Composición de especies y patrones de distribución de paralarvas
Anuncio

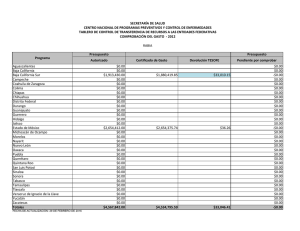
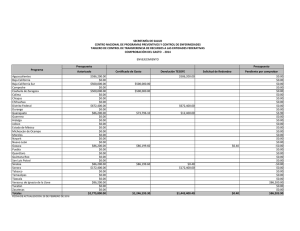
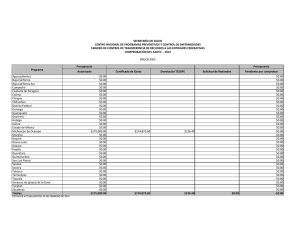
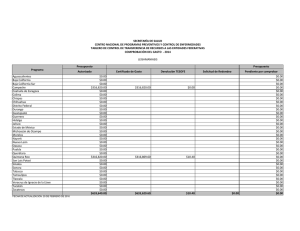
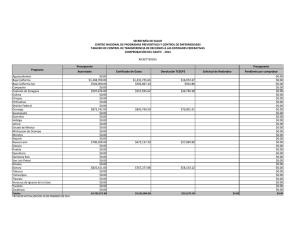
Composición de especies y patrones de distribución de paralarvas de calamar, 1996–1999 J Granados-Amores1, 2 , R De Silva-Dávila1*+ , S Camarillo-Coop1, 2, FG Hochberg3 , R Durazo4, R Avendaño-Ibarra1+, ME Hernández-Rivas1 Resumen Se encontró que la composición, abundancia y distribución de paralarvas de calamar registradas de 1996 a 1999 frente a la costa Pacífica de la Península de Baja California, México estuvo influenciada por las condiciones oceanográficas presentes durante El Niño Oscilación del Sur (ENOS) 1997–2001. Se recolectaron paralarvas durante siete cruceros oceanográficos mediante arrastres con una estructura Bongo estándar. Se identificaron 10 familias, 17 especies, 12 formas y dos complejos de especies. Las familias más importantes fueron Ommastrephidae, Onychoteuthidae, Cranchiidae y Gonatidae, que contribuye- Centro Interdisciplinario de Ciencias Marinas (CICIMAR-IPN). Departamento de Plancton y Ecología Marina. Apdo. Postal 592. La Paz, BCS 23000. [email protected], [email protected], [email protected], [email protected]. 2 Dirección actual: Centro de Investigaciones Biológicas del Noroeste (CIBNOR). Mar Bermejo # 195, Col. Playa Palo de Santa Rita. La Paz, BCS 23090. [email protected]. 3 Department of Invertebrate Zoology. Santa Barbara Museum of Natural History (SBMNH). 2559 Puesta del Sol Rd. Santa Barbara, California, USA, 93105-2936. [email protected]. 4 Universidad Autónoma de Baja California. Facultad de Ciencias Marinas. Carretera Tijuana-Ensenada km 107, Ensenada, Baja California. [email protected]. * Becarios COFAA y +EDI. 1 453 ron respectivamente con 72%, 10%, 7% y 6% de las paralarvas, totalizando un 95% del total colectado. Los taxones más abundantes fueron Dosidicus gigas, Sthenoteuthis oualaniensis y el complejo SD de la familia Ommastrephidae junto con los taxones de afinidad tropical-subtropical que se concentraron entre Punta Eugenia y Punta Abreojos durante El Niño 1997-1998, asociados a la masa de agua cálida Transicional-Subtropical Superficial. Durante La Niña 1999–2001 se registró un cambio en la composición de especies de calamar con el ingreso y predominancia de taxones de afinidad templada, principalmente de la familia Gonatidae que se distribuyeron en la masa de Agua Subártica. Palabras clave: paralarvas, cefalópodos, distribución, abundancia, ENOS 1997–2001. Abstract Species composition, abundance and distribution of squid paralarvae during 1996 to 1999 was found influenced by the oceanographic conditions of El Niño Southern Oscilation (ENOS) 1997–2001 off the Pacific coast of the Baja California Peninsula. Paralarvae were collected during seven oceanographic cruises by means of standard Bongo net tows. A total of 10 families, 17 species, 12 forms and 2 species complexes were identified. The Ommastrephidae, Onychoteuthidae, Cranchiidae, and Gonatidae families were the most important, contributing respectively 72%, 10%, 7%, and 6% paralarvae, accounting for 95% of all paralarvae identified. The most abundant taxa were Dosidicus gigas, Sthenoteuthis oualaniensis and the SD complex of the family Ommastrephidae, that were distributed between Punta Eugenia and Punta Abreojos in the cruises during El Niño 1997–1998, in association with the warm Transitional-Subtropical Surface Water. The remainder of the taxa of tropical-subtropical affinity showed the same distribution pattern. A change in species composition was recorded with the presence and predominance of the family Gonatidae of temperate affinity throughout the area in the cruises under the influence of La Niña 1999–2001, along with temperate taxa of other families that were distributed in the Subarctic Water mass. Keywords: paralarvae, cephalopod, distribution, abundance, ENOS 1997–2001. 