Distribución de Erycina crista-galli - CIIDIR Unidad Oaxaca
Anuncio

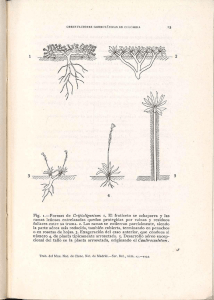
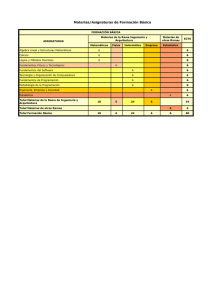
vol. 5 núm. 1 Naturaleza y Desarrollo enero - junio 2007 Distribución de Erycina crista-galli (Orchidaceae) sobre arbustos de café Citlalli Maldonado-Mijangos y Demetria Mondragón-Chaparro Laboratorio de Epífitas, CIDIR Unidad Oaxaca IPN, Hornos 1003, Col. Noche Buena, Santa Cruz Xoxocotlàn, C.P. 71230, Oaxaca, México. Tel: 01 951 70610, 70400 y 71199 Ext: 82743. E-mail: [email protected] Resumen Se evaluó el patrón de distribución espacial de 563 individuos Erycina crista-galli sobre 29 arbustos de café. Los cafetos fueron divididos en tres zonas en función de su altura (0 a 101 cm A1, 101.1 a 202 cm A2, > 202.1 cm A3) y del diámetro de sus ramas (0.22-1.79 cm D1, 1.80-3.36 cm D2, >3.37 cm D3); los individuos de E. crista-galli se clasificaron en cuatro estadios: plántulas, juveniles y adultos 1 y 2. Se determinó la abundancia y la estructura poblacional de E. crista-galli en cada zona. Se observó una baja correlación entre la altura de las ramas y la abundancia de E. crista-galli (r=0.4029, P=0.5744), y una alta correlación con el diámetro de las ramas (r= -0.8609, P= -0.0217). Las estructuras poblacionales en las zonas D1 y D2 fueron semejantes (G= 0.1086, P=0.05), difiriendo de D3 en donde no se encontraron plántulas (G= -1.2838, P=0.05; G= -1.3925, P=0.05 respectivamente). El patrón de distribución de E. crista-galli está fuertemente determinado por la presencia de ramas delgadas, la cual se ve favorecida por las podas anuales que se le aplican a los arbustos de café. Palabras clave: epífitas de ramillas, abundancia, diámetro de ramas. Abstract An evaluation was made of the spatial distribution of 563 individuals of Erycina crista-galli on 29 coffee shrubs. Each coffee shrub was divided into three zones based on height and branch diameter. The individuals of E. cristagalli were classified in four stages: seedling, juvenile, adult 1 and adult 2. The abundance and population structure of E. crista-galli in each zone was determined. A low correlation was found between height and abundance of E. crista-galli (r=0.4029, P=0.5744), and a high correlation with branch diameter (r= -0.8609, P= -0.0217). The population structure in zones D1 and D2 were similar (G= 0.1086, P=0.05), and differed from D3, where no seedlings were found (G= -1.2838, P=0.05; G= -1.3925, P=0.05). The pattern of distribution of E. crista-galli was found to be strongly determined by the presence of thin branches, which is enhanced by cultural management on the coffee plantation. Key words: twig epiphytes, abundance, twig diameter. Introducción 2004), determinan el patrón de distribución vertical de las epífitas sobre el forofito. Diferentes factores intervienen en la definición del patrón de distribución de una especie. En el caso de las epífitas, dicho patrón está fuertemente influido por la arquitectura y ciertas características del forofito, como son: el sistema de ramificación, el diámetro y el ángulo de inclinación de las ramas, la textura y el pH de la corteza, la transparencia de la copa (cantidad de luz que llega a la parte interna de la copa en función de la densidad de follaje), el tamaño y la edad de las ramas (Ter Steege y Cornelissen, 1989; Rudolph et al., 1998; Zotz et al., 1999; Isaza et al., 2004). Dichas características en conjunto con las condiciones microambientales de luz, humedad y temperatura de las diferentes partes del forofito y los requerimientos ecofisiológicos de las epífitas (Bennett, 1988; Zotz, 1997; Zotz et al., 1999; Graham y Andrade, 2004; Jácome et al., En general, el patrón de distribución vertical de las epífitas puede ser explicado con base en las zonas en el forofito propuestas por Johansson (1974), quien subdividió a los árboles en cinco zonas en función de la composición de especies epífitas encontradas: parte basal de tronco (zona I), tronco (Zona II) y tres zonas de igual longitud en la copa (base de la copa: Zona III, parte media: Zona IV y parte exterior: zona V) (Bøgh, 1992). Sin embargo, esta zonificación varía en función del tipo de vegetación evaluada y la arquitectura del hospedero, las cuales alteran las condiciones microambientales en el forofito (Bøgh, 1992; Benzing, 1995; Zotz y Vollrath, 2003). 