- Ninguna Categoria
Importancia de las Citocinas
Anuncio
UNIVERSIDAD VERACRUZANA
FACULTAD DE BIOANALISIS
“Importancia de las Citocinas”
M O N O G R A F I A
Que para obtener el Título de:
LICENCIADO EN QUIMICA CLINICA
i
i
Presenta:
.nrfCa
ROSA M ARIA P ER A LTA V A Z Q U E Z
Asesor:
Q.C. SAN D R A MURRIETA
X A LAPA, VER.
1999.
AGRADECIMIENTOS.
A DIOS
Por su amor, su fé y su compañía.
A MIS PADRES
Abel y Mercedes con mucho cariño, por haberme permitido concluir mis
estudios y por contar con ellos en todo momento.
A MIS HERMANOS
Abel y Claudia por crecer juntos y por estar conmigo cuando más los
necesito.
A RAFAEL
Con mucho amor, te agradezco el ser mi estímulo y tu apoyo
incondicional para la realización de esta tesis.
A SANDRA MURRIETA
Por ser una gran compañera y la asesora de este trabajo.
AL HONORABLE JURADO:
Q.C. HECTOR ESCOBAR HENRIQUEZ.
Q.C. SANDRA LUZ GONZALEZ HERRERA.
Q.C. MARIA TERESA CRODA TODD.
DR. FRANCISCO MALPICA.
INDICE:
CAPITULO UNO.
1 - CITOCINAS.
1 .1 - Definición de citocinas.
1.2 - Descubrimiento y caracterización.
1 .3 - Propiedades generales.
1.4 - Funciones generales.
1.5 - Nomenclatura y clasificación.
1.6 - Bioquímica y fisiología.
Pag.
i
n
■OliOT? >
r i*
\I »
CAPITULO DOS.
2 - CITOCINAS QUE MEDIAN LA INMUNIDAD NATURAL.
2.1 - Interferón tipo 1.
2.2 - Factor de necrosis tumoral.
2.3 - Interleucina-1.
2.4 - Interleucina - 6 .
2.5 - Quimiocinas.
CAPITULO TRES.
3 - CITOCINAS QUE REGULAN LA ACTIVACIÓN, PROLIFERACIÓN Y
DIFERENCIACIÓN LINFOCITARIA.
3.1 - Interleucina-2.
3.2 - Interleucina -4.
3.3 - Factor transformador de crecimiento |3
CAPITULO CUATRO.
4 - CITOCINAS QUE REGULAN LA INFLAMACIÓN DE ORIGEN
INMUNITARIO.
4.1 - Interferón y
4.2 - Linfotoxina.
4.3 - Interleucina-10.
4.4 - Interleucina -5.
4.5 - Interleucina-12.
4.6 - Factor de inhibición de la migración.
CAPITULO CINCO.
5 - CITOCINAS QUE ESTIMULAN LA HEMATOPOYESIS
5.1 - Ligando de c-KIT.
5.2 - Interleucina -3.
5.3 - Factor estimulador de colonias de granulocitos-macrófagos
5.4 - Factor estimulador de colonias de monocitos-macrófagos.
5.5 - Factor estimulador de colonias de granulocitos.
5.6 - Interleucina-7.
5.7 - Otras citocinas estimuladoras de colonias.
1
1
2
3
5
6
7
8
9
10
13
16
17
19
20
23
24
26
27
29
30
30
31
32
33
36
37
37
38
38
39
39
CAPITULO SEIS.
6 - ALGUNOS TEMAS CLÍNICOS RELACIONADOS CON LAS CITOCINAS
6.1 - Mielopoyesis.
6.2 - Oncología.
6.3 - Producción de citocinas por los tumores.
6.4 - Citocinas en el tratamiento de cáncer.
6.5 - Citocinas en el caso de sepsis.
6.6 - Otro rol: las vacunas.
CAPITULO SIETE.
7 - ENSAYOS Y BIOENSAYOS.
7.1 - Ensayos.
7.2 - Bioensayos.
7.3 - Inmunoensayos.
7.4 - Ensayos de Citocinas (ELISA y LIA).
7.5 - La necesidad de una estandarización.
7.6 - Aplicaciones clínicas.
“
40
40
41
41
41
42
44
44
45
45
47
48
CONCLUSIONES.
49
BIBLIOGRAFIA.
50
CAPITULO UNO.
1.1 - DEFINICION:
Las atocinas son proteínas de tamaño pequeño a mediano ó glicoproteínas
utilizadas por las células inmunes e inflamatorias para comunicarse entre sí y para
controlar el medio interior en el cual operan, con actividades potentes y biológicas
diversas. La evidencia reciente sugiere que éstas son de inmensa importancia para
controlar los sucesos locales y sistémicos de la respuesta inmune, la inflamación,
hemopoyesis, el desarrollo neonatal y óseo, reparación de heridas (respuesta
sistèmica a la lesión), y las interacciones entre las diferentes vías biológicas.
Las atocinas pueden ser producidas como moléculas precursoras grandes
que pueden ser procesadas mediante la proteólisis mediada por las enzimas hacia
las formas de peso molecular más bajo. En ciertos ejemplos la forma precursora más
grande es inactiva, mientras que las otras formas precursoras pueden tener actividad
biológica. Las mitades de carbohidratos de las citocinas glicosiladas no parecen ser
necesarias para la actividad biológica, pero pueden afectar su farm acodinam ia.(3,5)
Las citocinas son moléculas secretadas característicamente en varias
cantidades, las cuales son rápidamente sobrereguladas en respuesta a varios
estímulos ambientales (incluyendo las concentraciones aumentadas de otras
citocinas). Estas pueden tener efectos sobre sus células de origen (efectos
autócrinos), sobre células adyacentes de otros tipos (efectos parácrinos) ó
sistemáticamente (efectos endocrinos).
En general, las citocinas se sintetizan en respuesta a estímulos inflamatorios ó
antigénicos y actúan en forma local, de manera autócrina ó parácrina como se
mencionó anteriormente, uniéndose a receptores de alta afinidad de las células
diana. Ciertas proteínas pueden producirse en una cantidad suficiente para circular y
ejercer acciones endocrinas.
El término citocina describe a un grupo de péptidos con efectos
inmunorregulatorios, siendo estos péptidos distintos de los factores de crecimiento
que modulan la proliferación de células no inmunes.
Numerosas enfermedades pueden ser explicadas basándose en una
producción no balanceada de citocinas. Su secreción anormal puede prolongar los
estragos patológicos, mientras que ciertas enfermedades pueden ser exacerbadas ó
causadas por la secreción anormal de citocinas, como un fenómeno secundario
después de un suceso iniciador, tal como la infección viral ó bacterial.
Las anormalidades en la producción de las citocinas están involucradas en
las enfermedades mediadas por células inmunes, tales como las alergias, el asma
extrínseco, artritis reumatoide, el lupus eritematoso sistèmico, síndrome de Sjogrin, la
pancitopenia autoinmune y la psoriasis. Estas están involucradas en las
malignidades tales como el mieloma múltiple, la leucemia aguda, la leucemia
linfocítica crónica, la enfermedad de Hodking, el linfoma de Burkitt y la enfermedad
de Castleman.
La producción anormal de citocinas ocurre en las enfermedades inflamatorias
incluyendo la pancreatitis, el síndrome de fuga capilar, la mielodisplasia, el mixoma
cardiaco, la osteoporosis, rechazo de injertos, la curación de heridas, la falla renal y
la malaria cerebral.(3,5)'
1.2 - DESCUBRIMIENTO Y CARACTERIZACIÓN.
El descubrimiento de una citocina puede relacionarse con la investigación de
una enfermedad infecciosa ó de una respuesta inmunitaria inducida por un antígeno.
Los primeros estudios sobre las citocinas se realizaron a partir de 1950 y las
describían como “numerosos factores proteicos producidos por diferentes células que
mediaban funciones particulares en bioanálisis particulares ” . (12 20>En la segunda fase de investigación, en la década de los 70’s se llevó a cabo
la purificación y caracterización parcial de muchas citocinas y se observó primero
que diversos efectos mediados por citocinas, estaban a menudo producidos por las
mismas moléculas. Por ejemplo:
El Interferón (IFN-y) fue descubierto por virólogos como “una proteína antiviral
derivada de la célula T” y de forma independiente por inmunólogos como “activador
de las funciones de los macrófagos derivados de las células T . ( )'
Una importante hipótesis generada en ese momento fue que las citocinas eran
sintetizadas principalmente por leucocitos que actuaban básicamente sobre (otros)
leucocitos y por ello las denominaron interleucinas (IL). Por ejemplo:
A un coestimulador de la actividad de los timocitos derivados de los
macrógafos se le llamó IL-1, y a un factor de crecimiento derivado de la célula T se le
llamó IL-2.
No obstante las preparaciones de citocinas de las que se disponían en ese
tiempo solían ser impuras, y muchos de los anticuerpos anticitocina no eran
absolutamente específicos de esta citocina, por lo que fue una de las limitantes
metodológicas que evitaron la identificación firme de los factores activados como la
misma molécula ó moléculas distintas.
La edad de oro de las investigaciones de las citocinas fue en la década de los
80’s donde se realizó la clonación y expresión molecular de citocinas individuales y la
producción de anticuerpos neutralizantes completamente específicos, a menudo
3
monoclonales. Esto permitió una identificación
propiedades de la molécula de citocina. (12'20)-
definitiva
de
la estructura
y
En esta etapa de investigación, fue más que una culminación del trabajo
previo, porque además, se descubrieron muchas citocinas nuevas y se revelaron
muchas propiedades inesperadas de citocinas conocidas.
Los últimos 50 años han presenciado el reconocimiento y caracterización de
muchas hormonas producidas por células especializadas. Desde los experimentos
de Rich y Lewis en 1932, sin embargo, ha estado claro que las células con funciones
específicas tales como la inmunidad y la fagocitosis son capaces de producir
moléculas de señalamiento solubles.(20)El desarrollo de la tecnología de DNA en los últimos 10 años se ha convertido
en una serie de factores y actividades que ha demostrado que está formado por un
gran número de péptidos bien definidos que llevan los signos de actividad,
crecimiento y diferenciación de un amplio rango de células. Su importancia en el
diagnóstico y tratamiento de la enfermedad es probablemente enorm e.(9)'
1.3 - PROPIEDADES GENERALES.
Aunque las citocinas son un grupo de diversas proteínas, comparten varias
propiedades...
I)
Las citocinas se producen durante las fases efectoras de la inm unidad natural
y específica, y sirven para m ediar y regular las respuestas inm unitarias e
inflam atorias. En la inmunidad natural, los productos microbicidas, como el
lipopolisacárido (LPS), estimulan directamente a los fagocitos mononucleares
a secretar sus citocinas. Por el contrario las citocinas derivadas de las células
T se producen en respuesta sobre todo al reconocimiento específico de
antígenos extraños. Sin embargo, estas distinciones no son absolutas porque
las citocinas producidas por un tipo celular suelen regular la síntesis de
citocinas por otras células.
II)
La secreción de citocinas es breve, autolimitada. En general, las citocinas no
se almacenan como moléculas pre-formadas, y su síntesis se inicia por una
nueva transcripción genética. Esta activación suele ser transitoria, y los RNAm
que modifican las citocinas suelen ser inestables. La combinación de un
periodo corto de transcripción con un RNAm de vida corta, asegura que la
síntesis de citocinas sea transitoria. Algunas citocinas pueden controlarse
además por ciertos mecanismos, como la liberación proteolítica de un
producto activo a partir de un precursor inactivo. Una vez sintetizadas, las
citocinas suelen secretarse con rapidez, lo que dá lugar a un pico de liberación
de citocinas cuando se necesita.
4
III)
Muchas atocinas son producidas po r muchos tipos celulares. La fuente celular
de estas moléculas no suele ser una característica que las distinga. Pero
como ya se ha mencionado entre las células que intervienen en su formación
están los fagocitos mononucleares, linfocitos T activados y linfocitos.
IV)
Las citocinas actúan sobre diferentes tipos celulares. Esta propiedad se llama
pleiotrofismo. Al concepto de que las citocinas son básicamente moléculas
producidas por los leucocitos que actúan en particular sobre los leucocitos
(interleucinas) se le considera demasiado restringido.
V)
Las citocinas tienen a menudo m últiples efectos diferentes sobre la misma
célula diana. Algunos efectos pueden ser simultáneos, mientras que otros
ocurren en franjas de tiempo diferentes(es decir, minutos, horas o d ía s ).(3,5)
VI)
Las acciones de las citocinas son a menudo redundantes. Muchas funciones
atribuidas originalmente a un a citocina comparten propiedades de varias
citocinas diferentes. Esta observación se ha reforzado con el estudio de los
ratones =Knock-out= que carecen de genes de una citocina particular que
puede mostrar solo alteraciones sutiles en sus respuestas inmunitarias,
VII)
Las citocinas a menudo influyen en la síntesis de otras citocinas. Produciendo
cascadas en las que una segunda o tercera citocina pueden mediar los
efectos biológicos de la primera. La capacidad de una citocina de aumentar o
suprimir la producción de otras citocinas puede proporcionar importantes
mecanismos reguladores positivos y negativos para las respuestas
inmunitarias e inflamatorias.
VIII)
Las citocinas a menudo influyen en la acción de otras citocinas. Dos citocinas
pueden interaccionar para antagonizar sus acciones, producir efectos aditivos
ó, en algunos casos, producir efectos mayores de los previstos e incluso
distintos, un tipo de acción que se llama “sinèrgica”.
IX)
Las citocinas, como otras hormonas polipeptídicas, inician su acción
uniéndose a receptores específicos en la superficie de la célula diana. La
célula diana relevante puede ser la misma célula que secreta la citocina
(acción autócrina) una célula cercana (acción parácrina) o, como las
verdaderas hormonas, una célula distante que se estimula a través de las
citocinas secretadas a la circulación (acción endocrina). En consecuencia solo
necesitan producirse cantidades muy pequeñas de citocinas para
desencadenar efectos biológicos.
X)
La expresión de muchos receptores de citocinas está regulada p o r señales
específicas. Esta señal puede ser otra citocina ó incluso la misma citocina que
se une al receptor, permitiendo la amplificación positiva ó la retroalimentación
ne gativa .(3,5)-
5
XI)
La m ayor parte de las respuestas celulares a las atocinas precisan RNAm
nuevo y la síntesis proteica. El mecanismo por el que la unión de la citocina a
un receptor de superficie celular estimula transcripción se explicará más
detalladamente. Algunos estudios recientes han identificado secuencias de
nucleótidos en las regiones adyacentes a los genes, cuya transcripción se
activa por la acción de las citocinas. Se cree que las citocinas estimulan la
producción ó la unión de factores nucleares reguladores específicos a estas
secuencias diana, y que tal unión produce la transcripción.
