Aspectos generales de la microbiología del
Anuncio

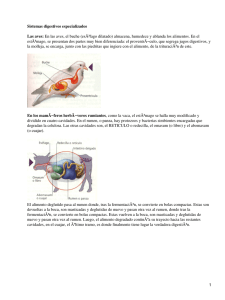
ASPECTOS GENERALES DE LA MICROBIOLOGÍA DEL RUMEN Grudsky P., Roberto y Arias B., José Luis. 1983. Monografías de Medicina Veterinaria, 5(2). *Laboratorio Biología Celular, Depto. Patología Veterinaria, Facultad Ciencias Agrarias, Veterinarias y Forestales, Universidad de Chile. www.produccion-animal.com.ar Volver a: Fisiología digestiva INTRODUCCIÓN El rumiante puede ser considerado como una fábrica fermentadora. Los vegetales que el animal ingiere, constituyen la materia prima para estas fermentaciones. Primariamente el proceso ocurre en la boca, con la particulación de este alimento por medio de la masticación, luego el material vegetal es deglutido y transportado al pre-estómago del rumiante llamado retículo rumen o en forma más simple rumen, lugar donde ocurre la fermentación. En el rumen habita una masiva comunidad de microorganismos, principalmente bacterias y protozoos, los que fermentan el material vegetal, entregando cómo productos principalmente ácidos grasos volátiles (AGV), metano y dióxido de carbono. Los AGV son removidos del rumen mediante el paso a circulación y son subsecuentemente usados por el rumiante como fuente primaria de energía y carbono. Los gases en tanto, son eliminados por la vía del eructo. La fermentación provee nutrientes y energía para el desarrollo y división de los microorganismos, éstos, junto al alimento no digerido (parcialmente fermentado o que escapó a la fermentación), son posteriormente removidos en forma más o menos continua del rumen por pasaje hacia las partes posteriores del tracto digestivo del rumiante. Los procesos digestivos que allí ocurren son similares a aquellos que se realizan en animales monogástricos e incluyen una digestión en abomasum e intestino delgado. La digestión de la masa microbial que ha salido del rumen, provee al rumiante de su mayor fuente de aminoácidos y vitaminas solubles. Los microorganismos no digeridos y los residuos alimenticios y microorganismos que habitan en el intestino grueso, salen del animal en las heces. A continuación se describirán las principales características de la población microbiana ruminal y se destacará la vital importancia que tiene la simbiosis microorganismos-rumiante. MICROAMBIENTE RUMINAL El rumen constituye un medio muy favorable para el desarrollo de determinados microorganismos y puede considerarse como un aparato de cultivo continuo y de gran eficacia pata el desarrollo de los microorganismos anaerobios. Existe una entrada relativamente constante de alimentos y una mezcla continua de estos, gracias a las contracciones ruminales, que ayudan a mantener a los microorganismos en contacto con la ingesta fresca o la comida rumiada, además, las condiciones de humedad son relativamente constantes y muy favorables para el desarrollo de numerosos microorganismos. De igual manera, la presión osmótica se mantiene próxima a la de la sangre, la temperatura va de 39°C a 41°C y el pH de 5.5 a 7, lo que es muy cercano al óptimo de numerosos sistemas enzimáticos. Cabe mencionar, que el pH es amortiguado por el paso de los AGV y amonio a circulación, por la entrada de grandes cantidades de saliva que contiene bicarbonato y fosfato y por la tendencia hacia el equilibrio iónico entre el contenido ruminal y el torrente sanguíneo. La fase gaseosa del contenido ruminal está constituida fundamentalmente por CO2 (65%), Metano (25%), Nitrógeno (7%) y trazas de Hidrógeno y Oxígeno. Bajo las condiciones descritas, una gran variedad de géneros bacterianos anaerobios y protozoos ciliados anaerobios, constituirán la población microbiana de mayor importancia y magnitud. Aproximadamente un 4% del volumen total del líquido ruminal corresponde a volumen microbiano, del cual un 50% corresponde a volumen bacteriano y el otro 50% a volumen, protozoario. En términos de actividad metabólica, a pesar de que en volumen los protozoos y las bacterias se encuentran en valores similares, la actividad metabólica de estos últimos es de mayor envergadura. MICROORGANISMOS RUMINALES Como se dijo anteriormente, son bacterias anaeróbicas y protozoos ciliados los microorganismos que se encuentran en mayor número, sin embargo, además se encuentran protozoos flagelados (especialmente en 1 de 13 Sitio Argentino de Producción Animal rumiantes jóvenes y antes de que se establezcan los protozoos ciliados), bacteriófagos y actualmente se han descrito hongos. Se han sugerido diversos criterios para caracterizar a un microorganismo como típico del rumen, esto se debe a que a veces resulta difícil determinar si el microorganismo es un habitante normal del rumen o bien si su presencia resulta de la ingestión del microorganismo por vía del alimento, agua u otro medio. Para ser considerado como típico del rumen, un microorganismo debe cumplir los siguientes requisitos: ♦ ser capaz de vivir anaeróbicamente ♦ tener un metabolismo compatible con las reacciones que ocurren en el rumen, es decir ser capaz de producir algunos de los productos finales que se encuentran en el rumen ♦ encontrarse en número no inferior a 1.000.000/ml de contenido ruminal (se excluyen los protozoos). ¿Qué impide una mayor variedad de microorganismos en el rumen? Aparte de las condiciones propias del ambiente ruminal, se han evidenciado ciertos mecanismos inhibitorios o líticos sobre formas microbianas no típicas del rumen. Enterobacterias como E. coli y Salmonellas se encuentran en pequeño número en el rumen, ya que son inhibidas por los AGV. Tampoco el Clostridium perfringens tipo D (patógeno) puede mantenerse en rumen, pero en caso de llegar a intestino delgado se multiplica rápidamente. También cierta actividad lítica sobre algunas especies bacterianas, se asocia a la presencia de bacteriófagos. Se ha observado que las poblaciones microbianas del rumen, tienen ciertos medios químicos que restringen el crecimiento