Núcleo interfásico
Anuncio

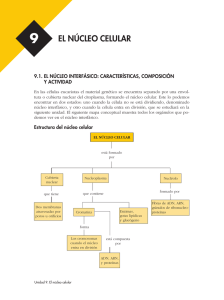
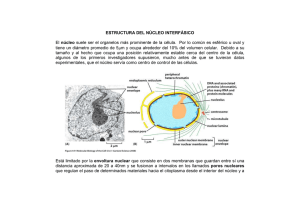
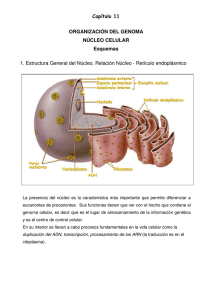
N úcleo interf ásico Núcleo interfásico Prof. Iván Rebolledo Introducción histórica Generalidades estructurales En 1876, Balbiani observó estructuras cilíndricas en el núcleo, previo a su división. En 1871, Miescher designó como nucleina al compuesto fosforado encontrado en el espermatozoide de salmón. En 1879, Flemming acuñó el término de cromatina para nombrar a la sustancia que se colorea intensamente con colorantes básicos. En 1888, Waldeyer usó el nombre de cromosoma para señalar las estructuras cilíndricas observadas años atrás. Con el uso de colorantes básicos se encontró que en los núcleos interfásicos, la cromatina puede encontrarse en estado condensada (heterocromatina) o en forma dispersa (eucromatina). La heterocromatina tiende a adherirse al lado interno de la envoltura nuclear. Recordemos que la envoltura nuclear es la estructura citológica que marca la clasificación de células procarióticas y eucarióticas. Los términos procarionte (pro, antes de y karyon, núcleo) para la vida pronuclear y eucarionte (eu, verdadero y karion, núcleo) para los organismos nucleados fueron sugeridos por Hans Ris en los años 60, siendo aceptados hasta ahora. El término interfásico proviene de un lapso de tiempo en que la célula no está en división. Creo que el esquema de abajo aclare esto. Mitosis Interfase Mitosis La envoltura nuclear es una cisterna perinuclear conformada por dos membranas, una externa y otra interna, entre las cuales se encuentra el espacio perinuclear y que se encuentra atravesada por los llamados complejos de poro nuclear. El espacio interior del núcleo suele llamarse nucleoplasma, el cual puede contener uno o varios cuerpos denominados nucléolos que poseen una alta concentración de moléculas de ARNr, proteínas y subunidades de los ribosomas. El nucleoplasma muestra también gránulos densos ricos en nucleoproteínas que contienen precursores de los ARN. El núcleo parece tener una estructura interna que organiza los materiales internos y localiza algunas funciones en sitios precisos, de aquí que se esté hablando de la matriz nuclear, como una especia de esqueleto estructural del núcleo. 111 Núcleo interfásico Envoltura nuclear(*) Las membranas llegan a fusionarse a nivel de los complejos de poro, la única vía que pueden utilizar las pequeñas moléculas polares y macromoléculas para entrar o salir del núcleo. La envoltura nuclear separa el contenido del núcleo del citoplasma, actúa como barrera que impide que moléculas no autorizadas puedan entrar o salir y así mantiene un compartimiento bioquímico muy regulado e independiente. Subyacente a la membrana interna de la envoltura nuclear se encuentra la lámina nuclear: una trama fibrosa que le da soporte a la envoltura nuclear. Esta lámina está conformada por proteínas fibrosas llamadas lámina (60-80 kd), de las cuales existen 4 variedades (A, B1, B2 y C) y que corresponden al tipo V de los filamentos intermedios. Además de proporcionar soporte a la envoltura nuclear se piensa que sirve como punto de unión a las diferentes asas del ADN. Por último, estas proteínas logran unirse a proteínas de la membrana interna de la envoltura nuclear. La envoltura nuclear está conformada por dos membranas nucleares, una lamina nuclear subyacente y complejos de poro nuclear. Las membranas son una externa y otra interna. La membrana externa se continúa con el RE de tal forma que el espacio perinuclear se comunica con el lumen del RE. Además, esta membrana externa es molecular y funcionalmente similar a la membrana del RER; aún más, existen ribosomas asociados a la superficie externa de la membrana externa. (*) no debe hablarse de membrana nuclear para referirse a la envoltura nuclear, pues ésta la conforman dos membranas. B C D E A Micrografía electrónica que muestra un sector de la envoltura nuclear. A, citosol; B, nucleoplasma; C, complejo de poro; D, ribosomas; E, membranas nucleares. 112 Núcleo interfásico Complejo de poro El complejo de poro es el único canal por el cual pueden atravesar las moléculas polares, los iones y las macromoléculas (proteínas, ARN, subunidades del ribosoma). Es una estructura muy compleja con un ∅ de 120 nm y un peso molecular estimado en 125 millones de daltons Al ME se visualiza una estructura octogonal alrededor de un gran canal central. El uso de otras técnicas han sugerido un modelo muy particular, que se muestra aquí. citoplasma Membrana externa Se ha establecido la presencia de una pequeñas hendiduras (≈ 10 nm ∅) entre los complejos octogonales que podrían representar pasos a través de los cuales podrían cruzar pequeñas moléculas no iónicas (lisozima, 14 kd; ovoalbúmina, 44 kd). En