454 Aspectos biológicos Introducción El análisis bibliográfico más reciente, que incluye registros en contenidos estomacales de peces depredadores de cefalópodos, registros de pesca exploratoria y de paralarvas, indica la presencia de 60 taxones de calamares distribuidos en aguas del Pacífico Mexicano (Granados-Amores 2008). De estos, 46 tienen registros en la costa occidental de la Península de Baja California y pertenecen a 18 familias de las cuales sólo la Loliginidae y Ommastrephidae tienen importancia comercial, particularmente el ommastréfido Dosidicus gigas (calamar Humboldt o jumbo del Pacífico). Con excepción de los estudios realizados sobre los adultos de esta especie en México (Morales-Bojórquez et al. 2001, Markaida 2006, Rodhouse et al. 2006), los antecedentes sobre el resto de la comunidad de cefalópodos en nuestro país son escasos. La falta de estudios sistemáticos sobre este conjunto de especies se atribuye tanto a la ausencia de un programa de muestreo como a la dificultad para capturar tanto a los adultos, sobre todo de especies de distribución meso y batipelágica, como a sus paralarvas planctónicas (Okutani y McGowan 1969). Las paralarvas registran generalmente poca abundancia alrededor del mundo (Haimovici et al. 2002), pero su distribución en los primeros 200 m de la columna de agua (Sweeney et al. 1992) las hace un excelente indicador de la riqueza de especies de un área determinada. El plancton en la costa occidental de la Península de Baja California ha sido muestreado sistemáticamente por diferentes instituciones. Los resultados de los muestreos indican una mezcla de faunas de diferente afinidad, asociadas a los flujos estacionales de las masas de agua Subártica, Central y Ecuatorial que conforman un ambiente variable donde es posible detectar el avance de los fenómenos de escala interanual como El Niño Oscilación Sur (ENOS) (Loeb et al. 1983, Lavaniegos et al. 2001, Palomares-García et al. 2003). En 1996 surge el interés por el análisis de las paralarvas de cefalópodos de la costa occidental de la península, abordándose de manera formal con el inicio del programa de Investigaciones Mexicanas de la Corriente de California (IMECOCAL). El esfuerzo inicial se ha enfocado hacia la taxonomía, enfatizando la relación entre taxones y masas de agua. Este esfuerzo ha contribuido con la incorporación de nuevos taxones al listado sistemático de cefalópodos en México y a la identificación de áreas de desove para algunas de las especies. Este trabajo resume los resultados sobre la composición de especies de calamar identificadas con base en sus paralarvas, así como sobre su distribución y abundancia obtenidas a partir del C omposición de especies y patrones de distribución de paralarvas de calamar 455 análisis de muestras de siete cruceros oceanográficos realizados frente a la costa occidental de la Península de Baja California, México. Metodología El material biológico provino de siete cruceros oceanográficos que cubrieron