de posibles competidores. Algunos microorganismos en el rumen producen toxinas y/o antibióticos, los cuales afectan otras formas microbianas. BACTERIAS DEL RUMEN El rumen contiene una gran variedad de bacterias, casi todas son anaerobias no esporuladoras, unas pocas especies son anaerobias facultativas y ocasionalmente se detectan bacterias anaerobias que forman esporas (Ej.: Oscillospira guillermondii). La densidad bacteriana va de 10.000.000.000 a 100.000.000.000/ml de contenido ruminal y se reconocen alrededor de 250 especies. Establecimiento en Rumiantes Jóvenes El desarrollo de una flora bacteriana en los rumiantes jóvenes, semejante a la encontrada en animales adultos, empieza a una edad muy temprana. Este desarrollo se ve afectado por el tipo de dieta suministrada y en cierta medida por el grado de aislamiento del animal joven de los rumiantes que albergan microorganismos típicos del animal adulto. Los rumiantes jóvenes adquieren su población microbial por contacto oral con animales de mayor edad y también por inhalación de bacterias temporalmente suspendidas en el aire. Estudios realizados en terneros indican que existe un gran desarrollo de lactobacilos en las tres primeras semanas de vida, población que tiende a declinar después de la tercera semana, alcanzando a los tres a cuatro meses, niveles similares a los encontrados en animales adultos. Los microorganismos celuloIíticos pueden encontrarse en niveles altos, incluso en la primera semana de vida, alcanzando a las tres semanas valores similares a los de los rumiantes adultos. Otros estudios, llevados a cabo en rumiantes jóvenes bajo condiciones relativamente normales, muestran que las bacterias cultivables predominantes de los terneros de 1 a 3 semanas de edad eran muy diferentes a las del ganado vacuno adulto, a las 6 semanas, muchos grupos de bacterias típicas de los animales adultos, se encontraban entre las bacterias predominantes, pero permanecían aún varios grupos no encontrados en los adultos. Entre 9 y 13 semanas de edad, las bacterias aisladas eran principalmente típicas de las de los animales adultos mantenidos con dietas similares. El aislamiento estricto de los terneros puede aplazar el establecimiento de una flora bacteriana, pero sin efectos muy drásticos para el animal. Distribución La mayoría de las especies bacterianas han sido encontradas sólo en el rumen, sin embargo, muchas especies similares o idénticas se han encontrado presentes en otros habitats como el aparato gastrointestinal de mamíferos no rumiantes, sedimentos orgánicos de aguas naturales y Iodos de aguas residuales. Por Ej.: Cepas del género Butyrivibrio han sido aisladas de heces humanas, de conejo y de caballo, también se han aislado Ruminococcus celulolíticos del ciego. del conejo. Especies del género Selenomonas cultivadas de la boca humana y del rumen, son similares. Se ha observado además, que Bacteroides humanos s (orales y fecales) están relacionados muy estrechamente con Bacteroides ruminícola. 2 de 13 Sitio Argentino de Producción Animal Factores que afectan el número y la naturaleza de las poblaciones bacterianas Hay numerosos factores que pueden afectar la población bacteriana ruminal, tanto cuantitativa como cualitativamente. 1) Efecto de la Alimentación-Dieta: La selectividad en el pastoreo, velocidad de ingestión del alimento, fertilidad del terreno, localización geográfica y clima (T°, luz, etc.), son todos factores que pueden influenciar la calidad y cantidad de los nutrientes presentados a los microorganismos bacterianos del rumen. Las variaciones cíclicas que presentan las poblaciones bacterianas del rumen a lo largo del día, son afectadas por la frecuencia de la alimentación. La variación en el número de microorganismos es menor cuando los animales son alimentados frecuentemente, el peak en el número ocurre 4 a 8 hrs. después de la alimentación, dependiendo de la naturaleza de la dieta. Con respecto al efecto de la composición de la dieta, las observaciones no son del todo concluyentes. Dietas muy ricas en carbohidratos solubles producen una depresión de los microorganismos celulolíticos. Valores adecuados de proteínas en la dieta, implican usualmente un aumento del número de bacterias en comparación con dietas bajas en proteínas. Normalmente, el número de bacterias/gr. del contenido ruminal tiende a ser mayor en animales alimentados con pastos verdes, en comparación con aquellos alimentados con raciones secas. 2) Efecto de la Defaunación: En rumiantes defaunados (sin protozoos ciliados) por métodos como por Ej.: la administración de sulfato cúprico, ácidos o sustancias químicas, como el dioctilsulfosuccinato de sodio o dimetridazole, la población bacteriana se encuentra muy incrementada. I d e n ti fi c a c i ó n . Existe una variedad de criterios utilizados para la identificación de las bacterias ruminales. La Morfología (forma-tamaño) se usa para identificar numerosas especies, sin embargo muchas especies son similares en la identificación en base a este criterio solamente, debido a que la mayoría de las especies son cocáceas. Los cocus y bacilos cortos miden de 0.4 a 1 μm de diámetro por 1 a 3 μ m de largo. Otras formas bacterianas que se encuentran son espiroquetas, rosetas, tetracocus, sarcinas, etc. Otro inconveniente para identificar sólo por morfología, es el hecho de que existen cepas que modifican su forma dependiendo del medio en que se encuentran. (Fig. 1 a y b). La motilidad es otra característica que permite identificar ciertas especies. En resumen, varias especies y cepas pueden identificarse en base a su forma, tamaño o apéndices móviles, sin embargo muchas otras deben ser identificadas en base a otras características. Tinción: La tinción Gram es también usada como método de identificación, pero más útil resulta el uso de varios procesos de tinción y la utilización del microscopio electrónico de transmisión (TEM) y el de barrido (SEM). La identificación también puede efectuarse parcialmente por estudios de tipo metabólico, como por ejemplo: principal(es) sustrato(s) atacado(s), fuentes de energía utilizada, nutrientes requeridos para desarrollarse en medios de cultivo, etc. Sin embargo, existe mucha superposición entre las diferentes especies al utilizar estos métodos. Otras técnicas, más complejas y no muy empleadas, son la detección de enzimas y test serológicos (anticuerpos fluorescentes). Clasificación. Las diversas clasificaciones actuales, se basan principalmente en los sustratos utilizados y en los productos principales de la fermentación. 