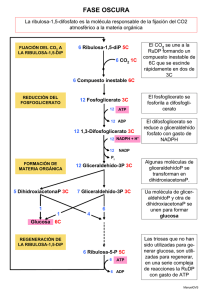
Asimilación del carbono en diferentes grupos microbianos.
Anuncio

Metabolismo del Carbono Microorganismos Clasificación Fuente de energía Oxido-reducción Quimiótrofas Luz Fotótrofas Compuestos orgánicos Quimioheterótrofos Fuente de carbono Compuestos orgánicos Fotoheterótrofos Fuente de carbono Compuestos orgánicos Quimioheterótrofos CO2 Compuestos inorgánicos Quimiolitótrofos Fuente de carbono Compuestos Orgánicos Mixótrofos CO2 Litoautótrofos CO2 Fotoautótrofos Crecimiento en compuestos de C1 Muchos procariotes pueden crecer en compuestos de un carbono como única fuente de carbono. Los compuestos más comunes son: •Dióxido de carbono (CO2) •Metano (CH4) •Metanol (CH3OH) •Metilamina (CH3NH2) Methylococcus capsulatus Pocos procariotes anaerobios (metanógenos) pueden utilizar el CO2 como fuente de carbono y también como aceptor de electrones, reduciendo a metano y produciendo ATP. Autótrofos Organismos que emplean el CO2 como fuente de carbono. Algas Cianobacterias Bacterias púrpura Bacterias halófilas Bacterias quimiolitótrofas Crecimiento Autotrófico Ribulosa difosfato carboxilasa (RubisCO). Convierte la ribulosa 1,5-difosfato en ácido 3-fosfoglicérico. http://www.efn.uncor.edu/dep/biologia/intrbiol/fotosint.htm RubisCO Se encuentra presente en: •Bacterias púrpura •Cianobacterias •Algas •Plantas verdes •Muchas bacterias Quimiolitótrofas •Algunas arqueobacterias halófilas e hipertermófilas Protein Data Bank •Carboxisomas Thiobacillus neapolitanus Prochlorococcus marinus Synechococcus sp. Ciclo de Calvin http://www.msu.edu/~smithe44/campbell_10.16 Ciclo de Calvin •Carboxilación. La enzima rubisco cataliza la combinación de la ribulosa difosfato con el CO2, formándose un compuesto intermedio e inestable (de 6 átomos de carbono), que se descompone en dos moléculas de fosfoglicerato (con 3 átomos). •Reducción. Mediante la energía que suministra el ATP y el poder reductor del NADPH, el fosfoglicerato se transforma en gliceraldehído 3-fosfato. •Recuperación. De cada 6 moléculas de gliceraldehído 3-fosfato, 5 se transforman en 3 moléculas de ribulosa difosfato (con consumo de ATP) y la otra se considera el rendimiento neto del ciclo. http://recursos.cnice.mec.es/biologi a/bachillerato/segundo/biologia/ud 04/02_04_04_02_023.html Ciclo de Calvin Emplea el ATP y el poder reductor para fijar el CO2. •Emplea una ferredoxina reducida proveniente de la fotosíntesis. •Del oxaloacetato se asimilan 3 moléculas de CO2 y se produce Piruvato que va a formar parte del material celular. •Hay gasto de ATP. Ciclo de Krebs reverso Bacterias que realizan el Ciclo de Krebs reverso •Bacterias verdes del azufre (Chlorobium). Bacterias Gram negativas que fueron aisladas en Nueva Zelanda y se desarrollan en aguas termales con presencia de sulfuros de hidrógeno. C. tepidum, es el modelo del grupo por su facilidad de cultivo, es una bacteria que realiza la fotosíntesis anoxigénica, utiliza H2, Sº y H2S como donadores de electrones además deposita en el medio gránulos de Sº. Bacterias que realizan el Ciclo de Krebs reverso •Desulfobacter sp Bacteria sulfato reductora, Gram negativa anaerobia estricta, móvil por la presencia de un flagelo polar, utiliza el acetato como donador de electrones y lo oxida a CO2 por el ciclo de Krebs. •Hydrogenobacter sp Bacterias que oxidan el hidrógeno para obtener energía, acidófilas y termófilas. Vía del Hidroxipropionato •Acetil-CoA/propionil-CoA carboxilasa es la enzima clave de la vía. •Un acetil-CoA y dos