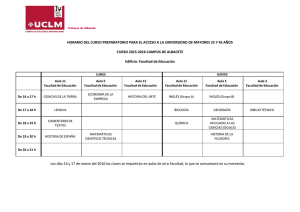
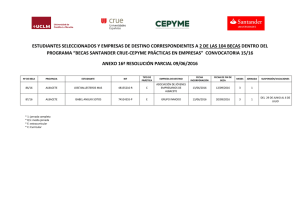
DETERMINACIÓN DE LA COBERTURA Y DENSIDAD DE LOS
Anuncio

DETERMINACIÓN DE LA COBERTURA Y DENSIDAD DE LOS ESPARTIZALES TIPICOS DEL SURESTE DE LA PROVINCIA DE ALBACETE MEDIANTE ANÁLISIS DIGITAL DE IMÁGENES E. JORDÁN GONZÁLEZ~ M. SELVA DENIAL E. OROZCO BAYO' J. M. LÓPEZ PÉREZ~ J. A. LÓPEZ-DONATE? 'Escuela Técnica Superior de Ingenieros Agrónonios de Albacete. Universidad de Castilla-La Mancha 'Centro de Experimentación Agraria de Alhacete. Junta de Coii~unidadesde Castilla-La Mancha 1 RESUMEN ABSTRACT Con este trabajo se pretende aportar cierta información a cerca de la cubierta vegetal de una comunidad herbácea muy frecuente en los terrenos calcáreos de la provincia de Albacete. Nos referimos a los espartizales, los cuales en nuestra provincia aparecen en gran extensión de terreno y ocupan generalmente exposiciones heliófilas, ya sea solana o zonas llanas. Para esta formación se ha obtenido información acerca de la diversidad florística, parámetros morfológicos de la parte aérea, cobertura vegetal y oferta de forraje aportado por el esparto. This work is intented to provide some info about the vegetable cover of the herbaceous community wich is very frecuent on the calcareous fields in Albacete. We are talking about the esparto fields wich in our province appear in large-sized lands. and generally occupy heliophiles exposition either suntraps or flat areas. In order to achieve this information, formation about the floral diversity, the morphological parameters of the aerial part, the vegetable cover and the offer o forage supplied by the esparto has been obtained. Palabras clave: Stipa telzacissima. pastos Keywords: Stipa telzacissima. perennial pastures. diversity, erosion. vegetation cover. vivaces, diversidad. erosión, cobertura vegetal. El esparto o atocha (Stipa te~zacissinza),gramínea vivaz. de carácter xerófilo. porte amacollado y hojas duras filifor~nes,es una especie de gran importancia. capaz de ocupar hábitats degradados (fundamentalmente de solana) donde la vegetación arbórea. y la mayoría de los matorrales tienen escasas posibilidades para desarrollarse, en aiilbientes seriliáridos del sureste peninsular. Por tanto, estas formaciones son importantes por los grandes beneficios indirectos (protección y estabilización de litosuelos. etc.) que pi-oducen, aunque éstos sean muy difíciles de cuantificar. Entre estos beneficios. se puede citar el efecto que tiene sobre el control de los recursos hídricos del suelo: la macolla de esta planta vivaz se comporta controlando la radiación solar que llega al suelo, y evitando la evaporación del agua al~ilacenadaen éste. Por otro lado. actúa como colchón reteniendo en su estructura parte del agua de lluvia y aportándola al sustrato edáfico de forma más uniforme, permitiéndose con ello un mayor volumen de infiltración del agua de lluvia. Este efecto sobre el control de la humedad edáfica se ve acentuado por el bajo ratio de transpiración por unidad de área cubierta por esta planta. ya que la mayoría de las veces cuenta en su estructura con abundante vegetación muerta. Es importante remarcar que esta fracción de cubierta muerta cumple sus funciones en el control de los recursos hídricos. y a veces de forma más eficaz que la viva. ya que el efecto de empape es mayor, y por ende su capacidad de almacenamiento de agua. Por tanto el efecto general que se produce es una disminución del agua de escorrentía. una disminución del calado de la lámina de escurrido. un incremento del tiempo de concentración de la cuenca y del embalse edáfico. gracias a la presencia y efectividad de esta herbácea vivaz. Además. el porte amacollado desde la base del suelo se convierte en un factor primordial en la defensa ante la erosión, permitiendo una estabilización del terreno y contribuyendo a evitar, o al iilenos reducir la emisión de elementos finos del suelo al agua de escorrentía. Debido a la intercepción del flujo de escorrentía, resultan frecuentes después de los incendios forestales las acumulaciones de sedimentos en la parte superior de la planta. a modo de minúsculas mesetas. En cuanto a beneficios directos, en épocas pasadas se mantuvo un aprovechamiento de su esparto. pero hoy en día prácticamente tiene interés econóinico en nuestra provincia por el aprovechamiento pastoral de su espiga (atochón). Por tanto. el objetivo fundamental de este trabajo fue evaluar y cuantificar la cobertura y densidad que presentan estas peculiares formaciones vegetales del sureste árido de la provincia de Albacete. comparando metodologías clásicas de muestreo (Point-Center Quarter: Passera el al., 1983) con nuevas tecnologías de análisis digital de imágenes. y obtener. por tanto, coeficientes de corrección que aproximen los muestreos clásicos a la realidad del monte. Los muestreos se realizaron en la finca de propiedad pública La Loma (Liétor). Entre los espartizales que aparecen en dicha finca se eligieron las diversas zonas de muestreo. de manera que fueran representativos de la estructura típica de los espartizales del sureste de la provincia de Albacete. 1.3. Corrección de la densidad aportada por Point Center Quarter 1. Determinación de la densidad 1.1. Muestreos de vegetación mediante bandas para obtener la densidad real Sobre estas formaciones representativas, se han replanteado sobre el terreno 6 bandas de 50 in de largo por 2 m de ancho, en las que se han contabilizado todos los individuos de las especies de arbustivas presentes. además del esparto, obteniendo de esta manera la densidad real (número de individuoslha). 1.2. Muestreos de la vegetación mediante el método Point Center Quarter (Passera, et al., 1993) Esta metodología se halla descrita en otra comunicación de estas mismas jornadas (Joru'cín et al., 2 0 0 1 ~ ) .aportando información de densidad (número pieslha). diámetro de copa y altura de planta. A partir de estos datos se pueden obtener, además, coberturas. y fitovolumen ideal de los diversos componentes de la formación vegetal. Los transectos realizados se han llevado a cabo en el interior de las bandas señalizadas anteriormente, para poder comparar ambos métodos y así poder obtener el coeficiente de corrección de la densidad. Se trata de un método de muestreo utilizado con bastante frecuencia para el muestreo de estratos arbustivos, sobretodo en estudios sudainericanos (Passera et al., 1993). En nuestro ámbito también se ha utilizado en importantes estudios pastorales (Robles, 1990; Jordán et al., 20016). pero no se ha comprobado la fiabilidad del método a la hora de aportar información referente a densidades de individuos por hectárea y cobertura vegetal real. Por tanto, en este trabajo se intenta acometer esta labor, llevándose a cabo sobre una formación muy representativa de la provincia de Albacete. como son los espartizales. Para este ajuste se utiliza la densidad de individuos obtenida por el método de las bandas, y se confronta con la aportada por el método de muestreo de arbustos Point Center Quarter. De esta comparación se obtiene el valor de la constante p, dependiente de la formación vegetal. Ecuación de densidad del método Point Center Quarter: D: 10.0001 P*d"m) D: densidad de individuos por hectárea. p: constante de corrección. d: distancia media desde el punto de inuestreo hasta la planta más cercana. 2. Determinación de la cobertura vegetal de los espartizales La determinación de la cobertura vegetal en estas comunidades se ha calculado mediante 2 procedimientos: mediante análisis digital de imágenes tomadas sobre el terreno. y a partir de la información aportada por el método Point Center Quarter. 2.1. Análisis de la cobertura con ER MAPPER Para ello, sobre los mismos transectos utilizados para la metodología clásica. se han extraído iinágenes digitales. que posteriormente se han analizado mediante el programa informático ER MAPPER v.6.0. Esta aplicación informática aporta como resultado el porcentaje de fracción de cubierta vegetal y de suelo desnudo de las imágenes tratadas. Toma de imágenes en campo En cada una de las zonas representativas que se han elegido. se ha realizado un transecto de 50 in de longitud. Este transecto se ha colocado en la dirección de la máxima pendiente para incorporar al inuestreo toda la variación posible del medio. Sobre este transecto, ubicado físicamente con una cinta métrica de 50 metros de longitud, se ha realizado la toma de una imagen digital, cada 1 metro. Gráfico 2. Frecuencia florística Especie Se observa (Gráfico 2) una clara dominancia de la frecuencia de individuos de Stipa tenacis.sima (56.9%). Le siguen especies de la familia Labiatae como son Thymus vulgaris (9.9%) y Fumana thymifolia (9.0%). En este trabajo la frecuencia de cada especie es importante. ya que junto a la cobertura individual se halla directamente correlacioi.iada con la contribución que cada especie realiza a la cobertura total. Según este inétodo clásico. el número de individuos por hectárea (Tabla 1) es de 5 1.149. siendo la especie Stipa terzacísima con una media de 28.550 individuos por hectárea la que inayor densidad posee. Le siguen en importancia la Furnana thqwiifblia (5.459 individuoslha) y Alztllyllis cytisoi(les (4.40 1 individuoslha). 2.2. Método de las bandas 2. Densidad de individuos por hectárea Tabla 2: Densidad según el niétodo de las bandas A continuación se exponen los resultados de densidad obtenidos mediante los métodos Point Center Quarter y por bandas. 2.1. Metodología Point Center Quarter Tabla 1: Densidad según el niétodo Point Ceiiter Quarter Especie Media Anthyllis cytisoides Satureja obovatcr Cistiis cliisii Dorycnium pentnplzq~llum Stipa tetzacissiina Filrnancr thyinifolin Heliaizthenzunz pilosunz Heliurlthern~rmlavand~ilifoli~~~n Litlzodorcr fr~iticosa Rosmar-irzus oflcirzalis Rhcrrnnir.5 1ycioicie.s Tvmus vlrlgaris Teucrilim poliuin 440 1 2017 49 1 1450 28550 5459 236 723 44 48 177 6889 665 Totaiha 51 149 Especie Media Anthyllis c.ytisoide.s Satilr-eja obovata Cistzls clu.tii Dorycnizim pentcr~~lzyllunz Stipa tenucissima Ficmnna thqlrnifolia Helianthernilm pilosurn Heliarzthemiim luva~zdulifoli~~~iz Lirhodolzr fiuticosa Rosinnl-ir1lis oficinalis RI7amrzu.s lycioides T\'nlus vlrlgcrris Teucrilim poli~irn Mediante este método de bandas también se aprecia la doininancia del núinero de individuos de Stipa telzcrcísima (10.850 individuos/ha), seguida muy de lejos por Ty1nus villgar-is (3.467 individuostha), Antlzyllis cytisoides ( 1 3 5 0 individuosíha) y Fin?zana tlq~mifolia(1.600 individuoslha). 2.3. Obtención de la constante de corrección de la densidad "b" Tabla 3: Valores medios de la constante Especie Media Anthyllis cytisoides Satureju obovata Cistus clusii Dorycrzium petltaphyllurn Stipcz terlacissima Fumuna thymijolia Heliarzthemum pilosirrn Heliarzthemum 1ai)urzdulifolium Litlzodoru fruticosa Rosnzarinus oficinalis Rhurnrzus lycioides n m u s vulgaris Teucrium poliurn tal, mediante la confrontación de resultados de una y otra metodología. De esta comparación de resultados por especie. se obtiene un valor medio aritmético de b para todas las especies de 3.1. Si se tiene en cuenta la frecuencia de cada especie la media ponderada que se obtiene de b es de 2.5. Por tanto el método clásico Point Center Quarter tiene un valor de P=2,5 cuando se aplique a las formaciones dominadas por Stipa tenacissima. 3. Cobertura Por una parte se calcula la cobertura a partir del método clásico. y por otra se calcula con el software inforinático ER Mapper de alto grado de precisión. De la comparación de ambos métodos se obtiene la constante de corrección (K) del método clásico. Media aritmética 3s Media ponderada 2.5 Según los valores de las tablas 1 y 2 , se observa que el método clásico de muestre0 Point Center Quadrat aporta una densidad de individuos por hectárea muy superior a la existente: por tanto. se propone corregir este método para esta formación vege- 3.1. Metodología Point Center Quarter 3.1.1. Diámetro de copa El valor medio del diámetro de copa es de 37.6 cm, (Tabla 4) pero existen claras diferencias significativas entre las diferentes especies (F: 63.74. a:O.OO). Ello da lugar a la formación 4 grupos homogéneos diferentes. Tabla 4: Valores medios de diámetro iiiedio de copa (crii) Intervalo de confianza al 95% Media Error típico Límite inferior Límite superior Gráfico 3. Diámetro medio de copa (cm). E C, E 'tu ii Especie **a,b,c,d,e: las especies que comparten la misma letra pertenecen al inisrno grupo hoinogéneo de diárrietro de copa. Rosmarinus oficinalis (79 cm) y Rhamrzus lycioides (105 cm) son las especies que mayor diámetro de copa presentan (grupo homogéneo d y e). Le sigue Stipa tenucissirna con 50 cm de diámetro de copa (grupo homogéneo c). Los valores más bajos son ofrecidos por Teucrium polium ( 1 2 cm, grupo a). 3.1.2. Cobertura media individual En cuanto al factor área de copa (F: 31.12, a:O.OO), la mayoría de las especies se hallan dentro del misino grupo (Gráfico 4), excepto Rosmarinus oficina1i.r (grupo b ) y Rhamnus lycioides (grupo c) con valores de área de copa bastaste superiores. El valor medio global de área es de 0.20 m'. (Tabla 5). Tabla 5: Valores iuedios de área de copa (m?). Intervalo de confianza al 95% Media Error típico Límite inferior Límite superior Gráfico 4. Cobe~íuramedia individual (m?). Especie **a,b,c: las especies que comparten la misma letra pertenecen al misino gmpo homogéneo de área (cobertura) media de copa. Las especies con mayor área de copa resultan ser Rosmarinus oficinalis y Rhainnus lycioides, pero debido a su escasa frecuencia su contribución a la cobertura vegetal es escasa. En cambio el mayor aporte a la cubierta lo produce Stipa tenacissima. tanto por su razonable área de copa. como por su elevada frecuencia. 3.2. Cobertura mediante ER MAPPER Tabla 7. Cobertura mediante ER MAPPER 1 N" transecto 1 Cobertura 1 1 MEDIA 1 49 1 3.1.3. Cobertura por transecto mediante el método clásico Tabla 6. Cobertura mediante Point Center Quarter 1 No transecto 1 2 3 4 1 Cobertura 42 40 102 73 1 La cobertura aportada por el software ER Mapper es inferior, en la mayor parte de los casos. a la ofrecida por el método clásico. 