Introducción
Anuncio

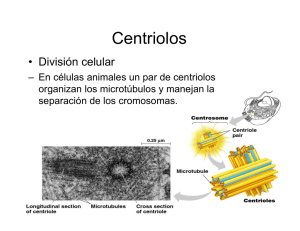
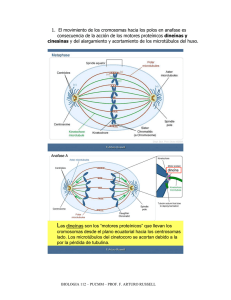
Introducción En eucariontes, los cromosomas de una célula somática son duplicados fielmente durante la fase S del ciclo celular y durante la mitosis aparecen como las dos cromátidas de los cromosomas, unidas solo por el centrómero. La mitosis es el proceso de distribución Interfase equitativa de las cromátidas entre las dos células hijas y está convencionalmente dividida en 4 fases : profase, metafase, anafase y telofase. Se dice que la división es convencional, pues este proceso es uno solo y por razones de estudio se le ha dividido en 4 fases. Además, los dos núcleos hijos son genéticamente idénticos al original. Profase Metafase Anafase Anafase Telofase Mitosis PROFASE 1. En el NÚCLEO. Se inicia la condensación de los cromosomas visibles al MO como finos filamentos entremezclados, los que van haciéndose cada vez más cortos y gruesos a medida que avanza la profase. De esta manera, se van organizando los cromosomas mitóticos visibles desde la mitad de la profase e integrados por 2 cromátidas paralelas y separadas, excepto por una unión en su constricción primaria. cromosoma centrómero cinetocoros Microtúbulos cinetocóricos cromátida Los centrómeros de mamíferos consisten en secuencias de ADN altamente repetidas. En humanos, cada secuencia consta de 171 pares de bases y ella está repetida varias veces. De aquí su gran tamaño al cual pueden unirse los cinetocoros. Cada cinetocoro está conformado por proteínas asociadas al centrómero. Al ME se observa como dos masa discoides de 0.5 – 1.0 µm de diámetro conteniendo un material finamente fibrilar. Detalles de mayor aumento muestran una capa densa interna unida a la cromatina, una capa densa externa que une los microtúbulos cinetocóricos y una capa clara entre ambas capas densas. El espesor de las capas electrón-densas es de 400 Å y la capa clara es de 300 Å. Mitosis Las proteínas asociadas a la capa externa parecen actuar como “motores moleculares” que guiarán el movimiento de los cromosomas a lo largo de las fibras del huso mitótico. Estudios inmunológicos muestran que las proteínas que forman los cinetocoros persisten en el núcleo interfásico asociadas a la envoltura nuclear. Los centríolos se encuentran en pares, cuyo conjunto se denomina centrosoma, dispuestos en ángulo recto y rodeados por un material electrondenso llamado material pericentriolar, que viene a constituir el centro organizador de los microtúbulos. Durante la interfase, el par de centríolos se duplica, así en la célula premitótica se encuentran 2 pares de centríolos perinucleares. El nucléolo desaparece progresivamente entre el inicio y la mitad de la profase. 2. En el CITOPLASMA Mientras está ocurriendo la condensación cromosómica y la desaparición nucleolar en el núcleo, en el citoplasma se inician los procesos tendientes a lograr una bipolaridad con la ubicación, en polos opuestos de la célula, de los 2 centríolos entre los cuales se extienden microtúbulos. El conjunto de microtúbulos se denomina huso mitótico y cada microtúbulo se llama fibra del huso. Se recordará que el centríolo es una formación microtubular cilíndrica (0.2 x 0.4 µm) que consta de 9 tríadas de microtúbulos (subfibrillas A, B y C, siendo la A la más interna). En la profase, el material pericentriolar se hace más denso y constituye el centro de nucleación para la formación inicial de los microtúbulos que constituirán el huso, es decir, es un centro inductor de polimerización de la tubulina. El material pericentriolar posee una proteína llamada gamma tubulina, la cual manifiesta estrecha relación molecular con las alfa y beta tubulinas, por ende, sirve como punto de nucleación para el desarrollo de los microtúbulos. Mitosis Los primeros microtúbulos se orientan de una manera irregular, pero luego, al ir aumentando en cantidad y longitud, los van haciendo a manera de rayos alrededor de cada complejo centriolar (material centriolar más el par de centríolos). Ahora se llaman microtúbulos del áster, que al MO conforman las fibras del áster. El conjunto formado por el complejo centriolar y las fibras del áster se llama áster, por esto este tipo de mitosis se llama mitosis astral, a diferencia de las mitosis anastrales observadas en vegetales superiores. Los ásteres migran progresivamente en direcciones