XII)
Para muchas células diana, las citocinas actúan como reguladores de la
división celular, es decir, como factores de proliferación. Algunos inmunólogos
prefieren que las citocinas se clasifiquen con los factores de proliferación
celular epitelial y mesenquimal en un grupo funcional mayor de moléculas
reguladoras polipeptídicas. Sin embargo ahora se prefiere separar a aquellas
moléculas cuyas acciones básicas son la mediación de la defensa del
huésped (es decir, las citocinas), de aquellas moléculas cuya función básica
reside en la reparación de tejidos (es decir, los factores de proliferación celular
polipeptídicos epitelial y mesenquimal). (3'5)'
1.4 - FUNCIONES GENERALES.
A continuación, se han organizado a las citocinas específicas en 4 clases de
funciones amplias:
A) M ediadores de la Inm unidad natural. Desencadenadas por agentes infecciosos a
partir de los fagocitos mononucleares. En donde se encuentran:
• Interferón tipo 1.
• Factor de necrosis tumoral.
• lnterleucina- 1 .
• lnterleucina- 6 .
• Quimiocinas.
B) Citocinas que regulan la activación, proliferación y diferenciación linfocitaria. Que
surgen en respuesta al reconocimiento de antígenos específicos por los linfocitos:
• lnterleucina- 2 .
• lnterleucina-4.
• Factor transformador del crecimiento (5
C) Reguladoras de la inflam ación de origen inmunitario. Que activan a células
inflamatorias inespecíficas en respuesta a antígenos específicos por los linfocitos T:
• Interferón y.
• Linfotoxina.
• lnterleucina- 10 .
• lnterleucina-5.
• lnterleucina- 12 .
6
D) a to c in a s que estim ulan la proliferación y diferenciación de los leucocitos
inm aduros (hematopoyesis).
• Ligando de c-KIT:
• lnterleucina-3.
• Factor estimulador de colonias de granulocitos-macrófagos.
• Factor estimulador de colonias de monocitos- macrófagos.
• Factor estimulador de colonias de granulocitos.
• lnterleucina-7.
• Otras citocinas estimuladoras de colonias.
Esta clasificación se basa en las que perecen ser las principales de las diferentes
acciones biológicas de una citocina, aunque como se verá más adelante, muchas
citocinas pueden actuar en más de una c la s e .<5)
1.5 - NOMENCLATURA Y CLASIFICACIÓN.
Como ya se mencionó él termino citocina ha sido utilizado para describir un
grupo de péptidos con efectos inmunoregulatorios, siendo estos péptidos distintos de
los factores de crecimiento que modulan la proliferación de células no inmunes.
Ahora se sabe que existe una considerable relación funcional entre las
citocinas y los factores de crecimiento. Por lo tanto, él termino factor regulador
péptido es usado para describir las moléculas de baja masa molecular (menor de 80
kD) que tiene un rango corto ó intermedio de acción, receptores específicos y la
habilidad para controlar la diferenciación y la proliferación de las células, sin
embargo, algunas hormonas clásicas particularmente entre el vientre y el cerebro
también caen dentro de esta categoría.
Para el interés de los Químicos Clínicos están las Interleucinas (IL), los
interferones (IFN), los factores de necrosis tum oral (TNFS), los factores de
crecim iento hem opoyéticos y los factores de crecim iento de colonias (CFS).
No hay evidencia alguna que indique que las mediciones de los otros factores
de crecimiento puedan ser aplicadas útilmente a los fluidos biológicos; a pesar de los
intentos para unificar la nomenclatura, la terminología no está bien definida.
El termino IL fue introducido para describir los productos de la conocida
secuencia de aminoácidos producida por linfocitos ó macrófagos, a pesar de esto,
varios mediadores encajan en esta descripción. Ejemplo: El factor de necrosis del
tumor, los interferones y los factores de estimulación colonial mantienen sus nombres
originales.(3)
1.6 - BIOQUÍMICA y f is i o l o g í a .
A pesar del hecho de que la mayoría de las citocinas han sido descritas e
inicialmente nombradas en base a una simple función biológica, muchas son
moléculas multifuncionales con diversas acciones biológicas y células objetivo
(diana). En general las funciones más importantes parecen ser los efectos locales,
modulando el comportamiento de las células adyacentes (parácrina) ó la célula que
la secreta (autócrina), pero en algunos casos notablemente (IL-1, TNF e IL-6 ) hacen
efectos significativos sobre órganos ó tejidos distantes (endocrina).
En muchos casos hay duplicación de función entre las citocinas, por ejemplo:
IL-1 comparte actividades con el factor de crecimiento derivado de plaquetas (FCDP)
sobre las células del músculo liso, con el factor de estimulación colonial del
macròfago (FEC-M) sobre las células hemopoyéticas, con IL-6 sobre la inducción de
proteínas de fase aguda, y con el TNF e IL-6 en la inducción de fiebre.
En general, las citocinas actúan a través de receptores de la superficie celular
de alta afinidad, muchos de los cuales han sido duplicados y secuenciales. La
mayoría de las citocinas son glicosiladas (dando un aumento a la considerable
heterogeneidad) y las mitades de azúcares pueden ser importantes para el enlace
receptor y la m odulación.(3)
Esto significa que los estudios funcionales de las citocinas recombinantes
producidas en las bacterias (y por lo tanto no glicosiladas) pueden proveer solo una
parte de la imagen. Algunos receptores de citocinas contienen un complejo de 7
dominios transmembranales que interactúan con las proteínas de enlace GTP. Las
proteínas de enlace GTP regulan las enzimas críticas en las diferentes vías de
transducción. Ejemplos son las enzimas fosfoionocitasa C, la cual lleva a la
movilización del calcio y a la activación de la proteina quinasa C, y la
adenilatociclasa, la cual cataliza la conversión de ATP a AMPc. El receptor IL-1 está
conectado a la adenilatociclasa y a la proteina quinasa-A por vía de la proteina de
enlace GTP en algunas células, la AMP es un segundo mensajero probable.
Otros receptores de citocinas tienen un dominio intracelular con la actividad de
la tirosinaquinasa. Muchas citocinas interactúan entre sí a nivel celular. Los efectos
de la citocina sobre una célula pueden depender del medio de otras citocinas y
hormonas. Esto lleva a la conclusión importante de que la regulación celular por las
citocinas depende de las circunstancias bajo las cuales la célula está viviendo en un
momento particular. Los experimentos in vitro con citocinas simples ó hasta con
combinaciones de citocinas pueden ser engañosos, y las mediciones de citocinas
simples pueden revelar solo parte de la historia.<3)
CAPITULO DOS.
CITOCINAS QUE MEDIAN LA INMUNIDAD NATURAL.
La inm unidad es la propiedad del organismo, en virtud de la cual este no
desarrolla una enfermedad cuando entra en contacto una bacteria, virus ó toxina. La
inmunidad puede ser natural ó adquirida (activa o pasiva).
La inm unidad natural es la que el individuo posee desde el nacimiento y se
basa en una serie de mecanismos que incluye la fagocitosis y las acciones de las
citocinas.
Las citocinas que median la inmunidad natural son aquellas que protegen
contra la infección viral y aquellas que inician reacciones inflamatorias que protegen
contra las bacterias. Las citocinas que veremos en este capítulo se resumen en la
siguiente tabla.
Citocina
describir
IFN tipo 1
Factores
necrosis
tumoral
a Num. de
genes.
20 IFN-a
1 IFN-p
de 1
Tamaño del
polipéptido.
18 KD
(monómero)
17 KD
Homotrímero
17 KD
Interlucina -1 2
(IL-tx.lL-P ) (monómero)
lnterleucia-6
Quimiocinas
Fuentq
celular.
Fagocito
mononuclear
Fibroblasto
Nuclear,
célula T
Fagocito
mononuclear,
otras
Fagocito
mononuclear
Célula
endotelial
célula T
20 genes 8-10
KD Fagocito
en relación (monómero)
mononuclear
Célula
endolial
Fibroblasto
Célula T
Plaquetas
1
26 KD
homodímero
Célula
diana.
Todas.
Efectos básicos en cada
célula diana
Antiviral antiproliferativa.
Célula.NK
Neutrófilo
C. endotelial
hipotálamo
hígado
músculo
timocito
Timocito.
C.encotelial
Hipotálamo
Hígado
Músculo.
Activación
Activación (inflamación)
Timocito
Célula
madura
Hígado
Leucocitos
Activación, inflamación coagulación
Fiebre
Reactantes de fase aguda.
Catabolismos (caquexia)
Co-estimulador.
Co-estimulador
Activación
(inflamación,
coagulación)
Fiebre
Reactantes de fase aguda.
Catabolismo (caquexia)
Co-estimulador
B Proliferación
Reactantes de fase aguda
Reactantes de fase aguda.
Quimiotáxis.y activación de
leucocitos
9
2.1 - INTERFERON TIPO 1.
Los ¡nterferones (IFN) son una familia de citocinas antivirales que pueden ser
reguladores naturales negativos de crecimiento para varios tipos de células.
Basándose en la secuencia de aminoácido, son reconocidos 2 grupos del Interferon
tipo 1 :
19) Interferon a para la cual hay 13 genes con código para 15 proteínas funcionales.
Es producto celular mononuclear.
29) Interferon (3 Viene principalmente de los fibroblastos y células epiteliales.
• ) Interferon y: Inmunitario o gamma, producido por las células T. Se mencionará
independientemente, también es conocido como Interferon II.
Los ¡nterferones a y p son inducidos por virus, bacterias y DNA de doble
enlace, mientras que el Interferon y es inducido por mitógenos y antígenos. A pesar
que solamente existen un 29% de homología, el Interferon a y p usan el mismo
receptor celular. Estos son antivirales, induciendo la sintetasa oligodenilatosa la cual
causa una ruptura en RNAm. Además son proliferativos para muchas líneas de
células transformadoras y cánceres. Estas son estimuladoras potentes de células T
naturalmente destructoras y citotóxicas y sobreregulan los antígenos de clase 1 ,
MHC (complejo mayor de histocompatibilidad), mejorando el reconocimiento del
antígeno.
El IFN tipo 1 tiene 4 funciones biológicas principales:
• Inhibe la replica viral: el IFN hacen que las células sinteticen varias enzimas,
como la 2, 5 oligodenilato sintetasa, que interfieren con la replica del RNA y DNA
viral. La Acción antiviral del IFN tipo l es básicamente parácrina, ya que las
células infectadas por virus secretan el IFN para proteger a las células vecinas
todavía no infectadas. De una célula que ha respondido al IFN y es resistente a la
infección viral, se dice que está en su estado antiviral.
• Inhibe la proliferación celular: Esto se debe a la inhibición de las mismas enzimas
que bloquean la replicación viral, pero también pueden afectar a otras enzimas
que evitan la síntesis de aminoácidos, especialmente de aminoácidos esenciales
como el triptófano. Aunque los mecanismos pueden ser parcialmente diferentes,
los efectos antivirales y antiproliferativos del IFN no pueden separarse. Se ha
propuesto que el IFN-(3 es un inhibidor fisiológico de la proliferación celular
normal. El IFN-a se utiliza como un agente proliferativo en ciertos tumores (por
ejemplo, la leucemia de células peludas y los hemangiomas infantiles).
•
Aum enta el potencial Utico de las células agresoras naturales (NK). Una de las
funciones principales de las células NK es matar a las células infectadas por
virus.
10
•
Modula la expresión de moléculas del MHC. En general, el IFN tipo 1 aumenta la
expresión de moléculas de clase 1 del MHC e inhibe profundamente la de
moléculas clase 2. Dado que la mayor parte de los linfocitos T citolíticos (LTC)
reconocen antígenos extraños unidos a moléculas de clase 1 del MHC, el IFN tipo
1 estimula la fase efectora de las respuestas inmunitarias mediadas por células
aumentando la eficacia de la lisis mediada por los LTC. Al mismo tiempo, el IFN
tipo 1 puede inhibir la fase de reconocimiento de las respuestas inmunitarias
evitando la activación de los linfocitos T cooperadores restringidos por la clase 2
del MHC. (57'12)
De este modo, las principales actividades del IFN tipo 1 son, principalmente la
inducción del estado antiviral, la activación de las funciones líticas de las células NK,
y el aumento de la expresión de las moléculas de clase 1 del MHC en las células
infectadas por virus, todas ellas coordinadas para erradicar las infecciones virales.
Hay muchas razones para creer que los interferones son agentes importantes
naturales antivirales y controladores negativos del crecimiento de células. Su
importancia en la enfermedad humana se relaciona principalmente a sus efectos
antiproliferativos. Una de las aplicaciones terapéuticas más dramáticas de las
citocinas han sido las remisiones completas obtenidas mediante el IFN-a en la
leucemia de las células velludas, desafortunadamente el IFN-a ha probado ser
menos exitosa en otras enfermedades mieloproliferativas, pero es efectiva en la
leucemia mieloide crónica y tiene alguna actividad en el mieloma y en los linfomas.
La respuesta de los tumores sólidos es desilucionante.(5,7,12)
2.2 - FACTOR DE NECROSIS TUMORAL. (FNT).
El factor de necrosis tumoral (TNF, del: inglés, tumor necrosis factor) es el
principal mediador de las respuestas del huésped frente a las bacterias
gramnegativas, y puede actuar también en la respuesta frente a otros
microorganismos infecciosos. Algunos investigadores se refieren al TNF como TNFa, y a la Linfotoxina (LT) como TNF-0; aquí utilizaremos los términos como TNF y LT.
Los componentes activos de las bacterias gramnegativas son las moléculas
de lipopolisacáridos (LPS), también llamada endotoxina, derivadas de la pared
celular bacteriana. El TNF se identificó originalmente y así se le llamó, como un
mediador de la necrosis tumoral presente en el suero de animales tratados con LPS.