3.3. Coeficientes de corrección de la cobertura del método clásico "K" MEDIA 58 En esta tabla se ofrecen los resultados de cobertura para los 6 transectos en que se ha calculado ésta. mediante las dos metodologías (clásica y ER Mapper). Se observa cómo en ocasiones. este méto.. do clásico ofrece una cobertura incluso superior al 100 %, debido a que tiene en cuenta los solapes. De la comparación de la cobertura aportada por ambos métodos se ha obtenido la constante de corrección del método clásico (k), siendo el valor de esta de 0.88. Este valor se aplica a la cobertura del método clásico cuando no se ha corregido aún la deiisidad, es decir. para la cobertura resultante de una densidad calculada para P= l . CONCLUSIONES Stipa tenacissirna es una especie que muestra una preferencia por las zonas heliófilas de solana. formando esta herbácea vivaz comunidades donde a veces deja escasa posibilidad de entrada a otras especies. tal y como demuestra el porcentaje de frecuencia de aparición (casi un 57%). Del análisis de resultados de densidad aportados por los dos métodos utilizados en este estudio. se desprende que el coeficiente P que se debe emplear en el método clásico (Point Center Quarter) para el cálculo de la densidad en comunidades de Sripa telzacissirna, tiene un valor medio (ponderado con la frecuencia media de cada especie de esa comunidad) de 2.5. En cuanto a la cobertura media de copa. el romero (Rosmarinus oflcinalis) y el espino negro (Rhamnus lycioides), seguidas del esparto. son las especies que presentan el mayor tamaño de copa. Pero las dos primeras poseen una representación escasa. Por tanto, debido al considerable tamaño de copa del esparto y su alta frecuencia en la comunidad estudiada, esta especie cumple una función crucial en la disminución de la erosión. ya que además del aporte a la cobertura vegetal, su macolla actúa a la vez reteniendo parte de los arrastres y disminuyendo la velocidad del flujo hídrico. Por tanto. es una especie con un valor hidrológico importante para ampliar zonas de la provincia de Albacete. En cuanto a la estimación de la cobertura global. se aprecia que el método clásico sobreestima los resultados. los cuales se pueden corregir aplicando el coeficiente de corrección (K) con valor 0.88 (habiendo calculado esta cobertura previamente con P=l). COTTAN, G. & CURTIS, J. T., 1949. A method for making survey of woodlands by selected trees. Ecology 30: 101- 104. COTTAN. G. & CURTIS. J. T.. 1956. The use of distance measures in phyto sociological sampling. Ecology 37: 45 1-460. JORDÁN, E.; SELVA, M.; OROZCO, E.; LÓPEZ. J. A. Y FERNÁNDEZ, J., 2001a. Determinación de la oferta forrajera consumible en aprovechamiento extensivo en un enclave característico del sur de la provincia de Albacete (Liétor). Actas II Jornadas de Estudios Albacetenses. Albacete. JORDÁN, E.: SELVA. M.; ARTIGAO, A. y GARCÍA. J., 2001b. Análisis de la coinposición y estructura de un sistema forestal del Sureste Ibérico (Sierra Segura. Albacete). III Congreso Forestal Español. Granada. PASSERA. C.: DALMASO, A. Y BORSETTO. O.. 1983. Método del point quadrat modificado. Taller de arbustos forrajeros para zonas áridas y semiáridas. FAO/IADIZA. Mendoza. Argentina, pp. 7 1-79. ROBLEDO. A.: RÍOS, S.; CORREAL, E., 1990199 1. Estimación de biomasa en los matorrales de albaida (Atzthyllis cytisoides) del Sureste de España. Pastos, 20-21, 107- 129. ROBLES A.B. 1990. Evolucicín de la oferta forrcljera y capacidad sustentadora de un agrosistema semiárirlo del Sureste Ibérico. Tesis Doctoral Facultad de Ciencias. Universidad de Granada. VEGETACIÓN DE LOS "VALLEJOS" DE ALATOZ Y CARCELÉN RESUMEN ABSTRACT En esta comunicación hacemos una aproximación al análisis y estudio de la vegetación de los valles que descienden de las montañas de Alatoz y Carcelén. conocidos localmente como "vallejos". El principal interés de estas comunidades radica en la presencia importante de elementos florísticos submediterráneos, muy localizados en este extreino meridional del Sistema Ibérico. In this comunication we approach to the analysis and study of the valley vegetation which go down the mountains of Alatoz and Carcelén. locally known as "vallejos". The inain interest of these communities is the important presence of floristics subinediterranean eleinents. located in this meridional extreme of Sistema Ibérico. Palabras clave: Sistema Ibérico. vallejos. vegetación submediterránea. Keywords: Sistema Ibérico, vallejos. submediterranean vegetation. Las últimas estribaciones del Sistema Ibérico alcanzan la provincia de Albacete formando una alineación montañosa que. desde Carcelén hasta Chinchilla por el norte y desde Alpera hasta HoyaGonzalo por el sur. se extiende de este a oeste a lo largo de unos 50 km. Formada por materiales calizos, las altitudes máximas se encuentran en el Molatón de Higueruela (1245 m. ), Puntal de Tortosilla ( 1 203 m.) y Malefatón (1 108 m.). A pesar de albergar esta zona interesantes valores naturales. se halla mucho menos estudiada que otras montañas de la provincia y es bastante desconocida por los aficionados a la naturaleza. es la mediterránea. La vegetación predon~i~lante En general, se encuentra más desarrollada y mejor conservada cuanto más nos acerquemos al sector oriental. En estas sierras destacan como formaciones vegetales: Matorrales; dominantes en el sector occidental. Aparecen los típicos de la serie de degradación del encinar manchego: espartizales. romerales y tomillares. Encinares; aunque más bien deberíamos hablar de chaparrales. ya que son escasos los enclaves con abundancia de pies de porte arbóreo. Se encuentran con mayor abundancia en las inmediaciones de Higueruela y en las muelas de Alatoz y Carcelén. Pinares; a los de repoblación. muy extendidos en el sector occidental, hay que sumar los aparentemente naturales que dominan en el sector oriental. Básicamente son formaciones casi monoespecíficas de pino carrasco (Pinirs Izalel~erisis).aunque también se encuentran mezclas con pino rodeno (Piillrs pillaster) y inás raramente con pino laricio (Pirzus rligrcr subsp. scrllzmutzii) como en el Malefatón. Qztejigares; vegetación de influencia subnlediterránea que aparece en uinbrías del sector oriental. Normalmente encontrainos quejigos dispersos entre pinares aunque existen algunos quejigares muy localizados, destacando el existente en la umbría del Molatón. donde sobre suelos arenosos. encontrainos alguna especie silicícola como Cisrlts laur(fo1ius. Resultan particularmente interesantes los pequeños valles. conocidos en la zona co~iio"vallejos" que descienden de las muelas de Alatoz y Carcelén en dirección norte. En éstos podemos encontrar una vegetación relicta de clara influencia subinediterráiiea y levantina. con elementos caducifolios, inarcescentes y lauroides. Las mejores rnanifestaciones de esta vegetación se localizan en orientaciones de umbría. al pie de roquedos y a una altitud de unos 1.O00 m. Por ejemplo. en el "vallejo de las Fuentezuelas" (Carcelén) encontramos: Especies de óptimo submediterr'aneo coino arce (Acer granatense). roble ( Q u e ~ u.frrginea), s guillomo (Arrzelarichier ovalis), gayuba (Arctos-t~~~?l?ylos idva-ursi) junto a otras como Catarlariche cuerulea y Globirlaria virlgaris. Especies mediterráneas con requerimientos hídricos elevados (considerados en el contexto local) coriio rnadroños (Arbutus errledo). durillos (Viburnuin tinus), brezos tardíos (Erica termirzalis), madreselvas (Lonicera iinplexa), iniestas (Sarnrhnmncrs scoparicrs subsp. rei~erchorzii).Buple~trirrn rigiduin, Vincero.uic~rr?ii~igrirrn,etc. Endemisinos levantinos como la pebrella (Tl~j~iiiirs piperella). Especies mediterráneas que denotan la intlluencia levantina sobre la flora local como la aliaga (Ulex parv@orus), brezo tempranq (Erica multiflora), Cephalariu le~caatha,etc. Otras especies acompañantes de distribución mediterránea comunes en la zona como pino carrasco (Piatus halepensis), pino rodeno (Pinus pinaster), carrasca (Quercus ilex subsp. rotundifolia), matarrubia (Quercus coccifera), sabina (Juniperus phoenicea), crujía (Digitalis obscura), vidarra (Clematis fiammula), espliego (Lavandula latifolia), etc. Estas formaciones vegetales tienen gran interés a nivel provincial, pues representan una vegetación de transición entre la vegetación mediterránea (encinares) y la submediterránea (robleddesg, con la presencia de elementos relícticos en la zona como arces, durillos o madroños. 3. Madroño (Arbutus unedo). 1. Arce (Acer granatense). 2. Quejigo (Quercusfaginea) 4. Gayuba (Arctostaphylos uva-ursi). QUEJIGARES EN EL NORTE DE ALBACETE Y SUR DE CUENCA (España) Arturo VALDÉS FRANZI José Reyes RUIZ GALLARDO Area de Biología Vegetal. E. U. de Magisterio de Albacete. Universidad de Castilla-La Mancha RESUMEN ABSTRACT Existen localizadas extensiones de quejigos en la cuenca del Júcar, en el norte de Albacete y sur de Cuenca. Han sido visitadas en diversos momentos del año, procediendo al estudio de su flora y a la toma de inventarios fitosociológicos. Aquí se presentan algunos resultados, se discute su encuadre sintaxonómico y se proponen medidas de protección al amparo de la legislación autonómica actual. Some populations of Querciis fagitzea have been located in the Júcar river basin, in the areas of Northern Albacete and Southern Cuenca. They have been visited in different seasons in order to study their flora and prepare phyto-sociological inventories. In this paper some results are presented, their syntaxonomic frame is discussed and protective measures suggested, based on current regional laws. Palabras clave: Fitosociolgía, Quercus fagineo, Albacete, Cuenca, España. Keywords: Phytosociology, Quercus faginea, Albacete, Cuenca, Spain. El quejigo (Quercusfaginea Lam.) es una cupuIífera marcescente de porte más grácil que la encina y hoja más grande. Fácilmente identificable cuando aparece en grupo. puede pasar desapercibido a pnmera vista si está mezclado en pequeña proporción con masas de encinar. En cambio, en otoño e invierno sus hojas marcescentes y de color rojizo destacan sobre la tonalidad verdoso oscura del encinar. Los quejigos (también llamados rebollos) de la subespecie tipo (faginea) prefieren sustratos calcáreos y están extendidos por la mayor parte de la Península aunque predominan en la región Mediterránea. Los quejigares mediterráneos basófilos se encuadran en la alianza Aceri granatensis- Quercion fagineae y se encuentran en los pisos bioclimáticos supra y mesomediterráneos con un ombroclima subhúmedo o húmedo. En espacios abiertos empieza a aparecer el quejigo cuando las precipitaciones se aproximan a los 500 mm. Existen diversos tipos de encinares en la región Mediterránea peninsular. Aquí nos referimos sólo a los mesomediterráneos basófilos con irradiaciones al supramediterráneo inferior. y a los termo-mesomediterráneos. Unos bien representados en la región estudiada (Asparago acutifolii-Quercetum rotuizdifoliae) característicos de ombroclima seco, otros de forma fragmentaria y finícola (Rubio loizgifoliaeQuercetum rotundifoliae) bajo ombroclima secosubhúmedo. Se han visitado diversas zonas donde aparecen qnejigm, en1 localidades situadas en 11 terminos municipales distintos. Desde hace años su presencia era conocida por nosotros directamente o por referencias de terceras personas (R. Molina y J. Fajardo, com. pers.). Se han levantado los inventarios correspoadientes y, en su caso, se han tomado muestras para su idmtifieación correcta con ayuda de claves. De acuerdo con la fisionomla de las formaciones, su altitud en m. s. n. m., su composición florística, ate., se han confeccionado tres tablas de inventamos &~istintascuya posición fitosociológica se mrneflh. En d a s se separan las especies características de Qwerco-Fagetea (incluidas las de RhamnoP m e t e a ) , las de Quexetea ilicis y las acompañantes que tienen muy diversa adscripción fitosociológica. Loa tzhones se denominan según los criterios nomencIatmdes de las revisiones monogr4ficas aparecidas hasta la fecha en Flora Iberica, para el resto $e sigue a mora Europea o diversas revisiones de g b m s más recientes existentes en la bibliografía (aparecen citados tal como se recogen en el Catálogo de Phntas Vasculares de la Provincia de Albacete (Vddés 83. al,, 2001). En su determinación se ha descendido al rango subespecífico. En su caso, si no se indica la subespecie, nos referimos a la tipo. FigUrYl1: Localidades estudiada%en las provincias de Albacete y Cuenca. RESULTADOS Y CONCLUSIONES (= Bupleuro rigidi-Quercetum rotundifo- Encinares (Quercus ilex L. subsp. ballota @esf.) Samp.; = Q. rotundijolia L a . ) con quejigos, del m~srrmedi'teniáneosuperior o supramediterráneo Seriar, escasos en elementos de Querco-Fagetea (incluidos los de Rhamno-Prunetea). Propios de espacios abiertos, horizontales o de ligera exposición d norte, ombrodlha seco o subhúmedo inferior. Falm los elementos termófilos de Quercetea ilicis, así Sasminum fncticans, Smilax aspera, Pistacia lentiscus, l! te~binthus, Arbutus unedo. Los referimos a la asociación: Asparaga ~Wiollt-Quereetumrohcradifoliae Rivas-Martínez, Fmández González, Loidi, Lousa & Penas 2001 liae Rivas-Martínez 1982; Quercetum rotundifoliae "alcarreño" Rivas Goday in Rivas Goday, Borja, Esteve, Galiano, Rigual & Rivas-Martínez 1959, no Quercetum rotundifoliae Br.-B1. O. Bolos in Vives 1956) variante con Quercus faginea Son los que ocupan mayores extensiones, tanto en el sur de Cuenca como norte de Alhacete, entre los 800 y 1.200 m. Entre las acompañantes figura. un buen elenco de plantas del Sideritido incanaeSalvion lavandulifoliae (Rosmarinateaoficinaiisis). Tabla N" 1 Asparago acutifolii-Qitercetuni rotuizdifoliae Rivas-Martínez, Feinández González, Loidi, Lousa & Penas 2001 variante con Quercus fagirzea Otras conipañeras: Eriiiacea anihyllis 1.2 en 1, + en 2; J~iniperusconiinuiiis hemisphaesica + en 4, 1.1 en 5; Aphyllanthes monspeliensis + en 4. 1.1 en 5 ; Catananche caesulea + en 4, 1.1 en 5 ; Avenula bromoides 1.1 en 5, + en 6; Crocus serotinus salziiiannii + en 1 , + en 5; Salvia lavandulifolia + en 2, + en 6; Geum sylvaticuin + en 3, + en 4; Dorycnium pentaphylluin + en 3, + en 7; Euphorbia nicaeensis + en 4, + en 5; Argysolobi~irnzanonii + en 4, + en 7; Leuzea conifera + en 5, + en 6; Ophrys scolopax + en 5 , + en 7; Teucri~im gnaphalodes + en 5, + en 7; Coronilla ininiina+ en 6, + en 7; Satureja intricata gracilis 1.1 en 2, + en 7; Silene nlitans + eri 1, + en 5 ; Helictotrichon filifoli~iin1.1 en 1; Brachypodium phoenicoides + en 1; Cynos~irusechinat~is + en 1; Thapsia villosa + en 1; Cephalaria leucantha + en 2; Pinus pinaster + en 3; Cistus laurifolius + en 3; Holcus lanatus + en 3; Lavandula stoechas pedunculata + en 3; Thyrn~is inastichiiia + en 3; Agrostis sp. + en 3; Cistus salviifolius + en 3; Corynephorus divaricatus + en 3; Alyssum serpyllifoli~im+ en 4; Cai-ex iiiairii + eri 4; Teucri~impse~idochaiiiaepitys+ en 4; Anthyllis vulneraria subsp. + en 5; Horinathophylla lapeyrousiana + eii 6; Linuin narbonense + en 6; Heliantheiiium violaceuni + en 6; Scabiosa turolensis + eii 6; Sideritis tragoriganuni iii~igronensis+ en 7; Heli~inthein~ini oelandicum incanum + en 7; Arenaria aggregata + en 7; Plantago lanceolata + en 7; Sanguisorba minor + en 7; Astsagal~issp. + en 7; Helianthemuni apennin~iin+ en 7. l . Albacete. Higueruela. Eiitre El Molatón y Cerro de la Punta o Punta Gira Valencia. XJ395 173. 2. Albacete. Higueruela. Punta Gira Valencia o Cerro de la Punta. XJ 408 174. 3. Cuenca. A 5 km. del Puerto de Las Tórdigas hacia Motilla del Palancar. Junto N-320. 4. Cueiica. Desviación a Monteagudo de Las Salinas. Junto N-320. 5. Cuenca. Desviación a Olmeda del Rey, a 37 km. de Motilla del Palancar. Jurito N-320. 6. Cueiica. A 3 km. de Motilla del Palancar hacia Albacete. WJ 9477 7. Cuenca. De Villan~ievade la Jara a Motilla del Palancar. WJ 947 1 Quejigares mesomediterráneos acantonados en ramblas o barrancas en las cercanías del río Jucar, en laderas expuestas al norte. Más ricos en elementos de Querco-Fagetea (incluidos los de RhamnoPrunetea). Medios más mesofíticos por una mayor humedad ambiental y frecuencia de nieblas; incluso, a veces, con una mayor proximidad del nivel freático a la superficie (aparece con cierta frecuencia Rubia tinctoncm y Brachypodium phoenicoides, por ejemplo). Situados en el dominio del encinar seco (Asparago acutifolii-Quercetum rotundifoliae S), suponen una inversión ombroclimática: Cephalanthero rubrae-Quercetum fagineae Rivas-Martínez in Rivas Goday, Borja, Esteve, Galiano, Rigual & Rivas-Marthez 1960 con: Rivas-Marthez 1972 Tales quejigares, plenamente ubicados en el mesomediterráneo, están empobrecidos puesto que presentan muchos menos elementos de QuercoFagetea que los quejigares existentes más al norte, en el supramediterráneo subhúmedo de la provincia de Cuenca (vd. Ginés López, 1976, Mateo Sanz, 1983) que pertenencen a las asociaciones Cephalanthero rubrae-Quercetumn fagineae o Violo wilkommiiQuercetumfagineae, faltando los más mesofíticos. Ocupan espacios muy reducidos. Pero llegan a presentar docenas de ejemplares de quejigo, algunos de hasta 50 cm. de diámetro de tronco a 1 m. del suelo. Al avanzar hacia el E, hacia la Comarca de Ves, se aprecia el influjo setabense con la aparición de elementos más termófilos y umbrófilos como transición hacia encinares y fresnedas de Rubio longifoliae-Quercetum rotundifoliae y Fraxino orniQuerceturn fagineae respectivamente. Fotografía 1: Vista de encinares con quejigos entre Viilanueva de la Jara y Motilla del Palancar (Cuenca). TABLA N" 2 Cephalunthero rubrae-Quercetum fagineae Rivas-Martínez in Rivas Goday, Borja, Esteve, Galiano, Rigual & Rivas-Martinez 1960 corr. Rivas-Martínez 1972 Otras compañeras: Erinacea anthyllis 2.2 en 5, 1.2 en 6; Helianthemum cinereum rotundifolium + en 5, + en 6; Genista pumila pumila + en 1, + en 6; Coronilla minima + en 5, 1.1 en 6 ; Aphyllanthes monspeliensis + en 5, + en 6 ; Ophrys scolopax + en 5, + en 6 ; Linum suffruticosum + en 5, + en 6; Bellis perennis + en 5, + en 6; Linum narbonense + en 5, + en 6; Cistus clusii 1.1 en 1; Carex distachya 1.1 en 8; Salvia lavandulifolia + en 1; Heiictotnchon filifolium + en 1; Satureja intncata gracilis + en 1; Fumana ericoides + en 1; Heiianthemum syriacum + en 1; Atractylis humilis + en 1; Pinus pinea + en 2; Stipa tenacissima + en 2; Phlomis lychnitis + en 5; Eryngium dichotomum + en 5 ; Euphorbia squamigera + en 5; Ailanthus altissima + en 5; Fumana scoparia + en 5; Hippocrepis squamata + en 5; Biscutella valentina + en 5; Rharnnus lycioides + en 5; Agrimonia eupatona + en 5; Cephalana leucantha + en 6; Avenula bromoides + en 6; Sanguisorba minor + en 6; Ficus carica + en 6; Teucrium webbianum + en 6; Gladiolus illyncus + en 8; Equisetum ramosissimum + en 8; Echinops ritro + en 8; Melica minuta + en 10; Cytisus scoparius reverchonii + en 10. 