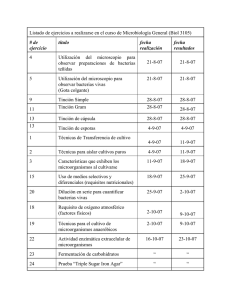
Curso completo - Universidad Pública de Navarra
Anuncio

MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Universidad Pública de Navarra Departamento de Producción Agraria MICROBIOLOGÍA CLÍNICA 1er CURSO DE DIPLOMATURA EN ENFERMERÍA CURSO 2008-2009 GRUPO 1 CÓDIGO 48108 Prof. Dr. Antonio G. Pisabarro Catedrático de Microbiología Pamplona, 2009 1 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) MICROBIOLOGÍA CLÍNICA CÓDIGO 48108 PROFESORES DE LA ASIGNATURA Prof. Dr. D. Antonio G. Pisabarro, Catedrático de Microbiología, Profesor responsable del grupo 1 Teléfono: 948 169 107. Tutorías: miércoles de 11 a 12 h en el despacho de la escuela de estudios sanitarios. E-mail: [email protected] (Indicar en el subject del mensaje MicroClinica) Dra. Dña. María M. Peñas, Profesora Ayudante Doctor de Microbiología, responsable de la docencia práctica Teléfono: 948 169 219 Tutorías: Es necesario pedir cita previa mediante un mensaje enviado a la dirección de e-mail. E-mail: [email protected] DESCRIPCIÓN DE LA ASIGNATURA La asignatura de MICROBIOLOGÍA CLINICA (Código 48108) consta de 4.5 créditos (3.0 teóricos y 1.5 prácticos) englobados dentro de la materia troncal de Enfermería comunitaria 1. El objetivo de la misma es presentar al alumno los fundamentos de la microbiología que influyen en la actividad profesional de los diplomados en enfermería que entran en contacto con pacientes de enfermedades infecciosas o especialmente susceptibles a ellas. La asignatura se centra más que en el diagnóstico y tratamiento de las enfermedades infecciosas, en estudiar cómo se limita la difusión de los microorganismos evitando su transmisión y contagio. En el Plan de Estudios de la Diplomatura en Enfermería existe una asignatura optativa titulada Descripción y Detección de Agentes Infecciosos enfocada en el estudio más detallado de los principales grupos de microorganismos patógenos de importancia sanitaria. EVALUACIÓN DE LA ASIGNATURA • • • • • Para la evaluación de la asignatura se tendrán en cuenta la asistencia a clase teórica, y los exámenes. Durante el curso, habrá dos exámenes parciales para todos los alumnos que no serán liberatorios. El primero comprenderá los temas 1 a 4 (ambos inclusive) y el segundo los temas 5 a 9. Cada uno de los exámenes parciales contribuye con un 20% a la nota de la asignatura. El examen final, se celebrará en las fechas publicadas oficialmente, comprende toda la asignatura y contribuye con un 60% a la nota de la asignatura. Las calificaciones de los exámenes parciales tendrán valor para las convocatorias ordinaria (junio) y extraordinaria (septiembre). 2 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) • • • Para aprobar la asignatura es necesario haber realizado correctamente las prácticas de la misma y alcanzar una nota mínima de 5.0. El examen final de junio incluirá unas preguntas de prácticas cuya superación será condición necesaria para los alumnos repetidores de la asignatura que por estar en otros cursos no puedan asistir a las prácticas de laboratorio. Para el resto de los alumnos que se encuentran en primero, las preguntas de prácticas computarán en el examen como el resto de las preguntas del mismo. A aquellos alumnos que hayan conseguido una nota de 5.0 o superior, se les podrá sumar hasta un punto por la asistencia a las clases teóricas. Para los alumnos repetidores, se contabilizará la asistencia del curso anterior 07-08. PROGRAMACIÓN A continuación se indican las fechas previstas para las clases y los temas que se tratarán en ellas. Los números de los temas son los de este volumen de Notas de Microbiología Clínica. Clases prácticas: De lunes a jueves de 17:00 a 20:00 y los viernes de 9:00 a 12:00 en las semanas indicadas para cada grupo en la guía y los tablones de anuncios. 3 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) TEMAS DE TEORÍA Tema 1.- Estructura de los microorganismos. Células procariotas y eucariotas. Tamaño y morfología de las bacterias. Observación microscópica de los microorganismos. Tinciones. Membrana bacteriana y peptidoglicano. Bacterias Gram-positivas y Gramnegativas. Ribosomas bacterianos. Elementos facultativos de la célula procariota. Cápsula y capas mucosas. Apéndices bacterianos: flagelos y fímbrias. Endosporas. Estructura de los virus. Tema 2.- Crecimiento y muerte de microorganismos. Concepto y expresión matemática del crecimiento bacteriano. Concepto de muerte de un microorganismo. ¿Qué necesita un microorganismo para crecer?. Detección y medida del crecimiento. Ciclo de crecimiento de poblaciones. Factores físicos y químicos que influyen en el crecimiento. Multiplicación de los virus. Tema 3.- Taxonomía microbiana. Principales grupos de Bacterias, Virus, Hongos y Protozoos de importancia clínica. Tema 4.- Genética microbiana. Elementos genéticos bacterianos. Transferencia horizontal de material genético en microorganismos: transformación, conjugación y transducción. Elementos genéticos de los virus. Expresión de la información genética. Tema 5.- Interacción con los microorganismos. Concepto de flora normal. Localización de la flora normal. Interacción patogénica entre huésped y bacteria. postulados de Koch. Poder patogénico y virulencia. Factores determinantes de la acción patógena Tema 6.- Dispersión de los microorganismos. Reservorios y fuentes de infección. El contagio y su prevención. Vías y modos de transmisión microorganismos. Tema 7.- Esterilización, desinfección y asepsia. Esterilización, desinfección, descontaminación y antisepsis. Desinfectantes y antisépticos. Evaluación de la actividad germicida. Desinfectantes de alta, media y baja actividad. Desinfección de endoscopios. Esterilización por calor. Esterilización por óxido de etileno. Tema 8.- Control de microorganismos. Inhibición del crecimiento. Grupos de antibióticos y quimioterápicos. Mecanismos de resistencia a antibióticos. Cepas multirresistentes. Precauciones en el uso de antibióticos. Tratamiento de las infecciones virales. Tratamiento de enfermedades causadas por protozoos y por hongos. Tema 9.- Defensa frente a microorganismos. Barreras a la entrada de organismos. Respuesta inespecífica a la infección. Respuesta inmune específica. Antígenos y anticuerpos. Células de la respuesta inmune específica. Mecanismo de la respuesta inmune específica. Ejemplo del desarrollo y función del sistema inmune: el sistema inmune intestinal. Reacciones de hipersensibilidad. Inmunodeficiencias. Enfermedades autoinmunes. Superantígenos. Inmunoterapia. 4 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) TEMAS DE LABORATORIO La asistencia a las prácticas de laboratorio y la superación de esta parte del programa es imprescindible para poder aprobar la asignatura conjunta. Práctica 1: Preparación de medios de cultivo y siembra de bacterias en medios de cultivo. Esterilización. Con esta práctica se pretende aprender las técnicas de preparación de diferentes medios de cultivo, su esterilización y su almacenaje, así como la importancia de los diferentes medios de cultivo y su uso. Además también se pretende que el alumno se familiarice con las técnicas empleadas en Microbiología para el cultivo y la manipulación de microorganismos en condiciones de esterilidad. Práctica 2: Obtención de cultivos puros. Con esta práctica se pretende estudiar las diferentes técnicas de aislamiento de un microorganismo a partir de una población mixta para obtener un cultivo puro. Práctica 3: Recuento del número de bacterias por mililitro de un cultivo líquido. El objetivo de esta práctica es dar a conocer al alumno otra de las técnicas de obtención de cultivo puros así como el aprendizaje para hacer diluciones y recuentos bacterianos. Práctica 4: Cultivo de bacterias anaerobias. El objetivo de esta práctica es conocer el fundamento y la técnica más comúnmente empleada en el aislamiento de bacterias anaerobias. Práctica 5: Observación de bacterias teñidas. Tinción Simple y Tinción de Gram. Con esta práctica se pretende familiarizar al alumno con el manejo del microscopio óptico para la visualización de microorganismos. Se observarán preparaciones bacterianas bajo diferentes técnicas de tinción que permitan establecer diferencias morfológicas y estructurales entre y dentro de los microorganismos. Práctica 6: Transformación bacteriana por resistencia a ampicilina. En esta práctica se hablará de los fenómenos de transformación, conjugación y transducción y se realizará un ensayo de transformación bacteriana por adquisición de plásmidos resistentes al antibiótico ampicilina. Práctica 7: Antibiograma. El objetivo de esta práctica es estudiar qué son los antibióticos y cómo se realiza la valoración de la actividad antimicrobiana de un compuesto químico (antibiótico) mediante la técnica de Kirby-Bauer. Práctica 8: Efecto de las altas temperaturas sobre el crecimiento microbiano El objetivo de esta práctica consiste en estudiar el efecto y la resistencia de un cultivo bacteriano a altas temperaturas. Práctica 9: Aislamiento y características de un bacteriófago (virus de bacterias) 5 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) El objetivo de esta práctica consiste en visualizar la presencia de bacteriófagos a través de las consecuencias de su ciclo infeccioso sobre bacterias de Staphylococcus aureus. Práctica 10: Observación de levaduras. El objetivo de esta práctica es el cultivo de diferentes levaduras y la posterior identificación de las mismas de acuerdo a sus características morfológicas a través de su observación al microscopio. Práctica 11: Tinción de esporas. En esta práctica se estudiará el género Bacillus por su capacidad para la formación de endosporas y se procederá a la realización de la correspondiente tinción diferencial para poder visualizar al microscopio dichas estructuras. Práctica 12: Tinción de cápsula. En esta práctica se estudiarán las cápsulas bacterianas y se procederá a una tinción diferencial para la visualización de la misma al microscopio. 6 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) BIBLIOGRAFÍA: 1.- Pisabarro, A.G. 2003. Notas de clase de Microbiología Clínica. (Revisión 2006) 2.- de la Rosa, M. 1997. Microbiología: Enfermeria - Ciencias de la Salud. Conceptos y Aplicaciones. Ed. Harcourt Brace. 3.- Stuart Walker, T. 1998. Microbiología. Ed. McGraw-Hill Interamericana. 4.- Ingraham, J.L. y C.A. Ingraham. 1998. Introducción a la Microbiología. Ed. Reverté, S.A. 5.- García-Rodríguez, J.A. y J.J. Picazo. 1996. Microbiología Médica. Ed. Mosby Pamplona miércoles, 17 de septiembre de 2008 7 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) TEMAS DE TEORÍA 8 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Tema 1.- Estructura de los microorganismos. Células procariotas y eucariotas. Tamaño y morfología de las bacterias. Observación microscópica de los microorganismos. Tinciones. Membrana bacteriana y peptidoglicano. Bacterias Gram-positivas y Gramnegativas. Ribosomas bacterianos. Elementos facultativos de la célula procariota. Cápsula y capas mucosa. Apéndices bacterianos: flagelos y fímbrias. Endosporas. estructura de los virus. 1.- INTRODUCCIÓN Las enfermedades infecciosas, aunque están relativamente controladas en los países desarrollados, siguen siendo una de las principales causas de mortalidad en los países en desarrollo. A este hecho se une la aparición de nuevas enfermedades infecciosas desconocidas conocidas como enfermedades emergentes (SIDA, Legionelosis, Ébola, Síndrome Respiratorio Agudo Severo o SARS, Gripe Aviar, etc.) y la reaparición de otras que habían sido controladas pero que surgen de nuevo presentado nuevas resistencias (Tuberculosis) conocidas como enfermedades re-emergentes. Por último, hay que señalar la alta incidencia de las infecciones hospitalarias (infecciones nosocomiales) producidas por patógenos portadores de múltiples factores de resistencia y el aumento de los segmentos de población especialmente susceptibles a enfermedades infecciosas, para poder tener una imagen completa de la incidencia de la microbiología en la sanidad contemporánea. Por otro lado, la interacción de los microorganismos que constituyen la flora (microbiota) normal con nuestro cuerpo es esencial para el desarrollo correcto de nuestro sistema inmune y vascular. Así mismo, la actividad microbiana es también esencial para un correcto procesamiento y absorción de los alimentos a nivel intestinal. LOS MICROORGANISMOS Y LA SALUD PÚBLICA Las enfermedades infecciosas tienen una relevancia especial en la salud pública. Las infecciones víricas o bacterianas más frecuentes suelen pasar desapercibidas para la administración nacional de salud y sólo se tienen datos razonablemente fiables de las enfermedades que, por sus especiales características, son de declaración obligatoria. Entre las primeras (sin declaración obligatoria) debemos contabilizar no sólo las enfermedades infecciosas comunes que o bien no están incluidas en brotes epidémicos o, si lo están, no requieren ser declaradas individualmente; sino que también hay que considerar, en conjunto, las infecciones producidas por lesiones que presentan normalmente un tratamiento fácil y pronóstico favorable. La incidencia de las enfermedades infecciosas puede valorarse mediante el seguimiento de los brotes epidémicos de enfermedades transmitidas a través de los alimentos y cuya declaración sea obligatoria y mediante el seguimiento general de las enfermedades de declaración obligatoria que en 2003 supusieron más de 1.7 millones de casos en toda España (Datos del Boletín Epidemiológico Semanal del Instituto Carlos III) de los 9 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) cuales la gripe (con cerca de 1,5 millones de casos en 20031) y la varicela (con más de 233.000 casos en 20032) son las de mayor incidencia. 2.- CÉLULAS PROCARIOTAS Y EUCARIOTAS. Organismos eucarióticos son aquellos en cuyas células puede diferenciarse un núcleo que contiene el material genético separado de un citoplasma en el que se encuentran diferentes orgánulos celulares. Los microorganismos eucarióticos más relevantes en clínica incluyen ciertos animales de pequeño tamaño productores de enfermedades parasitarias, protozoos y hongos unicelulares o pluricelulares. Organismos procarióticos son aquellos en los que no existe la separación entre núcleo y citoplasma. Dentro de este grupo se incluyen las bacterias, a las que dedicaremos la mayor parte del curso. Mención aparte merecen los virus, partículas inanimadas de material genético protegido por capas más o menos complejas de proteínas y lípidos. Carecen de actividad metabólica cuando se encuentran libres. En general, las células procarióticas son más simples que las eucarioticas ya que estas contienen membranas internas que diferencian órganos celulares (aparato de Golgi, retículo endoplásmico, vacuolas, etc.) no presentes en las células procariotas. En estas el citoplasma es continuo y se encuentra lleno de ribosomas encargados de llevar a cabo la traducción del mensaje genético en proteínas. Las células eucarióticas son el resultado de una simbiosis establecida hace muchos millones de años entre células procarióticas (que han dado lugar a las mitocondrias) y un núcleo eucariótico (el núcleo de nuestras células). A causa de esta simbiosis, ciertos agentes químicos que son activos frente a procariotas pueden resultar tóxicos para eucariotas al actuar sobre sus mitocondrias. Estas simbiosis estables entre organismos de diferentes orígenes son relativamente frecuentes y se dan en organismos patógenos como, por ejemplo, el Plasmodium productor de la malaria o paludismo. 3.- TAMAÑO Y MORFOLOGÍA DE LAS BACTERIAS. La forma de las bacterias puede ser esférica (cocos), cilíndrica (bacilos), de coma (vibrios) o helicoidal (espirilos). La forma de las bacterias viene determinada principalmente por la estructura de su pared celular y es una de las características que sirven para identificarlas. 1 La acumulación de casos de una enfermedad puede variar en distintos años, así, la acumulación de casos declarados de gripe en 2004 fue de poco más de medio millón de casos, un poco más de un tercio de los casos declarados en 2003 (datos de Boletín Epidemiológico Semanal 12: 292). 2 Incidencia declarada: 178.933 en 2003 y 233.934 en 2004. (Datos del Boletín Epidemiológico Nacional. 12: 289-296). 10 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Las bacterias pueden presentarse como células aisladas o formando grupos. Esta característica es también importante para poder identificarlas. En algunas casos la aparición de las bacterias formando agrupaciones no es una característica de estas in vivo sino un efecto de ciertas técnicas de tinción (como en el caso del género Staphylococcus que aparece formando racimos en preparaciones fijadas y teñidas; pero no en muestras vivas). Las principales formas de agrupamiento de las bacterias son las que se observan en estreptococos y estreptobacilos (cadenas de cocos o de bacilos, respectivamente), estafilococos (agrupaciones en forma de racimos de cocos), diplococos (parejas de cocos) sarcinas (agrupaciones en tétradas o en grupos de ocho cocos dispuestos en forma de cubo). El tamaño de las células bacterianas es variable oscilando entre una micra (µm) de diámetro y varias decenas de longitud en las especies más grandes. En cualquier caso, su tamaño es más reducido que el de una célula eucariótica típica. 4.- OBSERVACIÓN MICROSCÓPICA DE LOS MICROORGANISMOS. Debido a su pequeño tamaño, la observación de los microorganismos ha de realizarse mediante el uso de microscopios. En el caso de la mayoría de los microorganismos procarióticos, la observación microscópica se limita a determinar la forma, el modo de agrupamiento, el movimiento y la presencia de algunos elementos extracelulares tales como las cápsulas. En el caso de microorganismos eucarióticos, la información que se obtiene de la observación microscópica es mucho mayor porque también lo es su diversidad morfológica y anatómica. Una consideración especial requiere el método de tinción de Gram que permite clasificar la mayoría de las bacterias en uno de dos grandes grupos cuyas características anatómicas y fisiológicas son diferentes. Las técnicas microscópicas más frecuentes de uso en microbiología son las de microscopía óptica normal para observar células fijadas y teñidas, de contraste de fase para la observación de células vivas, de fluorescencia para observar células marcadas y de interferencia (Nomarski) para la observación de células vivas. En microscopía electrónica, se usan tanto microscopios de transmisión (MET) como microscopios de barrido (MEB). La resolución que se alcanza en microscopía óptica está en torno a 0,2 µm, en MET puede llegar a los 0,2 nm y en MEB entre 1 y 10 nm. 5.- TINCIONES. Como se ha indicado en el apartado anterior, la observación microscópica de bacterias proporciona una cantidad limitada de información debido a la reducida variabilidad morfológica de estos organismos. La observación de microorganismos vivos (contraste de fase) permite obtener información relativa a su movilidad y a su tamaño. Sin embar- 11 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) go, la mayor parte de la información microscópica deriva del examen de muestras fijadas y teñidas. La fijación consiste en la precipitación de las proteínas y desecación de las muestras mediante técnicas físicas o químicas. En la práctica, la fijación se suele realizar por calor con lo que se consigue la desnaturalización y desecación al mismo tiempo que se adhiere la muestra al portaobjetos en el que se realizará la observación. En la mayoría de los casos, la fijación se realiza en muestras que han sido extendidas sobre el portaobjetos produciendo lo que se denomina un frotis. Una vez fijada, la muestra se somete a uno o varios procesos de tinción a fin de revelar la morfología del microorganismo y algunas de sus características estructurales. En algunos casos (como el ya citado de Staphylococcus) la fijación puede dar lugar a agrupaciones características de bacterias que permiten su identificación. Para realizar la tinción es necesario usar colorantes específicos. Los colorantes se agrupan en básicos que tiñen especialmente bacterias ya que la membrana de estas suele tener una cierta carga eléctrica negativa (a este grupo pertenecen la safranina, fucsina básica, cristal violeta y azul de metileno); y ácidos más adecuados para teñir células animales en muestras de tejidos infectados. A este último grupo pertenecen la fucsina ácida, la eosina y el rojo Congo. Para ciertos tipos de tinciones es necesario aumentar la afinidad del colorante por el microorganismo (o estructura) utilizando substancias que actúan como mordientes. Los mordientes también se utilizan para aumentar el grosor de ciertas estructuras como los flagelos de forma que puedan ser observados al microscopio cuando se tiñen adecuadamente. Los principales tipos de tinción que se usan en microbiología clínica son los siguientes: TINCIÓN SIMPLE Permite observar la forma, tamaño y agrupamiento de las bacterias usando un único colorante (normalmente básico). TINCIÓN DIFERENCIAL Permiten distinguir diferentes tipos de microorganismos en función de sus características estructurales Tinción de Gram: es un sistema de dos tinciones simples sucesivas, separadas por una fase de decoloración selectiva. Permite diferenciar las bacterias que retienen el primer color (Gram-positivas) de las que no lo retienen (Gram-negativas). Esta diferencia en comportamiento refleja diferencias estructurales y fisiológicas entre ambos grupos de bacterias. 12 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Tinción de esporas (Wirtz-Conklin): ciertos tipos de bacterias producen esporas de resistencia cuya pared celular es característica. La tinción de esporas permite detectar este tipo de células lo que tiene utilidad en la identificación del microorganismo en estudio. Tinción de Ziehl-Neelsen (ácido-alcohol resistencia): es un tipo especial de tinción que permite la identificación de microorganismos de los grupos Mycobacterium y Nocardia de gran relevancia clínica Tinción de cápsulas: se trata de una tinción negativa usando tinta china que permite determinar la presencia de cápsulas polisacarídicas. Tinción de flagelos (Leifson): permite teñir flagelos usando un mordiente para incrementar su grosor y hacerlos visibles al microscopio óptico. 6.- MEMBRANA BACTERIANA Y PEPTIDOGLICANO. Las bacterias presentan una membrana interna que rodea el citoplasma bacteriano y presenta las características generales de las membranas plasmáticas. Hacia el exterior de la membrana interna, todas las bacterias (con excepción de los micoplasmas grupo de organismos al que pertenecen patógenos de los géneros Mycoplasma y Ureaplasma) presentan una pared celular formada por un polímero complejo denominado peptidoglicano. Algunos tipos de bacterias tienen una segunda membrana (membrana externa) que recubre la capa de peptidoglicano por su parte exterior. En las bacterias que tienen membrana externa (bacterias Gram-negativas), el espesor de la capa de peptidoglicano es más reducido que en las que carecen de ella (bacterias Gram-positivas). Se denomina espacio periplásmico al comprendido entre las membranas interna y externa en las bacterias Gram-negativas y al inmediatamente adyacente a la membrana interna en el caso de las bacterias Gram-positivas. ESTRUCTURA DE LA MEMBRANA INTERNA La membrana interna está formada por una bicapa lipídica. En las bacterias los lípidos que forman esta membrana son generalmente fosfolípidos y no se encuentran esteroles (salvo en el caso de los micoplasmas). Esto diferencia claramente las membranas bacterianas de las de células eucarióticas que sí tienen esteroles en sus membranas FUNCIONES DE LA MEMBRANA INTERNA 1.- Barrera de permeabilidad selectiva: La membrana lipídica que recubre las células es impermeable a las moléculas cargadas y a los iones, mientras que es permeable a los compuestos orgánicos y moléculas neutras. Por ello es una barrera de permeabilidad que restringe el paso de los nutrientes al interior de la célula, y el de compuestos intracelulares al exterior. La entrada de compuestos polares a través de la membrana lipídica se consigue de varias maneras: 13 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) a.- Poros a través de la membrana que permiten el paso selectivo de compuestos polares. Estos canales suelen estar dedicados a la entrada de solutos y su entrada está impulsada por la difusión libre a lo largo de un gradiente de concentración. b.- Transportadores que recogen del exterior moléculas y las introducen al interior consumiendo energía en el proceso. La energía motriz en este caso es ATP. c.- En el caso de ciertos iones intracelulares, éstos son transportados al exterior mediante sistemas de proteínas transportadoras de forma que se generan gradientes transmembranales. La energía que impulsa este transporte deriva de la oxidación de compuestos reducidos (potencial redox). d.- En el caso de los protones (H+) que se acumulan en el exterior de la célula estableciéndose un gradiente a través de la membrana, la entrada de estos cationes puede realizarse a través de canales especializados de forma que la energía que se libera como consecuencia de la entrada de los protones (destrucción del gradiente) se emplea para la síntesis de ATP. Estos canales son conocidos como protón-ATPasas. e.- En el caso de proteínas de membrana que han de exportarse al exterior de la célula, éstas tienen secuencias especiales en su extremo amino-terminal que las dirigen hacia la membrana y son procesadas (las secuencias especiales son eliminadas) cuando se produce el tránsito a través de la membrana celular. Los sistemas de transporte activo a través de la membrana pueden clasificarse en tres grandes grupos: uniportadores que translocan un solo tipo de moléculas, simportadores que translocan una molécula asociada a otra que se mueve a lo largo de un gradiente de concentración, y antiportadores que son similares a los anteriores pero en los que la energía liberada por el transporte a favor de gradiente de concentración se utiliza para translocar otra molécula en dirección contraria. 2.- Soporte ordenado de sistemas enzimáticos: Gran cantidad de reacciones metabólicas han de llevarse a cabo por conjuntos de varias enzimas que funcionan en cadena de forma que los productos de la reacción catalizada por una enzima son substrato de la siguiente y así sucesivamente. Para que estas reacciones puedan funcionar eficientemente, es necesario que los constituyentes de estos sistemas multienzimáticos estén colocados correctamente en el espacio de forma que la transferencia de substratos entre ellos sea la apropiada. Las membranas biológicas son un buen soporte de sistemas enzimáticos de este tipo ya que en ellas pueden ordenarse en dos dimensiones los constituyentes de la cadena de enzimas. Sistemas enzimáticos de membrana son los responsables de la cadena respiratoria, los de síntesis de pared celular, los de generación de ATP mediante sistemas de protón-ATPasa, los de recepción y transmisión de señales al interior celular, etc. En la membrana interna se encuentran los sistemas enzimáticos responsables de la síntesis de la propia membrana, los sistemas receptores de señales extracelulares (sistemas de dos componentes formadas por un receptor de la señal y un transmisor de la señal al interior celular), el sistema de transporte de electrones acoplado al transporte de protones que forma la cadena respiratoria, el sistema de protón ATP-asa responsable de 14 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) la síntesis de ATP, el sistema enzimático responsable de las últimas etapas de la síntesis de la capa de peptidoglicano y los sistemas de transporte a través de membrana. Debido a su naturaleza lipídica las propiedades de las membranas cambian con la temperatura pasando de una estructura de membrana fluida a temperaturas altas (similares a las ambientales o a las del interior de los organismos donde viven) a una estructura de membrana cristalina a temperaturas bajas (del entorno de 4ºC). La condición fluida o cristalina de una membrana afecta a su actividad y, por consiguiente, al metabolismo general de la célula. La integridad de la membrana interna es vital para la célula: si la membrana se rompe el contenido celular se pierde. Incluso pequeños poros no controlados que se produzcan accidentalmente en la membrana pueden ser fatales ya que la generación de la energía celular es sólo posible cuando la membrana interna está íntegra. Debido a sus especiales características, la membrana interna es diana para la acción de diferentes tipos de antibióticos tales como los inhibidores de la síntesis de peptidoglicano (antibióticos β-lactámicos, grupo al que pertenecen la penicilina y sus derivados) o antibióticos formadores de poros que destruyen los gradientes transmembranales. La membrana interna está empujada por la presión de turgor contra la capa de peptidoglicano. ESTRUCTURA DEL PEPTIDOGLICANO El peptidoglicano es una macromolécula que rodea completamente las células bacterianas proporcionándoles resistencia mecánica frente a la presión osmótica y confiriéndoles la forma característica de los diferentes grupos bacterianos. La membrana interna por sí sola no es capaz de soportar la presión de turgor (debida a la presión osmótica) de la célula bacteriana. El elemento de resistencia mecánica de la célula es la capa de peptidoglicano. La capa de peptidoglicano está formada por un polímero complejo denominado mureína que forma una macromolécula que recubre completamente la célula. Estructuralmente está formado por cadenas glucosídicas en las que se repite una unidad elemental de N-acetil-glucosamina unida por un enlace glicosídico β1→4 a ácido N-acetil-murámico. Las unidades elementales están también unidas entre ellas mediante enlaces glicosídicos β1→4. Estas cadenas glucosídicas pueden ser de longitud variable entre las diferentes especies bacterianas y entre diferentes momentos de la vida de la bacteria. Las cadenas glucosídicas están orientadas de forma paralela y están unidas entre sí mediante puentes peptídicos formados por cadenas de aminoácidos que están unidos al resto de ácido N-acetil-murámico. La orientación de las cadenas glicosídicas no se conoce con claridad todavía: en algunos modelos de peptidoglicano se propone que están orientadas de manera perpendicular al eje mayor de la bacteria, mientras que en otros se propone que se orientan de forma perpendicular a la superficie de la bacteria. 15 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Las cadenas peptídicas unidas al ácido N-acetil-murámico están formadas por aminoácidos en los que se observa una alternancia de restos con configuración L y con configuración D. La presencia de D-aminoácidos en estas cadenas peptídicas es de la mayor importancia porque prácticamente no se encuentran aminoácidos de este tipo en otras estructuras celulares procarióticas y están ausentes en las estructuras eucarióticas. La capa de peptidoglicano presenta un grosor variable según las especies bacterianas: aquéllas presentan una reacción Gram-positiva tienen una gruesa capa de peptidoglicano, mientras que las Gram-negativas tienen una capa prácticamente monomolecular. SÍNTESIS DE LA CAPA DE PEPTIDOGLICANO El peptidoglicano se sintetiza por la acción de un conjunto de enzimas situadas en la membrana interna de la bacteria. La actividad de estas enzimas tiene que estar perfectamente regulada porque simultáneamente han de abrir nuevos sitios de crecimiento de la pared celular cuidando de que no se produzcan fallos en la contención de la presión osmótica intracelular. Existe una gran cantidad de antibióticos cuya actividad se debe a la inhibición selectiva de estas moléculas de síntesis de pared celular que provocan rápidamente el estallido (lisis) de la célula bacteriana. Algunos actúan en etapas tempranas de la síntesis (cicloserina, vancomicina, bacitracina) mientras que otros, que contienen un núcleo químico conocido como anillo β-lactámico y, por ello, se denominan antibióticos βlactámicos de los que el ejemplo representativo es la penicilina, actúan en las etapas finales. Estos antibióticos sólo son efectivos cuando la célula está creciendo y, por consiguiente, las proteínas de síntesis peptidoglicano son activas. Dada la presencia de aminoácidos D en la capa de peptidoglicano, las enzimas responsables de su síntesis han de tener sus centros activos diseñados de forma que puedan reconocer los aminoácidos de esta configuración. Los antibióticos que inhiben estas enzimas son totalmente selectivos para bacterias ya que la estructura del peptidoglicano no se encuentra en otro tipo de células. Las proteínas responsables de las etapas finales de la síntesis de peptidoglicano se conocen como PBP's, del inglés Penicillin-Binding Proteins que hace referencia a su capacidad de unir penicilina. FUNCIONES DEL PEPTIDOGLICANO Protección osmótica: Las células bacterianas se encuentran en una situación de riesgo osmótico permanente: la alta concentración de los solutos intracelulares crea una gran presión osmótica que llevaría a las células a estallar si no contaran con una cubierta de protección. En este sentido, las funciones de la capa de peptidoglicano son dos: a.- preservar la integridad de la célula de manera que resista la presión de turgor b.- conferir una forma definida a la célula bacteriana 16 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) MEMBRANA EXTERNA Ciertas bacterias presentan una segunda membrana (membrana externa) por el exterior de la capa de peptidoglicano. Estas bacterias presentan una respuesta especial a la tinción de Gram y se clasifican como Gram-negativas. La membrana externa presenta ciertas diferencias en su estructura con las membranas clásicas. Las más relevantes son la presencia de un lipopolisacárido en su cara exterior y la presencia de porinas. El lipopolisacárido es una molécula compleja que proyecta hacia el exterior de la célula cadenas de polisacárido: su importancia radica en que es altamente antigénico (provoca una fuerte respuesta inmune, endotoxina) siendo uno de los agentes que se apuntan como responsables del llamado «shock endotóxico» producido cuando las células del sistema inmune entran en contacto con él. La función del lipopolisacárido en las bacterias es la de incapacitar las defensas del huésped, proporcionar carga negativa a la superficie de la membrana y estabilizarla. Las variantes de lipopolisacárido de diferentes cepas de una misma bacteria se pueden distinguir usando métodos serológicos. El antígeno de lipopolisacárido se conoce como antígeno O. En las bacterias Gram-positivas, carentes de membrana externa y, por tanto, de lipopolisacárido, una función equivalente (en la estabilidad de la membrana y en proporcionarle carga negativa) a la del lipopolisacárido la realizan los ácidos teicoicos. En la membrana externa se encuentran las porinas que abren vías de entrada de solutos al interior celular. Las porinas son complejos de varias moléculas de proteína que forman un canal por el que pueden atravesar la membrana externa moléculas de hasta 1000 Da de tamaño molecular. La membrana externa se encuentra unida al peptidoglicano a través de la proteína conocida como lipoproteína de Braun cuya parte lipídica se encuentra en la membrana y la proteica unida covalentemente a la mureína. La membrana externa tiene unos puntos de contacto con la membrana interna que se denominan uniones de Bayer y que se supone son importantes en el transporte a través e la membrana. ESPACIO PERIPLÁSMICO En las bacterias Gram-negativas, el espacio comprendido entre las membranas interna y externa se denomina espacio periplásmico y comprende un volumen que rodea a la célula conteniendo gran cantidad de enzimas que permiten procesar los nutrientes para que puedan ser transportados al interior celular a través de la membrana interna. Otras enzimas importantes presentes en el espacio periplásmico son las β-lactamasas responsables de la destrucción de los antibióticos β-lactámicos y, por tanto, de la resistencia de ciertas bacterias a ellos. 17 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) En las bacterias Gram-positivas no hay, en realidad, espacio periplásmico; pero se discute si la parte más interna del peptidoglicano puede desarrollar una función similar reteniendo mediante fuerzas electrostáticas moléculas de enzimas equivalentes a las periplásmicas de Gram-negativas. 7.- BACTERIAS GRAM-POSITIVAS Y GRAM-NEGATIVAS. Como se ha indicado anteriormente, la mayoría de las bacterias pueden clasificarse en uno de dos grupos diferenciables por medio de la tinción de Gram. Las bacterias Gram-positivas y las Gram-negativas. Estructuralmente ambos grupos se diferencian porque las Gram-positivas carecen de membrana externa, tienen una gruesa capa de peptidoglicano y presentan ácidos teicoicos en su superficie, mientras que las Gramnegativas tienen membrana externa portadora de lipopolisacárido y una capa de peptidoglicano más fina. En general, las bacterias Gram-negativas suelen presentar mayores problemas de permeabilidad a ciertos antibióticos que las Gram-positivas, debido a la doble membrana. Sin embargo, muchas bacterias Gram-positivas presentan otros mecanismos de resistencia (esporas) de los que carecen las Gram-negativas, y una mayor resistencia a antisépticos. Desde el punto de vista morfológico, las bacterias Gram-negativas suelen ser bacilos, comas (Vibrio) o espirilos, mientras que sólo en casos muy raros son cocos (Neisseria). Dentro de las bacterias Gram-positivas encontramos cocos y bacilos. 8.- RIBOSOMAS BACTERIANOS. Los ribosomas, tanto los procarióticos como los eucarióticos, están formados por proteínas y ARN; sin embargo, ambos tipos de constituyentes ribosómicos son diferentes en procariotas y eucariotas de suerte que puede disponerse de inhibidores (antibióticos) específicos de ribosomas procarióticos que no afectan a los eucarióticos y viceversa. 9.- ELEMENTOS FACULTATIVOS DE LA CÉLULA PROCARIOTA. Estas últimas cubiertas no están presentes en todos los microorganismos y, por consiguiente, no pueden considerarse elementos estructurales esenciales, aunque confieren a los microorganismos que las poseen características biológicas particulares. CÁPSULA Y CAPAS MUCOSA. Muchas bacterias presentan en la parte exterior de sus paredes celulares otras capas que sirven de protección y les facilitan la colonización de ambientes intracorporales. Entre estas capas se encuentran cubiertas proteicas que forman una especie de coraza denominada genéricamente capa S, y se encuentran capas de naturaleza polisacarídica denominadas genéricamente cápsulas. 18 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) La capa S está formada por proteínas y glicoproteínas y participa en la adhesión de las bacterias a superficies, la protección frente a la fagocitosis y actúa como barrera frente a enzimas o substancias que pudieran dañar a las bacterias que la poseen. Las cápsulas están formadas por polisacáridos o polipéptidos y participan en la adhesión de las bacterias a superficies, retardan la desecación de las bacterias en ambientes secos y proporcionan protección frente a la fagocitosis. No solo las bacterias presentan cápsulas sino que también han sido descritas en algunos hongos unicelulares (Cryptococcus neoformans). Una característica macroscópica fácilmente observable de los microorganismos con cápsula es que forman colonias de aspecto mucoso y liso. Las diferentes variantes de cápsula de distintas cepas de una misma especie se pueden identificar mediante métodos serológicos. El antígeno capsular se conoce como antígeno K. 10.- APÉNDICES BACTERIANOS: FLAGELOS Y FÍMBRIAS. Las bacterias pueden poseer una serie de apéndices celulares que desempeñan funciones diversas: FLAGELOS La mayoría de las bacterias móviles lo son por la acción de los flagelos: estructuras proteicas cuyas características pueden ser fácilmente detectadas por medios serológicos lo que permite la identificación de microorganismos o distintas cepas de una misma especie con facilidad. El antígeno flagelar se conoce como antígeno H. Las bacterias flageladas pueden tener entre uno y 20 flagelos por célula. Su composición es proteica y su tamaño es de unos 20 nm de diámetro y de entre 5 y 20 µm de longitud. La función de los flagelos es proporcionar movimiento a las bacterias. Cuando este movimiento se dirige hacia, o en dirección opuesta, a un punto determinado se denomina tactismo, distinguiéndose los tipos de tactismo por el agente atrayente o repetlente (fototactismo, quimiotactismo, etc.). Los flagelos pueden estar colocados alrededor de la célula (peritricos) o en los polos (polares o lofotricos). El tipo de localización de los flagelos se puede identificar estudiando la forma del movimiento de la célula. FIMBRIAS Son pequeñas fibras de naturaleza proteica denominadas que se encuentran en la superficie de muchas especies de bacterias. (entre ellas muchas patógenas). Su número varía entre 100 y 1000 por bacteria y su tamaño entre 2 a 9 nm de diámetro y 1 a 5 µm 19 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) de longitud. Estas estructuras son de gran importancia en la adhesión de la célula bacteriana a las superficies que van a colonizar. PELO F Es un tipo especial de fimbria producido por bacterias capaces de transmitir su información genética a otras mediante conjugación bacteriana. Cuando está presente hay sólo uno por célula. Su naturaleza es proteica. Su longitud llega a alcanzar las 10 µm. PROLONGACIONES DE ADHESIÓN Algunos tipos de microorganismos son portadores de prolongaciones con forma de ventosa que les permiten adherirse a las células animales que infectan. Esto ocurre, por ejemplo, en ciertos micoplasmas. 