A bajas concentraciones, El LPS estimula las funciones de los fagocitos
mononucleares del huésped que contribuyen a la eliminación de las bacterias
invasoras. Sin embargo, concentraciones altas de LPS producen lesión tisular,
coagulación intravascular diseminada (CID) y shock, a menudo provocando la
muerte, por lo que se puede decir que el TNF es uno de los principales mediadores
de estos efectos del LPS. (1-9’13)
11
La principal fuente celular del TNF es el fagocito mononuclear activado por el
IP S , aunque las células T activadas por el antígeno, las células NK activadas y los
mastocitos activados pueden secretar también esta proteína. El IFN-y producido por
las células T, aumenta la síntesis TNF por los fagocitos mononucleares estimulados
por el LPS. De este modo, el TNF es un mediador de la inmunidad natural y
adquirida, y un importante nexo entre las respuestas inmunitarias específicas y la
inflamación aguda. (1'9'22)
La síntesis del TNF es inducida por productos bacteriales tales como la
endotoxina, la proteína 1 de choque tóxico y las proteínas micobacteriales. En
general, actuando solo el TNF produce un rango de efectos proliferativos de células,
la activación de las funciones celulares, y la expresión de enzimas y
macromoléculas. Los aspectos más importantes del TNF son sus efectos citolíticos y
citoestáticos; los cuales son fuertemente expresados cuando el TNF actúa en
sinergia con el IFN-y. Esta combinación es citotóxica para un amplio rango de células
transformadas y para las células normales epiteliales y endoteliales. El TNF es uno
de los mejores ejemplos cuyas funciones son moduladas, inducidas y hasta
invertidas en la presencia de otras citocinas.(1,9)
Los efectos fisiológicos más importantes del TNF son los siguientes:
•
Produce la expresión en las células endoteliales vasculares de receptores de
superficie nuevos (moléculas de adhesión), que hacen a la superficie endotelial
adherente a los leucocitos, inicialmente para los neutrófilos y después para los
monocitos y linfocitos. El TNF actúa también sobres los neutrófilos para aumentar
su adherencia a las células endoteliales. Estas acciones contribuyen a la
acumulación de leucocitos en los lugares de inflamación, y probablemente son los
efectos fisiológicos más importantes del TNF.
•
Activa a los leucocitos inflamatorios para que maten a microorganismos. El TNF
es especialmente potente en la activación de los neutrófilos, pero también afecta
a eosinófilos y fagocitos mononucleares.
•
Estimula a los fagocitos mononucleares y a otros tipos de células a producir
citocinas, como la IL-1, la IL-6 , el propio TNF y a las quimiocinas.
•
Ejerce un efecto protector similar al del interferón contra los virus, y aumenta la
expresión de moléculas de clase I del MHC, potenciando la lisis de células
infectadas por virus por los LTC.(1,9,22)
Estos efectos del TNF son críticos para las respuestas inflamatorias frente a los
microorganismos. Si están presentes cantidades inadecuadas de TNF, por ejemplo,
en animales tratados con anticuerpos anti-TNF neutralizantes, puede que no se
contengan las infecciones. Si el estímulo para la producción de TNF es
suficientemente fuerte, se producen mayores cantidades de citocinas. En esta
12
situación, el TNF entra en el torrente sanguíneo, donde puede actuar como una
hormona endocrina. Las acciones sistémicas principales del TNF en las respuestas
fisiológicas del huésped a las infecciones son las siguientes:
•
El TNF es un pirógeno endógeno que actúa sobre células de regiones
reguladoras del hipotálamo para inducir fiebre. Comparte esta propiedad con la
IL-1, y ambas citocinas se encuentran en el suero de personas ó animales
expuestas al LPS, que actúa como un pirógeno exógeno. La producción de fiebre
en respuesta al TNF ó la IL-1 está provocada por una síntesis aumentada de
prostaglandinas en las células hipotalámicas, estimuladas por las citocinas. Los
inhibidores de la síntesis de prostaglandinas, como la aspirina, reducen la fiebre
al bloquear esta acción del TNF y la IL-1.
•
El TNF actúa sobre los fagocitos mononucleares y quizás sobre las células
endoteliales vasculares para estimular la secreción de IL-1 e IL-6 a la circulación.
Este es un ejemplo de una cascada de citocinas que comparten muchas
actividades biológicas.
•
El TNF actúa sobre los hepatocitos para aumentar la síntesis de ciertas proteínas
séricas, como la proteína A. Este espectro de proteínas del hepatocito inducidas
por el TNF es idéntica al de la IL-1, aunque difiera del inducido por la IL -6 (que se
describirá más adelante). La combinación de proteínas plasmáticas derivadas del
hepatocito inducidas por el TNF ó la IL-1, más las inducidas por la IL-6 , constituye
la respuesta de fase aguda a los estímulos inflamatorios.
•
El TNF activa el sistema de coagulación, básicamente alterando el equilibrio entre
actividades procoagulantes y anticoagulantes del endotelio vascular.
•
El TNF suprime la división de la célula madre de la médula ósea. La
administración crónica del TNF puede producir linfopenia e inmunodeficiencia.
•
La administración continua de TNF a animales de experimentación produce
alteraciones metabólicas o caquexia, un estado caracterizado por el desgaste de
las células musculares y grasas. La caquexia se produce en gran parte por la
supresión del apetito que induce el TNF. El TNF también suprime la síntesis de la
lipoproteina lipasa, una enzima necesaria para liberar ácidos grasos de las
lipoproteínas circulantes, para su utilización por los tejidos. Aunque el FNT puede
producir caquexia por sí mismo en animales de experimentación, otras citocinas
como la IL-1, puede contribuir también a la caquexia aue acompaña a ciertas
enfermedades crónicas como la tuberculosis y el cáncer, h-9'12-22)-
La combinación de fiebre, elevación de la IL-6 , la elevación de los reactantes de
fase aguda, la supresión de la médula ósea y la activación de la coagulación, se han
observado en pacientes tratados con TNF intravenoso en la quimioterapia
antineoplásica. La administración intravenosa continua de cantidades elevadas de
TNF es por sí sola mortal, provocando un síndrome similar al shock.
i3
Varias acciones específicas del TNF a concentraciones extremadamente altas
contribuyen a los siguientes efectos mortales:
•
Reduce el riego sanguíneo tisular al deprimir la contractilidad miocàrdica. El
mecanismo de acción implica la inducción de una enzima en los miocitos
cardiacos, la sintetasa de óxido nítrico (NOs), que convierte la arginina en citrulina
y en óxido nitroso (NO). El NO que produce esta enzima inhibe la contractilidad
miocàrdica.
• Reduce aún más la presión arterial y el riego sanguíneo tisular por la relajación
del tono muscular liso vascular. El TNF puede actuar directamente sobre las
células musculares lisas y también indirectamente estimulando la producción de
vasodilatadores, como la prostaciclina y el NO por las células endoteliales
vasculares.
•
Produce trombosis intravascular, lo que reduce el riego sanguíneo tisular. Esto se
debe a una combinación de alteraciones endoteliales y de los fagocitos
mononucleares, que promueven la coagulación y a la activación de neutrófilos
que producen taponamiento vascular.
•
El TNF produce varias alteraciones metabólicas, como una caída de las
concentraciones de glucosa sanguínea a valores incompatibles con la vida. Esto
se debe a la utilización excesiva de glucosa por el músculo y a la incapacidad del
hígado de reem plazarla.(1,22)
2.3 - INTERLEUCINA -1 .
La interleucina -1 (IL-1), es una citocina multifuncional, el aislamiento de los
duplicados de DNA para IL-1 ha demostrado que 2 genes son expresados para esta
citocina: IL-a e IL-p. Estas citocinas muestran sorprendentemente poca homología
(26%), aunque estas actúan sobre el mismo receptor de alta afinidad. La mayoría de
las células nucleadas parecen capaces de producir IL-1 cuando son dañadas; pero
los macrófagos producen las cantidades más grandes y son probablemente los
productores clave en la inflam ación.(16)'
La IL-1 actúa para promover las respuestas inmunes específicas de
antígenos, la inflamación y el remodelamiento de la matriz extracelular. Las acciones
comunes sobre los muchos tipos de células son para estimular el metabolismo,
produciendo prostaglandinas y la producción de proteasas neutrales. La IL-1 tiene
actividad de factor de crecimiento en las células T también en las células B. su
actividad proinflamatoria afecta casi a cualquier suceso inflamatorio, actuando en
forma parácrina.
14
Aunque, la IL-1 se descubrió como coestimulador de las células T, ahora
sabemos que su principal función, como la del TNF es como mediadora de la
respuesta inflamatoria del huésped en la inmunidad natural. La principal fuente
celular de IL-1, al igual que el TNF, es el fagocito mononuclear activado.(8)'
La producción de IL-1 por los fagocitos mononucleares puede estimularse con
productos bacterianos como el LPS, con citocinas derivadas de macrófagos como el
TNF ó la propia IL-1 y mediante el contacto con células T CD4+. Como el TNF, la IL -1
puede encontrarse en la circulación tras una sepsis por gram negativos, donde
puede actuar como hormona endocrina.
La síntesis de IL-1 difiere de la TNF en dos aspectos importantes. Primero, las
células T son más eficaces que el LPS en la estimulación de la síntesis de IL-1 por
los monocitos mononucleares. Segundo, la IL-1 produce en muchos tipos de células
endoteliales y epiteliales, lo que proporciona fuentes locales potenciales de IL-1 sin
infiltrados ricos en macrófagos.
La purificación bioquímica de la IL-1 secretada por fagocitos macrófagos
mononucleares, reveló que la actividad biológica de esta citocina residía en dos
especies de polipéptidos principales, cada unos de 17 kD pero con puntos
isoeléctricos distintos de 5.0 a 7.0. Ahora se sabe que estas dos formas
mencionadas anteriormente, llamadas IL-a e IL-p respectivamente, son producto de
dos genes diferentes. Las dos formas de IL-1 muestran una homología estructural
menor del 30%, pero ambas se unen a los mismos receptores de superficie celular y
sus actividades biológicas son esencialmente idénticas. Cabe mencionar que las
moléculas de IL-1 se relacionan también estructuralmente con las diferentes formas
del factor de crecimiento de los fibroblastos.
Los dos polipéptidos de la IL-1 se sintetizan como precursores de unos 33kD
que se inciden mediante proteólisis para generar las proteínas de 17 kD maduras.
Las formas de 17 kD se pliegan en una estructura similar a un barril, ricas en bandas
de hojas plegadas en (3 . El precursor de la IL-1 de 33 kD es biológicamente activo,
pero el de la IL-1 j3 debe procesarse a la forma de 17 kD antes de poder ejercer sus
funciones biológicas. En los fagocitos mononucleares, se ha identificado una
proteasa específica de IL-1 que es responsable en su mayor parte de la conversión
de la IL-1 (3 a su forma activa.
Se han caracterizado dos receptores de la membrana diferentes para la IL-1,
ambos miembros de la superfamilia de las inmunoglobulinas. El receptor de tipo I se
ha caracterizó inicialmente a partir de una línea de células T donde mediaba la
estimulación de la IL-1; tiene una afinidad algo mayor por la IL-1 (3 que por la IL-1 a. El
tipo II se caracterizó inicialmente a partir de la célula B; tiene mayor afinidad por la IL1a que por la IL-1 (3. (18)
15
Los efectos biológicos de la IL-1, similares a los del TNF, dependen de la
cantidad de citocina liberada. A concentraciones bajas de IL-1, los principales efectos
biológicos son como mediador local de la inflamación. De forma específica, la IL-1
actúa sobre los fagocitos mononucleares y el endotelio vascular para aumentar a un
más la síntesis de IL-1 e inducir la IL-6 . También comparte muchas de las
propiedades inflamatorias de TNF. Por ejemplo, la IL-1 actúa sobre las células
endoteliales promoviendo la coagulación y aumentando la expresión de moléculas
de superficie que median la adhesión de los leucocitos. La IL-1 no activa
directamente a los leucocitos inflamatorios, como los neutrófilos, pero hace que los
fagocitos mononucleares y las células endoteliales sinteticen quimiocinas que activan
a los leucocitos.
Cuando se secreta en grandes cantidades la IL-1 entra en el torrente
circulatorios y ejerce efectos endocrinos. La IL-1 sistèmica comparte con el TNF la
capacidad de producir fiebre, de inducir la síntesis de proteínas plasmáticas de fase
aguda (como la proteina A del amiloide sérico) por el hígado, y de iniciar el desgaste
metabòlico (caquexia).
Aunque las acciones de la IL-1 son muy similares a las del TNF, existen varias
diferencias importantes entre ellas ...
I)
II)
III)
IV)
La 11-1 no produce lesión tisular por si misma, aunque se secreta en respuesta
al LPS y puede potenciar la lesión tisular producida por el TNF. Además,
incluso a concentraciones sistémicas muy elevadas la IL-1 no es mortal.
Aunque la IL-1 imita muchas
de las propiedades inflamatorias y
procoagulantes de TNF, no produce necrosis hemorrágica en los tumores.
La mayor parte de las líneas de células tumorales no son lisadas
directamente por la IL-1 in vitro.
La IL-1 no comparte con el TNF la capacidad de aumentar la expresión de
moléculas del M H C .(8,1618)
El interés de la IL-1 ha aumentado dramáticamente durante la última década,
pero ha sido grandemente restringida a los inmunólogos, los biológos celulares y los
que estudian la inflamación y el cáncer. Sin embargo, recientemente se ha
reconocido que el cerebro controla y modula directamente muchos aspectos de la
función inmune, mientras que las moléculas asociadas clásicamente con el sistema
inmune tales como la IL-1 son sintetizadas dentro del cerebro y actúan directamente
en el SNC para modificarlas funciones locales y sistémicas, por lo tanto este tema es
relativamente nuevo para los neurobiólogos. (18'
16
2.4 - INTERLEUCINA - 6 .
La interleucina - 6 (IL-6 ), es una citocina de aproximadamente 26 kD que
sintetizan los fagocitos mononucleares, las células endoteliales vasculares, los
fibroblastos y otras células en respuesta a la IL-1 y en menor grado, al TNF. También
es sintetizada por algunas células T activadas. La IL-6 puede detectarse en la
circulación tras la infección de bacterias gram negativas ó la administración
intravenosa de TNF, y parece secretarse en respuesta al TNF ó la IL-1 en lugar de al
propio LPS. La IL-6 no produce la trombosis vascular ó la lesión tisular que se ven en
respuesta al LPS ó al TNF.
La IL -6 es una citocina pleiotrópica que actúa sobre la respuesta inmune
específica del antígeno; en la inflamación este es un mediador clave en la respuesta
de fase aguda.