1. Albacete. Rambla en proximidades de "Las Mariquillas". XJ077332. 2. Albacete.Villagordo del Júcar. Barranca, 200 m antes de Central eléctrica, yendo desde El Galapagar. WJ821461. 3. Albacete. Villalgordo del Júcar. Barranca, 100 m pasada Central eléctrica, yendo desde El Galapagar. WJ821463. 4. Albacete. Villalgordo de 1 Júcar. Barranca, 900 m pasada Central eléctrica, yendo desde El Galapagar. WJ822471. 5. Albacete. De Alcalá del Júcar hacia Villavaliente 3 km. XJ3638. 6. Albacete. De Alcalá del Júcar hacia Villavaliente 3 km. XJ3638. 7. Albacete. Alcalá del Júcar. De Tolosa a Molino de Don Benito. XJ4140. 8. Albacete. Alcalá del Júcar. De Tolosa a Molino de Don Benito. XJ4240. 9. Albacete. Casas de Ves. De Barranco de Mingo a Tolosa. XJ 4440. 10. Albacete. Casas de Ves. De Barranco de Mingo a Tolosa. XJ 4440. Tabla No 3 Fresnedas con quejigos en la comarca de Ves. En zonas pertenecientes al nivel inferior del piso mesomediterráneo con manifiesto influjo setabense. Se sitúan en barrancas o umbrías. En el dominio del encinar de Rubio longifoliae-Quercetum rotundifoliae S. Pertenecen a la comunidad termo-mesomediterránea de encinares, propios del ombrocliina seco superior-subhúmedo, en su variante con fresnos: Rubio longifoliae-Quercetum rotundifoliae Costa, Pens & Figuerola 1983 subasociación fraxinetosum orni Costa, Peris & Figuerola 1983 En la formación son frecuentes elementos termófilos de Pistacio-Rlzamnetalia alaterni y, entre las plantas acompañantes, táxones setabenses o de influ- jo setabense como Erica multiflora, Cistus creticus. Thymus piperella, Globularia alypum y Euphorbia segetalis. Comunidades muy emparentadas con Fraxino orrzi-Quercetum fagineae Rivas Goday & Rigual in Rivas Goday. Borja, Esteve, Galiano, Rigual & Rivas-Martínez 1960 corr. Rivas-Martínez 1972 subasociación "térmica lentisceto-cocciferetosum" Rivas-Martínez & Borja (vd. Rivas Goday & al. 1959: 376) definida para las serranías valentinas. Sin embargo. la mayor abundancia de elementos de Quercetea ilicis frente a los de Quercolfagetea y el ombroclima seco superior-subhúmedo del territorio nos inclina a ubicarlas entre los encinares de Rubio longifoliae-Quercetum rotundifoliae. Unas veces presentan la apariencia de encinares con quejigos y fresnos, otras de fresnedas con quejigos y encina, según la especie dominante. TABLA N" 3 Rubio longifoliae-Quercetum rotundifoliae Costa, Peris & Figuerola 1983 subas.fraxinetosurn orrti Costa, Pens & Figuerola 1983 Otras compañeras: Rosmarinus ofíicinalis 1.1 en 1; Sanguisorba nipicola 1.1 en 1; Dictainnus hispanicus 1.1 en 1; Aphyllanthes monspeliensis 1.1 en 3; Leuzea conifera + en 1; Globularia alypum + en 1; Genista scorpius + en 1; Dorycnium pentaphylluin + en 1; Arostolochia pistolochia + en 1; Linum suffruticosum + en 1 ; Melica niinuta + en 1; Pinus halepensis + en 2; Euphorbia segetalis + en 3. 1. Albacete. Villa de Ves. Casa del Zurdo. XJ5340. Fresneda. 2. Albacete. Villa de Ves. Casa del Zurdo. XJ5440. Fresneda. 3. Albacete. Villa de Ves. De Castillo de Don Sancho a Casa del Francés. XJ5540. Fresneda. PROPUESTA Al amparo de la actual Ley Autonómica de "Conservación de la Naturaleza" de fecha 26-V1999, proponemos se inicien las acciones legales conducentes a constituir Microreservas en las localidades siguientes: 1. Barranca situada en Villalgordo del Júcar. Cuenca. a la izquierda del cauce, paraje denominado "Molino del Francés" (UTM: WJ821463) a 650-690 m.s.n.m. Existe un pequeño quejigar en muy buen estado de conservación. donde hay ejemplares corpulentos, amenazado por fuerte presión antrópica. 2. En las bajadas hacia el Júcar desde Villavaliente. junto a la carretera comarcal CM3254 (XJ 3638). Ladera N y NW y fondo de barranco. a 720-700 m.s.n.m. Se encuentran pequeñas extensiones de quejigos de talla media. 3. En las umbrías situadas entre el Molatón y Cerro de la Punta (Higueruela) (XJ 39174017). Hay una banda de quejigar situada entre 1150 y 1080 m.s.n.m. a lo largo de centenares de metros. Zona amenazada por plantaciones de Pil~ushalepelzsis. 4. Algunas parcelas de fresnedas con quejigos o de encinares térmicos con fresnos seleccionadas en la Comarca de Ves susceptibles de dicha figura de protección. No obstante. esta comarca. por sus valores naturales y paisajísticos, puede hacerse merecedora de una protección más integral y extensa con la figura de Reserva natural. SINTAXONOM~ADE SERIES DE VEGETACI~NDE QUEJIGARES Y ENCINARES MESOMEDITERRÁNEOS BASÓFILOS EN ESPAÑA PENINSULAR A. Quejigares mediterráneos basófilos: C1. Querco-Fagetea Br.-Bl. &Vlieger in Vlieger 1937 O. Quercetalia pubescentis Klika 1933 Al. Aceri granatensis-Quercion fagineae (Rivas Goday, Rigual & Rivas-Martínez in Rivas Goday, Borja. Esteve, Galiano. ~ i & ~~ ~i ~ ~ 1960) l ~ ~ ~ á r t í n e z1987 Cephalanthero rubrae-Quercetum fagineae Rivas-Martínez in Rivas Goday, Borja, Esteve. Galiano, Rigual & Rivas-Martínez 1960 con. Rivas-Martínez 1972 (Serie supra-mesomediterránea castellano-alcarreño-manchega subhúmedo-húmeda basófila del quejigo) Violo willkommii-Quercetum fagineae Br.-BI. & O. Bolos 1950 corr. Rivas-Martínez 1972 (Serie supra-mesomediterránea tarraconense, maestracense y aragonesa subhúmedo-húmeda basófila del quejigo) Daphno latifoliae-Aceretum granatensis Rivas-Martínez 1965 (Serie supra-mesomediterránea bética subhúmedo-húmeda basófila del quejigo) Fraxino orni-Quercetum fagineae Rivas Goday & Rigual in Rivas Goday. Borja, Esteve, Galiano, Rigual & Rivas-Martínez 1960 corr. Rivas-Martínez 1972 (Serie meso-supramediterránea setabense subhúmedo-húmeda basófila del quejigo) B. Encinares mesomediterráneos basófilos: C1.- Quercetea ilicis ~ ~ Br.-Bl. ~ ex A.í O. Bolos ~ 1950~ O. Quercetalia ilicis Br.-Bl. ex Molinier 1934 em. Rivas-Martínez 1975 Al. Quercion ilicis Br.-B1. ex Moliner 1934 em. Rivas-Martínez 1975 Subal. Quercenion ilicis Viburno tini-Quercetum ilicis (Br.B1. ex Molinier 1934) Rivas-Martínez 1975 (Serie mesomediterránea catalana seca de la alsina (Q. ilex ilex) Subal. Quercenion rotundifoliae Rivas Goday in Rivas Goday. Borja. Esteve. Galiano. Rigual & Rivas-Martínez 1960 em. Rivas-Martínez 1975 Asparago acutifolii-Quercetum rotundifoliae Rivas-Martínez. Fernández González, Loidi. Lousa & Penas 2001 (Bupleuro rigidi-Quercetum rotundifoliae Rivas-Martínez 1982) (Serie mesomediterránea manchega y alcarreña seca basófila de la encina) ~ Al. Quercion broteroi Br.-Bl., P. Silva & Rozeira 1956 em. Rivas-Martínez 1975 corr. Ladero 1974 Subal. Paeonio broteroi-Quercenion rotundifoliae Rivas-Martínez in RivasMartínez. Costa & Izco 1986 Paeonio coriaceae-Quercetum rotundifoliae Rivas-Martínez 1965 (Serie mesomediterránea bética marianense y araceno-pacense seco-subhúmeda basófila de la encina) Castroviejo, S. et al. (eds. ) 1986-2000. Flora Ibérica. Vols. 1. 2, 3, 4, 5, 6, 7 y 8. Real Jardín Botánico-C.S.I.C. Madrid. Costa. M.; Peris, J. B. y Figuerola, R. 1983. Sobre los carrascales termomediterráneos valencianos. Lazaroa 4: 37-52. Madrid. Ley 911999 de 26 de mayo, de"Consewación de la Naturaleza".D.O.C.M. No 40. 12-VI- 1999 Toledo. López González, G. 1976. Contribución al conocimiento fitosociólogico de la Serranía de Cuenca. Anales Inst. Bot. Cavanilles 33: 5-87. Madrid. Mateo Sanz, G. 1983. Estudio sobre laflora y vegetación de las Sierras de Mira y Talayuelas. Publ. M.A.P.1- I.C.O.N.A. Monografías, No 31. 290 pp. Rivas Goday, S.; Borja, J.; Esteve, F.; FemándezGaliano, E; Rigual, A. y Rivas-Martínez, S. 1960. Contribución al estudio de la Quercetea Al. Querco rotundifoliae-Oleion sylvestris Barbéro, Quézel & Rivas-Martínez in Rivas-Martínez, Costa & Izco 1986 Rubio longifoliae-Quercetum rotundifoliae Costa. Peris & Figuerola 1983 (Serie termo-mesomediterránea setabense, valenciano-tarraconense y murciano-almeriense seco-subhúmeda basófila de la encina) ilicis hispanica.. Anales Inst. Bot. Cavanilles 17 (2) : 285-403 Madrid. Rivas Martínez, S.; Gandullo, J. M; Serrada. R. y col. 1987. Memoria del mapa de series de vegetación de España. 1:400.000. Publ. ICONA. Serie Técnica. 268 pp. Madrid. Rivas-Martínez, S.; Fernández González, F.; Loidi, J.; Lousa, M. y Penas, A. 2001. Syntaxonornical checklist of vascular plant communities of Spain and Portugal to association level. Itinera Geobotanica 14. 341 pp. Tutin, T. G. et al. (eds. ) 1964-1980. Flora Europaea, Vols. 1-5. Cambrigde University Press. Cambrigde. Valdés Franzi, A,; Alcaraz Ariza, F. y Rivera Núñez, D. 2001. Catálogo de plantas vasculares de la provincia de Albacete (España). Instituto de Estudios Albacetenses, Ser. 1, Estudios, No 127. Albacete. 304 pp. Fotografía 2: Panorámica del Cerro de la Punta desde El Molatón. En la ladera N, a la izquierda de la imagen, banda de encinar con quejigos entre los 1050-1100 m., por encima de repoblaciones de pino carrasco. Higueruela (Albacete). Fotografía 3: Quejigar denso en ladera N de barranca. Villalgordo del Júcar (Albacete). Fotografía 4: Bosque denso de fresnos con quejigos, madroños y algunas encinas. Ladera expuesta al N del Valle del Júcar. Villa de Ves (Albacete). Lavandula stoechas subsp. pedunculata, Rosmannus officinalis y Thymus vulgaris EN UN ÁREA RECREATIVA CON SUELO COMPACTADO *M. LÓPEZ *L. PULIDO *M. ANDRÉS **S. GATTUSO * E.T.S.I. Agrónonios. Universidad de Castilla-La Mancha. Albacete. España ** Facultad de Ciencias Bioquímicas g Farniacéuticas. Universidad Nacional de Rosario. Rosario. Argentina RESUMEN ABSTRACT El pisoteo es un impacto ambiental importante especialmente en relación al incremento del uso en zonas recreativas. Los efectos del pisoteo se han relacionado con la reducción de la cobertura vegetal. la pérdida de diversidad del ecosistema y la sustitución de especies sensibles por otras más resistentes. En este trabajo se realiza un estudio morfoanatómico de las raíces de tres especies de la familia Labiatae: Lavarldula stoechas subsp. peduizculata, Rosinarincis oficinalis y Thymus vulgaris. Son especies predominantes en el sotobosque del área mediterránea del sudeste peninsular. La zona de estudio está situada entre los términos municipales de San Clemente (Cuenca) y Villarrobledo (Albacete), y pertenece a pinares de Pinus pinea. Se recogieron muestras del material vegetal necesario para el estudio, tanto en el Área Recreativa como fuera de ella, en el Pinar, donde no hay influencia antrópica, con el objetivo de comparar si existen diferencias morfo-anatómicas entre los ejemplares sometidos o no a la compactación. Los ejemplares de las tres especies situadas en el Área Recreativa, presentan en las raíces adaptaciones morfológicas a los impactos mecánicos: una rápida bifurcación de la raíz principal y raíces laterales suculentas y superficiales. No se han encontrado notables diferencias en la anatomía de los órganos radiculares, tal como apuntan otros autores. sí se aprecia la peridermis desarrollada en un estadio temprano en los ejemplares sometidos a compactación del suelo. Trampling on plants results in negative environmental impact, especially in recreation areas. The effects of trampling have been linked to a reduction in plant cover, loss of diversity in ecosystems, and substitution of delicate species for more resistant ones. This morpho-anatomic study was carried out on the roots of three species of the Labiatae: Lavandula stoechas subsp. pedunculata, Rosmariizus oficiitalis and Thyrnus vulgaris. These species predominate in the understorey found in the Mediterranean area of the southeastern part of the Iberian Peninsula. The zone studied was made up of Pilzus pinea pine groves located near the town of San Clemente (Cuenca) and Villarrobledo (Al-bacete), Spain. Sainples necessary for the study were collected within a recreation area where trainpling was evident and also in the surrounding forest where no trampling had occurred in order to compare morpho-anatomic differences in the two groups of samples. The sainples from the three species in the recreation area showed morphological adaptations as a result of trainpling. These adaptations took the form of a rapid forking of the main root and also of the succulent and superficial lateral roots. As indicated previously by other authors, no substantial differences were found in the anatomy of the radicular organs. However. in samples subjected to trampling, the periderm was observed to develop at an early stage. Palabras clave: Cambios morfo-anatómicos; raíz: Familia Labiatae. Keywords: Morpho-anatomic changes: root: Labiatae Family. El pisoteo es un impacto ambiental importante debido al uso creciente de los espacios naturales para realizar actividades recreativas. En las últimas décadas, se han realizado diversos estudios que ponen de manifiesto la influencia de la intensidad de pisoteo humano sobre el suelo y la vegetación, analizando la reducción de la cobertura vegetal y la pérdida de diversidad del ecosistema. con la sustitución de especies sensibles por otras más resistentes (Liddle 1991: Cole 1987, 1995; Marion y Cole. 1996, etc.). La zona de estudio está situada entre los términos municipales de San Clemente (Cuenca) y Villarrobledo (Albacete), y pertenece a pinares de Piillrs piizea L. que señalan el límite de ambas provincias. En dichos pinares se encuentra el área recreativa "Pinar Nuevo" que está sometida a compactación del suelo por efecto del pisoteo de los visitantes (Fig. 1). OBJETIVOS En este trabajo se ha realizado un estudio morfo-anatómico de 3 especies de la familia Labiatae: Lava?zdula stoeclzas subsp. pedunculatll (Miller) Samp. ex Rozeira. Rosmarinus oficinalis L.y Thyin~isvulgaris L. Los ejemplares se han reco- gido en el Área Recreativa y en el Pinar, donde apenas hay influencia antrópica. De esta manera se puede comparar si existen cambios morfológicos o anatómicos en el sistema radicular a causa del impacto de compactación. Las técnicas llevadas a cabo en el laboratorio son las convencionales para este tipo de trabajos: fijación, deshidratación total, inclusión en parafina. realización de los cortes con micrótomo rotativo manual. tinción Gerlach (1969) y safranina fastgreen. RESULTADOS El sistema radicular de los ejemplares de lavándula del Área Recreativa presentan una rápida bifurcación de la raíz principal ocasionando raíces secundarias suculentas, con gran expansión lateral y muy superficiales; a diferencia de los ejemplares del Pinar que tienen una posterior bifurcación de la raíz principal. y predominancia de raíces verticales. La raíz principal de los ejemplares de romero del Área Recreativa se bifurca en 2 o más raíces laterales vigorosas, con amplia expansión lateral. En el Pinar. la raíz es axonomorfa de la que surgen raíces laterales. Las raíces de los ejemplares de tomillo del Área Recreativa presentan ramificación lateral, y carecen de una raíz principal. Raíces laterales suculentas y horizontales, próximas a la superficie. Los ejemplares del Pinar, carecen de raíz principal y sus raíces laterales son más delgadas y profundas (Fig. 2). Con respecto a los cambios anatómicos. entre los ejemplares del Pinar y del Área Recreativa no se han encontrado notables diferencias (QueirozVoltan et al., 2000) en estudios realizados con diseños experimentales y con niveles de compactación semejantes a los de nuestro estudio, se observa, no obstante, la peridermis muy desarrollada que permite a las raíces de estas especies soportar mejor los impactos mecánicos ocasionados por el pisoteo (Fig. 3). CONCLUSIONES Las raíces de las 3 especies de la familia Labiatae, situadas en el Área Recreativa. presentan adaptaciones morfológicas a los impactos ocasionados por el pisoteo: una rápida bifurcación de la raíz principal y raíces laterales suculentas y superficiales. El cambio anatómico más notable es la diferenciación temprana de la peridermis. Cole, D.N. 1987. Effects of three seasons of experimental trampling on five montane forest communities and a grassland in western Montana. USA. Biological Consewation 40: 2 19-244. Cole. D.N. 