11.- MATERIAL DE RESERVA Las bacterias acumulan materiales de reserva en forma de inclusiones de polihidroxibutirato, polifosfato, gránulos de azufre, etc. 12.- ENDOSPORAS Las endosporas (esporas) son formas de resistencia (a la temperatura, agentes químicos y físicos, desecación, etc.) que presentan algunas bacterias Gram-positivas (Bacillus, Clostridium). La producción de una endospora por una bacteria es un proceso lento que requiere un tiempo considerable (horas). Se inicia como respuesta a condiciones ambientales adversas y culmina con la muerte de la célula madre de la endospora y la liberación de esta. En las esporas el material citoplásmico se ha desecado al máximo y han acumulado ciertas moléculas (ácido dipicolínico) que les dotan de una gran termorresistencia. La pared celular de las esporas es de un tipo de peptidoglicano modificado. La forma y posición de la endospora en la célula productora proporciona información sobre qué especie es. Por consiguiente, tiene valor taxonómico. En general, puede decirse que las esporas son de forma esférica y suelen aparecer libres, aunque en ocasiones se pueden observar en el interior de las bacterias a las que deforman de una manera característica, lo que sirve para su identificación (Clostridium). 14.- ESTRUCTURA DE LOS VIRUS. Los virus son un complejo grupo de estructuras que sólo desarrollan una actividad metabólica cuando se encuentran en el interior de una célula huésped. Por consiguiente, se trata de parásitos obligados que no tienen vida libre. Las partículas de virus libres metabólicamente inactivas se conocen como viriones. Los virus están formados por una o varias moléculas de material genético (ADN o ARN, monocatenario o bicatenario) recubiertas de una capa de proteínas que forma la 20 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) cápsida que, a su vez, puede estar recubierta por otras estructuras membranosas en ciertos tipos de virus. La forma de los viriones viene determinada por la que adquiere la cápsida y, generalmente, adopta tres tipos: forma icosaédrica (por ejemplo, los adenovirus), forma helicoidal (por ejemplo, el virus del mosaico del tabaco) o alargada (por ejemplo, el virus Ébola). Dos excepciones a estas formas son algunos virus bacterianos (bacteriofagos) que tienen cápsulas complejas, y los rhabdovirus (virus de la rabia). Algunos virus tienen unas estructuras en forma acicular denominadas peplómeros que les permiten adherirse a las células huésped que infectan. 15.- PRIONES En ciertos tipos de enfermedades neurodegenerativas tales como el Alzehimer, Parkinson, corea de Huntington y encefalopatías transmisibles tales la enfermedad de Creufeldt-Jakob, el Insomnio Familiar Fatal, la encefalopatía bovina transmisible y el kuru) se observa la formación de depósitos de proteínas en el cerebro de los pacientes afectados. Estos depósitos se conocen como amiloides34, y están formados por agregados de proteínas derivadas del tejido nervioso del paciente que se agregan por tener una estructura terciaria (tridimensional) alterada (no nativa). Parece ser que la transición de la forma normal no ensamblable la proteína a la forma alterada patógena y ensamblable parece estar provocada por la presencia de esta última que induce el cambio conformacional en las proteínas nativas. Se denominan priones a estas proteínas infectivas producidas por un procesamiento anormal de algunas proteínas de la membrana de las células eucarióticas. Como consecuencia de dicho procesamiento anormal, los priones polimerizan en tejidos tales como el nervioso dando lugar a diferente patologías. Existen varios tipos de priones cuya secuencia de aminoácidos no tiene relación entre sí. Es decir: lo característico del prión es que puede sufrir la alteración estructural para dar lugar al agregado amiloide, no la secuencia en sí del prión. Un ejemplo de prión es la proteína PrP que en la forma nativa (PrPsen) no polimeriza y es sensible a proteasas y en la alterada (PrPres) polimeriza y es insensible. La presencia de PrPres induce el cambio de PrPsen a adoptar la forma PrPres y formar el amiloide. La proteína PrP es una glicoproteína de membrana codificada en nuestro cromosoma 20 y cuya función no está clara. La enfermedad de Creufeldt-Jakob es hereditaria y dominante, los afectados tienen una copia de los dos genes de PrP que codifica una forma variante en la que la substitución de un a minoácido da lugar al desarrollo de la enfermedad. La inyección de PrPres en animales sanos, también produce el desarrollo de la enfermedad, a no ser que se trate de animales en los que se ha eliminado experimentalmente el gen PrP (ver los Postulados de Kock). Inicialmente los priones eran considerados como virus de desarrollo extremadamente lento (lentivirus) puesto que las patologías que producían tardaban en manifestarse mu3 Nature 437: 197-198 Existen algunas otras situaciones en las que se producen depósitos amiloides, tales como la formación de fibrillas de β2-microglobulina en pacientes dializados. No jay que confundir esto con la patología causada por priones. 4 21 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) chos años. En la actualidad, su naturaleza proteica está claramente aceptada por la comunidad científica y deben ser estudiados independientemente de los virus. 22 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Tema 2.- Crecimiento y muerte de microorganismos. Concepto y expresión matemática del crecimiento bacteriano. Concepto de muerte de un microorganismo. ¿Qué necesita un microorganismo para crecer?. Detección y medida del crecimiento. Ciclo de crecimiento de poblaciones. Factores físicos y químicos que influyen en el crecimiento. Multiplicación de los virus. 1.- CONCEPTO Y EXPRESIÓN MATEMÁTICA DEL CRECIMIENTO BACTERIANO. Entendemos por crecimiento microbiano el aumento del número de microorganismos a lo largo del tiempo. Por tanto, no nos referimos al crecimiento de un único microorganismo (ciclo celular) sino al demográfico de una población. En este tema nos centraremos en el crecimiento de bacterias, el estudio que se hace puede servir también para entender el crecimiento de levaduras y de otros hongos. El crecimiento de los virus se produce de otra forma diferente y será tratado al final de este capítulo. Denominamos ciclo celular al proceso de desarrollo de una bacteria considerada de forma aislada. A lo largo del ciclo celular, tiene lugar la replicación del material de la bacteria, la síntesis de sus componentes celulares, el crecimiento para alcanzar un tamaño doble del inicial y su división por bipartición de la bacteria para dar lugar a dos células hijas. La duración del ciclo celular coincide con el tiempo de generación y depende, en general, de los mismos factores de los que depende este. El crecimiento de una población resulta de la suma de los ciclos celulares de todos sus los individuos. Este crecimiento suele ser asincrónico puesto que cada microorganismo se encuentra en un punto diferente del ciclo celular. Por consiguiente, en un momento determinado en una población se encuentran células que acaban de dividirse, otras que están replicando su ADN y elongándose, otras que están iniciando la división celular, etc. En un crecimiento sincrónico todas las células se encuentran simultáneamente en la misma fase del crecimiento celular. Los cultivos sincrónicos son muy difíciles de mantener por lo que su importancia está principalmente ligada a los estudios básicos de biología microbiana. Sin embargo, en la naturaleza, las bacterias del suelo se encuentran en condiciones de crecimiento próximas a la fase estacionaria (en la que se produce una cierta sincronización del cultivo) y, por consiguiente, durante cierto tiempo las poblaciones naturales probablemente se comporten como relativamente sincrónicas. Las poblaciones de bacterias pueden crecer de una forma explosiva acumulando grandes números en un periodo de tiempo muy reducido. Puesto que el efecto nocivo (infecciones o intoxicaciones) de los microorganismos depende de su número en la mayoría de los casos, entender cómo se produce el crecimiento microbiano es importante para poder evitar o reducir dichos efectos nocivos. Se denomina crecimiento equilibrado a aquél en el que todos la biomasa, número de células, cantidad de proteínas, de ADN, etc., evolucionan en paralelo. El crecimiento equilibrado probablemente ocurra en muy contadas ocasiones en condiciones naturales. Por tanto, es principalmente un concepto de aplicación en el laboratorio. Sin embargo, es útil porque permite estudiar el crecimiento microorganismos. 23 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Las bacterias crecen siguiendo una progresión geométrica en la que el número de individuos se duplica al cabo de un tiempo determinado denominado tiempo de generación (τ). De esta forma, podemos calcular el número de bacterias (N) al cabo de un número de generaciones (n) usando la ecuación siguiente: N = N0 2n siendo N0 el número de células en el momento actual. El número de generaciones se puede calcular de la siguiente forma: n=t/τ donde t es el tiempo transcurrido. Los tiempos de generación de bacterias creciendo en ambientes favorables pueden ser muy cortos (valores de τ de 20 min). Esto lleva a que una única célula (N0 = 1) creciendo con un τ = 20 min, llegue a poder producir 4.7 x 1021 células en 24 horas. Si la bacteria crece en un medio líquido, las células que se producen en cada división continúan su vida independientemente en la mayoría de los casos formando una suspensión de células libres. Cuando una célula aislada comienza a crecer sobre un substrato sólido, el resultado del crecimiento al cabo del tiempo es una colonia. Se denomina unidad formadora de colonia (UFC) a una célula viva y aislada que se encuentra en un substrato y en condiciones ambientales adecuadas y produce una colonia en un breve lapso de tiempo. Una UFC también puede corresponder a más de una célula cuando éstas forman parte de grupos unidos fuertemente (estreptococos o diplococos, por ejemplo) ya que cada grupo formará una sola colonia. Cuando algunos tipos de bacterias o de levaduras patógenas crecen sobre superficies forman biopelículas5 (biofilms) en los que las células se asocian entre sí mediante capas de polisacáridos que forman una película que recubre la superficie sobre la que se encuentran las células. Los biofilms son importantes porque los microorganismos que los forman resultan más resistentes a antibióticos y al ataque de células del sistema inmune y, por consiguiente, las infecciones que producen son más difíciles de tratar. Además, dentro de los biofilms (que pueden estar formados por microorganismos de diferentes especies) se facilita la transferencia horizontal de genes entre lo que facilita la dispersión de factores de virulencia6 o de resistencias a antibióticos7. La presencia de biopelículas es un problema serio en los implantes ortopédicos, catéteres, etc. El sarro de los dientes es un ejemplo de biofilm. 5 Cuando las bacterias crecen como unidades aisladas se habla de crecimiento planctónico para diferenciarlo del crecimiento formando un biofilm 6 Se denominan factores de virulencia a las características de un microorganismo que le permiten causar una enfermedad. Los factores pueden ser proteínas que faciliten el ataque al huésped, que permitan resistir la acción del sistema inmune, etc. 7 ASM News 71: 412-419 24 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) La formación de biofilms es importante en la génesis de diferentes patologías. Se están identificando genes que controlan el cambio de crecimiento libre (crecimiento planctónico) a crecimiento en forma de biofilm en distintos tipos de bacterias. Por ejemplo, se han identificado genes de este tipo en Pseudomnas aeruginosa que son responsables del aumento de la cronicidad de las infecciones con este patógeno8. En otros microorganismos (tales como Streptococcus mutans) la formación de biofilms se ha asociado a la presencia de señales de quórum (ver el párrafo siguiente) de manera que bacterias competidoras de S. mutans son capaces de eliminr estas señales y reducir la formación de la biopelícula. Algunas bacterias que comparten un mismo espacio físico pueden comunicase entre sí mediante lo que se denomina señales de quórum. Estas son moléculas simples excretadas por las células. Cuando el número de células aumenta, la concentración de la señal de quórum también lo hace. Las células tienen receptores para estas señales de quórum que reaccionan en función de la concentración de la señal. De esta forma, cuando la concentración de la señal de quórum es mayor, la respuesta es mayor. De esta forma, el comportamiento de una célula depende de que se encuentre sola o acompañada. Las señales de quórum pueden ser reconocidas por especies diferentes de las emisoras. También hay bacterias capaces de degradar las señales de quórum de otras especies para interferir su comunicación9. El estudio de las señales de quórum permite desarrollar productos que dificultan su formación. Por ejemplo, dentífricos con productos anticarro10. 2.- CONCEPTO DE MUERTE DE UN MICROORGANISMO. Desde el punto de vista microbiológico, un microorganismo muere cuando pierde de forma irreversible la capacidad de dividirse. El fundamento de esta definición es que si un microorganismo ha perdido la capacidad de dividirse no podrá formar una colonia sobre un medio de cultivo y no será posible detectar su presencia por los métodos microbiológicos tradicionales. Es decir: cuando no se produce aumento en el número de microorganismos no hay crecimiento. Sin embargo, un microorganismo puede estar muerto desde el punto de vista microbiológico y continuar desarrollando una actividad metabólica que se traduzca, por ejemplo, en liberación de toxinas. Por otra parte, hay que considerar que la capacidad de multiplicación (crecimiento) de un microorganismo puede verse transitoriamente afectada por lesiones o por las condiciones físicas o químicas del entorno. En estos casos, podríamos considerar como muertos microorganismos que pueden reanudar su crecimiento si las condiciones son de nuevo favorables. 3.- ¿QUÉ NECESITA UN MICROORGANISMO PARA CRECER? 8 J. Bacteriology 187: 1441-1454. Por ejemplo, parece ser que Escherichia coli es capaz de interferir la señal de quórum que permite a la bacteria patógena Vibrio cholerae calcular el tamaño de la población. Esto es importante porque cuando la población de V. Cholerae es grande, se inhibe la producción de la toxina colérica y el patógeno se desprende del intestino para buscar nuevos individuos que infectar. Por tanto, la acción de E. coli afecta la patogenicidad de V. Cholerae. 10 ASM News 71: 129-135 9 25 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) El aislamiento de bacterias a partir de muestras naturales se realiza, en la mayoría de los casos, mediante la producción de colonias aisladas en cultivos sólidos. El crecimiento explosivo de las bacterias produce un gran número a partir de una única célula inicial de forma que, tras un periodo de tiempo de incubación en las condiciones ambientales adecuadas, se produce una colonia de individuos iguales. Para crecer, un microorganismo necesita nutrientes que le aporten energía y elementos químicos para la síntesis de sus constituyentes celulares. Dependiendo de la fuente de carbono que utilizan, los microorganismos se pueden clasificar en autotrofos si es el CO2 atmosférico (microorganismos que fotosintetizan) y heterotrofos si utilizan carbono orgánico. Los microorganismos de importancia clínica son todos ellos heterotrofos. La fórmula elemental de un microorganismo es, aproximadamente, C4H7O2N lo que supone que los componentes de las células son: carbono que representa alrededor del 50% del peso seco, oxígeno (32%), nitrógeno (14%) y debe estar disponible, normalmente, en forma de NH4 o de aminoácidos a los que se pueda tomar su grupo amino; fósforo (3%) y debe estar en forma de PO43-, azufre que representa en torno al 1% y procede de aminoácidos sulfurados o de SO42-; y otros elementos traza entre los que se encuentran Fe, K, Mg, Mn, Co, Mb, Cu y Zn. La elaboración de medios de cultivo que permitan aislar microorganismos a fin de iniciar posteriores cultivos puro o axénicos requiere proporcionar los nutrientes antes citados y, en ciertos casos, algunos aminoácidos o vitaminas que determinados tipos de microorganismos no pueden sintetizar. Los medios de cultivo se pueden clasificar en definidos cuando su composición química se conoce totalmente y complejos cuando no es el caso porque están compuestos por mezclas de extractos de materiales complejos (extracto de levadura, extracto de carne, etc.). Por otra parte, los medios de cultivo pueden ser líquidos o bien sólidos si se añade algún agente solidificante que no sea consumible por los microorganismos (normalmente agar). En función de los microorganismos que pueden crecer en ellos, los medios pueden ser generales, selectivos cuando favorecen el crecimiento de ciertos microorganismos mientras suprimen el de otros (por ejemplo, el medio SPS para clostridios), diferenciales cuando alguno de sus componentes permite identificar las colonias de un tipo de microorganismos (por ejemplo medios con hematíes para identificar colonias de microorganismos hemolíticos), selectivo-diferenciales cuando combinan las dos características anteriores (por ejemplo, el agar de MacConkey para identificar Escherichia coli), y medios de enriquecimiento que permiten aislar un tipo determinado de microorganismo a partir de una mezcla una población mixta de gran tamaño. 26 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) 4.- DETECCIÓN Y MEDIDA DEL CRECIMIENTO Existen diferentes sistemas para detectar y medir el crecimiento de microorganismos. Los principales, se enumeran a continuación: Recuento directo: consiste en la observación al microscopio de volúmenes muy pequeños de suspensiones de bacterias. Se usan unos portaobjetos especiales denominados cámaras de Petroff-Hausser. Para que la medida sea correcta es necesario que la densidad de células sea del orden de 105 por ml. Medida de la masa de células: el sistema se basa en que las células en suspensión dispersan la luz causando la turbidez del cultivo. La turbidez depende de la masa en suspensión y, por tanto, midiendo esta se puede estimar aquella. Este es el parámetro de medida más fácil de usar en los cultivos de laboratorio. La densidad de células debe ser del orden de 105 por ml. Recuento de viables: consiste en sembrar un volumen determinado de cultivo o muestra sobre el medio de cultivo sólido adecuado para estimar el número de viables contando el número de colonias que se forman puesto que cada una de estas deriva de una UFC. Para que la medida sea correcta desde el punto de vista estadístico, es necesario contar más de 300 UFC. En ciertas ocasiones en las que la densidad de microorganismos es demasiado baja, éstos se pueden recolectar por filtración a través de una membrana (de 0.2 µm de tamaño de poro) y posterior colocación de la membrana en un medio de cultivo adecuado para que se formen las colonias. Medida del número de partículas usando contadores electrónicos de partículas. Estos sistemas no nos indican si las partículas corresponden a células vivas o muertas; pero nos pueden dar una idea del tamaño de las partículas. Medida de parámetros bioquímicos tales como la cantidad de ADN, ARN, proteínas, peptidoglicano, etc. por unidad de volumen. Medida de actividad metabólica de las bacterias como que respiran producen una disminución del potencial redox del medio en que se encuentran como consecuencia del consumo de oxígeno (utilización de colorantes sensibles a oxidación-reducción tales como el azul de metileno). 5.- CICLO DE CRECIMIENTO DE POBLACIONES. En un cultivo bacteriano en medio líquido, se pueden diferenciar cuatro fases en la evolución de los parámetros que miden el crecimiento microbiano: Fase lag o de adaptación: Durante la que los microorganismos adaptan su metabolismo a las nuevas condiciones ambientales (de abundancia de nutrientes) para poder iniciar el crecimiento exponencial. 27 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Fase exponencial o logarítmica: en ella la velocidad de crecimiento es máxima y el tiempo de generación es mínimo. Durante esta fase las bacterias consumen los nutrientes del medio a velocidad máxima. La evolución del número de células durante esta fase se explica con el modelo matemático descrito anteriormente. Esta fase corresponde a la de infección y multiplicación dentro del organismo del agente infeccioso. Fase estacionaria: en ella no se incrementa el número de bacterias (ni la masa u otros parámetros del cultivo). Las células en fase estacionaria desarrollan un metabolismo diferente al de la fase de exponencial y durante ella se produce una acumulación y liberación de metabolitos secundarios que pueden tener importancia en el curso de las infecciones o intoxicaciones producidas por bacterias. Los microorganismos entran en fase estacionaria bien porque se agota algún nutriente esencial del medio, porque los productos de desecho que han liberado durante la fase de crecimiento exponencial hacen que el medio sea inhóspito para el crecimiento microbiano o por la presencia de competidores u otras células que limiten su crecimiento. La fase estacionaria tiene gran importancia porque probablemente represente con mayor fidelidad el estado metabólico real de los microorganismos en muchos ambientes naturales. Fase de muerte: se produce una reducción del número de bacterias viables del cultivo. Las fases, parámetros y cinética de crecimiento discutidas para el caso de los medios líquidos se presentan también en los sólidos. La cinética de crecimiento, en este caso, sólo se puede seguir utilizando unos sistemas de detección especiales siendo el más sencillo, la medida del número de células viables por unidad de superficie o por unidad de masa. 6.- FACTORES FÍSICOS Y QUÍMICOS QUE INFLUYEN EN EL CRECIMIENTO. 28 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Temperatura: Cada microorganismo tiene una temperatura de crecimiento adecuada. Si consideramos la variación de la velocidad de crecimiento en función de la temperatura de cultivo, podemos observar una temperatura mínima por debajo de la cual no hay crecimiento; a temperaturas mayores se produce un incremento lineal de la velocidad de crecimiento con la temperatura de cultivo hasta que se alcanza la temperatura óptima a la que la velocidad es máxima. Por encima de esta temperatura óptima, la velocidad de crecimiento decae bruscamente y se produce la muerte celular. El aumento de la velocidad de crecimiento con la temperatura se debe al incremento generalizado de la velocidad de las reacciones enzimáticas con la temperatura. Se denomina coeficiente de temperatura a la relación entre el incremento de la velocidad de reacción y el de temperatura. En términos generales, la velocidad de las reacciones bioquímicas suele aumentar entre 1.5 y 2.5 veces al aumentar 10ºC la temperatura a la que tienen lugar. La falta de crecimiento a temperaturas bajas se debe a la reducción de la velocidad de las reacciones bioquímicas y al cambio de estado de los lípidos de la membrana celular que pasan de ser fluidos a cristalinos impidiendo el funcionamiento de la membrana celular. La muerte celular a altas temperaturas se debe a la desnaturalización de proteínas y a las alteraciones producidas en las membranas lipídicas a esas temperaturas. Es importante tener en cuenta que a temperaturas bajas, el metabolismo celular es lento y las células paran de crecer; aunque suelen morir. Sin embargo, cuando la temperatura es superior a la óptima, se produce la muerte celular rápidamente y las células no pueden recuperar su capacidad de división si baja posteriormente la temperatura. Esto permite esterilizar por calor y no por frío. Hay varios tipos de microorganismos en función de sus temperaturas de crecimiento mínima, máxima y óptima. Tipo de Temperatura Temperatura Temperatura microorganismo mínima óptima máxima Psicrófilo -5 +5 12 - 15 15 - 20 Psicrótrofo -5 +5 25 - 30 30 - 35 Mesófilo 5 - 15 30 - 45 35 - 47 Termófilo 40 - 45 55 - 75 60 - 90 Los microorganismos psicrótrofos son mesófilos que pueden crecer a temperaturas bajas. Por tanto, se les puede considerar como psicrófilos facultativos. Esto es importante desde el punto de vista aplicado porque cuando se encuentran contaminando alimentos, son capaces de crecer en condiciones de refrigeración (4 - 8ºC) y de producir infecciones en los consumidores del alimento (30 - 35 ºC). Desde el punto de vista clínico, los microorganismos capaces de producir infecciones en pacientes son los mesófilos y algunos psicrótrofos ya que sus temperaturas óptimas de crecimiento coinciden con las corporales. 29 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Actividad de agua (aw): Se denomina actividad de agua a la relación entre la presión de vapor de agua del substrato de cultivo (P) y la presión de vapor de agua del agua pura (P0). El valor de la actividad de agua está relacionado con el de la humedad relativa (HR). El valor de la actividad de agua nos da una idea de la cantidad de agua disponible metabólicamente. Por ejemplo: comparemos el agua pura donde todas las moléculas de agua están libremente disponibles para reacciones químicas con el agua presente en una disolución saturada de sal común (NaCl) donde una parte importante de las moléculas de agua participa en la solvatación de los iones de la sal disuelta. En este último caso, la actividad de agua mucho menor que en el primero. conforme aumenta la cantidad de solutos en el medio, disminuye su actividad de agua. El agua es un substrato en muchas reacciones bioquímicas (proteasas y lipasas, por ejemplo). Cuando no hay agua disponible, estas reacciones se detienen y el metabolismo se para. Esta falta de agua también detiene muchas de las enzimas que podrían degradar las estructuras biológicas. Por ello, las células que no crecen por falta de agua no mueren rápidamente: los sistemas de degradación tampoco funcionan y no las degradan. Es decir: cuando un microorganismo se encuentra en un substrato con actividad de agua menor que la que necesita, su crecimiento se detiene. Esta detención del crecimiento no suele llevar asociada la muerte del microorganismo, sino que éste se mantiene en condiciones de resistencia durante un tiempo más o menos largo. En el caso de las esporas, la fase de resistencia puede ser considerada prácticamente ilimitada. La gran mayoría de los microorganismos requiere valores de actividad de agua muy altos para poder crecer. Los valores mínimos de actividad para diferentes tipos de microorganismos son, a título orientativo, los siguientes: bacterias aw>0.90, levaduras aw>0.85, hongos filamentosos aw>0.80. Como puede verse, los hongos filamentosos son capaces de crecer en substratos con actividad de agua menor (más secos) de la que permite el crecimiento de bacterias o de levaduras. Por esta razón se puede producir la colonización de substratos con baja actividad de agua (por ejemplo, alimentos como el queso o almíbares o la piel) por mohos (hongos filamentosos) mejor que por bacterias. En función de su tolerancia a ambientes con baja aw, los microorganismos que pueden crecer en estas condiciones se clasifican en halotolerantes, halófilos y xerófilos según toleren o requieran condiciones salinas o hipersalinas, respectivamente. La reducción de la actividad de agua para limitar el crecimiento bacteriano tiene importancia aplicada en industria alimentaria. La utilización de almíbares, salmueras y salazones reduce la actividad de agua del alimento para evitar su deterioro bacteriano. pH: Es un parámetro crítico en el crecimiento de microorganismos ya que cada tipo de microorganismo tiene un rango de pH en el que puede vivir adecuadamente, fuera de este rango muere. El pH intracelular es ligeramente superior al del medio que rodea las células ya que, en muchos casos, la obtención de energía metabólica depende de la existencia de una diferencia en la concentración de protones a ambos lados de la membrana citoplásmica. El pH interno en la mayoría de los microorganismo está en el rango de 6,0 a 8,0. 30 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Los rangos de pH tolerables por diferentes tipos de microorganismos son, también, distintos. Hay microorganismos acidófilos que pueden vivir a pH=1.0 y otros alcalófilos que toleran pH=10.0 Hay que considerar que, como consecuencia del metabolismo, el pH del medio de crecimiento suele tender a bajar durante el cultivo. Por otra parte, la bajada del pH del medio que producen ciertos microorganismos les confiere una ventaja selectiva frente a otros competidores. Así, por ejemplo, las bacterias lácticas que producen grandes cantidades de ácido láctico como consecuencia de su metabolismo primario reducen el pH del medio a valores inferiores a los soportables por otras bacterias competidoras (llegan a bajar el pH del medio hasta 4.5). De esta forma, las bacterias competidoras mueren y las lácticas se convierten en la población dominante. La bajada del pH se puede deber a varios factores, uno de los cuales es la liberación de ácidos orgánicos de cadena corta (fórmico, acético, láctico) por ciertas bacterias. En este sentido, hay que tener en cuenta que la acción bactericida de estos ácidos orgánicos de cadena corta es más potente que la debida únicamente a la bajada del pH que producen. Esto es, los ácidos orgánicos de cadena corta son tóxicos para algunas bacterias por sí mismos. El efecto letal del pH ácido sobre los microorganismos tiene aplicación en la conservación de alimentos acidificándolos. De esta forma, la adición de ácido acético en forma de vinagre permite la conservación de alimentos perecederos (escabeches, por ejemplo) y la producción de ácidos en el curso de fermentaciones naturales permite alargar la vida de los alimentos (coles fermentadas, por ejemplo). Potencial redox: nos indica la capacidad del substrato para aceptar o donar electrones, esto es: sus características oxidantes o reductoras. Uno de los factores que intervienen en el potencial redox, aunque no el único, es la concentración de oxígeno [O2]. Hay microorganismos que requieren ambientes oxidantes para crecer, mientras que otros necesitan ambientes reductores. El metabolismo de ambos tipos de microorganismos presenta diferencias notables. El requerimiento de condiciones oxidantes o reductoras no debe confundirse con la necesidad de presencia o ausencia de oxígeno para que se produzca el crecimiento. En general, cuando un microorganismo requiere un ambiente oxidante se dice que desarrolla un metabolismo oxidativo (o respirativo) mientras que los microorganismos que requieren ambientes reductores (o menos oxidantes) realizan un metabolismo fermentativo. Un microorganismo es aerobio cuando necesita oxígeno para vivir y es anaerobio cuando o bien no lo necesita (anaerobios facultativos como las bacterias entéricas, o como Saccharomyces cerevisiae; o anaerobios aerotolerantes como las bacterias lácticas) o cuando muere en presencia de oxígeno (anaerobios estrictos como los clostridios). 31 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Hay microorganismos que, aunque viven en presencia de oxígeno, no son capaces de utilizarlo como aceptor final de electrones y deben desarrollar un metabolismo fermentativo (los estreptococos, por ejemplo.) Por otra parte, hay microorganismos que pueden desarrollar ambos tipos de metabolismo. Esto es: en presencia de oxígeno desarrollan un metabolismo oxidativo y en su ausencia, fermentativo. El rendimiento de los procesos fermentativos es menor que el de los respirativos: las bacterias y las levaduras crecen menos cuando lo hacen fermentando que cuando lo hacen respirando. En el curso de ciertas reacciones metabólicas redox se forman compuestos altamente reactivos (radicales libres, formas superóxido) que pueden dañar las proteínas, membranas y ácidos nucleicos produciendo la muerte de las células. Las células se defienden de estos compuestos reactivos mediante las enzimas siguientes: Superóxido dismutasa (SOD) y catalasa. Los anaerobios estrictos carecen de SOD y de catalasa o tienen niveles muy bajos de estas enzimas de forma que no pueden sobrevivir en presencia de oxígeno. La detección de estas enzimas tiene valor taxonómico. 7.- MULTIPLICACIÓN DE LOS VIRUS. Los virus tienen un ciclo celular diferente al de los microorganismos de vida libre y por consiguiente, su forma de crecimiento y la cinética del proceso es diferente. Una vez que un virus infecta una célula (fases de adsorción y penetración) puede comenzar su proceso de multiplicación que requiere la liberación del material genético del virus (desnudamiento) síntesis de una serie de enzimas, la multiplicación de su material genético y la síntesis de las proteínas de su cápsida (síntesis vírica), el ensamblaje de nuevos viriones (maduración) y la liberación de estos al exterior. Por consiguiente, desde el ingreso del virión en la célula huésped hasta la liberación de los nuevos viriones producidos como resultado de la infección, transcurre un periodo de tiempo en el que no es posible detectar la presencia de partículas víricas fuera de las células. A este periodo se le denomina fase de eclipse. Las fases de eclipse separan liberaciones explosivas de viriones. 32 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Tema 3.- Taxonomía microbiana. Taxonomía bactriana. Principales grupos de Bacterias, Virus y Hongos de importancia clínica. TAXONOMÍA BACTERIANA El Manual Bergey establece que el Reino Procariotae está formado por microorganismos caracterizados por carecer de membrana nuclear y por tener ribosomas 70S. A este Reino pertenecen las bacterias. El orden en que se presentan aquí los diferentes grupos es distinto del seguido para su descripción de los microorganismos de importancia clínica. El objetivo de este resumen es ayudar a organizar de forma complementaria a la anterior la información que se presenta allí. Se han señalado en amarillo los grupos bacterianos descritos particularmente en el tema de taxonomía. En Microbiología Médica interesan las siguientes secciones de bacterias: 1.- Espiroquetas Bacilos flexuosos en forma de espiral con movilidad debida a un filamento axial y semejante a un sacacorchos. Son Gram-negativos. Comprende los órdenes y familias siguientes: Orden Spirochaetales Familia Spirochaetaceae Género Treponema: T. pallidum Género Borrelia: B. recurrentis Familia Leptospiraceae: Género Leptospira: L. interrogans 2.- Bacterias Gram-negativas aerobias o microaerofilas (vibrioides): Bacilos espirales curvados y móviles. Parásitos del hombre y animales. Género Campylobacter: Especies: C. fetus, C. jejuni y C. coli Género Helicobacter. Especie: H. pylori. 3.- Bacilos y cocos Gram-negativos aerobios: Familia Pseudomonadaceae. Bacilos Gram-negativos, con flagelos polares. Producen citocromo y oxidasa. Género: Pseudomonas: P. aeruginosa Familia Moraxellaceae. Género Moraxella. Especies: M. lacunate, M catharralis. 33 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Género Acinetobacter Especie: A baunannii. Familia Xanthomonadaceae Género Stenotrophomonas. Especie: S. maltophila. Familia Burkholderia Especie B. cepacea Familia Legionellaceae. Bacilos móviles de vida libre. Patógenos para el hombre. Especie: L. pneumophila. Familia Neisseriaceae. Cocos agrupados en parejas con las zonas adyacentes planas. Especies patógenas. Género Neisseria Especies: N. meningitidis, N. gonorrhoeae. Género Kingella Especie: K. kingae Familia Bartonellaceae Género Bartonella Familia Brucellaceae. Cocobacilos Gram-negativos. Parásitos del hombre y de animales. Género Brucella. Especie: B. melitensis, B. abortus. Familia Bradyrhizobiaceae Género Afipia. Especie: A. felis Otros géneros de interés: Género Flavobacterium Especie: F. meningosepticum. Género Alcaligenes Especie: A. faecalis. Género Bordetella. Especie: B. pertussis. Género Francisella. Especie: F. tularensis. 4.- Bacilos Gram-negativos anaerobios facultativos: Familia Enterobacteriaceae. Bacilos Gram-negativos, no esporulados. Fermentan la glucosa con formación de ácido o de ácido y gas. Reducen los nitratos a nitritos y no poseen citocromo-oxidasa. Género Escherichia. Especie: E. coli Género Shigella. Especies: S. dysenteriae, S. sonney 34 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Género Salmonella. Especie: S. enterica, S. typhimurium, S. typhi, S. cholerasuis Género Citrobacter. Especie: C. freundii Género Klebsiella. Especie: K. pneumoniae Género Enterobacter. Especies: E. cloacae, E. aerogenes Género Serratia. Especie: S. marcescens Género Proteus. Especies: P. mirabilis, P. vulgaris Género Yersinia. Especies: Y. enterocolitica, Y. pseudotuberculosis, Y. pestis Género Morganella. Especie: M. morganii Género Providencia. Especies: P. rettgeri, P. stuartii Género Plesiomonas Otros géneros de interés Género Vibrio Especie: V. cholerae Género Aeromonas. Especie: A. hydrophila Género Pasteurella Especie: P. multocida Género Haemophilus. Especie: H. influenzae 5.- Bacilos Gram-negativos anaerobios: Bacilos Gram-negativos, anaerobios estrictos, no esporulados. Género Fusobacterium. Especie: F. nucleatum Género Bacteroides. Especie: B. fragilis Géneros Prevotella y Porphyromonas: antiguos bacteroides pigmentados 6.- Cocos Gram-negativos anaerobios: Son inmóviles. No esporulados. Familia Veillonellaceae. Género Veillonella Especie V. parvula 7.- Rickettsias y Chlamidias: Microorganismos pleomórficos que necesitan para su cultivo células vivas donde originan corpúsculos de inclusión. 35 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Género Rickettsia. Especies: R. Prowazecki, R. Connori, R. Typhi Género Coxiella. Especie: C. burnetti Familia Chlamydiaceae. Género Chlamydia. Especie: C. trachomatis. Género Chlamydiophila Especie: C. pneumoniae, C. psittaci 8.- Micoplasmas: Carecen de pared celular. Son los menores microorganismos cultivables en medios sintéticos. Familia Mycoplasmataceae. Género Mycoplasma. Especies: M. pneumoniae, M. hominis Género Ureaplasma. Especie: U. urealyticum 9.- Cocos Gram-positivos: Aerobios – anaerobios facultativos. Género Micrococcus Especie: M. Luteus. Género Staphylococcus. Especies: Staph. Aureus, Staph. Epidermidis, Staph. Saprophyticus Género Streptococcus. Especies: S. Pyogenes, S. Pneumoniae, S. Agalactiae Género Enterococus Especie: E. Faecalis. Otros Estreptococos anaerobios estrictos. Género Peptococcus. Especie: P. Niger. Género Peptostreptococcus. Especie: P. Anaerobius Género Sarcina 10.- Bacilos Gram-positivos esporulados: Género Bacillus. Especies: B. anthracis, B. cereus Género Clostridium. Especies: C. tetani, C. botulinum, C. perfringens, C. difficile 011.- Otros bacilos Gram-positivos: Género Listeria Especie: L. monocytogenes 36 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Género Corynebacterium: bacilos Gram-positivos que tienden a agruparse Especie: C. diphtheriae Género Actynomyces: forman hifas o micelios rudimentarios. Especie: A. israelii Género Nocardia. Especie: N. Asteroides Familia Mycobacteriaceae. Bacilos ácido-alcohol-resistentes. Género Mycobacterium Especies: M. tuberculosis, M. leprae 1.- PRINCIPALES GRUPOS DE BACTERIAS DE IMPORTANCIA CLÍNICA. Las bacterias patógenas pueden clasificarse en tres grupos: patógenos penetrantes extracelulares, patógenos intracelulares y patógenos toxigénicos. PATÓGENOS PENETRANTES EXTRACELULARES Los patógenos penetrantes extracelulares pueden producir toxinas; pero las enfermedades que causan se deben sobre todo a que invaden tejidos y crecen en su interior, más que a la producción diseminación de alguna toxina. Las bacterias penetrantes extracelulares en general producen infecciones agudas que incluyen la formación de exudado purulento (pus) y fiebre. En algunos casos la enfermedad se debe a la respuesta inflamatoria frente a la bacteria y en otros a eventos patógenos que se producen cuando la bacteria se disemina en la sangre o en los órganos que invade. En el caso de las bacterias Gram-negativas, el choque endotóxico debido a la presencia de lipopolisacárido es una complicación importante. Las bacterias penetrantes extracelulares más importantes son: Cocos y cocobacilos piógenos Streptococcus y Enterococcus Staphylococcus Neisseria Bordetella Haemophilus Moraxella Microorganismos entéricos Campylobacter Enterobacter Escherichia Helicobacter Klebsiella Proteus Salmonella Serratia Shigella Yersinia 37 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Pseudomonas A continuación describiremos brevemente las características de los grupos de microorganismos y las enfermedades que producen. ESTREPTOCOCOS Y ENTEROCOCOS Los estreptococos y los enterococos son un grupo grande de cocos Gram-positivos que se desarrollan en pares o en cadenas de cocos fuertemente unidos entre sí, lo que hace que en un recuento de viables las UFC correspondan en ocasiones a cadenas completas denominadas unidades estreptocócicas. Son bacterias anaerobias aerotolerantes que pertenecen al grupo de las bacterias lácticas por su capacidad para producir este ácido como resultado de la fermentación de glucosa. Los estreptococos se distinguen con claridad de otros tipos de cocos piógenos Grampositivos por su carácter de catalasa negativos. La mayoría de los estreptococos son inocuos; pero existen algunas especies que son intrínsecamente patógenas. Las especies menos virulentas pueden causar infecciones oportunistas. La clasificación de los enterococos y estreptococos se basa en su capacidad para producir hemolisis [bien de tipo α (verdosa) o β (clara)], y en las características de sus ácidos teicoicos que pueden ser determinadas mediante anticuerpos (sistema de clasificación de Lancefield). La mayoría de los patógenos humanos pertenecen al grupo A de Lancefield. Los patógenos más importantes del grupo son • Streptococcus pyogenes (β-hemolítico, Grupo A) • S. agalactiae (β-hemolítico, Grupo B) • S. pneumoniae (α-hemolítico, no presentan antígenos Lancefield) • Enterococcus faecalis (actividad hemolítica variable, Grupo D) 1.