaron definidos tanto por la distribución 65 corporal de los tipos funcionales como por los porcentajes de superficie corporal ocupados por cada uno de ellos. Posteriormente los grupos ecológicos de tiburones actuales propuestos por Reif (1985) fueron reexaminados y cada una de las especies estudiadas fue incluida en alguno de ellos. La relación entre el patrón de escamación y el modo de vida (grupo ecológico) fue analizada por dos vías. En primer lugar, mediante la descripción del patrón típico de cada grupo ecológico teniendo en cuenta tanto la presencia como la disposición corporal de cada tipo funcional y, en segundo lugar, mediante un análisis discriminante a partir de los porcentajes de cobertura de los tipos funcionales. En ambos casos se demostró una buena correspondencia entre el modo de vida y el patrón de escamación de tiburones actuales. Queda establecido, de esta manera, un marco de comparación donde incluir potencialmente un número importante de grupos extintos de vertebrados poco conocidos desde una perspectiva ecológica. Así, la identificación de los tipos funcionales de escamas, la descripción de su disposición y el cálculo del área corporal ocupada por cada uno de ellos en estos grupos puede aportar información muy relevante sobre su modo de vida. El trabajo se vuelve especialmente relevante para el caso de grupos fósiles sin representantes actuales donde las interpretaciones paleoecológicas son más difíciles de establecer. Agradecimientos. Este trabajo ha sido financiado por el proyecto CGL2011-24775 del MINECO y es una contribución al PIGC-596-IUGS. Bibliografía Raschi, W. & Tabit, C. 1992. Functional aspects of placoid scales: a review and update. Australian Journal of Marine & Freshwater Research, 43, 123-147. Reif, W.E. 1982. Morphogenesis and function of the squamation in sharks. Comparative functional morphology of shark scales, and ecology of sharks. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 164, 172-183. Reif, W.E. 1985. Squamation and ecology of sharks. Courier Forschungsinstitut Senckenberg, 78, 255 pp. Sansom, I.J., Davies, N.S., Coates, M.I., Nicoll, R.S. & Ritchie, A. 2012. Chondrichthyan-­‐like scales from the Middle Ordovician of Australia. Palaeontology, 55, 243-247. 66 Conodontos, biofacies y microfacies del Givetiense de Renanué (Devónico Medio, Pirineos Aragoneses) J-C. Liao1,2 y J.I. Valenzuela-Ríos2 1,2 Departamento de Paleontología, Universidad Complutense de Madrid, c/ José Antonio Novais, 2, 28040 Madrid. [email protected]. 2 Departmento de Geología, Universidad de Valencia, c/ Dr. Moliner 50, 46100 Burjasot. [email protected]. Los conodontos del área de Renanué fueron estudiados por primera vez por Boersma (1973) en dos secciones 08-A y 08-B, cuya edad conjunta comprendía desde el Eifeliense al Frasniense, pero ninguna de sus muestras proporcionó conodontos del Givetiense, por lo que su edad se dedujo por la posición estratigráfica de los estratos comprendidos entre las muestras datadas. Nuestra sección Renanué corresponde a la sección 08-A y los resultados preliminares ya demostraron una clara edad Givetiense para la mayor parte de la sección (Liao et al., 2001; Liao et al., 2008). La sección Renanué comienza con unos 4 m de pizarras negras que contienen dos niveles de nódulos arenosos. Sobre estos materiales se dispone, en contacto tectónico, una sucesión carbonática de unos 50 m, que comprende una variada distribución de biofacies de conodontos y de microfacies con un control temporal muy preciso. El límite Eifeliense/Givetiense se sitúa en la capa -8 con la entrada del indicador bioestratigráfico Polygnathus hemiansatus. Este nivel representa, a su vez, el límite inferior del Givetiense Inferior y de la biozona de hemiansatus. El límite inferior de la siguiente biozona, timorensis, se identifica en la capa 3 mediante la entrada del taxón nominal. El Givetiense Inferior comienza con la biofacies de Polygnathus como dominante. Sin embargo, en la suprayacente biozona timorensis, dominan los Icriodus. Las microfacies dominantes son wackestone bioclásticas con escasas packstone. La matriz consiste de micrita o pelmicrita con restos esqueléticos muy pequeños. En la biozona timorensis destaca un packstone con predominio de dacrioconáridos. El límite inferior del Givetiense Medio coincide con el de la biozona rhenanus/varcus que en Renanué se reconoce en la capa 5 con la entrada de los taxones índice (P. rhenanus y P. varcus). La siguiente biozona, ansatus, comienza en la capa 9 con Tortodus caelatus, que mundialmente comienza en esta biozona. El índice bioestratigráfico, P. ansatus entra retrasado en la capa 10. En general, en el Givetiense Medio, dominan las biofacies de Polygnathus, pero los porcentajes y composición varían. Así, p. ej., en la capa 16 los Belodéllidos alcanzan el 20% del total y los Icriodus descienden por debajo del 3%. La proporción entre elementos no polygnátidos, como acompañantes de los polygnátidos varía. Así en la capa 24 los icriódidos dominan con un 11,8%, mientras que en la capa 42 Belodella alcanza el 10,3% y en la siguiente capa (43) Icriodus domina coma taxón accesorio, alcanzando el 5%. La capa 21 muestra un pico de Icriodus, taxón que alcanza el 56%. El análisis de las microfacies en la parte baja del Givetiense Medio son similares a las del Givetiense Inferior, pero a partir de la capa 22 las microfacies cambian a calcisiltitas, mudstone y wackestone con escasa fauna que en general son de menor tamaño. El aporte siliciclástico aumenta, especialmente en las capas 36 y 37 hasta la 41. En la 44 es importante destacar una rudstone oncoidal. Por encima siguen calizas puras donde dominan microfacies de tipo grainstone con wackestone subordinadas. El límite inferior del Givetiense Superior no se puede reconocer por el momento en Renanué porque falta el índice P. latifossatus. Sin embargo, la asociación de conodontos sugiere que este límite se podría situar entre las capas 49-53. Por encima, se reconocen la mayor parte de las 67 zonas del Givetiense Superior. Así la Zona hermanni Superior comenzaría 260 cm por encima de la base del tramo 55 con la entrada de P. c. cristatus. El límite inferior de la Zona disparilis Inferior se situaría 40 cm por encima de la anterior. La Zona disparilis Superior comenzaría en la base de la capa 56 (unos 60 cm por encima de la base de la anterior zona) con la entrada del índice K. disparilis. La última biozona del Givetiense, norrisi, se reconoce en la parte superior de la capa 56, con