Inmunidad en el Deporte
Anuncio

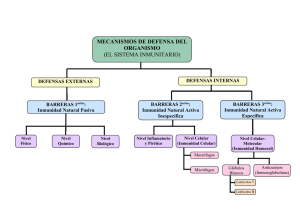
Universidad del País Vasco Euskal Herriko Unibertsitatea Departamento de Fisiología Programa de Doctorado en Fisiología 2003 - 2005 Inmunología Celular Inmunidad en el Deporte Natalia I. Wiereszen Inmunidad en el Deporte Indice Introducción 3 Características Generales del Sistema Inmune 4 El Sistema Inmune y el Ejercicio Físico 12 Ventajas y Desventajas del Ejercicio sobre la Capacidad de Respuesta Inmune 20 Conclusiones 22 Referencias Bibliográficas 23 Programa de Doctorado en Fisiología 2003 – 2005 Natalia I. Wiereszen 2 Inmunidad en el Deporte Introducción El sistema inmune comprende todos aquellos mecanismos fisiológicos que dotan al organismo de la capacidad de reconocer materiales extraños al mismo, e incluye mecanismos de defensa no específicos. El ejercicio conlleva una serie de demandas sobre el organismo que son dependientes de la forma, intensidad y duración del mismo y que, a su vez, tienen profundas repercusiones sobre la capacidad de respuesta inmune del deportista. De modo recíproco, la capacidad de respuesta inmune del deportista repercute sobre la salud y ésta sobre su rendimiento. El estudio de los efectos del ejercicio sobre el sistema inmune necesita de la comprensión de una red de interacciones entre el sistema nervioso central, el endocrino y el mismo sistema inmune, ya que serán los encargados de la respuesta al estrés originado por la actividad física. El componente nervioso incluye la inervación simpática de la corteza suprarrenal y los ganglios, el endocrino a las hormonas de estrés producidas por la estimulación del eje hipotálamo-hipófisissuprarrenal; y el sistema inmune participa produciendo mediadores inmunológicos que condicionan la respuesta hormonal. El ejercicio altera la distribución, el tráfico y las capacidades funcionales de los distintos tipos de células inmunes. Como también, causa alteraciones en los niveles locales y sistémicos de diversos mediadores moleculares1-2-5-9-13. El objetivo de este trabajo es: identificar los efectos del ejercicio sobre los distintos componentes del sistema inmune. Una revisión bibliográfica de los últimos años, junto a la consulta en las bases de datos Medline y Lilacs y la seleccionaron de los artículos originales cuyas fechas de publicación oscilaban entre los años 1994 – 2004; han sido la fuente del trabajo que a continuación se expone. Programa de Doctorado en Fisiología 2003 – 2005 Natalia I. Wiereszen 3 Inmunidad en el Deporte Características Generales del Sistema Inmune La inmunidad es la capacidad del organismo para luchar contra los microorganismos y las toxinas que intentan dañar los tejidos y órganos1. El sistema inmune reconoce y reacciona frente a moléculas de organismos capaces de provocar enfermedad, a los que agrede de varias formas. Para ello, este sistema discrimina las moléculas propias y extrañas (antígenos), y responde tolerando a las primeras y reaccionando contra las segundas. Las células del sistema inmune son capaces de reconocer distintos antígenos y desarrollar diferentes tipos de respuestas frente a ellos (Figura 1)2. Gran parte de la inmunidad depende de un especial sistema productor de anticuerpos y linfocitos activados que atacan y destruyen los microorganismos específicos o sus toxinas. Esta es la Inmunidad Específica o Adquirida. Otra parte de la inmunidad resulta de procesos generales, dado por los leucocitos y las células de los tejidos que originalmente procedentes de ellos. Esta es la Inmunidad Inespecífica o Innata. Figura 1. Células del Sistema Inmune Programa de Doctorado en Fisiología 2003 – 2005 Natalia I. Wiereszen 4 Inmunidad en el Deporte Inmunidad Inespecífica o Innata Los leucocitos o glóbulos blancos son “unidades móviles” del sistema protector del cuerpo. Se forman en la médula ósea y en el tejido linfoide, pero después de producidos son transportados por la sangre a diferentes partes del organismo, donde ejercen sus funciones. El valor fundamental de los leucocitos estriba en que son transportados específicamente a zonas donde hay inflamación, proporcionando así una defensa rápida y enérgica contra cualquier posible agente infeccioso3. Normalmente se encuentran en la sangre 6 tipos de glóbulos blancos3: 9 Polimorfonucleares neutrófilos 9 Polimorfonucleares eosinófilos 9 Polimorfonucleares basófilos 9 Monocitos (macrófagos tisulares) 9 Linfocitos 9 Células Plasmáticas Los 3 tipos de células polimorfonucleares tienen aspecto granuloso, por lo que se denominan granulocitos. Los granulocitos y los monocitos protegen el cuerpo contra gérmenes invasores captándolos por fagocitosis. Los linfocitos y células plasmáticas funcionan principalmente en relación con el sistema inmunitario. Estas células pueden responder de distinta manera a los patógenos: a) Los neutrófilos y los monocitos-macrófagos lo fagocitan y lo destruyen en su interior. b) Los eosinófilos, basófilos y linfocitos citotóxicos espontáneos (NK) exocitan productos tóxicos y pro inflamatorios al exterior. Así, los primeros destruyen los patógenos y, los segundos desencadenan la respuesta inflamatoria. Los neutrófilos constituyen el 62% de todos los leucocitos, son células maduras que pueden atacar y destruir bacterias y virus incluso en la sangre circulante. Proceso de Fagocitosis12 En cambio, los monocitos sanguíneos (5.3%) son células inmaduras