Mecanismos de regulación neuroendocrina del corazón
Anuncio

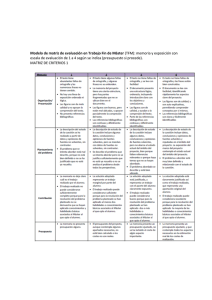
Facultad Biología UB Ginés Calín Máster Fisiología Integrativa MECANISMOS DE REGULACIÓN NEUROENDOCRINA DEL CORAZÓN. EJE DE COMUNICACIÓN CEREBROCARDÍACO. Tabla de contenido INTRODUCCIÓN ...................................................................................................... 2 OBJETIVOS .............................................................................................................. 4 DISCUSIÓN Y RESULTADOS .................................................................................. 5 MECANISMOS DE REGULACIÓN CARDIOVASCULAR POR EL SISTEMA NERVIOSO AUTÓNOMO. ..................................................................................... 5 MECANISMOS DE REGULACIÓN DEL SISTEMA NERVIOSO CARDÍACO INTRÍNSECO ........................................................................................................ 8 MECANISMOS DE REGULACIÓN DEL SISTEMA ADRENÉRGICO INTRÍNSECO CARDÍACO.......................................................................................................... 11 MECANISMOS DE REGULACIÓN ENDOCRINA DE LA FUNCIÓN CARDÍACA 11 HIPÓTESIS DE CONEXIÓN CEREBRO-CARDÍACA .......................................... 17 CONCLUSIONES ................................................................................................... 21 REFERENCIAS ....................................................................................................... 21 1 Facultad Biología UB Ginés Calín Máster Fisiología Integrativa INTRODUCCIÓN El estudio del corazón ha pasado por diferentes modelos y concepciones. Harvey en 1628 en su tratado “De Mortis Cordes” describió el funcionamiento cardíaco como una bomba contráctil. Claude Bernard en 1865 y Darwin en 1872 plantearon como hipótesis la influencia sobre el corazón, tanto de las vías eferentes vegetativas, como de las aferencias desde la periferia al sistema nervioso central (Alfonso & Báez 2008). Ningún paradigma ha superado el paso del tiempo a nivel conceptual, hemos pasado por diferentes modelos como el cardio-renal, luego el hemodinámico y el actual modelo neuroendocrino, para dirigir finalmente toda la atención al desarrollo de la proteómica y de la genómica en el fenotipo cardíaco (Bayés-Genís 2005). Una de las ideas principales de esta revisión es que cualquier intento de terapia selectiva en el eje neuroendocrino cardíaco requiere una mayor integración del rol que cada elemento juega en la coordinación de los valores cardíacos regionales, dada su naturaleza estocástica primaria, es difícil predecir su comportamiento sin sólidas evidencias in situ, ya que el sistema está abierto a múltiples influencias de control (Armour 2008). El centro regulador cardiovascular medular integra información sensorial e inputs desde centros cerebrales superiores, y aferencias de inputs cardiovasculares y de tensión arterial a través de vías simpáticas y parasimpáticas. Este artículo también discute sobre el sistema nervioso cardiaco intrínseco y la conexión cerebro-cardíaca a través de la cual la información aferente puede influenciar la actividad en las áreas subcortical, frontocortical y de la corteza motora (Shaffer et al. 2014). Este “Little brain” cardíaco está compuesto por aferencias sensitivas que interconectan un circuito local y eferencias motoras (adrenérgicas y colinérgicas) que comunican con otras en los ganglios intratorácicos extracardíacos, todo bajo la influencia tónica del comando neuronal central y de hormonas circulantes. Estas vías neurales conectan el corazón al córtex insular generando reflejos que determinan los índices cardíacos (Armour 2008). En cuanto a hormonas circulantes, CRFlike peptides llamadas urocortinas, tienen receptores en cerebro CRF1R y también CRF2R en cerebro y corazón. Tanto CRF 2 Facultad Biología UB Ginés Calín Máster Fisiología Integrativa como urocortinas producen importantes efectos hemodinámicos como respuesta de adaptación al estrés, vía acción en sus receptores, con importantes efectos cardíacos y nerviosos. Pueden suponer un lazo de comunicación endocrina entre cerebro y corazón (Yang et al. 2010). La importancia de la norepinefrina y epinefrina en el estímulo cardíaco está bien establecida. La solicitación cardiovascular durante la clásica reacción de lucha o huída se piensa que es debida a la liberación de catecolaminas desde las terminaciones nerviosas simpáticas y la médulosuprarrenal. Un mecanismo adrenérgico distinto del clásico eje simpático-adrenal está involucrado en la regulación cardiovascular. Se describe la existencia de un sistema adrenérgico intrínseco en el miocardio de mamíferos, con capacidad para sintetizar catecolaminas (Huang et al. 1996). Además se encuentran en el corazón neuronas histaminérgicas, otras sensibles a endotelina, con receptores ETA y ETB, también se reportan neuronas sensibles a neurotensina y otras postganglionares con capacidad de síntesis de óxido nítrico (ON) (Alfonso & Báez 2008). Niveles plasmáticos de norepinefrina, renina, vasopresina, endotelina-1, péptido natriurético atrial (ANP), péptido natriurético cerebral (BNP) y TNFα, están elevados en el fallo cardíaco. La actividad del sistema simpático reflejado por norepinefrina está correlacionada a la mortalidad y parece poseer información pronóstica independiente. Diversos estudios han documentado el beneficio de betabloqueantes en insuficiencia cardíaca crónica. ACE-inhibidores y antagonistas de angiotensina 2 han