las zonas costera y oceánica frente a la Península de Baja California (tabla 1), el primero de los cuales (SIMSUP IX 9603) fue realizado en marzo de 1996 en tres áreas de surgencia: de Ensenada a Punta Baja, de Punta Eugenia a Bahía Asunción y en el Golfo de Ulloa. Las estaciones de este crucero cubrieron la plataforma continental, el talud y la zona oceánica adyacente en cada una de ellas (fig. 2a). Los seis cruceros oceanográficos restantes fueron realizados por IMECOCAL de 1997 a 1999 (cruceros IMECOCAL 9710, 9801, 9807, 9810, 9901 y 9908) siguiendo la red básica de estaciones de muestreo (ver fig. 1 de Durazo y Gaxiola-Castro, este volumen). Las muestras de zooplancton se obtuvieron realizando arrastres con una estructura Bongo siguiendo la metodología estándar de colecta (Smith y Richardson 1979). Aunque la metodología fue diseñada y utilizada originalmente para la Tabla 1. Relación de los siete cruceros oceanográficos realizados en la costa occidental de la Península de Baja California: crucero, temporada, fechas, número de estaciones muestreadas y abundancias bruta y estandarizada (PL/1000 m3) de paralarvas de calamar. Cruceros/Año Temporada Fecha Estaciones muestreadas Abundancia Bruta Muestreada SIMPSUP IX 1996 9603 Invierno 6−22/03 66 53 88 1997 9710 Otoño 28/09−10/10 39 16 43 1998 9801 Invierno 24/01−14/02 69 64 195 9807 Verano 15−30/07 65 182 1791 9810 Otoño 28/09−1/11 63 2 4 9901 Invierno 14−31/01 55 41 145 9908 Verano 8−22/08 79 63 285 421 2551 IMECOCAL 1999 Total 456 Aspectos biológicos prospección de huevos y larvas de peces, ésta ha resultado uno de los mejores métodos de recolecta para paralarvas de cefalópodos (Saito 1994, Granados-Amores 2008). Las paralarvas de calamar se separaron en su totalidad de las muestras originales y se identificaron siguiendo diferentes criterios establecidos (Sweeney et al. 1992, Diekmann et al. 2002). En algunos casos la identificación requirió de disección y teñido de los ejemplares y de la elaboración de preparaciones temporales de rádulas. De las muestras de otoño de 1997 y 1998 sólo se separaron e identificaron los especímenes de la familia Ommastrephidae. Las abundancias por taxón se estandarizaron a 1000 m3 de agua filtrada (Postel et al. 2000) y se elaboraron mapas de distribución de la abundancia para los 3 taxones de las 3 familias más abundantes por crucero utilizando el programa Surfer (Golden Software V 8.0). Con base en los valores de spiciness (π), Durazo y Baumgartner (2002) determinaron la presencia de Agua Subártica (π < 0.6) y Agua Transicional Subtropical Superficial (π > 1.5) frente a la Península de Baja California durante 1997–1998. Valores de π > 1 indican que el agua es más caliente y salada mientras que valores bajos de π indican lo contrario. El valor de π ~ 1 representa el límite entre el Agua Subártica y el Agua Transicional-Subtropical Superficial. Con estos resultados se elaboraron los mapas de distribución de las masas de agua mencionadas durante los cruceros realizados y se analizó la distribución de paralarvas asociada a la presencia de las masas de agua. Resultados El elenco sistemático de paralarvas de calamar registradas incluyó la presencia de 10 familias, 17 géneros, 17 especies, 12 formas y dos complejos de especies (tabla 2) que fueron ordenados con base en el Sistema de Información Taxonómica Integrada (ITIS por sus siglas en inglés, http://www.itis.usda.gov). Abundancia Las familias Ommastrephidae, Onychoteuthidae, Cranchiidae y Gonatidae fueron las más