con muy poca capacidad para luchar contra los gérmenes infecciosos. Sin embargo, una vez que entran en los tejidos comienzan a hincharse, aumentando su diámetro hasta cinco Programa de Doctorado en Fisiología 2003 – 2005 Natalia I. Wiereszen 5 Inmunidad en el Deporte veces, ahora estas células se llaman macrófagos y tiene una gran capacidad para combatir a los agentes patógenos. Estas células fagocíticas poseen un mecanismo de reconocimiento de patógenos cuya base molecular son receptores no antígeno específicos. Estos receptores guían a éstas células hacia los patógenos que deben fagocitar. Tienen receptores para lipopolisacárido bacteriano (CD14) y para otros componentes de la pared bacteriana. También expresan receptores para componentes activados del complemento (receptores de C3b) y para los anticuerpos (receptores de IgG) unidos a los patógenos2-12. Los eosinófilos constituyen del 2 al 3% de todos los leucocitos, con frecuencia se producen en gran cantidad cuando existe una infección parasitaria3. Vierten el contenido de sus gránulos que contienen varias proteínas como peroxidasas, ribonucleasas y la proteína básica principal del eosinófilo (BMP) activadora de neutrófilos, basófilos, mastocitos y macrófagos. También contienen proteínas que son capaces de lesionar tejidos como la cationica (ECP) y la neurotoxina. Asimismo, tienen enzimas anti-inflamatorias (histaminasa, arilsulfatasa, fosfolipasa D) que atenúan o interrumpen reacciones inflamatorias. También producen citocinas IL-3, IL-5 y GM-CSF que prolongan su supervivencia, citocinas pro-inflamatorias como TNFα, IL-6, IL-8 y otras anti-inflamatorias como TGFα y TGFβ. Estas células expresan receptores de baja afinidad para la región constante de la IgE (CD23) y de la IgG (CD32)2. Los basófilos (0.4%) y los mastocitos son células con receptores para la IgE y derivan de precursores comunes. Los basófilos se encuentran en la sangre y los mastocitos en el tejido conectivo, concretamente en la dermis y en los tejidos submucosos del tracto respiratorio y gastrointestinal, donde dependen de los factores de crecimiento producidos por los fibroblastos. Entre los mediadores que contienen sus gránulos podemos destacar aminas vasoactivas como la histamina y la serotonina, los proteoglicanos, la heparina, las carboxipeptidasas, las serinaproteasas y las sulfatasas. También producen PGD2, leucotrienos y factor activador de plaquetas (PAF). Su activación aumenta el flujo sanguíneo local y la extravasación de fluído en los tejidos adyacentes, favoreciendo la extravasación de células y proteínas activas frente a los patógenos. Los mastocitos sintetizan, tras el entrecruzamiento con sus Ig, mediadores lipídicos derivados del ácido araquidónico: prostaglandinas (PGD-2), tromboxanos y leucotrienos. Estos compuestos mantienen la inflamación. A estas células se las vincula con procesos alérgicos. Dentro de las células exocíticas, las NK tienen receptores para IgG, mientras que los basófilos y los eosinófilos tienen receptores para IgE2. Programa de Doctorado en Fisiología 2003 – 2005 Natalia I. Wiereszen 6 Inmunidad en el Deporte Inmunidad Específica o Adquirida El cuerpo humano tiene capacidad de desarrollar una inmunidad específica muy poderosa contra agentes invasores concretos (bacterias, virus, toxinas, proteínas extrañas). Existen dos tipos básicos, estrechamente relacionados, de inmunidad. En uno de ellos, los linfocitos B producen anticuerpos circulantes, moléculas de globulina capaces de atacar al agente invasor. Este tipo de inmunidad se llama Inmunidad Humoral. En el segundo tipo de inmunidad adquirida, los linfocitos T, forman linfocitos activos, programados de manera específica para destruir al agente extraño. Este tipo de inmunidad se llama Inmunidad mediada por células T o Inmunidad Celular3. Los linfocitos no sólo están en la sangre, también se encuentran en los ganglios linfáticos y en tejidos linfoides (bazo, médula ósea, submucosa del tracto gastrointestinal, amígdalas y adenoides). Los linfocitos de la médula ósea constituyen una reserva que regenera los linfocitos circulantes1. Los linfocitos T y B, tienen receptores con los que reconocen antígenos específicos. Los linfocitos T reconocen antígenos presentados por otras células, mientras que los linfocitos B reconocen antígenos libres. El reconocimiento antigénico induce en los linfocitos B su diferenciación en células plasmáticas secretoras de anticuerpos, mientras que en los linfocitos T induce su diferenciación en linfocitos T efectores. Los linfocitos T CD4 se diferencian en efectores cooperadores (Th) y los T CD8 en efectores citotóxicos (Tc)2. Ambos tipos de linfocitos T efectores (Th y Tc) responden de distinta forma a la estimulación con el antígeno2: 9 Los linfocitos T citotóxicos secretan proteínas que matan a las células en las que reconocen el antígeno. 