demostrado reducir la mortalidad y morbilidad en insuficiencia cardíaca (Kjaer & Hesse 2001) Ampliamos entonces la atención en el sistema renina-angiotensina, ya que se han identificado áreas en el Núcleo del tracto solitario que específicamente participan en la acción de la aldosterona y parecen tener un importante rol en la estimulación vagal con repercusiones cardiovasculares y centrales (Murck et al. 2012) Otras hormonas centrales consiguen modular al SNA con influencia cardíaca, como oxitocina y vasopresina, asociadas a la percepción del entorno en contextos de peligro o de interacción social (Sue Carter 1998). La regulación neuroendocrina cardíaca permite mecanismos adaptativos al entorno en los conocidos sistemas de lucha-huída, pero también en los mecanismos de 3 Facultad Biología UB Ginés Calín Máster Fisiología Integrativa inmovilización reptiliana o de involucramiento social. Será revisada la perspectiva polivagal de regulación (Porges 2007b). Otros autores en cambio proponen un modelo de integración neurovisceral central, Central Autonomic Network (CAN), donde se establece una vía de comunicación dinámica integrada en el Núcleo del tracto solitario (NTS) desde aferencias cardíacas neurovegetativas con centros superiores en amígdala y córtex prefrontal, que analizan el entorno para establecer una respuesta adecuada (Thayer & Lane 2009) En ambos modelos la variabilidad cardíaca (HRV) es propuesta como medida fisiológica de control (Shaffer et al. 2014) Las disfunciones fisiopatológicas de las glándulas hipófisis, tiroides, paratiroides, suprarrenales y sus repercusiones en el sistema cardiovascular superan el objetivo de este estudio. Para una mayor comprensión ver (Rhee & Pearce 2011) OBJETIVOS Como Osteópata siento un gran interés por el estudio de las relaciones entre los diferentes sistemas a diferentes niveles de profundidad, y los mecanismos de adaptación-compensación que se establecen entre ellos en la búsqueda de un equilibrio alostático y homeostático. La regulación cardíaca es altamente compleja y la literatura científica nos ofrece una visión parcelada de los diferentes mecanismos reguladores. El objetivo de este estudio, lejos de ser una revisión sistemática de la bibliografía publicada sobre el tema, dará una visión global integrativa de la participación de los sistemas nervioso, endocrino, inmune y cardiovascular. Manteniendo la idea del premio Nobel de química Ilya Prigogine: El todo es más que la suma de las partes, con propiedades emergentes que derivan de su relación con el entorno, no predecibles. El objetivo del estudio es analizar las relaciones entre los diferentes sistemas a diferentes niveles de profundidad, con conciencia epistemiológica. (Prigogine 1997) 4 Facultad Biología UB Ginés Calín Máster Fisiología Integrativa Finalmente se expondrán diversas teorías publicadas que proponen hipótesis sobre la organización de dichos sistemas en el control de la función cardíaca. DISCUSIÓN Y RESULTADOS MECANISMOS DE REGULACIÓN CARDIOVASCULAR POR EL SISTEMA NERVIOSO AUTÓNOMO. Una red de áreas neuronales específicas ubicadas en el tallo cerebral controla al SNA en su regulación cardiovascular, denominada “Centro vasomotor”, localizado en un área bilateral, en la sustancia reticular del bulbo y en el tercio inferior de la protuberancia, con un rol integrador de informaciones: Sensitivas viscerales, procedentes de barorreceptores de cavidades cardíacas y grandes vasos. También de quimiorreceptores. Del centro respiratorio bulbar. De estructuras suprasegmentarias como zonas del córtex cerebral, complejos amigdalinos, el hipotálamo y sustancia reticular del diencéfalo, mesencéfalo y protuberancia en diferentes estados emocionales y conductuales. (Alfonso & Báez 2008) Este centro vasomotor es altamente dependiente de los centros superiores, quienes tienen además vías directas, particularmente desde el hipotálamo, que sin hacer relevo en el centro vasomotor llegan a las neuronas preganglionares simpáticas, por lo que no sólo el centro determina el flujo de salida final del SNA (Coote 2007). Conecta con el núcleo ambiguo del vago con efecto eferente parasimpático, y con neuronas preganglionares ortosimpáticas del asta intermedio-lateral medular. Se han identificado las siguientes áreas funcionales (Fig.1): 5 Facultad Biología UB Ginés Calín Máster Fisiología Integrativa Fig. 1 (Alfonso & Báez 2008) Área sensorial (aferente parasimpática) Área vasodilatadora (área moduladora, recibe de la sensorial e inhibe la vasoconstrictora). Área vasoconstrictora (área rostro-ventro-lateral (ARVL) y área rostro- ventromedial (ARVM), neuronas presimpáticas que proyectan a neuronas preganglionares simpáticas en la medula espinal) El nervio vago tiene un 80/85% de fibras aferentes sensitivas proyectadas al NTS, principal área sensorial del centro vasomotor, a través de sus dos ganglios yugular y plexiforme. El principal neurotransmisor del NTS es glutamato con receptores de tipo NMDA (N-acetil-D-aspartate) y otros receptores sensibles a AMPA (α-amino-3-hidroxi5-methyllisoxazole-4-propionico). La serotonina (5HT) también influye en el NTS, con roles para algunos subtipos de receptor 5HT1, 5HT2, 5HT3, 5HT4, 5HT7, y efectos cardiovasculares asociados a termorregulación, sueño-vigilia y alerta al entorno. Otros receptores purinérgicos (A2) y a vasopresina (V1, A2a) operan en NTS con influencia de adenosina, ATP, y vasopresina con efectos cardiovasculares y respiratorios. Ciertos experimentos desmienten la idea de que el vago no pueda