Regulación neuro-hormonal de la función reproductora
Anuncio

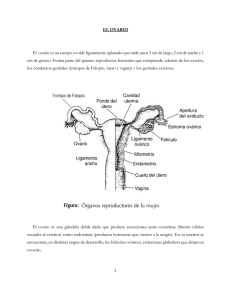
Clases de Residentes 2008 Regulación neurohormonal de la función reproductora Servicio de Obstetricia y Ginecología Hospital Universitario Virgen de las Nieves Granada REGULACIÓN NEUROHORMONAL DE LA FUNCIÓN REPRODUCTORA. María de la Torre Bulnes 1. INTRODUCCIÓN El proceso reproductivo depende de la ovulación regular de un óvulo maduro a mitad de cada ciclo menstrual; a su vez, este complejo proceso depende del eje hipotálamo-hipófisis-ovario, y de los estímulos aferentes de otros sistemas. 2. CONCEPTOS BÁSICOS Componentes del eje hipotalamo-hipofisiario-gonadal Hipotálamo GnRH Hipófisis FSH, LH Ovario Estrógenos, progesterona Endometrio Dra. de la Torre/ Dr. Fontes -1- Clases de Residentes 2008 Regulación neurohormonal de la función reproductora Todos estos órganos y glándulas van a comunicarse y a regularse entre sí por distintos sistemas de retroalimentación, además de recibir otros estímulos reguladores externos, a través de hormonas y neurohormonas, con el fin de conseguir dos objetivos básicos para la reproducción: 1-Obtención de un ovocito en la ovulación. 2- Preparar al endometrio para acoger un posible embarazo. Tipos de vías de retroalimentación • Asa larga: Efecto de retroalimentación de las hormonas esteriodeas del ovario sobre el hipotálamo y la hipófisis. • Asa corta: Efecto de retroalimentación negativa de las hormonas hipofisiarias sobre el hipotálamo. • Asa ultracorta: Inhibición de una hormona de su propia síntesis. Concepto de hormona Sustancia que se produce en un tejido especial desde donde se libera hacia el torrente circulatorio y viaja a células sensibles distantes en las que ejerce sus efectos característicos. Son sustancias que proporcionan un medio de comunicación. Otros tipos de comunicación más inmediata son: • Paracrina: Comunicación intercelular que supone la difusión local de sustancias reguladoras desde una célula a células próximas o contiguas. • Autocrina: Comunicación intracelular mediante la cual una célula aislada produce sustancias reguladoras que a su vez actúan en receptores situados sobre o en el interior de la célula. • Intracrina: Cuando se unen sustancias no secretadas a receptores intracelulares. Los efectos biológicos y metabólicos de una hormona se encuentran determinados por la capacidad de una célula de recibir y retener la hormona, Dra. de la Torre/ Dr. Fontes -2- Clases de Residentes 2008 Regulación neurohormonal de la función reproductora gracias a receptores específicos, que ayudan a transmitir el mensaje de la hormona a la transcripción génica nuclear. El resultado es la producción de ARN mensajero, que desencadena la síntesis proteica y una respuesta celular activa de la célula. Concepto de neurohormona Son proteinas que se sintetizan en un cuerpo neuronal y se transportan a través del axón hasta la terminación nerviosa para secretarse a la sangre. Las neurohormonas del hipotálamo se vierten a los vasos de la circulación portal hipotálamo-hipofisiaria, constituida por una densa red de capilares en la eminencia media del hipotálamo que se vacían en los vasos portales que descienden a lo largo del tallo hipofisiario hasta la adenohipófisis. Si se secciona el tallo hipofisiario y se interrumpe esta circulación portal se produce la inactividad y atrofia de las gónadas, junto con el descenso de la actividad suprarrenal y tiroidea. Además se ha comprobado que si se transplanta la hipófisis a zonas ectópicas, incluso dentro del mismo encéfalo, también se produce el mismo efecto, a excepción de la secreción de prolactina, que se mantiene. De ahí la importancia de este sistema vascular. 