- Ninguna Categoria
Los tiempos precámbricos
Anuncio
La vida en el pasado y la escuela Fundamentación Se calcula que han existido unas 700.000.000 especies en nuestro planeta. ¿Cómo se llega a esta cifra? En la actualidad, los científicos estiman la biodiversidad entre 10 y 30 millones de especies (de las cuales sólo cerca de dos millones están clasificadas). También se sabe que los restos fósiles, único testimonio de la vida pasada, se hicieron abundantes a partir de los últimos 1.000 millones de años, con una tasa de recambio de especies de 700.000 cada millón de años. Sin embargo, sólo se conocen unas 700.000 especies fósiles, lo que representa una pequeñísima fracción de la biodiversidad que realmente existió, aproximadamente el 0,001%. Ahora bien, pese a esta pequeñísima fracción de la vida pasada, todavía es posible trazar un panorama claro y completo a partir del examen de los fósiles, tarea que compete a la paleontología o ciencia de las cosas antiguas. A los chicos de edad escolar les fascinan los fósiles, en particular los dinosaurios pero no mucho menos otros organismos del pasado, muchos de ellos gigantescos, y la visita a los museos y sitios de excavación constituye otra meta central en el estudio de los fósiles. Por todas las razones esgrimidas, y para realizar un aprendizaje significativo e interactivo de una temática tan interesante, se ofrece este curso para captar las principales característias y cambios de la historia de la vida en la Tierra, con un énfasis en los yacimientos paleontológicos y del patrimonio de nuestro país. Síntesis El presente curso que abarca el estudio de los organismos del pasado y el patrimonio paleontológico se ha dividido en una introducción y siete módulos: En la Introducción general se analiza la clasificación de los fósiles y los procesos de fosilización. En el módulo 1 se presenta un estudio general sobre los fósiles. El módulo 2 analiza la edad de la tierra, la geocronología y la datación, los cambios paleogeográficos y paleoclimáticos, En el módulo 3 se analizan las principales características de los tiempos precámbricos y se discute sobre el origen de la vida. En el módulo 4 se explican las principales características de la era Paleozoica. En el módulo 5 se analizan las características de los fósiles de la era Mesozoica con énfasis en los dinosaurios y los reptiles marinos. En el módulo 6 se analizan las características de los fósiles de la era Cenozoica, con una introducción al origen y evolución humanas. . Marco curricular Como se señala desde el currículo de educación primaria, “si bien por lo general, los alumnos/as de estas edades pueden imaginar que la Tierra no siempre ha sido igual, no todos tienen conciencia de la magnitud de los cambios que han ocurrido y que seguirán ocurriendo, ni de los tiempos implicados en esos cambios. Por otra parte, los cambios violentos (terremotos, vulcanismo, maremotos) son tomados como catástrofes y sus conocimientos sobre los mismos son aislados y no suelen establecer relaciones entre unos y otros. A partir del estudio de la estructura de la Tierra y de los cambios visibles y actuales que se producen en ella se propone ofrecer herramientas para abordar el estudio de su historia y sus cambios. Estudiar la historia de la Tierra implica manejar magnitudes de tiempo muy alejadas de lo cotidiano e imaginar paisajes notablemente diferentes de los actuales, contrariando la percepción de estabilidad respecto de la fisonomía general del planeta. Para ello es necesario que los alumnos/as se aproximen a la noción de tiempo geológico y se retrotraigan miles de millones de años. Teniendo en cuenta que esto puede representar un obstáculo, se propone ayudarlos a imaginar semejantes tiempos mediante la noción de magnitudes características. Trabajar sobre las magnitudes características significa comparar el orden de magnitud de los tiempos en que se desarrollan distintos procesos (minutos, horas, meses) y establecer relaciones entre ellos. Por ejemplo, podrán comparar primero procesos más conocidos por los alumnos/as, que puedan medirse en meses como la duración de las vacaciones o el tiempo que tarda la luna en volver a comenzar su ciclo; en años, tales como el tiempo promedio de la vida humana o del movimiento de traslación de la Tierra. Estos contenidos se podrán articular con los del subnúcleo “Las relaciones entre los seres vivos”, pues los alumnos/as podrán reinterpretar la actual diversidad de ambientes y de organismos, y las relaciones que se establecen entre ellos, recurriendo a conocimientos sobre los cambios que ocurrieron a lo largo de la historia de la Tierra. En este nuevo contexto, podrán trabajar la noción de fósil e interpretar mapas filogenéticos sencillos para aproximar a los alumnos/as a la idea del origen común de los seres vivos y de que unos organismos están emparentados con otros. Se podrá retomar la noción de extinción de las especies, por ejemplo, estableciendo relaciones entre especies extinguidas y actuales Módulo 1. Introducción: los fósiles Objetivos • Determinar la importancia de los fósiles como testimonios del pasado de la Tierra • Analizar los principios fundamentales de los estudios paleontológicos • Comparar y clasificar los procesos de fosilización • Determinar los biocronos y acmés de diferentes grupos de organismos • Analizar la relación entre fósiles y biodiversidad pasada • Evaluar la información que proporcionan los fósiles • Relevar la importancia de los fósiles guía para la correlación • Explicar los principios de la exploración paleontológica Contenidos Cambios evolutivos y fósiles Paleontología y su estudio Los procesos de fosilización Rocas que son fósiles Biocrono y acmé Fósiles vivientes Fósiles guía y fósiles faciales El paleoADN La tafonomía Cambios evolutivos y vida en la Tierra Muchos cambios evolutivos tienen lugar lo suficientemente rápido como para ser estudiados directamente y manipulados experimentalmente. La cría de plantas y animales por los agricultores, y la evolución de la resistencia a los pesticidas son buenos ejemplos de evolución rápida y de corto plazo. Otros cambios como la aparición de nuevas especies y los linajes evolutivos, usualmente toman lugar en encuadres de tiempo mucho más largos. Para comprender los patrones de largo plazo del cambio evolutivo que trazaremos a lo largo de la Parte Cinco y Seis de este libro, debemos pensar en marcos de tiempo que abarcan muchos millones de años y debemos imaginar sucesos y condiciones muy diferentes a los actuales. La Tierra del pasado distante representa para nosotros un planeta foráneo habitado por extraños organismos. Los continentes no se hallan donde se encuentran ahora, y los climas eran, a veces, drásticamente diferentes a los actuales. Casi todas las rocas antiguas sufrieron los procesos exógenos de erosión y la acción de las altas temperaturas y presiones que las transforma (metamorfismo), por lo que resulta muy difícil hallar rocas muy antiguas. Sin embargo, hacia 1985, el geólogo estadounidense Samuel Bowring, del Instituto Tecnológico de Massachusetts, exploró un área de los Estados Unidos cuyas rocas fueron datadas en 3.960 millones de años (m.a.), mientras que poco después, los científicos de la Universidad Nacional de Australia dirigidos por el geólogo William Compston, encontraron otras rocas todavía más antiguas, de cerca de4.100 m.a. Estas dataciones sugieren que la vida no pudo haber comenzado en el planeta antes de haber transcurrido 4.200 m.a., ya que no existían antes rocas sólidas. En 1996, Steve Mojzsis, de la Universidad de California, dio a oconocer las evidencias más antiguas de testimonios de vida del planeta, una mezcla de hidrocarburos y apatita (fosfatos), probablemente formado por la acción de células en rocas descubiertas en la isla Akilia, en el sur de Groenlandia Occidental, que de no menos de 3.850 m.a. La suma de estas evidencias indica que durante la etapa de la historia terrestre que va de los 4.300 a los 3.900 m.a. resulta fundamental, pues en ella tuvo lugar la transformación de los compuestos simples (dióxido de carbono, agua, metano, sulfuro de hidrógeno) en otros más complejos que forman parte de los seres vivos (aminoácidos, azúcares, lípidos). En 1987, William Schopf y Bonnie M. Packer, de la Universidad de California, hallaron las primeras células fósiles del Arqueano Temprano, en el oeste de Australia, de entre 3.300 y 3.500 m.a. Estos eran, por entonces, los fósiles más antiguos conocidos. En la década de 1990, se descubrieron restos celulares todavía más antiguos, de 3.650 m.a, denominadas Eobacterium, en Sudáfrica. Ambos depósitos provenían de de ambientes litorales, no profundos. Recreación de Eobacterium isolatum, la primera bacteria. Fósiles y Paleontología La Paleontología es la ciencia que estudia los restos de organismos del pasado o su actividad, conservados en rocas de millones de años y que varían según el terreno estudiado (su localidad y época de formación). Los estratos más antiguos contienen fósiles que no se parecen a ningún ser vivo actual y para los cuales es difícil definir un parentesco. Otros fósiles no existen en el presente, aunque se parecen bastante a algunos organismos actuales. E incluso pueden hallarse como fósiles algunos organismos vivientes. Diversos especialistas en la geografía, el clima, el suelo y los ecosistemas del pasado brindan información adicional para interpretar y reconstruir su aspecto e indicar la forma de vida de estos organismos y sus relaciones con el entorno. Para realizar todos estos estudios, los científicos requieren un arsenal de instrumentos y técnicas. los fósiles–restos preservados de organismos antiguos o de su actividad–nos pueden brindar mucha información acerca de la forma del cuerpo, o morfología de un organismo que vivió hace mucho tiempo, y también de cómo y dónde vivía. Pero para comprender los patrones de los cambios evolutivos, debemos entender también cómo cambió la vida a lo largo del tiempo. La historia de la Tierra está ampliamente registrada en sus rocas. No podemos señalar las edades de las rocas sólo observándolas, aunque sí podemos determinar las edades relativas de una respecto de otra. La primera persona que formalmente reconoció que eso podría hacerse fue el naturalista danés del siglo XVII Niels Stenen (obispo de Titiópolis y nautalista, latinizado como Steno), Steno comprendió que en las rocas sedimentarias (formadas por la acumulación de granos en el fondo de los cuerpos de agua) que no habían sido perturbadas, las capas, o estratos más viejos, yacen en el fondo y que, sucesivamente, los estratos más altos son progresivamente más jóvenes. Steno postuló una serie de principios fundamentales para la Paleontología. 1. El Principio de Estratificación horizontal. En una pila de estratos de sedimentos, los que primero se depositan son (y contienen los fósiles) más antiguos. 2. El Principio de Similitud suficiente. Un fósil se parece a todo organismos emparentado con él. Así, si se hallan dientes de tiburones fósiles, la similitud morfológica es indicio de un parentesco. 3. El Principio de Cementación. Todo fósil es más antiguo que el estrato que lo contiene o lo cubre. Los geólogos, particularmente el científico inglés del siglo dieciocho, William Smith a continuación combinaron la visión de Steno con su observación de los fósiles contenidos dentro de las rocas sedimentarias. Concluyeron que: Los fósiles de organismos similares son encontrados en lugares ampliamente separados de la Tierra. Ciertos organismos se encuentran siempre en rocas más jóvenes que otros. Los organismos hallados en los estratos más altos y recientes, eran más similares a los organismos modernos que aquellos encontrados en los estratos más bajos y antiguos. Cráneo de tiranosaurio. Los procesos de fosilización Cuando un organismo muere y cae al lecho marino o a un pantano, comienzan a operar los procesos de descomposición y desintegración que pueden hacer desaparecer en poco tiempo todo resto del organismo original. Por ejemplo, la erosión puede disolver o fragmentar un resto, y la actividad de otros organismos, como los escarabajos y los hongos, puede desarticularlos y descomponerlos. La Tafonomía es la disciplina que estudia los procesos biológicos, geológicos y fisicoquímicos que tienen lugar en la transformación de un cadáver en fósil. Compara la forma de preservación de los fósiles con lo que sucede realmente con los cadáveres de animales en distintos ambientes (muchos son rápidamente destruidos mientras que otros logran enterrarse y preservarse de alguna manera). Por otra parte, los organismos comúnmente fosilizados son criaturas marinas, como caracoles, almejas, corales, erizos de mar, etc., y se conservan en aguas someras, de plataforma, donde las condiciones ambientales resultan óptimas para que actúen los procesos de fosilización. En el ambiente aeroterrestre, la preservación es más difícil y se requieren condiciones especiales de deposición, de baja energía (la sedimentación predomina sobre la erosión), como la que tiene lugar en pantanos y lagos. Por otra parte, en la selva o en el desierto, las probabilidades de que un resto quede enterrado y permanezca más o menos protegido de la destrucción por microorganismos resulta sumamente baja. Queda claro entonces que la formación de un fósil, o proceso de fosilización, ocurre raramente. Existen dos tipos principales de procesos de fosilización: los de compresión (en una pila de sedimentos) y los de relleno. En la compresión, cuando se conservan tanto las partes duras como las blandas, en medios especiales, se habla de momificación, y en el caso del relleno, de preservación total o in toto. Continuando con la secuencia, los restos blandos desaparecen y se conservan solo las partes duras. De hecho, casi todos los fósiles que conocemos se conservan como partes duras (huesos, dientes, caparazones, etc., que contienen sustancias como fosfatos y carbonatos de calcio). Pero también, cuando los restos son reemplazados por procesos químicos y físicos, se tienen las carbonizaciones (en el caso de las compresiones), y las recristalizaciones (la transformación de un resto desde un estado cristalino de un mineral, como la calcita, en otro, como la aragonita) o petrificaciones (la transformación de las sales originales en otras circulantes en el medio, por ejemplo, carbonato de calcio por sílice) en el caso de los rellenos. En el otro extremo, raramente se conservan partes blandas (piel, músculos, etc.) en ámbar, hielo, alquitrán o condiciones especiales de preservación (por ejemplo, ambientes anaerobios o sin oxígeno que impiden la acción bacteriana). Cuando se pierden las estructuras o partes de los fósiles (sean las originales o las que ya se convirtieron en piedra), todavía existe la posibilidad de la formación de fósiles. Se habla de impresiones o improntas (por presión de la columna sedimentaria que elimina los restos orgánicos) o cementaciones o moldes (por el relleno sedimentario). En ambos casos persiste sólo la forma de los organismos originales. Pero también se consideran fósiles los restos de actividad de los organismos, como como las icnitas (trayectos de locomoción y pisadas fósiles), los coprolitos (restos de excrementos) y los gastrolitos (contenidos estomacales). Rocas que son fósiles La era Paleozoica señala la formación de abundantes rocas sedimentarias de origen biológico, como las coquinas, constituidas por caparazones de moluscos y otros organismos que depositaron carbonato de calcio, las diatomitas, formadas por arenas silíceas de diatomeas y las radiolaritas, constituidas por sílice que depositaron los radiolarios. Todos estos tipos de rocas se usan para la fabricación de objetos, adornos y revestimientos. La formación de estos caparazones y esqueletos en diferentes organismos, tanto caracoles, como algas y protozoos, fue posible gracias a la creciente acumulación y migración superficial de iones como el sodio, potasio, magnesio, fosfato, sulfato y cloro, que sólo pudo durante la era Paleozoica en los extensos mares de plataforma. Por esta razón, como los animales pudieron formar partes duras, al eón se conoce como Fanerozoico (del griego faneros, cubierta, y zoicos, de vida). Radiolarita. Biocrono y acmé La mayoría de los grupos de organismos que conocemos actualmente, o que vivieron alguna vez en el planeta experimentan períodos de origen y diversificación inicial, de radiación adaptativa o abundancia y riqueza evolutiva, y de extinción o declinación y desaparición. La representación gráfica de la distribución vertical o temporal de las especies o de los grupos o taxones en la columna geológica se conoce como biocrono. Si se produce la extinción final, como en el caso de los trilobites, se habla de biocrono cerrado, y si el grupo todavía persiste, como en el caso de los merostomados (cangrejos bayoneta) se habla de biocrono abierto. Por su parte, se conoce como acmé el período de mayor expansión geográfica y la diversidad de un grupo determinado. Por ejemplo, los trilobites fueron más abundantes durante el Cámbrico y el Ordovícico que en otros períodos posteriores de la era Paleozoica, mientras que los reptiles marinos alcanzaron su máxima expansión durante el Jurásico. Biocrono completo de los grupos de trilobites. Observen que durante el Cámbrico y en parte el Ordovícico, los dos primeros períodos de la era Paleozoica, es cuando se alcanza su acmé o másima expansión. Se trata de un biocrono cerrado porque a final del Pércmio se extinguen completamente sus últimos representantes. Fósiles y biodiversidad Se calcula que en nuestro planeta existieron unas 700 millones de especies. Algunas surgieron en diferentes momentos (evolucionaron), otras desaparecieron progresivamente o por catátrofes, como los dinosaurios al final del Cretácico, y algunas más permanecieron sin cambios, los fósiles vivientes, como el celacanto y el cangrejo bayoneta. En la actualidad, los científicos estiman que la biodiversidad ronda los 10 a 20 millones de especies (de las cuales únicamente un millón y medio está clasificada). Se conoce asismismo que los restos fósiles, único testimonio de la vida pasada, se hicieron abun dantes a partir de los últimos 1.000 millones de años y se calcula la tasa de recambbio de especies en 700.000 cada millón de años. Sin embargo, sólo se conocen unas 700.000 especies fósiles, lo que representa una pequeñísima fracción de la biodiversidad que realmente existió, de aproximadamente el 0,001 %. Fósiles vivientes La comparación de las características de los organismos a través del tiempo ha cobrado gran interés en los estudios evolutivos. En especial, llama poderosamente la atención que algunas especies se conservan prácticamente sin cambios desde su aparición en el registro fósil, como los tiburones y las cucarachas, mientras que otras han experimentado una rápida evolución y radiación adaptativa, como los insectos y los caballos. Ejemplos extremos de persistencia en el tiempo son el celacanto (Latimeria chalumnae), extraño pez de aguas profundas, y la neopilina (Neopilina galatheae), molusco bentónico sumamente primitivo. El celacanto, que se creía completamente extinto desde hace más de 100 milones de años, fue redescubierto por la ciencia en 1938; actualmente se sabe que vive en el océano Índico, cerca de las islas Comores. La neopilina, por su parte, es uno de los escasos representantes de los monoplacóforos, clase fósil de moluscos muy primitivos, con numerosas branquias pares, que era abundante hace unos 500 millones de años. Las especies que pertenecen a grupos completamente extintos, como el celacanto y la neopilina, se conocen como fósiles vivientes. Otros ejemplos de fósiles vivientes son la tuátara, extraño reptil de Nueva Zelandia; el ginkgo, árbol que ya no se conoce en estado silvestre sino únicamente en cultivos, es el único representante de un antiguo y exitoso linaje de la era Mesozoica; el ornitorrinco, extraño mamífero australiano ovíparo y con pico de pato, y el cangrejo bayoneta, un pariente marino de las arañas que ha cobrado gran importancia médica pues de él se obtiene una sustancia coagulante que permite detectar bacterias mortales en vacunas, medicamentos y dispositivos médicos. Celacanto de las Comores (Latimeria chalumnae). En las Molucas existe una segunda especie recientemente descubierta. Nautilus pompilius (una de las cinco especies sobrevivientes de este antiguo y en otras épocas exitoso grupo, los nautiloideos). Limulus polyphemus, el cangrejo bayoneta de la costa pacífica de Norteamérica. En la costa atlántica existe una segunda especie. Neopilina galatheae, una de las 25 especies sobrevivientes de galeroconchos, un antiguo grupo de moluscos casi totalmente extinto. Hojas de Ginkgo biloba, único representante actual de los ginkgófitos. ¿Qué información proporcionan los fósiles? Los fósiles son una valiosa fuente de información. A partir de su estudio se puede conocer: Los fósiles son los únicos “documentos” que poseemos sobre los organismos que poblaron la Tierra en otras épocas. Analizándolos puede conocerse la anatomía del organismo, su modo de vida, sus relaciones con otros organismos, el ambiente en que vivió, su distribución geográfica, etc. El ambiente en que se formó la roca que lo contiene. Cada organismo ocupa un hábitat determinado, que puede ser marino o continental, de clima cálido o frío, húmedo o seco. El sedimento que enterró al organismo se transformó en la roca que hoy contiene su fósil, por eso el ambiente en que vivió suele ser aquel en que se formó la roca. Esto permitirá saber si una zona, hoy continental, fue en otra época oceánica o el clima que existía. Cuándo se formó la roca que lo contiene. Como los seres vivos que han poblado la Tierra han cambiado de unos períodos a otros, si conocemos cuándo vivió un determinado organismo podremos saber la edad de la roca en la que fosilizó, o lo que es lo mismo, datarla. No todos los fósiles son igualmente útiles para datar una roca. Los mejores reciben el nombre de fósiles característicos o fósiles guía y deben reunir tres características: – Haber vivido durante un período de tiempo geológicamente corto, de manera que permitan precisar la edad de la roca. Las especies que existieron durante extensos períodos de tiempo resultan poco útiles para datar. – Haber tenido amplia distribución geográfica, para que su interés no sea solo local. – Ser abundantes en las rocas sedimentarias, para que resulten útiles con frecuencia. Además de los fósiles, las rocas poseen ciertas características determinadas por el lugar y las condiciones en que se formaron. Así, un sedimento transportado por el viento tendrá un tamaño de grano muy homogéneo, mientras que otro transportado por un glaciar tendrá materiales de tamaños muy diferentes. Se denomina facies al conjunto de características litológicas (forma, tamaño y disposición de los granos, composición mineralógica, etc.) y paleontológicas (fósiles) que ayudan a conocer dónde y cómo se originó la roca. En los casos en que se alude solo a las características litológicas se habla de litofacies, y si son las paleontológicas, biofacies. Fósiles guía y fósiles faciales Los fósiles guía son aquellos utilizados para definir e identificar periodos geológicos. Caracterizan a organismos de vida breve en el tiempo geológico pero amplia distribución geográfica. A la rama de la Paleontología que se encarga del estudio de los fósiles guía y de su utilización para la datación de terrenos se la llama Bioestratigrafía. Su utilización parte de la premisa de que, aunque los diferentes sedimentos pueden parecer muy distintos dependiendo de las condiciones bajo las cuales se originaron, deben incluir restos de las mismas especies que existían en la época de su formación. Si dichas especies tuvieron una vida corta (en términos geológicos), podrán ser utilizadas para determinar con bastante exactitud la edad del estrato en el que se encuentran. Cuanto más corta haya sido la duración de la especie, con más precisión podrá hacerse la datación. Por tanto, aquellos tipos de fósiles de organismos que evolucionan rápidamente son los más valiosos. Para que un fósil funcione como guía debe ser común, fácil de identificar hasta el nivel de especie y tener una amplia distribución mundial. Para las diferentes etapas del tiempo geológico, se utilizan diferentes fósiles guía. Por ejemplo, mientras que los trilobites son los fósiles guía característicos de la era Paleozoica, los ammonoideos lo son para la era Mesozoica y los foraminíferos, entre otros, para la era Cenozoica. Además de los fósiles guía, erxisten grupos de organismos que caracterizan determinados ambientes, los fósiles faciales, como los graptolites, típicos del plancton de la era Paleozoica, o los corales tabulados, característicos de mares tropicales de plataforma en la misma era. Estos son organismos de escasa distribución geográfica o ecológica, pero con una prolongada representación temporal. Un trilobite, fósil guía característico de la era Paleozoica. Un graptolite, fósil facial característico del plancton antiguo. Cuando los fósiles no alcanzan: el paleoADN Una nueva disciplina ha surgido en el campo de la Paleontología, la Paleontología molecular, que trata de hallar residuos de ADN en el material genético conservado en restos de pieles o partes blandas de especies extinguidas. Pero esta es una tarea de verdaderos detectives, ya que generalmente sólo se conservan partes duras. Las técnicas de secuenciación de ingeniería genética permiten, a su vez, reconstruir todo el código genético de la especie extinta a partir de los fragmentos de ácido nucleico. Sin embargo, esto implicaría no sólo la recuperación de todo el genoma del organismo, sino la inserción del material genético en un óvulo fecundado de una especie similar, del cual fue extraído el núcleo. Esto parece posible en el caso del mamut, ya que es un pariente muy cercano (una especie del mismo género) del elefante asiático, pero resultaría extremadamente difícil en el caso de los dinosaurios, a no ser que pensemos en las aves, supuestamente sus más próximos descendientes. Se ha encontrado ADN en un mosquito de un ámbar cretácico del Líbano y también se ha informado del aislamiento de ADN de un hueso de dinosaurio. Pero todos estos resultados se han criticado y cuestionado. La puerta al gran descubrimiento de Jurassic Park queda abierta. Insecto hematófago conservado en ámbar. La Tafonomía La Tafonomía (del griego taphos, sepulcro, y nomía, ley) es la disciplina que estudia los procesos biológicos, geológicos y fisicoquímicos que tienen lugar en la transformación de un cadáver de organismo en un fósil. Los organismos del pasado ocuparon diferentes ecosistemas (paleoecosistemas), estableciendo múltiples relaciones con su entorno mientras vivieron. Cuando un organismo muere y cae al lecho marino o a un pantano, comienzan a operar los procesos de descomposición y desintegración, que pueden hacer desaparecer todo resto del organismo original en poco tiempo, aunque se conservan las partes duras. La erosión puede disolver o fragmentar el resto, mientras que la actividad de otros organismos, como los escarabajos y los hongos, suele desarticularlos y descomponerlos. Con el transcurrir del tiempo, se depositan sucesivas pilas de estratos que comprimen los restos o estos se petrifican (son reemplazados por sales minerales) o desaparecen completamente aunque dejan su forma estampada en la roca. Finalmente, al cabo de millones de años, los restos son excavados o afloran las rocas fosilíferas al perderse los estratos superiores. La exploración paleontológica En el campo, los paleontólogos estudian la sucesión de los estratos, recolectan muestras de rocas, localizan los restos fósiles mediante análisis por computadoras y fechan los estratos por el análisis de la radiactividad. Cuando localizan un macrofósil, fósil mayor o macroscópico (por ejemplo, la impronta de una fructificación, un caparazón o un esqueleto), proceden a la separación del material de la roca para su estudio en el laboratorio (fósiles mayores o macroscópicos) que indican ambientes y edades geológicas. Estos estudios se complementan o son correlativos con los análisis del subsuelo para localizar los yacimientos minerales y de combustibles fósiles, obtener muestras de microfósiles o fósiles microscópicos y explorar las características de las formaciones geológicas. En estas múltiples tareas, los científicos utilizan toda clase de herramientas para la exploración directa del terreno, como los cinceles y picos, o instrumentos más sofisticados como computadoras, satélites con sensores remotos y radares para localizar los estratos y analizar relieves y rocas de diferentes edades y otros diversos métodos de análisis, como el monitoreo sísmico, del campo gravitatorio y magnético o la reflexión de ondas de luz, para trazar mapas tridimensionales del terreno. Las perforaciones realizadas mediante instrumentos diversos permiten, además, obtener perfiles de la columna geológica y recolectar muestras para analizar su contenido, su composición química y otras características que indican variaciones del ambiente y el clima. Para el estudio de los macrofósiles, se excavan directamente los estratos de rocas y sedimentos con herramientas especiales, como palas, martillos, cinceles y punzones, se desentierran los caparazones y esqueletos y se los separa de las rocas para su estudian en el laboratorio. También se forman moldes de yeso y en el museo se realizan maquetas, además de limpiarlos y acondiconarlos para su exhibición. Una de las labores más fructíferas, luego de interpretar otros datos ambientales y basados en el estudio de los microfósiles indicadores de suelos y vegetación, son las reconstrucciones de los organismos en su ambiente. Para el estudio de la topografía del terreno y las formaciones geológicas se utilizan métodos de prospección o exploración de yacimientos de minerales y combustibles, entre los cuales sobresale la contribución de los palinólogos, que estudian los granos de poley esporas, y los micropaleontólogos, que realizan importantes análisis de muestras de rocas, que permiten localizar con facilidad los depósitos sedimentarios en que se acumulan el carbón y el petróleo. Luego de localizado el depósito, se procede a la obtención de muestras de microfósiles mediante perforadoras y sondas. Es preciso recordar que el carbón se forma por la contribución de macrofósiles vegetales (troncos, raíces, etc.) y microfósiles (polen y esporas), mientras que el petróleo tiene un origen mixto (a partir de la acumulación de foraminíferos, diatomeas y otros microfósiles). A lo largo de los miles de millones de años transcurridos desde la formación del planeta Tierra hasta el desarrollo de la civilización moderno, la biota (conjunto de todos los orgnaanismos vivos, desde las bacterias al ser humano) se fue transformando progresivamente a medida que los ambientes cambiaban. Los fósiles incluyen un amplio muestrario de la biodiversidad pasada, incluidos restos de organismos (generalmente partes duras, como esqueletos y caparazones), grabados en las rocas (moldes e improntas), Muestra de microfósiles. Actividades iniciales 1. ¿Qué métodos de fosilización conocen? ¿Cuáles son más probables en la conservación de restos de plantas y de vertebrados? =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 2. Reflexione sobre lo siguiente. ¿Es frecuente que se formen fósiles? ¿Los fósiles llegan intactos a nosotros? ¿El registro fósil es un indicador fiel de la biodiversidad pasada. =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 3. Analice las siguientes secuencias y explíquenlas de acuerdo a lo planteado en la Introducción. =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 4. Defina biocrono y acmé. ¿Cómo describiría el biocrono de los dinosaurios? ¿Y de las aves? ¿Y si las aves fueran los sobrevivientes del linaje de los dinosaurios (como de hecho se afirma actualmente)? =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 5. Analice la importancia de la tafonomía y de la bioestratigrafía como dos caras de una misma moneda, el conocimiento de la vida en el pasado. =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== Foro Paseando con dinosaurios La película Dinosaurio de los estudios Walt Disney Pictures, logró una excelente presentación de los ecosistemas del pasado mediante técnicas de fotorrealismo, utilizando animaciones generadas por computadoras en tres dimensiones, que permiten que los personajes se muevan sobre un fondo de escenarios reales. Dinosaurio transcurre durante el Cretácico superior, más exactamente durante el límite Cretácico-Paleoceno, hace aproximadamente 65 millones de años, cuando un gran asteroide hizo impacto con el planeta Tierra. Intenta representar los paisajes cretácicos a partir de escenas de diferentes lugares como California, Australia, Hawaii, Florida, Venezuela y Samoa Occidental. El realismo alcanzado por estas técnicas es asombroso ya que permiten recrear con sus más mínimos detalles no solamente el exterior de la piel de los dinosaurios sino también una buena representación de la musculatura de los animales, su comportamiento reproductor y cuidado de las crías, su forma de alimentación, la velocidad alcanzada en la carrera, las técnicas de defensa y ataque, etcétera. Por supuesto que tan sólo se logra una interpretación de la forma en que estos animales se comportaban, ya que sólo se conocen generalmente unos pocos restos fosilizados. En la película, una familia de lémures primitivos descubre un huevo que, casualmente, fue dejado caer por un pterosaurio o reptil volador. Del huevo nace un Iguanodon, dinosaurio que es adoptado y criado por los lémures como si fuera una mascota. Al crecer Aladar, el Iguanodon, reconoce que él pertenece a otro especie. Tras la caída del asteroide y la destrucción de grandes extensiones de bosques, Aladar y su familia adoptiva se unen a una manada de dinosaurios muy heterogénea, que van en busca de un sitio fuera de peligro para anidar. Durante el éxodo son atacados por dinosaurios carnívoros, primero por una manada de Velociraptor y luego por una pareja de carnotaurios (los villanos sin nombre en la película). El Carnotaurus sastrei (el carnotaurio más conocido ya que otras especies se conocen de Madagascar y la India) fue encontrado por la familia Sastre en su estancia, situada en la región de Bajada Moreno, Departamento de Telsen, Provincia de Chubut, en la Argentina. Los restos fósiles de este dinosaurio fueron excavados en 1984 por un equipo del Museo Argentino de Ciencias Naturales "Bernardino Rivadavia", dirigido por el paleontólogo Dr. José F. Bonaparte. El esqueleto fue encontrado articulado, en posición natural, dentro de una roca que realmente protegió al especimen de la meteorización. También se conservaron impresiones fosilizadas del cuero o de la piel, que fueron descubiertas debajo de varias partes del esqueleto. Los restos originales del Carnotaurus sastrei se encuentran depositados en el Museo Argentino de Ciencias Naturales “Bernardino Rivadavia” (MACN), de la Ciudad de Buenos Aires. En la Sala de Paleontología de dicho museo se encuentra montada una reproducción del esqueleto. En la película aparecen otros dinosaurios, pterosaurios (reptiles voladores), aves, lagartos y primates lemuriformes. ¿Qué se puede decir acerca de los padres adoptivos de Aladar? Los ldmures son mamíferos primates que habitan actualmente en la isla de Madagascar, pero que durante el Paleoceno y Eoceno se extendieron por otros continentes. Sin embargo, no se conoce la existencia de lémures del Cretácico, aunque sí de otros mamíferos que coexistieron con los dinosaurios, como Vincelestes neuqueanus, y también fueron registrados otros mamíferos pequeños similares a los roedores que se dominan los gondwanatherios (o mamíferos de Gondwana), del Cretácico superior de la Provincia de Río Negro, Argentina. 1. Más allá de los errores e imprecisiones, ¿creen que alguna vez los hechos descritos en Jurassic Park podrían ser una realidad? 2. ¿Por qué es importante plantear correctamente la reconstrucción de comunidades fósiles? 3. ¿Es posible recrear los ecosistemas del pasado? ¿Cómo pueden los paleontólogos conocer las características del comportamiento y adaptaciones de los dinosaurios y otros animales extintos? 4. Busquen información sobre la dieta, defensas y otras características de los distintos grupos de dinosaurios. ¿En qué época vivieron los dinosaurios? 5. La teoría de la extinción por un gran asteroide cuenta actualmente con muchas evidencias. ¿Qué otras teorías explicarían la extinción de los dinosaurios? Organicen un debate para defender el punto de vista de la extinción que presenta la película Dinosaurio. ¿Creen que sobrevivieron dinosaurios a la caída del asteroide? Módulo 2. Datación y cambios paleogeográficos Objetivos • Analizar brevemente la historia de los descubrimientos paleontológicos • Determinar las edades relativas • Distinguir entre datación relativa y absoluta • Explicar los principios de la datación absoluta • Determinar la columna geológica patrón • Comparar las etapas de la historia terrestre con un reloj cósmico • Analizar los métodos de datación sedimentológicos y biocronológicos • Examinar los principios de correlación estratigráfica • Analizar la forma de ordenar la secuencia de fósiles • Comparar el estudio de microfósiles y microfósiles • Analizar las características y la evolución de los supercontinentes • Evaluar la importancia de los cambios paleoclimáticos • Comparar la teoría de la deriva continental con la tectónica de placas • Correlacionar los cambios paleogeográficos y paleoclimáticos con la aparición y extinción de organismos • Comparar extinciones graduales y masivas Contenidos Edad de la Tierra en foco La datación como proceso central Geocronología y edad de los fósiles Datación radimétrica Columna geológica patrón Otros métodos de datación Correlación estratigráfica Techo y muro Criterios de polaridad Concordancias y discordancias Dificultades en la correlación El estudio de los microfósiles Columna geológica patrón Cambios paleogeográficos Teoría de Wegener Cambios paleoclimáticos Teoría de la tectónica de placas Movimiento de las placas Ciclo supercontinental El oxígeno y la historia del clima terrestre Los eventos extraterrestres y la extinción Los comienzos de la datación Durante el siglo XVIII, no existían medios tecnológicos para determinar la antigüedad de las rocas, y la edad de la Tierra era un tema de especulación. Pero el descubrimiento, a fines del siglo XIX, de los fósiles guía, organismos representativos de distintas épocas geológicas que se conservaban en los estratos de rocas sedimentarias, así como el descubrimiento de la radiactividad a fines del siglo XIX y comienzos del siglo XX, revolucionó la comprensión de la historia del planeta. Tres grandes científicos se relacionan con estos avances: el geólogo inglés James Hutton (1726-1797), el físico irlandés Lord Kelvin (1824-1907) y el físico inglés Ernest Rutherford (1871-1937). Kelvin estimó que la edad de la tierra oscilaba entre 20 y 400 millones de años, mucho más que los 4.000 años que se consideraban por entonces hasta el Diluvio en base a la edad de los patriarcas bíblicos. Sus cálculos se basaron en los efectos de la convección del manto terrestre y de su reloj radiométrico. Poco después, en 1896, Rutherford, descubrió el núcleo atómico y comenzó las primeras mediciones de la desintegración de los isótopos radiactivos para medir la edad de las rocas. La edad de la Tierra en foco La sensación de estar habitando en un planeta recién creado flotó por el imaginario de nuestros antepasados durante mucho tiempo. Desde que el cristianismo se impuso con su cronología bíblica, pareció necesario determinar cuál había sido el momento exacto de la creación, para lo cual se hicieron los cálculos más inverosímiles. El ejemplo más famoso es el del arzobispo James Ussher, del Trinity College de Dublín, quien mediante un minucioso análisis de las escrituras, concluyó que la Creación había tenido lugar a las nueve de la mañana del día 26 de octubre del año 4004 a. de C. Si bien las cifras eran discutibles, en ese entonces, no diferían demasiado: un contemporáneo de Ussher, John Lightfoot, de la Universidad de Cambridge, propuso el año 3928 a. de C.; mientras el astrónomo polaco Johannes Hevelius calculaba la posición exacta del sol en el jardín del Edén a la hora de la creación, que, según él, había sido a las 18 horas del 24 de octubre de 3963 (a. de C.). El mismísimo Newton dedicó buena parte de su tiempo al problema de calcular el momento de la Creación, del Diluvio y del Juicio Final. Hacia el siglo XIX, sin embargo, la idea de una tierra reciente empezó a desmoronarse cuando emergió con fuerza una vieja teoría de Descartes (1596 – 1650): según él, la Tierra no era sino un trozo que se había desprendido del Sol y que se había enfriado lentamente. Obviamente, una esfera incandescente del tamaño de la Tierra no podía enfriarse en sólo cuatro o cinco mil años. Necesitaba mucho más tiempo. El primero en arriesgar una cifra superior fue el naturalista y precursor de la ciencia popular Buffon (1707-1788). Buffon adoptó la teoría de Descartes de que la Tierra no era más que un fragmento desprendido del sol e ideó un curioso y sencillo procedimiento para ver cuánto tardaría en enfriarse: moldeó 24 bolitas de 2 centímetros y medio de diámetro y las calentó en el horno. Después, sólo restaba medir el tiempo que transcurría hasta que se pudieran agarrar con la mano desnuda. La cifra que obtuvo, finalmente, fue de 74.832 años. Las repercusiones de este osado cálculo fueron estrepitosas: resultaba difícil de creer que la Tierra fuera tan espantosamente antigua. Y eso que todavía faltaba mucho. En 1830 Charles Lyell publicó su Geología (que dicho sea de paso inspiraría a Darwin), donde sostenía que los procesos geológicos eran lentísimos y que habían sido siempre así: evidentemente, para cavar un cañadón por donde pasa un río no alcanzaba con los setenta y cinco mil años de Buffon. Además, ya circulaban las primeras hipótesis evolucionistas que necesitaban como ingrediente fundamental el tiempo en grandes cantidades. El asunto es que, inspirado por Lyell, el geólogo John Philips, hizo una estimación de la edad de la corteza terrestre estudiando a fondo los estratos rocosos y estimando cuánto habían tardado en formarse y le dio ¡noventa y seis millones de años! A lo largo del siglo XIX la cifra aceptada por los científicos para la edad de la Tierra rondó los cien millones de años. En 1863 Lord Kelvin, el gran físico inglés, calculó como edad mínima para la Tierra veinte millones de años, y, como edad máxima, ¡doscientos millones! Y sin embargo seguía siendo poco: cuando se pudo empezar a medir la antigüedad por medio de métodos radiactivos, un geólogo inglés, Arthur Holmes, en 1931 aseguró que la tierra tenía al menos 1460 millones de años. Un poco más tarde, gracias al refinamiento de los cálculos, elevó la edad de la Tierra a cuatro mil quinientos millones de años. Y esa es la cifra que (con respeto, con incredulidad) manejamos hoy. La datación, proceso central para conocer la historia de la vida Si hablamos de fósiles, nos interesa conocer la edad (absoluta, pero también relativa, es decir, en relación con depósitos similares) de las rocas que los contienen. Estos patrones revelaron mucho acerca de las edades relativas de las rocas sedimentarias así como patrones en la evolución de la vida. Pero los geólogos aún no podían decir cuán antiguas eran realmente las rocas. No hubo disponible un método de datación de rocas hasta que la radiactividad fue descubierta a comienzos del siglo veinte. La columna geológica es la serie de estratos que se depositan a lo largo de miles a millones de años. Muchas veces estos sedimentos afloran en el terreno, y otras exigen laboriosas perforaciones profundas para muestrear los sedimentos, suelos, rocas e incluso capas de hielo antártico de diferentes estratos y edades. Estas muestras individuales son los testigos, ya que evidencian cambios de la fauna, la flora, el clima o los ecosistemas de una época de deposiciòn. Los perforadores de muestras consisten básicamente en instrumentos cortantes, como un sacabocados gigante, o aquellos que utilizan el principio del tornillo de Arquímedes para retirar las muestras. Luego del muestreo, las porciones de cada nivel se ubican en uno o más recipientes para su análisis posterior en el laboratorio. La Geocronología La Geocronología es la ciencia que se ocupa de la medición y la determinación del tiempo geológico. Los diversos intervalos de tiempo determinados a partir de las formaciones rocosas acumuladas en la columna de estratos son las unidades geocronológica, de las cuales las mayores son el eón, la era, el período y la época. Cada unidad se caracteriza por determinadas condiciones geográficas, del relieve, el clima, los ambientes y, por supuesto, los seres vivos diferentes que entonces vivieron. Los isótopos radiactivos de los átomos se desintegran bajo un patrón predecible a lo largo de amplios períodos de tiempo. Durante cada lapso sucesivo, o vida media, la mitad de los materiales radiactivos restantes de los radioisótopos se desintegran, bien cambiando en otro elemento, o volviéndose el isótopo estable del mismo elemento Cada radiosiótopo presenta una vida media característica. Para emplear un radioisótopo para fechar un suceso del pasado, debemos conocer o estimar la concentración de dicho isótopo en el momento del evento. En el caso del carbono, la producción de nuevo 14C en la alta atmósfera (mediante la reacción de neutrones con el 14N) equilibra la desintegración radiactiva natural del 14C. Por lo tanto, la proporción de 14C respecto al isótopo estable 12C es relativamente constante en los organismos vivientes y en su ambiente. Sin embargo, tan pronto como un organismo muere, cesa de intercambiar compuestos de carbono con su ambiente. Su 14C en desintegración ya no es recuperado, y la proporción de 14C a 12 C en sus restos, decrecen a lo largo del tiempo. Los paleontólogos (científicos que estudian los fósiles) pueden usar la relación 14C a 12C en los organismos fósiles para fechar a aquellos que tienen menos de 50.000 años de antigüedad (y, por lo tanto, a las rocas sedimentarias que los contienen) con un grado de certeza fiable. Después de este tiempo, las trazas de 14C que permanecen son tan pequeñas que alcanzan los límites de su detección. Datación radimétrica Cuando se precisa conocer los datos exactos del tiempo que ha transcurrido desde un determinado suceso geológico en la historia de la Tierra, el método más exacto al que puede recurrirse es la datación radimétrica, que utiliza la radiactividad natural de los elementos presentes en las rocas. Por lo general, las rocas contienen una cierta cantidad de elementos radiactivos, como el uranio, el radio y el torio, entre otros, y sus respectivos isótopos. Los isótopos son átomos con el mismo número atómico (igual número de protones) pero diferente número másico (protones más neutrones). A medida que transcurre el tiempo, estos isótopos se desintegran espontáneamente y se transforman en otros elementos, como el plomo o el helio. Algunos ejemplos son el uranio 235 ( 235U), que se transforma en plomo 207 (207Pb); el uranio 238 (238U), que se convierte en plomo 206 ( 206Pb); el torio 232 (232Th), que da lugar al plomo 208 (208Pb) y el carbono 14 (14C) que da lugar a nitrógeno 14 (14N). Los factores ambientales no influyen en este proceso y, en la mayoría de los casos, el fenómeno se produce a lo largo de miles de años. Sin embargo, independientemente del elemento de que se trate, la transformación sigue siempre una regla “de oro”: la mitad de los átomos del elemento que existen en un momento inicial, se descomponen en una determinada cantidad de años, específica para cada isótopo. Un análisis minucioso de la composición de una roca permite establecer la cantidad presente de átomos del elemento radiactivo no descompuesto y las del isótopo estable que han aparecido desde su formación. De este modo, se puede calcular la edad de la roca estudiada. Algunos ejemplos de isótopos utilizados en la datación radimétrica Isótopo Isótopo estable Vida media (en Materiales que pueden inestable años) datarse Uranio 238 Plomo 206 4,50 . 109 Circón y uranitita 9 Uranio 235 Plomo 207 0,70 . 10 Circón y uranitita Torio 232 Plomo 208 13,90 . 109 Circón y uranitita 9 Potasio 40 Argón 40 1,25 . 10 Moscovita Calcio 40 Biotita Hornblenda Rocas volcánicas 9 Rubidio 87 Estroncio 87 47 . 10 Moscovita Biotita Feldespato (potásico) Rocas ígneas y metamórficas Carbono 14 Nitrógeno 14 5,750 +/- 30 Restos orgánicos Aguas subterráneas Glaciares Pinturas rupestres y otros elementos asociados a culturas prehistóricas e históricas En las últimas décadas se comenzó a utilizar el método del potasio - argón. Este método se basa en que la radiactividad transforma el isótópo del potasio (K), de peso atómico 40, en gas argón (A), del mismo peso atómico. Para datar sucesos recientes, se recurre al método del radiocarbono. La atmósfera contiene isótopos radiactivos del carbono 14, 14C, cuyo período de semidesintegración es de 5.568 años. La presencia de este isótopo se debe a la acción de los rayos cósmicos en la atmósfera, que generan la producción de neutrones. Estos reaccionan con el isótopo del nitrógeno, 14N, y dan origen al isótopo radiactivo 14C. El 14C es absorbido por las plantas como dióxido de carbono del aire y, en consecuencia, pasa los organismos que se alimentan de ellas. Mientras el organismo vive, la relación entre el carbono no radiactivo y el radiactivo no varía; cuando muere, la cantidad de radiocarbono comienza a disminuir. Averiguando la relación entre ambos y conociendo el período de semidesintegración, puede hallarse la edad de un material orgánico de hasta 10.000 años de antigüedad. La precisión no es menor de 200 años, pero es suficiente para múltiples propósitos. Evolución de la proporción de elementos padre e hijo. La datación de las rocas con radioisótopos, combinada con los análisis de los fósiles constituyen los métodos más poderosos para la determinación de la edad geológica. Pero en los sitios donde las rocas sedimentarias no contienen intrusiones ígneas apropiadas y pocos fósiles se hallan presentes, los paleontólogos deben recurrir a otros métodos de datación. Un método conocido como datación paleomagnética, que relaciona las edades de las rocas con los patrones del magnetismo terrestre, que cambia con el tiempo. Los polos magnéticos de la Tierra se mueven y ocasionalmente revierten su posición. Debido a que tanto las rocas sedimentarias como las ígneas preservan un registro de los campos magnéticos terrestres en el momento en que se formaron, el paleomagnetismo ayuda a determinar las edades de dichas rocas. Otros métodos de datación que serán descritos en capítulos posteriores utilizan la deriva continental, los cambios en el nivel de los mares y los relojes moleculares. Mediante el uso de tales métodos, los geólogos dividieron la historia de la vida en eras geológicas, que a su vez están divididas en períodos (Tabla 21.2). Los límites entre estas divisiones se basan en diferencias sorprendentes observadas por los científicos en los conjuntos de organismos fósiles contenidos en los estratos sucesivos de rocas, de ahí el sufijo zoic (“ de vida”) para las eras. Los geólogos establecieron y nombraron a estas divisiones antes de haber conocido las edades de las eras y los períodos, y continuamente estamos refinando las fechas usadas para dichos límites a medida que se realizan nuevos descubrimientos. Otros métodos de datación Existen otros procesos naturales rítmicos que ocurren a una velocidad constante y, en consecuencia, pueden ser utilizados para datar. Entre ellos destacan: A. Las varvas glaciares. Se forman en lagos cubiertos por una capa de hielo en invierno que en verano se funde. Cada año se depositan dos capas de sedimentos, una clara y una oscura. La clara corresponde al sedimento de verano y está formada por los materiales más gruesos (son los primeros en depositarse). Los materiales muy finos permanecen en suspensión y se depositan en invierno, cuando el hielo superficial deja las aguas en calma. Su color oscuro se debe a la presencia en ellos de materia orgánica no oxidada. Bastará contar el número de pares de bandas para conocer el número de años durante los cuales se ha estado formando este sedimento. Este método de datación solo es útil para materiales de edades muy recientes. Utilizando las varvas glaciares se ha podido establecer en Suecia una cronología de los últimos 9000 años. B. Anillos de crecimiento de los corales. Algunos corales antiguos con forma de campana originaban una fina capa diaria de carbonato cálcico. Una pequeña estrangulación marca la separación entre un año y otro, de manera que es posible contar el número de láminas (días) de cada año. La atracción lunar está frenando el giro terrestre a razón de 20 segundos cada millón de años. Por eso durante la historia de la Tierra los días han sido cada vez más largos y, en consecuencia, se han reducido los días que tiene un año, manteniéndose constante el total de horas anuales. Así, hace 550 M.a. el día duraba 21 horas, y el año tendría 417 días. Si se cuentan los anillos del coral, se sabrá los días que tenía aquel año y, por tanto, su antigüedad. Varvas. La correlación estratigráfica y la edad relativa Los sedimentos se depositan en estratos o capas, que se forman a lo largo de millones de años por la acumulación de partículas. Al endurecerse, los sedimentos forman los estratos de las rocas sedimentarias (proceso conocido como litificación). La Estratigrafía es la ciencia que estudia los estratos de rocas sedimentarias. Para analizar como se produce la sedimentación, pueden realizar la siguiente experiencia en el aula. Consigan dos vasos de agua y agreguen en el primero, lentamente, arena; luego, trozos de piedras, y, finalmente, otra capa de arena. Repitan el mismo proceso con el segundo vaso, pero agreguen los elementos mezclados. Observen cómo se depositan los sedimentos en ambos casos. El ya mencionado Niels Stenen debió pensar en un experimento similar cuando enumeró algunos principios que rigen la depositación de los estratos. Como señaló, al depositarse los estratos, van englobando restos fósiles, como los esqueletos y dientes de vertebrados y los caparazones de moluscos. Si el enterramiento es rápido (no actúan las bacterias de la descomposición), el ambiente es de baja energía (escaso transporte y erosión), y hay poco oxígeno (no ocurre la oxidación), los restos fósiles se conservan mejor. El contenido de fósiles sirve para determinar la edad relativa de los estratos de rocas sedimentarias, es decir, indican qué resto se depositó antes o después en la columna geológica. Si las rocas de dos localidades contienen los mismos fósiles, puede afirmarse que se conservaron en la misma época. Es decir, comparando el contenido de fósiles de diferentes localidades, es posible correlacionar los estratos (correlación estratigráfica) de la misma edad. Correlación de estratos de dos localidades geográficas. En particular, Niels Stenen y a continuación Adam Segdwik y James Hutton, establecieron los principios de la estratigrafía y la correlación: A. Principio de horizontalidad original de los estratos: los sedimentos se depositan formando capas horizontales. De acuerdo con este principio, si encontramos un conjunto de estratos que no se disponen horizontalmente se concluirá que, después de su formación, han sido sometidos a algún tipo de esfuerzo que les ha cambiado su disposición original. B. Principio de continuidad lateral de los estratos: originalmente, los estratos se extienden lateralmente y terminan adelgazándose en sus bordes. La edad es la misma en toda la superficie de un estrato. C. Principio de superposición de los estratos: los sedimentos se depositan unos sobre otros, de tal manera que en una serie que se encuentre en su disposición original el estrato situado más abajo es el más antiguo y el de arriba, el más moderno. Para representar por orden cronológico los materiales se utiliza la columna estratigráfica. En ella se representan horizontalmente los materiales, abajo los más antiguos, arriba los más modernos y, además de los tipos de rocas, se indica la presencia de fósiles y otras características. Techo y muro Las superficies que limitan un estrato reciben el nombre de planos de estratificación. La parte más alta y moderna de un estrato se denomina techo. La base de un estrato será el muro. Los mismos términos se utilizan para referirse a una serie de estratos: la parte superior y más moderna sería el techo de la serie, mientras que la parte más baja y más antigua correspondería al muro de la serie. La distancia medida en vertical entre el techo y el muro de un estrato recibe el nombre de potencia. Criterios de polaridad El principio de superposición es suficiente para ordenar una serie que mantenga la horizontalidad original. Sin embargo, en muchas ocasiones los estratos han abandonado esa disposición, encontrándose verticales e incluso invertidos, es decir, con el techo situado por debajo del muro. En estos casos se utilizan los criterios de polaridad o criterios de techo–muro, que son un conjunto de estructuras sedimentarias que permiten orientar los estratos. Estas estructuras a veces se presentan en la superficie de los estratos y otras, en su interior. Concordancias y discordancias Se dice que dos materiales son concordantes si la superficie que los separa es paralela a los planos de estratificación de ambos, en caso contrario serán discordantes. La discordancia implica que entre el depósito de un material y el siguiente ha ocurrido algún proceso. Si lo que ha sucedido es una erosión se habla de discordancia erosiva. Si ha habido un plegamiento será una discordancia angular, y si se han producido un plegamiento y una erosión la discordancia será angular y erosiva. ¿Cómo ordenar los acontecimientos? Los materiales A, B, C y D que aparecen en la figura de la derecha están plegados. En aplicación del principio de sucesión de acontecimientos, un proceso geológico (plegamiento) es posterior a los materiales que afecta (estratos A, B, C y D). Además, como estos pliegues están erosionados, deberá concluirse que el proceso de erosión es posterior al de plegamiento. Aplicando los principios de horizontalidad original, superposición de los estratos y sucesión de acontecimientos, se puede deducir la secuencia completa de acontecimientos que ha dado lugar a la estructura final. Dificultades en la correlación La correlación estratigráfica se ve dificultada por los procesos endógenos y exógenos que actúan en la superficie terrestre, por ejemplo, la erosión puede quitar una capa o estrato de sedimentos de la columna geológica, y un pliegue o falla puede invertir el orden de deposición de los sedimentos. Cuando falta un estrato o una serie de estratos sedimentarios se habla de hiato o discontinuidad. También es importante tener en cuenta que en distintas épocas una región continental pudo haber sido invadida extensamente por el mar (transgresión) iniciándose la sedimentación, y otra oceánica pudo secarse y aflorar las rocas (regresión) iniciándose una etapa de erosión. Es posible que más tarde, un sedimento oceánico ascienda por los fenómenos tectónicos y forme una montaña, conservándose allí los fósiles marinos. Estudio de microfósiles y prospección geológica La prospección geológica es la exploración de recursos geológicos por diferentes métodos. La exploración de nuevas reservas de combustibles fósiles, en especial petróleo, implica un conocimiento muy detallado del subsuelo, y en estos estudios sobresalen las contribuciones de los palinólogos y los micropaleontólogos. Los palinólogos, que estudian los granos de polen y esporas de plantas fósiles y los micropaleontólogos, estudiosos de los microfósiles o fósiles de microorganismos, realizan importantes análisis de las muestras de rocas que permiten situar con facilidad los depósitos sedimentarios en que se han acumulado el carbón y el petróleo. Es preciso recordar que el carbón se forma no sólo por la contribución de macrofósiles vegetales (troncos, raíces, etc.) sino también por microfósiles (polen y esporas) mientras que el petróleo presenta un origen mixto, a partir de la acumulación de foraminíferos, diatomeas y otros microorganismos. Estos estudios complementan aquellos que aportan la gravimetría, el magnetismo, la sísmica y otras técnicas geológicas muy precisas para realizar la prospección. La columna geológica patrón Los distintos métodos de datación han permitido establecer una columna geológica. Los distintos intervalos de tiempo determinados a partir de las formaciones rocosas acumuladas en la columna son las unidades geocronológicas: el supraeón, el eón, la era, el período, la época, la edad y el crono, del más extenso al más breve. En Geología se distinguen dos tipos de unidades. Las unidades cronoestratigráficas se basan en secuencias de estratos (litológicas) y por lo tanto tienen existencia real caraterizadas por su contenido de fósiles. Por otra parte, las unidades geocronológicas son unidades paratemporales basadas en lls métodos de datación radimétrica. Unidades cronoestratigráficas Unidades geocronológicas Eonotema Eón Eratema Era Sistema Período Serie Época Piso Edad Zona Crono El eón es el mayor intervalo en la escala del tiempo geológico. Se distinguen el eón Chaotiano, desde los 4.560 a los 4.350 m.a., el eón Hadeano, desde los 4.350 m.a. (cuando se consolida el planeta Tierra) hasta los 3.850 m.a., el eón Arqueano, hasa los 2.500 m.a., el eón Proterozoico, hasta los 543 m.a. y el eón Fanerozoico, hasa la actualidad, dividido en tres grandes eras: Paleozoica, Mesozoica y Cenozoica. Los tres primeros eones constituyen los Tiempos precámbricos (antes del Cámbrico, que es el primer período de la era Paleozoica). Las eras del Fanerozoico están divididas en intervalos de tiempos cada vez más pequeños, como los períodos, las épocas y las edades. El crono es la unidad inferior que representa el tiempo correspondiente al rango d eun fósil o grupos de fósiles detemrinados y su denominación se efectúa tomando el nomre de la localidad geográfica y/o del fósil que sirvió para su caracterización. ¿Cómo cambiaron con el tiempo los continentes y climas de la Tierra? Los mapas y globos terráqueos con los que adornamos nuestras paredes, estantes y libros nos brindan una impresión de una Tierra estática. Sería fácil para nosotros suponer que los continentes han estado siempre en el mismo lugar que en el que están. Pero estaríamos equivocados. La idea de que las masas continentales de la Tierra han cambiado de posición en el transcurso de los milenios y que lo continúan haciendo de esta manera, fue primeramente planteado en 1912 por el meteorólogo y geofísico alemán Alfred Wegener. Su libro El Origen de los Continentes y Océanos fue tomado inicialmente con escepticismo y resistencia. Hacia la década de 1960, sin embargo, la evidencia física y la mejor comprensión de los geofísicos de la tectónica de placas han convencido virtualmente a todos los geólogos de la realidad de la visión de Wegener. La corteza terrestre consiste en un número de placas sólidas de aproximadamente 40 kilómetros de espesor, que colectivamente constituyen la litosfera. Las placas litosféricas flotan sobre una capa fluida de rocas fundidas, o magma. El magma circula porque el calor producido por la desintegración radiactiva en el núcleo terrestre establece corrientes de convención en el fluido. Las placas se mueven porque el magma se eleva y ejerce tremendas presiones. Donde las placas son empujadas entre sí, o bien, se desplazan lateralmente entre sí, o una de ellas se hunde debajo de la otra levantando cadenas montañosas y excavando profundos valles de rift. (Cuando esto sucede debajo del agua, dichos valles son conocidos como fosas). Donde las placas se separan, las cuencas oceánicas pueden formarse entre ellas. El movimiento de las placas litosféricas y de los continentes que ellas contienen se conoce como deriva continental. Las primeras ideas movilistas Los continentes no solo están sometidos a los movimientos verticales que describe la isostasia sino también a otros horizontales. Las teorías que defienden estas tesis se denominan movilistas. Sus antagonistas son las teorías fijistas. La historia de las ideas movilistas está inevitablemente unida al libro El origen de los continentes y océanos publicado en 1915. Su autor, un meteorólogo alemán llamado Alfred Wegener, defendía que los continentes se desplazaban. La complementariedad entre las costas de África y Suramérica había sido detectada tras los grandes viajes de los siglos XVI y XVII, cuando se elaboraron los primeros mapas del mundo que ya incluían América. Así, Francis Bacon en 1620 comentaba este llamativo parecido. No obstante, sería excesivo considerarlo un predecesor de Wegener. A principios del siglo XIX, Alexander von Humboldt no solo se sorprende de la complementariedad de las costas a uno y otro lado del Atlántico, sino que señala la continuidad entre algunas formaciones geológicas africanas y suramericanas. A su juicio, debía investigarse este ajuste, que no le pareció casual. Pero el precedente más inmediato y claro de Wegener fue Frank Taylor. En 1910 publicó un extenso trabajo en el que exponía la hipótesis de la movilidad continental. El punto de partida de Taylor no era el ajuste de los continentes, sino la forma y distribución de las cordilleras en Asia y Europa. Supuso que la corteza se había desplazado de norte a sur. Este desplazamiento y las ocasionales colisiones originarían las cordilleras. Alexander von Humboldt. Estudios paleogeográficos Cuando se encontraron los primeros restos fósiles de hipopótamos y cocodrilos en lugares como Noruega o Gran Bretaña, así como corales en Siberia, los científicos se vieron en apuros para encontrar una explicación coherente y lógica del hecho. ¿Se trataba de testimonios del diluvio? ¿O tal vez todos los continentes habían estado unidos en el pasado? La Paleogeografía es el estudio de la distribución de los continentes y los océanos a lo largo de las eras geológicas. Trata de explicar, en un contexto espacial, la distribución de las faunas y floras en la historia terrestre. Según los estudios de dispersión, la franja de tierra que separa dos océanos actúa como una barrera efectiva para la migración marina, mientras que los océanos profundos impiden la dispersión de organismos entre dos islas o entre una isla y el continente. Pero una vez que queda eliminada la barrera geográfica, el puente formado (oceánico o continental) permite el libre intercambio de especies a modo de un corredor, lo que conduce a una presión de selección y a la extinción de las especies menos adaptadas al nuevo ambiente o de aquellas que no pueden competir en igualdad de condiciones con los recién “llegados”. Pero, ¿cómo realizan los investigadores las reconstrucciones geográficas del pasado? Recurren a datos paleomagnéticos (el magnetismo remanente de las rocas y su variable orientación respecto de los polos magnéticos terrestres en cada época), litológicos (depósitos de tillitas o rocas glaciarias que indican un clima frío; de evaporitas, rocas sedimentarias que se depositan en condiciones desérticas y cálidas; etc.), geofísicos (el perfil de las líneas de costa entre dos continentes, la ubicación y distribución de las cadenas montañosas submarinas, etc.) y, por supuesto, paleontológicos (presencia de fósiles similares en dos zonas continentales alejadas) o biogeográficos (presencia de grupos de organismos vivientes en áreas muy alejadas, las que pudieron haber estado conectadas en el pasado geológico). Se pueden analizar dos ejemplos muy famosos de cómo se explican estas antiguas distribuciones de los organismos. 