participar de las aferencias involucradas en el control inhibitorio central de nocicepción y dolor, aunque las vías aferentes de la sensibilidad dolorosa del corazón pertenecen a la división simpática del sistema autónomo, integradas en los ganglios cervicales, estrellado y 6 Facultad Biología UB Ginés Calín Máster Fisiología Integrativa medio. Otras fibras aferentes se dirigen directamente desde el plexo cardiaco, hacen sinapsis en el asta dorsal de los primeros cinco segmentos torácicos y ascienden, junto con el haz espinotalámico, hasta el tálamo y la corteza. En el bulbo raquídeo se controla la actividad de las neuronas preganglionares simpáticas en el área vasodilatadora o área caudal ventro-lateral (ACVL) y localizada bilateralmente en las porciones anterolaterales de la mitad inferior del bulbo raquídeo, centro cardiomodulador. Las neuronas gabaérgicas de esta área son activadas por neuronas glutaminérgicas provenientes del núcleo del tracto solitario, así como por fibras con neurotransmisores como la angiotensina II e inhibida en forma tónica por el GABA y los opioides. Las fibras de estas neuronas se proyectan hacia arriba, al área vasoconstrictora e inhiben la actividad de esa área, por lo que causan vasodilatación. Se han encontrado una alta concentración de axones desde esa área a todo lo largo de la formación reticular bulbar lateral, que incluyen: el núcleo retrofacial, el NA, el ARVL, el hipogloso, el núcleo intercalado, y el núcleo facial. (Alfonso & Báez 2008) Estos datos sugieren que existe una interacción funcional entre sitios bulbares en el control del sistema nervioso parasimpático y simpático, como plantea Porges en su teoría polivagal (Porges 2001). El área vasoconstrictora o área rostral ventro-lateral (ARVL), juega un papel esencial en el control estimulante y reflejo del tono simpático vasomotor. Estas neuronas son glutaminérgicas, pero al igual que en otras áreas, su actividad está modulada por diferentes neurotrasmisores. Las neuronas de esta área excitan a las neuronas preganglionares del simpático, que a su vez inervan al corazón, vasos sanguíneos y glándulas adrenales. La estimulación de los nervios simpáticos libera norepinefrina en los receptores β-1 adrenérgicos, aumentando la permeabilidad de membrana a los iones sodio y calcio, excitando al nodo sinusal, nodo aurículo-ventricular (AV) y en los haces AV con efectos inotrópicos y cronotrópicos positivos. La acetilcolina liberada en las terminales nerviosas del vago, en los receptores muscarínicos (principalmente M2), incrementa enormemente la permeabilidad de las membranas a los iones de potasio, provocando hiperpolarización, lo cual las hace mucho menos excitables. En el nodo sinusal, el estado de hiperpolarización aumenta 7 Facultad Biología UB Ginés Calín Máster Fisiología Integrativa el potencial de membrana de reposo de las fibras del nodo sinusal, con efectos inotrópicos y cronotrópicos negativos. Si la estimulación del vago es suficientemente fuerte, entonces se logra detener enteramente la auto-excitación rítmica de éste. En el nodo A-V, un estado de hiperpolarización, atrasa la conducción del impulso, pero una disminución intensa puede bloquear completamente la conducción. En este punto las fibras de Purkinje, usualmente en la porción ventricular del septo en el haz A-V, desarrollan un ritmo propio y causa contracción ventricular con un ritmo de 15 a 40 pulsaciones por minuto y una disminución de la fuerza contráctil del 30%. La respuesta a estímulos vagales es casi instantánea (<1”) mientras que el estímulo ortosimpático es algo más diferido (5’’ a 30’’) en su respuesta. Los potenciales de acción conducidos por el sistema nervioso cardíaco intrínseco dispara norepinefrina y epinefrina hacia los receptores β-1 adrenérgicos de las fibras musculares cardíacas (Shaffer et al. 2014) Se ha evaluado el papel que desempeñan las neuronas de los núcleos ambiguo (fibras B mielínicas) y NMDX (motor dorsal, con fibras C amielínicas) en el control de la actividad cardíaca en la adaptación al entorno desde un punto de vista filogenético, pero este punto será desarrollado más adelante en este trabajo. MECANISMOS DE REGULACIÓN DEL SISTEMA NERVIOSO CARDÍACO INTRÍNSECO Los datos actuales indican que las propiedades emergentes que aparecen en la mayoría de las neuronas que componen el neuroeje cardíaco tendrán que ser tomadas en consideración. Resultados derivados de modelos animales en grandes mamíferos han ayudado a clarificar la hipótesis de que las neuronas del plexo ganglionar de la aurícula derecha únicamente controlan al nodo sinoauricular mientras que los de la vena cava inferior (auricular) controlan sólo el nodo aurículo-ventricular. 