La primera función identificada de este molécula fue la inducción de la
diferenciación terminal de las células B y la producción de inmunoglobulinas. La
preactivación mediante IL-4 es necesaria. La IL -6 tiene un rol auxiliar en la activación
de las células T y sinergetiza con la IL-3 ó con el granulocito-macrófago para mejorar
el crecimiento de las células progenitoras hemopoyéticas. Se duda que la IL-6 posea
cualquier actividad antiviral. La IL-6 es probablemente la mayor inductora fisiológica
de la síntesis de proteínas de la fase aguda en el hígado, con la IL-1 y el TNF
tomando roles solamente subsidiarios.(7)
Las 2 acciones mejor descritas de la IL-6 son sobre los hepatocitos y las
células B ...
I)
II)
La IL-6 hace que los hepatocitos sinteticen varias proteínas plasmáticas como
el fibrinógeno, que contribuyen a la respuesta de fase aguda.
La IL -6 sirve como factor de proliferación para las células B activadas en fases
avanzadas de su diferenciación. La IL-6 actúa de forma similar al factor de
proliferación de muchas células plasmáticas neoplásicas (plasmocitomas ó
mieolomas), y muchas células de plasmocitomas que proliferan de forma
autónoma pueden secretar en realidad IL-6 como un factor de proliferación
autócrino. Además la IL -6 puede promover el crecimiento de los híbridos de
células somáticas producidas por la fusión de células normales con células de
plasmocitoma, es decir los hibridomas que producen lo anticuerpos
monoclonales.
La IL -6 puede inducir fiebre y la síntesis de IL -6 mediante el estroma
endometrial ha sido sugerido como mediadores de las fluctuaciones de temperatura
en el ciclo menstrual. La IL -6 actúa sobre células de tejido para mejorar o inhibir la
proliferación e induce la liberación de prostaglandina en el fibroblasto. Al igual que la
IL-1, esta puede por lo tanto tener efectos locales en la inflamación y en la
reparación. <3,7)'
17
Entre las características que hacen a la IL-6 tan interesante e importante están
su inducción mediante otras citocinas y su sinergismo con la IL -1 y el TNF para
inducir la síntesis de proteina de fase aguda y para estimular el crecimiento de los
timocitos. Estas observaciones delinean la complejidad de la cadena de citocinas.
Los roles posibles de la IL-6 en la enfermedad han sido dramáticamente
demostrados por un grupo de Kishimoto, quien creó ratones transgénicos,
expresando constitutivamente altas cantidades de IL-6 . Los ratones desarrollaron
plasmocitomas policlonales mortales con glomerulonefritis membranosa proliferativa
e infiltración masiva de la médula mediante los megacariocitos maduros. La IL -6 ha
sido fuertemente implicada como un factor de crecimiento autócrino para varios
tumores.
La mayoría de los pacientes con sepsis que han sido estudiados, han
mostrado aumentos en la IL-6 . Waage investigó a pacientes con enfermedad
meningofocal. La concentración media de suero con IL-6 fue 1000 veces mas alta en
los pacientes con choque séptico que en los pacientes con meningitis o leucemia
solamente. Hank tuvo descubrimientos similares mostrando que las concentraciones
de IL -6 en el plasma se correlacionaron con la severidad del choque y el resultado
clínico.
El también observó un rango muy amplio de los valores de IL-6 . Los pacientes
con 40 U/ml o menos sobrevivieron mientras que el 89 % de aquellos con
concentraciones excediendo las 7500 U/ml murieron. En otros estudios la IL-6 se
correlacionó con las concentraciones en suero de las proteínas de fase aguda, como
la proteína C reactiva, haptoglobina y la a1-ácidoglicoproteína y también se
relacionaron con la fie b re .(3,7''
2.5 - QUIMIOCINAS.
Un descubrimiento reciente en el campo de las citocinas es la existencia de
una gran familia de citocinas con estructuras homologas, de aproximadamente 8 a 10
kD. Estas moléculas comparten la capacidad de estimular el movimiento de los
leucocitos (quimiocinesis) y el movimiento dirigido (quimiotaxis) y se han denominado
quimiocinas. Todas estas moléculas contienen 2 asas de disulfuro internas. Algunos
investigadores separar estos factores en dos subfamilias, ya sea, que los dos
residuos, cisteína del extremo amino terminal estén juntos (cis-cis) o separados por
un aminoácido (cis-X-cis). Estas diferencias se correlacionan con la organización de
las 2 subfamilias en grupos de genes separados.(12,21)
Las quimiocinas de la subfamilia cis - x - cis son producidas en gran parte por
los fagocitos mononucleares, así como por células tisulares (endotelio, fibroblastos) y
megacariocitos (que originan plaquetas que contienen quimiocinas almacenadas).
Estas moléculas actúan de forma predominante sobre los neutrófilos como
18
mediadores de la inflamación aguda. El miembro mejor caracterizado de esta
subfamilia es la interleucina - 8 . (12, 1)
La subfamilia cis-cis la produce en gran parte la célula T activada. Estas
moléculas actúan de forma predominante en subgrupos de células inflamatorias
mononucleares. Por ejemplo: una quimiocina llamada RANTES actúa sobre las
células T memoria CD4+ y los monocitos. Una excepción a esta generalización es la
proteína quimiotáctica del monocito -1 (MCP-1 del inglés monocyte chemotactic
protein- 1 ), una quimiocina cis-cis que actúa solo sobre los monocitos pero es
sintetizada por fagocitos mononucleres activados y células tisulares, así como por las
células T.
Las quimiocinas de las 2 subfamilias se unen a los proteoglicanos heparan
sulfato de la superficie de la célula endotelial, y pueden principalmente estimulñar la
quimiocinesis de los leucocitos que se unen al endotelio activado por citocinas, a
través de la inducción de moléculas de adhesión.
CAPITULO TRES
CITOCINAS
QUE
REGULAN
DIFERENCIACIÓN LINFOCITARIA.
LA
ACTIVACIÓN.
PROLIFERACIÓN
Y
Algunas citocinas actúan principalmente para regular la proliferación y
diferenciación de los linfocitos, y mediar la fase de activación de las respuestas
inmunitarias. La mayor parte de estas citocinas son producidas por las células T.
especialmente linfocitos T CD4+ específicos para el antigeno. Estas células T
cooperan en las respuestas inmunitarias mediadas por células en las humorales. Las
citocinas que actúan básicamente para regular a los propios linfocitos son las
interleucinas -2 , la interleucina - 4 y el factor transformador del crecimiento (TGF-3).
Las propiedades de las citocinas a comentar en este capítulo se resumen en el
cuadro siguiente...
:
CÍTQC1NA
H úm m
fU ShlTE
© f e ADA ^ É U J L A
■
Proliferación,
producción de
citocinas
Activación
Síntesis de
anticuerpos
Activa la unión al
isotipo IgE.
Innibe la activación
Proliferación
Células T
lnterleucina-2
i
14-17kD
(monómero)
Células T
Células NK
Células B
Células B
lnterleucina-4
Factor
transformador
del crecimiento
i
i
20 kD
(monómero)
14 kD
(homodlmero)
Células TCD4
Mastocitos
Células T.
fagocito
mononuclear.
Otras
Fagocito mononuclear
Células T
Células T
Inhibe la activación y
la proliferación
Inhibe la activación
Regulación de la
proliferación
Fagocito mononuclear
Otros tipos de células
■
Abreviaturas NK, agresora natural; kD. kiloDalton; Ig, inmunoglobulina
20
3.1 - INTERLEUCINA-2.
La interleucina-2 (IL-2), originalmente llamada factor de proliferación de la
célula T (TCGF del inglés, T cell growth factor), es la principal citocina responsable
de la progresión de los linfocitos T desde la fase G a la S del ciclo celular. La IL-2 es
producida por las células T CD4+ y en menor cantidad por las células T CD8+. La IL-2
actúa sobre la misma célula que la produce, es decir, funciona como un factor de
proliferación autócrino.
La IL-2 también actúa en los linfocitos T cercanos, incluidas las células CD4+ y
CD8+, y por lo tanto es también un factor de proliferación parácrino. Durante las
respuestas inmunitarias fisiológica, la IL-2 no circula en la sangre para actuar a
distancia, por lo que no se le considera factor de proliferación endocrino.
La IL-2 secretada es una glucoproteína de 14 a 17 kD codificada por un solo
gen del cromosoma 4 en los seres humanos. El tamaño heterogéneo de la proteína
madura se debe a la extensión variable de la glucosilación de un polipéptido de
aproximadamente 130 aminoácidos. La IL-2 nativa se pliega en una proteína globular
que contiene dos grupos de hélices paralelas en parejas, estando orientada cada
hoja en una leve angulación respecto a la otra. Normalmente se transcribe, sintetiza
y secreta por las células T, solo tras la activación de los antígenos. La síntesis de IL2 suele ser transitoria, con un pico precoz de secreción que se produce
aproximadamente cuatro horas después de la activación.
Las acciones principales de la IL-2 son sobre los linfocitos:
•
La IL-2 es el principal factor de proliferación autócrino de los linfocitos T, y la
cantidad de IL-2 sintetizadas por las células T CD es un determinante importante
de la magnitud de las respuestas inmunitarias que depende de las células T. La
IL-2 estimula también la síntesis de otras citocinas derivadas de las células T. A
estas otras citocinas se les conoce con el nombre de IFN-yy la linfotoxina (LT). La
incapacidad para secretar cantidades adecuadas de IL-2 se ha descrito como
causa de anergia de células T específica para el antígeno. La acción de IL-2
sobre las células T esta mediada por la unión a su receptor. Este sistema es
quizás el mejor conocido de todos los receptores de citocinas. Existen dos
proteínas de superficie celular diferentes que se unen a la IL-2 que son: IL-2Ra e
IL-2Rp.
La ll-2Ra fué la primera en identificarse, es un polipéptido de 55 kD que aparece
tras la activación de la célula T y al que se llamó en un principio antígeno Tac (por
activación T). La unión de la IL-2 a las células que expresan IL-2Ra no produce
ninguna respuesta biológica detectable.
21
La segunda proteina que se une al IL-2 “IL-2RP” tiene aproximadamente 70 a 75
kD. La afinidad de la unión por este receptor es mayor que por el IL-2Ra. La IL-2R(3
se expresa junto con un polipéptido de 64 kD llamado IL-2Ry formándose un
complejo denominado “IL-2R(3y".
La IL-2 provoca la proliferación de las células que expresan solo IL-2Rpy,
produciendo la mitad de la estimulación proliferativa máxima con la misma
concentración de IL-2 que produce la mitad de unión máxima.
Las células que expresan IL-2Ra e IL-2 RPy pueden unirse a la IL-2 con mayor
fuerza. La estimulación de la proliferación de estas células se produce a una
concentración de IL-2 igualmente baja.
La unión a la IL-2 y la estimulación de la proliferación puede bloquearse con
anticuerpos frente a IL-2Ra ó IL-2Rp, y más eficazmente mediante una combinación
de anticuerpos frente a las subunidades del receptor.
La IL-2Ra forma un complejo con la IL-2RPy, aumentando la afinidad de este
último por la IL-2, y permitiendo por tanto, producir una señal de proliferación con
concentraciones significativamente menores de IL-2. Se cree que la IL-2 se une
primero a la IL-2Ra ya que esto facilita la asociación de IL-2RPy
Las células T en reposo expresan IL-2Rpy pero no IL-2Ra y pueden ser
estimuladas por niveles elevados de IL-2. Tras la activación de las células T por el
antígeno, se expresa con rapidez la IL-2Ra, reduciendo así la concentración de IL-2
necesaria para la estimulación de la proliferación. De hecho, la propia IL-2 puede
aumentar la síntesis de IL-2Ra.
•
La IL-2 estimula la proliferación de las células NK y aumenta su función citolítica,
produciendo las también conocidas como células agresoras activadas por
linfocinas (LAK, del inglés lymphokine-activated killer). Las células NK, como las
células T en reposo, expresan IL-2Rpy y pueden ser estimuladas por niveles
elevados de IL-2. Las células NK, sin embargo no expresan IL-2Ra y, por lo tanto
no reducen la cantidad de IL-2 necesaria para estimular su proliferación. La IL-2
actúa sinérgicamente con otras citocinas, especialmente con la IL-12, para inducir
la secreción de IFN-y por las células NK.
•
La IL-2 actúa sobre las células B humanas como factor de proliferación y como un
estímulo para la síntesis de anticuerpos.
22
APCPRESENTADO
EL ANTIGENO
CONCENTRACION
DE IL-2 NECESARIA
LINFOCITO T EN
REPOSO (l_-2Rp7 bajo,
IL-2Ra ausente)
PARA LA RESPUESTA
PROLIFERATIVA.
1 x 10_9M
ÜNFOCITO T
PARCIALMENTE
ACTIVO <IL-2R0y alto,
IL-2Ra bajo)
1 x 10U M
Estimulación I
airtocrina/paracrina I
por IL-2 I
LINFOCITO T
TOTALMENTE
ACTIVO (IL-2RPY alto,
IL-2Ra alto; síntesis
aumentada de IL-2))
h, IL-2RP7
■“ *.- 1 x 10"
^
M IL-2
IL-2R a
K .-J x TO'* M IL-2
EDd
1 I i o 15m
COMPLEJO lL-2Rpy, IL-2Roc,
HL-1 x 10” M IL-2
IL-2
El receptor de alta afinidad de la IL - 2 (IL - 2R) se compone de un complejo de dos pollpéptldos
separados (IL - 2R (3ye IL - 2R a) que Interaccionan para unirse a la IL - 2 con alta afinidad. Las
células T en reposo expresan solo IL - 2R fty, que se une a la IL - 2 con baja afinidad.
La activación de la célula T por el antígeno y una célula presentadora (ADC) conduce a la síntesis y
expresión de IL - 2R a, aumentando la afinidad de receptores IL - 2R Pyy permitiendo la estimulación
de la proliferación a concetraciones de IL - 2 fisiológica. La IL - 2 producida por la célula T activada,
aumenta aún mas la expresión de IL - 2 R a y estimulan la síntesis de IL - 2, proporcionando un
sistema de amplificación positivo.
23
3.2 - INTERLEUCINA-4.
La interleucina 4 (IL-4) se identificó inicialmente como una citocina derivada de
la célula T de aproximadamente 20 kD y su principal función fisiològica es la
regulación de las reacciones alérgicas. La IL-4 es un miembro de la familia de las
citocinas helicoidales, y su receptor es una proteína de 130 kD. Las principales
fuentes celulares de IL-4 son los linfocitos T CD4+. Los mastocitos y los basófilos
activados, como algunos T CDa+, también son capaces de producir IL -4 .(5,20)'
La IL-4 tiene importantes acciones sobre varios tipos de células:
I)
La IL-4 es necesaria para la producción de Ig E, y es la principal citocina de
switching ó cambio de células B a este isotipo de cadena pesada. La Ig E es el
principal mediador de las reacciones de hipersensibilidad inmediata
(alérgicas), y se cree que la producción aumentada de IL-4 es central para el
desarrollo de reacciones alérgicas. Los anticuerpos Ig E también actúan en la
defensa frente a las infecciones con helmintos, siendo esta una importante
función. Los ratones con el gen de la IL-4 anulado no producen Ig E. La IL-4
también inhibe el switching a Ig G2a y a Ig G3 en los ratones, que están
potenciando en cambio por el IFN-a. Esta es una de las diversas acciones
antagónicas recíprocas de la IL-4 y el IFN-y.
