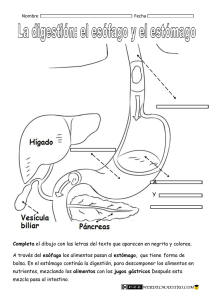
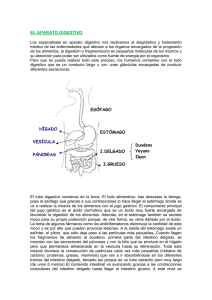
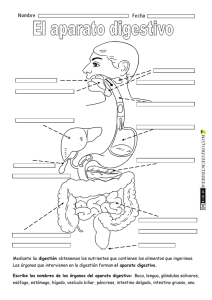
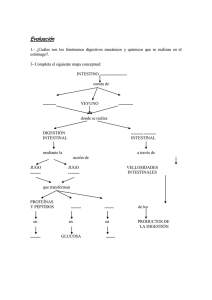
FISIOLOGIA II FUNCIONES GASTROINTESTINALES
Anuncio

FISIOLOGIA II FUNCIONES GASTROINTESTINALES Introducción. Control de la función digestiva (1) A pesar de la gran divergencia entre las dietas de las diferentes especies de animales, el aparato digestivo realiza unas funciones similares en cada una, y a pesar de estar adaptado a la dieta correspondiente, también presenta una estructura similar. Las funciones digestivas son: ingestión del alimento; transporte del alimento a una velocidad que permita una digestión y absorción óptimas; secreción de líquidos, sales y enzimas digestivas; digestión; absorción de los productos resultantes de la digestión; y expulsión de los restos digeridos fuera del organismo (defecación). Las estructuras digestivas están adaptadas anatómicamente a cada especie (carnívoros, omnívoros y herbívoros), sin embargo son las mismas en cada una de ellas: el compartimiento gástrico (estómago) tiene la función de secretar ácidos, continuar la digestión de los alimentos y rebajar la carga bacteriana; el primer compartimiento intestinal (intestino delgado) tiene la función de absorber y digerir los nutrientes absorbibles; y el segundo compartimento intestinal (intestino grueso) tiene la función de recuperar agua y electrolitos y realizar la excreción de lo sobrante. En cada órgano se dan tanto funciones secretoras como motoras, pero su independencia entre los unos y los otros provoca que su regulación sea muy compleja. La diversidad de funciones y las diferencias en la composición y en la cantidad de la dieta en un mismo animal requieren unos sistemas de control peculiares. Además, el tracto digestivo está colonizado por microorganismos, lo que hace que haya la mayor concentración de células inmunes de todo el organismo; estas células inmunes también regulan la actividad del sistema digestivo. La función digestiva tiene una regulación muy compleja que requiere diversos mecanismos de control: · Control nervioso: El aparato digestivo se controla principalmente por el sistema nervioso autónomo, concretamente el sistema nervioso entérico, y por los plexos intrínsecos encontrados en las paredes intestinales. Estos plexos intrínsecos donde se encuentran las neuronas y los ganglios se denominan plexo submucoso (interno) y plexo mientérico (externo). Hay más cantidad de neuronas que en el cerebro. · Control hormonal: También se encuentra regulado bajo control hormonal por las células endocrinas situadas en el tracto digestivo y que son secretoras de hormonas peptídicas. · Control humoral: Y además está controlado por el sistema inmunitario debido a la gran cantidad de bacterias que habitan en el aparato digestivo. Este control humoral está producido por mastocitos secretores de histamina, células enterocromafines secretoras de serotonina, células epiteliales secretoras de interleucinas, células musculares secretoras de factores de crecimiento… El sistema inmune reconoce la microbiota intestinal y modula los reflejos nerviosos que se dan en el tracto digestivo; por ejemplo: controlan la secreción de HCl en el estómago. Para que se produzcan adecuadamente estos controles se requiere: · Una innervación extrínseca · Un sistema nervioso entérico, localizado en la propia pared del tracto digestivo con una gran diversidad de neuronas, tanto por su función (sensitivas, interneuronas y motoneuronas) como por los múltiples neurotransmisores que liberan. Esto permite realizar tanto respuestas inhibitorias como respuestas excitantes para las secreciones y la actividad motora. Además, la autonomía de cada segmento garantiza la respuesta adecuada a cada situación particular. El control nervioso de la función gastrointestinal se realiza bajo la influencia de 4 niveles distintos: · En el nivel 1 se encuentra el sistema nervioso entérico, el cual recibe información sensorial (vía aferente) desde el tracto digestivo, y envía información motora (vía eferente) hacia el tracto digestivo. · En el nivel 2 se encuentran los ganglios prevertebrales, los cuales reciben información sensorial (vía aferente) desde el sistema nervioso entérico (1), y envían información motora (vía eferente) hacia el sistema nervioso entérico y el tracto digestivo. · En el nivel 3 se encuentran las áreas autónomas centrales. Las áreas parasimpáticas centrales reciben información sensorial (vía aferente por el nervio vago) desde el tracto intestinal, y envían información motora (vía eferente por el nervio vago) hacia el sistema nervioso entérico (1); las áreas simpáticas centrales reciben información sensorial (vía aferente) desde el tracto intestinal, y envían información motora (vía eferente) hacia los ganglios prevertebrales (2), nunca directamente a las células efectoras.. · En el nivel 4 se encuentra el SNC y los centros superiores, los cuales reciben información sensorial (vía aferente) desde las áreas parasimpáticas centrales (por el nervio vago) y áreas simpáticas centrales, y envían información motora (vía eferente) hasta las mismas. Las neuronas aferentes se comunican con sus órganos diana a través de la sustancia P y del CGRP (péptido relacionado con el gen de la calcitonina); y las neuronas eferentes se comunican a través de una gran variedad de neurotransmisores excitantes (acetilcolina, sustancia P…) y inhibitorios (NO, VIP, ATP…). La motilidad digestiva está controlada tanto por una innervación extrínseca como por una innervación intrínseca. Los péptidos reguladores gastrointestinales son secretados por células polarizadas endocrinas que son sensibles a la composición de los alimentos gracias a unos quemoreceptores situados en la membrana apical de las células que reconocen cada tipo de nutriente. Además, para la detección de la composición química del contenido intestinal, existen a lo largo del tracto gastrointestinal, unas células endocrinas que también regulan tanto las secreciones como la motilidad digestiva. Estas células captan estímulos mecánicos (deformación, distensión, estiramiento…) y químicos (pH…) y secretan gran variedad de hormonas digestivas con funciones variadas y que se distribuyen de manera diferente: Hormona Gastrina Secretina Colecistocinina (CCK) Péptido inhibitorio gástrico (GIP) Motilina Producción Acción Estímulo de liberación Estómago distal Estimula la secreción ácida de las Proteínas en el glándulas gástricas. estómago; un pH Estimula la motilidad gástrica y gástrico alto; la crecimiento del epitelio estomacal. estimulación vagal. Duodeno Estimula la secreción de bicarbonato Ácido en el por el páncreas. duodeno. Estimula la secreción biliar de bicarbonato. Duodeno hasta el Estimula la secreción de enzima del Proteínas y grasas en íleon, con la mayor páncreas. el intestino delgado. concentración en el Inhibe el vaciamiento gástrico. duodeno Duodeno y parte Inhibe la motilidad gástrica y la Carbohidratos y superior del yeyuno. actividad secretora. grasas en el intestino Estimula la secreción de insulina, delgado. siempre y cuando exista suficiente glucosa presente; puede ser la acción más importante en muchas especies. Duodeno y yeyuno. Es probable que controle el patrón Acetilcolina. de motilidad del intestino en los periodos interprandiales. Puede regular el tono del esfínter esofágico inferior. Secreciones digestivas (2) Toda la mucosa digestiva, desde la cavidad oral hasta el recto, necesita de secreciones para evitar lesiones cuando circule el material sólido por su luz. Las secreciones digestivas más importantes son la secreción salivar, gástrica, biliar y pancreática. Todas estas secreciones son mayoritariamente acuosas y contienen electrolitos y enzimas que les confieren sus características especiales a cada una de ellas para realizar funciones como el tamponar (excepto gástrica), el transportar enzimas (excepto biliar)... Además, en todo el tracto gastrointestinal hay células caliciformes cuya función principal es producir una secreción rica en mucopolisacáridos muy ramificados (moco) que presenta diferencias en cada órgano, pero siempre tiene como función la protección de las mucosas digestivas gracias a su dificultad por ser digerido. Secreción salivar (2.1) Las glándulas salivares son exocrinas. La composición de la saliva varía según las especies para adaptarse a la dieta. Las funciones generales de la secreción salival son: · Lubricación de la ingesta con la ayuda de la mucina o el moco, lo que facilita el proceso de deglución. · Protección de la boca manteniendo húmedo el epitelio bucal; además diluye y tampona las sustancias nocivas y controla el crecimiento bacteriano gracias a sus sustancias antibacterianas. · Iniciación del proceso de digestión gracias al contenido de los enzimas digestivos. · Facilita las sensaciones gustativas al disolver las sustancias gustativas y producir un lavado de las papilas gustativas. · Excreción · Termorregulación y regulación del equilibrio hídrico. Las glándulas salivales están formadas por diferentes tipos de células: las células secretoras acinares que forman los acinos y generan la saliva primaria; las células mioepiteliales que forman el conducto intercalar; y las células ductales que forman el conducto granular y modifican la saliva primaria a medida que circula por el conducto. La saliva está compuesta por: · Enzimas: Son enzimas no imprescindibles para el proceso digestivo. La α-amilasa es una enzima que hidroliza los enlaces carbonados α1:4 y degrada almid ón generando dextrinas; su actividad es sensible al pH por lo que en el ambiente ácido de la zona distal del estómago se inactiva (tarda 1 o 2 horas en llegar a esa zona); no se secreta en carnívoros ni bóvidos; y es especialmente importante en humanos y suidos. Las lipasas son muy importantes en rumiantes lactantes porque colaboran en la digestión de la grasa de la leche. Las lisozimas son enzimas no digestivos formados por muramidasa y con actividad