3 de 13 Sitio Argentino de Producción Animal 1. Bacterias Celulolíticas: Estas bacterias tienen la habilidad bioquímica de producir celulasas, enzimas que pueden hidrolizar la celulosa. También pueden utilizar celobiosa (disacárido) y otros carbohidratos. Especies celulolíticas de importancia son: Bacteroides succinogenes, Ruminococcus flavefaciens, Ruminococcus albus, Clostridium loch headii y Cillobacterium cellulosolvens. 2. Bacterias Hemicelulolíticas: La hemicelulosa difiere de la celulosa en que aquella contiene tanto pentosas como hexosas y usualmente contiene ácidos urónicos. La hemicelulosa es un importante constituyente de las plantas. Los organismos que son capaces de hidrolizar celulosa, habitualmente también pueden utilizar hemicelulosa. Sin embargo, algunas especies hemicelulolíticas no pueden utilizar la celulosa. Dentro de las especies que digieren hemicelulosa tenemos: Butyrivibrio fibrisolvens, Lachnospira multíparus y Bacteroides ruminícola. 3. Bacterias aminolíticas: Todas las bacterias celulolíticas son también capaces de digerir almidón, sin embargo algunos microorganismos amilolíticos no pueden utilizar celulosa. Especies importantes que digieren almidón son: Bacteroides amylophilus, Succinomona amylolítica, Butyrividrio fibrisolvens, Lachnospira multíparus y Bacteroides ruminícola. 4. Bacterias que utilizan Azúcares: La mayoría de las bacterias que son capaces de utilizar polisacáridos, son también capaces de utilizar disacáridos o monosacáridos. 5. Bacterias que utilizan Ácidos: Un gran número de bacterias utilizan ácido láctico, no obstante este ácido no está presente en cantidades apreciables en el rumen, excepto en condiciones anormales. Otras bacterias utilizan ácido succínico, málico y fumárico, así omo también utilizan ácido fórmico y ácido acético, pero probablemente no como fuentes primarias de energía. También el ácido oxálico es descompuesto por bacterias ruminales. Ejemplo de bacterias que utilizan lactato: Veillonella gazogenes, Veillonella alacalescens, Propionibacterium sp., Desulphovibrio y Selenomona lactilytica. 6. Bacterias Proteolíticas: Cierto número de bacterias ruminales utilizan aminoácidos como fuente primaria de obtención de energía. Ej.: Bacteroides amylophílus, Clostridium sporogenes y Bacilus licheniformis, son tres especies que tienen reconocida capacidad proteolítica. 7. Bacterias productoras de Amonio: Algunas especies bacterianas producen amonio a partir de distintas fuentes. Ej.: Bacteroides ruminícola, Selenomona ruminantium, Peptostreptococcus elsdenii y algunos Butyrivibrios. 8. Bacterias que producen Metano: Las principales son Methanobacterium ruminantium y Methanobacterium formicum, de menor importancia son, Methanobacterium sohngenii, Methanobacterium suboxydans y Methanosarcina sp. 9. Bacterias Lipolíticas: Existen bacterias que utilizan glicerol y lo hidrolizan. Otros microorganismos hidrogenan ácidos grasos insaturados y algunos metabolizan ácidos grasos de cadena larga a cetonas. Ejemplos de bacterias ¡¡políticas son: Selenomona ruminantium y Anaerovibrio lypolítico. 10. Bacterias sintetizadoras de Vitaminas: Especial importancia tienen las bacterias sintetizadoras de vitaminas del complejo B. Ej.: Selenomona ruminantium. TABLA I.- Características de las principales bacterias ruminales cultivadas in vitro oo Go Medidas, Especie Morfología Mo Función importante Algunas fuentes μm de Energía Bacilar a - 0.3 - 0.4 x 1) Bacteroides succinogenes - Celulolítica G, C, a cocoide 1-2 2) Ruminococcus flavefaciens Cocos ± 0.8 - 1 - Digestión de fibra g, c, x 3) Ruminococcus albus Cocos ± 0.8 - 2 - Digestión de fibra g, c, x Bacilar a - 0.9 - 1.6 x 4) Bacteroides amylophilus - Amilolítica G, x, a cocoide 1.6 - 4 Cocoide a - 1.0 - 1.5 x 5) Succinomonas amylolítica + Amilolítica G, A bacilar 2.2 - 3 - Fermentadora de 6) Veillonella alcalescons Cocos - 0.3 - 0.6 L lactato 7) Methanobacterium Bacilos + 0.7 - 0.8a - Producción de CO2, H2, F, ruminantium curvados 1.8 metano AGV 0.4 x 1.2 8) Anaerovibrio lypolítica Bacilos + Lipolítica G Ly, (fructosa) - 3.6 9) Peptostreptococcus elsdenii o - Fermentadora de Cocos - 1.2 - 2.4 G, L, gly Megasphaera elsdenii lactato 4 de 13 ooo Productos Finales S, A, F A, S, F, H A, E, F, H A, S, F, L S A, P. H CH4 A, P, B, H CO2 Sitio Argentino de Producción Animal Bacilos curvados Cocoide a bacilar 0.7 - 1.7 x 2.6 1 x 7 - 12 o 2,3 x 7 - 0.3 - 0.5 x 4-7 + 0.4 - 0.6 x -2-4 0.5 - 0.7 x 1-2 15) Butyrivibrio fi brisolvens Bacilos curvados 16) Butyrivibrio alactacidigens 10) Clostridium lochheadii Bacilos 11) Clostridium longisporum Bacilos 12) Borrelia sp. 13) Lachnospira multiparus Espiroqueta - Celulolítica G, C, A + ? G, C + ? G, L, gly A, E, F, L, H2, CO2 + Digieren pectina G, a, (pectinas) + Celulolítica G, C - 0.4 - 0.6 x 2-5 + Amilolítico a muy adaptable G, c, x, a Bacilos curvados 0.5 - 1 x 1.5 - 8 + Amilolítico a muy adaptable G, X, A Cocoide a bacilar Bacilos curvados - 0.8 - 1 x 0.8 - 30 - 0.8 - 2.5 x 2-7 - Muy adaptado G, x, a A, S, F, L, + Muy adaptado G, a, I, gly A, P, L, B, CO2 19) Selenomonas lactilytica Bacilos curvados 0.4 - 0.6 x 1.8 - 3 20) Succinivibrio dextrinesolvens Espirales - 0.3 - 0.5 x 1 - 1.5 Cocos + 0.7 - 0.9 Cocoide a bacilar + 0.4 - 0.7 x 0.7 - 1.5 23) Barcina bakeri SSS Cocos 1-4 24) Lactobacillus