1995. Experimental trampling of vegetation 1. Relationship between trampling intensity and vegetation response. Jounzal of Applied Ecology 32:203-214. Gerlach, D. 1969. A rapid Safranin-Crystal. VioletLight, Green staining sequence for paraffin sections of plant material. Staiiz Techizology 44:210-211. Liddle. M.J. 1991. Recreation ecology: effects of trampling on plants and coral. Treizds iii Ecology and Evolution 6: 13-17. Marion. J. L. and Cole, D.N. 1996. Spatial and temporal variation in soil and vegetation impacts on campsites. Ecological Applications 6: 250-530. Queiroz-Voltan, R. B.; Dos Santos Sevá Nogueira, S. and Coelho de Miranda. M. A. 2000. Aspectos da estrutura da raiz e do desenvolvimento de plantas de soja em solos compactados. Pesquisa Agropecuaria Brasileira 35(5):929-938. Fig. 1: Área Recreativa "Pinar Nuevo" Fig. 3: C. T. de raíz de Lavancdula stclechas subespecie pedunrula#a. Ecología PONENCIA MARCO ENTENDIDOS COMO ECOSISTEMAS Carlos MONTES Departamento Interuniversitario de Ecología. Universidad Autónoma de Madrid Las Zonas húmedas o Huinedales están en la cresta de ola de la conservación de los espacios naturales. Prácticamente todos los gobiernos y asociaciones ambientalistas incluyen en sus agendas de actuación programas específicos encaminados a su protección. Los Humedales constituyen el único tipo de ecosistema en el planeta que posee un Convenio Internacional para su conservación: El Convenio de Ramsar. De todas formas su futuro se encuentra seriamente amenazado. Históricamente, las culturas humanas han estado estrechamente relacionadas y dependientes de sus recursos (alimentación, minerales, fuente de agua, materiales para la construcción, etc.) pero este diálogo hombre-humedal ha ido cambiando y se está produciendo una intensa y progresiva pérdida y degradación de sus funciones. Como ejemplo, en España hemos perdido más del 70% de nuestras tierras encharcables. A pesar de que se han dictaminado leyes específicas para su conservación se sigue degradando y perdiendo superficie encharcable. De esta forma, aunque un determinado humedal sea objeto de una figura legal de protección. incluso como la de Parque Nacional. no tiene asegurado su conservación a medio o largo plazo. Son los modelos de gestión basados en el conocimiento y protección de los procesos ecológicos y especies esenciales de organismos los que garantizan el mantenimiento de su integridad y por tanto el suministro de servicios a los sistemas socioeconómicos. Una mala gestión legal de un humedal protegido puede llegar a ser peor que mantenerlo desprotegido. Por otra parte, hoy en día. la conservación de los humedales constituye algo más que un conjunto de programas y medidas para la protección de unos paisajes singulares. Suponen unos excelentes escaparates de la viabilidad o fracaso de filosofías de conservación que actualmente están de moda y que se desarrollan sobre nuestro medio natural. Si estos programas de conservación se aplican sin una base científica sólida. nos podemos encontrar con una nueva forma de destrucción de estos ecosistemas. Esta conferencia intenta poner de manifiesto en primer lugar la personalidad ecológica de las tierras encharcables, justificando su conservación por su capital natural es decir, por los bienes y servicios que podemos obtener. En segundo lugar, se pretende mostrar las patologías más importantes que padecen nuestros humedales naturales protegidos que disminuyen o limitan el desarrollo de estrategias efectivas de conservación. En último término se intenta defender y promover una aproximación ecosistémica a su gestión como la vía más racional y eficaz de conservar la integridad ecológica y "salud" de nuestros paisajes del agua. COMUNICACIONES ORALES NOTAS SOBRE LA ECOLOGÍA DE LA FLORA MEDICINAL DE LA PROVINCIA DE ALBACETE Alonso VERDE LÓPEZ Diego RIVERA NÚÑEZ José FAJARDO RODRIGUEZ Concepción OBÓN DE CASTRO RESUMEN ABSTRACT Las 386 especies de plantas medicinales de la provincia de Albacete se distribuyen en nueve tipos de hábitats diferentes. Se presentan de forma mucho más numerosa en los hábitats transformados por el hombre (huertos, caminos, etc.). Las familias de plantas no contribuyen en la misma proporción a la flora medicinal por lo que las Labiatae, Cistaceae y Rosaceae se encuentran bien representadas mientras que Gramineae y Leguminosae no. The 386 medicinal plant species living in the province of Albacete grow in nine different habitat types. These are more numerous in secondary habitats deeply transformed by human activities (orchards, fíelds, roadsides,...). The relative contribution of each plant family to the medicinal flora of Albacete is not uniform. Labiatae, Cistaceae and Rosaceae are over-represented whereas Gramineae and Leguminosae are under-represented. Palabras clave: Farmacoetnobotanicidad. Hábitats. Principios activos. Keywords: Habitat. Medicinal Plants. Active principies. Existen en Albacete casi 400 especies de plantas medicinales (386). pertenecientes en gran parte a familias como las Labiadas, Compuestas. Leguminosas, Rosáceas y Umbelíferas. lo que se traduce en una rica y variada flora medicinal. constituyendo un 13.27% del total de la flora provincial. Dada la gran diversidad geográfica de la provincia de Albacete. existen numerosos ambientes que albergan a esta rica flora medicinal. Por esta razón cabría preguntarnos si existe alguna relación entre el número de especies medicinales y hábitat. así como si las plantas medicinales que se desarrollan en cada hábitat presentan un denominador común en principios activos. Se ha elaborado el catálogo de la flora medicinal de la provincia de Albacete que comprende datos publicados como los de Cantero, Gaudens y González, (1987); Verde (1997). Verde y Fajardo (1998), Verde. Rivera y Obón. (1997 y 1998). Verde y cols. (1998). Fajardo y cols (2000). Fajardo y Verde (2001). Obón y Rivera (1997) y Sánchez y cols (1994). y se ha comparado con el catálogo florístico de la provincia de Albacete de Valdés. Alcaraz y Rivera (2001). Los datos sobre los hábitats preferentes de las diversas especies provienen de Castroviejo y cols. (1986, 1990. 1993a, 1993b. 1997a, 1997b. 1998, 1999, y 2000) y para las especies no incluidas en la citada obra se recurrió a Bolós y Vigo (1984. 1990 y 1995) y Mateo y Crespo (1995). En el cálculo de la Fármaco-Etnobotanicidad se consideraron como faiililias "numerosas" aquellas que tenían 10 o más especies en la flora medicinal o 45 o más especies en la flora general. Estos números se establecieron considerando el rango de variación mostrado por las diferentes familias. En la Tabla 1 se recogen los nueve hábitats preferentes de las diferentes familias. así como el número de especies medicinales usadas en la provincia de Albacete. En total comprende 80 familias y 386 especies que se emplean en la medicina popular en las diferentes localidades de nuestra provincia. 'I$bEri 1.Riqueza en especies de flora medicinal de los diferentes tipos de hábitats. * 1.Espacios abiertos, pedregales y roquedos. 2. Zonas umbrosas protegidas de vientos, cuevas y abrigos del ganado. 3. alas' faestales y bosques de ribera 4. Ríos y arroyos, lagos, acequia y humedales. 5. es muy ralos y dehesas. 6, Bosques densos caducifolios y peremifolim. 7. Estepas y campos de cereales. 8. Huertos y jardines. 9. Linderos, ribazos, metas y caminos. Si observamos las especies medicinales existentes en cada uno de los hábitats estudiados (Fig. l), podemos apreciar que los espacios profundamente alterados por el hombre (huertos, caminos, pastizales,...) constituyen los mayores reservorios de flora medicinal. E?igu9 1.Número de especies medicinales @terid6fitos,gimnospermas y angiospemasts)en cada tipo de Mbitat. Muchas de estas especies medicinales no son exclusivas de un solo hábitat, pudiendo ocupar más de uno. Así, vemos en la Tabla 2 la riqueza medicinal detallada para las familias m8s representativas del total de las estudiadas en cada uno de los diferentes hábitats. Tabla 2. Hábitats preferentes de las especies pertenecientes a las familias más relevantes de la flora medicinal. Especies totales HABITATS FAMILIA BENACEAE 3 3 GERANIACEAE GLOBULARIACEAE 1 GRAMINEAE 1 1 1 1 6 8 2 10 APAVERACEAE Si estudiamos los principios activos que caracterizan y justifican la utilización medicinal de cada una de las familias seleccionadas, para lo que se han utilizado los trabajos de Mulet (1997) y Evans (1989), podemos observar que dependiendo de los diversos ambientes la presencia mayoritaria de especies. familias y sustancias activas varía. Esta variación de la composición mayoritaria relacionada con los hábitats había sido puesto de manifiesto desde antiguo. Harborne (1 972. 1978, 1993) muestra numerosos ejemplos relacionados con la presencia de sustancias como respuesta a factores ambientales o consecuencia de la coevolución de plantas y herbívoros. 1. ESPACIOS ABIERTOS, PEIIREGALES Y ROQUEDOS Los hábitats generalmente soleados. pedregosos y secos se presentan en espacios iná5 o menos dependientes de la acción humana (ganadería extensiva. talas. rozas. incendios, etc.). Por 10 común aparecen ocupados por plantas ricas en aceites esenciales y flavonoides como los tomillos (Labiadas). o con secoiridoides amargos como la hiel de la tierra (Gencianáceas) (Tabla 3). Tabla 3. Abundancia de especies medicinales en los espacios abiertos Familia Cistaceae C'onil7osit:ie Especies medicinales 8 6 Ephedi-aceae Gentinilacene L,ahiatae 2 3 29 Liliaceae 4 Principios inmediatos Oleoresinas, terpenos y f1:ivonoides Aceites esenciales, lactonas sesq~iiterpénicas,c~imarinas, Ilavonoides Alcaloides (efedrina) Secoiridoides amargos y icidos fenólicos polifc.i~olcs Aceites esenciales: saponin:is: c~iri~:iri~~as; flavonoides Alcaloides. cardenólidos. flavonoides, sustancias cianogenéticas; esteroles 11. ZONASUMBROSAS PROTEGIDAS DE VIENTOS, CUEVAS Y ABRIGOS DEL GANADO Son espacios poco soleados, donde pueden desarrollarse especies que no soportan la acción directa de los rayos solares (ver Figura 6. Por lo general como consecuencia de la acumulación de polvo y de su uso como abrigos del ganado, suelen presentar una fuerte carga de sales minerales en el suelo. por lo que albergan especies de las denominadas "nitrófilas". En este tipo de hábitat encontramos en general plantas alcaloideas especialmente de la familia Solanáceas (Tabla 4). Tabla 4. Abundancia de especies medicinales en los espacios umbrosos. Familia Rhamnaceae Solanaceae :II. ORLAS FORESTALES Y Principios inmediatos Glucosidos quinonaz alcaloides peptidicos Alcaloides saponinas cumarinas flavonoides Especie3 medicinales 3 2 con otros C O ~ ~ U ~ Sfenólicos ~ O S precursores de 10s propóleos como en los chopos (Rivera y cols., 1997), o con sustancias tan conocidas como el ácido salicílico de las cortezas de los sauces y de las raíces del género Spiraea (Rosáceas) (Tabla 5). O BOSQUES DE RIBERA Las orlas forestales y bosques de ribera (ver Figura 8) suelen estar dominadas por especies con flores ricas en flavonoides como las del espino albar, Tabla 5. Abundancia de especies medicinales en las orlas forestales y bosques de ribera. Familia Equisetaceae Liliaceae Especies medicinales 2 -1 Moraceae Plantaginaceae Rhamnaceae Rosaceae 2 3 9 Salicaceae Ulmaceae 2 2 1 Principios inmediatos Acidos fenólicos, flavonoides, sales minerales Alcaloides, cardenolidos, flavonoides, sustancias cianogenéticas, esteroles Enzimas proteolíticos. furocumarinas fotosensibilizantes. Alcoholes y rnucilagos Glucósidos, quinonas, alcaloides peptidicos Flavonoides. glucósidos cianiogenéticos: saponinas, taninos, flavonoides, terpenos y mucilagos. Glucósidos fenblicos (salicina, populina) Mucílagos IV. =OS Y ARROYOS, LAGOS, ACEQUIAS Y HUMEDALES En estos ambientes (ver Figura 3) permanentemente inundados crecen plantas con mucfiagos y flavonoides como el malvavisco (Malváceas), plantas con aceites esenciales como el apio (Umbelíferas) y plantas ricas en sílice como la cola de caballo (Equisetáceas) (Tabla 6). Tabla 6. Abundancia de especies medicinales en los ríos, arroyos, lagos y acequias. En estos ecosistemas intensamente modificados por el hombre destacan las plantas ricas en glucósidos cianogenéticos, alcaloides y principios amargos. directamente relacionados con la defensa frente a los herbívoros (Harborne, 1972, 1978. 1993) (Tabla 7). Tabla 7. Abundancia de especies medicinales en los pastizales y dehesas. Alcaloides, casdenólidos, flavonoides, sustancias VI. BOSQUES DENSOS CADUCIFOLIOS Y PERENNIFOLIOS Son ambientes propicios para el desarrollo de especies ricas en glucósidos como las de la familia Liliáceas, Peoniáceas o Asclepiadáceas. Estas sustancias se acumulan por lo general en los órganos de reserva subterráneos (Tabla 8). Tabla 8. Abundancia de especies medicinales en los bosques. VII. ESTEPAS Y CAMPOS DE CEREALES En las estepas (ver Figura 5) dominan plantas ricas en resinas, flavonoides, lactonas sesquiterpénicas. La mayor parte del territorio albaceteño está ocupado por campos de cultivo, en su mayoría dedicados a cereales y viña. En estos parajes dominan plantas alcaloideas como los ababoles (ver Figura 4), las lechitreznas ricas en poliisoprenoides, plantas ricas en sílice como la avena (Tabla 9). Tabla 9. Abundancia de especies medicinales en las estepas y campos de cereales. Familia Convolvulaceae Especies medicinales 1 Cruciferae Gramineae 3 6 Papaveraceae Ranunculaceae 1 1 Principios inmediatos Glucí~sidos,alcaloides, Bcidos fenólicos, resinas, saponinas triterpénicas Mucilagos, glucósidos Saponinas, minerales, ácidos fenólicos, flavonoides, principios cianogenéticos Alcaloides Alcaloides VIII. HUERTOS Y JARDINES Son espacios reservados (ver Figura 7) a especies introducidas o a plantas recogidas del campo y cultivadas cerca de la casa por su uso frecuente: té de huerta, romero, poleo, hierba buena. manzanilla, i~iaravillas,la hierba de Santa María. el sanalotó. Aquí no solainente incluimos las especies cultivadas. sino también las que crecen silvestres en los huertos de regadío (Tabla 1 0). Tabla 10. Ab~indanciade especies medicinales en los huertos y jardines. Solanaceae Umbelliferae 7 IJrticaceae 2 1X. LINDEROS, RIBAZOS, 5 Alcaloides, saponinas. cuiiiai-inas, flavonoides Aceiies esenciales, alc:iloides, curnai-inas, flcivonoides, terpenos, fui-ocuinni-inas scsquiteipéiiicas Acidos 01-gúnicosy cni-otcnoides CUNETAS Y CAMINOS Estos espacios ocupados en su mayoría por plantas nitrófilas, ricas en mucílagos como la malva, plantas con flavonoides como el marrubio, plantas con lactonas sesquiterpénicas como las arzollas, plantas con saponósidos como los verbascos o las hierbas de la virgen, con aceites esenciales como el hinojo, con hipericina como el pericón (Tabla 1 1). Tabla 1 1 . Abundancia de especies inedicinales en los rriárgenes de caminos. Familias Bosaginaceae Especies medicinales Cni~ophy1lace:ic Cc>r~ipositae -I 16 Guttifcsae 2 Labiatae 19 Mal~.aceae Polygonaceae Iiaiiunculaccac Scsophulasiaceae 3 6 -I 6 Umhellifcrac -I IJi-iicaceae 3 -I Principios inmediatos Naftoquinonas, tciiiinos, Ucidos i'cri0licos, alcaloidcs pin-olizidinicos S~iponiiins Aceites esenciales. I:icion:is sc<]iiiici-pénicac,cumai-inns, Il~ivoiioidcs Aceites tciybiiicos, pigmcnios i-c)josdiaiitr6nicos fotoscnsil~iliza~itcs, alcaloides Aceites escncialcs, s;ipoiiiiins, cuiiiarinas, poliknolcs y Iln\,onoidcs Mucilagos, snpoiiiiins, toninos, Ucidos ICn0licc)s Aiiiociaiios, Ilnvonas Alc:iloides 1letcrósidos cal-diolfiiiicos, glucósidos cianogenL:ticos, sapoiiiiias 11-iicrpbiiicas. Aceites esencinlcs, cilcaloidcs, cuniai-inas, glucorónidos de Ilavonoidcs, Icilxiios, f~isoc~iri-iasirias scsq~iitci-pbnicns Acidos oi-ginicos y cnrotciioiilcs DIFERENCIAS EN LA RIQUEZA EN ESPECIES MEDICINALES DE LAS FAMILIAS MÁs IMPORTANTES DE LA FLORA VASCULAR DE LA PROVINCIA DE ALBACETE Se puede apreciar que no existe una relación constante entre el número de especies totales de cada familia y el de especies medicinales (Tabla 12). Así se aprecia que las Labiadas, Cistáceas y Rosáceas contribuyen proporcionalmente más a la flora medicinal que familias muy bien representadas en la flora como las Gramíneas o las Leguminosas. Tabla 12. Fármaco-Etnobotanicidad relativa de las diferentes familias de plantas más relevantes en número de la flora de Albacete. Cyperaceae Orchidaceae 1 56 57 1 O Esto es bien conocido desde antiguo y ha sido puesto de manifiesto de forma cuantitativa por Moerman (1998) utilizando una recta de regresión como elemento que permite diferenciar de manera clara las familias sobreutilizadas de las infrautilizadas. Hemos aplicado el mismo tratamiento gráfico a nuestros datos (Figura 2) y son muy diversas las razones que pueden explicar estas diferencias aunque la primera que se nos ocurre es la distinta composición química de unas y otras. Generalmente la riqueza en principios activos de utilidad medicinal es algo peculiar de cada familia de plantas y no de géneros 1 0.017 O concretos, por lo que cabe pensar que se utilizara un mayor número de especies de entre las familias mejor dotadas de un repertorio de sustancias medicinales. Esta explicación puede convenir con la escasa proporción de gramíneas medicinales, pero no así con la de leguminosas. En este caso, es bien conocido que las leguminosas son muy relevantes en la flora medicinal mundial, cabe recordar el problema de la toxicidad. Es posible que la toxicidad extrema de algunas leguminosas haya llevado a la población local a descartar su uso. Figura 2. Relación entre el número de especies totales y el número de especies medicinales. Relación de Moerman. O 100 - 200 300 Totales 400 500 CONCLUSIONES La flora medicinal de la Provincia de Albacete es relativamente rica con 386 especies, que son algo más de un 13% de la flora provincial. Las especies medicinales se encuentran distribuidas por muy diversos tipos de hábitats, se presentan de forma mucho más numerosa en los hábitats transformados por el hombre (huertos, caminos, etc.), por el contrario, aparecen menos especies medicinales en zonas umbrosas y protegidas y campos de cereales. Las familias de plantas no contribuyen en la misma proporción a la flora medicinal por lo que las Labiadas, Cistáceas y Rosáceas se encuentran bien representadas, con un índice de Farmaco-etnobotanicidad más elevado, mientras que Gramíneas y Leguminosas no. No existe una relación constante entre el número de especies totales de cada familia y el número de especies medicinales. La riqueza medicinal en principios activos, probablemente es algo peculiar de cada familia por lo que se utilizan mayor número de especies de entre las familias mejor dotadas de un repertorio de principios activos. Bolós, O. y Vigo, J. (1984). Flora dels pai'sos catalans. Volumen I. Edit. Barcino. Barcelona. 