Ciclo de Krebs Destino del piruvato. Descarboxilación oxidativa
Anuncio

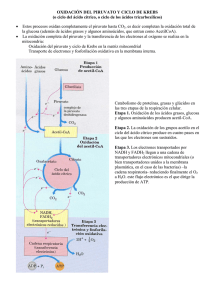
Ciclo de Krebs Destino del piruvato. Descarboxilación oxidativa. Aspectos estructurales y mecanismos de la piruvato deshidrogenasa. Relevancia del ciclo de Krebs dentro del metabolismo en general. Descripción y características de las reacciones que lo componen. Sitios de conservación de energía. Regulación metabólica, integración con la glucólisis. Efecto Pasteur. Ciclo del glioxilato. Reacciones que lo componen. Reacciones anapleróticas. Los mamíferos no pueden convertir los ácidos grasos en glucosa Del acetil-CoA no se puede ir a gluconeogénesis. Respiración es la oxidación completa del Pyr a CO2 y H2O. Respiración celular: procesos moleculares mediante los cuales las células consumen O2 y producen CO2. Etapas de la respiración celular. Etapa 1: oxidación de combustible orgánico (ácidos grasos, glucosa y algunos AA) a acetil-CoA. Etapa 2: Oxidación de grupos acetilos en el ciclo del ácido cítrico/de los TCA/de Krebs, a CO2 y 4 electrones son extraídos (NADH y FADH2). Etapa 3: NADH y FADH2 son llevados a la cadena respiratoria mitocondrial (o en bacterias a la membrana plasmática) para finalmente reducir el O2 a H2O. Este flujo electrónico lleva a la producción de ATP. Etapa 1: oxidación de ácidos grasos, glucosa y algunos AA que dan acetilCoA. Etapa 2: Oxidación de grupos acetilos en el ciclo del ácido cítrico, que incluye 4 etapas en la cual los electrones son extraídos. Etapa 3: Los electrones aportados por el NADH y FADH2 son llevados a la cadena respiratoria mitocondrial (o en bacterias a la membrana plasmática) para finalmente reducir el O2 a H2O. Este flujo electrónico lleva a la producción de ATP. Reacción global catalizada por el complejo de la Piruvato deshidrogenasa. Cinco coenzimas y 3 enzimas participan de este complejo que catalizan una decarboxilación oxidativa. Reacción irreversible 4 vit. esenciales: tiamina, riboflavina, nicotinamida y pantotenato Localización mitocondrial y prototipo de α−DH Coenzima A (CoA o CoASH) Tioéster de alto poder de transferencia de grupos acilos No se encuentra en la molécula de ADP Lipoato transportador de electrones (hidrógeno) y de acilos (acetilo) Complejo de la PDH por microfotografía crioelectrónica de riñón de bovino Más de 5 veces el tamaño de un ribosoma Imagen tridimensional del complejo de la PDH: E1: PDH (TPP) E2: dihidrolipoil transacetilasa E3: dihidrolipoil DH (FAD) 60 moléculas de E2 ordenadas en 20 trímeros para formar un dodecahedro pentagonal Sitio de unión del grupo lipoilo al dominio lipoilo de E2 Varias subunidades E3 también están unidas al centro, donde el brazo oscilante de E2 puede alcanzar el sitio activo. Dominio lipoilo que toca el sitio activo de E1 ordenada alrededor de E2 E2 complejo de la PDH E2 tiene 3 tipos de dominios unidos por polipéptidos de unión cortos: un dominio catalítico acetiltransferasa, un dominion de unión de E2 a E1 y E3 y uno o más dominios lipoilícos dependiendo de las especies. Descarboxilación oxidativa del piruvato a acetil-CoA por complejo PDH. Un ejemplo de canalización de sustratos El destino del Piruvato está marcado en rojo 1: decarboxilación (más lento, limita la veloc. reacción y ejerce especificidad de sustrato; 2: transferencia de electrones y grupo acetilo; 3: transesterificación; 4: transferencia de 2H; 5: transferencia de H+ de FADH2 a NADH Reacciones del ciclo del ácido cítrico El destino del acetato del AcetilCoA está marcado en rosa. Reacciones 1, 3 y 4 son irreversibles en la célula •4 de las 8 reacciones son oxidaciones (conservación de la E) •Precursores de otros productos Formación de citrato (condensación de Claisen) Estructura de la Citrato sintasa Forma abierta Forma cerrada con OAA unido, que provoca aparición de sitio de unión de acetil-Coa En mamíferos, el OAA se une primero, el orden de reacción es estrictamente ordenado. Centro Fe-S de la aconitasa, participa tanto en la unión al sustrato como en la catálisis Isocitrato deshidrogenasa Semejante al complejo de la PDH, E1 es diferente Fosforilación a nivel de sustrato Inhibidor competitivo de SDH FUMARASA: Áltamente estereoespecífica trans cis Citrato: molécula simétrica que reacciona asimétricamente Unión del citrato al sitio activo de aconitasa Naturaleza proquiral del citrato Productos de una vuelta del ciclo TCA incompleto en bacterias anaeróbicas (no poseen 2-OGDH) Reacciones “de relleno” que permiten reponer los intermediarios del ciclo resultando en un equilibrio dinámico. Así, las concentraciones de los intermediarios del ciclo permanecen prácticamente constantes. Fosforilación de E1 Inactiva PDH Inh. alostérico de FFK1 Efecto Pasteur Louis Pasteur fue el primero en describir el gran incremento, más de 10 veces, en el consumo de glucosa por un cultivo de levadura cuando pasaba de condiciones aeróbicas a anaeróbicas. Este “efecto Pasteur” se produce sin un cambio significativo de la conc. de ATP o de la mayoría de los centenares de intermediarios metabólicos y productos derivados de la glucosa. Plantas, ciertos invertebrados y algunos microorganismos (incluidas E.coli y levaduras) convierten acetato en glúcidos por el ciclo del glioxilato 2 acetil-CoA + NAD+ + 2 H2O succinato + 2 CoA + NADH + H+ Semillas de pepino en germinación Evita las 2 decarboxilaciones y por eso da síntesis neta de succinato, OAA y otros intermediarios Asp Regulación coordinada de ambos ciclos por IDH