5 Citllali Maldonado-Mijangos y Demetria Mondragón-Chaparro Entre las epífitas, existe un pequeño grupo que está restringido a habitar las ramas más pequeñas y delgadas de los forofitos (zona V), en las cuales existen altas radiaciones solares y poca humedad (Johansson, 1974; Benzing, 1995). Este tipo de epífitas, conocidas como «epífitas de ramillas», al parecer presentan adaptaciones anatómicas y fisiológicas que les permiten sobrevivir en este ambiente, pero que al mismo tiempo, pudieran estar restringiendo su distribución en otras partes del árbol (Chase, 1987; Benzing, 1995). A pesar de que los estudios demuestran la presencia de este tipo de especies epifitas en la zona V (Chase, 1987; Benzing, 1995), no existen estudios que documenten su distribución espacial a lo largo de todo el forofito y menos aún aquellas que evalúen el cambio en la estructura poblacional en las diferentes partes del árbol. Es por ello que analizamos el patrón de distribución espacial de Erycina crista-galli, la que pudiera ser clasificada como epífita de ramilla (Chase, 1987), evaluando cómo varía su abundancia y su estructura poblacional en las diferentes zonas del forofito. fasciculadas; flores amarillas, sépalos agudos, pétalos obtusos, labelo lobulado provisto de manchas rojizas en el centro, cápsulas menores de tres cm de largo. La especie se distribuye en bosques húmedos desde México hasta Sudamérica y las Antillas (Ames y Correll, 1953; Conzatti, 1982; Hágsater y Salazar, 1990). Coffea arabica L. es una planta arbustiva menor que 2.5 m de altura. El sistema de ramificación de los arbustos de café es variable, comenzando generalmente a menos de 30 cm de altura; desde esta base pueden observarse más ramificaciones secundarias, terciarias, etc.- a lo largo de las ramas primarias. Presenta toda clase de ramas, principalmente delgadas distribuidas tanto en la periferia y en la parte interna del arbusto, como en las partes más altas y bajas de éste (Standley, 1926; obs. pers.). A fin de conocer la distribución espacial de Erycina cristagalli sobre arbustos de café, en febrero de 2004 se eligieron aleatoriamente 29 cafetos (Coffea arabica), sobre los cuales se marcaron todos los individuos presentes de la orquídea bajo estudio. Para cada orquídea marcada se midió el diámetro y la altura de la rama sobre la cual se encontraba posada. Para determinar si existía alguna relación entre el diámetro de las ramas y su altura, se realizó una prueba de correlación de Pearson (Zar, 1984). Materiales y métodos El estudio se llevo acabo en Santo Domingo, municipio de Unión Juárez, de Chiapas, México, entre 92°0621" longitud Oeste, y 15°01 48 latitud Norte, y a una altitud de 850 msnm. El clima es semicálido-húmedo AC(m), con una temperatura media anual de 27.8°C, lluvias periódicas en los meses de mayo a octubre y una precipitación total anual de 2,478 mm. Sto. Domingo cuenta una vegetación de selva alta perennifolia combinada con plantaciones de café de sombra (INEGI 2000). De acuerdo con la clasificación propuesta por Moguel y Toledo (1999) para los sistemas de producción de café en México, las plantaciones en la zona de estudio se clasifican como un sistema de producción «moderno» de monocultivo con sombra donde