sp. Bacilo + 0.7 - 1 x 1-6 14) Cilliobacterium cellulosolvens 17) Bacteroides ruminícola 18) Selenomona ruminantium 21) Streptococcus bovis 22) Eubacterium ruminantium CLAVE: + Fermentadora de lactato a muy adaptada - Fermentadoras de dextranos A, S, L - Amilolítico a varios G, A L - Azúcares, xilosa G, x B, F, L - ? - Muy adaptado en condiciones ácidas G, a GRAM M°: Motilidad oo: G (glucosa); C (celulosa); X (xilosa); A (almidón) - L (lactato); Gly (glicerol); F (formiato) - X: Todos las cepas lo fermentan - x: La mayoría de las cepas lo fermentan - x: Algunas cepas lo fermentan - G, L, Gly, a G G°: ooo: B, F, H, L, CO2 A (acetato); B (butirato); P (propionato); L (I actato) S (succinato); E (etanol); F (formiato) PROTOZOOS DEL RUMEN Sólo se encuentran protozoos ciliados. Los protozoos flagelados que se han descrito en rumiantes corresponden a formas móviles de hongos. La densidad de los protozoos va de 200.000 a 2.000.000/ml de contenido ruminal y sus medidas se encuentran en un rango que va de los 38 a 195 µm de largo por 15 a 109 µm de ancho. Se han descrito alrededor de 40 especies de protozoos. Clasificación. Clase: Ciliados Sub-clase: Holotricos (tienen su cuerpo cubierto de cilios) (Fig. 2a y b). 5 de 13 Sitio Argentino de Producción Animal Género y Especie: Isotricha intestinalis Isotricha prostoma Dasytricha ruminantium Blepharocerys bovis (raro) Charon equi (raro, también encontrado en caballo) Charon ventriculi (raro) Buetschilla (raro) también se conoce como Buetschlia parva. Sub-clase: Spirotricos. Orden: Entodiniomorfos (comúnmente llamados Oligotricos, tienen solamente un penacho de cilios en el polo anterior). (Fig. 3a y b). Género y Especie: Caloscolex (Camello) Diplodinium Sub-Especie: Diplodinium dentatum Diplodinium posterevesiculatum Diplodinium crista-galli Diplodinium psitaceum Diplodinium elongatum Diplodinium polygonale Eudiplodinium neglectum (sin. Eremoplastron bovis) Eudiplodinium magii (sin. Metadinium medium) Eudiplodinium medium (sin. Metadinium tauricum) Eudiplodinium bursa (sin. Diplodinium neglectum) Eudiplodinium affine Eudiplodinium rostratum Polyplastron multivesiculatum Elystroplastron bubali 6 de 13 Sitio Argentino de Producción Animal Ostracodinium obtusun Ostracodinium gracile Ostracodinium uninucliatum Ostracodinium dentatum Enoploplastron triloricatum Entodinium caudatum Entodinium bursa Entodinium simplex Entodinium longinucleatum Entodinium minimum Entodinium rostratum Entodinium elongatum Entodinium ogimotoi Entodinium bubalum Entodinium fujitai Entodinium tsunodai Epidinium ecaudatum Ophryoscolex purknynei Ophryoscolex caudatus Ophryoscolex inerminis Opisthotrichum (encontrada en antílopes africanos) Establecimiento en Rumiantes Jóvenes En contraste a la situación que ocurre con las bacterias ruminales, el establecimiento de la población ciliada en el rumiante joven requiere del contacto con los animales de mayor edad, que contengan las poblaciones protozoarias típicas. Lo anterior podría deberse en parte, a que no existe una fase resistente en el ciclo de vida de los protozoos, por lo cual no se producen formas resistentes que permanezcan viables cuando se expongan al medio o a otras condiciones adversas durante períodos largos. Además, el contenido ruminal de los animales jóvenes tiende a ser ácido bajo la alimentación láctea o de concentrado de iniciación, situación que previene el establecimiento de la microfauna. Se ha observado que manteniendo un pH adecuado, los protozoos empiezan a establecerse alrededor de las primeras tres semanas de vida, alcanzando niveles adultos a las 6 semanas y más concretamente entre los 3 y 6 meses de edad. Los Entodiniomorfos (Entodinium seguido de Diplodinium) son los primeros en establecerse, a continuación lo hacen los Holotricos. Distribución Los ciliados del rumen parecen encontrarse sólo en los rumiantes, de forma que en contraste con algunas especies bacterianas del rumen, no pueden obtenerse de otras fuentes naturales más que de las ingestas del rumen o materiales tales como saliva, recién contaminada con ingestas del rumen. Sin embargo existen excepciones, protozoos ciliados del género Ophrioscolex han sido encontrados en primates y Charen es comúnmente encontrado en caballos. Factores que afectan el número y naturaleza de la población protozoaria ciliada Uno de los factores más importantes es la dieta, la cual se refleja principalmente en el pH del contenido ruminal, siendo este pH el factor "crítico". De hecho, la población ciliada disminuye e incluso a veces desaparece, cuando valores de pH 5,5 o menores permanecen durante gran parte del día. También existen variaciones cíclicas diurnas en la densidad de los ciliados, produciéndose un aumento de esta, cuando la frecuencia de suministros de una ración es mayor. Al entregar la ración una sola vez al día, se produce una gran baja de pH con la consiguiente baja del número de ciliados. La inclusión de ciertos aditivos químicos tales como clortetraciclina, tilosina, aureomicina, penicilina o dietilbestrol en la dieta, pueden causar un incremento de los protozoos ruminales. Parásitos de los Protozoos del Rumen Se han descrito hongos, Sphaerita, hoari y Sphaerita sp., parasitando protozoos entodiniomorfos, principalmente Eremoplastron bovis y Entodinium simplex de rumiantes domésticos en la India. También se ha observado al Sagittospora cameroni parasitando a Eudiplodinium magii y otros entodiniomorfos de la cabra en la 7 de 13 Sitio Argentino de Producción Animal India. Además, organismos que podrían ser Rickettsias han sido observadas en el macronúcleo de Isotricha intestinalis. METABOLISMO MICROBIANO RUMINAL METABOLISMO DE LOS CARBOHIDRATOS EN EL RUMEN Generalidades El rumiante provee los nutrientes que permiten el crecimiento y desarrollo de los microorganismos ruminales. Todo el C, N, P, S y elementos trazas necesarios son aportados por el alimento que consume el