9 Los linfocitos T cooperadores secretan moléculas de comunicación intercelular (citocinas) que estimulan a otras células inmunes. Ejercen funciones reguladoras sobre macrófagos y linfocitos B. Activan a los primeros para que secreten, productos pro inflamatorios y antimicrobianos (radicales oxidantes, óxido nítrico, lisozima), al tiempo que cooperan con los linfocitos B para que éstos sinteticen y secreten anticuerpos. Estos anticuerpos (Ac) son moléculas proteicas solubles que reconocen y se unen a los antígenos (Ag) de los patógenos. Estas uniones Ac-Ag (llamadas inmunocomplejos) atraen a otras moléculas y células del sistema inmune que contribuirán a la eliminación del antígeno (Figura 2)2. Programa de Doctorado en Fisiología 2003 – 2005 Natalia I. Wiereszen 7 Inmunidad en el Deporte Figura 2. Los anticuerpos dirigen el antígeno hacia las células el sistema inmune Tras la estimulación con el antígeno, las células B que lo reconocen también sufren procesos de diferenciación celular hacia células plasmáticas que se especializan en la secreción de anticuerpos de un único isotipo. Así, cada célula plasmática produce un único isotipo de inmunoglobulina: IgA, IgM, IgD, IgE, IgG1, IgG2, IgG3 o IgG4. Los distintos isotipos tienen funciones inmunes especializadas, pues reclutan a diversos tipos de moléculas y células en la lucha contra el antígeno que reconocen. La unión de anticuerpos de ciertos isotipos al patógeno desencadena la activación del complemento exclusivamente en el espacio vascular (IgM), o indistintamente en el espacio vascular o el extravascular (IgG1, IgG2 e IgG3). La de otros isotipos como la IgE provoca la liberación de mediadores pro inflamatorios en los tejidos desde los mastocitos, los basófilos y los eosinófilos2. Comunicación Intercelular Las células del sistema inmune se comunican entre sí mediante moléculas de sus membranas plasmáticas, y a través de la secreción de moléculas de comunicación intercelular solubles, las citocinas. Las células del sistema inmune también están sometidas a regulación por células de otros sistemas, como son el sistema nervioso y el endocrino2. Las citocinas se dividen en una serie de familias (Tabla 1). Los mecanismos de acción de las citocinas son usualmente yuxtacrinos (sobre células adyacentes) o paracrinos (sobre células del mismo tejido), aunque algunas citocinas como la IL-1, Programa de Doctorado en Fisiología 2003 – 2005 Natalia I. Wiereszen 8 Inmunidad en el Deporte la IL-6 y el factor de necrosis tumoral α (TNFα) pueden además ejercer una función endocrina. Las citocinas participan en la regulación de las respuestas inflamatorias secundarias al ejercicio2. FAMILIA ESTRUCTURAL Citocinas de Tipo 1 CITOCINAS QUE INCLUYE IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-9, IL-11, IL-12, IL-13, IL-15 y algunos factores de crecimiento hematopoyéticos Citocinas de Tipo 2 Interferones, IL-10 Familia del TNF TNF, Leucotrienos (LT) Miembros de la superfamilia de Ig IL-1, Quimiocinas Tabla 1. Familias estructurales de citocinas Las citocinas producidas por los linfocitos T cooperadores son decisivas en la determinación del tipo de respuesta inmune. El interferón γ estimula respuestas inmunes del tipo 1 en las que participan anticuerpos fijadores de complemento y células efectoras fagocíticas (neutrófilos y monocitos). Por el contrario, la IL-4 provoca respuestas inmunes de tipo 2 en las que participan anticuerpos de isotipo IgE y células exocíticas (basófilos y eosinófilos)2. Mecanismos Celulares Efectores de la Respuesta Inmune La respuesta inmune pretende eliminar el agente patógeno, o al menos, controlar su dispersión y virulencia. Para ello dispone de una variedad de tipos celulares con distintas armas. Los linfocitos citotóxicos (T y NK) matan células infectadas o estresadas. Aunque los mecanismos por los que Tc y NK reconocen a sus víctimas (células diana) son distintos, los mecanismos efectores por los que matan a sus células diana son comunes e incluyen dos grandes tipos2: A. Secreción de gránulos que contienen perforinas y proteínas inductoras de la apoptosis. B. Moléculas de membrana (Fas ligando) que inducen apoptosis en células diana que expresan la molécula Fas. Los linfocitos T citotóxicos reconocen con su receptor antigénico a un antígeno de un patógeno determinado y matan a las células infectadas que lo presentan. Programa de Doctorado en Fisiología 2003 – 2005 Natalia I. Wiereszen 9 Inmunidad en el Deporte Las células citotóxicas espontáneas, o células NK (natural Killer), reconocen a las células que tienen que matar por distintos mecanismos2: A. Citotoxicidad dependiente de anticuerpo, que dirige a las células NK contra las células cuyo antígenos son reconocidos por anticuerpos de isotipo IgG. B. Receptores para la proteína de estrés HSP72, que les sirve para reconocer y eliminar a células sometidas a un elevado nivel de estrés. C. Receptores para moléculas de histocompatibilidad de clase I, que detectan a las células con una anómala expresión de moléculas de histocompatibilidad. De este modo reconocen y lisan a las células que han perdido expresión de moléculas de clase I. Existen otras células con receptores para la fracción constante de la IgG (FcIgG), los leucocitos mononucleares (monocitos, macrófagos y células dendríticas) y leucocitos polimorfonucleares (neutrófilos) que, por medio de estos y otros receptores (de complemento y de lipopolisacárido bacteriano), reconocen y fagocitan los patógenos que serán destruidos en sus fagolisosomas. Las células fagocíticas mononucleares no sólo destruyen patógenos, sino que procesan los antígenos fagocitados y los presentan en moléculas de histocompatibilidad a loslinfocitos T. Los linfocitos mononucleares producen citocinas, radicales libres, óxido nítrico y enzimas proteolíticas. Finalmente, otras células efectoras como eosinófilos, basófilos y mastocitos tienen receptores para la FcIgE que inducen