antibacteriana y antivírica. · Inmunoglobulinas: La saliva contiene inmunoglobulinas A que se concentran en las mucosas y tienen actividad antibacteriana y antivírica. · Urea: Su secreción es poco importante en condiciones normales, sin embargo la saliva es una fuente de excreción en estados de insuficiencia renal. Es un componente significativo en rumiantes porque actúa como agente tampón del pH ruminal y es la base para la síntesis de proteínas bacterianas ruminales. La secreción de agua y electrolitos tiene lugar en los acinos pero la composición se modifica al paso por los conductos. La formación de la secreción salival se distribuye en dos fases: · La saliva inicial o primaria se genera en los acinos glandulares formada por un filtrado plasmático isotónico. Las células acinares secretan electrolitos mediante mecanismos de transporte activo y difusión a partir de la sangre. También secretan agua como flujo osmótico a favor de gradiente generado por la secreción de iones. Finalmente se adicionan compuestos específicos de las células acinares. · La saliva inicial sufre una modificación en los conductos. Esta modificación no incluye movimientos de agua debido a la baja permeabilidad del epitelio tubular, sin embargo, sí secretan activamente bicarbonato con alta actividad de anhidrasa carbónica. El epitelio reabsorbe iones cloruro pasivamente debido a los gradientes eléctricos generados por el transporte activo de Na+ y K+, el primero de los cuales se reabsorbe y el segundo se secreta debido a la actividad de la bomba Na+/K+ encontrada en la membrana basolateral y regulada por la aldosterona. La mayor o menor actividad de los túbulos modificadores se regula en función de la aldosterona y la ADH, las cuales modulan la sensación de ser que será provocada por la saliva según su concentración en Na+. El producto final de la secreción es un fluido supuestamente hipotónico y alcalino. Sin embargo, la composición final de la saliva depende del grado de estimulación de los acinos, es decir, la tonicidad depende del flujo. Cuanto mayor sea el flujo de saliva producido, menor importancia tiene la modificación realizada por el conducto modificador, por lo que el líquido quedaría isotónico. En los rumiantes se puede observar como siempre está estimulada la secreción de saliva, por lo que su composición es muy parecida a la secreción primaria (isotónica). La secreción salivar se regula bajo la actividad del SNA parasimpático, el cual se estimula mediante pensamientos (estimulación cefálica), mediante el contacto del bolo alimenticio con la cavidad bucal (detectado por el nervio glosofaringeo que envía información sensorial eferente hacia el centro de la salivación que responde con una activación de la glándula salivar de forma aferente), mediante el miedo, tras hacer ejercicio físico… El SNA parasimpático activa las células acinares mediante su secreción de acetilcolina para aumentar la su secreción y producir hipersalivación; y mediante unas neuronas inhibidoras que secretan VIP se produce una vasodilatación de los capilares que rodean las células acinares para disminuir el flujo de sangre y permitir mejor su absorción. Secreción pancreática (2.2) Los acinos pancreáticos exocrinos funcionan del mismo modo que los acinos salivares. La parte exocrina del páncreas está formada por una estructura acinar donde se encuentran célulares acinares polarizadas que realizan una secreción proteica; por una zona de transición entre el acino y el conducto donde se encuentran células centroacinares que secretan agua rica en iones; y un sistema de conductos formados por células ducturales que secretan agua rica en iones. Desde que la secreción se forma hasta que se expulsa circula por el acino, los conductos acinares, los conductos intralobulares, los conductos interlobulares y el conducto pancreático. La secreción pancreática exocrina es una secreción acuosa isotónica compuesta por electrolitos (Na+, K+, Cl- y HCO3-) y muchas proteínas de tipo enzimático digestivas esenciales para el proceso normal de digestión y absorción. Su pH siempre es alcalino por la presencia de HCO3- y su osmolaridad es constante y ronda los 300 mOs / L, siendo 150 mOs / L de los 300 la concentración de Cl- y de HCO3-. Los acinos pancreáticos están modulados por la acción de la colecistoquinina, la cual estimulará con mayor o menor intensidad los acinos según sea alta o baja (respectivamente) la cantidad proteica y lipídica de la comida ingerida. Por otro lado, los conductos están modulados por la acción de la secretina, la cual se activa si el pH a nivel del duodeno es inferior a 5 generando que los conductos secreten gran cantidad de HCO3- para tamponar el pH y evitar la desactivación de los enzimas digestivos. Secreción gástrica (3) La mucosa gástrica no tiene una distribución homogénea, sino que presenta diferentes áreas. La localización de estas áreas (mucosa aglandular, mucosa cardiaca o fúndica, mucosa gástrica, y mucosa pilórica o antral) varía entre especies. El gran tamaño de la mucosa aglandular del estómago de cerdos y caballos (en comparación con el perro) permite que los enzimas de la saliva tengan larga actividad predigestiva de algunos carbohidratos estructurales (fibra) de las plantas antes de ser degradadas en contacto con el HCl. En el píloro, además, se producen contracciones para degradar el alimento que llega. La secreción gástrica tiene varios componentes. Los más importantes son el HCl y el factor intrínseco secretados (a la vez) por las células parietales o oxínticas encontradas en la mucosa cardiaca, el pepsinógeno inactivo (precursor de la pepsina activa) secretado por las células principales, y el moco gástrico compuesto principalmente por mucina y HCO3- y secretado por las células caliciformes intercaladas entre las células gástricas de la mucosa pilórica. Las glucoproteínas (carbohidratos ramificados) tienen la función de retrasar la digestión de proteínas uniéndose a ellas, protegiéndolas de los ácidos y permitiendo su paso hacia el intestino delgado. El factor intrínseco es un tipo de glucoproteína que absorbe la vitamina B12 de la luz gástrica y la protege hasta llegar al íleon donde será absorbida y transportada al plasma; otros ejemplos de glucoproteínas son las inmunoglobulinas A y la membrana celular intestinal con glucocálix. Las cirugías gástricas pueden bloquear la acción captadora de vitamina B12 del factor intrínseco y provocar anemias. La barrera gástrica es un concepto que permite entender por qué el HCl junto a la pepsina en condiciones normales no provocan úlceras; está compuesta por una parte extrínseca y otra parte intrínseca: la parte extrínseca de la barrera gástrica está formada por un moco resistente a la digestión ácida gracias a su alta concentración de HCO3-, el balance ácido-base del tejido, y el flujo sanguíneo; y la parte intrínseca de la barrera gástrica está formada por la membrana apical de las células del epitelio gástrico, las uniones estrechas entre estas células, la regulación del pH, y la elevada y rápida renovación de todas las células epiteliales (total renovación de células cada 3 días). Cuando se producen úlceras, éstas estimulan el nervio vago y hacen que el estómago responda inadecuadamente secretando aún más HCl y disminuyendo aún más el pH, fenómeno especialmente dañino cuando el estómago se encuentra vacío y no hay sustancias que puedan tamponar la acidez; la alta incidencia de úlceras en el pasado ha llevado a la necesidad de conocer los mecanismos de secreción de HCl y al desarrollo de fármacos para su control como es el caso del omeprazol, ya que antiguamente se separaba el cardias del estómago y se empalmaba con el yeyuno lo que generaba anemias por falta de factor intrínseco. Los mecanismos de control de la secreción ácida están generados bajo la interacción de varios mecanismos endocrinos y nerviosos. La secreción de HCl está inducida por unas células endocrinas con receptores de la distensión del estómago y a través del nervio vago informan de forma aferente al SNC sobre la distensión, y éste mismo responde (reflejo vago-vagal) a ella con el aumento de la secreción de acetilcolina, la cual aumenta la generación de ácidos en el estómago; por otro lado, la gastrina circulante en sangre estimulada por el contacto de las células con las proteínas del alimento estimula sobre las células de la mucosa fúndica la producción de ácido cuando se detecta el incremento de comida en el interior del estómago, y se realiza un feed-back negativo debido a que el ácido suprime la producción de gastrina; las células D son unas células secretoras de somatostatina, la cual inhibe la secreción de gastrina y HCl de sus células vecinas y que es estimulada cuando el pH estomacal es inferior a 2 (cuando no hay comida en su interior); en las células de la mucosa gástrica hay unos receptores histamínicos para las histaminas de los mastocitos, la cual suprime la liberación de somatostatina de las células D. Con el paso del tiempo, finalmente se han llegado a conocer los mecanismos intracelulares de secreción de HCl, lo que nos ha llevado a la obtención de fármacos que controlan de forma muy eficaz la secreción de HCl, uno de ellos es el omeprazol. La célula parietal obtiene los H+ que utiliza para fabricar HCl a partir de la hidrólisis del agua: los H+ obtenidos son expulsados en contra de gradiente hacia la luz estomacal utilizando una bomba K+/H+ que introducirá K+ en contra de gradiente; los OH- obtenidos se unirán a CO2 obteniéndose HCO3- que será expulsado hacia la sangre para que actúe como tampón. El Cl- utilizado para el ácido se obtiene directamente de la sangre y se expulsa al estómago de forma activa. El K+ utilizado para ser expulsado a favor de gradiente hacia la luz estomacal y que luego se utiliza para expulsar H+ mediante la K+/H+ ATPasa se obtiene mediante el uso de una Na+/K+ ATPasa de la membrana basal de las células que expulsará Na+ hacia la sangre y introducirá K+ hacia la célula. Finalmente, el Na+ utilizado para la