- S. pyogenes El hábitat natural de S. pyogenes son las vías respiratorias superiores de entre un 5 y un 20% de la población general. Las enfermedades producidas por este patógeno incluyen: 1) Enfermedades supurativas a) respiratorias i) faringitis ii) sinusitis b) piel i) escarlatina ii) piodermia iii) erisipela iv) fascitis necrosante c) síndrome estereptocócico tóxico 38 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) 2) Enfermedades no supurativas a) fiebre reumática b) enfermedad cardiaca reumática c) glomerulonefritis El tratamiento de S. pyogenes suele estar basado en Penicilina. A veces, este tratamiento se continúa hasta la edad adulta para prevenir recaídas en fiebres reumáticas. 2.- S. agalactiae El hábitat natural de S. agalactiae es el aparato digestivo, vagina y algunas fuentes animales tales como la leche cruda procedente de ovejas o vacas con mastitis. También puede comportarse como patógeno oportunista. Las enfermedades que produce se relacionan con la infección de los recién nacidos durante el parto a causa de bacterias presentes en la vagina y región perineal de la madre. Las enfermedades que producen son: 1) enfermedad neonatal (meningitis, neumonía y sepsis) 2) sepsis puerperal 3) faringitis El tratamiento se basa en Ampicilina administrada a madres portadoras en el momento del parto. 3.- S. pneumoniae El hábitat natural de S. pneumoniae (neumococo) son las vías respiratorias superiores de entre el 5 y el 70% de la población. El ser humano es el único reservorio conocido de este patógeno. La forma de del neumococo es muy característica: diplococos en forma de punta de flecha. Un factor de virulencia principal de este microorganismo es la presencia de una cápsula polisacarídica que permite la serotipificación de los aislados (serotipo K.) Hay descritos en torno a 20 serotipos de los que siete son los más frecuentes y representan en torno al 83% de los casos de infección en EE.UU. La infección por neumococos es especialmente grave en niños que no tienen completamente desarrollado su sistema inmune, así como en pacientes inmunodeprimidos. Se estima que se producen en el mundo en torno a un millón de muertes de niños menores de dos años al año por infecciones causadas por el neumococo. Este patógeno se transmite con facilidad por lo que puede producir brotes epidémicos. Incidencia (casos/ 100.000 año) 200 150 Incidencia de la infección por neumococo según las edades 39 100 50 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Las enfermedades de las que son responsables son: 1) 2) 3) 4) 5) neumonía meningitis otitis media sinusitis conjuntivitis Tratamiento: el tratamiento suele ser con cefalosporinas ya que hay muchas cepas resistentes a penicilinas. Existe una vacuna conjugada denominada Prevnar® que consiste en una mezcla de polisacáridos correspondientes a los siete serotipos K más frecuentes del patógeno conjugados sobre una proteína transportadora aislada de Corynebacterium diphteriae. La vacuna se aplica a los 2, 4 y 6 meses seguida de una dosis de recuerdo a una edad comprendida entre los 12 y 15 meses. 4.- E. faecalis El hábitat natural de E. faecalis (enterococo o estreptococo fecal) es el aparato digestivo, la cavidad oral, la vesícula, uretra y vagina. Es un patógeno oportunista. Es responsable de las siguientes enfermedades: 1) endocarditis, causada por miembros del grupo viridans y relacionada con pacientes con problemas cardiacos que han sido sometidos a tratamientos dentales. (S. mutans, miembro del grupo viridans es uno de los responsables de la caries dental.) 2) bacteremia 3) infecciones de las vías urinarias 4) infecciones abdominales 5) sepsis neonatal e infecciones de los tejidos blandos 6) Infecciones hospitalarias en pacientes con catéteres y otros dispositivos fijos. ESTAFILOCOCOS Los estafilococos son los cocos piógenos más peligrosos y la infección por ellos se suele acompañar de grandes cantidades de pus. Cuando se observan al microscopio for- 40 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) man masas arracimadas. Son catalasa positivos lo que permite distinguirlos con facilidad de los estreptococos y de los enterococos. Son organismos anaerobios facultativos. Hay más de 20 especies de Staphylococcus pero sólo tres se relacionan con enfermedades: • S. aureus • S. epidermidis • S. saprophyticus 1.- S. aureus Sólo S. aureus es coagulasa positivo (coagula el plasma sanguíneo) y sólo esta especie es considerada francamente patógena. S. aureus es una de las causas más importantes de infección nosocomial. Es un residente normal de la nariz y el intestino de entre el 30 y el 50% de la población general y cerca del 90% del personal clínico porta el microorganismo. Es muy resistente a la desecación y se disemina con facilidad por el personal clínico a partir de la cabeza (pelo, caspa) y de fomites tales como sábanas, ropa y equipo. S. aureus es también muy resistente a los agentes antisépticos (desinfectantes) y a muchos antibióticos. Todo esto facilita su rápida diseminación en el medio hospitalario, sobre todo en poblaciones susceptibles tales como recién nacidos, pacientes con quemaduras, sujetos inmunodeprimidos y pacientes con catéteres u otros dispositivos permanentes. Las infecciones causadas por este patógeno ocurren cuando se produce una lesión cutánea que favorece la penetración del microorganismo desde la piel hasta los tejidos más profundos. Entre las infecciones que producen se cuentan: 1) Enfermedades de la piel b) forúnculos y ántrax c) foliculitis d) impétigo 2) neumonía a) endocarditis b) pioartrosis c) bacteremia en quemaduras d) enteritis aguda e) intoxicación alimentaria producida por una enterotoxina termorresistente f) síndrome de escaldamiento cutáneo g) síndrome de choque tóxico producido en mujeres jóvenes que usan incorrectamente tampones vaginales durante la menstruación. Tratamiento: se usan penicilinas resistentes a β-lactamasas (Meticilina); sin embargo, la creciente aparición de cepas resistentes a este antibiótico (cepas MRSA) que normalmente presentan otras resistencias más (cepas multirresistentes) reduce las posibilidades de elección a los antibióticos Vancomicina y Teicoplanina. 2.- S. epidermidis 41 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) S. epidermidis es un habitante habitual de la piel humana. Se distingue del S. aureus por ser coagulasa negativo. La mayor parte de las infecciones que produce se deben a algún dispositivo implantado (válvulas cardiacas, prótesis de cadera, marcapasos, etc.) El microorganismo es muy resistente a los fármacos y su tratamiento es complicado. Es un microorganismo que forma biofilms (biopelículas) lo que dificulta su tratamiento. El tratamiento se basa en la substitución de los implantes con frecuencia y en la vigilancia de los mismos y de sus puntos de inserción. Las cepas suelen ser multirresistentes. Los antibióticos de elección son la Vancomicina y la Teicoplanina. 3.- S. saprophyticus S. saprophyticus es un habitante ocasional de la piel humana. Es uno de los principales causantes de infecciones urinarias en mujeres con actividad sexual. En mujeres produce cistitis o pielonefritis. En hombres produce de forma ocasional infecciones urinarias en pacientes de más de 50 años. NEISSERIA Las neiserias son diplococos Gram-negativos (principales cocos Gram-negativos.) Muchas están encapsuladas y tienen gran cantidad de fimbrias. Estas bacterias se distinguen de otros cocos por su respuesta a la tinción de Gram y por su carácter oxidasa positivos (cambian rápidamente a negro el color del reactivo de oxidasa dimetil o trimetilparafenilendiamina). Hay dos especies patógenas: • N. gonorrhoeae • N. meningitidis Los requerimientos de cultivo de las neiserias son complicados que contienen sangre o sangre calentada (medio color chocolate). En muchos casos es necesario usar medios selectivos. 1.- N. gonorrhoeae11 N. gonorrhoeae (gonococo) sólo habita en seres humanos. No forman parte de la flora normal (por lo que ha de considerarse como intrínsecamente patógeno); pero pueden estar presentes sin producir síntomas, por lo general en mujeres. Las enfermedades que producen son de transmisión sexual: 1) gonorrea 2) enfermedad pélvica inflamatoria 3) infección gonocócica diseminada 4) oftalmia neonatal 11 Incidencia de la infección gonocócica: 1.063 casos en 2003 y 866 en 2004 en toda España (Datos del Boletín Epidemiológico Nacional. 12: 289-296). 42 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Los gonococos eran antes muy susceptibles a la Penicilina; sin embargo, el uso de este antibiótico ha favorecido la aparición de cepas resistentes y hace necesario buscar nuevos antibióticos (cefalosporinas y quinilonas). Un problema adicional es que muchos pacientes de gonorrea tienen otros agentes infecciosos productores de la uretritis, por lo que es necesario tratar ambos agentes patógenos. 2.- N. meningitidis N. meningitidis (meningococo) se porta asintomáticamente por entre el 5 y el 20 % de la población normal y por tanto puede considerarse componente de la flora normal. Sus factores de patogenicidad más importantes son la cápsula [lo que permite su serotipificación en grupos A (el más frecuente en Europa), B y C], las fimbrias y el lipopolisacárido. La enfermedad producida por el meningococo es la meningitis meningocócica que se produce poco después de que el paciente enferme de una infección (normalmente respiratoria) producida por el meningococo. Esta es la primera causa de meningitis bacteriana en los niños y la segunda en los adultos. El contagio puede producirse por contacto estrecho entre el enfermo y el personal sano. La infección puede propagarse de forma epidémica con rapidez en recintos cerrados como colegios, guarderías, cuarteles, etc. Se presenta de forma más frecuente en invierno y primavera. Es una enfermedad de declaración obligatoria. Es importante el diagnóstico y tratamiento rápido porque esta enfermedad puede volverse rápidamente mortal con tasa de mortalidad en niños que van del 5 al 15%. Esta enfermedad es peligrosa porque los meningococos liberan el lipopolisacárido altamente endotóxico. Si el diagnóstico no es rápido, la enfermedad puede progresar con mucha rapidez. El tratamiento solía ser con penicilina; pero en la actualidad se recurre a cefalosporinas de tercera generación (cefriaxona, comercializada con el nombre de Rocephin®.) Se puede hacer un tratamiento quimiprofiláctico con Rifampicina a aquellas personas en contacto estrecho con enfermos. En casos de brotes epidémicos puede ser conveniente una terapia preventiva con antibióticos. También existen vacunas. MORAXELLA Las moraxelas están muy relacionadas con las neiserias, de hecho, el principal patógeno del grupo (M. catarrhalis) se clasificó durante mucho tiempo dentro del género Neisseria porque es un diplococo Gram-negativo oxidasa-positivo que habita en las mucosas. La separación del grupo se ha hecho, posteriormente, atendiendo a criterios de taxonomía molecular. M. catarrhalis se encuentra en las vías respiratorias de cerca del 5 % de la población adulta sana (y en el 50% de los niños sanos). Las enfermedades que produce son: 43 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) 1) sinusitis 2) bronquitis 3) laringitis (en algunos casos) El tratamiento suele ser con β-lactámicos e inhibidores de β-lactamasa (Augmentine .) ® ACINETOBACTER A. baumannii es una bacteria que pertenece a un grupo presente frecuentemente en el suelo, en el agua y en la piel de individuos sanos. Esta especie da cuenta del 80% de las infecciones producidas por este género que causa problemas en unidades de cuidados intensivos y similares. Normalmente no produce infecciones fuera de establecimientos sanitarios. Acinetobacter es un patógeno oportunista que causa, en pacientes con las defensas deprimidas, varios tipos de enfermedades que van desde neumonías hasta infecciones de la sangre o de heridas (especialmente en sitios de traqueotomías y en heridas abiertas.) El tratamiento se hace con los antibióticos comunes; pero el tratamiento es complicado porque los enfermos sueles encontrarse en situación grave por otras causas. A. baumanii es potencialmente muy peligrosa por la gran capacidad de adquirir resistencia a antibióticos (algo parecido a lo que ocurre con los MRSA) por lo que el uso de antibióticos de amplio espectro es problemático porque la aparición de resistencias frente a estos antibióticos limita su utilidad en el tratamiento posterior. Se estima que la tasa de mortalidad en los casos de infecciones de pacientes de riesgo es cercana al 40%. La prevención de las infecciones por Acinetobacter se basa en la higiene del personal sanitario ya que la bacteria puede vivir en la piel y en el ambiente durante varios días. Por tanto es necesaria una buena limpieza de las manos y de las instalaciones para limitar la dispersión del patógeno. HAEMOPHILUS Son pequeños bacilos Gram-negativos cuyo cultivo es complicado por ser muy exigentes. Son anaerobios facultativos, oxidasa y catalasa positivos. Se conocen muchas especies de este género, de las que muchas son patógenas. La más importante de ellas es • H. influenzae Esta bacteria está presente en entre el 40 y el 80% de la población adulta en las vías respiratorias superiores12. Por tanto, puede considerarse flora normal (estas cepas suelen ser no-encapsuladas, a diferencia de las virulentas que suelen ser encapsuladas). 12 Esta bacteria no produce la influenza o gripe que es una enfermedad vírica. 44 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Es un patógeno especialmente importante en niños pequeños (menores de cinco años). Las enfermedades que produce son: 1) bronquitis 2) celulitis en la mucosa bucal que se disemina a cara, cuello y ojos. 3) epiglotitis 4) faringitis aguda 5) meningitis (principal causa de meningitis bacteriana en niños antes de que se dispusiera de una vacuna). La enfermedad en niños se presenta hasta los cuatro años con una mayor incidencia entre los 6 y 9 meses. Puede llegar a tener unas tasas de mortalidad del entorno del 3 al 5%; pero en un gran número de casos deja secuelas en audición y desarrollo intelectual. 6) otitis media 7) neumonía 8) sepsis 9) sinusitis aguda Los principales factores de virulencia de esta especie son la cápsula, el lipopolisacárido y la presencia de fimbrias. La tasa de mortalidad de los pacientes de meningitis varía entre el 3 y el 17% en pacientes tratados. El tratamiento se ha hecho tradicionalmente con β-lactámicos; pero están apareciendo muchas cepas resistentes. La prevención se realiza mediante inmunización frente a las cepas encapsuladas (vacunas frente a H. influenzae de tipo b), y quimioprofilaxis (tratamiento preventivo de personas que vivan en contacto con pacientes infectados.) BORDETELLA Son bacterias Gram-negativas de crecimiento muy lento y cuyo cultivo requiere el uso de medios especiales. Especies de Bordetella patógenas • B. pertusis • B. bronchiseptica • B. parapertusis B. pertusis crece sobre el epitelio mucoso nasofaríngeo humano. Su principal característica es la de producir la toxina pertusis causante de los síntomas de la tos ferina que es la enfermedad causada por este patógeno. La prevención de la tos ferina se realiza principalmente por vacunación. BACTERIAS ENTÉRICAS Familia de las enterobacteriáceas Las bacterias entéricas pertenecen a la familia Enterobacteriaceae y se denominan así porque con mucha frecuencia se encuentran en el tracto intestinal (entérico) de ani45 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) males superiores. Sin embargo, se conocen enterobacteriáceas que también se encuentran en vida libre en el agua y en el suelo. Las enterobacteriáceas son bacilos Gram-negativos pequeños, anaerobios facultativas, son fermetadores de glucosa, oxidasa negativos, catalasa positivos y con flagelación peritrica (cuando están flagelados). Con excepción de los géneros Salmonella, Shigela y Yersinia (que son patógenos profesionales), las enterobacteriáceas son parte de la flora normal o son bacterias ambientales. Las enterobacterias pueden llegar a alcanzar concentraciones de 109 UFC/g en la materia intestinal. Este valor representa aproximadamente el 1% del contenido bacteriano intestinal que puede llegar a 1011 UFC/g, la mayoría de los microorganismos del intestino adulto son anaerobios estrictos del género Bacteroides. En la mayoría de los casos, las patologías producidas por estas bacterias son causadas porque el microorganismo invade zonas del cuerpo normalmente estériles. Casi todos los microorganismos entéricos son oportunistas y, como tales, son causa común de enfermedades nosocomiales. En pacientes de hospitales, las colonizaciones por enterobacterias suelen ser rápidas y están agravadas por tratarse de cepas multirresistentes. El principal factor de virulencia de estas bacterias es el lipopolisacárido (endotoxina.) Las enterobacteriáceas son tolerantes a las sales biliares y pueden crecer en medios de cultivo que las contengan, siendo esta una característica que permite elaborar medios de cultivo selectivos para ellas. La identificación de los cultivos de enterobacteriáceas se realiza en función de sus características bioquímicas (usando ensayos múltiples como los que constituyen las galerías API) y por pruebas serológicas en función de los antígenos O, H y K. 1.- Escherichia coli Escherichia coli (coli o colibacilo) es parte de la flora normal humana (hasta 109 UFC/g en la materia intestinal.) Es una bacteria fermetadora de la lactosa, y por esta característica puede ser identificada en medios de cultivo diferenciales (Agar de MacConkey). Los factores de virulencia en E. coli son los siguientes: • Las fimbrias que participan en la adhesión • El lipopolisacárido (antígeno O) • Enterotoxinas que son exotoxinas que afectan al funcionamiento del aparato digestivo • La verotoxina que daña las células del endotelio • Otros productos tóxicos, entre ellos una hemolisina Todos estos factores de virulencia, que pueden presentarse en unas u otras cepas, son responsables de los diferentes tipos de patologías causadas por E. coli: 46 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) 1) Infecciones oportunistas, especialmente neumonía, sepsis e infecciones postoperatorias. En recién nacidos se han descrito casos de meningitis. 2) Patologías de las vías urinarias: cistitis y pielonefritis en mujeres jóvenes. 3) Diarreas: a) Diarreas acuosas: i) E. coli enterotoxigénica (ETEC) causada por el consumo de alimentos contaminados. (1) El factor de virulencia en estas cepas es una toxina similar a la de Vibrio cholera pero de menor potencia que aquella. (2) Es más característica de países subdesarrollados donde causa la diarrea del viajero (o enfermedad de Hamburger) consistente en una intoxicación alimentaria cuyo periodo de incubación es de 24 a 72 h. Normalmente en adultos no es severa; pero en niños puede llegar a ser mortal por la deshidratación. (3) La enfermedad suele autolimitarse porque se elimina el patógeno al cabo de 2 o 3 días. Es necesario vigilar la hidratación del paciente y no es conveniente administrar antidiarreicos que limitan la eliminación del patógeno o productos lácteos que pueden ser no tolerados durante la infección. (4) En algunos casos, esta infección puede complicarse con el síndrome urémico-hemolítico. ii) E. coli enteropatógena (EPEC) ocasiona epidemias y diarreas entre lactantes. (1) Produce una enterotoxina citotóxica similar a la de Shigella dysenteriae13. (2) Está prácticamente erradicada de los países desarrollados. b) Diarreas con sangre i) E. coli enterohemorrágica (EHEC) causada por consumo de alimentos de origen animal en mal estado. (1) Produce una enterotoxina citotóxica para las células Vero (verotoxina) (2) Es especialmente importante en épocas recientes E. coli O157:H7. (3) En un gran número de casos, la infección se complica con el síndrome urémico-hemolítico. ii) E. coli enterinvasiva (EIEC) causada por el consumo de agua o alimentos contaminados con materia fecal. Posiblemente produce una enterotoxina citotóxica. En el tratamiento de E. coli se pueden usar diferentes antibióticos y la selección del más adecuado debe hacerse en función de la sensibilidad que presente el patógeno aislado y de la localización de la infección. Los antibióticos más usuales son β-lactámicos, quinolonas sulfamidas y Ciprofloxacina. 13 Las enterotoxinas citotóxicas Shiga de Shigella dysenteriae y Shiga-like (SLT) o Verotoxina de E. coli y las citotoxinas A y B de Clostridium difficile son similares en el modo de acción citotóxico por inactivación de las subunidades 60S de los ribosomas. Sólo son activas sobre células de humanos o primates. 47 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) En el tratamiento es muy importante velar por la rehidratación del paciente. El síndrome urémico-hemolítico es una complicación grave de ciertas infecciones producidas por E. Coli (O157:H7), Shigella y Salmonella (principalmente.) Es una enfermedad más común en los niños, que era infrecuente; pero cuya incidencia ha ido aumentando. En adultos es rara, aunque se puede presentar en pacientes oncológicos. La enfermedad se caracteriza por insuficiencia renal, anemia hemolítica y trombocitopenia. La enfermedad es grave y el tratamiento es solo de soporte. En torno al 60% de los afectados tratados se recuperan. No se conocen muchos factores determinantes de esta enfermedad, aunque se ha comprobado una asociación con el consumo de carne picada contaminada no suficientemente cocinada. 2.- Shigella Shigella es una bacteria muy similar a E. coli de la que se diferencia porque Shigella no es capaz de fermentar la lactosa. También presenta características similares a Salmonella de la que se diferencia por su falta de motilidad y por su incapacidad para producir H2S. Shigella dysenteriae produce disentería bacilar (enfermedad de declaración obligatoria) causada por su capacidad para producir la enterotoxina Shiga. Además de esta toxina, Shigella presenta una serie de proteínas que facilitan la invasión celular (patógeno invasivo penetrante) y una hemolisina. Shigella es muy infecciosa y la ingestión de un número reducido de microorganismos es suficiente para causar la enfermedad. El contagio suele ser por el consumo de alimentos contaminados con materia fecal y por contagio oro-fecal. El tratamiento de la enfermedad se puede hacer con antibióticos, aunque en adultos, el propio curso de la diarrea tiende a eliminar el patógeno. 3.- Salmonella Salmonella es una bacteria similar a las anteriores responsable de muchas infecciones alimentarias. Además de contagio a través de los alimentos, se puede originar la infección por contacto con material fecal de individuos enfermos o de portadores sanos y con material fecal de algunos tipos de animales de compañía (tortugas y reptiles, por ejemplo) o de animales de corral. Por consiguiente, se puede considerar una infección zoonótica. Las enfermedades producidas por salmonelas son: 1) Enteritis causada por S. typhimurium. 2) Fiebre entérica (fiebre tifoidea o tifus) causada por S. typh14i. a) Es una enfermedad de declaración obligatoria b) En este caso la enfermedad es mucho más grave y puede dar lugar a alteraciones importantes del intestino. 14 No hay que confundir esta enfermedad con el tifus producidlo por una Rickettisa 48 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) c) Una característica de S. typhi es que el patógeno puede refugiarse en la vesícula biliar y sobrevivir allí mucho tiempo siendo eliminado por las heces. En este caso, se habla de portadores sanos que son capaces de transmitir la enfermedad sin presentar síntomas. 3) Septicemia primaria en pacientes anémicos infectados por S. cholerasuis. El tratamiento es con antibióticos y existen vacunas contra la fiebre tifoidea. 4.- Klebsiella Klebsiella patógeno oportunista causantes frecuentes de infecciones nosocomiales. Son microorganismos sin movilidad y fermetadores de lactosa. Su principal factor de virulencia es la presencia de cápsula (antígenos K.) Su hábitat más frecuente es la parte superior del aparato respiratorio y en el aparato digestivo. La especie principal es K. pneumoniae. Las enfermedades que produce son 1) Neumonía 2) Rinitis 5.- Enterobacter Enterobacter como Klebsiella es un patógeno oportunista frecuente en infecciones nosocomiales. Es una bacteria de la flora entérica comensal. Se diferencia de Klebsiella por su hábitat y por ser un microorganismo móvil. Las principales especies son E. aerogenes y E. cloacae. Las enfermedades que producen son 1) Infecciones de las vías urinarias 2) En ocasiones: neumonías, infecciones de heridas y sepsis. 6.- Serratia Serratia, al igual que las dos especies anteriores, es un patógeno oportunista causante frecuente de infecciones nosocomiales. Son fermentadores lentos de la lactosa y son móviles. La principal especie es S. marcescens y las enfermedades que causa son 1) Neumonías y sepsis. 2) Infecciones de las vías urinarias e infecciones de heridas 7.- Proteus Proteus son bacterias de la flora intestinal normal y se encuentran como microorganismos saprófitos en agua y suelo. Son microorganismos móviles que tienen una gran actividad ureasa y son incapaces de fermentar lactosa. Sólo hay dos especies patógenas: P. vulgaris y P. mirabilis. Los factores de virulencia son el lipopolisacárido, la presencia de fimbrias, la cápsula y la producción de ureasa. 49 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Las enfermedades en las que participan son: 1) Infecciones de las vías urinarias tales como cistitis y pielonefritis 2) Sepsis 3) Cálculos renales e infecciones de vejiga 4) Infecciones de heridas y neumonías 8.- Yersinia Yersinia son bacterias productoras de enfermedades zoonóticas. Son lactosanegativas. Sus factores de virulencia son el lipopolisacárido y la producción de proteínas que facilitan su actividad invasiva del tejido que infectan Hay tres especies, dos móviles (Y. pseudotuberculosis y Y. enterocolitica) que se transmiten a los humanos por ingestión de alimentos contaminados y que causan una enfermedad de tipo diarréico; y una inmóvil (Y. pestis) causante de la peste y que es transmitida por picaduras de pulgas portadoras del bacilo (esta última es una enfermedad de declaración obligatoria a la OMS y requiere la cuarentena de los posibles contagiados). Otras bacterias entéricas de intrés que no son enterobacteriáceas 9.- Campylobacter Las bacterias de este género son bacilos Gram-negativos cortos y curvados, microaerófilos y móviles por medio de un único flagelo polar. Sus principales factores de virulencia son la producción de capas S en algunas especies y la liberación de una enterotoxina en otras. Son, junto con las salmonelas, las bacterias causantes del mayor número de procesos intestinales de origen alimentario. La principal especie del grupo C. jejuni se adquiere principalmente por consumo productos de ave contaminados, de leche no tratada o de agua contaminada. También puede contagiarse por animales de compañía como los perros. Recientemente, se ha comprobado que puede transmitirse a través de las moscas15 ya que un número muy reducido de bacterias es capaz de causar enfermedad. Por otra parte, se están encontrando un gran número de portadores asintomáticos. Las enfermedades producidas por Campylobacter son: 1) Enteritis aguda y diarrea que se produce de 2 a 5 días después de la exposición al patógeno. Esta infección es también una de las causas de la diarrea del viajero. La infección se elimina por sí sola y se previene con una buena higiene. 2) Bacteremia febril aguda 3) Meningitis 4) Enfermedad respiratoria 15 EID 11: 361-364 50 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) 10.- Helicobacter Es una bacteria microaerfílica con forma bacilar un poco curva. Es Gram-negativa. Vive en la mucosa gástrica humana y produce úlceras gástricas. La especie de este grupo es H. pylori. Es interesante señalar que hay mucha gente que tiene H. pylori y no desarrolla la enfermedad, lo que sugiere que debe de haber otros factores que contribuyan a su patogenicidad. Los mecanismos de patogenicidad se basan en la presencia de adhesinas, ureasa, flagelos, producción de una citotoxina vacuolante H. pylorii se encuentra en las heces secreciones orales y placa dental. Por tanto, se cree que la vía de contagio es la oral. También puede transmitirse a través del agua. Las enfermedades que produce son: 1) Gastritis crónica 2) Úlceras gástricas y duodenales 3) Es un factor de riesgo para el adenocarcinoma gástrico. PSEUDOMONAS Las pseudomonas son un grupo grande de bacilos Gram-negativos generalmente aerobios, que pueden tener flagelos polares, son oxidasa-positivos y crecen en medios de cultivo simples. La taxonomía del grupo es muy complicada y varía con frecuencia debido a avances en los estudios moleculares y ecológicos lo que supone que se han producido cambios de nombres en los últimos años. Son bacterias causantes de enfermedades como patógenos oportunistas. 1.- Pseudomonas aeruginosa Es una pseudomona cuyas colonias producen un color y olor característicos. Es una bacteria frecuente en el agua y en el suelo y que puede contaminar algunos utensilios tales como líquidos de desinfección (es bastante resistente) esponjas, material clínico, etc. Puede colonizar al hombre sin causar enfermedad. Esta se produce cuando el patógeno alcanza localizaciones normalmente estériles por medio de una herida. En personas sanas no suele causar problemas, no así en pacientes inmunocomprometidos. Es una causa importante de infecciones hospitalarias. Las patologías más frecuentes asociadas a Ps. aeruginosa son 1) infecciones de heridas quirúrgicas y quemaduras 2) infecciones de las vías respiratorias (neumonía) 3) Infecciones de oído externo (oído de nadador) 51 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) 4) bacteremias (graves por la posibilidad de shock séptico debido al LPS.) 5) infecciones respiratorias en pacientes de fibrosis cística Con frecuencia aparecen cepas multirresistentes asociadas con brotes nosocomiales. El tratamiento es difícil por la presencia de múltiples resistencias y se suele basar en el uso de cefalosporinas de tercera generación. BACTERIAS PATÓGENAS INTRACELULARES MICOBACTERIAS Son bacterias que tienden a formar largas cadenas filamentosas que les dan un aspecto fúngico. Son bacterias de crecimiento muy lento. Bacilos Gram-positivos resistentes al ácido-alcohol debido a la presencia de una pared celular de gran complejidad. Son bacterias presentes en el medio ambiente. Como patógenos son intracelulares ya que se multiplican dentro de células del sistema retículo-endotelial. Las enfermedades que producen son crónicas con periodos de latencia muy largos. Actualmente, además de las formas patológicas clásicas producidas por estas bacterias, han aparecido otras de incidencia en pacientes inmunodeprimidos. 1.- Mycobacterium tuberculosis Se conoce con el nombre de bacilo de Koch y es el causante de la tuberculosis. El hombre es el único reservorio de esta bacteria. Es una patología cuya incidencia había seguido un descenso paulatino en los países desarrollados hasta que con la llegada del SIDA su incidencia ha aumentado en los últimos tiempos. En países subdesarrollados, la tuberculosis es una patología muy importante por su incidencia. Por otra parte ha aumentado también la incidencia de tuberculosis causadas por micobacterias atípicas diferentes de M. tuberculosis, principalmente en pacientes inmunodeprimidos. El contagio se produce por aeerosoles emitidos por pacientes infectados (tos, esputos, etc.) aunque también puede producirse por ingestión de alimentos contaminados (M. bovis y consumo de leche contaminada.) El contagio por contacto a través de la piel es muy infrecuente. La enfermedad tuberculosa tiene múltiples formas: 1) tuberculosis pulmonar 2) tuberculosis extrapulmonar 3) tuberculosis diseminada 52 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) 4) meningitis tuberculosa El diagnóstico de la tuberculosis comprende pruebas de laboratorio microbiológico para identificar el patógeno. Todo el personal de laboratorio que trabaje con muestras susceptibles de contener M. tuberculosis deben extremar las medidas de seguridad para evitar la infección. Hoy en día, debido a la asociación de SIDA con tuberculosis hay que considerar que cualquier muestra de esputo puede contener la micobacteria y, por consiguiente, debe ser tratada como potencialmente peligrosa. Una prueba de diagnóstico muy usada es la prueba de la tuberculina o prueba de Mantoux en la que se inyecta intradérmicamente un extracto de micobacterias y se observa la reacción inmunológica que se produce. La reacción positiva solamente indica que se ha estado en contacto con una micobacteria relacionada. Su mayor interés es para la aplicación a la población infantil. El tratamiento es complejo y requiere el uso de varios fármacos combinados (fármacos de 1ª y 2ª línea) durante largos periodos de tiempo (6 a 9 meses). Es especialmente preocupante la aparición de cepas multirresistentes De forma general, existen dos estrategias por las que una bacteria puede hacerse persistente: (1) multiplicándose más rápidamente de lo que es destruida por antibióticos y células del sistema inmune o (2) reduciendo mucho su tasa de crecimiento y protegiéndose de la acción del sistema inmune. M. tuberculosis la estrategia es la estática (no multiplicarse) por lo que para combatir la infección es necesario usar fármacos dirigidos frente a microorganismos que no estén creciendo16. Existe una vacuna (BCG) basada en micobacterias atenuadas de origen animal (M. bovis) cuyo uso es controvertido y que se recomienda sólo en los casos de grandes endemias. La BCG no protege de la enfermedad; pero sí previene su progresión. La tuberculosis es una enfermedad de riesgo en el personal sanitario por lo que recomienda el seguimiento del mismo mediante la prueba de Mantoux y la quimioprofilaxis en casos de conversión de la respuesta. 2.- Mycobacterium leprae Es la bacteria causante de la lepra. Esta es una enfermedad endémica en Europa en la Edad Media, aunque ahora está muy contenida en los países desarrollados (en España hay unos 5000 enfermos.) Sin embargo, es una enfermedad importante en países subdesarrollados. Al igual que en el caso de M. tuberculosis el reservorio es únicamente humano, la vía de transmisión principal son los aerosoles y cualquier persona es potencialmente susceptible a la enfermedad. Los enfermos eliminan un gran número de bacilos por las secreciones nasales, por lo que estas y los aerosoles son las principales vías de contagio. El contagio piel-piel o ambiente-piel se considera poco importante. 16 Infect. Immun. 73: 546-551 53 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) El cultivo en el laboratorio de este microorganismo no es posible en medios sin células. Existen dos variantes de la enfermedad 1) lepra tuberculosa 2) lepra lepromatosa (productora de deformaciones) El diagnóstico se hace por análisis microscópico de las secreciones y por el sintomatológico. El tratamiento es muy complicado porque debe ser combinado y de larga duración (2 años.) BRUCELAS Son bacterias intracelulares zoonóticas productoras de la fiebre de Malta. Hay varias especies de este género que causan enfermedades en diferentes tipos de animales. Son cocobacilos Gram-negativos que crecen con lentitud. Se identifican microbiológicamente por sus características de fermentación, la producción de H2S y la producción de ureasa. Son aerobios estrictos. Las vías de contacto de las brucelas animales son las heridas producidas por los correspondientes animales (arañazos, mordeduras) o por contacto con sus heces. El tratamiento suele basarse en la penicilina. 1.- Brucella melitensis Es la productora de la fiebre de malta o brucelosis. Se trata de un microorganismo muy contagioso cuando se manipula en el laboratorio. Es enfermedad de declaración obligatoria. Es una zoonosis17 porque el animal infectado primariamente son las ovejas o las cabras. El contagio a humanos se realiza a través del consumo de leche de estos animales que no ha sido tratada correctamente. Es una enfermedad profesional del personal en contacto frecuente con estos animales. Con frecuencia la enfermedad se transforma en crónica y son frecuentes las complicaciones en forma de focalizaciones (artritis en rodilla, hombro, etc.) El diagnóstico sintomatológico se completa necesariamente por pruebas de laboratorio y se basa principalmente en pruebas serológicas. El tratamiento con antibióticos debe ser prolongado y basado en antibióticos bactericidas que tengan penetración intracelular (tetraciclina, estreptomicina.) 17 Se denomina zoonosis a una enfermedad infecciosa que puede afectar a animales y a humanos. 54 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) La prevención se realiza mediante la eliminación de los animales infectados y mediante el control higiénico de los productos que puedan transmitir la infección. No existe una vacuna efectiva contra la brucelosis. OTRAS BACTERIAS INTRACELULARES 1.- Legionella pneumophila Es el agente causante de la legionelosis o enfermedad de los legionarios. Se trata de una bacteria Gram-negativa (aunque la tinción de Gram es muy difícil de realizar en esta especie), aerobia de vida acuática cuyo cultivo en el laboratorio es muy difícil (microorganismo exigente). El contagio se produce por la inhalación de aerosoles que contienen la bacteria por personas susceptibles. Los aerosoles se producen por ciertos equipos de refrigeración, duchas, etc. que usan agua contaminada. No se transmite por contacto. Se han detectado casos de infección nosocomial y suele producir brotes epidémicos. Es una bacteria poco patógena. La población de riesgo está formada por ancianos y por jóvenes muy fumadores. La infección que causa es principalmente respiratoria (neumonía atípica.) El diagnóstico microbiológico es importante porque los antibióticos comúnmente empleados para el tratamiento de las neumonías convencionales son ineficaces frente a Legionella. El antibiótico de elección para tratar este patógeno es la eritromicina. 2.- Listeria monocytogenes Es una bacteria Gram-positiva aerobia no esporulante catalasa-positivo. Es un microorganismo psicrotrofo. Las bacterias de este género (Listeria) están ampliamente distribuidas en la naturaleza en el suelo, polvo, etc. y coloniza un gran número de animales. La infección por la especie patógena (L. monocytogenes) se produce por ingestión de alimentos contaminados y la enfermedad (listeriosis) que puede causar 1) abortos 2) meningitis en los recién nacidos 3) sepsis en ancianos, recién nacidos y pacientes inmunodeprimidos En muchos casos, la infección transcurre de forma asintomática sin mayor trascendencia. El tratamiento es con ampicilina y gentamicina. Las medidas de control preventivo se dirigen hacia los alimentos. PATÓGENOS TOXIGÉNICOS 55 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) A continuación vamos a estudiar aquellas bacterias que producen su acción patógena principalmente a través de la producción de toxinas más que por su acción piógena o por la invasión de células del paciente. CORINEBACTERIAS Son bacterias pleomórficas anaerobias facultativas, catalasa-positivas y habitualmente no hemolíticas. La mayoría de ellas son sapróftas inofensivas y forman parte de la flora normal de las mucosas y la piel; aunque pueden comportarse como patógenos oportunistas y causar enfermedad en pacientes inmunodeprimidos. 1.- Corynebacterium diphteriae Es el agente causal de la difteria. El hombre es el único reservorio conocido de este patógeno. La vía de entrada del patógeno es la respiratoria donde el patógeno coloniza y se multiplica en la faringe y libera la exotoxina diftérica que causa la necrosis de los tejidos. La reacción inmunológica causa la producción de una membrana que va recubriendo las zonas afectadas y termina por obstruir las vías respiratorias causando dificultades respiratorias y asfixia. La toxina es absorbida por la sangre y causa efectos a nivel de corazón y de sistema nervioso produciendo fallos cardiacos y parálisis nerviosas. El diagnóstico se realiza por los síntomas y el tratamiento debe iniciarse inmediatamente. El tratamiento es mediante una antitoxina destinada a inactivar la toxina antes de que entre en las células y mediante eritromicina para eliminar el patógeno. Puede ser necesaria la traqueotomía o intubación. El contagio es por inhalación de aerosoles producidos por los pacientes infectados y por contacto con las mucosas y membranas de los enfermos. La enfermedad es de declaración obligatoria. Actualmente está erradicada en nuestro país gracias a los programas de vacunación. Sin embargo puede haber población susceptible entre los adultos de más de 40 años. Hay un programa de vacunación (vacunación triple: difteria, tos ferina y tétanos) de niños que previene las epidemias. La vacuna se realiza mediante la administración de un toxoide (forma inactivada de la toxina diftérica.) BACILOS GRAM-POSITIVOS AEROBIOS ESPORULANTES 56 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) En este apartado se trata de bacilos Gram-positivos aerobios o anaerobios facultativos, catalasa-positivos y esporulantes. Se trata de microorganismos muy abundantes en el medio que nos rodea y raramente causan enfermedad. Las especies patógenas principales son 1.