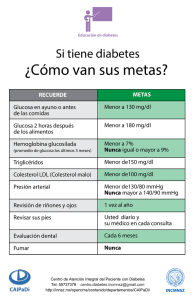
Promotor del gen ABCA1 en relación a la diabetes
Anuncio

“ESTUDIO DEL PROMOTOR DEL GEN ABCA1 EN RELACION A DIABETES TIPO 2 EN POBLACION GENERAL ESPAÑOLA” M.U. “APROXIMACIONES MOLECULARES EN CIENCIAS DE LA SALUD” Trabajo Fin de Máster Ana Paulina Arévalo J. Director: Dr. Felipe Javier Chaves Martínez, Fundación Investigación Clínico de Valencia – INCLIVA Tutor: Dr. José Enrique O’Connor, Departamento de Bioquímica y Biología Molecular, Universidad de Valencia 2012 – 2013 ÍNDICE Resumen/Abstract 2 1. Introducción 4 Proteínas ABC Subfamilia ABCA Proteína ABCA1 Funciones Regulación ABCA1 y diabetes tipo 2 4 5 5 6 8 9 2. Objetivos 11 3. Materiales y métodos 12 Muestras Polimorfismos – ABCA1 Genotipado Análisis estadístico 12 12 12 16 4. Resultados 17 5. Discusión 27 6. Conclusiones 31 7. Anexos 32 8. Bibliografía 34 1 RESUMEN ABCA1 desempeña un papel clave en el mantenimiento de las concentraciones celulares de colesterol. La regulación de su expresión es vital, ya que tanto el déficit como el exceso de colesterol podrían ser perjudiciales para la célula. Variantes genéticas en ABCA1 se han asociado con diversas alteraciones metabólicas, entre ellas alteraciones de los niveles de lípidos en plasma, aumento de niveles de glucosa e, incluso el riesgo a desarrollar diabetes. El posible rol que ABCA1 desempeña en el desarrollo de diabetes tipo 2 podría deberse a alteraciones en los niveles de colesterol en las células β pancreáticas. El principal objetivo de este trabajo es identificar polimorfismos presentes en el promotor del gen ABCA1 y evaluar su asociación a diabetes tipo 2, o a parámetros bioquímicos y clínicos relacionados con alteraciones lipídicas en población española. Se han identificado ocho polimorfismos en la región comprendida entre -500 y +250pb en 5’ y 3’, respectivamente, del inicio del exón 1. El polimorfismo rs2740483 mostró relación estadística con niveles de colesterol total y LDL. Los individuos G/G presentaron concentraciones mayores de colesterol y LDL respecto de los G/C-C/C. Al analizar por sexo, se observaron asociaciones diferentes en cada grupo. Los genotipos C/T-T/T y G/C-C/C de los polimorfismos rs2422493 y rs2246293 respectivamente, mostraron asociación a niveles altos de colesterol total y LDL, y a valores bajos de HDL en hombres. Mientras que en mujeres se asociaron a niveles bajos de LDL. Los alelos T y C de los polimorfismos rs2246298 y rs1800976, respectivamente, se relacionaron con niveles altos de colesterol total y LDL en hombres. En mujeres el alelo T del polimorfismo rs1800977 se asoció a valores bajos de colesterol total y LDL. Al analizar la asociación de haplotipos encontramos que el haplotipo TCCCGCC (rs2422493, rs2246293, rs2246298, rs1800976, rs2740483, rs1800977, rs111292742) se asociaba con la disminución del riesgo de padecer diabetes tipo 2 en la población. Y el haplotipo CGCGGCC se asoció a niveles altos de glucosa, colesterol total y LDL. Se identificaron nuevas variantes de secuencia en la región promotora del gen, aunque aparentemente no tienen efecto sobre el fenotipo, se debería validar su asociación y función en otros estudios. Los resultados obtenidos en este trabajo muestran que existe una posible asociación de estos SNPs con parámetros bioquímicos de importancia clínica. Es necesario analizar otras poblaciones y realizar estudios funcionales a fin de validar los resultados. 2 ABSTRACT ABCA1 plays a key role in maintaining cellular cholesterol concentrations. The regulation of its expression is vital, since both the deficit and excess cholesterol could be harmful to the cell. Genetic variants in this gene have been associated with various metabolic disorders, including lipid levels in plasma, glucose levels and even the risk of developing diabetes. The possible role that ABCA1 plays in the development of type 2 diabetes could be due to alterations in cholesterol levels in β cells. The main objective of this study was to identify single nucleotide polymorphisms in the ABCA1 gene promoter and evaluate its association with type 2 diabetes, and clinical or biochemical parameters related to lipid abnormalities in Spanish population. Eight polymorphisms have been identified in the region between -500 and +250 bp of 5 'and 3', respectively, in the start of exon 1. The rs2740483 polymorphism was associated to the levels of total cholesterol and LDL in general population, G/G individuals presented higher levels of cholesterol and LDL compared to G/C-C/C. The analysis according to sex showed differences in each group. The genotypes C/T-T/T and G/C-C/C of the rs2422493 and rs2246293 polymorphisms, respectively, showed association with high levels of total cholesterol and LDL, also with low levels of HDL in men. While in women were associated with low levels of LDL. T and C alleles of the rs2246298 and rs1800976 polymorphisms, respectively, were associated with higher levels of total cholesterol and LDL in men. In women, the T allele of rs1800977 polymorphism was associated with low levels of total cholesterol and LDL. In the haplotype analysis we found that TCCCGCC haplotype(rs2422493, rs2246293, rs2246298, rs1800976, rs2740483, rs1800977, rs111292742)was associated with decreased risk of type 2 diabetes in the population. And CGCGGCC haplotype was associated with high levels of glucose, total cholesterol and LDL. New sequence variations were identified in the promoter region of this gene, and although they have no apparent effect on the phenotype, their association and function should be investigated in further studies. The results obtained in this study show the possible effect of these SNPs in clinical and biochemical parameters. It is necessary to analyze other populations and conduct functional studies in order to validate these results. 3 1. INTRODUCCIÓN Proteínas ABC Las proteínas ABC (ATP-binding cassette) constituyen una de las más grandes súperfamilias de transportadores conocida. Los sustratos transportados por las proteínas ABC incluyen lípidos, péptidos, aminoácidos, carbohidratos, vitaminas, iones, glucurónidos, conjugados de glutatión, xenobióticos, entre otros. En los eucariotas, los transportadores ABC se encuentran en la membrana plasmática, y en las membranas de compartimentos intracelulares, tales como el aparato de Golgi, endosomas, cuerpos multivesiculares, retículo endoplásmico, peroxisomas, y en las mitocondrias. En humanos se han identificado 48 miembros de esta familia de transportadores. En base a su relación estructural se subdividen en 7 familias, designadas ABC A – G (Kaminski et al. 2006; Dean, et al. 2001). Gráfico 1.1. Genes ABC humanos, ubicación cromosómica y función. (Dean et al. 2001) 4 Subfamilia ABCA La subfamilia ABCA está formada por 12 proteínas transportadoras designadas ABCA1 ABCA13. La comparación de la secuencia de aminoácidos de los 12 miembros revela homologías que van desde 28% (ABCA8/ABCA12) al 72% (ABCA8/ABCA9). El análisis filogenético sugiere que los miembros de esta subfamilia han evolucionado a partir de un gen ancestro primordial y se dispersaron posteriormente a través del genoma (Kaminski et al. 2006). Los transportadores ABCA se dividen en dos subgrupos, basados en el análisis filogenético y la estructura intrónica. El primer grupo incluye siete genes dispersos en seis cromosomas diferentes (ABCA1, ABCA2, ABCA3, ABCA4, ABCA7, ABCA12 y ABCA13), mientras que el segundo grupo contiene cinco genes (ABCA5, ABCA6, ABCA8, ABCA9yABCA10) dispuestos en grupo en un solo cromosoma. ABCA11 no se considera, ya que esta fue una asignación errónea que se dio inicialmente a un pseudogen (Stefková et al. 2004, Kaminski et al. 2006). Gráfico 1.2. Árbol filogenético y dispersión cromosómica de genes ABC humanos (Kaminski et al. 2006). Proteína ABCA1 ABCA1 es el integrante que define la subclase A de proteínas ABC. Se expresa en numerosos órganos humanos con altos niveles de expresión en la placenta, hígado, pulmón, glándulas suprarrenales, los tejidos fetales y macrófagos. Es una proteína de 2261 aminoácidos, con un peso molecular de 220kDa (Kaminski et al. 2006, Santamarina-Fojo et 5 al. 2000). El gen que codifica esta proteína se ubica en el cromosoma 9 (9q31.1) y contiene 50 exones (NCBI Reference Sequence: NG_007981.1). ABCA1 es una proteína integral de membrana, localizada principalmente en la membrana plasmática, pero también se encuentra a nivel de compartimentos intracelulares (Nagao et al. 2011). Los transportadores ABC muestran una organización global común, que comprende dos mitades de estructura similar (Gráfico 1.3). Cada mitad tiene un dominio transmembrana que contiene seis hélices y un dominio de unión a ATP, también conocido como dominio de unión a nucleótido. Este último contiene dos motivos peptídicos conservados conocidos como Walker A y Walker B, separados entre sí por aproximadamente 90 a 110 aminoácidos presentes en muchas proteínas que utilizan ATP, y un dominio Walker C característico de los transportadores ABC. ABCA1 tiene su extremo NH2 terminal orientado hacia el citosol y posee dos bucles extracelulares que están altamente glicosilados y unidos por uno o más enlaces de cisteína (Oram & Heinecke 2005, Singaraja et al.2003, Kaminski et al. 2006). Gráfico 1.3.Estructura de ABCA1 (Bungert et al. 2001) Funciones Una de las principales funciones de ABCA1, es la de transportar colesterol y fosfolípidos celulares hacia las apolipoproteínas, como a apoA-I para la formación de HDL. Esto provee a la célula de un método eficaz para la liberación del exceso de colesterol (Liu y Tang 2011). Existen varios modelos para explicar el mecanismo de formación de HDL mediada por ABCA1 (Nagao et al. 2011). Uno de ellos, propuesto por varios estudios, sugiere que ABCA1 genera dominios lipídicos, en la cara externa de la bicapa lipídica, incluso en ausencia de apolipoproteínas. Estos dominios sirven para aliviar la tensión que generan los fosfolípidos densamente empaquetados en la cara interna de la membrana. La generación de estas superficies curvadas ricas en lípidos en la membrana favorece la interacción con las apolipoproteínas. La interacción de ABCA1 con las apolipoproteínas activa vías de señalización, como la JAK2, que a su vez favorecen la unión de apolipoproteinas a ABCA1. Estas interacciones facilitan el contacto de apoA-I con los dominios lipídicos, la solubilización de éstos y su salida de la célula (Vedhachalam et al. 2007, Liu y Tang 2011). 