respuestas inflamatorias alérgicas. Estas células producen mediadores y citocinas pro inflamatorias y son muy importantes en los procesos inflamatorios de las vías respiratorias. Al contrario que los receptores para FcIgG que en su mayoría son de baja afinidad, la mayor parte de las células con receptores para FcIgE expresan receptores de alta afinidad capaces de unir moléculas libres de IgE. Sin embargo, las células B, células de Langerhans, monocitos y eosinófilos expresan receptores de baja afinidad2. Eje Neuro – Endocrino – Inmunológico Los sistemas de comunicación interna del organismo (el sistema nervioso y el sistema endocrino) interaccionan entre si y modulan la fisiología del sistema inmune, que no sólo emplea los sistemas de comunicación intercelular que le son propios, sino también otros que participan en las respuestas al estrés. Programa de Doctorado en Fisiología 2003 – 2005 Natalia I. Wiereszen 10 Inmunidad en el Deporte Varios tipos de estrés fisiológico, incluyendo al ejercicio físico y el estrés emocional, influyen en la respuesta inmune4. Las hormonas de estrés (catecolaminas, cortisol y hormona de crecimiento) movilizan las reservas y aumentan la capacidad del individuo para afrontar las necesidades de una situación crítica2-4-12. Estas hormonas también modulan el número y actividad de los leucocitos circulantes, que pueden comprometer, la capacidad de respuesta del sistema inmune y predisponer a infecciones12. Los efectos del ejercicio sobre la respuesta inmune con mediados por circuitos neuro-endocrinos (hormonas de estrés) y por los circuitos paracrinoendocrinos del sistema inmune constituidos por las citocinas (Figura 3). Estas moléculas actúan como señales de emergencia del sistema inmune, integrando y coordinando la señalización local y sistémica durante las reacciones inmunes e inflamatorias. En general actúan localmente, pero algunas pueden producirse en cantidades suficientes para alcanzar una concentración en la circulación que le permite ejercer efectos sistémicos. Figura 3. Relación entre los sistemas nervioso, endocrino e inmune en la respuesta de estrés originada por el ejercicio En la interrelación entre los tres sistemas participan diversos mecanismos y mediadores: 9 La acción del sistema nervioso es desempeñada fundamentalmente por las terminaciones simpáticas, que estimulan tanto la respuesta endocrina como ciertas acciones de los órganos linfoides. 9 Los mediadores endocrinos incluyen las hormonas: de crecimiento (GH), la hipotalámica Programa de Doctorado en Fisiología 2003 – 2005 Natalia I. Wiereszen liberadora de ACTH (CRH), la 11 Inmunidad en el Deporte adenocorticotropa (ACTH), el cortisol, las β-endorfinas y las catecolaminas. 9 Por parte del sistema inmune intervienen mediadores pro inflamatorios producidos por macrófagos residentes en los tejidos y por los linfocitos T, así como mediadores antiinflamatorios que participan en la resolución de la inflamación en respuesta al ejercicio. Algunos de estos mediadores inmunes actúan a nivel sistémico estimulando la respuesta de fase aguda, además de intervenir sobre el sistema neuro-endocrino. Las moléculas mediadoras paracrinas y endocrinas que intervienen en las respuestas de estrés y procesos inflamatorios subsecuentes se muestran en la Tabla 2. Grupo de Mediadores Fuente de los Mediadores Mediadores Mediadores endocrinos de la Hipotálamo CRH, GH, ACTH, respuesta de estrés Hipófisis anterior β-endorfinas, cortisol, Corteza suprarrenal catecolaminas Mediadores paracrinos del Citocinas pro inflamatorias IL-1, IL-6, IL-8, TNFα, sistema inmune Macrófagos titulares IFNγ, IL-5 Linfocitos T Citicinas con efecto Diversas células inmunes y inmunosupresor y no inmunes IL-10, IL-1ra, TGFβ antiinflamatorio Tabla 2. Mediadores en la respuesta de estrés al ejercicio Programa de Doctorado en Fisiología 2003 – 2005 Natalia I. Wiereszen 12 Inmunidad en el Deporte El Sistema Inmune y el Ejercicio Físico El ejercicio conlleva una serie de demandas sobre el organismo que son dependientes de la forma, intensidad y duración del mismo y que, a su vez, tienen profundas repercusiones sobre la capacidad de respuesta inmune del deportista. De modo recíproco, la capacidad de respuesta inmune del deportista repercute sobre su salud y ésta, sobre su rendimiento. El estudio del efecto del entrenamiento sobre la inmunidad, requiere dos definiciones operativas: la cantidad y la intensidad incluida en las tandas de entrenamiento, y el nivel inicial de forma física de los sujetos estudiados. EL ejercicio intenso, especialmente el que requiere mayor proporción de contracciones excéntricas, induce respuestas inflamatorias transitorias en los músculos ejercitados microtraumatismos más musculares intensamente. y participa Esta en inflamación los procesos responde de a reparación, hipertrofia y angiogénesis muscular secundarios al ejercicio. Por tanto, la inflamación es un proceso esencial en la adaptación del músculo al ejercicio2-5-7-13. Sin embargo, no todas las consecuencias de la inflamación muscular son beneficiosas. La repetición de reacciones inflamatorias intensas, provocadas por cargas diarias excesivas de entrenamiento, puede provocar una afección inflamatoria local de carácter crónico o recurrente que produce dolores musculares y disminución del rendimiento físico. Ya que la intensidad de la respuesta inflamatoria local es proporcional