entrada de K+ a partir de la Na+/K+ ATPasa es introducido en la célula de forma activa a partir de la absorción en el estómago. La función del omeprazol es bloquear la bomba K+/H+ y bloquear consecuentemente la secreción de ácido. Secreción pancreática y digestión de hidratos de carbono y de proteínas (4) En el intestino no se produce ninguna secreción a excepción de la expulsión de agua y electrolitos (pero no cuenta como secreción) que se utiliza como mecanismo defensivo para la eliminación de bacterias; la única función de los enterocitos es la de absorber y terminar de digerir algunos elementos de la dieta, pero no la de secretar enzimas digestivos hacia la luz intestinal. Por lo tanto, las únicas secreciones que circulan por el intestino vienen producidas por el páncreas, el cual secreta gránulos de zimógeno donde se acumulan enzimas digestores de la gran mayoría de sustancias (grasas, hidratos de carbono y proteínas) que contiene la dieta. Sin embargo, la mucosa intestinal utiliza sus bordes con forma de cepillo de su superficie para mantener entre las vellosidades a las enzimas. Como ya se ha explicado anteriormente, el 70% de la secreción pancreática exocrina está controlada por el reflejo vago-vagal. Los mecanorreceptores y quimiorreceptores del intestino detectan el tipo y la cantidad de alimento que circula por el intestino, iniciando el reflejo vago-vagal que responde secretando acetilcolina que activa la secreción de enzimas por parte del páncreas; por otro lado, las células S miden el pH del duodeno y secretan secretina cuando el pH es inferior a 5, estimulando la secreción de HCO3- por parte de las células del conducto pancreático lo que tampona la acidez obteniéndose un pH neutro ligeramente alcalino para mantener activas las proteïnas; finalmente, hay un tipo de células endocrinas que se estimulan a causa de los componentes de la dieta (ácidos grasos provenientes de lípidos y aminoácidos provenientes de proteínas) provocando que secreten colecistoquinina, una hormona que estimula los acinos y su secreción. Los elementos (sobretodo enzimas) secretados en las diferentes especies van en función de su tipo de dieta: Origen Glándulas salivales Enzima Activador α-amilasa salival Cl (única en humanos) Substrato Almidón Estómago Pepsinas (pepsinógenos) HCl Proteínas y polipéptidos Páncreas exocrino Tripsina (tripsinógeno) Enteropeptidasa Proteínas y polipéptidos Mucosa intestinal Citoplasma de las células de la mucosa Quimotripsinas Tripsina (quimotripsinógenos) Proteínas y polipéptidos Elastasa (proelastasa) Tripsina Elastina, algunas otras proteínas Carboxipeptidasa A Tripsina (procarboxipeptidasa A) Proteínas y polipéptidos Carboxipeptidasa B Tripsina (procarboxipeptidasa B) Proteínas y polipéptidos Lipasa pancreática … Triglicéridos Esterasa pancreática … α-amilasa pancreática Ribonucleasa Desoxirribonucleasa Fosfolipasa A (profosfolipasa A) Enteropeptidasa Aminopeptidasas Cl- Esteroles colesterol Almidón … … Tripsina RNA DNA Lecitina … … Tripsinógeno Polipéptidos Dipeptidasas Maltasa … … Lactasa Sacarasa α-destrinasa limitante Nucleasa y enzimas relacionadas Diversas peptidasas … … … Dipéptidos Maltosa, maltotriosa Lactosa Sacarosa α-dextrinas limitantes Ácidos nucleicos Di-, tri- y tetra- péptidas … … del Función o productos Hidroliza los enlaces α1,4 produciendo α-dextrinas limitantes, maltotriosa y maltosa Rompen los enlaces peptídicos adyacentes a los aminoácidos aromáticos Rompen los enlaces peptídicos adyacentes a la arginina o la lisina Rompen las uniones peptídicas adyacentes a los aminoácidos con cadenas aromáticas Rompe los enlaces adyacentes a los aminoácidos con cadenas alifáticas laterales Separa los carboxiaminoácidos terminales con cadenas laterales aromáticas o alifáticas ramificadas Separa los carboxiaminoácidos terminales con cadenas laterales básicas Monoglicéridos y ácidos grasos Colesterol Igual que la salival Nucleótidos Nucleótidos Lisolecitina α-amilasa Tripsina Separa el aminoácido Nterminal del péptido Dos aminoácidos Glucosa Galactosa y glucosa Fructosa y glucosa Glucosa Pentosas y bases púricas y pirimidínicas Aminoácidos Las enzimas que degradan proteínas (proteasas) se secretan de forma inactiva, mientras que las enzimas sin actividad proteolítica se secretan de forma activa. Todas las enzimas se acumulan en el interior de los grános de zimógeno del páncreas, y posteriormente (las inactivas) se activan cuando llegan a la luz intestinal y contactan con la tripsina. La activación de las enzimas proteolíticas está regulada por 3 factores: la existencia del propio tripsinógeno, el inhibidor de tripsina que secretan las propias células y que se degrada en el intestino, y la falta de Ca2+ que hay en la luz del páncreas y que también es necesario para deshacer los gránulos de zimógeno contenedores de las enzimas. Un cúmulo de enzimas digestivas activas en el páncreas pueden desencadenar una pancreatitis, la cual provoca la degeneración del órgano y el paso de los enzimas a la sangre; además, los islotes endocrinos también se verán afectados, lo que provocará que no se produzca insulina y la consecuente diabetes y pancreatitis crónica. Organización funcional del intestino (4.1) El intestino delgado es una unidad funcional con la función de digerir y absorber agua, electrolitos y principios inmediatos; se separa en tres áreas anatómicas: · Duodeno: Zona de reequilibrio de soluciones gástricas, de vertido de secreciones digestivas y de escasa absorción · Yeyuno: Zona de digestión enzimàtica y de mayor absorción (100% de los lípidos) · Íleon: Zona de absorción de vitaminas y sales biliares (para su reciclaje) que antes no estaban disponibles debido a la protección que les otorgaban las proteínas; el transporte se realiza mediante transportadores específicos. Digestión de carbohidratos (4.2) La α-amilasa pancreática tiene la función de hidrolizar los enlaces α1,4 produciendo αdextrinas limitantes, maltotriosa y maltosa; sin embargo, grandes glúcidos como el almidón y el glucógeno le presentan limitaciones por ser glúcidos ramificados. Para ofrecer ayuda a la α-amilasa, el intestino realiza contracciones que colaboran en la rotura de los grandes glúcidos para obtener sacáridos pequeños y fáciles de digerir por la α -amilasa y las otras enzimas que se encuentran en la superficie (borde en cepillo) de los enterocitos. El conjunto de enzimas consiguen transformar los carbohidratos en azúcares (monosacáridos). Una vez digeridos los azúcares podrán ser absorbidos por los enterocitos, los cuales utilizan transportadores activos específicos para introducir la glucosa y la galactosa (SGLT1) y transportadores pasivos para introducir la fructosa (GLUT5). Digestión de proteínas (4.3) La digestión enzimàtica de proteínas se produce en tres lugares diferentes: · La digestión gástrica es dependiente de la pepsina, una enzima pH-dependiente (se activa a pH inferior a 3) que rompe los enlaces peptídicos adyacentes a los aminoácidos aromáticos y provoca una disgregación de la proteína. · La digestión intestinal luminar es dependiente de enzimas proteolíticos pancreáticos, unos enzimas que digieren enzimáticamente los restos de proteína y obtienen un 30% de aminoácidos libres y un 70% de oligopéptidos de 2 a 6 aminoácidos. Estas enzimas se secretan en forma de pro-enzimas (inactivas) y se activan en la luz por la acción de la enteroquinasa que transforma el tripsinógeno en tripsina, el cual realiza un feedback positivo sobre la enteroquinasa. · La digestión en el borde en cepillo es dependiente de oligopeptidasas de los enterocitos, unos enzimas que digieren enzimáticamente los restos y obtienen aminoácidos libres y oligopéptidos de 2 o 3 aminoácidos. La absorción de proteínas se realiza mediante unas proteínas de transporte inespecíficas de muy pequeños péptidos que permite su absorción cuando no se produce su completa digestión, y mediante unas proteínas activas específicas para según cual sea el tipo de aminoácido: básico, neutro o ácido. Los pequeños péptidos serán degradados en el interior de la célula por peptidasas intracelulares y junto al resto de aminoácidos libres se transportarán hacia los capilares sanguíneos a través de un transportador activo. Secreción biliar y digestión de las grasas (5) La secreción biliar o bilis está compuesta por agua en su gran mayoría, sales biliares (o ácidos biliares, pero es más correcto sales debido a hay mas Na+ que H+), bilirrubina conjugada para ser mas soluble y polar, colesterol cuya cantidad depende del tipo de dieta, lecitina (o fosfatidilcolina) que actúa como emulsionante y como componente de las membranas celulares y un conjunto de electrolitos que se encuentran en concentraciones parecidas a las del plasma (Na+ y Cl- en grandes cantidades, K+ y Ca+ en bajas cantidades, y HCO3-. Esta secreción biliar es constantemente expulsada por el hígado con un flujo constante (diferente entre especies). Las especies que disponen de vesícula biliar la utilizan para almacenar esta bilis secretada y poder expulsarla por el conducto colédoco de forma rápida; la expulsión a través del conducto colédoco está regulada bajo la actividad de la colecistoquinina secretada por el intestino delgado (sobretodo duodeno) cuando la cantidad lipídica y proteica de los alimentos es elevada, aumentando la secreción de bilis. Esta hormona permite sincronizar la secreción pancreática con la secreción biliar. Los ácidos biliares son los componentes de la bilis que tienen función digestiva. El precursor de éstos es el colesterol, a partir del cual se obtienen los ácidos biliares primarios, que en su mayoría son el ácido cólico y el ácido quenodeoxicólico, los cuales han ganado polaridad y orientación al sufrir una hidroxilación en diversos puntos de la molécula. Estos ácidos acompañarán a los alimentos a lo largo del intestino y en el íleon se reabsorberán y volverán al hígado vía portal para proceder a su posterior reutilización; sin embargo, el paso del tiempo puede llevar a que éstos sufran