3. EL HIPOTÁLAMO Y LA GnRH. Los agentes neuroendocrinos que se originan en el hipotálamo tienen efectos estimuladores positivos sobre la hormona del crecimiento, la tirotropina, la corticotropina y las gonadotropinas de la hipófisis. La neurohormona que controla las gonadotropinas de denomina hormona liberadora de gonadotropinas, gonadoliberina o GnRH. Inicialmente se pensaba que había 2 hormonas de liberación distintas, una para la FSH y otra para la LH; actualmente se admite que hay una sola neurohormona y que los distintos patrones de liberación de FSH y LH en respuesta una única GnRH se deben a los efectos de retroalimentación de los esteroides sobre la adenohipófisis. Dra. de la Torre/ Dr. Fontes -3- Clases de Residentes 2008 Regulación neurohormonal de la función reproductora Núcleos hipotalámicos Los núcleos hipotalámicos se encuentran a ambos lados del III ventrículo. De delante hacia atrás se distinguen: - La parte anterior o quiasmática, formada por los núcleos preóptico, paraventricular, supraóptico, supraquiasmático y anterior. - La parte media o túber cinereum, formada por los núcleos dorso medial, ventromedial e infundibular o arqueado. - La parte posterior o mamilar, formada por los cuerpos mamilares y los núcleos premamilares. - El núcleo hipotalámico posterior, situado por encima de los cuerpos mamilares. Las neuronas que fabrican la GnRH se encuentran principalmente en el núcleo arqueado, que se sitúa por fuera de la barrera hemato-encefálica (lo que le permite responder a estímulos aferentes originados en distintos órganos). Estas células se originan en el área olfativa, en una placa engrosada del ectodermo, y migran durante la embriogénesis a lo largo de los pares craneales que conectan la nariz y los núcleos septales preópticos del encéfalo. Este trayecto explica la clínica que aparece en el síndrome de Kallman, una asociación entre la falta de GnRH y un defecto del olfato, por fallo en la migración axonal olfativa y neuronal de GnRH. Además, al igual que las células olfativas de la cavidad nasal, las neuronas de GnRH poseen cilios. El origen olfativo y la semejanza estructural de estas neuronas y las células epiteliales nasales denotan una evolución de la reproducción controlada por feromonas, que son sustancias químicas transmitidas por el aire que libera un sujeto y pueden afectar a otros miembros de la misma especie. Secreción de GnRH Diversos experimentos han demostrado que la secreción normal de gonadotropinas requiere una secreción pulsátil de GnRH dentro de un intervalo Dra. de la Torre/ Dr. Fontes -4- Clases de Residentes 2008 Regulación neurohormonal de la función reproductora crítico de frecuencia y amplitud, y que varía sustancialmente a lo largo del ciclo ovárico: la actividad pulsátil es más frecuente pero de menos amplitud durante la fase folicular comparada con la fase lútea. Además se ha observado que si disminuye la frecuencia de los pulsos, aumenta la secreción de FSH, mientras que frecuencias mayores favorecen la secreción de LH. Por eso la disminución de la frecuencia de los pulsos de GnRH en la fase lútea tardía es un factor importante que favorece la síntesis y la secreción de FSH para en siguiente ciclo. Tanto una disminución en la frecuencia de los pulsos, como una exposición prolongada y continua a GnRH (como ocurre con el uso farmacológico de agonistas de la GnRH) van a provocar anovulación y amenorrea. Efectos de la GnRH Las acciones de la GnRH sobre sus receptores en la hipófisis activa múltiples mensajeros, provocando: 1. Liberación inmediata de gonadotropinas. 2. Síntesis y almacenamiento de gonadotropinas (pool de reserva) 3. Movimiento de gonadotropinas desde el pool de reserva al pool de secreción. Estas 3 acciones varían a lo largo de ciclo, en respuesta a los esteroides ováricos. Además por un fenómeno de autocebamiento estimula la síntesis de nuevos receptores que permiten a las células responder a cantidades menores de GnRH, lo cual es importante para la aparición del pico de LH previo a la ovulación. Control de los pulsos de GnRH Aunque la actividad rítmica pulsátil en una propiedad intrínseca de las neuronas de GnRH, puede modularse por la acción de diversas hormonas y neurotransmisores: Dra. de la Torre/ Dr. Fontes -5- Clases de Residentes 2008 • Regulación neurohormonal de la función reproductora Dopamina: Actúa suprimiendo directamente la actividad de las neuronas de GnRH, además de suprimir la secreción de prolactina en la hipófisis anterior. • Noradrenalina: Su acción favorece