Metabolismo de hidratos de carbono II. Gluconeogénesis
Anuncio

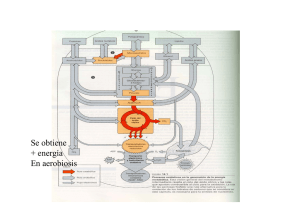
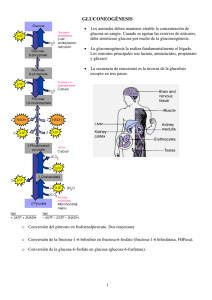
Metabolismo de hidratos de carbono II. Gluconeogénesis. Reacciones propias. Mecanismos de la piruvato carboxilasa y fosfoenolpiruvato carboxiquinasa. Requerimientos energéticos. Relación con el ciclo de Cori y el ciclo de la alanina. Ciclos fútiles: reciclado de sustrato. Ciclo oxidativo de las pentosas fosfato. Síntesis de glúcidos a partir de precursores sencillos Variaciones de energía libre de reacciones glucolíticas en los eritrocitos Rutas opuestas de la glucólisis y de la gluconeogénesis en el hígado de rata. Las reacciones de la glucólisis se muestran en el lado izq en rojo. Síntesis de fosfoenol piruvato a partir de piruvato Papel de la biotina en la reacción de la piruvato carboxilasa. El cofactor biotina está unido de forma covalente a la enzima a través de un enlace amida con el grupo ε-amino de un residuo Lys formando un biotinil-enzima. Residuo de Lys Rol de la biotina en la reacción catalizada por la Piruvato carboxilasa Primera enzima reguladora en la gluconeogénesis que requiere acetil-CoA como efector positivo Rutas alternativas desde el piruvato al fosfoenolpiruvato. La importancia relativa de las dos rutas depende de la disponibilidad de lactato o piruvato y las necesidades citosólicas de NADH para la gluconeogénesis. La ruta de la derecha predomina cuando el lactato es el precursor, debido a que el NADH citosólico se genera en la reacción de la LDH y no se ha de transportar fuera de la mitocondria. Reacciones secuenciales de la gluconeogénesis empezando a partir del piruvato ΔG´o 0,9 kJ/mol ΔG -25 kJ/mol ΔG´o -16,3 kJ/mol ΔG´o -13,8 kJ/mol ΔG´o 0,9 kJ/mol ΔG -16 kJ/mol Aminoácidos glucogénicos agrupados según el sitio de entrada Regulada por transcripción por factores que exigen mayor producción de Gluc (baja Gluc sanguínea, señalización por glucagón) PFK-1 + FBPasa-1: ATP + H2O ---- ADP + Pi + calor Si operaran de manera simultánea, disiparía calor: CICLO FÚTIL. Sin embargo, pueden ser ventajosos para controlar rutas, con lo que el término CICLO DEL SUSTRATO es más adecuado. Dos destinos alternativos para el piruvato. El piruvato se puede convertir en glucosa y glucógeno via gluconeogénesis o ser oxidado a acetil-CoA para la producción de enegía. La primera enzima de cada ruta está regulada alostéricamente; el acetil-CoA producido por oxidación de los ácidos grasos o por el complejo de la PDH, estimula la PC e inhibe la PDH. Regulación de FBPase-1 y PFK-1 Continúa con el rol de la F2,6BP Rol opuesto de la F2,6BP en la regulación de gluconeogénesis y glucólisis Demostraron la conversión del glucógeno en lactato en tejidos, el paso del lactato de la sangre al hígado y, dentro del hígado, la reconversión del lactato en glucógeno, ruta que se conoce como CICLO DE CORI Cooperación metabólica entre el músculo esquelético y el hígado Ciclo de Cori Ciclo de la glucosaalanina. La Ala Pir y Lac se incorporan en Gluc, que es devuelta al músculo y el amoníaco se convierte en urea actúa como transportador de amoníaco y del esqueleto carbonado del piruvato desde el músculo esquelético al hígado. El amoníaco se excreta y el piruvato se utiliza para producir glucosa, que vuelve al hígado. Esquema general de la ruta de las pentosas fosfato Papel del NADPH y del glutatión en la protección de las células frente a derivados de oxígeno altamente reactivos. El GSH protege a las células destruyendo el peróxido de hidrógeno y los radicales hidroxilo libres. La regeneración del GSH a partir de GSSG necesita NADPH producido por la reacción de la glucosa 6-fosfato deshidrogenasa. Reacciones oxidativas de la ruta de las pentosas fosfato. Los productos finales son la D-ribosa 5-fosfato, CO2 y NADPH Reacciones no oxidativas de la ruta de las pentosas fosfato (a) Estas reacciones convierten pentosas fosfato en hexosas fosfato, lo que permite que continúen las reacciones oxidativas. Las enzimas transcetolasa y transaldolasa son específicas de esta ruta; las otras enzimas también actúan en las rutas glucolíticas o gluconeogénicas.(b) Cada reacción es reversible, aquí se muestran en la dirección de la oxidación continua de la glucosa 6-fosfato. La reacción general catalizada por la transcetolasa es la transferencia de un grupo de dos carbonos, transportando de forma temporal sobre TPP ligado a la enzima, desde un dador cetosa a un aceptor aldosa. Conversión de dos pentosas fosfato en una triosa fosfato y un azúcar fosfato de siete carbonos, la sedoheptulosa 7-fosfato Reacción catalizada por la transaldolasa Segunda reacción catalizada por la transcetolasa Intermediarios carbanión estabilizados por interacciones covalentes con la transcetolasa y la transaldolasa Papel del NADPH en la regulación del reparto de la glucosa 6-fosfato entre glucólisis y ruta de las pentosas fosfato. Cuando se forma NADPH más rápidamente de lo que se utiliza para la biosíntesis y reducción del GSSG, aumeta la conc. de NADPH con lo que inhibe la primera enzima de la ruta de las pentosas fosfato. El resultado es que hay más glucosa 6-fosfato disponible para la glucólisis.