deshidroxilaciones por su larga utilización, lo cual los convierte en ácidos (secundarios, terciarios…) no tan polares, ni tan fáciles de reabsorber ni con tanta capacidad de emulsión (como el ácido deoxicólico obtenido por la deshidroxilación del ácido cólico, o el ácido licocólico obtenido por la deshidroxilación del ácido quenodeoxicólico). En la bilis, por lo tanto, podremos encontrar gran diversidad de ácidos, desde los recién creados a partir del colesterol como los reutilizados pocas y muchas veces. Los ácidos biliares son secretados de forma conjugada con aminoácidos para aumentar la polaridad de los ácidos, facilitar su orientación y consecuentemente facilitar el transporte hacia la luz; estos aminoácidos serán la taurina o la glicina, los cuales se encuentran en diferentes proporciones según la especie y su dieta. Al llegar a la luz intestinal, éstos se desconjugan para realizar su actividad mientras que los aminoácidos serán reabsorbidos o serán captados por las bacterias intestinales. El contacto de los ácidos biliares con las grasas procedentes de la dieta genera unas micelas biliares donde los ácidos biliares junto a los fosfolípidos (con la cabeza polar hacia el exterior) rodean los cúmulos de grasas. Estas grasas o lípidos dietéticos son compuestos caracterizados por su baja o nula solubilidad en el agua, por lo que es necesario polarizarlos con los ácidos biliares; los lípidos dietéticos se introducen en forma de triglicéridos, ésteres de colesterol, fosfolípidos o vitaminas liposolubles también esterificadas (K, A, D y E). La función pues, de la emulsión por parte de los ácidos biliares es la de producir gotillas de grasa emulsionada (10.000 Å) a partir de la separación de los glóbulos líquidos de grasa producidos en el estómago por la acción del calentamiento y del mezclado con el HCl y la pepsina, y así aumentar la superficie de ataque para las lipasas, co-lipasas y componentes biliares dando lugar a la formación de micelas (40 Å) más pequeñas y más estables. En el yeyuno, la difusión de las micelas a través de la capa de agua no agitada permite el transporte directo de la mayoría de los componentes de la micela, con excepción de los ácidos biliares, hacia el interior de los enterocitos. En el íleon, las proteínas de co-transporte especializado para el Na+ son las responsables de la absorción de los ácidos biliares. Las gotillas de grasa emulsionadas tienen una membrana formada por ácidos biliares y fosfolípidos, contienen en su interior lípidos desorientados. La función de las enzimas es adherirse a estas gotillas de grasa para degradarla, sin embargo hay poca superficie de ataque por lo que tendrán el máximo de actividad cuando las gotillas se conviertan en micelas. Las enzimas pancreáticas que actúan sobre las gotillas y las micelas son las lipasas que actúan sobre los triglicéridos para romper el enlace de los extremos y obtener 2 ácidos grasos y 1 monoglicerol; las fosfolipasas que actúan sobre las lecitinas (fosfolípido) para romper el enlace central y obtener 1 ácido graso y 1 lisolecitina; y las esterasas que actúan sobre los ésteres de colesterol para degradar el enlace ester y obtener una molécula de colesterol y 1 ácido graso. Finalmente, los lípidos son absorbidos e incorporados a los enterocitos de forma sencilla a través de la membrana apical de la célula. Una vez en el interior, los lípidos sufren los procesos inversos a los que habían sufrido previamente para ser absorbidos, es decir, se forman de nuevo triglicéridos, ésteres de colesterol y fosfolípidos que serán incorporados en unas estructuras de muy baja densidad (mucha grasa y poca proteína) llamadas quilomicrones, mientras que el 95% de los ácidos biliares se reincorporarán más adelante, en el íleon, siendo incorporados en la circulación enterohepática y reutilizados por el hígado para ahorrar colesterol. Los quilomicrones son unas partículas de lipoproteínas formadas por triglicéridos en su interior (80-90%), colesterol en la membrana y ésteres de colesterol en el interior (2%), fosfolípidos en el exterior (89%), vitaminas liposolubles y apolipoproteínas en el exterior (2%). Estos quilomicrones se ensamblan en el retículo endoplasmático rugoso y el aparato de Golgi de los enterocitos; tienen un tamaño de unos 750 – 5000 Å y se expulsan por exocitosis a través de la membrana basolateral. Debido a su gran tamaño, no caben por las fenestraciones de los capilares sanguíneos, por lo que penetran en los capilares linfáticos y llegan hasta la vena cava sin pasar por la vena porta ni el hígado. Finalmente se acumularán en el tejido adiposo y de ellos se extraerán los lípidos necesarios para el organismo. Absorción de agua y electrolitos. Bases fisiológicas de las diarrea (6) El flujo y movimiento de agua y electrolitos se produce de forma bidireccional, es decir, en ambos sentidos. Se produce una absorción y una secreción de éstos generando, sin embargo, un balance neto positivo de absorción a lo largo del intestino; este movimiento bidireccional es utilizada para equilibrar el agua y los electrolitos a los niveles fisiológicos adecuados. La absorción de la mayor parte de agua y solutos (que además van acompañados de agua) se produce en el intestino delgado (66%), seguido del intestino grueso (18%) y finalmente el colon (13%), por lo que las heces tienen un contenido hídrico realmente bajo (3%); sin embargo, en la parte proximal del intestino grueso aún hay agua y encontramos bacterias supervivientes. El factor determinante de los flujos de agua es la tonicidad del contenido intestinal, ya que ésta debe ser de unos 300 mOsmolar constantemente. Cuando la presión osmótica de los alimentos ingeridos es alta (como alimentos muy salados), se debe secretar agua para equilibrar la presión osmótica, lo que genera la sensación de sed y la necesidad de beber agua; cuando la presión osmótica después de absorber los electrolitos es baja, no se requiere secretar agua y pero sí se debe absorber para equilibrar la tonicidad y evitar el descenso de la tonicidad luminal. Cuando en la luz intestinal se encuentra una solución hipotónica se absorbe velozmente el agua para aumentar la tonicidad; cuando en la luz se encuentra una solución hipertónica se secreta agua hacia la luz para rebajar la tonicidad, sin embargo luego se reabsorbe junto a los electrolitos cuando éstos son absorbidos; sin embargo, cuando en la luz intestinal se encuentran sustancias no ionizables, no fraccionales o no absorbibles (como los cationes divalentes), el agua secretada no puede ser reabsorbida, lo que genera trastornos diarreicos; estas sustancias purgantes deben ser ingeridas junto a la cantidad de agua suficiente como para no formar una solución hipertónica de sales y no provocar la secreción de agua por parte del organismo. En el intestino delgado se produce la mayor absorción de solutos utilizando un co-transporte que aprovecha las (muy) bajas concentraciones de Na+ de la célula intestinal (entericito). Durante el co-transporte, el Na+ se mueve siguiendo su gradiente electroquímico. El gradiente favorable para el movimiento del Na+ se mantiene por la acción continua de la bomba Na+/K+ y la permeabilidad para K+ y Na+ de las uniones densas entre los enterocitos, generando una corriente constante de Na+. Durante la absorción, la diferencia de concentración del soluto a absorber (como por ejemplo glucosa) parece que se vuelve desfavorable para el transporte, pero la absorción continúa debido al gradiente de Na+. Además, con cada soluto y Na+ que entran en la célula también entran moléculas de agua. En el intestino delgado la mayor o menor absorción de agua genera un contenido isotónico respecto a la sangre debido a la corriente de entrada y salida de Na+; sin embargo, este contenido se reajusta (disminuye) en el intestino grueso debido a la absorción de agua, ya que no quedan (no deberían quedar) solutos por absorber, y al impedimento de salida de Na+ por parte de las uniones densas entre los enterocitos que son impermeables, por lo que las concentraciones de Na+ serán bajas al estar absorbiéndose constantemente y las de K+ serán altas al ser secretado por la célula constantemente. En las células hay receptores para la aldosterona y para la ADH que controlan los niveles de recuperación de agua. En el intestino grueso, además, se encuentran diferencias entre las células de las criptas y las células de las vellosidades. Las células de las criptas tienen muy poca capacidad de absorción, mientras que las células de las criptas son las encargadas de ello de forma tan estricta que su mecanismo no se ve afectado ni por procesos diarreicos. La secreción de agua y electrolitos en las criptas se efectúa debido a la secreción de Cl- por los canales de Cl- de las membranas apicales de los enterocitos de las criptas. El Na+ se mueve hacia la luz por la vía paracelular y equilibra eléctricamente la secreción de Cl-. El agua lo sigue osmóticamente y el efecto neto es el de la secreción de una solución de NaCl hacia el interior de la luz de la cripta. En las criptas, los mecanismos para la absorción acoplada de NaCl parece que existen en la membrana basolateral con compuertas de Cl- las cuales se hallan presentes en la membrana apical. La apertura de las compuertas de Cl- en las membranas de las células apicales presentes en las criptas inicia la secreción en la cripta. La posición de la membrana de los procesos para el transporte acoplado Na+/Cl- se ponen en reversa por sí mismos, moviéndose desde la membrana basolateral hacia la membrana apical a medida que las células maduran y se mueven hacia arriba por los vellos. Son los mecanismos endógenos los que favorecen el inicio de los mecanismos intracelulares que generan la abertura de los canales de Cl- en las criptas intestinales. La enterotoxina producida por E. coli estimula la abertura de estos canales de Cl- en las criptas y la consecuente secreción de agua junto al Cl-, lo que genera deshidratación en el organismo. Las diarreas en lechones y terneros