la liberación de GnRH. • Serotonina: Tiene efecto inhibidor sobre la secreción de GnRH. • Neuropéptido Y: Se produce en el hipotálamo en respuesta a los esteriodes gonadales. Estimula los pulsos de GnRH y potencia la respuesta gonadrópica de la hipófisis a dicha hormona; sin embargo, en ausencia de estrógenos, inhibe la secreción de gonadotropinas. Como la desnutrición se asocia a un incremento de neuropéptido Y, y se han medio mayores cantidades de dicha sustancia en el líquido cefalorraquídeo en mujeres con anorexia y bulimia nerviosa, se considera que este neuropéptido representa un vínculo entre la nutrición y la función reproductora. • Prolactina: Inhibe la secreción pulsátil de GnRH, lo que explica la amenorrea asociada con la hiperprolactinemia. • Opioides endógenos: Suprimen la liberación hipotalámica de GnRH, y además favorecen la secreción de prolactina, que también va a suprimir la liberación de GnRH. Provienen de un péptido precursor común con la ACTH, la proopiomelanocortina (POMC). Mientras que en la hipófisis la expresión del gen POMC produce sobre todo ACTH y está controlada por la CRH, en el hipotálamo predomina la producción de B-endorfinas, y está bajo el control de los esteriodes sexuales. Los estrógenos son los principales responsables del mantenimiento del tono opioide, ya que, aunque tanto el estradiol como la progesterona incrementan el tono opioide, los estrógenos potencian la acción de la progesterona, lo que explica la máxima supresión de GnRH y de gonadotropinas durante la fase lútea. De este modo, los niveles de B-endorfina en el sistema portal hipofisiario progresan lentamente desde un nadir en el momento de la menstruación, aumentando desde la mitad del ciclo folicular hasta alcanzar el acmé en la fase lútea. Dra. de la Torre/ Dr. Fontes -6- Clases de Residentes 2008 • Regulación neurohormonal de la función reproductora CRH (hormona liberadora de corticotropina): Inhibe la secreción de GnRH en el núcleo arcuato de forma directa, además de actuar indirectamente a través de un aumento de opiodes endógenos; esta podría ser la vía a través de la cual el estrés afecta a la función reproductora. El estrés, la depresión, los transtornos alimenticios, etc, producen la liberación de CRH, con la consiguiente activación del eje hipotálamo-hipófisissuprarrenal, además de incrementar el neuropéptido Y, que va a estimular la secreción de ACTH. Estas situaciones van a provocar un incremento de los corticoides, que además van a inhibir tanto la secreción de GnRH como la secreción hipofisiaria de LH y tornan los tejidos estrógeno-sensibles resistentes a los mismos. 4. HIPÓSIS Y GONADOTROPINAS La hipófisis ocupa el hueco de la silla turca, en el cuerpo del esfenoides. Se encuentra perfectamente encapsulada por una estructura fibrosa colágena que se continúa con la duramadre del diafragma. Las gonadotropinas se van a sintetizar en la adenohipófisis o hipófisis glandular, situada en el lóbulo anterior de la glándula, por debajo del diafragma. A diferencia del lóbulo posterior de la hipófisis, que tiene origen en el tubo neural, esta porción de la hipófisis tiene su origen embriológico en la cavidad bucal primitiva, por delante de la membrana faríngea: se forma a partir de la bolsa de Rathke, que es una invaginación que va desde el techo del estomodeo en dirección ascendente y antero-posterior hacia el suelo del III ventrículo. Las gonadotropinas Son las hormonas fabricadas por la adenohipófisis en respuesta a la GnRH, encargadas de vehiculizar la información desde el bloque central hasta las gónadas. Se denominan hormona foliculoestimulante (FSH) y hormona luteinizante (LH). Ambas son hormonas glucoproteicas constituidas por 2 subunidades, alfa y beta. La subunidad alfa es común para FSH, LH, TSH y gonadotropina Dra. de la Torre/ Dr. Fontes -7- Clases de Residentes 2008 Regulación neurohormonal de la función reproductora coriónica placentaria, mientras que la subunidad beta es específica de cada hormona y en ella residen sus propiedades biológicas. Sus funciones son: • FSH: Es necesaria para el desarrollo folicular más allá de la fase preantral: - Estimula la multiplicación celular en la granulosa, junto con los estrógenos y la LH. - Aumenta la cantidad de receptores para LH en la granulosa, preparándola así para la producción de progesterona durante la fase lútea. - Incrementa la capacidad de aromatización del folículo, favoreciendo la transformación de los precursores androgénicos en estrógenos. • LH: - Promueve la esteroidogénesis, aumentando la producción de precursores androgénicos en la teca, y también en la granulosa después de la ovulación. - Favorece el desarrollo folicular. - Favorece la ovulación. 