ovocitos, espermatocitos y células embrionarias se han encontrado en el citosol las llamadas laminillas anulares, que son unas formaciones membranosas apiladas con abundantes complejos de poro. Se postula que podrían representar una forma de almacenamiento de complejo de poro preformados. Importación selectiva Es evidente de las diferentes funciones que suceden dentro del núcleo, la presencia de muchas proteínas las cuales son todas sintetizadas en el citoplasma. nucleoplasma Membrana interna Puede verse la presencia de dos anillos de 8 unidades cada uno: uno citosólico y otro nuclear. Además hay proteínas filamentosas que se extienden desde estos anillos hacia cada compartimiento. Por último, se ve el tapón central, cuya naturaleza está en estudio. Como Ud. ya sabe, las proteínas sintetizadas en el citosol disponen de una secuencia señal que permite dirigirlas hacia el RER, en caso contrario se quedarán en el citosol. Como una extensión de este principio, le resumiré unos resultados experimentales de Alan Smith(*) sobre la identificación de la secuencia señal nuclear. (*) Cell 39:499-509 (1984) 113 Núcleo interfásico Utilizó como modelo una proteína llamada antígeno T del virus SV40, de 94 kd y necesaria para que el virus pueda replicarse dentro del núcleo de las células infectadas. Cuando se obtuvo una mutación de esta proteína en la lisina ubicada en la posición número 128, la proteína permanecía en el citosol y no ingresaba al núcleo. Para probar la hipótesis de una secuencia de señalización nuclear, otros investigadores fraccionaron la proteína en 3 sectores: 1-126; 127-133 y 134 hasta el C terminal. Encontraron que el primer y tercer sector podía entrar al núcleo, no así el segundo. Parecía que este sector era la señal para que la proteína entrara al núcleo. Ahora viene lo grandioso: tomaron este sector intermedio (127-133) y lo añadieron a proteínas que normalmente permanecen en el citosol. ¿Qué cree Ud. que sucedió? Pues sí señor, esas proteínas (βgalactosidasa; piruvato kinasa) pudíeron entrar al núcleo. Conclusión: esta secuencia de 7 aminoácidos representa la secuencia señal nuclear para las proteínas que deben ingresar al núcleo. Posteriores estudios determinaron que esta secuencia la integran PKKKRKV (averigue a cuáles aminoácidos corresponden). 114 La importación de la proteína a través del complejo de poro puede ser operacionalmente dividida en dos etapas. En la primera etapa la secuencia de localización nuclear es reconocida por un receptor citosólico y el complejo se une al complejo de poro. Este receptor se denomina importina y posee dos cadenas polipeptídicas: alfa, que se une a la secuencia y beta, media la unión del complejo con el complejo de poro. Secuencia señalización nuclear Importina Membrana nuclear externa Membrana nuclear interna Núcleo interfásico La segunda etapa corresponde a la translocación de la proteína a través del complejo de poro. Esta etapa requiere de energía proporcionada por hidrólisis del ATP. El complejo proteína-receptor se une a las proteínas del anillo proteico externo, cambia su configuración y desprende la subunidad beta. Con ayuda energética el resto penetra. Una proteína necesaria para la translocación es la llamada Ran que maneja ciclos de GTP-GDP. Se considera que intervenga en la disociación de la otra subunidad. Como se aprecia en la tabla, las histonas y el ADN se encuentran en una relación aproximada de 1:1. Esto sugiere una relación molecular. Las histonas son proteínas pequeñas con alto contenido de arginina y lisina. Existen 5 variedades de histonas denominadas: H1, H2A, H2B, H3 y H4. Estas proteínas son básicas y pueden unirse fuertemente al ADN que es ácido (por su contenido en PO4). Las histonas ricas en arginina son la H3 y H4; las ligeramente ricas en lisina son las H2A y H2B; y la rica en lisina es la H1. 2 moléculas de c/u H2A, H2B, H3 y H4 Cromatina El ADN es la principal componente genético de la célula, que se halla en el núcleo de las células eucarióticas, formando un complejo con proteínas denominado cromatina. La siguiente tabla resume la composición porcentual de los componentes de la cromatina en núcleos de hepatocitos. COMPONENTE % ADN 31 ARN 5 Histonas 32 Proteínas no-histonas 28 Hay más de mil proteínas no histónicas diferentes, comprometidas en una serie de actividades nucleares, principalmente la replicación y la transcripción. Como ejemplos recordamos: ADN polimerasas, ARN polimerasas, helicasa, primasa, topoisomerasa, etc. etc. 115 Núcleo interfásico La unidad estructural básica de la cromatina es el nucleosoma (mostrado en la figura anterior) descrito por Roger Kornberg (1974). Propuso que cada nucleosoma estaba formado por un sector de ADN de 146 pares de bases que daban casi dos vueltas (1.75) alrededor de un centro de 8 histonas Las histonas que integran dicho centro son dos de cada una de las siguientes histonas : H2A, H2B, H3 y H4. La histona H1 se ubica en el sitio en que el ADN entra y sale del nucleosoma. Entre los nucleosomas hay trozos de ADN de ≈60 pares de bases. Al ME esta configuración puede verse como si fuera un “collar de cuentas”. 