la entrada del taxón índice Skeletognathus norrisi. Al comienzo del Givetiense Superior dominan las biofacies de Polygnathus, pero la capa 51 representa la biofacies de Icriodus. En los niveles últimos del Givetiense, siguen dominando los Polygnathus pero cambia la composición de los taxones accesorios, desaparecen los Icriodus y entran Skeletognathus, Klapperina y Mesotaxis. Las microfacies de la parte inferior (capas 49-52) contienen wackestones bioclásticos y mudstones, parcialmente bioturbados. Hacia arriba continuan las wackestones que en el Givetiense tardío contienen fauna más pelágica y menos diversa que antes. Agradecimientos. Este trabajo ha sido financiado por el MINECO proyecto CGL2011-24775, y el M100131201 “Hi-Res correlation Mid-Paleozoic”. Representa una contribución al IGCP-596 y ACI 20091037. La Fundación Alexander von Humboldt apoyó trabajos iniciales de J.I.V-R y el DAAD de J-C.L. Bibliografía Boersma, K.TH. 1973. Devonian and Lower Carboniferous conodont biostratigraphy, Spanish Central Pyrenees. Leidse Geologische Mededelingen, 49, 303-377. Liao, J.-C., Königshof, P., Valenzuela-Ríos, J. I. & Schindler, E. 2008. Depositional environment interpretation and development of the Renanué section (Upper Eifelian- Lower Frasnian; Aragonian Pyrenees, N. Spain). Bulletin of Geosciences, 83 (4), 481-490. Liao, J.-C., Valenzuela-Rios, J.I. & Rodríguez, S. 2001. Descripción de los conodontos del Givetiense y Frasniense inferior (Devónico) de Renanué (Pirineos Aragoneses). Coloquios de Paleontología, 52, 1345. 68 Kozlowskia (Productidina, Brachiopoda) del Bashkiriense superior/ Moscoviense inferior (Carbonífero), Cordillera Cantábrica (N de España) M.L. Martínez Chacón1 y C.F. Winkler Prins2 1 Departamento de Geología, Universidad de Oviedo, c/ Jesús Arias de Velasco, s/n, 33005 Oviedo, España. [email protected]. 2 NCB Naturalis, PO Box 9517, 2300 RA Leiden, Holanda. [email protected]. Los prodúctidos son, junto con los espiriféridos, los braquiópodos más abundantes en las rocas carboníferas. Kozlowskia, género cosmopolita, se encuentra frecuentemente desde el Pensilvánico hasta el Guadalupiense (Pérmico medio). Shi et al. (2008) han contabilizado 48 especies incluidas en el género, en todo el mundo. La revisión de los Productidae del Bashkiriense/Moscoviense emprendida por los autores nos permitió reconocer en estos tramos siete especies distintas: K. bediae Martínez Chacón, 1979, Bashkiriense tardío; K. barroisi Martínez Chacón, 1982, Moscoviense inferior (Vereyense); Kozlowskia cf. postsetosa (Rotai, 1952), Bashkiriense tardío; y Kozlowskia sp. nov., Kozlowskia sp. 1, Kozlowskia sp. 2 y Kozlowskia sp. 3, del Bashkiriense tardío y del límite Bashkiriense/Moscoviense. K. bediae y K. barroisi ya han sido ampliamente descritas y discutidas. K. cf. postsetosa (Fig. 1.7-9): Kozlowskia pequeña, de contorno rectangular redondeado, disco ventral moderadamente convexo, surco medio débil, que no alcanza el margen anterior, en donde es sustituido por un aplanamiento medio; ornamentación semirreticulada, con costillas y arrugas bien desarrolladas sobe el disco; parte anterior de la cola, sin costillas, frecuentemente con unos pocos pliegues pequeños y bajos. Esta especie fue erróneamente atribuida por nosotros a Eomarginifera setosa, pero la presencia de una fuerte cresta marginal en el interior dorsal, correspondiente a una serie de colas sucesivas, y la fila de espinas cerca de la charnela obligan a excluir estos ejemplares de Eomarginifera y a incluirlos en Kozlowskia, siendo K. postsetosa la especie más próxima. Kozlowskia sp. nov. (Fig. 1.1-4): pequeña especie de Kozlowskia, de contorno subcircular, cavidad del corpus alta, cola relativamente larga, aurículas bien diferenciadas y, en ocasiones, muy extendidas, sin surco medio; ornamentación semirreticulada, con arrugas bien desarrolladas y costillas débiles, ausentes en la cola, la cual a veces lleva unos pocos pliegues bajos y anchos. Interior dorsal con cresta marginal ancha y lamelosa, correspondiente con muchas colas dorsales. Los ejemplares considerados previamente por nosotros como Eomarginifera praecursor son incluidos ahora en el género Kozlowskia. Las características de la especie la separan de las conocidas, lo que nos ha llevado a considerarla como nueva, pendiente de descripción. Kozlowskia sp. 1 (Fig. 1.5-6) tiene tamaño medio, contorno semicircular, valva ventral muy inflada, aurículas pequeñas; carece de surco medio, las costillas se borran en la parte anterior de la cola, que normalmente lleva algún pliegue pequeño e irregular. Kozlowskia sp. 2 (Fig. 1.11-12) tiene tamaño medio, es transversa, con aurículas bastante grandes, con surco medio que alcanza el frente de la valva; costillas bien desarrolladas en toda la valva, cruzadas sobre el disco por arrugas débiles. Kozlowskia sp. 3 (Fig. 1.10) tiene concha pequeña, transversa, con aurículas grandes, con un débil surco medio que alcanza el margen anterior; costillas finas y débiles, cruzadas en el disco por arrugas débiles. 69 Figura 1. 1-4. Kozlowskia sp. nov., DGO 4223. 5-6. Kozlowskia sp. 1. DGO 3647 y 3801, respectivamente. 7-9. Kozlowskia cf. postsetosa (Rotai, 1952). 7- DGO 4238, sección longitudinal que muestra la cresta marginal dorsal y una endospina. 8 y 9, DGO 3818 y 4239, respectivamente. 10. Kozlowskia sp. 3. DGO 3828. 11-12. Kozlowskia sp. 2. DGO 3823 y 3824, respectivamente. Agradecimientos. A Joaquín Vázquez Varela (Universidad de Oviedo) por tomar las fotografías. El trabajo se ha beneficiado de la subvención del Principado de Asturias SV-PA-13-ECOEMP-46, y se incluye en el PICG 596. Bibliografía Shi, G.H., Chen, Z.Q. & Tong, J.-N. 2008. New latest Carboniferous brachiopods from the Hexi Corridor Terrane, North China: faunal migrations and palaeogeographical implications. Proceedings of the Royal Society of Victoria, 120, 277-304. 