II)
La IL-4 inhibe la activación del macròfago y bloquea la mayor parte de los
efectos activadores sobre el macròfago del IFN-y incluida la producción
aumentada de citocina como la IL-1, el óxido nítrico y las prostaglandinas.
III)
La IL-4 es un factor de proliferación y de diferenciación de las células de un
subgrupo llamado TH2. La IL-4 promueve el desarrollo de las células TH2
secretoras de IL-4 + IL-5, a partir de células T nativas estimuladas por el
antígeno. De este modo, los estímulos que favorecen la producción de IL-4
precozmente tras la exposición al antígeno, favorecen la aparición de las
células TH2. La IL-4 también actúa como factor de proliferación autócrino para
las células T H2, promoviendo más la expansión de este subgrupo. Los ratones
que carecen del gen de la IL-4 muestran un déficit en el desarrollo y
mantenimiento de las células TH2, incluso tras estímulos (como las infecciones
por helmintos que suelen ser potentes inductores de este grupo.)
IV)
La IL-4 estimula la expresión de ciertas moléculas de adhesión, sobre todo de
las moléculas de adhesión celular vascular 1(VCAM-1) sobre las células
endoteliales, lo que produce una mayor unión de los linfocitos, monocitos y
especialmente los eosinófilos.Las células endoteliales tratadas con IL-4
también secretan la proteína quimiotáctica de monocitos 1 (MCP-1, del inglés,
monocyte chemotactic protein- 1 ), y una quimiocina todavía no definida que
actúa específicamente sobre los eosinófilos. Como resultado de ello,
concentraciones locales elevadas de IL-4 inducen reacciones inflamatorias
ricas en monocitos y eosinófilos.
24
V)
La IL-4 es un factor de proliferación para los mastocitos y actúa
sinèrgicamente con la IL-3 en la estimulación de la proliferación de los
m astocitos.(5,20)
De este modo, la IL-4 desempeña una función crítica en las reacciones
inflamatorias medidas por la Ig E y los eosinófilos. En la actualidad se están
probando antagonistas de la IL-4 en pacientes para controlar reacciones alérgicas
graves.
3.3 - FACTOR TRANSFORMADOR DE CRECIMIENTO (5
La descripción original del factor transform ador del crecim iento [3 se hizo en el
campo de la Biología Tumoral. Se observó que ciertos tumores producían llamados
factores transformadores del crecimiento que permitirían a células normales proliferar
en agar blando, una característica de las células malignas (transformadas).
Posteriormente se observó que la estimulación de la proliferación estaba causado
por un polipéptido llamado TG Fa (transforming growth factor a); este factor es un
polipéptidico de proliferación para las células epiteliales y mesenquimales y no entra
dentro del grupo de las citocinas.(3)
Existe un segundo factor que se requería en la supervivencia en el agar
blando al que se le llamó TGF(3 el cual es una familia de moléculas muy
relacionadas, codificadas por genes diferentes, que se clasifican en TGF-(31, TGF-J52
y TGF-P3. Las células del sistema inmunitario (por ejemplo, las células T y los
monocitos) sintetizan principalmente TGF-(31, aunque ciertas localizaciones
anatómicas (por ejemplo, el SNC) puede contener cantidades elevadas de TGF-p3.
El TG F-pi nativo es una proteína homodímera de aproximadamente 28 kD y se
sintetiza en una forma latente que debe activarse por medio de proteasas.
Sintetizan
Células T y
monocitos
*
Principalmente
TGF-p1
Se activa
Inhibe la proliferación
__________ ^ de células.
Por proteasas
Tanto las células T activadas por el antígeno como los fagocitos
mononucleares activados por el LPS secretan TG F-pi activo. Los receptores del
TGF-p1 no están bien definidos, aunque una de las proteínas que se unen al TGF-p
puede ser una serina /treonina cinasa.
25
Las acciones del TGF-p son muy pleiotrópicas. Este inhibe la proliferación de
muchos tipos de células y estimula la proliferación de otras. A menudo el TGF-p
puede inhibir o estimular la proliferación del mismo tipo de célula, dependiendo de
condiciones del cultivo como el grado de confluencia.
El TGF-p produce la síntesis de proteínas de la matriz extracelular como las
colágenas, y de receptores celulares para las proteínas de la matriz. La capacidad
del TGF-p de inducir la matriz extracelular probablemente está relacionada con su
capacidad de promover la proliferación celular del agar blando. In vivo, el TGF-p
estimula la proliferación de vasos sanguíneos nuevos, un proceso llamado
angiogénesis.
Como citocina, el TGF-p es potencialmente importante, porque...
•
Antagoniza muchas respuestas de los linfocitos. Por ejemplo: el TGF-p inhibe la
proliferación de la célula T frente a mitógenos policlonales ó en cultivos mixtos de
leucocitos e inhibe la maduración de los LTC.
•
Inhibe la activación de los macrófagos.
•
Actúa sobre otras células, como los leucocitos polimorfonucleares y las células
endoteliales, de nuevo antagonizando los efectos de las citocinas
proinflamatorias.
Aunque el TGF- es un regulador en gran medida negativo de las respuestas
inmunitarias, puede tener efectos positivos, ya que se encarga de generar
respuestas inmunitarias en algunas mucosas.(3,5)-
VA*
26
CAPITULO CUATRO.
CITOCINAS QUE REGULAN LA INFLAMACIÓN DE ORIGEN INMUNITARIO.
La inflamación es un estado que se produce en el organismo, debido a la
respuesta natural defensiva de los tejidos contra estímulos nocivos, sean físicos,
químicos o bacterianos. Se caracterizan por ingestión, aumento local de temperatura,
edema, dolor y alteración temporal de la funcionalidad de la parte afectada. La
inflamación que acompaña a menudo al proceso de reconstrucción de los tejidos
dañados, sirve también para limitar la expanción de una infección, construyendo una
barrera alrededor; aunque esta barrera puede convertirse en un obstáculo durante el
tratamiento.(21)
La inflamación incluye una serie de cambios celulares que facilitan finalmente
la fagocitosisy ya sea la destrucción de microorganismos ó la digestión de desechos
celulares. Esto es seguido por la proliferación de las células del tejido conectivo y la
reparación de la matriz intracelular. Ambos sucesos requiren el control complejo de
varios tipos de células a un nivel local. La evidencia aumentada indica que, aunque
otras moléculas puedan ser de efectos mediadores tales como la permeabilidad
vascular y la quimiotaxis, las citocinas ejercen control de una forma muy sofisticada
con la duplicación remarcable de elementos clave y la interacción compleja negativa
y positiva entre sí y con otras hormonas.(12,21)
Los procesos locales de inflamación son facilitados por los cambios
metabólicos sistémicos que movilizan la energía en forma de glucosa ácidos grasos y
bloques de construcción de aminoácidos. La fiebre sobreregula varias reacciones
enzimáticas en las células inflamatorias y de tejidos que tal vez al ser potencialmente
peligrosos, operan a un índice suboptimal en la temperatura normal del cuerpo. Los
efectos contraregulatorios tales como la liberación de cortisol entra en efecto y
detienen los efectos potencialmente destructivos en la inflamación. Todos estos
sucesos son en parte, controlados por las mismas citocinas pleiotrópicas que ejercen
controles locales claves.(12,21)
En este capítulo se comentará a este grupo de citocinas que son derivadas
principalmente de los linfocitos T CD4+ y CD8+ activados por el antígeno, que sirven
básicamente para activar las funciones de las células efectoras ¡nespecíficas. De
este modo, estas citocinas actúan en la fase efectora de las respuestas inmunitarias
mediadas por células. Las moléculas a describir en esta sección se resumen en la
siguiente tabla...
FÜEfsTE
ru a, fUAllíL
POt-iPcPTlOO
......
Interferón-y
1
21-24 kD
(homodímero)
Linfotoxina
1
24 kD
(homodímero)
lnterleucina-10
1
lnterleucina-5
1
20 kD
(homodímero)
20 kD
(homodímero)
lnterleucina-12
2
35-40 kD
(heterodímero)
Factor
de
inhibición de la
migración.
?
?
.....
Célula T
Célula NK
.......
, ...... '
EFECTOS BASICOS EN . í
..............
F.M.
Cél. endotelial
Célula NK
Todas.
Activación
Activación
Activación
Aumento de moléculas
de clase 1y II del MHC
Célula T
Neutrófilo
Activación
Cél. endotelial Activación
Célula NK
Activación
CéluIaT
F.M.
Inhibición
Célula B
Activación
Célula T
Eosinófilo
Activación
Célula B
Proliferación y
activación
Macròfago Células NK
Activación
Células T
Activación (proliferación
y diferenciación)
Célula T
F M:
Conversión de estado
de movilidad al de
inmovilidad
Abreviaturas NK. agresora natura!, KD. kiloDalton. MHC, complejo mayor de histocompatibilidad ;F M .fagocito mononuclear
4.1 - IN TE R FE R O N -y.
El interferón-y (IFN-y) también denominado “interferon tipo II" es una
glucoproteína homodímera que contiene subunidades de aproximadamente 21 a 24
kD El tamaño variable de las subunidades se debe a los grados variables de
glucosilación, pero cada subunidad contiene un polipéptido idéntico de 18 kD
codificado por el mismo gen.(24)
El IFN -7 lo produce tanto células T cooperadoras CD4" nativas como T CD°".
La transcripción se inicia directamente como consecuencia de la activación del
antígeno y está aumentada por las IL-2 e IL-12. El IFN-y también lo producen las
células agresoras naturales (NK), que son la principal fuente de esta citocina en los
ratones que carecen de células T; en esta situación el IFN-y puede actuar como un
mediador de la inmunidad natural.
Como su nombre implica, el INF-y comparte muchas actividades con el IFN tipo I
De forma específica el IFN-y, induce a un estado antiviral y es antiproliferativo No
obstante, el IFN-y se une a un receptor de superficie celular único, diferente aunque
estructuralmente relacionado con el que utiliza el IFN tipo I. Lo más importante, es
que el IFN-y tiene varias propiedades relacionadas con la inmunorregulación que las
separan funcionalmente del IFN tipo l .(3 24)
28
•
El IFN-y es un potente activador de los fagocitos mononucleares. Induce
directamente la síntesis de enzimas que median el respiratory burs't (en español,
estallido respiratorio, aludiendo al metabolismo oxidativo que generan radicales
tóxicos), que permite a los macrófagos matar a los microorganismos fagocitados.
Junto a segundas señales, como el LPS y quizás el TNF, permite a los
macrófagos matar células tumorales. A las citocinas que producen cambios en los
fagocitos mononucleares se les llama “factores activadores de macrófagos” (MAF,
del inglés macrophage, activating factors). El IFN-y es el principal MAF y es el
medio por el que las células activan a los macrófagos. Es importante señalar
aquí, que la activación del macròfago implica en realidad varias respuestas
diferentes, y se dicen que los macrófagos están activados cuando realizan una
función particular que se mide. Por ejemplo, el IFN-y activa completamente a los
macrófagos para que maten microorganismos fagocitados pero solo parcialmente
para que maten células tumorales.
•
El IFN-y aumenta la producción de moléculas de clase I del MHC y al contrario
que el IFN-y tipo I, también provoca la expresión de moléculas de clase II de MHC
con diversas células. De este modo, el IFN-y amplifica la fase de reconocimiento
de la respuesta inmunitaria promoviendo la activación de las células T
cooperadoras CD4+ restringidas por la clase II. In vivo, el IFN-y puede aumentar
las respuestas inmunitarias humorales y celulares a través de acciones en la fase
de reconocimiento
•
El IFN-y actúa directamente sobre los linfocitos T y B para promover su
diferenciación. IFN-y promueve la diferenciación de la células T CD4+ nativas e
inhibe la proliferación de otras células. También es una de las citocinas
necesarias para la maduración de los LTC CDa+. También actúa sobre las células
B para promover el =switching= a las subclases lgG2a e lgG3 en ratones, e
inhibir el =switching= a lgG1 e IgE.
•
El IFN-y activa a los neutrófilos, activando su =respiratory burs't=. Es un activador
de neutrólitos, menos potente que TNF ó la linfotoxina.
•
El IFN-y estimula la actividad citolítica de la células NK, más que el IFN tipo I.
•
El IFN-y es un activador de las células endoteliales vasculares, promoviendo la
adhesión de la linfocitos T CD4+ y los cambios morfológicos necesarios para
facilitar su extravasación. Como se mencionó antes, el IFN-y también potencia
muchas de las acciones del TNF sobre las células endoteliales.*3'2124>
El efecto neto de estas actividades del IFN-y es promover las reacciones
inflamatorias ricas en macrófagos, a la vez que suprimen las reacciones ricas en
eosinófilos. Los ratones en los que se han anulado los genes del IFN-y o de sus
receptores muestran varios defectos inmunitarios, incluida la mayor susceptibilidad a
las infecciones de microorganismos intracelulares (que no pueden eliminarse por el
fallo en la activación de los macrófagos), la producción reducida de ácido nítrico en
29
los macrófagos procedentes de ratones infectados por micobacterias, la
concentración sérica reducida de anticuerpos lgG2a e lgG3, la expresión reducida de
moléculas de clase II del MHC en los macrófagos de ratones infectados por
micobacterias, y fallos en la función de la célula NK. ^ 1,24>
4.2 - LINFOTOXINA
La linfotoxina es una glupoproteína de 21-24 kD que presenta una homología
de aproximadamente un 30% con el TNF, y compite con él por la unión a los mismos
receptores celulares (como ya se mencionó, la LT se denomina a veces TNF-(3). En
los seres humanos, los genes de la LT y del TNF se localizan dentro del MHC del
cromosoma 6 .
La LT la producen exclusivamente los linfocitos T activados, y a menudo se
produce junto al IFN-y en tales células. La LT, humana, al contrario que la TNF,
contiene 1 ó 2 oligosacáridos unidos por enlaces N (responsables de la variedad del
tamaño molecular). Además, la LT se sintetiza como una verdadera proteína
secretora sin región transmembranosa. Recientemente se ha descrito un tercer
miembro de la familia TNF/LT. El producto genético llamado provisionalmente LT-(3
es una proteína de superficie celular que se une a la LT para formar un complejo de
superficie que media los efectos de la LT sobre otras células.