provocadas por estas enterotoxinas son fáciles de producir por la baja cantidad de defensas; un tratamiento efectivo es el compense la secreción de agua aumentando su absorción en el intestino delgado, uno de los cuales pueden ser los preparados de cereales, el almidón de los cuales genera poca carga osmótica, sin embargo su destrucción va ofreciendo gran cantidad de glucosas que se absorben junto al agua. Microorganismos en el tracto digestivo (7) El tracto digestivo está poblado por gran cantidad de microorganismos que en conjunto forman la llamada microbiota del aparato digestivo, y entre ellos hay protozoos, bacterias… Éstos con el organismo establecen una relación de simbiosis. Según la especie animal de la que se trate, sus tres compartimentos (estómago, intestino delgado e intestino grueso) actuarán de diferente forma: · Carnívoros: Al igual que en el resto de especies, el intestino delgado tiene la carga bacteriana más baja debido a la gran cantidad de secreciones y movimientos que impiden su proliferación; este hecho es muy importante porque es necesario que los nutrientes sean absorbidos por el organismo y no por las bacterias. La zona proximal del estómago, donde no llegan las secreciones de HCl proliferarán las bacterias que realizarán pequeñas (pero no muy significativas) fermentaciones. En el intestino grueso sólo se encontrarán bacterias que fermentarán los pocos compuestos que queden sin digerir por parte del intestino delgado, por lo que tienen un efecto limitado. · Omnívoros (tipo cerdo): Las bacterias del intestino delgado son pocas y ocurre lo mismo que en carnívoros. En la zona proximal del estómago, donde no llega la acidez estomacal, las bacterias realizan el inicio de la digestión de la celulosa (carbohidrato estructural), un compuesto muy difícil de digerir, y de forma diferente según el tiempo que permanezca allí el alimento y el tamaño del estómago. En el intestino grueso las bacterias continúan degradando la celulosa y sacando provecho de ella debido a que el intestino delgado no ha sido capaz de absorberla, al igual que las células que quedan intactas después de la masticación; estas bacterias crecerán y producirán a través de esos compuestos ácidos grasos volátiles que se difundirán pasivamente a través de las membranas celulares. · Herbívoros: Las bacterias del intestino delgado son pocas y ocurre lo mismo que en carnívoros y omnívoros. · Típico (tipo caballo): Tiene un intestino grueso especialmente adaptado para que las bacterias tengan el tiempo y el tamaño necesario para digerir completamente la celulosa y obtener energía para su propio metabolismo, a través del cual crearán gran cantidad de ácidos grasos volátiles que los enterocitos tendrán la oportunidad de absorber adecuadamente. · Lagomorfo (tipo conejo): Su intestino grueso no está demasiado adaptado a contener bacterias que digieran la gran cantidad de celulosa que ingiere, sino que son especies especializadas en realizar cecotrofia, es decir, la liberación en horas nocturnas de masas fecales envueltas de moco por parte del ciego, y su inmediata ingesta (sin llegar a caer al suelo) que le permitirá realizar una segunda vuelta y darle al estómago una segunda oportunidad para digerir los carbohidratos estructurales. · Rumiantes: Tienen un intestino grueso desarrollado pero no tanto como los caballos, por lo que la zona encargada de mantener la mayor cantidad de bacterias es el estómago proximal (retículo, rumen y omaso, sobretodo rumen); esta grandiosidad en cantidades, crecimientos, desarrollos y tipos de microorganismos le dan la capacidad a los rumiantes de digerir el 70% de la celulosa (carbohidrato estructural por excelencia), el almidón (carbohidrato no estructural), las proteínas… mediante fermentación en la que se obtienen ácidos grasos volátiles. En el rumen se obtienen ácidos grasos volátiles, CH4 (metano) y CO2 de los carbohidratos estructurales, ácidos grasos volátiles, CH4 y CO2 de los carbohidratos no estructurales, y ácidos grasos volátiles y NH3 de las proteínas; en el abomaso e intestino delgado se obtienen monosacáridos de los carbohidratos no estructurales, aminoácidos de las proteínas, y ácidos grasos de los lípidos; y en el ciego y el colon se obtienen ácidos grasos volátiles, CH4 y CO2 de los carbohidratos estructurales, ácidos grasos volátiles, CH4 y CO2 de los carbohidratos no estructurales, y ácidos grasos volátiles y NH3 de las proteínas. En el rumen hay gran cantidad de especies bacterianas que se diferencian según el tipo de sustrato al que fermentan. Las más importantes son las bacterias celulolíticas, bacterias con enzimas celulasas en la pared celular que les permitirá digerir la celulosa y poner a disposición de otras células no celulolíticas ciertos compuestos útiles. La digestión (fermentación) de carbohidratos siempre dará CH4, CO2 y los ácidos grasos más volátiles (de cadena más corta): el ácido acético, el butírico y el propiónico. Las bacterias fermentarán los carbohidratos no estructurales (almidón) y estructurales (celulosa) de la misma forma, sin embargo, los primeros serán mucho más rápidos de digerir. Los ácidos grasos volátiles en su forma asociada al H+ (forma más apolar) se absorberán en el rumen por difusión; y los pequeños gases como el CO2 y el CH4 se expulsarán mediante eructos. Los aminoácidos obtenidos por la digestión de proteínas serán secuestrados por las bacterias, las cuales fabricarán a partir de ellos todo tipo de aminoácidos y al ser digeridas aportarán al rumiante todos los aminoácidos necesarios. En esta gráfica se puede observar la digestión de los elementos de la alfalfa a lo largo del tiempo: los carbohidratos solubles como el almidón se digieren totalmente a una alta velocidad por la gran capacidad de las bacterias; la pectina (hemicelulosa), un compuesto con los mismos enlaces que la celulosa pero con una mayor solubilidad, es rápida de digerir totalmente pero tiene requiere más tiempo y energía por parte de las bacterias; y la celulosa (carbohidrato estructural) requiere de una actividad duradera por parte de las bacterias para conseguir ser degradada en parte (no totalmente). La microbiota de los organismos se adaptarán al tipo de dieta que ingiera el individuo, por lo que predominarán células celulolíticas si la solubilidad de la dieta es baja, y predominarán otro tipo de células si la solubilidad es alta. Cuando la actividad celulolítica es alta debido a una dieta poco soluble, el pH del rumen aumenta debido a que los ácidos grasos volátiles recogen H+ del medio para aumentar su solubilidad y ser absorbidos por los enterocitos; cuando la dieta es muy soluble, la falta de ácidos grasos volátiles obtenidos por fermentación celulolítica provoca que no se adopten H+ y no se pueda aumentar el pH, por lo que la acidez será lo suficientemente elevada para que los Lactobacillus tengan la capacidad de producir ácido láctico, lo cual supone un problema. Las proteínas obtenidas de la dieta serán degradadas a péptidos y aminoácidos por parte, en parte, de las bacterias gástricas; las proteínas provenientes de la saliva también se descompondrán y proporcionarán y urea y otros compuestos nitrogenados como el NH3. Todos los compuestos serán adoptados por las bacterias y protozoos que los utilizarán para que más tarde lleguen al omaso y a partir de ahí sean digeridos y recuperados todos los compuestos. El ciclo del nitrógeno entre los órganos en los rumiantes se produce según muestra este diagrama. Muestra los efectos de la concentración de amoniaco en el rumen sobre la formación y la utilización de urea. Cuando la concentración de amoniaco en el rumen se encuentra elevada, entonces el movimiento neto del nitrógeno no proteico se realiza hacia el hígado, dando lugar a velocidades altas en al producción de urea y a una muy baja conservación del nitrógeno. Cuando las concentraciones de amoniaco en el rumen son bajas, el movimiento neto de nitrógeno no proteico se realiza desde el hígado hacia el rumen, dando lugar a la producción de proteínas a partir de la urea endógena. La eficiencia con la cual la energía de la dieta se utiliza para la síntesis de proteínas en el rumen depende del equilibrio que existe entre las fuentes de energía y de nitrógeno. La proporción de energía utilizada para la síntesis de proteína y para el mantenimiento celular cambia la relación que existe en el equilibrio de los aportes de péptidos (nitrógeno) y de glucosa. Motilidad digestiva: Deglución y llenado gástrico (8) La motilidad digestiva de todo el tracto intestinal, es decir, desde el esófago hasta el colon, está producida por músculo estriado esquelético y por músculo liso, cada uno de los cuales está presente en una zona determinada y en mayor o menor medida según la especie. El músculo estriado se encuentra únicamente en los extremos del tracto digestivo, es decir, en la faringe, en el esfínter esofágico superior y en una parte (más o menos grande según la especie) del esófago; el músculo liso se encuentra en el resto del tracto digestivo, es decir, en una parte (más o menos grande según la especie) del esófago, en el esfínter esofágico inferior, en el estómago… El músculo estriado está regulado por neuronas somáticas, sin embargo el músculo liso está inervado por el SNA entérico. Este sistema está formado por neuronas eferentes que detectan la distensión de los compartimentos digestivos, y por neuronas aferentes intrínsecas o extrínsecas que se rigen bajo el control del SNA parasimpático (utilizando acetilcolina como neurotransmisor) y de hormonas. El 70% de las motoneuronas que inervan el músculo liso son inhibitorias y utilizan, mientras que el 30% son excitatorias. Las motoneuronas inhibitorias utilizan NO, ATP y el Péptido Intestinal Vasoactivo (VIP) como neurotransmisores que provocan una hiperpolarización en las células musculares y su consecuente relajación; las motoneuronas excitatorias utilizan acetilcolina (de efecto rápido y corto) y la Sustancia P (de efecto lento y sostenido) como neurotransmisores