los árboles utilizados son especies del género Inga (Fabaceae); sin embargo, existen parcelas en donde además están presentes otras especies arbóreas lo que las clasificaría como policultivo comercial. Posteriormente se crearon categorías de rangos tanto de diámetro como de altura (cuadro 1), determinándose la abundancia de individuos de E. crista-galli presentes en cada una de ellas. Diferentes autores mencionan que la abundancia de individuos epífitos está influida por la disponibilidad de área colonizable (Yeaton y Gladstone, 1982; Rudolph et al., 1998; Tremblay et al., 1998). Con el propósito de calcular el área disponible por categoría de ramas se procedió a medir el diámetro y la longitud de ramas y troncos de cinco arbustos. Con las 15 áreas calculadas y el número de individuos de E. crista-galli localizados en dichas áreas, se realizó una prueba Erycina crista-galli (Rchb.f.) N.H.Williams y M.W.Chase, es una orquídea de menos de 10 cm de alto, con pseudobulbos menores a cuatro centímetros de alto, de hojas planas, Cuadro 1. Categorías de ramas del hospedero en función de los diámetros y alturas. Febrero de 2004, Sto. Domingo, Unión Juárez, Chiapas. Categoría por Intervalo de diámetros Categoría por Diámetros Intervalo de diámetros alturas D1 0.22-1.79 cm A1 0 a 101 cm D2 1.80-3.36 cm A2 101.1 a 202 cm D3 >3.37 cm A3 > 202.1 cm 6 vol. 5 núm. 1 Naturaleza y Desarrollo enero - junio 2007 de correlación entre el diámetro y las áreas de las diferentes categorías de ramas. por lo cual se decidió excluir a dicho parámetro del resto del estudio. Para evaluar si las abundancias de individuos de E. cristagalli presentes en las diferentes categorías de ramas diferían estadísticamente, se procedió a realizar un ANOVA de una sola vía, para lo cual se transformaron los valores de abundancia aplicando logaritmos naturales. Se aplicaron pruebas pareadas (LSD) para determinar cuáles categorías diferían entre sí (Zar, 1984). Los análisis de correlación entre la abundancia de E. cristagalli y el diámetro de las ramas fue estadísticamente significativo (r= -0.8609, P= -0.0217), mostrando una relación inversa entre el grosor de la rama y la abundancia de individuos. Dada la baja correlación entre el diámetro de la rama y el área colonizable de las mismas (r= -0.0157 P=0.575), se puede suponer que la abundancia de los individuos de E. crista-galli está efectivamente correlacionada con el diámetro de las ramas del forofito, lo cual se ve reforzado por la mayor abundancia de individuos en las ramitas del café (F= 48.578, 2, P< 0.001). Los individuos de E. crista-galli se clasificaron en cuatro estadios (cuadro 2) de acuerdo a características de tamaño y número de pseudobulbos, así como en la presencia/ausencia de estructuras reproductivas, con el fin de agrupar individuos con tasas demográficas similares entre ellos y diferentes a individuos pertenecientes a otros estadios. Posteriormente se calculó la proporción de individuos de E. crista-galli para cada estadio. Con estos datos se determinó la estructura poblacional para cada categoría de ramas, las cuales fueron comparadas a través de pruebas G para determinar si existían diferencias estadísticamente significativas (Zar, 1984). En cuanto a las estructuras poblacionales en D1 y D2, se observó que éstas fueron muy similares entre sí (G= 0.1086, P=0.45) (figura 1). Para ambas categorías, cabe resaltar la presencia tanto de plántulas y juveniles, como una gran cantidad de individuos adultos. Por su parte la estructura poblacional de D3, difirió estadísticamente de las de D1 y D2 (G= -1.2838, P=0.05; G= -1.3925, P=0.05, respectivamente), ya que en D3 no se observan plántulas y la proporción de juveniles en relación con las otras dos categorías fue mínima. Cuadro 2. Categorías y características