8 Facultad Biología UB Ginés Calín Máster Fisiología Integrativa Esas neuronas cardíacas intrínsecas están bajo la influencia tónica de hormonas circulantes y otros neurotransmisores liberados directamente al corazón por las terminales de los nervios autonómicos eferentes postganglionares. Es importante recalcar que el corazón está enmarcado en un pericardio fibroso que no sólo previene el desplazamiento del órgano sino también limita la distensión diastólica ventricular latido a latido. La evidencia indica que neuronas intratorácicas extracardíacas y neuronas intrínsecas cardíacas componen una red local distributiva que procesa tanto información centrípeta como centrífuga respecto al control cardíaco. Su función integrativa está bajo la influencia tónica del troncoencéfalo y de la médula espinal, así como de hormonas circulantes. Muchos de los somas neuronales en los ganglios intratorácicos, incluyendo los intracardíacos, proyectan axones solamente hacia otras neuronas dentro del mismo ganglio. Otros somas proyectan axones hacia neuronas en diferentes ganglios intratorácicos mientras otros proyectan axones hacia neuronas centrales. Así envían la información transducida por los quimiorreceptores y barorreceptores del corazón así como de los principales vasos sanguíneos intratorácicos. Colectivamente las poblaciones de neuronas intratorácicas presentan capacidad de memoria relativa a eventos cardiovasculares transducidos durante cada ciclo cardíaco, para influenciar a los outputs neuronales eferentes al corazón durante el propio o los subsiguientes ciclos cardíacos. Como consecuencia las perturbaciones cardiacas iniciadas durante un latido arrítmico simple, pueden afectar a los outputs neuronales durante algunos ciclos cardíacos más. Este complejo sistema nervioso intracardiaco, junto con neuronas autonómicas extracardíacas, regulan integrativamente la actividad cardiaca (el ritmo, la conducción de los impulsos y la fuerza contráctil del miocardio), tanto en condiciones fisiológicas, como patológicas como sucede en los diversos trastornos del ritmo cardiaco, comportándose como un sistema de control estocástico, al que han venido a llamar “Little Brain” .(Armour 2008) 9 Facultad Biología UB Ginés Calín Máster Fisiología Integrativa Fig. 2 Representación de una vista posterior del corazón humano y vasos principales ilustrando las localizaciones de los plexos ganglionares posteriores atrial y ventricular. Atención a los nervios mediastínicos pasando adyacentes al cayado aórtico y uniendo los dos plexos ganglionares superiores atriales. Son mostradas las ubicaciones de la vena cava superior (SVC), vena cava inferior (IVC), ventrículo derecho (RV), ventrículo izquierdo (LV) (Armour et al. 1997) Fig. 3 Descripción de una vista superior del corazón humano ilustrando la distribución de los plexos ganglionares en la superficie de atrias y ventrículos. (Armour et al. 1997) 10 Facultad Biología UB Ginés Calín Máster Fisiología Integrativa MECANISMOS DE REGULACIÓN DEL SISTEMA ADRENÉRGICO INTRÍNSECO CARDÍACO. El corazón no sólo funciona como un centro de procesamiento y codificación de intrincada información, sino también como glándula endocrina que puede producir y secretar sus propias hormonas y neurotransmisores. Por ejemplo, miocitos atriales pueden secretar el péptido natriurético atrial (ANP) de cuyas funciones hablaré más adelante. Por otro lado han sido identificadas células con capacidad para la síntesis de catecolaminas como dopamina, epinefrina y norepinefrina, además de altas concentraciones de oxitocina (Shaffer et al. 2014), llamadas Intrinsic Cardíac Adrenergic cells (ICA), presentes en los corazones de fetos humanos en una etapa de desarrollo (semana 20) antes de la inervación simpática del corazón. Estos hallazgos apoyan la idea de que estas células constituyen un sistema de señalización ICA capaz de participar en la regulación cardíaca que parece ser independiente de la inervación simpática. Pacientes transplantados de corazón mantinenen una función cardíaca adecuada en ausencia de reinervación simpática. Las catecolaminas identificadas en las células ICA primarias aisladas no son debidas a la contaminación de las terminaciones nerviosas simpáticas ya que ni los análisis de inmunohistoquímica ni la microscopía electrónica han detectado ninguna terminación nerviosa simpática o estructuras axonales en aislamientos celulares de ICA. La presencia de esta población diferente de células adrenérgicas es esencial durante la ontogenia cardíaca y tiene importantes implicaciones en la regulación postnatal de la función cardíaca. (Huang et al. 1996) MECANISMOS DE REGULACIÓN ENDOCRINA DE LA FUNCIÓN CARDÍACA CRF Y UROCORTINAS Es bien conocida la función del neuropéptido Corticotropin Releasing Factor (CRF) en la función mediadora neuroendocrina de regulación del eje Hipotálamo-HipofisarioAdrenal en los mecanismos de adaptación al estrés. Posteriormente fueron descubiertos unos neuropéptidos similares llamados CRFlike o bien Urotortinas (Uc). 11 Facultad Biología UB Ginés Calín Máster Fisiología Integrativa Ambos tienen efectos centrales y periféricos hemodinámicos vía la activación de CRFreceptores tipo 1 (sistema nervioso) y tipo 2 (sistema nervioso y cardíaco), y son importantes hormonas cardioactivas y de adaptación al estrés. Los receptores CRF1 son abundantes en el sistema nervioso central, cerebro y médula espinal, así como a través del sistema nervioso autónomo. Tiene una expresión predominante en la pituitaria, córtex, amígdala, hipocampo, cerebelo y bulbo olfatorio. En cambio están muy débilmente expresados en el corazón, en la piel, gónadas o las suprarrenales. Los receptores CRF2 tienen 3 variantes (a,b,c). CRF2a es expresado tanto en cerebro como en órganos periféricos, CRF2b principalmente en cerebro, mientras que CRF2c se expresa en el sistema límbico hipocampo y en menor proporción en amígdala. También se han descrito tres estructuras diferentes para las urocortinas Uc1, Uc2, Uc3 donde Uc2 y Uc3 tienen afinidad específica por los receptores tipo 2, mientras que Uc1 tiene afinidad por receptores tipo 1. Estudios realizados por inyecciones centrales de CRF y sus análogos aumentan el ritmo cardíaco y causan un efecto inotrópico positivo a través de la activación del sistema nervioso ortosimpático y médula suprarrenal. Sin embargo, el