1. El gran escritor francés Julio Verne, en su novela Viaje al centro de la Tierra, hablaba de un verdadero mar interior, pero desconocía que este mar existió aunque no en las “entrañas” del planeta, sino en otras épocas. En efecto, esto ocurrió a fines del Triásico, cuando se formó el océano interior Tethys, que separaba los antiguos continentes de Gondwana al sur y de Laurasia al norte. Este acontecimiento produjo un cambio en el patrón de circulación marina, de este a oeste (en vez de norte a sur) y las especies de invertebrados y peces pudieron intercambiarse sin problemas a ambos lados de los continentes. (distribución de organismos comunes en Gondwana, tomar ilustración de p. 177 de Historical Geology, en parte, pero en base a paleomapa de Les mondes disparus) 2. Cuando Charles Darwin visitó nuestro país en el transcurso de su famoso viaje en el Beagle descubrió mamíferos más antiguos, que eran muy diferentes a los que vivieron en otros continentes, y otros con características similares a los del Hemisferio Norte. ¿Cómo era posible esta mezcla? La explicación la dio el gran investigador argentino Florentino Ameghino: Sudamérica había sido una isla con su propia fauna endémica, pero al levantarse el puente centroamericano, se hizo posible la migración de la fauna a ambos lados del corredor. Los “vencedores” fueron los llegados del Hemisferio norte, como los zorros y las vicuñas, que compitieron exitosamente con las especies de carnívoros y herbívoros locales (por ejemplo, los borhiénidos, con aspecto de zorros y los astrapoterios, con apecto de vicuñas) por los mismos recursos o predaron sobre ellos. La idea de la deriva continental fue captada por Wegener cuando estudiaba el estrecho encaje entre las costas del oeste de África y del este de Sudamérica. La evidencia geológica que relaciona las formaciones de rocas en los Apalaches con formaciones similares en las tierras altas de Escocia, así como fenómenos geológicos hallados tanto en Sudáfrica como en Brasil, alentó su visión de que los continentes estuvieron unidos alguna vez. Ahora sabemos que, en ocasiones, la deriva de las placas ha unido los continentes y que en otros tiempos, dichos continentes se han separado entre sí. Las posiciones y los tamaños de los continentes influyen sobre los patrones de circulación oceánica, los niveles del mar y los climas globales. Las extinciones masivas de las especies, en especial de los organismos marinos, han acompañado generalmente cambios importantes en el nivel del mar, que expusieron amplias áreas de las plataformas continentales, eliminando a dichos organismos que vivían en los mares someros que éstas cubrían. Evidencias de la deriva continental Evidencias geográficas. El borde de los continentes encaja entre sí, tal como se observa en el caso de Sudamérica y África, lo que implica que se separaron en algún momento de la historia geológica, más concretamente en el Jurásico. Evidencias fósiles Los paleontólogos han encontrado restos fósiles de ciertas especies tanto en Sudamérica como en el sur de África. Es un hecho curioso que una especie fósil fuera hallada en ambos lados del océano Atlántico. Nadie cree que estas especies hubieran cruzado tal distancia nadando, por lo tanto, éste sería un hecho que apoya la idea de que estas regiones estuvieron alguna vez unidas. El hallazgo del reptil de agua dulce, Mesosaurus, en el este de Sudamérica y el sur de África, el helecho Glossopteris en todo el hemisferio sur y el reptil terrestre Cynognathus en la Argentina y el sur de África, que vivieron hace 250 millones de años, indican que en ese entonces estas masas continentales debieron estar unidas, lo que permitió la dispersión de la flora y la fauna de la época. Evidencias geológicas. En ambos lados del Atlántico se encuentran montañas de edad y estructura comparables. Esto significa que los continentes no sólo se complementan en sus bordes, sino que también concuerdan en el tipo de rocas que presentan. Esta es una evidencia muy fuerte en favor de la antigua unión de estos continentes. Por ejemplo, el tipo de rocas que se encuentran en la zona del Cabo de Buena Esperanza (sur de África) también pueden hallarse en la provincia de Buenos Aires. Evidencias climáticas. Los registros de los climas de hace millones de años (estudiado por la paleoclimatología) indican que las regiones que se encuentran actualmente en la zona ecuatorial estuvieron alguna vez cubiertas por glaciares. Estas evidencias son difíciles de explicar con la disposición actual de los continentes. Pero si se considera que los continentes estuvieron en ese momento de la historia juntos, con la región de Sudáfrica centrada en el polo sur, estos datos paleoclimáticos podrían ser justificados satisfactoriamente. Valoración actual de la Teoría de Wegener Aunque las primeras ediciones de El origen de los continentes y océanos no despertaron demasiado interés, en 1922 el prestigioso geofísico H. Jeffreys analizó la teoría de Wegener y la calificó de “hipótesis imposible”. Con todo, el momento clave hay que situarlo en el Congreso de la Asociación Americana de Geólogos del Petróleo celebrado en 1926. Asistieron Wegener y Taylor y, con alguna notable excepción, se quedaron solos en defensa del movilismo. La teoría de la deriva continental fue rechazada. Alfred Wegener murió en 1930 durante una expedición a Groenlandia de la que esperaba aportar pruebas definitivas de que América y Europa, aún hoy, se estaban separandoiento de la corteza terrestre hacia el oeste. Ninguna de las dos era suficiente. Resulta sorprendente la cantidad abrumadora y el acierto de los datos y argumentos aportados por Wegener en defensa de la movilidad continental, la mayor parte de los cuales continúan considerándose válidos hoy, si bien el desplazamiento es mucho más lento de lo que suponía el científico alemán. Los desaciertos de su teoría eran básicamente dos: Las causas de los movimientos continentales no son las sugeridas por Wegener. Los fondos oceánicos no constituyen un “tapete” fijo sobre el que se desplazan los continentes, como suponía Wegener. Los fondos oceánicos también se mueven, a veces junto con los continentes y a veces de manera autónoma. No puede extrañar ninguno de estos dos errores, ya que se conocía muy poco acerca del interior terrestre y casi nada sobre los fondos oceánicos. El camino hacia la Tectónica Aunque minoritarias, las ideas movilistas siempre contaron con algún defensor. Uno de los apoyos más destacables lo recibió Wegener un año antes de su desgraciado accidente. Se lo proporcionó Arthur Holmes al indicar que el manto terrestre se encontraba agitado por corrientes de convección. El resurgimiento del movilismo en los años cincuenta está marcado por avances tecnológicos. Así, el sónar permitió conocer mejor la topografía de los fondos oceánicos y descubrir la existencia de un relieve submarino de más de 60 000 km de longitud: la dorsal oceánica. La utilización del ordenador permitió a Bullard en el año 1964 comprobar que el encaje de los continentes era casi perfecto Dorsales océanicas El estudio de los fondos oceánicos deparó notables sorpresas, entre ellas hay tres que destacan: la existencia de la dorsal oceánica, la relativa escasez de sedimentos en los fondos oceánicos y la juventud de esta corteza. El océano Atlántico está recorrido de norte a sur por un relieve submarino que se eleva de 2 a 3 km sobre las llanuras circundantes y que emerge en Islandia, es la dorsal oceánica. Esta dorsal se bifurca hacia el océano Índico y el océano Pacífico, superando los 60 000 km de longitud. La dorsal atlántica tiene un surco central, limitado a ambos lados por fallas normales, que se denomina rift. Otras dorsales, como la del Pacífico, no tienen este rift central. Periódicamente, la dorsal se encuentra interrumpida por fracturas transversales, que se denominan fallas transformantes. Una de las razones por la que los científicos estaban muy interesados en el conocimiento de los fondos oceánicos, es que esperaban hallar, capa sobre capa, materiales depositados a lo largo de ¡toda la histori terrestre! Puesto que en el océano se están depositando materiales continuamente, se pensaba que encontrarían sedimentos, con sus correspondientes fósiles, desde que existen los océanos, es decir, desde hace más de 4000 m.a. Por ello, la ausencia de sedimentos en las dorsales y su relativa escasez en el resto de los fondos, resultó un hallazgo sorprendente. Por otra parte, los sedimentos no se distribuyen homogéneamente, sino que su potencia aumenta a medida que nos alejamos de la dorsal. Al hacer un sondeo en el fondo oceánico, se encuentran diversas capas de sedimentos; los más superficiales son actuales y su antigüedad aumenta con la profundidad. La edad de los más antiguos de cada zona es similar a la de los basaltos situados inmediatamente debajo de ellos; es la edad de la corteza oceánica en ese lugar. Hay tres datos que se repiten regularmente en todas las cuencas oceánicas: – En las dorsales, los basaltos son actuales, es decir, se han formado en el último millón de años. – La antigüedad de los basaltos situados bajo los sedimentos se incrementa al distanciarnos de las dorsales. – Esa edad nunca supera los 180 millones de años. Los sorprendentes datos recogidos generaban muchas preguntas. Así, si hay océanos desde hace al menos 4000 M.a., ¿por qué no se encuentran fondos oceánicos con edades superiores a los 180 M.a.? ¿Qué ha pasado con los fondos más antiguos? ¿Por qué las edades se reducen a medida que nos acercamos a la dorsal? ¿Qué ocurre en la dorsal? Teoría de la Expansión del fondo océanico Durante más de cuarenta años desde la publicación del trabajo de Wegener, la comunidad científica se había mantenido escéptica a la propuesta de la deriva de los continentes. Hacia 1950 se desarrollaron nuevas técnicas que permitieron realizar un detallado estudio del fondo de los océanos. Estos estudios revelaron la existencia de un sistema de cordilleras, denominadas dorsales, que se extendía a lo largo de toda la región central del océano Atlántico. Estas regiones se caracterizaban por una intensa actividad volcánica. Así, en 1960, el geólogo Harry Hess elaboró la Teoría de Expansión del fondo oceánico. En ella proponía que el fondo marino se expande y se mueve simétricamente hacia ambos lados de la dorsal. Es decir, las observaciones geológicas y geofísicas indican que las cordilleras meso-oceánicas o dorsales funcionan como centros donde se genera nuevo piso oceánico conforme los continentes se alejan entre sí. Este útlimo concepto fue propuesto por el geólogo John Tuzo Wilson. En 1965, Tuzo Wilson introdujo el término placa para referirse a grandes fragmentos de litosfera que se movían de forma unitaria. La escasez de sedimentos y la distribución de las edades de los fondos oceánicos no fueron las únicas sorpresas que proporcionó su investigación. En efecto, también se descubrió que los basaltos de la corteza oceánica muestran una llamativa magnetización en bandas paralelas al eje de la dorsal, que alternativamente tienen polaridad normal (la actual) y polaridad invertida. Para explicar el bandeado magnético, F. Vine y D. Matthews elaboraron en 1963 la teoría de la extensión del fondo oceánico, según la cual las dorsales son lugares en los que se genera nueva litosfera oceánica a partir de materiales magmáticos procedentes del interior. La litosfera recién creada se aleja, a uno y otro lado, de la dorsal y el espacio dejado lo ocupa nuevo magma. El fondo oceánico se comporta como una cinta grabadora que registra la orientación del campo magnético terrestre en el momento en que se incorpora cada masa magmática. Además del bandeado magnético, esta teoría permite explicar la actividad volcánica que hay en las dorsales, el incremento de la edad de los fondos oceánicos al alejarse de ellas, así como la distribución de los sedimentos en las cuencas oceánicas. También las características de la actividad sísmica en las dorsales encajan perfectamente en la dinámica propuesta para estas zonas: terremotos de foco somero, de poca magnitud pero muy frecuentes. Por todo ello, constituye una parte esencial de la teoría de la tectónica de placas. Las ideas propuestas por Vine y Matthews se conocen también como teoría de la expansión del fondo oceánico. Sin embargo, hay autores que consideran inadecuado el término “expansión” ya que significa dilatación, o incremento de volumen sin incremento de materia, y no es eso lo que ocurre en los fondos oceánicos. Subducción La subducción de placas es un proceso de hundimiento de una placa bajo otra en un límite de placas convergentes. Generalmente es la litosfera oceánica, de mayor peso específico, la que subduce bajo la litosfera continental, de menor peso específico debido a su mayor grosor cortical. La zona de subducción es una zona larga y estrecha donde una placa desciende por debajo de otra. Dado que la temperatura y la presión aumentan con la profundidad, una parte de los materiales de la placa en subducción son liberados (en especial el agua) lo que conlleva a la fusión del manto, que a su vez, asciende a través de la corteza continental originando erupciones volcánicas. Las zonas de subducción constituyen una parte muy importante dentro de la dinámica de los materiales terrestres. Los materiales subducidos han cambiado posiblemente las propiedades del manto, y permitido que la convección se mantenga. Sismológicamente las zonas de subducción son caracterizadas por las zonas de Benioff. El desplazamiento de una placa con respecto a la otra no es continuo sino que se produce a saltos, como consecuencia de los cuales se generan terremotos. Las zonas de subducción presentan la mayor actividad sísmica del planeta. Los terremotos, atendiendo a la profundidad del foco sísmico, se clasifican en los siguientes tipos: • Someros, si la profundidad es menor de 70 km. • Intermedios, si el foco sísmico está entre 70 y 300 km de profundidad. • Profundos, cuando el foco sísmico se encuentra localizado entre 300 y 700 km de profundidad. La litosfera oceánica que subduce está fría y contiene ciertas cantidades de agua. El rozamiento con la litosfera continental incrementa la temperatura, y el agua baja el punto de fusión de los minerales. Esto permite que se produzca una fusión parcial de los minerales más ricos en sílice, que funden a menor temperatura. Se originan así magmas que alimentarán erupciones volcánicas de andesitas (el nombre de esta roca se debe a que es frecuente en los Andes). Teoría de la Tectónica de placas La Teoría de la tectónica de placas fue forjada principalmente en las décadas de 1950 a 1970 y se le considera la gran teoría unificadora de las Ciencias de la Tierra, ya que analiza una gran cantidad de observaciones geológicas y geofísicas de una manera coherente y elegante. La Geología clásica ofrecía explicaciones independientes para interpretar procesos como el origen de los volcanes, el origen y distribución de los terremotos, la formación de las cordilleras o los cambios en la distribución de tierras y mares a lo largo de la historia de la Tierra. Una de las grandes virtudes de la teoría de la tectónica de placas es que establece relaciones entre todos estos procesos y los incluye dentro de la dinámica global del planeta, razón por la que también es denominada teoría de la tectónica global. En síntesis, la Tectónica de placas señala que la litosfera se mueve, y explica cuáles son las causas de estos movimientos y cuáles son sus consecuencias. Esta teoría puede resumirse en las siguientes ideas básicas: 1. La litosfera se encuentra dividida en un conjunto de fragmentos rígidos denominados placas. Las placas son fragmentos de litosfera cuyo grosor oscila entre 50 y 200 km y poseen una extensión superficial muy variable. La mayoría de ellas contienen litosfera continental y litosfera oceánica. Existen siete grandes placas litosféricas: Euroasiática, Africana, Indoaustraliana, Pacífica, Norteamericana, Suramericana y Antártica. Entre ellas se sitúan una docena de placas de menor tamaño, como las de Nazca, Caribe, Cocos, Arábiga y Filipina. 2. Los límites o bordes de las placas litosféricas pueden ser de tres tipos: – Dorsales, o límites divergentes en los que se genera nueva litosfera oceánica. – Zonas de subducción, o límites convergentes en los que se destruye litosfera. – Fallas transformantes, o límites conservadores en los que no se crea ni se destruye litosfera, sino que se produce un desplazamiento lateral de una placa con respecto a otra. 3. Las placas litosféricas se desplazan sobre los materiales plásticos del manto sublitosférico. Lo hacen a una velocidad variable que oscila entre 1 y 12 cm/año, siguiendo distintas direcciones. Dado que entre las placas no existen huecos, el movimiento de cualquiera de ellas afecta también a las que le rodean y provoca alejamiento o aproximación de continentes e, incluso, colisiones entre ellos. Por esta razón los límites de las placas litosféricas son las zonas de la Tierra en las que existe mayor actividad geológica. En ellos se originan las cordilleras y la mayor parte de los terremotos y volcanes. 4. Los desplazamientos de las placas litosféricas son causados por la energía térmica del interior terrestre ayudada por la energía gravitatoria. Este calor, en parte originado por la desintegración de los elementos radiactivos y en parte procedente del generado durante la formación del planeta, impulsa las corrientes de convección en el interior terrestre que causan el movimiento de las placas. La dinámica que afecta a la superficie terrestre es el resultado de la interacción entre una litosfera fría y rígida y el interior agitado térmicamente. 5. La litosfera oceánica es renovada continuamente, mientras que la litosfera continental tiene un carácter más permanente. La formación de nueva litosfera en las dorsales y su destrucción en las zonas de subducción explica que la antigüedad de los fondos oceánicos sea siempre inferior a 180 millones de años, mientras que en los continentes se han encontrado rocas de casi 4000 millones de años. 6. A lo largo de la historia de la Tierra ha cambiado no solo la posición de las placas litosféricas o su forma y tamaño, sino también el número de estas. La creación de litosfera en las dorsales, su destrucción en las zonas de subducción, junto con procesos de divisiones y uniones de placas son las causas de ello. Continúa habiendo debate en torno al peso relativo de cada proceso natural involucrado en la orogénesis (fuerzas tectónicas, deformación de la litosfera, erosión y transporte de sedimento, clima, magmatismo, etc.) en determinar la estructura actual de los orógenos. Desde finales de los años 90, por ejemplo, se ha desarrollado la idea de que el crecimiento del orógeno y su deformación interna es sensible a la distribución superficial de la erosión, controlada por el clima, pero no existe aún consenso sobre la relevancia de este efecto. Bordes de placas En los bordes o límites de las placas se concentra actividad sísmica, volcánica y tectónica. Se consideran tres tipos de bordes, aunque en determinadas circunstancias, se forman zonas de límite o borde, donde se unen tres o más placas formando una combinación de los tres tipos de bordes. • Divergentes: son límites en los que las placas se separan unas de otras y, por lo tanto, emerge magma desde regiones más profundas. En la mayoría de los casos son constructores del fondo oceánico y de las dorsales, por ejemplo, la dorsal mesoatlántica, formada por la separación de las placas de Eurasia y Norteamérica y las de África y Sudamérica. • Convergentes: son límites en los que una placa chocan entre sí. Son también conocidos como "bordes activos o destructivos". Si chocan dos placas océanicas, o una océanica con una continental, se forma una zona de subducción (la placa oceánica se hunde bajo de la placa continental o de la otra océanica). Las fosas tectónicas son grandes depresiones del terreno, que se forma en el lugar donde la placa entra en subducción. La más profunda es Las Marianas, al noroeste del océano Pacífico. También se forman volcanes que emergen sobre la superficie del océano, como una cadena de islas. Si chocan dos placas continentales, no se se produce subducción. Los materiales que componen las placas no pueden penetrar en la astenosfera y por lo tanto ambas quedan “a flote”, colisionando hasta fusionarse completamente y convertirse en un único bloque. El resultado es la formación de un cinturón orogénico (al chocar y comprimirse las placas). • Transformantes: son límites donde los bordes de las placas se deslizan una con respecto a la otra a lo largo de una falla de transformación. Los materiales calientes del interior terrestre no solo se mueven, desplazando así las placas litosféricas, sino que salen al exterior en unos lugares llamados dorsales oceánicas y hacen que las placas incrementen su tamaño. Como la superficie terrestre es limitada, si unas placas incrementan su tamaño otras lo pierden introduciéndose en el interior terrestre en las denominadas zonas de subducción. Así, los bordes de las placas litosféricas, o límites que separan unas de otras, pueden ser de tres tipos: El movimiento de las placas Una de las posibles causas del movimiento son las corrientes de convección del manto. Éstas transportan material caliente desde zonas internas hacia las capas más externas, donde el material comienza a desplazarse lateralmente, se enfría, desciende y comienza nuevamente el interminable ciclo. Otro modelo considera la formación de plumas. Estas son largas columnas verticales de materiales calientes, que ascienden desde el límite núcleo-manto por tener menos densidad. Algunas plumas se encuentran en las dorsales oceánicas, otras, en el medio de las placas, debajo de océanos y continentes. Las plumas tienen un lugar fijo, y cuando las placas litosféricas pasan por encima de ellas pueden generar la deformación de la corteza. Si bien las plumas no se han observado directamente, existen evidencias indirectas de su existencia. Los científicos han identificado numerosos puntos calientes, zonas de la superficie terrestre que se encuentran por encima de las plumas. Gracias a nuevas tecnologías que utilizan rayos láser y satélites se han podido realizar mediciones precisas de distancias entre puntos situados en placas diferentes, constatándose que las placas litosféricas se mueven. Así, entre Londres y Nueva York se han registrado desplazamientos de 1,7 cm/año y de 4,6 cm/año entre Hawai y Australia. No hay duda de que el calor del interior terrestre tiene un papel determinante en la dinámica litosférica. Si el interior estuviese frío no habría tectónica de placas. Por eso se dice que la Tierra es una máquina térmica. Si bien la energía gravitatoria parece desempeñar también un papel clave. Sin embargo, los científicos tienen aún ciertas dudas sobre el papel que desempeña cada uno de los procesos que intervienen y el tipo de interacciones que se dan entre ellos. Las ideas más aceptadas son las siguientes: La energía térmica del interior terrestre genera corrientes de convección en el manto sublitosférico y constituye la causa que “pone en marcha” el movimiento de las placas. Sin embargo, dado que los materiales del manto se encuentran en estado sólido, los movimientos de convección son complejos y no responden al modelo idealizado de celdillas de convección perfectamente cerradas. En algunas zonas de la base del manto, en la capa D’’, se originan columnas ascendentes de materiales muy calientes que pueden alcanzar la superficie. Son los puntos calientes, como Islandia y las islas Hawai. Estas columnas ascendentes forman parte del proceso convectivo general del manto. Las placas litosféricas se ven arrastradas por los movimientos convectivos del manto que, en todo caso, son muy lentos, entre 1 y 12 cm/año. La gravedad desempeña también un papel importante entre las causas del movimiento de las placas. Y lo hace a través de dos mecanismos complementarios: – La mayor altura de la dorsal favorece el deslizamiento hacia abajo del fondo oceánico. – La litosfera subducida es densa y fría y las presiones reinantes en el interior del manto la hacen aún más densa. Así el extremo de la placa subducida tira de ella y la arrastra. El proceso es comparable a lo que le ocurre a una toalla que tiene una parte sumergida en el agua: a medida que se empapa pesa más y arrastra al resto de la toalla hacia abajo. Por esta razón, al tirón motivado por la densificación de la placa subducida se le conoce como “efecto toalla”. Ciclo supercontinental En primer lugar hay que suponer que los procesos característicos de la tectónica global arrancaron en un determinado momento de la evolución planetaria, y que no se han mantenido totalmente uniformes en su intensidad y, hasta cierto punto, su mecánica. De la misma manera sabemos que, a medida que el planeta va perdiendo energía interna, por su disipación como calor en el espacio, las fuerzas necesarias para la orogénesis se van debilitando. Los procesos volcánicos provocan la desgasificación del interior, que va perdiendo agua y otros volátiles, lo que da lugar a una progresiva evolución de sus propiedades mecánicas hacia una mayor rigidez; a la larga los procesos geotectónicos mayores, como la expansión oceánica y la orogénesis, tendrán que detenerse. El ciclo supercontinental (o ciclo de Wilson), propuesto por John Tuzo Wilson por postula que cada 400-500 millones de años todas las masas de tierra emergidas se unen, formando un gran supercontinente. Hace 250 millones de años los continentes actuales estaban unidos en un supercontinente, la Pangea de Wegener, que suele denominarse Pangea 2 para diferenciarla de otra que existió hace 650 millones de años, Pangea 1. Si se considera que los continentes se desplazan en una superficie esférica, su colisión y, por tanto, su unión se hacen inevitables. Pero ¿por qué se disgrega un supercontinente? Para explicarlo, debe recordarse que la litosfera continental es más gruesa que la oceánica y no dispone de zonas, como las dorsales, a través de las cuales evacuar el calor del interior terrestre. Un supercontinente funciona como una enorme “manta” que dificulta la salida tanto del calor generado por la desintegración de los elementos radiactivos como del evacuado desde el núcleo. La acumulación de calor bajo el supercontinente hace que se eleven determinados lugares de este y la litosfera se adelgace y fragmente por ellos, originándose un rift que dará lugar a un océano interior. El océano se extenderá y los continentes se separarán hasta que la formación de zonas de subducción invierta el proceso. La fragmentación y el reagrupamiento de los continentes serían las fases principales de un ciclo conocido como ciclo de Wilson que, según algunas hipótesis, se repetiría cada 400 ó 500 millones de años. De tal manera, Pangea 1 no habría sido el primer supercontinente, sino que habrían existido otros hace 1100, 1600 y 2100 millones de años. Conviene aclarar que, si bien existe un acuerdo mayoritario acerca de que a lo largo de la historia de la Tierra se han producido numerosas uniones y divisiones continentales, no todos los científicos consideran que este proceso haya ocurrido siguiendo ciclos regulares, ni que los supercontinentes hayan sido en todos los casos pangeas que integrasen casi toda la corteza continental del momento. Tanto si los ciclos de Wilson se han repetido de manera regular como si no, los continentes muestran numerosas “cicatrices” o suturas, resultado de otras tantas colisiones, como por ejemplo la cordillera de los Urales, entre Europa y Asia. Antes de Pangea, debió de haberse formado un supercontinente llamado Pannotia, a fines del Proterozoico, durante el período Véndico (hace unos 800 millones de años). Su desintegración y los consecuentes choques obductivos entre placas se relacionan como la orogenia hercínica, de gran importancia en la formación de los relieves más antiguos del planeta. Surgimiento y transformación de supercontinentes y continentes La Paleogeografía es el estudio de la distribución de los continentes y los océanos a lo largo de las eras geológicas. Sobre la base de toda la evidencia disponible, los paleogeógrafos han determinado la distribución continental y océanica en otras épocas y han llegado a conclusiones muy interesantes. Se conocen ciclos de unos trescientos millones de años, aproximadamente, en los que los continentes se fragmentaron y volvieron a unirse en supercontinentes. Los más conocidos en la historia geológica fueron Rodinia, que se formó hace unos mil millones de años, durante el eón Proterozoico, y Pangea, que se formó hace trescientos millones de años, al final de la era Paleozoica, y que, poco después, durante el Jurásico, se dividió en Gonwana y Laurasia. Por otra parte, hubo épocas de fragmentación continental extrema, verdaderos “mundos marinos”, como desde el Proterozoico superior al Ordovícico, cuando existían numerosos continentes islas como Báltica, Siberia, Laurentia, Sinocorea, etc., y uno mayor, Nigritia. También sabemos que en todos los continentes actuales hay sectores que antes estaban muy alejados entre sí. Por ejemplo, la Argentina está formada por fragmentos que vienen de la Antártida, como el sur patagónico, fragmentos que provenían de Laurentia, como la Puna y la Precordillera, y otros sectores que ocupaban posiciones más alejadas. Durante el amplio intervalo de tiempo geológico denominado eón Hadeano no se registra la formación de continentes ni océanos extensos. 1. Hace 4.300 m.a. se inició la formación de los cratones o núcleos continentales, entre los cuales el más primitivo, Yilgarn, corresponde a Australia occidental, y al cual pronto se sumarían otros como Kalahari, Arqueoplata (escudo del Rìo de la Plata), Arqueosiberia y Arqueogroenlandia. La única evidencia de vida que puede asociarse a esta época estaría formada por virus de ARN y luego “metacélulas y eocélulas”. 2. Las primeras rocas sólidas de estos núcleos arqueogondwánicos y arqueolaurénticos emergieron hace 4.005 m.a., al cesar el bombardeo meteórico..Al iniciarse el Arqueano (Paleoarqueano), hace 3.800 m.a., continuó la deposición de rocas sólidas y se formaron dos nuevos protocontinentes mayores, Kapvaal (equivalente a Sudáfrica) y Pilbara (en Australia). Esta época concuerda, aproximadamente, con el surgimiento de LUCA (el ancestro universal) y, luego, de las primeras eubacterias (las bacterias verdes no azufrosas o cloroflexas). 3. Hace 3.600 m.a., se formó el más antiguo supercontinente 1. Vaalbara, por el agregado de los núcleos arqueogondwánicos y arqueolaurénticos mencionados en 1. y la expansión continental que agregó corteza a través de los núcleos arqueogondwánicos y arqueolaurénticos. Hace 3.450 m.a. comenzó la fragmentación de Vaalbara en bloques continentales (primera evidencia de fragmentación continental) surgiendo el cratón de Zimbawe, y poco después el cratón Komatii. Durante esta época dilatada de la historia de la Tierra, probablemente se agregaron las radiobacterias o bacterias resistentes a las radiaciones, otro grupo de bacterias. Vaalbara 4. Al promediar el Arqueano, hace 3.100 m.a., se formó el supercontinente 2. Ur. A continuación, casi al final de dicho eón, hace 2.800 m.a., se agregaron dos nuevos núcleos continentales independientes, Árctica y Protobáltica y continuó la acreción continental global de Ur. La fecha aproximada corresponde con las primeras evidencias de las arqueas, sobre la base de los marcadores de metano, y la segunda con las cianobacterias, sobre la base de los metilhopanoides y la formación más abundante de estromatolitos o depósitos de carbonato de calcio, según modelos bandeados. Ur. 5. Hace 2.600 m.a. (finalizaciòn del Arqueano o Neoarqueano), Ártica se fusionó con Ur y conformó el nuevo supercontinente 3. Kennorlandia. En su interior se formó una importante pluma tectónica magmática que se mantuvo activa durante el comienzo del eón Proterozoico inferior (Paleoproterozoico), hasta hace 2.500 m.a. Hace 2.450 m.a., comenzaron a separarse los cratones Kola y Karelia. Hacia los 2.300 m.a. se agregaron los continentes de Protolaurentia, Protobáltica, Kalahari y Australia occidental. Se especula que en esta fase temprana debieron haber divergido las primeras firmibacterias, Gram positivas (bacilos y clostridios). Kennorland. Comparación de Kennorland (a) con los supercontinentes posteriores, Columbia (b) y Rodinia (c) 6. Hace 2.200 m.a. se formó un gran núcleo septentrional, supercontinente 4. Atlántica (que representaría el núcleo protolauréntico principal) aunque se mantuvieron separadas Protobáltica y Protolaurentia. Hace 2.100 m.a. se agregaron otros cratones menores, como Amazonia, Protosiberia, Ucrania y Mongolia. Para la misma época surgieron también al N. las placas de Ártica y Antártica oriental En esta etapa continuó la radiación de las cianobacterias, y se hizo mayor la oxigenación de los océanos, que culminó con los depósitos de hierro ferroso. Por la misma época surgieron las proteobacterias (bacterias púrpura) y las chlorobias (bacterias verdes azufrosas). Como resultado de toda esta actividad tectónica y biológica, se produjo la Snowball Earth paleoproterozoica (el primer gran antecedente de la glaciación global). 7. Hace entre 2.000 y 1.900 m.a. Atlántica se fusionó con Protobáltica (todavía libre) y con las placas de Ártica y Antártica oriental) para dar lugar al supercontinente 5. Nena, si bien todavía se mantuvieron fragmentados los núcleos protogondwánicos (Amazonia), Ucrania y Protolaurentia. Durante esta época se conocen las primeras evidencias de ecuariotas (los gripánidos) y surgen nuevos tipos de cianobacterias, y probablemente entre ellas los procloròfitos (solo con clorofilas pero sin ficobilinas, a diferencia de las cianobacterias). Nenna. 8. Finalmente, hace 1.800 a 1.750 m.a., Nena se fragmentó en varias microplacas incluidas entre las principales Congo, Mato-Grosso, Escandinavia, Protosiberia y Protochina Esta época concuerda con el surgimiento de las primeras algas rojas y nuevos tipos de eucariotas fósiles (tawuídos y chuáridos). 9. Al promediar el Proterozoico, entre 1.750 y 1.600 m.a. se volvió a formar un supercontinente 6. Columbia, Hudsonia o Nuna, que además del antiguo núcleo reunió los fragmentos que permanecieron libres como Ucrania, Amazonia. Kalahari, Protosiberia, y Protolaurentia, aunque quedó ligeramente separado el núcleo de Zahertai (China N.). Con tan heterogénea composición no es de extrañar que en este supercontinente quedara la porción oriental de India unida al occidente de Norteamérica, en una extensión máxima de 4.900 km. Probablemente surgieron en esta época las primeras algas verdes (prasinófitos) y los protohongos (Tappania). 10. Hace entre 1.500 y 1.300 m.a., se fue produciendo la fragmentación de Columbia, con la formación de un rift y la separación entre los sectores de India y Norteamérica occidental para formar el cratón Purcell (Mesolaurentia), India oriental (Mahanadi), Mesobáltica y Rifeica, que se agregaron a las todavía independientes desde su formación, Zhaertai (China N.) y Kalahari. Esta época concuerda con la glaciación mesoproterozoica y la evidencia de los primeros dinofagelados. 11. Entre 1.100 y 1.000 m.a. se formó un nuevo supercontinente a partir de las placas derivadas de Columbia, el supercontinente 7. Rodinia o Protopangea, bañada por el gran océano Mirovia, y si bien se unieron la mayor parte de las placas independientes, todavía permaneció separado aun el pequeño núcleo continenental de Zaherti (China N.) y se agregó Indochina. Esta época marca la probable radiación de los coanoflagelados, los cercozoos (testamebas filosas) y los amebozoos (tecamebas o testamebas lobosas). Posición de masas continentales en Rodinial. 12. Entre los 900 y 750 m.a., Rodinia comenzó a fragmentarse en varios (hasta ocho) microcontinentes: Neoamazonia, Namibia, Prelaurentia, Presiberia, Prebáltica, Protogondwana y Welsworth-Whitmore o Antártica occidental. Como resultado de esta fragmentación se diferenciaron tres grandes océanos meridionales: Adamastor, Brazilides y Mozambique. En esta época se registran las primeras algas doradas (somatocitos) y también los primeros animales (poríferos primitivos). Fragmentación de Rodinia. 13. Hace 750 millones de años, se formaron dos grandes sectores continentales, uno alrededor del Polo Norte, 8. Neoártica, y otro en el Polo Sur, 9. Prenigritia, y pronto se formó una masa menor, en posición central, Congo o Centroáfrica. En este momento de la historia terrestre se produjo la mayor glaciación global, la Varangiana (neoproterozocia), la segunda o tercera Snowball Earth (Tierra bola de nieve). Probablemente surgieron por entonces los primeros ancestros de los radiales (cnidarios) y vendozoos (relacionados con los ctenóforos y placozoos). 14. Hace 650 millones de años, al final del Proterozoico, el núcleo de Protolaurasia (Neoártica) rotó al hacia el polo sur y Protogondwana (Prenigritia) hacia el polo Norte en sentido de las agujas del reloj, fusionándose en un movimiento de pinzas con Congo. Como resultado se formó el nuevo supercontinente 10. Pannotia. Este supercontinente migró hacia el ecuador y quedó bañado por el océano Panafricano, probable antecedente del Panthalassa paleozoico. Esta época concuerda con la radiación de los radiales (cnidarios) y la aparición de los primeros bilaterales (protostomados), los lofotrocozoos. Pannotia. 15. Pannotia tuvo breve duración, ya que su fragmentación se fue completando prematuramente hasta hace 550 m.a., un poco antes de los comienzos del Cámbrico. Entonces se formaron en posición ecuatorial: Laurentia, Báltica, Siberiana, Kazhakstana, Sinocorea, Centrochina, China sepentrional y, el mayor, Nigritia, formándose el océano Iapetus y el océano Rheico, así como ell nuevo océano Panthalassa, heredero del Panafricano. Por entonces, la península de Florida, Puna y Precordillera constituían placas independientes, insulares. Esta época concuerda con la aparición de radiolarios, foraminíferos y los primeros ecdizoos y deuterostomados, entre los bilaterales. A este período se lo conoce como el Carnaval de los Animales debido a la intensa radiación evolutiva que dio origen a numerosos phyla, la mayoría extintos, así como todos los grupos con representantes actuales.. Fragmentación de Gonwana. Separación de Nigritia (Protogondwana). 16. Durante el Ordovícico se agregó una nueva placa, Avalonia, separada de Larurentia por el mar de Tornquist, mientras que ambas quedaron separadas de Nigritia por el océano Rheico que se ensanchaba. También se formaron nuevos océanos: Prototethys, separando Báltica, Norchina y Siberia de Nigritia, y el Protouraliano, separando Siberia de Kazakhstania. Iapetus continuó como un océano intermedio entre Laurentia y Báltica. Esta época concuerda con la aparición de los briozoos y la radiación de gasterópodos y pelecípodos junto con los ostracodermos (peces acorazados sin mandíbulas),, mientras en los continentes se agregaban los musgos y ascomicetes además de los oligoquetos o lombrices. 17. Durante el Silúrico, mientras México e Irán-Tíbet, así como Afganistán formaron pequeñas placas independientes, el océano Protoetethys se ensanchó, y el Iapetus y el Rheico comenzaron a cerrarse. Avalonia quedó soldada a Báltica luego del cierre del Mar de Tornquist, y China septentrional se unió a Nigritia. En el mar se incorporaron los acantodios, junto con los actinopterigios y condrictios (tiburones primitivos), mientras en el agua dulce prolieraron los ostracodermos. Las primeras plantas vasculares (riniófitos) se incorporaron en los continentes, junto con los primeros escorpiones terrestres. 18. Para el Devónico, hace 300 m.a., Laurentia colisionó con Báltica y se formó el supercontinente menor 11. Laurrusia, cerrándose definitivamente el Iapetus, mientras que el Rheico profundizó su cierre progresivo, acercando a este continente con Nigritia. Donde se ubicaba el Prototethys, se formó ahora el Paleotethys, meridional. Esta época es de intensa radiación evolutiva continental, con la invasión de los condrictios al agua dulce y el predominio de los peces con aletas lobuladas (sarcopterigios), la incorporación de los primeros anfibios (ictiostegalios), los primeros hexápodos (colémbolos), los primeros arácnidos verdaderos (seudoescorpiones y ácaros) y las primeras gimnospermas o plantas con semillas, las pteridospermas. 19. En el Carbonífero, el cierre del océano Rheico fusionó Laurrusia con Nigritia y se forma entonces el supercontinente menor 12. Micropangea, antecedente de Pangea, mientras que al N., Euramérica formó una placa continental independiente, en tanto que Norchina, Kazkhastania y Siberia se aproximaron para formar otro supercontinente menor 13. Angara. Esta época contó con extensos bosques pantanosos que resultaron importantes depósitos de carbón, formados especialmente por licófitos y equisetófitos gigantes, además de los helechos y gimnospermas que comenzaron a ser abundantes, con la incorporación de filicófitos o helechos verdaderos y las primeras coníferas. Otro evento crucial fue la aparición de los amniotas, mejor adaptados al ambiente aeroterrestre, tanto parareptiles (anápsidos) como los primeros eureptiles con forma de lagartos,los primeros lisanfibios (anuros), pero especialmente notable fue la incorporación de los primeros insectos, inicialmente los tisanuros no alados y luego diversos grupos de insectos alados como la gigantesca Meganeura, así como las arañas y otros grupos entre los arácnidos. 20. Finalmente durante el Pérmico, al colisionar Angara con Euramérica y Micropangea, se formó al gran supercontinente 14. de Pangea, el propuesto por Wegener para su teoría de la deriva continental. En esta época los boques retrocedieron, aunque en zonas intermedias se incorporaron nuevos grupos de plantas como los ginkgófitos y cidacófitos y los sinápsidos (ancestros de los mamíferos) así como los primeros insectos holometábolos con metamofosis completa, los coleópteros (escarabajos).Hacia el final de la época se produjo una gran extinción (casi el 95% de toda la fauna y la flora terrestres y marinas. La Pangea. 21. Durante el Triásico, continúo Pangea aunque en proceso de fragmentación transversal, y hacia el final, la apertura definitiva del océano Tethys, que reemplazó al Paleotethys, dividió a la gran masa continental en dos, el supercontinente 15. Laurasia al N. y el supercontinente 16. Gondwana al S, heredero de Nigritia. Ahora el gran océano comenzó a denominarse Pacífico, como sucesor de la Panthalassa paleozoica. Esa época habrá de marcar la incorporación de importantes novedades: las madréporas o corales típicos (escleractinios), los belemnoideos (ancestros de sepias y calamares), los peces teleósteos entre los actinopterigios, los primeros reptiles marinos, como los placodontos y notosaurios,los dípteros (moscas y mosquitos), los primeros pterosaurios o reptiles alados, los primeros dinosaurios, las tortugas o quelonios, los primeros saurios o lagartos y los primeros mamíferos verdaderos, entre otros. 22. Durante el Jurásico, se formó el océano Atlántico central, al alejarse África de Norteamérica, y se separó como supercontinente menor 17. Mesoasia del resto de Larurentia, que formó el otro supercontinente menor 18. Neolaurentia. Al sur, comenzó la separación progresiva entre el supercontinente menor 19. Gondwana oriental, mientras África se separaba de Sudamérica en 20. Gondwana occidental. Las novedades evolutivas más importantes de esta época incluyen la aparición de los primeros mamíferos marsupiales, de las primeras aves (Archaeornithes), de varios grupos de dinosaurios como los estegosaurios y anquilosaurios, de los urodelos o salamandras entre los lisanfibios, los ofidios o serpientes y varios grupos de reptiles marinos incluidos los plesiosaurios.Pero tal vez la incorporación más notable incluyó a las primeras angiospermas o plantas con flores y frutos, con uno de los primeros fósiles, Archaeofructus También surgieron los lepidópteros (mariposas y polillas). 23. En el Cretácico inferior, se abrió el Atlántico sur al separarse África de Sudamérica. Luego, comenzó a formarse el océano Índico al fragmentarse Gondwana oriental en India, Madagascar y Paleantártica (Antártica más Australia, Nueva Guinea y Nueva Zelandia). Una gran lengua marina, el mar de Sundance, fragmentó al sector de Norteamérica en dos, y Mesoasia se unió al extremo occidental de ese continente. En esta época se incorporaron los primeros mamíferos placentarios, los mosasaurios entre los reptiles marinos y las cecilias o ápodos entre los mamíferos,y el grupos de las Enaliornithes entre las aves, mientras se producía una intensa radiación de las angiospermas con la incorporación de las rósidas. 24. Al finalizar el Cretácico, quedó formado entonces un nuevo supercontinente menor 21. Neogondwana, resultante de la unión de Paleantártica y Sudamérica, mientras ésta se separaba de Norteamérica. África quedó dividida a su vez en dos placas, y en el sector septentrional se formó el Archipiélago europeo, un gran conjunto de islas separadas. Esta época marcó la evolución final de los dinosaurios, con la aparición de tiranosaurios y ceratopsios, de las primeras aves modernas (Neornithes), de los ancestros de los primates (los plesiadápidos) y de las primeras angiospermas tubifloras (labiadas y otras familias). Igualmente se incorporaon los himenópteros (hormigas, avispas, abejas). 25. En el Terciario inferior o Paleógeno, las masas continentales se separaron cada vez más, y esto se hizo más acentuado en el sector gondwánico o meridional. De esta manera, Sudamérica quedó separada de Norteamérica y unida a Antártica, posición que mantendría hasta finales del Oligoceno. Nueva Zelandia y Australia se separaron definitivamente del resto de Neogondwana (es decir, Sudamérica y Antártica). En cambio, Asia y Norteamérica (supercontinente 22. Asiamérica) por un lado, y Norteamérica y Europa por el otro (supercontinente 23. Neoeuramérica), quedaron unidas entre sí, aunque separadas por el océano Uraliano (donde hoy están los montes Urales). Hacia fines del Oligoceno, India colisionó con Asia y se levantaron los montes Himalaya. Esta época asiste a una intensa radiación de mamíferos, aves y angiospermas, y de los teleósteos en el mar. Surgen entonces los quirópteros o murciélagos, las primeras ballenas (cetáceos), los primeros pájaros (passeriformes), los colibríes o trochílidos polinizadores y los ancestros de los hómínidos. 26. En el Terciario superior, la distribución continental cada vez se hizo más parecida a la actual. Europa se unió a Asia al cerrarse el océano Uraliano, y en cambio, Norteamérica y Groenlandia quedaron separadas de Europa formándose el Atlántico Norte y también el océano glacial Ártico. De esta manera, quedó finalmente formado el supercontinente moderno 24. Eurasia. 27. Hacia 20 m.a., Antártica se separó de Sudamérica, que terminaría colisionando con Norteamérica a medida que se formaba el puente centroamericano en el Plioceno, hace 15 m.a. Así se formó el supercontinente moderno 25. América. Finalmente, también se elevaron los Andes en Sudamérica y, hace 5 m.a., se formó un nuevo supercontinente moderno 26. Afroeurasia. Para esta época surgieron los ancestros directos del hombre, los autralopitecinos, los bosques retrocedieron frente a sabanas y praderas con la incorporación de gramíneas o pastos y de compuestas. 27. Desde el comienzo del Cuaternario, la posición continental y oceánica resultaba similar a la actual, aunque grandes masas de hielo continental ocupaban los continentes septentrionales y amplias zonas de los meridionales como Nueva Zelandia, Australia y Patagonia. Finalmente, con la retirada de los hielos, en el Holoceno, el mundo adquirió su perfil geográfico actual. Esta época asistió a intensas glaciaciones seguidas por períodos de retirada del hielo, lo que originó la evolución final del género Homo (con diferentes especies) que convivieron con mamíferos gigantescos como mamuts y tiges dientes de sable. 28. Dentro de 60 m.a. (en el futuro), Australia colisionará con el este de Asia y se creará un nuevo cinturón de montañas comparable a los Himalayas. Por entonces se formará el supercontinente futuro 27. Autralo-afro-eurasia. 29. Dentro de 130 m.a. en el fuituro se formará un conjunto mayor, el supercontinente futuro 28. Australo-antártico-afro-eurasia, al chocar Antártida con esa masa continental. 