736 pp. Bolós, O. y Vigo, J. (1990). Flora dels pai'sos catalans. Volumen II. Edit. Barcino. Barcelona. 921 pp. Bolós, O. y Vigo, J. (1995). Flora dels pai'sos catalans. Volumen III. Edit. Barcino. Barcelona. 1230 pp. Cantero, A., M. V. Gaudens y P. González (1987). Plantas Medicinales en la Sierra de Albacete. En anónimo (Eds.) Actas de las IV Jornadas de Etnología de Castilla-La Mancha, Albacete 1986: 348-358. Junta de Comunidades de Castilla-La Mancha. Toledo. 390 pp. Castroviejo, S. y cols. (1986). Flora Ibérica. Tomo 1. Real Jardín Botánico. CSIC. Madrid. 575 pp. Castroviejo, S. y cols. (1990). Flora Ibérica. Tomo 11. Real Jardín Botánico. CSIC. Madrid. 897 pp. Castroviejo, S. y cols. (1993a). Flora Ibérica. Tomo 111. Real Jardín Botánico. CSIC. Madrid.730 pp. Castroviejo, S. y cols. (1993b). Flora Ibérica. Tomo IV. Real Jardín Botánico. CSIC. Madrid. 730 pp. Castroviejo, S. y cols. (1997a). Flora Ibérica. Tomo V. Real Jardín Botánico. CSIC. Madrid. 320 pp. Castroviejo, S. y cols. (1997b). Flora Ibérica. Tomo VIII. Real Jardín Botánico. CSIC. Madrid. 730 pp. Castroviejo, S. y cols. (1998). Flora Ibérica. Tomo VI. Real Jardín Botánico. CSIC. Madrid. 592 pp. Castroviejo, S. y cols. (1999). Flora Ibérica. Tomo VI1 (1). Real Jardín Botánico. CSIC. Madrid. 578 pp. Castroviejo, S. y cols. (2000). Flora Ibérica. Tomo VI1 (11). Real Jardín Botánico. CSIC. Madrid. 730 pp. Evans, W. C. (1989). Trease and Evans' Pharmacognosy. Baillikre Tindall. London. 832 pp. Fajardo, J. y cols. (2000). Las plantas en la cultura popular de la provincia de Albacete. Instituto de Estudios Albacetenses. Albacete. 264 pp. Fajardo, J. y Verde, A. (2001). Catálogo, ecología y usos populares de las Rutáceas presentes en la provincia de Albacete. Sabuco 1: 163-179. Harborne, J. B. (ed.) (1972). Phytochemical Ecology. Academic Press, Londres. Harborne, J. B. (ed.) (1978). Biochemical Aspects of Plant and Animal Evolution. Acad. Press. Nueva York. Harborne, J. B. (ed.) (1993). Introduction to Ecological Biochemistry, 3rd edn., Academic Press. Londres. Mateo, G. y Crespo, B. (1995). Flora abreviada de la Comunidad Valenciana. Editorial Gamma. San Vicente (Alicante). 483 pp. Moerman, D. (1998). Native North American Food and Medicinal Plants: Epistemological Considerations: 69-74. En Prendergast. H. et al. (eds.). Plants for Food and Medicine. Royal Botanic Gardens. Kew. 438 pp. Mulet, L. (1997). Flora tóxica de la Comunidad Valenciana. Diputació de Castelló. Castelló. 630 PP. Obón, C. y Rivera, D. (1997). Usos tradicionales de rabogatos y zahareñas. Quercus, 142: 36-38. Rivera, D. y cols. (1997). Study of the flavonoids as chemotaxonomic markers in Populus (Salicaceae) of Spain. Preliminary results. Lagascalia 19(1-2): 813-818. Sánchez, M. D. y cols. (1994). Plantas útiles de la comarca de La Manchuela (Albacete). Colectivo de Escuelas Rurales de La Manchuela. Casas Ibáñez. 190 pp. Valdés, A., Alcaraz, F. y Rivera, D. (2001). Catalogo de plantas vasculares de la provincia de Albacete (España). I.E.A. Albacete. 304 pp. Verde, A. (1997). Estudio etnofarmacológico de las sierras de Segura y Alcaraz. Universidad de Murcia. Murcia. 176 pp. (Tesis de licenciatura inédita). Verde, A., Rivera, D. y Obón, C. (1997). Etnobotánica de las sierras de Alcaraz y Segura. Quercus, 132: 36-37. Verde, A., Rivera, D. y Obón, C. (1998). Etnobotánica de las sierras de Segura y Alcaraz: las plantas y el hombre. IEA. Albacete. 35 1 pp. Verde, A. y Fajardo, J. (1998). Usos tradicionales del torvisco. Quercus, 144: 38-39. Verde, A. y cols. (1998). Las plantas medicinales en la provincia de Albacete. Usos, creencias y leyendas. Zahora no 28. Diputación de Albacete. Figura 3. En los arroyos crecen plantas ncas en mucflagos, aceites esenciales y sílice. Figura 4. En los campos de cereales dominan plantas alcaloideas como los ababoles. Figura 5. En las estepas dominan plantas ncas en resinas, flavonoides y lactonas sesquiterpénicas. F i p á En las zooas nmbrosas y abrigos se desarroillrn plantas que no soportan la acci6n directa de los rayos solares. 1 F " i 7. Los peq11e5os huertos acogen una p variedad de plantas medicinales. Figura 8. Los bosques de ribera están dominados por especies con flores ricas en flavonoides. ESTUDIO Y CONTROL POBLACIONAL DE LOS DIFERENTES FOCOS DE LANGOSTA MARROQUÍ (DOCIOSTAURUS MAROCCANUS T.) EN EL CAMPO DE MANIOBRAS DE CHINCHILLA DE MONTEARAGÓN (ALBACETE, ESPANA) GÓMEZ LADRÓN DE GUEVARA, R.' CATALÁN HERNÁNDEZ, AOZNÚÑEZ ZORNOZA, A.Z VÉLAZ AZPILICUETA, M.2 PICAZO CÓRDOBA, M. 1.' ' E. T. S. de Ingenieros Agrónomos. Universidad de Castilla-La Mancha ' Delegación de Agricultura y Medio Ambiente de Albacete RESUMEN ABSTRACT La principal zona de aparición de langosta marroquí en la provincia de Albacete se localiza en el Campo de Maniobras de Chinchilla de MonteAragón. Para controlar sus poblaciones se han realizado durante los últimos años tratamientos fitosanitarios indiscriminados sobre grandes superficies de terreno, sin llevar a cabo ningún tipo de estudio previo que justificase su necesidad, lo que ha ocasionado fuertes agresiones medioambientales sobre la fauna. suelo y agua. Con el fin de reducir este tipo de impacto se propone la realización de un control racional de la población de langosta, en función del estudio de la distribución superficial de sus focos, la densidad de individuos. su etapa de desarrollo, su capacidad de desplazamiento a nuevas áreas y la existencia de cultivos próximos. The main appearance area of maroccan grasshoppers (D. maroccanus T.) in the province of Albacete is situated in the maneuvering field in Chinchilla de Monte-Aragón. In order to control their population, phytosanitary treatments have been carried out during the last years on great ground areas without maiung any previous study which justifies its necessity, what has caused strong environmental aggressions on fauna, ground and water. With the aim of reducing this kind of impact the canying out of a rational control of the grasshopper population in function of the study of the surface distribution of its focuses, the density of individuals, its growing phases, its moving capacity to new areas and the existence of nearby farming fields is proposed. Keywords: Control, population, grasshop- Palabras clave: Control, población, langosta. El presente trabajo "Estudio y control poblacional de los diferentes focos de langosta marroquí (Docio,stuurus marocca~zusThunberg. 18 15) en el Campo de Maniobras de Chinchilla de MonteAragón" nace del acuerdo de colaboración entre la Delegación de Agricultura y Medio Ambiente de Albacete (Junta de Comunidades de Castilla-La Mancha) y el Departamento de Ciencia y Tecnología Agroforestal (Universidad de Castilla-La Mancha). ante la necesidad de conocer la ubicación de los principales focos de langosta existentes en el Campo de Maniobras y proponer. cuando así sea necesario. mecanisnlos de control de dichos focos en el momento más adecuado. siempre que por el estudio de sus densidades poblacionales y desplazamientos podamos inferir que pueden constituir áreas potenciales de gregarización o bien ocasionar daños severos en la agricultura circundante. pers. Hasta ahora. la lucha tradicional contra la langosta marroquí en el Campo de Maniobras de Chinchilla de Monte-Aragón. donde el abandono del cultivo de las tierras debido a su utilización militar es un hecho incontestable, ha consistido en tratamientos fitosanitarios indiscriminados sobre grandes superficies de terreno. sin realizar estudios previos que justificasen su necesidad. ni delimitar la exacta localización de los focos ni la superficie a tratar y sin evaluar el estado de desarrollo de la población y el riesgo potencial de su gregarización. Este tipo de acción contra la langosta, por el tipo y la dosis del insecticida utilizado. ha ocasionado fuertes agresiones rriedioainbientales sobre las enormes superficies de terreno tratadas. Por ello. hemos considerado que sería de gran importancia implantar algún otro mecanismo de actuación que permita minimizar este tipo de impacto. En la provincia de Albacete, la principal zona de aparición de la langosta se localiza en el Campo de Maniobras de Chinchilla de Monte-Aragón. que se extiende sobre una superficie superior a las 14.300 hectáreas, a las que hay que sumar las ocupadas por la denominada Zona de Seguridad Militar. El citado Campo está situado entre las localidades de La Felipa, Casas de Juan Núñez, Higueruela. HoyaGonzalo y Chinchilla de Monte-Aragón. Además, se da la circunstancia de que debido a la extensión, características y régimen especial de protección que goza el terreno, es numerosa la fauna (principalmente aves) que sobre esta superficie se asienta. Entre la fauna alada que habita el Campo de Maniobras de Chinchilla de Monte-Aragón, junto a las aves esteparias, aparece una de las rapaces más emblemáticas del territorio europeo, el cernícalo primilla (Falco naumanni). La rápida regresión de sus poblaciones en nuestro país, debido a su carácter gregario, junto a la masiva e indiscriminada utilización de potentes insecticidas durante las últimas décadas, por parte del hombre, convierten a las colonias de cernícalo primilla del Campo de Maniobras de Chinchilla de Monte-Aragón en reductos de la especie de incalculable valor ecológico. Por ello, durante los últimos años, a pesar de los enormes esfuerzos que se han hecho desde la Delegación de Agricultura y Medio Ambiente de Albacete para potenciar el crecimiento de sus colonias, el número de individuos que las componen apenas se ha visto incrementado. Es un hecho constatado que la alimentación de esta rapaz es fundamentalmente insectívora, y que a lo largo del ciclo vital de la langosta (abril a julio) el sustento de los pnmillas adultos y sus crías se basa en gran medida en su consumo. Sin embargo, y a pesar de la protección de que gozan las colonias al estar ubicadas en construcciones situadas dentro de la zona de seguridad militar, en los últimos años no se ha detectado apenas aumento en el número de parejas que las componen debido, probablemente. a la falta de alimento causada por los tratamientos fitosanitarios que se han venido realizando de forma periódica contra la langosta. Ante esta perspectiva, poco favorable ni siquiera para garantizar el mantenimiento de las colonias de cernícalo primilla a medio plazo, desde la Delegación de Agricultura y Medio Ambiente y la Universidad Regional nos hemos propuesto realizar un control racional de la población de langosta, en función de: la distribución superficial de los distintos focos existentes, su proximidad a zonas de cultivo, sus desplazamientos a nuevas áreas, la densidad de individuos en cada momento del ciclo biológico y la fase en que se encuentran. Esperamos que el conocimiento de estas variables permita reducir la frecuencia tanto del número de tratamientos como de superficie tratada, minimizando el efecto nocivo que provocan sobre el resto de fauna, suelo y agua, y que favorezcan, a su vez. la aparición de nuevas colonias de cernícalo primilla en la zona. Sin embargo, las consideraciones anteriores sólo representarían una esperanzadora declaración de intenciones si no disponemos previamente de las herramientas necesarias para su ejecución. Para que nuestro proyecto de control de los focos de langosta del Campo de Maniobras de Chinchilla de MonteAragón sea efectivo y alcance el rango de iniciativa pionera en nuestro país, debe permitir, de forma paralela, el crecimiento de las poblaciones de cernícalo primilla. Para ello, tenemos que dotarlo de unos objetivos claramente definidos y de una metodología científica y rigurosa que permita su consecución. Finalmente, como epílogo a nuestra introducción sobre el tema de trabajo, debemos tener en cuenta en todo momento que las actuaciones que se deriven de este estudio no deben ir nunca encaminadas a la erradicación definitiva de la presencia de la langosta marroquí en el Campo de Maniobras de Chinchilla de Monte-Aragón; objetivo, por otra parte, prácticamente imposible de conseguir sin ocasionar previamente daños medioambientales de primera magnitud. No olvidemos que la superficie de terreno a que nos referimos representa una zona endémica de langosta, derivada del uso a que se ha sometido la tierra durante las últimas décadas, donde algo más de 6.000 hectáreas pueden ser consideradas como biotopos adecuados para la existencia y proliferación del insecto. Por ello, antes que la erradicación de la langosta, consideramos de mayor importancia su correcto control. Con el conocimiento de la densidad de individuos existentes en los diferentes focos y sus desplazamientos a nuevas áreas, en cada momento del ciclo biológico, podemos mantener controlada la población total del insecto utilizando las opciones que se ponen a nuestra disposición. y que más adelante se detallan. De esta forma, podemos hacer compatible la presencia controlada de la langosta en el Campo de Maniobras de Chinchilla de MonteAragón con la protección de los cultivos adyacentes. Además, sólo así podremos disponer de un eslabón en la cadena trófica en el que puedan basar gran parte de sus necesidades alimenticias una numerosa fauna, entre la que se incluye el cernícalo primilla. OBJETIVOS El trabajo realizado en la zona de estudio, que comprende tanto al Campo de Maniobras de Chinchilla de Monte-Aragón como la Zona de Seguridad Militar que la circunda, se ha enfocado con vistas a la consecución de los siguientes objetivos: 1. Delimitar previamente las áreas de distribución de Dociostaurus maroccanus, con el fin de tener conocimiento exacto de la ubicación de los focos de langosta existentes. 2. Efectuar el seguimiento periódico de los focos de langosta detectados a lo largo del ciclo biológico del insecto, con el fin de delimitar las zonas de avivamiento ninfa1 (eclosión de los huevos), sus posibles desplazamientos a nuevas áreas y la superficie final ocupada por el ortóptero en la zona de estudio. 