70 Aplicación del Synchrotron X-Ray tomographic microscopy al estudio funcional de los elementos conodontales C. Martínez-Pérez1 y P.C.J. Donoghue1 1 School of Earth Sciences, University of Bristol, Wills Memorial Building, Queen's Road, Bristol BS8 1RJ, United Kingdom. [email protected]; [email protected]. Los conodontos son un grupo de animales marinos extintos que conforman posiblemente el primer miembro de nuestro linaje evolutivo que desarrolla partes esqueléticas mineralizadas. Como tal, el estudio de los elementos conodontales es de gran importancia, debido a la información que éstos pueden aportar sobre la biología y la función del esqueleto más primitivos de los vertebrados. Desde su descubrimiento por Pander (1856), además de sus afinidades biológicas, su posible función ha sido centro de discusión, estando las primeras hipótesis limitadas a simples comparaciones morfológicas (ver por ejemplo Jeppsson, 1979). Recientemente, un enfoque más cualitativo en base al análisis de su oclusión, microdesgaste y microestructura, ha facilitado la interpretación de los elementos conodontales como análogos funcionales de los dientes de vertebrados (Purnell, 1995; Donoghue & Purnell, 1999; Donoghue, 2001). Sin embargo, hasta la fecha, debido en parte a su pequeño tamaño y a la falta de herramientas adecuadas, no se ha realizado ninguna aproximación cuantitativa al análisis de la función de los conodontos. El objetivo del presente trabajo es profundizar en el estudio de la función de los elementos conodontales mediante la aplicación de nuevos métodos de análisis cuantitativo basados en el uso del Synchrotron X-Ray Tomographic microscopy para el desarrollo de modelos tridimensionales de resolución submicrométrica. Estos modelos son la base para un posterior análisis computacional, permitiéndonos, entre otras cosas, la reconstrucción tridimensional del aparato conodontal a partir del estudio de agrupaciones naturales para el estudio de su función con la ayuda de software especializado. De esta forma, el análisis digital de la oclusión mediante el software Occlusal Fingerprint Analyser, la impresión de los modelos digitales en 3D a escala para obtener modelos físicos tangibles (rapid-prototypes), o el análisis biomecánico de los mismo mediante el desarrollo de modelos de Elementos Finitos, ha permitido una nueva aproximación a su estudio, obteniendo resultados concluyentes a favor del uso de los conodontos como elementos que procesaban activamente el alimento. Sin embargo, nuestros resultados revelan también que los modelos oclusales existentes son demasiado simplistas y tal vez sobregeneralizados, ocultando una mayor diversidad de estilos de oclusión y grados de precisión. Estos datos han sido además apoyados por otros análisis mas tradicionales sobre microdesgaste y microestructura del elemento conodontal, confirmando de nuevo que muchos elementos P1 habrían desarrollado una función similar a los dientes del resto de gnatostomados. Por último, los datos tomográficos de alta resolución obtenidos nos han permitido una mejor caracterización de la estructura interna de los elementos, lo que nos está ayudando a profundizar en la propia biología del organismo, confirmando por ejemplo la hipótesis de que los elementos conodontales no eran reemplazados durante la vida del animal. Agradecimientos. CMP es beneficiario de un contrato postdoctoral Marie Curie FP7-People IEF 2011299681 del séptimo Programa Marco de la Unión Europea. 71 Bibliografía Donoghue, P.C.J. 2001 Microstructural variation in conodont enamel is a functional adaptation. Proceedings of the Royal Society of London, Series B, 268, 1691-1698. Donoghue, P.C.J. & Purnell, M.A. 1999. Mammal-like occlusion in conodonts. Paleobiology, 25 (1), 58-74. Jeppsson, L. 1979. Conodont element function. Lethaia, 12,153-171. Purnell, M.A. 1995. Microwear on conodont elements and macrophagy in the first vertebrates. Nature, 374, 798-800. Pander, C.H. 1856. Monographie der fossilen Fische des silurischen Systems der Russisch- Baltischen Gouvernements. Akademie der Wissenschaften, St. Petersburg, 91 pp. 72 Patrones de integración morfológica en el género Polygnathus (Conodonta) C. Martínez-Pérez1 y S. de Esteban Trivigno2 1 School of Earth Sciences, University of Bristol, Wills Memorial Building, Queen’s Road, Bristol BS8 1RJ, United Kingdom. [email protected]. 2 Transmitting Science, Gardenia 2, Can Claramunt, 08784 Piera, Barcelona, Catalonia, Spain. [email protected]. Los conodontos poseen un excelente registro fósil abarcando más de 300 millones de años, desde el Cámbrico Superior hasta el Triásico Superior. Durante este intervalo temporal el grupo se caracterizó por presentar unas altas tasas de cambio morfológico, convirtiéndose por tanto en excelentes herramientas para dataciones y correlaciones geológicas. Esta importante variabilidad morfológica representa una fuente ideal de información para abordar cuestiones fundamentales sobre los procesos evolutivos. Sin embargo, este potencial no ha sido explotado hasta el momento, y sólo un pequeño número de estudios cuantitativos utilizan los conodontos como herramienta de trabajo para abordar problemas micro y macroevolutivos (ver Jones, 2009 y referencias allí citadas). Dada su posición en la región oral del animal y su similitud morfológica con los dientes de otros grupos de vertebrados, esta gran variabilidad morfológica se ha relacionado estrechamente con la forma de procesar el alimento o con adaptaciones específicas a diferentes tipos de alimentos. El hallazgo de numerosas agrupaciones naturales, con los elementos preservados en su posición de vida, unido a la morfología molariforme de muchos grupos de conodontos, permitió sugerir a varios autores (Purnell, 1995; Donoghue & Purnell, 1999, entre otros) que los elementos plataforma o P1 del orden de los Ozarkodinidos (el grupo más abundante de conodontos) procesaban el alimento de forma similar a los molares de los mamíferos, mediante un proceso de oclusión relativamente sofisticado. Debido a que los conodontos no poseen mandíbulas, muchos grupos desarrollaron una compleja morfología que, de algún modo, ayudaría a un mayor control de este movimiento (Donoghue & Purnell, 1999). Este fenómeno, que fue descrito para el género Ideognathodus, parece ser común a otros grupos de conodontos de tipo plataforma, como Gnathodus o Polygnathus, siendo éste último uno de los géneros más abundantes de conodontos del Devónico y Carbonífero inferior. En todos estos grupos de conodontos es evidente el incremento de la complejidad morfológica de la superficie oral, pudiéndose identificar claramente dos regiones, la parte anterior y posterior respectivamente, con adaptaciones morfológicas distintas. En este contexto, el presente trabajo pretende profundizar, utilizando métodos de morfometría geométrica, en las tendencias de cambio morfológico y patrones de integración morfológicas del género Polygnathus durante la Emsiense (Devónico), intervalo temporal en el que el grupo experimentó su primera radiación morfológica. La hipótesis propuesta es que, dado que el elemento P1 presenta diferentes funciones en la parte