animal. También el rumiante contribuye sustancialmente a mantener las condiciones fisicoquímicas apropiadas para este medio fermentativo, por ejemplo; contribuye al control de la temperatura y pH y al control de la dinámica de reciclaje de los compuestos en el rumen. En respuesta a la entrega de este excelente hábitat, los microorganismos proveen de actividades y productos que son esenciales para el animal. El principal aporte son las celulasas microbianas. La celulosa es la más importante fuente de carbono y energía en la dieta del rumiante, pero el animal por sí mismo no produce las enzimas necesarias para digerir la celulosa. Como los microorganismos usan celulosa y otros carbohidratos presentes en las plantas como sus propias fuentes de energía y carbono, ellos aportan gran cantidad de productos (AGV y otros), los cuales son usados por el animal como fuente de carbono y energía. La planta contiene un 75% de un carbohidrato u otro, la cantidad y distribución dependen de la edad del vegetal, factores del medio y agronómicos y de la especie vegetal en cuestión. Los carbohidratos que se encuentran en los tejidos vegetales son, en su gran mayoría, aptos para ser fermentados en el rumen. Estos son principalmente polisacáridos -celulosa, hemicelulosa, pectinas y almidón- y cantidades menores de una gran variedad de compuestos carbohidratados y sus derivados -glucosa, sucrosa, sacarosa, propilenglicol, xilosa, celobiosa, arabinosa, galactosa, manosa, lactato, glucurunato, piruvato, succinato, citrato, etc.-. De estos carbohidratos, la celulosa es lejos la más abundante, sólo en unas pocas plantas hay una apreciable concentración de disacáridos como la sucrosa. Degradación de los Carbohidratos en el Rumen El catabolismo de los polisacáridos implica la degradación extracelular hasta monosacáridos u oligosacáridos de cadena corta, en tanto que el catabolismo intracelular supone la hidrólisis o demolición fosforilativa de los polisacáridos a monosacáridos y ulterior catabolismo del piruvato producido por glicólisis hasta AGV, lactato, succinato, formiato, etanol, CO2, metano e hidrógeno (Fig. 4). Figura 4.- Vías del metabolismo de los carbohidratos en el rumen. La habilidad de un microorganismo ruminal para fermentar algún carbohidrato específico es dependiente de la presencia de la enzima requerida para poder utilizar este carbohidrato. Numerosas enzimas han sido aisladas a 8 de 13 Sitio Argentino de Producción Animal partir de varias especies bacterianas del rumen, entre ellas, hemicelulasas, celulasas, amilasas, isomaltodextrinasas, alfa galactosidasas, sucrasas, fosforilasas, isomaltasas, etc. Existen algunas enzimas hidrolíticas extracelulares di fusibles, que ejercen su acción a cierta distancia de las colonias bacterianas que las produjeron. La mayoría de las bacterias, utilizan uno o más tipos de los principales carbohidratos dietéticos de los rumiantes como fuente de energía para su crecimiento. Las que no utilizan estos carbohidratos, utilizan los productos carbohidratados más simples de aquellos o los principales productos finales del metabolismo. Por lo tanto es corriente que exista una considerable superposición de funciones entre las diversas especies bacterianas (también entre especies bacterianas y protozoarias). Otro punto de interés al respecto, es el grado de versatilidad que presentan las bacterias en cuanto a los compuestos utilizados como fuente de energía. Por ejemplo, el Bacteroides amylophilus obtiene energía únicamente del almidón, sin embargo, varias de las especies más numerosas e importantes son muy versátiles, es el caso de Butyrivibrio y especialmente Selenomonas, que fermentan muchos carbohidratos e incluso algunas cepas además fermentan glicerol y lactato. Formación de Productos Finales El metabolismo de las hexosas hasta piruvato por la microflora ruminal, se realiza principalmente por la vía glicolítica, mientras que el de las pentosas presentes en las hemicelulosas ocurre vía síntesis de hexosas por la ruta ciclo de las pentosas. Del punto de vista del rumiante, los productos finales más importantes son los AGV, los que son absorbidos a través de la pared ruminal a una velocidad similar a su velocidad de producción. Alrededor de un 80% de la energía fermentada en el rumen es convertida en AGV, el resto es perdida como calor y metano. Las vías terminales que intervienen en la formación de los productos finales de la fermentación de los carbohidratos son muy complejas y entrañan distintos mecanismos de reacción, los cuales conducen desde el piruvato o el fosfoenelpiruvato (productos de la glicólisis), al CO2, metano y ácidos acético, propiónico, butírico y valeriánico. El acetato y el CO2 son producidos, a partir de los carbohidratos, por numerosas especies individuales de microorganismos del rumen, mientras que otras especies los producen a partir de los ácidos propiónico, butírico y valeriánico. Sin embargo, muchas de estas especies individuales también tienen uno o más productos finales tales como succinato, lactato, fermiato, hidrógeno y etanol, que no son productos finales de la fermentación. Es evidente que estos compuestos, con la posible excepción del etanol, son metabolizados ulteriormente por otras especies que dan lugar a los productos finales típicos (Fig. 4). La mayor parte del acetato del rumen se produce vía piruvato por dos reacciones generales: 1) descarboxilación oxidativa, que suministra acetato, CO2 e hidrógeno. 