5. EL OVARIO Y LOS ESTERIODES SEXUALES Las responsabilidades fisiológicas del ovario son la liberación periódica de gametos y la producción de hormonas esteroideas. Ambas actividades se integran en un proceso repetitivo y contínuo de maduración del folículo, ovulación y formación y regresión del cuerpo lúteo. Es un órgano en constante cambio. Histología del ovario Consta de 3 partes principales: • 1- La corteza: Parte externa del ovario. La parte más externa de la corteza se denomina túnica albugínea y su superficie está coronada por una sola capa de epitelio cúbico que recibe el nombre de epitelio superficial del ovario o mesotelio ovárico. Dra. de la Torre/ Dr. Fontes -8- Clases de Residentes 2008 Regulación neurohormonal de la función reproductora Los folículos se encuentran en la parte interior de la corteza, incrustados en el estroma, que está formado por tejido conjuntivo y células intersticiales que derivan de células del mesénquima y son capaces de responder a la LH o a la hCG produciendo andrógenos. • 2- La médula: Zona central del ovario, que procede de células mesonéfricas. • 3-Hilio del ovario: Es el punto de unión con el mesovario. Contiene células nerviosas, vasos sanguíneos y células hiliares, que son muy parecidas a las células de Leydig de los testículos. Ciclo folicular El ciclo típico de maduración de folículos, ovulación y formación del cuerpo lúteo durante los años reproductores de la vida de la mujer es el resultado de la secuencia compleja de interacciones hipotalámicas-hipofisiarias-gonadales, en las que se integran las hormonas esteriodeas de los folículos y el cuerpo lúteo, las gonadotropinas de la hipófisis, y distintos factores autocrinos y paracrinos. • Ovario fetal: A las 6-8 SG comienza una rápida multiplicación mitótica de las células germinales del ovario primitivo, que alcanzan los 6-7 millones de ovogonias entre las 16-20 SG; a partir de este momento su número irá disminuyendo. Estas ovogonias se transforman en ovocitos cuando entran en la primera división meiótica y se detienen en la profase; este proceso comienza a las 1112 SG. Posteriormente estos ovocitos darán lugar a un solo óvulo tras completar las dos divisiones meióticas: una justo antes de la ovulación y la 2ª que forma el óvulo haploide, cuando penetra el espermatozoide. A las 18-20 SG los ovocitos son rodeados por células de pregranulosa originadas en el mesénquima: la unidad resultante es el folículo primordial. Estos folículos experimentan grados variables de maduración: - Folículo primario: la capa de pregranulosa se transforma en una capa cúbica de células de granulosa. Dra. de la Torre/ Dr. Fontes -9- Clases de Residentes 2008 Regulación neurohormonal de la función reproductora - Folículo preantral: hay una proliferación de las células de la granulosa. Se detectan en el 6º mes de gestación. - Folículo antral o de De Graaf: Aparece un espacio lleno de líquido en el interior del folículo. Aparecen al final de la gestación, pero no en grandes cantidades. Debido a la reducción prenatal de ovocitos, el contenido total de células germinales de la corteza disminuye a 1-2 millones en el momento del nacimiento; las recién nacidas comienzan su vida habiendo perdido el 80% de sus ovocitos. Al principio de la pubertad, la masa de células germinales se reduce hasta 300.000-500.000, de las cuales sólo 400-500 serán seleccionadas para la ovulación. • Crecimiento folicular: El tiempo que dura la progresión desde folículo primario hasta la ovulación es de aproximadamente 