al daño muscular provocado por el ejercicio, las cargas excesivas con componente excéntrico que provocan daño muscular, elevan la intensidad de la inflamación hasta un grado en el que se pueden tener repercusiones sistémicas en el organismo del deportista. Esta afectación sistémica se traduce en forma de respuesta de fase aguda a la inflamación, que cuando es intensa y mantenida a lo largo del tiempo, altera la capacidad inmune del deportista y puede conducir a situaciones de inmunosupresión, aumentando la susceptibilidad a infecciones y poniendo en riesgo su salud5-9. Efectos del Ejercicio sobre las Células y los distintos Mediadores del Sistema Inmune Es bien conocido que el ejercicio altera la distribución, el tráfico y las capacidades funcionales de los distintos tipos de células inmunes. Como también, causa alteraciones en los niveles locales y sistémicos de diversos mediadores moleculares1-2-5-9-13. Programa de Doctorado en Fisiología 2003 – 2005 Natalia I. Wiereszen 13 Inmunidad en el Deporte Leucocitos Circulantes El ejercicio provoca una leucocitosis transitoria, cuya magnitud está relacionada directamente con la intensidad del mismo, es más pronunciada en respuesta a ejercicios máximos, e inversamente con el nivel de forma física ya que es más acusada en sujetos sin entrenar que en sujetos entrenados. El valor de leucocitos aumenta hasta cuatro veces y puede mantenerse hasta 24 horas después del ejercicio1-2-4-5-9-12 -13. La leucocitosis afecta principalmente a los neutrófilos, aunque también lo hace sobre monocitos y linfocitos, especialmente sobre linfocitos B y células NK2-4. Estas respuestas a la actividad física disminuyen con el entrenamiento, posiblemente debido a la disminución de catecolaminas en respuesta al ejercicio. El ejercicio prolongado aumenta el recuento de neutrófilos, pero provoca una reducción en el número de linfocitos. Por otra parte, el ejercicio no tiene efecto a largo plazo sobre las cifras de leucocitos en reposo que, por lo tanto, son normales en los individuos entrenados2-4-5-11-13. Población Leucocitaria % de los Leucocitos Cifra Absoluta/µL Granulocitos 60-70 6000-9000 Neutrófilos >90 de los granulocitos 6000-9000 Eosinófilos 2-5 200-500 Basófilos 0-1 20 Agranulocitos 30-40 2000-400 Monocitos 2-10 300-1000 Linfocitos 20-25 2000-3000 Tabla 3. Poblaciones Leucocitarias Circulantes2 El ejercicio provoca alteraciones en el tráfico de los leucocitos de unos compartimientos corporales a otros (Tabla 3). Los leucocitos sanguíneos son un pequeño porcentaje del total de leucocitos maduros del organismo, el resto permanece en la microcirculación con baja perfusión en los pulmones, hígado y bazo2-5. El aumento del gasto cardíaco, con el subsiguiente aumento de la perfusión microvascular, provoca la movilización de estos leucocitos compartimentales. Por otro lado, el ejercicio estimula la liberación de leucocitos inmaduros de la médula ósea, que pueden distinguirse de los maduros por los antígenos de diferenciación celular expresados en sus membranas plasmáticas. Aunque el aumento de formas jóvenes de neutrófilos y monocitos, sólo se observan tras ejercicio prolongado. Programa de Doctorado en Fisiología 2003 – 2005 Natalia I. Wiereszen 14 Inmunidad en el Deporte El ejercicio de corta duración aumenta las cifras de linfocitos (Tabla 4), pero dicho aumento es menor al observado en las cifras de otros leucocitos como los neutrófilos2-4-5. Aunque la magnitud de la linfocitosis es proporcional a la intensidad del ejercicio, la duración de éste se relaciona inversamente con la linfocitosis, de tal forma que el ejercicio de larga duración provoca su disminución. Aunque tras carreras de larga distancia la cifra de linfocitos disminuye, en reposo se mantiene dentro de los límites de la normalidad, en los individuos que entrenan para prueba de fondo. Subpobl. Linfocitarias % de los Linfocitos Cifra Absoluta/µL Linfocitos T 60-75 1700-2500 CD4 60-70 de los Linfocitos T 800-2000 CD8 30-40 de los Linfocitos T 400-1200 Linfocitos B 5-15 100-450 Células NK 10-20 200-600 Tabla 4. Poblaciones y Subpoblaciones Linfocitarias Circulantes2 El recuento de linfocitos T aumenta hasta un 150% tras el ejercicio breve de intensidad máxima. Este aumento es mayor en los linfocitos CD8 que en los CD4, por lo que el cociente CD4/CD8 disminuye1-2-4-9. En el ejercicio intenso la concentración de linfocitos decrece y la duración de esta supresión depende de la intensidad y duración del ejercicio4. La cifra de Linfocitos B aumenta durante el ejercicio, pero retorna rápidamente a los niveles basales1-2-4-9. La cifra de las células NK aumenta entre un 150-300% en respuesta a ejercicio submáximo, máximo y submáximo prolongado, aunque puede disminuir tras ejercicio de resistencia intenso. Las células Nk tienen alta afinidad por los receptores β2-adrenérgicos, lo que explica porque aumenta significativamente su nivel en sangre durante los ejercicios intensos, cuando las concentraciones de adrenalina son elevadas11. Parece que las células NK son movilizadas a la circulación durante el ejercicio y posteriormente retiradas de ella, normalizándose sus tasas en menos de 24 horas tras cualquier tipo de ejercicio1-2-4-5-9-11-13. El número de monocitos en reposo es normal en atletas, pero aumenta rápidamente en respuesta al ejercicio de corta y/o larga duración, estando la magnitud del aumento relacionada