- Bacillus cereus Causante de intoxicaciones alimentarias por consumo de alimentos ricos en hidratos de carbono y de baja actividad de agua (arroz cocido, por ejemplo) que ha sido mantenido a temperaturas incorrectas. La toxina actúa en el intestino y la enfermedad cursa con vómitos y ocasionalmente con diarreas. 2.- Bacillus anthracis Causante del carbunco o ántrax cutáneo (no confundir con el ántrax causado por S. aureus.) Se trata de una zoonosis en la que los primeros animales afectados son ovejas o vacas. También se puede considerar una enfermedad profesional para el personal en contacto con estos animales infectados. La resistencia y perdurabilidad de las esporas de B. anthracis hacía muy difícil su eliminación de los campos afectados y era necesaria la destrucción por fuego de los animales afectados para evitar la propagación de la enfermedad. En humanos es poco frecuente y se manifiesta como ántrax cutáneo que se adquiere por contacto directo con el animal infectado o con las esporas del patógeno. La infección puede diseminarse desde la piel y causar una alta mortalidad (hasta el 20% de los pacientes sin tratar). Existen formas pulmonares e intestinales mucho más graves causadas al inhalar o ingerir esporas (unas 10.000) del bacilo. La causa de la enfermedad es la toxina producida por las formas vegetativas. El tratamiento con antibióticos se basa en penicilina y en ciprofloxacina. Existe también una vacuna. Es una bacteria que puede usarse como arma biológica por lo que hay algunos programas de vacunación entre el personal militar. BACILOS GRAM-POSITIVOS ANAEROBIOS ESPORULANTES Se trata de bacilos anaerobios estrictos (catalasa-negativos) que se encuentran muy distribuidos en el suelo y el polvo. Algunos forman parte de la flora intestinal de animales superiores y se comportan como patógenos oportunistas. La mayoría de las enfermedades que causan son producidas por causas endógenas, aunque también hay casos de infección. Una de las vías de infección es a través de las heridas en las que se producen condiciones anaerobias (heridas profundas). Debe sospe57 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) charse de heridas que huelan mal porque pueden indicar la presencia de estos microorganismos anaerobios. Hay varias especies y condiciones causantes de patologías: 1.- Clostridium perfringens Es una especie frecuentemente aislada de muestras clínicas y puede encontrarse en muchas ocasiones en procesos de: 1) 2) 3) 4) contaminación de heridas celulitis gangrenas gaseosa bacteremia La patología más grave es la gangrena gaseosa a la que están más expuestos aquellos pacientes de edad avanzada con problemas circulatorios, sobre todo en los miembros inferiores. Esta bacteria también es causante de intoxicaciones alimentarias principalmente en alimentos cárnicos maltratados térmicamente. 2.- Clostridium botulinum Causa el botulismo: intoxicación muy grave a consecuencia de la ingestión de la toxina producida por este microorganismo al crecer en alimentos de pH alto que no han sido tratados térmicamente de forma adecuada. La toxina botulínica es una de las más potentes que se conocen. Es una toxina termosensible y por consiguiente se destruye en la cocción de los alimentos. La sintomatología se inicia con diarrea y vómitos seguidos de parálisis que llevan a la muerte por insuficiencia respiratoria. Como consecuencia de la acción de la toxina se produce una parálisis flácida Hay una variante infantil (botulismo infantil) causada por la ingestión de un número alto de esporas que se desarrollan en el intestino del niño y producen la toxina. Se produce, entonces, un aletargamiento del niño (bebé flojo.) Una de las formas más frecuentes de contagio del botulismo infantil es por la ingestión de miel que contiene las esporas (no se debe dar miel a los recién nacidos.) El tratamiento del botulismo se basa en el vaciamiento del estómago para eliminar la fuente de la toxina y la administración de una antitoxina. La tasa de mortalidad ha sido históricamente muy alta (70%); pero ahora se ha reducido a valores del entorno del 25%. 3.- Clostridium tetanii 58 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Es el agente causante del tétanos. C. tetanii es habitante habitual del intestino de humanos y de animales superiores y la infección se produce por la entrada de esporas del patógeno presentes en objetos que producen heridas profundas. La neurotoxina tetánica es muy fuerte y produce una contracción muscular generalizada (risa sardónica y postura en arco rígido.) La muerte se produce por asfixia. El tratamiento requiere la administración de relajantes musculares y de antitoxina tetánica. Hay una vacuna basada en un toxoide tetánico que se administra a los niños en el conjunto de la vacuna triple. En casos de heridas se suele administrar una dosis de recuerdo y, en casos de grandes heridas sucias, se complementa con gammaglobulina y una vacunación inmediata. La vacunación de recuerdo se administra en el momento de producirse la herida sospechosa, al mes, al año y a los diez años. 4.- Clostridium difficile Es el agente causante de la colitis post-antibiótica. Se trata de una bacteria presente en el intestino de muchas personas sanas en las que su número es reducido. Cuando un tratamiento con antibióticos reduce el número de bacterias competidoras, su presencia puede aumentar y la producción de toxina dar lugar a la enfermedad. La enfermedad puede variar desde una diarrea a una colitis pseudomembranosa de carácter muy grave y con alta mortalidad. El contagio puede producirse por diseminación de las esporas por lo que es necesario extremar las medidas de higiene y de contención cuando se ha estado en contacto con estos enfermos. El tratamiento requiere detener la administración del antibiótico exacerbante y administrar un antibiótico específico para C. difficile (Vancomicina.) BACTERIAS ENTÉRICAS TOXIGÉNICAS En este apartado trataremos de los vibrios que son bacilos Gramnegativos que comparten as características de las enterobacteriáceas (fermentadores, oxidasa-positivos) y presentan motilidad por un flagelo polar. La forma es característica de coma. 1.- Vibrio cholera Es el causante del cólera. Esta es una toxiinfección originada por el consumo de agua contaminada con materia fecal en la que se encuentra el patógeno. El reservorio de V. cholera es humano; pero puede haber alguno animal todavía no identificado completamente. 59 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) La enfermedad se causa por la toxina colérica que altera intensamente la permeabilidad de la membrana intestinal produciendo una fuerte diarrea acuosa (agua de arroz) que supone una gran pérdida de agua y de electrolitos para el paciente. La enfermedad cursa en brotes epidémicos cuando las condiciones de higiene no son suficientes y las deyecciones de los enfermos contaminan agua o alimentos de los sanos. El curso de la enfermedad es muy rápido (pueden producirse las muertes en 24 a 48 horas.) El tratamiento se basa en la rehidratación por vía intravenosa y en la administración de antibióticos (tetraciclina o doxiciclina) para eliminar el patógeno. Es también importante tratar el agua (cloración, adición de lejía) para eliminar el patógeno y controlar la difusión de la enfermedad. Es una enfermedad extremadamente contagiosa, de declaración obligatoria y que requiere el establecimiento de medidas de cuarentena. La presencia de anticuerpos contra V. cholera reduce mucho la severidad de la enfermedad. Existen dos vacunas disponibles que ofrecen protección corta y limitada contra la bacteria del cólera, aunque ninguna de las dos se encuentra disponible en España pero se puede obtener a través del Servicio de Sanidad Exterior. Se suministran dosis de refuerzo cada 6 meses a las personas que permanecen en un área donde hay riesgo de cólera. OTRAS BACTERIAS PATÓGENAS ESPECIALES ESPIROQUETAS Treponema pallidum Es la causante de la sífilis. Se trata de un microorganismo de pequeño tamaño que no es observable al microscopio óptico con los sistemas de tinción normales. No ha podido ser cultivado en el laboratorio. El único reservorio es el humano. La sífilis es una enfermedad de transmisión sexual y de declaración obligatoria. La enfermedad pasa por diferentes estados (periodo de incubación, sífilis primaria, secundaria y terciaria) y puede atravesar la barrera placentaria y causar la sífilis congénita. El síntoma característico de la sífilis es la aparición de una úlcera plana no dolorosa, principalmente en el área genital, que se conoce con el nombre de chancro de inoculación. El tratamiento de la sífilis se basa en la inyección de altas concentraciones de penicilina de acción retardada o doxiciclina en el caso de pacientes alérgicos a los βlactámicos, y la prevención del contagio se basa en las medidas de protección frente a las enfermedades de transmisión sexual. 60 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) El patógeno escapa del sistema inmune y es difícil de tratar en pacientes con SIDA. MICOPLASMAS Mycoplasma pneumoniae Es el microorganismo causante de la neumonía atípica primaria. Los microplasmas no tienen pared celular de peptidoglicano y, por consiguiente, son insensibles a los βlactámicos y son muy pleomórficos Su cultivo en el laboratorio es muy complicado. Hay especies que son parte de la flora normal; pero M. pneumoniae es intrínsecamente patógeno. La infección se presenta en adultos jóvenes y el contagio es persona-persona por medio de contacto directo y siendo necesario un contacto estrecho. La enfermedad se presenta como faringitis, bronquitis o neumonía siendo por lo general leve, con fiebre y tos no productiva. El tratamiento normalmente no es necesario porque la infección se autolimita. En caso de ser necesario el uso de antibióticos, la elección es un antibiótico macrólido, tetraciclina o eritromicina. CLAMIDIAS Son pequeñas bacterias Gram-negativas que se comportan como parásitos obligados ya que carecen de sistemas de producción de energía. Se multiplican dentro de las células que invaden produciendo los llamados cuerpos reticulados y cuerpos de inclusión Hay tres especies patógenas para el hombre: 1.- Chlamydia trachomatis Causante de infecciones de transmisión sexual que se manifiestan como uretritis en el varón, enfermedad pélvica inflamantoria en la mujer y pueden causar conjuntivitis neonatal y neumonía en el recién nacido. La enfermedad conjuntiva es grave y se conoce con el nombre de tracoma siendo esta una de las principales causas de ceguera, sobre todo en países subdesarrollados. El tratamiento se basa en la utilización de doxiciclina o de eritromicina 2.- Chlamydia pneumoniae Es uno de los principales causantes de las neumonías atípicas y de las bronquitis en el mundo. La enfermedad se contagia por contacto con las secreciones nasales de los enfermos y puede dar lugar a brotes epidémicos. 61 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) 3.- Chlamydia psittaci. Es causante de la psitacosis, tipo de neumonía que se presenta en personas que conviven con aves. RICKETTISAS Son bacterias con características particulares que causan enfermedades endémicas o epidémicas de gran difusión y transmitidas frecuentemente por artrópodos. Son parásitos celulares estrictos que pueden infectar no sólo fagocitos sino también otros tipos de células. 1.-Rickettsia prowazekii. Es el ejemplo de este tipo de microorganismos. Causa el tifus epidémico (o enfermedad de Brill-Zinsser.) Esta enfermedad se transmite a través de piojos (y, por consiguiente, está asociada a condiciones de baja higiene) cuando la temperatura es fría. La enfermedad produce fiebres extremadamente altas (de hasta 40,5 - 41ºC), cefaleas, mialgias, fotofobia y la aparición de un sarpullido que cubre todo el cuerpo. Pueden aparecer trastornos neurológicos (psicosis) causados por meningitis. La enfermedad puede producir una mortalidad de entre el 10 y el 60%. Los pacientes mayores de 60 años son quienes presentan el mayor riesgo de muerte. El tratamiento se dirige a eliminar la infección y a tratar los síntomas con antibióticos tales como tetraciclina, doxiciclina o cloramfenicol. Puede ser necesario administrar líquidos intravenosos y oxígeno para ayudar a estabilizar el paciente. Existe una vacuna que se administra a individuos con riesgo (por ejemplo, personal sanitario de zonas endémicas.) 2.-Rickettsia typhi. Es la causante del tifus murino o tifus endémico. Es una forma más leve de tifus que rara vez es mortal (menos del 2%.) Se ve con frecuencia en el verano y el otoño y por lo general tiene de dos a tres semanas. Al cabo de 15 días sin tratamiento los síntomas más agudos remiten aunque el abatimiento general puede durar varios meses. Entre los factores de riesgo para el tifus murino se puede mencionar la exposición a pulgas o heces de rata o la exposición a animales como gatos, y ratas. 2.- PRINCIPALES GRUPOS DE VIRUS DE IMPORTANCIA CLÍNICA. Los virus son agentes infecciosos que presentan una serie de características especiales: • Pequeño tamaño (10-300 nm) • Genoma con un solo tipo de ácido nucleico • Carecen de actividad metabólica 62 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Requieren medios de cultivo especiales en los que haya células a las que infectar • Son resistentes a los antimicrobianos habituales • Los virus siempre han tenido una gran importancia como agentes patógenos; sin embargo, en los últimos años esta se ha incrementado aún más debido a enfermedades víricas graves de diferentes tipos • • • • • Enfermedades primarias de nueva aparición (SIDA) Enfermedades asociadas a pacientes inmunodeprimidos Enfermedades asociadas a pacientes con otras patologías crónicas Infecciones víricas banales que presentan complicaciones graves Infecciones nosocomiales. Los virus se clasifican en función de la naturaleza de su genoma, de la simetría de la cápsida y de la presencia o no de envuelta. La multiplicación de los virus presenta varias etapas; adsorción, penetración, decapsidación (desnudamiento), replicación, ensamblaje (maduración) y liberación del virus. La fase de replicación suele requerir la participación de enzimas codificadas por el propio virus y es una diana de acción de agentes antivirales. El diagnóstico de las infecciones víricas se lleva a cabo por tres procedimientos básicos: • Detección de las partículas víricas o alguno de sus componentes • Cultivo de los virus en medios con células vivas • Serología (detección de anticuerpos.) Las muestras tomadas para la detección de virus en el laboratorio deben procesarse lo más rápidamente posible. En caso de que esto no fuera posible, es necesario conservarlas en congeladores de -80 ºC, y nunca a temperaturas superiores a los -20ºC. La prevención y tratamiento de las enfermedades víricas se basa en los siguientes puntos: • Inmunización activa mediante vacunas de virus muertos, atenuados o antígenos específicos • Inmunización pasiva mediante antisueros • Inmunomoduladores que de forma inespecífica hacen al huésped menos receptivo a la infección por el virus (interferón) • Antivíricos que intervienen actuando sobre alguna de las etapas del ciclo de multiplicación del virus. Los interferones son glicoproteínas que se sintetizan como parte de la respuesta a la infección. Dependiendo del tipo de células que los producen se clasifican en • Interferón α producido por leucocitos • interferón β producido por fibroblastos • Intrerferón γ producido por linfocitos T 63 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Los interferones α y β se comportan como antivíricos al impedir el ensamblaje de las partículas víricas. VIRUS DE ADN ADENOVIRUS Son uno de los dos tipos de virus con más incidencia en nuestro entorno. Se conocen más de 40 serotipos de adenovirus causantes de patologías respiratorias, diarreas y conjuntivitis. En general las patologías por adenovirus son de evolución benignas. Algunos adenovirus se están usando como vectores para la fabricación de vacunas para otros virus (como en el caso del virus Ébola18, o en la construcción de vectores para terapia génica. HERPESVIRUS Son el segundo tipo de virus más importante en nuestro entorno. Son virus de ADN envueltos que se replican en el núcleo de las células que parasitan y son observables por las acumulaciones de viriones que producen (inclusiones intranucleares.) Son específicos de especie. Producen infecciones crónicas porque quedan en latencia en el genoma de las células infectadas y después pueden activarse en diferentes etapas de la vida del paciente. En general las infecciones que producen son benignas, aunque pueden surgir complicaciones en pacientes comprometidos. Patologías causadas por herpesvirus: Herpes Simple (VHS) Causa infecciones primarias en la piel o en las mucosas (calenturas), aunque puede diseminarse a otras localizaciones. La característica más notable del VHS es la recurrencia de la enfermedad después de una primera infección asintomática en la mucosa oral (VHS de tipo 1) o genital (VHS de tipo 2). Destacan por la gravedad el herpes neonatal, la encefalitis herpética y el herpes diseminado. Sin embargo, los más frecuentes son el herpes labial y el herpes genital. Este último es la tercera causa de infección de transmisión sexual después de las infección por Chlamydia trachomatis y por Neisseria gonorrhoeae. 18 Nature 424:681 64 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) El diagnóstico sintomatológico de los tipos más frecuentes es sencillo. En casos más complicados es necesario tomar muestras de los tejidos infectados en el curso de la manifestación de la infección. El fármaco de tratamiento es el aciclovir que se administra en casos graves como cuando se produce encefalitis. En el caso de madres portadoras, la cesárea reduce el riesgo de infección del niño al nacer. Algunos investigadores han relacionado la infección crónica por herpesvirus con enfermedades neurodegenerativas como el Alzheimer. Varicela-Zoster Es el causante de la varicela como infección primaria y del Herpes-Zóster como manifestación recurrente. La varicela es una enfermedad de la infancia19 consistente en un proceso febril leve, con exantema vesicular característico que afecta generalmente a cabeza, tronco y cuello. El periodo de incubación es de unos 14 días y es contagiosa desde unos 2 días antes de la manifestación hasta unos 5 días después. No suele tener complicaciones. Sin embargo, es especialmente peligrosa la infección de mujeres embarazadas que se encuentran en la última semana de embarazo o la infección en las primeras semanas postparto por el alto riesgo de que se produzca una infección grave del neonato. El herpes Zóster surge habitualmente en adultos como resultado de la reactivación del virus que quedó latente en los ganglios neurológicos dorsales o craneales. Se manifiesta por una erupción vesicular dolorosa en las áreas de la piel inervadas por los nervios afectados. En huéspedes inmunodeprimidos puede presentarse una varicela o Zóster diseminado que causa una mortalidad elevada (más del 50%.) El tratamiento es con aciclovir, valaciclovir o famciclovir para los casos más graves. Citomegalovirus. Es muy frecuente y afecta hasta cerca del 80% de la población. La mayoría de las infecciones son asintomáticas; sin embargo, la importancia de este virus radica en que es el que más frecuentemente aparece implicado en infecciones congénitas (es la causa viral más frecuente de defectos congénitos) y en infecciones en pacientes inmunodeprimidos donde se comporta como un patógeno oportunista. La infección congénita (afecta a entre el 0,5 y el 2% de los recién nacidos) se adquiere en el útero y el caso puede ser más grave si se da en las primeras semanas de ges19 Incidencia: 178.933 casos en 2003 y 233.934 casos en 2004. En Navarra 47 casos en 2004. (Datos del Boletín Epidemiológico Nacional. 12: 289-296). 65 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) tación o si es la primera infección de la madre. En estos casos, el desarrollo del feto se ve muy comprometido (en torno al 10% de los niños infectados presentan microcefalia y otros problemas neurológicos entre los que destacan la sordera y el retraso mental.) En la mayoría de los casos, sin embargo, cursa como una infección benigna. La infección perinatal se produce (1) en el parto, aproximadamente el 50% de los partos de mujeres portadoras del virus (en torno al 20%); o (2) como consecuencia de la lactancia. Esta infección hace a los niños difusores del virus; pero no provocan síntomas graves en niños sanos nacidos a término. El CMV se transmite por vía sexual con lo que alcanza la prevalencia en la población del entorno del 20%. Su prevalencia está asociada a niveles socioeconómicos bajos. En la mayoría de los casos la infección es asintomática; pero puede presentarse también un síndrome de mononucleosis heterófila negativa. La infección en inmunodeprimidos es frecuentemente pulmonar y renal. El problema es más grave cuando se trata de una infección primaria y cuando el órgano transplantado procede de un donante con infección latente. El diagnóstico se basa en general en la presencia de antígenos. El tratamiento es con ganciclovir y con globulinas anti-citomegalovirus (anti-CMV.) Virus Epstein-Barr Como en el caso anterior, es un virus muy frecuente en nuestro entorno (80%) que causa infecciones sintomáticas. El caso clínico más representativo de este virus es la mononucleosis infecciosa caracterizada por fiebre y astenia prolongadas, faringitis, adenopatías múltiples y hepatitis moderada. Este virus tiene poder oncológico y se ha asociado al linfoma de Burkitt (carcinoma nasofaríngeo y linfoma) en pacientes inmunodeprimidos. El diagnóstico es principalmente de laboratorio. POXVIRUS La viruela es una de las enfermedades víricas más graves que actualmente está erradicada desde el punto de vista epidemiológico; pero que constituye un riesgo en términos de guerra biológica. Sus síntomas son parecidos a los de la varicela pero de mayor gravedad. Se distingue la viruela maior con mortalidad próxima al 35% que produce cicatrices en la cara y la viruela minor de baja mortalidad (0.25%) y que no suele producir cicatrices. 66 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Profilaxis: la vacuna de la viruela se desarrolló a partir de una cepa de virus procedente de vacas (vacuna) que se modificó ligeramente para dar lugar al virus vaccinia que no es patógeno para el hombre. La vacuna produce un alto grado de protección. Es una enfermedad de declaración obligatoria y requiere el establecimiento de medidas de cuarentena. VIRUS DE ARN VIRUS INFLUENZA20 Son los causantes de la gripe de la que se conocen tres variantes humanas (A, B y C). Son virus de ARN entre cuyas características estructurales se encuentran la presencia de la proteína M en la membrana (virus envueltos) con la que se diferencian los "tipos", la hemoaglutinina y la neuraminidasa de la envuelta con la que se diferencian los "subtipos". El origen de las epidemias21 de gripe suele estar en animales infectados (generalmente aves) desde los que el virus “salta” a la población humana22. En muchos casos el origen es asiático y se producen pandemias de gripe. El tipo A está sometido a modificaciones antigénicas lo que causa las sucesivas epidemias de gripe de los distintos años. Las vacunas deben prepararse cada año y su fabricación es complicada porque requiere el cultivo de los virus en embriones de pollo con varios meses de anticipación23. La gripe es una enfermedad de declaración obligatoria24 y cursa con fiebre, cefaleas, malestar general, dolores musculares generalizados y tos seca e improductiva. El periodo de incubación es de 1 a 4 días y la enfermedad dura entre 4 y 7 días. La infección puede causar algunas complicaciones de tipo neumonía vírica muy grave, traqueobronquitis, miopatía, cardiopatía o encefalopatías. También puede facilitar la aparición de neumonías bacterianas por S. pneumoniae, Staph. aureus o H. influenzae. 20 Boletín epidemiológico Semanal. Semanas 13-14, 2004. Semanas 18-21, 2004. Se producen epidemias de gripe todos los años. Se denomina pandemia cuando la epidemia se extiende por todo el mundo. Hay registradas tres grandes pandemias de gripe: 1918, 1957 y 1968. 22 Los virus pueden adquirir nuevas características infecciosas cuando varios tipos diferentes infecta a un mismo individuo (animal o humano) combinándose en el transcurso de la infección los genomas de los distintos virus. En el caso del virus de la gripe, esta combinación por coinfección se puede producir en cerdos (infectados por virus que afectan a aves y a humanos) o en humanos directamente (hay unos 15 subtipos diferentes de virus de influenza aviar que pueden infectar humanos). 23 La última variante que está causando preocupación por sus posibles efectos pandémicos es la H5N1 de virus influenza A cuyo origen se encuentra en poblaciones asiáticas de aves y que ha causado varios casos fatales en humanos. Hasta ahora (octubre 2005) no se han detectado casos claros de transmisión entre humanos, aunque sí hay algunos casos sospechosos en el sudeste asiático. Esta falta de transmisión entre humanos ha impedido, por el momento, que se produzca una pandemia de este virus. Sin embargo, la alta mutabilidad del virus influenza podría favorecer cambios en el patógeno que dieran lugar a la epidemia humana. Esta alta mutabilidad del setotipo H5 hace también poco efectivas las vacunas contra este serotipo que están disponibles. 24 El número de casos declarados en España en fue de 1.462.174 en 2003 y 584.574 en 2004. En Navarra, el número de casos declarados en 2004 fue de 1.633) (Datos del Boletín Epidemiológico Nacional. 12: 289-296). 21 67 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) El contagio se produce por contacto con las secreciones de personas enfermas. La enfermedad es más grave en niños menores de ciinco años, ancianos u otras personas con enfermedad respiratoria subyacente. Por tanto es muy importante evitar la infección nosocomial especialmente en salas de enfermos crónicos y en salas de pediatría. El diagnóstico suele ser sintomatológico, aunque se puede complementar con pruebas serológicas. Para el tratamiento se han usado cuatro antivirales (los inhibidores de la replicación del virus25 amantadita y rimantadina, y los inhibidores de la neuraminidasa oseltamivir26,27 y zanamivir) con una efectividad del 70-90% en adultos sanos. Si se toman dentro de los dos primeros días de gripe, se mitigan los síntomas y se acorta 1 ó 2 días la duración de la enfermedad, al mismo tiempo que disminuyen la capacidad del paciente para transmitirla. Estos fármacos se usan bajo control médico en situaciones especiales tales como instituciones donde hay un contacto estrecho (asilos de ancianos, hospitales), en buques cruceros o en entornos similares. No está autorizada la administración de estos antivirales a niños menores de 1 año de edad. Es muy importante la profilaxis mediante la vacunación que debe administrarse cada año debido a las variaciones antigénicas. Esta vacunación es importante para los siguientes grupos de poblaciones: alto riesgo (ancianos con problemas, internos en asilos, crónicos, niños, mujeres embarazadas) • riesgo moderado (ancianos, niños sometidos a terapias de larga duración) • potenciales transmisores a grupos de riesgo (sanitarios en contacto con el primer grupo) • La gripe y el resfriado común son enfermedades que presentan algunos síntomas similares y que tienden a ser confundidas por la población general. Ambas son producidas por virus diferentes. En general, los síntomas de la gripe son más agudos que los del resfriado. Por su parte, los pacientes con resfriado suelen tener mayor congestión nasal o flujo nasal (moqueo.) Los resfriados no suelen presentar complicaciones VIRUS TOSCANA28 (TOSV) Es un virus cuyo material genético consiste en tres moléculas de ARN monocatenario de sentido negativo (RNA-), que se transmite por un insecto (mosca del género Phlebotomus) y es uno de los tres principales agentes causantes de meningitis en España. La 25 Ambos son activos sólo frente a virus influenza A El oseltamivir es comercializado por Roche bajo el nombre de TAMIFLU 27 En el caso del virus H5N1, el antiviral de elección parece ser el oseltamivir ya que el virus estuvo expuesto originariamente a amantadina y rimantadina utilizada de forma negligente por granjeros chinos en la zona de origen del virus (ASM News 71: 420-424). Sin embargo, se ha descrito un caso de virus resistente a oseltamivir en una niña que, por otra parte, puede corresponder a un caso de transmisión de este virus (de pollo) entre humanos. 28 Emerging Infectious Diseases 11: 1657-1663 26 68 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) enfermedad es más abundante en áreas rurales donde hay una gran actividad exterior. Se calcula que el porcentaje de seropositivos puede llegar a ser del 26%. La enfermedad es en la mayoría de los casos benigna con pronóstico favorable 8aunque hay algunos casos con complicaciones). El periodo de incubación va desde unos pocos días a dos semanas y cursa con dolor de cabeza, fiebre, náuseas y vómitos y rigidez del cuello (como síntomas más frecuentes). En la mayoría de los seropositovos la enfermedad ha cursado como asintomática o paucisintomática. La meningitis causada por TOSV se incluye dentro de las denominadas asépticas. En la actualidad se está considerando la conveniencia de realizar análisis para detectar este virus en las donaciones de sangre para evitar esta vía de contagio. PAROTIDITIS El virus de las paperas afecta a las glándulas parótidas causando un cuadro febril con inflamación bilateral, malestar general y otalgia. Puede haber, con menor frecuencia, inflamación de otras glándulas salivares, ovarios o testículos. En general, la evolución es benigna. El periodo de incubación es de 16 a 18 días. El contagio se produce por la saliva infectada. El diagnóstico es esencialmente clínico. No hay un tratamiento específico. La profilaxis se realiza con una vacuna atenuada que se administra de forma obligatoria a los 15 meses de edad en una dosis única. La inmunidad adquirida dura toda la vida. SARAMPIÓN Es una infección de declaración obligatoria. Era una infección muy frecuente en la población infantil hasta la implantación de la vacunación obligatoria. Actualmente en España se considera que se ha eliminado29 y su incidencia es menor un caso por millón de habitantes. Normalmente se manifiesta por unos pródromos respiratorios (secreción nasal, congestión ocular), seguidos de la aparición de unas manchas en la mucosa bucal (enantema de Koplik), fiebre alta y exantema maculopapular que dura entre 2 y 5 días. El periodo de incubación es de 14 días. La transmisión se efectúa por vía aérea a partir de secreciones de los enfermos. 29 Eliminación según los criterios de la OMS que son una incidencia menor de un caso por millón, ausencia de casos durante un periodo de tiempo superior al de la transmisión en todo el territorio nacional, variabilidad en los genotipos circulantes y tasa de reproducción R<1. (Boletín Epidemiológico Semanal 13: 49-56) 69 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Las complicaciones más frecuente son la neumonía vírica, e infecciones bacterianas de oído o vías respiratorias. En aproximadamente 1/1000 casos se producen complicaciones neurológicas en forma de encefalitis. En pacientes inmunodeprimidos, las complicaciones son más frecuentes e importantes. Como en otros casos de infecciones producidas por virus ARN, la enfermedad tiene un pronóstico más favorable en niños que en adultos. El diagnóstico es sintomatológico salvo en los casos complicados. La profilaxis es muy eficaz, la vacuna consistente en un virus atenuado se administra en una dosis única a los 15 meses de edad. Es recomendable administrar una dosis de recuerdo a los 4-5 años para cubrir un 1-5% de fallos que se producen en la primera inmunización. VIRUS RESPIRATORIO SINCITIAL Es la causa más frecuente de hospitalización por problemas respiratorios en los niños de menos de 2 años de edad. Es el agente causante de los cuadros de bronquilotis que causa dificultades respiratorias, rinorrea e hiperventilación en niños pequeños. Esta infección puede tener mal pronóstico si afecta a niños lactantes con problemas o a prematuros. Es un agente causante de infección nosocomial por lo que es necesario identificar rápidamente a los pacientes que ingresen por este motivo para evitar la propagación de la enfermedad. En este caso es muy importante el aislamiento de los pacientes para evitar la transmisión de la enfermedad. En los casos de complicaciones más graves, el antiviral que se usa es la ribavirina en aerosol (nucleósido sintético, antiviral de amplio espectro.) CORONAVIRUS Los coronavirus son virus de ARN de cadena simple, envueltos y caracterizados por presentar unas proyecciones de glicoproteína que les dan un aspecto de corona. Se asocian principalmente a infecciones leves de las vías respiratorias superiores, especialmente en jóvenes. Se calcula que entre el 10 y el 20 % de los resfriados comunes de invierno son producidos por coronavirus. En algunos casos, la infección puede producir complicaciones intestinales (enteritis necrosante) en niños de corta edad. En 2003 se describió el síndrome Respiratorio Agudo Grave (SARS)30, conocido como neumonía atípica asiática, producido por un coronavirus de origen desconocido. El SARS es una enfermedad emergente de origen probablemente zoonótico ya que los coronavirus son patógenos importantes de animales (cerdos, aves) aunque hasta ahora no se había descrito ningún caso de problemas serios en humanos. 30 Boletín Epidemiológico Nacional. Semanas 15-16. 2004 70 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Los síntomas son parecidos a los de una gripe (fiebre, escalofríos y temblor, dolores musculares, tos, dolor de cabeza) con una diarrea inicial. La enfermedad es grave, con unas tasas de mortalidad de en torno al 15% en los casos diagnosticados y de más del 50% en pacientes mayores de 60 años. La vía de transmisión del SARS es el contacto directo. La enfermedad se manifiesta en 2/3 de los afectados por una diarrea fuerte que libera muchos virus, lo que sugiere que la vía oro-fecal puede servir para la transmisión en condiciones de hacinamiento. La tasa de mutación del virus no es muy alta, lo que favorece la producción de vacunas. El sistema inmune tarda 1 –2 semanas en controlar la enfermedad. Es importante el aislamiento de los casos sospechosos. El personal sanitario es población de riesgo. PICORNAVIRUS Los picornavirus son pequeños virus de ARN que incluyen rinovirus (causantes de infecciones respiratorias no graves de las vías superiores) y enterovirus. Los enterovirus se agrupan en varios serotipos (poliovirus, coxsackievirus, ecovirus y virus de la hepatitis A) que penetran en el organismo por vía oral, se multiplican en el tubo digestivo desde donde invaden otros tejidos, permanecen en el intestino o se eliminan por las heces. Dada la gran variedad de serotipos los cuadros que producen son también muy variados, entre ellos están la poliomielitis, meningitis linfocitaria (de curso benigno) infección respiratoria aguda, exantemas víricos y hepatitis A. El diagnóstico se basa en el aislamiento del virus y su cultivo. Poliovirus La poliomielitis es una grave infección vírica de declaración obligatoria causada por un enterovirus que se multiplica en la garganta, el intestino y de allí pasa a la sangre donde da lugar a una infección sistémica. En la mayoría de los casos la infección produce síntomas inespecíficos, dolor de garganta y cefaleas que son superados sin mayores problemas y la enfermedad termina en esta fase. Sin embargo, el virus puede producir una enfermedad paralizante grave como consecuencia de la infección de la médula espinal o el tronco cerebral. En los casos más graves la parálisis bulbar puede ser fatal. El hombre es único reservorio conocido del virus y los enfermos lo excretan en grandes cantidades en las heces, por lo que el contagio es oro-fecal. La enfermedad es más grave en los países desarrollados que en los subdesarrollados, probablemente porque en estos se producen muchas infecciones en niños muy pequeños en los que el virus produce una enfermedad benigna sin mayores complicaciones. 71 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) La profilaxis se realiza con la vacuna de Salk (basada en un virus muerto inactivado) y con la vacuna de Sabin (basada en un virus atenuado) que se administra en varias dosis. No existe un tratamiento específico contra este virus. Virus de la hepatitis A Hepatitis A. Es el principal tipo de hepatitis contraído por contagio oro-fecal. Es una enfermedad de declaración obligatoria. En los niños la enfermedad suele cursar sin síntomas aparentes mientras que en los adultos se manifiesta como una hepatitis evidente. Por esto, como en el caso de la polio, los casos de hepatitis A clínica son más frecuentes en los países desarrollados que en los subdesarrollados. El periodo de incubación es de un mes y la duración de uno a dos meses. No parece producir enfermedad hepática crónica, aunque puede haber casos de recaídas. El contagio se produce por el consumo de alimentos lavados con aguas contaminadas y por el consumo de moluscos que han acumulado el virus por filtración de aguas contaminadas. El enfermo elimina virus por las heces durante la primera fase de la enfermedad. La prevención se basa en las medidas higiénicas. También puede usarse la inmunización pasiva con inmunoglobulinas. VIRUS DE LA RUBÉOLA Es una enfermedad exantemática benigna propia de la infancia. Su importnacia está en el poder teratógeno del virus cuando infecta a una mujer embarazada. En estos casos, el feto se infecta también por el virus, especialmente si la enfermedad aparece durante los primeros tres meses de embarazo, y el riesgo de malformación fetal es muy alto: semana de infección % infección feto menos de 4 84 5-8 70 9-12 50 13-16 >16 - % malformación feto > 90 > 90 > 90 35 prácticamente nulo El diagnóstico de la enfermedad es fundamentalmente serológico. Toda mujer en edad gestacional debe conocer su estado inmune frente a la rubéola, sobre todo si no se ha vacunado previamente. La vacuna consiste en virus atenuados y 72 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) se administra en dos dosis, una a los 15 meses y una segunda dosis a las niñas a los 11 años. ROTAVIRUS Son la causa más frecuente de gastroenteritis en niños menores de dos años. Producen muchos brotes epidémicos en guarderías y salas de pedriatría. La incidencia mundial de esta enfermedad se resume en el cuadro siguiente que muestra los caso por año: casos tratados en el domicilio visitas clínicas hospitalizaciones Muertes 111 millones 25 millones 2 millones 400.000 – 500.000 Se estima que a los cinco años prácticamente todos los niños han tenido un episodio de rotavirus. Los niños de países subdesarrollados dan cuenta del 80% de las muertes por esta enfermedad La transmisión es oro-fecal. El periodo de incubación dura entre uno y cuatro días. La época más frecuente para la enfermedad es el invierno. La infección produce diarrea, vómitos y fiebre. La pérdida de líquidos puede hacer necesaria la hospitalización. El tratamiento está basado en la rehidratación por vía oral y en medidas de apoyo. En los países subdesarrollados el desenlace puede ser fatal por la deshidratación del enfermo. En las salas de pediatría son frecuentes las infecciones epidémicas porque se llegan a liberar 1010 viriones por gramo de heces y la dosis mínima infectiva está en torno a los 10 viriones para iniciar la infección. No hay ninguna vacuna disponible frente a rotavirus. HEPATITIS VÍRICAS La hepatitis es una inflamación hepática caracterizada por ictericia, hepatomegalia, elevación de los marcadores bioquímicos de daño hepático y síntomas inespecíficos. Las causas pueden ser tanto infecciosas como no infecciosas. Existen varios tipos de hepatitis infecciosa. En el cuadro siguiente se indican las principales características de las mismas: Virus Tipo A ARN B ADN 73 C ARN D ARN E ARN GB ARN MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Periodo de incubación (días) Edad habitual (años) Transmisión fecal Transmisión sexual Transmisión parenteral Gravedad Cronicidad 15-45 <45 +++ + ++ 0% 7-190 15-160 todas todas +/+++ + ++ +++ +++ ++ 10% 50% 15-90 todas +++ +++ - 15-45 <25 +++ + 0% ? ? +/? + + ? El virus de la hepatitis A es un picornavirus ya descrito. HEPATITIS B Es una enfermedad de declaración obligatoria causada por el virus de la hepatitis B (VHB.) En España se calcula que se producen unos 12.000 casos anuales de hepatitis B y que un 1% de la población adulta es seropositiva para el antígeno de superficie o antígeno Australia. El periodo de incubación es de unos tres meses, la mayoría de las personas infectadas no presentan síntomas y en el 90% de los casos la enfermedad se resuelve mientras que en el 10% de los casos evoluciona hacia la cronicidad. En un pequeño porcentaje de casos (1%) se produce hepatitis fulminante, sobre todo si hay coinfección con el virus delta. Las vías de contagio son principalmente la parenteral, la sexual y la perinatal. También es importante la transmisión intrafamiliar a causa del estrecho contacto de los miembros. El personal sanitario tiene alto riesgo por la incidencia de pinchazos accidentales y por el contacto con la sangre de los pacientes. Deben aplicarse normas de conducta en este personal para evitar el contagio. La profilaxis se realiza mediante vacunación y administración de gammaglobulina. La vacunación está indicada para el personal sanitario, pacientes de diálisis, recién nacidos de madres portadoras, personas con contacto sexual con personas portadoras o con prácticas sexuales de riesgo. Es necesario el control del VHB en la sangre de las transfusiones. El curso de la enfermedad puede seguirse mediante serología de los diferentes antígenos. El tratamiento con antivirales puede ayudar a controlar los casos crónicos. HEPATITIS C 74 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) El virus de la hepatitis C (VHC) es un virus ARN que puede transmitirse por vía parenteral y es el causante del 80 - 90% de las hepatitis post-transfusionales. Actualmente, se controla la presencia de este virus en la sangre empleada en las transfusiones. Es muy frecuente en drogadictos, transplantados y hemodializados. El riesgo de contagio por pinchazo con material infectado es relativamente bajo (4%.) También es bajo el de transmisión vertical (5%), y el de transmisión sexual (5 -10 %). Alrededor del 40% de los infectados no pertenece a ningún grupo de riesgo claro. La enfermedad es asintomática en la mayoría de los casos. Sólo en algunos casos es aguda y la hepatitis fulminante es muy inusual. La infección crónica evoluciona de forma diferente según los pacientes. En aproximadamente el 50% evoluciona hacia cirrosis y de estos el 15% desarrolla hepatocarcinoma. Este virus no ha podido ser cultivado y su detección se basa en métodos moleculares de PCR. El tratamiento se basa en ribavirina en combinación con interferón α. Es necesario comprobar la presencia de VHC en la sangre usada en transfusiones. HEPATITIS D El virus es de ARN, de pequeño tamaño (virus delta) y necesita la presencia del virus B para multiplicarse. Las vías de transmisión son similares a las del VHB; pero el riesgo es mucho menor. En general la presencia de este virus empeora el pronóstico de la enfermedad causada por VHB. HEPATITIS E Y GB El virus de la hepatitis E es de ARN y cadena simple, de transmisión principalmente oro-fecal. Es más frecuente en los países subdesarrollados donde afecta principalmente a adultos jóvenes. La hepatitis GB se ha descrito recientemente y está causada por un virus parecido al de la hepatitis C. Parece responsable de cuadros de hepatitis de evolución generalmente benigna donde no se ha podido encontrar otro agente etiológico. La transmisión parece ser por vía parenteral y oro-fecal. VIRUS DE LA INMUNODEFICIENCIA HUMANA (VIH)31 31 Boletín Epidemiológico Semanal. Semana 47. 