85 días. La mayor parte de ese tiempo se dedica a un desarrollo que es independiente de las gonadotropinas, hasta alcanzar una disposición que permitirá que continúe el crecimiento en respuesta a la estimulación de FSH. Encontramos al ovocito rodeado de una capa de células de granulosa, que están separadas del estroma que las envuelve por una membrana basal. Estas células están privadas de riego vascular hasta después de la ovulación y dependen de uniones comunicantes especializadas que conectan las células. Se comunican con el ovocito para el intercambio metabólico y el transporte de moléculas de señalización, estimulando al ovocito para que la meiosis se produzca en el momento correcto. En estos folículos aparece una cavidad (antro) donde se acumula líquido que es un trasudado de sangre filtrada a través de la granulosa desde los vasos de la teca (que proviene del tejido conjuntivo que rodea al folículo). Con la formación del antro la teca interna se desarrolla de forma más completa, expresado por un aumento de masa celular, una mayor vascularización y la formación de vacuolas citoplasmáticas ricas en lípidos. Conforme el folículo se expande, el estroma que lo rodea se comprime, recibiendo el nombre de teca Dra. de la Torre/ Dr. Fontes - 10 - Clases de Residentes 2008 Regulación neurohormonal de la función reproductora externa. Con el estímulo de las gonadotropinas, estos folículos continuarán su crecimiento, aunque muchos de ellos sufrirán un proceso apoptótico, denominado atresia. • Ovulación: Si el estímulo de las gonadotropinas es suficiente uno de los folículos progresará hasta la ovulación. Se produce la dilatación del antro por aumento de líquido antral y compresión de la granulosa contra la membrana basal que la separa de la teca vascularizada y luteinizada. El folículo se encuentra distendido y prominente, provocando el adelgazamiento de la teca sobre la superficie del folículo, y creándose una zona avascular que debilita la cápsula del ovario. Finalmente y por distintos mecanismos, se produce una distensión aguda del antro con la rotura folicular y la extrusión del ovocito. La expulsión física del ovocito depende de un aumento vertiginoso de la síntesis de prostaglandinas en el folículo antes de la ovulación. La inhibición de esta síntesis de prostaglandinas produce un cuerpo lúteo con un ovocito atrapado. Parece que tanto las prostaglandinas como el aumento de gonadotropinas a mitad del ciclo provoca un incremento en la concentración y la actividad de proteasas locales y la conversión del plasminógeno a plasmina. Todo ello provoca la pérdida de la integridad de las uniones intercelulares comunicantes y la rotura de fibras elásticas, lo que conlleva un debilitamiento tisular generalizado. • Formación del cuerpo lúteo: Poco después de la ovulación se producen notables alteraciones de la organización celular del folículo. Cuando se recupera la integridad y la continuidad del tejido, las células de la granulosa experimentan una acusada hipertrofia llenando la cavidad quística del cuerpo lúteo precoz. Además por primera vez, la granulosa se luteiniza notablemente por la incorporación de vacuolas ricas en lípidos al interior de su citoplasma. Dra. de la Torre/ Dr. Fontes - 11 - Clases de Residentes 2008 Regulación neurohormonal de la función reproductora Por su parte la teca pierde prominencia y sólo se aprecian vestigios de la misma en el cuerpo lúteo maduro. Como resultado se forma el cuerpo amarillo, dominado por la granulosa hipertrófica, rica en lípidos y totalmente vascularizada. En los 14 días de su vida, dependiendo de las cantidades de LH en la fase lútea, esta unidad produce estradiol y progesterona. Al faltarle una fuente creciente de hCG procedente de una implantación satisfactoria, el cuerpo lúteo envejece con rapidez. Su vascularización y su contenido de lípidos desaparecen y se produce su cicatrización (cuerpo albicans) Esteroides sexuales del ovario Todas las hormonas esteriodeas tienen una estructura semejante; la estructura básica es la molécula de ciclopentanoperhidrofenantreno, que deriva del colesterol. Los