30. Entre 250 y 400 m.a. en el futuro se habrá de formar el supercontinente futuro 29. Neopangea o Amasia, que será el último gran conjunto continental predicho por la Teoría de la Tectónica de placas. El oxígeno se ha incrementado en forma incesante en la atmósfera de la Tierra Los continentes se han movido en forma irregular sobre la superficie terrestre, aunque algunos cambios físicos tales como el incremento en el oxígeno atmosférico, han sido mayormente unidireccionales. La atmósfera de la Tierra primitiva probablemente contenía poco o nada de gas oxígeno (O 2). El incremento del oxígeno atmosférico se produjo en dos pasos hace más de mil millones de 7años. El primero tuvo lugar aproximadamente hace 2.400 millones de años, cuando ciertas bacterias evolucionaron la capacidad de utilizar agua como fuente de iones hidrógeno para la fotosíntesis. Mediante la descomposición química del H2O, estas bacterias generaron O 2 atmosférico como producto de desecho. También produjeron electrones disponibles para reducir el CO 2 para formar compuestos orgánicos. Un grupo de bacterias generadoras de oxígeno, las cianobacterias, formaron estructuras similares a rocas llamadas estromatolitos, que se preservan en forma abundante en el registro fósil. Las cianobacterias aún forman estromatolitos en unos pocos sitios salados del planeta. Las cianobacterias liberaron suficiente O 2 para abrir el camino a la evolución de las reacciones de oxidación como fuente de energía para la síntesis del ATP. Su capacidad para descomponer el agua sin duda contribuyó a este extraordinario acontecimiento. Por lo tanto, la evolución de la vida cambió irrevocablemente la naturaleza física de la Tierra. Dichos cambios físicos, a su vez, influyeron en la evolución de la vida. Cuando apareció por primera vez en la atmósfera, el oxígeno resultó venenoso para los procariotas anaerobios que habitaban la tierra en ese tiempo. Aquellos procariotas que evolucionaron la capacidad de metabolizar el O 2 no sólo sobrevivieron, sino que también ganaron una serie de ventajas. El metabolismo aerobio avanza a velocidades mayores y acumula energía en forma más eficaz que el metabolismo anaerobio (véase Sección 7.5). En consecuencia, los organismos con metabolismo aerobio reemplazaron a los anaerobios en la mayoría de los ambientes de la Tierra. Una atmósfera rica en O2 también hizo posible la existencia de células más grandes y de organismos más complejos. Los pequeños organismos acuáticos unicelulares pueden obtener suficiente O 2 mediante difusión simple aún cuando las concentraciones de O2 sean muy bajas. Los organismos unicelulares más grandes presentan una relación superficie-volumen menor. Para lograr obtener suficiente O2 por difusión simple, deben vivir en un ambiente con una elevada concentración relativa de O2. Las bacterias pueden prosperar con un 1% de los niveles del O2 atmosférico normal; las células eucariotas requieren niveles de oxígeno que son por lo menos de 2 o 3% de las concentraciones atmosféricas usuales. (Para que las concentraciones de O 2 disuelto en los océanos alcancen dichos niveles, se requieren concentraciones atmosféricas mucho más elevadas). Probablemente debido a que tomó muchos millones de años para que la Tierra desarrollara una atmósfera oxigenada, sólo los procariotas unicelulares vivieron en la Tierra durante más de dos mil millones de años. Hace aproximadamente mil millones de años (1.000 m.a.), las concentraciones de O2 atmosférico se volvieron suficientemente elevadas para que las grandes células eucariotas florecieran. Los incrementos ulteriores de los niveles de O 2 atmosférico hace 700 a 570 millones de años (m.a.), permitieron que los organismos multicelulares evolucionaran. En contraste con este cambio mayormente unidireccional en las concentraciones de O2 en la atmósfera, la mayoría de las condiciones físicas terrestres han oscilado como respuesta a los procesos internos planetarios, tales como la actividad volcánica y la deriva continental. Los sucesos extraterrestres, tales como las colisiones con meteoritos, también han dejado su marca. En algunos casos, como veremos en este capítulo, dichos sucesos causaron extinciones masivas, durante las cuales una gran proporción de especies vivientes en ese tiempo desaparecieron. Luego de cada extinción masiva, la diversidad de la vida volvió a florecer, aunque la recuperación tomó millones de años. El clima de la Tierra ha cambiado entre condiciones cálidas y húmedas, y frías y secas A través de gran parte de su historia, el clima de la Tierra resultó considerablemente más cálido que en la actualidad, y las temperaturas disminuían más gradualmente hacia los polos. En otros momentos, sin embargo, la Tierra fue más fría que hoy en día. Grandes áreas fueron cubiertas con glaciares hacia finales del Precámbrico y durante partes de los períodos Carbonífero y Pérmico. Estos períodos de enfriamiento se hallan separados por largos intervalos de climas más moderados. Para que la Tierra permaneciera en un estado frío y seco, los niveles de CO2 debieron haber sido inusualmente bajos, si bien los científicos no saben que causó estas bajas concentraciones. Debido a que estamos viviendo en uno de los períodos más fríos en la historia de la Tierra, es difícil para nosotros imaginar los climas moderados que se encontraban en altas latitudes durante gran parte de la historia terrestre. Por ejemplo, durante el período Cuaternario, se ha dado una serie de avances glaciales, separados por intervalos interglaciales más cálidos durante los cuales se retiraron los glaciares. Si bien el tiempo meteorológico a menudo cambia rápidamente, los climas generalmente cambian con lentitud. Sin embargo, los cambios climáticos principales han tenido lugar durante períodos tan breves como de 5.000 a 10.000 años, principalmente, como resultado de cambios en la órbita terrestre alrededor del Sol. Unos pocos cambios climáticos han sido incluso más rápidos. Por ejemplo, durante el período interglacial del Cuaternario, el océano Antártico cambió de estar cubierto de hielo a estar libre de él en menos de cien años. Tales cambios rápidos generalmente son causados por variaciones repentinas en las corrientes oceánicas. Algunos cambios climáticos han sido tan rápidos que las extinciones causadas por ellos resultan “instantáneas” en el registro fósil. En la actualidad estamos viviendo en un tiempo de rápido cambio climático causado por una acumulación del CO 2 atmosférico, principalmente por La utilización de combustibles fósiles. La concentración normal de CO 2 atmosférico es mayor que en cualquier otro momento desde hace miles de años, excepto por un intervalo cálido de hace 5.000 años, cuando el nivel pudo haber sido ligeramente superior al actual. Una duplicación de la concentración de CO 2 atmosférico, que puede tener lugar durante el presente siglo a menos que se realicen esfuerzos mayores para reducir el consumo humano de combustibles fósiles, probablemente incrementaría la temperatura promedio de la Tierra, causaría sequías en las regiones centrales de los continentes, incrementos de las precipitaciones en las áreas costeras, derretimiento de los glaciares y las calotas de hielo, y resultar en aumentos del nivel del mar que inundaría las ciudades costeras y las tierras agrícolas vecinas. En cuanto a la mayoría de las erupciones volcánicas producen sólo efectos locales y de corto plazo, pero las grandes erupciones volcánicas han tenido consecuencias fundamentales para la vida. La colisión continental durante el período Pérmico (hace aproximadamente 275 m.a.) para formar una única masa de tierra gigantesca, llamada Pangea, causó erupciones volcánicas masivas. La ceniza arrojada por los volcanes hacia la atmósfera redujo la penetración de la luz solar en la superficie terrestre, disminuyendo las temperaturas, reduciendo la fotosíntesis y provocando una glaciación gigantesca. Las erupciones volcánicas masivas también tuvieron lugar en los continentes que se separaron durante el período Triásico tardío y también al final del Cretácico. Los eventos extraterrestres han inducido cambios en la Tierra Al menos 30 meteoritos del tamaño entre pelotas de béisbol y fútbol, golpean la Tierra cada año. Las colisiones con meteoritos más grandes son raras, pero los meteoritos grandes probablemente fueron responsables de varias extinciones masivas. Varios tipos de evidencia señalan estas colisiones. Sus cráteres y las rocas drásticamente desfiguradas que resultaron de esos impactos se encuentran en diversos sitios. Los geólogos han descubierto también gigantescas moléculas que contienen helio y argón atrapados en proporciones características de meteoritos y que son muy diferentes a las presentes en la Tierra. Un meteorito causó o contribuyó a la extinción masiva a fines del período Cretácico (hace 65 m.a.). La primera pista de que un meteorito fue el responsable provino de la concentración anormalmente elevada del elemento iridio en una fina capa que separa las rocas depositadas durante el Cretácico de aquellas depositadas durante el Terciario. El iridio es abundante en algunos meteoritos aunque resulta excesivamente raro en la superficie terrestre. Los científicos a continuación descubrieron un cráter circular de 180 km de diámetro enterrado bajo la costa septentrional de la península de Yucatán en México. Cuando colisionó con la Tierra, el meteorito liberó una energía equivalente a 100 millones de megatones de explosivos fuertes, causando gigantescos tsunamis. Una nube masiva de detritos se elevó hasta un diámetro de 200 km, se extendió sobre la superficie de la Tierra y descendió luego. Los residuos en descenso calentaron la atmósfera hasta cientos de grados, originando incendios masivos y bloqueó la luz solar, impidiendo que las plantas fotosintetizaran. Los detritos depositados produjeron la capa rica en iridio. Aproximadamente mil millones de toneladas de hollín, con una composición similar al humo de los incendios forestales, se depositó también. Muchas especies fósiles, en particular los dinosaurios, encontrados en las rocas cretácicas no se observan en las rocas terciarias del próximo estrato. ACTIVIDADES 1. Investigue y explique cuál es la razón por la que el método radimétrico no puede utilizarse para datar algunas rocas y minerales. =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 2. Busque información acerca de distinto tipo de rocas y minerales que no figuren en el cuadro e investiguen si pueden datarse por métodos radimétricos. Analice cuál de los isótopos mencionados en el texto utilizaría para calcular su antigüedad. El método del carbono 14 es fundamental para las investigaciones arqueológicas. Discutan la razón de este tipo de estudios. =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 3. Piense. ¿En qué época solidificaron las primeras rocas? ¿Con qué método de datación radimétrica se podría estimar la edad de estas rocas antiguas? =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 4. Describa la secuencia representada. ¿Cuáles fueron los principales supercontinentes del pasado gelógico? =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 5. Explique cuáles son las principales evidencias del cambio climático en el pasado geológico. ¿De qué clima son indicadores las tillitas, las evaporitas y las calizas? =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 6. Busque información adicional y explique qué tipos de organismos se extinguieron en los principales episodios de extinción masiva, a fines del Ordovício, del Devónico, del Pérmico, del Triásico y del Cretácico. =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 7. Lea la siguiente información y determine la diferencia entre extinciones masivas y graduales. ¿Cuál cree que sería el punto de vista más acertado para postular la extinción de los dinosaurios? Justifique. Uno de los naturalistas más grandes de todos los tiempos fue el barón Georges Léopold Cuvier (1769-1832), especialista francés en anatomía comparada y paleontología de los vertebrados, y fundador de ambas disciplinas. Trabajó en los ricos depósitos de fósiles de la cuenca de París y fue un pionero a la hora de desarrollar secuencias geológicas y de dilucidar la historia de la vida sobre la Tierra. Gran parte de las investigaciones de Cuvier son tan válidas en la actualidad como lo fueron hace 150 años. Vale destacar que Cuvier trabajó varias décadas antes de la publicación, en 1859, de la obra de Charles Darwin, Origin of Species. Cuvier escribía en Recherches sur les ossaments fossiles (1812): "Estos repetidos avances y retiradas del mar no han sido ni lentos ni graduales; casi todas las catástrofes que los han ocasionado han sido súbitas; y esto, es fácilmente demostrable, en especial, en lo que atañe a las últimas, cuyos rastros son muy visibles...La vida, por lo tanto, se ha visto a menudo alterada en la Tierra a causa de terribles acontecimientos, calamidades que, en su inicio, tal vez, hayan movido y derrumbado hasta una gran profundidad toda la corteza externa del planeta...innumerables criaturas vivientes han sido las víctimas de estas catástrofes...Sus razas se han extinguido..." (Traducción del francés). Nota: Conviene recordar en este punto que se distinguen cuatro tipos de catástrofes. a) Topológicas (originadas por cambios en el relieve del terreno y la variedad de los materiales que lo componen; por ejemplo, avalanchas, aluviones y aludes); b) Tectónicas (originadas por fenómenos que tienen lugar en el interior de la corteza terrestre; comprenden maremotos, terremotos y fenómenos volcánicos); c) Meteorológicas (originadas por los factores climáticos, tales como tornados, huracanes, tifones e inundaciones); d) Astronómicas (originadas por la caída de meteoritos, explosiones de supernovas, colisión de cometas, etc.). Esta visión de la historia de la vida sobre la Tierra fue discutida por Charles Lyell (1797-1875), el gran géologo británico. Este interpretó las mismas secuencias de forma muy distinta a Cuvier. Por ejemplo, en la primera edición de su manual de geología (1833) escribió las siguientes palabras, bastante rebuscadas: "Oímos hablar de repentinas y violentas revoluciones del planeta, de la elevación instantánea de cadenas de montañas, de paroxismos de energía volcánica...También se nos habla de catástrofes generales y de una sucesión de diluvios, de la alternancia de períodos de reposo y desorden, de la refrigeración del planeta, de la súbita aniquilación de razas enteras de animales y plantas y otras hipótesis, en las que vemos el antiguo espíritu de la especulación redivivo y un deseo manifiesto de cortar, en lugar de desatar pacientemente, el nudo gordiano...". "En nuestro intento de desentrañar estos difíciles problemas seguiremos un camino distinto, limitándonos a los efectos conocidos o posibles de causas existentes, con la seguridad de que aún no hemos agotado los recursos que el estudio del presente curso de la naturaleza puede ofrecer y, por tanto, de que estamos autorizados, en los inicios de neustra ciencia, a recurrir a agentes extraordinarios". (Traducción del inglés) Comentario: De la lectura de estos textos queda claro que Lyell estaba en desacuerdo con Cuvier...y esto lo expresaba de una manera bastante desagradable. La frase "antiguo espíritu de la especulación redivivo" no es en absoluto un cumplido. Y lo que dice sobre desatar pacientemente el nudo gordiano supone que sólo gracias a la honradez y el trabajo duro podrán los géologos llegar a alguna parte. La primera postura es el catastrofismo y la segunda el actualismo, gradualismo o uniformitarismo. Esta última postura ha sido omnipresente en geología ya que evita apelar a agentes extraordinarios para interpretar las hitoria terrestre. Lyell no dudaba en transformar a la catástrofe en una cuestión de moral. Parecía afirmando ésto más un abogado (su carrera original) o un político que un científico. Indudablemente el dogma lyelliano ha hecho posible mantener una clara separación entre la ciencia y la religión. Más aún, ha sido un puntal para el enunciado de la teoría de la evolución por Charles Darwin y Alfred Wallace. Sin embargo, luego de dos guerras mundiales, en pleno siglo XX, cuando cobraron auge las especulaciones sobre el efecto invernadero, el invierno nuuclear y la lluvia de meteoritos sobre distintos astros del Sistema Solar, la situación política y social parece preparar el escenario para una recuperación plena del catastrofismo o es mejor decir, tal vez, neocatastrofismo. Durante un siglo, el pensamiento geológico y paleontológico convencional ha estado dominado por las teorías de Lyell. A menudo se citaba a Cuvier como algo cómico en las clases de Introducción a la Geología. con demasiada frecuencia se rechazaba la postura catastrofista afirmando enfáticamente que Cuvier estaba equivocado. Pero en realidad, el mayor ataque a Cuivier no vino de los geólogos sino de los biólogos evolucionistas ya que, aparentemente, Cuvier creía en la inmutabilidad de las especies. Sin embargo, a medida que se sabe más sobre las extinciones periódicas de organismos, emimentes paleontólogos como Otto Schindewolf, Digby Mc Laren, David Raup y otros han documentado con innumerables pruebas que las extinciones al final de distintas eras pudieron ser realmente hechos catastróficos de tipo astronómico y topológico combinados, debido al impacto de grandes volúmenes (meteoritos, cometas, etc.) y erupciones volcánicas inmensas. Los meteoritos, literalemnte, "rocas que caen del cielo", sin duda cumplen los requisitos de "agentes extraordinarios" según las palabras de Lyell, o de agentes catastróficos, según la noción de Cuvier. En un principio, cuando se abogó por primera vez por los meteoritos como causantes de extinciones, los seguidores de Lyell absorbieron los nuevos hechos y conceptos sin cambiar seriamente la docrina uniformitarista. Esto queda ilustrado en las palabras de George Weherhill y Eugene Shoemaker, en una fecha tan tardía como 1982: "Aunque el encuentro físico de objetos tales como meteoritos o cometas con la Tierra se pueda calificar apropiadamente como catastrófico, por la magnitud de los efectos que producen, al mismo tiempo son uniformitaristas, porque representan la extención de procesos geológicos observados actualmente en una era geológica anterior”. Actualmente, en esta época posmoderna de grandes angustias existenciales, la teoría catastrofista sienta muy bien aunque sin dejar de lado algunos principios básicos gradualistas en apoyo de la teoría de la evolución. =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 8. Discuta los datos del siguiente calendario cósmico y relaciónelos con la evolución de la vida en la Tierra. CALENDARIO CÓSMICO Millones de años transcurridos desde el suceso hasta nuestros días Suceso Años Gran explosión 15.000 Galaxias 14.999 Sol 5.000 Planetas 4.600 Vida 3.600 Vida pluricelular 800 Peces 505 Anfibios 360 Reptiles 286 Dinosaurios 248 Aves y mamíferos 213 Extinción de los 65 dinosaurios Primates 55 Caballos primitivos 47 Ballenas y monos 25 antropomorfos Carnívoros y 5 homínidos Homo sapiens 1,5 Aparición de la 0,05 escritura Civilización occidental 0,003 Descubrimiento de 0,0005 América El hombre pisa la luna 0,00003 Momento hipotético; si todo el tiempo se hubiera comprimido en un año Día y mes Hora 1 de enero 12:00:00 a.m. 1 de enero 12:35:42 a.m. 1 de septiembre 12:00:00 a.m. 10 de septiembre 10:04:47 p.m. 4 de diciembre 05:16:47 p.m. 12 de diciembre 03:50:24 a.m. 19 de diciembre 11:25:26 a.m. 23 de diciembre 01:43:40 a.m. 24 de diciembre 09:46:22 p.m. 25 de diciembre 08:23:25 p.m. 26 de diciembre 05:13:20 p.m. 30 de diciembre 09:18:43 a.m. 30 de diciembre 30 de diciembre 31 de diciembre 03:15:50 p.m. 08:01:32 p.m. 09:07:11 a.m. 31 de diciembre 09:01:26 p.m. 31 de diciembre 31 de diciembre 11:06:25 p.m. 11:58:12 p.m. 31 de diciembre 31 de diciembre 11:59:53 p.m. 11:59:58 p.m. 31 de diciembre 11:59:59 p.m. http://www.memo.com.co =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== Foro Impacto profundo La espectacular y reciente colisión del cometa P/Shoemaker Levy 9 o más bien de sus fragmentos sobre la superficie del planeta Júpiter ha llevado a las primeras páginas de todos los medios de información un fenómeno bien conocido, pero al que hasta ahora no se le había prestado gran antención: el choque periódico de asteroides o cometas contra los planetas y satélites del Sistema Solar. Se sabe, por ejemplo, desde hace tiempo, que los cráteres lunares han sido formados por colisiones con objetos errantes, en general de dimensiones no superiores a las montañas terrestres. También, la mayoría de los científicos, coinciden en que cometas y asteroides que chocaron en otras épocas con la Tierra fueron responsables de fenómenos de extinción masiva de la fauna y la flora de la época. Por ejemplo, las investigaciones submarinas en el golfo de México ya no dejan dudas que el cráter sumergido Chixulub, resto de la caída de un gran objeto, meteorito o cometa, que chocó con la Tierra hace 65 millones de años, es la evidencia de la extinción de los dinosaurios y otros muchos seres que convivían con ellos en el planeta por entonces. Tanto los cometas como los asteroides representan los últimos restos de una población extinta de pequeños objetos que, en los inicios de la historia del Sistema Solar, se agregaron para formar los actuales planetas y sus satélites. Estas dos poblaciones poseen características físicas y dinámicas muy diferentes, de modo que resulta útil distinguirlos. Los asteroides son objetos básicamente rocosos, y más precisamente formados por rocas ígneas. Algunos, conocidos como sideritas, tienen un elevado contenido metálico, principalmente hierro y níquel o aleaciones de ambos metales. Otros, llamados lititas, están compuestos sobre todo de silicatos formados por combinaciones de silicio, oxígeno, magnesio y hierro (que forman principalmente dos clases de minerales, el olivino y el piroxeno), como en las rocas terrestres; otro grupo, las litosideritas o siderolitas, tiene un carácter mixto de metal y silicatos, y un último grupo, las condritas carbonosas, está compuesto además de silicatos o aleaciones de metales, mayormente por carbonatos. La mayoría de los asteroides gira en torno al Sol, en la zona del espacio comprendida entre la órbita de Marte y la de Júpiter. A diferencia de los asteroides, los cometas son objetos muy frágiles, compuestos principalmente de hielos de diferente naturaleza, en los que predomina el agua sólida, así como el hielo seco (dióxido de carbono congelado), el metano, etcétera. La mayor concentración de cometas se halla más allá de los confines de los planetas exteriores, formando la nube Oort. Pero investigaciones recientes muestran que debajo de la capa de hielo, existen similitudes con los meteoritos rocosos. La interacción gravitatoria con los planetas mayores como Júpiter, puede hacer que estos objetos sea fluctuante, por lo que resulta difícil estimar su número y evaluar la probabilidad de que uno de ellos venga a caer sobre nosotros, lo que constituye uno de los mayores escollos de la organización de una vigilancia permanente. Lo realmente escalofriante acerca de estos objetos es que uno de tan solo 10 km de diámetro, lanzado a una velocidad de 20 km por segundo, casi no “vería” la atmósfefa, emplearía uno o dos segundos en atravesarla, dejando un tubo de vacío tras de sí, y explotaría en el suelo con una potencia estimada de mil millones de megatones, produciendo un cráter de 100 km de diámetro: continentes enteros se incendiarían, lanzando a la atmósfera tal cantidad de cenizas y hollín que impedirían la llegada de la luz solar al menos durante un par de años. Un impacto en el mar, como el que destruyó a los dinosaurios, levantaría marejadas de hasta 600 km de alto que invadirían los continentes destruyendo las ciudades. Estos catastróficos episodios han sido dramatizados en dos películas de estreno reciente: Deep Impact (Impacto profundo) y Armaggedon. Pero todavía es difícil imaginar las consecuencias de tales choques, mientras persisten las dudas y los temores...hasta encontrar una forma de detección, prevención y...evacuación global. 1. A partir del análisis de este texto, ¿qué diferencias existirían entre cometas y asteroides, y qué tipos de asteroides se conocen en cuanto a su composición química? ¿Cuáles son los elementos químicos más abundantes en la corteza terrestre? ¿Por qué? 2. ¿Qué sentido tiene la afirmación de que los meteoritos más abundantes tienen una composición similar a la corteza terrestre? 3. ¿Qué opinan sobre la caída de grandes objetos sobre nuestro planeta? ¿Creen que existe una posibilidad real de que algo catastrófico ocurra en los años venideros? 4. ¿Habría alguna posibilidad de sobrevir para el ser humano? Si pueden, vean en grupos alguna de las películas mencionadas y extraigan su conclusión. 5. En la película Deep Impact (Impacto profundo) se relata un ELE (Extinción level event, o evento de nivel de extinción) y que alude a la caída de un asteroide de más de 10 km de diámetro y sus consecuencias para la vida en el planeta, similar a la que mató a los dinosaurios. Según David Raup, uno de estos meteoritos puede caer a la Tierra cada 25 millones de años. ¿Creen que el ser humano ha desarrollado una tecnología suficiente para poder eludir o superar uno de estos eventos? Módulo 3. El origen de la vida y los tiempos precámbricos Objetivos • Discutir la importancia de los tiempos precámbricos • Explicar la formación de la primera atmósfera • Analizar el surgimiento de los mares y océanos • Explicar la constitución de cratones en las etapas iniciales de la historia terrestre • Analizar las etapas en el origen de la vida terrestre • Realizar simulaciones sobre el origen de la vida • Examinar las características de la vida en el eón Arqueano • Examinar las características de la vida en el eón Proterozoico • Comparar el origen exógeno y endógeno de la vida terrestre • Examinar las características de la evolución de la vida celular Contenidos Tiempos precámbricos en foco Formación de la atmósfera Los primeros océanos La formación de las primeras rocas La historia primitiva y el inicio de la evolución prebiológica Persistencia de las condiciones extremas en la superficie terrestre Hipótesis original de Operain y Haldane El origen de la vida Reacciones de polimerización Las panspermias Intraterrestres y tipos de metabolismo Los coacervados El mundo de ARN El origen y la evolución de las células Del progenota a LUCA Los tiempos precámbricos en foco El período de la historia terrestre que se extiende entre los 4650 millones de años y los 570 millones de años se conoce, comúnmente, como tiempos precámbricos (por haber transcurrido antes del período Cámbrico) o supraeón Criptozoico (de kriptos, oculto, y zoikos, ser). La Geología moderna (Atlas de la Asociación Geológica internacional, 2008) divide este dilatado lapso en cuatro eones, el eón Chaotiano, en relación con la formación de la Tierra a partir de la protonebulosa original y de los plantesimales en colisión perpetua, es decir, hasta 4.450 m.a.; el eón Hadeano o Katarqueano (de kata, más allá de, y archaios, antiguo) o Hadeano (por Hades, dios del infierno de la mitología griega), desde los 4.450 a los 3.850 millones de año; el eón Arqueozoico o Arqueano (de archaios, antigua), entre los 3.850 y los 2.500 millones de años, y el eón Proterozoico (de proteros, primitiva), entre los 2.500 y 543 millones de años. Porcentaje de la historia terrestre que corresponde a cada eón 17,4% (Hadeano) 28,2% (Arqueano) 42% (Proterozoico) 12,4% (Fanerozoico) ------- (comprende tres eras) era Paleozoica (7,1%) era Mesozoica (3,9%) era Cenozoica (1,4%) Los eones son las unidades mayores en que se divide el tiempo geológico y son cuatro: Hadeano, Arqueano, Proterozoico y Fanerozoico). Los tiempos precámbricos (los tres primeros eones) abarcan entonces un 87% de toda la historia terrestre, lo que representa aproximadamente más de 21 horas de un reloj imaginario que hiciera equivaler los estratos a las horas. Durante la mayor parte de los dos primeros eones, la Tierra fue una masa viscosa y fundida bombardeada continuamente por grandes asteroides, hasta los 4.000 millones de años. Al promediar esta etapa, se fueron formando los océanos primitivos y con ello descendió la temperatura superficial, solidificando el magma original y formándose las primeras rocas. Debido a las extremas condiciones imperantes no se conocen testimonios de vida tan antiguos, pero esta etapa es muy importante por haberse realizado la evolución química o prebiótica hasta la formación de las primeras células. Eón Fanerozoico.543 m.a. a la actualidad. 4a. Era Paleozoica. Aparecen los esqueletos y caparazones y se produce la conquista del medio aeroterrestre, con la aparición de anfibios, insectos, helechos y gimnospermas. 4b. Era Mesozoica. Se desarrollan los vertebrados modernos, incluidos los dinosaurios, los mamíferos y las aves. Finalmente, evolucionan las angiospermas y los insectos polinizadores, como las mariposas. 4c. Era Cenozoica. En la tierra firme, se produce el triunfo de los mamíferos placentarios, los pájaros (Passeriformes) y las angiospermas. Finalmente llegó la radiación evolutiva de los primates (monos) y la aparición del ser humano. Los eones son las unidades mayores en que se divide el tiempo geológico y son cinco en la actualidad: Chaotiano, Hadeano, Arqueano, Proterozoico y Fanerozoico. Los tiempos precámbricos (los cuatro primeros eones) abarcan entonces un 87% de toda la historia terrestre, lo que representa aproximadamente más de 21 horas de un reloj imaginario que hiciera equivaler los estratos a las horas. Los tres primeros eones de la historia terrestre se conocen, tradicionalmente, con el nombre de tiempos precámbricos. Al período de la historia terrestre que se extiende desde los 4.600 m.a. hasta los 570 m.a. se lo denomina, comúnmente, tiempos precámbricos (antes del Cámbrico, primer período de la era Paleozoica) o eón Criptozoico (de kriptos, oculto, y zoon, ser animado o animal), porque se pensaba que no existían fósiles. Los eonses que se incluyen en este amplio intervalo representan el 87 % de la historia de la vida terrestre, lo que representa aproximadamente más de 21 horas (de las 24 de un reloj imaginario que hiciera equivaler los estratos a horas) como se vio en el módulo anterior. La formación de la atmósfera y los primeros océanos Durante el amplio intervalo de tiempo geológico que incluye los dos primeros eones, es decir, el eón Chaotiano (4650 a 4250 m.a.) y el eón Hadeano (4250 a 3.850 m.a.) no se registraba la formación de una atmósfera suficientemente densa ni de océanos extensos. Y sin estos dos componentes, todavía no era posible la formación de la vida. Así que debió transcurrir algo más de 500 m.a. para que se iniciara la historia de la vida en la Tierra. Los cálculos que realizó Claude Allègre, de la Universidad de París, sugieren que la Tierra empezó a formar la atmósfera, más o menos al mismo tiempo en que comenzó a diferenciarse su núcleo después de transcurridos entre 120 y 150 millones de años desde su origen, hace unos 4.450 m.a.. La Tierra primitiva estaba sujeta a los bombardeos que perduraron hasta hace 4.005 millones de años, los océanos iniciales se evaporaban y volvían a condensarse, tal vez en numerosas ocasiones. El hidrógeno y helio (además de nitrógeno) de la atmósfera primaria (barrida en sus gases ligeros por los vientos solares hacia la los planetas exteriores durante la acreción de los planetesimales) fue reemplazado paulatinamente por una atmósfera secundaria, que comenzó a aumentar su concentración y proporción de gases de invernadero y otros que resultarían muy tóxicos para la mayor parte de los seres vivos, no sólo metano y amoníaco, sino también monóxido y dióxido de carbono, sulfuro de hidrógeno y óxidos de nitrógeno y azufre, además de cierta proporción de vapor de agua, lo que contribuyó al calentamiento del planeta y la fusión de los materiales rocosos; Gradualmente, se fue constituyendo lo que se conoce como atmósfera inicial reductora y de invernadero extremo, lo que significa que carecía de oxígeno libre y se mantenía a temperaturas por encima de los 200 °C, debido al metano y otros gases raros. La atmósfera reductora con metano y sulfuro de hidrógeno, especialmente proveniente de los volcanes, sería muy parecida a la que actualmente presenta Titán, la luna mayor de Saturno. El bombardeo meteórico continuo impedía tanto la formación de una corteza de rocas sólidas como la acumulación de agua líquida. La evidencia suministrada por la datación radimétrica en cristales de zircón de rocas sedimentarias muy antiguas señala que la interacción entre roca fundida y vapor de agua formó los primeros océanos (protocéanos) recién cerca de los 4.200 m.a. Sin embargo, ambos aportes comenzaron a cambiar rápidamente la composición de la atmósfera terrestre al agregar agua y aumentar el efecto invernadero: el vapor de agua desprendido en los volcanes al iniciarse la actividad tectónica terrestre, el desprendido de los cometas (que están formados por hielos de diferente composición, incluida el agua sólida) y el condensado desde la atmósfera caliente. El hidrógeno, el helio y los gases predominantes en la composición de la nebulosa que originó el Sistema Solar, no participaron en el proceso que condujo a la formación de los distintos planetas interiores de manera que, establecidos estos, una atmósfera con una constitución química similar a la de la nebulosa primordial quedó reunida alrededor de cada uno de dichos astros como consecuencia principalmente de fuerzas gravitatorias ejercidas por sus respectivas masas. Sin embargo, la atmósfera terrestre actual tiene una composición distinta pues carece de hidrógeno libre y de helio, siendo muy escasos en ella los gases nobles. El neón supera al nitrógeno en el universo, de manera que debería hallarse en cantidades importantes en la atmósfera terrestre si ésta fuera primitiva. En consecuencia, la única explicación razonable es aceptar que la atmósfera original terrestre fue reemplazada por otra similar a la actual, la que así resultaría secundaria. ¿Cuándo desapareció la atmósfera original terrestre y cuál pudo haber sido la causa? Si realmente existió una atmósfera primitiva terrestre, ello debió acontecer hace más de 3.350 m.a., pues las rocas del Supergrupo Swaziland (Sudáfrica) contienen carbonatos precipitados inorgánicamente, lo que muestra la abundancia de CO2 en la atmósfera de esos tiempos. En general, se adjudica el proceso de reemplazo atmosférico a la actividad solar. El sol es una estrella en evolución. Al estadío de Protosol su temperatura era muy inferior a la actual, por lo que debió ejercer escasa influencia sobre los planetas circundantes. Pero a medida que las reacciones termonucleares de su interior tomaban importancia, la temperatura se incrementó y en determinado momento, al adquirir un estadío comparable al de la estrella T-Taurus, comenzó a emitir gran cantidad de partículas ionizadas en forma de un fuerte viento solar que barrió a la atmósfera primitiva de los planetas interiores, no así la de los exteriores que, por hallarse más lejos, retuvieron la mayoría de la atmósfera original, que aún consevarían. En ella, la abundancia de hidrógeno dio lugar al hidrógeno molecular, metano, amoníaco y agua, dominando el hidrógeno y el helio, pero faltando el carbono libre, el nitrógeno y el oxígeno. En consecuencia, la atmósfera de los planetas exteriores es reductora, a diferencia de la oxidante (rica en oxígeno) que ahora tienen los interiores (excepto Mercurio, que carece de atmósfera). Comúnmente se admite que la atmósfera terrestre actual se originó por un proceso de desgasificación ocurrido en el interior de la Tierra como consecuencia de la diferenciación del núcleo, el manto y la corteza. Dicho proceso habría sido enérgico y, con liberación de gran cantidad de energía a través de un intenso volcanismo. Así, inmensas cantidades de Gases fueron expelidas desde el interior del planeta hacia el exterior donde se acumularon hasta constituir una nueva atmósfera. Algunos autores sostienen que el mencionado proceso de desgasificación se produjo progresivamente a lo largo del tiempo geológico, mientras que otros opinan que la casi totalidad del mismo (80%) tuvo lugar posiblemente en el lapso 4.500 - 3.800 m.a., (cuando aparecen las rocas sedimentarias más antiguas conocidas), y desde entonces se ha estado produciendo una desgasificación residual. Inicialmente, debido al intenso vulcanismo, el CO2 habría sido abundantes en la atmósfera, a pesar de que ahora se lo halla en una proporción baja. Ello obedecería a que el excedente se combinó con el silicato de calcio de las rocas produciendo cuarzo y carbonato de calcio (calizas). Esto sucedió principalmente a partir del Neoproterozoico y desde entonces, el intercambio gaseoso en la interfase aire-agua contribuyó a mantener en equilibrio el contenido de CO2 atmosférico, pues en esa región dicho compuesto es tomado por los organismos marinos para la elaboración de sus esqueletos calcáreos y por el fitoplancton, para su fotosíntesis. La actual proporción de CO2 atmosférico permite ejercer un efecto invernadero al regular la temperatura de la superficie terrestre, manteniéndola entre límites que hacen posible la vida en el planeta. La aparición de microbiotas fotosintetizadoras a partir del final del Arqueozoico resultó de fundamental importancia para la evolución química de la actual atmósfera terrestre ya que, el oxígeno producido por dicha actividad la fue convirtiendo de reductora en oxidante. La presencia de ciertos minerales y compuestos (hierro férrico, yeso, anhedrita) en rocas precámbricas muy antiguas ha hecho pensar que, con anterioridad al momento que dichas microbiotas pudieron empezar a liberar oxígeno, este elemento ya existía (hasta un 25%) en la atmósfera, posiblemente como consecuencia de la disociación fotolítica del vapor de agua por los rayos UV, aunque ello es poco probable, ya que el oxígeno proveniente de la desintegración del vapor de agua, siempre tiende a situarse por arriba de éste y así, al interceptar a los rayos UV, detendría su propia formación. Este sistema autoregulador se conoce como Efecto Urey. Los primeros organismos en liberar oxígeno mediante la fotosíntesis fueron las bacterias verdes no azufrosas cianobacterias, y más tarde las cianobacterias o algas azul-verdosas. Primeramente, este elemento comenzó a acumularse en los ambientes acuáticos y se utilizó en la oxidación de las formaciones ferríferas zonadas; posteriormente, su excedente fue transferida a la atmósfera, transformándose en ozono por la acción de los rayos UV. Hace unos 2.000 m.a., el contenido de oxígeno en la atmósfera y los océanos era suficiente como para que, en la superficie terrestre se formaran los estratos rojos, y en las aguas se depositaran sulfatos y carbonatos. Según L.V. Berkner y L.C. Marshall (1965) la concentración de oxígeno en la atmósfera reductora nunca superó el 0,1% de su concentración actual. Entonces, la vida sólo habría podido desarrollarse en el agua (lagos, mares someros protegidos), entre los 10 y 13 m de profundidad, donde llegaría la luza pero no los rayos UV. Cuando el oxígeno atmosférico alcanzó la concentración del 1% actual (Primer Nivel Crítico o Punto de Pasteur) o según otros autores, el 3%, se estableció la posibilidad de la respiración, mecanismo que permitió disponer entre 30 y 50 veces más energía a los organismos que con los procesos químicos de la fermentación. La vida aún permaneció confinada al agua, pero ahora podría desenvolverse a profundidades menores debido a la protección ofrecida por una capa de ozono cada vez más efectiva. Ello ocurrió en el Neoproterozoico, favoreciendo el surgimiento de los organismos metazoos (multicelulares). En el Paleozoico Temprano todavía la radiación UV dificultaba a los organismos habitar las aguas más superficiales pero, cuando la concentración del oxígenos llegó al 10% de la actual (Segundo Nivel Crítico) fue posible la vida en las aguas superficiales y surgió la respiración aérea, lo que habría acontecido al Silúrico. A partir del Devónico, la formación de grandes bosques contribuyó al aumento de la capa de ozono, y muchas formas orgánicas que hasta entonces sólo habrían podido vivir en el agua, comenzaron a invadir el ambiente terrestre, utilizando el aire para su respiración. Los valores actuales del oxígeno atmosférico datarían del Paleozoico Tardío si bien, no se puede descartar la posibilidad de que se hayan producido fluctuaciones de los mismos durante el Fanerozoico. La hidrosfera se formó por la condensación del vapor de agua proveniente del intenso vulcanismo relacionado con la diferenciación magnética del interior terrestre. Inicialmente el agua integraba la composición de los minerales hasta que la elevada temperatura interior del planeta, rompió los enlaces químicos y las moléculas de agua quedaron liberadas, siendo expulsadas por los volcanes en estado de vapor. Los océanos fueron adquiriendo su salinidad a través del aporte de los cursos de agua que, recorriendo la superficie terrestre, erosionaban las rocas, y también desde las crestas y dorsales medias oceánicas en las que continuamente, se está llevando a cabo un intenso hidrotermalismo y el agua oceánica es reciclada y salinizada. En un comienzo, el agua de estos océanos primitivos era clorobicarbonatada y la mineralización fue aumentando hasta el Neoproterozoico, cuando habría alcanzado valores similares a los actuales. Los cloruros y sulfatos fueron incrementándose mientras que los bicarbonatos, hierro ferroso, calcio y bario, disminuían y la presión del oxígeno se elevaba. Además, la creciente abundancia de fosfato, permitió la primera síntesis biológica. La formación de las primeras rocas Si bien hace 4.300 m.a. se inició la formación de los cratones o núcleos continentales, como se analizó en el módulo anterior, entre los cuales el más primitivo, Yilgarn, corresponde a Australia occidental, y al cual pronto se sumarían otros como Kalahari, Arqueoplata (escudo del Rìo de la Plata) y Groenlandia, sin embargo, las primeras rocas sólidas de estos núcleos de la corteza continental emergieron recién hace entre 4.100 y 3.900 m.a., aproximadamente al cesar el bombardeo meteórico. A pesar de que casi todas las rocas antiguas se han destruido por diferentes procesos, tales como la erosión y los conocidos como metamorfismo –transformación de las rocas por acción de altas temperaturas y presiones –, a partir de la década de 1980 se realizaron varios descubrimientos importantes. El grupo de Stephen Moorbath, de la Universidad de Oxford, Inglaterra, halló en el oeste de Groenlandia un terreno que tiene cerca de 3.800 millones de años. Samuel Bowring, del Instituto Tecnológico de Massachusetts, exploró una pequeña área de los Estados Unidos, llamada Acasta, que está formada por una roca denominada gneiss que tiene 3.960 millones de años. Pero las dataciones que dieron mayor antigüedad fueron realizadas por un equipo de la Universidad Nacional de Australia, en Canberra, dirigido por William Compston, sobre un mineral llamado circón, proveniente del este de ese país; su antigüedad estaba comprendida entre 4.100 y 4.300 millones de años. Algunas conclusiones sobre las características de la Tierra primitiva y el inicio de la evolución prebiológica Las dataciones en conjunto sugieren que la vida más primitiva posible (incluidos los virus), en la Tierra no pudo haber comenzado antes de unos 4.200-3.900 millones de años. Obviamente, las condiciones extremas detalladas hasta aquí impidieron inicialmente la formación y la perdurabilidad de las células o de cualquier forma de vida (debido a la esterilización completa del ambiente, como si se tratara de un gigantesco autoclave, con temperaturas que en ocasiones se acercaban a las de la superficie de Mercurio). Sin embargo, hay que destacar tres características que contribuyeron a la formación de las primeras moléculas orgánicas y su ensamblado posterior para formar las macromoléculas de la vida (principalmente las proteínas y los ácidos nucleicos). • La ausencia de oxígeno en la atmósfera inicial resultaba esencial para impedir la destrucción de las moléculas orgánicas que se formaban (al no producirse la oxidación), ya que estaban ausentes los sistemas catalíticos (que aceleran la velocidad de las reacciones químicas) y son protectores de la oxidación. • La gran acidez de los océanos contribuía a la formación de depósitos de este material. • La radiación ultravioleta y las elevadas temperaturas suministraban la energía necesaria para el ensamblado de moléculas. Entonces, estas extremas condiciones resultaron esenciales para que tuviera lugar la evolución prebiótica, química o prebiológica (antes de la formación de las células). Esta evolución química conduciría a que los elementos simples que formaban moléculas sipmles (hidrógeno, nitrógeno, oxígeno, además de azufre y fósforo elementales) se combinaran para formar moléculas organógenas: metano, monóxido de carbono, amoníaco, dióxido y trióxido de azufre, dióxido de carbono, vapor de agua, cianuro de hidrógeno (ácido cianhídrico). Persistencia de las condiciones extremas en la superficie terrestre Más allá del Hadeano (el primer eón), durante el segundo eón de la historia terrestre, el eón Arqueano (de los 3.900 a 2.500 m.a.), los océanos primitivos se conservaron ácidos y calientes, con temperaturas que oscilaban entre 60 y 80 °C, y la atmósfera, muy rica en dióxido de carbono, mantuvo temperaturas superficiales elevadas, similares a las que se observan en el planeta Venus. Esta etapa proliferó la formación de cratones base de las futuras placas litosféricas. Por otra parte, los océanos permanecieron calientes (con temperaturas cercanas al punto de ebullición) y ácidos (debido al aporte de sulfuro de hidrógeno y óxidos de azufre y nitrógeno que reaccionaban con el agua), al menos durante unos 500 m.a. más. La continua radiación ultravioleta letal (UVL) que llegaba hasta la superficie de los océanos y continentes iniciales, actuaba como un poderoso esterilizador. Esto se debía no sólo a la ausencia de oxígeno libre, sino también de ozono, que recién apareció hace 500 m.a., durante el período Ordovícico (el ozono está formado por tres moléculas de oxígeno en una capa de la estratosfera y logra filtrar la radiación UV). La hipótesis original de Oparin y Haldane: la sopa prebiótica y el puente entre el mundo químico y el viviente En 1924, el bioquímico ruso, Alexandr Ivanovich Oparín (1894-1940) expuso en su pequeño libro El origen de la vida una novedosa teoría acerca de la aparición de los organismos primitivos (protocélulas) a partir de la combinación de compuestos inorgánicos simples, como los gases que formaban la atmósfera primitiva. En esencia, Oparin postuló que, si la atmósfera primitiva carecía de oxígeno, las reacciones químicas podían sintetizar espontáneamente los constituyentes orgánicos o biomoléculas de las primeras células, que funcionarían como unos organismos heterótrofos simples que se nutren en el caldo primordial del cual emergieron. Esta teoría, conocida como hipótesis del caldo o sopa primordial, suponía que el ensamblado de moléculas se produjo en un medio acuoso caliente y ácido (como el de la Tierra primitiva) y sujeto a continua evaporación, que permitió la acumulación de aminoácidos, ácidos grasos, monosacáridos y nucléotidos. Diez años más tarde, el biólogo inglés John Burdon Sanderson Haldane llegó a ideas similares a las de Oparin aunque sin tener conocimiento de sus escritos, que recién se tradujeron al inglés en 1938. No obstante, en la hipótesis original de Haldane se otorgó mayor importancia al ambiente volcánico y al papel de la radiación ultravioleta. La ventaja del ambiente propuesto por Oparin, sin embargo, resultaba en la mayor facilidad para lograr y mantener una concentración adecuada de moléculas por la evaporación continua producida por el calentamiento (en estos ambientes se forma un tipo de rocas sedimentarias especiales, las evaporitas). En la elaboración de sus ideas, Oparin retomaba ideas anteriores expuestas ante la Sociedad Botánica Rusa en 1922 y basadas en las ideas darwinianas transmitidas por su ilustre maestro, el botánico K. Timiryazev (1843-1920) quien sí había conocido a Charles Darwin. En su famosa obra El Origen de las especies por selección natural, Darwin todavía postulaba la existencia de organismos iniciales sencillos formados por el Creador que luego comenzaron a evolucionar en formas más complejas. Sin embargo, en su correspondencia privada a geólogos y naturalistas de la época, Darwin adhería a un origen prebiológico (antes de la vida) y abiótico (a partir de sustancias químicas sencillas). No es casual que ya en 1871, Charles Darwin afirmara que: "[...] Si pudiéramos concebir (¡y cuán gran suposición implica este si!) la posibilidad de la formación química de un compuesto proteico, en algún estanque cálido y pequeño, que contuviera toda clase de sales fosfóricas y amónicas, que recibiera luz, calor, electricidad, etc., compuesto que una vez formado podría sufrir otros cambios posteriores de mayor complejidad, en las condiciones actuales este material sería devorado o absorbido al instante, lo cual no podría haber sido el caso previamente a la formación de los primeros seres vivos". Obviamente, los bioquímicos de comienzos de siglo rechazaron las teorías de Oparin y Haldane por un hecho fundamental: retrocedían a la época en que se consideraba la realidad de la generación espontánea. El dogma de la generación espontánea, según el cual los seres vivos podían surgir de la materia inanimada, había prevalecido hasta el siglo XIX, cuando Louis Pasteur fue el encargado de desechar la aparición de formas de vida espontáneas con sus investigaciones experimentales y planteó la existencia independiente de las diferentes formas de vida. Sin embargo, lo que no comprendieron los detractores de Oparin y Haldane, era que los mecanismos del origen espontáneo de la vida se relacionaban con las condiciones que prevalecían en el planeta hace más de 4.000 m.a., muy diferentes de las ensayadas por Louis Pasteur en su laboratorio. Las teorías de Oparin y Haldane parten de una atmósfera reductora rica en metano, amoníaco, agua y descargas eléctricas para llegar a la síntesis de los monómeros. Esta es la base de las pruebas de laboratorio que siguieron al planteo teórico y demostraron la veracidad del plateno. Acumulación de los monómeros a partir de los gases de la atmósfera reductora (las pruebas de laboratorio) La primera etapa en el origen de la vida consistió, entonces, en la formación de los monómeros orgánicos (sillares fundamentales de las macromoléculas, como los aminoácidos) a partir de moléculas más simples y organógenas (como el metano). Sin embargo, esto todavía debía ser probado en forma experimental. Hay que aclaar que la primera síntesis analógica d euan reacción prebiológica digna de ser nombrada fue realizada en 1828 por el químico alemán Friedrich Wöhler. Para obtener una sustancia orgánica, partió de una mineral, el cianato de amonio, y demosró por primera vez que era posible franquear la barrera uqe separaba la química mineral de la viviente. En particular, una importante reaccion prebiótica que surgía del modelo de Oparin era la de la fomración del ácido cianhídrico (HCN), muy soluble en agua. En presencia de radiación UV, el nitrógeno molecular gaseoso se disocia en dos átomos: N2 ⇒ 2N Mientras que la molécula de nitrógeno es inerte, los átomos aislados pueden unirse a gran variedad de otros átomos y moléculas, en particular el metano paa producir la siguiente reacción: N + CH4 ⇒ HCN + 3/2 H2 Otra síntesis prebiótica posible involucra al metano y al agua y permite sintetizar formaldehído: CH4 + H2O ⇒ HCHO + 2H2 En septiembre de 1951, en una conferencia que dio el profesor Harold Clayton Urey, de la Universidad de Chicago, premio Nobel en 1934 por su descubrimiento del deuterio y quien tenía un conocimiento muy acabado del origen del Sistema Solar, y a la que asistió el químico estadoundense Sanley Lloyd Miller de la Universidad de Berkeley, científico que conocía muy bien las deas de Oparin y Haldane, así como la experiencia de Wöller, y concluyó que: • la atmósfera primordial de la Tierra era favorable para la formación de las biomoléculas complejas, por la abundancia de hidrógeno; • los gases que envolvieron a la Tierra primitiva eran reducidos (con este mismo gas hidrógeno) y estaban representados, principalmente, por metano, amoníaco y vapor de agua; • en este medio, las reacciones químicas, alimentadas por descargas eléctricas de los relámpagos además de las radiaciones UV (ultravioletas) de alta energía, producirían las moléculas orgánicas rocas en hidrógeno que conforman los seres vivos. En 1953, Miller y Urey se reunieron para probar esta teoría. Utilizaron un frasco de cristal con una mezcla invisible de gases (sin oxígeno). Agregaron un sistema complejo de circulación continua, con matraces, condensadores y colectores, y simularon una atmósfera de metano, amoníaco, vapor de agua e hidrógeno. En presencia de descargas eléctricas que imitaban la acción de los relámpagos obtuvieron, se formaron compuestos intermedios clave, como el formaldehído y el cianuro de hidrógeno. Las principales reacciones químicas que se investigaron fueron las siguientes: HCHO+NH3+HCH ==NH2CH2CN (aminonitrilo) + H20 NH2CH2CN + 2H2O=NH2CH2COOH (glicina) + NH3 Se formó el gas aminonitrilo por la presencia de amoníaco. El aminonitrilo reaccinó consecutivamente con el amoníaco y el cianuro de hidrógeno y formó el aminoácido más sencillo, la glicina. Finalmente, al cabo de un tiempo, una sustancia marrón que chorreaba por las paredes del frasco ya contenía una colección de moléculas orgánicas sencillas, principalmente los aminoácidos alanina, valina, prolina, ácido aspártico y ácido glutámico. Igualmente aparecieron en cantidades más pequeñas los otros aminoácidos que forman las proteínas naturales. Experimento original de Urey y Miller En el matraz superior, Urey y Miller recogieron, a través de la llave para toma de muestras, los gases necesarios de la atmósfera primitiva circulante, los que sometieron a descargas eléctricas continuas o relampagueos (provocados por la interacción atómica entre dos electrodos), y a medida que se producían las reacciones químicas en el medio de solución, las sustancias solubles formadas eran recogidas en un condensador. Los gases se disolvían cuando el caldo resultante se enfriaba y las gotitas de agua caían en un colector y llegaban al matraz inferior que se calentaba con un mechero Bunsen. Esto simulaba una lluvia primordial sobre el protocéano, lago o charco caliente inicial. Es importantes destacar que, en todas las muestras de laboratorio en las que se obtuvieron aminoácidos, las proporciones de la forma levógira y la dextrógira (los dos estereoisómeros), por ejemplo, L y D-glicinas, eran idénticas, mientras que en todos los seres vivos actuales, salvo ciertas proteínas de las arqueas, sólo se presentan L-aminoácidos y D-azúcares. Para explicar esta diferencia de concentración se postuló una superficie cristalina asimétrica en el ambiente de formación de los aminoácidos. La publicación de estos hallazgos unos meses más tarde causó gran revuelo en la opinión pública, si bien en una encuesta realizada por la famosa agencia de encuestas Gallup, el 78% de la gente consideró que el resultado del experimento era imposible. Sin embargo, pronto más investigadores confirmaron y complementaron estos descubrimientos con nuevas mezclas. En 1961, el español Juan Oró mezcló cianuro de hidrógeno con amoníaco en solución acuosa sin introducir formaldehído (que se había agregado al experimento de Urey) y encontró que de todas maneras se formaban los aminoácidos y, también, inesperadamente, la adenina, una purina compleja. Otros experimentos posteriores permitieron obtener las restantes bases: la purina guanina, y las pirimidinas citosina y timina, a partir de compuestos intermedios como el cinanógeno (C2N2) y el cianoacetileno (HC3N). De esta manera, quedaba planteada entonces la formación de aminoácidos y bases (pirimidinas y purinas), los constituyentes respectivos de las proteínas y los ácidos nucleicos, en una atmósfera reductora, condición imperante en la Tierra primitiva. Más allá de los aminoácidos y las bases, la formación de los azúcares, como la tetrosa (por ejemplo eritrosa) es posible a partir de dos moléculas de formaldehído, lo que se ha logrado con temperaturas cercanas a los 100 °C. Reacciones de polimerización en la Tierra primitiva y formación de los prebiontes La síntesis de moléculas más complejas a partir de monómeros más simple es la polimerización. En todos los casos, la formación de los biopolímeros resulta de las reacciones de condensación de los biomonómeros mediante la eliminación de una molécula de agua en presencia de energía (ATP o adenosín trifosfato, la moneda energética de las células). Por ejemplo, en la formaciíon de las proteínas, el agua proviene del grupo carboxilo de un aminoácido y del grupo amino del próximo; en la formación de los polisacáridos, el agua proviene de un oxhidrilo en posición 1 de un molécula, y de un hidrógeno en posición 4 o 6 de la otra; mientras que en los lípidos comunes (los triglicéridos), por la unión del oxhidrilo del glicerol con el grupo carboxilo del ácido graso. La formación de los ácidos nucleicos requiere de la partida de tres moléculas de agua mediante el agregado de la base (pirimidina o purina) al azúcar (ribosa para el ARN-acido ribonucleico, o desoxirribosa para el ADN-ácido desoxirribonucleico): dos para formar un nucleótido (base+azúcar+grupo fosfato, con una molécula de agua que deriva del oxhidrilo del azúcar en psición 1 más el hidrógeno de la base, y el otro del hidroxilo del azúcar en posición 5 y de un hidrógeno del fosfato) y la tercera para la formación secuencial del dímero mediante otro enlace de fosfato en posición 3 (polímero). A partir de los monómeros, en particular los aminoácidos, se debieron ensamblar inicialmente dipéptidos (dos aminoácidos unidos) y oligopéptidos (secuencias de unos pocos aminoácidos ensamblados), aunque también debieron originarse oligonucléótidos (pocos nucléótidos) antes de los polinucléotidos (muchos nucléotidos). Este sería el comienzo de las reacciones de polimerización que permitieron la formación de las principales macromoléculas orgánicas de los seres vivos, las proteínas y los ácidos nucleicos. La presencia de agua, tan importante para la vida, llevó a los científicos a plantearse el modo en que se formaron los polímeros iniciales a partir de los monómeros. Más allá de estas posibilidades y limitaciones, queda claro que las reacciones de polimerización son reversibles en agua y esta inversión es favorecida termodinámicamente (ocurre espontáneamente). Debilidades inherentes a la hipótesis de la sopa primordial La química prebiológica planteada por Oparin, Haldane, Miller y Urey supone unas condiciones determinadas: • Que la concentración de los productos iniciales sea suficientemente importante para que no se dispersen en la sopa prebiótica del protocéano; esto sólo se habría logrado sólo en un sistema de lagunas o mares someros aislados, o en suelos húmedos y salinos. • La química del agua conduce la mayor parte del tiempo la hidrólisis esontánea de compustos, por lo que la polimierización es desfavorecida termodinámicamente en solución acuosa, en beneficio de la despolimerización. Esto implica que se requeriría una cierta activación “artificial” de los compuestos intervinientes. • También sería primordial que las reaccions periféricas no afecten los procesos nacientes ni produzcan tóxidos metabólicos. Por lo tanto, se requería un proceso de selección de metabolitos en el medio original. Para superar estas dificultades, se propusieron etapas intermedias y soluciones alternativas. En el laboratorio se determinó la existencia de dos maneras para lograr la cotinuación de los procesos de polimerización y la iniciación del metabolismo: la concentración de los sustratos o reactivos, lo que resulta muy difícil no ser que tena lugar en pequeños estanques de agua dulce (sin sales) sometidos a evaporación y desecación o congelación o, que se produzca el acoplamieno del proceso a alguna reacción en que se facilita la liberación y transferencia de energía. Lamentablemente, el ATP es una molécula orgánica formada por ribosa y, en ausencia de enzimas, es difícil la síntesis de este azúcar que también está presente en las moléculas de ARN (ácidos ribonucleicos). Más allá de la dificultad de sintetizar ribosa, en el laboratorio se había logrado la síntesis de nucléotidos al combinar fosfato con nucléosidos, aunque estas condiciones controladas no podían existir en la naturaleza, en la atmósfera primitiva, y probablemente se originarían mezclas ensambladas con nucléosidos incorrectos o con otras moléculas similares, que impedirían la catálisis y replicación de las moléculas iniciales de los ácidos nucleicos. La acumulación a partir de las panspermias y el aporte exobiológico Algunos científicos suponen que el tiempo de evolución prebiológica resulta demasiado pequeño si se tiene en cuenta el cese del bombardeo meteórico y los primeros testimonios de marcadores bioquímicos (de 4.000 a 3.800 m.a.) para que pudiera ocurrir un aporte de materiales del tipo de los experimentos de formación de los coacervados. Por esta razón, recurrieron a una explicación alternativa: el origen exógeno o extraterrestre de los monómeros apoyados en ciertas evidencias meteoríticas. Fue postulada inicialmente por el Premio Nobel de origen sueco Svante Arrhenius a comienzos del siglo XX, aunque su mayor notoriedad se alcanzó con el descubrimiento de probables microorganismos en los meteoritos provenientes de la superficie marciana (aunque de dimensiones menores a las bacterianas) y todavía es apoyada por importantes científicos como H.C. Crick, uno de los descubridores de la estructura tridimensional del ADN, y Carl Woese, el creador del sistema de tres imperios de vida. A esto se suma el conocimiento de la evolución del Sistema Solar condujo a los geoquímicos a plantear un modelo de atmósfera terrestre primitiva poco reductora (diferente al planteado en el modelo de Oparin y Haldane), que contendría más dióxido de carbono que metano. En estas condiciones, la biomasa no pudo haber formado la Tierra primitiva únicamente a partir de de fuentes disponibles endógenamente, y parte debió venir desde el espacio exterior. Las panspermias son consideradas las unidades de dispersión de la vida a través de la galaxia y, aunque cuenta con muchos adeptos, no contradice el origen endógeno sino que lo complementa. La química orgánica es particularmente activa en el medio interestelar donde los radioastrónomos han descubierto moléculas de ácido cianhídrico y formaldehído, así como una centena adicional de compuestos orgánicos, incluidos etanol, acetona e incluso glucosa. Las evaluaciones recientes muestran que existen canttidades improtantes de esta materia orgánica, cerca de 20 g por cm2 que proviene de cometas y meteoritos y habría sido depositada en la Tierra primitiva. La hipótesis de las panspermias le otorga un papel fundamental al bombardeo meteórico y cometario, y no la simple fusión de la corteza y la vaporización oceánica, ya que lo relaciona con la siembra orgánica en la Tierra primitiva, con sustancias como las pirimidinas, los aminoácidos y otros monómeros importantes, aunque también podría explicar, por ejemplo, la presencia de molibdeno, un metal de escasa presencia terrestre aunque clave para muchos sistemas enzimáticos celulares e, incluso, de las primitivas células. El 28 de septiembre de 1969, varios fragmentos de un meteorito que cayó en Murchison (estado de Victoria, Australia) fueron sometidos a análisis de laboratorio. Todos los constituyentes de la atmósfera primitiva de Miller y Urey, incluido el cianuro de hidrógeno, el tioformaldehído (CH 2S), el cianocacetileno, los aminoácidos e incluso las pirimidinas. Esto se une a las claras evidencias espectroscópicas de testimonios de moléculas complejas en cometas, asteroides e incluso el polvo cósmico o las galaxias. La sonda europea Giotto ha revelado, además, la presencia de moléculas orgánicas complejas en el cometa Halley (se encuentra en él un 14% de carbono en los granos cometarios) además de ácido cianhídrico y polímeros del formaldehído, y junto con ellos 17 de los 20 aminoácidos, muchos de los cuales aparecieron en la experiencia de Miller y Urey. La hipótesis original consideraba la existencia de radiopanspermias que llegan a la superficie terrestre por la radiación solar (aunque esta hipótesis fue desechada ya que la radiación UV destruiría rápidamente las formas de vida). Más tarde fue reemplazada por las litopanspermias que postulan la existencia de estructuras cubiertas por una vaina de silicio luego de la expulsión planetaria, lo que las protege de las radiaciones. Los intraterrestres y los tipos de metabolismos Recientemente se ha descubierto la formación de una biosfera bacteriana y arqueana en los sedimentos del océano Pacífico a profundidades superiores a 500 m, así como otros microorganismos que viven en la corteza terrestre. A partir de sondeos profundosefectuados en Carolina del S., en la localidad de Savannah River, se han hallado microorganismos subterráneos a profudnidades superiores a 2.800 m, en el seno de rocas ígenas (redes solidificadas a partir del magma en fusión) como el basalt. Y también se han descubierto las primeras bacterias vivients en los granitos profundos. Todas estas bacterias, como Thiboacillus, Plectonema e Hyphomicrobium, presentan un metabolismo litoautotrófico (quimiolitótrofo), ya que utilizan la energía química, al igual que las bacterias y arqueas de las fuentes hidrotermales. Estos microorganismos de las profundidades deben haber resistido a estas duras condiciones debido a la adaptación extrema y a su lento metabolismo: algunos se dividen una sola vez por siglo según cálculos