La mayor parte de los estudios han encontrado pocas diferencias entre los
efectos biológicos del TNF y de la LT, compatible con su unión al mismo receptor. La
más importante distinción entre estas citocinas parece ser que la LT la sintetizan solo
las células T, deriva predominantemente de los fagocitos mononucleares. En
general, las cantidades de LT, sintetizadas por las células T son mucho menores que
la de TNF sintetizadas por los fagocitos mononucleares estimulados por el LPS. Por
lo tanto, la LT habitualmente actúa como un factor parácrino y no como mediador de
lesión sistèmica. Aunque ni el TNF ni la LT son tóxicos para las células normales (no
neoplásicas), ambos pueden participar en la lisis de células diana mediada por los
LTC.<5'9)
Como el TNF, la LT es una activadora potente de neutrófilos y de este modo
proporciona a los linfocitos un medio de regular las reacciones inflamatorias agudas.
Es un activador de neutrófilos más potente que el IFN-y, y este último potencia las
acciones de la LT.
La LT es también un activador de las células endoteliales vasculares
aumentando la adhesión leucocitaria, la producción de citocinas y los cambios
morfológicos que facilitan la extravasación de los leucocitos. Estos afectos como los
del TNF, son también potenciados por el IFN-y.(5,9)
30
4.3 - INTERLEUCINA - 10.
La ¡nterlucina 10 (1L- 10) es una citodna de 18 kD producida por el subgrupo
Th2 de las células cooperadoras CD4+. También es producida por algunas células B
activadas, por algunas células Th 1 (en seres humanos), por macrófogos activados y
por algunas células no linfocíticas (por ejemplo, los queratinocitos). La IL-10 es un
miembro de la familia de las citocinas a-helicoidales, y probablemente actúa en
forma de homodímero.
Las 2 actividades principales de la IL-10 son inhibir la producción de citocinas
(es decir, TNF, IL-1, quimiocina e IL-2) por los macrófagos, e inhibir las funciones
accesorias de los macrófagos en la activación de la célula T. Este último efecto se
debe a la expresión reducida de moléculas de clase II del MHC, y a la expresión
reducida de ciertos coestimuladores. El efecto neto de estas acciones es inhibir la
inflamación mediada por la célula T. Además de sus efectos inhibidores sobre los
macrófagos B, la IL-10 tiene acciones estimuladoras sobre las células B. Puede ser
un factor de =switching= para la producción de lgG4 en seres humanos (homólogo a
la IgG 1 en ratones).
Estudios en ratones con el gen de la IL-10 anulado por una cierta tecnología,
revelan pocas alteraciones inmunitarias. Estos ratones presentan lesiones
inflamatorias intestinales, de origen desconocido. Es interesante resaltar que el
genoma del virus de Epstein-Barr contiene un gen homólogo al de la IL-10, y que la
IL-10 viral comparte actividades in vitro con la citocina derivada de la célula T. Esto
plantea la posibilidad intrigante de que el virus haya adquirido el gen humano como
un medio de inhibir la inmunidad an tiviral.(3,5)
4.4 - INTERLEUCINA- 5 .
La ¡nterlucina-5 (IL-5) es una citocina homodimérica de aproximadamente 40
kD producida por el subgrupo TH2 de las células T CD4+ y por los mastocitos
activados. Pertenece a la familia de citocinas de 4 hélices, aunque cada haz de 4
hélices consta de 3 tramos de un monómero y uno del otro.
La principal acción de la IL-5 es estimular la proliferación y diferenciación de
los eosinófilos y activar a los eosinófilos maduros para que puedan matar helmintos.
En los ratones, los anticuerpos neutralizantes frente a la IL-5 inhiben la eosinofilia
que se vé en respuesta a la infección por helmintos. La actividad de la IL-5 se
complementa con las de la IL-4 (por ejemplo el =swtching= a IgE y el reclutamiento
de eosinófilos) y las de la IL-10 (por ejemplo el =swtching= a lgG4), contribuyendo a
las reacciones alérgicas mediadas por el T h2.
La IL-5 también actúa como un coestimulador para la proliferación de las
células B de ratón activadas por el antígeno y previamente se le denominó factor del
crecim iento de la célula B ó factor reem plazador de la célula T. La IL-5 puede actuar
31
sinérgicamente con otras citocinas, como la IL-2 y la IL -4 para estimular la
proliferación y diferenciación de las células B. También la IL-5 actúa sobre las células
B maduras para aumentar la síntesis de inmunoglobulinas, especialmente de Ig A .(3,5)
4.5 - INTERLUCINA - 12.
La interleucina -1 2 es un heterodímero de 70 kD que consta de 2 cadenas
polipépticas unidas de forma covalente, una de 35 kD (p35) y otra de 40 kD (p40). La
subunidad p35 la producen muchos tipos de células, incluidos los linfocitos T y B, las
células NK y los monocitos. La cadena p40 es producida principalmente por
monocitos activados y células B, por lo que estos son las fuentes principales de la
citocina completa (en esta categoría funcional de citocinas, la IL-12 es la única que
no producen las células T, y por su acción por las células NK, puede ser considerada
un mediador de la inmunidad natural).
La proteína p35 tiene una estructura de 4 hélices a, similar a la de otras
citocinas. Es interesante señalar que el componente p40 de la IL-12 es homólogo al
del receptor de la IL-6 . De este modo el heterodímero intacto parece estar
compuesto de una proteína similar a una citocina y de una proteína similar a un
receptor de citocinas. Estudios de la unión indican que el verdadero receptor de la IL12 se expresa en las células T y NK activadas, pero todavía no se ha caracterizado
completamente.
La IL-12 es un importante regulador de las respuestas inmunitarias mediadas
por células y por sus efectos sobre células NK y linfocitos T.
1) La IL-12 es el más potente activador conocido de la célula NK. Induce la
transcripción de IFN-y por las células NK, y muestra fuerte sinergia con la IL-2.
Además de estimular la producción de IFN-y, la IL-1 2 aumenta la actividad
citolítica de las células NK y es un factor de crecimiento para estas células.
2) La IL-12 estimula la diferenciación de las células T CD4+ nativas al subgrupo T H1
frente a una TH2, puede controlarse por la producción relativa de IL-12 e IFN-y,
que favorece la diferenciación a TH1. y de IL-4 e IL- 1 0 , que promueve la
diferenciación a T h2.
3) La IL-12 estimula la diferenciación de las células T CD8+ a LTC activos y maduros.
Debido a este efecto, la IL-12 puede ser un tratamiento potencial del cáncer
diseminado.
De este modo, la IL-12 es un importante regulador de la fase efectora de las
reacciones inmunitarias mediadas por células. Sirve a esta función activando
directamente a algunas células efectoras, y regulando el desarrollo de otras.(4,6)
32
Por otro lado, desde su descubrimiento al final de los 80 s, la potente citocina
IL-12 ha disfrutado las trampas de una molécula caliente. Su rol crucial para darle
forma a los momentos más tempranos de la respuesta inmune ha llevado a una
explosión de artículos en los periódicos inmunológicos. Como droga, esta ya ha
entrado en los juicios clínicos humanos en contra del cáncer y el sida. Los
investigadores recientemente otorgaron el sobrenombre de bala mágica a la IL-12 a
causa de su poder para inhibir a los vasos sanguíneos que alimentan a los tumores
en los experimentos naturales, poderes anotados en el Wall Street Journal.
Recientemente fué el tumo de los expertos en enfermedades infecciosas en
los Institutos Nacionales de Salud, quienes patrocinaron los encuentros para exaltar
a la IL-12 como un agente terapéutico potencial y una vacuna adyuvante. Los
representantes de las compañías farmacéuticas dejaron claro en el encuentro que
aunque la IL-12 ha mostrado una promesa en contra de las enfermedades
infecciosas prevalentes en los países en desarrollo, las presiones económicas
sugieren que esta probablemente será desarrollada primero para mercados
lucrativos cercanos.
Debemos remarcar que desde 1989, cuando apareció el primer reporte en la
literatura, la IL-12 ha demostrado ser producida por los monocitos, macrófagos,
neutrófilos y las células dendríticas (las primeras células en encontrar un antigenó
extraño durante una infección) tanto como las células B que producen anticuerpos.
La IL-12 a su vez activa a las células destructoras naturales y las células T y parece
particularmente potente en su habilidad para inducir la producción del IFN-y La
combinación del IFN-y y la IL -12 envía un poderoso signo hacia las células
precursoras sencillas del lineaje auxiliar T, cambiando una respuesta inmune no
duplicable hacia la inmunidad mediada por las células, una de las dos grandes
extremidades de la inmunidad adquirida (la otra siendo una forma humoral ó
asociada con el anticuerpo). (4’6’15)
4.6 - FACTOR DE INHIBICION DE LA MIGRACION
El factor de inhibición de la migración (MIF, del inglés, migration inhibition
factor) es otra de las citocinas que regulan a las células efectoras. Una de las
primeras investigaciones de las reacciones inmunitarias mediadas por células
propuso que la acumulación del fagocito mononuclear en los tejidos dependía de la
retención de tales células en respuesta a citocinas producidas a nivel local, que
inhibían la motilidad.
Ahora parece más probable que la retención de los leucocitos en los tejidos
esté controlada básicamente por la expresión de receptores específicos para las
moléculas de la matriz extracelular. Sin embargo, una de las principales actividades
de citocina que se identificó, fué una que inhibía la motilidad de macrófagos in vitro
llamada Factor de Inhibición de la M igración.(3,5)
33
CAPITULO CINCO.
CITOCINAS QUE ESTIMULAN LA HEMATOPOYESIS.
Algunas de las citocinas generadas durante la inmunidad natural y las
respuestas inmunitarias inducidas por antígenos específicos, tienen potentes efectos
estimuladores para la proliferación y diferenciación de las células progenitoras de la
médula ósea. De este modo, las reacciones inmunitarias e inflamatorias, que
consumen leucocitos, también estimulan la producción de leucocitos nuevos que
reemplacen a las células inflamatorias.
Todas las poblaciones de leucocitos maduros surgen a consecuencia de una
expansión progresiva, y de una diferenciación irreversible de la progenie de células
madre pluripotenciales autorrenovables.
La maduración de las células implica el compromiso de una línea particular, y
ocurre junto a la pérdida de la capacidad de evolucionar hacia otros tipos de células
maduras. Este proceso se ha representado como un árbol.(2,3)
Las citocinas que estimulan la expansión y diferenciación de las células
progenitoras de la médula ósea, se denomina Factores Estim uladores de Colonias
(CSF del inglés, colony- stimulating factor), porque se suelen analizar en función de
su capacidad para estimular la formación de colonias celulares de los cultivos de
médula ósea. Estas colonias de células maduran durante el análisis in vitro,
adquiriendo características de líneas de células específicas (por ejemplo,
granulocitos, fagocitos mononucleares).
Diferentes CSF actúan sobre células de la médula ósea en diferentes estados
de maduración y promueven de forma preferencial el desarrollo de colonias de
diferentes líneas. Todo esto se lleva a cabo como se presenta en la siguiente
figura....
34
MADURACION DE LAS CÉLULAS SANGUINEAS.
CELULA MAORE f
a u t o -r e n o v a b l e
y
\
1Ligando da c-kit 1
CELULAMADRE
PROGENITORA UNFOIDE
Pl.URIPOTENCIAL
|
L _ -i IL-7, otras
TIMO; IL-2.
1------ ^ ------1 IL-4. IL ^ . o u a s j
PROGENITORA
MIELOIDE
P 'lL-3 . GM-CSF: IL-1 .IL-8 ~l
V
*
| e r it r o p o y e t in á ]
UNFOCITOSB
CFU eritroide
Megacariocito
CFU de
basófilos
I erítropoyéiina}
CFU de granulocítos-monocilos
CFU de
eosinófilos
m
/
llt -3 GM-CSF. G-CSf I
T
r
* •
•
ERITROCITOS
PLAQUETAS
BASOFILOS
EL ARBOL HEMATOPOYETICO
EOSINOFILOS
NEUTROFILOS
'
{IL-3 GM-CSF,
o
MONOCITOS
La m a d u r a ^ c t ó l a s
o n .a s T I n t e r l e u a n a ; ' e s t e l a n t e
cró fago s.(2,312>.
UNFOCITOST
de colonias de granuloatos -
Los nombres asignados al CFS reflejan los tipos de colonias que surgen en
estos análisis. Es interesante señalar que muchos de los CFS se localizan en un
grupo de genes en el cromosoma 5 humano, incluyendo a la IL-3 y el G M -CbK La
IL-4 y la IL-5 se han localizado en este mismo complejo.
Algunas de las acciones de los CSF están influidas por otras atocinas Por
ejemplo, el TNF, la LT, el IFN-y y el TGF-p inhiben la proliferación de las células
progenitores de la médula ósea. Por el contrario, la IL-1 y la IL-6 aumentan la
respuesta a los CSF
En general, se cree que las citocinas son necesarias para la normal función de
la médula ósea y para proporcionar un medio de ajuste fino en respuesta a la
estimulación. Algunos de los CSF específicos se enumeran a continuación
;s
A A ^ 'r it
i
C E L U L A O lA N A
C E L U L A f?
Activación
lntertucina-3
CSF
granulocitosmacrófagos
24 Kd (monómero)
1
Ligando de c- KIT
CSF de granulocitos
lnterteucina-7
Célula
madre
pluripotencial
Progenitora
inmadura
Proliferación
y
diferenciación
de
todas
las
lineas
celulares.
1
22 Kd
(dimero)
1
22 KD
(dimero)
Célula T, fagocito
mononuclear Célula
endotelial. fibroblasto
Progenitora
inmadura
diferenciación
de 1
todas
las
lineas
celulares.
1
40kD
(dimero)
Fagocito.
mononuclear,
endotelial
Progenitora
inmadura
diferenciación
de
todas
las
lineas
celulares
1
19 kD
(dimero)
1
25 kD
(monómero)
célula
célula
Progenitora
inmadura
Proliferación
i diferenciación
todas
las
celulares
Fibroblasto,
células
estromales
de
la
médula ósea
Progenitora
inmadura
Proliferación
diferenciación
linfocito.