anterior y posterior, esto quizás puede haber llevado a que ambas partes hayan evolucionado de manera independiente, a pesar de encontrarse en una única estructura la cual está habitualmente altamente integrada debido a los procesos de desarrollo. De encontrarse que parte anterior y posterior evolucionaron como dos módulos diferentes (es decir, de manera independiente), esto sugeriría un alto grado de plasticidad en el desarrollo de los elementos conodontales. El uso de la Morfometría Geométrica permite la captura de la forma de un objeto, estudiándolo de manera cuantitativa y facilitando la comparación con otros individuos. Para ello, en el presente trabajo, nos hemos centrado en el estudio de 18 especies del género Polygnathus 73 (definiendo 8 landmarks en 213 ejemplares), abarcando los primeros 15 millones de años de evolución del género. El estudio morfométrico de la plataforma de las distintas especies del género Polygnahus muestra una tendencia evolutiva hacia plataformas más anchas (aunque siempre más estrechas en la parte anterior), con ciertas modificaciones en la forma en que la lámina libre se una a la plataforma, así como a una reducción de la longitud de la lámina libre. Estos cambios están posiblemente relacionados con una mejora en el control de la oclusión y la función de masticación que realizarían, en concordancia con lo sugerido en la literatura para las tendencias evolutivas de los conodontos. En relación al análisis de modularidad, la hipótesis propuesta se corrobora, teniendo ambas zonas funcionales (anterior y posterior) un comportamiento modular y un bajo nivel de integración, a pesar de ser parte de una misma estructura. La existencia de ambos modules corrobora la idea de que las diferencias morfológicas entre ambas regiones pueden ser consecuencia de una selección funcional, estando los cambios de la region anterior relacionados con una mejora en el control de la oclusión y la función de masticación que realizarían. Agradecimientos. CMP es beneficiario de un contrato postdoctoral Marie Curie FP7-People IEF 2011299681 del séptimo Programa Marco de la Unión Europea. Este trabajo representa una contribución al IGCP 596. Bibliografía Donoghue, P.C.J. & Purnell, M.A. 1999. Mammal-like occlusion in conodonts. Paleobiology, 25 (1), 58-74. Jones, D. 2009. Directional evolution in the conodont Pterospathodus. Paleobiology, 35, 413 - 431. Purnell, M.A. 1995. Microwear on conodont elements and macrophagy in the first vertebrates. Nature, 374, 798-800. 74 Polygnáthidos del Emsiense en los Pirineos Centrales Españoles y su importancia para la comprensión de la evolución temprana del grupo C. Martínez-Pérez1 y J.I. Valenzuela-Ríos2 1 School of Earth Sciences, Univeristy of Bristol, Wills memorial Building, Queen’s Road, Bristol BS8 1RJ, United Kingdom. [email protected]. 2 Departamento de Geología, Universidad de Valencia, c/ Dr. Moliner 50, 46100 Burjasot, Valencia, España. [email protected]. El Sistema Devónico está subdividido en siete Pisos; de ellos, el Emsiense es quizás uno de los que más interés ha despertado en los últimos años, siendo los problemas de la redefinición de su límite inferior y su posible subdivisión en dos subpisos, algunos de los objetivos principales que se ha planteado la Subcomision Internacional de Estratigrafia del Devónico (SDS). Teniendo en cuenta estas intenciones se realizó un estudio en profundidad de los materiales de edad Emsiense que afloran de forma abundante en los Pirineos Centrales Españoles. Este estudio permitió la descripción en detalle de 9 secciones estratigráficas (Isábena 1, Baliera 6, Compte-I top, LGA, LGA-X, LGA-XI, Vi-IA, Vi-IB y Vi-IE) pertenecientes a las Subfacies Baliera y Compte del Área de Facies Sur de Mey (1967). Estas secciones libraron una abundante fauna de conodontos, principalmente especies del género Polygnathus, que son adecuadas para realizar estudios bioestratigráficos detallados. Estos registros comprenden desde el Praguiense más alto al Emsiense superior y han permitido establecer una subdivisión detallada para los materiales del Emsiense en los Pirineos Centrales Españoles. El registro más bajo del género es P. pireneae de edad Praguiense y procedente de los estratos del Miembro Ponferrat (miembro inferior de la Formación Basibé). Los siguientes registros del género provienen del miembro superior (Miembro Llaviero) de dicha Formación o de la parte alta de la equivalente Formación Castanesa, que proporcionaron la siguiente secuencia en orden estratigráfico: P. pireneae, P. kitabicus, P. excavatus excavatus, P. excavatus 114, P. sokolovi, P. rosae, P. pannonicus, P. gronbergi, P. nothoperbonus y P. luciae, junto con 3 especies nuevas del género. En la Fm. Villech la secuencia continúa con P. mashkovae, P. laticostatus, P. vigierei, P. gilberti, P. linguiformis bultyncki y P. serotinus. La secuencia pirenaica es de gran relevancia mundial ya que permite el reconocimiento de taxones cosmopolitas que son de gran importancia para correlaciones suprarregionales. Además, se identificaron algunos taxones que hasta la fecha se consideraban como endémicos de la región donde se ubica el estratotipo del Emsiense (Zinzilban/Uzbekistán). Por otro lado, es importante destacar que la secuencia de polygnátidos descrita en los Pirineos contiene los taxones índices que definen las diferentes biozonas del Praguiense final y del Emsiense, desde la Zona pireneae hasta la Zona serotinus, incluyendo tanto los índices del límite inferior del Emsiense actual (P. kitabicus), como el taxón elegido por la SDS para su próxima redefinición (P. excavatus 114). Asimismo, la riqueza y la buena sucesión estratigráfica de esta fauna de conodontos nos ha permitido proponer diferentes hipótesis filogenéticas de los taxones de polygnátidos estudiados, identificándose hasta 8 linajes diferentes, o partes de ellos. Asimismo, el establecimiento de los diferentes linajes evolutivos ha permitido corroborar la hipótesis de una radiación evolutiva de este género en las proximidades del límite Praguiense/Emsiense. Por último, cabe destacar que el presente estudio ha permitido aumentar el número de secciones relevantes con conodontos en los Pirineos, contribuyendo de esta manera a mejorar el conocimiento general de las faunas de conodontos del Emsiense. Esto ha posibilitado el establecimiento de una escala bioestragráfica local que es de gran utilidad para correlaciones tanto regionales como suprarregionales. 