con la forma física y la duración del ejercicio9-11. EL estrés físico aumenta los niveles de las hormonas de estrés en sangre, incluyendo catecolaminas, GH, β-endorfinas, ACTH y cortisol5. Las hormonas de Programa de Doctorado en Fisiología 2003 – 2005 Natalia I. Wiereszen 15 Inmunidad en el Deporte estrés son elementos importantes que participan en la regulación de los cambios en las cifras de poblaciones leucocitarias y subpoblaciones linfocitarias2-5-11-12. Las concentraciones de adrenalina y cortisol aumentan en relación a la capacidad individual y en función de la intensidad del ejercicio explicando las variaciones leucocitarias secundarias al mismo. La leucocitosis en ejercicios de corta duración es causada por el aumento de la concentración de adrenalina, sin embargo, en ejercicios mantenidos durante más de una hora, el cortisol actúa de manera sinérgica2-5-12. Al final de los ejercicios de larga duración, la disminución de cifras sería causada por el descenso de la concentración de adrenalina a pesar de que se mantenga elevada la de cortisol, hecho que, por otra parte, provoca el mantenimiento o descenso más lento de las cifras de leucocitos tras la finalización del ejercicio. Efectos del Ejercicio sobre la Inmunidad Inespecífica o Innata Células Fagocíticas La actividad fagocítica de macrófagos y neutrófilos aumenta inmediatamente tras la realización del ejercicio. Estas respuestas son menos marcadas en atletas de elite que en individuos no entrenados1-2-12. Por el contrario, la actividad fagocítica de los monocitos sanguíneos desciende tras el ejercicio máximo de corta duración1. Macrófagos La capacidad fagocítica de los macrófagos se mide por su capacidad para ingerir microesferas de látex, y su contenido lisosómico se estudia mediante técnicas de inmunohistoquímica. Durante la realización de ejercicio estas actividades aumentan en los macrófagos extraídos del tejido conectivo, aunque disminuyen en lo de sangre periférica y en los macrófagos alveolares pulmonares1-211-12 . Neutrófilos Existe una creciente evidencia que, tanto la práctica de ejercicio moderado como el intenso, está asociado con una prolongada mejora de la capacidad microbicida de los neutrófilos sanguíneos y macrófagos peritoneales9. La actividad microbicida de los neutrófilos se estima por su capacidad por su capacidad para medir compuestos oxidantes (H2O2, O2-)1-2-9-12. La capacidad oxidativa de los neutrófilos aumenta en respuesta al ejercicio, tanto en individuos entrenados como en desentrenados, siendo la magnitud del aumento mayor en estos últimos2-12. Asimismo, en situaciones de reposo, la actividad oxidativa de los Programa de Doctorado en Fisiología 2003 – 2005 Natalia I. Wiereszen 16 Inmunidad en el Deporte individuos entrenados esta disminuida con respecto a la de los individuos sedentarios2-12-13. Esta supresión crónica de la actividad oxidativa puede ser una adaptación para reducir la respuesta inflamatoria al daño tisular de baja intensidad, como el producido por el ejercicio en el tejido muscular, es decir, se trataría de un mecanismo de defensa natural. Eosinófilos, Basófilos y Mastocitos Estas células parecen no estar afectadas por los efectos del ejercicio1-2-9. Células Citotóxicas Espontáneas (NK) La intensidad del ejercicio es la responsable del grado de incremento del número de NK, aunque los ejercicios de más de una hora de duración ejercen una acción supresora1-4-5. Durante la recuperación posterior al ejercicio, el número de NK puede permanecer elevado o bien descender y continuar bajo de 21 a 24 horas1-10. La actividad de las NK puede aumentar más en los ejercicios de corta duración; no obstante, tras ejercicios intensos o agotadores y prolongados, la actividad de las NK se suprime1-10. El ejercicio tiene un efecto dual sobre la actividad citotóxica espontánea de las células NK. El ejercicio máximo o submáximo produce un aumento transitorio de la actividad NK que se mantiene unas tres horas y se sigue de una disminución de la misma tras el cese del ejercicio2. Estas alteraciones son paralelas a los efectos que tiene el ejercicio sobre la concentración de las células NK. Parece que en la fase de supresión intervienen el aumento de concentración de la prostaglandina plasmática y el aumento de monocitos en circulación2-9-11. Citocinas El ejercicio, cuando induce daño muscular, ya sea por microtraumas adaptativos, isquemia/hipoxia local, contusiones, o bien por el tipo de ejercicio desarrollado, se asocia 2-10 antiinflamatorias a elevación de los niveles de citocinas pro y . Cuando el ejercicio es ligero produce aumento de la concentración sérica de IL-6, pero no de TNFα ni de IL-1b1-2-4-5-13. En el ejercicio intenso la elevación de IL6 se correlaciona con la de la actividad de la creatin kinasa, que es un marcador indirecto del daño muscular2-5-9. Las citocinas se liberan en respuesta al ejercicio extremo de modo secuencial, del mismo modo que en la respuesta a estímulos sépticos. La concentración de IL-6 aumenta hasta cien veces inmediatamente después del ejercicio2. Programa de Doctorado en Fisiología 2003 – 2005 Natalia I. Wiereszen 17 Inmunidad en el Deporte La producción de IL-1 por los macrófagos aumenta durante y después del ejercicio prolongado. El nivel de IL-1 en reposo es mayor en los corredores de resistencia que en las personas desentrenadas1-10. Las citocinas antiinflamatorias están