2004 75 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) El VIH es un retrovirus que posee una única cadena de ARN y que para poder expresarse necesita integrarse en forma de ADN en el genoma de la célula huésped. El paso de ARN a ADN lo realiza mediante la acción de una enzima denominada transcriptasa reversa. Hay dos tipos de VIH (tipo 1 y tipo 2). El VIH 1 es el más ampliamente distribuido y el que se encuentra en España. El síndrome de inmunideficiencia adquirida es la manifestación de la infección por VIH y se produce por una inmunosupresión de tipo celular muy grave. Como consecuencia de ella, se producen infecciones oportunistas y desarrollos tumorales que normalmente no serían un problema al ser reprimidos por el sistema inmune celular. La infección primaria da lugar a un cuadro de tipo gripal que luego remite y permanece latente durante años. Posteriormente surgen infecciones oportunistas como candidiasis bucales. A partir de este momento las infecciones y tumores se van haciendo más frecuentes y el paciente sobrevive un tiempo variable que, por lo general, no es inferior a dos años. Actualmente, con el uso de tratamientos antirrretrovirales, el tiempo de supervivencia ha aumentado mucho y no se puede fijar un límite máximo. Las enfermedades más frecuentes que presentan los enfermos de SIDA son: enfermedades fúngicas como candidiasis y criptococosis enfermedades bacterianas como tuberculosis y salmonelosis enfermedades parasitarias como criptosporidiasis (diarrea crónica), toxoplasmosis y neumonía producida por Pneumocystis carinii. • enfermedades virales producidas por herpesvirus • tumores como el sarcoma de Kaposi • • • Las vías de transmisión son la sexual parenteral y vertical. En la prevención es importante no realizar prácticas sexuales con personas desconocidas sin usar preservativo, evitar contacto con las mucosas o piel sangrante, no compartir agujas, cuchillas, etc. El diagnóstico es por serología de los anticuerpos. Los resultados positivos deben confirmarse por técnicas moleculares. El tratamiento está basado en antirretrovirales inhibidores de la transcriptasa reversa (AZT) o de su procesamiento (inhibidores de la proteasa viral.) Actualmente se utilizan terapias combinadas usando varios antirretrovirales simultáneamente para evitar la aparición de cepas resistentes. No hay vacuna disponible. 3.- MEDIDAS DE SEGURIDAD PARA EL PERSONAL SANITARIO 76 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Los virus VIH, VHB y VHC pueden contagiarse por inoculación con sangre infectada, por lo que deben tomarse las máximas precauciones para evitar el contagio. El mayor riesgo de contagio es por el VHB por el alto número de viriones presentes en la sangre de los infectados; el riesgo de infección por VHC es más bajo. En el caso del VIH el riesgo es relativamente pequeño debido al bajo número de viriones circulantes en sangre incluso en las fases de infección muy aguda (donde se recuentan entre 10 y 1.000 partículas invectivas por ml de sangre). Sin embargo, aunque el riesgo de contagio por VIH sea más bajo que por VHB, el riesgo de muerte es prácticamente el mismo a causa de la mayor letalidad del VIH. El uso de guantes debe ser constante mientras se manipula material posiblemente contaminado, y debe evitarse: • manipular material cortante contaminado • encapuchar agujas • arrojar objetos cortantes o punzantes a la basura • usar pipetas con la boca • comer, fumar o maquillarse en zonas posiblemente contaminadas En caso de accidente, debe comunicarse inmediatamente y si este se ha producido con material contaminado puede ser necesario iniciar un tratamiento de inmunización activa contra VHB o de quimioprofilaxis frente a VIH (aunque la eficacia de este último no está totalmente clara). 4.- PRINCIPALES GRUPOS DE HONGOS DE IMPORTANCIA CLÍNICA. Los hongos son microorganismos eucarióticos que tienen una pared celular de composición diferente a la de las bacterias. Se pueden distinguir dos tipos de organización de los hongos: • los hongos filamentosos que forman unas estructuras tubulares microscópicas denominadas hifas que se agregan dando lugar a una estructura denominada micelio que les da es aspecto de mohos. • hongos unicelulares denominados levaduras que presentan características de crecimiento similares a las de las bacterias. En ciertos casos los hongos pueden ser dimórficos y pasar de una forma micelial a otra levaduriforme en función de las condiciones ambientales. Este cambio de morfología puede estar relacionado con el aumento de la patogenicidad o de la virulencia del hongo. Los hongos producen cuatro tipos de infecciones: • Micosis superficiales o dermatomicosis • Causadas por dermatofitos o tiñas. • El origen del hongo puede ser diverso y el contagio es por contacto. • El tratamiento es por antimicóticos tópicos y por antifúngicos orales como la griseofulvina. • Micosis subcutáneas. 77 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Las más frecuentes son las esporotricosis y los micetomas producidos por la infección de heridas superficiales. Micosis profundas • Producidas por hongos que se multiplican dentro del cuerpo. • Estas infecciones pueden variar desde asintomáticas hasta mortales. • Entre las más representativas está la criptococosis (Cryptococcus neoformans) que produce meningitis en los enfermos de SIDA Micosis oportunistas (que pueden ser superficiales o profundas) • Producidas por hongos que causan infección cuando el sistema inmune se encuentra debilitado. • Entre las más importantes se encuentran • Criptococosis (C. neoformans) • Candidiasis (Candida albicans) • Aspergilosis (Aspergillus fumigatus). • Otras infecciones fúngicas graves se producen al infectar los microorganismos las prótesis quirúrgicas, especialmente las válvulas cardiacas (endocarditis fúngicas sobre válvulas protésicas). • Mucormicosis (hongos de los géneros Mucor, Rhizopus, etc.) que infectan pacientes hematológicos y pacientes con cetoacidosis diabética. • • • Hay algunos hongos que pueden producir más de un tipo de infección (C. neoformans produce meningitis en un enfermo de SIDA y a la vez es un patógeno oportunista.) El tipo de enfermedad producido por un hongo depende de: En tropismo del hongo el estado inmunológico del paciente • • En los últimos años ha aumentado la incidencia de micosis debido al aumento de pacientes inmunodeprimidos. En general, las micosis son infecciones poco graves; pero crónicas, aunque en los pacientes inmunodeprimidos su gravedad aumenta. El método de diagnóstico es el cultivo del material infectado en medios adecuados como el agar de Sabouraud. Los tratamientos se basan en antimicóticos tópicos y en fungicidas orales como la griseofulvina. REFERENCIAS A continuación se relacionan los principales libros de Microbiología Médica disponibles en castellano. Para el curso de Microbiología Clínica se recomiendan especialmente los de de la Rosa (1997) y García Rodríguez (1996.) 78 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) 1. Ballows A, Hauser-Jr WJ, Herrmann KL, Isenberg HD y Shamody HJ. 1991. Manual of clinic Microbiology 5th Edition. American Society for Microbiology. Washington D.C. 2. de la Rosa M. 1997. Microbiología. Enfermería-Ciencias de la Salud. Conceptos y Aplicaciones. Harcourt Brace de España S.A. Barcelona. 3. García-Rodríguez JA y Picazo JJ. 1996. Microbiología Médica (Vols. 1 y 2.) Mosby/Doyma Libros S.A. Madrid Ingraham JL y Ingraham CA. 1998. Introducción a la microbiología. Ed. Reverté. Barcelona 4. Murray PR, Rosenthal KS, Kobayashi GS y Pfaller MA 2002. Microbiología Médica. 4ª Edición.. Mosby Inc. (Elsevier). Madrid 5. Pumarola A, Rodríguez-Torres A, García-Rodríguez JA y Piédrola-Angulo C. 1987. Microbiología y parasitología médica. Ediciones Científicas y Técnicas S.A. Barcelona Stuart Walker T. 2000. Microbiología. MacGraw-Hill Interamericana. México D.F. 79 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Tema 4.- Genética microbiana. Elementos genéticos bacterianos. Transferencia horizontal de material genético en microorganismos: transformación, conjugación y transducción. Elementos genéticos de los virus. Expresión de la información genética. conclusión 1.- INTRODUCCIÓN ESTRUCTURA DE LOS ÁCIDOS NUCLEICOS Hay dos tipo de ácidos nucleicos (AN) en las células: el ácido desoxirribonucleico (ADN) y el ribonucleico (ARN). El ADN está formado por bases nitrogenadas de tipo púrico (adenina y guanina) y pirimidínico (citosina o timina) unidas al azúcar (desoxirribosa) que forma una cadena en la que se alterna con el ácido fosfórico. La composición del ARN se diferencia de las del ADN en que el azúcar es la ribosa y en que la base pirimidínica timina es substituida por el uracilo. Al conjunto de una base nitrogenada, una molécula de azúcar y un grupo fosfato se le denomina nucleótido, y a una cadena de nucleótidos (una molécula de AN) polinucleótido. Se denomina estructura primaria de un AN a la secuencia de sus nucleótidos. Dos cadenas de AN pueden unirse de forma no covalente mediante el establecimiento de puentes de hidrógeno entre sus bases nitrogenadas. Esta asociación se produce de forma específica de manera que se forman cuatro tipo de parejas de nucleótidos A:T, T:A, C:G y G:C. Cuando dos cadenas de AN con secuencia complementaria se asocian de la forma descrita, se forma una doble hélice de AN que es la forma en la que se encuentra por lo general el ADN y algunas partes de las moléculas de ARN. La estructura tridimensional de los AN se denomina estructura secundaria. En las células la información genética se conserva en moléculas de ADN que se transmiten de generación en generación. Durante el ciclo celular, las moléculas de ADN de la célula se replican mediante la actividad de un conjunto de enzimas entre las que destaca la ADN polimerasa que sintetiza una molécula de ADN complementaria a la que usa como molde. La información almacenada en el ADN se transfiere al ARN en un proceso denominado transcripción en el que un conjunto de enzimas entre las que destaca el sistema de la ARN polimerasa que sintetiza una molécula de este AN complementaria a una de las cadenas de la moléculas de ADN que utiliza como molde. Existen varios tipos de ARN en las células: • ARN mensajero que resulta de la transcripción de aquellas regiones del genoma que codifican genes funcionales que van a ser traducidos en proteínas. Este tipo de ARN constituye sólo una pequeña parte del ARN celular total (en torno al 10%). • ARN estable que constituye la mayor parte del ARN celular e incluye el ARN ribómico que forma parte de los ribosomas (orgánulos encargados de la síntesis de proteínas), el ARN de transferencia que participa en la síntesis de proteínas leyendo 80 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) la información del ARN mensajero e insertando los aminoácidos correspondientes, y el ARN heterogéneo nuclear que cumple diferentes funciones en el núcleo de las células. 2.- ELEMENTOS GENÉTICOS BACTERIANOS Se denomina nucleoide la zona de la bacteria donde se encuentra su material genético. En el caso de las bacterias, éste está formado generalmente por un cromosoma cerrado y por elementos extracromosómicos denominados plásmidos. CROMOSOMAS BACTERIANOS Los cromosomas de las bacterias están formado por ADN, pequeñas cantidades de ARN y por proteínas que sirven para mantener su estructura. La longitud de los genomas bacterianos varía en torno a las 3-6 Mbp32. Actualmente (28/3/2005) están disponibles en la base de datos los genomas de 355 especies bacterianas entre los que se encuentra un buen número de bacterias patógenas a las que hay que añadir varios genomas de protozoos y hongos patógenos. Teniendo en cuenta la velocidad de secuenciación de genomas en la actualidad, este número se incrementará de forma muy importante durante los próximos años de forma que conocer la secuencia de un microorganismo patógeno (al menos de patrones para los diferentes patógenos) será algo habitual en la mayoría de las enfermedades infecciosas comunes. La secuenciación de los genomas de microorganismos patógenos es importante desde el punto de vista clínico porque permite: • Identificar los factores de virulencia del patógeno y las bases moleculares de su fisiología, lo que permite diseñar agentes quimioterápicos más específicos y eficientes. • Identificar las bases genéticas de la variabilidad de los patógenos que les permiten escapar de los sistemas de defensa inmune del organismo huésped. • Diseñar nuevos sistemas de detección de microorganismos más sensibles y específicos al estar basados en la detección de secuencias genéticas del patógeno. REGULACIÓN DE LA EXPRESIÓN GÉNICA En el genoma bacteriano se encuentran regiones que codifican genes estructurales que se transcriben y traducen en proteínas y otras regiones que codifican genes reguladores cuya función es controlar la expresión ordenada de los genes estructurales. 32 Como una generalización simplista, los tamaños de los diferentes elementos genéticos son los que se indican en la siguiente tabla: Elemento o genoma Tamaño Plásmido 3x103 3 Kpb Virus bacteriano (fago λ) 50x103 50 Kpb 3.5 Mpb Célula procariótica (E. coli) 3.5 x 106 14 Mpb Célula eucariótica (S. cerevisiae) 14 x 106 3.5Gpb Genoma humano 3.5x 109 Es decir, la relación de tamaño entre un plásmido y una célula bacteriana es de 1 a 1000, y la que hay entre una célula bacteriana y una humana es de 1 a 1000 también. 81 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Esta expresión génica regulada por los genes reguladores se produce como respuesta a señales ambientales que causan la modulación (inducción o represión) de la expresión de grupos de genes que se controlan de forma simultánea (operones). Según la forma de respuesta a las condiciones externas, los operones pueden ser inducibles represibles (ver más adelante). Los sistemas de regulación de la expresión génica en eucariontes son más sofisticados, aunque en ciertos aspectos recuerdan el modelo básico del operón. En células eucarióticas, sin embargo, los genes se regulan de forma individual (no por grupos) y el ARNm transcrito debe procesarse para eliminar los intrones antes de ser exportado al exterior del núcleo celular para ser traducido. VARIACIONES EN LA INFORMACIÓN GENÉTICA La información genética de una bacteria puede variar por dos mecanismos diferentes: mutación o cambio en la secuencia que producen cambios en la información genética; y por transferencia horizontal de material genético proceso por el que se comparte información genética entre dos organismos que comparten un mismo ambiente. La variación en la información genética es una causa importante de aparición de nuevas resistencias a antibióticos, de variaciones en los mecanismos de escape de los sistemas de defensa inmunológica y de variaciones en los factores de virulencia del patógeno. Las mutaciones se producen espontáneamente en el material genético de cualquier organismo con una frecuencia muy baja (en torno a 10-6 por posición y ciclo de división celular)33. Una nueva mutación, si no confiere ninguna ventaja selectiva al organismo que la presenta, suele desaparecer de la población en unas pocas generaciones. Esta desaparición es más rápida, todavía, si la mutación es deletérea para el organismo que la porta. Sin embargo, si la mutación produce algún tipo de ventaja adaptativa para el organismo que la tiene, su frecuencia aumentará en la población hasta llegar a hacerse determinante. Por ejemplo, una mutación que confiera resistencia a un antibiótico puede perderse en un entorno libre de antibióticos porque o bien no confiere ninguna ventaja al microorganismo resistente en ausencia del antibiótico o, incluso, porque puede causar al microorganismo mutado alguna deficiencia en relación con los competidores que lo lleve a la desaparición. Sin embargo, ese mismo microorganismo mutado en un ambiente en el que esté presente el antibiótico frente al que es resistente, tendrá una ventaja selectiva frente a los competidores de forma que se convertirá rápidamente en el dominante (o en el único) en la población. Por esto, es muy importante que los tratamientos con antibióticos se realicen correctamente de forma que se evite la selección de cepas resistentes que inutilizan el antibiótico en el futuro. Puesto que muchas de estas mutaciones de resistencia se producen en elementos genéticos transmisibles (plásmidos) su diseminación entre diferentes poblaciones micro33 Es decir, se produce una célula de cada millón es portadora de una mutación en un punto dado del genoma. 82 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) bianas pude ser muy rápida lo que causa graves problemas con las cepas hospitalarias que suelen ser portadoras de resistencias múltiples (por ejemplo cepas multirresistentes de Staph. aureus). PLÁSMIDOS BACTERIANOS Se denominan plásmidos a los elementos genéticos extracromosómicos que pueden llevar ciertas cepas bacterianas y que codifican información genética prescindible para la supervivencia del microorganismo en ambientes naturales. Los plásmidos son moléculas de ADN normalmente cerrado (aunque también los hay de ADN abierto) que se replican independientemente del cromosoma o cromosomas bacterianos. El tamaño de los plásmidos es mucho menor que el de los cromosomas y puede oscilar (como ejemplo general) en torno a las 3 a 10 kbp. El número de plásmidos por célula es variable, y pueden coexistir varios tipos de plásmidos en la misma célula. También es variable el número de copias de un determinado plásmido, aunque este factor está controlado genéticamente (hay plásmidos de bajo número de copias y plásmidos de alto número de copias). Los plásmidos deben llevar entre sus genes un origen de replicación que permita controlar su multiplicación en la célula en la que se encuentran Se puede utilizar el perfil de plásmidos de un microoganismo para identificar a qué cepa pertenece. Cuando no se tiene evidencia de qué tipo de función realizan los genes que lleva un plásmido determinado, se dice que se trata de un plásmido críptico; sin embargo, en muchos casos, sí puede asignarse una función a dichos genes plasmídicos. Entre estas funciones destacan las siguientes: 1. Codificación de factores de virulencia en algunas especies. • Plásmidos inv que confieren invasividad intestinal a microorganismos enteroinvasivos • Plásmidos ent que codifican enterotoxinas 2. La codificación de factores de resistencia a antibióticos. P. ej., la producción de β-lactamasas plasmídicas. 3. Los plásmidos sexuales que confieren a la bacteria que los porta la capacidad para transferir información genética de forma horizontal mediante la conjugación bacteriana. Este plásmido se denomina plásmido F y es portador, entre otros, de los factores genéticos responsables de la producción del pelo F.) 3.- TRANSFERENCIA HORIZONTAL DE MATERIAL GENÉTICO EN MICROORGANISMOS Se denomina transferencia horizontal de información genética a la transmisión de material genético entre dos células que conviven en un mismo ambiente o que no están relacionadas entre sí por un vínculo de descendencia directa. A la transferencia de in- 83 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) formación genética de una célula madre a las hijas se le denomina transferencia vertical. Existen tres mecanismos de transferencia horizontal de información genética: la transformación, la transducción y la conjugación. TRANSFORMACIÓN La transformación genética consiste en la captación de material genético que esté libre en el medio por una célula bacteriana que lo incorpora posteriormente a su propio material genético y lo expresa como propio. El material genético libre puede proceder de diferentes orígenes tales como restos celulares de bacterias lisadas, plásmidos liberados por otras células, etc. Para que una célula pueda tomar material genético del medio en el que se encuentra y, de esta forma, transformarse, debe encontrarse en un estado fisiológico especial denominado competencia. Por tanto, una célula competente es aquella que puede ser transformada genéticamente. Existen especies bacterianas que son competentes naturales (por ejemplo, el neumococo) y que, por tanto, siempre pueden captar ADN del medio externo; mientras que otras pueden ser competentes inducidas cuando se les somete a un tratamiento que les confiere esta capacidad. Probablemente las especies competentes inducidas puedan comportarse como competentes naturales en determinadas circunstancias por lo que la característica de la competencia puede estar muy extendida entre las bacterias. Una vez que el ADN externo ha sido introducido dentro de la célula competente, este material puede ser destruido por la célula receptora mediante sus mecanismos de restricción consistentes en endonucleasas que digieren el ADN extraño en sitios en los que hay una secuencia determinada. (endonucleasas de restricción). Estas enzimas constituyen, por tanto, un sistema por el que las células bacterianas son capaces de eliminar el material genético extraño que entra en su interior. Para evitar que las endonucleasas de restricción degraden el material genético propio, existe otro sistema enzimático denominado de enzimas de modificación que colocan grupos metilo (-CH3) en las secuencias del ADN que reconocen las enzimas de restricción correspondientes y, de esta forma, evitan que la secuencia modificada sea degradada por ellas. Al conjunto de los dos sistemas se les conoce como sistema de modificación /restricción de la célula. Si una molécula de ADN que ha entrado en una bacteria por transformación no ha sido eliminada por el sistema de modificación/restricción, puede replicarse autónomamente si posee un origen de replicación, puede integrarse por recombinación en alguno de los elementos genéticos de la bacteria o perderse al dividirse la célula. En cualquiera de los dos primeros casos, la nueva información genética se podrá expresar y, si confiere ventajas selectivas a la célula transformada, establecerse en la población. La eficiencia del proceso de transformación es baja; sin embargo, dado el elevado número de microorganismos que se pueden encontrar en ciertas poblaciones bacterianas (más de 1011 células por gramo de materia intestinal, por ejemplo) la transformación es 84 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) un mecanismo de diseminación de información genética importante entre bacterias en ambientes clínicos y en ecosistemas humanos. CONJUGACIÓN La conjugación es la transferencia directa de información genética desde una bacteria donadora a otra aceptora utilizando el Pelo F como canal de paso del material genético. Las especies de las bacterias donadora y aceptora pueden ser diferentes. Para que una bacteria sea donadora en un proceso de conjugación debe llevar un plásmido conjugativo (sexual) que se suele llamar plásmido F. Una bacteria portadora de plásmido F, se dice que es F+. Una bacteria F+ es capaz de pasar su plásmido F a otra que carezca de él (F-) convirtiendo a esta aceptora en una nueva célula F+ y transmitiéndole todos los genes que se encuentren en el plásmido conjugativo F. Puesto que cada bacteria aceptora se convierte en una nueva donadora F+, el proceso de conjugación es muy eficiente y da lugar a un cambio rápido en la población de forma que los genes presentes en el plásmido F se diseminan por toda la población. En ciertas ocasiones, el plásmido F se puede introducir por recombinación en el cromosoma bacteriano dando lugar a una célula Hfr. Cuando una célula Hfr transmite información genética por conjugación a una célula aceptora F-, en vez de transmitir únicamente los genes contenidos en el plásmido F, transferirá la información genética presente en el cromosoma bacteriano en el que se integró el plásmido F. En este caso puede llegar a transferirse toda la información genética de la célula donadora a la aceptora. La conjugación puede producirse entre microorganismos de diferentes especies. TRANSDUCCIÓN La conjugación bacteriana se produce cuando un virus bacteriano (bacteriofago) infecta una célula bacteriana y en el momento de formar los nuevos viriones resultantes del ciclo de vida del fago, éstos incluyen en su material genético parte del material genético de la bacteria. De esta forma, cuando un fago que lleva material genético de una bacteria infecta a otra, esta pequeña cantidad de material genético de la primera bacteria se transmite horizontalmente a la segunda. La transducción por fagos puede ser generalizada cuando el fago es capaz de incluir en su partícula vírica un fragmento del cualquier parte del genoma de la bacteria que infecta o restringida cuando sólo se incluyen ciertas secuencias determinadas en la partícula vírica. Los fagos (de forma similar a como ocurre con otros tipos de virus) pueden desarrollar dos tipos de ciclo biológico: un ciclo lítico en el que se forman inmediatamente después de la infección nuevos viriones que salen de la célula infectada rompiéndola (lisis) para proseguir la infección; o un ciclo lisogénico en el que el material genético del fago se integra en el genoma de la bacteria infectada y se mantiene como un grupo de genes bacterianos más que se multiplica conjuntamente con el resto del material genético de la célula huésped (lisógeno o fago atemperado). Cuando se produce un cambio en las 85 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) condiciones ambientales en las que se encuentra la bacteria, el fago lisogénico puede activarse de nuevo para pasar a un ciclo lítico y reiniciar el proceso infectivo. En este momento, se puede producir la captación de material genético de la bacteria que producirá la transducción. La importancia de la transducción puede ser grande a nivel global. En ambientes naturales (acuáticos y terrestres) el la abundancia de virus excede a la de procariotas en un factor de 10 a 1, y en secuenciación de metagenomas34 se han encontrado numerosos casos de virus portadores de elementos móviles (plásmidos y transposones). Ciertos fagos son portadores de factores de virulencia responsables de patologías graves; por ejemplo, la toxina diftérica está codificada en un fago presente en Corynebacterium diphteriae. 4.- ELEMENTOS GENÉTICOS DE LOS VIRUS Dentro de los virus se encuentran todas las posibilidades de utilización de los AN como vectores de la información genética: hay virus ADN, y virus de ARN. En ambos casos, el material genético puede estar presente como cadena simple o como cadena doble y en el caso de que el material genético sea de cadena simple, esta cadena puede ser la codificante (cadena +) o la no codificante (cadena -). En todos los casos, es necesario que la información genética llegue a la producción de muchas copias de la cadena de ARN mensajero (cadena +), y para esto es necesario que existan los sistemas enzimáticos correspondientes a las diferentes actividades. En ciertos casos, estas actividades enzimáticas son particulares de los ciclos biológicos víricos y, por consiguiente, dianas para la acción de agentes quimioterápicos antivirales. Los virus pueden producir en las células eucarióticas que infectan diferentes tipos de efectos tales como la transformación oncogénica, infección lítica, infección persistente en la que la célula infectada libera de forma permanente nuevos viriones sin morir, e infección latente equivalente al ciclo lisogénico de los bacteriófagos. 5.- EXPRESIÓN DE LA INFORMACIÓN GENÉTICA CICLO DEL MATERIAL GENÉTICO El material genético de cualquier organismo (procarionte o eucarionte) está sometido a una serie de procesos cíclicos que aseguran la realización de sus dos funciones esenciales: la transmisión entre generaciones de la información genética con fidelidad y la expresión de esa información genética con precisión para que pueda cumplir la misión para la que ha sido seleccionada. Por consiguiente, el ciclo del material genético comprende tres procesos: replicación para su transmisión en copias idénticas a la descendencia, trascripción para sintetizar el material que asegura la expresión de la información en la célula y traducción de la información genética de la forma de ácido nucleico a la forma de proteínas. Estos tres procesos ocurren independientemente de cuál sea la molécula transmisora principal de la información genética entre generaciones (ADN en 34 Secuenciación global de los genomas presentes en un ambiente determinado sin separar individualmente las distintas especies de organismos que se secuencian. 86 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) la mayoría de los casos y ARN en el caso de los virus de ARN) y son similares en procariontes y en eucariontes. 5.1.- Replicación La replicación del material genético es un proceso complejo en el que participa un gran número de proteínas que reconocen la molécula de material genético y llevan a cabo la polimerización de otras moléculas complementarias cada una de sus hebras de forma que, al final, se logra disponer de dos moléculas idénticas (salvo errores denominados mutaciones) a la inicial. El proceso presenta ciertas diferencias según el material genético que se transmite entre generaciones sea ADN o A RN y, en el primer caso, si el organismo es eucarionte o procarionte. 5.1.1.- Replicación del ADN procariótico En este proceso interviene un grupo de enzimas conocido genéricamente como ADN polimerasa. Este complejo multienzimático va incorporando nucleótidos a la cadena que se va sintetizando utilizando como molde una de las hebras de la cadena antigua. Esta polimerización se produce añadiendo nuevos nucleótidos al extremo 3' de una hebra de ADN en crecimiento. Por ello se dice que la síntesis del ADN se produce de forma que la cadena va creciendo hacia el extremo 3’ (dirección 5'→3') mientras la ADN polimerasa va leyendo la hebra molde en dirección 3'→5'. Por otra parte, para que se produzca la síntesis de ADN por la ADN polimerasa, además de la enzima que realiza la polimerización en sí, es necesario otro juego de proteínas que van desenrollando la doble hélice de ADN delante de la ADN polimerasa para permitir que ésta funcione. Estas enzimas se denominan genéticamente topoisomerasas y girasas. La acción de las topoisomerasas y girasas permite abrir la doble hélice de ADN para que entre el complejo de la ADN polimerasa y realice la polimerización. Sin embargo, la ADN polimerasa no es capaz de iniciar la polimerización del ADN sino que solamente es capaz de continuarla. Para que la ADN polimerasa funcione es necesario que exista un extremo 3' libre al que añadir un nuevo nucleótido. ¿Quién pone ese extremo 3' libre?. En todos los casos (salvo algunas excepciones de ciertos virus) ése extremo 3' viene de una pequeña cadena de ARN que se ha sintetizado como paso inicial de la replicación del ADN. Por consiguiente, la replicación del ADN comienza con la síntesis de una pequeña molécula de ARN que luego continúa como molécula de ADN. Esta pequeña molécula de ARN se denomina cebador o primer, y es sintetizada por otro complejo multienzimático denominado ARN polimerasa. La síntesis de ADN (replicación) va procediendo detrás de la horquilla que se forma en el punto en que se van abriendo las dos hebras de ADN molde por acción del complejo de girasas y topoisomerasas. De esta forma, la hebra que se va sintetizando es continua. Sin embargo, la hebra que se sintetiza usando como molde la complementaria de la anterior no puede ser continua porque la acción de las topoisomerasas y girasas se produce detrás del punto de origen de la transcripción. Por esto, una de las dos hebras hijas será continua y la otra discontinua. Los fragmentos de ésta última se unirán entre sí mediante la acción de otra enzima denominada ADN ligasa que une los fragmentos anteriores. 87 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) En las bacterias, cuyo material genético es cerrado, no tiene extremos, sólo existe un origen de replicación del ADN en cada molécula de ADN. Esto es válido para los plásmidos y para los cromosomas bacterianos. Asimismo, esto se cumple también en el ADN de los orgánulos celulares procarióticos presentes en las células eucarióticas. 5.1.2.- Replicación del ADN eucariótico. La replicación del ADN eucariótico es esencialmente igual a la del procariótico aunque presenta ciertas diferencias los cromosomas eucarióticos son abiertos, en lugar de cerrados, y el número de orígenes de transcripción es múltiple en cada cromosoma en lugar de tener uno solo por cromosoma. 5.2.- Trascripción Es el proceso por el que se transmite la in formación contenida en el ADN al ARN. Este proceso se lleva a cabo por la ARN polimerasa que utiliza como molde una de las dos hebras del ADN, la denominada hebra codificante. Durante el proceso de transcripción se reconoce un sitio específico de la molécula de ADN en el que se van a unir las enzimas del complejo de ARN polimerasa. Este sitio específico se denomina promotor del gen y permite que se produzca la transcripción. Genes sin promotores no pueden ser transcritos aunque un promotor puede servir en procariontes para transcribir varios genes seguidos que forman lo que se denomina un operón. En células procarióticas existe una serie de proteínas que pueden un irse a la ARN polimerasa controlando a qué promotores se puede unir ésta y regulando así la expresión génica. Estas proteínas se denominan factores sigma. En células eucarióticas la unión de la ARN polimerasa a un promotor o a otro viene regulada por la unión de otras muchas proteínas que, de alguna forma, dirigen la unión de la polimerasa al promotor conveniente. No todas las moléculas de ARN que se sintetizan en una célula van a ser traducidas en proteínas. La gran mayoría de las moléculas de ARN tienen otras funciones estructurales o enzimáticas diferentes de la transmisión de la información genética. Son las moléculas de ARN transferente (que sirve para activar los aminoácidos de manera que puedan ser polimerizados en proteínas) y las de ARN ribosomal que sirven para que estos orgánulos celulares puedan desarrollar su función. Constantemente se van encontrando nuevas moléculas de ARN con función diferente a la de transmisión de la información genética. En cualquier caso, éstas moléculas especiales de ARN son sintetizadas por un procedimiento de transcripción similar al descrito aunque el complejo enzimático de la ARN polimerasa pueda ser algo diferente. 5.2.1.- Trascripción en los virus Dentro de los virus se pueden encontrar especies con material genético basado en ADN y otras en ARN, tanto de cadena simple como doble. La transcripción del material genético vírico depende de cuál su naturaleza. 88 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) • Material genético es ADN de doble cadena (dsDNA, virus de clase I como son los herpesvirus) la transcripción ocurre como se ha descrito en eucariontes. Este es el caso de • Si es ADN de cadena simple (ssDNA, virus de clase II) se sintetiza un intermediario de dsDNA y este se transcribe como se ha descrito para eucariontes. • Si el material genético es ARN bicatenario (dsRNA, virus de clase III como son los reovirus, grupo en el que se encuentran los rotavirus) se produce la trascripción usando una RNA polimerasa vírica específica35. • Si es ARN de cadena simple (ssRNA) y esta cadena puede ser leída directamente (cadena codificante) por el ribosoma (ssRNA+, virus de clase IV como el poliovirus)) el propio virus puede servir inicialmente como ARN mensajero. • Si se trata de un ssRNA cuya cadena no es la codificante sino su complementaria (ssRNA-, virus de clase V como el virus de la gripe) se produce una transcripción mediante RNA polimerasa lectora de ARN (caso similar al de los virus del grupo III). • Hay virus con genoma de ssRNA+ que se replican usando un intermediario de ADN (virus de clase VI como los retrovirus entre los que se encuentra el HIV). La síntesis del ADN en este caso se realiza por una transcriptasa reversa que lee ARN para sintetizar ADN. Este ADN es posteriormente transcrito normalmente. • Por último, hay virus de dsDNA, similares a los del grupo I, en los que el ADN vírico se replica mediante un intermediario de ARN [virus de clase VII como el virus de la hepatitis B (VHB)]. 5.3.- Traducción Es el proceso por el que la información genética con tenida en el ADN y transcrita en una ARN mensajero va a ser útilizada para sintetizar una proteína. El proceso de lleva a cabo en los ribosomas. Para que un ribosoma pueda reconocer una ARN mensajero y utilizarlo para sintetizar una proteína, el ARN tiene que contener, además de la serie de nucleótidos que llevan la secuencia de la proteína, otras secuencias que permitan al ribosoma unirse al ARN, saber dónde ha de iniciarse la traducción de la secuencia (el denominado codón de iniciación), saber dónde debe terminar la traducción (codón de terminación) y saber en qué punto de la molécula de ARN mensajero deben separarse éste y el ribosoma. Los ribosomas de las células procarióticas son diferentes de los de las células eucarióticas como puede comprobarse por su diferente susceptibilidad a antibióticos ribosomales. Por otra parte, la traducción en células procarióticas ocurre simultáneamente a la transcripción del gen, mientras que en las células eucarióticas, la traducción se produce sólo una vez que el ARN mensajero ha llegado al citoplasma y a la zona donde se encuentran los ribosomas, por lo que nunca es simultánea con la transcripción. La traducción del mensaje genético desde el ARNm hasta una proteína no es suficiente, en algunos casos, para que se exprese correctamente. Es necesario que el polipéptido que se va sintetizando por el ribosoma se pliegue de forma adecuada para que 35 Las RNA polimerasas sintetizan ARN, la peculiaridad de las descritas aquí es que leen ARN para sintetizar nuevo ARN, mientras que las ARN polimerasas eucarióticas normales leen ADN para sintetizar ARN. 89 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) pueda desarrollar su función biológica correctamente; sin embargo, en muchos casos esto no ocurre de una manera completamente espontánea: es necesario el concurso de un grupo de proteínas esenciales denominadas chaperoninas (anglicismo que viene a significar proteínas tutoras o, si se quiere, tutorinas) que ayudan al péptido naciente a adquirir la configuración tridimensional correcta. Estas proteínas son muy importantes, por otra parte, para corregir errores pequeños que se produzcan en proteínas maduras y que causan su inactivación o bajada de rendimiento. 5.3.1.