abundantes a lo largo del período de estudio, contribuyendo con el 72%, 10%, 7% y 6% respectivamente, para un total conjunto del 95% de las paralarvas de calamar en el área. Durante el crucero del invierno de 1996 las familias Onychoteuthidae y Loliginidae fueron las más abundantes (48% y 32%, respectivamente; fig. 1). La segunda no C omposición de especies y patrones de distribución de paralarvas de calamar 457 Tabla 2. Listado sistemático de paralarvas de calamar identificadas en el área de estudio (Sistema de Información Taxonómica Integrada, http://www.itis.usda.gov). PHYLUM MOLLUSCA CLASE CEPHALOPODA SUBCLASE COLEOIDEA SUPERORDEN DECAPODIFORMES ORDEN TEUTHIDA SUBORDEN MYOPSIDA FAMILIA LOLIGINIDAE Loligo opalenscens Berry, 1911 SUBORDEN OEGOPSIDA FAMILIA ANCISTROCHEIRIDAE Ancistrocheirus cf lesueuri (D´Orbigny, 1842) FAMILIA CRANCHIIDAE Cranchiidae no identificados Cranchiidae tipo 1 SUBFAMILIA CRANCHIINAE Cranchia scabra Leach, 1817 Leachia danae (Joubin, 1931) Leachia dislocata Young, 1972 Leachia pacifica (grupo) Leachia spp. SUBFAMILIA TAONIINAE Helicocranchia beebei Robson, 1948 FAMILIA ENOPLOTEUTHIDAE Abraliopsis felis McGowan y Okutani, 1968 Abraliopsis spp. Abraliopsis sp. 1 Abraliopsis sp. 2 FAMILIA GONATIDAE Gonatidae no identificados Gonatus sp. 1 Gonatus sp. 2 Gonatopsis borealis Sasaki, 1923 458 Aspectos biológicos Tabla 2. Continúa. FAMILIA HISTIOTEUTHIDAE Histioteuthis heteropsis (Berry, 1913) FAMILIA OCTOPOTEUTHIDAE Octopoteuthis deletron Young, 1972 FAMILIA OMMASTREPHIDAE SUBFAMILIA OMMASTREPHINAE Ommastrephidae no identificados Ommastrephes bartramii (Lesueur, 1821) Dosidicus gigas (D’Orbigny, 1835) Eucleoteuthis luminosa (Sasaki, 1915) Sthenoteuthis oualaniensis (Lesson, 1830) Hyaloteuthis pelagica (Bosc, 1802) Morfotipo A Morfotipo B Complejo EHO Complejo S-D FAMILIA ONYCHOTEUTHIDAE Onychoteuthidae no identificados Onychoteuthis banksi sp. 1 Onychoteuthis banksi sp. 2 Onychoteuthis banksi sp. 3 Onychoteuthis borealijaponica Okada, 1927 FAMILIA PYROTEUTHIDAE Pyroteuthidae no identificados Pterygioteuthis sp.1 Pterygioteuthis sp. 2 se registró en el resto de los cruceros analizados. En los cruceros realizados durante El Niño 1997–1998 la familia Ommastrephidae registró el mayor porcentaje de abundancia relativa. En los cruceros de otoño de 1997 y 1998 sólo se separaron e identificaron paralarvas de esta familia, por lo que el porcentaje está influenciado por la falta de otros taxones. Sin embargo, durante invierno y verano de 1998 la importancia de los omastréfidos en la comunidad de cefalópodos alcanzó 72% y 89% respectivamente (fig. 1). En los cruceros de invierno y verano de 1999, realizados en C omposición de especies y patrones de distribución de paralarvas de calamar 459 condiciones de La Niña 1999–2001, la composición taxonómica cambió resultando más abundantes las familias Gonatidae, Onychoteuthidae y Cranchiidae (fig. 1). En el crucero de verano de 1999 también se registró la familia Ommastrephidae. Distribución En la primavera de 1996 se registraron paralarvas de Onychoteuthis borealijaponica en las 3 áreas de surgencia muestreadas, particularmente entre Punta Eugenia y Bahía Asunción (fig. 2a) y también en la zona oceánica (fig. 2g). Loligo opalescens se distribuyó entre Ensenada y Punta Baja y entre Punta Eugenia y Bahía Asunción, mientras que Abraliopsis spp. se registró sólo en la primera zona. En los cruceros de IMECOCAL analizados, excepto el de invierno de 1999, se identificaron paralarvas de las cinco especies