6 Gráfico 1.4. Formación de HDL mediada por ABCA1. (i) Traslocación de fosfolípidos y colesterol a través de la membrana por ABCA1 (ii) apoA-I recepta los lípidos transportados (iii) las curvaturas de la membrana creadas por ABCA1 interactúan con apoA-I facilitando la captación de fosfolípidos y colesterol (Nagao et al. 2011) Otra de las funciones atribuidas a ABCA1, es su participación en la organización de la membrana celular. En la membrana plasmática varios subdominios como balsas lipídicas, suponen una organización y reorganización dinámica, involucrada en varias funciones celulares como en la transducción de señales, y al parecer ABCA1 participa en la formación de estos dominios. Estudios en ratón muestran que macrófagos deficientes de ABCA1 expresan un incremento de balsas lipídicas en la superficie celular, esto de manera independiente a la presencia de apoA-I (Nagao et al. 2011). Algunos estudios sugieren que ABCA1 también tiene propiedades anti-inflamatorias. Estudios en ratones que carecen de ABCA1en todos los tejidos o de forma selectiva en macrófagos, tienen una reacción más fuerte en respuesta a llipopolisacárido (LPS) como estímulo inflamatorio, postulando que ABCA1 en los macrófagos cumple también una función antiinflamatoria. Análisis realizados en cultivos celulares indican que la actividad anti-inflamatoria de ABCA1 es secundaria a su capacidad para modular el contenido de colesterol de las balsas lipídicas de la membrana. Macrófagos de ratón que carecen de ABCA1 aumentan el contenido de colesterol en las balsas lipídicas, y aumenta el estímulo del LPS, a través de su receptor Toll-like receptor 4 (TLR4), aumentando la producción de citoquinas inflamatorias. La incubación de monocitos humanos con apoA-I reduce la capacidad de los estímulos inflamatorios, efecto que se suprime en células que carecen de ABCA1. Todas estas respuestas parecen depender del contenido de colesterol de las balsas lipídicas de la membrana plasmática. Otros estudios indican que la interacción de apoA-I/ABCA1activa el factor de transcripción STAT3. La vía de STAT3 tiene un efecto antiinflamatorio conocido en macrófagos. La interacción de apoA-I con ABCA1 en macrófagos suprime además la expresión de citoquinas inflamatorias, sin embargo, la activación de STAT3 no se requiere para el eflujo de lípidos mediado por ABCA1 (Liu y Tang 2011). 7 Regulación La regulación de la expresión de ABCA1, ocurre a distintos niveles. A nivel transcripcional, ABCA1 parece responder principalmente a metabolitos lipídicos. En macrófagos y fibroblastos la expresión de ABCA1 está regulada por los receptores nucleares LXRα y β (Liver X receptor α y β), cuyos ligandos son principalmente hidroxicolesterol y epoxicolesterol. LXRβ es de expresión ubicua y se expresa de forma estable incluso en ausencia de colesterol, mientras que LXRα se expresa a nivel de hígado, tejido adiposo, glándulas suprarrenales, pulmones, riñones y células de origen mieloide, y su expresión está altamente regulada por la presencia de colesterol. En estado basal, LXRβ y RXR (retinoid X receptor) forman un heterodímero, que se une a elementos de respuesta (LXREs, liver X response elements) en la región promotora de sus genes diana. Cuando el colesterol se acumula en las células, aumentan las concentraciones de oxiesteroles, la unión de estas moléculas a LXRβ/RXR favorece la transcripción de ABCA1, y también de LXRα. Se ha identificado también una región promotora que responde a SREBP-2 (Sterol Regulatory Element Binding Proteins), esta se ubica en el primer intrón de ABCA1, y se ha indicado su participación en la regulación de la transcripción de ABCA1 a nivel hepático (Nagao et al.2011). Activadores de PPARs (peroxisome proliferator-activated receptors) también se han visto relacionados en la regulación de la transcripción de ABCA1 en diferentes células. PPARγ estimula el eflujo de colesterol desde las células mediante la activación de la transcripción de LXRα, el cual induce la transcripción de ABCA1. El receptor nuclear TR (thyroid hormone receptor) se presenta como un supresor de la transcripción de ABCA1, al formar un complejo con RXR evitando la formación del complejo LXR/RXR y la unión a sus elementos de respuesta en el gen (Oram & Heinecke 2005). Mir-33, también regula la expresión de ABCA1. Este es un micro RNA ubicado en el gen SREBF2. Uno de los principales reguladores de la síntesis de colesterol pero también se ha definido como un inhibidor de la expresión de ABCA1 (Rayner et al.2010). Debido al papel primordial del colesterol como componente esencial de las células y principalmente su función estabilizadora de membranas, su eliminación o acumulo excesivos podrían causar la muerte celular. De ahí la importancia de regular el eflujo de colesterol y por ende la actividad de ABCA1. ABCA1 tiene una vida media de 1-2 horas. Se proponen varias vías para la degradación de este transportador, así: ABCA1 de la superficie celular es endocitado y reciclado a la superficie celular, o entregado a los lisosomas mediante endosomas tempranos y tardíos para su degradación. Otra vía es la degradación mediada por calpainas, proteasas que degradan a ABCA1 a nivel de la membrana plasmática e intracelularmente, especialmente en ausencia de la unión con apoA-I. Finalmente, otra vía propuesta para la degradación de ABCA1 es la mediada por proteasoma (Nagao et al. 2011). 8 Una forma de regular la degradación de este transportador es mediante la fosoforilación en diferentes residuos. Varios estudios revelan que la unión de apoA-I evita la fosforilación de los residuos de treonina-1286 y treonina-1305 de la secuencia PEST de ABCA1, consecuentemente disminuye la degradación de la proteína mediada por calpainas (Liu y Tang 2011). ABCA1 y diabetes tipo 2 ABCA1 se ha relacionado principalmente con variaciones en los niveles de lípidos en sangre, sobre todo con los niveles de HDL. Defectos genéticos en el gen ABCA1causan alteraciones como la enfermedad de Tangier e hipoalfalipoproteinemia familiar. Los individuos con alteraciones en este transportador presentan niveles muy bajos de HDL, acumulación de colesterol en tejidos periféricos, e incremento de aterosclerosis (Singaraja et al.2003, Liu y Tang 2011). Algunos estudios indican además, que ABCA1 podría estar asociado al desarrollo de diabetes tipo 2, principalmente por el papel que desempeña en la homeostasis de la célula β pancreática. La diabetes tipo 2 se caracteriza por la incapacidad de la célula β del páncreas de secretar insulina suficiente para satisfacerlas demandas metabólicas asociadas con resistencia a la insulina y la obesidad. La disfunción de las célulasβes un elemento clave en la patogénesis de la diabetes. La disminución en la secreción de insulina mediada por glucosa y un gradual deterioro en la función de la célula β son características en los pacientes con diabetes tipo 2. Se plantean varias propuestas para explicar la disminución en la secreción de insulina por los islotes pancreáticos, algunas de éstas son: el deterioro de la célula β, la desensibilización a la estimulación por glucosa, y la lipotoxicidad. Esta última hace referencia al efecto deletéreo de los niveles altos de lípidos sobre la célula β, considerando que en éstas existe un pequeño margen de colesterol en las membranas celulares para que esta funcione correctamente (Brunham et al. 2008, 2010). Estudios en ratones a los cuales se inactiva selectivamente ABCA1 en las células β muestran intolerancia grave a la glucosa y una secreción deficiente de insulina. Además, los islotes aislados de estos animales, presentan reducción en la secreción de insulina mediada por glucosa, y una gran acumulación de colesterol, sugiriendo que ABCA1 es esencial para la salida de colesterol en la célula β y su correcto funcionamiento (Brunham et al. 2007). El colesterol forma parte de los gránulos de secreción como de las membranas, y juega un papel importante en la formación y tráfico de gránulos. Los islotes pancreáticos deficientes de ABCA1 presentan una disminución en su capacidad secretora, sin embargo tienen un alto contenido de insulina intracelular, sugiriendo que en estos existe una alteración en el proceso de exocitosis más que en la sensibilidad a la glucosa (Brunham et al. 2008, 2010). LXRβ es uno de los reguladores de la expresión de ABCA1. En ratones LXRβ-/- se reduce los niveles de RNAm de ABCA1, y esto se asocia con acumulación de lípidos en los islotes, intolerancia a la glucosa y reducción en la secreción de insulina. Mientras que su activación 9 se traduce en la inducción de la expresión de ABCA1 y un incremento en la secreción de insulina (Gerin et al. 2005, Brunham et al. 2008). Por otro lado, las lipoproteínas aterogénicas, tales como LDL y VLDL inducen la muerte por apoptosis de islotes en cultivo. Este efecto es bloqueado por las HDL, proponiendo que la carga de colesterol en los islotes pancreáticos puede ser citotóxica. Las LDL oxidadas reducen la secreción de insulina mediada por glucosa en la línea celular HIT-T15 (células β de hámster). Estas lipoproteínas oxidadas reducen también la insulina y el RNAm de la preproinsulina, además de aumentar la apoptosis. Aparentemente, el colesterol afecta directamente la función de la célula β y la estimulación de la secreción de insulina mediada por glucosa (Hao et al. 2007, Brunham et al. 2008). En un estudio se mostró que la expresión de ABCA1 y sus funciones se ven disminuidas en pacientes con diabetes tipo 2, la expresión y función de ABCA1 estuvo además asociada a niveles de HDL (Patel et al. 2011). Al parecer, alteraciones en los lípidos plasmáticos y en el metabolismo del colesterol a nivel de los islotes pancreáticos puede contribuir a la patogénesis de la diabetes tipo 2. El transportador ABCA1, que regula la salida de colesterol, desempeña un papel importante en el normal funcionamiento de la célula β, y su alteración resulta en una sobrecarga de colesterol a nivel celular y una secreción defectuosa de insulina. Dada la importancia que tiene ABCA1 en el metabolismo celular, su relevante participación en el metabolismo del colesterol y su posible efecto sobre la funcionalidad de las células beta pancreáticas y sobre el desarrollo de diabetes tipo 2 en el presente trabajo se ha propuesto estudiar el promotor de este gen mediante secuenciación de última generación, con la finalidad de identificar polimorfismos que pudieran estar relacionados con el desarrollo de diabetes tipo 2 o alteraciones en los parámetros bioquímicos relacionados al metabolismo lipídico. 