2) una reacción que suministra acetato y formiato. En este último caso, el formiato es metabolizado a dióxido de carbono e hidrógeno dentro de las células que lo producen o excretado al líquido ruminal, donde otros microorganismos pueden catabolizar el formiato con producción de CO2 e hidrógeno (Fig. 4). Se conocen dos vías generales de producción de propionato en el rumen: descarboxilación del succinato (éste es el principal precursor del propionato) y reducción del lactato, vía acrilato. Los mayores productores de propionato en el rumen son Selenomonas ruminantium y Megasphaera elsdenii (Fig. 4). La producción del butirato procede de forma principal vía producción de Acetil CoA a partir del piruvato y del acetato extracelular (Fig. 4). El metano del rumen se produce principalmente por reducción de CO2 mediante hidrógeno gaseoso. El formiato también es un muy buen sustrato para la producción de metano por parte de las bacterias metanogénicas. La contribución de los AGV al metabolismo del rumiante es considerable. Así, el acetato producido en la fermentación ruminal proporciona el mayor porcentaje de la energía requerida por el rumiante, constituye alrededor del 90% de la concentración de AGV en la sangre periférica. Con respecto al propionato ruminal, sólo un 5%.de este es metabolizado por el epitelio ruminal a lactato, de esta manera la mayor parte del propionato, es transportado al hígado, donde junto a los aminoácidos no esenciales es' la principal fuente de glucosa para el rumiante (vía gluconeogénesis). El 50% del butirato es metabolizado por el epitelio ruminal. La pequeña cantidad que pasa a sangre periférica es metabolizada en el hígado, dando principalmente Acetil CoA, este compuesto regula la utilización de energía por el rumiante, dado que por su concentración puede limitar la oxidación del propionato hacia Acetil CoA, reservando propionato para gluconeogénesis. Los Protozoos y el Metabolismo de los Carbohidratos en el Rumen Holotricos: Estos protozoos metabolizan preferentemente carbohidratos solubles como fuente de energía y carbono. 9 de 13 Sitio Argentino de Producción Animal Los Isotricha fermentan rápidamente glucosa, fructosa, sucrosa, rafinosa, pectina y además ingieren pequeños gránulos de almidón intacto. Sus principales productos son el almacenamiento de amilopectina (como polisacárido de reserva), acetato, butirato, lactato, CO2, hidrógeno y trazas de propionato. Los Dasitricha son más versátiles en cuanto al metabolismo de los carbohidratos y aparecen fermentando celobiosa, galactosa, maltosa, rafinosa, glucosa, fructosa y sucrosa a gran velocidad y en forma más lenta pectina. Como resultado de este metabolismo, ellos acumulan amilopectina y producen acetato, butirato, lactato, CO2 e hidrógeno. Además, bajo ciertas condiciones aparecen produciendo formiato y metano. Particularmente son Holotricos los que almacenan polisacáridos de reserva en forma de amilopectina. Durante la fermentación de los carbohidratos exógenos, este material es catabolizado a los mismos productos que los carbohidratos exógenos, pero a menor velocidad. Además, estos protozoos producen mayor cantidad de lactato y menos AGV que los Entodiniomorfos. Algunos protozoos ingieren tal cantidad de azúcares, que en un momento dado estallan. Al respecto hay dos teorías: Una de ellas afirma que el protozoo sintetiza una cantidad máxima de amilopectina y que sería la acumulación de lactato la causa de que la célula protozoaria reventara. La segunda teoría sugiere que existiría una síntesis incontrolada de amilopectina y esto llevaría a la ruptura de la célula. Entodiniomorfos: Utilizan muy poco los carbohidratos solubles, sólo en caso de no disponer de alimento particulado. Principalmente ingieren alimento particulado como células bacterianas y vegetales, siendo el almidón su principal fuente de carbono y energía. Sus productos de fermentación son acetato, butirato, CO2, hidrógeno y pequeñas cantidades de lactato y propionato. Algunas especies son capaces de fermentar celulosa, pero esta actividad celulolítica es indirecta, por cuanto se debe a la presencia y actividad de bacterias celulolíticas en el endoplasma protozoario (por esta característica suele denominarse a estos protozoos como microrumiantes). METABOLISMO DEL NITRÓGENO Existe una gran variedad de compuestos nitrogenados disponibles para los microorganismos presentes en el rumen. Dichos compuestos incluyen: proteínas de diversa naturaleza, las que varían en solubilidad y contenido aminoacídico, nucleoproteínas que contienen gran variedad de bases púricas y pirimídicas, varios compuestos nitrogenados no proteicos pomo péptidos, aminoácidos, amoníaco, amidas, aminas, aminas volátiles, sales de amonio, nitritos y nitratos, así como también compuestos como la urea (que además entra al rumen a través del torrente circulatorio y de la saliva) y biuret, los cuales pueden ser incluidos en forma de aditivos en la ración de los rumiantes. El esquema general del metabolismo nitrogenado microbiano es el siguiente: 1. Las proteinasas y peptidasas hidrolizan las proteínas a péptidos y aminoácidos libres. 2. Los aminoácidos se utilizan directamente para la síntesis de proteínas y otros constituyentes celulares microbianos, tales como los constituyentes de la pared celular y ácidos nucleicos. 3. Los aminoácidos son también catabolizados a AGV y otros ácidos, CO2 y amoníaco. 4. La urea es hidrolizada a amoníaco por la potente actividad ureásica. 5. Los compuestos tales como los nitratos son reducidos a amoníaco. 