de omastréfidos que se distribuyen como adultos en aguas del Pacífico mexicano, dos complejos denominados SD y EHO (paralarvas <3 mm de longitud de manto pertenecientes a Sthenoteuthis oualaniensis y/o Dosidicus gigas, y a Eucleoteuthis luminosa, Hyaloteuthis pelagica y/o Ommastrephes bartramii respectivamente), y dos morfotipos (morfotipo A y morfotipo B). De los taxones anteriores, D. gigas, S. oualaniensis y el complejo SD fueron los más abundantes concentrándose principalmente al sur del Figura 1. Abundancia relativa de paralarvas de calamar agrupadas por familia, registradas de 1996 a 1999 en la costa occidental de la Península de Baja California. En otoño de 1997 y 1998 sólo se separó e identificó a la familia Ommastrephidae. 460 Aspectos biológicos área de estudio entre Punta Eugenia y Punta Abreojos (figs. 2e, h; 3a). El resto de los taxones (figs. 2d; 3a, e) se distribuyeron con muy bajas abundancias frente a Punta Banda, Bahía Vizcaíno y Punta Abreojos. Leachia pacifica (grupo), de afinidad tropical y perteneciente a la familia Cranchiidae, se distribuyó con abundancias importantes en la zona sur entre Punta Eugenia y Punta Abreojos (fig. 2i) y también frente a Punta Colonet (fig. 3c), mientras que L. dislocata (fig. 3c, f) y todos los taxones de la familia Gonatidae de afinidad templada se registraron en toda el área de estudio asociados al avance de la masa de Agua Subártica (figs. 2j; 3b, c, f). La distribución de los taxones de las familias Onychoteuthidae (figs. 2g; 3d, g) y Pyroteuthidae (fig. 2f) no mostró en conjunto un patrón característico, pero la afinidad de cada taxón de manera individual, reflejó su afinidad por alguna de las masas de agua presentes en el área. Discusión El presente estudio aporta 12 taxones diferentes de calamar (2 especies y 10 formas) a los 60 ya registrados para el Pacífico Mexicano, incrementando en 20% el total y en 26% los reportados en el área frente a la costa occidental de la Península de Baja California. Se presenta también el primer listado de especies de calamares en México identificados por la presencia de sus paralarvas. Las paralarvas denominadas complejo SD y complejo EHO de la familia Ommastrephidae (<3 mm de longitud de manto) sólo pueden separarse morfológicamente en dos complejos de especies con base en el tamaño relativo de las ventosas laterales de la proboscis: el complejo SD correspondiente a paralarvas de S. oualaniensis y D. gigas con ventosas laterales del mismo tamaño que el resto, y el EHO que comprende las que tienen ventosas laterales del doble del tamaño que el resto y que incluye a E. luminosa, H. pelágica y O. bartrami. En términos de abundancia los resultados coinciden con reportes previos que establecen que las familias Ommastrephidae, Cranchiidae, Enoploteuthidae y Onychoteuthidae son las más importantes en los diferentes océanos alrededor del mundo (Vecchione et al. 2001, Diekmann et al. 2002). A nivel mundial se ha encontrado que usualmente el número de paralarvas de cefalópodos capturadas en una muestra de zooplancton va de 1 a 5 (Vecchione 1987), lo que coincide con las abundancias registradas en la mayoría de las muestras recolectadas en el área de estudio; sin embargo, en el caso de las que contenían omastréfidos su abundancia C omposición de especies y patrones de distribución de paralarvas de calamar 461 Figura 2. Distribución de las paralarvas de calamar más abundantes registradas durante primavera de 1996, otoño de 1997 e invierno y verano de 1998 en la costa occidental de la Península de Baja California. Los tonos de gris muestran la distribución de las masas de agua: gris claro = Agua Transicional-Subtropical Superficial; gris oscuro = Agua Subártica. La isolínea indica el límite entre ambas (tomado de Durazo y Baumgartner 2002). 