10 2. OBJETIVOS General - Identificar polimorfismos en el promotor del gen ABCA1 y evaluar su asociación a diabetes tipo 2 y/o alteraciones lipídicas en población española. Específicos - Secuenciar la región del promotor del gen ABCA1 en individuos de población general española. - Identificar los polimorfismos presentes en la región de estudio. - Analizar posibles asociaciones entre los SNPs encontrados y parámetros bioquímicos y clínicos relacionados. 11 3. MATERIALES Y MÉTODOS Muestras Este trabajo es un estudio transversal, observacional en población general. Las muestras forman parte del banco de ADN y base de datos del “Estudio Hortega” (Mena et al. 2003). Forman parte de este trabajo 1430 individuos con un rango de edades entre 21 – 96 años de edad, 709 mujeres y 721 hombres de la población de Valladolid – España. Las muestras de ADN se diluyeron hasta obtener una concentración de trabajo ≈25ng/ul. Polimorfismos – ABCA1 En este caso el interés es identificar SNPs en el promotor del gen ABCA1 y estimar su asociación a variables clínicas y/o bioquímicas de interés. La secuencia de referencia fue tomada del NCBI. Número de acceso NG 007981, versión NG_007981.1 G: 189458805. La región a amplificar consideró 500 y 250pb en 5’ y 3’, respectivamente, del inicio del exón 1 según la secuencia de referencia. Genotipado Para la secuenciación y genotipado se utilizó la tecnología aportada por el sistema 454 GS Junior de Roche. Este sistema utiliza secuenciación de última generación y permite preparar, amplificar y secuenciar una librería de fragmentos de ADN de forma masiva y paralela a partir de fragmentos de ADN amplificados por PCR. Preparación de amplicones (PCR1) Para la amplificación se dividió a la región de interés en tres amplicones. Los oligonucleótidos utilizados para la amplificación específica se diseñaron utilizando el programa Primer 3 (versión 0.4.0) Tabla 3.1. Las condiciones de la PCR se ajustaron a las características de las enzimas y oligonucleótidos utilizados. Tabla 3.1. Oligonucleótidos ABCA1 Nombre Secuencia ABCA1_Prom_a_5’ AGGCAGTAGGTCGCCTATCA ABCA1_Prom_a_3’ GCGCCGCAGACTCTCTAGT ABCA1_Prom_b_5’ TTGGCTGCCGGGAACGTG ABCA1_Prom_b_3’ GCAGAGGTTACTATCGGTCA ABCA1_E1_5’ TTTCTGCTGAGTGACTGAACTACA ABCA1_E1_3’ GACAAGCCTTCCGGAGAA Tamaño del producto (pb) Tm (ºC) 308 62 322 68 318 62 Los oligonucleótidos utilizados en este paso llevan una secuencia adicional “cola”, que será reconocida por los oligonucleótidos utilizados en el siguiente paso. 12 El producto de esta amplificación se verificó mediante electroforesis capilar utilizando el equipo QIAxcel, utilizando una dilución de 1:5 con agua. Adición de MIDS (PCR2) Una vez obtenidos los amplicones es momento de añadir a cada muestra una secuencia específica que servirá para identificar a cada una de ellas como un individuo particular. La figura siguiente representa los oligonucleótidos utilizados y sus partes. Figura 3.1. Estructura de los oligonucleótidos usados en la PCR2. En el extremo 3’ una secuencia complementaria a la “cola” de los oligonucleótidos usados en la primera PCR, y un MID o secuencia identificadora, específica para cada individuo. La porción 5' contiene una secuencia específica para hibridar con la “bead” o bola de captura de ADN y para el reconocimiento por los oligonucleótidos de amplificación emPCR y el oligonucleótido de secuenciación. Esta secuencia es diferente para el oligonucleótido “forward” y “reverse” (secuencias A y B, Figura 3.2.), lo que permite secuenciar en los dos sentidos. Además, esta parte 5' contiene la “key” secuencia de nucleótidos "TCAG" que se utiliza para la secuenciación del amplicón (454LifeSciences Corp. 2012). En este punto se ha obtenido ya la “librería” de ADN a secuenciar (Figura 3.2.). Figura 3.2. Estructura de los amplicones Purificación y cuantificación El producto de la PCR2 es purificado mediante perlas magnéticas Agencourt AMPurexp, posteriormente se toman 3ul de cada producto de PCR para unirlos en un “pool” de muestras. 13 Al “pool” de muestras obtenido en el paso previo se purifica una vez más mediante la separación en gel de agarosa al 2%, utilizando el equipo y cassettes PippinPrep de SageScience. La cuantificación de la librería se realizó utilizando el kit Quant-iT™ PicoGreen® dsADNssay Kit (Invitrogen™). Los datos obtenidos en este paso se utilizan para diluir la muestra a una concentración de 2x106 moléculas/ul, determinada por el protocolo de secuenciación. Amplificación de la librería (emPCR) En este paso se realiza una amplificación clonal de los fragmentos de ADN en microesferas generadas a partir de una emulsión (Figura 3.3.). El éxito de este paso consiste en que en cada microesfera haya una sola “capture bead” con una única molécula de ADN unida, esta unión se da a través de los oligonucleótidos añadidos en la PCR2, forward (secuencia A) o reverse (secuencia B). Para esto es necesario colocar la concentración adecuada de muestra. La cantidad de muestra a añadir se calcula con la siguiente fórmula: 6 Volumen de muestra (µl) = (moléculas deseadas por bead) (5x10 beads) 6 2x10 moléculas/ul Finalizada la emPCR se realizan pasos sucesivos para enriquecer la librería, es decir eliminar aquellas “capture beads” sin muestra. Para esto se desnaturaliza el ADN de doble cadena unido a las bolas, utilizando NaOH, dejando así moléculas de cadena simple unidas a las bolas. Luego ocurre una hibridación con los oligonucleótidos de enriquecimiento que contienen biotina. Posteriormente estos oligonucleótidos se unirán a las “enrrichtment beads” bolas magnéticas de enriquecimiento que contienen estreptavidina mediante unión biotina-estreptavidina (Figura 3.3.). Finalmente, enriquecida la librería se destruye la unión biotina-estreptavidina, se recolecta la muestra y se añaden los oligonucleótidos de secuenciación para continuar con el proceso. Figura 3.3. Esquema de emPCR y enriquecimiento de la librería de ADN Secuenciación El sistema 454 GS Junior de Roche utiliza pirosecuenciación, tecnología basada en la secuenciación por síntesis, acoplando la síntesis de DNA a una reacción 14 quimioluminiscente, permitiendo la determinación de secuencias en tiempo real. Esta técnica se resume en los siguientes pasos: - ADN de simple cadena amplificado por PCR hibrida con el oligonucleótido de secuenciación y se incuba con las enzimas ADN polimerasa, ATP sulfurilasa, luciferasa y apirasa, más los sustratos adenosina-5’-fosfosulfato (APS) y luciferina. - La adición de uno de los cuatro dNTPs inicia el segundo paso. La polimerasa cataliza la incorporación del dNTP al molde si es complementario, la cantidad de PPi liberado es equivalente a la cantidad de dNTP incorporado. - La ATP-sulfurilasa convierte el PPi en ATP en presencia de APS. El ATP generado permite la conversión de la luciferina en oxiluciferina por acción de la luciferasa, generando luz visible en cantidades proporcionales a la cantidad de ATP presente. La luz emitida es detectada por una cámara CCD y puede ser analizada por el equipo. - Para continuar con la secuenciación, es esencial la degradación y eliminación mediante lavado de aquellos dNTPs que no han sido incorporados, de esto se encarga la enzima apirasa. - Se añaden nuevos dNTPs para iniciar un nuevo ciclo. Todo esto ocurre dentro de los pocillos de una placa de fibra óptica, que permite el paso de luz, y que está diseñada de tal forma para que en cada pocillo se incorpore una “bead” (454LifeSciences Corp. 2012). Figura 3.4.Reacción de secuenciación llevada a cabo en la placa multipocillo Análisis El análisis de la secuenciación se realizó utilizando el programa Amplicon Variant Analyzer (AVA) de Roche, cuyo principal objetivo es detectar y cuantificar variantes conocidas o no conocidas en mezclas complejas de ADN. Los resultados iniciales de este análisis indican el número de lecturas por amplicon en cada dirección y por muestra. La cuantificación de las variantes es expresada en porcentaje de presencia del nucleótido alternativo, tomando como referencia la secuencia indicada por el usuario. 15 A partir de estos datos se realiza una primera selección de muestras, se dejan fuera del estudio aquellas con menos de 10 lecturas. Posteriormente se identifican las variantes indicadas por el AVA y que ya están reportadas en las bases de datos. Aquellas variantes reportadas por el AVA y que no están descritas deben ser revisadas para definirlas como tal, para esto se consideró el número de lecturas, cercanía a un homopolimero y porcentaje. Con estos datos iniciales se realizó la asignación de genotipos, considerando como homocigotos para el alelo de referencia a los valores ≤5% y homocigoto para el alternativo a los ≥95%, y heterocigotos a los valores entre 30-70%. Se realizó la secuenciación capilar de varias muestras para verificar que la asignación de genotipos era correcta. Para evaluar si las variantes nuevas indicadas por el AVA tienen algún efecto sobre los factores de transcripción que se unen al promotor del gen ABCA1 se utilizó el programa MatInspector (Genomatix Software GmbH 2013). Análisis estadístico Se utilizó el paquete estadístico SPSS v15.0 para determinar los estadísticos descriptivos de la población. Los datos se presentan como media y desviación estándar para las variables cuantitativas y porcentajes para las categóricas. El cálculo de frecuencias alélicas y genotípicas, el equilibrio de Hardy-Weinberg, desequilibrio de ligamiento, estimación de frecuencia de haplotipos, y análisis de asociación entre SNP individuales o de haplotipos con las variables de respuesta se realizó utilizando el software libre SNPStats v.0.95 (http://bioinfo.iconcologia.net/SNPstats). Para definir el modelo genético bajo el cual se presentarán los resultados se tomó en cuenta los valores medios de cada genotipo y se agruparon los heterocigotos con el homocigoto correspondiente según su similitud, en consecuencia se escogió el modelo dominante o recesivo, si los heterocigotos se unían al homocigoto minoritario o mayoritario, respectivamente. Sexo, edad, índice de masa corporal, horas de ayuno se utilizaron como variables de ajuste en los análisis de asociación. Valores de p < 0.05 se consideraron estadísticamente significativos. Al aplicar la corrección de Bonferroni, valores de p < 0.002 se consideran significativos. Tomando en cuenta que el análisis incluye siete polimorfismos y tres variables principales de comparación: diabetes, hipercolesterolemia, hipoalfalipoproteinemia. Existen otras variables en el estudio, pero al estar en relación directa con estas no se han considerado para la corrección. 