conservadores. Estos descubrimientos, que amplían mucho lo que actualmente se considera el campo de la biosfera o esfera de vida de la Tierra, podrían mostrar las condiciones geoquímicas que estaban presentes durante el origen de la vida. La energía necesaria para el crecimiento de los seres vivos provienem primariamente, de la luz solar (fotolitótrofos); en segundo lugar, se inicia en las reacciones de compuestos inorgánicos como el sulfuro de hidrógeno el ion ferroso (Fe2+) y el hidrógeno (quimiolitótrofos), y en tercer lugar, de la energía química de oxidoreducción que implica compuestos orgánicos (a partir de la fermentación o de la respiración aerobia) en los quimiorganótrofos (incluidos los fotorganótrofos, que también utilizan la luz solar). Por otra parte, la única fuente de carbono de los organismos heterótrofos (que son los quimiorganótrofos y fotorganótrofos) es la materia orgánica. Dependen, por lo tanto, de otros seres vivos. Se especula entonces que los primeros organismos (o preeorganismos) debieron ser quimiolitótrofos (o, directamente, fotolitótrofos) y de metabolismo anaerobio, más que quimiorganótrofos y heterótrofos. El origen de la vida a partir de un heterótrofo primordial que consumió la sopa primordial es visualizada por otros investigadores. En particular, Hyman Hartman en 1975 visualizó un origen autotrófico de la vida y del protometabolismo o metabolismo autotrófico primordial, probablemente quimiolitótrofo, aunque no realizó experiencias de laboratorio para confirmarlo. Origen endógeno de los monómeros a partir de fuentes hidrotermales Si bien la mayoría de la comunidad científica consideró más importante el aporte endógeno (y no exógeno) de los monómeros iniciales, se propusieron también diversas alternativas al ambiente de ensamblado original que incluían no sólo el caldo primordial, las charcas calientes (ambientes lacustres sujetos a evaporación copntinua) de Oparin y los ambientes volcánicos terrestres de Haldane, sino también los ambientes marinos superficiales a temperaturas de ebullición e incluso ambientes congelados y con géiseres. Sin embargo, todos estos escenarios presentaban la desventaja de no hallarse suficientemente protegidos de la radiación ultraviolea letal por la ausencia de la capa de ozon. Así surgía un nuevo escenario: era posible que los compuestos necesarios para la síntesis prebiótica hayan provenido no de la atmósfera (o del espacio exterior) sino de las fuentes hidrotermales submarinas, fuentes calientes y chimeneas volcánicas al nivel de las dorsales océanicas, a más de 2.500 m. de profundidad, protegidas suficientemente de la radiación UV. La exploración de estas fosas se inició también en la década de 1970. En las fosas hidrotermales, la nueva corteza terrestre se forma a partir de las rocas fundidas del magma. Verdaderos géiseres y fumarolas arrojan gases ricos en hidrógeno, incluido el metano, el sulfuro de hidrógeno y el amoníaco, a temperaturas que pueden alcanzar los 400 °C y a elevadísimas presiones. En estas fuentes (que suelen clasificarse en las arenas de metano, donde dominan las arqueas metanógenas o formadoras de este gas, y las colonias de gusanos gigantes, donde viven las bacterias productoas de sulfuro de hidrógeno) se agrega entonces un origen biogénico de estos gases al proveniente de los volcanes. Además, estos organismos que son resistentes a temperaturas, presiones e incluso radiaciones, resultan muy similares a los microfósiles más antiguos conocidos, como Eobacterium. Los primeros catalizadores La exploración de las fosas hidrotermales submarinas permitió la formulación de una nueva hipótesis por el químico alemán Günter Wächterhäuser. Para Wächasterhäuser, en estos sitios se produce la reacciòn continua de sulfuro de hidrógeno y óxidos de hierro para formar pirita (FeS 2). Además, para este científico, sólo tal ambiente suministra una red continua de superficies catalíticas (para acelerar las reacciones químicas) y compartimientos tridimensionales asimétricos que permiten la concentración sostenida de los reactantes o reactivos que podrían haber ayudado en la formación de las primeras moléculas complejas y en las reacciones de polimerización. El ambiente hidrotermal exhibe otras diversas ventajas: suministran continuamente otras moléculas orgánicas sencillas, como monóxido de carbono, dióxido de carbono, cianuro de hidrógeno y varios óxidos metálicos además de los mencionados amoníaco, sulfuro de hidrógeno y metano, que provienen de las chimeneas volcánicas, todos necesarios para la síntesis de aminoácidos y nucléotidos, así como también de los intermediarios energéticos, como el ATP (adenosíntrifosfato); estos gases pueden estabilizarse sobre la superficie de minerales de hierro y azufre (pirita o monosulfuro de hierro y esfalerita o sulfuro de zinc, respectivamente) descubiertos en las chimeneas en la década de 1990, y que reaccionan fácilmente con el hidrógeno molecular; Además, es probable que las reacciones de oxidorreducción (redox) asociadas con la pirita y compuestos similares, se forme suficiente energía libre y electrones suficientes para la fijación de los carbonos primordiales de los monómeros en reacciones acopladas. El mundo del tioéster y la autotrofia primitiva La hipótesis hidrotermal del origen de la vida propone también la existencia de un grupo de compuestos activadores intermediarios, los tioalcoholes, tioles o mercaptanos, análogos de los alcoholes y en los cuales el azufre reemplaza al oxígeno: R-SH en vez de R-OH. Christian de Duve propouso que la formación de tioésteres de aminoácidos y de otros ácidos, como el del ácido acético para formar la coenzima A, son la fuente energética fundamental del metabolismo primordial. El mundo del tioéster emana de diversas observaciones que provienen del matabolismo contemporáneo. En particular, se destaca la presencia del azufre en el sitio activo de numerosas enzimas, y el metabolismo azufroso de numerosas eubacterias y arqueas. Ya desde la década de 1970 se lograron formar en el laboratorio mezclas experimentales de polipéptidos a partir de aminoácidos a temperaturas situadas por encima del punto de ebullición del agua con mezclas adecuadas de aminoácidos, y que arrojaron mejores resultados en presencia de los catalizadores inorgánicos del tipo de la pirita (FeS) y la esfalerita (FeS 2). La autotrofia primitiva pudo consistir en la incorporación de dióxido de carbono a pequeñas cadenas carbonadas, mediante la utilización de hidrógeno sulfuroso (sulfuro de hidrógeno) y los minerales de hierro mencionados. FeS + H2S ⇒ FeS2 + H2 Por su parte, la presencia de FeS y la formación de FeS 2 libera la energía necesaria para favorecen la formación de succinato y otras moléculas orgánicas que constituyen intermediarios metabólicos. Superficies de adsorción, arcillas y obtención de péptidos catalíticos En la década de 1950, Desmond Bernal propuso nueas soluciones para resolver las dificultades inherentes al modelo de la sopa prebiótica, en particular a la fuerte deisolución que surgía. Según su opinión, para poder unirse en polímeros, el encuentro de las moléculas debía ser facilitado por la adsorción de los monómeros sobre calizadores sobre una superficie mineral, por ejemplo, la arcilla. Muy ampliamente distribuida en la Tierra y en los meteoritos, las arcillas presentan una estrutura compuesta de aniones oxigenadas y azufrados que portan cationes metálicos capaces de retener sustancias por intermedio de uniones electrostáticas. Las capas de arcilla se organizan en láminas constituidas por silicatos e hidróxidos de aluminio y hierro formando un tetraedor u octaedor. • Un primer modelo de organización de un tetraedro de sílice alternando con un octaedro de hidróxido de aluminio corrpresponde a la kaolinita. • Un segundo modelo que consta de una lámina octaédrica acompañanda de dos octaédricas es la montmorillonita, que posee una gran capacidad para dilatarse. El centro del tetraedro puede estar ocupado por un catin Si4+, que puede ser eremplazado por Al 3+ o Fe3+, mientras que el centro del octaedro puede ester ocupado por un catión Al 3+ que puede ser sustituido por Mg2+, Fe2+, y en menor proporción Li, Ti, V, Cr, Mn, Cu, Ni y Zn. Las láminas no son neutras y se compensan mediante iones intercambiables como el Na+, el Ca+2 y el K+. Estas características químicas de las arcillas les otorgan gran capacidad de absorción y adsorción y, probablemente les otorgó la capacidad de participar de los ciclos de hidratadión y deshidratación en la Tierra primitiva, así como la regulación de cada ciclo (pH, humedad, temperatura y fuerza iónica del medio de reacción). La adsorción sobre la arcilla permite facilitar la polimerización. Los aminoácidos y las bases pirimidínicas y purínicas se fijaron entre las láminas cargadas y se aproximaron entre sí facilitando la reactividad. En 1970, Abaron Katchlksy y Mella Paecht Horowitz realizaron la ilsutración experimental de esta tesis y demostarron que la montmorillonita, en particular, funciona como un reactor capaz de concentrar y posicionar los aminoácidos y adenilatos entre las láminas favoreciendo dicha polimerización. Además, varios trabajos muestran la eficacia de la montmorillonita en el transcurso de la condensación de los aminoácidos simulando ciclos de humidificación y desecación. Los péptidos obtenidos de esta manera a partir de las mezclas de aminoácidos presentan, a su vez propiedades catalíticas. De igual manera, los químicos estadounidenses Lawless, Edelson, Burton, Gibbs y Ferris condensaron mononuclétidos sobre una superficie arcillosa. La región de especificidad del enlace fosfodiéster (2’-5’ o 3’-5’) sirve de matriz para la condensación en la arcilla de la síntesis de oligoadenilatos. Los péptidos simples de composicion aleatoria obtenidos a partir de los aminoácidos se pueden calentar a 100 °C. Leslie Orgel investigó la síntesis rápida de estos péptidos a partir de aminoácidos tiolados en su función carboxílica, sin intervención de agentes extrínsecos. La activación del metabolismo de superficie Además de los factores mencionados, se descubrió la acción de una nueva serie de moléculas, las carbodimidas, la cianamida y el cianógeno todas las cuales, en contacto sucesivamente con dos monómeros o polímeros, permiten la eliminación de agua y la formación de oligonucléotidos similares al ATP. Por otro lado, quedó demostrado que para evitar que el agua obstaculice el proceso, se pueden utilizar iones fosfato (cargados negativamente) que se unen las moléculas poliméricas en formación. En la década de 1980, el químico alemán Albert Eschenmoser reemplazó uno de los intermediarios normales de la reacción anterior por formaldehído y finalmente logró sintetizar la cadena correcta de oligonucléotidos de desoxirribosa con pocas dificultades. La síntesis prebiológica de oligonucléotidos y polifosfatos explicaría la activación del metabolismo al permitir la glucólisis o fermentación anaeróbica (en ausencia de oxígeno) ligada a moléculas de ADP (adenosindifosfato) y que producen ATP por unión con el ion fosfato. El interés del metabolismo naciente en la superficie de las arcillas recide en la eficacia química en el curso de la catálisis de reacciones específicas. Se sabe que la mayoría de las moléculas vivientes se cargan negativamente (es el caso, por ejemplo, COO- o PO43-). Las uniones salinas también sn favorcidas sobre soportes cargados positivamente. La pirita aparece entonces como un soporte ideal, si bien no exclusivo, para este tipo de intercambios, y la montmorillonita como un molde de adsorción. El ensamblado de los prebiontes Pasar a la siguiente etapa en la evolución prebiológica, el ensamblado de las macromoléculas en agregados organizados, los prebiontes o proteinoides, estructuras no vivas (no celulares), sin membranas, aunque con identidad bioquímica propia, tal como propuso Sidney Fox en 1960, y más tarde Leslie Orgel en 1968. A partir de este punto, se sugirieron tres posibilidades, de la más simple a la más compleja, para la continuación del proceso: a) Después de calentadas y sacadas por contacto con lavas volcánicas, las soluciones de aminoácidos y polipéptidos se condensaron en glóbulos o microesferas denominados prebiontes o proteinoides. b) Los aminoácidos se polimerizaron sobre arcillas en los márgenes de lagunas someras para formar estos compuestos proteinoides mientras lo nucleótidos formaban simultáneamente polinucléotidos estables. c) Mezclados en agua con pequeña cantidades de agentes de condensación como el cianocetileno (de los experimentos de Miller) se pudieron formar polipéptidos y también polisacáridos. Para Fox, el estadio de prebiontes fue el paso inicial para la formación de las células primitivas. Los coacervados como antecesores de las primeras células Los polímeros originales debieron separarse de alguna manera del medio externo para ser viables. El próximo paso en la evolución prebiológica consistió en el agregado de los prebiontes, conjuntos todavía mayores (aunque no vivientes) de un tamaño de 1 a 2 micras de diámetro, principalmente los coacervados, propuestos inicialmente por Oparin, aunque también diversas estructuras similares como los liposomas, las microsferas e incluso las primeras monocapas y, gradualmente, bicapas de fosfolípidos fijados a la superficie por grupos fosfato. Los coacervados son gotas de solución orgánica “protegidas”: se diferencian las de proteína y carbohidrato (histona y goma arábiga), las de proteína y ácido nucleico (histona y ADN o ARN) y las de proteína y proteína (histona y albúmina). Estas esferas de formación espontánea se hallan separados por una membrana que se forma por un engrosamiento de la pared más externa del polímero. Cuando los coacervados se calientan a temperaturas de 130 a 180 ˚C, las imperantes en la Tierra de hace 4000 millones de años, forman espontáneamente microesferas de una a dos micras de diámetro. Oparin descubrió que si se agregaba la enzima fosforilasa a una solución de coacervados, se concentraba en el interior de las gotitas. Si posteriormente, se añadía glucosa 1-fosfato al agua del medio, esta difundía hacia el interior de las gotitas y se polimerizaban Los coacervados de histona y ARN presentan una capacidad adicional, el autocrecimiento. Además de los coacervados, se investigaron otras estructuras esferoidales: Los liposomas, esferas de moléculas orgánicas rodeadas por películas lipídicas monocapa cuyo ambiente interno se mantenía diferenciado y formaban otras esferas más pequeñas por fisión. Las microsferas, esferas proteicas, selectivamente permeables y excitables, rodeadas de agua, o bien formadas por pequeños volúmenes de agua encerrados en capas de polipéptidos. Finalmente, en parte también se lograron formar bicapas lipídicas estables en soluciones acuosas, y esta podría ser también una condición alternativa para la formación de las primeras estructuras precelulares. A pesar de la gran ventaja catalítica que tendrían los coacervados, y la permeabilidad selectiva que ofrece la formación de bicapas lipídicas, todavía faltaba la capacidad de replicación necesaria para la continuidad de la vida, así como la batería enzimática para ensamblar macromoléculas, que parecía no estar presente en la Tierra primitiva (donde no había células ni un código genético). Coacervados y liposferas. El origen de la replicación y la síntesis dirigida por una matriz Richard Dakwins elaboró sobre la definición de la vida de la Nasa su imagen acerca de la naturaleza de la entidad viviente más temprana que existió en la Tierra en su obra El gen egoísta: " En algún momento, una molécula sorprendente se formó por azar, a la que llamaremos el replicador. Puede no ser la más grande ni la más compleja a nuestro alrededor, aunque sí posee la propiedad extraordinaria de crear copias de sí mismo". Cuando Dawkins escribió estas palabras, el ADN formado por los bloques constituyentes, los desoxirribonucléotidos, era el candidato más seguro para cumplir este rol. Sin embargo, las proteínas y los ARN pronto lo sustituyeron como el candidato más adecuado. Los primeras sistemas de replicación debieron experimentar la intervención de una síntesis dirigida por una matriz. El principio de esta síntsis ha sido estudiado por Leslie Orgel y sus colaboradores. Los mononucléotidos previamente activados por el agregado de un núcleo de imidazo sobre el grupo fosfato se posicionan espontáneamente por las reglas de apareamiento de bases de Watson y Crick (los creadores del modelo del ADN) en una matriz preformada y se ligan unos a otros para formar la cadena primitiva. La síntesis es más sencilla si los monómeros de partida tienen la unión poco común fosforamida en vez del fosfodiéster. Sin embargo, al producirse este tipo de síntesis en el laboratorio se producen mezclas inestables de compustos levógiros y dextrógiros, además de diferentes isómeros con el nitrógeno en posición 1, 3, 9, 6 o 7 de la ribosa. En la década de 1970, el químico scocés Graham Ciarns-Smith sugirió que los cristales de arcillas pudieron constituir, por su organización tridimensional específica, la matriz informacional inicial de la replicación. Es decir, el precursor genético de los ácidos nucleicos habría sido de naturaleza mineral o geoquímica. En este mundo inicial de rocas, las moléculas orgánicas poliméricas como el ARN funcionaron como catalizadores para la síntesis de arcilla. Las cadenas de ARN cargadas negativamente se habrían asociado a las partículas de arcilla cargadas positivamente y habrían podido leer la información de los genes de la arcilla. Además, los cristales de arcilla se replican espontáneamente y proporcionan una plantilla que permite que los átomos de silicio y los iones metálicos crezcan capa a capa. El relevo genético recién tuvo lugar cuando el ARN adquirió la capacidad de autorreplicarse utilizando monómeros de nucleótidos en vez de cristales de arcilla. Paulatinamente se habría producido un relevo genético de materiales cada vez más similares a los ácidos nucleicos (polímeros más eficaces en la replicación). Se han sugerido intermedios de compuestos acíclicos en que el glicerol reemplaza a la ribosa, y también con la acroléina y el erytritol. El ancestro de estos preácidos nucleicos no presentaba un esqueleto uniforme, sino que podría consistir en un copolímero de diferentes familias monoméricas. Los químicos daneses Elgolm y Nilsen describieron una nueva molécula alternativa posible, el PNA (péptido-nucleico-ácido) en que se produce la formación de una cadena peptídica. Estos PNA están constituidos por unidades de 2-aminoetil glicina que forman un esqueleto poliamídico y peptídico sobre el cual se insertan las bases estándar de los ácidos nucleicos. Los PNA son capaces de aparearse con oligodesoxirribonucléotidos según las reglas de Watson y Crick. Otros compuestos genéticos alternativos se sintetizaron a comienzos del siglo XXI en los laboratorios, como el hexitol ácido nuleico y el alritol ácido nucleico. Todos estos compuestos presentan propiedades geométricas notables que permiten los apareamientos y la síntesis dirigida. La transición entre sistemas diferentes pudo establecerse, de esta maera, sin pérdida de la información genética. Wächterhäuser propuso tambíen la existencia de uniones hemiacetálicas establecidas entre triosas fosfatos, gliceraldehído-fosfato y dihidroxiacetona fosfato ligadas a la pirita por su grupo fosfato. Las triosas fosfato serían precursoras de las ribosas de los ARN (fosfotribosas) y habría podido fijar un núcleo imidazo para formar una bbase de Schiff (en el curso de la condensación de las funciones aldehído y cetona con las aminas primarias). Como resultado se abría obtenido el ácido tribonucleico (ATrN) formado sobre la superficie del mineral por uniones salinas con la pirita y las fuerzas del apareamiento entre bases. Wächterhäuser propuso también un precursor origina totalmente purínico. Este tipo de aapreamiento, aunque raro, existen todavía en os ácidos nucleicos fuertemente favorecido en cieras condiciones poar las fuerzas de las bases purínicas. En este contexto, la xantina es el equivalente del uracilo, la isoguanina, de la citosina y la hipoxantina a algunos ácidos nucleicos constituiría un vestigio de los ácidos nucleicos originales. Mundo de ARN, priones y controversias entre las hipótesis del proteinoide y el protogen Desde un principio, Oparin y Haldane mantuvieron puntos de vista divergentes con respecto a las condiciones iniciales y a los primeros polímeros que resultan de mayor importancia para la evolución de la vida y que surgieron en el ambiente de polimerización. En cierta medida, este desacuerdo subsiste hasta la actualidad entre sus seguidores y se relaciona con la importancia concedida a dos propiedades fundamentales de las células: la capacidad de metabolismo y la capacidad de reproducción. La capacidad de metabolismo se relaciona con las proteínas funcionales conocidas como enzimas. La capacidad de reproducción se relaciona con la presencia de los ácidos nucleicos. La hipótesis metabólica de Oparin, postula la existencia de protobionte, con el desarrollo del primer metabolismo funcional proteico protegido por algún tipo de membrana igualmente proteica (similar a los coacervados) que impedía la dilución y la destrucción por parte del medio circundante. En cambio, según la hipótesis reproductiva de Haldane, la macromolécula primordial debió permitir la obtención de copias de sí misma a partir de materiales procedentes del medio ambiente, un protogen. Indudablemente, los ácidos desoxirribonucleicos actuales son incapaces de replicarse sin la presencia de enzimas y no pueden sintetizarse las enzimas sin ellos. Por lo tanto, debió existir un origen y evolución conjunta de ambos catalizadores o, tal vez, la existencia de alguna molécula con ambas propiedades. En la década de 1980, A. Cairns Smith postuló en apoyo a Oparin que, en vez de pensar en el ADN, tenía que prestarse atención a las proteínas, que de alguna manera debieron formar un primer código genético inverso. La existencia de los priones, sustancias proteicas que producen infecciones, podrían apoyar fragmentariamente este punto de vista. La Teoría Priónica actual propone que el prión (PrPsc) es una proteína idéntica en su secuencia de aminoácidos a una proteína sintetizada por el propio organismo afectado (PrPc) aunque diferente en su conformación. La infección es debida a que la presencia de la variante patógena induce a las del organismo a plegarse adoptando la forma PrPsc. Èstas, a su vez, inducen el cambio de las restantes PrPc transmitiéndose la infección en cascada. Los análisis bioquímicos handesvelado que la PrP es una proteína globular de 253 aminoácidos, que arazón de 50-100 aa por dominio la convierten, en mi opinión, en una proteína demasiado pequeña para querérsele atribuir todas las actividades especiales que se suponen. Los priones son capaces de llegar al núcleo celular debiendo atravesar para ello los poros celulares que admiten exclusivamente un reducido grupo de moléculas. Una vez allí, tienen el poder de replegar determinadas secuencias de ADN y hacer saltar retrotransposones (segmentos de ADN), ARN diferentes o retrovirus constituyéndose únicamente en promotor para esta acción. Así, los priones pueden formar parte parte de un gran mecanismo que permite a estas proteínas alternar conformaciones teniendo cada una de ellas una función diferente. Esto permite al organismo adaptarse a una nueva situación alterando a todas las proteínas del mismo tipo dentro de la vida del individuo con la posibilidad de transmitirle a la descendencia esa nueva característica. Casi al mismo tiempo, en 1978, Carl R. Woese, el iniciador de la clasificación moderna en tres imperios, junto con Francis Crick y Leslie Orgel, los creadores del modelo molecular del ácido desoxirribonucleico de doble cadena, postularon que el ARN podría haber reemplazado al ADN en el mundo primitivo para cumplir las funciones de molde de las macromoléculas como las proteínas. En 1982, los químicos Thomas R. Cech y Sidney Altman observaron que el ARN ribosómico podía cortar segmentos (intrones) de su propia molécula sin necesidad de la presencia de enzimas, y sus propiedades catalíticas se mantenían en ciertos ARN de los ciliados y otros microorganismos (por lo tanto, actuaban como ribozimas). En esta línea de pensamiento, el ARN (ácido ribonucleico) y no las proteínas o el ADN, serían las primeras macromoléculas de la vida y formarían estadios protocelulares primitivos que consisten en vesículas de ARN como material catalítico y replicante, los protogenes. En este mundo de ARN, se puede considerar que el ADN sería un ARN modificado. Este relevo genético sería el lógico de un procesos eovlutivo en el que otras moléculas hubieran precedido al ARN y transmitido información hereditaria. En síntsis, el mundo de ARN se sustenta en tres hipótesis: • En cierta época de la evolución, la continuidad genética fue asegurada por la replicación del ARN. • La replicación descansaba en el apareamiento de bases del tipo de Watson y Crick aplicado al ADN. • Las proteínas codificadas genéticamente no estaban implicadas en la catálisis (sobre la base de la función de ribozima del ARN ribosómico). Antes de las células: de la eocélula a la metacélula Antes de la célula se consideran actualmente tres estadios evolutivos: las eocélulas (arquibios), las metacélulas (nanobios) y las precélulas (virus de ADN) como estadios intermedios que exhiben varias (aunque no todas) las propiedades vitales. Aunque no existe evidencia alguna acerca del ambiente en el que surgió la vida, es evidente que se trataría de uno caliente y ácido como el propuesto por Oparin, si bien los compuestos orgánicos que eventualmente dieron origen a los primeros seres vivos habría tenido vidas medias muy cortas. Por otra parte, mientras que las filogenias moleculares están basadas en secuencias de aminoácidos o nucleótidos, los primeros seres vivos seguramente carecían de proteínas, ribosomas, y de otras las estructuras cuyos componentes macromoleculares aparecieron en el curso de la evolución biológica; es decir, se trata de productos de un régimen evolutivo en el que operan mecanismos de evolución diferentes a los que operarían en el mundo fisicoquímico de las moléculas sintetizadas abióticamente, en donde la persistencia (o no) de los compuestos depende únicamente de su estabilidad. Los protoorganismos iniciales no fueron verdaderas células aunque ya contaban con todos los materiales de la vida, pues el código genético basado en el ADN no existía, y tampoco los mecanismos de transferencia y traducción. Esta etapa inicial de un mundo de ARN parece quedar evidenciada en los virus de ARN y, en particular, en los retrovirus. Por lo tanto, a diferencia de las células actuales, las eocélulas o ribocélulas probablemente estaban basadas en el ARN. En algún punto de la evolución prebiológica, el ARN logró sintetizar las primeras moléculas de ADN cuando las condiciones resultaron favorables para la acumulación de desoxinucléotidos (con desoxirribosa). Para que el ADN logre reemplazar al ARN, el modelo requiere la formación inicial de dobles cadenas en los ácidos nucleicos (presentes en el ADN). Eschenmoser creó en su laboratorio la molécula de piranosil-ARN, muy parecida al ARN aunque con una ribosa diferente que tenía cinco carbonos (y no cuatro) y que permitió la formación catalítica de los dobles enlaces. De esta manera, queda abierta la posibilidad de reemplazar el ARN por el ADN, con su propia capacidad de replicación y duplicación. Además, el ADN resultó más estable que el ARN. Probablemente, en la evolución celular temprana surgió un escollo insalvable: no había forma de contener el material genético global (ADN y ARN) en una protocélula muy pequeña, del tamaño de pocos nanómetros. Se postuló, entonces, la existencia inicial de metacélulas, un tipo de células que portan una parte del genoma basado en el ADN o incluso un único gen y que, obligatoriamente, deben asociarse para vivir. Además, tal vez estas metacélulas compartían como material replicativo y de herencia tanto el ADN como el ARN. El remanente de estas protocolonias de metacélulas podría hallarse en la actualidad representado por los nanobios. Al comienzo, existió cierta contradicción entre la hipótesis de Oparin y la teoría darwiniana, ya que la evolución de la vida seguía un curso aparentemente vertical, desde lo más simple a lo más complejo (y se consideraba los productores, o autótrofos, como las plantas, los organismos más simples), mientras que Oparin postulaba la existencia inicial de organismos consumidores o heterótrofos, al estilo de los animales. En la actualidad, los criterios de simple y complejo resultan obsoletos y han dado lugar a criterios basados en los genes que explican la verdadera naturaleza de la vida inicial. Sin embargo, todavía persiste la duda acerca de la naturaleza de los primeros organismos y, algunos científicos insisten en que dichos organismos eran heterótrofos, tal vez como los deinococos o bacterias resistentes a la radiación (Halobacteria), mientras que otros afirman que eran autótrofos, como las cloroflexas o bacterias verdes filamentosas (Chlorobacteria). El progenota o protocélula: primer ancestro universal (precelular) Como argumentaron Woese y Fox a partir de 1977, resulta evidente que los tres linajes celulares y los virus de ADN (incluidos los mimivirus) están separados por una enorme distancia evolutiva, todos ellos provenientes de un ancestro común (aunque la existencia de diferencias muy importantes, por ejemplo en las modificaciones que sufren las bases de los ácidos ribonucleico ribosómico (ARNr) y de transferencia (ARNt) en cada uno de estos linajes, llevó a sugerir que se trataba de una entidad mucho mas simple que cualquier procariota actual, en donde operaba una versión aún rudimentaria de la expresión de la información genética). En algún punto de la evolución, el metancestro (las metacélulas) fue obteniendo por transferencia horizontal un genoma completo (de más de 500 bases) y contó con una maquinaria enzimática suficiente que le permitió contar con un metabolismo autónomo. Inmediatamente, surgieron los ribosomas como organelas en reemplazo del ARN ribosómico (riboenzimas) libres. Se habla entonces de protocélulas o precélulas. La protocélula es considera el verdadero ancestro universal en la base del árbol filogenético (antes de la divergencia de los tres imperios y también de los virus de ADN). Se postula, además, que el ancestro precelular es común a los representantes de las dos biotas “conocidas”: Marciania (de Marte) y Terrestria (de la Tierra). Los análisis preliminares de la sonda Viking hicieron suponer que no existían organismos marcianos, pero los últimos análisis y relevamientos podrían hacer suponer que sí existe vida y, al menos, se habría clasificado una especie “celular” de metabolismo activo. Woese y Fox supusieron que en el punto la trifurcación de los tres linajes celulares había existido la mencionada entidad celular hipotética a la que ellos denominaron progenota y en la cual, a diferencia de lo que ocurre con los organismos contemporáneos, la separación evolutiva entre genotipo y fenotipo aún no se había completado. Por otra parte, de este progenota también debió haberse separado el ancestro de los virus de ADN, por la vía de los mimivirus. Estos primeros organismos celulares, junto con los mimivirus, podrían haber surgido entonces a comienzos del eón Arqueano, hace entre 4.000 y 3.800 millones de años (m.a.) luego de una prolongada evolución prebiótica a finales del eón Hadeano. Se estima que estas protocélulas fueron de tipo mesófilo (a 50 °C). Uno de los argumentos más sólidos en contra de la naturaleza termofílica de la precélula proviene de un modelo de evolución molecular desarrollado por Nicolas Galtier y sus colegas de las universidades de Montpellier y de Lyon. Aunque todas las filogenias moleculares están construídas con algoritmos que suponen que la probabilidad de substitución de los nucleótidos es esencialmente la misma, Galtier y sus colaborados hicieron uso de análisis estadísticos muchos más refinados y supusieron un proceso en donde en cada rama de cada linaje las substituciones son variables. Por otra parte, este grupo francés ya se había percatado de que el ARNr de los hipertermófilos es mucho más rico en enlaces G:C, que forman tres puentes de hidrógeno, que en el parejas A:U, en donde sólo hay dos puentes de hidrógeno. Al analizar sus bases de datos extrapolando al pasado con su modelo de substitución de ribonucleótidos, pudieron reconstruir una secuencia ancestral de ARNr 16S para el último ancestro común que tenía valores de G:C típicamente mesófilos En el esquema se observa la transición entre la protocélula (de origen mesófilo) y el cenancestro (LUCA) de características secundariamente termófilas. El árbol filogenético, muestra a LUCA cerca de la raíz. En el estadio inicial de protocélulas, los miembros de sus poblaciones cambiaban y recombinaban su material genético con gran frecuencia. Supuestamente, la capacidad metabólica y genética de las primeras entidades celulares era más simple y rudimentaria que la de LUCA (el ancestro celular) y de aquella con la que cuentan las modernas células (según estableció Karl Woese en 1998) aunque por otra parte se habría caracterizado por la ausencia de barerras genéticas (panmixia) en la transferencia genética inicial. LUCA, el cenancestro terrestre de los tres imperios celulares LUCA (del inglés Last Universal Common Ancestor, que significa, ancestro común universal último) es cenancestro o ancestro celular más reciente y común de todas células (excluidos los nanobios y probablemente también los virus de ADN). La comparación de las diferencias y similitudes que existen entre los tres linajes o dominios, es decir, los imperios Eubacteria, Archaea y Eucarya, permitió, en principio, conocer no sólo la relación evolutiva que guardan entre ellos, sino también las características de su ancestro terrestre, al que Walter Fitsch designó como cenancestro, un neologismo acuñado haciendo uso de un prefijo griego derivado de la palabra koiné, que quiere decir común. LUCA fue, probablemente, un organismo termófilo moderado (de 70 a 80 °C), a diferencia de las precélulas que originaron a los virus de ADN. Tal vez ejerció alguna suerte de quimiosíntesis en la Tierra primitiva hace 4.000 a 3.800 m.a., posiblemente en un ambiente oceánico relativamente profundo, donde el magma permitía la circulación de sistemas hidrotermales. Todo parece indicar que este cenancestro poseía, entre otras carcterísticas: (a) un sistema de transcripción y traducción de tipo moderno, con ribosomas y proteínas, factores de transcripción y una ARN polimerasa ADN-dependiente oligomérica); (b) metabolismo energético dependiente de ATPasas asociadas a membranas (proteasas); (c) posibilidad de biosíntesis completa de aminoácidos, nucleótidos y coenzimas; y (d) presencia de un genoma de ADN (heredado de las precélulas). Todo indica que el último ancestro común a los tres linajes celulares (y, por lo tanto, a los todos los seres vivos) tenía la complejidad equivalente a la de cualquier procariota contemporáneo, y no era un progenota. La reconstrucción de estadios ancestrales ha adquirido una perspectiva totalmente novedosa con la disponibilidad, a partir de 1995, de un número creciente de genomas celulares completamente secuenciados. Como habían afirmado desde 1965 Zuckerkandl y Pauling, la historia evolutiva de un organismo está contenida en su genoma –pero a menudo esta información es difícil de interpretar, debido a una serie de fenómenos biológicos que van desde la falta de preservación de la estructura primaria de los genes, hasta la existencia de niveles de redundancia de las secuencias cuya naturaleza noentendemos del todo, pasando por el transporte horizontal, una de las peores pesadillas que pueden enfrentar los biólogos– Los genes del cenancestro estarían definidos por el conjunto de secuencias presentes en la intersección de los conjuntos que representan los genomas de las Archaea, las Eubacteria y los Eucarya. Sin embargo, en la práctica esta reconstrucción se ha visto limitada por (a) el hecho de que los genomas secuenciados no representan la diversidad biológica real; (b) los distintos niveles de conservación de los genes, que distan mucho de ser los mismos para todas las secuencias; (c) los problemas para anotar, es decir, identificar a las secuencias presentes en las bases de datos; (d) la presencia de secuencias altamente conservadas cuya función es aún completamente desconocida por no disponer de datos experimentales; (e) la pérdida polifilética independiente de secuencias, funciones y rutas metabólicas que han sufrido diversos organismos, sobre todo parásitos y simbiontes; y (f) el movimiento lateral de los genes, que en algunos casos pueden traspasar las fronteras que separan a los grandes dominios. La evolución horizontal o transferencia "horizontal" de genes es el proceso mediante el cual uno o más genes abandonan a una especie y son transferidos a otra, puede llevar a nuevos errores. Es probable que haya sido un proceso común, no sólo en las metacélulas sino también en la línea progenota-LUCA y, tal vez, hayan participado o sean sus eslabones los virus de ADN. Patrick Forterre, de la Université de Paris-Sud, en Francia, propuso en la década de 1990 un complejo esquema de transporte lateral de genes que involucra a los tres linajes y a sus virus de ADN. La transferencia horizontal de secuencias génicas es entonces un fenómeno real (que subyace, por ejemplo, la bien conocida resistencia a antibióticos que se observa con una frecuencia cada vez más alarmante entre muchos patógenos de humanos y animales) y que puede oscurecer la reconstrucción del pasado evolutivo. • Si los genes que se transfieren son muy numerosos, el "árbol de la vida" se parece más a una "red de la vida", y puede no ser posible distinguir las ramas. • Además, si los genes son altamente móviles, los genes verdaderamente "universales" (o sea, luquianos de origen) no podrán ser diferenciados de aquellos que tienen más éxito para transferirse de una especia a otra. Se conoce, por ejemplo, que incluso los operones del ARNr pueden llegar a sufrir este fenómeno, pero el hecho de que se mantenga la identidad de los grupos de microorganismos (es decir, las espiroquetas siguen siendo indentificables como tales, no hay bacterias Gram positivas que se hayan transformado en Gram negativas, etc.) sugiere que existen limitaciones biológicas al intercambio de genes. En 1999, por ejemplo, tres grupos de trabajo demostraron, en forma independiente, que la representación gráfica del contenido total de secuencias (es decir, fenogramas) en los genomas secuenciados mostraba como, a pesar del transporte horizontal, no sólo los tres dominios, sino incluso las distintas líneas biológicas que las constituyen, se pueden identificar sin problemas. Es decir, a pesar de la promiscuidad con la que los seres vivos han intercambiado genes a lo largo de la evolución, la reconstrucción del pasado es posible. El problema de la transferencia de genes fue estudiado en profundidad: en 1998, Jeffrey Lawrence y Howard Ochman compararon los genomas de dos bacterias comunes, Escherichia coli y Salmonella. Las dos especies se separaron hace 100 millones de años a partir de un antepasado común, pero sus genomas se formaron de manera muy distinta, ya que sufrieron no menos de 200 aportes de genes ajenos a ellas mediante el proceso de transferencia horizontal. ¡Eso representa el 10% del genoma de cada una! La preocupación de los lucabiólogos es que LUCA fuese totipotencial (como un huevo fertilizado, que puede generar músculo, neuronas, células óseas, etc.), lo que lo convertiría en la fuente última de la diversidad biológica de nuestro planeta. Por el contrario, si todos los genes se mueven horizontalmente todo el tiempo, es posible que LUCA no fuese totipotencial como un óvulo fecundado. Sin embargo, así no lo parecería, porque muchas características que se han transmitido horizontalmente no parecen universales y transmitidas por herencia de generación en generación. Los resultados de los estudios sobre las dos bacterias pueden falsearse por completo si se las proyecta un par de miles de millones de años en el pasado. ¿Y si el mecanismo original era horizontal, aunque hoy la mayoría de los genes se heredan verticalmente? Karl Woese desarrolló una idea original: si los genes primigenios eran completamente libres de migrar de una especie a otra, entonces no existían diferentes linajes. Así, ni siquiera sería posible distinguir distintas especies en aquellos días, porque los mismos genes se mudaban de una a otra libremente. A medida que la complejidad de los organismos aumentaba, y a que la precisión de las funciones de los genes se hacía cada vez mayor, los genes comenzaron a hacerse interdependientes unos de otros, y la transferencia horizontal dejó paso a la herencia vertical. Un gen no podía simplemente irse a otra especie, porque dependía de otros genes a los que no podía dejar atrás, y otros más dependían de él. ¿Cómo fue quedando fijado el patrimonio genético de cada especie? Woese afirma que lo primero que quedó "establecido" (en genética, "cristalizado") fue el proceso de traducción, por el que la información guardada en el ARN codifica a una proteína. Las demás funciones celulares cristalizaron más tarde. Hay científicos que opinan que la transferencia horizontal constituye un punto álgido en la discusión de este problema, y que se transformará en un escollo que confundirá durante mucho tiempo a quienes intentan reconstruir a LUCA. El asunto es muy complejo: * Los métodos para reconstruir árboles evolutivos son limitados, y pueden entregar evidencia falsa de transferencias horizontales que en realidad no existieron. * Para estudiar relaciones genéticas se utilizan métodos que no incluyen el análisis de la información evolutiva. En muchos casos, los datos se utilizaron para argumentar que la transferencia horizontal era muy poco notable. * La fiabilidad de los métodos para determinar si hubo o no transferencia horizontal goza hoy de muy poco consenso entre los científicos. * No quedan claros realmente los datos necesarios para comprobar una transferencia horizontal. * Si se tiene en cuenta la selección natural, probablemente la mayoría de las transferencias horizontales resultarán en la eliminación del gen transferido. Ejemplo: los genes de la resistencia al antibiótico no se extenderán, a menos que el organismo sea atacado por ese antibiótico específico. ¿Para qué migrar los genes en un ambiente donde no hay antibiótico? Muy a menudo los investigadores ignoran esta última circunstancia, y casi no se han hecho esfuerzos por establecer un patrón de referencia, como por ejemplo: "¿Presentan todos los genes igual tendencia a la migración o al nomadismo?". Entonces, la pregunta es: ¿Cómo deben tratar los lucólogos a la transferencia horizontal? * Si se acepta la existencia de un furioso y masivo intercambio horizontal de genes entre los tres imperios se debe aceptar que las herramientas actuales para observar el pasado de los árboles evolutivos ya no resultan válidas, lo que significa que se debe abandonar la investigación de LUCA. Se sabe que la transferencia horizontal presenta casos bien demostrados, aunque este caso extremo aún no se ha comprobado. * Si se toma como punto de partida el caso extremo opuesto, es decir, que el efecto horizontal ha sido despreciable, la posición es mejor. No hay duda de que se puede atacar desde una postura intermedia, pero en medio de la lucha acerca de la transferencia horizontal, muchos investigadores hicieron uso de genomas completos y llegaron a la conclusión de que la horizontalidad tuvo un efecto mínimo sobre nuestra capacidad de reconstruir árboles evolutivos. Estos resultados sugieren que sería posible reconstruir el árbol de la vida y, por último, llegar a la conclusión de que la estructura del árbol (tres imperios) está apoyada por las estructuras de los genomas completos de los organismos que lo componen. La pérdidas de genes, además de la transferencia horizontal, constituye un problema crucial para dilucidar la estructura genómica de LUCA. Si un gen en efecto estaba en LUCA, pero hoy en día sólo uno de los tres imperios lo conserva, entonces quedará fuera de la lista de "genes universales" y por supuesto, fuera del "genoma de LUCA". A pesar de estas grandes y evidentes limitaciones, el concepto del genoma mínimo posiblemente sea el único método para la aproximación a la lista central de genes que han formado parte del patrimonio de LUCA. Es también el único marco útil disponible actualmente. Un trabajo de Koonin recientemente publicado lista un grupo de genes mínimos para LUCA. Esta vez, en vez de comparar los genomas de dos especies lo ha hecho con 21. Sorprendentemente, de los 256 genes de la primera lista, sólo 81 han llegado a la segunda. La nueva lista de Koonin no sólo es claramente escasa para formar el genoma de LUCA, sino que es también insuficiente para que una célula cualquiera llegue siquiera a funcionar. Para algunos científicos, la conclusión es evidente: LUCA no almacenaba su información en ADN. Y coherente con este concepto, el grupo de Arkadt y Koonin sostiene que este último ancestro común poseía todavía un genoma de ARN, o tal vez de ARN/ADN (al igual que las ribocélulas y tal vez también las metacélulas y los nanobios). Independientemente de si LUCA realmente ya tenía ADN completo además de ARN, en esta etapa de la evolución que ya puede considerarse biológica, se fijó el código genético universal en base al ADN. Un grupo de la Facultad de Ciencias de la UNAM, que ha podido identificar la presencia de dominios catalíticos de distintas ADN polimerasas en los tres linajes, cree haber identicado partes de la ADN polimerasa ancestral, que probablemente se derivó, como lo sugiere su homología con algunas ARN polimerasas, de una molécula que originalmente estaba involucrada en la replicación de genomas de ARN que poseían estas células más sencillas que precedieron al cenancestro. Estas conclusiones, de hecho, son congruentes con los resultados preliminares obtenidos por Fredj Tekaia, Bernard Dujo y Antonio Lazcano, que muestran que las secuencias más conservadas comunes a los genomas celulares disponibles codifican para secuencias que sintetizan, degradan o interaccionan con ARN –es decir, que se pueden reconocer, al menos en parte, evidencias de un pasado biológico anterior a la aparición del ADN mismo–. La replicación y la traslación evolucionaron en esta fase y formaron el núcleo del cual evolucionaron los genomas de los ancestros de cada imperio. Se ha propuesto un período de evolución denominada “época monofilética”. Durante este periódo, existían diferentes líneas independientes (que carecían de mecanismos de transferencia) y la evolución tenía lugar por mutación y selección. Un grupo de hipótesis supone que hubo un evento catastrófico gigantesco que condujo al efecto cuello de botella en los sobrevivientes. Así, estos sobrevientes podrían haber sido los representantes de los tres imperios de vida de Woese. Esto explicaría la dispersión de los organismos termófilos e hipertermófilos entre los grupos más primitivos de cada uno de los imperios. ¿Cuáles son los principales sucesos en la historia celular? A partir de LUCA, la vida celular evolucionó en la Tierra hace, aproximadamente desde 3.850 m.a., es decir, a comienzos del eón Arqueano. Y ya cerca de los 1.500 m.a., en pleno Proterozoico, los organismos eucariotas ya habían evolucionado (anque su origen tal vez se remonte a 2.000 m.a.). Y si continuamos hacia los comienzos, hace 2750 m.a. hicieron su aparición las cianobaterias o algas azulverdosas (procariotas) fomentando una expansión y diversificación de los estromatolitos (que inicialmente producían las bacterias verdes no azufrosas). Hacia 3.100 m.a., ya habían surgido las primeras arqueas tal como indican los marcadores de metano (ya que las arqueas son los únicos organismos metanógenos o productores de metano). En cambio, las bacterias debieran remontarse directamente a LUCA, al menos desde hace 3.500 m.a., y entre las primeras se contaron las bacterias verdes no azufrosas (como se mencionó), es decir, las primeras formadoras de estromatolitos, y las radiobacterias, o bacterias resistentes a las radiaciones. No obstante, el registro fósil de los organismos que vivieron antes de los 550 millones de años es fragmentario, aunque es suficientemente bueno como para mostrar que el número total de especies y de individuos se incrementó radicalmente en el Precámbrico tardío. Como vimos anteriormente, los geólogos pre-darwinianos dividieron la historia geológica en eras y períodos basados en sus conjuntos fósiles distintivos Varios procesos contribuyen a la escasez de fósiles Sólo una pequeña fracción de los organismos que alguna vez se convierten en fósiles, y sólo una ínfima parte de dichos fósiles son alguna vez estudiados por los paleontólogos. La mayoría de los organismos viven y mueren en ambientes ricos en oxígeno en los que rápidamente se descomponen. No es probable que se vuelvan fósiles a menos que sean transportados por el viento o el agua a sitios sin oxigenación, en los que la descomposición avanza lentamente o no se produce. Más aún, los procesos geológicos a menudo transforman las rocas y destruyen los fósiles que contienen, y numerosas rocas fosilíferas se hallan profundamente enterradas e inaccesibles. Los paleontólogos han estudiado sólo una muy pequeña fracción de aquellos sitios que contienen fósiles, pero encuentran y describen nuevas especies fósiles cada año. El número de especies fósiles conocidas resulta especialmente elevado para los animales marinos que poseían esqueletos duros (que resisten la descomposición). Entre los nueve grupos de animales con representantes que poseen partes duras, aproximadamente 200.000 especies se han descrito como fósiles – apenas el doble de las especies marinas vivientes de estos mismos grupos. Los paleontólogos se basan fuertemente en estos grupos para sus interpretaciones de la evolución de la vida. Los insectos y las arañas también están relativamente bien representados en el registro fósil. El registro fósil, aunque incompleto, es suficientemente bueno para demostrar claramente que organismos de tipos particulares se encuentran en rocas de edades específicas y que los organismos más nuevos aparecen secuencialmente en rocas más jóvenes. Al combinar la información de los cambios geológicos durante la historia terrestre con la evidencia del registro fósil, los científicos han compuesto representaciones de cómo pudieron ser la Tierra y sus habitantes en diferentes tiempos. Sabemos en general dónde se hallaban los continentes y cómo la vida cambió durante el tiempo, si bien muchos detalles son poco conocidos, especialmente aquellos sucesos del pasado más remoto. Chaotiano y Hadeano en foco El sistema solar tuvo su origen en una nebulosa compuesta de polvo cósmico, átomos y moléculas de elementos químicos pesados y gases (hidrógeno, neón, criptón, argón, etc.) por los efectos de su movimiento rotacional, esta nebulosa tomó la forma de un enorme disco en cuyo interior reinaba gran turbulencia. La acción de las fuerzas rotacionales motivó que las partículas más pesadas se agruparan en el centro de la nebulosa y cosntituyeran un protosol. Otras condensaciones de materia ocurrieron en el resto del disco y pronto, el protosol se halló rodeado por una concentración de partículas sólidas de distinta masa, denominadas planetesimales. La colisión entre planetesimales motivó una reducción de su número ya sea por pulverización o por fusión entre ellos, lo que condujo a la formación de los planetas interiores o terrestres ((Mercurio, Venus, Tierra, Marte). La región más externa, gaseosa, de la nebulosa dio lugar a los planetas jovianos o mayores (Júpiter, Saturno, Urano, Neptuno), de poca densidad y gran tamaño. Es probable que especialmente Júpiter y Saturno tengan una composición media similar a la del Sol, y por lo tanto, muy semejante a la que tenía la nebulosa primordial hace unos 5.000 m.a. el hidrógeno sería el principal componente de estos planetas, junto con cierta proporción de helio, metano y amoníaco. Los planetas terrestres, más densos que los jovianos, se habrían formado a temperaturas iniciales relativamente bajas si bien posteriormente las mismas se elevaron por la radioactividad interior. La edad del Sistema Solar se puede conocer mediante la determinación de la edad de los meteoritos, pues estos se originaron juntamente con el resto del Sistema Solar. Según los cálculos basados en las relaciones uranio/helio, rubidio/estroncio y uranio/plomo de los meteoritos, llegados a la Tierra, el Sistema Solar tendría una edad cercana a los 4.600 m.a. La elevación de la temperatura interior terrestre dio lugar a la diferenciación de un núcleo de hierro y níquel (Nife) y una capa periférica más liviana, de silicatos que constituyó la primera corteza terrestre, en la que predominaron rocas basálticas, gábricas y anortosíticas. Restos de esta corteza primitiva se hallarían conservados en los núcleos arqueanos de los continentes. El Arqueano en foco Durante el Arqueano, los océanos primitivos se mantuvieron ácidos y calientes, pero ahora con temperaturas que oscilaban entre 60 y 80 °C, mientras que la atmósfera, todavía muy rica en dióxido de carbono, mantenía temperaturas superficiales elevadas sobre los continentes, similares a las que se observan en el planeta Venus (al menos 200 ºC). Los terrenos arqueanos contienen dos asociaciones rocosas principales: una muy metamorfizada y otra de escaso metamorfismo, a la que pertenecen los "greenstones" o rocas verdes, en su mayoría originadas entre los 2.800 y los 2.600 m.a., y que se conocen principalmente en Sudáfrica, Rhodesia, Australia Occidental, Canadá, Estados Unidos de América, India y Finlandia. Estratigráficamente, en las rocas verdes se reconocen tres unidades: 1- basal, de lavas máficas y ultramáficas 2- media, de andesitas y vulcanitas silíceas de tipo calco-alcalino 3- superior, de conglomerados, areniscas, turbiditas, depósitos ferríferos, ftanitas (cherts) y calizas precipitadas químicamente. Estas rocas contienen gran riqueza de oro, plata, cromo, níquel, cobre, hierro y zinc, y de ahí su importancia económica. La mayoría de las rocas arquezoicas sufrieron importantes procesos de deformación y metamorfismo, por lo que se conocen pocos lugares en los que su edad supera los 3.200 m.a. como los gneisses de Groenlandia y labrador (3.750 m.a.) de Minesota (USA), Noruega, Peninsula de Kola y Africa (Rhodesia, Sudáfrica). Los datos paleomagnéticos indican que la primitiva corteza terrestre arqueano ya estuvo sometida a los procesos de las derivas continentales. Las rocas arqueanas contienen a las más antiguas evidencias de vida, la cual se habría originado apenas el planeta se constituyó. El límite Arqueozoico-Proterozoico (2.500 m.a.) tiene extraordinaria significación en la evidencia de la corteza, pues refleja un momento de gran disminución del calor radiogénico, lo que derivó en una mayor estabilidad y menor importancia de las deformaciones tectónicas y el metamorfismo. Así, las rocas volcánicas en su mayoría se redujeron a basaltos toleíticos, andesitas, riolitas y piroclastitas, mientras que las rocas sedimentarias adquirieron mayor preponderancia y se acumularon en cuencas en constante ampliación, constituyendo depósitos de gran espesor, dado que la corteza ya era suficientemente espesa y rígida para soportarlos. Entre las secuencias sedimentarias proterozoicas que revisten particular interés están las "Formaciones ferríferas zonadas" (FFZ) y los "estratos rojos" ("red beds"). Las primeras aparecen a partir del Arqueozoico, pero su principal desarrollo tuvo lugar en el lapso 2.600 - 1.800 m.a.. se caracterizan por la alternancia de capas de ftanitas criptocristalinas con otras ferríferas, en las que el hierro se halla como carbonato, silicato, óxido o hidróxido. Suelen alcanzar centenares de metros de espesor y su depositación se habría producido en cuerpos acuosos (marinos?) a los que el hierro llegó en estado ferroso para allí precipitar como férrico mediante el agregado de oxígeno proveniente de la fotólisis o más probablemente, de los primeros procesos fotosintéticos. El oxígeno de origen fotosintético también oxidó a los estratos rojos, cuyos granos de arena están cubiertos por hematita. Este tipo sedimentario comenzó a formarse hace unos 2.500 m.a. en Sudáfrica y luego se extendió a otras regiones. En el Proterozoico, los núcleos arqueanos quedaron rodeados por los cinturones móviles o fajas orogénicas de gran complejidad estructural, cuyas rocas poseen edades de 2.500 m.a. o menos. Originalmente fueron áreas tectónicamente muy activas hasta que se fusionaron a los núcleos antiguos, transformándose en regiones estables (cratones). Su origen estuvo vinculado a la formación de los primeros geosinclinales típicos, ubicados en los bordes de las placas y no en el interior de los mismos como eran los geosinclinales intracontinentales arqueanos. Esta etapa se caracteriza por la formación de los núcleos continentales graníticos, base de las futuras placas litosféricas. Los primeros testimonios fósiles fueron estromatolitos, depósitos bandeados producidos por la actividad de las primitivas bacterias fotoiltótrofas y quimilitótrofas anaerobias (no respiradoras de oxígeno), asociadas con las hadobacterias (como Deinococcus y Thermus) y las chloroflexibacterias o bacterias verdes no azufrosas (como Anaerolinea y Chloroflexus). Como ya se mencionó, el primer fósil que se descubrió en un chert, roca metamórfica bandeada es Eobacterium, de Sudáfrica, probablemente una hadobacteria. Otros restos fósiles podrían corresponder a bacterias verdes, y según algunos autores como Knoll y Woese también a las primeras cianobacterias o algas azules, o tal vez sus ancestros, todas unicelulares. El Proterozoico en foco La primera parte del eón Proterozoico, de los 2.500 a los 1.500 millones de años (era Proterozoica I) está marcada por la convergencia de los antiguos núcleos continentales hasta formar escudos mayores, como las placas Atlántica y Columbia. Entre los 1.950 y 1.590 millones de años se produjo el primer fenómeno de rift océanico o formación de un oceáno interior y el depósito de las primeras rocas carbonatadas, indicando un descenso de la acidez oceánica. En el Proterozoico, las tillitas documentan la existencia de ambientes glaciales. Las colisiones continentales, en el proterozoico originaron importantes orogenias, como la Grenville, de hace unos 1.100 m.a., coincidente con las ocurridas en otras regiones del mundo, lo que habría hecho posible para esos tiempos, la reunión de la mayoría de los continentes en un supercontinente, Rodinia. La existencia de un Supercontinente en el Mesoproterozoico Tardío se hallaría avalada por los datos paleomagnéticos, la similitud de las historias tectónicas de sus partes constituyentes, la distribución de los orógenos y de ciertos tipos de rocas (granulitas, anortositas, FFZ, tillitas). T.H. Torsvick et al., (1996) consideran que Rodinia se originó hace unos 1.100 m.a. y que su desintegración comenzó hace unos 750 - 725 m.a.. Su reconstrucción de Rodinia (750 m.a.) difiere de la de Dalziel (777 m.a.) en la posición de la Placa Báltica (Escandinavia + Spitzbergen + Rusia + Ucrania) con respecto a Laurentia y en la ubicación paleogeográfica total de Rodinia, la que se encontraría mayormente en posición ecuatorial y girada en sentido horario. En el lapso 750-725 m.a., la parte meridional de los continentes (India + Antártida Oriental + Australia) inició su separación del borde occidental lauréntico y se originó el Protopacífico. El resto de Rodinia rotó en sentido horario y se movió hacia el sur, por lo que en el Véndico Temprano (aproximadamente 650 m.a.) la glaciación varangiana afectó a la región báltica, groenlándica y africana occidental. Mientras se reconstruía un nuevo continente hacia 600 m.a., la Pannotia. Las partes centrales de Laurentia se hallaban en la zona templada, pero, al continuar el desplazamiento hacia el sur se produjo la glaciación Brook (aproximadamente 610 - 590 m.a.). Partes de Báltica permanecieron glaciadas si bien, en Siberia no se conocen depósitos glacígenos neoproterozoicos. Otra separación importante ocurrió entre Báltica - Amazonia y Laurentia cuando, hace unos 600-580 m.a., se produjo la apertura del Océano Iapetus, aunque ambos continentes continuaron en latitudes australes (altas). Sin embargo, hacia los 550 m.a., Laurentia se hallaba en latitudes bajas y permaneció allí hasta el Ordovícico. Entre Báltica y Amazonia se abrió el mar