esteroides sexuales se dividen en 3 grupos, según el número de átomos de carbono: -21 C: Corticosteroides y progestágenos, basados en el núcleo de pregnano. -19C: Todos los andrógenos. Se basan en el núcleo de androstano. -18C: Estrógenos, basados en el núcleo del estrano. El ovario difiere de otros órganos que también sintetizan esteroides sexuales, como el testículo y las glándulas suprarrenales, en su dotación enzimática, y por tanto en los productos secretados. • Síntesis de esteroides sexuales en el ovario: el sistema bicelular: Es una explicación lógica de los acontecimientos que intervienen en la esteroidigénesis folicular ovárica; fue propuesta por 1ª vez por Falck en 1959. Para comprenderlo es importante destacar los siguientes puntos: - En las células de la granulosa existen receptores de FSH, cuya síntesis se estimula por la propia FSH. Dra. de la Torre/ Dr. Fontes - 12 - Clases de Residentes 2008 Regulación neurohormonal de la función reproductora - En las células de la teca hay receptores de LH, que inicialmente están ausentes en las células de la granulosa, aunque conforme crece el folículo, la FSH induce la aparición de receptores de LH en las células de la granulosa. - La LH estimula la síntesis de andrógenos en las células de la teca, que pasarán a las células de la granulosa. - La FSH estimula en las células de la granulosa la aromatización de los andrógenos de la teca a estrógenos. - Tanto las células de la teca como las de la granulosa secretan péptidos que actúan como factores autocrinos y paracrinos: Factor de crecimiento insulinoide (IGF): Se secreta en la teca y potencia el estímulo de la LH para la producción de andrógenos en las células de la teca, así como la aromatización mediada por la FHS en la granulosa. Inhibina y activina: Se generan en la granulosa en respuesta a la FSH. La activina potencia las acciones de la FSH, sobre todo la producción de receptores FSH; además inhibe la síntesis de andrógenos en la teca. La inhibina potencia el estímulo de la LH para la síntesis de andrógenos en la teca, a fin de aportar un sustrato para la aromatización a estrógenos en la granulosa. Por tanto, antes de la ovulación, la capa granulosa se caracteriza por su actividad de aromatización y conversión de los andrógenos de la teca en estrógenos, una actividad mediada por la FSH. Dado que la tasa de aromatización en la capa granulosa depende del sustrato de andrógenos que aportan las células de la teca, la secreción de estrógenos por el folículo antes de la ovulación es el resultado de la estimulación combinada de LH y FSH tanto en la teca como en la granulosa. Tras la ovulación, el predominio de la capa granulosa luteinizada depende de la inducción preovulatoria de un número adecuado de receptores de LH, y por consiguiente de una acción adecuada de la FSH. En este momento del ciclo, la capa granulosa secreta estrógenos y progesterona al torrente circulatorio; una actividad mediada por la LH. Dra. de la Torre/ Dr. Fontes - 13 - Clases de Residentes 2008 Regulación neurohormonal de la función reproductora Este sistema bicelular de la síntesis de estrógenos, explica porqué cuando estimulamos los ovarios con FSH pura se produce crecimiento folicular, pero la producción de estrógenos está limitada, debido a una menor concentración de LH. 6. REGULACIÓN DEL CICLO Fase folicular • Fase folicular temprana: Partimos de una serie de folículos que han sido “reclutados” en respuesta a la FSH generada al final del ciclo anterior. Es importante recordar que el desarrollo precoz de los folículos se produce durante los ciclos menstruales previos por un mecanismo independiente de gonadotropinas. Esta cochorte de folículos alcanza un estadio en el que, a menos que sean rescatados por la FSH, sufrirán un proceso de atresia. Este proceso se produce invariablemente y por mecanismos aun desconocidos tanto en ciclos ovulatorios, como en épocas de anovulación, como la infancia, el embarazo o durante la toma de ACO; por eso la paridad, la edad de la menarquia o la toma de ACO no van a influir en la edad de la menopausia de la mujer. Esto también explica