aumento del antagonismo con el parasimpático genera un aumento en la variabilidad cardíaca causada por receptores CRF1, con efectos en los núcleos motor dorsal del vago en el tronco cerebral. Cuando CRF o urocortinas son inyectadas periféricamente, inducen a través de los receptores CRF2 aumentos en el gasto cardíaco y la fracción de eyección ventricular izquierda junto con un aumento en la contractilidad cardíaca, disminución de la resistencia vascular (mediada por óxido nítrico) y significativos aumentos sistemáticos en péptido natriurético atrial (ANP) y péptido natriurético cerebral (BNP) y adrenomedulina. La activación del receptor CRF2 también ejerce directamente efectos inotrópicos positivos más efectos lusitrópicos en los miocitos cardíacos por estimulación de PKA., el incremento de Calcio sarcoplásmico conduce a un incremento de liberación de calcio durante la sístole resultando en un incremento de la contractilidad del miocito. (Yang et al. 2010) Se propone una vía de comunicación cerebro cardíaca neuroendocrina (ver fig 4). 12 Facultad Biología UB Ginés Calín Máster Fisiología Integrativa Fig. 4 Resumen esquemático de las acciones de la CFR y urocortins entre el corazón y el cerebro. Estrés hipotalámico (HYP), CRF induce la liberación de la hormona adrenocorticotrópica (ACTH) de la pituitaria (PIT), que a su vez estimula la liberación de glucocorticoides por la corteza suprarrenal, mejorado la tasa de la contractilidad vascular así como el aumento en la presión arterial, mediada por β-endorfina tras liberación de catecolaminas de la médula suprarrenal. CRF central puede modular la actividad del sistema nervioso autónomo a través del núcleo motor dorsal del vago (DMV), lo que resulta en la disminución de la frecuencia cardíaca y aumento de la variabilidad cardíaca debido a un mayor antagonismo simpatovagal. CRF y urocortins periféricas median la relajación vascular y la posterior disminución de la presión sanguínea, mientras que la acción directa sobre receptor CRF2 en el corazón genera una mayor contractilidad de los miocitos. Efecto barorreflejo (aumento de la frecuencia cardíaca) en condiciones de baja presión arterial están mediadas a través del núcleo del tracto solitario (NTS) y eferentes simpáticas. Urocortinas periféricas inyectadas cruzan la barrera hematoencefálica puede provocar efectos secundarios por acciones centrales. (Yang et al. 2010) 13 Facultad Biología UB Ginés Calín Máster Fisiología Integrativa SISTEMA RENINA-ANGIOTENSINA-ALDOSTERONA (RAAS) En el sistema renina-angiotensina-aldosterona (RAAS), la renina (de liberación renal), convierte angiotensinogeno (hepático) en angiotensina 1 y la enzima convertidora de angiotensina (ACE) convierte a la forma activa de angiotensina 2, un potente vasoconstrictor que incrementa la liberación de aldosterona. Los receptores de angiotensina AT1 participan de la hipertrofia del miocardio y los receptores AT2 participan de la apoptosis de las células cardíacas mientras que la aldosterona participa de la fibrosis del miocardio además de la retención de sodio y agua con el consiguiente aumento de la tensión arterial. ACE contribuye a la degradación del vasodilatador bradikinina (Kjaer & Hesse 2001). Áreas anatómicas en el NTS han sido identificadas con acción específica de la aldosterona, con un importante rol similar a la acción del vago. Hallazgos por RNM apoyan que las hiperintensidades de materia blanca subcortical, que son frecuentemente consecuencia de factores de riesgo cardiovascular, son también un factor de riesgo para la depresión en ancianos, y están asociadas a altas concentraciones de aldosterona en pacientes con polimorfismos genéticos del sistema renina-angiotensia-aldosterona (RAAS). RAAS está íntimamente ligado a la regulación del sueño en sujetos sanos, con un incremento de su actividad durante las fases de sueño de ondas lentas (SWS). Durante SWS las secreciones de cortisol están suprimidas, tanto en sujetos sanos como en pacientes con depresión, y está asociado a la secreción de renina y en algunas circunstancias de aldosterona. RAAS también está asociado a la regulación del estado de ánimo, a través de la secreción de monoaminas como serotonina, norepinefrina y dopamina, el sistema de receptores NMDA (en condiciones normales bloqueado por magnesio e implicados en procesos de aprendizaje y memoria), y el control de procesos de inflamación. Hay una relación importante entre la acción de los mineralocorticoides y el eje HPA, cuando se liberan en el área del hipocampo conducen a una reducción del eje HPA, habiendo una interacción entre los receptores de mineralocorticoides y glucocorticoides a nivel central. 14 Facultad Biología UB Ginés Calín Máster Fisiología Integrativa En situaciones patológicas, la elevación de aldosterona conduce a disforia e hiperaldosteronismo (Sd. Conn) asociado a asiedad y somatización. La reducción de la HRV asociada al estrés parece estar también asociada al metabolismo del magnesio, determinado por sus niveles intraeritrocitarios. La actividad del ortosimpático incrementada puede generar una mayor excreción de magnesio. La administración de aldosterona conduce a una mayor excreción renal de magnesio en animales de experimentación. Es ya bien conocido el principal efecto de la aldosterona para retener sodio. La activación de RAAS ha sido asociada a cambios inflamatorios, con un efecto de la alosterona en las células endoteliales induciendo daño vascular. La acción proinflamatoria de la aldosterona incluye la activación de