XIV Jornadas de Divulgación Técnico-Científicas 2013. Jornada Latinoamericana
Facultad de Ciencias Veterinarias – Universidad Nacional de Rosario
Distribución de los somas y fibras neuronales GnRH en el
encéfalo de bovinos pre-púberes (Bos taurus)
1
1
2
Soto, A.T. ; de La Sota, R.L. ; Zucolilli, G.O.
1
2
Laboratorio de Reproducción Animal. Laboratorio de Neurociencias.
Facultad de Ciencias Veterinarias, Universidad Nacional de La Plata
[email protected]
La cantidad y distribución de las neuronas y fibras GnRH fueron
caracterizadas por técnicas de inmunohistoquímica en cortes de tejidos
1,2,3
encefálicos de la región hipotalámica de los rumiantes adultos . La
cantidad total de somas neuronales GnRH (snGnRH) en encéfalos de
1,3
rumiantes adultos fue de 1300 a 4000 , los que se distribuyeron
principalmente en el área preóptica (POA), septal medial (MSA) y medial
preóptica (MPOA). Desde el POA, la población de snGnRH se extiende
tanto hacia craneal como caudal decreciendo progresivamente en el
número. La extensión rostral alcanza el MSA y la banda diagonal de Broca
(DBB). La proyección caudal se extiende hacia el hipotálamo mediobasal y
alcanza el núcleo arcuato (AC). Los restantes snGnRH se encuentran en
1,2,3
áreas extrahipotalámicas en forma aislada . Las fibras GnRH (fGnRH) se
encontraron principalmente en la región basal del hipotálamo desde la DBB
hasta el complejo mamilar. Las fGnRH alcanzan primariamente a la
eminencia media (EM), en la cual forman una densa red, y al órgano
vasculoso de la lámina terminal (OVLT). En el área hipotalámica anterior
(AHA) las fGnRH constituyen dos vías que se extienden caudoventralmente
hasta la EM y el tallo infundibular. Una vía recorre la pared del tercer
ventrículo (TV) hacia el infundíbulo atravesando sucesivamente los núcleos
periventricular, paraventricular y el AC. La otra vía desciende hacia la EM
1,2,3
inmediatamente dorsal al núcleo supraóptico (SON) . El objetivo del
presente trabajo fue describir la distribución del sistema de fibras y neuronas
GnRH en encéfalos de bovinos prepúberes (Bos Taurus). Se sacrificaron
bovinos prepúberes Holando Argentino (n=2), de un peso vivo de
104,5±8,5kg para lo cual fueron tranquilizados con clorhidrato de xilacina
(0,2mg/Kg/IM), anestesiados con pentotal sódico (20 mg/kg/EV) y ketamina
(15mg/Kg/EV) y finalmente sangrados a blanco. La fijación de las cabezas
se realizó con una solución de Zamboni mediante perfusión de la arteria
carótida. Se separó el tálamo e hipotálamo, se postfijaron durante 4 h a
4ºC., lavados con PBS y conservados a -20ºC en una solución de Olmos. El
material fue incluido en una solución de gelatina/glicerol y los bloques se
endurecieron por inmersión en formol-calcio durante 6 h y conservados en
una solución de sacarosa al 30%. El material se seccionó en láminas
coronales de 50 μm con un micrótomo manual de congelación. La técnica
de inmunohistoquímica (IHQ) se realizó por el método de secciones
flotantes y se incubó durante 48hs con el anticuerpo monoclonal LHR13
XIV Jornadas de Divulgación Técnico-Científicas 2013. Jornada Latinoamericana
Facultad de Ciencias Veterinarias – Universidad Nacional de Rosario
(Biosignal Research Center, Gunma University, Japón), utilizando el sistema
peroxidasa-antiperoxidasa (Vector Labs®) y diaminobenzidina como
cromógeno. Dos series de secciones se utilizaron como testigo: una sin
someterla a la IHQ y la otra sin la incubación con el anticuerpo primario. Los
cortes montados fueron observados y fotografiados mediante un
microscopio binocular conectado a una video cámara. El conteo de snGnRH
se realizó de manera manual sobre las imágenes capturadas (20x). Los
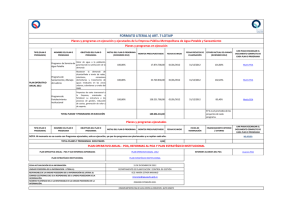
snGnRH (2083±223) se observaron principalmente en distintos núcleos y
áreas hipotalámicas que incluyeron MSA, DBB, POA, AHA y SON. Estas
regiones tuvieron el 93,03% del total de los snGnRH. El área de mayor
concentración de snGnRH fue el POA (45,2%). Entre el POA y dorsal al
SON, en un área de escasa densidad neuronal, se observó un 4% de
snGnRH. La población restante se ubicó en rostral al POA (20,6%), o bien
en caudal de esta área (29,39%). En la extensión rostral, los snGnRH se
observaron principalmente en el MSA (11,21%), en la DBB (8,52%) y en
núcleos extrahipotalámicos (0,33%). En la proyección caudal al POA, los
snGnRH se encontraron en el AHA (12,89%), SON (11,21%), entre el AHA y
el SON (1,65%), en el AC y la región periacuata (3,64%) y en regiones
extrahipotalámicas (1,3%). Las fGnRH-ir se observaron en gran cantidad en:
DBB, MSA, POA, SON, OVLT, EM, AC y en el tallo infundibular de la
hipófisis. La mayor concentración de fGnRH-ir se encontró en la EM y el
OVLT. Las fGnRH formaron una densa red en el OVLT y se distribuyeron en
toda la extensión de la EM, observándose material inmunorreactivo
rodeando a los capilares primarios del sistema porta. Tanto en el POA como
en el AHA los snGnRH aparecieron rodeados de fGnRH que finalizaron
agrupándose en dos tractos. Un fascículo caudo-ventral que cursó
acompañando al fascículo basal del prosencéfalo inmediatamente dorsal del
SON y se extendió hasta la EM y el tallo infundibular. El otro fascículo de
fGnRH recorrió el hipotálamo por las áreas periventriculares que rodean el
TV hasta alcanzar el AC donde finalizaron en una red en el interior de la EM.
Finalmente, se visualizaron escasas fGnRH en regiones extrahipotalámicas.
El número de snGnRH estimados y la distribución tanto de snGnRH y
fGnRH en el hipotálamo de los bovinos prepúberes fue similar a lo descripto
en rumiantes adultos.
BIBLIOGRAFÍA
1. Dees, W.L.; Sorensen, A.M.; Kemp, W.M.; McArthur, N.H.
Immunohistochemical localization of gonadotropin-releasing hormone (GnRH) in
the brain and infundibulum of the sheep. Cell Tissue Res. 215(1):181-91, 1981.
2. Dees, W.L. and McArthur, N.H. Immunohistochemical localization of
gonadotropin releasing hormone (GnRH) in the bovine hypothalamus and
infundibulum. Anat Rec. 200(3):281-5, 1981.
3. Hamada, T.S.; Ichikawa, M. and Mori, Y. Immunohistochemical study on
gonadotropin-releasing hormone neurons in the shiba goat brain. J Reprod Dev.
38:133 -42, 1992.
 0
0