3. Establecer la duración de las diferentes etapas del ciclo biológico de Dociostaurus maroccanus en la zona de estudio. 4. Calcular la densidad de individuos por unidad de superficie en los focos de langosta existentes, en diferentes momentos de su ciclo biológico (ninfa e imago). 5. Determinar, por medio del estudio biométrico de los individuos capturados, la fase en que se encuentra la población de langosta (solitaria gregaria). 6. Delimitar. al final del ciclo biológico del ortóptero, las zonas de mayor concentración de hembras (posibles "rodales" de puesta), con el fin de realizar intensos controles sobre las poblaciones de ninfas emergentes en la generación posterior. 7. Realizar un cálculo aproximado. en los "rodales" de puesta, de la densidad de ninfas emergentes el próximo año (2002), en función del número de canutos depositados por metro cuadrado y el número medio de huevos por canuto. La consecución de dichos objetivos, junto a la experiencia acumulada durante años anteriores, nos permitirá aplicar un Método de Control Integrado para la correcta gestión del área de estudio. en función de: (1) la etapa del ciclo biológico en que se encuentre el insecto, (2) su distribución superficial, (3) su densidad poblacional, (4) la capacidad de desplazamiento a nuevas áreas y (5) la existencia de cultivos próximos. Finalmente. la interacción de los datos obtenidos en las diversas fases del estudio nos va a permitir, para un año concreto y en función de las variables antes mencionadas, proceder al correcto control de las poblaciones de langosta, a través de la elección de una de las siguientes opciones: Opción O Ámbito de aplicación: En focos de densidad inferior a 15 ninfas e11 estado NIV-NV o 12 iinagos, por metro cuadrado. Actuaciórz: No realizar tratamiento químico alguno contra la población de langosta en general, o alguno de sus focos en particular. Opción 1 Ámhito de al9licación: En focos con densidad de 15 a 40 ninfas en estado NIV-NV o 12-32 imagos, por metro cuadrado. Actuación: Empleo de tratamientos químicos localizados, utilizando medios terrestres, sobre determinadas áreas de proliferación dentro de los focos de langosta con el fin de reducir la densidad de individuos presentes. Opción 2 Ámhito de ul~licacióri:En "rodales" de puesta localizados con densidad de hembras superior a 10 individuos por inetro cuadrado. Actuaciórz: Tratainientos culturales consistentes en el arado superficial de los llamados "rodales" de puesta. Opción 3 Ámhito de aplicacicín: En focos con densidad superior a 40 ninfas en estado NIV-NV o 32 in~agos,por metro cuadrado. Actuación: Empleo de tratamientos quí~nicos aéreos sobre superficies concretas. preferiblemente en el momento preciso en que la población de langosta se encuentra mayoritariamente en las dos últimas etapas ninfales (NIV NV). - Distribución y cartografía A partir de la información obtenida en los inuestreos ubicamos, con la ayuda de mapas del Servicio Geográfico del Ejército (E. 1 :50.000), la situación exacta de la población de Dociostaurus maroccanus. Esta población, constituida por los diferentes focos de langosta detectados. se representó posteriormente sobre mapas de proyección U.T.M. de 1 Km de lado (Planos 1 a 4), realizados a partir de fotografía por satélite (E. 1:75.000). El volcado desde los mapas del Servicio Geográfico del Ejército a la fotografía obtenida por satélite, y su posterior manipulación (introducción de datos), se realizó utilizando el programa informática de diseño asistido AutoCAD 2000. día, cuando los insectos presentan mayor actividad, la superficie de los cuadros se fijaba entre 2-4 m2 aproximadamente, con el fin de que el conteo fuese más exacto. c) Batida con manga entomológica Con este método se obtuvo la proporción de sexos y edades (determinado por el estado ninfal). Kemp (1992), propuso efectuar tantas pasadas de red como son necesarias para capturar 100 ejemplares, lo que finalmente realizamos. Parte del material recolectado se conservó vivo. mediante su colocación en tarros con alimento, para su posterior estudio biométrico en laboratorio. Estimación de la densidad Métodos de muestreo utilizados Las diferentes prospecciones fueron realizadas entre el veinte de abril de 2001 y el tres de agosto de este mismo año. momento en que constatamos la ausencia total de ejemplares adultos de langosta marroquí en toda la superficie del Campo de Maniobras. Para llevar a cabo el estudio de la población de langosta marroquí se han establecido una serie de métodos de muestreo para los estados de ninfa e imago que, a su vez. nos han permitido conocer el desarrollo de los focos y su evolución en el tiempo. a) Transecto Este método consiste en recorrer una superficie rectangular, donde la anchura es la distancia que se bate con la red entomológica al desplazarla hacia un lado y otro del cuerpo, a una distancia lo suficientemente cercana al suelo como para poder levantar y capturar las langostas. b) Cuadros marcados en el suelo Este método está basado en el utilizado por Waloff en Chipre, aunque este autor utilizó cuadros móviles de alambre de un pie cuadrado (0,093 m') y que situaba 24 horas antes del recuento, para evitar posibles distorsiones. Onsager y Henry ( 1977), desarrollaron un método para la estima poblacional de ortópteros semejante al anterior utilizando anillo de aluminio de O, 1 m2,y estudiaron estadísticamente su eficacia al ser utilizado por diferentes operarios. Basándonos en lo expuesto, en nuestro estudio, se marcaron una serie de cuadros por medio de piedras pintadas de blanco y situadas en los vértices. Estos cuadros presentaban una superficie variable, de 10 a 24 m2en la mayoría de los casos. realizando entre 35 y 50 medidas cada día. Sin embargo, si las prospecciones se realizaban durante las horas centrales del Para estimar la densidad de ninfas se enlpleó el método de conteo directo por unidad de superficie conocida. en nuestro caso de 1 metro cuadrado (Duranton et al., 1987; Duranton & Lecoq. 1990). Se realizaron. sobre cuadros marcados en el suelo de superficie variable. al menos 50 conteos para determinar la densidad final de cada una de las cuadrículas de cada Plano (1 a 4). La estima de la densidad del número de ninfas o imagos por hectárea se realizó mediante la fórmula: donde n = número de ninfas o imagos contados y S = superficie prospectada en m2. Etapas nirifales Para la estimación exacta de la duración cada una de las etapas ninfales (proceso de por sí frecuentemente complicado ya que las mudas se pueden producir a lo largo de un periodo dilatado de varios días. dependiendo de lo sincronizada que se encuentre la población), hemos adoptado la metodología propuesta por Dennis & Keinp (1988). donde se establece la duración de cada etapa como el tiempo que transcurre desde que el 50% de la población está en determinado estadio hasta que ese porcentaje alcanza el siguiente. Biometría En el material capturado se realizaron una serie de medidas con el fin de caracterizar en cada inoinento la fase de agregación en que se encontraba la especie. Para todo ello se procedió al adecuado manejo de los individuos mediante su montaje, su etiquetado individualizado. su medición y la posterior conserva- ción de alguno de los ejemplares por los métodos habituales (cajas entomológicas). En el presente estudio se han medido una serie de paráinetros, con objeto de comprobar su variación. Los parámetros estudiados son: longitud de la cabeza (LC), longitud del pronoto (LP), longitud de la teginina izquierda (LTI). anchura del esternón (ST) y longitud del fémur posterior izquierdo (LF). A partir de los valores de la tegmina y féinur se ha obtenido. para los cálculos correspondientes, el valor medio del índice élitro-fernoral. Para el estudio de la variación de los datos biométricos en relación con las fases se realizó un análisis estadístico mediante el programa informático SPSS, a partir de las medidas efectuadas sobre 100 individuos de cada uno de los sexos. Estimación de la cantidad de rziizfas de laitgosta emergentes erz el ciclo siguiente Una vez delimitados los rodales de hembras ovipositoras se extrajeron 100 ootecas de cada uno de los focos considerados, excavando varios cuadrados de 0,25 in2 durante las fechas que duró el estudio ( 19 a 3 1 de julio de 2001). A partir de este inuestreo se obtuvieron los siguientes datos: número medio de huevos por "canuto" y de "canutos" por metro cuadrado. RESULTADOS Ciclo biológico Según los datos de campo recogidos en el presente estudio. el avivamiento de las ninfas se produjo entre el cuatro y el siete de mayo, tomando como referencia los dos focos más importantes existentes en el Campo de Maniobras de Chinchilla de MonteAragón: Verlupe-Chinchilla de Monte-Aragón (Foco 1) y Loma de Enmedio (Foco 2). Los adultos se mantuvieron hasta el 3 1 de julio en prácticamente todas las localidades de muestreo. El total de días de presencia de langosta en la zona de estudio en el año 2001, considerando su paso por las diferentes etapas ninfales y su presencia corno imago, ha sido de 88 días (Figura 1). Se ha establecido una duración aproximada de 33 días, para las diferentes etapas ninfales de desarrollo: NI (7 días), NI1 (6 días), NI11 (9 días), NIV (6 días) y NV (5 días). Esta menor duración se asemeja en gran medida a la encontrada por otros autores para poblaciones de Europa oriental y asiáticas (Bodenheimer, F.S.. 1944: Latchininsky, A.V. & Launois-Luong, M.H., 1992). Localización de los focos de langosta marroquí en el Campo de Maniobras y su dinámica poblacional La localización de los diferentes focos de langosta detectados. su evolución a lo largo del tiempo (vías de dispersión) y la densidad de individuos por unidad de superficie en cada uno de ellos, queda reflejada en los Planos 1 a 4. En ellos se representa la situación de los focos de langosta detectados en diferentes fechas, tal coino se indica en la leyenda correspondiente: Estudio biométrico Coino se ha comentado anteriormente, en el correspondiente apartado de material y métodos, se incluyó la medida de los siguientes parámetros: loti- gitud de la cabeza (LC), longitud del pronoto (LP). anchura del esternón (ST). longitud de la tegiiiina izquierda (LTI) y loiigitud del fémur posterior izquierdo (LFI). Los resultados. a partir del exaiiien de 100 individuos de cada uno de los sexos. se muestran en la Tabla l . Por otra parte, se obtuvo el valor del índice élitro-femoral (EíF) en ambos sexos. junto a su desviación típica. cuyos resultados se muestran a continuación: - Índice E/F para machos = 1,50 I0,01 mm. Índice EíF para hembras = 1,56 t 0,01 mm. La comparació~i de estos resultados con los obtenidos con anterioridad por otros autores en diferentes regiones del inundo (Jannone, 1934: Guetzova. 195 1, etc.), nos llevó a la conclusión de que la población de langosta adulta objeto de estudio se encontraba en fase solitaria. Sin embargo. y este dato parece ser de gran importancia, hemos observado un aumento generalizado en las medidas de los cinco paráinetros considerados, con respecto a las obtenidas en el estudio biométrico realizado el año anterior sobre la misma población. Así. el índice élitro-fenioral en machos ha pasado de 1,47 inin a 1,50 miii y en hembras de 1,53 inni a 1,56 rnrn. A pesar de estar incluidos los índices para ambos sexos dentro del rango correspondiente a fase solitaria, no podemos pasar por alto el crecimiento experiiiientado durante el último año. hasta el punto que los acerca peligrosamente a los valores máximos permitidos para seguir considerando su presencia en fase solitaria. Este aumento detectado, con respecto al año anterior, en los valores del índice élitro-Semoral en ambos sexos, puede ser debido a las favorables condiciones climatológicas de los dos últimos años. Sea cuál fuere la razón del auiiiento del índice antes referido. esta situación tios obliga a adoptar para la próxima campaña una serie de precauciones con el fin de detectar posibles tendencias a la gregarización de la población en estudio. Zonas de máxima densidad de puestas En el Plano 4 aparecen las zonas que presentaban las mayores concentraciones de individuos al final del ciclo biológico, entre el 19 y el 31 de julio. Las densidades de ortópteros en esta última fase, con respecto a las existentes en el momento de su máxima expansión (Plano 3). se mantuvieron prácticamente constantes tanto en el Foco 1 (Verlupe-Chinchilla de Monte-Aragón) como en el Foco 2 (Loma de Enmedio). Fue en el citado momento del ciclo cuando se realizó un muestre0 en ambos focos, arrojando los siguientes porcentajes de hembras: 51,87% y 44,16%, para el Foco 1 y 2, respectivamente. Densidad de langosta estimada, para el año 2002, en los dos focos principales Hemos intentado hacer una pequeña estimación del posible número de ninfas de langosta emergentes en los dos Focos principales (1 y 2), en el ciclo biológico correspondiente al año 2002. Siguiendo la metodología descrita en el apartado de Material y Métodos, se obtuvieron los siguientes datos: Foco 1: 2 1,7 huevos/canuto; 2,4 canutoslm2 Foco 2: 19,8 huevos/canuto; 1,6 canutos/m2 Si suponemos que para el próximo ciclo (año 2002) se mantienen las favorables condiciones ambientales de los dos últimos años (fuertes lluvias primaverales seguidas de elevadas temperaturas), y tenemos en cuenta porcentajes de un 20% de pérdida de huevos ocasionado por circunstancias ambientales adversas y parasitismo de dípteros y coleópteros (Arias et al., 1997), más otro 15% de reducción en la población hasta alcanzar la fase NIV (observación personal), se pueden alcanzar, en el peor de los casos, densidades máximas de 33,8 individuos/m2 (en el Foco 1) y de 20,6 individuos/m2 (en el Foco 2). 4 mayo de 2001 10 mayo de 200 1 16 mayo de 2001 25 mayo de 2001 3 1 mayo de 200 1 7 junio de 2001 A partir de 14 de junio de 2001 A partir de 11 julio de 2001 Hasta 3 1 julio de 2001 Figura l. Duración de las distintas etapas del ciclo vital de Dociostaurus maroccanus en el Campo de Maniobras de Chinchilla de Monte-Aragón (Albacete). Tablsi 1. Valores medios y desviaciones típicas (en mmJ de las variables biom&i~%smedidas en 108 individuor de mbos sexos de Dociostarrm mroccanlar, en el Campo de Maniobras de C b x iw m de MM&Amgón (Aib;rem). DINAMICA POBLACIONAL DE LOS FOCOS DE LANGOSTA Plano 1: Plano 3: Corresponde al estado de la población en el inicio del periodo de avivamiento ninfal, ocurrido entre el 4 y el 7 de mayo de 2001. En él se representan las vías de dispersión de las primeras ninfas de langosta desde su emergencia hasta el momento de aparición de los adultos. Corresponde a la situación que presentaban los distintos focos de langosta en el momento de su máxima expansión, fenómeno ocumdo entre el 13 de junio y el 18 de julio de 2001. Plano 4: Plano 2: Representa el periodo de transición de la población entre el último estado ninfal (NV) y su transformación en imagos, entre el 2 y el 7 de junio de 2001. En él aparecen detalladas las vías de dispersión de los individuos adultos. LEYENDA Aparecen detalladas las zonas de máxima concentración de hembras adultas al final del ciclo biológico, lo que puede representar una notable concordancia con la situación de los posibles rodales de puesta. Este hecho ocurrió entre el 19 y el 3 1 de julio de 200 1. Plano 1 Plano 2 Plano 3 Plano 4 AGRADECIMIENTOS Este trabajo ha sido realizado gracias al convenio existente, en materia de investigación, entre la Junta de Comunidades de Castilla-La Mancha y la Universidad Regional. ARIAS, A,, JIMÉNEZ, J. & PÉREZ, A. 1997. Observaciones sobre el desarrollo embrionario y el avivamiento de Dociosruurus rnaroccanus en una finca de La Serena (Extremadura). Bol. San. Veg. Plagas. 23: 1 13-132. BODENHEIMER. F. S.. 1944. Studies on the ecology and control of the Moroccan locust (Docio.stauru,s rnaroccanus T.) in 'Iraq. 1. Results of a mission of the 'Iraq Department of Agriculture to N. 'Iraq in sprig 1943. Bull. Dir. Gen. Agric. Imq.,29: 12 1 pp. DENNIS. B.. KEMP. W. P.. 1988. Further stadistical infeence methos for a stochastic model of insect phenology. Environ. Entonzol., 17 (5): 887-893. DURANTON, J. F., LAUNOIS-LUONG. M. H.. LECOQ. M & RACHARDI. T., 1987. Glride antiacridien du Salzel. Ministere de la Cooperation et C.1.R.A.D.