opuestas en el citoplasma de la célula, produciéndose elongación de los microtúbulos que se extienden entre los ásteres, denominándose ahora microtúbulos polares (contínuos); algunos de ellos se apoyan en la envoltura nuclear. Por último, los dos pares de centríolos alcanzan puntos opuestos en la célula quedando determinados los dos polos del huso mitótico. La segunda parte de la profase, que suele llamarse prometafase, se caracteriza por el inicio de la ruptura de la envoltura nuclear en varios puntos. Se sabe que previo a la ruptura visible de la envoltura ha ocurrido una despolimerización de las proteínas de la lámina nuclear, como consecuencia de la fosforilación de las mismas por acción del MPF activo. Después de esta ruptura, que es progresiva, se evidencian los microtúbulos cinetocóricos (cromosómicos) desde los cromosomas, orientándose éstos hacia el cinetocoro más cercano; de esta manera, se mezclan los microtúbulos cinetocóricos con los polares, siendo sus trayectos paralelos. Los cromosomas duplicados tienen una posición al azar con respecto a los complejos centriolares y a los microtúbulos polares, pero al avanzar la prometafase se van orientando perpendicularmente a los polos, con sus microtúbulos cinetocóricos paralelos a los microtúbulos polares. Así, al final de la profase llegan a coexistir 3 grupos de microtúbulos : (a) microtúbulos cinetocóricos o cromosómicos, que se extienden desde el cinetocoro de una cromátida hasta el centrosoma que está más cercano a ella; (b) microtúbulos polares o entrelazados, se extienden desde el centrosoma hasta antes o después del plano ecuatorial. Sólo el 30 % de ellos alcanzan el polo contrario; y (c) microtúbulos libres, parecen corresponder a segmentos aislados de microtúbulos cinetocóricos y polares. Mitosis Estudios con luz polarizada muestran una alta birrefringencia en los semihusos, los que también muestran una alta fluorescencia al aplicar anticuerpos antitubulinas con fluoresceína. METAFASE La principal característica de la metafase es el ordenamiento de los cromosomas en el plano ecuatorial del huso. En esta estadio, los cromosomas se encuentran en su máxima condensación y comienzan una migración hacia el plano ecuatorial del huso. Este huso metafásico está integrado por las 3 categorías de los microtúbulos descritos anteriormente. La distribución de los microtúbulos es variable en el huso, ya que es más elevado en los semihusos que en la interzona. Para las células humanas se han reportado a ME unos 1500 microtúbulos en los semihusos y unos 900 en la interzona. Si consideramos que la célula humana posee 46 cromosomas, cada uno de ellos puede tener de 20 a 40 microtúbulos cromosómicos. Se considera que los cromosomas se mantienen en el plano ecuatorial debido a una tensión balanceada ejercida por la unión de los cinetocoros hermanos a los microtúbulos orientados a polos opuestos. De hecho, si se destruye selectivamente uno de los cinetocoros de un cromosoma con un microrayo laser, el cromosoma es retraído inmediatamente hacia el polo conectado con el cinetocoro intacto. Mitosis ANAFASE El comienzo de la anafase está indicado por la separación de las cromátidas a nivel de la constricción primaria. Cada cromátida se desplaza hacia el polo al que mira su cinetocoro y, simultáneamente, los microtúbulos cinetocóricos se acortan. Este desplazamiento es sincrónico : 0.2 a 5.0 µm/min aproximadamente y es independiente de la forma y tamaño de los cromosomas. A menudo el movimiento es más rápido al inicio de la anafase y se hace más lento posteriormente. El final de la anafase está indicada por la reunión de cada lote de cromátidas en un polo, las cuales constituirán los cromosomas de las futuras células hijas. Anafase A Migración de los cromosomas La proteína motora del complejo cinetocórico libera tubulinas Desensamble microtúbulo Cromosoma Conforme los cromosomas se mueven a sus respectivos polos, éstos se alejan entre sí y el huso se alarga (semihusos se acortan e interzona se alarga). Debido a estos dos movimientos la anafase suele dividirse en anafase A en la que los cromosomas se separan y anafase B en la que los polos se alejan. Los microtúbulos constituyen la estructura esencial en ambos movimientos, puesto que el empleo de la colchicina altera a ambos por igual. Se recordará que los microtúbulos son estructuras polarizadas debido