que cuando administramos FSH a una mujer para provocar un desarrollo multifolicular, simplemente estamos rescatando de la atresia a toda la cohorte de folículos de ese ciclo, pero no vamos a disminuir la reserva folicular de la mujer, ya que de todas maneras, dichos folículos estaban destinados a la atresia. Una vez que los folículos alcanzan el estadio preantral, la FSH actúa sobre ellos promoviendo la proliferación y diferenciación celular, de modo que el ovocito aumenta de tamaño y se rodea de una membrana (la zona pelúcida), al mismo tiempo que las células de la granulosa sufren una proliferación en múltiples capas y la capa de la teca se va organizando a partir del estroma circundante. En estos folículos se sintetizan principalmente estrógenos. Las concentraciones aún bajas de estrógenos actúan a nivel de la hipófisis estimulando la síntesis y el almacenamiento de gonadotropinas, creando un suministro de gonadotropinas para satisfacer las necesidades del pico a mitad del ciclo. Además aumentan los receptores de GnRH en la hipófisis, para Dra. de la Torre/ Dr. Fontes - 14 - Clases de Residentes 2008 Regulación neurohormonal de la función reproductora preparar a las células gonadotropas para responder posteriormente a la GnRH y producir el pico de FSH y LH a mitad del ciclo. Por otro lado, estas concentraciones bajas de estrógenos van a actuar inhibiendo la secreción de FSH. A pesar de que la concentración de FSH es relativamente baja al comienzo del ciclo, la aparición precoz de estrógenos en el folículo y el aumento de receptores de FSH permiten el desarrollo de un número determinado de folículos, aunque en cualquier momento de este desarrollo, algunos de ellos sufrirán un proceso de atresia. • Selección del folículo dominante: Es el resultado de 2 acciones: - Interacción de los estrógenos y la FSH en el seno del folículo dominante, que es más sensible a la FSH porque tiene un mayor contenido de receptores de FSH (adquiridos por una mayor proliferación de la granulosa). - Efecto de los estrógenos sobre la secreción de FSH: mientras que favorecen la acción de la FSH en el folículo que madura, la retroalimentación negativa sobre la FSH en el eje hipotálamo-hipofisiario sirve para retirar el soporte gonadotrópico de los demás folículos menos desarrollados. El descenso de FSH que se produce alrededor del 5º día del ciclo provoca un declive en la actividad de la aromatasa, limitando la producción de estrógenos de los folículos menos maduros. Es decir, la selección del folículo dominante se establece durante los días 5-7 del ciclo, y por consiguiente, las concentraciones periféricas de estradiol empiezan a aumentar significativamente desde el día 7º del ciclo. Estas concentraciones de estrógenos, derivadas del folículo dominante, aumentan de manera contínua y por retroalimentación negativa ejercen una influencia progresivamente mayor sobre la secreción de FSH. Además, el desarrollo de las células de la granulosa, se acompaña de un desarrollo avanzado de la vascularización de la teca, de modo que hacia el día 9º del ciclo la vascularización del folículo dominante es el doble que la de los Dra. de la Torre/ Dr. Fontes - 15 - Clases de Residentes 2008 Regulación neurohormonal de la función reproductora demás folículos , lo que permite una liberación preferente de gonadotropinas al folículo. Conforme aumentan los estrógenos, la FSH comienza a generar nuevos receptores de LH en las células de la granulosa, que permiten responder al folículo al pico ovulatorio y convertirse posteriormente en un cuerpo lúteo viable. Por otro lado, cuando los niveles de estradiol llegan a una concentración crítica y se mantienen durante un tiempo determinado, su acción inhibidora sobre la secreción hipofisiaria de LH pasa a ser activadora debido a un aumento en la concentración de receptores y de la sensibilidad hipofisiaria a la GnRH. De este modo comienzan a aumentar las concentraciones de LH, lo que estimula la producción de andrógenos en la teca (que servirán de sustrato para la producción de estrógenos) y optimiza la maduración y la función final del folículo