-Prifas. 344 pp. DURANTON, J. F. & LECOQ, M.. 1990. Le críquet pelerin au Suhel. Ministkre des Affaires Etrangkres des Pays-Bas et C.I.R.A.D. / Prifas (France) 183 pp. GUETZOVA. A. B., 195 1. Les ucridierzs rclvageurus en Moldavie. Exposé de thkse de 1- degrk, ~ e n i n ~ r a d s k~i o~s u d a r s t v e n n ~Pedagogitl chenskil Institut: Leningrad. 13 pp. (Doc. Multigr). JANNONE. C., 1934. Aspecti bio-inorfologici e soinatometrici del problema delle fasi nel Dociostal4rus maroccuizus Thunb. In Italia e fuori, con particolare riguardo alla provincia di Napoli. (Secondo contributo). Boll. R. úrb. Ent. Agc Portici, 1: 261.328. KEMP, W. P., 1992. Temporal variation in rangeland grasshopper (Orthopteru: Acridue) coininunities in the steppe region of Montana. USA. Can. Ent., 124: 437-450. LATCHININSKY. A. V. & LAUNOIS-LUONG. 1992. Le criqet ~narocuin,Dociostaurus ~narocannus (Thunberg, 1815), dans la partie orientale de son aire de distribution. tud de nzonogrlrphique relative irl'ex URSS et uuxpays proches. CYRAD.GERDAT-PRYFAS, Montpellier, IVIZIR, Saint-Petersbo~irg,270 pp. ONSAGER. J. A. & HENRY. J. E.. 1977. A inethod for stiinating the densiy of rangeland grasshoppers (Ortlzoptem, Acrididae) en experimental plots. Acrida, 6: 23 1-237. LA CONTAMINACIÓN DE LAS LAGUNAS DE RUIDERA M. ÁLVAREZ COBELAS1 R. CASTELLÓ~ S. CIRUJANO2 C. ROJO3 M. A. RODRIG03 'Centro de Ciencias Medioambientales (CSIC) 'Real Jardín Botánico de Madrid (CSIC) 'Instituto Cavanilles de Biodiversidad y Biología Evolutiva. Universidad de Valencia Las Lagunas de Ruidera comienzan a descubrir sus claros. azules. sosegados, limpios espe-joos JosÉ MART~NEZ RUIZ"AZOR~N" Lu ruta de Don Quijote ( 1905) RESUMEN ABSTRACT Realizamos aquí la primera evaluación global de la contaminación en las Lagunas de Ruidera, basándonos en trabajos de campo realizados durante los años 1997, 1998, 2000 y 2001 y en análisis preexistentes de la Confederación Hidrográfica del Guadiana. Las causas de la contaminación son los vertidos de aguas residuales de Ossa de Montiel y Ruidera. las filtraciones de las fosas sépticas de las viviendas, hoteles, restaurantes y campings en el entorno de las lagunas y los nitratos presentes en el acuífero, procedentes de la fertilización agrícola mediante abonos nitrogenados en la zona de las lagunas altas. Las consecuencias han sido las siguientes: disminución de la transparencia del agua. anoxia en las capas profundas en algunas lagunas durante el verano, aumentos de la concentración de nitrógeno y fósforo en el agua y acuinulación de aquéllos en los sedimentos de todas las lagunas y expansión de especies vegetales indicadoras de eutrofia. Estos procesos son recientes. pues se remontan a finales de la década de 1970. Las lagunas, no obstante, están en su mayoría en buen estado todavía y deben tomarse algunas medidas urgentes para impedir que continúe su previsible evolución hacia una mayor eutrofia, tales como impedir que continúen llegando vertidos de aguas residuales a ellas, limpiar con frecuencia las fosas sépticas y dragar las lagunas en peor estado. Debe intentar favorecerse también la expansión de la vegetación sumergida de carófitos y los procesos naturales de desnitrificación. La principal conclusión de este trabajo es la interconexión entre las aguas superficiales y las aguas subterráneas en este espacio natural protegido, tanto cuantitiva como cualitativamente. Una gestión ambiental razonable de las lagunas debe contemplar todos estos aspectos de modo global. An assessment of water pollution in Ruidera lakes. a Natural Park in Castilla-La Mancha county (Central Spain), is carried out. The study is based upon field work in 1997, 1998, 2000 and 2001 and data by the Guadiana Water Authority. Water pollution resulted from wastewater disposal in the catchment, septic tank filtration and the discharge from the underlying nitrate-rich aquifer, which was polluted by heavy nitrogen fertilization in the late seventies of the XX century. A decline in lake water transparency, anoxic waters in the deeper layers of some lakes. increases in nitrogen and phosphorus contents in waters and sediments and the spreading of plant species indicating water eutrophy in recent years, al1 have been the main outcomes of increasing water pollution. However, most lakes are still in good condition, and so some measures must be taken in the near future to arrest the ongoing eutrophication process, such as wastewater reclamation and efficient treatment. frequent cleaning of septic tanks and sediment dredging of less healthy lakes. Furthermore, charophyte beds must be preserved and natural denitrification processes must be enhanced. As an overall conclusion, we want to stress the fact that surface and groundwaters are tightly. quantitatively and qualitatively, connected in this catchment. A reasonable-sound, environmental management must take these facts into account on a global basis. Palabras clave: aguas residuales, nitratos, aguas subterráneas, organismos indicadores. Keywords: wastewaters, nitrate, groundwater, indicator species. Las Lagunas de Ruidera son un espacio natural muy característico de Albacete y su importancia puede considerarse muy grande, no sólo para España. sino en el contexto europeo. Que sepamos, en Europa sólo hay un lugar comparable, que es el Parque Nacional de Plitvice. en Croacia (www.np-plitvice.com). Gran parte de los valores naturales y culturales de las lagunas (Ecohábitat, 1997: García del Cura et al.. 1997a: Jiménez Ram'rez, 1994,2000: Planchuelo, 1954) son bien conocidos y no los repetiremos aquí. Lamentablemente. apenas existen datos sobre su ecología acuática. a pesar del obvio interés de la misma en un país semiárido donde un paisaje como Ruidera resulta tan excepcional. En particular. sólo conocemos algunos trabajos relativos a la ictiofauna (Almodóvar y Elvira, 1994. 1997: Elvira y García Utrilla. 1991; Elvira et al., 1996; Nicola et al. 1996). La concesión de un proyecto de la Junta de Comunidades de Castilla-La Mancha a S. Cirujano permitió un primer avance en el conocimiento de la flora acuática (Cirujano et al., 2002). Un proyecto de investigación financiado con fondos Feder promovió el inicio de las investigaciones limnológicas en las lagunas, en el que participarnos los arriba firmantes. Dado el candente interés del estado ambiental de las lagunas para la opinión pública, que se plasma en la frecuente aparición de noticias sobre las mismas en los medios de comunicación regionales y nacionales durante los veranos, y su catalogación por la Unión Europea como lugar no apto para el baño en 200 1. nos ha parecido oportuno comenzar la referencia de nuestros estudios con alguna información preliminar sobre la contaminación de las lagunas. fruto de la cual es este pequeño trabajo. LUGAR DE ESTUDIO Bien conocido de los posibles lectores, no lo describiremos aquí más que muy sucintamente, remitiendo a los trabajos de la editorial Ecohábitat (1997) y a la monografía de Montero (2000). Las Lagunas de Ruidera son un paisaje sobre materiales de la Era Secundaria donde la cantidad y la calidad de las aguas se debe a procesos superficiales y subterráneos que interactúan. La descripción más reciente de su origen geológico se encuentra en García del Cura et al. (1997b). En total. existen en la actualidad quince lagunas y un embalse dentro del Parque Natural (Figura 1). La extensión total de las lagunas. incluyendo el e~nbalse de Peñarroya, es pequeña. no superando los 7 km' (menos de la mitad sin el embalse), lo cual supone una cifra inferior a una quinta parte de la extensión total del Parque. Su cuenca superficial puede estimarse en unos 880 km2,inientras que la subterránea asciende a 2.572 km2, pero ambas cuencas no son coincidentes y hay porciones de la subterránea que descargan hacia las cuencas del Júcar. del Segura y del Guadalquivir. La profundidad máxima de las lagunas es variable, pues oscila entre el metro y medio de la Cenagosa y los 21 metros de la San Pedro (30 metros en Peñarroya). La capacidad total de las lagunas asciende a unos 23 Hm3. mientras que la del embalse es de 48.7 Hm3. Los flujos superficial y subterráneo de agua tienen lugar en dirección SE-NW, hacia la Llanura Manchega. Los datos químicos de este trabajo proceden de varios muestreos trimestrales que realizamos en todas las lagunas durante los años 2000 y 2001. En general, en lo que seguirá se hablará siempre de promedios. Los muestreos en las lagunas se hicieron siempre en el centro de cada una. en el área más profunda. deducida del estudio de GOA (1989). La transparencia del agua se midió con un disco de Secchi de 25 cms de diámetro. También se midió en varias ocasiones la extinción vertical de la luz mediante un fotómetro Li-Cor 188B dotado de un sensor escalar Li- 193SA. Paralelamente se registró, siempre de metro en metro, la concentración de oxígeno disuelto en el perfil vertical de cada laguna, usando un aparado YSI-59. La torna de muestras de agua de distintas profundidades se llevó a cabo mediante una botella oceanográfica. Las muestras se llevaron refrigeradas a Madrid y su análisis comenzó inmediatamente, siguiendo los protocolos de la APHA (1992). Las sustancias medidas fueron: carbono. nitrógeno y fósforo en sus fracciones particulada y disuelta, metales pesados. pesticidas y otros compuestos orgánicos. Durante el verano de 2000. tomamos una muestra del sedimento de cada laguna. usando una draga van Veen. Sobre estas muestras sedin~entarias,se midieron metales pesados, carbono, nitrógeno y fósforo, siguiendo los protocolos ya citados. También realizamos varias tornas de muestras a lo largo del año en los principales aportes de aguas residuales a las lagunas. particularmente en la salida de la depuradora de Ruidera (a un kilómetro de la laguna Cueva Morenilla) y en el río Alarconcillo a dos kilómetros de la San Pedro. Simultáneamente con la toma de muestras para los análisis químicos, medimos el caudal con un caudalímetro a fin de esti- mar las cargas de contaminantes. que no son más que el producto de concentración y caudal durante un periodo de tiempo determinado (Meybeck, 1979). En este trabajo daremos sólo los promedios anuales de las cargas para el periodo de estudio. a partir de muestras estacionales, lo cual es una medida más razonable, puesto que incluye épocas del año sin turismo (caso de Ruidera) o en las que la descarga de origen subterráneo en el entorno del Alarconcillo diluye mucho la contaminación proveniente de Ossa de Montiel. Sabiendo que las viviendas de los alrededores de las lagunas disponen sólo de fosas sépticas para el tratamiento de sus aguas residuales y que este sistema de tratamiento siempre presenta fugas (Tchobanoglous. 1995). hemos realizada estimaciones de los aportes de nitrógeno y fósforo que. desde dichas viviendas, se hacen a las lagunas donde el número de viviendas es más importante (San Pedro, Colgada. Cueva Morenilla). Ese número de viviendas del entorno de cada laguna se ha extraído del informe de la Consejería de Política Territorial de la Junta de Comunidades (1991). Hemos asumido que en cada vivienda residen cuatro habitantes equivalentes durante un mes y medio al año. Un habitante-equivalente produce una DBO, de 6 0 g O2 por día (Tchobanoglous. 1995), lo cual supone 10,90 y 2.18 gldía de nitrógeno y de fósforo, respectivamente. A las cifras de aguas residuales por vivienda les hemos añadido las del uso de campings (San Pedro y Los Molinos). conocida por los datos de visitantes al Parque Natural. Una fosa séptica con la menor filtración posible exporta el 45% del nitrógeno y el 60% del fósforo que recibe. siguiendo a Ryding & Rast (1991). Estas suposiciones. muy conservadoras. nos han permitido calcular unas cifras preliminares de aportes de nitrógeno y fósforo a las lagunas. que seguramente serán subestimaciones. Los datos de la Confederación Hidrográfica del Guadiana sobre concentraciones de nitratos en 2000 en el acuífero nos permitieron confeccionar un mapa de isolíneas. mediante el programa SURFER (Golden-Software. 1995). Los datos de nitratos en la estación de La Cubeta, que prospecta mensualmente la CHG desde 1973. situada entre la laguna del Rey y la Cueva Morenilla. se han usado para averiguar el aumento de su concentración durante los casi 30 años transcurridos desde el comienzo de la toma de datos por este organismo oficial. La metodología de Vighi & Chiaudani (1985) nos sirvió para extrapolar hacia el pasado las concentraciones de fósforo total. que es el indicador clásico de eutrofización (Vollenweider, 1968), y averiguar cuánto se han incrementado en años recientes. Brevemente. estos italianos asumen que la concentración de fósforo en los lagos que no han sufrido ningún proceso de eutrofización antropógeno depende del índice inorfométrico. es decir. del cociente entre la conductividad (o la alcalinidad) y la profundidad media de cada lago. Y demuestran que esa relación se cumple para un amplio conjunto de lagos en Europa y América. Así. su relación de regresión puede utilizarse para estimar el promedio de las concentraciones pasadas de fósforo total en función de esas dos variables de fácil conocimiento. Durante 1997 y 1998. mediante buceo y transectos longitudinales en los distintos lagos extrayendo la vegetación de fondo con plomadas revestidas de ganchos, hemos obtenido una idea razonablemente precisa de la vegetación sumergida. realizando una estimación preliminar de su cobertura. pues conocíamos también hasta qué profundidad llegaba la luz, ya que la habíamos medido con el fotómetro. Este proceder nos ha permitido esbozar una primera aproximación de cuánta superficie ocupan los carófitos en los fondos de Ruidera. Por último, las observaciones de la vegetación litoral prolongadas durante varios años nos han indicado "grosso modo" cómo se propagaban algunas plantas indicadoras por las distintas lagunas. Las aguas residuales de Ruidera son las que más sustancias están incorporando a las lagunas Cueva Morenilla. Coladilla y Cenagosa (Fotografías 1-2; Figura 2). si bien las de Ossa también suponen una contribución importante a la laguna San Pedro y. cuando ésta se desborda. hacia las situadas por debajo de ella. El problema se acentúa en las lagunas bajas porque albergan mucha menos agua que la San Pedro y, así. la dilución es inferior. No obstante. la relación entre el nitrógeno y fósforo indica que la procedencia del vertido de Ossa es eminentemente agrícola, mientras que el de Ruidera parece típicamente urbano. Las aportaciones de las viviendas situadas en el entorno de las lagunas son. a grandes rasgos. diez veces menores que las de Ossa en el caso del nitró- geno (Figura 3) y unas cien veces menores que las de Ruidera. Como es lógico, son proporcionales al número de habitantes de chalets. hoteles. adosados. etc. La contribución de este impacto humano es más importante en las lagunas más urbanizadas. siguiendo de nuevo la Cueva Morenilla la peor suerte por su escaso volumen. El tercer motivo de contaminación de las lagunas son los fertilizantes nitrogenados que se han usado y se usan en su cuenca. parte de los cuales se infiltran hacia el acuífero. El flujo subterráneo se dirige hacia las lagunas. como atestiguan los mapas de isopiezas existentes (Montero, 1994) y el nitrato que poseen procede de las aguas subterráneas en gran medida. Las Lagunas de Ruidera están situadas dentro de un campo de isolíneas de nitrato que oscila entre 30 y 40 mg/L, a excepción de la laguna Blanca, cuyo entorno subterráneo presenta concentraciones más elevadas (Figura 4). Los efectos de estos contaminantes son múltiples. Aquí sólo destacaremos algunos de ellos. La transparencia del agua, por ejemplo, se resiente en los ambientes más contaminados. Es el caso de las lagunas San Pedro. Cueva Morenilla y Coladilla (Figura 5). El resto de las lagunas presenta una elevada penetración de la luz que permite el desarrollo de extensas praderas de carófitos en muchas zonas del fondo de las lagunas (Figura 6). Ese desarrollo es proporcional a la extensión de los niveles a los que llega la luz y al tamaño de las lagunas y. así, aunque en las lagunas altas penetre más la luz, es la Colgada la que presenta más cobertura de carófitos en razón de su extensión. Otra respuesta a la contaminación es la distribución del oxígeno disuelto en el agua. cuyo momento más crítico es la época estival. La Figura 7 muestra los perfiles verticales en tres de las lagunas estudiadas. En ella. podemos apreciar que San Pedro y Colgada presentan zonas anóxicas cerca del fondo. de más espesor en la primera. Esta anoxia también se aprecia en las lagunas Salvadora, Batana y del Rey, aunque con distintos espesores. El resto de las lagunas carecen de esos ambientes sin oxígeno. pero sí pueden llegar a tener sedimento anóxico (en Cueva Morenilla, Coladilla. Peñarroya). La entrada de nitratos por vía subterránea (y. menos frecuentemente. superficial) es la responsable principal de la distribución del nitrógeno total en las lagunas. Existe un gradiente apreciable de este conlpuesto desde las lagunas altas a las bajas, que es mayor en las lagunas Blanca, Conceja, Tomilla y Tinaja (Figura 8). Ese gradiente decreciente es sostenido hasta la Batana y la Colgada. donde vuelve a subir el nitrógeno, por efecto de los manantiales subacuáticos del Pinar (caso de la Batana) y de las Hazadillas y los flujos de las fosas sépticas del entorno (caso de la Colgada): véase que las isolíneas de nitrato son superiores en los alrededores de estas lagunas (Figura 4). Sin embargo, la concentración en los sedimentos es relativamente baja en todas las lagunas hasta la Batana. ascendiendo especialmente en Cueva Morenilla y Coladilla (Figura 9). El nitrógeno, pues. parece acumularse especialmente en la Batana. quizá porque rara vez descargue superficialmente hacia la Colgada (y apenas hay contacto subacuático con la Colgada; Montero. 