- Modificaciones del ARN mensajero Tiene que producirse una serie de modificaciones en el ARN mensajero para que pueda ser traducido correctamente. En las células eucarióticas las moléculas de ARN deben ser trasladadas del núcleo, donde ocurre la transcripción, al citoplasma, donde ocurre la traducción. Por otra parte, los genes de organismos eucarióticos suelen tener fragmentos que no se traducen pero sí se transcriben, son los denominados intrones por contraposición a los exones o secuencias del ARN que van a ser traducidas. Cuando se transcribe un ARN eucariótico éste es una secuencia de intrones y exones que debe ser procesada para eliminar los intrones y dejar sólo los exones colocados ordenadamente. Esta eliminación selectiva de intrones se denomina técnicamente procesamiento o splicing, y ocurre en el núcleo. En los genes procarióticos no existen (salvo alguna excepción muy rara en un grupo especial de arqueobacterias) intrones y, por lo tanto, no hay procesamiento. Además de la eliminación de los intrones, los ARN mensajeros eucarióticos deben ser modificados en sus extremos 5’ y 3’ mediante la adición de las estructuras conocidas como CAP y cola de poliadenina, respectivamente. 5.4.- Aplicaciones biotecnológicas Los procesos y sistemas enzimáticos involucrados en la transmisión y expresión de la información genética entre generaciones pueden ser aislados y utilizados en la Biotecnología para lograr la expresión y manipulación de información genética por microorganismos e, incluso, por organismos superiores. Como ejemplo citaremos dos aplicaciones derivadas de las propiedades y enzimas involucradas en la replicación del ADN. 5.4.1.- Detección se secuencias de ADN o ARN mediante hibridación de ácidos nucleicos. Estas técnicas están basadas en la especificidad de la unión de hebras de ácidos nucleicos con secuencias complementarias. Dependiendo de la perfección del acoplamiento de las dos secuencias (del grado de complementariedad) la unión entre las dos cadenas será más o menos fuerte. Esto significa que podrá resistir temperaturas más elevadas cuando la complementariedad es más alta o se separarán las hebras (se producirá la fusión) cuando la complementariedad es demasiado baja. En cualquier caso, las hebras se mantendrán unidas o no dependiendo del binomio grado de complementariedad-temperatura. La especificidad de la unión de hebras nos permite detectar la presencia de secuencias en una molécula, o en una mezcla de moléculas, de ácido nucleico (ADN o ARN) 90 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) usando como sonda una molécula con secuencia complementaria y convenientemente marcada para facilitar su detección. Estos procedimientos se utilizan en hibridación in situ, experimentos de Southern (detección de secuencias de ADN) o en experimentos de Northern (detección de secuencias de ARN). 5.4.2.- Amplificación de secuencias de ácidos nucleicos mediante la reacción en cadena de la polimerasa En esta técnica se utiliza la capacidad de la ADN polimerasa de sintetizar ADN a partir de una secuencia cebadora determinada. En ella se usan dos cebadores que flanqueen una secuencia de ADN que se quiere detectar o de la que se desea disponer de un gran número de copias; estos cebadores deben servir para iniciar la síntesis de ADN en cada una de las dos hebras del fragmento que se quiere amplificar y, por consiguiente, deben estar (orientados hacia el interior de la secuencia que se quiere amplificar. Mediante la adición de ADN polimerasa y de los cofactores y substratos necesarios es posible realizar la síntesis de la molécula de ADN flanqueada por los dos cebadores in vitro. Si, además, se realizan varios procesos de síntesis de ADN usando los mismos cebadores en procesos consecutivos el número de copias de l a molécula, de ADN que se quiere amplificar aumentará exponencialmente permitiendo disponer de grandes cantidades de la misma. En la práctica, esta reacción de polimerización se lleva a cabo con la ADN polimerasa de una bacteria termófila (Thermus aquaticus u otras similares) capaz de resistir múltiples ciclos a elevada temperatura (Taq-polimerasa). La técnica también se puede utilizar para detectar ARN siempre y cuando se realice un paso previo de transcripción reversa del mismo. 6. CONTROL DE LA EXPRESIÓN GÉNICA La transmisión de la información genética entre generaciones y la expresión en forma de proteínas de dicha información genética no son suficientes para permitir que el programa de desarrollo de un organismo se lleve a cabo correctamente. Es necesario, además, asegurar que los genes que constituyen la información genética transmitida se expresen en momentos adecuados cronológica o espacialmente (procesos de desarrollo) o como respuesta a cambios en las condiciones ambientales en las que se encuentra la célula o el individuo. Por otra parte, las respuestas al ambiente o los procesos de desarrollo no suelen producirse como con secuencia de la activación de un único gen en un momento, sitio o condición determinado, sino que suele ser necesaria la expresión coordinada de un conjunto de ellos para que tenga lugar el efecto. Estos dos hechos hacen que el control de la expresión génica sea central en el proceso de la biología molecular de los seres vivos. El modo de regulación de la expresión génica más frecuente en procariontes se conoce con el nombre de Modelo del Operón que explica, como veremos, la expresión coordinada de genes. Por otra parte, veremos a continuación cuál es el proceso por el que llega al material genético la información ambiental o de desarrollo que permita la expresión coordinada; estudiaremos el proceso de transducción de la señal. MODELO DEL OPERÓN 91 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) La expresión de un grupo de secuencias codificantes de diferentes péptidos cuya presencia debe ser coordinada se logra colocando todas las secuencias bajo el control de un mismo promotor. La transcripción de todas estas secuencias darán lugar a un ARN mensajero de gran tamaño que codifica todos los genes contenidos en el operón (ARN policistrónico) de forma que siempre se podrán traducir coordinadamente las diferentes proteínas del operón. La cantidad de cada una de las proteínas del operón que se produce como consecuencia de la traducción no siempre es equimolecular porque existen elementos de regulación de la traducción que provocan la separación del ribosoma y el ARNm en ciertos momentos controlando así más finamente la coordinación de la expresión. ¿Cómo se regula de la expresión génica?. Entre el promotor del operón y el inicio del primer gen estructural del fragmento policistrónico existe una secuencia de ADN denominada Operador. A esta secuencia se puede unir una proteína denominada Regulador de forma que cuando se encuentra unido el regulador a la secuencia del operador impide el paso de la ARN polimerasa en su trayecto de transcripción del ADN para sintetizar el ARN policistrónico. Por consiguiente, la regulación de la expresión se logra mediante un impedimento físico de la actividad de la ARN polimerasa causado por la unión de una proteína al ADN. Existen dos clases de operones según sea su respuesta a las condiciones ambientales: los operones inducibles y los operones represibles. 6.1.- Operón inducible Es aquél en el que como consecuencia de la presencia de una molécula inductora se produce la expresión de los genes del operón. El ejemplo más clásico es el del Operón de la lactosa que regula la síntesis de las proteínas que permiten el metabolismo de la lactosa por las bacterias. Cuando una bacteria se encuentra en un ambiente donde puede disponer de lactosa los genes que hacen posible la utilización de este azúcar por la bacteria se inducen. Estos genes, agrupados en el Operón de la lactosa, están constitutivamente reprimidos porque a la región del operador del operón se ha unido una proteína inhibidora que impide el paso de la ARN polimerasa. Cuando hay lactosa en el medio de crecimiento alguna molécula de este azúcar puede llegar al interior de la célula usando alguno de los sistemas de transporte presentes en la membrana bacteriana para azúcares. Una vez en el interior de la célula, la lactosa puede unirse a la proteína inhibidora unida a la región reguladora del operón de la lactosa de suerte que, cuando la lactosa está unida, la proteína inhibidora se separa de la región operadora permitiendo el paso de la ARN polimerasa y la transcripción de los genes correspondientes. Cuando desaparece la lactosa del medio la proteína inhibidora queda de nuevo libre de forma que puede volver a unirse al fragmento operador del operón impidiendo, de nuevo, la transcripción de los gen es que lo forman. El resultado de este proceso es que, como respuesta a la presencia de lactosa en el medio, se induce la expresión de los genes que permiten su catabolismo. Este sistema es extremadamente sensible porque la acción de las enzimas que permiten el paso de la 92 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) lactosa a través de la membrana (transportadoras de lactosa) permite que la concentración intracelular de lactosa sea suficientemente alta hasta el momento en el que la concentración extracelular es ya tan baja que no puede utilizarse el azúcar eficientemente. 6.2.- Operón represible En otros casos es necesario que la expresión de ciertos genes se detenga tan pronto como sea metabólicamente posible, porque su actividad no sea necesaria, para lograr una mayor economía de la energía celular. Es el caso, por ejemplo, de los operones de la arginina y del triptófano. Estudiaremos con más detalle el primero de ellos. Si la cantidad de arginina presente en la proteínas que se encuentran en el medio en que crece una bacteria es suficientemente alta como para satisfacer las necesidades bacterianas de este aminoácido, no es necesario que la bacteria lo sintetice sino que, simplemente, lo asimila del medio de crecimeitno. Ahora bien, si la cantidad de arginina exógena no fuera suficiente, las bacterias pueden sintetizarla a partir de otros precursores mediante el concurso de una serie de enzimas codificadas en el operón de la arginina. De nuevo, nos encontramos con un operón y a su secuencia operadora se une una proteína represora específica. Sin embargo, en este caso la proteína represora se mantiene unida siempre que a ella se encuentre unida, a su vez, una molécula de arginina o de un análogo de arginina. Cuando la concentración celular de arginina desciende por debajo de unos niveles críticos (que vienen determinados por la constante de afinidad de la molécula represora por la arginina o su análogo) el represor se separa de la arginina y queda inactivo separándose, también, de la secuencia operadora y permitiendo la transcripción de los genes que codifican las proteínas de síntesis endógena de arginina. El efecto final es la expresión de los genes del operón como respuesta a los niveles de arginina presentes en la célula. MECANISMOS DE TRANSDUCCIÓN DE SEÑAL Se conocen con este nombre los mecanismos que permiten a las células sentir condiciones exteriores (ambientales o señales de desarrollo) y activar como consecuencia de ello la expresión de algunos genes u operones. Los dos ejemplos de operón descritos anteriormente responden a las condiciones ambientales de concentración de lactosa y arginina, respectivamente; sin embargo, no se consideran incluidos dentro de los mecanismos de transducción de señal porque la detección del estado ambiental no se realiza a través de la membrana bacteriana sino que se produce directamente por interacción del inductor con el represor modulando su unión al operador. En el caso de los sistemas de transducción de seña, un complejo de proteínas de membrana recibe la señal externa. Para ello, este complejo ha de tener una proteína, al menos, dirigida hacia el exterior de la célula de manera que actúe como receptor de la señal. Como con secuencia de la detección de la señal (lo que, en términos bioquímicos suele significar "cuando se ha unido el receptor de membrana la molécula que actúa como inductor") se produce un cambio conformacional en la molécula del receptor; cambio que, a su vez, causa otros en las otras proteínas del complejo de membrana con las que interacciona. Como consecuencia de estos cambios estructurales el complejo de 93 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) proteínas, por su cara interna, es capaz de modificar químicamente proteínas citoplásmicas activando o inactivando así su función como represores de operones o genes (esto suele ocurrir mediante procesos de fosforilación) o bien sintetiza moléculas que son inductores que regulan la actividad de represores presentes en la célula (estas moléculas inductoras son lo que se suele denominar colectivamente "segundos mensajeros"). Los sistemas de transducción de señal son muy importantes porque de ellos depende gran parte del control de la expresión génica y porque son manipulables genéticamente de forma que podemos utilizarlos para provocar la expresión génica usando señales inductoras diferentes de las que normalmente las controlan en la naturaleza pero que son poco manejables en procesos industriales biotecnológicos. REQUERIMIENTOS ESTRUCTURALES PARA LA EXPORTACIÓN DE PROTEÍNAS Si se desea utilizar el potencial de los microorganismos como productores de proteínas en la industria es necesario, además de proceder al clonaje de los genes codificantes de las proteínas de interés y de estudiar y manipular el control de la expresión de los gen es correspondientes, conocer cómo se puede lograr que la proteína se localice en una fracción del cultivo donde sea más fácil proceder a su purificación. En la mayoría de los casos es interesante que la proteína se exporte al exterior de la célula y para ello hay que conocer el mecanismo por el que esto ocurre. Las proteínas sintetizadas en las células están destinadas, como norma, a quedarse en el citoplasma celular. Para que una proteína se integre en la membrana celular o para que la atraviese y sea exportada al exterior, es necesario que la proteína contenga unas señales que dirijan este proceso. Estas señales son componentes de la secuencia de aminoácidos de la propia proteína que se localizan en el extremo amino-terminal (el primero que se sintetiza por el ribosoma) del péptido naciente. Existen varios tipos de señales: unas son las señales de exportación que sirven para que el péptido que va sintetizándose sea exportado al exterior de la membrana citoplásmica. Estas señales son secuencias que se insertan y atraviesan la membrana plasmática y, una vez que se ha conseguido el paso del péptido naciente al exterior celular, son eliminadas mediante una enzima denominada peptidasa señal (signal peptidase, en inglés). Si no funciona la peptidasa señal, la proteína queda unida por su extremo aminoterminal a la membrana citoplásmica. Otro tipo de señales sirven para que la proteína se ancle en la membrana celular: son secuencias que están diseñadas para quedarse atrapadas en la bicapa lipídica de la membrana celular de manera que las proteínas que las contienen pasan a ser proteínas integrales de membrana y no pueden purificarse con métodos convencionales. Por último, en los organismos eucarióticos se puede producir otro procesamiento adicional de las proteínas: la glicosilación, consistente en la adición ordenada de cadenas polisacarídicas a los péptidos nacientes. La presencia de estas modificaciones polisacarídicas es esencial para la función de muchas proteínas eucarióticas. La adición de las cadenas de azúcar se produce en el Aparato de Golgi y tiene lugar en proteínas que contienen una secuencia dos tipos de señales: una que las dirige hacia dicho orgánulo celular (especializado en modificación de las proteínas y su posterior secreción al medio 94 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) extracelular) y una segunda señal que indica en qué posiciones de la secuencia de la proteína va a producirse la unión del polisacárido. 7.- CONCLUSIÓN Desde el punto de vista biológico, la expresión de la información genética de un organismo y su transmisión entre generaciones es un proceso complejo en el que interviene un gran número de proteínas y de mecanismos. Estos mecanismos aseguran la fidelidad de la copia de la información desde el ADN al ARN para asegurar la expresión, y del ADN a una nueva molécula de ADN para asegurar la transmisión. Sin embargo, además de la traducción del ARN por los ribosomas, para que se produzca una correcta expresión del mensaje genético ha de estar controlado el momento del dicha expresión, ha de asegurarse que el mensaje (proteína) adquiera la conformación correcta para su funcionamiento y se coloque en el lugar celular correspondiente. Desde el punto de vista biotecnológico, todos estos procesos son manipulables para poder conseguir la expresión artificial de proteínas de interés industrial en procesos llevados a cabo por microorganismos. 95 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Tema 5.- Interacción con los microorganismos. Concepto de flora normal. Localización de la flora normal. Interacción patogénica entre huésped y bacteria. postulados de Koch. Poder patogénico y virulencia. Factores determinantes de la acción patógena 1.- CONCEPTO DE FLORA NORMAL Un cuerpo humano está formado por alrededor de 1014 células36, de las que sólo aproximadamente el 10% son humanas, el resto son microorganismos asociados. En su gran mayoría estos microorganismos no son patógenos. La flora normal puede llegar a ser localmente muy abundante como en el intestino grueso donde se alcanzan niveles de 1011 microorganismos por gramo. Se denomina microbiota normal, flora normal o flora nativa al conjunto de microorganismos que viven de forma habitual en un cuerpo sano. Los lugares donde se encuentran pueden ser muy variados y en ellos desarrollan tareas beneficiosas para el ecosistema general del cuerpo. Estas tareas incluyen la participación en los procesos de digestión de alimentos y de síntesis de vitaminas en el intestino, la producción del pH ácido de la vagina o la protección competitiva frente a patógenos. Por consiguiente, en la mayoría de los casos, la interacción entre la flora normal y el ser humano es beneficiosa; pero pueden producirse circunstancias en que esto cambie y la flora normal se torne patógena oportunista. Los tratamientos con antibióticos de amplio espectro o la acción antiséptica de algunos productos de limpieza (jabones, por ejemplo) pueden alterar la flora normal lo que, en ocasiones, deja la puerta abierta para el desarrollo de procesos infecciosos oportunistas que pueden llegar a ser graves (por ejemplo, la colitis post-antibiótico producida por C. difficile). La flora normal puede inducir una respuesta inmune en los tejidos lo que facilita el control de los microorganismos. Es más, en algunos casos la flora normal es esencial para el desarrollo de los tejidos37. Por lo tanto, no debe verse a las bacterias de la flora normal como algo extraño que los tejidos humanos aprenden a soportar sino, más bien, parece ser que los tejidos pueden promover en ciertas ocasiones asociaciones con bacterias que resultan favorables. En este sentido, los receptores de nuestras células pueden participar en el mantenimiento de estas relaciones saludables con nuestra flora habitual y en el establecimiento de nuevas relaciones38. La flora normal está formada por microorganismos que se han adaptado a la permanencia en organismos superiores. Se puede pensar que la interacción patógena puede ser la excepción y no la norma en las relaciones entre microorganismos y organismos superiores. Un ejemplo de coevolución de microorganismos y huésped es el del virus de la 36 Hay una cierta controversia sobre este número y sobre si en realidad las células humanas suponen un 10% o más; pero, en cualquier caso, suponen sólo una parte del total de las células del organismo considerado como una comunidad. 37 Por ejemplo: algunos elementos del sistema inmune, incluyendo las placas de Peyer, la lámina media y el espacio intraepitelial no se en ratones gnotobióticos (libres de gérmenes) a no ser que se permita su colonización con microorganismos. 38 Ver ASM News 69: 186. 96 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) mixomatosis y los conejos en Australia: este virus se introdujo para controlar las poblaciones de conejos asilvestrados y produjo una alta mortalidad entre estos; sin embargo, en pocos años apareció espontáneamente una variante atenuada del virus que no causaba la muerte del conejo y que con unos pocos años más se hizo predominante en la población. Ejemplos similares podrían explicar la presencia de muchos reovirus en el intestino humano.39 2.- LOCALIZACIÓN DE LA FLORA NORMAL MICROORGANISMOS NATURALES DE LA PIEL Aunque la piel está expuesta al contacto con un gran número de microorganismos, la mayoría de ellos no puede crecer sobre ella debido a la sequedad y a la baja actividad de agua (aw) de las secreciones de sudor. Los microorganismos de la piel son generalmente Gram-positivos (Staphylococcus, Streptococcus, Corynebacterium, Bacillus) que son, además, más resistentes a agentes antisépticos. Se encuentran en menor proporción Gram-negativos (Pseudomonas) y bacterias entéricas. También pueden encontrarse algunas levaduras y hongos productores de tiña. FLORA NORMAL DE LAS MANOS Dado que la mayoría de las infecciones nosocomiales son transmisibles por las manos, el lavado de las manos es una práctica de gran importancia en el control de estas infecciones y es una de las prácticas más descuidadas por el personal sanitario. La flora nativa de las manos está compuesta por microorganismos transeúntes y microorganismos residentes. Al primer grupo pertenece la mayoría de los microorganismos patógenos. Se trata de microorganismos que se quedan en nuestra piel durante poco tiempo y son fácilmente eliminados por lavado. Llegan a nosotros a través del contacto con material o instrumental contaminado. Los microorganismos residentes son habitantes habituales de la piel y, en general, son de baja virulencia por lo que no suelen ser peligrosos. Es más difícil eliminarlos por lavado. Incluyen diferentes tipos de estafilococos, corinebacterias y coliformes. En ciertas ocasiones es necesario eliminar ambos tipos de flora normal (operaciones quirúrgicas, tratamiento de pacientes con inmunodepresiones severas). Se ha comprobado que un número significativo de pacientes con catéteres arteriales (1%) o venosos (7%) llegan a tener bacteremias causadas por microorganismos de la piel que acceden al interior del paciente a través de la herida de inserción del catéter. Para la limpieza de las manos del personal sanitario es necesario usar jabones antisépticos y jabones de arrastre. Debido a la alta frecuencia de uso de estos jabones es necesario prestar atención a su calidad cosmética. 39 Nature 433: 360. 97 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) MICROORGANISMOS DE LA CAVIDAD ORAL En la boca se desarrollan muchos microorganismos (se llega a niveles de 1010 por gramo en el sarro). Así mismo, se encuentra una gran diversidad de microorganismos colonizando los diferentes ambientes bucales. Hay estreptococos que son parte de la flora habitual tanto de los dientes como de la saliva, una gran cantidad de bacterias anaerobias estrictas y especies de los grupos de neiserias, estafilococos e, incluso, Vibrio. También se encuentran herpesvirus La presencia de elevados números de microorganismos anaerobios estrictos se debe a la alta tasa metabólica que tiene lugar en la boca, lo que genera los ambientes anaerobios necesarios para estos microorganismos. En la cavidad oral se producen biofilms. Entre estos son especialmente importantes los de Streptococcus mutans (estreptococo del grupo viridans) que es el agente productor de la caries dental40. El cepillado sólo produce una alteración temporal de las floras habituales de la boca. Algunas bacterias de la cavidad oral (S. salivarius, estreptococos α-hemolíticos, corinebacterias y Stomatococcus) pueden participar en la oxidación del alcohol a acetaldehído. Este último compuesto es cancerígeno. La actividad de estos microorganismos podría ser la causa del aumento de la incidencia de cáncer de esófago en los bebedores habituales y la relación entre la higiene dental deficiente y la aparición de cánceres de esófago y boca41. También se ha visto este efecto de producción de acetaldehído en la flora intestinal (principalmente en la anaerobia facultativa) de forma que antibióticos que eliminan la flora anaerobia facultativa y la anaerobia estricta (ciprofloxacina) reducen la acumulación de aceltaldehido en el colon mientrans que antibióticos que estimulan este tipo de flora (metronidazol) favorecen su acumulación42. MICROORGANISMOS DEL TRACTO GASTROINTESTINAL Muchos microorganismos bucales son arrastrados hacia el interior del tracto gastrointestinal. El bajo pH del estómago elimina la mayoría de ellos; pero el intestino, con la temperatura y el aporte de nutrientes constante es una ambiente muy favorable para el desarrollo de altas poblaciones microbianas: por encima de 109 bacterias por gramo pertenecientes a más de 500 especies diferentes. La mayoría de los microorganismos se encuentran en el intestino grueso. Las poblaciones de microorganismos intestinales cambian con la alimentación y son diferentes en los lactantes (bacterias lácticas) y los adultos (bacterias lácticas, anaerobias estrictas y enterobacterias). Aproximadamente el 99% de la flora intestinal de un adulto lo forman bacterias anaerobias estrictas (Bacteroides, Fusobacterium, Clostri- 40 La presencia de otros microorganismos competidores como S. gordonii dificulta la formación del biofilm de S. mutans reduciendo su acción como agente productor de caries (aem 71: 354-362) 41 La pobre higiene bucal favorece el desarrollo de Candida que también es productora de acetaldehido 42 ASM News 69: 164 98 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) dium) y bacterias lácticas (Streptococcus, Lactobacillus); las bacterias anaerobias facultativas (enterobacterias) sólo representan el 1% (Escherichia, Proteus, Klebsiella). Hay una presencia importante de enterovirus, rotavirus, adenovirus y herpesvirus localizados en el intestino grueso. Las poblaciones gastrointestinales cambian notablemente con la dieta y con los tratamientos con antibióticos. Los intestinos de los niños contienen inicialmente grandes números de microorganismos anaerobios facultativos que incluyen Escherichia coli y estreptococos. Al terminar la lactancia estas esñpecies disminuyen rápidamente y las especies anaerobias estrictas como Bacterioides y Clostridium son finalmente las más abundantes43. La flora intestinal es esencial para el desarrollo correcto del sistema inmune intestinal. Se ha visto que ratones que no tienen flora intestinal requieren un 30% más de calorías para mantener su peso correcto en comparación con ratones normales. La necesidad de controlar una flora intestinal tan compleja ha llevado al desarrollo de un sistema inmune intestinal que se va poniendo a punto como consecuencia del contacto con esta flora20. Conceptos de alimentos prebióticos y probióticos. Se denominan probióticos aquellos alimentos que contienen bacterias cuya presencia en el intestino es beneficiosa porque favorecen la digestión de alimentos y eliminan competidores. La ingesta de ciertas bacterias como Lactobacillus y Bifidobacterium tiene efectos particularmente favorables para la salud. Se denominan alimentos prebióticos aquellos que estimulan el desarrollo de las poblaciones bacterianas intestinales beneficiosas. Normalmente estos alimentos contienen azúcares complejos que no son digeridos en la parte superior del intestino y llegan a la región del colon donde alimentan estos tipos de bacterias. No está muy claro todavía qué hace de un bacteria un buen probiótico y de un alimento un buen prebiótico. Este es un campo de investigación en desarrollo actualmente. MICROORGANISMOS DE VIAS RESPIRATORIAS Es una microbiota muy diferente de la del tracto gastrointestinal. Los microorganismos habituales del tracto respiratorio superior son Streptococcus, Neisseria, Staphylococcus, Haemophilus, Bacteroides y Fusobacterium. En esta región se encuentran adenovirus y herpesvirus. El tracto respiratorio inferior, por el contrario, no tiene ninguna flora asociada normal porque los microorganismos que llegan allí son rápidamente eliminados por los sistemas de fagocitos del huésped. MICROORGANISMOS DEL OIDO EXTERNO 43 AEM 71: 77-83 99 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Presenta un flora similar a la de la piel en la que predominan los cocos Grampositivos, bacilos Gram-positivos y una pequeña proporción de bacilos Gram-negativos (enterobacterias y pseudomonas). MICROORGANISMOS DE LAS CONJUNTIVAS La mucosa conjuntiva se contamina en el momento del nacimiento y contiene una flora similar a la de la piel con presencia adicional de Neisseria, Haemophilus y algunos virus. MICROORGANISMOS DE VIAS GENITOURINARIAS La mayor parte del tracto genitourinario (desde los riñones a la vejiga) están libres de microorganismos y no hay flora normal asociada. Sin embargo, sí hay flora normal asociada en las regiones genitales externas masculinas y femeninas. Debido a su amplia superficie, a la cantidad de sus secreciones y a su pH ácido, la vagina es un nicho muy rico en flora normal formada por especies de Streptococcus, Lactobacillus, Bacteroides y Clostridium. También se encuentran hongos (levaduras) del género Candida e incluso protozoos del género Trichomonas. También se detectan pequeñas cantidades de herpesvirus. La población microbiana de la vagina está muy equilibrada de manera que si se producen alteraciones en ella ciertos microorganismos que la componen (Candida, Trichomonas) pueden comportarse como patógenos oportunistas y dar lugar a infecciones (vaginitis) o intoxicaciones (choque tóxico por estafilococos). La población normal vaginal cambia durante el embarazo. 3.- INTERACCIÓN PATOGÉNICA ENTRE HUÉSPED Y BACTERIA La interacción de los microorganismos con el hombre no siempre es beneficiosa o inocua sino que también puede ser una interacción patogénica. Esto puede ser debido bien a la actividad de microorganismos intrínsecamente patógenos o a la de microorganismos patógenos oportunistas bien de la microbiota normal o de otras poblaciones bacterianas que por heridas o por descenso de las defensas inmunitarias llegan a colonizar sitios no permitidos desarrollando allí su acción patógena. El término infección hay que distinguirlo, también, del de enfermedad ya que aquel se refiere únicamente al desarrollo de un microorganismo dentro de un huésped, mientras que este hace referencia también a la respuesta del huésped al crecimiento y factores de virulencia de un microorganismo. La infección puede adquirir varios grados: 100 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) • • • Colonización que es el grado mínimo de la infección. Las bacterias colonizan las mucosas y se multiplican allí sin que haya una respuesta clínica o inmune por parte del huésped. (Por ejemplo, la presencia de estafilococos potencialmente patógenos en la cavidad nasal.) Infección inaparente en la que el huésped no muestra una respuesta clínica específica, pero sí se observa una respuesta inmune. Es una infección asintomática o subclínica. Enfermedad infecciosa en la que se producen síntomas clínicos y respuesta inmune. La infección subclínica se suele producir cuando se produce el contagio por una número pequeño de microorganismos o estos son poco virulentos; la enfermedad se produce cuando los microorganismos son muy virulentos o su número es mayor de forma que interfieren con los mecanismos de resistencia del huésped. La enfermedad infecciosa y la subclínica siguen una evolución similar. Sin embargo, en ocasiones se pueden producir infecciones crónicas el las que el huésped se convierte en un portador asintomático del patógeno (por ejemplo, el caso de la fiebre tifoidea). 4.- POSTULADOS DE KOCH Las infecciones se caracterizan por su especificidad. Esto es, cada microorganismo patógeno produce una infección determinada causando un cuadro clínico característico. La especificidad de la infección se comprueba mediante los postulados de Koch que permiten identificar el agente etiológico de un proceso infeccioso. Los postulados son los siguientes: 1. El microorganismo debe encontrarse en todos los casos de la enfermedad 2. Debe aislarse y obtenerse como cultivo puro a partir de las lesiones 3. Debe reproducir la enfermedad cuando se inocula, a partir de un cultivo puro, en un animal de experimentación susceptible (modelo animal) 4. Debe aislarse el mismo microorganismo en cultivo puro a partir de las lesiones producidas en el animal. 5. El microorganismo debe inducir una respuesta inmune con la aparición de anticuerpos específicos en la sangre del hombre o animal infectado que puedan demostrarse por pruebas serológicas. No siempre pueden comprobarse todos los postulados, principalmente por razones técnicas (hay microorganismos no cultivables o hay una gran cantidad de microorganismos de la flora normal o no hay modelo animal adecuado). 5.- PODER PATOGÉNICO Y VIRULENCIA Es necesario distinguir el concepto de patogenicidad que se refiere a la capacidad o incapacidad de un microorganismo para producir una enfermedad del de virulencia que 101 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) indica el grado de patogenicidad de el grado de patogenicidad del parásito y normalmente se indica por la dosis o número de microorganismos que son necesarios para que se desencadene la enfermedad en un tiempo dado. La patogenicidad es una característica intrínseca del microorganismo. Por tanto, la patogenicidad nos permite comparar diferentes especies microbianas. La virulencia, por el contrario, es una característica de las diferentes cepas de una misma especie y permite realizar comparaciones entre ellas. La patogenicidad no depende únicamente del patógeno, sino también del huésped ya que microorganismos patógenos o muy patógenos para un tipo de huésped pueden ser poco patógenos o apatógenos para otro. Por ejemplo, la patogenicidad de Clostridium botulinum como habitante intestinal es diferente en niños lactantes y en adultos a causa de las diferencias en la microbiota intestinal. El virus de la influenza presenta diferente patogenicidad en personas de diferentes edades, etc. La virulencia tampoco es una característica totalmente estable dentro de una cepa, sino que esta puede variar como consecuencia de mutaciones, transferencia horizontal de genes, etc. En función de su patogenicidad, podemos diferenciar dos tipos de microorganismos: MICROORGANISMOS INTRÍNSECAMENTE PATÓGENOS Son los patógenos verdaderos o estrictos. Tienen la capacidad de colonizar de producir enfermedad en huéspedes normales sanos en los que superan las barreras de defensa normales. Son los patógenos clásicos: Staph. aureus, Streptococcus del grupo A, Neisseria gonorrhoeae, Salmonella, Shigella, Brucella, Corynebacterium diphteriae, Vibrio cholerae, etc. Estos patógenos se caracterizan porque: • Proceden de una fuente exógena y es necesario adquirirlos por contagio. • Su acción patógena es debida principalmente a factores dependientes del mismo propio microorganismo (factores de virulencia) • Producen un cuadro clínico más o menos específico de la enfermedad lo que facilita el diagnóstico. En muchos casos, los genes que codifican factores de virulencia están agrupados en lo que se denominan islas de patogenicidad. Así, por ejemplo, en Salmonella hay un grupo de genes (islas de patogenicidad) que no aparece en E. coli y que permiten a Salmonella invadir y sobrevivir dentro de las células del huésped. Este tipo de grupo de genes virulentos está presente en los patógenos y ausente en las bacterias benignas estrechamente emparentadas con ellos. PATÓGENOS OPORTUNISTAS O PATÓGENOS POTENCIALES 102 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Son capaces de colonizar el organismo sólo cuando fallan sus defensas y cambian sus condiciones ecológicas. Por tanto, las condiciones alteradas del huésped son el factor determinante de las enfermedades infecciosas producidas por patógenos oportunistas. A este grupo pertenecen muchos microorganismos de la flora normal o de la flora ambiental tales como Klebsiella, Enterobacter, Serratia, Proteus, Pseudomonas, Staph. epidermidis, estreptococos del grupo D, Bacteroides, herpesvirus, Pneumocystis carinii, Candida, etc. Estos patógenos se caracterizan porque: • En general proceden de una fuente endógena (microorganismos de la flora normal o de la flora externa que se han integrado como parte de la flora normal por un periodo de tiempo antes de desencadenar la enfermedad). • Su acción patógena es debida principalmente a las condiciones deficitarias del huésped. • Producen un cuadro clínico atípico que se añade al estado que presenta el enfermo lo que dificulta el diagnóstico. PATÓGENOS INTRACELULARES Y PATÓGENOS EXTRACELULARES En relación con su interacción con las células, las bacterias pueden agruparse en tres clases (los virus son todos patógenos intracelulares): • Las bacterias extracelulares se multiplican en los espacios intercelulares y son eliminados con facilidad por los fagocitos. Sólo producen enfermedad si pueden escapar de la acción de los fagocitos (por ejemplo, cápsulas) o si el huésped tiene el sistema inmune deficitario. Son ejemplos Staphylococcus, Neisseria, Clostridium, Cryptococcus • Las bacterias intracelulares facultativas se multiplican en el medio extracelular y si son fagocitadas desarrollan mecanismos para inhibir la digestión y pueden permanecer vivas largo tiempo en el interior del fagocito o invadir desde él otras células. Por ejemplo, Mycobacterium tuberculosis, Listeria, Brucella. • Bacterias intracelulares obligadas. Sólo pueden multiplicarse en el interior de las células. Por ejemplo: M. leprae y las clamidias. 6.- FACTORES DETERMINANTES DE LA ACCIÓN PATÓGENA • • • • Para que las bacterias puedan desarrollar su acción patógena es necesario que: Lleguen a la superficie del huésped por una puerta adecuada, colonicen el epitelio y resistan la acción de los sistemas fagocitarios Penetren el epitelio para llegar a los tejidos internos. Se multipliquen en los tejidos del huésped Produzcan alteraciones o lesiones en las células o tejidos del huésped. COLONIZACIÓN La vía de colonización de los microorganismos exógenos es generalmente la piel o, con mayor frecuencia, las mucosas de las vías gastrointestinal, genitourinaria o respira103 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) toria. En el caso de muchas infecciones oportunistas, la fuente de los microorganismos es la propia flora normal. En el caso de infecciones de origen exógeno, las bacterias deben resistir los sistemas de defensa de las mucosas y de la acción de ciertos tipos de anticuerpos. Muchos patógenos desarrollan sistemas de defensa o de competencia con las bacterias de la microbiota normal para asegurar esta fase de colonización. Otro factor de gran relevancia es la adhesión a la superficie de los epitelios que logra por medio de moléculas de la superficie celular (fimbrias, etc.) que actúan como vías de anclaje. La adhesión por medio de adhesinas y de fimbrias es un fenómeno específico en el que estas moléculas de la superficie bacteriana interaccionan de forma específica con componentes de la superficie de algunos tipos de células del huésped. Esta especificidad del tipo de célula al que una bacteria es capaz de adherirse es la base molecular del tropismo de ciertas células por diferentes tejidos u órganos. En el caso de la flora normal, la adherencia es también la responsable de su localización específica en el cuerpo y facilita su multiplicación por la formación de microcolonias y de biopelículas. Así mismo, en el caso de las infecciones patógenas, la presencia de adhesinas facilita el establecimiento del patógeno y la colonización del tejido. Esta adhesión puede producirse también a materiales inertes que actúan como soporte para la colonización (por ejemplo, prótesis de cadera o cardiacas). La adhesión puede prevenirse mediante inhibidores de la adherencia que son análogos de bajo peso molecular de las moléculas que reconocen los sistemas de adherencia y también con antibióticos que inhiben la producción de adhesinas. Las bacterias de la flora normal compiten con las patógenas por los sitios de adhesión en los nichos que colonizan. PENETRACIÓN Algunas bacterias son capaces de realizar su actividad patógena sin atravesar el epitelio (en general las bacterias toxigénicas como C. diphteriae, V. cholerae o Bordetella pertusis). Un segundo grupo penetra de una forma pasiva bien mediante la acción de vectores tales como mosquitos, pulgas, etc, o cuando se produce una alteración funcional o física del epitelio (bacterias que penetran por heridas, por ejemplo.) Por último, hay bacterias con sistemas activos de penetración mediante un sistema de endocitosis inducida (Salmonella, por ejemplo). 