de los estadios del ciclo de vida de O. crista galli. Febrero de 2004, Sto. Domingo, Unión Juárez, Chiapas. Estadio Características Plántulas (p) 1 pseudobulbo de 0.01-0.3 cm; 1 o 2 hojas presentes; estructuras reproductivas ausentes. Juveniles (j) 1 pseudobulbo de 0.31-0.4 cm; 1 o 2 hojas presentes; estructuras reproductivas ausentes. Adultos 1 (a1) 1 pseudobulbo >0.4 cm; con o sin hojas; estructuras reproductivas presentes. Adultos 2 (a2) Individuos con 2 o más pseudobulbos; con o sin hojas; con o sin estructuras reproductivas. Resultados Discusión Se marcaron 563 individuos de E. crista-galli los cuales se encontraron distribuidos de la siguiente manera: 479 en D1, 39 en D2 y 45 en D3. Mientras que en los rangos de altura fue de: 14 individuos en A1, 218 en A2 y 102 en A3. En varios estudios realizados para conocer la abundancia de las epífitas sobre sus forofitos, diversos autores sugieren que mientras más grande y/o viejo sea el forofito, éste proporciona una mayor área y un mayor tiempo para la colonización (Rudolph, et al. 1998; Zotz et al., 1999; Merwin et al., 2003; Zotz y Vollrath, 2003; Mondragón et al., 2004), lo cual da como resultado una mayor abundancia de individuos epífitos en las ramas o troncos más grandes y viejos (Yeaton y Gladstone, 1982; Zotz et al., 1999; Isaza et al., 2004). Considerando esto, se esperaría encontrar la mayor abundancia de individuos de E. crista-galli sobre los troncos y ramas gruesas principalmente, ya que éstos tienen mayor Al realizar la prueba de correlación, se observó relación negativa entre los diámetros y la altura de las ramas (r= 0.266), sin embargo esta no fue estadísticamente significativa (P=0.063), por lo que en el resto de los análisis las variables se analizaron independientemente. Dado que al aplicar la prueba de correlación entre el número de individuos y la altura de ramas, los valores de r resultaron menores de 0.08 (r=0.4029), además dicho valor no fue significativo estadísticamente (P=0.5744), se determinó que la altura no era un buen indicador de la abundancia de E. crista-galli, 7 promedio de cada estadio Proporción de individuos Citllali Maldonado-Mijangos y Demetria Mondragón-Chaparro 1 .4 1 .2 1 0 .8 0 .6 0 .4 0 .2 0 D1 D2 D3 C a te g o ria s d e ra m a s Figura 1. Estructuras poblacionales de E. crista-galli por categoría de ramas en arbustos de café. Plántulas columna negra, juveniles columna gris oscuro, adultos 1 columna gris claro, y adultos 2 columna blanca. X + 1 EE. Feb. 2004, Sto. Domingo, Chiapas. superficie de contacto y han estado expuestos durante más tiempo a la colonización; no obstante, los datos muestran que E. crista-galli presenta un patrón de distribución diferente sobre los arbustos de café, lo que se sugiere que no es necesariamente, la superficie disponible la que influye directamente en el patrón de distribución de esta especie. Johansson (1974) pueden cambiar o fusionarse dependiendo de la de la arquitectura de los forofitos dada la estructura irregular de algunos de ellos, lo que hace posible encontrar todo tipo de ramas en todas las zonas del árbol. Para que una población pueda mantenerse en un sitio, en este caso las ramillas, es necesario que los individuos puedan completar su ciclo de vida (Harper, 1977). En el caso de E. crista-galli, esto sólo puede llevarse a cabo en las ramas medianas y ramillas (D2 y D1), donde se pudieron encontrar individuos de todos los estadios del ciclo de vida de esta especie; no así en las ramas gruesas y tronco (D3), en donde la estructura poblacional estuvo compuesta básicamente por individuos adultos, los cuales probablemente se hayan establecido cuando los diámetros del tronco y de las ramas gruesas eran menores, permitiendo las condiciones propicias para el establecimiento de plántulas, o