16 4. RESULTADOS La población estudiada incluye 1430 individuos, de los cuales el 50,4% (721) son hombres. La Tabla 4.1. muestra la media y desviación estándar (DE) de las principales características de la población. La prevalencia de obesidad en la población total fue del 24,7%; se encontró un 42,2% de hipertensión, 17,2% de hipercolesterolemia, 21% de hipoalfalipoproteinemia, 7,4% de diabetes y un 33,2% de síndrome metabólico. Tabla 4.1. Características clínicas y bioquímicas de la población Variables Mujer (709) Hombre (721) Media DE Media DE p Edad (años) 54,33 19,32 54,28 19,28 NS IMC (kg/m2) 26,12 4,87 26,83 3,41 <0,001 Cintura (cm) 83,26 ICC 0,81 0,08 0,93 PAS (mmHg) 128,93 23,75 132,58 19,02 <0,001 PAD (mmHg) 78,81 10,72 79,58 10,73 NS CT (mg/dL) 202,03 36,86 198,86 37,56 NS LDL (mg/dL) 115,08 32,75 112,16 33,79 NS HDL (mg/dL) 57,54 13,66 45,94 11,58 <0,001 Triglicéridos (mg/dL) 147,01 79,46 203,78 113,05 <0,001 Glucemia (mg/dL) 91,23 18,69 93,98 Obesidad (%) 26,3 23,3 NS Sobrepeso (%) 29,6 46,3 <0,001 35 26 <0,001 Hipertensión (%) 40,3 44,1 NS Diabetes tipo 2 (%) 5,8 9 <0,05 Hipercolesterolemia (%) 14,8 19,5 NS Hipertrigliceridemia (%) 52 50,6 <0,05 9 33 <0,001 31,3 35,1 NS Obesidad central (%) Hipoalfalipoproteinemia (%) Síndrome metabólico (%) 12,66 95,93 10,02 <0,001 0,07 22,4 <0,001 <0,05 p=p-valor de la diferencia entre géneros, U de Mann Whitney para variables continuas/estadístico Fisher para categóricas. NS= no significativo. IMC= índice de masa corporal. ICC= índice cintura cadera. PAS= presión arterial sistólica. PAD= presión arterial diastólica. CT= colesterol total. Los criterios que se tomaron en cuenta para la determinación de los parámetros clínicos fueron: Obesidad: IMC≥30. Sobrepeso: IMC≥25 y <30. Obesidad central: cintura≥88cm en mujeres y ≥102cm en hombres. Hipertensión: ≥140/90mmHg. Diabetes tipo 2: glucemia≥126mg/dL. Hipercolesterolemia: colesterol_LDL ≥160mg/dL. Hipertrigliceridemia: TG ≥200mg/dL. Hipoalfalipoproteinemia: HDL<40mg/dL. En estos casos además de los valores indicados se consideró el diagnóstico previo por un médico y la existencia de un tratamiento. Síndrome metabólico: tener tres o más de los factores de riesgo que incluyen, obesidad central, presión arterial elevada, niveles bajos de HDL, niveles altos de TG y glucemia 17 elevada o padecimiento de diabetes. Además del diagnóstico previo (OMS, NCEP-ATPIII 2002, EGIR 1999). En la región analizada del gen se han descrito varios polimorfismos de un solo nucleótido, sin embargo en este trabajo se hará referencia únicamente a aquellos encontrados en la población estudiada. En la Tabla 4.2. se detallan las frecuencias genotípicas y alélicas de los polimorfismos analizados, todos estos polimorfismos se mantuvieron en equilibrio de Hardy-Weinberg. El polimorfismo rs56064613 se encontró solamente en una persona, por lo que no se incluye en los análisis de asociación. Tabla 4.2. Frecuencias genotípicas y alélicas SNP rs2422493 rs2246293 rs2246298 rs1800976 rs56064613 rs2740483 rs1800977 rs111292742 FRECUENCIA GENOTÍPICA C/C 0.24 C/T 0.53 T/T 0.23 G/G 0.24 G/C 0.53 C/C 0.23 C/C 0.68 C/T 0.28 T/T 0.04 G/G 0.31 G/C 0.52 C/C 0.17 C/C 0.999 C/T 0.001 T/T --G/G 0.47 G/C 0.44 C/C 0.09 C/C 0.41 C/T 0.45 T/T 0.14 C/C 0.98 C/G 0.019 G/G 0.001 FRECUENCIA ALÉLICA C 0.51 T 0.49 G C 0.51 0.49 C T 0.82 0.18 G C 0.57 0.43 C T 0.999 0.001 G C 0.69 0.31 C T 0.64 0.36 C G 0.99 0.01 A continuación se presentan los resultados del análisis de asociación entre los polimorfismos y las variables clínicas y bioquímicas en la población general (Tabla 4.3. y 4.4.). 18 Para realizar el análisis de asociación con el perfil lipídico (colesterol total, LDL, HDL y triglicéridos) se eliminó aquellos individuos que se encontraban en tratamiento con fármacos hipolipemiantes, para valorar la asociación con glucemia se eliminó a aquellos que tomaban medicamentos hipoglucemiantes, para evitar que los resultados de asociación se vean alterados por el efecto farmacológico. Tabla 4.3. Análisis de polimorfismos como factor de riesgo frente parámetros clínicos en población general. SNP rs2422493 Genotipo N 1.00 1.00 C/T-T/T 823 0.81 (0.46-1.44) 0.87 (0.60-1.27) 1.10 (0.81-1.50) 1.09 (0.74-1.59) 0.47 0.48 0.54 G/G 260 1.00 1.00 1.00 0.66 1.00 G/C-C/C 822 0.80 (0.45-1.43) 0.94 (0.65-1.37) 1.07 (0.79-1.45) 1.09 (0.75-1.60) 0.46 0.75 0.66 0.65 C/C-C/T 791 1.00 1.00 1.00 1.00 T/T 35 1.69 (0.94-3.05) 1.63 (0.73-3.60) 1.80 (0.82-3.96) 1.04 (0.42-2.56) 0.085+ 0.25 0.14 0.94 G/G 237 1.00 1.00 1.00 1.00 G/C-C/C 539 0.82 (0.43-1.54) 0.78 (0.52-1.19) 1.01 (0.73-1.41) 0.92 (0.61-1.38) 0.53 0.25 0.94 0.68 G/G 599 1.00 1.00 1.00 1.00 C/G-C/C 670 1.08 (0.69-1.70) 0.88 (0.65-1.20) 0.94 (0.74-1.19) 0.92 (0.69-1.24) 0.73 0.42 0.62 0.6 P rs1800977 lipoproteinemia 1.00 P rs2740483 Hipoalfa- 1.00 P rs1800976 HTG 259 P rs2246298 tipo 2 HCT C/C P rs2246293 Diabetes C/C-C/T 535 1.00 1.00 1.00 1.00 T/T 150 1.26 (0.63-2.50) 0.89 (0.56-1.44) 1.11 (0.76-1.62) 1.08 (0.69-1.68) 0.52 0.64 0.58 0.75 1179 1.00 1.00 1.00 1.00 29 1.26 (0.63-2.50) 1.38 (0.55-3.48) 0.54 (0.23-1.27) 0.78 (0.28-2.19) 0.52 0.51 0.15 0.63 P rs111292742 C/C C/G-G/G P Valores representan OR (IC 95%). Ajustado por sexo, edad, IMC. HCT= hipercolesterolemia. HTG= hipertrigliceridemia. += modelo dominante C/C vs C/T-T/T Estatus (individuos que lo presentan) (individuos que no lo presentan)= Diabetes tipo 2: (106) (1324), HCT: (245) (1182), HTG: (683) (648), Hipoalfalipoproteinemia: (298) (1130) Como indican los datos mostrados en la Tabla 4.3. no se encontró asociación entre ningún polimorfismo individual y el riesgo de padecer diabetes tipo 2, hipercolesterolemia, hipertrigliceridemia e hipoalfalipoproteinemiaen la población general. Al realizar el análisis por sexo tampoco se encontró que estos polimorfismos actuaran como factores de riesgo para estas alteraciones (datos no mostrados). Cabe indicar que se revisaron también las asociaciones entre polimorfismos y obesidad, sobrepeso, obesidad central y las variables cuantitativas asociadas. Con la finalidad de descartar alguna asociación a estos factores, por la influencia que tienen sobre el riesgo de padecer diabetes. 19 Tabla 4.4. Asociación entre polimorfismos y variables cuantitativas en población general. SNP Genotipo rs2422493 C/C C/T-T/T P rs2246293 G/G G/C-C/C P rs2246298 C/C-C/T T/T P rs1800976 G/G G/C-C/C P rs2740483 G/G C/G-C/C P rs1800977 C/C-C/T T/T P rs111292742 C/C C/G-G/G P Glucosa (mg/dL) 90.9±1.3 (234) 90.15±0.56 (736) 0.41 91.03±1.29 (234) 90.29±0.57 (735) 0.41 90.68±0.66 (722) 97.66±3.89 (32) 0.07 91.11±1.33 (219) 90.9±0.8 (485) 0.88 90.71±0.76 (543) 90.2±0.66 (598) 0.43 89.93±0.51 (849) 91.99±1.65 (136) 0.21 90.05±0.48 (1056) 93.78±4.42 (27) 0.19 CT (mg/dL) 198.39±2.46 (224) 201.28±1.4 (710) 0.46 198.98±2.45 (225) 202.05±1.39 (710) 0.4 200.94±1.43 (697) 211.42±8.07 (31) 0.31 196.72±2.6 (208) 202.53±1.68 (476) 0.06 202.27±1.55 (526) 198.21±1.57 (578) 0.036 201.89±1.32 (818) 195.95±3.08 (129) 0.089 200.87±1.16 (1023) 201.04±8.19 (23) 0.85 Triglicéridos (mg/dL) 167.53±6.69 (224) 176.7±4.12 (710) 0.16 169.06±6.72 (225) 177.29±4.1 (710) 0.21 173.02±3.92 (697) 190.42±20.33 (31) 0.59 167.87±6.71 (208) 178.65±4.9 (476) 0.12 170.84±4.21 (526) 173.13±4.4 (578) 0.64 175.13±3.82 (818) 173.22±8.13 (129) 0.43 174.93±3.32 (1023) 143±13.77 (23) 0.14 LDL (mg/dL) 112.51±2.29 (222) 114.06±1.22 (709) 0.73 112.87±2.29 (223) 114.68±1.23 (709) 0.63 113.9±1.28 (695) 124.2±7.04 (31) 0.21 111.77±2.36 (207) 114.14±1.52 (475) 0.51 115.95±1.41 (526) 111.77±1.39 (576) 0.02 114.59±1.18 (815) 111.44±2.82 (129) 0.35 113.71±1.04 (1020) 122.25±6.75 (23) 0.15 HDL (mg/dL) 52.9±0.98 (223) 51.92±0.51 (709) 0.13 52.83±0.98 (224) 51.94±0.51 (709) 0.18 52.66±0.54 (696) 49.14±2.19 (31) 0.2 52.01±1 (208) 52.71±0.65 (475) 0.56 52.15±0.62 (526) 52.08±0.59 (577) 0.72 52.45±0.5 (816) 49.87±1.08 (129) 0.16 52.32±0.45 (1021) 50.19±2.18 (23) 0.35 Los valores representan la media ± desviación estándar (número). p=p-value. Ajustado por sexo, edad, IMC, horas de ayuno. Los análisis realizados muestran que el genotipo G/G del polimorfismo rs2740483 se asocia a niveles más altos de colesterol total y de LDL (p= 0.036 y p= 0.02, respectivamente) Tabla 4.4. Al realizar el análisis por sexo, se mantiene esta tendencia de asociación a los niveles de colesterol total y LDL, sin embargo solo alcanza significancia estadística con los niveles de LDL en hombres (p=0.039) (Tabla 4.5.). 