que provocan una despolarización en las células musculares y su consecuente contracción. Los esfínteres esofágicos, el superior controlado por el SNC y el inferior controlado por el SNE, son utilizados para prevenir el flujo de sustancias cuando no es el momento; el superior previene la aerofagia, es decir, la introducción de aire hacia el estómago al respirar, y el inferior previene el reflujo del ácido proveniente del estómago y que dañaría constantemente el esófago. Estos esfínteres se encuentran basalmente contraídos, por lo que lo siempre tienen un mínimo tono muscular que impide el paso de sustancias de un lado al otro. La relajación de ambos esfínteres se induce a través del reflejo de deglución, el cual además, provoca una onda peristáltica primaria a lo largo del esófago asociadas a la deglución. El reflejo de deglución provoca un aumento de presión en la faringe y la consecuente generación de unas ondas de relajación-contracción en el esfínter esofágico superior que permiten el paso del bolo alimenticio al esófago; el bolo alimenticio ayudado por las ondas peristálticas (y la gravedad en caso de animales verticalizados) recorre el esófago en sentido distal y se dirige hacia el estómago; el descenso de presión detectada a lo largo del esófago después de pasar el bolo alimenticio provoca que el esfínter esofágico inferior se relaje y deje pasar el bolo alimenticio, lo que generará seguidamente un aumento de presión. Además, el esófago dispone de un mecanismo que garantiza el paso del contenido alimenticio hacia el estómago; este mecanismo actúa mediante la generación de una onda peristáltica secundaria a lo largo del esófago cuando los receptores de distensión detectan una lentitud en el paso del bolo alimenticio y envían esta información vía vagal eferente hacia un ganglio nervioso del plexo mientérico, el cual responde generando una onda peristáltica secundaria. Existen enfermedades como la Achalasia, en la que se producen alteraciones en la deglución a causa de la degeneración de los ganglios del SNE y la consecuente degeneración de las motoneuronas inhibitorias (la mayoría de veces), la cual cosa genera una coordinación de la onda peristáltica haciendo que todo el esófago se contraiga a la vez (efecto rebote del tono excitatorio) en vez de contraerse por partes para hacer avanzar el bolo alimenticio; consecuentemente el bolo alimenticio no avanzará. El fallo de las motoneuronas inhibitorias también produce que el esfínter esofágico inferior pueda relajarse, abrirse y dejar circular el bolo alimenticio hacia el estómago. El tono inhibitorio está determinado por la actividad de las monoteuronas inhibitorias. Este tono inhibitorio es esencial para conseguir mantener relajado fisiológicamente el tracto intestinal, el cual se mantiene así hasta que el bolo alimenticio le obliga a contraerse para permitir su circulación; la excepción en el tracto digestivo de esta situación es la de los esfínteres, los cuales están constantemente contraídos hasta que el bolo alimenticio provoca la activación de las células inhibitorias y la consecuente relajación de ellos. El estómago tiene una estructura histológica igual al resto del tracto digestivo, y dispone de varias zonas funcionales según los patrones motores y según los patrones secretores. Según los patrones motores el estómago dispone de una región proximal receptiva de alimento y una región distal con actividad mecánica intensa y disgregación del alimento; según los patrones secretores el estómago dispone del fundus donde se produce la secreción mucosa, el cuerpo donde se encuentra la mucosa oxíntica secretora de HCl, y el antro pilórico donde se produce secreción mucosa y hay actividad endocrina. En el estómago, la motilidad es distinta según el área determinada, por lo que la motilidad del fundus, es decir, la parte más proximal del estómago, es independiente a la parte distal. Cuando el fundus se encuentra vacío, las paredes se retraen y contactan las unas con las otras, manteniendo así una presión estomacal y poco superior a la atmosférica; cuando se produce la ingesta y la circulación por el esófago de alimentos, las paredes del estómago se relajan (disminución de la presión) para permitir la entrada del bolo alimenticio a su interior y seguidamente se contrae para adaptarse al bolo alimenticio y mantener la presión estomacal relativamente constante. El tono inhibitorio es el encargado de relajar el fundus a medida que entran los alimentos ingeridos, y luego el SNE lo contraen para adaptar el tamaño al bolo alimenticio. El llenado gástrico se produce por un reflejo asociado a la llegada de alimento y generado en la región proximal del estómago, donde hay presencia de tono muscular pero no de sin motilidad. Este reflejo implica una relajación muscular asociada al proceso de deglución para facilitar el paso del bolo del esófago al estómago, y una acomodación al aumento de volumen minimizando los cambios de presión. El mantenimiento de la presión intragástrica es un reflejo vago-vagal, ya que experimentos con animales vagotomizados demuestran que la presión gástrica de éstos aumenta sin parar a medida que aumenta el volumen de alimentos en su interior, mientras que los animales normales mantienen una presión gástrica más o menos constante. Motilidad digestiva: vaciamiento gástrico (9) En la parte proximal del estómago (fundus), la única actividad que realizan las motoneuronas es producir la variación del tono muscular para adaptarse a la llegada de alimentos y mantener la presión gástrica constante; sin embargo, en la parte distal (cuerpo y antro), los músculos inervados por las motoneuronas se encargan de favorecer la destrucción del alimento mediante una actividad mecánica intensa para obtener partículas de hasta 2 mm de diámetro, la medida óptima para pasar por el píloro. El desarrollo muscular del antro es más importante en algunas especies como los humanos o lo perros, que en otras como los rumiantes. El píloro es el esfínter situado al final del estómago por donde deben circular los alimentos para llegar al duodeno; tiene un tono muscular basal que lo mantiene casi cerrado y ofrece resistencia al paso del bolo alimenticio aunque el estómago sufra contracciones. Las contracciones gástricas provocan que los alimentos choquen repetidamente contra el píloro y retrocedan de nuevo para volver a chocar contra el píloro, generando un ataque mecánico contra el bolo alimenticio; además, los ácidos secretados por el estómago facilitan la degradación del alimento, generando un ataque químico; estos ataques degradan el alimento y lo preparan para circular por el intestino. La motilidad del cuerpo y el antro pilórico está producida por contracciones fásicas que generan los llamados movimientos peristálticos, los cuales empujan el contenido alimenticio contra el píloro. La frecuencia de las contracciones es especie-específica, en el perro se producen 4 o 5 cada minuto. Estas contracciones se originan en la parte media del cuerpo gástrico, y se propagan en dirección aboral (distal) con cada vez mas potencia debido a la coordinación que hay en la masa muscular debido a que las fibras musculares están unidas por sincitios. Estos músculos se contraen mediante ondas lentas. Constantemente sus células están hiperpolarizadas por la acción del tono inhibitorio, pero cuando éste disminuye, la acumulación de ondas pueden llegar al umbral de descarga y generar la contracción del músculo. El número máximo de contracciones que estos músculos es igual a la frecuencia de las ondas lentas, así las contracciones se producen de forma ordenada y el músculo no se tetaniza. Cualquier parte del músculo puede convertirse en marcapasos gracias al sincitio; las células marcapasos por excelencia serán las que tengan una frecuencia mas alta de ondas lentas, ya que la primera generará la despolarización que se va propagando por todas las fibras de manera ordenada y coordinada. La onda lenta del píloro es muy grande, por lo que cuando le llega la despolarización la mantendrá durante largo tiempo y se contraerá permaneciendo así un largo periodo. Como hemos dicho antes, la actividad motora gástrica genera el empuje del contenido gástrico hacia el píloro debido a las fuertes contracciones que sufren el antro y el esfínter pilórico (desde oral hasta distal). Las moléculas menos de 2 mm serán la parte del contenido gástrica que pasará al duodeno, mientras que el resto sufrirán una retropropulsión hacia el cuerpo. Las consecuencias funcionales de este proceso son la mezcla de los alimentos, la trituración mecánica y el vaciamiento gástrico. El vaciamiento gástrico se produce por orden de facilidad, es decir, primero se vacía de líquidos, ya que los sólidos necesitan romperse hasta los 2 mm; luego se vacía de los sólidos menos pesados, es decir, los sólidos más atacables por las enzimas; finalmente conseguirá vaciarse de los sólidos más pesados, los cuales permanecen mucho tiempo en el estómago y ralentizan el vaciamiento gástrico. Los objetos indigestibles como el plástico, el metal… se podrán expulsar del estómago cuando éste esté vacío por completo y se produzca una motilidad de limpieza (y no digestiva), en la que el píloro se abre considerablemente. El control del vaciamiento gástrico se produce mediante un control hormonal y un control nervioso que pueden originarse en diferentes lugares: en el estómago o en el intestino. El control gástrico mediante la distensión de las paredes (lo cual hace secretar acetilcolina procedente del reflejo vagovagal, y gastrina) y las proteínas digeridas (lo cual hace secretar gastrina) aceleran el vaciado gástrico; el control intestinal mediante un pH duodenal bajo (lo cual hace secretar secretina), una solución hipertónica (lo cual provoca un reflejo hormonal), una digestión grasa (lo cual hace secretar colecistoquinina, neurotensina y GIP), la llegada de L-triptófano (lo cual hace secretar colecistoquinina), y la distensión o irritación del duodeno (lo cual hace activar el