75 Agradecimientos. Este trabajo ha sido financiado por el MINECO proyecto CGL2011-24775, y el M100131201 “Hi-Res correlation Mid-Paleozoic”. Representa una contribución al PIGC 596 “Climate change and biodiversity patterns in the Mid-Palaeozoic (Early Devonian to Late Carboniferous)”. CMP es beneficiario de un contrato postdoctoral Marie Curie FP7-People IEF 2011-299681. La Fundación Alexander von Humboldt apoyó trabajos iniciales de J.I.V-R Bibliografía Mey, P.H.W. 1967. The geology of the Upper Ribagorzana and Baliera Valleys, Central Pyrenees, Spain. Leidse Geologische Mededelingen, 41, 153-220. 76 Cirtospiriféridos del Devónico de la Zona Centroibérica meridional M.V. Pardo Alonso 1 Departamento de Geología, Universitat de València, c/ Dr. Moliner, 50, 46100 Burjassot, Valencia, España. [email protected]. En la mayor parte de los niveles de areniscas y limolitas arenosas frasnienses de la Zona Centroibérica Meridional, especialmente en su parte central (Herrera del Duque, Almadén, Guadalmez y Cabeza del Buey), es frecuente encontrar formas atribuibles al género Cyrtospirifer. Pese a esta abundancia, el estudio de los ejemplares a nivel específico resulta muy problemático, debido en gran parte a la amplia variabilidad que presentan corrientemente las especies del género, a lo que hay que añadir las peculiaridades del material disponible, generalmente fragmentado y desarticulado. Autores anteriores clasificaron estos cirtospiriféridos asignándolos a diferentes especies, una de ellas descrita por vez primera en la propia región; la variedad de determinaciones sobre material procedente de los mismos yacimientos es un reflejo de la especial dificultad aludida. Así, se han citado las siguientes especies: Cyrtospirifer verneuili, Cyrtospirifer e. g. verneuili, C. disjunctus (como sinónimo de C. verneuili), C. almadenensis (descrita con material de la región), C. cf. canaliferus, C. syringothyriformis, C. cf. brodi, C. cf. schelonicus. En nuestra opinión, los cirtospiriféridos de la región pueden agruparse en cinco formas. Una de ellas, C.? archiaci, nunca fue citada anteriormente; otra, muy próxima a la anterior, por algunas características no comunes a ambas y la escasez de material queda separada de momento como C.? cf. archiaci. Bajo el nombre de C. almadenensis, Paeckelmann (1942) describió una especie que por su gran variabilidad, a la vista del material que hemos recogido en la región, o se trata de un único taxón muy variable, como supuso Paeckelmann (op. cit.) o más bien abarca dos especies distintas; de momento preferimos separar las dos morfologías, dejando en nomenclatura abierta las formas más alejadas del holotipo, designado por el autor original, y dejando la forma restante con la denominación provisional de Cyrtospirifer sp. 1. El estudio del material centroibérico lleva además a plantearnos la revisión de la diagnosis del género. La presencia de lamelas apicales dorsales ha llevado a diversos autores (ver p. ej. Brice, 1970: p. 115) a excluir del género a aquellas especies que las poseen, dado que estas estructuras están ausentes en la especie tipo, Cyrtospirifer verneuili (Murchison), y en C. syringothyriformis (Paeckelmann), una forma próxima con la que muchas veces se ha confundido. Sin embargo, dos de nuestros cirtospiriféridos, C. almadenensis y C. aff. almadenensis, cuya morfología coincide con la especie tipo en casi todos los caracteres considerados diagnósticos del género por anteriores autores, sí que presentan placas apicales dorsales, parcialmente embebidas por el espesamiento apical. La coincidencia en casi todos los caracteres diagnósticos, y el hecho de que las diferencias encontradas entre especies con lamelas apicales dorsales y especies sin ellas se sitúen a nivel de caracteres específicos, justifican el plantearse una enmienda a la diagnosis de Cyrtospirifer de modo que incluya también las especies portadoras de dichas estructuras. Agradecimientos. Este trabajo ha sido financiado por el proyecto MINECO CGL2011-24775. Representa una contribución al PIGC-596. Bibliografía Paeckelmann, W. 1942. Beitrage zur kenntnis devonischer Spiriferen. Abhandlungen des Reichsamts für bodenforschung, N.F., 197, 1-188. Brice, D. 1970. Étude paléontologique et stratigraphique du Dévonien de l'Afghanistan. Contribution a la connaissance des brachiopodes et des polypiers rugueux. Notes et Mémoires Moyen-Orient, 11, 1-364. 77 78 Corales rugosos del Misisípico de Alabama (USA) S. Rodríguez1 y D.C. Kopaska-Merkel2 1 Instituto de Geociencias (Consejo Superior de Investigaciones Científicas), Departamento de Paleontología, Universidad Complutense de Madrid, c/ José Antonio Novais 2, 28040 Madrid, España. [email protected]. 2 Geological Survey of Alabama, Box 869999, Tuscaloosa, Alabama 35486 (USA). [email protected]. Los corales rugosos son conocidos en Alabama y sus estados limítrofes desde mediados del siglo pasado, cuando algunas de las monografías clásicas sobre el tema incluyeron algunos ejemplares de esta edad y área (D’Orbigny, 1850; Milne-Edwards & Haime, 1851). Posteriormente, nuevas descripciones de corales fueron realizadas por Butts (1917, 1922, 1926) y, más recientemente, por Easton (1943, 1944) y Ehlers & Stumm (1953). Sin embargo, no se han desarrollado estudios recientes, aunque los hallazgos de corales son frecuentes y los museos de la región poseen colecciones importantes. Por ello se ha procedido a revisar las colecciones de tres instituciones de Alabama, así como a recolectar nuevo material en localidades escogidas. Las colecciones estudiadas son las del McWane Science Center de Birmingham, el Alabama Museum of Natural History de Tuscaloosa y, especialmente, el Geological Survey of Alabama, también en Tuscaloosa. Asociaciones de corales rugosos. Se han identificado cuatro asociaciones diferentes, correspondientes a distintas formaciones y edades, que van desde el Tournaisiense hasta el Serpujoviense. Corales rugosos de la Formación Chert de Fort Payne.- Los corales no son abundantes en esta formación, pero hay niveles con concentraciones interesantes. Sin embargo, debido a la especial litología la preservación suele ser muy mala, ya que los corales suelen estar fuertemente silicificados. Sólo se identifican corales sin disepimentos, típicos de ambientes no apropiados para el desarrollo de corales coloniales o solitarios de gran tamaño. La edad de la asociación es Tournaisiense: Barylasma tennesseense (Miller and Gurley, 1893) Baryphyllum ovale (Bassler, 1937) Zaphrentites centralis, (Milne-Edwards and Haime, 1851) Zaphrentites cliffordianum (Milne-Edwards and Haime, 1851) Corales rugosos de la Formación Calizas de Tuscumbia.