20 Tabla 4.5. Asociación entre polimorfismos y variables cuantitativas por sexo. SNP Genotipo P rs2422493 C/C T/T-C/T P rs2246293 G/G C/C-G/C P rs2246298 C/C-C/T T/T P rs1800976 G/G G/C-C/C P rs2740483 G/G C/G-C/C P rs1800977 C/C-C/T T/T P rs111292742 C/C C/G-G/G P Mujeres Glucosa (mg/dL) 89.54±1.39 (114) 89.22±0.81 (359) CT (mg/dL) 203.54±3.4 (107) 199.1±1.88 (355) LDL (mg/dL) 117.98±3.2 (107) 112.27±1.62 (355) HDL (mg/dL) 58.28±1.38 (107) 58.3±0.7 (355) Hombres Glucosa (mg/dL) 92.25±2.17 (120) 91.03±0.78 (377) CT (mg/dL) 193.68±3.49 (117) 203.47±2.07 (355) LDL (mg/dL) 107.42±3.22 (115) 115.85±1.82 (354) HDL (mg/dL) 47.95±1.24 (116) 45.51±0.57 (354) 0.94 0.13 0.046 0.82 0.29 0.031 0.032 0.045 89.75±1.38 (113) 89.1±0.81 (360) 204.8±3.44 (107) 199.48±1.87 (355) 119.37±3.22 (107) 112.45±1.63 (355) 58.16±1.37 (107) 58.42±0.71 (355) 92.23±2.15 (121) 91.42±0.81 (375) 193.69±3.43 (118) 204.62±2.06 (355) 106.87±3.17 (116) 116.91±1.84 (354) 47.95±1.23 (117) 45.45±0.56 (354) 0.88 0.1 0.024 1,00 0.37 0.014 0.009 0.035 88.65±0.81 (368) 94±3.39 (16) 202.63±1.93 (360) 205.13±11.45 (15) 115.19±1.73 (360) 122.24±10.15 (15) 58.89±0.71 (360) 54.03±3.41 (15) 92.78±1.04 (354) 101.31±7.02 (16) 199.14±2.13 (337) 217.31±11.52 (16) 112.51±1.91 (335) 126.04±10.07 (16) 45.99±0.64 (336) 44.55±2.35 (16) 0.23 0.98 0.51 0.21 0.21 0.008+ 0.016+ 0.49 88.66±1.27 (115) 88.64±1.04 (241) 201.74±3.48 (111) 202.25±2.3 (243) 116.73±3.1 (111) 114.05±2.09 (243) 57.19±1.36 (111) 59.11±0.86 (243) 93.81±2.41 8104) 93.13±1.19 (244) 190.97±3.82 (97) 202.83±2.47 (233) 106.03±3.53 (96) 114.24±2.21 (232) 46.09±1.23 (97) 46±0.77 (232) 0.83 0.95 0.33 0.32 0.74 0.009 0.046++ 0.79 88.89±0.97 (274) 88.98±0.75 (299) 202.34±2.12 (266) 199.81±2.16 (297) 115.76±1.88 (266) 113.24±1.91 (297) 58.49±0.87 (266) 57.82±0.81 (297) 92.57±1.16 (269) 91.41±1.08 (299) 202.19±2.27 (260) 196.51±2.3 (281) 116.15±2.11 (260) 110.2±1.03 (279) 45.66±0.69 (260) 45.99±0.68 (280) 0.98 0.23 0.23 0.59 0.35 0.077 0.039 0.84 88.73±0.71 (419) 91.48±2.29 (65) 203±1.78 (412) 192.38±4.43 (60) 115.54±1.59 (412) 109.18±3.92 (60) 58.78±0.7 (412) 57.03±1.52 (60) 91.1±0.72 (430) 92.46±2.4 (71) 200.76±1.95 (406) 199.06±4.27 (69) 113.62±1.75 (403) 113.41±4.04 (69) 45.99±0.57 (404) 43.63±1.07 (69) 0.2 0.024++ 0.034++ 0.43 0.55 0.62 0.88 0.16 88.6±0.61 (527) 95.46±8.21 (13) 201.29±1.58 (516) 203.33±11.66 (12) 114.11±1.41 (516) 121.62±10.22 (12) 58.76±0.63 (516) 56.5±2.79 (12) 91.48±0.74 (529) 92.21±4.13 (14) 200.45±1.71 (507) 198.55±12 (11) 113.3±1.54 (504) 122.95±9.16 (11) 45.74±0.51 (505) 43.31±1.86 (11) 0.08 0.64 0.27 0.63 0.77 0.93 0.33 0.37 + Los valores indican la media ± desviación estándar (número). p=p-value. = p-value modelo dominante. ++ = p-value modelo aditivo. Ajustado por edad, IMC, horas de ayuno. Los hombres portadores del alelo C del polimorfismo rs1800976 presentaron niveles más altos de colesterol total frente a los portadores del genotipo G/G (p=0.009). Además, este polimorfismo mostró asociación con valores de LDL al considerar el modelo aditivo, donde cada copia del alelo C se relaciona con niveles 5.49mg/dL (IC 95% 0.12 - 10.87) más altos de LDL, respecto del genotipo G/G (p= 0.046) (Tabla 4.5.). El polimorfismo rs2246298 se relacionó con valores de colesterol total y LDL en hombres. Los hombres con genotipo C/T-T/T presentaron niveles más altos de colesterol y LDL al comparar frente a los hombres C/C (p= 0.008 y p= 0.016, respectivamente) (Tabla 4.5.). 21 En mujeres, cada copia del alelo T del polimorfismo rs1800977 se asoció con valores 5mg/dL y 4 mg/dL más bajos de colesterol total y LDL, respectivamente (p= 0.024 y p= 0.034, respectivamente) (Tabla 4.5.). Los polimorfismos rs2422493 y rs2246293 mostraron asociaciones en hombres y mujeres, principalmente para hombres en relación con LDL. La asociación de estos polimorfismos es diferente para cada sexo (Tabla 4.5.), por lo que su efecto se anula al analizar toda la población conjuntamente (Tabla 4.4.). Los genotipos C/T-T/T del polimorfismos rs2422493 mostraron asociación a niveles altos de colesterol total (p=0.031) y LDL (p=0.032), y valores bajos de HDL (p=0.045) en hombres. Mientras que en mujeres se asociaron a niveles bajos de LDL (p=0.046). Los genotipos G/C-C/C del polimorfismo rs2246293 mostraron asociación a niveles altos de colesterol total (p=0.014) y LDL (p=0.009), y a valores bajos de HDL (p=0.035) en hombres. Mientras que en mujeres se asociaron a niveles bajos de LDL (p= 0.024). Aplicando la corrección de Bonferroni, ninguna de las asociaciones indicadas alcanza la significancia estadística (p corregida = 0.002). Al realizar el análisis de ligamiento se observó que los polimorfismos estudiados presentaban desequilibrio de ligamiento. El gráfico siguiente representa en colores oscuros mayor desequilibrio de ligamiento tomando en cuenta el valor de D, que representa la diferencia entre la frecuencia observada y la esperada, asumiendo equilibrio. Figura 4.1. Desequilibrio de ligamiento entre los polimorfismos En la Tabla 4.6. se muestran los haplotipos encontrados en la población con una frecuencia superior al 5%, estos haplotipos son los más frecuentes también en mujeres y hombres pero con frecuencias distintas entre los haplotipos 2 y 3 (Tabla 4.7. y 4.8.). Posteriormente se realizó el análisis de asociación entre haplotipos frecuentes y las variables en la población general y por sexo. Al realizar el análisis de asociación entre haplotipos y parámetros clínicos no se encontró ninguna asociación a hipercolesterolemia, hipertrigliceridemia e hipoalfalipiproteinemia, tanto 22 en la muestra total como al separarla por sexo. Tampoco se encontró asociación con obesidad y sobrepeso (datos no mostrados). El haplotipo, TCCCGCC, se mostró como factor protector frente a la diabetes tipo 2, con un OR de 0.45 (IC 95% 0.21 - 0.98) y p= 0.045, en población general (Tabla 4.6.). Al evaluar por sexo se pierde la significación estadística. El haplotipo CGCGGCC, se asoció a niveles altos de glucosa (Diferencia 2.02, IC 95% 0.07 - 3.97 y p= 0.042), colesterol total (Diferencia 5.67, IC 95% 1.18 - 10.16 y p= 0.013) y LDL (Diferencia 5.9 IC 95% 1.84 - 9.97 y p=0.004) en toda la muestra (Tabla 4.6.). Esta asociación se mantiene con significancia estadística para el grupo de hombres en el caso de la glucosa (p= 0.048) y en mujeres para colesterol (p=0.009) y LDL (p= 0.018)(Tabla 4.7 y 4.8.). Solamente en hombres TCTCGTC, se asoció a niveles más altos de colesterol y LDL (p= 0.02) (Tabla 4.8.), y también con el riesgo a desarrollar obesidad central con un OR 1.97 (1.10 - 3.53) y p= 0.023, respecto del haplotipo más frecuente (datos no mostrados). Los niveles plasmáticos de HDL se ven disminuidos en hombres portadores del haplotipo TCCCGTC (p= 0.04) Tabla 4.8. Tabla 4.6.Análisis de haplotipos en población general Haplotipo Frec 1 CGCGCCC 0.29 2 CGCGGCC 0.20 3 TCTCGTC 0.196 4 TCCCGTC 0.16 5 TCCCGCC 0.12 ******* 0.03 Diabetes tipo 2 OR P (95% CI) 1.00 --1.25 0.32 (0.80 - 1.93) 1.16 0.53 (0.73 - 1.85) 0.87 0.65 (0.48 - 1.58) 0.45 0.045 (0.21 - 0.98) 0.46 0.37 (0.09 - 2.49) Glucosa (mg/dL) Diferencia p (IC 95%) 0.00 --2.02 0.042 (0.07 - 3.97) 1.08 0.29 (-0.92 - 3.09) 0.24 0.84 (-2.02 - 2.5) 0.15 0.9 (-2.18 - 2.49) 3.66 0.4 (-4.95 - 12.28) CT (mg/dL) Diferencia p (IC 95%) 0.00 --5.67 0.01 (1.18 - 10.16) 3.52 0.14 (-1.1 - 8.14) -1.16 0.66 (-6.33 - 4.02) 2.76 0.32 (-2.64 - 8.15) 2.74 0.62 (-8.23 - 13.71) LDL (mg/dL) Diferencia p (IC 95%) 0.00 --5.9 0.004 (1.84 - 9.97) 3.47 0.1 (-0.69 - 7.63) 0.86 0.72 (-3.86 - 5.58) 3.4 0.17 (-1.47 - 8.28) 7.45 0.15 (-2.64 - 17.54) SNPs:rs2422493, rs2246293, rs2246298, rs1800976, rs2740483, rs1800977, rs111292742. Frec.= frecuencias. p= p-Value. 23 Tabla 4.7.Análisis de haplotipos en mujeres Haplotipos Frec 1 CGCGCCC 0.29 2 CGCGGCC 0.21 3 TCTCGTC 0.19 4 TCCCGTC 0.16 5 TCCCGCC 0.12 ******* 0.03 Glucosa (mg/dL) Diferencia P (IC 95%) 0.00 --0.63 0.6 (-1.73 - 3) 0.12 0.92 (-2.27 - 2.51) 1.4 0.29 (-1.2 - 4.01) 0.13 0.93 (-2.61 - 2.86) 4.24 0.17 (-1.83 - 10.3) CT (mg/dL) Diferencia p (IC 95%) 0.00 --7.26 0.009 (1.81 - 12.71) -0.6 0.84 (-6.3 - 5.09) -3.11 0.32 (-9.27 - 3.05) -0.58 0.86 (-7.24 - 6.08) 3.77 0.53 (-8.12 - 15.66) LDL (mg/dL) Diferencia p (IC 95%) 0.00 --6.48 0.018 (1.11 - 11.85) -1.03 0.71 (-6.39 - 4.33) -2.2 0.47 (-8.14 - 3.73) 0.49 0.88 (-5.91 - 6.88) 7.82 0.25 (-5.41 - 21.05) HDL (mg/dL) Diferencia p (IC 95%) 0.00 ---0.1 0.94 (-2.51 - 2.31) -1.03 0.42 (-3.56 - 1.49) 1.08 0.46 (-1.77 - 3.93) -0.04 0.98 (-2.91 - 2.83) -2.85 0.32 (-8.47 - 2.78) TG (mg/dL) Diferencia (IC 95%) 0.00 4.17 (-6.97 - 15.31) 4.9 (-7.01 - 16.81) -7.46 (-20.27 - 5.36) -1.95 (-15.54 - 11.63) -6.87 (-8.83 - -4.91) p --0.46 0.42 0.25 0.78 <0.00 SNPs: rs2422493, rs2246293, rs2246298, rs1800976, rs2740483, rs1800977, rs111292742. Frec.= frecuencias. p= p-Value. Tabla 4.8.Análisis de haplotipos en hombres Haplotipos Frec 1 CGCGCCC 0.29 2 TCTCGTC 0.21 3 CGCGGCC 0.19 4 TCCCGTC 0.16 5 TCCCGCC 0.11 ******* 0.04 Glucosa (mg/dL) Diferencia P (IC 95%) 0.00 --1.79 0.27 (-1.38 - 4.96) 3.07 0.05 (0.03 - 6.11) -1.31 0.48 (-4.91 - 2.29) -0.24 0.9 (-3.95 - 3.46) 1.93 0.59 (-5.13 - 8.98) CT (mg/dL) Diferencia p (IC 95%) 0.00 --7.81 0.02 (1.14 - 14.48) 2.68 0.4 (-3.53 - 8.9) 0.71 0.86 (-6.94 - 8.35) 4.63 0.24 (-3.07 - 12.34) 4.08 0.54 (-9.03 - 17.2) LDL (mg/dL) Diferencia p (IC 95%) 0.00 --7.48 0.02 (1.47 - 13.5) 3.87 0.18 (-1.83 - 9.58) 3.34 0.35 (-3.7 - 10.38) 4.16 0.24 (-2.78 - 11.1) 8.78 0.16 (-3.49 - 21.05) HDL (mg/dL) Diferencia P (IC 95%) 0.00 ---0.64 0.52 (-2.58 - 1.3) -0.38 0.71 (-2.36 - 1.6) -2.39 0.04 (-4.69 - -0.09) -0.53 0.66 (-2.86 - 1.8) -1.44 0.45 (-5.2 - 2.32) TG(mg/dL) Diferencia p (IC 95%) 0.00 --0.83 0.85 (-7.67 - 9.32) -7.8 0.2 (-19.77 - 4.17) -4.58 0.05 (-9.15 - -0.01) 1.31 0.16 (-0.54 - 3.15) -16.07 <0.00 (-16.5 - -15.65) SNPs: rs2422493, rs2246293, rs2246298, rs1800976, rs2740483, rs1800977, rs111292742. Frec.= frecuencias. p= p-Value. 24 Polimorfismos poco frecuentes: Como se indicó anteriormente, el polimorfismo rs56064613 solo se encontró en un individuo. A continuación se detallan las características clínicas y bioquímicas de esta persona. Mujer de 75 años de edad Talla: 156cm, Peso: 46Kg, IMC: 18.9Kg/m2 Cintura: 71cm, Cadera: 90cm, Índice cintura-cadera 0.79 Presión arterial sistólica/diastólica: 107/64mmHg, Hipertensa en tratamiento Glicemia: 85mg/dL (sin tratamiento) Colesterol total: 208mg/dL, LDL: 130.4mg/dL, HDL: 60mg/dL, Triglicéridos: 88mg/dL, no padece hipercolesterolemia ni hipertrigliceridemia. No se han indicado asociaciones entre este polimorfismo y variables de interés clínico. Finalmente, cabe indicar que se encontraron algunas variantes de secuencia no descritas en las bases de datos, en la Tabla 4.9.se detallan la ubicación de éstas y las características de los portadores. Para evaluar el posible efecto de estas variantes se considera el fenotipo del portador, respecto de las características clínicas y bioquímicas que presenta la población general. Además de analizar si influyen sobre la unión de factores de transcripción. Como se puede observar en la Tabla 4.9., todos los valores se encuentran dentro de los percentiles 25 a 75 en hombres y mujeres (datos no mostrados), excepto algunos casos que se mencionan a continuación. Los individuos del cambio +29 A/T no presentan un patrón bioquímico y clínico similar. Por lo que estos valores de triglicéridos y HDL, que se ubican bajo el percentil 10 y sobre el percentil 90, respectivamente, probablemente no sean efecto de la variante. En la mujer portadora de +233 C/A los triglicéridos se encuentran bajo el percentil 10 respecto del grupo total de mujeres, hay que anotar además, que en este caso se trata de una mujer joven respecto a la media del grupo. Si bien ABCA1 no presenta un efecto directo sobre los triglicéridos, las concentraciones de HDL podrían estar relacionadas con este tipo de lípidos. Los resultados del análisis con el programa MatInspector mostraron que las variantes de secuencia -264 G/T, +29 A/T y +233 C/A promueven un cambio en el patrón de factores de transcripción que reconocen la secuencia. En la Tabla 4.10 se indican aquellos factores de transcripción que difieren respecto de los que se unen en la secuencia de referencia. Se han descrito relaciones de importancia clínica con algunos de estos factores, tal es el caso de la lactoferrina, MYB, GRHL1. Se han publicado en este sentido relaciones con el metabolismo de lípidos, diferenciación celular y mantenimiento de la integridad de tejidos (Videm et al. 2012, Yagi et al. 2008, Wilanowski et al. 2008, Lorenzo et al. 2011). 