moléculas de CO2 son reductivamente convertidas vía 3-hidroxipropionato a succinil-CoA. Este intermediario es reducido a 4-hidroxibutirato y convertido en dos moléculas de acetil-CoA por la 4-hidroxibutiril-CoA dehidratasa. Bacterias que realizan la vía del Hidroxipropionato •Bacterias verdes no del azufre (Chloroflexus). Bacteria filamentosa, anoxigénica, no todas las especies pueden emplear los sulfuros como donador de electrones. Chloroflexus generalmente es fotoheterótrofo y utiliza H2 y sulfuros como donadores de electrones. Cuando crece autotróficamente fija CO2 por la vía del hidroxipropionato. Chloroflexus tiene un crecimiento óptimo entre 50 y 60ºC por lo que se encuentra en la naturaleza en aguas termales, lagos, ríos y sedimentos, y en ambientes hipersalinos. Bacterias que realizan la vía del Hidroxipropionato •Arqueas hipertermófilas Acidianus sp Metallosphaera prunae Hipertermófila, acidófila, que posee capa cristalina S. Fue encontrada en una mina de uranio. Se observa la capa cristalina S y una partícla viral en la parte superior derecha. Mixótrofo o autótrofo, crece anaeróbicamente en CO2 y Sº y aeróbicamente en medios orgánicos. Sulfolobus sp Hipertermófila, acidófila, aerobia, heterótrofa que puede emplear el Sº como fuente de energía. Vía Acetil CoA reductiva •Un CO2 es capturado por un cofactor especial (tetrahidrofolato: T ) y reducido a grupo metil. •La otra el CO2 es reducido a un grupocarbonil (C=O) por la monóxido deshidrogenasa. •El grupo carbonil se une con el grupo metil para formar Acetil CoA por el complejo enzimático Acetil CoA sintasa. •La vía requiere H2 como donador de electrones. http://lecturer.ukdw.ac.id/dhira/Metabolis m/CarbonAssim.html Vía Acetil CoA reductiva •Bacterias acetogénicas (Clostridium thermoaceticum, Acetobacterium woodii). •Metanogénicas (Methanobacterium thermoautotrophicum). •Autótrofas sulfato reductoras (Desulfobacterium autotrophicum). Microorganismos Metilótroficos •Bacterias aerobias que pueden crecer en compuestos que no contienen puentes de C-C, diferentes al CO2. •Puede fabricar sus propios compuestos orgánicos empleando como fuente de carbono dichas pequeñas moléculas, y además puede oxidarlas para obtener energía metabólica. Methylobacterium oryzae Microorganismos Metilótroficos •Monóxido de carbono CO •Metano (CH4) •Metanol (CH3OH) •Formaldehído (HCHO) •Formato (HCOOH) •Metilamina (CH3NH2) •Trimetilamina [(CH3)3N]* •Dimetil éter [(CH3)2O]* •Dimetil carbonato (CH3OCOOCH3)* •Dimetil sulfóxido [(CH3)2SO]* Methylamine-enriched community of Lake Washington sediment featuring Methylotenera cells. *Compuestos con múltiples carbonos que no contienen enlaces C-C. Puede utilizar compuestos como al amonio, etileno, clorometano, bromometano e hidrocarburos de más carbonos como etano y propano como sustratos oxidables pero no utilizables para crecer. Clasificación de las bacterias metilótroficas Metilótrofos Obligados No pueden crecer en compuestos con multicarbonos y pueden crecer en metanol o metilamina Facultativos No pueden crecer en compuestos con multicarbonos y pueden crecer en metanol o metano Pueden crecer en cualquier compuesto de C1 Metanótrofos Bacterias metilótroficas Obligadas No metanótrofas Facultativas No pueden crecer en compuestos con multicarbonos y pueden crecer en metanol o metilamina, no en metano. Pueden crecer en cualquier compuesto de C1 Bacterias aeróbicas Gram negativas: Methylophilus y Metanobacillus. Bacterias Gram positivas y Gram Negativas: Bacillus, Acetobacter, Mycobacterium, Arthrobacter, Hyphomicrobium, Methylobacterium y Nocardia. Bacterias metilótroficas •Obligadas Metanótrofas. No pueden crecer en compuestos con multicarbonos y pueden crecer en metanol o metano. Grupo Organismo Formas de reposo Ciclo del ácido cítrico& Asimilación de carbono Fijación de N2 I Methylomonas Quistes Incompleta Ribulosa monofosfato No I Methylomicrobium No tiene Incompleta Ribulosa monofosfato No I Methylobacter Quistes Incompleta Ribulosa monofosfato No I Methylococcus * Quistes Incompleta Ribulosa monofosfato Si II Methylosinus Exospora Completa Serina Si II Methylocystis Exospora Completa Serina Si II Methylocellae Exospora Completa Serina Si * Contiene las enzimas del ciclo de Calvin. &Organismos con un ciclo del ácido cítrico incompleto carecen de la enzima alfa-cetoglutarato deshidrogenasa y no pueden oxidar acetato a CO2. Metanótrofos •Poseen un sistema de enzimas específico, la metanomonooxigenasa. •Forman un complejo sistema de membranas internas cuando crecen donde se lleva a cabo la oxidación del metano. Contienen alta concentración de esteroles. Tipo I, grupos de vesículas con forma de disco distribuidas por todo el organismo. Tipo II, membranas organizadas en la periferia de la célula. Methylomonas sp Methylosinus sp Oxidación del metano •Metanomonooxigenasa. •Metanol deshidrogenasa. •Formaldehido deshidrogenasa. •Formato deshidrogenasa. Vía de la ribulosa monofosfato Metanótrofos Tipo I Vía de la Serina Metanótrofos Tipo II Bacterias carboxidotróficas CO + H2O → CO2 + 2H+ + 2e•Enzima monóxido de carbono deshidrogenasa. •CO como donador de electrones para la síntesis de ATP. •Fija el CO2 generado mediante el Ciclo de Calvin. Nature Reviews Microbiology 5, 107-118 (February 2007) Catabolismo de compuestos orgánicos Enzimas extracelulares Las exoenzimas son enzimas producidas por microorganismos y excretadas al medio ambiente. Son ampliamente utilizadas en diversos factores industriales. Estas enzimas son producidas naturalmente por los microorganismos para su sobrevivencia. Lipasa Celulasa b-galactosidasa Degradación de polímeros de C Sustancias Composición Fuentes Enzimas catabólicas Celulosa Polímero de glucosa (b-1,4-) Plantas (hojas, tallos) Celulasa (b-1,4glucanasas) Almidón Polímero de glucosa (a-1,4-) Plantas (hojas, semillas) Amilasa Glucógeno Polímero de glucosa (a-1,4- y b-1,6-) Animales (músculo) Amilasa, fosforilasa Laminarina Polímero de glucosa (b-1,3-) Algas marina b-1,3-Glucanasa (laminarinasa) Paramilón Polímero de glucosa (b-1,3-) Algas b-1,3-Glucanasa Polímero de galactosa y ácido galacturónico. Algas rojas Agarasa Agar Degradación de polímeros de C Sustancias Composición Fuentes Enzimas catabólicas Quitina Polímero de Nacetilglucosamina (b-1,4) Hongos (paredes celulares), insectos (exoesqueleto) Quitinasa Pectina Polímero de ácido galacturónico Pectina (hojas, semillas) Pectinasa (poligalacturonasa ) Polímero de glucosa Cápulas o capas mucoides Dextranasa Heteropolímero de xilosa y otros azúcares (b-1,4- y otros laterares a-1-2 o a-1,3) Plantas Xilanasa Dextrano Xilano Utilización de lactosa •La lactosa puede ser utilizada por diferentes microorganismos por dos mecanismos: •La inducción bajo el control del operón lac para una permeasa y una b-galactosidasa. •Sistema PTS con una enzima II específica para lactosa. Operón lac. Con lactosa (alolactosa) •En presencia de lactosa extracelular, bajos niveles de b-galactosidasa que se encuentra en la célula convierten lactosa en alolactosa (grupo