reflejo vago-vagal y las fibras inhibitorias locales) disminuyen el vaciado gástrico. La distensión a nivel del estómago provoca un aumento de la excitabilidad en el músculo gástrico y consecuentemente un aumento de la velocidad de vaciado. Según que estímulo generen los alimentos en el intestino se provocará la secreción de diferentes hormonas que tienen efecto inhibitorio sobre las fibras aferentes nerviosas que modulan los reflejos nerviosos intrínsecos vago-vagales que actúan sobre el estómago; el aumento de nutrientes en el intestino provoca un ralentizamiento del vaciado para así poder amortizar las enzimas de las que se disponen en el intestino. Estímulo Control Gástrico Distensión Proteínas digeridas Control intestinal Bajo pH duodenal Solución hipertónica Digestión grasa L-triptófano Distensión o irritación del duodeno Mediador Efecto en el vaciado Acetilcolina (reflejo vago-vagal) y gastrina Gastrina Aumento Aumento Secretina Reflejo hormonal Colecistoquinina, neurotensina, GIP Colecistoquinina Reflejo vago-vagal y fibras locales inhibitorias Disminución Disminución Disminución Disminución Disminución Otro mecanismo regulador del vaciamiento gástrico es el llamado ileal brake, el cual se produce cuando hay presencia de nutrientes en el ileon, lo que genera que el ileon libere hormonas como neurotensina y active vías nerviosas que disminuyan el vaciamiento gástrico debido a que se calcula que en el duodeno y el yeyuno no está dando tiempo a realizar la absorción de estos nutrientes. Este mecanismo sigue siendo estudiado. Motilidad del intestino delgado (10) La motilidad del intestino delgado está provocada por varios tipos de motilidad con diferentes patrones, los cuales están regulados por los ganglios intrínsecos que pertenecen al sistema nervioso entérico; el nervio vago y el SNA simpático únicamente modulan, no regulan. Motilidad interdigestiva (10.1) La motilidad interdigestiva se produce en ayunas y tiene la función de mantener la luz intestinal libre de bacterias. Es independiente al sistema nervioso central. Las despolarizaciones de las células (debido a las ondas lentas) provocan contracciones al azar de segmentos del intestino, las cuales generan movimientos de segmentación. Sin embargo, también forma parte la motilidad interdigestiva unas despolarizaciones secuenciales que generan de forma coordinada la contracción del músculo a lo largo de todo el intestino. Movimiento de segmentación (10.2) El movimiento de segmentación que se produce por las contracciones del músculo en cada segmento del intestino, está regulado por el tono inhibitorio durante el ayuno, el cual mantiene activamente el intestino en relajación. Sin embargo, en determinados puntos y durante cortos periodos de tiempo, estas células inhibitorias se inactivan y permiten la contracción del músculo, lo que genera un movimiento que produce la mezcla de enzimas con el contenido residual intestinal para favorecer su descomposición. Motilidad digestiva (10.3) Esta motilidad postprandial está presente desde la ingestión del alimento hasta el final de la digestión intestinal, con el paso del contenido al colon. Se regula en función de la cantidad de alimentos que haya en el lugar y el momento concreto dentro del intestino. La despolarización de estas fibras se mantiene largo tiempo y genera una actividad continua de ellas, las cuales generan movimientos peristálticos. Movimiento peristáltico (10.4) Es el otro tipo de movimientos producidos en el intestino; su función es propulsar el contenido intestinal en sentido aboral. El estímulo local que lo provoca viene dado por la distensión generada por el bolo alimenticio en el intestino (estímulo mecánico) y el contenido luminal de nutrientes (estímulo químico); este estímulo local actúa únicamente en la zona donde se encuentra el bolo alimenticio y no se transmite al resto del órgano, sino que es el propio bolo quien va generando movimiento tras movimiento. Esta motilidad coordinada de tipo propulsivo es dependiente del SNE y en ella participan las capas musculares del intestino (circular y longitudinal). Su consecuencia funcional es la propulsión del contenido intestinal hacia el colon; están generados y controlados por el plexo mientérico; el estímulo desencadenante es la distensión local del intestino; las contracciones generan que el bolo alimenticio recorra segmentos limitados del intestino (1-4 cm). Como ya hemos dicho (perdón por repetirme), el movimiento peristáltico se inicia con la distensión intestinal del segmento donde se encuentre el bolo alimenticio, la cual provoca una estimulación en los mecanorreceptores. Esta distensión será detectada por los receptores de distensión de las capas musculares y generará que las neuronas intrínsecas aferentes provoquen la inactivación de las neuronas inhibitorias de la zona oral (anterior) y consecuentemente la contracción, produciendo un estrechamiento de la luz por contracción de la capa muscular circular; además, generará que las neuronas intrínsecas aferentes provoquen un aumento de activación de las neuronas inhibitorias de la zona aboral (posterior) y consecuentemente la relajación, produciendo un aumento en el diámetro luminal por contracción de la capa muscular longitudinal. Es un movimiento un poco impredecible. Las neuronas colinérgicas tienen una actividad secundaria. Patrón de ondas lentas intestinal (10.5) Las ondas lentas se transmiten a lo largo del intestino a partir de la primera que las ha provocado, sin embargo, su frecuencia desciende a lo largo del intestino debido a la (siempre inevitable) descoordinación que se produce en los sincitios. Las ondas lentas de un sincitio tomarán como célula marcapasos a la célula con mayor frecuencia de ellas, la cual en este caso son las células del duodeno, ya que las ondas lentas se iniciar a partir del píloro (consecuentemente las células con menor frecuencia de ondas lentas son las del íleon). Sin embargo, in vivo, el retraso de ondas lentas que se produce en el sincitio genera que haya diversas células marcapasos a lo largo del intestino, las cuales tienen una frecuencia de ondas lentas mayor por si solas que la que le correspondería si fuera una más del sincitio. A lo largo del intestino podemos encontrar 3 o 4 células marcapasos. La frecuencia de las ondas lentas determinará la frecuencia de las contracciones intestinales. Complejo motor migratorio (10.6) El complejo motor migratorio es consecuencia de la intensa actividad a lo largo del intestino que hay en ayunas. Este complejo se produce cada 2 horas aproximadamente, y no se inicia uno hasta que no se terminado el anterior. El complejo motor migratorio tiene la función de limpiar el intestino de bacterias. Se compone de 3 fases: una primera fase de quiescencia en la que no hay actividad motora y no se producen contracciones porque las ondas lentas no generan potenciales de acción; una segunda fase en la que se producen contracciones irregulares o aleatorias por una actividad motora aislada en la que algunas ondas lentas generan potenciales de acción para mezclar el contenido intestinal; y una tercera fase llamada frente de actividad en la que hay una actividad regular e intensa que se propaga a lo largo del intestino en sentido aboral y con todas las ondas lentas generando potenciales de acción, produciendo una función de limpieza del intestino arrastrando el contenido residual del periodo interdigestivo hacia el colon. El segmento que acaba de realizar la tercera fase, entrará en la primera fase durante un periodo hasta que entre de nuevo en la segunda fase; estas dos fases se irán alternando; a las 2 horas de haber pasado por la tercera fase, ésta volverá a producirse. Este complejo motor migratorio se encuentra inhibido durante la ingesta de alimentos por la acción de la colecistoquinina. El complejo moto migratorio está regulado por las neuronas del plexo mientérico. Este plexo está formado por una cadena de neuronas que dirigen sus señales en sentido aboral hasta el íleon; su función es inhibir las neuronas inhibitorias, produciendo así la contracción muscular del intestino. Allí donde estén inhibiendo localmente a esas neuronas se estará produciendo el frente de actividad. La primera de ellas es la que se convierte en el marcapasos de la cadena. Según la especie animal, el complejo motor migratorio se interrumpe durante la digestión o no. Las especies monocavitarias de omnívoros (perros, cerdos, humanos) y herbívoros (ratones, caballos) sufrirán una interrupción postprandial variable del complejo motor migratorio; mientras que las especies policavitarias de herbívoros (rumiantes) y aves no sufrirán ninguna iterrupción y el complejo motor migratorio estará siempre presente. Unión ileocecocólica (10.7) La unión ileocecocólica está cerrada siempre por el llamado esfínter ileocecocólico, el cual tiene la función de permanecer cerrado y evitar el paso de bacterias desde el colon hacia el íleon. Únicamente se abre cuando hay presión en el lado del íleon y se detecta una distensión para permitir la salida del bolo alimenticio (lo que queda de él) hacia el colon. Cuando ingerimos alimentos, la gastrina y la colecistoquinina asociadas a la llegada del nuevo alimento al estómago inducen la abertura de la unión ileocecocólica y el vaciamiento completo del intestino para preparar un nuevo proceso digestivo. Motilidad del intestino grueso (11) El intestino grueso tiene la función de recuperar durante el tiempo que permanezca el bolo en su interior, la máxima cantidad de agua y electrolitos posible; además, según la especie, en él también se produce cierta digestión bacteriana. Frecuencia de ondas lentas (11.1) Las ondas lentas del colon se originan, como siempre, en el lugar con mayor frecuencia de ellas. Esta localización, donde se concentran las células marcapasos, se sitúa en la parte medio-distal del colon, por lo que es aquí donde se generará el inicio de la motilidad intestinal. El recto por su parte, es relativamente independiente de estas ondas