2000). La laguna Batana se comporta como un sumidero de nitrógeno. Pero son la Cueva Morenilla y. en especial, la Coladilla las lagunas con mayores concentraciones nitrogenadas en agua y sedimento. El gradiente de nitrógeno en el agua y la similitud de sus concentraciones en el sedimento de las lagunas hasta la Santos Morcillo, junto con las elevadas concentraciones en el acuífero subyacente, apuntan a que debe haber algún mecanismo de pérdida de nitrógeno a lo largo de ese rosario de lagunas. El ii~ás probable es la desnitrificación bacteriana en el sedimento que da lugar a nitrógeno molecular, el cual -al ser un gas- se escapa hacia la atmósfera. La laguna del Rey también debe presentar procesos de esa índole, a juzgar por la baja concentración de nitrógeno en el sedimento. pero ese elemento vuelve a aumentar en las lagunas bajas, no sólo por los vertidos de aguas residuales (Figura 2) y por las elevadas concentraciones de nitrato en el agua subterránea (Figura 4). sino por la enorme producción de materia orgánica vegetal (masiega. carrizo) en sus orillas que, cuando se descompone. añade muchas compuestos nitrogenados al agua y a los sedimentos. La distribución del fósforo total en las lagunas es compleja. Contra lo que podría esperarse, no es demasiado alta en la San Pedro (Figura 10). la cual sí almacena más fósforo en el sedimento que las lagunas altas (Figura 11). Es probable, entonces. que la citada anoxia de la San Pedro (Figura 7) aumente el fósforo en el agua al final del verano, incrementando la eutrofia de esta laguna, pero -al ser nuestros inuestreos trimestrales- no hemos podido detectar ese hecho. La laguna Colgada y el embalse de Peñarroya son los ambientes con más fósforo superficial, aunque no por ello los que presenten más fitoplancton (datos inéditos). Esto pudiera deberse a la captación de gran parte de ese fósforo por las praderas sumergidas de carófitos (Figura 6). Los mayores almacenes de fósforo sedimentario se hallan en la San Pedro, la Cueva Morenilla y la Coladilla. Carecemos de datos antiguos de nitrógeno o fósforo en las lagunas que nos permitan conocer con absoluta certeza el grado de contaminación que hayan podido experimentar. Indirectamente, es posible deducir algo a partir de los datos de nitrato en la estación de La Cubeta, situada aguas abajo de la laguna del Rey. Coino muestra la figura 12. durante los años de la década de 1970 dicha sustancia fue creciendo paulatinamente hasta alcanzar un máxiino a comienzos de la década siguiente. Luego. se estabilizó para volver a incrementarse hacia 1996. Tomando valores promedio para años hidrológicos, podemos deducir que las concentraciones de nitrato en La Cubeta se han multiplicado en 1116s de un 250% desde el año hidrológico 1973- 1974 hasta ahora. En cuanto al fósforo, los cálculos realizados mediante la ecuación de Vighi & Chiaudani (1985) nos indican que todas las lagunas han ganado fósforo, algunas en una proporción del 1.100% (Colgada y Peñarroya; Figura 13). El resto de los análisis efectuados indica que sólo los metales pesados en el sedimento de la laguna Cueva Morenilla alcanzan unos niveles elevados. particularmente eil cuanto se refiere a aluminio (28 140 mgíkg). zinc (43,5 mglkg), plomo (30 mgíkg) y níquel (14 mgíkg). Hay una pequeña presencia de pesticidas organoclorados en las lagunas altas. pero es residual. Por último, sólo nos resta mencionar algunas breves informaciones sobre la respuesta de los organisiiios a estos aumentos generalizados de la contaiiiiiiación en las lagunas. Poco podemos decir. ya que son escasísiinas las observaciones disponibles sobre la tlora y la fauna acuáticas. Unicainente señalaremos que otro indicador del auiiiento de la eutrofia de las lagunas es el iiicreinento de la densidad en muchas lagunas de Scirpzrs litoi-crlis Schrader (juncia de agua). descubierta a finales de la década de 1980 (Cirujano et al., 1990) y que no ha hecho más que extenderse desde entonces. y T\phcr do~lzirlgelzsis(Pers.) Steudel (enea), dos plantas de aguas ricas en nutrientes. También heinos apreciado en los últimos años el creciiiiiento cada vez más conspicuo de algas filanientosas recubriendo a los carófitos. otro síntoma de eutrofia (Phillips et al.. 1978). La contaminación de las Lagunas de Ruidera se debe principalmente a los efluentes de aguas residuales de Ossa de Montiel, Ruidera (Figura 2). a las filtraciones del agua residual de las fosas sépticas de viviendas, hoteles, restaurantes. campings y chalets situados en el entorno de las lagunas (Figura 3) y a los aportes de nitrato desde el acuífero (Figura 4). procedentes del abonado intensivo con sustancias nitrogenadas en la zona de las lagunas altas. Como consecuencia de estos hechos, otros tipos de contaminación pueden considerarse como despreciables: en particular, la de pesticidas y metales pesados. lo cual corrobora los resultados previos de la Confederación Hidrográfica del Guadiana (1994) y de la tesis doctoral de Rodríguez Martín-Doiineadios (2000). Estas causas han engendrado. entre otros, los efectos siguientes: disminución de la transparencia del agua en algunas lagunas. aumento del nitrógeno y fósforo de las lagunas y aumento de la densidad y expansión de algunas especies indicadoras de eutrofia, como la juncia de agua y la enea. Además, hay un aumento de las algas filamentosas que recubren las praderas subacuáticas de carófitos. El proceso de aumento de nitrógeno y fósforo es iinportante (Figuras 12-13). pero -coino se partía de una situación de aguas muy liinpias- aún no es muy grave. Sí hace preciso ir tomando medidas paliativas y preventivas: la) limpiar frecuentemente las fosas sépticas. 2") depurar eficazmente los etluentes de aguas residuales de Ruidera y Ossa de Montiel. 3") dragar las lagunas San Pedro. Cueva Morenilla y Coladilla. Sin embargo. las lagunas taiiibién tienen algunos medios de limitar la extensión de la eutrofización. En primer lugar. desnitrifican (Figuras 8-9). es decir. convierten el nitrato en nitrógeno molecular que pasa a la atmósfera (Seitzinger, 1988). En segundo lugar. los carófitos del fondo se aprovechan de que aún la penetración de la luz es alta en la mayor parte de las lagunas (Figuras 5-6). El desarrollo de los carófitos consume e ininoviliza nutrientes (nitrógeno. fósforo) en el sedimento (Jeppesen. 1998) y genera unas sustancias antibióticas que inhiben en buen medida el desarrollo del fitoplancton en la columna de agua (Wiuin-Andersen et al.. 1982). con lo cual la transparencia de la inisina aumenta y los síntoinas de eutrofia -debidos al aumento del fósforo- son menos patentes. Nuestro estudio constata sin lugar a dudas la estrecha relación entre la calidad de las aguas superficiales y la de las aguas subterráneas en las lagunas de Ruidera. A SLI vez. se coinpleinenta con un inforine del CEDEX (1997) donde se asocia la cantidad del agua en anibos ambientes. Resulta. por tanto. ii-isoslayable el analizar conjuntamente la cantidad y la calidad del agua en las Lagunas de Ruidera en relación con las del acuífero si se quiere conocer con cierta exactitud lo que sucede en ellas. No obstante. esto no es fácil y precisa de una estrecha colaboración entre hidrogeólogos y ecólogos acuáticos. Una gestión ambiental razonable de las lag~inas debe contemplar todos estos aspectos de modo global. AGRADECIMIENTOS Este trabajo se ha podido hacer merced a la financiación del proyecto 1-FD97- 18 12. Además. se ha beneficiado extraordinariamente de las aportaciones de Federico Grande Pinilla y Marisa Colmenero Pérez (directores del Parque Natural). Ángel Nieva y José Ramón Aragón (Confederación Hidrográfica del Guadiana). Esperanza Montero (Dept. Geodinámica, Univ. Coinplutense de Madrid) y la Delegación de Sanidad de Albacete. además de Valentín Molina Rodríguez. Eusebio Reinosa Bascuñana. Kika Muñoz Oliver y Manuel López Sánchez (personal que trabaja en el Parque Natural). Palmira Riolobos, Salvador Sánchez Carrillo. José María Alonso y Jerónimo Sánchez fueron fundamentales en muestreos y análisis. Los inuestreos de la laguna Colgada se hicieron tomando como base de partida las instalaciones del Club Náutico Privado Las Lagunas de Ruidera, cosa que agradecemos desde aquí. Alrnodóvar, A. & Elvira. B. 1994. Further data oi-i the fish fauna ca~alogiie of the Natural Park of Ruidera Lakes (Guadiana River basin. central Spain). Verhandlungen der Internationcrle Vereinigrrng,fiir Liriznologie. 25: 2 173-2177. Almodóvar, A. y Elvira, B. 1997. Ictiofauna. En: P a q ~ r eNat~ircrl Lag~tnci.~ (le Ruiclercr, 133- 164. Ecohábitat. Talavera de la Reina. 395 pp. APHA. 1992. Métodos ~zornializcidos~)eireiel cinNlisi~ de cigrras yotcrbles y 1-esicl~icrles.Editorial Díaz de Santos. Madrid. CEDEX, 1997. Estudio de Icr Izidrologíci i.sotópicu en la cuerlca del Alto Cuadicinci. 1. Unirkrd hielrogeológic ~ 04-06 i L ~ ~ g ~ i de n a Rrtidem, s Ccrnlpo de Montiel. Informe técnico no 5 1-493-1-0 12. Centro de Estudios de Técnicas Aplicadas. Madrid. 155 pp. Cirujai-io, S., Velayos, M. & Carrasco. M. A. Notas sobre higrófitos peninsulares. 111. Anales del Jardín Botánico de Madrid. 47: 520. Cirujano, S., Medina, L., Álvarez Cobelas. M. y Moreno. M. 2002. Flora y vegetación acuática de las Lagunas de Ruidera. Actas II Joniadas del Meciio Natural Albacetense (en prensa). Confederación Hidrográfica del Guadiana, 1994. Microcontarninar~tes orgúnicos en las unidcides hidrogeológicas 04.04 Mancha Occidental y 04.06 Carizpo de Montiel. Centro de Publicaciones del MOPTMA. Madrid. Consejería de Política Territorial. 199 1 . Infornie sobre la situaciól? urbanística del Parque Natural de lcrs Lag~irlusde R~iidera.Junta de Com~inidadesde Castilla-La Mancha. Toledo. Elvira, B. y García Utrilla, C. 1991. La ictiofauna de las Lagunas de Ruidera: revisión bibliográfica y proyecto de catalogación actual. Actas de las 1 Jorrzadas sobre el Medio Naturcil Albacetense: 395-399. Elvira, B., Nicola, G. G. & Alinodóvar. A. 1996. Pike and red swamp crayfish: a new case on predatorprey relationship between aliens in central Spain. Journal of Fish Biology, 48: 437-446. Ecohábitat, 1997. Parque Natural Lag~rnuscle Ruiclerrr. Talavera de la Reina. 395 pp. García del Cura. M. A.. García Ráyego, J. L. y González Cárdenas. E. (eds.), 1997a. Elen~entos del medio rzcitural en la provincia cle Ciudad Real. Ediciones de la Universidad de Castilla-La Mancha. Cuenca. 266 pp. García del Cura. M. A., González, J.A. y Ordóñez, S. 1997b. Geología y geon-iorfología. En: Parque Natural h g u n a s de Ruiclera, 19-50. Ecohábitat. Talavera de la Reina. 395 pp. GOA, 1989. Estrldio de la bntitnetríu de las Lagui1ci.s (le Ruidera. MOPU, CHG, DGOH. Madrid. Golden Software, 1995. SURFER for Windows, Version 6 . Golden Software Inc., Golden (Colorado). Jeppesen, E. 1998. Tlze ecology of .shc~llow lcikes Trophic iriteractioils iii the pelcrgial. Tesis Doctoral. NERI Technical Report 247. Silkeborg (Dinamarca). 358 pp. Jiménez Raiiiírez. S. 1994. Lagirncrs de R l l i d e ~ ~el i : río q1re ptrsa por /ni p~iehlo. Perea Ediciones. Toinelloso. 191 pp. Jiménez Ramírez. S. 2000. Reul Sitio dp Ririderci. Panorcrrnci sociul y de la pr-(jpiedcrd desde Icr Prel~istoria.Edicio~iesSoubriet. Toirielloso. 255 PP. Meybeck. M. 1979. Concentrations des eaux fluviales en éléments inajeurs et apports en solution aux océans. Rev~ie de Céologie Dyncin~iq~ieet d~ Géographie Pli),.siq~ie.2 1 : 2 15-246. Montero, E. 1994. Fu~lcioncrnzic~rlto Iiidlvgeológico r l ~ l sistet~i~i de lcis Lcrgunas cie Ruiclere~.Tesis Doctoral. Facultad de Geología. Universidad Con~plutense. Madrid. 275 pp. + anexos. Montero, E. 2000. Coritribucióri e11 estudio (le la geometrí~ry los Iírnites clel ucltTero del Ccrnzpo de Montiel. Institiito de Estudios Albacetenses "Doi-i Juan Manuel". Albacete. 177 pp. Nicola. G. G.. Almodóvar. A. & Elvira. B. 1996. The diet of introduced larger-ilouth bass, Micro/~ter~r.s salinoides, in tl-ie Natural Park of the Ruidera HydroLakes, Central Spain. Polskie At*chi~t>~in1 biologii, 43: 179- 184. Phillips, G. L., Eminson, D. & Moss, B. 1978. A mechanism to account Sor macrophyte decline in progressively eutrophicated freshwaters. Aqlicrtic Botan)), 4: 103- 126. Planchuelo. G. 1954. Estudio del Alto Gucrciiarzri y de Icr altiplanicie del Ce11npo de Molztiel. Instituto de Estudios Manchegos. Madrid. 189 pp. Rodríguez Mastín-Doimeadios, R. 2000. Aportacioi1e.s al conocimierito del estado ~nedioun~bienral de hidrosi,stenzas de interés internacioiiul situeldos el1 Castillli-La Manclzcr. Tesis Doctoral. Ediciones de la Universidad de Castilla-La Mancha. Cuenca. 584 pp. Ryding, S.-O. y Rast. W. 1992. El coiltrol de Icr eutrofizaciórl err lagos y />antanos. Ediciones Pirámide. Madrid. 375 pp. Seitzinger, S. P. 1988. Denitrification in freshwater and coastal marine ecosysteins: ecological and geocheinical significance. Linlnology cind Ocecinograplzj,. 33: 702-724. Tchobanoglous, G. (Ed.). 1995. Mptcuíf-Edd.~:Ingeniería cle IrgLias re.siclucile.r, redes de alccrntcrrillado y bonlbeo. Madrid. 461 pp. Vighi, M. & Chiaudani, G. 1985. A simple method to estimate lake phospl-iorus concentrations resulting fr011-i natural, background, loadings. Wclter Resecircli, 19: 987-99 1. Vollenweider, R. A. 1968. Scient$c firndurnentcils of the eutropliiccrtion c$ lakrs crrzd jloi.ving warers, wirlz .special referente to nitmgen cincl pl7o.s/~l~orir.s as factors in eritroplzication. OECD. Paris. 154 pp. Wium-Andersen, S., Anthoni, U.. Christophersen. C. & Houen, G. 1982. Allelopathic effects on phytoplankton by substances isolated from aquatic macrophytes (Charales). Oikos, 39: 187- 190. Figura 10. Fósforo total promedio (mg P h ) en el agua super& cial de las Lagunas de Ruidera Figura 7. Oxígeno disuelro a disdntas profcmdidades en varias l a p a s de Ruidera a principios de septiembre de 2000. La situación en 2001 era muy simüaf. Figura 11. Fósforo total (mglkg)en el seckwntn de des Lagunas de Ruidera. Figora 81. Nitrbgeno total promedio (mg N&) en el agua superficial de las Lagunas de Ruidera. I 9. Nitrógeno total (mgkg)en los sedimentos de las Lagunas de Ruidera. Figura mal 7w4 9081 1 AÑO HIDI#K~~KX) U Figura 12. Concentración a lo fargo del tiempo de los nitratos en la estación de La Cubeta, situada entre la laguna del Rey y la laguna Cueva Morenilla. Los datos fueron obtenidos por la Confederaci6n Hidrográfica del Guadiana. Figura 13. Aumento del fósforo total en las Lagunas de Ruidera desde una situación sin entradas de fósforo de origen antrópico, que puede situarse a comienzos de la década de 1970. Fofograña l.Espumas en e1 Guadiana, aguas abajo del Salto del Hundimiento. Obsdrveme las acumulaciones de detergente en el centro de la imagen. FotopaEa 2. Sistema de tratamiento de Ruidera, consistente en un sencillo agitador.