- La asociación está compuesta principalmente por corales coloniales que pueden ser localmente muy abundantes, pero poco diversos. La edad de la asociación es Viseense inferior: Acrocyathus floriformis d’Orbigny 1850 Acrocyathus proliferus (Hall in Hall and Whitney, 1858) Zaphrentites centralis (Milne-Edwards and Haime, 1851) Corales rugosos de las formaciones Calizas de Monteagle y Pizarras y Calizas de Pride Mountain.- Ambas formaciones son sincrónicas y representan cambios laterales de facies. La asociación es bastante pobre y presenta predominio de corales sin disepimentos, pues las condiciones no eran muy favorables para el desarrollo de los corales, aunque otros grupos fósiles como equinodermos y briozoos son muy abundantes. La edad de la asociación es Viseense superior: 79 Siphonodendron martini (Milne-Edwards and Haime, 1851) Zaphrentites spinulosum (Milne-Edwards and Haime, 1851) Zaphrentites sp.nov. Amplexizaphrentis sp. Corales rugosos de la Formación Calizas de Bangor.- La asociación más diversa (sin llegar a serlo mucho) y más abundante. Constituida principalmente por corales coloniales fasciculados del género Lublinophyllum que forman biostromos que pueden seguirse a lo largo de centenares de kilómetros y montículos de pequeño tamaño (Kopaska-Merkel & Haywick, 2001). La edad de la asociación es Viseense terminal-Serpujoviense: Zaphrentites spinulosum (Milne-Edwards and Haime, 1851) Palastraea sp.nov. Arachnolasma sp. Lublinophyllum flaccidum (Easton, 1943) Siphonophyllia tennesseensis (Easton, 1943) Siphonophyllia sp. nov. Las asociaciones son altamente endémicas, presentes sólo en América del Norte, y sólo hay formas que las relaciona con cuencas del Paleotethys en las asociaciones del Viseense superior. Agradecimientos. Los autores agradecen su colaboración en los trabajos de campo a James Lacefield y las facilidades proporcionadas por los conservadores del Mc Wane Center, GSA y Alabama Museum of Natural History. Los fondos para la investigación han sido proporcionados por el proyecto CGL201230922. Este artículo es parte del PIGC 596. Bibliografía Butts, C. 1917. Description and correlation of the Mississippian formations of western Kentucky. Kentucky Geological Survey, 119 pp. Butts, C. 1922. The Mississippian series of eastern Kentucky. Kentucky Geological Survey, 5 (7), 1-188. Butts, C. 1926. Geology of Alabama; the Paleozoic rocks. Alabama Geological Survey, Special Reports, 14, 41-230. D’Orbigny, A. 1850. Prodrome de paleontology stratigraphique universelle des animaux mollusques et rayonnés, etc. Victor Masson, Paris, 394 pp. Easton, W.M. 1943. Revision on Campophyllum in North America. Journal of Paleontology, 18, 119132. Easton, W.M. 1944. New Chester corals from Alabama and Tennessee. Journal of Paleontology, 17, 276280. Ehlers, G.M. & Stumm, E.C. 1953. A new species of the tetracoral genus Palastraea from the Mississippian of Kentucky. Papers of the Michigan Academy of Science, Arts and Letters, 38, 383387. Kopaska-Merkel, D.C. & Haywick, D.W. 2001. A lone biodetrital mound in the Chesterian (Carboniferous) of Alabama? Sedimentary Geology, 145, 253-268 Milne-Edwards, H. & Haime, J. 1851. Monographie des polypiers fossils des terrains Paléozoiques, precede d’un tableau general de la classification des Polypes. Archives du Museum d’Histoire Naturelle de Paris, 5, 1-502. 80 Revisión del criterio de correlación mediante conodontos para el límite Misisípico/Pensilvánico J. Sanz López1 y S. Blanco Ferrera1 1 Departamento de Geología, Facultad de Geología, Universidad de Oviedo, c/ Arias de Velasco s/n, 33005 Oviedo. [email protected]; [email protected]. La mejora en el conocimiento del registro fósil a lo largo de los últimos años ha puesto a prueba los criterios de reconocimiento y correlación de gran parte de las secciones y puntos en los estratotipos globales (GSSPs) seleccionados desde la aceptación en 1972 de la base del Sistema Devónico en el Congreso Geológico Internacional de Montreal. La respuesta desde las diferentes subcomisiones de la Comisión Internacional de Estratigrafía ha sido distinta y en unos pocos casos ha incluido llamamientos a la revisión de los GSSPs después de una moratoria de 10 años desde su definición. Así, el límite inferior del Carbonífero se encuentra en discusión como consecuencia de la incertidumbre en la taxonomía del criterio primario de correlación y la escasa idoneidad de la sección tipo. También, la base del Emsiense está en revisión, ya que la unidad definida comprende un intervalo de tiempo mayor y distinto a la unidad cronoestratigráfica original que se pretendía limitar (Becker, 2009). Tal vez para evitar este problema, la Subcomisión para la Estratigrafía del Cámbrico ha optado por definir primero las unidades cronoestratigráficas operativas, y seguidamente nombrarlas. Sin embargo y en muchos otros casos, la publicación de trabajos en los que se modificaban los criterios de correlación, o se refinaba el margen de incertidumbre de la correlación, ha sido seguida por otros estudios en los que se aceptaban los mismos sin que hasta ahora esos cambios hayan sido necesariamente ratificados por la IUGS. Este es el caso de los límites de los pisos del Silúrico fijados entre 1980 y 1984, donde se están utilizando otros taxones a los propuestos inicialmente en los GSSPs como indicadores primarios (Melchin et al., 2012). Otros estudios sugieren la necesidad de confirmar la correlación de la primera aparición del marcador primario mediante un marcador geoquímico complementario, como es el caso de la base del Fanerozoico (Landing et al., 2013). En nuestro caso, Sanz-López et al. (2006) señalaron la baja resolución cronoestratigráfica del indicador primario seleccionado para la base del Pensilvánico y propusieron en su lugar la primera aparición del conodonto Declinognathodus inaequalis, complementado por la posición relativa de otros horizontes significativos. El establecimiento del GSSP para la base del Piso Bashkiriense, y por tanto del Subsistema Pensilvánico y de la Serie Pensilvánico Inferior (Heckel & Clayton, 2006) fue aprobado por la Comisión Internacional de Estratigrafía y ratificada por el Comité Ejecutivo de la IUGS en enero de 1996. El horizonte seleccionado está localizado a 82,90 m sobre la base de la Formación Bird Spring en la sección de Arrow Canyon (Nevada), en la primera aparición del conodonto Declinognathodus noduliferus (Ellison & Graves, 1941) sensu lato, según el concepto derivado de Lane et al. (1999), y en la línea evolutiva desde Gnathodus girtyi simplex Dunn, 1965. La utilización de este taxón como criterio de correlación cronoestratigráfica supone un grado alto de imprecisión. D. noduliferus sensu lato no es una especie, sino un grupo de especies en la evolución temprana del género. Por tanto, su primera aparición aglutina a varios procesos de especiación separados razonablemente en el tiempo. Además, el holotipo de D. noduliferus tiene una edad incierta, ya que proviene de una capa de la Formación Marathon, que incluye turbiditas con asociaciones mezcladas de conodontos del Frasniense a la base del Moscoviense. Finalmente, los primeros declinognátidos parecen evolucionar desde Gnathodus postbilineatus, o de una especie del grupo de G. bilineatus. 