Paleotornquist. Gondwana (incluyendo al Gondwana Occidental que se había separado de Laurentia) casi completo, se extendía desde las altas latitudes australes (Gondwana Occidental) hasta las intermedias de más al norte (Gondwana Oriental). La integración del Supercontinente Pannotia está poco esclarecida. En general, se la ubica en el Neoproterozoico, finalizando a los comienzos de Paleozoico, y este suceso pudo estar asociado a la previa acreción de numerosos bloques menores y al cierre de muchos océanos y mares. Finalmente, ocurrió una acreción a gran escala entre la región conocida como Nigritia o Protogondwana Oriental y Occidental a lo largo del Cinturón orogénico de Mozambique, constituyéndose el gran continente Nigritia. Las señales características de esta etapa fueron las condiciones estables y la meteorización prolongada en climas húmedos y cálidos y las formaciones de hierro bandeado por la actividad de las las algas azules que producían oxígeno que inmediatamente oxidaba los depósitos océanicos, y más tarde continentales, del ion ferroso para transformarlo en ion férrico. Este cambio estuvo marcado por el registro de un nuevo tipo de estromatolitos, cupulares o en forma de domo, que acompaña la expansión de las cianobacterias o algas azules, productoras de oxígeno. Estaba gestándose de esta manera el inicio del gran cambio que sobrevendría en la atmósfera terrestre, al desplazar el oxígeno al dióxido de carbono y otros gases de invernadero en forma paulatina. En este agitado marco geográfico tuvo lugar una diversificación inicial y luego una gran extinción de los primeros organismos anaerobios (arquibacterias y bacterias primitivas) por la acumulación de oxígeno. En estos antiguos mares se diversificaron los procariotas aerobios, representados por las bacterias azufrosas verdes y las cianobacterias, alcanzando formas extrañas, como Eostrion, que parece una estrella, y Kakabekia, que parece un paraguas, además de las primeras formas filamentosas pluricelulares. Entre 1.800 y 1.500 millones de años, aparecieron los primeros testimonios de organismos eucariotas y de sexualidad, al evidenciarse tétrades de células formadas por meiosis. La segunda parte del eón Proterozoico (era Proterozoica II), de los 1.500 a los 900 millones de años, se carateriza por la culminación de la convergencia de los antiguos núcleos continentales, hasta formar un gran supercontinente, la Rodinia, bañado en toda su extensión por un gigantesco océano, Mirovia. En Rodinia se comienzana distinguir dos ramas, una septentrional y otra meridional (esta última dará lugar a Nigritia, gran continente en que quedan englobadas África y Sudamérica). Hacia el final, se formó una placa, la Protoantártica, que se separó del sector de la Nigritia y quedó formada de esta manera la cuenca del Pantalasa, antecedente del océano Pacífico. Además de este gran supercontinente, existían pequeñas islas, como la Arqueoplata, donde se encuentra actualmente la provincia de Buenos Aires, separada de Sudáfrica por el antiguo océano interior, el Adamastor. En este marco geográfico, el oxígeno comenzó a acumularse en la atmósfera al haberse agotado los depósitos de hierro en forma de ion ferroso. Y como resultado se produjo un intenso enfriamiento del clima que culminó hacia los 900 millones de años con la glaciación Varangiana. En este marco se expandieron los eucariotas apareciendo los primeros protozoos, los dinoflagelados, los primeros hongos, las algas verdes y rojas, y hacia el final los primeros animales, las esponjas. En la porción terminal del eón Proterozoico, la era Ediacariana, o era Proterozoica III) se distinguen dos períodos, Rifeano, hasta los 700 millones de años, y el Vendeano o Edicariano, hasta el final. Se inició hacia 850 m.a. una nueva fusión continental, para formar el continente transversal conocido como Pannotia. Pero a partir de 570 m.a., con una nueva fragmentación continental y un calentamiento del clima, hizo que se volvieran a fragmentar las placas, que incluían la Nigritia, la mayor, y otras siete menores, principalmente al norte del ecuador, Lauréntica, Báltica, Siberiana, Kazhakstana, Centrochina, Sinocoreana e Indochina. Un nuevo océano, antedecente del Atlántico, el Iapetus, se formó entre Lauréntica y Nigrita. El derretimiento de los depósitos glaciares y el aumento del nivel de oxígeno condujeron a la formación de mares de plataforma, el depósito de rocas carbonatadas y la acumulación de sales minerales, como los fosfatos. Al incorporar nuevos nutrientes al mar, pudo diversificarse la vida animal y el plancton. De esta etapa es notable la fauna de Ediacara, caracterizada por organismos muy primitivos, los Vendozoa, de gran diámetro y escaso espesor, con un sistema acolchado de filtros, como Dickinsonia y Pteridinium. También aparecieron los primeros Metazoa (metazoos o animales verdaderos) representados inicialmente sólo por los cnidarios (pólipos y medusas) y más tarde platelmintos y grupos primitivos de celomados. Representación de la biota de finales del Proterozoico. El Proterozoico contiene mucha información sobre la evolución de la vida y los procesos físicos que condujeron a la consolidación de la corteza terrestre. En muchos lugares estos tiempos finalizaron con intensos movimientos orogénicos que originaron marcadas discordancias angulares con los cámbricos suprayacentes, pero en otras áreas, como Marruecos, namibia, Siberia, SE de China, Australia, América del Norte, etc., se conocen secuencias sedimentarias proterozoico-cámbricas aparentemente ininterrumpidas, en la que resulta casi imposible determinar con precisión el límite entre las rocas de ambas edades. En Meishuncun, SE de la China, se ha propuesto como estratotipo de dicho límite a un nivel donde ocurre una marcada diversificación de los organismos con conchillas pequeñas, aunque también se lo ha hecho coincidir con un horizonte en la base de la Formación Quiangzhusian, donde se pasa abruptamente de depósitos carbonático-fosfáticos a arcillitas y silstitas y que además, muestra uhna aparente anomalía de iridio y un cambio importante del 13 ªC hacia valores negativos. También se han propuesto otras localidades como tipos de este límite, en la Plataforma Siberiana, América del Norte y Norte de Africa. Los estromatolitos son utilizados para el reconocimiento de biozonas proterozoicas. Son estructuras biosedimentarias cuyo origen se debe a algas cianofíceas (cianobacterias) que habitaron las costa áridas, con gran evaporación. Sobre los fondos sedimentarios, las bacterias generan una película orgánica en la que habitan y se reproducen. Rápidamente esta capa bacterial es cubierta por los mares, depositándose sobre ellas una lámina sedimentaria que a su vez, es cubierta por otra capa orgánica y así sucesivamente. Todo el conjunto resulta cementado por carbonato tomado por las bacterias desde el agua, en la que se encuentra como bicarbonato. Los estromatolitos más antiguos conocidos son los de Australia occidental (Serie Warrawoona, 3.500 m.a.) y Sudáfrica (Supergrupo Swaziland, 3.500 - 3.300 m.a.), pero también se los encuentra en rocas más jóvenes y actualmente se están formando en las zonas marinas litorales de las bahamas, Bermudas, Australia, etc., como también en otras no marinas (fluviales, lacustres termales). Generalmente los estromatolitos son calcáreos (excepcionalmente silíceos) y de morfología variada, diferenciándose en estratiformes, nodulares y columnares. Se discute si estos rasgos se deben a características ambientales o están controlados genéticamente, resultando así útiles para las correlaciones estratigráficas. Hasta ahora, sólo los columnares parecerían importantes en tal sentido, siendo empleados en biozonaciones del Proterozoico en Australia, Rusia, Siberia, China y América del Norte. Las glaciaciones arqueanas y proterozoicas La presencia de horizontes tillíticos en secuencias arquezoicas y proterozoicas indicaría la existencia de episodios glaciales en esos tiempos. Las evidencias más antiguas se hallarían en el Arqueozoico sudafricano, donde el Supergrupo Witwatersrand tendría varios niveles tillíticos con edades entre 2.720 y 2.340 m.a. Otros depósitos dudosos se citan en América del Norte, Carelia y los Montes Baikal. Tillitas paleoproterozoicas se conocen en América del Norte, India y Australia Occidental. En América del Norte habrían existido dos períodos glaciales. La depositación de las tillitas paleoproterozoicas fue seguida por un período sin glaciaciones hasta que en el lapso 1.000-600 m.a., éstas volvieron a producirse con intensidad registrándose en todos los continentes actuales, excepto Antártida. En Africa se habrían producido 5 acontecimientos glaciales y en Asia, por lo menos 3. En Australia, la secuencia tillítica más desarrollada es la del geosinclinal Adelaida (Montes Flinders) donde alcanzaría 6.000 m de espesor, diferenciándose dos períodos glaciales: Sturtiano (800 m.a.) y Marinoano (700 m.a.). La mayoría de las glaciaciones neoproterozoicas europeas corresponden a la glaciación Varangiana o Laplandiana, cuya localidad clásica se halla en Noruega, y que ocurrió principlamente durante el Vendiano, aunque algunas tillitas son rifeánicas. Esta glaciación sería equivalente a la Glaciación Brook (600 m.a.) de Laurentia. Gran parte de las tillitas o diamictitas sudamericanas están en Brasil (Grupos Macaubás, Jangada, Bambuí). Algunas tillitas se ubican cerca del límite con el Cámbrico. Otras tillitas se hallan en el Paraguay Oriental (Formación Itapocumi) y Bolivia (Grupo Boqui). Mientras que para algunos autores (Young, 1995) las glaciaciones proterozoicas se vincularían con el tectonismo y la fragmentación de Rodinia o por la meteorización de un supercontinente tropical (proceso que habría consumido grandes cantidades de dióxido de carbono de la atmósfera), aunque otros autores autores sostienen que ellas habrían estado controladas climáticamente. No obstante, parecería que algunos depósitos glacígenos ubicados en latitudes bajas pudieron originarse en los márgenes elevados de los rifts. La vida en evolución durante los tiempos precámbricos Durante la mayor parte de su historia, la vida se hallaba confinada a los océanos, y todos los organismos eran pequeños. Durante las prolongadas edades del Precámbrico – hace más de tres mil millones de años – los mares someros lentamente comenzaron a bullir de vida. Durante la mayor parte del Precámbrico, la vida consistió en procariotas microscópicos; los eucariotas probablemente evolucionaron transcurridos los dos primeros tercios de este prolongado “período”. Los eucariotas unicelulares y los pequeños animales multicelulares se alimentaban de microorganismos fotosintéticos flotantes. Estos pequeños organismos flotantes, conocidos colectivamente como plancton, eran consumidos por animales ligeramente mayores que los filtraban del agua. Otros animales ingerían sedimentos del lecho marino y digerían restos de organismos dentro de sus cuerpos. Hacia fines del Precámbrico (hace cerca de 650 m.a.), muchas clases de animales multicelulares de cuerpo blando habían evolucionado. Algunos de ellos eran muy diferentes de cualquier animal actual y podrían ser miembros de grupos sin descendientes vivos en la actualidad.. Existe bastante disparidad de opiniones con respecto al momento en que la atmósfera comenzó a tener suficiente oxígeno como para posibilitar la existencia de la célula eucariota. Según algunos especialistas, una atmósfera oxidante habría existido hace 3.750 m.a., a juzgar por la presencia en rocas de esa edad, de minerales de uranio, cobre, cinc, etc., que no podrían haberse formado bajo una atmósfera reductora. Sin embargo, indiscutibles eucariotas recién aparecen en rocas de unos 900 m.a. de antigüedad, mientras que las evidencias fósiles y bioquímicas mostrarían que la célula eucariota era una parte imporante de los ecosistemas hace, por lo menos hace 1.900-1.700 m.a.. Con anterioridad los datos son más imprecisos y, en la Formación Guntflint (2.000 m.a.) habría restos de posibles eucariotas. Según A.H. Knoll (1992) las filogenias moleculares basadas en las proteínas y el rRNA indican que los eucariotas emergieron tempranamente en la historia de la vida y que la ausencia de evidencias paleontológicas se debería a la pobreza del registro fósil del Arqueano y Proterozoico más temprano y al escaso potencial de preservación de los eucariotas más primitivos. El procedimiento por el cual se logró el primer eucariota es desconocido. Lynn Margulis (1971) emitió una ingeniosa teoría al respecto basada fundamentalmente en las endosimbiosis hereditarias (Figura): un procariota ameboidal por ingestión de bacterias aeróbicas habría dado lugar a un nuevo procariota en el que los organismos incorporados se habrían convertido en mitocondrias. Seguidamente se habría producido la inclusión de bacterias filiformes o espiroquetas que se transformaron en flagelos y centríolos, y la diferenciación de una membrana nuclear dio lugar a un eucariota unicelular incoloro primitivo. Tal organismo, por evolución, habría conducido a los animales mientras que los vegetales fueron el resultado de una tercera etapa ingestiva con incorporación de cianofíceas al eucariota primitivo. La aparición de la membrana nuclear permitió la distribución adecuada del ADN recién sintetizado entre las células hijas, favoreciendo el desarrollo de los cromosomas y además, sirvió para proteger al delicado material genético de la acción del oxígeno requerido para la respiración celular. A partir del momento de su aparición y durante el resto del Proterozoico, los eucariotas unicelulares efectuaron una rápida evolución que incluyó el perfeccionamiento de la sexualidad y la adquisición del grado multicelular de organización. Es probable que los primeros organismos con reproducción sexual hayan tenido alternancia de generaciones asexuales y sexuales cumpliendo cada una de ellas una función diferente. La reproducción sexual permitiría al organismo afianzar su dominio en un medio previamente conquistado por otros individuos pertenecientes a la fase asexual. La reproducción sexual produce gran cantidad de gametas y cigotas ligeramente diferenciadas unas de otras, de manera que en la invasión de un nuevo hábitat la selección natural favorecerá a aquellos mejor adaptados a éste. Seguidamente, la reproducción asexual será la encargada de saturar el medio ambiente con formas similares a los progenitores. La causa principal que debió favorecer el surgimiento de la multicelularidad probablemente fue el valor adaptativo del aumento de tamaño lo que permitió una mayor variabilidad de las formas y un incremento de la especialización de las distintas partes del organismo que condujo a la formación de los tejidos y la elaboración de una cavidad interior o celoma. Evidencias de multicelularidad en organismos microscópicos simlares a las algas rojas y verdes, existen a partir del Proterozoico Medio, siendo abundantes en el Proterozoico tardío. Al final del Proterozoico también hay indicios de simetría bilateral, una característica que pudo estar asociada con la de un hábito reptador y una alimentación llevada a cabo sobre el sustrato, lo que finalmente condujo a la diferenciación, en el cuerpo, de una región anterior o cefálica que facilitaba el movimiento en una determinada dirección. Los platelmintos son seres que presentan sistemas circulatorio y excretor y se desplazan por contracciones musculares y ondulaciones que se propagan de adelante hacia atrás a lo largo de todo el cuerpo, el que así se halla bien adaptado para repatr sobre un fondo sóllido, pero no pueden penetrar enlos sedimentos excepto cuando estos se hallan muy disgregados. Los nemertinos son semejantes a los platelmintos, aunque el cuerpo es engrosado en vez de aplanado. Su denso mesodermo se comprime fácilmente, lo que les permite atravesar grietas estrechas efectuando ondulaciones que reecorren hacia atrás el cuerpo mientras el organismo se desliza hacia delante. Este sistema peristáltico posibilita a los nemertinos introducirse enel sustrato haciendo a un lado los sedimentos confinantes, aunque no solo pueden perforar profundamente debido a la limitada eficiencia hidráulica de los tejidos, la existencia de un sistema circulatorio poco avanzado y la ausencia de un sistema respiratorio. Estas limitaciones desaparecieron con el desarrollo del celoma o cavidad interior llena de fluído, cuyo origen podría haber estado relacionado con la locomoción peristáltica. El celoma facilitó la proliferación de la vida infaunal, la que habría comenzado hace unos 1.000 m.a., según lo sugieren ciertas perforaciones horizontales en sedimentos de dicha edad. Otras trazas no vuelven a encontarrse hasta niveles de 700 m.a. y desde entonces, los metazoarios muestran una continua y creciente diversificación. Por lo general, se admite que los metazoos aparecen recién en la Fauna de Ediacara, hace unos 600 m.a., pero algunos especialistas opinan que este acontecimiento pudo ocurrir antes, hace unos 1.000- 900 m.a., Según Wray et al., (1996) basados en la secuencia de nucleótidas, la divergencia invertebrados/vertebrados habría acontecido hace unos 1.200 m.a., mientras que los deuterostomados (equinodermos) lo hicieron hace unos 1.000 m.a., y los agnatos y gnatóstomos, hace unos 600 m.a. G. Vermeij (1996) opina que un origen de los animales entre 700-900 m.a. (Neoproterozoico Temprano a Medio) parece probable sobre base geoquímicas. Este fue un momento en el que los niveles del oxígeno se elevaron, tanto en los océanos como en la atmósfera, quizá ligado a cambios en el régimen tectónico terrestre y la formación de los continentes. La ausencia de metazoos vendianos podría deberse a su diminuto tamaño, pero fue en ellos donde evolucionó la gran diversidad de planos básicos característicos de los fila de animales vivientes. La elaboración de sistemas circulatorios, respiratorio y alimenticios activos, capacitó a algunos animales véndicos y muchos cámbricos a aumentar de tamaño, crear musculaturas sofisticadas para la alimentación y locomociójn y cubrir el cuerpo con un esqueleto protector. A la finalización del Véndico ocurrió una importante extinción en masa que afectó a la diversidad de determinados grupos, entre ellos, los estromatolitos, los que casi desaparecieron. También declinaron severamente al fitoplancton eucariota s) y la biota de Ediacara, la que desapareció casi totalmente. La transición al Fanerozoico significó un cambio mayor en la composición biótica, quizás provocada por los factores ambientales, como pudo ser el incremento del oxígeno atmosférico o una nueva configuración geográfica motivada por la tectónica de placas que permitió reducir la estacionalidad y aumentar la estabilidad de las fuentes tróficas. Los fila celomados experimentaron una mayor radiación adaptativa al explorar nuevos nichos y, entre los fila novedosos surgidos, algunos tendrían partes esqueletales duras. En la primera parte del Véndico ya se hallan tubos quitinoideos de Sabellidítodos y, posteriormente se presentan otros calcáreos de Claudina. Este último sería el organimso más antiguo conocido con esqueleto mineralizado. Son tubos calcáreos que en namibia, aparecen encima de la biota de Ediacara, pero también se encontraron en Brasil, Argentina y otras partes del mundo. Los Anabarítidos también desarrollaron un esqueleto tubiformes carbonáticos, quizás aragoníticos, pero fueron muy escasos al final del Proterozoico, mientras que mostraron su mayor diversidad al comienzo del Cámbrico. Las trazas fósiles véndicas indicarían que el sustrato marino de estos tiempos alojó una asociación infaunal que habitaba en escasos centímetros debajo de la interfase, en fondos blandos. Las perforaciones verticales fueron escasas. En los últimos años también se han mencionado evidencias que avalarían la posibilidad de existencia de vida terrestre en el Proterozoico. Tal es el caso de los microfósiles filamentosos, bacteriales, hallados en paleosuelos de la Caliza Mescal (Arizona, 1.200 m.a.) y la Dolomita Beck Spring (California, 800 m.a.), mencionados por R.J.Horodyski y L.P. Kout (1994). Las biotas precámbricas sudamericanas El conocimiento de las biotas precámbricas sudamericanas es muy deficiente. En Brasil, el Grupo Bambuí (1.350 - 620 m.a.) es portador de algas (Kinnevia, Bambuites, Asteriacites, Solenoporáceas) y de estromatolitos (Conophyton metula, Linella avis) conocidos en rocas de menos de 850 m.a. de antigüedad. Los microfósiles de estos estromatolitos son formas esferoidales y filamentosas de naturaleza carbonosa. Las esferas tienen entre 3 y 81 micrones y pertenecerían a cianobacterias, pero otros podrían ser eucariotas. En Brasil, el Grupo Jacadigo (que también penetra en Bolivia) contiene a las formaciones ferríferas zonadas de Urucum-Mutum, portadoras de microfósiles similares a lóricas de tintínidos o de quitinozoarios, cuya edad sería superior a los 850 m.a., aunque otras formas similares a Cloudina y Bavlinela indicarían una edad más joven e incluso podrían llegar al Fanerozoico. El Grupo Corumbá, cuya relación con el Grupo Jacadigo no se conoce (y que también se extiende por Bolivia y Paraguay) contiene en el techo algas (Sphaerocongregus variabilis), Cloudina lucianoi y al único metazoo precámbrico conocido de América del Sur, Corumbella werneri, los que seríand e Véndico, mientras que el vendoténido Tyrasotaemia indicaría Cámbrico Inferior. Los Grupos Jacadigo-Corumbá del brasil, Bolivia y Paraguay se correlacionarían con el Grupo Arroyo del Soldado, del Uruguay, el que también incluye formaciones ferríferas zonadas y microfósiles similares. En general, la depositación de las formaciones ferríferas zonadas se relacionan con una severa estratificación de los océanos vinculada a las glaciaciones y, además, se ha postulado otra pequeña glaciación en el límite ProterozoicoCámbrico (550 m.a.) con la que coincidirían las formaciones ferríferas zonaas arriba mencionadas. En la parte superior del Grupo Arroyo del Soldado (Véndico Superior- Cámbrico Inferior), la Formación Cerro Victoria tiene estromatolitos e icnofósiles (Gordia, Planolites, Selloulichnus) Véndico Superior- Cámbrico Inferior. En la base del Grupo (Formaciones Caliza Polanco y Cerro Espuelitas) existen otras microbiotas con Sphaerocongregus variabilis (= Bavlinella faveolata), Oscillatoriopsis rhomboidalis (característica del Véndico de la Plataforma de Europa Oriental), Leiosphaeridia bicrura (del Véndico europeo pero que llegó al Cámbrico Temprano); todas ellas indican una depositación entre 560-545 m.a. El Grupo Sierras Bayas, del sur de la provincia de Buenos Aires (Tandil) proporcionó estromatolitos y microfósiles que serían probables cianofíceas (Paleorivularia ontarica, conocida de la Formación Guntflint de Canadá) y acritarcas (Chuaria, Protoleiosphaeridium,, Leiosphaeridia), que indicarían el Véndico - Cámbrico Inferior. En el noroeste argentino, el Proterozoico Superior proporcionó impresiones medusoides atribuidas a Beltanelliformis y Sekwia (Aceñolaza y Durant, 1985). ACTIVIDADES 1. Caracterice brevemente los acontecimientos principales que tuvieron lugar durante los eones Chaotiano y Hadeano. =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 2. Explique la formación de la atmósfera primitiva. ¿Qué contribución tuvieron las cianobacterias? ¿Qué efecto produjeron? =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 3. Defina los siguientes términos: Snowball Earth – Estromatolitos – Coacervados – Panspermias =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 4. Explique brevemente el origen de la hidrosfera y de la litosfera terrestres. =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 5. Analice los aportes científicos de Arrhenius, Oparin, Haldane, Urey y Miller, Oró y Wächterhauser. ¿Es posible complementarlos? =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 6. Describa y sitúe correctamente en la columna geológica la siguiente escena de la vida precámbrica. =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 7. Explique este espiral del tiempo geológico de acuerdo con lo visto al comienzo del módulo. Compare con el reloj de las eras geológicas. =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 8. Describa brevemente las características de la vida arqueana y de comienzos del Proterozoico. =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== Foro Los biólogos se refieren al conjunto de todos los organismos de todas las clases en un tiempo o lugar particulares como biota (de ese tiempo y lugar). Todas las plantas vivientes en un tiempo o lugar particular constituyen la flora, mientras que todos los animales representan la fauna. Se han descrito cerca de 300.000 especies de organismos fósiles y el número crece incesantemente. Sin embargo, esta cifra es sólo una diminuta fracción de las especies que han existido alguna vez. No sabemos cuántas especies vivieron en el pasado, pero tenemos medios para hacer estimaciones razonables. De la biota presente se han nombrado aproximadamente 1,7 millones de especies. El número real de especies vivientes es probablemente al menos de 10 millones, porque la mayoría de las especies de insectos y ácaros (los grupos de animales con los mayores números de especies) todavía no han sido descritos. Así el número de especies fósiles descritas es menos de 3% del número mínimo probable de especies vivientes. La vida ha existido en la Tierra durante cerca de 3,8 mil millones de años. Las especies duraron, en promedio, menos de 10 millones de años. Por lo tanto, la biota terrestre debe haber cambiado en muchas ocasiones durante la historia geológica. Así, el número total de especies que han existido durante el tiempo evolutivo debe ampliamente exceder el de especies que viven actualmente. 1. ¿Por qué existen tan pocas especies descritas como fósiles? 2. ¿Por qué el registro de fósiels de los tiempos precámbrios es especialmente escaso? 3. ¿En qué testimonios se basan los científicos para determinar la existencia de una biota tan antigua? Marte y la colonizacion terrestre Marte, el planeta rojo, es el cuarto planeta terrestre del Sistema Solar, y muestra en su superficie claros indicios de un paisaje fluvial y aluvional tal como revelan las fotogrfías del Mariner y del Viking, y luego comprobadas por explorador marciano. Esto indicaría que en el pasado geológico de Marte, fue un planeta muy activo, con procesos de erosión y sedimentación, y con gran cantidad de agua líquida. En la actualidad, las diferencias de temperatura entre el día y la noche en la superficie marciana son muy importantes (de -15 a -80 °C) y existe una atmósfera con una presión menor que la terrestre, compuesta principalmente de dióxido de carbono (95%), seguiddo de nitrógeno, argón, trazas de oxígeno, monóxido de carbono y vapor de agua. El vulcanismo en Marte fue extremdamente abundante, al menos, en el 75% del planeta, y la tectónica, fue muy activa en ciertos sitios, tan importante como en Venus o la Tierra, aunque en la actualidad parece haberse detenido. El 27 de diciembre de 1984, un grupo de científicos de la NASA encontraron el oscuro fragmento marciano semienterrado por el hielo del sur, en un lugar denominado Allan Hills. El meteorito marciano se iba a convertir en la principal evidencia acerca de un notable hecho que cambiará, si las sospechas se confirman, la concepción de la vida tal cual se la conoce. El 16 de agosto de 1996, un equipo dirigido por el doctor David McKay, publicó en la revista Science el artículo científico más sorprendente del siglo XX, desbancando incluso al clásico de Watson y Crick de 1953. Lo que venía a decir el ya famoso texto de McKay era que la Ciencia había hallado, por primera vez, evidencias de la existencia de vida extraterrestre, más concretamente, marciana. En efecto, el meteorito ALH84001 contenía pequeñas estructuras (esférulas y bastones) que muy bien podrían ser microfósiles, restos de la existencia de minúsculas formas de vida marcianas. Una pretensión tan sorprendentemente revolucionaria no fue acogida por la comunidad científica con los brazos abiertos, sin antes haber originado altas dosis de polémica. Según una conocida máxima científica, cuanto más difícil de creer sea una afirmación, mayor debe ser el peso de la evidencia necesaria para tomarla en serio. Además, según la regla de Occam, la explicación más sencilla de un hecho suele ser la verdadera. Y la existencia de vida extraterrestre es una de las afirmaciones más increíbles y complejas que fueron enunciadas por un científico del siglo XX. La mayoría de los geólogos opinaba que el origen de las estructuras filamentosas que aparecían en el interior del meteorito podría muy bien tener una explicación mineralógica. El principal problema que surgía con estos microfósiles marcianos era que, en realidad, se trataba de nanofósiles, demasiado pequeños, mucho menores que cualquier bacteria viva de las que pudieran encontrarse en nuestro planeta y, probablemente por debajo del límite inferior de tamaño necesario para albergar la complicada maquinaria que necesita una célula para llevar a cabo sus funciones vitales y puede que, incluso, ser demasiado pequeños como para poder almacenar la información necesaria para su replicación. Las estructuras presentes en la roca marciana tienen una longitud de 20 a 200 nanometros (nm), de cien veces menos que la longitud de una típica bacteria como Escherichia coli (que mide unos 2 µm, o sea, 2000 nm). Las menores bacterias terrestres conocidas en aquel momento (década de 1980) pertenecían al género Mycoplasma, y su célula era una esfera de unos 150 nm de diámetro. Los micoplasmas (regnum Aphragmabacteria), con su dotación genética mínima, son considerados sin duda células vivas independientes, aunque al igual que los virus son parásitos intracelulares obligados y, además, degenerados, que sólo pueden hacerse crecer in vitro si se añaden al medio de cultivo numerosas sustancias orgánicas procedentes de otros organismos a los que normalmente infectan y causan enfermedades, incluido el ser humano. Además de las esférulas y bastones, en el meteorito marciano se encontraron hidrocarburos aromáticos policíclicos (HAP), de indudable origen biológico. A partir de 2.000 se inciaron nuevos estudios con métodos analíticos más precisos para estudiar las eventuales marcas de la actividad biológica en el planeta Marte. Se calcula que las conclusiones finales sobre la vida marciana habrían de llegar entre 2.012 y 2.015, si bien ya en 2007 un grupo de científicos argentinos, sobre la base de los cálculos de la NASA, habría demostrado la existencia de organismos marcianos. En este caso, cabría especular si la biota marciana (Marciania) se halla conectada con la terrestre (Terrestria), y de qué manera se estableció dicha conexión a través del modelo de las panspermias. 1. ¿Por qué esta presencia de microorganismo marcianos no es aceptada en todos los círculos de investigadores? ¿Les parece suficiente la evidencia? 2. ¿Creen que existe vida fuera del planeta Tierra? ¿Por qué? 3. Imaginen de qué manera podría ser concebida la evolución de la vida en otro planeta? 4. La exobiología es el estudio de la vida fuera del planeta Tierra. ¿Suponen que existen fundamentos para el desarrollo de esta disciplina? Módulo 4. La Era Paleozoica Objetivos • Evaluar las características de la era Paleozoica en general • Analizar la vida en el Cámbrico y justificar la explosión o big bang en ese período • Examinar la evolución de la vida marina y continental en el Ordovícico • Correlacionar los cambios paleogeográficos con la evolución de la vida silúrica • Analizar las características de la vida continental durante el Devónico • Examinar el desarrollo de la flora y de la fauna durante el Carbonífero • Explicar las características de la conquista del medio aeroterrestre • Analizar la evolución del ambiente durante el Pérmico • Analizar las primeras grandes extinciones y el impacto de la tardío-ordovícica, tardíodevónica y tardío-pérmica Contenidos La era Paleozoica y su división La era Paleozoica temprana La vida cámbrica La vida ordovícica El Silúrico La conquista del medio aeroterrestre El Devónico El Carbonífero El Pérmico Las grandes extinciones de la historia terrestre División de la era Paleozoica Con la era Paleozoica se inicia el eón Fanerozoico, hace 543 m.a., caracterizado por la presencia de los primeros fósiles de caparazones y esqueletos, de fosfatos, carbonatos y sílice amorfo. La primera era de este eón es justamente la era Paleozoica y su primer período, donde se inicia la revolución de la vida animal acuática, es el período Cámbrico. Claro que se discute realmente si hubo una transición gradual o un cambio explosivo en la fauna y flora de la época, y lo más probable es que se combinaran ambos factores en distinta proporción. Así, si hubo previamente una presencia de animales sin esqueletos ni caparazones, como los cnidarios y poríferos además de varios grupos fósiles ediacarianos, es ahora en el Cámbrico cuando se inicia una verdadera revolución de estas estructuras duras y, por lo tanto, el registro fósil se hace abundante. Era Paleozoica temprana El Eratema Paleozoico se inició con la depositación del Sistema Cámbrico. Durante el tiempo comprendido por este acontecimiento, se acentuó la dispersión de las placas continentales originadas por la disgregación de Rodinia, las que tendieron a ubicarse en las bajas latitudes. Este desplazamiento fue acompañado por la invasión marina de las plataformas y una intensa sedimentación en las áreas geosinclinales. Sobre las primeras se formaron depósitos carbonáticos y terrígenos que alojaron a numerosas comunidades bentónicas. Estas plataformas carbonáticas alcanzaron su máximo desarrollo al final del Cámbrico y principios del Ordovícico. Desde hace 540 millones ya no existía en la Tierra un supercontinente, sino varios continentes separados, rodeados de inmensos mares de plataforma, lo que determinó que la vida se hiciera extremadamente abundante en el mar, aunque en la tierra firme continuaba habitado únicamente por bacterias, en particular cianobacterias debido a la radiación (y la ausencia de una capa protectora de ozono que solo comenzó a formarse en el Ordovícico). La placa continental mayor de este tiempo era Nigritia, además de otras siete menores, ubicadas principalmente al norte del ecuador, Laurentia, Báltica, Siberiana, Kazhakstana, Centrochina, Sinocoreana e Indochina (estas tres últimas se fusionarían más tarde para formar China). Un nuevo océano, antecedente del Atlántico, el Iapetus, se había formado entre Laurentia y Nigritia. El continente mayor que aparece debajo del ecuador y con parte inmersa en el Paleopolo Sur es Nigritia (a veces también conocido como Gondwana, aunque este nombre induce a error y es preferible reservarlo para el continente que se habría de formar a partir de finales del Triásico, ya en la era Mesozoica). La vida se expandió rápidamente en el Cámbrico El gran desarrollo de las plataformas submarinas favoreció la proliferación y diversidad de la vida en el Cámbrico. Predominan las formas bentónicas sobre las planctónicas (radiolarios, acritarcas, algunos trilobites) y el 70% de la fauna de todo el período se compuso de trilobites . En efecto, los trilobites fueron los fósiles guía del Cámbrico, no cabe duda que fueron los trilobites, el grupo más importante, aunque al final del períopdo experimentaron una drástrica reducción, si bien continuaríann floreciendo hasta finales del Pérmico. Las concentraciones de O2 de la atmósfera del Cámbrico se estaba aproximando al nivel actual. En los mares de plataforma de los nuevos continentes se evidenció una rápida diversificación de la vida, lo que ahora se conoce como la explosión Cámbrica (aunque de hecho la evolución creciente comenzó antes del Cámbrico). La mayoría de los grupos de animales que tienen especies vivientes actualmente aparecieron durante el Tommotiano, el primer piso del Cámbrico inferior, cuando se produjo una radiación epibentónica con aparición de formas esqueletales que fueron reemplazando a las trazas dominantes y la la vida infaunal (sumergida en el fondo) que predominaba hasta entonces. En este momemnto aparecieron todos los grupos de animales así como también muchos grupos fósiles completamente extintos (por ejemplo, los cambroclávidos y los mitrosagóforos). Igualmente surgieron diversos grupos de protistas, en particular dos que serían muy importantes en la historia terrestre, los foraminíferos y los radiolarios. En su mayor parte, los fósiles representan sólo las partes duras de los organismos, aunque en los lechos fosilíferos del Cámbrico – como Burgess Shale, en la Columbia Británica, Sirius Passet en el norte de Groenlandia y Chengjiang en el sur de China–las partes blandas de numerosas especies de animales se han preservado también. Reconstrucción de la fauna de Chengjiang, en las calizas Maotianshan de los mares de plataforma chinos del Cámbrio inferior. En el fondo marino son abundantes onicóforos y priapúlidos, además de trilobites y braquiópodos, y en el cuerpo de agua aparecen radiodontos y craneados (como los milokummíngidos). Pero fueron los artrópodos (cangrejos, langostinos y sus parientes) el grupo más diverso en la fauna china. Esqueletos y caparazones, clave de la explosión cámbrica Los primeros esqueletos y caparazones debieron servir para aumentar la eficiencia de la locomoción del epibentos, donde no existían los efectosconfinantes de los sedimentos (en las formas infaunales). Los esqueletos rígidos para estos fines resultarían más ventajosos que los mecanismos hidrostáticos utilizados por la biota infaunal predecesora. También pudieron representar una reacción defensiva contra el ataque de formas depredadoras como los radiodontos (fósiles). En el Cámbrico temprano existieron grupos con parientes vivos y otros con afinidades sistemáticas conocidas junto a otros de afinidades desconocidas y que no superaron el Paleozoico Inferior. Entre los primeros se hallan representados los -Espongiarios o pofíeros: espículas hexactinélidas y de esponjas córneas o demosponjas -Cnidarios: perforaciones de actinias, hidrozoos (Scenella) y tetracorales (en Australia del Sur) -Braquiópodos: inarticulados (a partir del Tommotiano) y articulados (a partir del Atdabaniano) -Priapúlidos -Anélidos poliquetos -Moluscos: monoplacóforos (galeroconchos), arqueogastrópodos, rostroconchos (solo el primer grupo sobrevive) -Artrópodos: onicóforos (como Xenusium), merostomados (como Palaeomerus), ostrácodos primitivos y trilobites (desde el Atdabaniano) correspondientes a los agnóstidos, corynexóchidos, odontopléuridos, ptychopáridos, redlíchidos. -Quetognatos, como Protoherzina -Equinodermos: lepidocistoideos, helicoplacoideos, edrioasteroideos y camptostromatoideos (todos fósiles). -Hemicordados: Yunnanozoon de China, y otros géneros similares, habrían sido hemicordados primitivos. Los filos de afinidades desconocidas incluyen los arqueociátidos (a partir del Tommotiano), hiolitos y numerosos tubiformes, como los anabarítidos, hiolitelmintos, mitrosagóforos, etc. Los arqueociátidos fueron formadores de arrecifes e importantes fósiles guía del Cámbrico. La diversidad de la biota se acentuó en el Cámbrico mMedio, como lo revela el yacimiento excepcional de las Lutitas Burgess (Formación Stephen) del Cámbrico mMedio de la Columbia Británica, Canadá. Estos sedimentos de grano muy fino conservaron gran cantidad de impresiones de organismos marinos en las que se pueden apreciar detalles de las partes blandas más delicadas. Se conocen más cien géneros de esta fauna, la mayoría perteneciente a clases extinguidas, correspondiendo la representación principal a artrópodos (25% excluidos los trilobites, 10%) y anélidos (11%), priapúlidos (6%), poliquetos (5%) y otros grupos minoritarios. Muchas especies tienen gran importancia para la filogenia de los artrópodos. También en esta biota se halla el crinoideo más antiguo conocido y los únicos onicóforos seguramente identificados hasta el presente, como Aysheaia con solo diez segmentos en el cuerpo (en vez de 15-45 como los vivientes) y marinos. Los crustáceos contaron con ostrácodos y conchóstracos. La forma de la vida de la ma yoría de estos organismos es difícil de deducir, aunque habrían existido formas bentónicas, planctónicas y nectónicas con diferentes hábitos alimenticios, incluso depredadores como Esmeraldella. En general, esta asociación se habría aproximado mucho a la normal de esos tiempos. En la fauna del Cámbrico medio también existieron foraminíferos arenáceos, graptolites dendroideos, probablemente moluscos aplacóforos (Wiwaxia) y equinodermos primitivos. En el Cámbrico tardío aparecieron las conchillas asimétricas de los gastrópodos, preferentemente pleurotomáridos (llegan hasta la actualidad) y maclurítidos (se extinguen en el Devónico). Al final del Cámbrico surgieron los cefalópodos nautiloideos, con formas muy pequeñas de ellesmocerátidos, y los quitones (Palaeoloricata) que eran muy similares a los actuales. Cordados, craneados y vertebrados en el Cámbrico La fauna cámbrica contiene a los más antiguos representantes del phylum Chordata y también a ciertas formas que podrían ser asignadas a los Craneados o directamente los vertebrados. Los Cordados se caracterizan por tener: una notocorda o tubo viscoso flexible de posición dorsal que corre a todo lo largo del cuerpo, recubierto de colágeno, un cordón nervioso dorsal (sobre la notocorda) expandido anteriormente, faringe (fisuras o hendiduras branquiales), endostilo (equivalente a la glándula tiroidea de los vertebrados) y una verdadera cola (región simétrica extendida detrás de la cavidad visceral y técnicamente situada detrás del ano). Entre las formas fósiles cámbricas de Chordata se citan: - Palaeobotrylus, del Cámbrico inferior de Nevada, con aspecto de tunicado. - Pikaia, del Cámbrico medio de las Lutitas Burgess de Canadá, con aspecto de cefalocordado conservado como una película de aluminosilicato, que tiene 10 miótomos (bloques de músculos en forma de V). Se asemeja mucho al Amphioxus. - Anatolepis, del Cámbrico tardío de Wyoming. Son fragmentos de placas óseas, con superficie con nodosidades, que podrían ser algún tipo de escama. Dado que estas placas óseas contienen dentina (exclusiva de los Vertebrados), ahora se lo considera un Vertebrado. - Haikouella lanceolata, del Cámbrico inferior de China (530 m.a.). Muy similar al Amphioxus, el cefalocordado actual más abundante. Tendría corazón, aorta ventral y dorsal, arteria branquial anterior,filamentos branquiales, una proyección caudal, un cordón neural con un cerebro relativamente grande, posibles ojos laterales y una cavidad bucal situada ventralmente con tentáculos cortos, por lo que algunos autores suponen que sería un craneado del inicio del Cámbrico (JunYuan et al., 1999, Nature 402: 518-522). Los craneados se caracterizan por la presencia de un cráneo que protege un encéfalo (órgano complejo formado por el cerebro y otras estructuras). En el yacimiento de Chengjiang se hallaron otras dos formas de craneados, los milokummíngidos. - Myllokummingia, fusiforme, de unos 28 mm de largo, con región cefálica y tronco, 25 miómeros (en W), pliegues en forma de aletas, canal alimenticio con faringe, intestino, notocorda y probablemente, cavidad pericárdica. Carecería de esqueleto dermal (escamas). - Haikouichthys, muy parecido al anterior pero más evolucionado. Con región cefálica y tronco, aleta dorsal prominente con radios, pliegue-aleta ventro-lateral. Ambos géneros se hallan con trilobites y los artrópodos más cercanos a crustáceos, Naraoia y Waptia y se relacionarían más con los mixinos o anguilas glutinosas. Muchos paleontólogos sostienen que los vertebrados divergieron de una forma similar al Amphioxus, hace unos 545 m.a., aunque los estudios moleculares basados en las similitudes de los genes, indicarían que los Vertebrados se iniciaron hace 750 m.a. (Proterozoico tardío). Ello estaría avalado por estudios neuroanatómicos. El elenco de organismos se completaría con los primeros Vertebrados, representados por los ostracodermos, peces acorazados sin mandíbulas que hicieron su tímida aparición el Cámbrico superior. Los vertebrados se caracterizan además del cráneo por la presencia de vértebras. Mientras tanto, otro grupo relacionado con los vertebrados, aunque nno incluidos entre ellos, los conodontos, representarían también el primer paso en la aparición del esqueleto mineralizado pues, contrariamente a lo pensado, la mineralización no comenzó en la piel sino en la boca de los conodontos, lo que los convirtió en feroces depredadores. Sus elementos dentiformes son de fosfato de calcio y serían los más primitivos indicios del hueso, aunque también se menciona que Yunnanozoon y Haikouella (del Cámbrico inferior) tendrían dientes en la faringe por lo que, de confirmarse, representarían la más primitiva biomineralización en los cordados. Los conodontes vivieron desde el Cámbrico tardío hasta el Triásico y su verdadera naturaleza es conocida recién a partir de 1993, cuando se halló un animal completo (Clydagnathus) en el Carbonífero inferior de Edimburgo. Este ejemplar tiene unos 40 mm de largo (pero en el Ordovícico de Sud Africa se hallaron formas gigantes), forma de anguila, simetría bilateral, cabeza, ojos, notocorda, miótomos (segmentos musculares que por contracciones facilitaban un poderoso movimiento natatorio), cordón nervioso y aleta caudal con soportes radiales. Los elementos dentiformes, llamados conodontes, de naturaleza fosfática, yacían detrás de la región cefálica y servían para la alimentación, seccionando y triturando los alimentos, y estaban dirigidos hacia atrás posiblemente para ayudar a mantener el alimento en la boca, evitando su escape. Durante el lapso Cámbrico medio-Devónico medio existieron también equinodermos (del grupo de los Cornuta) asimétricos, como Ceratocystis, con esqueleto externo de carbonato de calcio, los que según Jefferies serían cordados, por lo que los agrupa en la denominación informal de Calcichordata, (calcicordados) pero la mayoría de los autores lo niega. En 1920, Walter Garstang notó las semejanzas entre la larva de un tunicado, un Amphioxus adulto y los Vertebrados. La cola del tunicado sería un apéndice transitorio que evolucionó como un crecimiento externo desde el cuerpo para asegurar la amplia dispersión de la larva antes de su fijación, y llamó “pedomorfosis” al nexo evolutivo entre los tunicados y los cordados superiores o sea, el desarrollo completo de gónadas y habilidades reproductivas en un estadio esencialmente juvenil. Hace unos 540 m.a. o más, una larva de tunicado aparentemente fracasó en metamorfosearse y se convirtió en un individuo adulto, reproductivamente maduro. Este juvenil retuvo su cola, notocorda y cordón nervioso, y perdió el estado sedentario del tunicado adulto. Esta teoría tiene poca evidencia directa pero se la acepta por la frecuencia de la pedomorfosis en la evolución de los Vertebrados. Diversos grupos de organismos se diversificaron en el resto de la era Paleozoica Los geólogos dividen el resto de la era Paleozoica en los períodos Ordovícico, Silúrico, Devónico, Carbonífero (a veces dividido en Missisìpico y Pennsilvánico) y Pérmico. Cada período se halla caracterizado por la diversificación de grupos específicos de organismos. Las extinciones en masa marcaron los finales del Ordovícico, Devónico y Pérmico. El Ordovícico y sus organismos (488-444 m.a.) Durante el período Ordovícico, los continentes, que se hallaban localizados principalmente en el Hemisferio Sur, aún carecían de plantas multicelulares. La radiación evolutiva de los organismos marinos fue espectacular durante el Ordovícico temprano, especialmente entre animales tales como los braquiópodos y los moluscos, que vivieron en el lecho marino y filtraban pequeñas presas del agua. Al final del Ordovícico, glaciares gigantescos se formaron sobre Nigritia, los niveles del mar descendieron cerca de 50 metros y la temperatura de los océanos descendió. Cerca del 75% de las especies animales se extinguieron, probablemente debido a estos grandes cambios ambientales. El zooplancton del Ordovícico estuvo integrado por radiolarios y graptolitoideos. El bentos marino experimentó una gran radiación con la participación de poríferos, conuláridos, corales, briozoarios, moluscos, equinodermos y artrópodos. Un hecho destacable fue la proliferación de graptolites dendroideos en el piso Tremadociano del Ordovícico, y de sus descendientes graptoloideos en el Arenigiano. En el Ordovício temprano persistieron las ricas comunidades de trilobites, pero paulatinamente fueron siendo reemplazadas por las de braquiópodos, y, como consecuencia, los trilobites se desplazaron a ambientes bentónicos profundos y pelágicos. Al final del período desaparecieron los agnóstidos y odontopléuridos. Otra explosión evolutiva acompañada de una amplia radiación ocurrió entre los nautiloideos al iniciarse el Ordovícico, abundando los ortocerátidos, los endocerátidos y otros órdenes con conchillas de diversas formas. Los bactrítidos, aparecidos en estos tiempos, fueron intermediarios entre los nautiloideos y los ammonoideos. A partir del Tremadociano fueron frecuentes los bivalvos epifaunales. Los gastrópodos siguieron mostrando características primitivas y los braquiópodos inarticulados alcanzaron su máxima diversificación, mientras que los articulados se diversificaron particularmente al final del período. Los mares ordovícicos alojaron también a los primeros arrecifes fanerozoicos de importancia, integrados por briozoarios trepostomátidos y criptostomátidos, corales tetracorálidos y tabulados y esponjas lithístidas. En el Ordovícico aparecieron los artrópodos xifosúridos y euriptéridos y los equinoideos, ofiuroideos y asteroideos; abundaron los hiolítidos, tentaculítidos y ostrácodos. En el Ordovícico, la diversidad biótica se incrementó notablemente y junto con el Cámbrico, ambos períodos produjeron la totalidad de los grupos actuales de animales, y la mayoría de las clases y órdenes. En los ambientes de plataforma de los mares ordovícicos, la disponibilidad de oxígeno habría variado según los lugares, influyendo en el desarrollo de las comunidades de los fondos. En los ambientes oxigenados la vida bentónica estuvo dominada por los organismos de esqueletos inorgánicos (braquiópodos, moluscos, trilobites, etc.) y a esta asociación se la conoce como “Facies Shelly”. Las “Facies Graptolíticas” se componen casi exclusivamente de graptolitos contenidos en lutitas negras. Estas rocas corresponden a ambientes euxínicos originados en los fondos con circulación muy escasa o nula. El poco oxígeno allí existente era consumido rápidamente sin llegar a ser repuesto. Así, la vida bentónica allí era nula y la única materia orgánica acumulada en esos fondos provenía del plancton que, como los graptolitos, habitaba los estratos superiores de la masa de agua. Los primeros vertebrados, Anatolepis, los arandáspidos y los astráspidos (todos ostracodermos) eran peces pequeños, de unos 10 cm de longitud, con una cola móvil cubierta por placas pequeñas, puntudas, y un escudo cefálico óseo compuesto de varias placas grandes que cubrían la cabeza y parte delantera del cuerpo o hasta la región anal en algunas formas primitivas. Arandaspis (Ordovícico medio de Australia) tenía un escudo dorsal y otro ventral; entre ellos se disponía una hilera de aberturas branquiales; además habría tenido una “línea lateral” de poros que alojabanterminaciones nerviosas para detectar movimientos del agua producidos por posibles depredadores.La superficie de las placas lleva pequeños tubérculos rómbicos Los principales grupos de algas cámbricas continuaron en el Ordovícico, agregándose las codiáceas, receptaculitos y otros grupos. El Ordovícico finalizó con una de las crisis más importantes del Fanerozoico. Ella ocurrió durante el piso Ashgilliano y la diversidad se redujo a un 20%. Los grupos más afectados fueron los corales, braquiópodos, trilobites, ostrácodos, equinodermos, graptolites, conodontes, quitinozoarios y acritarcas. La crisis afectó tanto al plancton como al bentos y se produjo en dos fases: la primera coincidió con una regresión motivada por la iniciación de la glaciación y el consiguiente descenso del mar. La temperatura disminuyó y las aguas frías se extendieron hasta los trópicos, lo que permitió el desarrollo de la Fauna de Hirnantia (braquiópodo) de distribución mundial, incluyendo el norte africano y Argentina. Esta Fauna de Hirnantia caracteriza al Piso Hirnantiano y el final del mismo. La segunda etapa en la crisis se debió a que, posiblemente, una vez terminada la glaciación, la fusión de los hielos originó una transgresión acompañada por un aumento en la temperatura, que ocasionó la desaparición de la Fauna de Hirnantia y muchos otros organismos. A pesar de lo profundo de esta crisis biológica, parte de los organismos ashgillianos llegaron a ser dominantes en las primeras faunas silúricas, cuyas asociaciones típicas recién aparecen en el Llandovery Tardío. Vida ordovícica. Destacan los crinoideos (equinodermos) y los gigantescos nautiloideos ortocerátidos. El Silúrico y sus organismos (444-416 m.a.) Durante el período Silúrico, los continentes sepentrionales se unieron, pero las posiciones generales de dichos continentes no cambiaron demasiado. Debido al acercamiento o la colisión entre grandes placas, muchos geosinclinales continuaron estrechándose y algunos se cerraron total o parcialmente al final del período, formándose elevaciones montañosas y ambientes continentales con abundantes estratos rojos y evaporitas. Las transgresiones más importantes ocurrieron al inicio y final del período, mientras que una amplia regresión caracterizó su parte media. A partir del Wenlock se incrementaron las transformaciones paleogeográficas y la colisión de Laurentia con Báltica originó al Supercontinente Laurusia. Avalonia se hallaba casi adosada a Laurentia, quedando el Iapetus muy reducido y suprimido en su parte norte, por lo que allí sus faunas marinas fueron reemplazadas, a partir del Downton, por el ambiente de las “Viejas Areniscas Rojas” (“Old Red Sandstones”), distribuídas por el norte de los Apalaches, Groenlandia, Gran Bretaña y Noruega. Tanto en la planicie central como la parte occidental de América del Norte se originaron depósitos dolomíticos, principalmente en el Silúrico Medio, y la región de los Grandes Lagos alojó extensos arrecifes coralinos. Entre Gondwana y Laurusia, se extendía un amplio océano, Rheico, en proceso de estrechamiento por la convergencia de ambos supercontinentes. Gran parte de Gondwana se ubicaba en el área polar sur y sólo el norte africano, Arabia, región himalayana y Australia gozaban de un clima cálido a templado. También en Laurentia, Siberia (Angara), Báltica y Avalonia las aguas eran cálidas y en ellas se depositaron carbonatos. Nigritia continuó internándose en la región polar sur y se formaron glaciares en varias regiones del mismo durante el Silúrico Temprano. Así, sus aguas adyacentes comenzaron a enfriarse, mientras en otras regiones continuaban predominando aguas cálidas o templadas. El Silúrico también se caracterizó por su brevedad, ya que habría durado apenas unos 25 m.a., lo que se debió a las rápidas transformaciones geográficas y biológicas ocurridas en dicho lapso. La vida marina se recuperó de las extinciones masivas de fines del Ordovícico. Por primera vez hicieron su aparición los animales que fueron capaces de nadar y alimentarse sobre el fondo oceánico, aunque ningún grupo nuevo de organismos marinos evolucionó en este período. El mar tropical no estaba interrumpido por barreras y la mayoría de los organismos se hallaban distribuidos ampliamente. Los peces experimentaron una rápida evolución, apareciendo las formas mandibuladas y comenzó la invasión de los ambientes continentales a través de algunos grupos de invertebrados y peces, lo que habría sido favorecido por el desarrollo de las primeras plantas vasculares. Los graptolitos y conodontos son los organismos más utilizados en la biozonación y como fósiles guía del Sistema Silúrico. Sobre la tierra firme, las primeras plantas vasculares hicieron su aparición en el período Silúrico tardío (hace cerca de 420 m.a.). Estas plantas fueron de menos de 50 cm de altura y carecían de raíces y hojas. Los primeros artrópodos terrestres– escorpiones y milpiés–aparecieron casi al mismo tiempo. Existió una moderada diversidad biótica e importantes innovaciones evolutivas. Los graptolitos diplográtidos no superaron el Llandovery y fueron reemplazados por los monográptidos. Los braquiópodos con braquidio espiralado completan su elenco con el agregado de los rostrospiroideos, espiriféridos y punctospiroideos y al final del período los terebratuloideos desarrollaron el braquidio en bandeleta. Por otra parte, los braquiópodos inarticulados declinaron. Los bivalvos incrementaron su presencia pero continuaron siendo primitivos; entre los gastrópodos, surgieron los Patellaceae y los nautiloideos continuaron diversificados. Los artrópodos contaron con los primeros picnogónidos y cirripedios; los euriptéridos estuvieron muy difundidos por América del Norte y Europa; abundaron los ostrácodos y decrecieron los trilobites en representatividad. Los arrecifes aumentaron su distribución, participando los coralestabulados y rugosos, estromatoporoideos, briozoarios, espongiarios litístidos y algas. Las biofacies “shelly” y graptolítica continuaron bien diferenciadas Contiene excepcionales preservaciones de partes blandas principalmente de artrópodos, seguidos en importancia por vermes, anélidos, graptolites y conuláridos, pero las formas esqueletales son raras o faltan. Entre los artrópodos hay trilobites, crustáceos, quelicerados y un miriápodo; los crustáceos incluyen a los filocáridos, a los ostrácodos y al más antiguo de los representantes de la enigmática clase de los tilacocéfalos (extinguidas en el Cretácico Superior). También hay una especie de Protoscolex o Palaeoescolex (Cámbrico SuperiorSilúrico Superior), cestode primitivo de los platelmintos; se encuentran también formas que recuerdan a las sanguijuelas, mientras los poliquetos están representados exclusivamente por escolecodontes. En el Silúrico, los invertebrados comenzaron a invadir el ambiente continental a partir de la aparición de grupos como los miriápodos y los escorpiones. Los miriápodos (centípedos más millipodos) habrían sido originalmente acuáticos y se los encuentra en rocas del Llandovery-Wenlock. Los vertebrados silúricos estuvieron representados por peces (incluyendo a los conodontes) los que fueron exclusivamente marinos hasta el Wenlock, pues luego comenzaron a invadir los ambientes dulciacuícolas o salobres. Carecieron de esqueleto interior pero tenían placas mineralizadas que cubrían todo o parte de su cuerpo, formando como un escudo, que habría servido de protección, por lo que se los denomina “ostracodermos”. Esta presencia de un escudo óseo ya se observa en los peces ordovícicos. Además, todos estos peces primitivos carecían de mandíbulas y sólo tenían una fisura o abertura cerca de la parte frontal. En el Silúrico, los peces evolucionaron rápidamente, apareciendo formas mandibuladas o Placodermos, los más antiguos serían los acantodios que aparecieron en el Silúrico Temprano. Los peces osteictios también habrían estado presente, pero son poco frecuentes. Fueron los primeros peces mandibulados y tuvieron una rápida evolución durante el Silúrico tardío y Devónico temprano. Los más antiguos constituyen la Clase Acanthodii (“tiburones espinosos”)1 que aparecen en el Silúrico Temprano pero que fueron abundantes sólo en el Devónico, llegando hasta el Pérmico Temprano. Pequeños (menos de 20 cm de longitud). Cuerpo alargado, cabeza grande cubierta con placas óseas, ojos grandes rodeados por placas escleróticas; algunos poseían dientes. Aletas pectorales y pélvicas modificadas como largas espinas, pudiendo existir hasta 6 pares de espinas a lo largo del vientre en las formas primitivas. Una o dos aletas dorsales, una aleta anal y cola heterocerca. Otras aletas sostenidas por una espina sobre el borde interno. Nadaban con la cola y el cuerpo. Según la teoría más aceptada, pero actualmente puesta en dudas, la mandíbula se habría originado por modificación de los arcos branquiales anteriores. Esta teoría se basa en la anatomía de los Vertebrados vivientes y particularmente, en aspectos de la embriología del desarrollo de la boca y estructuras faringeales, pero no hay fósiles que muestren tal transición, y las evidencias anatómicas mostrarían que la realidad no habría sido tan sencilla. Los actinopteriogios aparecieron en el Silúrico Tardío pero son poco conocidos antes del Devónico Tardío. Las plantas primitivas mejor conocidas son del Silúrico medio de Laurentia, pero también se conocen de Australasia, Asia