bien, puede ser el resultado de eventos esporádicos de reclutamiento, como lo sugiere la presencia de algunos juveniles en estas ramas. Existen pocos trabajos publicados que explican la distribución vertical de la estructura poblacional de las epífitas sobre los forofitos, como los realizados por Rudolph et al. (1998), Zotz y Vollrath (2003) y Mondragón et al. (2004). Este patrón de distribución vertical de los estadios del ciclo de vida de E. crista-galli difiere del reportado para Tillandsia brachycaulos Schltdl. (Bromeliaceae), una especie epífita no clasificada como de ramilla, en donde la mayor proporción de individuos adultos se localizó en la parte media del árbol, donde las ramas fueron clasificadas como D2 y D3. Esta distribución pudiera atribuirse a que este tipo de plantas, que se reproducen asexualmente, llegan a formar genets de Chase (1987) menciona que para las epífitas de ramilla, el diámetro de las ramas es el factor que determina el patrón de distribución vertical de este tipo de especies; éste parece ser el caso para E. crista-galli, por la alta correlación negativa entre la abundancia y el diámetro de las ramas. Por lo tanto, la arquitectura de los arbustos de café, mantenida mediante las labores de poda que los cafeticultores realizan anualmente con el fin de favorecer el crecimiento de nuevas ramas , propicia la presencia de «ramillas» a todo lo alto y ancho del hospedero. Este patrón de distribución vertical de E. crista-galli no concuerda con lo reportado por diversos autores, quienes señalan que la mayor abundancia de epífitas se encuentra en las zonas intermedias y/o altas (Yeaton y Gladstone, 1982; De Souza y Do Espírito-Santo, 2002; Castaño-Meneses et al., 2003; Isaza et al., 2004; Mondragón et al., 2004), zonas III y IV según la clasificación de Johansson (1974). Esta abundancia en las zonas medias la relacionan con una mayor disponibilidad de ramas colonizables y una mayor diversidad de microclimas y tipos de sustratos (Johansson, 1974; Bøgh, 1992; Benzing, 1995). Sin embargo, como hacen notar Bøgh (1992) y De Souza y Do Espírito-Santo (2002) , las zonas propuestas por 8 vol. 5 núm. 1 Naturaleza y Desarrollo enero - junio 2007 De Souza, M., y M. M. Do Espirito-Santo. 2002. Species diversity and abundance of vascular epiphytes on Vellozia piresiana in Brazil. Biotropica 34: 51-57. hasta 6 ramets, lo que hace que su peso sólo pueda ser soportado por ramas gruesas (Mondragón et al., 2004). Sin embargo, es posible encontrar ciertas similitudes en el patrón de distribución entre ambas especies: a) En T. brachycaulos, al igual que en E. crista-galli, las plántulas, individuos infantiles y juveniles se localizan en su mayoría en las ramillas (D1); A diferencia de E. crista-galli, no es raro encontrar estos estadios en la zona de ramas gruesas del árbol. b) La estructura poblacional de T. brachycaulos en el tronco es muy similar a la encontrada para E. crista-galli, ya que básicamente carece de plántulas (Mondragón et al., 2004). Graham, E. A., y J. L. Andrade. 2004. Drought tolerance associated with vertical stratification of two co-occurring epiphytic bromeliads in a tropical dry forest. American Journal of Botany 91(5): 699-106. Conclusiones Harper, J. L. 1977. Population Biology of Plants. Academic Press. Londres. 