galactósido en la posición 4 en lugar de la posición 6). •La alolactosa se une a la proteína represora y la desacopla de su lugar en el operador. •La ARN polimerasa transcribe los genes para el trasporte y utilización de la lactosa. Operón lac. Efecto de la glucosa •AMPc •CRP (Receptora de AMPc o •En un medio sin glucosa pero con lactosa los niveles de AMPc proteína activadora de catabolitos) son altos. La proteína CRP tiene sitios de unión para el ADN y para el AMPc. El complejo AMPc-CAP se une a una zona cercana al promotor lac, y favorece la unión de la ARN polimerasa al promotor, aumentando la transcripción hasta 50 veces. Se ha demostrado que el AMPc y la CAP contribuyen a la iniciación de la transcripción, ayudando al reconocimiento del sitio de iniciación por la ARN polimerasa. Simporte de lactosa •Lactose can be transported into E. coli against a concentration gradient using galactoside permease, one of the proteins encoded by the lac operon. This protein uses a proton-motive force to pump lactose into the cell. •The proton gradient is http://employees.csbsju.edu/hjakubowski/clas ses/ch331/signaltrans/olsignalenergy.html created by an electron transport complex in the membrane which is inhibited by cyanide, reminiscent of the cytochrome C oxidase complex in oxphos. Crecimiento Heterotrófico ATP Glucosa ATP NADPH Glucosa 6P Fructosa 6P ATP Gluconato 6P Pentosas-P ATP Gluconato CO2, NADPH Fructosa 1,6 biP Glucólisis Pentosa Gliceraldehído P ATP, NADH Fosfoenolpiruvato ATP Piruvato NADPH, CO2 Acetil CoA Entner Doudoroff Las vías del metabolismo central: Glucólisis (EmbdenMeyerhof-Parnas EMP), de las Pentosas fosfato y EntnerDoudoroff proveen de precursores metabólicos para otras vías, son las vías para el metabolismo de carbohidratos y ácidos carboxílicos. Las tres vías convierten por diferentes rutas la glucosa en gliceraldehído, que es oxidado por la misma reacción para formar piruvato. Glucólisis •La vía de la glucólisis (Embden-MeyerhofParnas EMP), se encuentra en todos los eucariotes y en muchas especies de bacterias. •En la primera etapa hay gasto de 2 moléculas de ATP por molécula de glucosa. •En la segunda parte se forman 4 moléculas de ATP por fosforilación a nivel de sustrato y 2 de NADH. Ciclo de las pentosas fosfato Importancia: •Produce los precursores de la ribosa, desoxirribosa en los ácidos nucleicos y provee de eritrosa fosfato como precursor de aminoácidos aromáticos. •Se produce en está vía, NADPH, la mayor fuente de electrones de la biosíntesis y varias de sus reacciones las comparte con el ciclo de Calvin. Vía EntnerDoudoroff •Muchos tipos de bacterias no poseen la enzima fosfofructocinasa-1 y no pueden convertir la glucosa-6P a Fructosa 1,6-difosfato. •Una vía alterna es EntnerDuodoroff, en la cual la glucosa 6P es convertida a piruvato y Gliceraldehído-3-fosfato por la desidratación poco usual del 6-fosfogluconato para formar 2-ceto-3-desoxi-6-fosfogluconato (KDPG). Oxidación/Fermentación •En la prueba de Oxidación- fermentación se emplea el medio de Hugh and Leifson en condiciones aerobias y anaerobias para determinar el metabolismo oxidativo o fermentativo de un hidrato de carbono. •La fermentación requiere de una fosforilación inicial, utiliza principalmente la vía de la Glucólisis (Embden-Meyerhof-Parnas EMP), aunque también puede utilizar las vías de las Pentosas fosfato o Entner-Doudoroff (ED) en combinación con EMP. •La vía metabólica de Embden-Meyerhof-Parnas también se denomina glucolítica o anaerobia o fermentativa. La