6. El amoníaco se utiliza en la síntesis de los componentes celulares microbianos como las proteínas y otros. La mayoría del Nitrógeno proteico de la dieta se convierte en proteína microbiana, también se excreta como urea (por orina) o proteína fecal no digerida. Relativamente poca proteína dietética escapa al metabolismo de los microorganismos del rumen. La proteína microbial subsecuentemente abandona el rumen para ser digerida en el abomasum e intestino delgado. Esta proteína microbial es muy digestible para el animal y tiene un alto valor biológico. Hidrólisis Proteica Normalmente, los productos intermediarios de la hidrólisis proteica (aminoácidos y péptidos), están presentes en grandes cantidades sólo durante un corto tiempo después de la ingestión de la proteína, lo que implica una rápida degradación que produce amoníaco o su fijación rápida en el interior de las células microbianas. Parece ser que la actividad proteolítica está influenciada principalmente por el pH, siendo los valores óptimos entre 6 y 7. Sólo han sido parcialmente caracterizadas las enzimas proteolíticas, las que se encuentran tanto en bacterias como en protozoos del rumen. Estas enzimas parecen ser principalmente intracelulares o ligadas a la superficie de las células microbianas, ya que en el líquido ruminal hay poca actividad proteolítica. El nivel de actividad proteolítica parece ser independiente de la dieta consumida. Esto se explica quizás parcialmente al menos, por el hecho de que las cepas de muchas especies de bacterias del rumen que tienen 10 de 13 Sitio Argentino de Producción Animal propiedades proteolíticas, utilizan diversos carbohidratos como fuentes energéticas, por lo tanto la mayoría de ellas pueden estar presentes en gran parte de los regímenes dietéticos. Aunque muchas especies bacterianas son proteolíticas, ninguna de las numerosas especies estudiadas hasta ahora son particularmente activas a este respecto, la mayoría requiere carbohidratos para obtener energía y sólo muy pocas utilizan aminoácidos como fuente energética para el crecimiento. Catabolismo de los Aminoácidos Lo realizan tanto bacterias como protozoos. Una parte relativamente grande de los aminoácidos dietéticos son catabolizados a productos finales antes de ser utilizados en reacciones biosintéticas. Los esqueletos carbonados de los aminoácidos son fermentados a sus ácidos derivados. Ácidos grasos a partir de aminoácidos alifáticos y ácidos aromáticos a partir de aminoácidos aromáticos. Los productos de fermentación de aminoácidos de cadena ramificada son ácidos grasos de cadena ramificada, con un carbono menos que el aminoácidos padre. En la producción de amoníaco a partir de los aminoácidos o los hidrolizados proteicos, sólo están implicadas unas pocas especies bacterianas entre las predominantes en los rumiantes adultos, Bacteroides ruminícola parece ser la más activa entre las especies dominantes usuales, también son activas ciertas Selenomonas, Peptostreptococcus elsdenii y cepas del género Butyrivibrio. Catabolismo de otros compuestos nitrogenados Los ácidos nucleicos y sus precursores púricos o pirimídicos son constituyentes comunes de las dietas de los rumiantes, muy poco se sabe de su catabolismo en el rumen, sin embargo, la demolición de las purinas entrega como productos principales amoníaco, CO2 y acetato. La urea entra continuamente en el rumen a través de la pared de éste, procedente del torrente circulatorio y a través de la saliva, si no lo hace como constituyente de la dieta y es una importante fuente de nitrógeno para el crecimiento y la síntesis de proteínas por parte de las bacterias del rumen. La urea se degrada rápidamente a amoníaco y CO2 por la enzima ureasa del contenido ruminal y es el amoníaco el compuesto nitrogenado utilizado realmente en la síntesis de la proteína bacteriana. El nitrato de la dieta es reducido por las bacterias del rumen a amoníaco, con nitrito como intermediario. El hidrógeno, formiato, succinato, lactato, malato, citrato, manitol y glucosa, son eficaces dadores de ión hidrógeno para la reducción del nitrato, disminuyendo con su reducción la producción de metano. Compuestos nitrogenados en el crecimiento microbiano Los principales compuestos nitrogenados extracelulares utilizados en la síntesis de la proteína microbiana ruminal y otros constituyentes celulares, son el amoníaco, aminoácidos y péptidos. Se ha observado que los péptidos son incorporados más eficazmente que los aminoácidos libres a la célula bacteriana. La mayor parte de los aminoácidos libres no pueden atravesar la pared de la célula bacteriana, en cambio péptidos que contienen entre 4 a 20 o más residuos de aminoácidos se utilizan rápidamente, al igual que el amoníaco. La mayor parte del carbono de aminoácidos como glutámico y aspártico, son más bien metabolizados a AGV que incorporados a los aminoácidos de la proteína microbiana. Un número importante de especies utiliza eficazmente aminoácidos exógenos libres como fuente de nitrógeno y carbono, sin embargo, el carbono peptídico exógeno y el nitrógeno peptídico, se convierten más eficazmente en proteína bacteriana que el carbono y nitrógeno de los aminoácidos exógenos libres. Los péptidos entran como tales a la célula bacteriana y después son hidrolizados a aminoácidos antes que se produzca su ulterior metabolismo. Numerosos aminoácidos preformados se catabolizan con producción de amoníaco y éste es utilizado como la principal fuente de nitrógeno. El amoníaco es esencial para un número importante de especies bacterianas ruminales. Incluso cuando las dietas contienen aminoácidos y péptidos en cantidades suficientes para toda la síntesis proteica microbiana y cantidades adecuadas de carbohidratos como fuente de energía, una cantidad considerable de los aminoácidos es catabolizada a amoníaco, CO2 y ácidos. La mayoría de las bacterias del rumen pueden sintetizar los constituyentes celulares principales, incluyendo a las proteínas, utilizando el nitrógeno amoniacal y fuentes carbonadas exógenas, tales como ciertos AGV y ácidos orgánicos, CO2 y carbohidratos. (Ver Figura 5 y Tabla Il). Los Protozoos ciliados y el Metabolismo Nitrogenado Algunas especies protozoarias hidrolizan proteínas. Ej.: Entodinium caudatum produce liberación de aminoácidos y péptidos y produce amoníaco, también Ophryoscelex produce amoníaco a partir de proteínas, sin 11 de 13 Sitio Argentino de Producción Animal embargo, este amoníaco no se produce por desaminación de los aminoácidos (el caso de las bacterias), sino por hidrólisis de los grupos amida. Tabla II.