104 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Las bacterias que son capaces de penetrar pueden multiplicarse destruyendo el epitelio sin penetrar al tejido submucoso (por ejemplo, Shigella, E. coli enteroinvasivo), o pueden penetrar hasta tejidos más internos (Staph. aureus, S. typhi). MULTIPLICACIÓN Una vez que el patógeno ha penetrado el interior del tejido, debe multplicarse para alcanzar un número crítico que le permita iniciar la infección, invadir el organismo y desarrollar su acción patógena. Para ello deben obtener del huésped los nutrientes mientras evitan la acción de los sistemas de defensa. Para el establecimiento de la enfermedad infecciosa no solo es necesaria la multiplicación sino que también es importante la velocidad de crecimiento ya que este factor condiciona el tiempo requerido para que el patógeno colonice el huésped. INVASIÓN En el curso de la infección, las bacterias producen como consecuencia de su metabolismo diversas substancias que dificultan o impiden la acción de los sistemas de defensa del huésped. Entre estos factores hay que considerar las alteraciones en la pared celular que hacen a las bacterias resistentes a la lisozima y a otros factores del sistema de defensas humorales, la presencia de cápsulas que impiden la fagocitosis (neumococo), la liberación de toxinas antifagocíticas y la producción de enzimas como la coagulasa que permiten a la bacteria (Staph. aureus) formar una cubierta protectora. Por último, hay bacterias capaces de inhibir su digestión dentro de los fagocitos. Las bacterias utilizan diferentes vías para lograr la invasión del cuerpo. Las principales son las siguientes: • Contigüidad. La difusión por contigüidad es especialmente frecuente en los epitelios y mucosas. Esta vía de difusión está favorecida cuando los patógenos producen sistemas enzimáticos que destruyan el tejido subepitelial. Por esta vía pueden diseminarse infecciones en las vías respiratorias que pueden llegar hasta el oído medio (otitis) o a los senos frontales (sinusitis). • Vía linfática. Las bacterias alcanzan el sistema linfático y llegan hasta los ganglios donde, si son capaces de resistir el ataque de los elevados números de fagocitos allí presentes pueden colonizarlos, quedarse en los ganglios linfáticos (peste) o utilizar los fagocitos como sistemas de transporte hacia otros lugares del cuerpo. • Vía sanguínea El sistema circulatorio es generalmente de difícil acceso para los microorganismos. Sin embargo, pueden llegar a la sangre a través de heridas, picaduras o catéteres. Al sistema sanguíneo también puede llegarse a través del sistema linfático; pero es una vía más larga. La vía sanguínea es muy rápida para la difusión del patógeno y este puede alcanzar todo tipo de órganos donde los microorganismos pueden establecerse y desarrollar su acción patógena. La bacteremia es un paso ocasional o fugaz de bacterias a través de la sangre. Se puede producir como consecuencia de extracciones, de la masticación o del cepillado de dientes. Es producida normalmente por bacterias de la microbiota normal que son rápidamente eliminadas por el sistema de fagocitos. Sin embargo cuando existe una 105 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) disminución general de los sistemas de defensa inespecíficos, las bacteremias pueden permitir a las bacterias localizarse e iniciar procesos patógenos (endocarditis). La septicemia o sepsis es un paso masivo de bacterias a la sangre a partir de un foco séptico. Generalmente se produce fiebre y están asociadas con un cuadro clínico grave. Para que se produzca una septicemia es necesario un foco de infección constante de tipo tromboflebítico o por una puerta de entrada gastrointestinal. • Vía nerviosa. No es frecuente en el caso de las bacterias aunque sí en el de los virus. Es también una vía de difusión importante de ciertas toxinas como la tetánica. CAPACIDAD LESIONAL Las bacterias patógenas se caracterizan porque producen alteraciones celulares y tisulares responsables del cuadro patológico. Estas alteraciones pueden producirse por la formación de substancias tóxicas, por acción directa como consecuencia del proceso inflamatorio o por mecanismos inmunológicos Acción tóxica Las toxinas son substancias bacterianas que presentan acciones nocivas y estimulan una respuesta inmunológica (lo que las diferencia de los venenos minerales, alcaloides y glucósidos). 1) Las toxinas pueden ser exotoxinas proteicas sintetizadas en el citoplasma bacteriano durante la fase de crecimiento exponencial y que se liberan al exterior. Su acción tóxica es muy elevada y con unas características determinadas responsables de un efecto específico que puede presentar una a) acción general como en el caso de la toxina diftérica, las toxinas de Pseudomonas aeruginosa, la toxina de Bacillus anthracis b) neurotoxinas como la tetánica y la botulínica c) enterotoxinas como la colérica, las enterotoxinas de E. coli y otras enterobacterias, de Staphylococcus y de Clostridium perfringens y C. difficile. 2) Endotoxinas que forman parte del lipopolisacárido de Gram-negativas. Se caracterizan por tener una menor toxicidad que las exotoxinas y tener una acción inespecífica con producción de fiebre, alteraciones en la composición celular de la sangre, alteraciones vasculares y shock. Mecanismo inflamatorio Se produce como consecuencia de que las toxinas antifagocíticas son capaces de destruir los fagocitos que llegan al punto de infección liberándose, como consecuencia de su lisis, substancias que aumentan el daño celular y causan muchos de los síntomas celulares, tisulares y vasculares que caracterizan el cuadro patológico de la infección (pus). Mecanismo inmunológico La respuesta inmune produce una serie de reacciones de tipo humoral (vasodilatación, edema, infarto ganglionar) que pueden influir en el cuadro patológico. 106 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) De forma añadida a esto hay que considerar la posibilidad de reacciones de tipo anafiláctico en las que se liberan substancias vasoactivas responsables de fenémenos patológicos (shock, broncoespasmo e inflamación local), reacciones citotóxicas que producen la lisis de células del huésped, reacciones por complejos inmunes que dan lugar a acciones irritativas sobre vasos y tejidos y reacciones mediadas por células de tipo inflamatorio y de formación de granulomas. 107 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Tema 6.- Dispersión de los microorganismos. Reservorios y fuentes de infección. El contagio y su prevención. Vías y modos de transmisión microorganismos. La cadena de infección (cadena epidemiológica) está formada por tres eslabones que son necesarios para que se pueda producir la infección y trasmitirse a nuevos huéspedes. A estos eslabones se les denomina factores epidemiológicos primarios y son : 1. El reservorio y la fuente de infección 2. El mecanismo de transmisión 3. La población susceptible 1.- RESERVORIOS Y FUENTES DE INFECCIÓN Se denomina reservorio al hábitat natural de un agente infeccioso y fuente de infección al hábitat ocasional a partir del que el microorganismo patógeno pasa rápidamente al huésped. En algunos casos el reservorio y la fuente de infección son el mismo organismo, como es el caso del sarampión en el que el hombre es reservorio y fuente; mientras que en otros casos (especialmente en las zoonosis) ambos factores son distintos (por ejemplo: en la peste, el reservorio son las ratas y la fuente de infección las pulgas). Los reservorios y fuentes de infección pueden ser el hombre, animales y materiales inanimados. EL HOMBRE COMO RESERVORIO Y FUENTE DE INFECCION El hombre enfermo Los enfermos infecciosos liberan una gran cantidad de microorganismos durante un periodo llamado periodo de transmisibilidad (o periodo en que la enfermedad es contagiosa) que es característico para cada enfermedad. En muchos casos el periodo de transmisibilidad no coincide con el de la enfermedad con síntomas clínicos y de ahí la poca eficacia de muchas medidas de aislamiento. En relación con su gravedad, se pueden presentar casos mortales, graves, moderados y leves; y en cuanto a sus manifestaciones clínicas los casos pueden ser típicos (con sintomatología clásica), atípicos (con sintomatología no clásica y poco expresiva) e inaparentes (subclínicos con un curso típico de la enfermedad pero sin que se manifiesten los síntomas. Tienen gran importancia epidemiológica porque contribuyen a una mayor difusión de la enfermedad y a la inmunización espontánea). Las formas leves y atípicas suelen ser las más peligrosas desde el punto de vista de la transmisión de la enfermedad porque no llegan a ser reconocidos hasta que ya se ha producido el contagio. El hombre como portador Se llama portador a la persona infectada que no muestra síntomas clínicos y que, sin embargo, puede eliminar una gran cantidad de microorganismos patógenos. 108 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Se consideran portadores: • Los portadores precoces o en periodo de incubación. Estos son importantes en casos como los de • difteria • sarampión • poliomielitis • tos ferina • hepatitis • Portadores convalecientes liberan gérmenes durante el periodo de convalecencia de la enfermedad. Pueden ser • Portadores temporales que liberan gérmenes durante 1-2 meses • difteria • escarlatina • fiebre tifoidea • Portadores crónicos • fiebre tifoidea, especialmente en mujeres de 40-50 años. • VIH • Portadores sanos o por contacto que son personas sanas que sin haber padecido la enfermedad de forma aparente liberan microorganismos patógenos. • meningitis meningocócica • difteria • poliovirus La infección es más frecuente que la enfermedad por lo que se dan casos de individuos portadores y diseminadores del patógeno que no presentan síntomas y que contribuyen a la diseminación del agente infeccioso al moverse. El caso más característico podría ser el de las mujeres portadoras asintomáticas de S. tiphy (fiebre tifoidea). Según dónde se encuentren los microorganismos patógenos, los portadores pueden ser nasales, faríngeos, cutáneos, fecales, urinarios, etc. y su eficacia está en relación con la capacidad de difusión. LOS ANIMALES COMO RESERVORIOS Y FUENTES DE INFECCIÓN Los vertebrados pueden ser reservorios y actuar como fuentes de infección cuando padecen enfermedades infecciosas (zoonosis). Las enfermedades que pueden transmitir son numerosas: rabia, tuberculosis, brucelosis, peste, etc. La importancia epidemiológica de las zoonosis está en que pueden afectar a un gran número de animales domésticos y peridomésticos. En algunos casos son consideradas como enfermedades profesionales que causan elevadas pérdidas económicas (carbunco, brucelosis). Un aspecto adicional de las zoonosis es que pueden permitir la transmisión de información genética entre diferentes microorganismos (o virus) que se encuentren simultá109 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) neamente en un mismo huésped. Esto facilita la adquisición de nuevas características patógenas por agentes que antes no las tenían o las tenían en menor grado.44 MATERIALES INANIMADOS El suelo, agua y fomites pueden ser reservorios de gérmenes patógenos, principalmente cuando éstos pueden presentar formas especiales de resistencia (esporas de Clostridium tetani, Bacillus anthracis), cundo las condiciones ambientales son favorables para su desarrollo (por ejemplo, leptospiras que permanecen en zonas húmedas) o cuando una parte del ciclo biológico del patógeno se desarrolla en un medio externo. En este apartado hay que incluir los microorganismos patógenos oportunistas de vida libre (Pseudomonas y algunas enterobacterias, por ejemplo). 2.- EL CONTAGIO Y SU PREVENCIÓN Se denomina contagio a la transmisión de una enfermedad por contacto directo o indirecto. En este sentido, una enfermedad contagiosa es aquella que puede adquirirse por contacto con el enfermo que la sufre, secreciones, fomites (substancia u objeto no alimenticio capaz de vehiculizar una enfermedad transmisible), etc. Muchas enfermedades infecciosas han sido eliminadas o casi eliminadas por inmunización, medidas de la salud pública y mejoras en las condiciones de vida. Es más difícil controlar la dispersión de microorganismos patógenos a través del aire que a través de las otras vías de transmisión. Sin embargo, hay medidas que reducen el nivel de esta transmisión tales como cubrirse la nariz y la boca al toser o estornudar, usar mascarillas cuando se visita a pacientes especialmente susceptibles a microorganismos (niños recién nacidos, etc.). Así mismo, disminuir o evitar la exposición de pacientes inmunodeprimidos a microorganismos patógenos es una medida muy importante para evitar las complicaciones causadas por las enfermedades infecciosas. Deben usarse mascarillas especialmente al tratar a pacientes enfermos de tuberculosis u otras enfermedades pulmonares infecciosas. En estos casos y en los de otras enfermedades transmitidas a través del aire, suele ser recomendable el aislamiento del paciente, sobre todo si se encuentran en lugares concurridos (salas de pediatría y enfermedades de tipo vírico como paperas, sarampión, varicela). También es importante evitar el contagio y la transmisión por contacto. En este sentido es importante evitar mantener relaciones sexuales con individuos infectados. En cuanto a las manos como vehículos de contagio, es necesario prestar atención a las prácticas de lavado antes y después de tratar a pacientes infecciosos o susceptibles, cuando 44 Por ejemplo: supongamos dos virus que coinfecten un mismo animal de los que uno también es patógeno para humanos mientras que el otro no. Por combinación del material genético en el huésped animal, el virus no patógeno para los humanos puede adquirir factores que lo conviertan en patógeno para el hombre del material genético del virus coinfectante que sí es patógeno para las personas. 110 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) se vaya a manipular implantes o realizar acciones que rompan la continuidad de piel o mucosas y cuando se participa en actividades quirúrgicas. El personal sanitario está especialmente expuesto a situaciones de riesgo de contagio por cualquiera de las vías tratadas en este capítulo. Es necesario extremar las precauciones al manipular no solo a los pacientes sino también al manejar fomites, muestras, residuos y cadáveres. Las vías de contagio de patógenos son: aire, agua y alimentos contaminados, contacto directo y heridas y vectores transmisores de la enfermedad. 3.- VÍAS Y MODOS DE TRANSMISIÓN MICROORGANISMOS. Se denomina modo de transmisión el mecanismo por el que un microorganismo patógeno llega a infectar un nuevo huésped susceptible. Hay varias vías generales de transmisión: vía aérea, secreciones de vías respiratorias, agua, alimentos, contacto directo, etc. TRANSMISIÓN POR VÍA AÉREA La transmisión aérea es una vía de transmisión estresante para el microorganismo puesto que el aire carece de los nutrientes y la humedad necesarios para permitir una larga supervivencia de muchos patógenos. Muchas bacterias son transmitidas a través del aire en gotas (gotitas de Pflüge de 150 µm de diámetro y microgotas de Wells de 1 a 10 µm de diámetro) o aerosoles producidos al toser, estornudar o hablar. Son especialmente importantes los aerosoles producidos por tos o estornudo porque la gran velocidad con la que se emiten las partículas en estas condiciones (hasta 100 m/s) reducen mucho el tiempo de trayectoria de la partícula hasta llegar al nuevo huésped y de esta forma se hace mínima la desecación. En general, esta vía requiere una estrecha proximidad entre la fuente y el receptor para que se produzca el contagio. El polvo es un coadyuvante para la transmisión de microorganismos por vía aérea porque permite a los microorganismos resistir más tiempo en suspensión en el aire, y facilitan la entrada en el huésped. El polvo es importante en la transmisión de infecciones nosocomiales por esta vía. Enfermedades bacterianas transmitidas por vía aérea • Difteria (Corynebacterium diphteriae) • Faringitis causada por Streptococcus pyogenes • Neumonía causada por Streptococcus pneumoniae • Tuberculosis (Mycobacterium tuberculosis) • Legionelosis causada por Legionella pneumophila • Tosferina causada por Bordetella pertusis • Enfermedad meningocócica 111 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Los patógenos de la tuberculosis y de la difteria tienen una gran resistencia a las condiciones exteriores, tienden a vehiculizarse por el polvo y a alcanzar grandes distancias de transmisión. Enfermedades víricas transmitidas por vía aérea • Varicela (virus Varicela-Zoster) • Rubéola • Sarampión • Gripe • Resfriado común • Virus respiratorio sincitial TRANSMISIÓN POR VEHÍCULO COMÚN Bajo este epígrafe se incluyen la transmisión a través de medios que o bien permiten la multiplicación del microorganismo (vehículo común activo que comprende no solo el agua y los alimentos que serán tratados en el siguiente apartado sino la sangre y las soluciones para administración intravenosa), y los vehículos que sólo transmiten el microorganismo (vehículo común pasivo). Los objetos contaminados que pueden ser vehículos pasivos de diseminación de microorganismos incluyen los utensilios de cocina, pañuelos, juguetes u objetos escolares, lápices, etc. que pueden ser vehículo de neumococo, rinovirus, enterobacterias, enterovirus, etc. Las manos sucias son una vía de dispersión especialmente importante: las manos del personal hospitalario son la vía más frecuente de transmisión de enfermedades nosocomiales. TRANSMISIÓN POR AGUA O ALIMENTOS CONTAMINADOS La vía de entrada de estos patógenos es el tracto gastrointestinal. Estos microorganismos pueden producir infecciones o intoxicaciones alimentarias y, por tanto, es común referirse a ellos como agentes de toxiinfecciones alimentarias. La mayoría de los microorganismos patógenos que usan esta vía de transmisión son vehiculizados por comida, manos, heces y moscas. Si el alimento o el agua es calidad dudosa, un método eficaz de prevención es hervir el agua y pelar o cocinar los alimentos. Las enfermedades causas por este tipo de patógeno son generalmente de tipo gastrointestinal. La contaminación del alimento suele comenzar por su manipulación en condiciones poco higiénicas seguida de una conservación deficiente (a temperatura superior a 4ºC). Toxiinfecciones bacterianas por vía de agua o alimentos contaminados • Infecciones Salmonelosis 112 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) • Infecciones por diferentes tipos de Escherichia coli virulentos. Fiebre tifoidea producida por Salmonella typhi Cólera causado por Vibrio cholera Intoxicaciones Causadas por Staphylococcus aureus Botulismo causado por la toxina de Clostridium botulinum Infecciones víricas transmitidas por el agua o por alimentos contaminados Virus de gastroenteritis viral aguda Rotavirus Hepatitis A Polio TRANSMISIÓN POR CONTACTO DIRECTO Se trata de una transmisión persona a persona y puede ser por contacto físico, transmisión vertical y por inoculación directa. Los microorganismos que requieren un contacto directo para su transmisión se deshidratan con facilidad, son sensibles al oxígeno o a ambientes oxidantes, son parásitos intracelulares obligados o presentan alguna combinación de estas características. Dentro de este grupo de patógenos se encuentran de forma destacada los causantes de enfermedades bacterianas de transmisión sexual tales como Gonorrea (Neisseria gonorrhoeae) Enfermedad pélvica inflamatoria y uretrititis causadas por Chlamydia trachomatis Sífilis causada por Treponema pallidum VIH y enfermedades de transmisión por contacto no sexual como Carbunco causado por contacto con esporas de Bacillus anthracis. Enfermedades víricas transmitidas por contacto directo Citomegalovirus (mononucleosis infecciosa) Herpesvirus de tipo I y II Virus del papiloma humano Virus de la rabia VIH Transmisión vertical Sífilis congénita Toxoplasmosis Hepatitis B Citomegalovirus Rubéola VIH TRANSMISIÓN POR HERIDAS Y POR VECTORES 113 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) 1. Las enfermedades transmitidas por un vector (animal que transporta un microorganismo causante de una enfermedad infecciosa). Este vector puede ser pasivo 8como las moscas que sólo dispersan el patógeno) o activo ( cuando parte del ciclo biológico del patógeno tiene lugar en el vector). Entre las enfermedades transmitidas por esta vía se incluyen Enfermedades de origen bacteriano Peste (causada por Yersinia pestis y transmitida por una pulga) Enfermedades de tipo parasitario Malaria o paludismo, causada por Plasmodium falciparum y transmitida por mosquitos del género Anopheles. Enfermedades de tipo viral Fiebre amarilla transmitida por mosquitos de la especie Aedes aegyptii Encefalitis víricas de origen tropical (fiebre del Nilo, etc.) transmitidas por artrópodos Dengue transmitida por mosquitos 2. Entre las enfermedades cuya vía de entrada son las heridas destacan Enfermedades de origen bacteriano Tétanos (causado por Clostridium tetani) Gangrena gaseosa causadas por Clostridium perfringens. Enfermedades de tipo vírico HIV causante del SIDA Virus de las hepatitis B y C 114 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Tema 7.- Esterilización, desinfección y asepsia. Esterilización, desinfección, descontaminación y antisepsis. Desinfectantes y antisépticos. Evaluación de la actividad germicida. Desinfectantes de alta, media y baja actividad. Desinfección de endoscopios. Esterilización por calor. Esterilización por óxido de etileno. 1.- ESTERILIZACIÓN, DESINFECCIÓN, DESCONTAMINACIÓN Y ANTISEPSIS. En todos los ambientes abiertos y en muchos lugares del cuerpo se puede encontrar una gran cantidad de microorganismos (principalmente bacterias y virus). Estos microorganismos, si tienen a su disposición nutrientes y condiciones ambientales adecuadas pueden crecer y multiplicarse produciendo en su caso enfermedades infecciosas. En los ambientes naturales hay mezclas complejas de muchos tipos de microorganismos que forman poblaciones que comparten el mismo ambiente. Con objeto de evitar los efectos nocivos de los microorganismos, es necesario disponer de métodos que permitan eliminarlos de manera que podamos conseguir ambientes limpios sin contaminación microbiana. Se dice que un ambiente es estéril cuando se han eliminado todos los microorganismos del mismo. La esterilidad se puede alcanzar usando procedimientos físicos (calor, radiaciones), químicos o mecánicos (filtración). Sin embargo, los procedimientos de esterilización son costosos y, en ciertas ocasiones, desaconsejables. Por ejemplo, la esterilización completa de ciertos alimentos no es posible sin destruir sus características nutritivas. Por su parte, la antisepsis es la técnica que permite eliminar los microorganismos de la piel o de los tejidos vivos. En la práctica es imposible afirmar la ausencia absoluta de microorganismos en una muestra determinada. En este sentido, se considera que un producto está estéril desde el punto de vista clínico cuando existe menos de una posibilidad ente un millón (10-6) de que existan en él microorganismos viables. También se usa el término desinfección referido a la técnica que empleando calor o substancias químicas reduce la carga microbiana o elimina los microorganismos de las superficies. Un término parcialmente sinónimo es el de descontaminación que es un poco más general ya que en él se aplican además del calor, otros métodos de eliminación de microorganismos. Se dice que un ambiente es aséptico cuando se han eliminado todos los microorganismos patógenos. Un ambiente aséptico no tiene porqué ser estéril. La asepsia también se puede conseguir por procedimientos físicos y químicos. LIMPIEZA La limpieza tiene por objetivo dejar un objeto libre de contaminación. En algunas ocasiones es suficiente con la limpieza en el medio hospitalario; sin embargo, en otras es necesario proceder a la desinfección o a la esterilización. Nada debe ser sometido a 115 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) procedimientos de desinfección o esterilización si no ha sido sometido anteriormente a limpieza manual o a limpieza mecánica (con lavadoras). En el medio hospitalario, los principales agentes de limpieza son: • Hipoclorito sódico • Derivados fenólicos • Complejos trialdehídicos quirúrgicos • Polvo abrasivo clorado Por otra parte, la limpieza en el medio hospitalario será diferente si se trata de: • Zonas sin contacto con enfermos • Zonas de hostelería y lavandería • Servicios y cuartos de baño • Áreas de hospitalización, consultas externas y servicios generales • Áreas críticas (quirúrgicas, intensivos, prematuros, hemodiálisis, quemados, dia- gnósticas intervencionistas) • Banco de sangre, laboratorios, hospital de día, urgencias, habitaciones con aisla- miento indicado. El personal de enfermería debe vigilar de forma personal la limpieza de su área asistencial. La limpieza del material hospitalario en muchas ocasiones requiere personal especializado adiestrado en el manejo de aquellos materiales que por ser huecos (botellas) o tubulares con diámetro pequeño (canales de endoscopía por ejemplo) presentan dificultades especiales. CLASIFICACIÓN DE LOS MATERIALES SEGÚN SU USO Los equipos que se reutilizan en los hospitales se pueden clasificar según su aplicación en: Elementos críticos que entran en contacto con el sistema vascular u otras zonas estériles del cuerpo (catéteres intravasculares) • Elemento semicrítico que entra en contacto con la mucosa (tubos endotraqueales) • Elementos no críticos que entran en contacto con la piel intacta (cuña de orina) • Todos los elementos que vayan a ser reutilizados deben ser limpiados previamente. Además, los elementos críticos deben ser esterilizados, los semicríticos sometidos a una desinfección de alto grado y los no críticos a una desinfección de bajo grado. 2.- DESINFECTANTES Y ANTISÉPTICOS Se denominan antisépticos aquellos compuestos químicos que desarrollan su acción letal inteaccionando de forma inespecífica con los componentes celulares de forma que no existe una acción selectiva frente a grupos de microorganismos sino que su acción es más general. Los antisépticos no suelen ser demasiado tóxicos y pueden aplicar116 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) se sobre tejidos vivos. Esta falta de especificidad de los antisépticos los diferencia de los antibióticos cuya principal característica es la selectividad. Los desinfectantes son productos químicos que matan los microorganismos y se aplican sobre objetos inanimados, mientras que los antisépticos, por su menor toxicidad, se emplean sobre tejidos vivos. Puesto que dependiendo de la forma como se realice el tratamiento un mismo agente puede utilizarse como antiséptico o como desinfectante, se suele usar el término germicida para englobar ambos conceptos. Los diferentes tipos de microorganismos o de sus formas de desarrollo (esporas vs. células vegetativas) tienen diferentes grados de sensibilidad a los tratamientos físicos o químicos. En muchos casos, el tratamiento con agentes desinfectantes no elimina completamente los microorganismos presentes, sino que simplemente se reduce mucho su número de forma que la acción indeseable de los microorganismos se retrasa. Las esporas bacterianas son las formas más resistentes a los antisépticos y desinfectantes y sólo mueren al ser tratadas con agentes con alta actividad germicida. En general las formas vegetativas de las bacterias son sensibles a todos los agentes desinfectantes, aunque algunos grupos de microorganismos tales como las micobacterias pueden presentar especial resistencia a los de baja actividad. Los hongos presentan, en general, mayor resistencia que las bacterias y resisten los desinfectantes de baja actividad. Por último, los virus presentan una sensibilidad similar a la de las bacterias, aunque es un poco más elevada en el caso de los virus desnudos que no presentan envueltas lipídicas. 3.- EVALUACIÓN DE LA ACTIVIDAD GERMICIDA. La determinación del efecto antiséptico o desinfectante de los diferentes productos es complicada porque este efecto depende de gran número de factores externos (temperatura, humedad, pH, etc.) así como de los diferentes tipos de microorganismos que se desea eliminar o controlar. Existen protocolos que regulan cómo se debe evaluar la eficacia de un compuesto germicida y entre ellas destaca la prueba del coeficiente del fenol (CF) en la que se toma como referencia de desinfectante el fenol y como referencia de microorganismos Staphylococcus aureus y Salmonella typhi. CF = dil fenol / dil desinf (Ecuación 17) donde dil fenol es el inverso de la mayor dilución del fenol que elimina completamente la bacteria de referencia en 10 min de tratamiento, y dil desinf es la mayor dilución del desinfectante que elimina el microorganismo de referencia en 10 min de tratamiento realizado el estudio de los supervivientes tras un cultivo de 48h45. El valor de CF es simplemente indicativo ya que se trata de una medida realizada sobre cultivos puros y en la realidad, los germicidas se usan sobre poblaciones mixtas. 45 Por ejemplo, si dil fenol es 1/90 y dil desinf es 1/450, el cf será de 50; esto es, el desinfectante es 50 veces más activo que el fenol. 117 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Para evaluar la acción de un germicida frente a un microorganismo en particular se realizan test de dilución similares a los realizados para los antibiogramas cualitativos o cuantitativos. En función de su CF, los germicidas se clasifican en de actividad alta, media o baja. Si se desea realizar una esterilización se deberán escoger germicidas de actividad más alta o a mayores concentraciones. Igual ocurre si en la muestra existen substancias que protegen a los microorganismos de la acción de los germicidas (como ocurre en el caso de la sangre o en las heces). 4.- DESINFECTANTES DE ALTA, MEDIA Y BAJA ACTIVIDAD. FACTORES QUE INFLUYEN EN LA DESINFECCIÓN Los factores que afectan la eficiencia de un proceso de desinfección son los siguientes: naturaleza del microorganismo, su número, la concentración del germicida,. el tiempo de actuación, la temperatura, pH, el deterioro del producto en agua o por almacenamiento, la inactivación por la presencia de materia orgánica (moco, pus, heces, agua dura, etc.), la presencia de otros compuestos que modulen la actividad del germicida (detergentes, corcho, etc.) y las características de la superficie sobre la que se aplica. PRINCIPALES FAMILIAS DE AGENTES ANTISÉPTICOS Agentes oxidantes • • Halogenados • Gas cloro, y Compuestos de cloro (500-5000 mg/l), agentes oxidante que se usa para desinfectar el agua. • Soluciones de iodo, agente oxidante que inactiva las proteínas, se usa en la piel, jabones, etc. • La tintura de iodo es una solución al 2% (o más) de yoduro potásico en agua y etanol (CF ≈ 4.0 - 5.0) • Solución de iodo, se usa en instrumental médico • Povidona yodada (Betadine) que es un complejo de iodo con polivinilpirrolidona de forma que se libera el germicida lentamente, se inactiva menos por materia orgánica y penetra mejor. Se utiliza en desinfección de la piel e incluso en lavados quirúgicos. • Bromo • Flúor Otros oxidantes • Permanganato potásico • Peróxido de hidrógeno (agua oxigenada) en disolución del 6 al 30% (para esterilización en este caso), es una agente oxidante que se usa sobre la piel. • Ácido peracético • Sales del ácido persulfúrico 118 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) Agentes reductores • Aldehídos tales como el formaldehído (6-8%) y el glutaraldehído (2%) que inactivan las proteínas y se pueden usar como agentes esterilizantes. • Se suelen usar para esterilizar instrumental como soluciones acuosas. • El glutaraldehído a pH alcalino (Cidex) permite una desinfección de alto grado y se aplica al material que no puede ser sometido al autoclave. Es un compuesto tóxico e irritante que hay que utilizar de acuerdo con unos procedimientos especiales. Alcoholes • • Etanol • Solución de alcohol al 70% en agua (CF ≈ 0.04), disuelve los lípidos y desnaturaliza las proteínas, se usa sobre la piel. Alcohol isopropílico Fenol y derivados Compuestos fenólicos (0.5-3%), agentes oxidantes que se usan para desinfectar superficies. • El fenol fue históricamente el primer agente antiséptico utilizado en clínica. Actualmente no se emplea en clínica sino que se usan en hospitales y laboratorios derivados fenólicos menos cáusticos. • Los cresoles (Lysol, CF ≈ 2.3) que tienen importancia por ser tuberculocidas y activos durante largo tiempo; sin embargo, su olor es desagradable y pueden llegar a ser tóxicos. • El hexaclorofeno (CF ≈ 5-15 para S. typhi y 15-40 para Staph. aureus) es una agente muy activo que perdura durante largo tiempo. Se usa solo para desinfectar departamentos infantiles después de brotes con estafilococos. Sin embargo, puede ser tóxico y por eso su uso es limitado. • Bifenoles, rompen la membrana celular y se usan en jabones y lociones. • Agentes tensoactivos • • Sales de amonio cuaternario Tensoactivos anfóteros Derivados minerales y organominerales Dicloruro de mercurio, inactiva las proteínas y se usa para desinfectar mesas, suelos y otras superficies. • Compuestos organomercuriales, actúan inhibiendo las proteínas, se emplean en uso tópico (por ejemplo, la mercromina o mercurocromo CF de 2.7 para S. typhi a 5.3 para Staph. aureus) • Sulfato de cobre, precipita las proteínas y se usa como alguicida y como antifúngico. • 119 MICROBIOLOGÍA CLÍNICA Curso 2008 - 2009 (grupo 1) • Nitrato de plata, precipita las proteínas. Se usa como solución al 1% en ojos de recién nacidos para evitar la ceguera producida por N. gonorrhoeae o por C. trachomatis. Clorhexidina antiséptico que combina la acción germicida de los compuestos yodados y la permanencia del hexaclorofeno. Es un bactericida eficaz, aunque tiene menos eficacia sobre micobactrias y sobre esporas. • Se usa en la desinfección de la piel al 5%, en la de la boca (0.5 - 0.1%) y en jabones. • N-duopropenida • compuesto formado por dos yoduros de amonio cuaternario con gran actividad y rapidez frente a bacterias y hongos, aunque menos activo frente a micobacterias. Es de baja toxicidad por lo que puede usarse para la esterilización de endoscopios. 5.- DESINFECCIÓN DE ENDOSCOPIOS La esterilización de endoscopios es muy importante porque pueden ser un elemento de transmisión de procesos infecciosos tales como la hepatitis B o C, VIH o micobacterias. El desinfectante que se emplee debe ser rápido y enérgico, capaz de destruir las micobacterias no tuberculosas y Mycobacterium tuberculosis sin dañar el instrumento. El desinfectante más aconsejado es el glutaraldehído alcalino o la N-duopropenida. 120 MICROBIOLOGÍA CLÍNICA Curso 2004 - 2005 (grupo 1) Antisépticos y Desinfectantes usados en aplicaciones relacionadas con la salud Agente Antisépticos Mercuriales orgánicos Nitrato de plata Uso en campos relacionados con la salud Modo de acción Piel Ojos de los recién nacidos para evitar la ceguera Piel Jabones, desodorantes y lociones corporales Se combina con grupos –SH de las proteínas Precipita proteínas Alcohol (etanol al 70% en agua) Piel Bifenoles (Hexaclorofeno) Peróxido de hidrógeno (solución al 3%) Jabones, lociones Piel Disolvente de lípidos y desnaturalizante de proteínas Rompe la membrana celular Agente oxidante Solución de iodo Desinfectantes Dicloruro mercúrico Solución de Yodo Gas cloro Compuestos de cloro Compuestos fenólicos Detergentes catiónicos (compuestos de amonio cuaternario) Óxido de etileno Ozono Mesas, superficie de los bancos, suelo Instrumental metálico Depuración de los suministros de agua Suministros de agua Superficies Instrumental médico Material de laboratorio sensible a la temperatura como los plásticos Agua de bebida Ioda los residuos de tirosina de las proteínas Agente oxidante Se combina con grupos –SH Ioda los residuos de tirosina Agente oxidante Agente oxidante Desnaturaliza proteínas Interacciona con los fosfolípidos de la membrana Agente alquilante Fuerte agente oxidante MICROBIOLOGÍA CLÍNICA Curso 2002- 2003 (grupo 1) 6.- ESTERILIZACIÓN POR CALOR. Los microorganismos mueren rápidamente cuando son sometidos a temperaturas superiores a su óptima de crecimiento. Esto permite utilizar altas temperaturas para eliminar microorganismos por termodestrucción. Los métodos basados en el calor son quizá los más utilizados para controlar el crecimiento microbiano. La sensibilidad de los diferentes tipos de microorganismos a los tratamientos térmicos es distinta. Las esporas y algunos virus son la formas más termorresistentes y las células vegetativas las más sensibles. Por otro lado, los microorganismos Grampositivos tienden a ser más resistentes que los Gram-negativos. Por consiguiente, desde un punto de vista práctico, la esterilización por calor está destinada a matar las esporas bacterianas. El medio en el que se encuentra un microorganismo influye en su sensibilidad al calor. Por lo general, los microorganismos son más sensibles a las altas temperaturas cuando se encuentran a pHs ácidos, mientras que las concentraciones altas de proteínas o azúcares en el medio disminuyen la efectividad del calor y protegen a las bacterias. Las altas concentraciones de sal tienen efectos variables según el tipo de microorganismo La esterilización por calor se puede hacer usando calor seco o calor húmedo. ESTERILIZACIÓN POR CALOR SECO Horno Pasteur: usa calor seco transmitido por el aire al objeto en esterilización. En este caso, la temperatura más usual suele ser de 160ºC y durante dos horas. El uso del horno Pasteur está limitado por las características del material a esterilizar y se usa de forma limitada para la esterilización de objetos metálicos y de vidrio y para algunos productos secos que no puden mzclarse con el agua. Flameado: tiene usos muy limitados. ESTERILIZACIÓN POR CALOR HÚMEDO El autoclave esteriliza usando el calor húmero transmitido por vapor de agua sobrecalentado debido al uso de altas presiones. El efecto del vapor de agua es facilitar la transmisión del calor al objeto en esterilización. El procedimiento usual es usar 121ºC para lo que es necesaria una presión de 1.1 kg/cm2. En estas condiciones un tratamiento de 15 min es suficiente para eliminar las esporas de Gram-positivos. También puede usarse un ciclo más rápido de 134ºC y 3 min. En los autoclaves modernos es posible secar el producto una vez esterilizado antes de sacarlo del autoclave. Una versión reducida del autoclave es una olla a presión doméstica que, sin embargo, tiene el inconveniente e que no es posible secar el producto. Para evitar que los productos esterilizados en el autoclave se humedezcan con el vapor de agua se deben empaquetar en bolsas impermeables de papel Kraft o de poliamida. 119 MICROBIOLOGÍA CLÍNICA Curso 2002- 2003 (grupo 1) Puesto que el tratamiento térmico puede alterar las características del producto tratado, en el caso de alimentos o de productos termosensibles, se han desarrollado diferentes tipos de tratamiento industriales entre los que destacan: La Pasteurización destinada a reducir las poblaciones bacterianas. Se emplea principalmente en el tratamiento de alimentos (por ejemplo, leche) y tiene muy poca utilidad en clínica. La pasteurización de la leche se realiza a 71ºC durante 15 seg. y el tratamiento UHT consiste en calentar el producto a 140-150ºC durante unos pocos segundos. Este tratamiento permite un tiempo de conservación del producto mucho más largo (hasta 8 semanas). CINÉTICA DE TERMODESTRUCCIÓN: VALORES D YZ. La muerte de microorganismos como consecuencia de un tratamiento a altas temperaturas sigue una cinética exponencial. Si representamos la variación del logaritmo del número de células supervivientes a un tratamiento térmico realizado a una temperatura dada en función del tiempo de tratamiento, se obtiene recta cuya pendiente nos indica la velocidad de termodestrucción. Se define el valor D como el tiempo necesario para que el número de supervivientes caiga al 10% del valor inicial (o, lo que es lo mismo, para que el logaritmo del número de supervivientes se reduzca en una unidad). Si consideramos N0 como el número de células al inicio del tratamiento y Nx el número de células supervivientes después de un tratamiento de t minutos a una temperatura T, el tiempo de termodestrucción se calcula de la siguiente manera: 120 MICROBIOLOGÍA CLÍNICA Curso 2002- 2003 (grupo 1) La magnitud de D es tiempo (en muchos casos se usan los min; pero ciertos tratamientos son tan efectivos que resulta más práctico usar los s, que, por otra parte, son unidades del SI). El tiempo de termodestrucción (D) varía para cada temperatura (de ahí el subíndice t) de forma que a mayores temperaturas el valor de D es menor, es diferente para distintos microorganismos, distintos entornos y diferentes condiciones fisiológicas. Si aumentamos la temperatura de tratamiento, el valor de D disminuye de forma logarítmica. De manera análoga a como el valor D indicaba el tiempo necesario para lograr que el número de supervivientes se redujera al 10% de la población inicial, el valor z indica el incremento en la temperatura (medida en número de grados) necesario para que el valor D se reduzca a la décima parte del inicial. z = ∆T / [log (Dt1 / Dt2)] donde ∆T es el incremento de temperatura, y DT1 y DT2 los valores de D a las dos temperaturas estudiadas. Los valores D y z varían para cada microorganismo y para cada condición. Las esporas, por ejemplo, tienen valores D mucho más altos que las células vegetativas de los mismos microorganismos. Cuando el valor tratamiento se realiza a 121.1ºC (250ºF) al valor D se le denomina Dr y, por tanto, representa el tiempo necesario para lograr la destrucción del 90% de las células del microorganismo tratado a esa temperatura. 7.- ESTERILIZACIÓN POR ÓXIDO DE ETILENO Óxido de etileno (OE), agente alquilante que se usa en la esterilización del material de laboratorio, material de plástico y en general para aquellos productos que no pueden ser esterilizados por calor. 