Fagocito,
mononuclear,
endotelial
Abreviaturas. CSF. ftjet or estimular te de colonias. kD kilodalton
-------------------------------
de
Célula T. fagocito
mononuclear Célula
endoteliai, fibroblasto
de
CSF de macrófagos
Célula estromal
médula ósea T
y!
de
lineas
y
a
36
5.1 - LIGANDO DE c-KIT
La célula madre pluripotencial expresa un receptor de tirosín cinasa en la
membrana que se ha identificado como el producto proteico del oncogén celular cKIT.
La porción extracelular de este receptor contiene 5 dominios de Ig. A la citocina
que interactúa con este receptor se le ha llamado Ligando de c-K IT y también se le
denomina factor de la célula madre.
El ligando de c-KIT se sintetiza en las células estromales de la médula ósea
(incluidos los adipocitos. Los fibroblastos y las células endoteliales) en dos formas:
•
•
Proteína transmembranosa de aproximadamente 27 kD
Proteína secretada de aproximadamente 24 kD.
Estos productos diferentes son el resultado de la religación alternativa del
mismo gen. La forma soluble de este ligando no existe en una cepa de ratones
mutantes llamada Factor A cero}3,5)
El ratón con el factor acero tiene huecos selectivos en la poblaciones celulares
derivadas de su médula ósea (por ejemplo, una producción inadecuada de
mastocitos y eosinófilos), que se llevado a la conclusión de que la forma superficial
celular del ligando de c-KIT es más importante que la forma soluble, para la
estimulación de la maduración de las células madre a varias líneas hematopoyéticas.
Todavía no es posible purificar un número elevado de células madre para su
análisis directo. Muchas de las conclusiones sobre el ligando de c-KIT y otros CSF
de acción precoz, derivan de experimentos en los que se exponen poblaciones
enriquecidas con células madre a citocinas en cultivo, y se analizan las tipos de
colonias que surgen. A partir de este tipo de experimentos, se cree que el ligando de
c-KIT es necesario para que la célula madre sea sensible a otros CSF, pero este no
produce la formación de colonias por sí solo.
Los cultivos de células de médula ósea que contienen células estromales no
necesitan un aporte exógeno del ligando de c-KIT ya que estas células no lo
expresan.(3)-
37
5.2 - INTERLEUCINA - 3
La interleucina 3 (IL-3) también conocido como factor estim ulante de colonias
de m últiples líneas (multi-CSF) es un producto de 20 a 26 kD de las células T CD4+
que actúa sobre la mayor parte de los progenitores de la médula ósea inmaduros, y
que promueve la expansión de células que se diferencian en todos los tipos de
células maduras conocidos. La IL-3 es un miembro de la familia de citocinas de 4
hélices. En el ratón la subunidad transductora de señales es única. La mayor parte
de los análisis funcionales de la IL-3 se han hecho en ratones.
Se ha encontrado que la IL-3 también promueve la proliferación y el desarrollo
de los mastocitos a partir de progenitores de médula ósea, una acción aumentada
por la IL-4. La IL-3 es producida por las células T cooperadoras CD4+ de los
subgrupos TH1 yTH2. La IL-3 humana se ha identificado mediante la clonación de
DNA complementario (DNAc) de una molécula homologa a la IL-3 de ratón.
Aunque la citocina 3 es sintetizada por algunos clones de células T humanas,
ha sido más arduo establecer la función de esta citocina en sistemas experimentales
de hematopoyesis en seres humanos. De hecho, muchas acciones atribuidas a la IL3 múrida parece que las realiza el CSF de granulocitos macròfago humano.
No se sabe si estos resultados experimentales reflejan diferencias entre
especies ó en la condiciones experimentales. Si se trata de una diferencia entre
especies, puede relacionarse con la diferente estructura del receptor de la IL-3.(3,11)
5.3 - FACTOR ESTIMULADOR DE COLONIAS DE GRANULOCITOSMACROFAGOS.
El factor estimulador de colonias de granulocitos macrófagos (GM-CSF) es
una glucoproteína de 22 kD sintetizada por las células T activadas y por los fagocitos
mononucleares activados, las células endoteliales y los fibroblastos.
El GM-CSF es un miembro de la familia de citocinas de 4 hélices. Su receptor
consta de una subunidad única y una subunidad transductora de la señal 150 KD.
Como se dijo antes, esta última subunidad la comparte el receptor de la IL-5, y en los
seres humanos pero no en los ratones, el receptor de la IL-3.
El GM-CSF del ratón actúa básicamente en los progenitores de la médula
ósea ya comprometidos en el desarrollo a leucocitos, y por ello actúa sobre
poblaciones más diferenciadas que la IL-3. En los sistemas humanos, sin embargo,
el GM-CSF también promueve la proliferación de células que todavía no se han
comprometido en evolucionar a leucocitos (por ejemplo, plaquetas y progenitores de
hematíes) reemplazando a la IL-3. El GM-CSF también activa a leucocitos maduros.
Por ejemplo, imita a algunas de las acciones del IFN-y como activador de
macrófagos, aunque es menos potente.
38
El GM-CSF no se detecta en la
pertérícos el GM-CSF
en los lugares en que se produc®_°® ®
pueden intervenir principalmente en la
nroducido por la células T y macrofagos p«teoen
respuestas inmunitanas
activación Se los leucocitos maduros e n ^ " / é . f c o s pueden estar mediados por
inflamatorias,
“ T |as células endoteliales ó los fibroblastos de la
el GM-CSF producido por la células i ,
médula ósea.
a He
Se ha utilizado
para e s S a /l^ r e T u p e r a c ió n de la
-
- —
e de 13 médu'a ~
5 .4 . f a c t o r e s t im u l a d o r d e c o l o n ia s d e m o n o c t o s - m a c r o f a g o s
El factor estimulador de co|0™a*
Sétulas endoteliales y los
£
llamado CSF-1. es sintetizado.por'a ^ " ^ a d a m e n t e 40 kD i * ™ * ™
fibroblastos. El polipetido secretado
re(aciona estructuraímente con el c-KIT.
celular normal del oncogen viral v-fms.
Fl M-CSF actúa básicamente sobre aquellos
en el desarrollo a monocitos, y
la cédulas sobre
&e\M C SF no circula, y el principal efecto
—
,oca' den,r° de *
medular.(5)
FACTOR ESTIMULADOR DE COLONIAS DE GRANULOCITOS.
55
'
E1 fa c o r estimulador de colones
las
mismas
células g ue
° £
C le m e n te forma un dímero. El G-C^ f ^
de'°a fe m to de citocinas de 4 hélices a. Al contrano que otros C
normalmente circula.
Actúa básicamente sobre
i r ^ a r r o llo a granulocitos, de nuevo
P
. . .
15.7)
médula .15,71
^
Un^ Q . CSF
„
comprometidos en
pob|aclón más madura que la que
P
Duecie actuar a distancia, la
5 6 - INTERLEUCINA
IN i tn L c u w n « »- 7■
La interleucina - 7
B T fm a y o rep a rte d rio semodelos de " S ^ c o m
—desarrollo
n que la IL-3 o el GM-CSF.
nivel de
¿
s
p
o ia
Estudios
recientes sugieren ^
: »
r o
b
, a m
e n
t e
s o
u y precozmente
b
r e
c é l u
, a
s
3 1
m
, s m
o
IL-7 puede también estimula
y q q 4+ y c ü 8+ inmaduras en el timo.
2 ¡: - »
rio la IL-7 en el timo.
ú n m
- “
e 81ori9e" ce,u,ar
on pxceso la IL-7 muestran un
en ,a módula ósea y en los „ i d o s
n ra rc a ^
linfoides periféricos.
s 7 . OTRAS CITOCINAS ESTIMULADORAS DE COLONIAS.
La IL-9 es una proteína de 3 ° a 4C> kD que
^
Sin embargo, no, »
~
d e ^o s mastocitos^
,ín6aS 3
« b e si
"
8U Pre° UrSOreS
'e M a ^ g u lic ió n de las
normales (diferentes e a
h matop0yesis ¡n vitro.
respuestas inmunttanas o en ,a hem
P ,
La IL-11 es un citocina de
la n z a r s e e x p e l—
^
^
-ras su activación (que puede
m e n te mediante agentes farmacologtcos).
ua IL-11 estimula la r P e g a c a n o p o y e t * t y d e X X T ' f
m a cró fa g o ^y quizás d e s o irá s H n e a s ^ ^ u la re s a p a rtirle precursores medulares.
40
CAPITULO SEIS
ALGUNOS TEMAS CLINICOS RELACIONADOS CON LAS CITOCINAS
6.1 - MIELOPOYESIS.
c n r 9r s F n MCSF v ^ N ^ ^ e c l a ^ a
proIrteraclóR1 y 'diferenciación de las
GMCSF, GCSF, MCSF y
.
sorDrendente número de factores de
células mieloides. En los humanos
aP una región sobre el brazo largo del
crecimiento mielopoyetico han :sido
^
eventos de translocalizacion o
cromosoma 5 frecuentemen
codificación de genes para GMCSF, MCSF,
desalojamiento vistas en ai euc
.
s¡do localizados en la región
IL-3, IL-4, IL-5, y el receptor celular pa a el m l
^ ,|n v |v 0 sug¡ere que estas
5P23-35. La evidencia de ambos * s,® breregu|ación de la m ie lo p o y e s is j^e n el
aumento d e M ic e de liberación de células sanguíneas hacia la circulación.
.
6 2 . ONCOLOGIA (CITOCINAS COMO FACTOR DE CRECIMIENTO AUTOCRINO)
Los sistemas experimentales indicari que 'a jr a n s .o m ja c ^
ocurrir mediante un circuito ae,oqr' ° '
Muchas lineas celulares malignas, y
ejemplo, el factor de < = ^ ' ^ 0slX t a ™ y responden a la citocinas y factores de
tumores extraídos de Pacientef ; . ^ ? ' as noS han nevado a la hipótesis de que las
crecimiento. Estas y otras cara
factores de citocina/crecimiento, y de sus
^
e
"
ee x p r e s ^ R e c e p to re s , pueden yacer bajas formas de malignidad.
Alguna evidencia indica que este m e c a m s m abtPó c rjt0 y e| sarcoma
d e ^a p o sL ^o m le "é tfa cto r decrecim iento de fibroblastos puede tomar este rol.
Matsuda ha demostrado q u e la ^
puede producir y responder a !a
como el origen de la IL-6 , pero co
pacientes con mieloma, las. f ulas
áP
f e
«
U'aha, ^
>h ^
¡madamente el 50% de los
responden a la IL -6 y que la anti IL-6
de ,a tim ,dina terciada, sugiriendo que la
C i m i e n t o a u to c h n o -
41
6.3 - PRODUCCIÓN DE CITOCINAS POR LOS TUMORES.
Ambos nodulos linfáticos, el de mixoma cardiaco y el hiperplásico de la
enfermedad de Kastleman puede producir altas cantidades de 11-6. Esto es
interesante porque ambas condiciones están asociadas con la activación de células
B policlonales, la hipergamaglobulinemia, la fiebre y la producción de proteínas de
fase aguda. En la enfermedad de Kastleman puede existir también el mieloma
múltiple. La removisión de los nodulos linfáticos involucrados resultan de la
recuperación completa.(25)
6.4 - LAS CITOCINAS EN EL TRATAMIENTO DEL CANCER.
Los interferones y el TGF-(3 tiene propiedades inhibidoras de crecimiento para
un amplio rango de tipo de células. La utilidad clínica de los IFN ha sido resumida
como = buena en la leucemia de células velludas, regular en otros cánceres
hematólogicos y mala en los tumores sólidos =.
La IL-2 ha sido infundida en pacientes con cáncer metastásico con algo de
mejoramiento en la carga de células de tumor. Cuando se combina con las células
destructivas activadas con la linfoquinas (las células destructivas naturales expuestas
a la IL-2), las respuestas son un tanto mejoradas pero, aún así, son
desilucionantes.(25)
6.5 - CITOCINAS EN LOS CASOS DE SEPSIS.
Las concentraciones aumentadas del TNF en el suero han sido observadas en
humanos y en animales después de inyección de endotoxinas, con un punto máximo
de 2 horas y un retomo a la línea de base a las 6 horas.
Varios estudios han medido el TNF y otras citocinas en la sepsis. En las
sepsis muy severa negativa de Gram, las concentraciones de TNF están
aumentadas en la mayoría de los pacientes, a veces hasta muy altos valores,
mientras que la IL-1 y el TNF están aumentados en 20% de los casos. En un grupo
de pacientes admitidos a una unidad de cuidados intensivos con choque séptico, el
TNF está aumentado en solamente el 25% de los casos. Sin embargo, todas las
concentraciones antes mencionadas indicaron una correlación reciente de las
concentraciones de TNF y la mortandad en el choque séptico.
Es difícil definir un valor por encima del cual la procnosis disminuye
marcadamente, particularmente porque la concentraciones vistas en la mayoría de
las pacientes con choque séptico severo son menores que aquellas en los
voluntarios humanos después de una inyección de endotoxina sin consecuencias
dañinas. La sepsis, la concentraciones de TNF, fueron mantenidas durante 2-6
horas sugiriendo que ia vibración sostenida de l a e n d o n a
duradón y .a
resultado. Ademas, el TNF circula
^
A p lica d o como el único
y¡J que ,a relaclon con otras
^
dt^'inas'y^a'en doto xina circulatoria misma debe ser importante.
Actualmente la
esta clase de información en el techa*
^
S
“
e, ¡ndicador de protección, dada la
'
—
Cabe mencionar que no solamenJ ^ ^
b
^
m
n
' ^
r c
T
“
.
i
—
*
quemaduras y en los recha.os
■ • _4.„ (1.22.25)
de injerto.'
6.7
OTROKUL.
ROL:i_no
LAS vVACUNAS
6
7 - -OTRO
/-ww
Además de la co n sid e ra ció n j*i la I M 2> c o n o, un
—
r
«
b = a e t ^
r ded°o e s « r fu " e SxIm fnadosV on el mismo
ratones mostraron una respuesta " £ ^ ^ ^ £
—
—
*
0
0
* »
de forma TH2 de
s r j ^ s ; »
*
te d o re s .
Varios otros modelos de « c u n a s ^
leishmaniasis el investigador
IL-6 , en los primeros no humanos en *
están
están siendo
siendo investigados
investigaos todavía.■
ca u s a -
"
r
s
S
En ,a
¡ da dPderep e n s i E a ha demostrado
—
do con
y
enfermedades
H
S
s
»
:
a l
, e
i i d
0
43
asociado con la esquistosomiasis (una infección helmíntica propia del agua). Este
daño es causado principalmente por la reacción del sistema inmune a los huevecillos
del parásito, los cuales son depositados en el hígado e inducen una vigorosa
respuesta TH2 asociada con la formación de masas de tejido inflamado, ó
granulomas, y el engrasamiento del tejido conectivo conocido como fib ro s is .(3)
44
CAPITULO SIETE
ENSAYOS Y BIOENSAYOS.