implicadas en la limitación espacial y la terminación temporal de la respuesta inflamatoria. A consecuencia del ejercicio se elevan pero con una cinética más retrasada que las pro inflamatorias2. El antagonista del receptor de IL-1 (IL-1ra) inhibe la actividad de la IL-1 bloqueando su receptor. El IL-1ra alcanza sus máximos niveles en una o dos horas después del ejercicio. La IL-10 es una citocina de gran potencial antiinflamatorio que suprime la síntesis de citocinas pro inflamatorias por macrófagos y linfocitos2. La producción de IL-2 se reduce durante el ejercicio, mientras que la actividad del interferón α aumenta durante una hora de ciclismo al 70% del VO2 máximo1-10. Efectos del Ejercicio sobre la Inmunidad Específica o Adquirida Linfocitos T Los ejercicios máximos de corta duración aumentan la población de células T en la circulación, principalmente en personas desentrenadas1. El número de células T citotóxicas/supresoras en la circulación aumenta considerablemente entre un 50 y 100% después del ejercicio de alta intensidad, mientras que las T 9 cooperadoras/inductoras de células B están relativamente inafectadas . Sin embargo este efecto es transitorio (alrededor de 30 minutos), los linfocitos de cada subpoblación salen a la circulación en gran número bajo la influencia del cortisol9. Durante el ejercicio prolongado, la respuesta es diversa, con un predominio del descenso de la población de células T1. La capacidad de proliferación de los linfocitos T refleja la intensidad de la respuesta de estas células frente a la estimulación antigénica, capacidad que es normal en reposo en los individuos entrenados. El ejercicio de corta duración (menos de una hora) no tiene efectos sobre las respuestas proliferativas de los linfocitos T, sin embargo, el ejercicio prolongado como la maratón si disminuye transitoriamente dicha respuesta a distintos estímulos. En animales de experimentación el ejercicio exhaustivo suprime las respuestas proliferativas de los linfocitos T, aunque este defecto es reducido a consecuencia del entrenamiento previo. Programa de Doctorado en Fisiología 2003 – 2005 Natalia I. Wiereszen 18 Inmunidad en el Deporte Linfocitos B Con el ejercicio aparece un pronunciado incremento de la población de células B con una rápida vuelta a los niveles basales1. Esta respuesta es mayor en personas desentrenadas en comparación con los deportistas1. El ejercicio tiene poco efecto sobre las concentraciones séricas de los distintos isotipos de anticuerpos. En reposo, la concentración de inmunoglobulinas de los deportistas es normal, ni los ejercicios intensos ni los máximos de corta duración o prolongados provocan cambios en el nivel sanguíneo de IgG, IgA, IgM e IgE1-2. No obstante, la producción in vitro de IgG, IgA e IgM por los linfocitos disminuyó cuando se extrajo una muestra de sangre tras 15 minutos de ciclismo1. A largo plazo y en deportistas de elite, la concentración de inmunoglobulinas puede disminuir durante el entrenamiento intensivo y la competición2. Por ejemplo, la IgA se restaura cuando el entrenamiento se reduce al final de la temporada, aunque los niveles en reposo y tras ejercicio disminuyen durante el entrenamiento intenso y la competición, encontrándose que los individuos con inmunosupresión asociada al ejercicio excesivo y, generalmente de resistencia, presentan una IgA anormalmente reducida (más del 50%)9 en sus secreciones corporales. Este fenómeno se ha asociado a un mayor riesgo de sufrir infecciones respiratorias de vías altas1-2-4-5-9-13. Programa de Doctorado en Fisiología 2003 – 2005 Natalia I. Wiereszen 19 Inmunidad en el Deporte Ventajas y Desventajas del Ejercicio sobre la Capacidad de Respuesta Inmune Efectos Positivos EL ejercicio moderado realizado en períodos largos de tiempo produce algunos efectos beneficiosos sobre la capacidad de respuesta inmune frente a la infección. A largo plazo induce un aumento sostenido de la actividad citotóxica espontánea de las células NK. El paseo diario reduce el número de días a la mitad con respecto a los individuos sedentarios5. La actividad física ligera estimula ciertas funciones inmunes, que por el contrario, son inhibidas por la actividad física extenuante. El efecto de la actividad citotóxica aumenta la resistencia a infecciones virales y contribuye a la eliminación de células preneoplásicas11. Un régimen regular de ejercicio moderado puede ayudar a los ancianos a mantener una buena calidad de vida, al contribuir a la preservación de la función inmune. Por lo tanto, el ejercicio moderado contribuiría a contrarrestar los efectos adversos del envejecimiento sobre el sistema inmune8. Efectos Negativos El ejercicio intenso y de larga duración, como es el deporte profesional de competición, produce un cuadro de respuesta de fase aguda, e incluso, una inmunosupresión que puede llegar a comprometer la salud del deportista y su rendimiento atlético. En los casos más severos, los atletas sufren, además de una disminución del rendimiento y una susceptibilidad aumentada a las infecciones, otros síntomas comunes al síndrome de respuesta inflamatoria sistémica como son la fatiga, la pérdida de peso y las alteraciones del estado anímico2-5-9-13. La inmunosupresión producida por ejercicios extremos es similar a la generada por el estrés físico severo como las situaciones de cirugía, traumatismos y quemaduras. En ellas, disminuye