dominante. • Fase folicular tardía (folículo preovulatorio): Durante la fase folicular tardía el folículo preovulatorio produce cantidades cada vez mayores de estrógenos, de modo que éstos alcanzan un pico aproximadamente a las 24-36 horas previas a la ovulación. Cuando se alcanza la concentración máxima de estrógenos se inicia el pico de LH, que promueve la luteinización de la granulosa en el folículo dominante, que comienza a secretar progesterona. Esta progesterona en concentraciones bajas, y en presencia de estrógenos ejerce una retroalimentación positiva en la hipófisis facilitando la aparición del pico de LH, y provocando la aparición del pico de FSH a mitad del ciclo, que realiza cambios intrafoliculares importantes necesarios para la expulsión física del ovocito, y garantiza que habrá suficientes receptores de LH para permitir una fase lútea normal y adecuada. En los demás folículos, la LH provoca un incremento de los andrógenos, lo que facilita el proceso de atresia y podría tener un efecto sistémico para estimular la libido en la mujer a mitad del ciclo. Dra. de la Torre/ Dr. Fontes - 16 - Clases de Residentes 2008 Regulación neurohormonal de la función reproductora Ovulación La ovulación se produce unas 10-12 horas después del pico de LH, que debe mantener una concentración máxima al menos durante 14-27 horas para permitir la maduración completa del ovocito. El pico de LH inicia la continuación de la meiosis del ovocito y la síntesis de prostaglandinas, además de favorecer la activación de enzimas proteolíticas, junto con la FSH y la progesterona. Fase lútea Tras la ovulación se produce la luteinización rápida y completa del folículo, que se convierte en cuerpo lúteo. En este momento la esteroidogénesis va a depender de la LH, que promueve la secreción y síntesis de progesterona a partir del colesterol de la sangre transportado por las LDL. Por tanto, en la fase lútea se produce un incremento notable de la concentración de progesterona, que alcanza su pico máximo unos 8 días después del pico de LH, y que en presencia de estrógenos ejerce una acción de retroalimentación negativa sobre la secreción de gonadotropinas, al inhibir los pulsos de GnRH y antagonizar la respuesta de la hipófisis a la GnRH. El inicio de un nuevo crecimiento folicular durante la fase lútea está inhibido por las bajas concentraciones de gonadotropinas por la retroalimentación negativa de los estrógenos, progesterona e inhibina A. Al final de la fase lútea, la degeneración del cuerpo lúteo provoca el descenso en la secreción de esteriodes e inhibina A, lo que provoca un incremento de la FSH unos 2 días antes de la menstruación. Además al desaparecer las altas concentraciones de esteriodes se produce un incremento progresivo de los pulsos de GnRH, que también favorece la secreción de FSH. Este aumento de FSH al final del ciclo es esencial para rescatar de la atresia a un grupo de folículos preparados, lo que permitirá que empiece de nuevo el ciclo y emerja un nuevo folículo dominante. Dra. de la Torre/ Dr. Fontes - 17 - Clases de Residentes 2008 Regulación neurohormonal de la función reproductora 7. BIBLIOGRAFÍA 1. Aznar F, Botija J, Lorente J. Regulación neurohormonal de la función reproductora. El eje diencéfalo-hipófisis-gónadas. En: Fundamentos de Obstetricia. Madrid: SEGO, 2007: 79-84. 2. Kujas M. Ontegenia de las hormonas hipotalamohipofisiarias. EMC (Elservier Masson SAS, París), Gynécologie, 5-A-15, 2007. 3. Martínez Hernández H. Histofisiología del ovario. EMC (Elservier Masson SAS, París), Gynécologie, 30-A-20, 1997. 4. Queenan J T. The Menstrual Cycle. In: Lewis V. Reproductive Endocrinology and Infertiliy. Texas: Landes Bioscience, 2007: 3-9. 5. Remohí Gutierrez J, Martín Valencia Llerena I. Regulación neurohormonal de la función reproductora. El eje diencéfalo-hipófisis- gónadas. En: SEGO. Tratado de Obstetricia, Ginecología y Medicina de la Reproducción. Tomo 1. Madrid: Panamericana, 2003: 6. Speroff L, Fritz M A. Endocrinología Ginecológica Clínica y Esterilidad. 2ª ed. Madrid: Wolters Kluwer Health, 2006. Dra. de la Torre/ Dr. Fontes - 18 -