7.1 - ENSAYOS DE CITOCINAS
Virtualmente todas las citocinas pueden ser mediadas con un apropiado
bioensayo, esto no es sorprendente ya que la mayoría de las citocinas fueron
originalmente
descritas como factores biológicamente activas y fueron
subsecuentemente purificadas y biológicamente caracterizadas. Sin embargo, los
bioensayos no son ideales y los sistemas apropiados de inmunoensayo serán
requeridos antes de que la medición de las citocinas pueda volverse un adjunto útil
para la Química Clínica.
7.2 BIOENSAYOS.
El problema más importante con los bioensayos es que estos no son
específicos. Como ejemplo se encuentran los intentos para desarrollar un bioensayo
específico para la IL-1,, aunque el ensayo general de proliferación de timocitos fué
descrita como un ensayo específico de IL-1, muchas citocinas han demostrado que
influyen en este. La IL -6 y el TNF pueden reemplazar a la IL-1, y la IL-2 e IL-4
pueden sinergizar con la IL-1. En un intento para evitar estos problemas, se han
desarrollado varias modificaciones en la metodología.
Utilizando anticuerpos neutralizadores específicos, uno puede demostrar que
la actividad reside en la IL-1 ó IL-6 . Saturando el ensayo con IL-2 ó IL -4 se puede
desechar la influencia de estas citocinas en el examen de muestra.
Cabe mencionar, que la actividad biológica medida en algunos bioensayos es
menos que el aparentemente medido por el inmunoensayo -ta l vez a causa de los
factores inhibidores- en las moléculas de enlace de las citocinas ó las citocinas
biológicamente activas pero inmunoreactivas. Las moléculas de enlace de las
citocinas en el suero y en la orina han sido descritas para la IL-1, la IL-6 y el TNF.
La mayoría de los bioensayos de citocinas son un trabajo intenso, caro y sujeto a
considerable influencia del operador. Aunque obviamente no es ajustable para los
exámenes de laboratorio y de rutina de Química Clínica, estos tienen algunas
ventajas que los hacen invaluables, al menos que el roll de los ensayos de citocinas
esté siendo determinado. Generalmente estos son muy sensitivos, detectando
citocinas en forma de monogramas por milímetro Además, los materiales requeridos
para el bioensayo son a menudo más
fácilmente aprendidos que los del
inmunoensayo; es particularmente difícil obtener buenos anticuerpos específicos
para algunas citocinas. Por lo tanto la aplicación de bioensayos puede identificar
cuales áreas garantizan la inversión en el desarrollo de un inmunoensayo.(5,23)
45
7.3 - INMUNOENSAYOS DE CITOCINAS.
Aunque los inmunoensayos tienen varias ventajas distintas sobre los
bioensayos (métodos simples para usar y controlar), la introducción de los
inmunoensayos de citocinas hacia los exámenes de laboratorio de Química Clínica
no es directa. Los inmunoensayos de varias citocinas están ahora disponibles de
fuentes comerciales, pero son de alto costo y en general han sido validados como
herramientas de investigación, a menudo solamente para los cultivos celulares. En
muchos casos, estos inmunoensayos carecen de sensibilidad para su uso con
plasma, y algunos están sujetos a la interferencia del factor reumatoide ó los
anticuerpos hetereofílicos. Estos son imprecisos y a menudo no demuestran perfiles
de recuperación aceptable de plasma, fluido cerebroespinal ó líquido sinovial. Por lo
tanto, cuando sea posible, los laboratorios que se interesan en la medición de
citocinas en número sustancial de tomas necesitarían establecer sus propios
ensayos. En esta forma ellos pueden asegurar la validez de sus resultados y motivar
a la producción de reactivos tales como los anticuerpos específicos de citocinas.(3,5)
7.4 - ENSAYOS DE CITOCINAS
La formación de colonias eritroides en los ensayos de colonia de la médula
ósea, la estimulación de la eritropoyesis en los ratones policitémicos post-hipóxicos y
la estimulación de la síntesis heme en las células de la médula ósea ratas- humanos
pueden ser analizadas mediante un examen de ELISA ó por un radioinmunoensayo
(RIA).
La GM-CSF estimula la proliferación de los granulocitos y el macròfago tanto
como la diferenciación y activación de la histamina. Este tiene un potente efecto
sobre varias líneas de células, es difícil de detectar en la sangre periférica aun en la
infección bacterial ó la endotoxemía, pero puede ser detectada en el fluido cerebro
espinal durante la meningitis bacterial. Este también puede ser examinado por el
examen ELISA Y RIA.
Los bioensayos para el G-CSF son realizados al examinar la formación de la
colonia de granulocitos en los ensayos de colonia de la médula ósea, el
mejoramiento de la producción de superóxidos en los neutrófilos, ó la proliferación de
una línea de células murinas. El G-CSF ha demostrado ser útil para movilizar las
células tallo de la médula ósea humana hacia la sangre periférica, poniéndola
disponible para su recolección, para uso posterior en la trasplantación de células
tallo. La G-CSF también ha sido utilizada para medir la toxicidad de la quimio y
radioterapia en médula ósea y puede inducir el reclutamiento de células malignas.
Otra de las funciones es la proliferación de líneas específicas de la células
leucémicas.
El factor de células de tallo es un cofactor de la hemopoyesis temprana,
producida por las células estromales. Este tiene una actividad relativamente pobre
sido desarrolladas.
Por otro lado, la IL-2
s
»
- - —
La
i l -3
estimula
"
los
ensayos de formac
ofreciendo examenes EL
—
res; ; '
" u e d e T e ' " “^ «
o
basófilos
^
por la
^ )L 3
P
r Aiuias j y NK y
La 1L 4 activa las células B, induce ' ^ P ^ f u n e T s d'e o é fu ts específicas
s
?
s
?
S
=
s s f ®
S
i H
S
2¿
=
S
S
«
5W ssE
de células. Los ELISA en est
^ ^ proliferación de
LA |L -6 estimula la secreción de an' ICU^ ° al¡zada al medir la proliferación de
IOS «mocitos y la hemop,cyesis J r t a p u r * • el mitógeno ó la línea p t e y * ™
líneas de células, la cél“ j f s de®ec,ada% o r su mejoramiento de la
tóboratórios^estón W ^a n d ^ d^ada^tarla^aj^examen ^
r S S m
q
* *
T '3
aaaaya
un factor de
pronósticos en Íos pacientes^oort míe,orna múltiple.
^
El factor de necrosis d® ^ d ^ r a c t i v a c i ó n T s t ó o c l á s i c a , la estim ularon
activación de
PT
T
0< o Z o ^ de factor decrecimiento
Sleucocitorhumano
H ^ LA?.eM(
S 3,f,23)s s s s s w
las moléculas de adhesión.
s s « - 4- .
47
7 5 C TO cS um Tn^S 3 rASETEsfDADEpORBSNTE0STANSARIZACIÓN.
I
mprliciones de laboratorio de las citocinas
r sr
pr
condusiones errén eas.A
analizar ciertas cltoc,n
xr¿ i
«
rpsDonder a diferentes atocinas y a su v
‘
P
,‘zzszsr¿£Z-rj,«
' " Z
Z
—
v
s
s
9
lo s estándares de la citocinas humanas y£
r| f 0^
disponibles en el Instituto Naciona de Estándar g
^
S
‘S
/ "
g
^
t
a
t t
TGF-
m.csf
s
,* ¿
r S
í:
” s s s v s - .« r ;" " rr® .
internacionales. La nat
. ,
0f
x
s
~
m
uso
s
~¡
¡dadoso del espécimen.
* * s z ó sse°n
« s r s
r s
(jc 0 con sus ensayos apropiados,
aumentado d e ja s e
preparado para apoyar en el
g
Las técnicas exactas son el requisito obvio para anaHaar es
¡ r s
m„ié c u la s <23)
7 .6
. APLICACIONES CLINICAS DE LA MEDICIONES DE CITOCINAS.
A un nivel de investigación,
ma^ r¡ | ^® s'V n muclros diferentes procesos
exitosamente
F
i S
S
S
S
y las c
cerebroespinal en la mfeccio
m e n in g o fo c a l
S
^
&
S
S
i
sistem a Nervioso Central.
^
Para entrar en el repertorio ^ Laboratorio de
citocina deben ya sea p ^ e e j™ e v a jn .o ™ tos .
examinan la
d ¡a g n ó s t¡c 0 6 el manejo
es
e n te m n e d a d jn ^
s e n s M d a d a e ¿ $e < ^ c t e d ^ v e l^ '^ a d ^ ® d'®apd ®Sp®¿dup i¿ n de c itrin a ® a a ^ a m s ,
Al contrano. es el W
están comenzand^
de los ^
^
u
mayona de los la c a d o r
,Aiido -Tales como proteínas d
. . . . d _ ¡ase aguda,
^
...
v (23)
y
de enzimas
CONCLUSIONES.
familia de mediadores
mismas atocina , y
redundantes e influyen
, as acciones de las diferentes atocinas w « »
An m síntesis de otras citocinas. En g
’
en la acción y
inflamatorios y antigemcos, y
ejercer acciones endócrinas.
son críticas para la D
y unen a la inmunida
$e sintetizan en
,a capacidad para actuar a
. alta afmidad en las
u n a cantidad suficiente
^
^ ^
jfica con ta natural.
P
al antíaeno.
>ara eliminar al antigen .
naturaleza de las respuestas
.
pxcesivas de las citocinas
, „ “ sss s s s s s s a s a . . - —
y asociarse con enfermedad.
—
en ,a detección y
rnediciónSdeaia^r^ á o |c in a s '^ i^ ^ o a ^ para^ambos e ^ s t u d i ^ ^ J ^ ^ ^
enfatiza.
-
^leggr
—
”
« „-¡ huos oara su medición están
Cabe mencionar
^ ^ n s a y o s .^ n m u rw e n s a y o s , medición de RNAm,
''desarrollados,
y p S m ° e n t o s inmunoquímicos.
siendo
ensayos de enlace de recepto y P
g su gfan
apoyar en el d,ag
. CITOCINAS.
este complejo mundo de las w
BIBLIOGRAFIA
s
2
s
Z
.
B. An endotoxln-
w illia m
:
-
£ *
°f —
pr oc Ma"
Acad sci !> g gi \
SC, Kamen R. The human hematopoietic colonystimulating factors. Science
'
anno
3 . Clinical Chemistry. Vol. 36,
M . Aste
4 . D Andrea A. R e n g a r a ju N £
rC han".
a
i
S
5
S
S
=
Chizzonite. 1994. Production o,
(1L-12).
„ af cStoo rr O
,t a )-
—
t t . »
^
^
%
'^
2
2
^° ;r r ^
7
science 234, —
r
.
oateiy, 1993.
r : : r : a
s“
_ na a 177Q-1782.
.
S .H
-
^
^
r
in 9
.
^
and secretion o,
8 . Giri JG L L 0 df di0 ° inan 9 9 5 134:346-9.
Interleukin 1. mm
and expression of DMA for human
•
9 . Gray PW.
lymphotoxin, a lymphokine wir
tO.Hirano
Taga ^ „ “
c h a ra c te ris a tio n of human
1
9
9
5
T.
‘
—
e ird ^ e le n ttio ^ tT o rp ro c Nati Acad Sci USA.
d
to rifin k in -3
i
a o o iiu la r b io lo g y of in te rle u k in
4 , . míe IN. The molecular and cellular bio
i
1 2 .Jam es T
gy
-o M e d ic a
B a rn * In m u n o lo g ia ^e d .
México, Ano 1990.
(LOnd° n)' 1" 4 h„ moaeneity and
o.
Y
e
a
r b
Im m u n o l, 1 9 9 5 .
pa¡s.
E d it. In te ra m e ric a n a . E d.
.
,p.
factor.
, 3 Jones E.Y. Stuart., O.I. and walker N.C.P. structu
13' Nature. 338, 225-228.
14.Me Kay D.B. (1994) Unraveling the structure of 1 ,
■ nce 257, 412-413.
see
51
D. T h , ™ r
, . »=
colony - stimulating factors blood. 1992.
h •
il
Kevacs EJ Mat sushima K, Durum SK. There is more than one
*01-15
interleukin-1. Inmmunol. Today. 1996,162.
1 6 .Oppenheim J.J. Kevacs
-r
17 S
S
d
bWa s J Ä
n
Hp Teresa R
v dr n 2 S Ä
et al Clinical correlates of cortical and
dementia. Neurol .994.
1 8.Schmit JA. Rhodes J. Bom.oid R. Manipulation of interleukin t syn.es,s secret,on
or action. P p- 303-19.
tg.S e lkoe DJ. Alzheimer's disease. A centra, neuropathol. Exp. Neur 1994.
Ä
ninrp Walter AM and Wlodawer (1994).
' Ä
I
ies Ot human recomOinan,
pS
interleukin-4 , Biol. 1, 301-310.
21 • moderno.
^
1991. P p. 379-387.
I,
-
S
Ä
op
r
Ä
Finari IS Dalladino MA, Shepard HM,
on pro,„era,ion ot nomna, and
transformed cells in vitro. Science, 1995.
» d -,h p r Mire Sluis AR. Detection and measurement o
23. Thorpe R. Wadwha M ^B ird OR. M, .
cytokines. Bload Rev. 1992.133 140.
RC Selective in vitro growth of T.
26.W. Morgan D.A., Ru* ° j W an bone marrow. Science 1995; 193: 1007-1008.
Lymhocytes from normal human Done m«*i i
cm
• . j
L.n o receotor Adv Immunol 1993,42: 1£>
27. W. Smith K. The interleukin - 2 receptor, «a
 0
0
Anuncio
Documentos relacionados








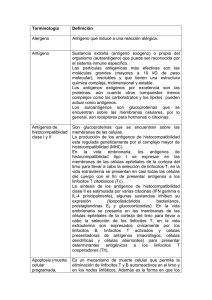
Descargar
Anuncio
Añadir este documento a la recogida (s)
Puede agregar este documento a su colección de estudio (s)
Iniciar sesión Disponible sólo para usuarios autorizadosAñadir a este documento guardado
Puede agregar este documento a su lista guardada
Iniciar sesión Disponible sólo para usuarios autorizados