oriental y central, Africa y América del Sur. Parecería que las plantas emigraron a la tierra en muchos lugares diferentes, alrededor de las costas del Océano Rheico, en las latitudes medias y altas. Ya en el Silúrico tardío existían muchas algas multicelulares con aspecto de plantas, como Parka y Prototaxites, cohabitando ambientes pantanosos junto a plantas vasculares más primitivas. Sin embargo, tales formas algales no tuvieron éxito en colonizar la tierra y se extinguieron al final del Devónico al no poder competir con las plantas vasculares mejor adaptadas. Ejemplo de una planta terrestre primitiva es Cooksonia del Silúrico y Devónico Inferior. Los ejemplos más antiguos son del Silúrico medio (425 m.a.) de Irlanda, pero también se la conoce en Gales, Bolivia, Checoslovaquia, Libia y Kazakhstan, alrededor del Rheico. Era de pocos milímetros de altura, tallo cilíndrico liso y bifurcado. En la vida silúrica marina destacaron los euriptéridos o escorpiones marinos gigantes, además de hacerse abundantes los erizos y estrellas de mar. Los primeros cursos de agua dulce con fauna adaptada a este ambiente y las primera splantas vasculares como Cooksonia, junto con hongos de sombrero (basidiomicetos), escorpiones y colémbolos, comenzaron a proliferar en el Silúrico. El Devónico y sus organismos (416-359 a.C.) Durante el Devónico, las áreas continentales se ampliaron y muchas cuencas marinas desaparecieron debido al encuentro entre placas, y en su reemplazo se constituyeron importantes elevaciones montañosas. Por primera vez se reconocen depósitos continentales cubriendo grandes extensiones de la superficie terrestre, los que contienen muchos fósiles animales y vegetales. En este período aparecieron los primeros cefalópodos ammonoideos, los goniatítidos y se produjo una gran diversificación de los peces, un grupo de los cuales, al terminar el período, originaría a los anfibios. En los ambientes continentales se difundieron las plantas vasculares incluidas las primeras gimnospermas, y aparecieron los primeros bivalvos de agua dulce. Al final del período se produjo una crisis evolutiva que originó una gran extinción. La fauna devónica de invertebrados fue muy variada, con importantes innovaciones evolutivas, pero al final del período desaparecieron muchos organismos hasta entonces característicos. Las Viejas Areniscas Rojas constituyen un excelente reservorio paleontológico de seres primitivos que habitaron el medio terrestre durante el Devónico Temprano. Un yacimiento muy conocido es el de Rhynie (Pragiano) de Escocia, que contiene miriápodos, arácnidos, crustáceos branquiópodos (Lepidocaris), ciertas formas de acáridos y Rhyniella praecursor, un collémbolo. La mayoría de los artrópodos son de muy pequeño tamaño. Los peces agnatos (ostracodermos) continuaron adaptados a diversas formas de vida y nichos ecológicos pero finalmente, quizás por la competencia de los peces mandíbulados, se extinguieron. Junto con diversos ostracodermos, en el Devónico predominaron los placodermos, cuando estuvieron representados por varios grupos, entre ellos los acanthodios como el gigantesco Dinichthys de 15 m de largo. En los placodermos, las aletas pectorales estaban bien desarrolladas pero las pélvicas eran reducidas; tenían una aleta dorsal y otra anal. La fauna de peces devónicos se completó con la presencia de los condrictios (= seláceos = peces cartilaginosos) y osteictios (= peces óseos), ambos originados a partir de placodermos, aunque se ignora cuál de ellos es más primitivo. Los osteictios se caracterizaron por su osificación interna y externa. Poseen un opérculo con la parte posterior libre para permitir la salida del agua procedente de las branquias. Todas las aletas tienen soportes espinosos interiores. En los depósitos de agua dulce del Devónico Medio se hallan sus representantes más primitivos, los paleoniscoideos, de dimensiones modestas, ojos grandes, boca alargada, cola heterocerca y “pulmones” funcionales en los momentos de sequía. Un ejemplo típico fue Cheirolepis del Devónico Medio de Escocia. Tenía unos 0,25 m de largo, ojos grandes, dientes filosos, escamas rómbicas articuladas, dispuestas en diagonal hacia atrás y abajo, compuestas de capas de hueso y dentina, cubiertas por una especie de esmalte. Cola muy heterocerca, aletas dorsal y anal grandes y triangulares, pectorales y pélvicas pares. Habrían sido rápidos nadadores y depredadores. El otro grupo que comenzó a diversificarse fueron los sarcopterigios se caracterizaban por sus aletas lobuladas carnosas soportadas por un único hueso basal y con músculos que podían modificar la postura de las mismas; las coanas (aberturas nasales internas) indicarían una respiración pulmonar. Aunque tenían algunas similitudes con los actinopterigios, las aletas pares se hallaban ubicadas en otras posiciones (aletas pectorales justo detrás de la cabeza; las pélvicas en la parte trasera del cuerpo), y poseían dos aletas dorsales (los actinopterigios tienen sólo una). Adicionalmente, llevaban una abertura pineal y escamas gruesas cosmoideas. En el Devónico, los sarcopterigios fueron más importantes que los actinopterigios pero luego se hicieron más raros. Durante el Devónico Temprano las plantas podían crecer bien hacia arriba pero sólo hasta cierto límite más allá del cual, debido a su propio peso o por el viento, colapsaban. Este problema fue superado con la producción de tallos leñosos, originados por la actividad de un meristema secundario (o cambium vascular) que rodeaba a los haces del xilema primario. Este tejido secundario, que aumenta el espesor del tallo, consiste de células fortalecidas con lignina y al permitir crecer más ancho lo puede hacer también más alto, mejorando la exposición a la luz. Se conocen restos de leño secundario desde el Devónico Medio y Superior. Estas plantas tenían xilema secundario (madera), floema o sistema vascular primario, hojas grandes y chatas, raíces verdaderas, mega y microesporas. Todo esto facilitó la aparición de las grandes plantas arbóreas en el piso Frasniano, como las Progimnospermas que habrían sido las antecesoras de las plantas con semillas. En las plantas primitivas del Silúrico, la fertilización debía realizarse necesariamente en un ambiente húmedo, pues el gametofito no estaba protegido y la gameta masculina debía nadar hasta las células femeninas en el arquegonio para que se produjera la fertilización. Este problema fue superado con la aparición de la semilla. En las plantas con semillas, el esporangio “masculino” (saco polínico) produce mucho polen pequeño que parece las esporas de las plantas primitivas, pero el esporangio “femenino” sólo contiene a una sola espora (megaspora), la cual es fertilizada mientras está aun en el esporangio unido a la planta. Hasta el momento de la fertilización, el esporangio y la espora constituyen al óvulo, pero después de la fertilización se llama semilla. Winfrenatia, de Rhynie, es la evidencia más antigua y real de líquenes; entre los hongos se hallaron quítridos, zigomicetes y ascomicetes. Durante el Silúrico y Devónico hubo una profunda transformación de la superficie terrestre, pues las plantas se adaptaron rápidamente a las condiciones más secas. No sólo la invadieron sino que también la alteraron. La erosión se fue reduciendo a medida que las plantas cubrían la tierra y originaban suelos. La masiva pérdida de vapor de agua por transpiración y evaporación alteró los tipos de lluvias. Particularmente, cuando las plantas llegaron a formar bosques se transformaron en un reservorio importante para el carbono, ayudando a reducir los niveles de CO2 en la atmósfera, por lo que declinó el “efecto invernadero” y el clima global se enfrió. La cubierta vegetal absorbió más calor desde la atmósfera que las rocas y suelos descubiertos, lo que también ayudó a disminuir la temperatura global. El desarrollo de la vegetación terrestre también ayudó a facilitar el camino para que los animales invadieran la tierra, quedando protegidos contra el calor o la deshidratación. Los primeros vertebrados terrestres son los del Devónico tardío de los hemisferios norte y sur. La mayoría de los anfibios primitivos eran acuáticos pero ciertas icnitas indicarían que algunos pudieron ser terrestres. Sin embargo, las plantas no constituyeron un incentivo como fuente alimenticia para que los animales migraran a la tierra, pues los primeros animales terrestres eran carnívoros; los herbívoros fueron minoría. Más importante habría sido la protección y el ocultamiento ofrecidos por las plantas. La caída del CO 2 en el Devónico (400-360 m.a.) según Retalleck se habría debido a la difusión de las plantas vasculares en las áreas elevadas continentales, con formación de los primeros suelos barrosos. Estas plantas con sus raíces aceleraron la meteorización química de las rocas provocando un incremento en la remoción del CO 2 atmosférico. En el límite entre los pisos Frasniano y Fammeniano, al final del Devónico, resultó afectada tanto la faun de los invertebrados como vertebrados, no así a las floras. Entre los invertebrados desaparecieron los tentaculítidos,, la mayoría de los hiolítidos, gran parte de los foraminíferos, briozoos, estromatoporoideos, corales y braquiópodos, varios órdenes de trilobites (facópidos, líchidos y odontopléuridos), diversos artrópodos trilobitoideos, ciertas clases de echinoideos (Cistoideos), muchos nautiloideos y los cefalópodos climénidos. La biomasa de los arrecifes estromatoporoideos y corales (tabulados y rugosos) resultó muy afectada. Los braquiópodos de aguas frías fueron más afectadas que los de aguas cálidas. Entre los peces marinos sobrevivió sólo el 35% de los placodermos y el 12% de los acanthodianos, mientras que entre los de aguas dulces lo hicieron el 77% y 70% respectivamente. La mayor caída de la diversidad habría ocurrido al final del Frasniano. Vida marina en el Devónico. Diversificación de la vida terrestre en el Devónico. El Carbónífero y sus organismos (359-297 m.a.) En el Carbonífero o Carbónico se concretó la reunión de las principales placas litosféricas en un supercontinente o Pangea. Dicho acontecimiento involucró el cierre de grandes océanos, la producción de intensas orogenias y la formación de importantes cordones montañosos. Grandes glaciares se formaron en las altas latitudes del continente meridional de Nigritia durante el período Carbonífero, pero gigantescos bosques pantanosos crecieron en el interior tropical de todos los continentes. Estos bosques no estaban compuestos de los tipos de árboles que conocemos actualmente, sino que estaban dominados por formas arbóreas gigantes de helechos y equisetos con pequeñas hojas, las calamitales. Los restos fosilizados de tales árboles formaron los depósitos de carbón que ahora excavamos para obtener energía. En muchas regiones se formaron ricos yacimientos carboníferos, favorecidos por la presencia de zonas climáticas cálidas y húmedas y el desarrollo de exuberante vegetación. La diversidad de los animales terrestres se incrementó enormemente durante el Carbonífero. Los caracoles, los escorpiones, los ciempiés y los insectos fueron abundantes y diversos. Los insectos evolucionaron alas, volviéndose los primeros animales voladores. El vuelo les dio acceso a plantas altas; las plantas fósiles de este período muestran evidencia de la actividad de ramoneo de los insectos. Por su parte, los anfibios se hicieron cada vez más y mejor adaptados a una existencia terrestre luego de separarse del linaje que condujo a los amniotas–vertebrados con huevos bien protegidos que pueden ser puestos en ambientes secos. En los mares, los crinoideos (lirios de mar y estrellas pluma) alcanzaron el pico de su diversidad, formando “praderas” del fondo marino. Las faunas marinas carbónicas de invertebrados estuvieron dominadas por fusulínidos (grupo de foraminíferos aparecidos en el Missisípico tardío), briozoarios trepostomátidos y criptostomátidos, corales tetracorálidos y tabulados, braquiópodos productáceos, espiriféridos y rinconéllidos, ammonoideos goniatítidos y equinodermos blastoideos y crinoideos. Los fusulínidos proliferaron en los mares cálidos de plataforma donde sus acumulaciones originaron calizas. Los moluscos marinos estuvieron representados por abundantes y diversificados bivalvos y gastrópodos además de los goniatítidos. Artrópodos marinos comunes fueron los merostomados, entre los que se hallan xifosúridos muy similares al actual cangrejo bayoneta y abundaron los ostrácodos. En el notable yacimiento de Mazon Creek (Pennsilvánico Medio de Illinois) muestra restos de isópodos (bichos bolita) y otros fósiles excepcionalmente preservados, como moldes de hidras, quetognatos, nemertinos, priapúlidos, insectos y peces, incluyendo entre los últimos a formas larvales y a una lamprea. Mientras, los trilobites muestran una marcada declinación y este hecho junto con la extinción de los graptolites dendroideos, constituyó uno de los acontecimientos más destacables de la evolución de las formas de estos tiempos. Los crinoideos llegaron al clímax de su desarrollo y sus restos contribuyeron a la formación de calizas. En los ambientes lacustres o húmedos y carboníferos habitaron nematelmintos, conchóstracos, crustáceos sincáridos, escorpiones y artropléuridos, gigantescos miriápodos de hasta 3 m de largo con tubérculos en toda su parte dorsal y apéndices birrámeos (a diferencia de los restantes miriápodos que son unirrámeos). Los bivalvos comenzaron a ser comunes en las aguas dulces y salobres y los gastrópodos pulmonados se distribuyeron en los diferentes ambientes terrestres. Grandes progresos experimentó la organización de los insectos, diversificándose los insectos alados (que tímidamente habían hecho su aparición al final del Devónico Los insectos de la primera parte del Carbonífero son todavía ápteros, y a los colémbolos se suman los dipluros y tisanuros. El origen de las alas se ha atribuido a la transformación ya sea de las branquias, los lóbulos paranotales o los estilos torácicos. Posiblemente, los insectos con alas se originaron de antecesores acuáticos que estaban asociados con una vegetación de pantanos a lo largo de la orilla de lagos, y no habrían sido totalmente terrestre sino formas semiacuáticas. En estas zonas debieron abundar las corrientes ascendentes que, junto a los remolinos, llevaban hacia arriba a los insectos ubicándolos en nubes lluviosas. Así nacieron las alas, para hacer posible un retorno exitoso al agua. Los insectos pterigotos más antiguos son los paleópteros Palaeodictyoptera, dominantes en el Pensilvánico y el Pérmico, que alcanzaron dimensiones gigantescas (como Meganeura). En el Pensilvánico aparecieron también los insectos con alas plegables (neópteros). Pertenecían a los ortópteros (grillos y saltamontes), con ojos compuestos, ocelos, y, a veces, un par de élitros no usados en el vuelo. Actualmente tienen alas reducidas o ausentes y entre sus representantes se hallan formas saltadoras (langostas, grillos). Se suele incluir en este grupo a los blattoideos o cucarachas, que fueron muy variadas en el Paleozoico Tardío. En el Carbónico, los arácnidos transformaron sus hábitos acuáticos en terrestres, pero al final del período muchos órdenes se extinguieron. La exuberante flora carbónica favoreció el desarrollo de muchos artrópodos, entre los que se hallaron numerosos miriápodos, algunos de hasta 30 cm de longitud, como los arquipolípodos que habrían vivido sobre las hojas. Durante la transición del Devónico al Carbónico en los ambientes marinos abundaron los tiburones hibodontoideos, con dentición adaptada a triturar conchillas, mientras que en los ríos y lagunas proliferaron los “tiburones” pleuracántidos, con aleta dorsal, cola dificerca y una larga espina poscraneal. Los peces paleoniscoideos lograron su clímax evolutivo en el Carbónico TardíoPérmico. En el Carbónico, los anfibios primitivos se adaptaron rápidamente a las condiciones terrestres, para lo cual debieron perfeccionar sus sistemas respiratorio y auditivo, y reforzar considerablemente su eje vertebral. Las aberturas nasales externas se fueron desplazando hacia la parte dorsal del cráneo, desarrollándose un definido pasaje nasal que las unía con la garganta. Destrás de los ojos una cavidad ótica u oido medio alojó a la membrana timpánica unida al oido interno por el estribo. Se definieron asimismo dos tipos de vértebras: las embolómeras compuestas por dos elementos (intercentro y pleurocentro), aproximadamente de igual tamaño; y las raquítomas, con el intercentro grande, en forma de cuña, y el pleurocentro pequeño. Los anfibios primitivos conservaron los dientes laberintodontos de sus antecesores ripidistianos y pertenecieron a los temnospóndilos. Eran organismos de hasta 1 m de longitud, con la cabeza grande y pesada y el cuerpo levemente elevado sobre el suelo; si bien fueron de hábitos terrestres, también podían frecuentar los ambientes acuáticos. Las formas más antigua se hallan en el Carbónico Inferior de Escocia, y sobrevivirían hasta el Cretácico del supercontinente Gondwana. Algunos especialistas opinan que los temnospóndilos originaron a los Lisanfibios o anfibios modernos (anuros, urodelos, ápodos) de pequeño tamaño y cuerpo delicado. Los anuros (sapos y ranas) experimentaron una reducción del esqueleto y una notable especialización de los hábitos, siendo Miobatracus (Pensilvánico) uno de los primeros. El resto de los anfibios carbónicos pertenece a los antracosáuridos, también surgidos en el carbónico temprano. Llegaron a ser animales de gran tamaño (hasta 4 m de largo) que vivieron en el Carbónico-Pérmico y se alimentaban de peces. Numerosos y diversificados, la mayoría tenía hábitos acuáticos y evolutivamente son de gran interés pues de ellos derivaron los reptiles en el Carbónico Tardío. En los reptiles primitivos, el cráneo se hizo más alto y estrecho que en los anfibios y desapareció la cavidad ótica de estos últimos, lo que facilitó la articulación mandibular, mientras que en la parte posterior, una membrana timpánica se ubicó en una nueva cavidad formada en el oído medio. Un paso importante en la evolución de los vertebrados fue la aparición del huevo Cleidoico (condición amniota), dotado de una cáscara protectora que le permitió ser depositado fuera del agua para su ulterior desarrollo y por dentro con varias membranas extraembrionarias (corion, alantoides y amnios). En el huevo amniota, el embrión se alimenta de la yema, y los desechos del metabolismo son acumulados en un saco alantoideo, que también participa en el intercambio gaseoso. El amnios es un saco con un líquido cuya composición es cercana a la del agua mar, en la que se ubica el embrión. Los tetrápodos que tienen este tipo de huevo se denominan amniotas y actualmente incluyen a los reptiles, aves y mamíferos. Tetrápodos indiscutiblemente amniotas aparecieron en el Pennsilvánico o Carbónico superior (Westfaliano, Moscoviano) de Nueva Escocia, donde se halló Hylonomus, un reptil anápsido pues su cráneo carece de aberturas temporales, condición que es aún retenida en los quelónidos. Los diápsidos aparecieronn en el Carbónico Superior con géneros como Petrolacosaurus R. Cowen (1994) destaca que la transición evolutiva desde los peces a los reptiles, pasando por los anfibios, fue un proceso rápido que involucró formas pequeñas y carnívoras, mientras que el hábito herbívoro recién habría surgido en el Pérmico. Las razones de esto último no están claras, aunque parecería que el pequeño tamaño y el hábito carnívoro resultaron propicios para habitar en medios boscosos, poblados de artrópodos y en el que la lignina de los troncos y las hojas podrían haber tenido sustancias químicas que resultarían tóxicas a estos primitivos tetrápodos. La flora carbónica se caracterizó por la proliferación de árboles de talla gigantesca y un frondoso foliage. El clima cálido y húmedo que reinó en Laurusia y otras regiones facilitó la depositación de materia vegetal en cuencas, las que posteriormente se transformarían en importantes minas de carbón ahora explotadas. Plantas notables por su porte fueron las licófitas Lepidodendron y las Calamites de tallo articulado. Otros grupos fueron de naturaleza herbácea, como las Sphenophyllales, probables lianas extinguidas al final del Paleozoico, igual que las formas arbóreas antes mencionadas, y de los que sólo quedan como relictos Selaginella y Equisetum. En esta flora también existieron líquenes, briofitas, helechos, progimnospermas y pteridospermas. Las progimnospermas continuaron siendo importantes árboles de los bosques al comienzo del Carbónico pero luego declinaron por la importancia que adquirieron las plantas con semillas. En las tierras bajas existieron los cordaites, pteridospermales de grandes hojas de tipo helecho y órganos reproductivos de tipo conífera, si bien las primitivas coníferas (Lebachiales) y quizá cycadales habrían comenzado su evolución en las tierras altas, donde también se habrían hallado otras formas enigmáticas como las Vojnovskyales, cuyos miembros posiblemente fueron árboles con grandes hojas en forma de abanico y conos que llevaban semillas y órganos polínicos. También habrían hecho su aparición en estas tierras altas, plantas de tipo Ginkgo, pero hay pocas evidencias directas. Gigantescos bosques carboníferos. Meganeura, pariente gigante fósil de las libélulas. Mares carboníferos: resaltan los tiburones primitivos y nautiloideos junto con briozoos y crinoideos. El Pérmico (297-251 m.a.) Durante el período Pérmico La colisión entre Laurusia y Nigritia culminó en el Pérmico, completándose la elevación total de los Varíscides, mientras que al finalizar el Período, la fusión de Laurusia con Siberia dio lugar al ascenso de los Montes Urales y a la constitución de Laurasia. De este modo quedaba reconstruida la Pangea de Wegener. Paleoclimáticamente, en el Pérmico Temprano continuaron diferenciándose un área cálida, en gran parte árida, distribuida a lo largo del paleoecuador, habitada por anfibios y reptiles, en la que se depositaron estratos rojos y evaporitas; y otra región húmeda que se prolongó hasta el Paleopolo Sur y en la que se habrían seguido produciendo episodios glaciales en los comienzos del período. En el Pérmico Tardío las condiciones se hicieron más uniformes y cálidas. En diversas cuencas prosiguió la formación de carbón. Las rocas pérmicas contienen representantes de la mayoría de los grupos modernos de insectos (alados). Hacia fines del período, los reptiles superarían en número a los anfibios. Tardíamente en el período, el linaje que condujo a los mamíferos divergió también. Mientras tanto, en las aguas dulces, el período Pérmico fue un tiempo de amplia diversificación de los peces de aletas con radios (actinopterigios). En los ambientes marinos continuó la evolución de muchos grupos de invertebrados ya presentes en el Carbónico. Los ambientes terrestres fueron poblados por una variada fauna de moluscos y artrópodos; los reptiles experimentaron una gran diversificación, mientras que las floras mostraban la misma exuberancia y riqueza de especies que en el Carbónico Tardío, si bien a nivel familiar y categorías inferiores se produjeron sustituciones importantes. No obstante, al finalizar el período, una profunda crisis evolutiva provocó numerosas extinciones. Las asociaciones de invertebrados dominantes fueron semejantes a las pensilvánicas, aunque se produjeron reemplazos importantes en los niveles inferiores a familia. Al comienzo del Pérmico, las biotas marinas aún se diferenciaban en comunidades de aguas cálidas y frías: mientras que en los primeros dominaban fusulínidos, corales, briozoos, goniatítidos, en los segundos lo hicieron braquiópodos y bivalvos. Los ammonoideos goniatítidos, que desarrollaron suturas más complejas. Los arrecifes fueron frecuentes en las aguas cálidas de poca profundidad. Muy escasos trilobites poblaron los mares pérmicos y todos fueron ptichopáridos opistoparios. Los equinodermos tendieron a la disminución del número de placas de los crinoideos, y aparecieron los equinoideos regulares con dos hileras de placas simples en cada ambulacro, como los actuales. En el ambiente continental proliferaron los bivalvos, gastrópodos y conchóstracos, apareciendo los insectos con metamorfosis compleja. En el Pérmico los peces estuvieron representados por los últimos acanthodianos además de representantes de los condrictios y osteictios. Los condrictios fueron muy variados aunque la mayoría de sus restos fósiles consisten en dientes o espinas, como el caso de los euglenodóntidos o edístidos, comunes en el Carbónico y Pérmico, conocidos sólo por sus dientes que crecían en forma espiralada. Las quimeras u holocéfalos, aun vivientes, ya se hallan en el Carbónico Inferior de Escocia, si bien las formas modernas datan del Jurásico. Los pleuracántidos (o Xenacántidos) fueron formas de agua dulce con aspecto de tiburón, mientras que los ctenacántidos (Devónico-Triásico) eran mucho más parecidos a los tiburones actuales. Los tiburones hibodóntidos (CarbónicoPaleoceno) tenían dientes delanteros con cúspides agudas, mientras que los traseros eran redondeados y achatados, funcionando como superficies trituradoras de moluscos. Los actinopterigios contaron con formas primitivas como los paleoniscoideos (Devónico-Cretácico). Una de las características del Pérmico en el sector meridional de Pangea, el núcleo de la futura Gondwana, es la flora de Glossopteris se compone de licópsidas arborescentes, equisetales en su mayoría endémicas, Marathiales, Progimnospermales y Pteridospermales. Las Glossopteridophyta eran pteridospermales arbóreas, su tronco tenía anillos anuales de crecimiento y la raíz (Vertebraria) estaba adaptada a habitar lugares pantanosos o cercanos a los ríos. Las hojas son de muy diferente tamaño y forma; se disponían en espiral alrededor de las ramas y generalmente, se las encuentra sueltas pues eran deciduas. Incluyen cuatro morfogéneros: Glossopteris, Gangamopteris, Palaeovittaria y Rubidgea, las que se disponían en grupos de hasta 15 de ellas, siendo la mayoría sésiles. Las hojas se ción entrecruzada formando una red; cuando existe una vena media se llama Glossopteris, si no la hay pero muestra una red de venas es Gangamopteris y si carece de vena media y tiene escaso entrecruzamiento, se denomina Palaeovittaria. En Glossopteris la vena media está formada por un manojo de venas muy apretadas, existiendo toda una gradación desde Glossopteris – Gangamopteris – Palaeovittaria. Las Glossopteridales llevan órganos reproductivos masculinos y femeninos y semillas con cápsulas aladas. Las correspondientes a las formas pérmicas argentinas se llaman Ottokaria, Lanceolatus y Arberia. El morfogénero Rubidgea se hallaría entre los más primitivos y ancestrales del grupo y Glossopteris representaría la culminación de una evolución que habría llevado hacia el aumento de tamaño de la hoja, desarrollo de nervaduras reticuladas y concentración de las nervaduras en el eje foliar central. Después del Pérmico, las hojas de Glossopteris son raras, hallándose algunas en el Triásico. Es difícil establecer si las Glossopteridales se extinguieron sin dejar descendientes o si originaron a grupos más modernos. Algunos especialistas sospechan que pudieron ser las antecesoras de las angiospermas. El origen de las Glossopteridales debió estar relacionada con la glaciación gondwánica del final del Carbónico, y cuando este episodio estaba llegando a su término, se constituyó una flora impura integrada por formas carbónicas del Hemisferio Norte y otras originadas durante la glaciación, pero que constituía más de la mitad de la flora de la región. Los anfibios característicos continuaron siendo los laberintodontes antracosaurios, mientras que entre los lisanfibios surgieron y prosperaron grupos enteramente fósiles como los Aistopoda, Nectridia y Microsauria se extinguieron al final del Pérmico Inferior. Tetrápodos pérmicos típicos fueron los seymouriomorfos y diadectomorfos y los reptiles primitivos. También destacaron los temnospóndilos (Pérmico-Triásico), entre ellos Eryops carnívoro y que medía 2 m de largo. También hicieron su aparición los primeros sinápsidos, en la línea d elos mamíferos, los pelicosaurios, grupo que en el Pérmico Temprano constituyó más del 70% de los sinápsidos. Los había carnívoros y herbívoros, pequeños y hasta de 3 m de largo, con “vela” y sin ella. La familia más numerosa de pelicosaurios fue la de los esfenacodontes, carnívoros del Carbónico Superior y Pérmico Inferior de América del Norte y Europa. Un ejemplo fue Dimetrodon.. En el Pérmico Temprano, los pelicosaurios esfenacodontes habrían dado origen a los Therapsida o “reptiles mamiferoides”, del Pérmico-Triásico de Rusia y Sud Africa, con varios grupos: - Teriodontes, los más primitivos de los therápsidos y que comprendieron a los: ·Gorgonopsianos, con caninos largo, en forma de hoja de sable, e incisivos fuertes; el resto de la dentición era débil. Exclusivos del Pérmico fueron los carnívoros dominantes en el Pérmico Tardío. ·Thenocephalianos, carnívoros del Pérmico Superior y Triásico. ·Cynodontes, con dientes con varias cúspides y corona trituradora desarrolladas como en los mamíferos. Aparecen en el Pérmico Tardío y tuvieron gran importancia en el Pérmico gondwánico. De ellos habrían descendido los mamíferos. - Dinocéfalos, conocidos sólo en el Pérmico Superior de Rusia y Sud Africa. Carnívoros y herbívoros. Moschops, de Sud Africa, midió 5 m de largo. - Dicinodontes, constituyen el 90% de los terápsidos del Pérmico Tardío; luego declinaron y se extinguieron en el Triásico. Herbívoros, sin dientes, excepto los caninos superiores; mandíbula terminada en un pico córneo. En el Pérmico, los reptiles típicos diápsidos (ancestros de saurios y ofidios) estuvieron estuvieron representados por formas primitivas pertenecientes a los Eosuquios, que habrían sido el tronco ancestral desde el que radiaron las restantes líneas filogenéticas diápsidas. Coelurosaurus haeckeli tenía una membrana plegable a ambos lados del cuerpo que le permitía “planear”, mientras su cola larga y flexible actuaba como “estabilizadora”. Eryops, típico anfibio carbónico. Dimetrodon, predador pérmico y pariente de los mamíferos. Fauna marina pérmica antes de la extinción. La flora de Glossopteris (extendida en el sur de la Pangea durante el Pémico) y reconstrucción de un árbol de Glossopteris y sus principales órganos reproductores. La crisis pérmica, madre de todas las extinciones de la historia terrestre La culminación del Pérmico se caracterizó por una profunda crisis (la de mayor importancia del Fanerozoico) que afectó más a los animales que a los vegetales, produciendo una drástica reducción de la diversidad total. Entre los invertebrados se extinguieron los fusulínidos, conuláridos (con una excepción triásica), corales tabulados y rugosos, briozoos trepo y criptostomátidos, braquiópodos órthidos, dalmanelláceos y productáceos, goniatítidos, euriptéridos, ostrácodos leperdítidos, trilobites, equinodermos blastoideos, crinoideos inadunata, flexibilia y camerata. Ocho de los 27 órdenes de insectos desaparecieron, como también muchos arácnidos. Las comunidades de los exitosos arrecifes del Pérmico resultaron muy disminuidas. Otros grupos de invertebrados, como los bivalvos y gastrópodos, fueron poco afectados. Los peces declinaron en su diversidad. Los tetrápodos de menos de 10 kg de peso, así como los de agua dulce, experimentaron extinciones de escasa importancia, no así los marinos y los grandes herbívoros terrestres. Entre los vertebrados desaparecidos se hallaron los anfibios raquítomos y los reptiles seymouriomorfos y captorrinomorfos, pelicosaurios y Mesosaurus. Alrededor del 49% de las familias de tetrápodos desaparecieron. Entre las plantas, importantes grupos no superaron la terminación del Pérmico, como Lepidodendron, Sigillaria, Cordaites, Coenopteridales (helechos primitivos), Calamites, y muchas pteridospermales. Esta extinción se habría llevado a cabo en dos etapas: al final del Guadalupiano, con la extinción del 58% de los animales, y a la culminación del Tatariano, con la eliminación del 80% de las especies marinas. Numerosas causas se han invocado para explicar esta crisis, en la que seguramente tuvieron particular influencia la recomposición de la Pangea y los cambios climáticos. Una primera es la caída de un gran asteroide, cuya evidencia sería el cráter Woodleigh de Australia occidental, pero también podría haber caído otro fragmento en la Antártica. El primer proceso redujo las áreas ocupadas por mares someros de un 43% en el Pérmico temprano al 13% en el Pérmico Tardío, con la consiguiente supresión de hábitats, disminución de la cantidad de alimentos y el aumento de la competencia. En el ámbito terrestre, se intensificaron las migraciones con aumento de la competencia y extinciones, a la vez que las condiciones climáticas se hacían más severas. Sin embargo, más probable es que esta crisis se haya debido a la concurrencia de una serie de circunstancias asociadas a la propia evolución de la corteza terrestre, como las regresiones, vulcanismo y transgresión de aguas pobres en oxígeno. Las causas extraterrestres parecen estar excluidas. Al final del Pérmico, el descenso del mar de unos 150 m ocasionó un brusco cambio climático, con aumento de la estacionalidad; los nichos ecológicos se redujeron notablemente. En el límite Pérmico–Triásico, en el norte asiático tuvo lugar el vulcanismo de los Siberian Traps que, en 1 millón de años originó unos 2 millones de km cúbicos de lavas. Los materiales eyectados habrían provocado una Edad del Hielo, mientras las aguas oceánicas se acidificaban por la incorporación del ácido sulfúrico y el exceso de dióxido de carbono (efecto invernadero) pudo dar lugar a un calentamiento nocivo para la vida. Inmediatamente después de la regresión, el mar invadió nuevamente las plataformas, cubriéndolas con aguas muy pobres en oxígeno procedentes de las profundidades oceánicas, en donde habría permanecido estancada. Esto afectó al microplancton y fueron eliminados muchos organismos que habrían sobrevivido a la regresión. Las erupciones volcánicas masivas resultaron en bocanadas de lava que cubrieron áreas extensas de la Tierra. La ceniza generada por los volcanes bloqueó la luz solar y enfrío el clima, dando por resultado los mayores glaciares de la historia terrestre. El oxígeno atmosférico gradualmente descendió del 30 al 12%. Con tales concentraciones de oxígeno, la mayoría de los animales no habrían sido capaces de sobrevivir a elevaciones de más de 500 metros; de esta manera, cerca de la mitad de la tierra pérmica habría sido inhabitable. La combinación de estos cambios resultó en el evento de extinción masiva más drástico de la historia terrestre. Muchas especies se extinguieron simultáneamente hacia fines del período; otras desaparecieron gradualmente durante un período de varios millones de años. La extinción del Pérmico pudo haber llegado peligrosamente a hacer desaparecer totalmente la vida de la Tierra. Los científicos estiman que cerca del 96% de todas las especies se extinguieron en ese tiempo. ACTIVIDADES 1. Explique las características de la transformación de la fauna marina desde el Cámbrico hasta el Pérmico. Relacionen con los cambios paleogeográficos y climáticos. =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 2. Señale al menos dos características biológicas globales que caracterizan al Cámbrico, al Silúrico y al Pérmico. =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 3. Analice el concepto de “Carnaval de los Animales”. Señale algunas de las características biológicas del Cámbrico inferior y medio a partir de la interpretación de los fósiles de Chengjiang y Burgess Shale. =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 4. Asocie con los períodos de la era Paleozoica los siguiente eventos: a. Aparición de los primeros “anfibios”: b. Aparición de los lisanfibios: c. Aparición de los vertebrados: d. Aparición de los artrópodos: e. Aparición de los bivalvos: f. Aparición de los cefalópodos: g. Aparición de las gimnospermas: h. Aparición de los musgos: i. Aparición de los hongos de sombrero: j. Aparición de los helechos: k. Aparición de los diápsidos: l. Aparición de los sinápsidos: m. Aparición de las plantas vasculares (traquéofitos primitivos). 5. Explique los principales cambios que se produjeron en los ambientes aeroterrestres a partir del Ordovícico. =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 6. Reconstruya a partir de los datos analizados en el módulo las características principales de la evolución de los vertebrados en la era Paleozoica. =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 7. Haga una lista de ganadores y perdedores en las siguientes extinciones: ordovícico final, devónico final, pérmico final. Tenga en cuenta además la siguiente información. ¿Por qué todos los niveles de radiación evolutiva de los animales se remontan, en definitiva, al Cámbrico? Se pueden considerar siete niveles en la evolución de los animales básicamente atendiendo al ambiente marino: a) El primero grupo de Eumetazoa está representado por la ediacarian fauna (fauna ediacariana) de 700 a 600 millones de años ya que junto con los Vendozoa, está representada por gran variedad de Radiata y grupos primordiales o basales de Bilateria como Sprigginia, Kimberella y Arkarua, en depósitos de Australia S. en las Ediacara Hillls, Inglaterra, Escandinavia, Namibia, Rusia, las sierras de Ventana en Argentina o Terranova. b) El segundo nivel está representado por la transición ediacariana-tommotiana (Cámbrico inferior) conocida como small phosphatic shelly fauna (fauna fosfática de pequeños caparazones) hasta los 600 a 550 millones de años situada básicamente en la plataforma de Siberia S., formación Petrotsvet, así como en otra importante sección china, Meishucun en Yunnan, e igualmente en Inglaterra, Australia C. y Escandinavia. Se caracteriza por la aparición de los primeros organismos con conchas microcópicas fosfáticas, como los Tommotozoa o los Hyolithelmintha, etc., y es interesante notar que muchos elementos de la fauna de la época que persistirían como los Brachiopoda presentaban por entonces conchas fosfáticas, debido a los altos niveles de fósforo, hasta que paulatinamente se establecieran otros tipos de conchas calcáreas más modernas, como los Cloudinozoa. Se sugiere que la aparición de los primeros elementos esqueletarios fue un hecho accidental y que luego mostró un gran desarrollo potencial, probablemente correlacionado con las piezas masticatorias de algunos organismos predadores que también aparecieron en esta época, como los Protoconodonta. c) El tercer nivel está representado por los sedimentos tambien del Cámbrico inferior (atdabanianos), de 550 a 530 millones de años, como en el Syrius Passet de Groenlandia, el Mount Cap de Canadá y el Emu Bay Sahle de Australia S., y de Sierra Morena en España. Se conoce como small carbonatic shelly fauna o fauna de caparazones carbonáticos pequeños, con ejemplos princiales en los Coelosclerithophora y Cambroclava. En esta fauna ya son más abundantes los elementos carbonáticos, lo que probablemente coincide con un descenso a los niveles actuales del nivel de fósforo. Junto con los animales de esqueletos carbonáticos microscópicos, se desarrolla una fauna sin partes duras, también altamente diversificada. Los elementos de esta fauna serán reemplazados drásticamente por los del cuarto nivel. d) El cuarto nivel esté representado por los sedimentos igualmente del Cámbrico inferior (botomianos) en China, Chengjiang, y el quinto nivel por la rica fauna del Cámbrico medio de Burgess Shale, Canadá, de Suecia o de Siberia N., reunidos con el nombre de soft cambrian fauna (para la fauna cámbrica blanda) y old big shelly fauna (fauna de grandes caparazones antigua). En ambos casos, los elementos carbonáticos de los Trilobita, Archaeocyathia, Brachiopoda, Hyolitha, Mollusca primitivos como los Galeroconcha o e) f) los primeros Echinodermata están acompañados por animales de partes blandas como los Kerygmachelea o los Banffimorpha. La diferencia entre ambas faunas es mayormente porcentual, y mientras que en la primera se asiste a la radiación de los Mollusca y Brachiopoda, en la del Cámbrico medio se produce una intensa radiación evolutiva de los Arthropoda. Estas faunas habrían de proliferar durante el Ordovícico, pero declinarán al final del período, y se mantendrán con variaciones hasta el Devónico, y con muy pocos elementos llegarán al Pérmico, cuyo episodio de extinción final no resistirán. El sexto nivel está representado por los yacimientos del Cámbrico superior al Pérmico, la big paleozoic shelly fauna o fauna paleozoica de grandes caparazones, con típicos esqueletos de grupos más modernos, como los Brachiopoda Articulata, Echinodermata Crinozoa, Anthozoa, Ostracoda, un grupo de Crustacea, Cephalopoda, un grupo de Mollusca modernos, Bryozoa y Graptolitha. Pero junto con la fauna de conchas, persistirán todavía en determinados sitios algunos grupos engimáticos y antiguos como se evidencia en otros depósitos posteriores, como del Silúrico de Wisconsin, el Devónico de Hunsrückschiefer de Alemania W, el Rhynie Chert del Silúrico de Escocia, o el Pensilvánico de Mazon Creek, Illinois, que evidencian organismos antiguos como los Palaeoscolecodonta o los Tullimonstra, además de otros muchos grupos cuyos primeros registros se observan en estos depósitos. El séptimo nivel se origina igualmente en el Cámbrico superior y durante el Ordovícico inferior, la modern fauna o fauna moderna, y está caracterizada por los grupos de animales modernos que dominan actualmente, como los Pelecypoda y Gastropoda entre los Mollusca, los Malacostraca entre los Crustacea, los Echinozoa entre los Echinodermata, y los distintos grupos de Vertebrata. Esta fauna fue experimentando distintos episodios de diversificación y extinción, hasta que a fines del Cretácico, luego de la segunda gran extinción terrestre, será la única bien representada en el fondo marino. =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 8. Señale de la siguiente lista de organismos aquellos que pasan de la transición Pérmico-Triásico (1), los que se extinguen completamente (2) y los que quedan muy menguados (3). Ammonoideos Nautiloideos Licófitos Helechos verdaderos (filicófitos) Gimnospermas Lisanfibios Insectos Trilobites Foro La proliferación marina, los comienzos de la evolución, los eslabones perdidos y la discusión con el creacionismo Existieron dos posturas filosóficas hasta el siglo XIX, creacionismo/fijismo y evolucionismo. La primera postula que no se produjeron cambios en la estructura de los organismos y que todos siguen siendo iguales al plan original de la creación. La segunda en cambio, postula que los organismos se transforman con el paso del tiempo. Durante el siglo XVII, la interpretación literal de la Biblia comenzó a ser cuestionada y resurgieron las explicaciones transformistas sobre las causas de diferentes fenómenos naturales, como la diversidad de organismos. Se considera que en ese siglo nació la “ciencia moderna”. Esta nueva ciencia se sustentó en que toda explicación debe validarse a través del método científico. Este modelo de conocimiento de las Ciencias naturales se caracterizó por privilegiar la observación y la experimentación, con un riguroso registro y cuantificación de los resultados, de manera de permitir la relación entre los hechos, las hipótesis y las conclusiones. Además de la importancia de la adquisición de un método sistemático de investigación, la Ciencia moderna no hubiera podido emerger sin el apoyo de la tecnología desarrollada en este siglo. La gran cantidad de inventos en instrumentos de medida y de observación resultó un aporte indispensable en la producción de nuevos conocimientos científicos. Uno de los mayores opositores a las ideas creacionistas y fijistas aún vigentes en ese siglo, fue Georges-Louis Leclerc, conde de Buffon (170 – 1788). Para responder a la pregunta sobre el origen de los fósiles, retomó la idea de selección natural de Empédocles y la propuso como la causa de la extinción de las especies: “[…] No es imposible que […] todos los animales del Nuevo Mundo fueran los mismos del Antiguo Mundo, derivados de ellos, podría decirse que, al quedar separados por mares inmensos o tierras impracticables, con el tiempo sufrieron los efectos de un clima… y al cabo de un determinado período, se transformaron […]”. En poco tiempo, estas ideas se impondrían a partir de los estudios de Charles Robert Darwin y Alfred Russel Wallace, quienes postularon la visión moderna de la Teoría de la evolución. No obstante, todavía algunos círculos discuten una teoría alternativa, el diseño inteligente, que no es científica ya que no puede ser refutada o falsada, y no difiere demasiado del creacionismo original, aunque no especula sobre la creación divina, aunque sí reconoce una naturaleza inteligente detrás de la evolución. Estos principios evolutivos pueden analizarse con las transiciones faunísticas observadas en la era Paleozoica. Durante la era Paleozoica, la vida marina continuó su proliferación a partir del período Ordovícico, y continuó durante los períodos restantes: Silúrico, Devónico, Carbonífero y Pérmico. Se desarrollaron todos los grupos de invetebrados marinos que viven en la actualidad y también los peces. Entre los invertebrados marinos, se destacaron los trilobites, los nautiloideos, de los cuales todavía persiste el género Nautilus, y los euriptéridos o escorpiones de mar. Desde el Silúrico, proliferaron diferentes grupos de peces, como los acantodios o tiburones espinosos; los diferentes grupos de ostracodermos o peces acorazados sin mandíbulas y los placodermos, con mandíbulas. En el período Devónico, se hicieron más abundantes los peces óseos, representados tanto por los actinopterigios de aletas radiadas, como los paleoniscoides, como los sarcopterigios de aletas lobuladas, los cuales darían origen a los primeros vertebrados terrestres o tetrápodos, los anfibios. Los científicos ya cuentan con fuertes evidencias de la transición entre peces y anfibios, comenzando por los elpistostegalios, como Panderichthys, que eran peces de aletas lobuladas en transición a respirar oxígeno, y los primeros “anfibios” fósiles devónicos (los ictiostegalios). Y si algunos autores estiman que estos fósiles fueron realmente “missing links” (eslabones perdidos), hoy se afirma que la evolución no funciona linealmente, sino en retículo y mosaico. Por esta razón, una objeción tradicional de los creacionistas acerca de la evolución, ya no tiene sentido, porque no existen eslabones perdidos reales, sino series de transiciones que por la suma de mutaciones van cambiando la fisonomía de los organismos hasta producir barreras reproductivas. Panderichthys, no eslabón perdido pero sí en transición hacia los anfibios. ¿Qué piensan ustedes del diseño inteligente o de la creación? ¿Qué diferencias se pueden postular entre ciencia y teología o religión? ¿Por qué el diseño inteligente no puede ser considerada una teoría científica? ¿Existen los eslabones perdidos? ¿Por qué se sigue afirmando esto desde los medios de comunicación? ¿Evolución gradual o evolución a saltos? En la conquista del medio aeroterrestre Muchos científicos adhieren a la perspectiva gradualista de Darwin. En su teoría, el naturalista inglés explica que la evolución de las especies se produce gradualmente, en una serie de pasos casi imperceptibles. El origen de nuevas especies sería el resultado, entonces, de la suma de pequeñas modificaciones acumuladas durante largos períodos de tiempo. En síntesis, esos científicos afirman que la macroevolución (origen de los grandes taxones por encima de la especie) se produce como resultado de la microevolución o especiación. Pero al estudiar los linajes de organismos fósiles como los trilobites, se encontró que, en ciertos casos, es posible constatar la gradualidad, y en otros, constatar discontinuidades. En 1972, los paleontólogos estadounidenses Nils Eldredge y Stephen Jay Gould, cuestionaron las ideas gradualistas de Darwin y propusieron una nueva explicación a las discontinuidades de los registros fósiles. Según estos científicos, la evolución no avanza por cambios continuos y graduales, sino a través de una serie de saltos bruscos y esporádicos. Este modelo explicativo, llamado saltacionista o de equilibrio puntual, sostiene que las especies se transforman rápidamente y que luego permanecen sin variaciones durante largos períodos de tiempo. Los científicos gradualistas se oponen a los defensores del equilibrio puntual alegando que las discontinuidades se deben a la imperfección en la búsqueda de “eslabones perdidos”. Evolución a saltos o evolución gradual es todo un tema de debate en la ciencia actual y se aplica por ejemplo a explicar las transiciones en el medio aeroterrestre durante la era Paleozoica. Hace 500 m.a., durante el período Ordovícico,, la atmósfera adquirió su composición actual de oxígeno, del 20 % (llamado nivel de Pasteur, en honor al gran científico) y disminuyó en consecuencia la incidencia de la radiación ultravioleta. De esta manera, la vida pudo desarrollarse fuera del agua. Los primeros invasores ordovícicos fueron los musgos, los hongos (incluidos los líquenes, o simbiontes entre hongos y algas) y seguramente las lombrices terrestres (anélidos). En este momento comenzó la formación de los paleosuelos o suelos antiguos. En el Silúrico, llegaron los caracoles pulmonados o terrestres, las primeras plantas vasculares o traquéotifos, como Cooksonia y Rhynia, los hongos formadores de micorrizas (asociaciones simbiontes con plantas vasculares), además de los colémbolos, pequeños insectos del suelo y los escorpiones y arañas primitivos. Durante el Devónico, la vida aeroterrestre se hizo muy abundante, con la incorporación de los helechos, las gimnospermas primitivas, los anfibios como Ichtyostega, y los insectos alados, como Meganeura, una libélula gigante. Durante el Carbonífero, en los extensos bosques pantanosos que proliferaron en un clima tropical y húmedo, surgieron árboles gigantescos, como los licopodios arborescentes. Igualmente, aparecieron los primeros reptiles anápsidos (sin aperturas en el cráneo), como Diadectes. Finalmente, en el Pérmico, comenzó el dominio de las coníferas y el actual Gingko biloba. Finalmente, durante el Pérmico y hasta su extinción final, proliferaron los reptiles mamiferoides (con una abertura temporal en el cráneo), antepasados de los mamíferos, como el gigantesco y voraz Dimetrodon, que tenía un velamen formado por una membrana alrededor de las vértebras muy alargadas. Desde el Ordovícico al Pérmico, las placas continentales se fueron uniendo progresivamente a medida que migraban hacia el norte o el sur. Estas colisiones origianaron cadenas montañosas, como los montes Apalaches en los Estados Unidos, y condujeron finalmente a la formación del supercontinente de Pangea, el más conocido y en que Wegener basó su Teoría de la Deriva continental. Esto produjo un deterioro progresivo del clima, con la desertización y el enfriamiento. Si tuvieran que destacar un evento principal en la evolución aeroterrestre, ¿cuáles elegirían? ¿Postularían una evolución gradual o una saltatoria? Justifiquen. Módulo 5. La vida en la era Mesozoica Objetivos • Explicar los cambios paleoclimáticos y paleogeográficos durante la era Mesozoica • Analizar las características de la fauna marina mesozoica • Explicar los cambios en los ecosistemas aeroterrestres durante la era Mesozoica • Analizar la evolución de diferentes grupos de organismos en las etapas de la era Mesozoica • Analizar las características de la gran extinción tardío-cretácica y evaluar diferentes hipótesis explicativas • Explicar las características faunísticas y florísticas del Triásico • Explicar las características faunísitas y florísticas del Jurásico • Explicar las características faunísticas y florísticas del Cretácico Contenidos La división de la era Mesozoica El período Triásico Vida triásica El período Jurásico Vida jurásica El período Cretácico Vida cretácica La gran extinción tardío-cretácica División de la era Mesozoica La era Mesozoica o edad intermedia de la historia de la vida en la Tierra se inició hace 251 m.a. y se divide en tres períodos: Triásico, Jurásico y Cretácico. Los escasos organismos que sobrevivieron a la extinción en masa del Pérmico se hallaron en un mundo relativamente vacío al comienzo de la era Mesozoica (hace 251 m.a.). A medida que Pangea lentamente se separó en continentes individuales, Laurasia y Gondwana, los océanos aumentaron de nivel y volvieron a inundar las plataformas continentales, formando gigantescos mares interiores someros. Las concentraciones de oxígeno atmosférico gradualmente se incrementaron a los niveles anteriores. Pero pronto, la vida comenzó nuevamente a proliferar y diversificarse, aunque diferentes grupos de organismos se hicieron dominantes en la Tierra, si bien una nueva extinción se cernió sobre los organismos en el Triásico superior (probablemente debido a un asteroide que hizo impacto en Manicougan, Canadá). Pero la gran extinción de la era Mesozoica ocurrió hace 65 millones de años, al final del Cretácico superior, cuando un asteroide chocó con la Tierra, se fragmentó en tres porciones, y la mayor chocó contra el golfo de México (el cráter Chixulub) provocando la extinción de los dinosaurios y otros organismos. Los tres grupos de fitoplancton (organismos fotosintéticos flotantes) que dominan actualmente los océanos–dinoflagelados, cocolitofóridos y diatomeas– se volvieron ecológicamente importantes en este tiempo. y otros grupos como los silicoflagelados y ébridos hicieron su aparición en el registro (aunque probablemente evolucionaron mucho antes). Nuevas plantas con semillas reemplazaron a los árboles que habían dominado en los bosques pérmicos. Durante la era Mesozoica, la biota terrestre que había sido relativamente homogénea hasta ese tiempo, se hizo crecientemente provincializada, es decir, evolucionaron distintas biotas terrestres en cada continente. Las biotas de las aguas costeras que bordean los continentes también divergieron de las restantes. El provincialismo que comenzó durante el Mesozoico continúa influenciando la geografía de la vida actual. Hacia fines de la era, los continentes se hallaban próximos a sus posiciones presentes, y muchos organismos se parecían a los que viven ahora. El Triásico (251-200 m.a.) El Triásico es el primer período de la era Mesozoica. La Pangea comenzó a fragmentarse y formó los dos grandes continentes, Laurentia al N. y Gondwana al S., y entre ambas masas continentales se interponía el Paleotethys. A pesar de los cambios tectónicos y geográficos, la mayor parte de la corteza continental se hallaba centrada sobre una faja paleoclimática cálida y árida, con frecuentes depósitos de evaporitas y sedimentitas rojas. A su vez, éste fue uno de los momentos en la historia de la tierra con mayor desarrollo de magmatismo y volcanismo. La crisis biológica del final del Pérmico fue seguida por una lenta renovación de la vida en el Triásico. La primera aparición de ammonoideos con sutura ceratítica (que reemplazarían a los goniatítidos) de la asociación de Otoceras y Ophioceras y el bivalvo Claraia generalmente se consideran el mejor indicio del comienzo del Triásico en los ambientes marinos del Paleotethys, el mar gigantesco que entonces dividía las dos masas continentales, aunque no formaba todavía un océano continuo como el Tethys jurásico. En los ambientes marinos, paulatinamente, los ammonoideos ceratítidos son reemplazados a su turno por formas con suturas más complejas y asi la base del Jurásico se toma con la aparición del amonoideo Psiloceras planorbis mientras en los ambientes continentales se hace abundante el polen de Classopollis. La crisis que afectó a la mayoría de los invertebrados marinos a fines del Pérmico fue seguida en el Triásico por una nueva expansión de la vida y nuevos taxones ocuparon los nichos ecológicos vacantes. Asi ya no se encuentran fusulínidos ni corales tabulados y rugosos ni briozoarios trepostomátidos y criptostomátidos. La aparición de los briozoarios queilostomátidos y los corales escleractínidos (madréporas) influyeron desde ese momento en la composición de los arrecifes. Los braquiópodos inarticulados fueron totalmente desplazados por los articulados, especialmente rinconélidos y terebratúlidos. Entre los moluscos se Destacarían ahora en los bivalvos aviculoidoes (Monotis), límidos, mytiloideos, ostreidos, trigonidos, etc.; entre los nautiloideos abundaban ahora los nautiliconos (espiralados) y los pocos ortocónidos que persistieron disminuyeron en tamaño y se extinguieron al final de este período. Los ammonoideos fueron ahora el grupo dominante de los cefalópodos marinos, y estuvieron netamente dominados por formas con suturas ceratíticas y al final del Triásico se extinguieron casi todos ellos, extendiéndose al Jurásico sólo los Phylloceratina de los cuales se derivaron luego los amonoideos que caracterizaron al resto de la era Mesozoica, con suturas más comlejas. Otras faunas de invertebrados presentes en este período comprenden crustáceos e insectos asi como crinoideos de características modernas. En ambientes dulceacuícolas proliferaron los conchóstracos y algunos bivalvos. Entre los peces, ahora predominaban netamente los actinopterigios paleoniscoideos para ser luego reemplazados por los "holósteos". Estos últimos aparecieron en el Triásico y alcanzaron su acmé en el Jurásico tardío. Dentro de los tetrápodos, se reconocían los anfibios laberintodontes temnospóndilos que se extinguirían en el Jurásico temprano. Otros anfibios incluían anuros primitivos (Triadobatrachus). Tanto en en los ambientes marinos como los continentales tuvo lugar una importante radiación de los reptiles, apareciendo también formas adaptadas a los ambientes marinos. A mediados del Triásico hicieron su aparición los dinosaurios y diversos grupos de reptiles marinos, en particular placodontos y nothosaurios, junto con los crocodilianos (cocodrilos y caimanes). Igualmente hicieron su aparición los quelonios o tortugas. Los restos de tortugas más antiguos son del Noriano de Alemania (Proganochelys y Proterochersis), sudeste de Asia (Proganochelys), Groenlandia (Proganochelys) y Argentina (Palaeochersis) . y arcosaurios. Entre los reptiles marinos aparecidos en el Triásico se hallan también los ictiosaurios, con forma de pez o delfín y vivíparos, y los primeros reptiles voladores, los pterosaurios (ranforincoideos). En el Triásico continental hicieron su aparición varios grupos de insectos, notablemente los tricópteros y dípteros. La fauna reptiliana triásica se completaba con los terápsidos o reptiles mamiferoideos. Dentro de éstos los cinodontes dieron lugar a los mamíferos al final del Triásico. Los teriodontes triásicos tenían muchos caracteres mamalianos tales como: dos cóndilos occipitales, paladar secundario, dentario alargado y dientes diferenciados. Apenas surgidos los mamíferos, al final del Triásico, de diferenciaron los prototerios y terios. La flora fue diezmada a fines del Pérmico y esto se ve reflejado en la casi ausencia de depósitos de carbón en el Triásico inferior. Pero en el Triásico medio a superior comenzaron a aparecer muchas familias de helechos y coníferas que perduran hasta la actualidad junto con otros grupos, como las bennetittales (cicadeoideas), ahora extinguidas pero que eran el alimento de los dinosaurios. La flora muestra ahora peculiaridades que anuncian a las asociaciones que habrán de predominar en el Mesozoico tardío. Un rasgo notable fue la disminución de tamaño de algunos grupos de plantas que llegaron a portes gigantescos en el Paleozoico superior. Las Lepidondrales desaparecieron y las Lycopodiales y Equisetales (Equisetites,Schizoneura) presentan tamaños modestos. La vegetación de las zonas bajas durante el Triásico inferior estuvo dominada por Pleuromeia. A fines del Triásico medio evolucionó una nueva asociación florística integrada por Osmundales y Matoniaceas (helechos) y Caytoniales, Corystospermales (Pteridospermales) asi como Bennettitales, Ginkgoales y Coniferales de hojas pequeñas, asi como el polen Classopollis (un tipo especializado de coníferas fósiles, las Cheirolepidáceas). Esta flora está presente en el Triásico medio y superior de América del Sur, América del Norte, Africa, Australia, Japón y Europa. Las floras más ricas del Triásico superior se encuentran en latitudes ecuatoriales, como en Alemanina y el sudoeste de Estados Unidos, con abundantes helechos, equisetales, pteridospermas, cycadales, bennettitales, ginkgoales y coniferales. Las de latitudes medias poseen características similares pero con menor diversidad específica. Placodontos, reptiles marinos típicos del Jurásico, con cierto aspecto similar a las tortugas