vía metabólica de Entner-Doudoroff también se denomina aerobia. Oxidación/Fermentación Glucosa Glucosa Deshidrogenasa Ácido glucónico Gluconato dehidratasa •La oxidación de la glucosa produce ácido pirúvico por la vía de Entner-Doudoroff (ED) o por una derivación. Ambas se han encontrado en procariotes. •La vía de ED ocurre en condiciones aerobias. •La vía de derivación no requiere de fosforilación inicial. Ácido2-ceto-glucónico Cinasa Ácido 2-ceto-6-fosfoglucónico (KDPG) Reacciones ED 2 Moles de ácido pirúvico • En la derivación se forma el intermediario KDPG que se incorpora a la vía de ED para formar piruvato. Ciclo de Krebs • Aunque la vía de los ATC se asocia a bacterias aeróbicas, no todas ellas poseen el ciclo completo. •También se presenta en muchas bacterias heterotróficas que crecen anaeróbicamente y en ciertos anaerobios que oxidan completamente los compuestos orgánicos a CO2. Fermentación Es definida como una ruta en la cual el NADH es reoxidado por metabolitos producidos por la vía. Las reacciones redox ocurren principalmente en el citoplasma y el ATP es producido por fosforilación a nivel de sustrato. La fermentación sucede en condiciones anaeróbicas. •Células eucariotes. Levaduras, fermentación alcohólica Células musculares y eritrocitos, fermentación láctica •Células procariotes. 1. Láctica 2. Alcohólica 3. Butírica 4. Ácido mixta 5. Propionica 6. Homoacética Fosforilación a nivel de sustrato El sustrato orgánico (donador de e-) pasa por una ruta catabólica (ej., la glucólisis), y uno de los intermediarios de esa ruta es oxidado por un coenzima, se origina un intermediario no fosforilado con una gran energía de hidrólisis. El intermediario experimenta enseguida una sustitución con un fosfato, para dar la correspondiente forma acilfosfato (enlace de alta energía). Este acil-fosfato dona su fosfato de alta energía al ADP, que pasa a ATP. Compuesto orgánico (reducido) Intermediario A NAD+ Pi NADH Intermediario B P Intermediario C P Acarreador electrónico ADP ATP Intermediario D (oxidado) NADH NAD+ Producto final (reducido) Fermentación •Se realiza en condiciones anaerobias. •Emplea como aceptor de electrones a una molécula orgánica. •Utiliza el NADH producido en la glucólisis. •Se conserva la energía de la fosforilación a nivel de sustrato (FNS). Acido láctico +2H Ácido pirúvico + ácido pirúvico -CO2 ATP Streptococcus Lactobacillus Staphylococcus Acido a-acetoláctico -CO2 H2 ADP +CO2 Acetoina +2H CO2 Acido oxaloacético +2H Acido málico 2,3-butanediol Shigella Ácido fórmico -H2O +2H Klebsiella Enterobacter Bacillus Acetil-S-CoA -CoA-SH +4H -CoA-SH Acido succinico -CO2 Acido propionico Etanol CO2 H2 E. coli Salmonella Bifidobacterium Propionibacterium Acido Acético +Acetil S-CoA -CoA-SH Clostridium Acetoacetil-S-CoA Acetona -CO2 +4H Butiril-S-CoA -CoA-SH +2H Isopropanol -CoA-SH Ácido Butirico +4H Butanol Efecto Pasteur Cuando microorganismos facultativos se están cultivando anaeróbicamente y se exponen al oxígeno, hay una inhibición del consumo de glucosa, esto es conocido como efecto Pasteur y refleja el incremento de rendimiento energético obtenido por el metabolismo respiratorio de la glucosa comparado con el obtenido por la fermentación de la glucosa. Efecto Crabtree Se refiere a la inhibición del consumo de oxígeno producido por aumento en la concentración de glucosa. Producción de energía Reacción de Stickland Reacción de Stickland Clostridium, emplea aminoácidos como aceptores y donadores de hidrógeno para obtener energía.