81 Nuestros estudios recientes de los conodontos de la Formación Barcaliente (norte de la península Ibérica) refuerzan las conclusiones adelantadas en Sanz-López et al. (2006) y soportan un esquema bioestratigráfico que a nuestro juicio enriquece la historia del grupo. En dicho esquema existe una separación en las dos provincias o faunas de conodontos diferenciadas por Higgins (1981), “Midcontinent” norteamericano y Euroasia. Los primeros declinognátidos (D. bernesgae y D. praenoduliferus) evolucionaron al final del Serpukhoviense y estuvieron restringidos a algunas áreas de la provincia Euroasiática durante el intervalo de la crisis biótica del final del Misisípico. En el Bashkiriense temprano expandieron su distribución durante la radiación de Declinognathodus y de otros géneros de la Familia Idiognathodontidae. Dos acontecimientos de aparición con distribución geográfica amplia están indicados por los taxones D. inaequalis y D. lateralis. Entre ambos horizontes se originaron Neognathodus higginsi y los primeros Idiognathoides (I. asiaticus e I. macer), aunque su presencia local está limitada por la condiciones paleoecológicas locales y la paleogeografía. Así, los primeros Idiognathoides parece que se encuentran en Asia Central, para más tarde llegar a los Urales y al norte de la península Ibérica, luego extenderse por otras cuencas durante pulsos transgresivos, y finalmente llegar al “Midcontinent” norteamericano. Por otro lado, la desaparición de las especies típicas del Misisípico también ocurre de forma diacrónica en el Bashkiriense temprano de la provincia Euroasiática. Resulta llamativo que Cavusgnathus haya persistido en algunas partes de Eurasia (Urales Meridionales y Japón) hasta antes de la entrada de D. lateralis, mientras parece extinguirse en el Serpukhoviense superior de la provincia del “Midcontinent” norteamericano en paralelo a la evolución y presencia de otros géneros de aguas someras, como Rachistognathus y Adetognathus. Agradecimientos. Por su soporte económico, a los proyectos CGL-2009-09109 (Ministerio de Economía y Competitividad, Fondos FEDER) y SV-PA-13-ECOEMP-46 (Principado de Asturias). Este trabajo representa una contribución al IGCP-596. Bibliografía Becker, R.T. 2009. Minutes of the SDS Business Meeting, Kitab Sate Geological Reserve, Uzbekistan. Subcommision on Devonian Stratigraphy Newsletter, 24, 12-15. Heckel, P.H. & Clayton, G. 2006. The Carboniferous System. Use of the new official names for the subsystems, series, and stages. Geologica Acta, 4, 403-407. Higgins, 1981. The distribution of conodonts in relation to the palaeogeography of late Visean-Namurian time. In: Neale, J.W. &. Brasier, M.D (eds.), Microfossils from recent and fossil shelf seas. Ellis Horwood Limited, Chichester, 37-51. Landing, E., Geyer, G., Brasier, M.D. & Bowring, S.A. 2013. Cambrian Evolutionary Radiation: Context, correlation, and chronostratigraphy - Overcoming deficiencies of the first appearance datum (FAD) concept. Earth-Science Reviews, 123, 133-172. Lane, H.R., Brenckle, P.L., Baesemann, J.F. & Richards, B. 1999. The IUGS boundary in the middle of the Carboniferous: Arrow Canyon, Nevada, USA. Episodes, 22, 272-283. Melchin, M.J., Sadler, P.M. & Cramer, B.B. 2012. The Silurian Period. In: Gradstein, F.M., Ogg, J.G., Schmitz, M. & Ogg, G. (eds.), The Geologic Time Scale 2012. Elsevier, Oxford, 525-558. Sanz-López, J., Blanco-Ferrera, S., García-López, S. & Sánchez de Posada, L.C. 2006. The MidCarboniferous boundary in Northern Spain: difficulties for correlation of the Global Stratotype Section and Point. Rivista Italiana di Paleontologia e Stratigrafia, 112, 3-22. 82 Correlación de las secuencias de conodontos del Lochkoviense (Devónico Inferior) de Nevada (USA) y de los Pirineos Centrales Españoles J.I. Valenzuela-Ríos1 y M.A. Murphy2 1 Departmento de Geología, Universidad de Valencia, c/ Dr. Moliner 50, 46100 Burjasot, Valencia, España. [email protected]. 2 Department of Earth Sciences, University of California, Riverside, Ca 92521, USA. [email protected]. Las secuencias de conodontos del Lochkoviense y Praguiense (Devónico Inferior) de Nevada Central (USA, Laurentia) consituyeron la base para el establecimiento de la zonación de la Cordillera Norteamericana, en el oeste de Estados Unidos de Norteamérica, que posteriormente se extendió globalmente. El estudio de las secuencias de conodontos del Lochkoviense de los Pirineos Centrales Españoles permitió identificar algunos de los puntos débiles de esta zonación y el establecimiento de una zonación más detallada que parcialmente podía corresponderse con la de Nevada. La comparación de ambas zonaciones permite el establecimiento de una correlación muy detallada para los estratos del Lochkoviense medio y superior. Esto se debe fundamentalmente a la presencia de taxones cosmopólitas en ambas regiones. Por el contrario, la marcada diferencia en la composición faunísitca del Lochkoviense inferior no permite, por el momento, correlaciones detalladas directas entre ambas regiones. El Lochkoviense medio comienza con Lanea omoalpha que se registra en numerosas secciones de Nevada y de los Pirineos. Por encima siguen los registros sucesivos de Ancyrodelloides transitans, A. trigonicus y A. kutscheri en ambas regiones que permiten una subdivisión más fina. Además, los registros de L. eleanorae, L. telleri, Flajsella schulzei, F. stygia, F. streptostygia, Kimognathus delta y K. limbacarinatus en similares posiciones estratigráficas en Nevada y en los Pirineos permiten aumentar la calidad de las correlaciones entre ambas regiones, así como subdivisiones temporales para algunos tramos con precisiones temporales