el número de linfocitos en sangre, y se reduce su capacidad proliferativa y citotóxica espontánea. Asimismo, se elevan los niveles séricos de citocinas pro-inflamatorias y anti-inflamatorias, además de producirse neutrofilia y alteración en la función de los neutrófilos. Los gérmenes patógenos, especialmente los virus del tracto respiratorio, pueden establecer infecciones durante esta inmunosupresión transitoria que se mantiene de 2 a 72 horas tras el ejercicio, aunque los efectos agudos se observan de 2 a 4 horas post-ejercicio2-5-9Programa de Doctorado en Fisiología 2003 – 2005 Natalia I. Wiereszen 20 Inmunidad en el Deporte 13 . Se ha sugerido que en los atletas de elite sobreentrenados, la mayor susceptibilidad a las infecciones puede deberse a una inmunosupresión más acusada y duradera provocada por la acumulación de un exceso de carga de entrenamiento. Otros factores como el estrés psíquico, la malnutrición o la pérdida de peso, pueden tener un efecto sinérgico sobre la inmunosupresión causada por el exceso de ejercicio. Si esta se combina con déficit en la higiene y con condiciones ambientales adversas, el riesgo de infección puede incrementarse. En conjunto, estos hallazgos proporcionan posibles mecanismos a las evidencias epidemiológicas de la mayor incidencia de enfermedades infecciosas en deportistas durante períodos de entrenamiento intensivo y alta competición. También el riesgo de sufrir infecciones de vías respiratorias altas se ha correlacionado con la disminución de los niveles de IgA, sobre todo en deportistas que practican ejercicios de resistencia. Programa de Doctorado en Fisiología 2003 – 2005 Natalia I. Wiereszen 21 Inmunidad en el Deporte Conclusiones El efecto de la actividad física sobre el sistema inmune está mediado por mecanismos que implican el eje neuro-endocrino-inmunológico, encuadrados dentro de las respuestas del sistema inmune a situaciones de estrés. Existe evidencia que el estrés físico induce a cambios en el sistema inmune. En esencia la respuesta inmune, especialmente la inmunidad innata, esta aumentada tanto en el estrés severo, moderado o leve, mientras que la respuesta inmune sólo esta suprimida después del estrés prolongado y severo. El ejercicio altera la función de varios componentes del sistema inmune, afecta tanto a mecanismos innatos como a mecanismos antígeno-específicos. Se elevan los niveles de citocinas pro y anti-inflamatorias, y se alteran tanto las concentraciones sanguíneas de las poblaciones leucocitarias como sus capacidades funcionales. Las poblaciones más afectadas por los efectos agudos del ejercicio son células no antígeno-específicas como las células citotóxicas espontáneas (NK), macrófagos y neutrófilos, características de las respuestas inmunes innatas. La respuesta del sistema inmune al ejercicio intenso generalmente es transitoria, pero se han descrito alteraciones inmunes positivas asociadas al ejercicio moderado durante largos períodos de tiempo. Asimismo, en respuesta a períodos prolongados de ejercicio intenso y entrenamiento mantenido, la función del sistema inmune también se altera a consecuencia de mecanismos sistémicos relacionados con la respuesta de fase aguda, que deprimen la función de los neutrófilos y ciertas clases de inmunoglobulinas en suero y secreciones. Programa de Doctorado en Fisiología 2003 – 2005 Natalia I. Wiereszen 22 Inmunidad en el Deporte Referencias Bibliográficas 1. Atko Viru. Análisis y Control del Rendimiento Deportivo. Human Kinetic Publishers, Inc. 2001. 2. A. Córdova Martínez y M. Álvarez de Mon. Inmunidad en el Deporte. Ed. Gymnos. 2001. 3. Guyton. Tratado de Fisiología Médica. Interamericana – McGraw Hill. 8º Edición (1991). 4. Pedersen, B.K. and col. The Immune System during Exposure to Extreme Physiologic Conditions. Int. J. Sports Med. 1994; 15:S116–S121. 5. Pedersen, B.K. and H. Bruunsgaard. How Physical Exercise Influences the Establishment of Infections. Sports Med. 1995; 19 (6): 393-400. 6. Northoff, H. and col. The Cytokine Response to Strenuous Exercise. Int. J. Sports Med. 1994; 15: S167-S171. 7. Nieman, D.C. and col. Cytokine Changes After a Marathon Race. J. Appl. Physiol. 2001; 91: 109-114. 8. Nieman, D.C. Exercise Immunology: Future Directions for Research Related to Athetes, Nutrition and the Elderly. J. Sports Med. 2000; 21 Supplement 1: S61-S68. 9. Nieman, D.C. Exercise, Infection and Immunity. It. J. Sports Med. 1994; 15: S131-S141. 10. Shephard, R.J., S. Rhind, P. Shek. Exercise and Training: Influences on Cytotoxicity, Interleukin-1, Interleukin-2 and Receptor Structures. Int. J. Sports Med. 1994; 15: S154-S166. 11. Woods, J., J. M. Davis, J. Smith and D.C. Nieman. Exercise and Cellular Innate Immune Function. Med. & Sci. In Sports & Exer. 1998; 57-66. 12. Rincón, E. Physiology and Biochemistry: Influence of Exercise on Phagocytosis. Int. J. Sports Med. 1994; 15: S172-S178. 13. Nieman, D.C. and B.K. Pedersen. Exercise and Immune Function. Sports Med. 1999; 27 (2): 73–80. 14. Rowbottom, D.G. and Green K.J. Acute Exercise effects on the Immune System. Med. Sci. Sports Exer. 2000; 32:S96–S405. 15. Nieman, D.C. and col. Immune Function in Female Elite Rowers and Nonathletes. J. Sports Med. 2000; 34: 181-187. Programa de Doctorado en Fisiología 2003 – 2005 Natalia I. Wiereszen 23 Inmunidad en el Deporte Programa de Doctorado en Fisiología 2003 – 2005 Natalia I. Wiereszen 24