892 pp. El patrón de distribución de E. crista galli sobre los arbustos de café, esta fuertemente influenciada por las labores de poda que sobre los cafetos se realiza, la cual favorece la presencia de ramas delgadas a todo lo alto de los cafetos. Agradecimientos Agradecemos a la Dra. Anne Damon, a ECOSUR de Tapachula y al CONACYT (Clave de proyecto SEMARNAT2002-CO1-697) por su apoyo. Al Dr. José García Franco y a la Dra. Ivón Ramírez por sus valiosos comentarios. Literatura Citada Ames, O. y Correll, D. S. 1953. Orchids of Guatemala. Fieldiana: Botany, Vol. 26, No. 2. Chicago Natural History Museum. Bennett, B. C. 1988. A comparison of life history traits in selected epiphytic and saxicolous Tillandsia (Bromeliaceae) from Florida and Peru. Tesis doctoral. University of North Carolina at Chapel Hill. U. S. A. Benzing, D. H. 1995. The physical mosaic and plant variety in forest canopies. Selbyana 16: 159-168. Bøgh, A. 1992. Composition and distribution of the vascular epiphyte flora of an Ecuadorian montane rain forest. Selbyana 13: 25-34. Castaño-Meneses, G., J. G. García-Franco, y J. G. PalaciosVargas. 2003. Spatial distribution patterns of Tillandsia violacea (Bromeliaceae) and support trees in an altitudinal gradient from a temperate forest in Central Mexico. Selbyana 24: 71-77. Chase, M. W. 1987. Obligate twig epiphytism in the Oncidiinae and other neotropical orchids. Selbyana 10:2430. Conzatti, C. 1982. Flora Taxonómica Mexicana. Tomo II. Instituto Politécnico Nacional. México, D. F. Hágsater, E., y G. A. Salazar. 1990. Icones Orchidacearum. Fascicle I. Orchids of Mexico Part I. Asociación Mexicana de Orquideología, A. C. INEGI. 2000. Anuario estadístico. Chiapas. Instituto Nacional de Geografía e Informática. México, D. F. Isaza, C., J. Betancur, y J. V. Estévez-Varón. 2004. Vertical distribution of bromeliads in a montane forest in the eastern cordillera of the Colombian Andes. Selbyana 25: 126-137. Jácome, J., G. Galeano, M. Amaya, y M. Mora. 2004. Vertical distribution of epiphytic and hemiepiphytic Araceae in a tropical rain forest in Chocó, Colombia. Selbyana 25: 118-125. Johansson, D. 1974. Ecology of vascular epiphytes in West African rain forest. Acta Phytogeographica Suecica. Acta Phyto 59: 1-136. Merwin, M. C., S. A. Rentmeester, y N. M. Nadkarni. 2003. The influence of host tree species on the distribution of epiphytic bromeliads in experimental monospecific plantations, La Selva, Costa Rica. Biotropica 35: 37-47. Moguel, P., y V. M. Toledo, 1999. Biodiversity conservation in traditional coffee system of Mexico. Conservation Biology 13: 11-21. Mondragón, D. L. Calvo-Irabién, y D. H. Benzing. 2004. The basis for obligate epiphytism in Tillandsia brachycaulos (Bromeliaceae) in a Mexican tropical dry forest. Journal of Tropical Ecology 20: 97-104. Rudolph, D., G. Rauer, J. Nieder y W. Barthlott. 1998. Distributional patterns of epiphytes in the canopy and phorophyte characteristics in a Western Andean rain forest in Ecuador. Selbyana 19:27-33. Standley, P. C. 1926. Trees and shrubs of Mexico. Smithsonian Institution on United States National Museum Vol. 23 Part 5, 1387 pp. Ter Steege, H., y J. H. C. Cornelissen. 1989. Distribution and ecology of vascular epiphytes in lowland rain forest of Guyana. Biotropica 21: 331-339. 9 Citllali Maldonado-Mijangos y Demetria Mondragón-Chaparro Tremblay, R. L., J. K. Z, L. Lebrón, P. Bayman, I. Sastre, F. Axelrod, y J. Alers-García. 1998. Host specificity and low reproductive success in the rare endemic Puerto Rican orchid Lepanthes caritensis. Biological Conservation 85: 297-304. Yeaton, R. I. y D. E. Gladstone. 1982. The pattern of colonization of epiphytes on calabash trees (Crescentia alata HBK) in Guanacaste Province, Costa Rica. Biotropica 14: 137-140. Zar, J. H. 1984. Biostatistical Analysis. Prentice Hall. Englewood Cliffs. 697 pp. Zotz, G. 1997. Substrate use of three epiphytic bromeliads. Ecogeography 20:264-270. Zotz, G., P. Bermejo, y H. Dietz. 1999. The epiphyte vegetation of Annona glabra on Barro Colorado Island, Panama. Journal of Biogeography 26:761-776. Zotz, G., y B. Vollrath. 2003. The epiphyte vegetation of the palm Socratea exorrhiza: correlations with tree size, tree age and bryophyte cover. Journal of Tropical Ecology 19: 81-90. 10