la expresión de la ciclooxigenasa-2(COX2) y la reducción intracelular de magnesio, por lo que los procesos inflamatorios inducidos por aldosterona pueden ser revertidos con la administración de magnesio. (Murck et al. 2012) PEPTIDO NATRIURÉTICO AURICULAR Y PEPTIDO NATRIURÉTICO CEREBRAL El Péptido natriurético auricular (ANP) se produce principalmente en las aurículas cardíacas y es liberado por la distensión auricular. ANP es un vasodilatador arterial, aumenta la natriuria y es un antagonista tanto del sistema ortosimpático como del sistema RAAS. Son conocidos tres receptores natriuréticos, A, B y C. ANP tiene mayor afinidad para el receptor A. El receptor de C es un receptor de aclaramiento. Además, los péptidos natriuréticos se degradan por una endopeptidasa, que también rompe otros péptidos, como por ejemplo angiotensina II, bradikinina y endotelina. ANP también tiene un efecto antihipertrófico en los cardiomiocitos. El péptido natriurético cerebral (BNP) principalmente se encuentra en los ventrículos del corazón y se libera principalmente desde el ventrículo izquierdo en respuesta al aumento de la presión arterial. La vida media de BNP es algo más larga (20 ± 25 min) que la de ANP (3 min), aunque su acción fisiológica es idéntica, es decir, vasodilatación y natriuresis. Ambos se encuentran aumentados en las fases tempranas del fallo cardíaco crónico (Kjaer & Hesse 2001) 15 Facultad Biología UB Ginés Calín Máster Fisiología Integrativa VASOPRESINA Y OXITOCINA La vasopresina sintetizada en el hipotálamo y almacenada en la neurohipófisis. Las acciones fisiológicas de la vasopresina son vasoconstricción mediada a través de receptores V1 y disminución de la diuresis mediada a través de receptores renales V2. La sensibilidad de la vasopresina para detectar cambios en la osmolaridad y el volumen menor en el fallo cardíaco crónico, donde aumenta independientemente de los valores osmolares. (Kjaer & Hesse 2001) La vasopresina también ha sido reportada como un elemento de activación ante situaciones potencialmente peligrosas del entorno activando al ortosimpático e inibiendo al parasimpático, así como activando a centros del sistema límbico como la amígdala y el hipocampo reclutando la memoria eventual de sucesos traumáticos. La Oxitocina por su parte estimula la activación del sistema parasimpático e inhibe el eje HPA favoreciendo los procesos de relajación y de involucramiento social con efectos vasodilatadores y aumentando la variabilidad cardíaca (Sue Carter 1998; Porges 2003) ENDOTELINA-1 Niveles de endotelina-1 se correlacionan fuertemente con el aumento de la presión pulmonar en el fallo cardíaco crónico. La endotelina-1 actúa a través de receptores selectivos (ETA o ETB) con fuertes efectos vasoconstrictores y, probablemente también un efecto trófico sobre los cardiomiocitos, encontrado en estudios in vitro. (Kjaer & Hesse 2001) ÓXIDO NÍTRICO El óxido nítrico (NO), se forma en las células endoteliales a partir de L-arginina por la enzima óxido nítrico sintetasa (NOS). A partir de las células endoteliales el NO puede expandirse localmente a células del músculo liso vascular produciendo vasodilatación. La actividad del NO endógeno a menudo se mide como dependiente de la acción de la acetilcolina. NO se puede aumentar de forma exógena, por ejemplo con nitroglicerina. 16 Facultad Biología UB Ginés Calín Máster Fisiología Integrativa NO inhibe el efecto inotrópico positivo de la estimulación β-adrenérgica. (Kjaer & Hesse 2001) CITOKINAS Varias citoquinas parecen estar implicadas en la regulación cardíaca especialmente el Factor de necrosis tumoral –α (TNF-α). Se ha demostrado en animales que el TNF-α provoca la dilatación y la remodelación del ventrículo izquierdo. Uno de los efectos de TNF-α es probablemente una inducción de la NOS y de ese modo NO, que tiene un efecto inotrópico negativo. (Kjaer & Hesse 2001) El IGF-1 estimula la quimiotaxis de macrófagos, la captación de colesterol ligado a lipoproteínas de baja densidad y la liberación de citokinas proinflamatorias. Se sabe poco acerca del papel de las IGFBP y sus proteasas en la aterosclerosis. La IGFBP-2 estimula la mitosis de células musculares lisas mientras que la IGFBP-4 inhibe de forma consistente las acciones mediadas por el IGF en todos los tipos celulares estudiados. Han aislado una nueva proteasa específica de la IGFBP-4 identificada como proteína plasmática-A asociada al embarazo (pregnancy-associated plasma protein-A [PAPPA]). Al degradar la IGFBP-4 y liberar el IGF-1 libre, la PAPP-A podría modular el crecimiento en procesos ateroscleróticos. La PAPP-A se expresa en las células musculares de la capa media vascular, así como en las células endoteliales y macrófagos espumosos del core lipídico del ateroma. (Bayés-Genís 2005) HIPÓTESIS DE CONEXIÓN CEREBRO-CARDÍACA TEORÍA POLIVAGAL El Dr. Stephen Porges propone un modelo de evolución filogenética del SNA asociada a la capacidad comunicativa social de los mamíferos, especialmente en primates y humanos. Esta evolución supera las antiguas concepciones de adaptación al estrés como mecanismo de activación del eje HPA en un sistema de lucha o huída, o bien la reacción de inmovilización reptiliana (congelación o síncope vasovagal), que es vehiculizada por la parte filogenéticamente más antigua del vago, la no mielínica (NMDX). La teoría describe una capacidad de establecer estrategias de autocontrol y 17 Facultad Biología UB Ginés Calín Máster Fisiología Integrativa comunicativas para resolver conflictos, vehiculizada por