25 Tabla 4.9. Nuevas variantes de secuencia Variante Sexo Edad (años) IMC 2 (Kg/m ) Cintura (cm) Sobre peso -264 G/T Hombre 42 28.4 96 Si No -215 G/T Mujer 74 28.3 90 Si Mujer 73 26.3 88 Hombre 63 27.7 Hombre 70 Mujer 30 +29 A/T +233 C/A Obesidad PAS central (mmHg) PAD (mmHg) HTA Glucosa CT (mg/dL) (mg/dL) LDL (mg/dL) HDL (mg/dL) TG (mg/dL) HTG SM 139.5 79.5 No 94 162 81.2 43.6 186 No No Si 135.7 90.3 Si 95 208 107.8 68.4 159 Yes Yes Si Si 138.5 66.5 Si 83 200 111.6 54.8 168 Yes No 101 Si No 139.5 89 Si 95 150 87.6 44.8 88 No No 28.0 104 Si Si 129.5 89 No 86 196 91.8 63.6 203 Yes Yes 23.6 76 No No 99.5 74.5 No 92 181 102 68.4 53 No No HTA= hipertensión arterial. HTG= hipertrigliceridemia. SM= síndrome metabólico. En azul los valores que se ubican sobre el percentil 90 o bajo el percentil 10 respecto del grupo al cual pertenecen. Tabla 4.10. Factores de transcripción que cambian según la secuencia Variante Factor de transcripción Función/Expresión La proteína muestra un amplio espectro de propiedades, incluyendo la regulación dela homeostasis del hierro, la defensa del huésped contra una amplia gama de infecciones microbianas, la actividad anti-inflamatoria, como factor de transcripción regula el crecimiento y la diferenciación celular y la protección contra el desarrollo del cáncer y la metástasis. -264 G/T Lactotransferrina y deltalactoferrinaª -215 G/T Ninguno +29 A/T AMYBª (v-myb myeloblastosis viral oncogene homolog) Esta proteína desempeña un papel esencial en la regulación de la hematopoyesis y puede desempeñar un papel en la tumorogénesis. OVOL1ª Esta proteína es probable que participe en la formación de pelo y la espermatogénesis en humanos, en base a la información conocida de su homóloga en Drosophila y ratón. AMYBª (v-myb myeloblastosis viral oncogene homolog) Esta proteína desempeña un papel esencial en la regulación de la hematopoyesis y puede desempeñar un papel en la tumorogénesis. GRHL1* (Grainyhead-like1) La isoforma 1 puede funcionar como un activador y la isoforma 2 como un represor en los tejidos donde se expresan ambas isoformas. La isoforma 1 se expresa en cerebro, páncreas, placenta, riñón. La isoforma 2 en cerebro e hígado. Puede desempeñar un papel en el desarrollo, y en la diferenciación epidérmica. +233 C/A Referencias: NCBI, GeneCards. *factor de transcripción que se une a la secuencia de referencia pero no cuando está presente el cambio. ªfactor de transcripción que se une cuando está presente el cambio y que no se encuentra en la secuencia de referencia. 26 5. DISCUSIÓN ABCA1 desempeña un papel clave en el mantenimiento de las concentraciones celulares de colesterol, por lo tanto, la regulación de su expresión es vital ya que tanto el déficit como el exceso de colesterol podrían ser perjudiciales para la célula. En las células β pancreáticas el papel puede ser aún más importante dado que el margen de colesterol que pueden tener sus membranas es más reducido para un correcto funcionamiento e incluso para la muerte celular. Variaciones en el funcionamiento del gen ABCA1 pueden relacionarse con los niveles de colesterol en partículas como las LDL y las HDL, los niveles de glucosa e, incluso el riesgo a diabetes. A nivel del promotor del gen actúan varios factores de transcripción que regulan su expresión ya sea promoviendo o inhibiendo su transcripción, la presencia de polimorfismos a este nivel podría alterar la unión de estos factores reguladores y por ende afectar el normal funcionamiento celular. Variantes genéticas en ABCA1 se han asociado con diversas alteraciones, entra ellas los niveles de HDL, la enfermedad cardiovascular y el riesgo a desarrollar diabetes tipo2 (Tang y Oram, 2009). Considerando esto, el principal objetivo de este trabajo fue identificar polimorfismos presentes en la región promotora y 5‘ UTR del gen ABCA1 y evaluar su asociación a diabetes tipo 2 o a parámetros bioquímicos y clínicos relacionados con alteraciones lipídicas en población española. En la población analizada se identificaron ocho SNPs, de los cuales ninguno mostró asociación individual a diabetes tipo 2 al analizar toda la población, tampoco se encontró asociación estadística a valores de glucosa en sangre (Tablas 4.3 y 4.4). Si bien, se ha reportado el posible efecto que ABCA1 desempeña en el desarrollo de esta enfermedad, también se propone que podría deberse a un efecto tóxico por el acúmulo de lípidos a nivel de los islotes β pancreáticos, ocasionando una disminución en la secreción de insulina (Gerin et al. 2005, Brunham et al. 2007, 2008). Es por eso que en el presente trabajo se evaluó también la asociación de los polimorfismos a variables relacionadas al metabolismo de lípidos. A pesar de no obtener resultados positivos de asociación entre los polimorfismos y la presencia de hipercolesterolemia, hipertrigliceridemia e hipoalfalipoproteinemia, si se encontraron algunas asociaciones con las variables cuantitativas relacionadas con el metabolismo de lípidos. El genotipo G/G del polimorfismo rs2740483 mostró asociación con niveles altos de colesterol total y LDL en la población (Tabla 4.4). En mujeres y hombres se mantiene la tendencia pero muestra asociación estadística únicamente para los valores de LDL en hombres (Tabla 4.5). Este polimorfismo junto con otros se ha estudiado en diferentes poblaciones, pero no se han encontrado asociado a valores lipídicos (Kiriakou et al.2007, Regieli et al.2011). Al analizar la población separando por sexo, se observaron asociaciones diferentes en cada grupo. Los genotipos C/T-T/T y G/C-C/C de los polimorfismos rs2422493 y rs2246293, respectivamente, mostraron asociación a niveles altos de colesterol total y LDL y valores bajos de HDL en hombres. Mientras que en mujeres se asociaron a niveles bajos de LDL (Tabla 4.5.). 27 Resulta llamativo el efecto contrario que presentaron los genotipos de los polimorfismos rs2422493, rs2246293 en ambos sexos, respecto a las concentraciones lipídicas. Estos polimorfismos inducen la unión de factores de transcripción diferentes respecto de la secuencia de referencia, sin embargo estos no muestran una relación directa con el sexo aunque sí podrían tener un efecto sobre la expresión del promotor (Anexo 1). El efecto de las condiciones fisiológicas propias de cada sexo podría estar modulando este efecto a través de diferentes señales y actuando de diferente forma mediante estos factores de transcripción. Los polimorfismos señalados anteriormente se han estudiado en varias poblaciones para evaluar su efecto sobre parámetros clínicos y bioquímicos. El polimorfismo rs2422493 se ha descrito como un factor que influye en la severidad de la aterosclerosis coronaria y en los niveles de expresión de ABCA1. Pacientes con esta enfermedad, portadores del alelo T presentaron mayor daño en sus arterias y bajos niveles de expresión de ABCA1 a nivel de la placa de ateroma. Pero en otro estudio se atribuye la disminución de complicaciones vasculares mortales en hombres con enfermedad coronaria al alelo T de este polimorfismo, cuando forma el haplotipo TGCC con los polimorfismos rs1800976, rs2740483 y rs1800977, respectivamente (Kiriakou et al.2005, Regieli et al.2011). rs2246293 ha sido relacionado con la edad de aparición de los síntomas en pacientes con enfermedad coronaria, apareciendo los síntomas 2.8 años más tarde en los portadores del alelo C respecto de los portadores del alelo de referencia (G), quienes además presentaban estenosis coronaria a temprana edad. Además, la región promotora con el alelo C mostró mayor actividad favoreciendo la transcripción del gen (Kiriakou et al.2007). El polimorfismo rs2246298 se ha estudiado en varias poblaciones. En población japonesa ha mostrado asociación con los niveles plasmáticos de HDL, efecto que no se presenta en otros grupos poblacionales (Shioji et al. 2004, Kiriakou et al. 2007). En nuestro caso tampoco se encontró asociación con los niveles de HDL, pero si se encontró asociación entre los genotipos C/T-T/T y las concentraciones altas de colesterol y LDL en hombres. El polimorfismo rs1800976 se ha vinculado con el padecimiento de enfermedad coronaria, con valores altos de VLDL, y también se ha indicado asociaciones a niveles de HDL en varias poblaciones (Shioji et al. 2004, Ergen et al. 2008). En nuestra población de estudio, los hombres con genotipos G/C-C/C del polimorfismo presentaron niveles altos de colesterol total y LDL. rs1800977 se ha relacionado con niveles altos de HDL en hombres y a valores altos de HDL en ambos sexos cuando se presenta en combinación con el polimorfismo R219K (Hodoglugil et al. 2005). En un estudio realizado en pacientes con enfermedad coronaria y controles sanos, rs1800977 mostró asociación a niveles bajos de triglicéridos y VLDL, aunque no se mostraron diferencias significativas entre los dos grupos de estudio (Ergen et al. 2008). En el presente trabajo el alelo T del polimorfismo rs1800977 estuvo asociado a niveles bajos de colesterol y LDL en mujeres. Los polimorfismos rs1800976 y rs1800977 se han postulado también como factores que aumentan el riesgo de eventos coronarios, esto sin que necesariamente tengan efecto sobre valores de lípidos plasmáticos (Zwarts et al. 2002). 