cercanas a los 200.000 años. El Lochkoviense superior comienza con Masaraella pandora beta. Este taxón se registra en secuencia evolutiva por encima de su ancestro (M. pandora alpha) y por debajo de su descendiente (M. pandora gamma) en numerosas secciones tanto en los Pirineos como en Nevada. La importante radiación de Pedavis acontecida en el Lochkoviense superior, permite la subdivisión del mismo en dos zonas, pandora beta-gilberti y gilberti-irregularis. Los registros en posiciones estratigráficas análogas de P. robertoi y su secuencia de morfotipos que evolucionan hacia P. gilberti, y de M. pandora pi, posibilitan correlaciones muy detalladas. El rico registro de conodontos en las secciones de Nevada y de los Pirineos Centrales Españoles permiten establecer unos intervalos temporales de correlación, que en ocasiones están próximos a 200.000 años. Estos intervalos serán la base para futuros estudios globales y correlaciones detalladas entre Laurentia y el Norte de Gondwana. Agradecimientos. Este trabajo ha sido financiado por los proyectos MINECO CGL2011-24775, el M100131201 “Hi-Res correlation Mid-Paleozoic” y MECyD Programa Salvador de Madariaga. Representa una contribución al IGCP-596. 83 Conodontos del Lochkoviense (Devónico Inferior) en los Pirineos Centrales Españoles y su correlación con las secuencias de Bohemia (República Checa) J.I. Valenzuela-Ríos1, L. Slavík2, H. Calvo1, A. Hušková2, J-C. Liao1,3 y L. Koptíková2 1 Departmento de Geología, Universidad de Valencia, c/ Dr. Moliner 50, 46100 Burjasot, Valencia, España. [email protected]. 2 Institute of Geology, AS CR, Rozvojova 269, 16500 Prague 6, Czech Republic. [email protected] 3 Departamento de Paleontología, Universidad Complutense de Madrid, c/ José Antonio Novais, 2, 28040 Madrid, España. [email protected]. Las sucesiones de carbonatos marinos del Devónico Inferior del Sinforme de Praga comprenden una amplia gama de ambientes marinos, desde plataforma a base de talud, y han servido para la elaboración de la escala cronoestratigráfica de los Pisos Hercínicos, dos de los cuales (Lochkoviense y Praguiense) son referentes mundiales y comprenden los estratotipos de referencia. Su contenido en conodontos es importante, pero solo recientemente se han comenzado a realizar estudios detallados que permitan compararlos con los existentes en otras regiones claves. Las sucesiones marinas del Devónico Inferior de los Pirineos Centrales Españoles corresponden fundamentalmente a ambientes de plataforma y contienen una de las secuencias de conodontos más ricas a nivel mundial, especialmente para el Piso Lochkoviense. La comparación de las secuencias de conodontos del Lochkoviense (Devónico Inferior) de los Pirineos Centrales Españoles y de Barrandia (República Checa) demuestra que estas dos áreas, que se ubicaron en las regiones periféricas al norte de Gondwana durante el Devónico, comparten muchos taxones. Así, los sucesivos registros de conodontos permiten establecer correlaciones detalladas en el Lochkoviense, que son más precisas en el Lochkoviense medio. En ambas regiones se registran taxones de los géneros cosmopolitas Ancyrodelloides, Lanea, Masaraella, Pedavis y Kimognathus y también de los géneros Icriodus y Pelekysgnathus cuyos taxones presentan una distribución más restringida. Es destacable la coincidencia en ambas regiones de los respectivos pasos evolutivos de diferentes géneros. En el Lochkoviense inferior taxones de los géneros Icriodus, Pelekysgnathus y Zieglerodina son comunes en ambas regiones. Los estratos del Lochkoviense superior comparten taxones de los géneros Pedavis, Masaraella y Wurmiella. En este trabajo se establece una primera correlación detallada basada en el contenido en conodontos, su distribución estratigráfica y los eventos evolutivos (innovación, especiación, extinción) más importantes en estas dos regiones. Este estudio permite la elaboración de una detallada base de datos que será la base sobre la que se apoyaran las correlaciones detalladas entre las secuencias del Lochkoviense en las regiones perigondwánicas de Europa y el Norte de África, incluyendo localidades importantes de los Alpes Cárnicos, Celtiberia, La Bretaña francesa y Marruecos. Nuestro estudio contribuye a aumentar la red de correlación mundial del Lochkoviense, que podrá servir de gran ayuda para la futura subdivisión del Lochkoviense en tres subpisos. Agradecimientos. Este trabajo ha sido financiado por el MINECO proyecto CGL2011-24775, y el M100131201 “Hi-Res correlation Mid-Paleozoic”. Representa una contribución al IGCP-596 y ACI20091037. 84 Formaciones Escalada y Fito (Unidad del Ponga, Zona Cantábrica): Contenido en fusulinas y correlación C. González Parra1 y E. Villa Otero1 1 Departamento de Geología, Universidad de Oviedo. [email protected]; [email protected]. En este trabajo, se aportan nuevos datos sobre el contenido en fusulinas de las formaciones Escalada y Fito (Unidad del Ponga, Zona Cantábrica, NW de España). Las muestras recogidas proceden de los cortes de Campo de Caso, Taranes I, Taranes II y Sebarga, representativos de la estratigrafía de dicha Unidad. Los nuevos hallazgos confirman la edad de ambas formaciones (van Ginkel, 1965; Villa, 1995) y precisan la de la parte alta de la Fm. Fito. Ambas formaciones representan en conjunto el Moscoviense superior (Podolskiense y Myachkoviense), si bien la parte baja de la Fm. Escalada del corte de Campo de Caso todavía pertenece al Moscoviense inferior (Kashiriense). Figura 1. Algunas fusulinas relevantes de las secciones estudiadas. FE: Fm. Escalada; FF: Fm. Fito. Gracias a estos datos ha sido posible correlacionar las secciones estudiadas en la Unidad del Ponga. Además, puesto que un buen número de especies son comunes con las descritas por van Ginkel (1965) en la Unidad del Pisuerga-Carrión (Fusulinella pandae, F. abismoe, F. branoserae, Fusulina agujasensis y F. cotarazoe), se facilita la correlación entre las sucesiones de las unidades del Ponga y Pisuerga-Carrión. 85 Figura 2. Correlación estratigráfica (K: Kashiriense; P: Podolsk.; M1: Myach. inf.; M2: Myach. medio). Agradecimientos. Proyecto SV_PA-13-ECOEMP-46, subvencionado por el Gobierno del Principado de Asturias. Representa una contribución al IGCP-596. Bibliografía Ginkel, A.C. van 1965. Carboniferous fusulinids from the Cantabrian Mountains. Leidse Geologische Mededelingen, 34, 1-225. Villa Otero, E. 1995. Fusulináceos carboníferos del este de Asturias (N de España). Biostratigraphie du Paléozoïque, 13, 1-261.