- Uso de amoníaco y aminoácidos como principal fuente de nitrógeno para las bacterias ruminales. Especies Bacteroides succinogenes B. amylophilus B. ruminicola Ruminococcus flavefaciens R. albus Butyrivibrio fibrisolvens Megasphaera eisdenii Selenomonas ruminantium Streptococcus bovis Methanobacterium ruminantium Fuente Nitrogenada Amoníaco Aminoácidos + + + + + + + + + + + + + - Wolín, 1979 Figura 5. Metabolismo de las proteínas en el rumen. (De Wolín, 1979). Los ciliados no utilizan amoníaco como fuente para biosíntesis de proteínas, como las bacterias, sino que utilizan aminoácidos de origen vegetal o bacteriano, por lo tanto, bacterias y proteínas del alimento son la principal fuente de nitrógeno para el crecimiento protozoario, lo anterior implica que gran parte del nitrógeno metabolizado por los ciliados entra a la célula como grandes moléculas proteicas. Muchos protozoos, particularmente Entodiniomorfos, ingieren bacterias, digieren sus proteínas a aminoácidos y resintetizan proteína protozoaria a partir de estos aminoácidos, Ej.: Isotricha es capaz de ingerir 3.000 bacterias (E. coli) por hora. Algunos protozoos también son capaces de utilizar aminoácidos libres exógenos, como fuente de nitrógeno. La proteína protozoaria es de mejor digestibilidad que la bacteriana, pero de similar valor biológico. Existiendo protozoos, es posible que una cantidad menor de proteínas de los alimentos se convierta en compuestos nitrogenados, tales como los presentes en los ácidos nucleicos y compuestos de la pared celular, que son relativamente abundantes en las bacterias y que probablemente no son utilizadas de manera eficaz por el rumiante. Se estima, a pesar de que el número y clase de protozoos ciliados en el rumen varía mucho, que podrían ser responsables de alrededor del 20% del requerimiento proteico y de los productos de fermentación ácida que quedan a disposición del rumiante. 12 de 13 Sitio Argentino de Producción Animal METABOLISMO DE LOS LÍPIDOS Los microorganismos del rumen tienen un gran efecto sobre los lípidos de la dieta. Los glicéridos y los fosfolípidos son hidrolizados y el glicerol es fermentado principalmente a propionato. Los ácidos grasos de la dieta y los liberados por lipólisis en el rumen no se degradan, sin embargo, los ácidos grasos insaturados tales como los ácidos linoléico, linolénico y oleico, son hidrogenados extensamente por las poblaciones bacterianas y protozoarias del rumen. Los Entodiniomorfos, constituyen un grupo muy importante en relación al metabolismo de los lípidos, debido a que ingieren material particulado de origen vegetal, incluidos los cloroplastos y pueden eventualmente proteger a los ácidos grasos insaturados, de la hidrogenación que ocurre en el rumen. Los ácidos grasos de cadena larga de los lípidos microbiales son también usados por medio de la digestión intestinal de las células microbianas, los ácidos grasas son directamente incorporados en los lípidos del animal. La hidrogenación de los lípidos de la dieta que ocurre en el rumen, tiene consecuencias importantes para la composición de las grasas de los tejidos del rumiante. Las grasas somáticas y láctea del rumiante, tienden a tener mayor cantidad de ácidos grasos saturados, que las de los animales no rumiantes. Vitaminas Después de haberse desarrollado la fermentación en el animal joven, el rumiante no tiene ningún requerimiento dietético para las vitaminas del complejo B o la vitamina K, debido a que los microorganismos del rumen sintetizan todas estas vitaminas. Estas son liberadas cuando los microorganismos son digeridos por el animal. También se encuentran en gran concentración en el fluido ruminal. Son más importantes las especies bacterianas que las protozoarias en relación a la síntesis de vitaminas en el rumen. Las especies bacterianas más numerosas del rumen contribuyen bastante a la síntesis de vitaminas del complejo B. Por Ej.: Selenomonas ruminantium tiene una habilidad particularmente grande para sintetizar vitamina B12 y factores relacionados con dicha vitamina. Muchas poblaciones bacterianas requieren vitaminas del complejo B para su crecimiento. La vitamina más requerida por las cepas ruminales estudiadas, es la Biotina, muchas necesitan Ac. p-amino benzoico y otros microorganismos, en menor número, requieren vitamina B12, Ac. fólico, riboflavina o tiamina. Los rumiantes pueden sufrir indirectamente deficiencias de vitaminas del complejo B, si es que la síntesis de vitaminas no toma lugar en el rumen. Ej.: cuando los animales ingieren una dieta deficiente en Cobalto, falla la síntesis de vitamina B12 por parte de los microorganismos y el animal sufre una enfermedad por deficiencia de vitamina B12. En cuanto a la vitamina C, esta no es requerida por las bacterias y de hecho es destruida por las bacterias del rumen, esta vitamina es sintetizada por el animal. Con respecto al Caroteno y vitamina A, no hay evidencias que demuestren que son sintetizadas en el rumen, sin embargo, hay cierto grado de destrucción de estos compuestos cuando son aportados por la dieta. BIBLIOGRAFÍA SELECCIONADA BRYANT, M.P_ (1981). Microbiología del Rumen. In: Fisiología de los Animales Domésticos. Dukes, H.H. y Swenson, M.J. Tomo I Ed. Aguilar, México. COLEMAN, G.S. (1980). Rumen Ciliate Protozoa. Adv. Parasitol 18: 121-173. CHURCH, D.C. (1976). Digestive Physiology and Nutrition of Ruminants. Vol. 1, 20 Ed., O & B Books, Corvallis, Oregon. DOUGHERTY, R.W. (1965). Physiology of Digestion in the Ruminant. Butterworth, Washington. HUNGATE, R.E. (1966). The Rumen and Its Microbes. Academic Press. N.Y. U.S.A. McDONALD, I.W.; Warner, A.C.I. (1975). Digestion and Metabolism in the Ruminant. Univ. New England Publ. Unit., Armidale U.S.A. PHILLIPSON, A.T. (1970). Physiology of Digestion and Metabolism in the Ruminant. Oriel Press, Newcastle upon Tyne. U.K. RUCKEBUSCH, Y.; THIVEND, P. (1980). Digestive Physiology and Metabolism in Ruminants. AVI Publi., Westport, Conn. U.S.A. WOLIN, M.J. (1979). The Rumen Fermentation: A Model for Microbial Interactions in Anaerobic Ecosystems. Adv. Microbial Ecol. 3: 49-77. Volver a: Fisiología digestiva 13 de 13