28 Si bien se ha indicado por varios estudios el efecto de los polimorfismos de ABCA1 sobre los valores de lípidos en plasma, existen discrepancias en los resultados, posiblemente debido a las características de las poblaciones analizadas, esto es origen, edad, características clínicas, alimentarias, entre otras. Las diferencias entre géneros en cuanto a la expresión fenotípica de un mismo polimorfismo probablemente sean el resultado de un efecto complejo y multifactorial que involucre las hormonas sexuales (Hodoglugil et al. 2005, Abellán et al. 2010). Tal como se indicó en los resultados, las asociaciones encontradas en esta población pierden significación estadística al aplicar la corrección de Bonferroni. El presente estudio es un estudio piloto para determinar si es interesante el estudio de esta región en una muestra amplia; si bien, los resultados obtenidos no son concluyentes a nivel estadístico sí que podría considerarse que confirman nuestras sospechas de un posible efecto en las variables de interés. Ninguno de los polimorfismos individualmente mostró relación con los niveles plasmáticos de glucosa o el padecimiento de diabetes tipo 2 en toda la muestra, sin embargo el haplotipo TCCCGCC se presentó como factor protector frente a la diabetes tipo 2 (OR de 0.45, IC 0.21 - 0.98, p=0.045), y el haplotipo CGCGGCC se asoció a niveles altos de glucosa (p=0.048) (Tabla 4.6.). Cabe señalar, que la significancia de esta asociación es débil, probablemente el número de individuos limita el poder estadístico, por lo que nuevos estudios son necesarios para validar estos resultados. El haplotipo CGCGGCC, además mostró asociación a valores altos de colesterol total y LDL (p=0.01 y p=0.004, respectivamente). Cabe indicar que el haplotipo CGCGGCC contiene el alelo G del polimorfismo rs2740483, el cual estuvo asociado a colesterol y LDL en la población cuando se evaluó su efecto individualmente, y es el único alelo que difiere respecto del haplotipo más frecuente. El análisis de haplotipos por sexo mostró que en mujeres CGCGGCC se asocia a concentraciones de colesterol total y LDL 7 y 6 mg/dL más altas, respectivamente (Tabla 4.7). En hombres el haplotipo TCTCGTC se asocia a niveles altos de colesterol y LDL, y el haplotipo TCCCGTC con niveles de HDL 2mg/dL más bajos respecto del haplotipo mayoritario (Tabla 4.8). El análisis de asociación de haplotipos aporta con información relevante principalmente cuando los alelos que lo conforman forman parte del mismo haplotipo; sin embargo pueden existir discrepancias en los resultados obtenidos, ya que es posible que asociaciones que se observaron con alguno de los alelos en forma individual puedan no observarse en el análisis de haplotipos. Este efecto podría deberse al hecho de que en el análisis de haplotipos se consideran los heterocigotos, lo que podría disminuir la significancia si la asociación se presentaba con los homocigotos minoritarios del polimorfismo (Hodoglugil et al.2005). Por otro lado, se pueden observar asociaciones que no se evidenciaron en el análisis de los polimorfismos. La presencia de polimorfismos del promotor de ABCA1 resulta en un cambio de los factores de transcripción que interactúan con la secuencia (Anexo 1), posiblemente la interacción de varios de ellos podría presentar un efecto significativo sobre una variable, efecto que no se manifiesta en el análisis de polimorfismos individualmente. 29 Finalmente, cabe indicar que se encontraron nuevas variantes de secuencia en el promotor del gen ABCA1 (Tabla 4.10 y 4.11). In silico los cambios -264 G/T, +29 A/T y +233 C/A modifican el patrón de factores de transcripción que reconocen el promotor del gen, considerando como referencia la secuencia NG 007981 del NCBI. Aparentemente, estos factores de transcripción no tendrían influencia directa sobre ABCA1. Harían falta estudios funcionales para descartar esta opción. Dado el importante papel que desempeñan algunos de estos reguladores (lactoferrina, MYB myeloblastosis viral oncogene homolog, GRHL1 Grainyhead-like1), en procesos como diferenciación celular, metabolismo lipídico, relación con ciertas manifestaciones clínicas, regulación de la expresión de otros factores transcripcionales, renovación celular, se constituyen en objetivos para el desarrollo de estudios en este sentido (Wilanowski et al. 2008, Lorenzo et al. 2011, Videm et al. 2012). Por otro lado, todos estos factores de transcripción se expresan en el páncreas, hígado, tejido adiposo, etc. lugares donde podrían regular la actividad del ABCA1, si bien este extremo habría que estudiarlo mediante estudios específicos. El análisis del efecto que tienen los polimorfismos rs2422493, rs2246293, rs2246298, rs1800976, rs2740483 y rs1800977 sobre la unión de factores de transcripción al promotor del gen ABCA1, indica algunos factores que podrían estar regulando su expresión (Anexo 1). En algunos casos se han descrito funciones importantes en el desarrollo y funcionamiento del páncreas, e incluso en el desarrollo de diabetes. PDX-1 (Pancreatic and Duodenal homeoboxprotein), miembro de los genes HOX, implicados en el desarrollo embrionario, está involucrado en el normal desarrollo y función del páncreas. También participa en la función endocrina del páncreas mediante la supervivencia de células beta en el adulto. La actividad de este regulador requiere de cofactores entre los que se incluyen proteínas MEIS, Myeloidecotropic viral integration site (Brustin et al. 2010). Un estudio indica que HOXA se expresa también en células precursoras de células β pancreáticas, y participa en procesos de transición del estado mesenquimal a epitelial o viceversa, planteando nuevas estrategias de estudio en el reemplazo, mantenimiento y función de células beta (Limbert et al. 2009). TGF-b/Smad3 se han reportado relacionados en la patogénesis de la obesidad y diabetes tipo 2. Alteraciones en TGF-b se han relacionado con obesidad mórbida y neuropatía diabética. Al parecer TGF-b/Smad3 regulan un amplio rango de eventos que involucran el metabolismo y la etiología de la obesidad y diabetes. Se ha descrito su papel en la diferenciación de adipocitos, metabolismo de lípidos y glucosa, función pancreática, inflamación, secreción de adipocitoquinas y producción de especies reactivas de oxígeno, con lo cual se plantean nuevas alternativas terapéuticas para estas condiciones metabólicas (Tan et al. 2012). Con los resultados de este trabajo se ha obtenido una referencia del posible efecto de estos polimorfismos sobre variables bioquímicas de interés clínico, sobre todo aquellas relacionadas con el metabolismo lipídico. Sin embargo es necesario que estos resultados se valoren en otras poblaciones de características similares, y se realicen estudios funcionales a fin de validar el efecto de estos polimorfismos. 30 6. CONCLUSIONES • • • • • • • • No se encontró asociación entre los polimorfismos estudiados y la presencia de diabetes tipo 2 o alteraciones en las concentraciones de glucosa sanguínea en la población. El genotipo G/G del polimorfismo rs2740483 se asoció a valores más altos de colesterol total y LDL en la población, respecto de los G/C-C/C. El efecto de los polimorfismos sobre los parámetros bioquímicos es diferente en hombres y mujeres. o Los genotipos C/T-T/T del polimorfismos rs2422493 mostraron asociación a niveles altos de colesterol total y LDL, y valores bajos de HDL en hombres. Mientras que en mujeres se asociaron a niveles bajos de LDL. o Los genotipos G/C-C/C del polimorfismo rs2246293 mostraron asociación a niveles altos de colesterol total y LDL, y a valores bajos de HDL en hombres. Mientras que en mujeres se asociaron a niveles bajos de LDL. o Los genotipos C/T-T/T del polimorfismos rs2246298 se asociaron a niveles altos de colesterol total y LDL en hombres. o Los genotipos G/C-C/C del polimorfismo rs1800976 se asociaron a niveles altos de colesterol total y LDL en hombres. o Los genotipos C/T-T/T del polimorfismo rs1800977 se asociaron a valores bajos de colesterol total y LDL en mujeres. Si bien estas asociaciones pierden significación estadística al aplicar la corrección de Bonferroni, si podría considerarse que confirman nuestras sospechas de un posible efecto en las variables de interés. El haplotipo CGCGGCC se asoció a niveles altos de glucosa, colesterol total y LDL en la población. El haplotipo TCCCGCC se relaciona con la disminución del riesgo de padecer diabetes tipo 2 en la población respecto del haplotipo mayoritario. Se identificaron las variantes de secuencia en la región analizada del gen ABCA1: 264 G/T, -215 G/T, +29 A/T, +233 C/A. Su efecto debería validarse en nuevos estudios. Los resultados obtenidos en este trabajo constituyen una referencia del efecto de estos SNPs sobre parámetros bioquímicos de importancia clínica. Es necesario analizar otras poblaciones y realizar estudios funcionales a fin de validar los resultados. 