462 Aspectos biológicos Figura 3. Distribución de las paralarvas de calamar más abundantes registradas durante otoño de 1998 e invierno y verano de 1999 en la costa occidental de la Península de Baja California. Los tonos de gris muestran la distribución de las masas de agua: gris claro = Agua Transicional-Subtropical Superficial; gris oscuro = Agua Subártica. La isolínea indica el límite entre ambas (tomado de Durazo y Baumgartner 2002). sobrepasó hasta en dos órdenes de magnitud al resto de las familias identificadas. De 1996 a 1999 se registró un cambio en la composición de especies de calamar en la costa occidental de la Península de Baja California, asociado a la presencia del ENOS. La gran abundancia de la familia Ommastrephidae durante El Niño 1997–1998 se correlacionó positivamente con las altas temperaturas y la masa de Agua Transicional-Subtropical Superficial presente en el área (Durazo y Baumgartner 2002), en particular la de D. gigas, S. oualaniensis y el complejo SD de afinidad C omposición de especies y patrones de distribución de paralarvas de calamar 463 tropical-subtropical (Camarillo-Coop 2006). La ausencia de estos taxones y la presencia de paralarvas de O. bartrami de afinidad templada y el complejo EHO, asociadas a la masa de Agua Subártica en invierno y verano de 1999, se relacionaron con el cambio a condiciones frías durante La Niña 1999–2001 (Granados-Amores et al. 2007). La concentración de paralarvas del complejo SD frente a Punta Eugenia indicó la presencia de desoves recientes de D. gigas o de S. oualaniensis (CamarilloCoop et al. 2007). Durante El Niño 1997–1998 los registros de capturas indicaron la ausencia de calamar (D. gigas) dentro del Golfo de California y su aparición en la costa occidental de la Península de Baja California (Morales-Bojórquez et al. 2001), lo que sugiere que ambos eventos están relacionados. La distribución de las paralarvas del resto de las familias analizadas fue el resultado del acoplamiento de cada taxón con una u otra masa de agua en función de su afinidad biogeográfica (Granados-Amores 2008). Las paralarvas de las familias Cranchiidae, Onychoteutidae y Pyroteuthidae (L. pacifica, O. banksi sp. 2 y Pterygioteuthis sp. 1), cuyos adultos son de afinidad tropical, se distribuyeron en la masa de agua cálida Transicional-Subtropical Superficial registrada durante 1997–1998 (Durazo y Baumgartner 2002), replegándose hacia el sur cuando la influencia de la masa de Agua Subártica invadió el área de estudio. Esta última permitió la presencia de la familia Gonatidae así como de L. dislocata, O. banksi sp. 1, O. borealijaponica y Pterygioteuthis sp. 2, todas de afinidad templada. La distribución y abundancia de las paralarvas fue modulada por las características físicas y el avance de las masas de agua como se ha establecido para otras especies (Yamamoto et al. 2002) con cambios estacionales cuyas abundancias se incrementan por el efecto del ENOS en la región. Los adultos y las paralarvas de Loligo opalescens, comercialmente importante en California, EUA, y potencialmente importante en México (Okutani y McGowan 1969, Roper et al. 1995), se distribuyen en zonas neríticas y en la plataforma continental (Sweeney et al. 1992, Hochberg 