aunque no relacionados con ellas. Lariosaurus, un nothosaurio del Triásico. Herrerasaurus, un primitivo dinosaurio triásico. El Jurásico (200-145 m.a.) Durante el período Jurásico, los continentes continuaron su división y ahora Laurasia al norte y Gondwana al sur estaban completamente separadas rodeando al océano Tethys (luego del cierre del Paleotethys previo del Triásico). Por otra parte, desde el Jurásico medio comenzó la apertura del Atlántico sur y la divergencia de dos sectores gondwánicos, Gondwana oriental y occidental. A partir del Jurásico temprano comenzó a formarse el Golfo de México y en el norte de América del Sur el desarrollo de fosas tectónicas en el Jurásico medio permitió la comunicación del Tethys con el Paleopacífico y condujo a la separación entre América del Norte y América del Sur. Estacomunicación formó parte del nacimiento del Atlántico Central, que contó con desarrollo de corteza oceánica alrededor de los 160 ma. El clima del Jurásico se caracterizó por su uniformidad, siendo principalmente templado cálido, aunque hay también amplios registros de estratos rojos y evaporitas. Dentro de los microorganismos, la vida planctónica resurgió durante el Jurásico. En el zooplancton participaron los tintínidos, radiolarios y a partir del Dogger (Jurásico medio), también los foraminíferos planctónicos (globigerínidos) y se diversificaba el nanoplancton calcáreo (los cocolitofóridos o hapófitos). Las comunidades bentónicas contaron con esponjas y corales, importantes integrantes de los arrecifes, y los braquiópodos estuvieron principalmente representados por terebratúlidos y rinconéllidos. Pero fueron los moluscos quienes dominaron todos los mares, con un predominio de los bivalvos se acentuó en el Jurásico y muchas especies desarrollaron grandes y gruesas conchillas. Frecuentemente se encuentran asociados a algas, briozoos, hexacorales, crinoideos y ammonoideos. Los géneros más comunes fueron: Cucullaea, Ostrea, Grypaea, Exogyra, Isognomon, Pholadomya, etc. En los gastrópodos aparecen las formas aladas (Harpagodes). Las faunas de cefalópodos se compusieron principalmente de ammonoideos, además de belemnoideos (ancestros de los calamares y sepias). Se conocen también los primeros registros de pulpos (octópodos). La gran mayoría de los ammonoideos jurásicos pertenecen a los Ammonitina, caracterizados por la complejidad sutural, surgidos de los Psilocératidos. Los Ammonitina experimentaron una enorme radiación en el Jurásico medio, con formas que persistieron hasta el Cretácico. En el Jurásico comenzaron a abundar los equinodermos regulares e irregulares (que hicieron su aparición ahora). Son comunes también los crustáceos decápodos. En los ambientes jurásicos continentales se encuentran restos de conchóstracos, ostrácodos, insectos y moluscos dulcícolas. Entre los insectos se incorporaron los lepidópteros (polillas y mariposas). En ambientes continentales se reconocen moluscos de agua dulce, conchóstracos, ostrácodos e insectos. Los peces actinopterigios comenzaron su gran radiación que los llevó a ser dominantes en los océanos. Pero entre ellos, todavía predominaban los “holósteos” aunque hicieron su aparición los teleósteos, que son los peces modernos. Es en esta época que las primeras salamandras (urdelos) y lagartos (saurios) hicieron su aparición y los reptiles voladores (pterosaurios) evolucionaron y se diversificaron, apariciendo junto a los ranforincoideos con larga cola y dientes los más conocidos pterodactiloideos sin cola y estuche córneo sin dientes (preanunciando la condición que luego desarrolarían las aves). Es en el Jurásico, por otra parte, que se ubica el origen de las Aves (Archaeornithes, como el género Archaeopteryx con dientes en las mandíbulas, garras en las extremidades, huesos compactos y tamaño pequeño (como una paloma). En los ambientes marinos predominan los ictiosaurios y plesiosaurios. Aparecieron los cocodrilos que en sus comienzos habitaron a lo largo de cuerpos de agua continentales, pero un grupo retornó a los ambientes marinos. Se aceleró la evolución de los dinosaurios, con gran adaptación de los ornitisquios (estegosaúridos y ornitópodos). Los saurisquios comprenden tanto formas carnívoras como herbívoras. A los primeras pertenecen los terópodos, de andar bípedo (Allosaurus). Los saurópodos, herbívoros y cuadrúpedos, fueron las formas más gigantes que habitaron la tierra (Brontosaurus,Diplodocus, Brachiosaurus). Los mamíferos jurásicos continuaron siendo de pequeño tamaño (como una rata). Son conocidos por fragmentos de mandíbulas y dientes de Europa, América del Norte, Sudáfrica y China. Se han descubierto recientemente en estratos del Jurásico medio en la provincia del Chubut. La Formación Morrison del Jurásico tardío de Norteamérica contiene multituberculados, triconodontes y eupantotherios. Hacia el final también hicieron su aparición los marsupiales. La evolución de las plantas continuó con la aparición de las plantas con flores que dominan la vegetación de la Tierra actual, y en particular las angiospermas o plantas con flores y frutos, representadas inicialmente por las Archaeofructáceas, de ambientes pantanosos. En la flora continental, a latitudes bajas había una vegetación diversa que incluia helechos, equisetales, cycadales (Nilssonia), bennetittales (Otozamites, Ptilophyllum, Dyctyozamites), ginkgoales y coniferales. La mayoría de los helechos pertenecían a familias que viven en la actualidad y lo mismo puede decirse de las coniferales. A este tipo de vegetación hay asociados depósitos de carbón, que reflejan lluvias copiosas y vegetación abundante. Una excepción a este patrón se dio en la parte occidental de este cinturón tropical donde prevalecieron condiciones más desérticas y las floras reflejan esta situación, son muy empobrecidas y están representadas por pocas coniferales y helechos. Cycadeoidea, una típica benetital fósil, con cierto aspecto de angiosperma. Archaeofructus, la primera angiosperma. Archaeopteryx, una de las primeras aves (Archaeornithes). Conjunto de belemnoideos. Estegosaurio, dinosaurio jurásico con púas y espinas. El Cretácico (145-65 m.a.) Hacia el período Cretácico temprano, Laurasia se separó completamente de Gondwana, que ya estaba fragmentándose. Un mar continuo rodeaba los trópicos. Los niveles del mar fueron altos y la Tierra era cálida y húmeda. La paleogeografía sufrió rápidas y marcadas transformaciones debido a la evolución de las principales cuencas (Ártica, Tethys, Atlántica central) y otras que se formaron en ese tiempo (Cuenca del Atlántico sur e Indico). La cuenca Ártica se desconectó del Paleopacífico y se convirtió en un ambiente practicamente cerrado. En el Cretácico temprano el Atlántico central y el Tethys dividían al mundo en dos amplias regiones continentales: América del Norte-Eurasia formando un único continente y Gondwana en proceso de desintegración. Sin embargo, mientras que el Océano Atlántico se fue ensanchando, el Tethys se consumía a lo largo de una zona de subducción situada en el norte de la cuenca, a lo largo de los Cáucasos, de manera que casi había desaparecido a fines del Mesozoico. Esto se debió al movimiento de Africa hacia el este a la vez que giraba en sentido antihorario. Este movimiento finalizó en el Campaniano con el choque de la placa de Arabia contra el sur de Europa. La parte occidental del Tethys se consumió más tarde cuando a partir del Oligoceno, se originaron varias cuencas que integran el actual mar Mediterráneo. Paleoclimáticamente predominó un clima templado cálido, más húmedo que en el Jurásico debido a la ruptura de la Pangea y al incremento de la influencia marítima por los ascensos del nivel del mar. A fines del Cretácico en las altas latitudes se estableció una tendencia hacia el enfriamiento y la inestabilidad climática. En general los organismos cretácicos muestran la presencia de caracteres modernos. En el medio acuático se desarrolló un plancton muy variado, cuando foraminíferos alcanzaron gran desarrollo. En el Cretácico superior, los globigerínidos y globotruncánidos constituyeron, junto con cocolitos y radiolarios, los componentes predominantes de la creta. En la formación de estos depósitos participaron además esponjas silíceas, braquiópodos, bivalvos, cefalópodos y equinodermos. Los organismos bentónicos dominantes fueron los moluscos. Los ammmonoideos fueron los invertebrados más característicos de todos los mares, acompañados por belemnoideos, los extraños pelecípodos rudistas de valvas asimétricas y equinodermos irregulares. Los arrecifes alojaron abundantes corales y briozoarios asi como bivalvos especializados (Rudistas). Muchos bivalvos, surgidos en el Jurásico continuaron su diversificación en el Cretácico, mereciendo especial atención los ostreidos (Ostrea, Gryphea, Exogyra, Aetostreom), Inoceramus, Trigonia y formas afines. Los ammonoideos continuaron representando los moluscos más característicos y entre ellos, algunos adquirieron enroscamientos irregulares hasta rectilíneos. Otras tendencias evolutivas consistieron en la simplificación de la sutura y el aumento del tamaño de las conchillas. El número de familias fue declinando paulatinamente hasta que al final del período el grupo se extinguió. Los belemnoideos fueron frecuentes hasta el Cretácico tardío inclusive, aunque en el Senoniano iniciaron una rápida decadencia que los llevó a su extinción. De los restantes grupos de cefalópodos, sólo los nautiloideos tienen cierta representatividad en el Cretácico. En ambientes dulcícolas se reconocen numerosos ostrácodos, conchóstracos, bivalvos, gastrópodos y esponjas de características modernas. Los insectos constituyeron los invertebrados terrestres más característicos del Cretácico y habrían tenido una gran influencia en la radiación de las angiospermas, al hacer su aparición avispas, abejas y hormigas (Himenópteros). También hay registros de arácnidos y crustáceos decápodos de agua dulce. El Cretácico se caracterizó por la radiación evolutiva de los teleósteos que aun perdura. Entre los dipnoos continúa siendo frecuente el género Ceratodus. En el ambiente marino existieron los últimos ictiosaurios y plesiosaurios frecuentemente de grandes dimensiones. En el Cretácico superior abundan los mosasaurios, otro grupo de reptiles marnios. En la tierra firme, los dinosaurios continuaron diversificándose y ahora hicieron su aparición grupos notables como los abelisaurios carnívoros con cuernos y yelmos, los tiranosaurios y los ceratopsios o dinosaurios herbívoros con yelmos y cuernos. Los pterosaurios estuvieron representados por formas tales como Pteranodon, con más de 6 metros de envergadura, aunque hay registros de formas mucha más gigantescas, como Quetzalcoatlus, de 15 m de envergadura. A pesar de la extinción de los dinosaurios al final del Cretácico, esta crisis no afectaría a ciertos grupos como las tortugas y los cocodrilos, los que persisten hasta la actualidad. Las primeras serpientes (ofidios) aparecieron durante el Cretácico, aunque los grupos modernos ricos en especies surgirían de una radiación posterior. Hacia fines del período, numerosos grupos de mamíferos habían evolucionado. La mayoría de ellos eran pequeños, si bien una especie recientemente descubierta en China, Repenomamus giganticus, era suficientemente grande como para capturar y consumir dinosaurios jóvenes. En los ambienes terrestres, las aves se hicieron más comunes (con la evolución de las Enaliornithes y la aparición de las Neornithes o aves modernas) y los mamíferos aumentaron su número y variedad con los primeros registros de euterios o placentarios en el Cretácico inferior de China. Los multituberculados fueron los mamíferos que más proliferaron en el Cretácico, mostrando numerosas adaptaciones. Los Triconodonta se extinguieron en el Cretácico tardío, mientras que los Pantotheria lo hicieron en el Cretácico temprano, probablemente originando a los marsupiales e insectívoros. Ya se conocen ya en el Barremiano los primeros placentarios de hábitos arbóreos, los plesiadápidos. Al comienzo del Cretácico, las plantas con flores iniciaron su radiación en particular entre las Magnoliales y Nymphaeales, así como las primeras monocotiledóneas, lo que las condujo a convertirse en el elemento dominante en la tierra firme. Las floras prosiguieron con una preponderancia de gimnospermas y ya en el Cretácico inferior aparecen las angiospermas. Las angiospermas tienen un origen debatido. La morfología y diversidad de las angiospermas vivientes y el aspecto predominantemente tropical húmedo a cálido templado de las floras del Cretácico tardío-Paleógeno temprano, hacen sospechar que las angiospermas se originaron como formas arbóreas en un ambiente boscoso tropical desde donde se difundieron hacia áreas más frías y secas. Muy probablemente el origen de estas plantas sea gondwánico, dada la abundancia y diversidad de polen de angiospermas en el Cretácico inferior. En general las megafloras del Cretácico inferior de Patagonia, Sudáfrica y la India son bastante similares y uniformes. En su conjunto constituyen la flora de Ptilophyllum, caracterizada por su bennetittales y otras gimnospermas. Elasmosaurus, plesiosauruio marino cretácico. Mosasaurus, el terror de los mares cretácicos. Evolución de los ceratopcios, dinosaurios típicos del cretácico. Tyrannosaurus rex. Plesiadápidos, mamíferos arborícolas en la línea evolutiva de los primates. Archaeanthus, un fósil cretácico de angiosperma relacionada con las magnólidas. Como se describió antes, un asteroide causó una extinción en masa que tuvo lugar hacia finales del período Cretácico. En el mar, muchos organismos del plancton así como invertebrados del fondo marino se extinguieron. En la tierra firme, todas las especies que superaban los 25 kilogramos en el peso corporal aparentemente se extinguieron. Muchas especies de insectos desaparecieron, quizás debido a que se redujo violentamente el crecimiento de las plantas de las que se alimentaban, luego del impacto. Algunas especies sobrevivieron en las partes septentrionales de Norteamérica y Eurasia, áreas que no estuvieron expuestas a los fuegos devastadores que devoraron a la mayor parte de las regiones de bajas latitudes. Sitio de impacto del asteroide que produjo la segunda gran extinción masiva de la historia terrestre, en Chicxulub, Yucatán. Tras la caída del asteroide se levantaron hojas gigantescas de más de 1000 m de altura, se incineraron los bosques y se produjo un verdadero “inveierno nuclear” que destruyó numerosas cadenas tróficas marinas y terrestres. En la imagen aparece en negro la capa de iridio (abundante en los meteoritos) que separa el Cretácico (abajo) del Terciario (arriba). Es el límite de la gran extinción. Actividades 1. Explique las principales características paleoclimáticas y paleogeográficas de la era Mesozoica y su influencia sobre los procesos biológicos, evolutivos. ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= 2. Analice las característias de la vida marina en los tres períodos de la era Mesozoica. Indique cambios en la fauna de invertebrados y vertebrados. ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= 3. Explique las características de la flora y de la fauna continentales durante el período Triásico. Evalúe la importancia de las faunas de mamíferos de la época. ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= 4. Explique las características de la flora y la fauna continentales durante el período Jurásico. Analice las características del auge de los dinosaurios. ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= 5. Explique las características de la flora y la fauna continentales durante el período Cretácico. Preste especial atención a los comienzos de la fauna y flora modernas. ¿Con qué grupos de organismos se relacionan? ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= 6. Analice las características de los grupos que se diversificaron a finales del Triásico y que no resultaron afectados por esta gran extinción. Relacione con lo aseverado en el punto anterior. ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= 7. Explique el concepto de límite K-T y las características de la gran extinción de finales del Cretácico. Busque información adicional y clasifique las explicaciones provisionales acerca de este proceso. ¿Cuáles son las principales evidencias de la caída de un gran meteorito en el límite K-T? ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= 8. Analice las siguientes imágenes y descríbalas de acuerdo con lo estudiado en el módulo. ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= ================================================================= Foro ¿Por qué las tasas evolutivas difieren entre los grupos de organismos? Los fósiles pueden revelar información acerca de las tasas de cambio dentro de linajes particulares de organismos. El cambio evolutivo en un linaje puede incorrectamente aparecer como rápido si el registro fósil es muy incompleto, pero algunos cambios rápidos están bien documentados por excelentes series de fósiles. Las modificaciones en los ambientes físicos y biológicos son probablemente el estímulo del cambio evolutivo. Los organismos que viven en ambientes que están sujetos a variaciones, probablemente evolucionan más rápido que aquellos que viven en los ambientes relativamente estables. Cuando se produce un cambio climático, las áreas de distribución de algunos organismos pueden cambiar y otros organismos pueden enfrentarse con predadores o competidores distintos. De un modo similar, los predadores pueden cambiar en respuesta a los cambios en sus presas. En contraste, la morfología de los organismos que habitan en ambientes relativamente estables, a menudo cambian muy lentamente, si es que lo hacen. Por otra parte, más del 90% de las especies que han vivido están extintas. Las especies se han extinguido a través de la historia de la vida, pero las tasas de extinción han fluctuado drásticamente. Los grandes eventos de extinción, a menudo fueron seguidos por elevadas tasas de evolución, ya que los organismos sobrevivientes respondieron a los nuevos ambientes con una composición diferentes de predadores, presas y competidores. Algunos grupos han experimentado altas tasas de extinción mientras que otros estaban proliferando. Por ejemplo, entre los moluscos de la planicie costera atlántica de América del Norte, las especies con amplias zonas de distribución tenían menor probabilidad de resultar extintas durante los períodos normales (cuando ninguna extinción masiva tuvo lugar) que aquellas que presentaban pequeñas áreas de distribución. Sin embargo, durante la extinción en masa de fines del Cretácico, grupos de especies de moluscos estrechamente emparentadas con amplias áreas de distribución sobrevivieron mejor que los grupos con áreas pequeñas, aún si las especies individuales dentro del grupo tenían zonas de distribución pequeñas. Las dietas de los organismos también pueden influir en sus tasas de extinción. El registro fósil muestra siempre que la selección natural a menudo favorece un mayor tamaño de los carnívoros (animales que consumen carne). Sin embargo, los carnívoros mayores típicamente presentan dietas más especializadas que los carnívoros menores y los animales con dichas dietas especializadas pueden ser más vulnerables a la pérdida de su fuente de alimentos, que aquellas con dietas más generalizadas. Una posible consecuencia de carnívoros grandes y especializados es que se extinguirían con más probabilidad que las especies pequeñas con dietas más generales. Un impresionante registro fósil de sucesivos linajes de cánidos (perros, lobos y sus parientes) permitió a Blaire Van Valkenburgh y sus colaboradores usarlos para un método comparativo (véase para testear y confirmar esta hipótesis. Hacia fines del período Cretácico, las tasas de extinción sobre la tierra firme fueron mucho más elevadas entre los grandes vertebrados que entre los pequeños. Lo mismo ocurrió durante la época del Pleistoceno, cuando las tasas de extinción fueron más elevadas sólo entre los mamíferos y las aves más grandes. Durante algunas de las extinciones en masa, los organismos marinos fueron fuertemente perjudicados, aunque los terrestres sobrevivieron bien. Otras extinciones en masa afectaron a los organismos que vivían en ambos ambientes. Estas diferencias no son sorprendentes, dado que los cambios principales en la tierra y en los océanos no siempre concuerdan. En síntesis, las tasas de cambio evolutivo varían grandemente entre linajes de organismos. Los cambios en los ambientes físicos y biológicos típicamente inducen cambios evolutivos. Las tasas de extinción han también fluctuado drásticamente a lo largo de la historia evolutiva. ¿Por qué los cambios graduales y lentos dominan el registro fósil? ¿De qué manera las dietas de los organismos influyen en las tasas de extinción? Los científicos no se ponen de acuerdo acerca de la extinción. Si bien la mayoría adhiere a la hipótesis (con mucha evidencia) de la caída del asteroide, otros prefieren las erupciones volcánicas. ¿Usted qué piensa? Argumente. Módulo 6. La Era Cenozoica y la vida moderna Objetivos • Analizar las características de la era Cenozoica • Explicar las diferencias entre Paleógeno y Neógeno • Correlacionar los cambios continentales y climáticos durante la era Cenozoica • Analizar la progresión de la fauna de aves y mamíferos durante la era Cenozoica • Examinar las características faunísticas y florísticas del Paleógeno • Examinar las características faunísticas y florísticas del Neógeno • Analizar la evolución humana en sus primeras etapas y la progresión de industrias líticas • Discutir sobre la microevolución, la especiación y sus mecanismos Contenidos La era Cenozoica y sus etapas El Paleógeno Características florísticas y faunísticas del Paleoceno al Oligoceno Características florísticas y faunísticas del Mioceno al Plioceno La evolución de los mamíferos durante el Cuartario Evolución de los homínidos y glaciaciones Cuevas e industrias líticas De la crisis a la expansión cenozoica Tras la crisis cretácica, hace 65 m.a., los continentes y océanos adquierieron, básicamente, sus características actuales y, a partir del Terciario Tardío, desarrolló la circulación oceánica ahora dominante. Se produjeron frecuentes encuentros entre placas que motivaron la elevación definitiva de los principales cordones montañosos ahora existentes (Andes, Alpes, Himalayas), lo que fue acompañado por un intenso plutonismo y volcanismo, con extrusión de enormes masas de andesitas y basaltos, además de importantes mineralizaciones. El campo magnético terrestre fue muy variable y el clima mostró gran inestabilidad, debido a la formación de áreas glaciadas en las regiones polares. La era Cenozoica se divide en dos períodos, el Terciario y el Cuartario y se extiende desde los 65 m.a. hasta la actualidad. Tradicionalmente comprende siete épocas, las dos últimas del Cuartario: Paleoceno, Eoceno, Oligoceno, Mioceno, Plioceno, Pleistoceno y Holoceno o Reciente. A veces se acepta otra división útil desde el punto de vista geológico y biológico: Paleógeno (del Paleoceno al Oligoceno) y Neógeno (del Mioceno al Holoceno). Durante el Terciario inferior (Paleoceno y Eoceno), las condiciones climáticas eran bastante uniformes, con un clima tropical a subtropical, que permitió el desarrollo de pluviselvas y, en los polos, bosques templados. En esta época aparecieron todos los grupos u órdenes de mamíferos placentarios, incluidas las ballenas primitivas o basilosaurios; los murciélagos primitivos, como Icaronycteris; los; los condilartros, herbívoros con molares imperfectos y carnívoros gigantes como Andrewsarchus. Al final del Eoceno y durante el Oligoceno, las temperaturas se hicieron más frías y secas, y las selvas se redujeron mucho en su extensión, lo que produjo una gran extinción de la fauna arborícola. Al finalizar el Terciario, a partir de las épocas Mioceno y Plioceno, se expandieron las gramíneas o pastos en las sabanas y las praderas, acompañando el cambio climático a medida que los continentes chocaban (India con Asia, y más tarde Sudamérica con Norteamérica). Esto marcó la evolución de grandes herbívoros, como los rumiantes, los caballos y los rinocerontes. Mientras tanto, al cambiar las corrientes marinas, se formaron gruesos casquetes de hielo en ambos polos y se desarrolló la fauna marina con la aparición de los pingüinos y las focas. La era Cenozoica se halla caracterizada por una amplia radiación de mamíferos, aunque otros grupos también experimentaron cambios importantes. En el transcurso del período Terciario, al haberse extinguido los reptiles gigantes, como los dinosaurios, los mamíferos fueron ocupando todos los ambientes del planeta, y junto con ellos, las aves, descendientes de los dinosaurios. El mayor desarrollo de los placentarios, que incluyen al ser humano, se favoreció por el desarrollo de la placenta (ya iniciado en el Cretácico), que es un órgano que permite la nutrición del embrión dentro del útero materno. Las plantas con flores se diversificaron extensamente y se volvieron dominantes en los bosques mundiales, con la excepción de las regiones más frías. Las mutaciones de dos genes en un grupo de plantas permitió que utilizaran el N 2 directamente mediante la formación de simbiosis con algunas especies de bacterias fijadoras de nitrógeno. La evolución de estas simbiosis entre ciertas plantas cenozoicas y estas bacterias especializadas resultó en la primera “revolución verde” e incrementó radicalmente la cantidad de nitrógeno disponible para el crecimiento de las plantas terrestres. El Terciario (65-1,8 m.a.) El Terciario se caracterizó por los cambios eustáticos del nivel del mar, habiéndose reconocido unos siete superciclos y veintidós ciclos. En cada uno de ellos se produjo una elevación general lenta del nivel del mar seguida por una rápida caída del mismo. Las posiciones de los continentes se asemejabam a la actual, pero Australia todavía estaba unida a la Antártida y el océano Atlántico era mucho más estrecho. Australia comenzó a derivar hacia el norte al promediar el Terciario, y hace 20 millones de años atrás, casi había alcanzado su posición actual. El Terciario temprano fue un tiempo cálido y húmedo, durante el cual la distribución de numerosas plantas cambió latitudinalmente. Los trópicos probablemente eran demasiado cálidos para las selvas, y en cambio estaban tapizados con vegetación baja. Hacia mediados del Terciario, sin embargo, el clima de la Tierra se hizo considerablemente más seco y frío. El notable desarrollo alcanzado en el Terciario por los foraminíferos planctónicos, radiolarios, cocolitofóridos y otros integrantes del zoo- y fitoplancton, constituye una excelente base para la biozonación del sistema. Los Nummulites entre los foraminíferos aparecen en el Paleoceno y resultan buenos fósiles guías en la región del Tethys, hasta el final del Eoceno, cuando se extinguieron. Igual que los restantes foraminíferos grandes, eran característicos de aguas cálidas, en las que llegaron a ser tan abundantes que sus acumulaciones dieron lugar a las compactas rocas calcáreas utilizadas por los faraones egipcios para construir sus pirámides. Otros foraminíferos igualmente importantes fueron los assilinas, operculinas, ortofragminas, alveolinas y orbitolítidos cuya evolución, durante el Terciario, proporciona gran ayuda en las correlaciones bioestratigráficas. Las faunas de foraminíferos planctónicos contaron con numerosos géneros y especies de amplia distribución en todos los mares terciarios, si bien estuvieron limitados por la temperatura de las aguas, las corrientes oceánicas, la salinidad y la distancia a la costa. Desde el punto de vista de la correlación estratigráfica, estos foraminíferos sustituyeron a los amonites mesozoicos en cuanto a su empleo como fósiles guías, siendo además importantes indicadores paleoecológicos. Los radiolarios, a diferencia de los foraminíferos, son protozoos de esqueleto silíceo (la mayoría de ellos) o de sulfato de estroncio (radiolarios acantarios); planctónicos. En los ambientes marinos terciarios el fitoplancton volvió a resurgir después de la profunda crisis del final del Cretácico y paulatinamente, alcanza sus valores actuales. Los cocolitofóridos, silicoflagelados, dinoflagelados y diatomeas se hallaron entre los principales grupos representados. Otras algas, importantes por su función secretora de CO3Ca y formación de depósitos calcáreos fueron Lithothammion, las algas dasicladáceas, las codiáceas y las cianoficeas, estas últimas constructoras de rocas traverténicas. A comienzos de la era Cenozoica, las faunas de invertebrados eran similares a las actuales. De hecho, el comienzo del Terciario se destacó por la aparición de nuevos elementos faunísticos y la creciente importancia que tomaron otros presentes con anterioridad. No obstante, las asociaciones del Terciario antiguo aún conservan integrantes que, en sus características principales, recuerdan fuertemente a sus antecesoras mesozoicas. Los ambientes arrecifales se hallan muy difundidos en los mares cálidos del Terciario, principalmente durante el Paleógeno pero, a partir del Mioceno, se restringieron a la región ecuatorial. Los corales hexacorállidos y los briozoarios queilostomátidos alcanzaron un neto predominio sobre los demás órdenes. Es entre los vertebrados que los cambios evolutivos durante el período Terciario resultaron más rápidos. Las regiones litorales marinas albergaron gran cantidad de moluscos cuya distribución estuvo controlada, principalmente, por los factores físicos (temperatura y profundidad de las aguas). Algunos géneros de moluscos de amplia distribución en la era Mesozoica y que no llegaron a extinguirse al final de dicha era, persistieron durante todo el Terciario y aún habitan, en calidad de relictos, nuestros mares. Tal es el caso de Aporrhais, Neotrigonia y Nautilus.Los cefalópodos terciarios, con excepción de ciertos géneros de nautiloideos que fueron relativamente frecuentes en el Paleoceno, pertenecieron a formas con conchilla interna (sepias, calamares) o ausente (pulpos), tal como ocurre en la actualidad. Los bivalvos y gastrópodos fueron los moluscos dominantes en el Terciario. Entre los primeros, algunos géneros como Ostrea llegaron a desarrollar individuos con conchilla de extraordinario tamaño y grosor (en Patagonia los hay de hasta 0.50 m de largo y varios centímetros de espesor valvar), siendo llamativo el pequeño espacio interior que queda libre entre las valvas y que alojaba los órganos del animal. Los neogastrópodos también alcanzaron notables dimensiones y esbeltez, desarrollando algunos una gran ornamentación. La abundancia de crinoideos y equinoideos regulares e irregulares revela la importancia que adquirieron estos equinodermos en los mares terciarios. Los ambientes continentales (terrestres, límmicos, fluviales) contienen faunas de bivalvos y gastrópodos muy similares a los actualmente vivientes y en Europa, la región del Paratethys (alrededor del antiguo océano Tethys) incluyó cuencas aisladas o semiaisladas propicias para la rápida evolución de nuevos géneros y especies. En los ambientes continentales y marinos del Terciario también fueron frecuentes los artrópodos, principalmente los ostrácodos, los que se adaptaron a las más diferentes condiciones de salinidad. Los balánidos fueron abundantes al igual que los cangrejos. Los peces teleósteos alcanzaron notable diversificación en los diferentes ambientes acuáticos del Terciario. En los mares también fueron comunes los tiburones y las rayas. En África, Australia y América del Sur, los sedimentos límmicos terciarios contienen restos de peces coanados similares a los que ahora habitan esos mismos continentes. La mayor parte de los anfibios terciarios conocidos corresponden a los anuros y, en menor proporción a las salamandras. Entre los reptiles, los fósiles más comunes pertenecen a las tortugas, cocodrilos, lagartos y serpientes que como los anfibios, poseían características modernas. Las serpientes y lagartos experimentaron extensas radiaciones durante este período, así como las aves y los mamíferos. Tres oleadas de mamíferos se dispersaron desde Asia a América del Norte a través de un puente terrestre que había conectado intermitentemente los dos continentes durante los últimos 55 millones de años. En los ambientes continentales, las mejores biozonaciones se basan en las palinofloras, mientras que la evolución de las faunas de mamíferos sirve para diferenciar las “edades-mamífero”. Los placentarios en particular mostraron gran variedad y consistieron en insectívoros, primates, creodontes, roedores y ungulados primitivos (condilartros, uintaterios y otros). Los roedores, los marsupiales, los primates y los ungulados artiodáctilos aparecieron en América del Norte durante este tiempo. Pero los proboscideos, cetáceos, desmostilios y otros grupos hicieron su aparición en los continentes meridionales, más precisamente en África, mientras que Sudamérica que se mantuvo como isla hasta el Plioceno, desarrolló una fauna particular, de grupos de meridiungulados como los toxodontes, los litopternos y los xenungulados. Los primates verdaderos (que reemplazan a los plesiadápidos cretácicos) aparecen en el Paleoceno Medio de Montana (Estados Unidos). Su tronco lo constituye los lemuroideos, actualmente habitantes de Madagascar y la India, donde son arborícolas y nocturnos; su origen se hallaría en algún insectívoro del Cretácico Superior o Paleoceno, En el Eoceno se produjo la segunda gran radiación de los placentarios, apareciendo órdenes que aún viven. Entre los grupos arcaicos, de origen paleoceno o anterior, los multituberculados disminuyeron mucho y los marsupiales fueron raros, excepto en América del Sur; los creodontes (carnívoros) fueron dominantes en el Eoceno, los Taeniodonta (herbívoros) desaparecieron n al final de este período; los condilartros, en cambio, se hicieron muy abundantes. En suma, el Eoceno marcó la aparición de los adentados, cetáceos, sirénidos, grupos avanzados de artiodáctilos (como los tilópodos o camellos y los suiformes o cerdos y pecaríes), perisodáctilos (ungulados con dedos impares, como caballo, tapir, rinoceronte), carnívoros y murciélagos. Al final del Eoceno Temprano, los perisodáctilos ya estaban bien diferenciados en équidos, tapires, calicóteros, titanóteros y rinocerontes. En el Eoceno continuarían los primates lemuroideos y aparecieron los Tarsioideos. El origen de los monos platirrinos sudamericanos, más primitivos que los catarrinos de Asia y África, no está aclarado. • Los monos platirrinos se caracterizan por sus aberturas nasales y separadas entre sí y dirigidas hacia los costados, cola larga y dentición con tres premolares. • Los monos catarrinos tienen las aberturas nasales próximas entre sí y abiertas hacia adelante y abajo, cola corta o ausente, dos premolares. El límite Eoceno-Oligoceno coincide con un hiato importante en la secuencia faunística de los grandes mamíferos herbívoros terrestres y roedores. En Europa Occidental el mismo está evidenciado por la extinción de formas eocenas adaptadas a condiciones locales y su reemplazo, a partir del Oligoceno, por inmigrantes asiáticos. Las faunas oligocenas de mamíferos están bien documentadas en todos los continentes, excepto Antártida. En Australia los restos de este edad son escasos y corresponden a grupos que todavía habitan esa región. Los perisodáctilos ya cuentan con rinocerontes y tapires típicos. Los équidos aumentaron de estatura, haciéndose tridáctilos. Entre los artiodactilos aparecieron los cerdos y pecaríes. La mayoría de las familias de felinos actuales ya existían en el Oligoceno y los arctoideos se vieron incrementados por la aparición de los mustélidos (zorrino, comadreja, tejón, nutria, etc.) y otros grupos (mapaches, oso panda y afines). Posiblemente, ciertos dientes del Eoceno Superior de Burma, ya serían de Pongidae (la familia de los orangutanes) pero la evidencia más cierta de la existencia de este grupo es el Propliopithecus del Oligoceno Inferior de Fayúm. Pasando al Neógeno, En el Mioceno se incrementó la modernización de las faunas de mamíferos y un alto porcentaje de las familias de entonces aún tienen representantes vivientes. En el Mioceno Temprano aún existían cetáceos primitivos (arqueocétidos) pero al final ya estaban presentes todas las familias de ballenas actuales. También aparecieron los leones marinos y focas. Los cánidos muestran ya gran diferenciación y aparecen los osos verdaderos (úrsidos). Los primeros roedores miomorfos (ratas y ratones) también son del Plioceno. Los équidos adquirieron dientes de crecimiento continuo (hipsodontia) y su alimento principal ya no fue el follaje sino las gramineas de las praderas. En el Mioceno Temprano, los proboscideos (Dinotherium, Gomphotherium, Mastodon), llegaron a Europa, mientras que a la América del Norte lo hicieron en el Mioceno Tardío, cuando en Eurasia ya existía un elefante primitivo (Stegomastodon). Los Driopithecidae, un grupo de primates en camino a la evolución humana, vivieron entre el Oligoceno-Plioceno desde España y Alemania hasta Italia y sur de China, y desde Egipto hasta Sudáfrica. Muchas especies provinieron del Mioceno Medio y Plioceno de los Siwalicks (Norte de India), como Driopithecus chinjiensis (Mioceno Medio). Los félidos adquirieron gran diversidad, apareciendo el género Felis, mientras que otros carnívoros desarrollaron caninos superiores muy grandes, en forma de hoja de sable, como Machairodus que, en el Plioceno Temprano habitó América del Norte, Europa, Asia y África. Los prociónidos, inicialmente confinados a la América del Norte, a partir del Plioceno Tardío invadieron América del Sur, donde todavía habitan el coatí y formas afines. Otra rama emigró al Asia, donde ahora viven los osos panda. Al final del Plioceno los bóvidos pasaron de Eurasia, su probable lugar de origen, al África. El bisonte migró a la América del Norte donde se mantuvo exitosamente hasta el arribo de los primeros hombres blancos que casi lo extinguieron. En el Plioceno surgieron los Australopitécidos, con caracteres simios y humanos. Comprenden a los géneros Australopithecus, Plesianthropus y Paranthropus del África Oriental, cuya capacidad craneana oscila entre 450 cm3 y 650 cm3, y el Meganthropus de Java, conocido por una mandíbula de grandes dimensiones. Comos se mencionó, una característica de la mayoría de las faunas de mamíferos terciarios sudamericano, es su endemismo, lo que se atribuye al aislamiento que experimentó el continente en éstos tiempos, con abundancia de marsupiales y de meridiungulados. Muchos linajes de plantas con flores evolucionaron al hábito herbáceo (no leñoso) y formaron praderas que se distribuyeron en grandes extensiones de la Tierra. La megaflora terciaria se caracterizó por el predominio de las angiospermas sobre las gimnospermas, si bien durante el Paleógeno las gimnospermas aún fueron importantes, debido al clima templado y uniforme y que facilitó que la flora templada de sequoia en el Eoceno se extendiera hasta Siberia y Groenlandia. En las regiones tropicales abundaron las palmeras, mientras cerca delas costas crecía Rhizocarpus. Esta flora templada hasta tropical persistió en el Oligoceno y en la región báltica, la resina de las coníferas atrapó a numerosos invertebrados, a los que ahora se los puede observar incluidos en trozos de ámbar disperso en las playas. El Cuartario o Cuaternario (1,8 m.a. a la actualidad) El período geológico actual es el Cuartario, subdividido en dos épocas, el Pleistoceno y el Holoceno (también conocido como Reciente). Durante el segundo período de la era Cenozoica, el Cuartario, hubo cuatro momentos en que los glaciares avanzaron sobre la Tierra, con el lógico descenso de temperatura y correspondientes cambios en la fauna y flora. Justamente, el límite Terciario-Cuaternario (Plioceno-Pleistoceno) coincidió con una brusca disminución de la temperatura, que se tradujo en un importante reemplazo entre las faunas de foraminíferos y radiolarios, ocurriendo algo similar con las floras. Sin embargo, este límite está menos definido que el CretácicoTerciario y algunos estratígrafos opinan que el Terciario debería ser extendido hasta nuestros días. La última glaciación ocurrió hace unos 10.000 años, y para muchos, la época actual tan sólo un período interglaciario (entre dos glaciares) a la espera de una nueva edad del hielo. La Beringia, un puente continental, permitió el intercambio faunístico entre Asia y Norteamérica también durante el Plioceno. En este período relativamente breve, de unos 2.5 m.a., se desarrolló el linaje del ser humano moderno. Sin embargo, en la edad del hielo vivieron también algunos de los mamíferos más grandes de todos los tiempos, como el oso de las cavernas, el rinoceronte lanudo, el megaterio o perezoso gigante y matutes y mastodontes, parientes del elefante. El Pleistoceno fue un tiempo de drástico enfriamiento y fluctuaciones climáticas. Durante cuatro “edades del hielo” principales y otras 20 menores, los glaciares masivos se distribuyeron a través de los continentes y la distribución de las poblaciones de plantas y animales cambió hacia el ecuador. Los últimos de estos glaciares retrocedieron desde las latitudes templadas hace menos de 15.000 años. Los organismos todavía se están ajustando a estos cambios. Muchas comunidades ecológicas de latitudes altas han ocupado sus distribuciones actuales desde no hace más que unos pocos miles de años. Resulta interesante que un número relativamente bajo de especies se extinguieron durante estas fluctuaciones climáticas. El Pleistoceno fue el tiempo de la evolución y radiación de los homínidos, que resultó en la especie Homo sapiens–humanos modernos (véase Sección 33.5). Muchas especies de aves y mamíferos gigantes se extinguieron en Australia y en América cuando H. sapiens llegó a dichos continentes hace, respectivamente, unos 40.000 y 15.000 años. Estas extinciones fueron probablemente el resultado de la caza por los humanos, aunque la evidencia existente no termina de convencer a todos los paleontólogos. Los mamuts lanudos sobrevivieron en la isla Wrangel de Siberia tan recientemente como hace 4.000 años, mucho después de que los cazadores los exterminaron del continente. Los mamuts pudieron haber mantenido su ambiente de estepa seca en la isla, alimentándose de los musgos de la tundra y reciclando el nitrógeno, mientas que el ambiente continental se convirtió en una tundra húmeda. las cuevas a las glaciaciones. Sobre cuevas y glaciaciones, e industrias culturales humanas La palabra deriva del bajo latín cava; y ésta, del latín clásico cavas, hueco. Con este nombre se designan cavernas, grutas y concavidades del subsuelo, naturales o artificiales. Las cuevas revelan la evolución geológica y la contextura de la corteza terrestre. A través de sus sedimentos y de las caprichosas formaciones de su interior, se pueden estudiar los acontecimientos geológicos y climáticos. Por otra parte, las cuevas son testigos de la vida orgánica que habita en los espacios subterráneos, insectos, reptiles y plantas, en lo que se refiere al presente. Respecto al pasado y como refugio de animales, tienen gran importancia en paleontología y, por supuesto en arqueología y antropología, ya que fueron los primeros hogares humanos. En las capas geológicas se han acumulado restos óseos o deyecciones (coprolitos) de diversas especies de animales, algunos extintos, incluido el linaje humano, que ocuparon estas cuevas en diferentes momentos. Igualmente, el polen de las plantas de cualquier estrato nos documenta sobre la flora de estas épocas remotas. Muchas de estas cuevas son verdaderas estaciones prehistóricas o arqueológicas, ya que el hombre desde los primeros tiempos de su existencia aprovechó estas formaciones naturales.. El estudio de las cuevas permite analizar entonces la forma de vida de los hombres de la Prehistoria, su modo de alimentación, las presas que cazaban, si complementaban su dieta con otros recursos, la forma en que se protegían del clima y de los animales, pero también sus representaciones simbólicas expresadas en el arte rupestre. Las sucesivas capas que cubren el suelo de las cuevas se formaron por la sedimentación de la roca junto con la litificación de los restos orgánicos e inorgánicos, incluidos los huesos y las herramientas utilizadas por los seres humanos. Junto a los sedimentos geológicos, se encuentran las capas arqueológicas formadas por las cenizas de los sucesivos hogares que marcaron las ocupaciones. Entre ellas, se mezclan desechos de cocina, útiles, armas e incluso huesos humanos. Muchas veces los objetos o desechos eran arrojados frente a la entrada, y su acumulación ha dado lugar a los tell o montículos. El estudio de los estratos, tanto artificiales (restos de ocupación), como naturales (geológicos), evidencian las etapas de la humanidad en relación con sus circunstancias. Los fenómenos geológicos y climáticos que sorprendieron a los ocupantes, rocas desprendidas, inundaciones, entradas bloqueadas, etc., tienen gran valor cronológico, no sólo al fechar un determinado nivel o yacimiento, sino al autentificar los restos de nuestros antepasados, especialmente las manifestaciones artísticas. Durante mucho tiempo se creyó que el hábitat en cuevas naturales fue común y único del Paleolítico, sinónimo de época del hombre de las cuevas y desde los tiempos clásicos se denominaron trogloditas a los hombres que las habitaron y a su género de vida. Hoy, los avances prehistóricos han desmentido, en parte, estas concepciones y se sabe que indistintamente se ocuparon las cuevas, los abrigos rocosos y las chozas o tiendas al aire libre, tanto en las glaciaciones como en tiempos más benignos. Sin embargo, son de gran valía estas rústicas habitaciones fáciles y cómodas en la vida nómada de los primitivos, y son numerosísimas en todo el mundo las cuevas que podrían citarse como estaciones prehistóricas, por ejemplo L'ermitage (Vienne) y otras muy diversas, con industrias paleolíticas, neolíticas, etc., e incluso con niveles que llegan hasta la época romana incluida. En las etapas históricas, y aun en la actualidad, se sigue aprovechando la ventaja de las cuevas naturales como vivienda, sobre todo en algunos países cálidos. Para comprender las características de las culturas humanas, conviene realizar un breve análisis de las “culturas” animales. En un reciente trabajo sobre el problema de la transmisión social de la conducta adquirida, Galef (1976) analizó y revisó múltiples ejemplos de procesos seudoculturales o protoculturales entre los animales, afirmando que «la interacción intraespecífica que resulta de la transmisión de pautas adquiridas de conducta de un individuo a otro dentro de una población, es un modo relativamente común e importante de adaptación en organismos vertebrados, tanto primates como no primates», Quedan, por supuesto, eliminadas aquellas pautas de conducta para cuya ontogenia es la condición necesaria la interacción social pero que sin: embargo forman parte de la «conducta específica de la especie»: en ellas no hay relación de aprendizaje puesto que en este caso el congénere no es otra cosa que la condición ,de aparición, el estímulo desencadenador de la pauta en un momento que suele ser crítico para su aparición. Y se excluye también así lo que McDougall llamó «inducción simpática», que nada tiene que ver con la imitación. Tinbergen (1964) lo define diciendo que son reacciones de hacer los mismos movimientos que el congénere, pero «compelidos a hacerlos movidos por una reacción puramente interna». Es el caso de muchas reacciones de huida, por ejemplo. Indudablemente este criterio puede resultar en la práctica un tanto ambiguo, fundamentalmente porque la apariencia de «conducta específica de especie» puede ocultar una pauta transmitida a partir de un aprendizaje cuyo origen se desconoce. Pero la eliminación virtual de casos interesantes de verdadera transmisión social se compensa con la seguridad de que los casos que entran en consideración están fundamentadamente elegidos. En este contexto, tanto las orcas y otras especies de delfínidos, incluido el delfín nariz de botella y el delfín común, tienen sociedades complejas y culturas definidas con su propio “idioma” y sus propias pautas de conducta, explotación de fuentes alimentarias e interacción dentro de la población. Las poblaciones de delfines y orcas tienen culturas caracterizadas por patrones alimentarios, sonidos específicos y pautas de conducta. Por su parte, entre los primates existen claros indicios de protoculturas, al menos entre los grandes monos (orangutanes, gorilas, chimpancés). Desde el principio, la familia de los homínidos poseía claramente lo que se denominan culturas rudimentarias, es decir, las especies en la línea evolutiva del ser humano tenían tradiciones sencillas que se habían transmitido de generación en generación, no heredando los genes de sus padres, sino aprendiendo como hacían las cosas. La diferencia fundamental entre culturas rudimentarias y culturas plenamente desarrolladas es de carácter cuantitativo. Simios y monos cuentan con escasas tradiciones, pero los humanos las tienen innumerables. Cuando nuestros antepasados traspasaron el umbral del despegue cultural, realizaron un avance tan decisivo como la transición de la energía a la materia o de los aminoácidos a la proteína viva. Resulta interesante analizar el sucesivo avance y retroceso de los hielos a lo largo de la era Cenozoica, porque esto se relaciona con los cambios climáticos clave para la evolución, así como la disponibilidad de recursos alimenticios. La glaciación más importante tuvo lugar desde fines del Plioceno y durante el Pleistoceno, con varios períodos de expansión y retracción de los glaciares. El avance glaciar determinó en el interior de los continentes, a partir de los trópicos, sequías y pasaje de las pluviselvas o los bosques a las sabanas y estepas, y en las áreas más próximas a los círculos polares, la extensión masiva del hielo y la formación de tundras o, directamente, de desiertos glaciales. Las cuatro fases de avance glaciar para el Viejo Mundo son las denominadas: Günz, Mindel, Riss y Würm (los nombres dados por los europeos) o sus equivalentes para Norteamérica: Nebraska, Kansas, Illinois y Wisconsin. Entre cada dos glaciaciones existió el correspondiente periodo Interglacial. El de la transición de Illinois a Wisconsin se llama Sangamon y culminó hace unos 70.000 años, muy anterior, por consiguiente, de la posible penetración del hombre en América, pero sí probablemente correspondiente a la primera expansión del hombre por Asia y Oceanía a partir de África. Por su parte, se denomina Posglacial a la serie de fases del último interglacial (que justamente corresponde a la época Holoceno, en la que vivimos actualmente). Como se vio en la Teórica de poblamiento, la glaciación Wisconsin comenzó con dos fases de avance de hielos que son las Alton y Farmdale hace unos 50.000 años. Además, dentro del extenso subcontinente de Norteamérica quedaron evidenciados varios focos de glaciación: los de Groenlania, Labrador, Keewtin y Piedmont. Tras un interestadio, continúa el avance Iowa, el interestadio Peoría, el avance Tazewell (ca. 20.000 a.C.), el interestadio Brady y el avance Cary. Este último empezó hace unos 13.600 años a.C. y se detuvo hacia los 12.000 años a.C. Durante el avance Tazewell, la masa de hielos concentrada en los focos del nordeste de Norteamérica se unió, al parecer, con lo que se desbordó desde el foco de Pledmont, situado en las Montañas Rocallosas del territorio canadiense, constituyendo una superficie helada extensa que llegaba desde la costa del Pacífico a la del Atlántico y al sur hasta San Louis, Missouri. Muy poco después, sin embargo, durante la retirada Haven (ca. 15.000 a.C.) el pasillo entre el foco de PIedmont y el resto de los hielos quedaría de nuevo expedito, permitiendo el paso de manadas de animales y bandas humanas desde Alaska. El interestadio Two Creeks o de retroceso de hielos estuvo situado entre dos estadios de avance de los mismos, que son el Mankato y el Valders. Entre 9.000 y 8.500 a.C. se inició la deglaciación o retirada definitiva de los hielos (es decir, con la fase final de la glaciación Wisconsin, los hielos comenzaron a retroceder): Corresponde justamente al Posglacial que iniciará la Revolución del Mesolítico y Neolítico (véase en períodos culturales). En el 8.000 a.C. se inició el Holoceno o Reciente (aunque otros autores prefieren la fecha anterior de 10.000 a.C.), y también se caracteriza por oscilaciones climáticas. El altitermal u optimun climático se dio hace justamente en 5.000 a.C. y duró hasta 2.000 a.C. La variabilidad térmica durante el Wisconsin influiría en la flora y fauna americanas creando obligados desplazamientos de algunas especies, e incluso la aparición y desaparición de otras. Período Cuartario Avance glaciar Años a.C. Época Pleistoceno 2.500.000 a 10.000 Glaciar Günz (Nebraska) x Primer interglacial Glaciar Mindel (Kansas) x Segundo interglacial Glaciar Riz (Illinois) x Último Interglacial 100.000 a 70.000 (Sangamon) Glaciar Würm x (Wisconsin) (sólo se indican las fases Fases Alton y Farmdale 50.000 para Norteamérica) Intermedio 30.000 Farmdale/Iowa Avance Iowa 25.000 Interestadio Peoria 23.000 Avance Tazewell 20.000 Interestadio Brady 14.000 Avance Cary 13.500 Estadio Mankato 11.000 Interestadio Twoo Creeks 10.000 Estadio Walders 9.000 Época Holoceno o Reciente Posglacial (Anatermal) 8.000 a 6.000 Posglacial (Altitermal) 5.000 a 2.500 Posglacial (Meditermal) 2.500 al presente Es importante tener en cuenta que las tres primeras fases de avances glaciarios corresponden desde el punto de vista cultural al Paleolítico inferior, mientras que la última fase (Würm) ocupa gran parte del Paleolítico medio y superior. La evolución humana se produce durante las etapas de las glaciaciones y se relaciona directamente con diferentes especies de homínidos que evolucionan durante este perìodo. La división tradicional es en Paleolítico o edad de la Piedra antigua y Neolìtico o Edad de la piedra Nueva sobre la base de herramientas e industrias culturales. El Paleolítico, a su vez, se divide en tres subperìodos: Inferio, Medio y Superior. A continuación se presenta la secuencia más aceptada que corresponde al Paleolítico inferior. Paleolítico inferior. (2,5 m.a.) 700.000 a 500.000 a.C. Corresponde a las industrias básicas del tallado, como la de guijarros y lascas radiales (unifacies y bifacies). a) Industria Pebble Cultures u Olduvayense (durante la glaciación Günz) Elementos característicos: guijarros (choppers o choppings tools) de la “pebble culture”. Es el modelo más básico y antiguo de tallado. Se trata de gujarros (choppers) o lascas sin retocar, que están talladas por una sola cara (unifaciales) e igualmente cantos tallados. Aparece en África hace 2,5 millones de año Corresponden a los Australopitecinos, básicamente, pero también podría asociarse a los primeros Homo, como Homo rudolfensis. b) Industria Kafuense (durante la glaciación Günz) Elementos característicos: guijarros mal tallados (kafuenses) de África meridional. Corresponden al Homo habilis. c) Industria Abbevilliense (Chelense) (en el Interglacial) Elementos característicos: bifacial abbevilliense (primeras herramientas trabajadas). Corresponden al Homo ergaster y probablemente al Homo erectus. d) Industria Clactoniense (probablemente en el segundo Interglacial, aunque podría ubicarse también antes o después del Achelense según otros autores, es decir, está en discusión). Descubrimiento del fuego. Elementos característicos: lascas y guijarros clactonienses de trabajo básico. Corresponden al Homo erectus y al Homo pekinensis (y linajes relacionados). e) Industria Achelense (durante la glaciación Mindel). Elementos característicos: amigdaloides y puntas achelenses trabajadas con técnicas más eficientes, es decir, bifacies. Corresponden al Homo antecesor. f) Piso Micoquiense/Tayaciense (durante la glaciación Riss) Elementos característicos: refinamiento de técnicas y lascas micoquienses más trabajadas. Corresponden al Homo georgicus y seguramente al Homo heidelbergenesis (eurántropo). El Paleolítico medio transcurre entre 500.000 a 150.000 a.C. y se relaciona básicamente con el hombre de Neanderthal. En el Paleolítico medio se produjo una gran innovación. Los seres humanos aprendieron a sacar del mismo volumen de piedra mayor cantidad de filo. El Paleolítico medio cubre un periodo comprendido entre los 200.000 y los 35.000 años. La industria es más diversificada que en el Paleolítico inferior, y se caracteriza por una alta proporción de industria sobre lascas: raederas, raspadores y denticulados. El Paleolítico medio está caracterizado por una forma de tallar la piedra denominada técnica Levallois. Esta técnica se caracteriza por efectuar una preparación del núcleo, que determina el tipo de lascas que se van a obtener a partir de él. Éste método que aparece hace aproximadamente 200.000 años, consiste en preparar el núcleo con una serie de extracciones que modelan la superficie del núcleo, de forma que posteriormente se obtienen lascas de una forma determinada. a) Industria Llevalloisiense (tercer Interglacial). Elementos característicos: lascas de trabajo perfeccionado fragmentos de roca levalloisiense con plano de percusión pulimentado. A la técnica para conseguir esto se la denomina Levallois. Primero se talla un canto al que se le da una forma estándar. A partir de ahí se van extrayendo las lascas con un percutor blando intermedio. Corresponden al Homo neanderthalensis. En Europa este modo de talla está íntimamente relacionado con los neandertales ya que con ellos aparece y con ellos desaparece. b) Industria Musteriense (durante la glaciación Würm). Elementos característicos: puntas, rascadores y cuchillos musterienses. Evidencias de enterramientos, símbolos religiosos e incluso, canibalismo ritual. Corresponden al Homo neanderthalensis. El Paleolítico superior cubre un periodo comprendido entre los 40.000 años y los 10-9.000 años, y aparece asociado a los humanos modernos. Esta época corresponde a la segunda parte de la última glaciación, el Würm, y termina con el calentamiento que se produce en el período Alleröd. El Paleolítico superior se caracteriza por el perfeccionamiento de la elaboración de útiles con la técnica laminar, y el aumento de la proporción de raspadores y buriles. Con el Paleolítico superior aparecen nuevos útiles y diseños que responden, con su variedad de formas, a distintos trabajos muy especializados: limpieza, curtido, preparación y perforado de pieles; trabajo y modificación del hueso; para grabar figuras en las paredes de las cuevas... El Paleolítico superior se extiende de 150.000 a 13.000 a.C. Se caracteriza por el desarrollo del arte Paleolítico y las representaciones figurativas. Arte rupestre y petroglifos, arte mobiliar y venus (esculturas). Tradicionalmente, de modo convencional y según un orden cronológico, se duele dividir al Paleolítico superior en tres períodos, llamados Auriñaciense (por Aurignac, Francia), Solutrense (por Solutré, Francia) y Magdaleniense (por La Madelaine, Francia), en los que se aprecia un perfeccionamiento de las primeras industrias humanas, en particular con relación a las armas de defensa y ataque. Pero en este análisis se incluye el Gravetiense. Los instrumentos del Paleolítico superior europeo variaron mucho dependiendo del lugar y la época. Algunos arqueólogos piensan que esto refleja diferentes grupos de individuos, mientras que otros creen que los distintos instrumentos reflejan diferentes escuelas o culturas. La cronología del Paleolítico superior de Europa occidental fue definida por H. Breuil y D. Peyrony a partir de las industrias líticas procedentes de abrigos y cuevas del sudoeste francés, reconociendo cinco tipos principales de industria: a) Industria Chatelperroniense (45.000-35.000 a.C.). Se solapa cronológicamente con el Auriñaciense y fue desarrollado por los últimos Neanderthales. b) Industria Auriñaciense-Perigordiense (durante el Würm) (35.000-25.000 a.C). Elementos característicos: se caracteriza por la presencia por primera vez de talla de tipo laminar así como puntas y rascadores auriñacienses de tipo lateral. Corresponde a razas básicas del Homo sapiens y probablemente también a Homo sapiens idaltu. c) Industria Gravetiense (27.000-19.000 a.C.). Se caracteriza por largas puntas afiladas, buriles, y azagayas de hueso. Elementos característicos: largas puntas afiladas finamente retocadas y buriles y azagayas de hueso (gravetienses). Corresponde al Homo sapiens sapiens (Combe Chapelle). l) Industria Solutrense (21.000 a 16.000 a.C.) Sólo se extiende por la Península Ibérica y Francia, y coincide con el momento de máxima extensión de los hielos, hacia 18.000 a.C. Elementos característicos: sierras y puntas de lanza de hojas muy delgadas y con retoques profundos llamadas “hojas de laurel”, así como cuchillos solutrenses. Corresponde al Homo sapiens sapiens (Cro-magnon). l) Industria Magdalaniense (16.000-12.000 años).. Elementos característicos: se hacen importantes los arpones, anzuelos y propulsores, las hachas de mano y otros elementos más perfeccionados, con huso del hueso magdaleniense. Corresponde al Homo sapiens sapiens (Cro-magnon). Actividades 1. Establezca la periodizaciòn de la era Cenozoica y distinga entre Paleógeno y Neógeno. =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 2. Delimite las características principales de las épocas del período Terciario. Tenga en cuenta principalmente los cambios en la fauna de mamíferos. =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 3. Explique la secuencia de faunas durante la era Cenozoica. Distinga entre las faunas y floras marinas y continentales. =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 4. Analice la secuencia evolutiva de los mamíferos durante la era Cenozoica. ¿Por qué se denomina a la era Cenozoica la edad de los mamíferos? Justifique. =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 5. Analice la importancia de las glaciaciones y de las cuevas en el estudio de los fósiles humanos de la era Cenozoica. =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 6. Explique el desarrollo cultural de los homínidos y asocie las diferentes industrias culturales con las etapas de la glaciación. =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 7. Analice las características del aislamiento y del intercambio faunístico en diferentes etapas del Terciario. Utilice como referencia el siguiente mapa. =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== 8. Explique el siguiente texto de acuerdo con lo leído en el módulo. Aunque los agentes del cambio evolutivo están operando en la actualidad como lo han estado haciendo desde que la vida apareció en la Tierra, los cambios principales están siendo conducidos por un incremento decisivo en la población humana de la Tierra. La predación (caza) por los humanos ha causado la extinción de numerosas mamíferos grandes en el pasado reciente, y continúa haciéndolo en la actualidad. Los humanos también están cambiando los ambientes físicos y biológicos al alterar en forma contundente la vegetación terrestre, al convertir los bosques y los pastizales en cultivos y pasturas. Deliberadamente, o sin advertirlo, estamos trasladando a miles de especies alrededor del globo, reduciendo el provincialismo de la biota terrestre que comenzó durante la era Mesozoica. Los humanos también se han encargado de la evolución de ciertas especies por medio de la selección artificial y la biotecnología. Como vimos en el Capítulo 16, modernos métodos moleculares permiten modificar a los organismos mediante el traslado de genes incluso entre especies lejanamente emparentadas. En resumen, los humanos se han vuelto un agente dominante del cambio evolutivo. Según cómo nos desempeñemos, nuestra influencia masiva afectará poderosamente el futuro de la vida en la Tierra. =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== =============================================================== Foro La selección sexual durante el Terciario Charles Darwin propuso que la selección sexual era responsable de la evolución de caracteres brillantes y conspicuos como las grandes astas de los ciervos y las grandes plumas de la cola de los pavos reales. Estos rasgos están especialmente exagerados en especies en las que los individuos de un sexo—en general los machos—compiten por las oportunidades de apareamiento con individuos del sexo opuesto—usualmente las hembras. Los caracteres exagerados evolucionan si les confieren una ventaja, tanto en la competencia entre machos por el acceso a las hembras, o mediante la estimulación y atracción de las hembras discriminatorias. Además de inducir los conspicuos ornamentos de los machos como la astas y el plumaje, la evidencia reciente sugiere que la selección sexual igualmente podría aumentar la tasa de formación de nuevas especies. La evidencia para dicho efecto de la selección sexual proviene de la comparación del número de especies que se encuentran en clados hermanos (grupos de parentesco), esto es, aquellos que comparten un ancestro común. Debido al ancestro común, los clados hermanos fueron evolucionando independientemente uno del otro en el mismo lapso. La tasa con la que se han formado las especies en los dos clados se puede estimar mediante la comparación del número de especies que poseen en la actualidad. Las aves con sistemas de apareamiento promiscuo ofrecen uno de los mejores ejemplos de ambos de los mencionados efectos de la selección sexual. En muchas especies, los machos se reúnen en territorios de cortejo y las hembras se dirigen a ellos para elegir con quien copular. Después de la cópula, las hembras de numerosas especies construyen sus nidos, ponen sus huevos y crían a su descendencia sin ninguna ayuda de los machos. Y, en la mayoría de estas especies, los machos han desarrollado plumajes brillantes y usualmente ornamentos tales como las colas con largas plumas. En contraste, en las especies monógamas, que forman parejas y comparten responsabilidades en la crianza de la descendencia, los individuos de ambos sexos tienden a exhibir un plumaje monótono y a parecerse entre sí. Como ejemplo de este fenómeno, durante la era Cenozoica se expandieron todos los grupos de aves conocidos, en especial los Passeriformes (pàjaros) y los Apodiformes (vencejos y colibríes). En la actualidad existen cerca de 320 especies de colibríes en América, todas las cuales son promiscuas. Sin embargo, sólo hay 103 especies de vencejos (clado hermano de los colibríes), a pesar de que los vencejos son de amplia distribución en el mundo. Los vencejos macho y hembra forman pareja monógamas y ambos se parecen. ¿Por qué la selección sexual estimula la divergencia de un linaje en muchas especies? Una razón probable es que las mutaciones aleatorias resultan en la elaboración de diferentes plumajes en diferentes zonas de distribución de una especie. Por los tanto las poblaciones locales de una especie puede desarrollar un único patrón de plumaje en los machos, cada uno de los cuales es favorecido por la selección sexual en dicha área. Las hembras pueden no responder positivamente a un macho inmigrante cuyo plumaje difiere del patrón hacia las cuales se sienten normalmente atraídas. El origen de nuevas especies —la división y la divergencia de un único linaje en distintas especies— es uno de los fenómenos más importantes en las ciencias biológicas. Charles Darwin reconoció esta preeminencia cuando escogió el título de su libro más famoso. Auque sin el conocimiento que aportaría la genética moderna, Darwin se hallaba observando principalmente las consecuencias de la especiación, y no las causas que subyacen a ella, y esto también lo reconoció él. Todavía estamos en la búsqueda de respuestas a muchos interrogantes acerca de la especiación, un proceso al que Darwin se refirió como “el misterio de los misterios”. • • • ¿De qué manera influye la selección sexual en la aparición de nuevas especies? ¿Por qué no siempre surgen nuevas especies? ¿Por qué se considera que junto con los mamíferos las aves se expandieron durante la era Cenozoica? ¿Cómo surgen nuevas especies? La distinción entre especies de homínidos durante la era Cenozoica y la expansión continua de los mamíferos en general nos lleva a preguntarnos acerca de la especiación. Queda claro que no todos los cambios evolutivos resultan en una nueva especie. Un único linaje puede cambiar a lo largo del tiempo sin originar una nueva especie. Si los cambios evolutivos resultan en una especie que se divide en dos o más especies hijas, entonces se ha producido la especiación. Posteriormente, dentro de cada pool de genes aislados, las frecuencias de los alelos y genotipos pueden cambiar como resultado de la acción de los mecanismos evolutivos. Si dos poblaciones se hallan separadas una de otra, y se acumulan suficientes diferencias en su estructura genética durante el período de aislamiento, entonces las dos poblaciones podrían no intercambiar genes cuando se juntan nuevamente. Como veremos, la cantidad de diferencia genética que se requiere para prevenir el intercambio genético es altamente variable. Por lo tanto, la especiación requiere que el flujo de genes dentro de una población cuyos miembros intercambian genes se interrumpa. El flujo de genes puede ser interrumpido en dos formas principales, cada una de las cuales caracteriza un modo de especiación. La especiación alopátrica requiere el aislamiento casi completo La especiación que resulta cuando una población es dividida por una barrera física se conoce como especiación alopátrica (allo, “diferente”; patris, “patria”) o especiación geográfica. Se considera que la especiación alopátrica es la forma dominante de especiación entre la mayoría de los grupos de organismos. La barrera física que divide la gama de una especie puede ser un cuerpo de agua, una cadena montañosa, para organismos terrestres, o la tierra seca para organismos acuáticos. Las barreras se pueden formar durante la deriva continental, el nivel de los mares se eleva, los glaciares avanzan y retroceden, y los climas cambian. Estos procesos continúan formando barreras físicas en la actualidad. Las poblaciones separadas por tales barreras son frecuentemente, aunque no siempre, inicialmente grandes. Estas poblaciones evolucionan diferencias por una variedad de razones, incluida la deriva genética, si bien especialmente porque los ambientes en los que viven son, o se vuelven, diferentes. La especiación alopátrica puede originarse también cuando algunos miembros de una población cruzan una barrera existente y encuentran otra población aislada nueva. Las 14 especies de pinzones del archipiélago de las Galápagos, a 1.000 kilómetros de la costa de Ecuador, se generaron por especiación alopátrica. Los pinzones de Darwin (como son llamados usualmente, porque Darwin fue el primer científico quien los estudiara) surgieron en las Galápagos a partir de una única especie sudamericana que colonizó las islas. En la actualidad, las 14 especies de los pinzones de Darwin difieren llamativamente de su pariente costero más cercano. Las islas del archipiélago de las Galápagos se hallan lo suficientemente distantes entre sí para que los pinzones se dispersen en ellas con escasa frecuencia. Además, las condiciones ambientales difieren entre las islas. Algunas son relativamente llanas y áridas, y otras presentan laderas montañosas forestadas. Las poblaciones de pinzones en las diferentes islas se han diferenciado lo suficiente a lo largo de millones de años que cuando los inmigrantes ocasionales llegan a otras islas, o bien no se cruzan con los residentes locales o, si lo hacen, la descendencia resultante frecuentemente no sobrevive tan bien como la generada por las parejas compuestas por los residentes de la isla. La distinción genética y la cohesión de las diferentes especies de pinzones se mantienen. Varias de las 800 especies de moscas de la fruta del género Drosophila en las islas de Hawai están restringidas a una única isla. Sabemos que estas especies son descendientes de nuevas poblaciones fundadas por individuos que se dispersan entre las islas porque el pariente más cercano de una especie en una isla es frecuentemente una especie de una isla vecina más que otra de la misma isla. Los biólogos que han estudiado los cromosomas de la especie de Drosophila de “alas pintadas” creen que la especiación en este grupo de moscas ha resultado en, por lo menos, 45 de estos eventos fundadores. La eficacia de una barrera física para prevenir el flujo de genes depende del tamaño y la movilidad de las especies en cuestión. Una autopista de ocho carriles que es, prácticamente, una barrera infranqueable para un caracol, no representa ninguna barrera para una mariposa o un pájaro. Las poblaciones de plantas polinizadas por el viento se hallan aisladas por la máxima distancia a la que el polen puede volar con el viento, si bien las plantas individuales están aisladas efectivamente por distancias mucho menores. Entre las plantas polinizadas por animales, la amplitud de la barrera es la distancia que pueden recorrer los animales mientras transportan los granos de polen o las semillas. Incluso animales con gran capacidad de dispersión son frecuentemente renuentes a cruzar trechos angostos de hábitats inadecuados. Para los animales que no pueden nadar o volar, las brechas estrechas y llenas de agua pueden ser barreras efectivas. Sin embargo, el flujo de genes puede, a veces, ser interrumpido incluso en ausencia de barreras físicas. La especiación simpátrica se produce en ausencia de barreras físicas A pesar de que el aislamiento físico es usualmente requerido para la especiación, bajo ciertas circunstancias la especiación puede darse sin ella. La separación de un pool génico sin aislamiento físico se denomina especiación simpátrica (sym, “junto con”). Pero si la especiación es usualmente un proceso gradual, ¿cómo puede desarrollarse un aislamiento reproductivo cuando los individuos tienen frecuentes oportunidades de aparearse entre sí? Lo que se requiere es alguna forma de selección disruptiva en la que ciertos genotipos presentan una gran adaptación para uno u otro recurso, como se describió en el caso de los pinzones cascanueces de vientre negro. La selección disruptiva en los pinzones cascanueces no ha resultado todavía en la formación de dos especies, si bien la especiación simpátrica a través de la selección disruptiva puede estar dándose en la mosca de la fruta (Rhagoletis pomonella) en el estado de Nueva York. Hasta mediados de la década del 1800, la mosca de la fruta Rhagoletis cortejó, se apareó y depositó sus huevos solo en los frutos del majuelo (especie de espinillo). Las larvas aprendieron a reconocer el olor del majuelo a medida que se alimentaban de las frutas. Cuando emergían de sus pupas como adultos, utilizaban esta pista para localizar otras plantas de majuelo en las cuales aparearse y poner sus huevos. Hace aproximadamente 150 años, grandes huertas comerciales fueron cultivadas en el valle del río Hudson. Los manzanos están estrechamente relacionados con los majuelos. Unas pocas hembras de Rhagoletis pusieron sus huevos en manzanas, quizás por error. Estas larvas no crecieron tan bien como las de la fruta del majuelo, si bien muchas sobrevivieron. Estas larvas reconocieron el olor de las manzanas, de modo que cuando emergieron como adultos, buscaron manzanos, donde se aparearon con otras moscas criadas en manzanas. Hoy en día hay dos grupos de Rhagoletis pomonella en el valle del río Hudson que podrían estar en camino de convertirse en especies diferentes. Una se alimenta principalmente de la fruta del majuelo, la otra de las manzanas. Las dos especies incipientes están aisladas reproductivamente en forma parcial porque se aparean principalmente con individuos criados en la misma fruta y porque emergen de la pupa en diferentes momentos del año. Además las moscas que se alimentan de manzanas han evolucionado de manera que ahora crecen en estas frutas más rápidamente de lo que lo hacían originalmente. La especiación simpátrica por medio del aislamiento ecológico (en diferentes recursos), como ocurre en Rhagoletis pomonella, puede estar ampliamente difundida entre los insectos, muchos de los cuales se alimentan de una única especie de planta, pero la forma más común de especiación simpátrica es la poliploidía, la producción dentro de un individuo de juegos duplicados de cromosomas. La poliploidía puede surgir ya sea por duplicación de cromosomas en una especie única (autopoliploidía) o por la combinación de cromosomas de dos especies diferentes (alopoliploidía). Un individuo autopoliploide se origina cuando (por ejemplo) las células que son normalmente diploides (con dos juegos de cromosomas) accidentalmente duplican sus cromosomas, resultando en un individuo tetraploide (con cuatro juegos de cromosomas). Las plantas tetraploides y diploides de la misma especie están reproductivamente aisladas debido a que su descendencia tetraploide híbrida es usualmente estéril. No pueden producir gametos viables porque sus cromosomas no hacen una sinapsis correcta durante la meiosis. Por lo tanto, una planta tetraploide no puede producir descendencia viable al cruzarse con un individuo diploide–pero puede hacerlo si se autofecunda o se cruza con otro individuo tetraploide. Por lo tanto la poliploidía puede resultar en el completo aislamiento reproductivo en dos generaciones–una importante excepción a la regla general de que la especiación es un proceso gradual. Los alopoliploides pueden también ser producidos cuando los individuos de dos especies diferentes (aunque estrechamente emparentadas) se entrecruzan, o hibridan. Los alopoliploides son frecuentemente fértiles debido a que cada uno de los cromosomas tiene un compañero prácticamente idéntico con quien aparearse durante la meiosis. La especiación por poliploidía ha sido importante en la evolución de las plantas. Los botánicos estiman que aproximadamente el 70% de las especies de plantas con flor y el 95% de los helechos son poliploides. La mayoría de ellos surgió como resultado de la hibridación entre dos especies, seguido de autofecundación. Nuevas especies surgen por medio de poliploidía mucho más fácilmente entre plantas que entre animales debido a que las plantas de muchas especies pueden autofecundarse. Además, si surge la poliploidía en varios descendientes de un único progenitor, los miembros de la progenie pueden fertilizarse los unos a los otros. Cuán fácilmente puede la alopoliploidía producir nuevas especies queda ilustrado en el ejemplo de los salsifies (Tragopogon), parientes de los girasoles. Los salsifies son malezas que proliferan en baldíos alrededor de las ciudades. La gente las ha diseminado inadvertidamente alrededor del mundo desde sus lugares de origen en Eurasia. Tres especies diploides de salsifies–Tragopon porrifolius, T. pratensis, y T. dubius– fueron introducidos en América del Norte tempranamente en el siglo veinte. Dos híbridos tetraploides–T. mirus y T. miscellus–entre las tres especies diploides originales fueron descubiertas primeramente en 1950. Los híbridos se han diseminado desde su descubrimiento y hoy están más ampliamente distribuidas que sus parientes diploides. Los estudios sobre su material genético muestran que ambos híbridos de salsifies se han formado más de una vez. Algunas poblaciones de T. miscellus–un híbrido de T. pratensis y T. dubius– tienen el cromosoma del cloroplasto de T. pratensis; otras poblaciones tienen el genoma del cloroplasto de T. dubius. Las diferencias genéticas entre entre las poblaciones locales de T. miscellus muestran que este alopoliploide se formó independientemente en, por lo menos, 21 oportunidades. T. mirus, un híbrido de T. porrifolius y T. dubius, se formó 12 veces. Los científicos raramente conocen bien las fechas y las localizaciones de la formación de especies. T. porrifolius tiene preferencia por lugares húmedos y sombreados; T. dubius prefiere lugares secos y soleados. T. mirus, sin embargo, puede crecer en ambientes parcialmente sombreados donde ninguno de sus progenitores prospera. El éxito de este nuevo híbrido de salsifies ilustra el modo en que tantas plantas con flor pudieron haberse originado como poliploides. La poliploidía, como hemos visto, puede resultar en una nueva especie que está completamente aislada reproductivamente de otras emparentadas durante dos generaciones, si bien la mayoría de las poblaciones separadas por una barrera física se convierte en reproductivamente aisladas muy lentamente. Veamos de qué manera, el aislamiento reproductivo puede establecerse una vez que dos poblaciones se han separado. Una vez que se establece una barrera para el flujo de genes, las poblaciones separadas pueden divergir genéticamente a través de la acción de los mecanismos evolutivos. Durante varias generaciones, se pueden acumular diferencias que reducen la probabilidad de que los miembros de dos poblaciones se apareen y produzcan una descendencia viable. De esta manera, el aislamiento reproductivo puede evolucionar como un subproducto incidental de los cambios genéticos de las poblaciones alopátricas. El aislamiento reproductivo parcial ha evolucionado de esta manera en cepas de Phlox drummondii. En 1835, Thomas Drummond, por quien esta planta a sido nombrada, recolectó semillas en Texas y las distribuyó a viveros en Europa. Durante los siguientes 80 años, estos viveros establecieron más de 200 cepas reproductivas reales de esta planta, que diferían en el tamaño y color de la flor y la forma de crecimiento de la planta. Los horticultores de estas plantas no las seleccionaron directamente por la incompatibilidad reproductiva entre cepas, pero en experimentos posteriores, en los que se midieron y compararon las tasas de fertilización de las flores de varias cepas por el polen de otras, se descubrió que la compatibilidad reproductiva entre cepas se había reducido de 14 a 50 por ciento, dependiendo de los casos. Sin embargo, el aislamiento geográfico no necesariamente lleva al aislamiento reproductivo, debido a que la divergencia genética no provoca la aparición del aislamiento reproductivo como un subproducto. Por lo tanto las poblaciones que han sido aisladas entre sí por millones de años pueden aún ser reproductivamente compatibles. Por ejemplo, el plátano americano y el plátano europeo han estado geográficamente aislados entre sí por, al menos, 20 millones de años. A pesar de ello, son morfológicamente muy similares, y pueden formar híbridos fértiles, a pesar de que nunca han tenido la oportunidad de hacerlo en la naturaleza. La incompatibilidad puede aparecer por muchos mecanismos diferentes, pero es útil agruparlos en dos tipos principales: barreras reproductivas precigóticas y postcigóticas. • • • • ¿Qué diferencias encuentra entre especiación simpátrica, alopátrica y parapátrica (por poliploidía)? ¿Cuál de estos tres mecanismos podría asociar con la evolución humana? ¿Podría explicar por qué una barrera efectiva al flujo de genes para una especie puede no aislar eficientemente a otra especie? ¿De qué manera la especiación por medio de la poliploidía se produce en dos generaciones? Epílogo Una ojeada retrospectiva La vida unicelular procariota y eucariota (progresión e hitos evolutivos) 1. Hace 4.000 m.a., una vez que el ancestro universal (LUCA) logró estabilizar su población inicial, el primer gran cambio fue la aprición del código genético y la síntesis de proteína, con el desarrollo del ADN o ádido desoxirribonucleico. En esta primera etapa, un primer grupo de organismos podrían haber sido los nanobios (si se confirma definitivamente su existencia). Como derivado del mundo de ARN y las ribozimas, surgieron los ribosomas, fundamentales para la síntesis de proteínas. 2. Hace 3.800 a 3.500 m.a., en un ambiente anaerobio (sin oxígeno) y un mar ácido, los factores de la evolución comenzaron a actuar y se produjo el surgimiento de organismos especializados, tanto quimiolitótrofoso como fotolitótrofos que podían elaborar sus propios nutrientes. Estos primeros organismos fueron bacterias verdaderas (imperio Eubacteria), en especial las cloroflexas o bacterias verdes no azufrosas y las clorobias o bacterias verdes del azufre. Ya estos organismos contaban con membranas plasmáticas bicapa anque tenían nucleoides circulares como todos los procariotas. Además, estos organismos cuentan en sus células con bacterioclorofilas (diferentes a la clorofila de las plantas y las algas) y no pueden utilizar el oxígeno (que, además, faltaba hace 3.500 m.a. en el planeta) aunque metabolizan el azufre y el sulfuro de hidrógeno. Un poco más tarde hicieron su aparición las bacterias púrpura, con un modo de nutrición fotoorganótrofo, como ya que pudieron aprovechan los materiales orgánicos complejos si bien continuaron utilizando la luz solar como fuente energética. 3. En una etapa final, hace 3000 a 2.800 m.a., hicieron su aparición las primeras arqueas, metanógenas o productoras de metano. Al englobar bacterias, surgieron los verdaderos cromosomas lineales con ADN e histonas de los eucariotas. Igualmente, las arqueas ya presentaban antecedentes de proteínas contráctiles, similares a las del citosesqueleto. 4. Hace 2.500 m.a. surgieron las cianobacterias que comenzaron a oxigenar el planeta ya que contaban con clorofila verdadera y cadena respiratoria. 5. Entre 1.900 y 2.100 m.a. comenzó la radiación de los eucariotas unicelulares, con núcleo celular y mitocondrias, tal vez representados con la primera proliferación de euglenas y las amebas. Más tarde surgieron los dinoflagelados, entre 1.700 a 1.500 m.a.. A esta época se la conoce como la del big bang o explosión de los eucariotas unicelulares, cuando estos organismos se hicieron abundantes en los ecosistemas marinos. 6. Entre los 1.500 y 1.200 m.a., la explosión de vida continuó en los mares de plataforma y se sumaron los primeross organismos pluricelulares, comenzando con las algas rojas y las algas verdes pluricelulares y carbonáticas, y probablemente los primeros hongos y los coanoflagelados, ancestros unicelulares de los animales. Entre los 900 a 800 m.a. se produjo una migración continental hacia los polos, y el oxígeno acumulado por la acción de las algas fue importante que la Tierra se congeló por completo, durante la glaciación Varangiana. A esta época de la Tierra congelada se la llama Snowball Earth (del inglés tierra bola de nieve). Al derreterise el hielo continental y aumentar el nivel de sales del mar, comenzó una segunda gran radiación de eucariotas pluricelulares, esta vez liderada por los animales, entre ellos los cnidarios y el grupo fósil de los vendozoos, a partir de los 600 m.a.. 7. Al comenzar la era Paleozoica, se inicia una segunda explosión de vida (animal) desde los 540 m.a., y surgen casi todos los ancestros de los fila o tipos animales, incluidos los bilaterales (con simetraí bilateral), existentes en la actualidad. Las novedades más importantes fueron los caparazones y los esqueletos (formados por diferentes sales presentes en el medio) y también se desarrollaron los ganglios y cadenas nerviosas. Hace 500 m.a., aparecieron representantes de la fauna moderna, como los gasterópodos y los peces, y se desarrollaron las vértebras y el esqueleto óseo. 8. Un paso adicional e importante fue la conquista del medio aeroterrestre, iniciada en el Ordovícico, hace 450 m.a. cuando se formó la capa de ozono en la atmósfera y se llegó al nivel de Pasteur (21 %) hacia finales del Silúrico. Entonces proliferaron los musgos, luego los helechos, ambos gracias a la actividad de los líquenes y los hongos micorrízicos y surgieron los esporangios (formadores de esporas), gametangios (formadores de gametos) y los tejidos vasculares. Enseguida llegó la invasión de los animales, que tuvo su mayor pico en el Devónico, con los anfibios y los insectos, con la adquisición de adaptaciones como las cubiertas y los pulmones (vertebrados) o las tráqueas (artrópodos). También por entonces surgieron las gimnospermas y las primera semillas. 9. Al final de la era Mesozoica, llegaron los mamíferos, los cocodrilos, las primeras aves y los dinosaurios. Con estos animales se impuso el huevo cleidoico (con cáscara), la placenta de los mamíferos, el control de la temperatura corporal y la tabicación completa del corazón. 10. Al final de la era Mesozoica, llegaron las angiospermas o plantas con flores y frutos y los insectos polinizadores y ya el mundo vegetal nunca más volvió a ser el que se conocía antes, proliferando los bosques y las sellvas. En la última etapa evolutiva, durante la era Cenozoica, los mamíferos evolucionaron en las selvas y los bosques que se formaron en esta época y se produjo la hominización, con el gran desarrollo de la corteza cerebral y la postura bípeda características del linaje humano. Yacimientos fosilíferos y colecciones de fósiles en el país Los yacimientos fosilíferos más importantes del país se encuentran en el sur argentino, aunque también otros períodos, como la era Paleozoica, se encuentran mejor representados en el norte, y la era Cenozoica en la provincia de Buenos Aires. Algunas de las colecciones de fósiles destacadas se encuentran en los siguientes: Museo Paleontológico Egidio Feruglio de la ciudad de Trelew, en Santa Cruz, Museo Provincial de Ciencias Naturales y Oceanográfico en la ciudad de Puerto Madryn, Santa Cruz, Museo de Ciencias Naturales Fray José Osvaldo Bersia, en Villa Allende, Córdoba, Museo Provincial de Ciencias Naturales Bartolomé Mitre y Museo de Mineralogía y Geología “Dr. Alfredo Stelzner”, ambos en Córdoba capital, Museo Municipal de Ciencias Naturales “Carlos Darwin” de Punta Alta, el afamado Museo de Ciencias Naturales de La Plata, Museo de Ciencias Naturales Florentino Ameghino de Las Flores, provincia de Buenos Aires, Museo de Ciencias Naturales "Domingo F. Sarmiento", de Mendoza, Museo de Ciencias Naturales "Dr. Angel Gallardo", en Rosario, Santa Fe, Museo Paleontológico y Petrolero Astra, en Comodoro Rivadavia, Río Negro, Museo de Ciencias Naturales de Chubut, Museo de Ciencias Naturales Bernardino Rivadavia de la Ciudad Autónoma de Buenos Aires y muchos otros. Proyectos y taller Visita a un museo de Ciencias naturales Lo que ofrecen los museos Muchas ciudades de nuestro país cuentan con grandes museos que exhiben importantes colecciones de fósiles. La ciudad de La Plata, capital de nuestra provincia, cuenta con uno de los museos más prestigiosos del mundo. Los fósiles son el principal testimonio del conocimiento que los científicos poseen sobre las eras geológicas de la Tierra. En general, existen fósiles característicos, los fósiles guía, típicos de los diferentes períodos geológicos. Esto encuentra explicación en la superposición de fenómenos de extinción y origen sucesivos. Características del museo general de La Plata El “museo general de La Plata” fue creado por un decreto del Gobierno de la provincia de Buenos Aires el 19 de septiembre de 1884, en base a un proyecto del coleccionista Francisco Pascasio Moreno (1852-1919) quien se desempeñó como Director hasta 1906. Las primeras colecciones provinieron del Museo Antropológico de Buenos Aires creado en 1877, con patrimonio donado por el propio Moreno. Dependiente del Ministerio de Obras Públicas provincial, abrió sus puertas al público en 1888. En sus primeros años, los objetivos de la Institución estuvieron ligados, por un lado, al montaje de exhibiciones de historia natural, antropología, arqueología y bellas artes para contribuir a la educación general de los habitantes de la provincia. Por otro lado, al desarrollo de expediciones en el interior del país, con el doble propósito de contar con colecciones de estudio e identificar recursos naturales pasibles de explotación económica por parte del Estado y los particulares. Hacia mediados de la década de 1890, con el apoyo financiero del Ministerio de Relaciones Exteriores, la Institución se comprometió con la cuestión de límites con Chile, poniendo a disposición del Gobierno Nacional los recursos humanos y materiales para realizar expediciones topográficas a lo largo de la Cordillera de los Andes. En ese entonces se crearon también las cinco secciones principales del Museo: Antropología, Geología, Zoología, Paleontología y Botánica. Estuvieron a cargo de estudiosos extranjeros, y tenían como función clasificar las colecciones para exhibición y estudio, realizar expediciones y publicar trabajos científicos en la Revista y los Anales del Museo de La Plata, creados en 1890. En 1906, el Museo, dirigido por Samuel Lafone Quevedo (1835-1920), pasó a depender administrativamente de la Universidad Nacional de La Plata, agregando a las funciones originales de exhibición pública e investigación, la de formación académica, bajo la denominación general de Instituto del Museo-Facultad de Ciencias Naturales. Desde ese entonces, el Museo y la Facultad funcionaron de manera imbricada compartiendo espacios En el caso de las actividades ligadas con el Museo, durante la década de 1920 se retomó el plan fundacional de las exploraciones científicas por el interior del país y se continuó con el envío de colecciones duplicadas de Ciencias Naturales a los museos escolares. Se modernizó el edificio, laboratorios, depósitos, salas y exhibiciones, editándose por primera vez una Guía para visitar el Museo de La Plata (1927). También se construyó la biblioteca en el lugar que aún ocupa. Se adquirieron importantes colecciones, como la serie arqueológica del Noroeste que perteneciera a Benjamín Muniz Barreto (1876-1933). Las mejoras edilicias y la creación de nuevos salones de exhibición continuaron durante la gestión de Joaquín Frenguelli (1883-1958), que se extendió en el período 1934-1946. Se inició posteriormente, en líneas generales, la etapa de los Directores-Decanos de la Facultad de Ciencias Naturales y Museo, caracterizada por gestiones breves marcadas por sucesivas intervenciones, en sintonía con los recurrentes cambios en el contexto sociopolítico del país. Hacia 1948, los Departamentos Científicos fueron reorganizados internamente con el nombre de Divisiones, agregándose a la estructura Institucional en las décadas siguientes numerosos Centros, Institutos y Laboratorios de investigación de la Facultad de Ciencias Naturales y Museo, que continúan funcionando hasta la actualidad. En esos años se crearon también nuevas orientaciones en la carrera, como la de Geoquímica y Ecología, y otras se dividieron en especialidades, como la de Paleontología en los campos de Vertebrados e Invertebrados, y la de Antropología, en los campos de Arqueología, Antropología Cultural y Antropología Biológica. Fuente: Procedimiento http://www.museo.fcnym.unlp.edu.ar/articulo/2012/10/25/taller 1. Organice la visita de a dos compañeros. Elijan un período de la historia terrestre y un grupo representativo cuyos orígenes y evolución desean conocer. 2. Con los datos que obtenga con sus alumnos, pídale que preparen fichas sobre las características, las relaciones de parentesco, la forma de vida, la dieta, la defensas, la distribución paleogeográfica, etcétera, de cada organismos. 3. Puede organizar la visita en grupos, y que cada uno elabore un informe para la sección estudiada. Las cinco secciones principales del Museo son: Antropología, Geología, Zoología, Paleontología y Botánica. Pero dentro de cada una puede observar diferentes períodos. En síntesis, el museo de la Plata cuenta con exhibiciones de aves, de mamíferos, de mamíferos fósiles, de dinosaurios e incluso acerca de la evolución humana y de pueblos originarios de Argentina y del mundo. Informe de la visita • Mencione y busque con los alumnos fósiles guía para algunos períodos geológicos. Trilobites: Cámbrico y Ordovícico. Arqueociatios: Cámbrico. Ostracodermos: Ordovícico y Silúrico. Placodermos: Devónico. Anfibios primitivos (laberintodontos): Carbonífero. Reptiles pareisauros: Pérmico. Tecodontos y phitosaurios: Triásico. Geosaurios y estegosaurios: Jurásico. Tiranosaurios y ceratopsios: Cretácico. Análisis 1. Luego de recorrer las salas donde se encuentran fósiles de distintos taxones, elabore una breve síntesis del resultado de la visita. 2. Pida a los alumnos que elijan un fósil y preparen una ficha modelo. Podrían responder las siguientes preguntas: ¿De qué lugar provienen? ¿A qué era o período pertenecen? ¿Cómo se determinó su antigüedad? ¿Cuáles son sus nombres científicos (recuerden la nomenclatura binomial) y a qué hacen referencia dichos nombres? (características particulares, descubridor, zona geográfica, etcétera). Si se realizó reconstrucción, ¿es veraz o responde a algún tipo de especulación? Exposiciones y museos en el aula laboratorio de Ciencias El aprendizaje de la ciencia requiere observar, experimentar y tocar. Y no hay nada mejor para ello que contar con un pequeño museo para el aula o la escuela. La creación del Museo Escolar debe incluir la organización y exposición de colecciones de objetos, documentos y otros materiales para facilitar a la comunidad escolar su exhibición y estudio. A través del Museo Escolar se motiva la conservación y protección del patrimonio cultural, científico, histórico y artístico y se refuerza la identidad comunitaria de la Institución, La creación del Museo Escolar contempla la organización y exposición de colecciones de objetos, documentos y otros que posea la Escuela. Supone el desarrollo de una actividad colectiva en donde participan profesores, alumnos y directivos en el diseño y montaje y que asuman en su totalidad el manejo. El Museo Escolar puede concentrar variedad de contenidos a partir del tipo de objetos que definan su centro de interés y servir de apoyo didáctico para el desarrollo de temas en distintas áreas curriculares. A través del Museo Escolar se motiva la conservación y protección del patrimonio cultural, científico, histórico y artístico desde el ámbito escolar y se refuerza la identidad comunitaria de la Institución. Inicialmente, busque la colaboración de personas e instituciones con amplia experiencia en montaje Museos. Esto debe ser sólo transitorio. La creación del Museo Escolar implica que este se convierta en un centro cultural y de investigación para la comunidad educativa y desarrolle diferentes actividades como son: • Generar investigaciones • Realizar talleres, conferencias y eventos • Divulgar la presencia del Museo Escolar en la localidad • Apoyar las actividades de educación formal y no formal de la Escuela. • Estimular el desarrollo de actitudes, conocimientos y habilidades • Integrar el Museo al Proyecto Educativo Institucional con base en los centros de interés de las áreas básicas del currículo. La organización de la colección del Museo escolar * La colección es la razón de ser del Museo. Si la Escuela no tiene una colección, se la puede iniciar. A partir de objetos naturales como rocas o plantas y aparatos y objetos del mundo de la fisica, etc., surgen ideas de acopio de elementos coleccionables, que aporten conocimiento a los integrantes del grupo escolar. Es vital complementar el registro e inventario de los objetos y elementos con que cuenta la Escuela y que tengan interés científico, artístico, histórico, documental, cultural, institucional, etc. El inventario de la colección se lleva a cabo en una ficha individual que contiene toda la información referente al objeto. Su desarrollo requiere de tiempo e investigación. Aparte del inventario, es conveniente realizar una clasificación u ordenamiento de los objetos a partir de características comunes, con la finalidad de proceder a su catalogación. Todos los objetos deben ir enumerados y marcados en alguna parte que no sea visible. Se debe promover, en esta etapa el interés, a que los estudiantes donen a la Escuela objetos que sirvan para ampliar la colección. Ficha técnica Número: Nombre del objeto Procedencia..................... Época……. Medida: Largo............................. Ancho......... Alto.............Peso......... Otros Técnica Estado del objeto Bueno....................... Regular ................... Malo Estado de Integridad Completo............................ Incompleto.......................... Unido Descripción Forma de adquisición Por donación............................ Por compra.................... Por canje Observaciones Si la la escuela no tiene una colección, se la puede iniciar. En esta oportunidad, se sugiere la colección de seres vivos acuáticos, sus huellas y señales o bien muestras de suelo, sedimento lacustre o fluvial y piedritas del agua. ¿Cómo organizar la colección? Materiales: • Cajas de zapato • Frascos de vidrio con tapa perforada • Botellas de plástico transparente o blanco • Papel de filtro • • • • • • • • • • • • • Aserrín Vidrios de reloj Pinzas Etiquetas Planchas de telgopor Chinches de colores Clips Tablas de madera Tarros Peceras o cajas de vidrio rectangular Microscopio Portabobjetos y cubreobjeto Papel de diario, hojas de cartulina y papel secante Procedimiento Recolección y reunión del material 1. En la salida al campo, busque ejemplares de plantas acuáticas, como repollitos de agua, saetas y camalotes. Colóquelas en bolsitas de plástico semiabiertas para conservarlas y llevarlas al museo escolar. 2. Capture pequeños organismos acuáticos, por ejemplo caracoles de agua dulce, camarones y escarabajos y ubíquenlos en sendos frascos con tapas perforadas para permitir el ingreso de oxígeno. 3. Tome piedras y otros elementos que indiquen la presencia de los organismos acuáticos. 4. Saque fotografías en el lugar visitado para armar carteles explicativos. 5. Obenga muestras de agua del ambiente natural o de un charco en el lugar. B. Colección (colección de preparados, herbario, caja entomológica, acuaterrario) 6. Prepare una infusión con la muestra en un frasco de vidrio con hojas de lechuga. Déjelo por una semana y luego obtengan preparados microscópicos para la observación de protozoos y algas. 7. Obtenga ejemplares de hojas y ramas de plantas más grandes y colóquenlas entre hojas secantes para armar el herbario. La idea de la conservación de las plantas secas para estudio surgió en el siglo XVI y a Lucca Ghini (1519-1556) se le considera el fundador del primer herbario con finalidad de científica. El herbario es muy importante en la investigación botánica. La confección del herbario se inicia en la salida de campo, al seleciconar una muestra de la planta en la que estén presentes los caracteres más destacados. El fragmento del vegetal es prensado entre papel secante (y entre diarios) para desecarlo lo más rápidamente posible. Después se dispone en un pliego de papel, al que se añade la etiqueta con todos los datos tomados en el campo en el momento de la recolección (localidad, altitud, coordenadas geográficas, ecología, fecha y nombre del recolector). Finalmente se traslada al lugar que le corresponde para su exhibición. 8. Si captura organismos vivos, colocarlos en los frascos de vidrio etiquetados o ubicarlos en el acuaterrario. Para ello usar la pecera, llenarla en parte con tierra y en parte con agua y acomodar los peces, insectos y plantas acuáticas que hayan capturado. 9. Prepare una caja entomológica para presentar la colección de insectos y otros organismos que hayan encontrado muertos. Para ello utilicen cajas rígidas de cartón, en la que se coloca en su interior, una plancha de telgopor de dos o tres cm de espesor, sobre la que se clavarán los insectos ya preparados. La caja entomológica debe reunir ciertas características como: - Una atmósfera cerrada como para colocar algún producto que proteja a los insectos de otros muy pequeños que los consumen, pero que no sea hermética (telgopor o plástico), Debe permitir la salida de la humedad al exterior. - Conservarse en un lugar oscuro, la luz altera los colores de los insectos. En el exterior, la caja debe llevar un rótulo que contenga información sobre el autor, cada insecto llevará, en lo posible un pequeño rótulo con los siguientes datos: 1Orden 2Nombre común 3- Autor 4-Lugar y fecha de recolección. Los rótulos irán clavados en el mismo alfiler en que va montado el insecto, por debajo de éste. En el interior de la caja, a los insectos se los puede acomodar sin un criterio en particular o en grupos de acuerdo al orden, a tamaños, dependiendo de sus hábitos alimenticios. Coloque bolitas de naftalina para proteger la colección 10. Para animales más grandes que no estén vivos, usen soluciones conservadoras, por ejemplo: - FORMOL al 10 % - ALCOHOL 70º Estos liquidos se preparan de manera sencilla, por disolución de las soluciones originales adquiribles en los comercios. Para la obtención del formol al 10%, vuelquen en un recipiente tres medidas de agua y una medida de formol comercial (al 40% ). Para obtener alcohol de 70ª, mezclen una medida de agua con tres medidas de alcohol 96ª (alcohol etílico de venta en farmacias). TIPO DE ANIMAL A CONSERVAR LIQUIDO CONSERVADOR De cuerpo blando: Peces, renacuajos, planarias, Formol al 10% sanguijuelas, lombrices de tierra, pequeños mamíferos, etc. Bichos bolita, libélulas, arañas, hormigas, ciempiés, etc. Alcohol 70ª Inventario y catalogación 11. Realice el inventario y la catalogación de la colección. El inventario de la colección se lleva a cabo en una ficha individual que contiene toda la información referente al objeto o al ser vivo coleccionado. Aparte del inventario, es conveniente realizar una clasificación u ordenamiento de los objetos a partir de características comunes, con la finalidad de proceder a su catalogación. Para ello enumerar y marcar en alguna parte que no sea visible todos los objetos y seres (o los cotenedores donde se exhiben). Ficha técnica Número: Tipo de piedra coleccionada Procedencia..................... Época……. Medida: Largo............................. Ancho......... Alto.............Peso......... Otros Técnica Estado del objeto Bueno....................... Regular ................... Malo Estado de Integridad Completo............................ Incompleto.......................... Unido Descripción Forma de adquisición Por donación............................ Por compra.................... Por canje Observaciones Guión museográfico y fichas técnicas 12. Realice un guión para el museo. Este guión museográfico puede ser la descripción de los objetos y seres coleccionados y también los comentarios de los visitantes (y usuarios). Los textos deben ser breves y sencillos y deben ser acompañados de dibujos, diagramas y mapas. 13. Elabore fichas técnicas para los siguientes elementos. Estas fichas deben ser breves descripciones de los elementos exhibidos. Una vez seleccionados todos los objetos del museo escolar se deben elaborar las fichas técnicas y rótulos de identificación para la exposición. La información debe ser clara, precisa y breve. Caracol de agua dulce (ampularia) Piedra canto rodado un lago Hojas o de de saeta Preparado con protoozos comunes (ciliados) Ditisco (escarbajo acuático) Montaje del museo Es la última etapa en la creación del museo escolar. El objetivo del montaje es poner directamente al espectador frente al objeto de la colección para que se establezca un dialogo valorativo y visual. 14. Ubique y disponga los objetos en la sala destinada a la exposición de los objetos coleccionados. Debe contar con buena iluminación y condiciones de seguridad. 15. Use las cajas de cartón, tarros o trozos de madera para construir bases o pedestales para exponer objetos tridimensionales. Estos pueden hacerse a partir cajas de cartón, tarros, trozos de madera, láminas y pintarlos para su presentación. 16. Ubique el microscopio con los preparados para exhibir, el herabario y la pecera y otros frascos con los orgnaismos acuáticos. 17. Arme la cartelera con las fichas técnicas y las descripciones. Puntos a considerar en las demostraciones prácticas Los siguientes puntos son considerados por Trowbridge (1969), como requisitos fundamentales para llevar a cabo la implementación de trabajos prácticos como estrategia de enseñanza y de aprendizaje, por lo tanto es de fundamental importancia que el docente tenga en cuenta que: • Los objetivos debe estar bien visible para los alumnos. • El docente debe explicar y ser claro respecto a las consignas iniciales. • Estar familiarizado con los aparatos e instrumentos simples. • Las demostraciones deben ser dinámicas y atractivas. • Se debe conseguir un interés general de participación por parte de los alumnos. • Comenzar la práctica con una pregunta disparadora “inductiva” para acaparar el interés. • Formular preguntas de interés respecto a lo que se hará • El docente debe tener siempre el propósito bien presente de lo que quiere demostrar. • Las respuestas de los alumnos debe ser bien recibidas. • Los objetivos deben ser anotados previamente en el pizarrón, por el profesor, antes de comenzar la práctica. • Al finalizar el TP, se debe pedir a los alumnos un resumen de lo realizado. • Evaluar los puntos más importantes de la experiencia. Entre las sugerencias para el museo se pueden destacar también las siguientes: - Colecciones: minerales, rocas, fósiles. Colección de minerales y rocas. Colección de fósiles. - Maquetas y maderas en relieve. Es conveniente su realización por los alumnos. - Modelos: esqueleto, hombre clásico, órganos, flor, pliegues. Análisis 1. Sugiera posibles objetos y seres vivos que podría incluir en una colección escolar. 2. Utilice el protocolo analizado en detalle para sugerir otros tipos de colecciones: por ejemplo, animales aeroterrestres como insectos, rocas, minerales, figuras y fotografías de animales. 3. Arme una simulación escolar para representar un diseño experimental en el museo. Por ejemplo, puede armar alguno de los aparatos y dispositivos sugeridos en los capítulos anteriores o exhibir preparados de organismos microscópicos con sus fotografías. Técnica de tratamiento de microfósiles. El método habitual que se sigue para el tratamiento de una muestra de 10 g de roca corriente para detectar microfósiles es el siguiente: a) rotura en fragmentos menores de 5 mm si se trata de una roca coherente; b) disolución de la fase carbonatada, sumergiendo la muestra en una solución de ácido clorhídrico (HCl) al 50% ; c) disolución de la fase silícea (cuarzos y silicatos) en ácido fluorhídrico (HF) al 70% (¡atención: sumamente peligroso!), y luego disolución de lso fluosilicatos formados con ácido clorhídrico (HCl) al 50%, 10 a 30 min; d) oxidación selectiva de las partículas carbonosas y de los compuestos húmicos con sosa cáustica (K OH) al 10% durante 30 min para su eliminación; e) eliminación de la pirita y limpieza del material palinológico mediante baño bree en unos minutos de duración en ácido nítrico (H NO3) concentrado; f) sometimiento a acetólisis en mezcla de ácido sulfúrico concentrado (H2SO4) y anhídrido acético (CH3C00-0-CH3). Después de cada ataque de los reactivos hay que proceder a uno o dos lavados con agua, seguidos de centrifugación. Los residuos se recogen en mallas de 5 a 20 µ o se filtran en mallas de 50 a 100 µ. Los ácidos concentrados en general (y, particularmente, el ácido clorhídrico al 100%) disuelven los carbonatos y permiten separar los microfósiles silíceos, fosfáticos y carbonáticos. El ácido fluorhídrico al 70% seguido de ácido clohídrico al 50% disuelve a los microfósiles silíceos y permite la separación de los microfósiles fosfáticos y carbonáticos. El ácido acético, monocloroacético o fórmico al 10 o 20% permite la extracción de los microfósiles fosfáticos. La mezcla de ácido sulfúrico y anhídrido acético permite la extracción de los microfósiles orgánicos. Clave dicotómica para microfósiles. A. De carbonato cálcico... 1. Caparazón mineralizado esferoidal a elíptico. Marinos. Protozoarios ameboides, con seudópodos retráctiles, granulosos, filamentosos o anastomosados en retículos. Biocrono: cámbrico superior (Bathysiphon)-reciente FORAMINÍFEROS. 1.1. Uniloculares (con una cámara). Aglutinados (con partículas de materiales diversos cementadas por quitina y/o carbonato cálcico). Cámbrico sup-Rec. TEXTULARIINOS. 1.2. Pluriloculares (con más de una cámara) pero sin tabiques. Aglutinados (con partículas de materiales diversos). Cámbrico sup.-Reciente.TEXTULARIINOS. 1.3. Pluriloculares tabicados. Bentónicos. 1.3.1. Caparazón microgranular o fibroso. Ordovícico sup-Jurásico sup. FUSULININOS. 1.3.2. Caparazón aporcelanado. Carbonífero-Reciente. MILIOLINOS. 1.3.3. Caparazón hialino o vítreo. Pérmico-Reciente. ROTALIINOS. 1.3.3.1. Pequeños, sin sistema canalicular. BULIMÍNIDOS. 1.3.3.2. Pequeños, con sistema canalicular, trocoespirales. ROTÁLIDOS. 1.3.3.3. Con sistema canalicular, planoespirales y de gran tamaño. PaleocenoPlioceno. NUMMULÍTIDOS. 1.3.3.4. Sin sistema canalicular, lenticulares o asimétricos, muy complejos, con cámaras laterales y ecuatoriales, de gran tamaño. Cretácico sup-Pleistoceno. ORBITÓIDIDOS. 1.4. Pluriloculares tabicados. Con cámaras globososas. Planctónicos. Triásico inferior-Reciente. GLOBIGERININOS. 2. Caparazones bibalvos. Con 7 pares de apéndices articulados. Lacustres, fluviales o marinos. Bentónicos o planctónicos. Cámbrico inferior-Reciente (las formas planctónicas aparecen en el ordovícico y las lacustres y fluviales en el carbonífero). OSTRÁCODOS. 3. Caparazones en forma de lóriga, con una apertura. Esferoidales, ovoides o en forma de urna. Fibroso-radiados. Protozoarios ciliados. Ordovício superiorReciente. Marinos y lacustres. CALPIONÉLIDOS. 4. Caparazones esferoidales, sin cámaras ni tabiques, grandes, de paredes gruesas. Morfología blanda desconocida. Devónico superior-Cretácico superior. Marinos. CALCISFÉRIDOS. 5. Tubos calcáreos aislados o compactos. Restos de estromatolitos de algas azules. Marinos. Precámbrico-Reciente. CIANÓFITOS. 6. Vainas pluriseriadas y ramificadas. Restos de algas rojas o verdes. Cámbrico inferior-Reciente.RODÓFITOS Y CLORÓFITOS. 7. Girogonitos. Con morfología en espiral. Devónico superior-Reciente. Lacustres o fluviales. CHARÓFITOS. 8. Caparazones en forma de conos reticulados, no tabicados o caparazones trocoespirales. Gasterópodos con un pie muscular. Eoceno-Reciente. PTERÓPODOS. 9. Caparazones en forma de conos estrechos, tabicados. Marinos. Silúrico superior-Cretácico superior. Afinidades desconocidas. TENTACULÁRIDOS. 10. Cocolitos. Caparazón formado por placas o corpúsculos calcáreos individuales. Unicelulares flagelados con pigmentos. Pérmico inferior-Reciente. Marinos. COCOLITOFÓRIDOS. 11. Anillos pentagonales o hexagonales con tabiques y puntas (como los radiolarios). Marinos. Triásico-Mioceno. Afinidades desconocidas. ESTEFANOLÍTIDOS. 12. Bastones cilíndricos, piriformes o aciculares. Cretácico inf-Plioceno superior. Afinidades desconocidas. MICRORHABDÚLIDOS. 13. Caparazones cilíndricos, globosos o piriformes, formados por plaquietas triangulares. Jurásico superior-Cretácico superior. Marinos. NANOCÓNIDOS. 14. Quistes calcáreos, esféricos ornamentados con espinas y cretas. Relacionados con las dinofíceas, algas unicelulares flageladas. Jurásico superiorReciente. Marinos. CALCIODINÉLIDOS. B. De sílice... 1. Con una cápsula de estructura compleja, con tabiques y trabéculas, formado por espículas. Protozoarios ameboides on filópodos o axópodos, expansiones citoplasmáticas rígidas. Ordovícico inferior-Reciente. Marinos. RADIOLARIOS. 2. Frústulo formado por dos valvas o tecas, ligeramente desiguales (epiteca e hipoteca). Cretácico inferior. BACILLARÓFITOS O DIATOMEAS. 2.1. Centrales, von valvas radiales redondas o poligonales, con poros dispuestos en filas radiales o concéntricas. Cretácico inferior-Reciente. Marinos. CENTRALES. 2.2. Pennales, con valvas elpticas o triangulares y bilaterales. Con un rafe. Eoceno-Reciente. Marinos, lacustres y fluviales. PENNALES. 3. Esqueleto formado por tubos huecos reticulados que dejan grandes aberturas. Marinos. Planctónicos. Cretácico inferior-Reciente. SILICOFLAGELADOS. 4. Cápsula formada por espículas tetraxonas o triangulares unidas (con tres o cuatro ejes) parecida a la de los radiolarios y silicoflagelados. Marinos Planctónicos. Paleoceno superior-Reciente. EBRIDÁCEOS. 5. Placas estrelladas de un grupo de dinoflagelados. Marinos. Cretácico superiorReciente. ACTINÍSCIDOS. C. De sulfato de estroncio... Con cápsula formada por espículas como las de los radiolarios. Marinos. Recientes. ACANTARIOS. D. De fosfatos... Estructuras mandibulares de un grupo fósil, los conodontofóridos. Marinos. Cámbrico 1/2 -Triásico superior. Marinos. CONODONTOS. E. Palinomorfos u orgánicos (de quitinas, esporopoleninas, lignindas, paraquitinas, etcétera). 1. En forma de copa o urna. Ordovícico inferior-Devónico superior. Marinos. QUITINOZOARIOS. 2. Quistes de dinoflagelados. Silúrico superior-Reciente. Marinos, lacuestres y fluviales. DINOFLAGELADOS. 2.1. Proximales. Formados en contacto con la teca celulósica. Con la típica tabulación de aquella. 2.2. Corados o Distales. Formados en el interior de la teca. Con apéndices característicos terminados en puntas estrelladas. 3. Esferoidales con paredes gruesas dobles. Marinos. Cámbrico superior-Pérmico superior. TASMANÍTIDOS. 4. Mandíbulas de poliquetos. Marinos. Cámbrico inf-Reciente. ESCOLECODONTOS. 5. Cónicos con estrías. Prosículos de graptolitos. Ordovícico superior-Devónico superior. Marinos. GRAPTOLITOS. 6. En forma de tapiz con dibujos geométricos. Silúrico superior-Reciente. Lacustres. HIDRODICTIÁCEOS (Pediastrum). 7. Esferoidales a elípticos. Con aperturas y ornamentaciones variables. Aeroterrestres. Silúrico superior-Reciente. ESPORAS Y GRANOS DE POLEN. En algunas películas aparecen inexactitudes o errores sobre las eras geológicas. Busque información adicional y responda. a) ¿Es posible que el ser humano haya conocido a los dinosaurios? b) ¿Pudo el ser humano convivir con los mamuts y dientes de sable? c) ¿El ser humano moderno conoció al Carcharodon megalodon, el gran tiburón comeballenas? d) En Parque Jurásico I de Steven Spielberg, aparecen el tiranosaurio y el velociraptor que caza en mandas junto con grandes reptiles saurópodos herbívoros. ¿Esto es posible? e) En Parque Jurásico I de Steven Spielberg, aparecen el tiranosaurio y el velociraptor que caza en mandas junto con grandes reptiles saurópodos herbívoros. ¿Esto es posible? f) En la película Dinosaurio aparecen primates modernos (lemures) viviendo con dinosaurios. ¿Esto es posible? Indique el período geológico (y etapa de la historia evolutiva según la lámina) en que se desarrollaron los organismos. Belemnoideos o belemnites: un tipo de cefalópodos. Trilobites: artrópodos primitivos. Teleósteteos. Radiolarios. Granos de polen. Tiburones. Diatomeas. Primates. Complete las frases del texto en base a la información de las páginas 70 y 71. Los registros fósiles más antiguos se remontan a - - - - - - - - - - - y corresponden a - - - - - - - - . Las - - - - - - - - fueron las responsables de la producción de - -- - - - - - que cambió la atmósfera terrestre. Los fósiles característicos de los tiempos precámbricos son los - - - - - - - - . La primera crisis en la atmósfera terrestre tuvo lugar hace aproximadamente de 2.200 a 1.800 millones de años cuando los niveles de oxígeno treparon hasta el - - - - - - -. Para enfrentar este aumento del nivel de oxígeno, muchos - - - - -- y - - - - - - - debieron desarrollar la capacidad de - - - - - - - - - - - - - - -. El período - - - - - - - de la era - - - - - - - es considerado el momento de mayor riqueza evolutiva de la historia biológica. Al final de esta era, sin embargo, se extinguió el - - - - - - - - - - en el período - - - - - -- - . Los dinosaurios evolucionaron en el - - - - - -- - - , conformaron un grupo muy diverso en el - -- - - - - - y finalmente se extinguieron en el - - - - - - - -. - - -- - - - - - significa vida reciente. El límite entre los períodos - - - - - - - - y - - - - - - - - esta marcado por el origen del ser humano. Clave de respuestas: 3.500 millones de años, bacterias, cianobacterias, oxígeno, estromatolitos, 15%, procariotas, protistas, respiración celular, Cámbrico, Paleozoica, 90% de la biota, Pérmico, Triásico, Jurásico, Cretácico, Cenozoica, Terciario, Cuaternario. Bibliografía Obligatoria Carretero. A, Sáez Brezmes. M. J (1998) “La cultura científica, un reto educativo” Editorial. La Muralla S. A. Madrid. España. Fesquet. A. (1970) “Manual de la UNESCO para la enseñanza de las ciencias”. Editorial Sudamericana. Buenos Aires. 3ª edición. P. 19- 34 Fumagalli. L. (1997) “El desafío de enseñar Ciencias Naturales” Editorial Troquel S. A. Buenos Aires. 1ª edición Marcelo, C. (1992). Aprender a enseñar: “Un estudio sobre el proceso de socialización de profesores principiantes”. C.I.D.E. Madrid. España. Optativa Souto. Marta y otros (1996) “La formación de formadores (entre la teoría y la práctica)”. Ediciones Novedades Educativas Facultad de filosofía y letras UBA. Trowbridge. L. W, Sund. H. (1969). “La enseñanza de la ciencia en la escuela secundaria”. Editorial Paidos. Buenos Aires. p. 40 - 116 Del capacitador y del curso - Aceñolaza, F.g. y F.R. Durand. 1987. Paleontología del límite PrecámbricoCámbrico en Argentina. Xº Congr. Geol. Arg. (S. M. de Tucumán), Actas I: 315320. - Conway, S. y H.B. Whittington. 1979. Los animales de Burgess Shale. Investigación y Ciencia 36: 88-99.
 0
0
Anuncio
Documentos relacionados

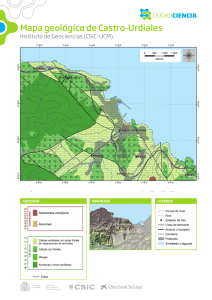
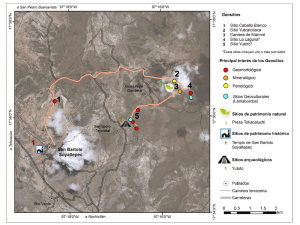





Descargar
Anuncio
Añadir este documento a la recogida (s)
Puede agregar este documento a su colección de estudio (s)
Iniciar sesión Disponible sólo para usuarios autorizadosAñadir a este documento guardado
Puede agregar este documento a su lista guardada
Iniciar sesión Disponible sólo para usuarios autorizados