la parte mielinizada del vago (núcleo ambiguo), en conexión con otros pares craneales V, VII, IX, XI, que le dan capacidades para la comunicación, deglución, masticación, fonación, audición, cuando esta conexión funciona correctamente, pero que genera un estado congelado e inexpresivo cuando se produce una disevolución en el control de todas estas funciones, con repercusiones sobre la frecuencia respiratoria y el ritmo cardíaco, medibles a través de la HRV. Por tanto, las funciones del Vago mielinizado (núcleo ambiguo), el cual sirve para apoyar estados de conducta calmados, inhibiendo las influencias simpáticas al corazón e inhibiendo al eje Hipotálamo–Pituitario-Adrenal (HPA). El sistema de movilización (lucha o huída) depende del funcionamiento del Sistema Nervioso Simpático. El componente filogenéticamente más primitivo del hombre, el sistema de inmovilización, es dependiente del Vago amielínico (NMDX), el cual está presente en la mayoría de los vertebrados. La teoría polivagal propone un sistema de control jerárquico, donde las estructuras filogenéticamente más evolucionadas controlan a las más antiguas, proporcionando así un control adaptativo a situaciones de estrés mediante estados calmados y resolución de problemas a través de la anticipación del córtex prefrontal y la inhibición de la amígdala, el hipocampo, el eje HPA o el NMDX, además de la activación del núcleo ambiguo. Considera el estado congelado como una situación de disevolución patológica, presente en los pacientes con estrés postraumático y que predispone a una hiperactivación del eje HPA en estado de alerta constante o bien una incapacidad para establecer comunicaciones sociales con su entorno. Todo ello acompañado de implicaciones clínicas de morbilidad y mortalidad cardíacas. (Porges 2001; Porges 2003; Porges 2007a; Porges 2007b). 18 Facultad Biología UB Ginés Calín Máster Fisiología Integrativa EL MODELO DE INTEGRACIÓN NEUROVISCERAL CENTRAL Fig. 5 Un diagrama esquemático compuesto que muestra las vías por las que la corteza prefrontal puede influir en el control de la frecuencia cardiaca. La corteza prefrontal, córtex del giro cingular, e ínsula forman una red interconectada en comunicación bidireccional con la amígdala. La amígdala está bajo control inhibitorio tónico través de vías prefrontales vagales de células intercaladas en la amígdala. La activación del núcleo central de la amígdala (CEA) inhibe el núcleo del tracto solitario (NTS), que a su vez inhibe al centro medular caudal ventrolateral (CVLM), conexiones al centro medular rostral ventrolateral (RVLM) y neuronas excitatorias simpáticas, e inhibe simultáneamente las neuronas motoras vagales en el núcleo ambiguo (NA) y el núcleo motor dorsal vagal (DVN). Además, el CEA puede activar directamente las neuronas excitatorias simpaticas en la RVLM. Una inhibición de la corteza prefrontal causaría una desinhibición del CEA que conduciría a la desinhibición de los circuitos cardio-aceleradores medulares y un aumento en la frecuencia cardíaca.(Thayer & Lane 2009) 19 Facultad Biología UB Ginés Calín Máster Fisiología Integrativa El Dr. Thayer propone un modelo de integración central neurovisceral que describe como un conjunto de estructuras neurales implicadas en funciones cognitivas, afectivas y en la regulación autonómica, relacionan la HRV y el rendimiento cognitivo. En este modelo de sistemas complejos, una red autonómica Central (CAN) enlaza el NTS en el tronco cerebral con las estructuras del cerebro anterior (incluyendo la corteza cingulada anterior, la ínsula, la corteza prefrontal ventromedial, la amígdala y el hipotálamo) a través de bucles de retroalimentación y bucles directos. Propone que esta red es un sistema integrado para la regulación interna del sistema por el cual el cerebro controla el input visceromotor, neuroendocrino, y las respuestas de comportamiento que son críticas para el comportamiento dirigido a un objetivo, la adaptabilidad y la salud. Cuando el córtex frontal reduce su control inhibitorio sobre el núcleo central de la amígdala, causa una desinhibición amigdalar generando un incremento del ritmo cardíaco vía una disminución en la inhibición de las neuronas simpáticas excitadoras en el RVLM al mismo tiempo que inhibición del output motor vagal en el NA y NMDX. Las funciones inhibitorias del córtex prefrontal derecho que facilitan la memoria de trabajo, capacidad de atención, respuestas de inhibición, regulación emocional y afectiva, han sido ya ampliamente demostradas. Dada la predominancia del hemisferio derecho sobre la inervación del nodo sinoatrial (vago derecho), Thayer propone que las conocidas habilidades del hemisferio derecho para gestionar el aspecto emocional pueden ser secundarias a la interacción con la inervación cardíaca por el mismo hemisferio. En el modelo de integración neurovisceral central se propone la relación entre HRV e importantes regulaciones fisiológicas, cognitivas y emocionales, debida a la capacidad de HRV para indicar la actividad en una red flexible de estructuras neurales que son organizadas dinámicamente en respuesta a retos ambientales (Thayer & Lane 2009; Thayer et al. 2012) 20 Facultad Biología UB Ginés Calín Máster Fisiología Integrativa CONCLUSIONES A lo largo de toda esta revisión hemos constatado la compleja red de relaciones que se establecen como mecanismos de control nervioso, endocrino, e incluso inmune, a diferentes niveles de profundidad, con propiedades emergentes difícilmente predecibles ya que dependen de la adaptación a un