1998). Su registro en el crucero costero durante el mes de marzo confirma su presencia en la temporada fría en la península, coincidiendo con los desoves de invierno registrados para el sur de California (Hastings y MacWilliams, 1999), y su ausencia en los cruceros IMECOCAL se relacionó con la falta de cobertura de muestreo en la zona costera. Es evidente que aún existe poca información sobre los cefalópodos en México. Este trabajo cubre sólo una porción del área marítima mexicana, por lo que la riqueza de especies puede ser mayor. Ecológicamente los calamares son uno de los grupos más complejos dada su amplia distribución horizontal y 464 Aspectos biológicos vertical, y por su comportamiento, uno de los más difíciles de estudiar. A pesar de ello, los resultados indican que las aguas frente a la costa occidental de la Península de Baja California albergan una gran riqueza de especies de calamar con diferente afinidad biogeográfica. La distribución de sus paralarvas a lo largo del año parece estar asociada con el flujo estacional e interanual de las masas de agua predominantes en la región. Referencias Camarillo-Coop S. 2006. Variación espacio-temporal de paralarvas de calamares (Cephalopoda: Ommastrephidae) de importancia comercial en la costa occidental de la Península de Baja California. Tesis de Maestría. CICIMAR-IPN, México. 92 pp. Camarillo-Coop S, De Silva-Dávila R, Hernández-Rivas ME, Durazo-Arvizu R. 2007. Distribution of Dosidicus gigas paralarvae off the west coast of the Baja California Peninsula, Mexico. En: Olson RJ, Young JW (eds.), The role of squid in open ocean ecosystems. Rep. GLOBEC-CLIOTOP/PFRP workshop, 16–17 November 2006, Honolulu, Hawaii, USA. GLOBEC Report 24: vi. Pp 7–8. Diekmann R, Piatkowski U, Schneider M. 2002. Early life and juvenile cephalopods around seamounts of the subtropical eastern North Atlantic: illustrations and a key for their identification. Ber. Inst. Meereskd. Kiel. 326: 1–42. Durazo R, Baumgartner T. 2002. Evolution of oceanographic conditions off Baja California: 1997–1999. Progr. Oceanogr. 54: 7–31. Durazo R, Gaxiola-Castro G. 2010. Introducción. En: Gaxiola-Castro G, Durazo R (eds.), Dinámica del Ecosistema Pelágico frente a Baja California, 1997–2007: Diez años de Investigaciones Mexicanas de la Corriente de California. Pp. 13–18. Granados-Amores J. 2008. Calamares epiplanctónicos de la costa occidental de la Península de Baja California, México. Tesis de Maestría. CICIMAR-IPN, México. 129 pp. Granados-Amores J, De Silva-Dávila R, Hernández-Rivas ME. 2007. Epiplanktonic squids from the west coast of the Baja California Peninsula, México. En: Olson RJ, Young JW (eds.), The role of squid in open ocean ecosystems. Rep. GLOBEC-CLIOTOP/PFRP workshop, 16–17 November 2006, Honolulu, Hawaii, USA. GLOBEC Report 24: vi. Pp. 22–24. Haimovici M, Piatkowski U, Aguiar dos Santos R. 2002. Cephalopod paralarvae around tropical seamounts and oceanic islands of the northeastern coast of Brazil. Bull. Mar. Sci. 71: 313–330. Hastings S, MacWilliams S. 1999. Multi-species and multi-interest management: an ecosystem approach to market squid (Loligo opalescens) harvest in California: Marine SanctuaC omposición de especies y patrones de distribución de paralarvas de calamar 465 ries Conservation Series MSD-99-1. U.S. Department of Commerce, National Oceanic and Atmospheric Administration, Marine Sanctuaries Division, Silver Spring, Md. 24 pp. Hochberg FG. 1998. Class Cephalopoda. En: Scott PV, Blake JA (compiladores), Taxonomic