lentas, ya que su sensibilidad es mucho mayor y actúa autónomamente cuando entra un mínimo de contenido en él, con la función de inducir la excreción de heces (defecación). Motilidad del intestino grueso (11.2) Esta motilidad asociada a las ondas lentas del intestino grueso con origen intermedio tiene una propagación oral y aboral, en ambos sentidos, por lo que se producen movimientos en masa (peristálticos) propulsivos en la zona aboral que empujan el contenido hacia zonas distales favoreciendo la defecación, y movimientos retroperistálticos en la zona oral que empujan el contenido hacia zonas proximales y permiten el llenado del ciego favoreciendo la retención del contenido colónico, su mezcla, su fermentación por digestión bacteriana y la absorción de agua y electrolitos. También en el intestino grueso se producen aquellos movimientos de segmentación, llamados en esta zona movimientos haustrales. Son movimientos de segmentación producidos en el colon proximal y el ciego que requieren más fuerza y energía que los del intestino delgado, ya que el contenido no es líquido, sino que cada vez se va haciendo más sólido; estos movimientos son, además, más lentos y afectan a segmentos más largos, para así poder mezclar el contenido, favorecer el proceso de digestión bacteriana y favorecer la absorción de agua. En vacas, donde las heces son inmensamente más grandes que las de las ovejas, necesitan movimientos en masa más constantes y grandes. El tipo de dieta influye en la motilidad del colon. Esto se vio en un experimento realizado en cerdos a los cuales les dieron primeramente comida normal (concentrados) o leche, y en el que se observó como con su comida normal se producían más movimientos en masa (con propagación) que haustrales, y sin embargo, cuando la dieta era leche se producían más movimientos haustrales (localizados) que en masa; este aumento de actividad de los movimientos haustrales (no fisiológicamente sanos) provocan un aumento de la absorción de agua y un consecuente estreñimiento. Sin embargo, cuando a la dieta de la leche le añadimos fibra, se puede observar como aumentan los movimientos en masa y disminuyen en proporción los movimientos haustrales, disminuyendo entonces el riesgo de estreñimiento. El intestino de los seres humanos está acostumbrado a una dieta más rica en fibra, con mayor cantidad de movimientos en masa que los que tenemos ahora; en nuestra sociedad cagamos poco, deberíamos cagar más. La motilidad del colon se ha observado que está relacionada con los ritmos circadianos y las comidas. Durante las horas de actividad sin ingestión, el colon tiene una actividad basal en la que se producen los movimientos haustrales antes vistos; durante las ingestas grandes (comida y cena), la actividad aumenta y se producen grandes movimientos en masa, los cuales aceleran las ganas de ir a defecar; sin embargo, durante las hora de sueño, la actividad basal desciende aún más y queda anulada durante ese periodo de tiempo. Defecación (11.3) El recto siempre debe encontrarse fisiológicamente vacío. Cuando se produce la llegada de elementos a su luz provenientes del colon y éstos generan una mínima distensión, se produce en consecuencia la activación de los movimientos peristálticos de la pared rectal, la relajación (y apertura) del esfínter anal interno (estimulado por el SNA parasimpático) y la posterior defecación. Sin embargo, esto puede no suceder así porque los animales disponemos del esfínter anal externo, el cual tiene control voluntario y permite impedir el paso de las heces eventualmente y así defecar en el sitio más adecuado para tu supervivencia. Particularidades de la función digestiva en las aves (12) Las aves utilizadas en granjas son granívoras, es decir, se alimentan únicamente de granos vegetales; entre éstas encontraríamos las gallinas y los pavos. Unas de las características de la función gástrica en aves domésticas son su peculiar ingesta de alimento de forma continua, y consecuentemente su motilidad gástrica continua que se produce bajo el llamado ciclo gastro-duodenal que les permite reciclar el alimento y llevarlo de nuevo al estómago una vez está en el duodeno; además, otra característica es su falta de dientes y sus reflujos duodenogástricos fisiológicos regulados por la colecitoquinina. Anatomía aviar (12.1) Debido a su adaptación para volar, las aves tienen un aparato digestivo muy pequeño con respecto al de los mamíferos, y es por ese motivo que han tenido que adaptarse y realizar según que funciones (que para otros animales no serían fisiológicas como el ciclo gastro-duodenal) para hacerlo igual de eficiente. Al igual que en los mamíferos, podemos separar el aparato digestivo en tres partes: la parte gástrica proximal donde se almacenan los alimentos, la parte del intestino delgado donde se digieren y se absorben los alimentos, y la parte del intestino grueso donde se recuperan líquidos y se forman las heces. En el esófago encontramos una estructura (2) llamado buche, que es un divertículo esofágico agrandable donde se almacenan alimentos para irlos expulsando poco a poco hacia el estómago; este divertículo se relaja durante la deglución y se va adaptando al contenido. Ya en el estómago, encontramos dos compartimentos. El compartimento glandular (proximal) o proventrículo donde se secreta pepsina y HCl; y el compartimento muscular (distal) o molleja donde se acumulan piedras ingeridos que luego favorecen la digestión y la trituración de los alimentos. En este último compartimento encontramos una musculatura muy desarrollada formada por dos músculos delgados y dos de gruesos, los cuales se comportan como un ventrículo; su contracción se produce de forma alternada entre gruesos y delgados coordinadamente. Las contracciones realizadas por los músculos gruesos pueden llegar a provocar lesiones en su interior, por lo que intercaladas podemos encontrar unas glándulas que secretan mucopolisacáridos que se endurecen al estar en contacto con el pH ácido del estómago, generándole protección. El intestino delgado, al igual que en los mamíferos está formado por el duodeno, el yeyuno y el íleon. En este caso, el duodeno es desproporcionalmente más grande que el resto; sobre el duodeno, se van drenando las secreciones de los diferentes conductos pancreáticos y del colédoco. A lo largo del intestino delgado podemos encontrar un resto de lo que fue el conducto (16) que unía al embrión con el saco vitelino. Ciclo gastroduodenal (12.2) El ciclo gastroduodenal se inicia en los músculos delgados de la molleja; debido a esta contracción, el contenido gástrico que sea inferior a los 2 mm de diámetro llegará al duodeno, por lo que allí se producirán seguidamente unas ondas peristálticas; seguidamente y a causa de la actividad de los músculos delgados, se produce la contracción de los músculos gruesos, los cuales actuarán sobre el contenido sólido del estómago para triturarlo y expulsarán los líquidos rápidamente; finalmente el estómago glandular será el que realice las últimas contracciones del ciclo. En el duodeno, al igual que en el de los mamíferos, también podemos registrar diferentes tipos de motilidad. El ciclo anterior muestra una motilidad preistáltica, en la que los alimentos van en dirección aboral. Sin embargo, las aves disponen de una motilidad antiperistáltica fisiológica, en la que realizan reflujos duodeno-gástricos controlados por la colecitoquinina que devuelven el contenido intestinal al estómago, sobretodo cuando hay nutrientes difíciles de absorber (como lípidos). Este mecanismo permite alargar el tiempo que dura el tránsito intestinal y así aumentar la eficiencia en ese espacio tan reducido del que disponen las aves para su aparato digestivo. Función intestinal (12.3) El intestino presenta complejos motores migratorios sin interrupción, e incluso en las horas de sueño se siguen produciendo movimiento antiperistálticos para digerir los alimentos. Estos animales, a diferencia de los mamíferos presentan un intestino corto, por lo que el proceso digestivo es más rápido que en los mamíferos. Otra peculiaridad es que los vasos linfáticos no actúan en esta especie como transportadores de quilomicrones, sino que éstos no llegan a formarse y son los VLDL los que se absorben y se distribuyen por vía sanguínea. Función del intestino grueso en aves (12.4) Las aves presentan dos ciegos que contactan con el colon (o recto, hay discusión sobre como llamarlo) a través de la unión ileocecocólica (17 en el dibujo anterior), por la cual entran muy limitadamente los sólidos (únicamente los componentes solubles de la fibra vegetal) pero no los líquidos. En estos ciegos se favorece la fermentación de estos componentes solubles gracias a la población bacteriana que le corresponde y que le proporcionan al vitaminas y elementos, aunque no de forma muy significativa. La función principal del recto y ciegos es favorecer la recuperación de agua procedente de la orina; esta orina está compuesta principalmente por uratos (ácido úrico), el cual es muy poco soluble y es necesario que sedimente y precipite sobre los restos (heces) que se expulsan por el intestino. Los ciegos tienen movimientos antiperistálticos frecuentes y contracciones tónicas; el vaciamiento de éstos está sujeto a un ritmo circadiano, por lo que la expulsión de heces a partir del colon se realiza sobretodo en las horas de sueño. [Mensaje para los que aprovecháis estos apuntes: Gracias a todos los que a lo largo del semestre han agradecido mi trabajo (a pesar de la gran cantidad de errores que he ido detectando, os pido perdón por ellos), que sepáis que los he hecho con toda mi buena intención de ayudarme primeramente a mí, y ya de paso poder ayudar a los demás; la verdad es que han sido bastantes horas de arduo trabajo y que para hacerlos he sudado mucho…así que si insistís en invitarme a una cerveza el día que salgamos de este examen para que no me muera deshidratado no me importará aceptárosla en absoluto, y es más, os lo agradeceré a muerte. Gracias. Alejandro]