121 MICROBIOLOGÍA CLÍNICA Curso 2002- 2003 (grupo 1) La esterilización se lleva a cabo en un esterilizador similar a un autoclave que controla la concentración de óxido de etileno, la temperatura (60ºC) y la humedad (esterilización en condiciones de alta humedad). El OE puede usarse puro (la duración del tratamiento en este caso es de 3-4 horas) o como mezcla de OE al 10 - 20% con CO2 u otro gas reductor porque el OE es explosivo. El tratamiento dura varias horas. Se usa también en disoluciones de 450-500 mg/l. El OE es muy tóxico; pero se diluye rápidamente en el aire, lo que permite su eliminación fácil después del tratamiento. 122 MICROBIOLOGÍA CLÍNICA Curso 2002- 2003 (grupo 1) Tema 8.- Control de microorganismos. Inhibición del crecimiento. Grupos de antibióticos y quimioterápicos. Mecanismos de resistencia a antibióticos. Cepas multirresistentes. Precauciones en el uso de antibióticos. Tratamiento de las infecciones virales. Tratamiento de enfermedades causadas por protozoos y por hongos. 1.- INHIBICIÓN DEL CRECIMIENTO Se denomina antibiótica a aquella substancia que interfiere el crecimiento y la supervivencia de los microorganismos mediante una interacción específica (toxicidad selectiva) con alguno de sus componentes celulares. Debido a esta especificidad, los antibióticos tienen un espectro de acción limitado; esto es, en general son activos frente a ciertos microorganismos e inactivos frente a otros, lo que permite usarlos como agentes selectivos. Se denomina antiséptico aquel compuesto químico que desarrolla su acción letal interaccionando de forma inespecífica con los componentes celulares de forma que no existe una acción selectiva frente a grupos de microorganismos sino que su acción es más general. Los antisépticos no suelen ser demasiado tóxicos y pueden aplicarse sobre tejidos vivos. Los diferentes tipos de microorganismos o de sus formas de desarrollo (esporas, células vegetativas) tienen diferentes grados de sensibilidad a los tratamientos físicos o químicos. Además, el uso de antibióticos supone una presión selectiva sobre las poblaciones bacterianas que puede llevar a la substitución de las poblaciones del microorganismo sensibles al antibiótico por otras que han desarrollado mecanismos de resistencia frente al mismo. Según su modo de acción los antibióticos pueden ser Bacteriostáticos o Bactericidas. Un antibiótico bacteriostático causa una parada del crecimiento microbiano. Los microorganismos no crecen en presencia del antibiótico; pero tampoco mueren de forma inmediata. Si se elimina el antibiótico, los microorganismos pueden recuperarse y volver a crecer. Los microorganismos cuyo crecimiento está detenido por acción de un antibiótico bacteriostático van muriendo con el paso del tiempo en presencia del antibiótico; sin embargo, este proceso de muerte es lento. Un antibiótico es bactericida cuando la presencia del antibiótico produce la muerte del microorganismo afectado rápidamente. Esta muerte puede ir acompañada de la lisis de las células (y se habla entonces de antibiótico bacteriolítico) o no. Se denomina concentración mínima inhibitoria (CMI) de un antibiótico sobre un microorganismo, a la concentración mínima de dicho antibiótico que es capaz de impedir que dicho microorganismo siga multiplicándose. Cuando se consigue que la concentración de un antibiótico en sangre sea mayor que la CMI frente a un patógeno causante de una enfermedad infecciosa, se puede considerar que el antibiótico será capaz de curar la infección. 123 MICROBIOLOGÍA CLÍNICA Curso 2002- 2003 (grupo 1) Se denomina antibiograma al estudio de la efectividad de diferentes antibióticos sobre un microorganismo concreto que se quiere tratar. El antibiograma es un metido empírico para determinar el procedimiento de tratamiento de una enfermedad infecciosa. 2.- GRUPOS DE ANTIBIÓTICOS Y QUIMIOTERÁPICOS Las substancias antibióticas pueden clasificarse atendiendo a varios criterios: CLASIFICACIÓN POR SU ESPECTRO DE ACCIÓN Según su espectro de acción (tipo de microorganismos a los que afectan), los antibióticos pueden ser activos frente a Procariontes: en general presentan baja toxicidad frente a los eucariontes (células humanas) aunque hay que considerar que existen en estas ciertos orgánulos celulares de origen procariótico que pueden verse afectados y, por consiguiente, afectada la actividad celular general. Eucariontes: suelen presentar más problemas de toxicidad frente a células humanas ya que estas son eucarióticas. Ambos grupos. A ellos se aplica también el comentario del grupo anterior. Antivirales son compuestos que boquean el proceso de replicación y multiplicación vírica. Puesto que los virus son parásitos intracelulares que utilizan los componentes celulares del huésped para su multiplicación, los antivirales usados en clínica presentan pueden presentar un cierto grado de toxicidad para las células eucarióticas. Los antibióticos pueden usarse para evitar se desarrollen bacterias o microorganismos eucarióticos (hongos) en un determinado ambiente. Sin embargo, el uso preventivo de antivirales no tiene sentido puesto que los virus sólo pueden desarrollarse en el interior de una célula viva y en un ambiente sin células sólo pueden permanecer inactivos. CLASIFICACIÓN POR SU DIANA DE ACCIÓN Según su sitio de acción a nivel celular, los antibióticos pueden ser: Antibióticos que actúan bloqueando la síntesis y acción del ácido fólico. Este ácido es necesario para la síntesis de bases nitrogenadas. A este grupo de antibióticos pertenecen las sulfamidas y el trimetoprim. Antibióticos que actúan sobre la pared celular. En general bloquean la síntesis del peptidoglicano de manera que las bacterias pierden su protección osmótica y lisan. Por consiguiente, son antibióticos, en general, bacteriolíticos. 124 MICROBIOLOGÍA CLÍNICA Curso 2002- 2003 (grupo 1) El mecanismo de acción más común es el de impedir la incorporación de nuevos componentes en el peptidoglicano mediante la inhibición de las enzimas de transporte de los componentes o de síntesis de la pared celular. Estos antibióticos son activos únicamente frente a bacterias en crecimiento e inactivos frente a esporas y a eucariontes. A este grupo pertenecen los β-lactámicos (penicilinas como la penicilina, amoxicilina, y ampicilina; cefalosporinas como cefalotina, cefuroxima, y cefotaxima; y carbapenems como el imipenem y el meropenem), la vancomicina y teicoplanina, etc. Dentro de las cefalosporinas se distinguen generaciones que difieren en su eficacia frente a ciertos patógenos: primera generación son las iniciales, segunda generación presentan una mayor actividad frente a estafilococos, tercera generación aumentan su acción frente a Gram-negativos, cuarta generación que poseen actividad antianaerobia y frente a Pseudomonas. Antibióticos que actúan sobre la membrana celular. En general su actividad se debe a que forman poros en la membrana plasmática de forma que se rompe su integridad, se destruyen los gradientes de iones que son necesarios para la obtención de energía y se produce la pérdida de solutos celulares. A este grupo pertenece la Nisina, antibiótico cuyo uso como aditivo alimentario está autorizado. Hay antibióticos de este grupo que actúan sobre procariontes y otros que lo hacen sobre eucariontes. Debido a que las membranas de los dos tipo de células son ligeramente diferentes, los antibióticos de este grupo no suelen ser activos simultáneamente sobre los dos tipos de células. Antibióticos que interfieren las funciones del cromosoma bacteriano. A este grupo pertenecen las quinolonas: ácido nalidíxico, norfloxacina, ciprofloxacina y oxfloxacina. Antibióticos que inhiben la transcripción en bacterias como la Rifampicina46 Antibióticos que actúan bloqueando la síntesis de proteínas. Son conocidos como antibióticos ribosomales ya que actúan sobre estos orgánulos. Puesto que los ribosomas procarióticos y eucarióticos son diferentes en ciertos aspectos, existen antibióticos antibacterianos (tipo I), anti-eucarióticos (tipo II) y antibióticos que actúan sobre los elementos comunes a ambos tipos de ribosoma y, por tanto, son activos frente a procariontes y a eucariontes (tipo III). Muchos de estos antibióticos son bacteriostáticos y su efecto es reversible si se eliminan del medio. Otros, sin embargo, son bactericidas porque inducen la formación de proteínas incorrectas o porque inactivan irreversiblemente los ribosomas. 46 La Rifampicina es utilizada principalmente en el tratamiento de la tuberculosis. No debe usarse de forma aislada porque suelen aparecer resistencias. 125 MICROBIOLOGÍA CLÍNICA Curso 2002- 2003 (grupo 1) A este grupo pertenecen antibióticos tales como el cloranfenicol, los aminoglicósidos (gentamicina, tobramicina, amikamicina), los macrólidos (eritromicina y claritromicina), y las tetraciclinas. 3.- MECANISMOS DE RESISTENCIA A ANTIBIÓTICOS. Existen varios mecanismos de resistencia a los antibióticos. Las resistencias aparecen en las poblaciones por mutación y se establecen por la selección que ejerce la presencia del antibiótico en las poblaciones de microorganismos. Los genes de resistencia pueden transmitirse de forma horizontal. Esta transmisión es más frecuente en el caso de resistencias codificadas en genes plasmídicos. Los mecanismos e resistencia más generales son cuatro: modificación de la diana del antibiótico (por ejemplo: mutación que hace a una PBP insensible a un β-lactámico), modificación del antibiótico (por ejemplo, las β-lactamasas que inactivan los βlactámicos o la cloranfenicol-acetil transferasa que inactiva el cloranfenicol), modificación de la vía de entrada del antibiótico (por ejemplo, cambios en las porinas que afectan a la entrada de antibióticos en Gram-negativos); y sistemas de bombeo del antibiótico al exterior de la célula. 4.- CEPAS MULTIRRESISTENTES. El hecho de que las mutaciones de resistencia plasmídicas sean fácilmente transmisibles de forma horizontal y de que los plásmidos tiendan a acumular genes de resistencia a diferentes antibióticos, ha generalizado el problema de las cepas multirresistentes. Hay ciertas evidencias de que la transferencia horizontal de resistencia a antibióticos puede ser una respuesta adaptativa del microorganismo a la presencia misma del antibiótico. Así, se ha visto que la transferencia de plásmidos con resistencia a tetraciclina en Bacteroides está estimulada por la presencia de este antibiótico47. El problema de la resistencia a antibióticos es de gran importancia porque estos pierden su valor como agentes quimioterápicos muy rápidamente. Por esta razón es imprescindible el establecimiento de una política de antibióticos tanto a nivel del hospital como a niveles más altos de la política sanitaria. El mal uso indiscriminado de antibióticos favorece la aparición y la diseminación de las resistencias. 5.- PRECAUCIONES EN EL USO DE ANTIBIÓTICOS. Para evitar la aparición de resistencias, una estrategia quimioterapéutica es la utilización de combinaciones de antibióticos. En ciertos casos estas asociaciones son más activas que la suma de los dos antibióticos por separado, en este caso se dice que la ac47 ASM News 71: 412-419. 126 MICROBIOLOGÍA CLÍNICA Curso 2002- 2003 (grupo 1) ción es sinérgica. Sin embargo, la combinación de antibióticos presenta en algunos casos problemas de aumento de la toxicidad y de efecto antagónico de los antibióticos entre sí. Por otra parte, hay que considerar el posible efecto tóxico de los antibióticos debido a que la diana para ciertos antibióticos está también presente en células eucarióticas, a que el antibiótico puede tener efecto sobre sistemas celulares del enfermo diferentes de la diana, a posibles reacciones de hipersensibilidad al antibiótico (alergias), y a efectos secundarios debidos a la alteración de la flora normal (por ejemplo, el efecto causante de la diarrea post-antibiótico). 6.- TRATAMIENTO DE LAS INFECCIONES VÍRICAS. Actualmente se dispone todavía de un número limitado de agentes antivirales que presentan, además, una cierta toxicidad para el paciente ya que los mecanismos enzimáticos de los virus son comunes a las células eucarióticas. Además de los fármacos que veremos a continuación, hay que considerar que en la terapia de las enfermedades víricas juegan un papel importante los sistemas de inmunoterapia basados en el uso de inmunoglobulinas y en el empleo de interferón (en el caso del tratamiento de virus del papiloma). Los agentes antivirales están diseñados para impedir la replicación del material genético del virus. Entre los fármacos antivirales se encuentran los siguientes: Inhibidores de la ADN polimerasa vírica. Entre estos se encuentran: foscarnet usado en el tratamiento tópico de herpesvirus y como inyectable para el tratamiento de citomegalovirus en pacientes inmunocomprometidos. Se acumula en la médula donde inhibe la replicación del ADN. • aciclovir es un profármaco que se activa sólo en células infectada por virus herpes. Por tanto no es tóxico para células sanas. Se usa para tratar virus herpes y varicelazóster. • ganciclovir está muy relacionado con el anterior. Se usa para tratar citomegalovirus. • ribavirina. Inhibe la replicación de muchos virus de ADN y de ARN. Se usa como aerosol para tratar el virus respiratorio sincitial. • Inhibidiores de la transcriptasa reversa • AZT o azidotimidina o zidovudina. Es un profármaco que se activa en las células infectadas por un retrovirus como el HIV. Este fármaco sólo inhibe la multiplicación del virus; pero no lo elimina cuando está en forma latente. Bloqueadores de canales iónicos • Amantadina se utiliza para prevenir la influenza A y para reducir sus síntomas tempranos. No es activa frente a la influenza B. Inhibe la replicación del virus interaccionando con proteínas de su cápsida. 127 MICROBIOLOGÍA CLÍNICA Curso 2002- 2003 (grupo 1) • Rimantadina semejante a la anterior; pero menos tóxica. Inhibidores de proteasa • Indinavir, ritonavir y saquinavir. Inhiben el procesamiento de algunas proteínas esenciales para la replicación del HIV. Son unos antivirales efectivos cuando se usan en combinación con inhibidores de la transcriptasa reversa; pero se detectan resistencias con rapidez. Otros inhibidores de la multiplicación vírica • Oseltamivir. Es un inhibidor de la neuraminidasa del virus influenza con lo que impide su crecimiento y propagación por el cuerpo. 7.- TRATAMIENTO DE ENFERMEDADES CAUSADAS POR PROTOZOOS Y POR HONGOS. Los agentes antiparasitarios se clasifican en antiprotozoarios y antihelmínticos. El mecanismo de acción más común de los antiprotozoarios es la inhibición de la síntesis del ácido nucleico; mientras que en el caso de los antihelmínticos el mecanismo de acción puede ser por acción sobre el sistema nervioso del gusano o por bloqueo del sistema de transporte de glucosa en el gusano. Existe una larga lista de productos antiparasitarios. Hay que tener en cuenta que todos ellos tienen efectos adversos en los pacientes. Los agentes fungicidas se han desarrollado principalmente para interferir con la membrana del hongo o con su síntesis. Su especificidad está basada en la presencia de ergosterol en estas membranas y, por tanto, la mayoría de ellos bloquean su síntesis. También hay algunos otros que actúan sobre otras dianas fúngicas. Hay que considerar que muchos de los fármacos fungicidas son tóxicos para los pacientes si se emplean de forma sistémica. Los principales antifúngicos son: antibióticos poliénicos como la anfotericina B y la nistatina. La anfotericina B es tóxica porque también actúa sobre el colesterol de las membranas humanas. La nistatina se usa para tratar candidiasis cutáneas o intestinales. • azoles: imidazoles y bistriazoles. Son fungistáticos. Se usan de forma tópica y algunos de forma sistémica. • análogos de nucleósidos como la flucitosina que se emplea para tratar infecciones urinarias de Candida. • antibióticos alilamínicos se usan para tratar infecciones por dermatofitos como la tiña o el pie de atleta. • griseofulvina es una antibiótico fungistático que se emplea para tratamiento de infecciones por dermatofitos resistentes a tratamiento tópico. Se administra de forma • 128 MICROBIOLOGÍA CLÍNICA Curso 2002- 2003 (grupo 1) oral para tratar infecciones del estrato córneo porque parece ser que se elimina por el sudor. Es un agente bloqueante de la mitosis fúngica. Se tolera bastante bien. 129 MICROBIOLOGÍA CLÍNICA Curso 2002- 2003 (grupo 1) Tema 9.- Defensa frente a microorganismos. Barreras a la entrada de organismos. Respuesta inespecífica a la infección. Respuesta inmune específica. Antígenos y anticuerpos. Células de la respuesta inmune específica. Mecanismo de la respuesta inmune específica. Ejemplo del desarrollo y función del sistema inmune: el sistema inmune intestinal. Reacciones de hipersensibilidad. Inmunodeficiencias. Enfermedades autoinmunes. Superantígenos. Inmunoterapia. 1.- BARRERAS A LA ENTRADA DE MICROORGANISMOS El cuerpo humano presenta una serie de barreras que evitan la colonización de las partes estériles por microorganismos y que contienen la flora normal en sus ambientes específicos. Hay barreras mecánicas (piel, las mucosas y las células ciliadas del epitelio respiratorio), sistemas mecánicos de expulsión de microorganismos (tos, expectoración, orina, sudor) y barreras químicas (pH ácido de la vagina, lisozima, complemento). La respuesta a una infección puede ser inespecífica o específica. 2.- RESPUESTA INESPECÍFICA A LA INFECCIÓN La respuesta inespecífica a la infección se produce mediante diversos procesos: INFLAMACIÓN Es un mecanismo de respuesta del huésped al daño producido en los tejidos. Se produce porque estos tejidos dañados liberan substancias químicas activas (histamina, prostaglandinas) que inducen una vasodilatación y un aumento de la permeabilidad de los vasos sanguíneos. Por su parte, los fagocitos salen de los vasos sanguíneos (diapedesis) mediante un movimiento quimiotácticos hacia el foco de inflamación donde eliminan los agentes infecciosos. Finalmente se produce la reparación del tejido dañado. Los síntomas de la inflamación son rubor, calor, tumor y dolor. FAGOCITOSIS Los fagocitos son un tipo de leucocitos especializados en la eliminación por fagocitosis de los cuerpos extraños que invaden el cuerpo. La fagocitosis está inducida por anticuerpos específicos que marcan el objeto a fagocitar mediante un proceso denominado de opsonización. Hay varios tipos de fagocitos: 1.- Leucociotos polimorfonucleares producidos en la médula ósea. Son de vida relativamente corta y acción muy rápida. Forman las células del pus lo que las hace las principales defensas frente a bacterias piógenas. 130 MICROBIOLOGÍA CLÍNICA Curso 2002- 2003 (grupo 1) 2.- Macrófagos son células de larga vida que viajan por la sangre en forma de monocitos que se convierten en macrófagos cuando salen del torrente circulatorio. Pueden ser macrófagos libres o macrófagos fijos, estos últimos están localizados en los ganglios linfáticos. Los macrófagos son las células más importantes del sistema de defensa conocido como sistma retículo endotelial. CELULAS ASESINAS (Natural Killer o NK) Son un tipo especial de linfocitos especializados en destruir las células infectadas por virus. Probablemente también actúen como defensas inespecíficas frente a protozoos y parásitos. SISTEMA DEL COMPLEMENTO Es un conjunto de proteínas que facilita la actuación de determinados anticuerpos producidos durante la respuesta específica. Por otra parte, es el principal mediador de la respuesta inflamatoria inespecífica. Se activa mediante una cascada (cascada del complemento) en la que van activándose los diferentes componentes que participan en la opsonización, en la estimulación de la respuesta inflamatoria y en actividad líticas que destruyen microorganismos. CITOCINAS Son proteínas solubles que comunican entre sí las células que participan en las respuestas inflamatoria e inmune. Sus actividades son variadas: quimioatrayentes de macrófagos, producción de lesiones en células durante el proceso inflamatorio, colaboración en la destrucción de células infectadas, etc. Entre las citocinas se cuentan las interleucinas, las linfocinas y los interferones INTERFERONES Son proteínas de pequeño tamaño producidas por las células eucarióticas como respuesta a la infección por virus. Son secretadas por las células infectadas y estimulan en otras células (son citocinas) la producción de proteínas con acción antiviral que bloquean los sistemas transcripción del ARN mensajero viral y destruyen el ARNm viral. Los interferones son específicos de especie. Su utilización como agentes antivirales se basa en su producción por técnicas de ADN recombinante. FIEBRE Es una respuesta general a la infección consistente en una elevación anormal de la temperatura. Se produce como una consecuencia de la alteración de los sistemas de control de la temperatura a nivel del hipotálamo por ciertos agentes (pirógenos) entre los que se en131 MICROBIOLOGÍA CLÍNICA Curso 2002- 2003 (grupo 1) cuentran el lipopolisacárido y el pirógeno endógeno (interleucina 1) producido como respuesta a la inflamación. 3.- RESPUESTA INMUNE ESPECÍFICA La respuesta inmune específica es aquella que se pone en marcha como consecuencia de la invasión del organismo por un agente extraño y se caracteriza porque es específica del agente extraño o de partes de él sin reconocer otros agentes diferentes. En un cuerpo sano normal, pueden producirse muchas respuestas inmunes específicas simultáneamente que están dirigidas contra diferentes agentes extraños o contra diferentes partes de un determinado agente extraño. Se denomina inmunidad activa natural a aquella que se produce como consecuencia de un contacto accidental entre el agente extraño y el cuerpo. Por ejemplo, la que se produce como consecuencia de una infección. La inmunidad activa artificial es la que se induce como consecuencia de la puesta en contacto forzada entre el cuerpo y el agente extraño. Por ejemplo, en el caso de una vacunación. La inmunidad pasiva natural se produce cuando algunos elementos de la respuesta inmune (los anticuerpos) entran en el feto a través de la placenta. Por último, la inmunidad pasiva artificial es la que se adquiere como consecuencia del uso de inmunoglobulinas inyectados (por ejemplo, el uso de antitoxinas tetánicas). La respuesta inmune se basa en la acción de dos tipos de elementos: la inmunidad humoral es la consecuencia de la acción de los anticuerpos y la inmunidad celular la resultante de la acción de sistemas de células. La respuesta inmune, por último, tiene como propiedades características la memoria, la especificidad y la discriminación entre lo propio y lo extraño. 4.- ANTÍGENOS Y ANTICUERPOS Se denomina antígeno a aquella molécula o parte de una molécula que es susceptible de ser reconocida por el sistema inmune específico y provoca una reacción específica por parte de este. Se denomina anticuerpo a una proteína que es capaz de unirse de forma específica a un antígeno y que es sintetizada por una célula del sistema inmunitario. La reacción antígeno-anticuerpo consiste en la unión específica del anticuerpo al antígeno en una zona determinada de este. Los anticuerpos neutralizan toxinas y virus, bloquean las adhesinas, activan el complemento (IgG e IgM) y opsonizan antígenos de forma específica. Los antígenos pueden tener varios epítopos (o determinantes antigénicos) que son las partes de las moléculas capaces de inducir la producción de un solo tipo de anticuerpos. Un antígeno determinado (por ejemplo, una proteína extraña) tiene un gran número de determinantes antigénicos por lo que es capaz de inducir la producción de una gran variedad de anticuerpos que reconocen cada uno de los epítopos que tiene. 132 MICROBIOLOGÍA CLÍNICA Curso 2002- 2003 (grupo 1) Para ser antigénica, una molécula debe contener epítopos, ser biodegradable y tener un cierto tamaño (normalmente mayor de 10 KDa). Una molécula menor de ese tamaño puede inducir una respuesta inmune si se asocia con otras para formar un complejo de tamaño molecular suficiente. A las moléculas pequeñas que por sí mismas no son antigénicas y sí lo son en asociación con otras moléculas portadoras se les denomina haptenos. En este sentido, una molécula antigénica con varios epítopos puede considerarse como una molécula portadora de diferentes haptenos. Dentro del sistema inmune hay dos tipos de proteínas que pueden reconocer específicamente antígenos: las inmunoglobulinas que constituyen los anticuerpos solubles que se encuentran en el plasma sanguíneo, y los receptores de los linfocitos T que son proteínas especializadas de la membrana de este tipo de células. Por último, existe en todas las células un tercer grupo de proteínas especializadas en una interacción similar a la que se establece entre antígeno y anticuerpo que es necesaria para la identificación de células infectadas con virus. Son las proteínas del sistema mayor de histocompatibilidad (MHC). ESTRUCTURA Y TIPOS DE ANTICUERPOS Los anticuerpos están formados por dos cadenas proteicas pesadas (cadenas H) y dos cadenas ligeras (cadenas L) unidas por puentes disulfuro. Uno de los dos extremos de las cadenas ligeras y de las pesadas son muy variables (región variable) mientras que las partes centrales y el otro extremo de cada cadena es constante. En la región variable se encuentra la zona en la que se producirá la interacción con el antígeno y esa variabilidad es responsable de la variabilidad de los anticuerpos. Los anticuerpos pertenecen a un tipo de proteínas que forma la familia de las inmunoglobulinas. A esta familia pertenecen también los receptores de los linfocitos T y las proteínas del sistema mayor de histocompatibilidad (MHC). Existen cinco clases diferentes de inmunoglobulinas: IgA que se encuentran como monómeros circulantes en el suero y como dímeros de inmunoglobulina en las mucosas IgD, IgE, IgG tienen la estructura monomérica descrita. La IgG es capaz de atravesar la placenta y puede encontrarse en el feto causando una inmunidad natural pasiva. IgM forma pentámeros que no son capaces de atravesar la placenta. Estos anticuerpos son los que aparecen principalmente en la respuesta inmune primaria (la primera vez que se reacciona frente a la infección) Se denomina respuesta primaria a la que tiene lugar cuando se produce por primera vez una infección. En esta respuesta se sintetizan inicialmente IgM y al cabo de un tiempo IgG. En la respuesta secundaria que tiene lugar en sucesivas infecciones (o cuando la infección se cronifica) se produce principalmente IgG. 5.- CÉLULAS DE LA RESPUESTA INMUNE ESPECÍFICA. 133 MICROBIOLOGÍA CLÍNICA Curso 2002- 2003 (grupo 1) La respuesta inmune depende de la actividad de una serie de células producidas en la médula ósea durante el proceso hematopoyético. Las diferentes estirpes celulares del sistema inmune y su genealogía es la siguiente: a partir de una célula madre de la médula ósea se producen dos tipos de células precursoras, el precursor linfoide que dará lugar a los diferente tipos de linfocitos y a las células de memoria, y el precursor mieloide que dará lugar a las diversas células sanguíneas (plaquetas, eritrocitos, leucocitos polimorfonucleares y macrófagos). Las células de la rama mieloide participan en la respuesta inespecífica a la infección. Las células de la rama linfoide son de dos tipos: los linfocitos T, y los linfocitos B. Estos últimos, más adelante, pueden diferenciarse en células plasmáticas productoras de anticuerpos y células de memoria Como se ha dicho, las células del sistema inmune se producen en la médula ósea de los adultos y se desplazan por el cuerpo por medio de la sangre y del sistema linfático. En los ganglios linfáticos hay una gran densidad de células inmunitarias encargadas de eliminar agentes extraños y de inducir la respuesta inmune específica. Además de la médula ósea, hay dos órganos del cuerpo que participan de forma especial en el proceso de maduración de las células del sistema inmune: el bazo y el timo a los que hace referencia el nombre de los dos tipos células iniciales de la rama linfoide. 6.- MECANISMO DE LA RESPUESTA INMUNE ESPECÍFICA En el proceso de inducción de la respuesta inmune participan diferentes tipos de células. 1.-Activación de los linfocitos B. Los linfocitos B son una población de células que circula por nuestra sangre y sistema linfático. Dentro de esta población, hay muchas subpoblaciones que se diferencian porque en su membrana llevan una variante distinta de molécula de anticuerpo. Cuando un linfocito B encuentra en la sangre un antígeno que reconoce la inmunoglobulina que lleva en su membrana el linfocito se activa y se transforma en una célula plasmática que segrega grandes cantidades de su anticuerpo característico48. La activación del linfocito B también puede producirse como consecuencia de las señales que reciba de linfocitos T activados. Mientras haya antígeno presente, la célula plasmática continuará produciendo anticuerpos. Cuando el antígeno ya no esté más presente, algunos linfocitos B específicos se diferenciarán en células de memoria que tienen una larga vida y son fácilmente diferenciables en nuevas células plasmáticas si el antígeno volviera a estar presente, produciéndose, de esta forma, una respuesta inmune más rápida. 2.- Activación de los linfocitos T. Los linfocitos T maduran en el timo y se caracterizan por tener en su membrana unas proteínas relacionadas ontogénicamente con los anticuerpos que se denominan receptores de linfocitos T (TCR). Estos TCR son capaces de unirse específicamente a las proteínas de tipo MHC presentes en la superficie de los macrófagos cuando en estas proteínas, a su vez, hay restos de proteínas extrañas. Al 48 Inicialmente la célula plasmática recién diferenciada produce anticuerpos de tipo IgM. Posteriormente puede cambiar de clase y pasar a producir otros tipos de anticuerpos (IgA, G, D). 134 MICROBIOLOGÍA CLÍNICA Curso 2002- 2003 (grupo 1) activarse, los linfocitos T emiten señales moleculares (citoquinas) que estimulan a otros linfocitos de tipo B y a sí mismos entrando en un proceso de multiplicación y de diferenciación en sucesivas subpoblaciones de linfocitos T que reconocen el antígeno presentado por la proteínas MHC y se ocuparán de distintas funciones específicas de la respuesta inmune celular. Cada antígeno es capaz de activar sólo un tipo de células T y, además, una pequeña proporción de estas (en torno a 10-5). Sin embargo, hay ciertas moléculas denominadas superantígenos que son capaces de estimular una proporción mayor de linfocitos T (hasta una proporción de 0.2) de manera inespecífica. Esto da lugar a la producción de una gran cantidad de interleucinas y a síntomas tales como vómito, náuseas, shock. Entre los antígenos más conocidos están el de Staph. aureus responsable de shock tóxico y las toxinas estreptocócicas. 3.- Función de los macrófagos. Los macrófagos fagocitan inespecíficamente antígenos opsonizados y otros productos extraños o restos celulares. Cuando los macrófagos digieren proteínas, exportan algunos fragmentos de estas proteína digeridas al exterior de la célula uniéndolas a las proteínas del sistema MHC. De esta forma se presentan al exterior celular los restos de las proteínas digerida. Si la proteína digerida es extraña, también lo serán sus restos. Estos restos de proteínas extrañas presentadas al exterior por las proteínas del MHC son reconocidos por linfocitos T mediante sus receptores TCR. 4.- Función de las células plasmáticas Producir anticuerpos circulantes. estas células se producen como resultado de la diferenciación de linfocitos B activados. 5.- Función de las células de memoria Son individuos de una población de linfocitos B de larga vida que pueden ser activados rápidamente como consecuencia de una nueva infección. 6.- Las células de otros tejidos no macrófagos también tienen moléculas MHC en su superficie y pueden activar de esta forma linfocitos T. 7.- EJEMPLO DEL DESARROLLO Y FUNCIÓN DEL SISTEMA INMUNE: EL SISTEMA INMUNE INTESTINAL Para controlar las grandes poblaciones de microorganismos que hay en el intestino se desarrolla un sistema inmune específico que comprende varias barreras: 1.- El epitelio intestinal supone una barrera física y, además, segrega grandes cantidades de antibióticos peptídicos por las células de Paneth. Entre estos antibióticos peptídicos destaca la angiotensina 4 que actúa sobre Listeria y sobre Enterococus faecalis; pero no sobre las bacterias Gram-negativas. La cantidad de angiotensina 4 aumenta cuando cambia la dieta de la lactancia a la dieta adulta. 2.- La respuesta inmune específica (adaptativa) está mediada por células B y T parte de cuyo desarrollo tiene lugar en órganos linfoides específicos del intestino conocidos como placas de Preyer. El desarrollo de estas células depende del contacto con las bacterias intestinales de forma que las bacterias comensales ayudan al desarrollo de las placas de Preyer. 135 MICROBIOLOGÍA CLÍNICA Curso 2002- 2003 (grupo 1) 3.- Además de los linfocitos derivados de las placas de Preyer en el intestino existe una gran cantidad de células T que se denominan linfocitos intraepiteliales. Las bacterias también contribuyen al desarrollo de estos linfocitos en la superficie del epitelio. Se piensa que un inadecuado desarrollo del sistema inmune en la infancia por falta a la exposición a estimuladores (hipótesis de la higiene) puede dar lugar posteriormente al desarrollo de patologías alérgicas o, en el caso del intestino. Al desarrollo de enfermedades inflamatorias intestinales49. 8.- REACCIONES DE HIPERSENSIBILIDAD Las reacciones de hipersensibilidad (alergias) son un tipo de respuesta inmune en la que hay un daño en los tejidos del propio organismo. Hay varios tipos de reacciones alérgicas que dependen, en algunos casos, de anticuerpos y en una variante de células T. En los casos más graves (reacción anafiláctica) se puede producir shock y muerte del paciente como consecuencia de la liberación masiva de compuestos vasoactivos que terminan por producir un fallo circulatorio. Las reacciones anafilácticas son poco frecuentes; pero deben tenerse en cuenta en casos como los de alergia a la penicilina (que actúa como un hapteno) o en casos de terapia por vacunación. REACCIONES DE HIPERSENSIBILIDAD RETARDADA En este tipo de reacción participa un grupo de linfocitos T de hipersenxsibilidad retardada y se produce entre unas horas y unos días (24 – 48 h) del contacto del paciente con el agente inductor de la respuesta. Este tipo de reacción está dirigida contra microorganismos .y es el resultado del contacto de la piel con ciertos productos químicos sensibilizantes lo que produce dermatitis de contacto. Son ejemplos de esta reacción la alergia a la hiedra, a ciertos cosméticos y a ciertos fármacos. Los síntomas de esta hipersensibilidad son el prurito, eczema y respuesta inflamatoria (hinchazón). Como consecuencia de la reacción se produce una cierta destrucción de tejidos. Un ejemplo práctico de este tipo de reacción es la que se produce durante la prueba de la tuberculina (prueba de Mantoux) en la que el resultado positivo es consecuencia de una hipersensibilidad retardada. Hay algunas infecciones bacterianas que producen este tipo de respuesta. Entre ellas se encuentra la tuberculosis, lepra, brucelosis, parotiditis y ciertas infecciones fúngicas. En todas ellas se puede usar un ensayo de inyección del antígeno para comprobar la exposición previa al patógeno de forma similar a lo que se hace con la prueba de la tuberculina. 49 ASM News 71: 77-83 136 MICROBIOLOGÍA CLÍNICA Curso 2002- 2003 (grupo 1) REACCIONES DE HIPERSENSIBILIDAD INMEDIATA Esta respuesta está mediada por anticuerpos producidos por linfocitos T activados y comprende las alergias. La respuesta puede ser desde leve a grave (choque anafiláctico50). Los antígenos que causan respuestas alérgicas se denominan alergenos. Entre el 10% y el 20% de la población presenta algún tipo de respuesta alérgica. La respuesta anafiláctica se produce como sigue: un antígeno alergénico desencadena la producción de IgE51 que se unen por su parte constante a células cebadas (mastocitos52) y a leucocitos basófilos53. Después de una segunda exposición del organismo al antígeno las IgE se unen al él y estimulan la producción de mediadores alérgicos por los mastocitos y los basófilos. Entre estos mediadores están la histamins y la serotonina que provocan vasodilatación y contracción del músculo liso causando dificultades respiratorias, eritema, abundante producción de moco, estornudo, prurito y enrojecimiento ocular. Las razones por las que los alérgicos producen grandes cantidades de IgE en respuesta a substancias que no inducen este tipo de respuesta en otros individuos son todavía desconocidas. De hecho, un tercio de la gente que produce un exceso de IgE no desarrolla síntomas alérgicos. Hay algunos datos epidemiológicos curiosos tales como que las alergias son menos frecuentes en niños que viven en entornos rurales (hipótesis de que la higiene hace “vago” al sistema inmune), o que las alergias son más frecuentes en poblaciones que trabajan en oficinas. También se constata una pequeña diferencia en la incidencia por sexos: las alergias son ligeramente más frecuentes en niños que en niñas, aunque estas diferencias se compensan posteriormente en la edad adulta. El choque anafiláctico se caracteriza por una grave dificultad para respirar debido a una vasodilatación masiva con caída de la tensión arterial, prurito y eritema. Estos casos hay que tratarlos rápidamente con adrenalina para evitar la muerte del paciente. 9.- INMUNODEFICIENCIAS Pueden ser primarias cuando son debidas a anomalías genéticas o a defectos en el desarrollo embrionario (por ejemplo, niños que carecen de timo) y secundarias que pueden ser, a su vez, transitorias (durante ciertas enfermedades como el sarampión) o irreversibles (pérdida de la médula ósea, del bazo, etc.). Por otra parte, hay enfermedades infecciosas (como el SIDA) en las que las células del sistema inmune son eliminadas por el patógeno produciendo la inmunodeficiencia. Los pacientes inmunocomprometidos requieren un cuidado especial para prevenir las infecciones. 10.- ENFERMEDADES AUTOINMUNES 50 El término anafilaxis es etimológicamente sinónimo de alergia. La IgE es la respuesta natural del cuerpo humano a las infecciones por parásitos. 52 Los mastocitos son células del tejido conectivo adyacentes a los capilares en todo el organismo. 53 Los basófilos representan el 1% de la población leucocitaria. 51 137 MICROBIOLOGÍA CLÍNICA Curso 2002- 2003 (grupo 1) Se producen por la reactivación de linfocitos T o B autoreactivos. La mayoría de estas enfermedades son de curso lento; pero con el tiempo resultan graves. Hay enfermedades autoinmunes debidas a procesos mediados por anticuerpos y otras debidas a procesos mediados por células. Ejemplos de enfermedades autoinmunes: Diabetes juvenil o diabetes mellitus insulino-dependiente causada por anticuerpos autoreactivos que actúan sobre los islotes de Langerhans. Lupus eritematoso sistémico (SLE) producido por una respuesta masiva de anticuerpos contra diferentes componentes celulares, ADN, nucleoproteínas, factores de coagulación, etc. En esta enfermedad, los complejos antígeno-anticuerpo circulantes pueden depositarse en diferentes tipos de tejidos o en diferentes órganos (riñón, bazo) donde activan el sistema del complemento y se produce un daño local grave. Esclerosis múltiple causada por autoanticuerpos que reaccionan con componentes del sistema nervioso central Cuando el daño se localiza en ciertos órganos (diabetes mellitus, hipotiroidismo autoinmune) la enfermedad se puede controlar suministrando los productos de las células afectadas (insulina o tiroxina, respectivamente). Cuando la enfermedad es sistémica (esclerosis múltiple, SLE, miastenia grave) el tratamiento se basa en la utilización de inmunosupresores. Parece ser que el componente genético en el desarrollo de las enfermedades autoinmunes es muy importante. 11.- SUPERANTÍGENOS Se denominan así ciertas toxinas producidas por estafilococos y estreptococos responsables del síndrome de shock tóxico, enfermedad producida por la enterotoxina estafilocócica y por la toxina exfoliativa estreptocócica. En ambos casos, el desarrollo de la enfermedad depende de la acción del sistema inmune. Estos antígenos se unen a una de las cadenas de los receptores de linfocitos T (TCR) en un sitio diferente al normal. De esta forma se puede llegar a estimular al 10% de las células T del individuo afectado, mientras que un antígeno normal no estimula a más del 0.1% de los linfocitos T. Esta activación masiva de los linfocitos T causa un efecto inflamatorio generalizado y shock. 12.- INMUNOTERAPIA La inmunoterapia puede ser pasiva cuando se trata de estimular este tipo de respuesta inmune mediante la administración de anticuerpos o activa mediante la vacunación consistente en estimular la respuesta inmune mediante la administración de un antígeno protector capaz de inducir la producción de anticuerpos efectivos en el control del patógeno. 138 MICROBIOLOGÍA CLÍNICA Curso 2002- 2003 (grupo 1) La inmunoterapia es el mecanismo más efectivo de supresión de la infección a largo plazo. No siempre es posible, aunque ha permitido controlar un gran número de enfermedades. Sin embargo, aún hoy se estima que mueren cada año en torno a un millón y medio de niños menores de cinco años por enfermedades frente a las que hay vacunas y que en torno a 27 millones de niños pasa los cinco primeros años de vida sin acceso a vacunas disponibles54. En España, las enfermedades sometidas al programa nacional de vacunación son las siguientes (agrupadas por las vacunas que se administran simultáneamente): • poliomielitis • difteria, tétanos, tos ferina • sarampión, rubéola, parotiditis • hepatitis B • H. influenzae • Meningococo 54 Nature 433: 345. 139