a la orientación de sus unidades de tubulina y al modelo dinámico de ensamble y desensamble. Además, se ha comprobado que todos los microtúbulos de cada semihuso, tanto cinetocóricos como polares, tienen una orientación paralela; en cambio, los microtúbulos de un semihuso tienen orientación opuesta con los del otro semihuso. Mitosis Los microtúbulos cinetocóricos, al igual que todos los microtúbulos del huso, están orientados con su extremo (–) hacia el polo y el extremo (+) está unido al cinetocoro de una cromátida. El extremo (+) de los microtúbulos polares se extiende libre en la zona ecuatorial. Tanto el ensamble como el desensamble de ambos microtúbulos ocurre en los extremos (+). Experimentos in vivo e in vitro han demostrado que el movimiento de los cromosomas (anafase A) se acompaña por un desensamble coordinado de los microtúbulos cinetocóricos desde sus extremos (+) a nivel de los cinetocoros. Por otro lado, al producirse el acortamiento de los microtúbulos cinetocóricos, los cromátidas van acercándose progresivamente a cada polo y aumenta la cantidad de moléculas de tubulina en el citosol. Es importante señalar aquí que este movimiento NO requiere de ATP, lo que parece ser que el propio desensamble es suficiente para producir el movimiento. ANAFASE METAFASE Eliminación en el polo En este punto del proceso es necesario indicar que actúa la topoisomerasa II para separar las moléculas de ADN que se encuentran entrelazadas en esta región. Adición en el cinetocoro Microtúbulos cinetocóricos mantienen longitud constante Leve eliminación en el polo Rápida eliminación en el cinetocoro Microtúbulos cinetocóricos se acortan por despolimerización en ambos extremos Mitosis Anafase B Elongación del huso En la mitad de la anafase, cuando los cromosomas se aproximan a los polos, se produce un alargamiento del huso derivado de un aumento en la longitud de los microtúbulos polares. Así, el huso se hace más largo y más estrecho. En algunas células, este alargamiento es considerable : doblan su longitud (de 10 µm a 20 µm). Se considera que este alargamiento de los microtúbulos se debe a una agregación de nuevas unidades de tubulina (ensamble) en los extremos ecuatoriales de los microtúbulos polares. Recordemos que la mayoría de los microtúbulos polares llegan hasta un poco más allá del ecuador; por tanto, se establece una zona en que se solapan los microtúbulos provenientes de ambos polos. Además, la orientación de estos microtúbulos es con su extremo (–) ubicado en un polo y su extremo (+) en la zona de solapamiento. Existen moléculas llamadas kinesina que manejan ciclos ATP-ADP que permiten empujar los microtúbulos hacia el extremo (–); por esta razón la anafase B requiere de la hidrólisis de moléculas de ATP. Experimentos en ausencia de moléculas de tubulinas libres han demostrado que al agregar ATP, la elongación del huso está limitado a la longitud de la zona de solapamiento. Sin embargo, esto no explicaría el hecho que la elongación llega a doblar la longitud de la célula De aquí que si en el experimento anterior se añaden moléculas de tubulinas libres, la elongación del huso llega a ser varias veces la longitud de la zona de solapamiento original. Las kinesinas empujan los microtúbulos hacia los polos Mitosis Al final de la anafase se inicia la citodiéresis o citocinesis, que corresponde a la división en partes iguales del contenido citoplasmático y se ha demostrado que es un fenómeno contráctil. De hecho, se ha evidenciado por inmunofluorescencia y por ME que se acumulan microfilamentos de actina en la región ecuatorial, por debajo de la membrana plasmática. Además, en experimentos que utilizan citocalasina B que disgrega los microfilamentos, se inhibe la citodiéresis; en cambio, el uso de colchicina que disgrega los microtúbulos, no tienen efecto sobre la progresión del surco de división. TELOFASE La telofase se inicia cuando comienza a reconstruirse la envultura nuclear, que envuelve cada lote de cromosomas ubicados en los polos. La envoltura nuclear inicia su formación a partir del final de la anafase B, cuando comienza la polimerización de las láminas proteicas de la lámina nuclear. Sobre esta lámina se colocan segmentos cortos de RE, los cuales inician progresiva coalescencia que conduce a la formación de una envoltura nuclear contínua. Los poros aparecen muy precozmente y la cisterna