116 Esta secuencia de nucleosomas conforma una fibra de ≈ 10 nm ∅, llamada fibra delgada. Esta forma de empaque del ADN acorta su longitud en unas seis veces. Luego, esta fibra delgada se pliega conformando una fibra gruesa de ≈30 nm ∅, en la cual se ubican unos 6 nucleosomas por vuelta de hélice. Las fibras gruesas se observan en los cromosomas metafásicos y en parte del núcleo interfásico. Eucromatina y heterocromatina Cuando las células no están en división, gran parte de la cromatina está descondensada y distribuida en todo el núcleo, suele llamarse eucromatina. Durante este período del ciclo celular, los genes son transcritos y ADN puede replicarse. Gran parte de la eucromatina se encuentra en forma de fibras gruesas de 30 nm, las cuales suelen organizarse en grandes asas. Núcleo interfásico Asa gruesa de ADN Proteína andamio La cromatina interfásico posee un 10 % de heterocromatina: un estado muy condensado similar a la cromatina de las células que están en mitosis. La heterocromatina es transcripcionalmente inactiva y contiene muchas secuencias repetidas, tales como las que se encuentran en los centrómeros y telómeros (averigüe qué son estas estructuras). Cuando las células entran en mitosis, su cromatina llega a condensarse a un nivel máximo para conformar los cromosomas, visibles al MO. Nucléolo La subestructura más prominente dentro del núcleo es el nucléolo, el cual es el sitio de transcripción y procesamiento del ARNr y del ensamble de las subunidades del ribosoma. Las células que realizan intensa síntesis de proteínas muestran un notable nucléolo. El nucléolo es la “fabrica” de las subunidades del ribosoma, las cuales abandonan el núcleo a través del complejo de poro para ir a unirse al ARNm e iniciar la síntesis de proteínas. Morfológicamente, el nucléolo comprende tres regiones concéntricas: centro fibrilar laxo, medio fibrilar denso y externo granular. Se consideran que estas regiones representan los sitios de estados progresivos de la transcripción, procesamiento y ensamble de las subunidades de los ribosomas. Así, los genes para los ARNr están localizados y se transcriben en la región central y media. El procesamiento y ensamble de las subunidades de los ribosomas, es decir, la unión de los ARNr con las proteínas, se inicia en la región media y continúa en la región externa, desde donde las subunidades están listas para ser exportadas hacia el citosol a través de los complejo de poro. 117 Núcleo interfásico Matriz nuclear Aplicación médica La matriz nuclear es el esqueleto estructural del núcleo. Varios científicos llegaron a este concepto luego de aplicar ADNasas para digerir gran parte del ADN y buffer muy salinos para extraer gran parte de las histonas y otras proteínas. Como comprenderá, después de extraer todo el contenido del núcleo, debiera variar la forma e integridad del núcleo. Pues no ocurrió así, sino que quedó una trama residual que mantuvo la forma y tamaño original del núcleo. Esto nos lleva a que la matriz nuclear es comparable al citoesqueleto que mantiene la forma celular. Además, se ha confirmado su importancia en movimientos direccionales de ciertos productos, tales como los ARNm, las subunidades ribosomales, etc. Existe una enfermedad llamada lupus eritematoso sistémico, del tipo de las enfermedades autoinmunes, que produce erupciones en la piel, artritis y alteraciones renales. Es más frecuente en mujeres que en varones, con edades promedio entre 20 y 60 años. Como es un punto de activa investigación, se empieza a considerar que estos filamentos de la matriz estarían constituidos por microfilamentos de actina. Se sugiere que la orientación de estos microfilamentos sería desde el centro del núcleo hasta sitios más periféricos. Servirían como guía de los ARN desde su lugar de transcripción y procesamiento hasta los complejo de poro. 118 Una enfermedad autoinmune sucede cuando el organismo produce anticuerpos contra sus propios constituyentes celulares normales. En el caso del Lupus, se producen anticuerpos contra componentes del núcleo: el ADN, histonas, proteínas cromosómicas y ribonucleoproteínas pequeñas nucleares (RNPsn). El diagnóstico histológico se hace precisamente detectando los anticuerpos antinucleares con sustancias fluorescentes. Debido a que se producen muchos autoanticuerpos se considera que la falla puede encontrarse a nivel de los linfocitos B, que al transformarse en células plasmáticas son las que producen los anticuerpos.