31 7. ANEXOS ANEXO 1. Factores de transcripción que cambian según el polimorfismo Variante rs2422493 C/T Factor de transcripción Sma- and Mad-related proteinsª Meishomeobox 1ª rs2246293 G/C rs2246298 C/T Member of the vertebrate HOX - cluster of homeobox factors* Huntington's disease gene regulatory region-binding protein 1 and 2* Estrogen response elementsª rs1800976 G/C CCCTC-binding factor* cAMP-responsive element binding proteins * rs2740483 C/G Función Esta familia de proteínas son moduladores transcripcionales que median múltiples vías de señalización. Funciona como transductor de señales de los miembros de la familia TGF-beta, se cree que desempeñan un papel en la regulación de la carcinogénesis. Miembro de la familia de genes HOX. Se lo vincula en la hematopoyesis. Puede funcionar como un cofactor junto con otros factores HOX en la inducción de leucemias mieloides. Se expresa en células precursoras de células beta y en páncreas. Los genes HOX desempeñan un papel crucial en el normal desarrollo embrionario. Varias homeoproteinas están involucradas en el desarrollo de neoplasias. Es un factor de transcripción nuclear implicado en la activación del transportador de glucosa SLC2A4. Interactúa con otro factor de transcripción, myocyte enhancer factor 2, para activar la transcripción de este gen. El receptor de estrógeno (ER) es miembro de la superfamilia de receptores nucleares. ER se une a secuencias específicas de ADN denominadas elementos de respuesta a estrógenos y activa la expresión génica en respuesta a estradiol. El estrógeno y sus receptores son esenciales para el desarrollo sexual y la función reproductiva, también juegan un papel en otros tejidos como hueso. Los receptores de estrógenos también están implicados en procesos patológicos, como el cáncer de mama, cáncer de endometrio, y la osteoporosis. Regulador transcripcional con once dominios dedos de zinc. Dependiendo del contexto puede unirse una histona acetiltransferasa, o histona desacetilasa y actuar como activador o represor transcripcional, respectivamente. Se lo ha asociado con los cánceres invasivos de mama, cánceres de próstata, y tumores de Wilms. Factor de transcripción que se une como un homodímero al elemento de respuesta a AMPc, un palíndromo octamérico. La proteína se fosforila por varias cinasas, induce la transcripción de genes en respuesta a la estimulación hormonal por la vía del cAMP. GABP: GA binding proteinª Factor de transcripción capaz de interactuar con repeticiones ricas en purina (GA repeticiones). Puede estar implicado en la activación de la expresión de la citocromo oxidasa y el control nuclear de la función mitocondrial. Insulator protein CTCF (CCCTC-binding factor)ª Regulador transcripcional recononce dominios dedos de zinc. Dependiendo del contexto puede unirse una histona acetiltransferasa, o histona desacetilasa y actuar como activador o represor transcripcional, respectivamente. Se lo ha asociado con los cánceres invasivos de mama, cánceres de próstata, y tumores de Wilms. Regulador transcripcional. Reconoce y se une a la secuencia de ADN 5'-CGCCCCCGC-3 '(EGR-site). Activa la transcripción de genes diana, cuyos productos se requieren para la mitogénesis y diferenciación. EGR1, early growth 32 response 1ª Kruppel-like factor 6ª v-Myb* rs1800977 C/T Smad4 transcription factor involved in TGF-beta signalingª Miembro de la familia de factores de transcripción Kruppel. Es un activador transcripcional, y funciona como un supresor tumoral. Se han descrito isoformas implicadas en la carcinogénesis. Esta proteína desempeña un papel esencial en la regulación de la hematopoyesis y puede desempeñar un papel en la tumorogénesis. Esta proteína se une al ADN y reconoce una secuencia palindrómica8-bp (GTCTAGAC) llamada elemento de unión a Smad. Forma complejos con otras proteínas Smad activadas, y regulan la transcripción de genes en respuesta a TGF-beta. Sus alteraciones se han relacionado con cáncer de páncreas, síndrome de poliposis juvenil, y el síndrome de telangiectasia hemorrágica hereditaria. Referencias: NCBI, GeneCards.*factor de transcripción que se une a la secuencia de referencia pero no cuando está presente el cambio. ªfactor de transcripción que se une cuando está presente el cambio y que no se encuentra en la secuencia de referencia. 33 8. BIBLIOGRAFÍA 1. Abellan R., Mansego M. L., Martinez-Hervasa S., Martin-Escudero J. C., et al. Association of selected ABC gene family single nucleotide polymorphisms with postprandial lipoproteins: Results from the population-based Hortega study. Atherosclerosis (2010) 203–209 2. Brunham L., Kruit J., Pape T., Timmins J., Reuwer A., et al. β-cell ABCA1 influences insulin secretion, glucose homeostasis and response to thiazolidinedione treatment. Nature Medicine (2007) vol.13 3. Brunham Liam R., Kruit Janine K., Hayden Michael R., Verchere Bruce. Cholesterol in βcell Dysfunction: The Emerging Connection between HDL Cholesterol and Type 2 Diabetes. CurrDiabRep (2010) 10:55–60 4. Brunham Liam R., Kruit Janine K., Verchere C. Bruce, and Hayden Michael R. Cholesterol in islet dysfunction and type 2 diabetes. J. Clin. Invest. 118 (2008) 403–408 5. Bungert Stefanie, Molday Laurie L., and Molday Robert S. Membrane Topology of the ATP Binding Cassette Transporter ABCR and Its Relationship to ABC1 and Related ABCA Transporters. TheJournal Of BiologicalChemistry (2001) 23539–23546 6. Dean M., Hamon Y. and Chimini G. The human ATP-binding cassette (ABC) transporter superfamily. Journal of Lipid Research (2001) Volumen 42. 7. Ergen Arzu, Isbir Selim, Tekeli Atike and Isbir Turgay. Investigation of ABCA1 C69T and G-191C Polymorphisms in Coronary Artery Disease. in vivo (2008) 22: 187-190. 8. European Group for the Study of Insulin Resistance (1999). 9. Genomatix Software GmbH (http://www.genomatix.de/) 10. Gerin Isabelle, Dolinsky Vernon W., Shackman Jonathan G., Kennedy Robert T., et al. LXRβ Is Required for Adipocyte Growth, Glucose Homeostasis, and β-Cell Function. The Journal Of Biological Chemistry (2005) Vol. 280, 23024–23031 11. Hao Mingming, Head Steven, Gunawardana S., Hasty Alyssa H., and Piston David W. Direct Effect of Cholesterol on Insulin Secretion. A Novel Mechanism for Pancreatic β-Cell Dysfunction. Diabetes (2007) vol.56 12. Hodoglugil Ugur, Williamson David W., Huanga Yadong, Mahley Robert W. Common polymorphisms of ATP binding cassette transporter A1, including a functional promoter polymorphism, associated with plasma high density lipoprotein cholesterol levels in Turks. Atherosclerosis (2005) 199–212 13. http://bioinfo.iconcologia.net/SNPstats 14. http://www.lifesequencing.com 15. Kaminski Wolfgang E., Piehler Armin, Wenzel Jürgen J. ABC A-subfamily transporters: Structure, function and disease. Biochimica et Biophysica Acta 1762 (2006) 510–524 16. Kyriakou T., Hodgkinson C., Pontefract D., Iyengar S. et al. Genotypic Effect of the 565C>T Polymorphism in the ABCA1 Gene Promoter on ABCA1 Expression and Severity of Atherosclerosis Arterioscler Thromb Vasc Biol (2005) 418-423 17. Kyriakou T., Pontefract D., Viturro E., Hodgkinson C. et al. Functional polymorphism in ABCA1 influences age of symptom onset in coronary artery disease patients. Human Molecular Genetics (2007) 1412–1422 18. Limbert C, Ebert R., Schilling T. et al. Functional Signature of Human Islet-derived Precursor Cells compared to Bone Marrow-derived Mesenchymal Stem Cells. StemCells and Development 2009. 19. Liu Yuhua, Tang Chongren Regulation of ABCA1 functions by signaling pathways Biochimica et Biophysica Acta 1821 (2012) 522–529 34 20. Lorenzo Petra I., Brendeford Elen M., Gilfillan Siv, Gavrilov Alexey A., et al.Identification of c-Myb Target Genes in K562 Cells Reveals a Role for c-Myb as a Master Regulator. Genes & Cáncer (2011) 805–817 21. Mena Martín F. J., Martín Escudero J. C., Simal Blanco F., Carretero Ares J. L., Herreros Fernández V. Factores de riesgo cardiovascular en pacientes diabéticos. Estudio epidemiológico transversal en población general: estudio Hortega. An. Med. InternaMadrid (2003) Vol. 20, 292-296 22. Nagao Kohjiro, Tomioka Maiko and Ueda Kazumitsu. Function and regulation of ABCA1 – membrane meso-domain organization and reorganization. FEBS Journal 278 (2011) 3190–3203 23. National Center for Biotechnology Information http://www.ncbi.nlm.nih.gov/ 24. Oram John F. and Heinecke Jay W. ATP-Binding Cassette Transporter A1: A Cell Cholesterol Exporter That Protects Against Cardiovascular Disease. PhysiolRev85 (2005) 1343–1372 25. OMS (http://www.who.int) 26. Patel DC, Albrecht C, Pavitt D, Paul V, Pourreyron C, et al. Type 2 Diabetes Is Associated with Reduced ATP-Binding Cassette Transporter A1 Gene Expression, Protein and Function. PLoS ONE (2011). 27. Rayner Katey J., Suárez Yajaira, Dávalos Alberto, Parathath Saj, Fitzgerald Michael L., et al. MiR-33 Contributes to the Regulation of Cholesterol Homeostasis. Science (2010) 28. Regieli Jakub J., Doevendans Pieter A., Grobbee Diederick E., Zwinderman Aeilko H. et al. ABCA1 impacts athero-thrombotic risk and 10-year survival in a contemporary secondary prevention setting. Atherosclerosis (2011) 457–463 29. Santamarina-Fojo S., Peterson K., Knapper C., Qiu Y., Freeman L., et al. Complete genomic sequence of the human ABCA1 gene: Analysis of the human and mouse ATPbinding cassette A promoter. PNAS (2000) vol. 97-14 7987–7992 30. Shioji Keisuke, Nishioka Junko, Naraba Hiroaki, et al. A promoter variant of the ATPbinding cassette transporter A1 gene alters the HDL cholesterol level in the general Japanese population. J HumGenet (2004) 49:141–147 31. Singaraja Roshni R., Brunham Liam R., Visscher Henk, Kastelein John J.P. and Hayden Michael R. Efflux and Atherosclerosis: The Clinical and Biochemical Impact of Variations in the ABCA1 Gene Arteriosclerosis. Arterioscler Thromb Vasc Biol.23 (2003) 1322-1332 32. Stefková J., Poledne R., Hubáček J. ATP-Binding Cassette (ABC) Transporters in Human Metabolism and Diseases. Physiol. Res 53 (2004) 235-243 33. Tan CK., Chong HC., Tan EHP and Tan NS. Getting ‘Smad’ about obesity and diabetes. Nutrition and Diabetes (2012) 2, e29; 10.1038. 34. Tang Chongren, Oram John F. The cell cholesterol exporter ABCA1 as a protector from cardiovascular disease and diabetes. Biochimica et Biophysica (2009) 563–572 35. Third Report of the National Cholesterol Education Program (NCEP) Expert Panel on Detection, Evaluation, and Treatment of High Blood Cholesterol in Adults (Adult Treatment Panel III) Final Report. American Heart Association. (2002) 36. Vedhachalam CH., Ghering A., Davidson W., Lund-Katz S., et al. ABCA1-Induced Cell Surface Binding Sites for ApoA-I. Arterioscler Thromb Vasc Biol. (2007) 27:1603-1609 37. Videm Vibeke, Dahl Hildegunn, Walberg Lise Eid, Wiseth Rune. Functional polymorphisms in the LTF gene and risk of coronary artery stenosis. Human Immunology 73 (2012) 554–559 38. von Burstin J, Reichert M, Wescott MP, Rustgi AK. The Pancreatic and Duodenal Homeobox Protein PDX-1 Regulates the Ductal Specific Keratin 19 through the Degradation of MEIS1 and DNA Binding. PLoSONE (2010) 5(8): e12311. 35 39. Wilanowski T., Caddy J., Ting S., Hislop N., et al. Perturbed desmosomal cadherin expression in grainy head-like 1-null mice. The EMBO Journal (2008) 27, 886–897 40. Yagi M., Suzuki N., Takayama T., Arisue M., et al. Lactoferrin suppress the adipogenic differentiation of MC3T3-G2/PA6 cells. J Oral Sci (2008) 50:419–25. 41. Zwarts KY, Clee SM, Zwinderman AH, Engert JC, Singaraja R. et al. ABCA1 regulatory variants influence coronary artery disease independent of effects on plasma lipid levels. Clin Genet (2002) 61: 115–125 36