perinuclear es al inicio de dimensiones variables haciéndose uniforme al final de la telofase. Simultáneamente, los cromosomas inician un proceso de descondensación hasta adquirir la disposición del núcleo interfásico. Los extremos polares de los microtúbulos polares se disgregan, quedando sólo los cercanos al ecuador del huso, los cuales se acercan unos a otros hasta fusionarse en un haz único, a medida que crece el surco de división. Mitosis Existe un modelo que intenta explicar la invaginación del surco de división, es el anillo contráctil. Los microfilamentos de actina se encuentran unidos a la membrana plasmática mediante la alfa-actinina. Los microfilamentos se disponen concéntricamente al surco de división, quedando entre ellos microfilamentos de miosina de tipo I. Las interacciones entre actina y miosina provocan la invaginación de la membrana en la región del surco de división. El anillo contráctil sigue contrayéndose y el surco subyacente profundizándose, hasta el punto en el cual solo queda un estrecho puente citoplasmático, por el cual se extiende el haz único de microtúbulos. Luego estos puentes se rompen quedando independientes las dos células hijas. En las células animales, la citodiéresis se inicia por una invaginación de la membrana plasmática, que va formar un surco circular que rodea la célula, es el llamado surco de división. En las células vegetales, que están protegidas por una pared celulósica rígida, las células hijas no pueden separarse más allá de los límites de la pared celular. De aquí que la citodiéresis sea diferente a la descrita para las células animales. En la región ecuatorial del huso se acumulan vesículas originadas del Golgi, cuyo conjunto se denomina fragmoplasto. Las vesículas se fusionan progresivamente hasta llegar a formar las membranas plasmáticas de las dos células hijas. Surco de división Mitosis MPF y la progresión hasta Metafase La mitosis incluye drásticos cambios en compuestos celulares que conducen a una reorganización de toda la célula. Estos cambios son iniciados por la activación del MPF (cdc2 y ciclina B). El MPF actúa como un regulador maestro de la transición hacia la fase M del ciclo celular. Además, fosforila algunas proteínas relacionadas con la reorganización celular y también activa otras protein kinasas (ver esquema abajo). Al comienzo de la anafase, el MPF es degradado. Esto promueve la actividad de las fosfatasas que van a desfosforilar todas aquellas proteínas que habían sido fosforiladas por el MPF activo. Cdc2 Condensación de la cromatina mediante fosforilación de la histona H-1. Rompimiento de la envoltura nuclear, mediante fosforilación de las láminas. Fragmentación del RE y del Golgi. Formación del huso microtúbulos con inestabilidad dinámica El siguiente esquema resume los “blancos” de la MPF activa Mitosis Muerte celular En todos los organismo multicelulares, los ritmos de mitosis y muerte celular son similares. Este equilibrio, que suele llamarse homeostasis, puede esquematizarse a continuación. Mitosis Muerte Homeostasis Este equilibrio puede alterarse cuando ocurra un trastorno por pérdida de células. Ejemplos patológicos de este caso son: el SIDA, las enfermedades de Alzheimer y Parkinson, la anemia aplasica, infarto al miocardio. En todos estos casos el esquema sería: Mi tos is Mu Este equilibrio puede alterarse por razones fisiológicas, por ejemplo, durante el desarrollo embrionario ó por razones patológicas, por ejemplo un traumatismo. Durante el desarrollo embrionario existe una enorme tendencia a producirse nuevos tejidos y a crecer organizadamente los órganos y estructuras ya formadas. Pero al mismo tiempo, otros tejidos deben desaparecer, por ejemplo, los que constituyen la cola que tuvimos durante una etapa del desarrollo. Esta es una muerte programada que se denomina apoptosis. Las células inician su propia muerte mediante la activación de un programa genético de suicidio, que se caracteriza por una autodigestión, utilizando sus lisosomas. ert e En otras patologías ocurre un trastorno por acumulación de células, como es el caso del cáncer, del lupus eritematoso, de glomerulonefritis. En estos casos el esquema tendría el siguiente aspecto: Mu s osi t i M e ert

![División Celular o Mitosis [Modo de compatibilidad]](http://s2.studylib.es/store/data/004563189_1-c93df7da2f4a17dc627669b68f110660-300x300.png)