Atlas of the Benthic Fauna of the Santa Maria Basin and the Western Santa Barbara Channel, Vol. 8: The Mollusca, Part 1. Santa Barbara Museum of Natural History: Santa Barbara, CA. Pp. 175–236. Lavaniegos B, Durazo R, Gaxiola-Castro G, Baumgartner T, Jiménez-Pérez LC, GarcíaCórdova J, González-Esparza M. 2001. El Niño y el mar. En: Escobar E, Bonilla M, Badán A, Caballero M, Winckell A (compiladores). Los efectos del fenómeno El Niño en México 1997–1998. CONACYT México. Pp. 181–198. Loeb VJ, Smith PE, Moser HG. 1983. Geographical and seasonal patterns of larval fish species structure in the California Current area. 1975. Calif. Coop. Oceanic Fish. Invest. Rep. 24: 132–151. Markaida U. 2006. Population structure and reproductive biology of jumbo squid Dosidicus gigas from the Gulf of California after the 1997–1998 El Niño event. Fish. Res. 79: 28–37. Morales-Bojórquez E, Cisneros-Mata MA, Nevárez-Martínez MO, Hernández-Herrera A. 2001. Review of stock assessment and fishery biology of Dosidicus gigas in the Gulf of California, Mexico. Fish. Res. 54: 83–94. Okutani T, McGowan J. 1969. Systematics, distribution, and abundance of the epiplanktonic squid (Cephalopoda, Decapoda) larvae of the California Current, April, 1954– March, 1957. Bull. Scripps Inst. Oceanogr. Univ. Calif. 14: 1–90. Palomares-García R, Martínez-López A, De Silva-Dávila R, Funes-Rodríguez R, CarballidoCarranza MA, Avendaño-Ibarra R, Hinojosa-Medina A, López-Ibarra GA. 2003. Biological effects of El Niño 1997-98 on a shallow subtropical ecosystem: Bahía Magdalena, Mexico. Geofís. Int. 42: 455–466. Postel L, Fock H, Hagen W. 2000. Biomass and abundance, En: Harris RP, Wiebe PH, Lenz J, Skjoldal HR, Huntley M (eds.), Zooplankton Methodology Manual. Academic Press. San Diego, California. Pp. 83–92. Rodhouse PGK, Waluda CM, Morales-Bojórquez E, Hernández-Herrera A. 2006. Fishery biology of the Humboldt squid, Dosidicus gigas, in the eastern Pacific Ocean. Fish. Res. 79: 13–15. Roper CFE, Sweeney MJ, Hochberg FG. 1995. Cefalópodos. En: Fischer W, Krupp F, Schneider W, Sommer C, Carpenter KE, Niem VH (eds.), Guía FAO Para la Identificación de Especies Para los Fines de la Pesca. Pacífico Centro-Oriental. Vol. I. Plantas e Invertebrados. FAO. Roma. Pp. 305–353. 466 Aspectos biológicos Saito K. 1994. A sampling method for rhynchoteuthion paralarvae of neon flying squid Ommastrephes bartramii. Bull. Hokkaido Nat. Fish. Res. Inst. 58: 25–34. Sistema de Información Taxonómica Integrada (ITIS). 2008. http://www.itis.usda.gov Smith PE, Richardson SL. 1979. Técnicas modelo para prospección de huevos y larvas de peces pelágicos. FAO. Doc. Tec. Pesca. 175 pp. Sweeney MJ, Roper CFE, Mangold KM, Clarke MR, Boletzky SV. 1992. “Larval” and juvenile cephalopods: a manual for their identification. Smith. Contrib. Zool. Smithsonian Institution Press, Washington D.C. 282 pp. Vecchione M. 1987. Juvenile ecology. En: Boyle PR (ed.), Cephalopod Life Cycles. Vol. 2. Academic Press. London, UK. 475 pp. Vecchione M, Roper CFE, Sweeney MJ, Lu CC. 2001. Distribution, relative abundance and developmental morphology of paralarval cephalopods in the Western North Atlantic ocean. NOAA Tech. Rep. NMFS 152. Washington, U.S.A. 54 pp. Yamamoto J, Masuda S, Miyashita K, Uji R, Sakurai Y. 2002. Investigation on the early stages of the ommastrephid squid Todarodes pacificus near the Oki islands (Sea of Japan). Bull. Mar. Sci. 71: 987–992. C omposición de especies y patrones de distribución de paralarvas de calamar 467