entorno abierto y cambiante. Se han propuesto dos modelos teóricos de integración ordenada de toda esta red de comunicación bidireccional entre cerebro y corazón, probablemente los sistemas más importantes del cuerpo humano. No obstante nuevos paradigmas y nuevos estudios pueden emerger a la luz de toda la evidencia creciente en el terreno de los mecanismos de control neuroendocrino del corazón. Cualquier intervención terapéutica mecanicista que no tenga en cuenta la compleja red de adaptacionescompensaciones que se establece en estos sistemas, por su propia naturaleza estocástica, tiene altas probabilidades de fracasar o causar considerables efectos adversos. La variabilidad cardíaca emerge en este contexto como una potencial herramienta de diagnóstico, pronóstico e incluso de tratamiento por biofeedback (Shaffer et al. 2014), para trabajar sobre las vías neuroendocrinas de regulación del eje cerebro-cardíaco. REFERENCIAS Alfonso, J. & Báez, M., 2008. Regulación del Sistema Cardiovascular por el Sistema Nervioso Autónomo. librosabiertos, p.36. Available at: http://mail.fbio.uh.cu/ginvest/mesna/vfc_docs/RegulacionAutonomicaCardiovascul ar.pdf. Armour, J. a, 2008. Potential clinical relevance of the “little brain” on the mammalian heart. Experimental physiology, 93(2), pp.165–76. Available at: http://www.ncbi.nlm.nih.gov/pubmed/17981929 [Accessed November 23, 2014]. 21 Facultad Biología UB Ginés Calín Máster Fisiología Integrativa Armour, J.A. et al., 1997. Gross and Microscopic Anatomy of the Human Intrinsic Cardiac Nervous System. The anatomical record, 298(September 1996), pp.289– 298. Bayés-Genís, A., 2005. El corazón como órgano endocrino. ¿Cuál es su relevancia? Endocrinología y Nutrición, 52(6), pp.273–276. Coote, J.H., 2007. Landmarks in understanding the central nervous control of the cardiovascular system. Experimental physiology, 92(1), pp.3–18. Available at: http://www.ncbi.nlm.nih.gov/pubmed/17030558 [Accessed January 18, 2015]. Huang, M.H. et al., 1996. An intrinsic adrenergic system in mammalian heart. Journal of Clinical Investigation, 98(6), pp.1298–1303. Kjaer, A. & Hesse, B., 2001. Heart failure and neuroendocrine activation: diagnostic, prognostic and therapeutic perspectives. Clinical physiology (Oxford, England), 21(6), pp.661–672. Murck, H., Schüssler, P. & Steiger, A., 2012. Renin-angiotensin-aldosterone system: The forgotten stress hormone system: Relationship to depression and sleep. Pharmacopsychiatry, 45(3), pp.83–95. Porges, S.W., 2007a. A phylogenetic journey through the vague and ambiguous Xth cranial nerve: a commentary on contemporary heart rate variability research. Biological psychology, 74(2), pp.301–7. Available at: http://www.sciencedirect.com/science/article/pii/S0301051106001906 [Accessed November 9, 2014]. Porges, S.W., 2007b. The polyvagal perspective. Biological psychology, 74(2), pp.116– 43. Available at: http://www.sciencedirect.com/science/article/pii/S0301051106001761 [Accessed July 10, 2014]. Porges, S.W., 2003. The Polyvagal Theory: phylogenetic contributions to social behavior. Physiology & Behavior, 79(3), pp.503–513. Available at: http://www.sciencedirect.com/science/article/pii/S0031938403001562 [Accessed July 12, 2014]. 22 Facultad Biología UB Ginés Calín Máster Fisiología Integrativa Porges, S.W., 2001. The polyvagal theory: phylogenetic substrates of a social nervous system. International Journal of Psychophysiology, 42(2), pp.123–146. Available at: http://www.sciencedirect.com/science/article/pii/S0167876001001623 [Accessed November 15, 2014]. Prigogine, I., 1997. Nonlinear Science and the Laws of Nature. International Journal of Bifurcation and Chaos, 07(09), pp.1917–1926. Rhee, S.S. & Pearce, E.N., 2011. Update: Systemic Diseases and the Cardiovascular System (II). The endocrine system and the heart: a review. Revista española de cardiología, 64(3), pp.220–31. Available at: http://www.sciencedirect.com/science/article/pii/S0300893210000813 [Accessed December 5, 2014]. Shaffer, F., McCraty, R. & Zerr, C.L., 2014. A healthy heart is not a metronome: an integrative review of the heart’s anatomy and heart rate variability. Frontiers in Psychology, 5, p.1040. Available at: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=4179748&tool=pmcentr ez&rendertype=abstract [Accessed October 7, 2014]. Sue Carter, C., 1998. NEUROENDOCRINE PERSPECTIVES ON SOCIAL ATTACHMENT AND LOVE. Psychoneuroendocrinology, 23(8), pp.779–818. Available at: http://www.sciencedirect.com/science/article/pii/S0306453098000559 [Accessed November 15, 2014]. Thayer, J.F. et al., 2012. A meta-analysis of heart rate variability and neuroimaging studies: implications for heart rate variability as a marker of stress and health. Neuroscience and biobehavioral reviews, 36(2), pp.747–56. Available at: http://www.sciencedirect.com/science/article/pii/S0149763411002077 [Accessed July 11, 2014]. Thayer, J.F. & Lane, R.D., 2009. Claude Bernard and the heart-brain connection: further elaboration of a model of neurovisceral integration. Neuroscience and biobehavioral reviews, 33(2), pp.81–8. Available at: http://www.sciencedirect.com/science/article/pii/S0149763408001255 [Accessed July 16, 2014]. 23 Facultad Biología UB Ginés Calín Máster Fisiología Integrativa Yang, L.-Z. et al., 2010. Corticotropin-releasing factor receptors and urocortins, links between the brain and the heart. European journal of pharmacology, 632(1-3), pp.1–6. Available at: http://www.ncbi.nlm.nih.gov/pubmed/20132811 [Accessed November 26, 2014]. 24