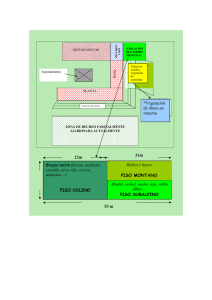
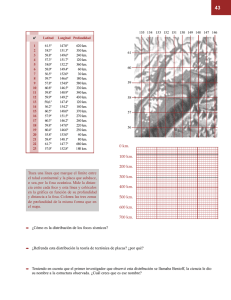
Operaciones oceanográficas en aguas profundas
Anuncio

Operaciones oceanográficas en aguas profundas: los retos del pasado, del presente y del proyecto TALUD en el Pacífico mexicano (1989-2009) Michel E. Hendrickx1 Exploración de las aguas profundas en el mundo Las aguas profundas de los océanos mundiales han sido consideradas como uno de los pocos ambientes naturales de la Tierra que todavía requieren de un fuerte esfuerzo exploratorio. Este ambiente constituye una de las últimas fronteras del planeta Tierra, extraordinariamente vasto, misterioso y de muy difícil acceso. Los océanos cubren un 71 % de la superficie de la Tierra. De estos, 76 % corresponden a aguas con una profundidad de entre 3000 y 6000 m. El océano Pacífico es, en promedio, el más profundo de todos, con una profundidad media de 4282 m. A nivel mundial, es en la plataforma continental, con una profundidad media de 130 m pero que alcanza en algunas zonas casi los 200 m, que se realiza la mayoría de las actividades humanas en los mares y donde el efecto de la contaminación y del deterioro ambiental es más notorio. Sin embargo, corresponde solo a un 9 % de la superficie total de los océanos (Herring y Clarke 1971, Smith 1992, Facebook 2010, Farlex 2010). 1 Laboratorio de Invertebrados Bentónicos. Instituto de Ciencias del Mar y Limnología, Unidad Académica Mazatlán, Universidad Nacional Autónoma de México, Joel Montes Camarena s/n, Mazatlán 82040, Sinaloa, México. Correo-e: [email protected]. 23 Las investigaciones más recientes han podido demostrar que las comunidades de macroinvertebrados de las aguas profundas se caracterizan por una alta diversidad (Grassle 1989, Smith et al. 1998, Zamorano et al. 2007). Este concepto vino a reemplazar la idea muy anclada hasta el siglo XIX de que los fondos marinos, más allá de la zona donde penetra la luz, no eran propicios para la vida, tanto por la ausencia de plantas (i.e., las algas y el fitoplancton) que conforman el primer eslabón de las cadenas alimenticias, como por las fuertes presiones ocasionadas por la columna de agua. Se necesitaron muchos esfuerzos e inversiones en personal científico y en infraestructura para llegar a modificar este concepto, así como aproximadamente 120 años de atención específica y sostenida en la exploración progresiva de estos ambientes para poder acumular una cantidad razonable de información acerca de las comunidades animales que lograron establecerse allí permanentemente (Soule 1970, Gage y Tyler 1992). Los primeros descubrimientos relacionados con la fauna de aguas profundas se remontan al principio del siglo XIX. Sin embargo, estos primeros datos fueron anecdóticos y son el resultado de muestreos incidentales o ligados a las exploraciones de los fondos para la instalación de cables submarinos trans-oceánicos (Soule 1970). Si bien se consiguieron muestras de representantes de los diferentes grupos animales que habitan las zonas batiales (entre 200 y 2000 m de profundidad) y abisales (entre 2000 y 6000 m), estos no fueron adecuadamente registrados y estudiados (Gage y Tyler 1992). La ausencia de luz, aunada a las enormes presiones que existen a grandes profundidades mantuvieron por mucho tiempo la noción de que la vida no era posible en estos ambientes, calificados de “zonas azoicas” (Gage y Tyler 1992). Sin embargo, basándose en los primeros descubrimientos incidentales que demostraban la presencia de vida en aguas profundas, como por ejemplo la colección montada por G.O. Sars a partir de muestras profundas obtenidas en aguas del Fjord de Lofoten, en Noruega, se organizaron dos expediciones a bordo de los buques H.M.S “Lightning” y H.M.S. “Porcupine” (1868-1870) en el Atlántico este. Según Thomson (1873), se efectuaron con éxito 57 arrastres a más de 500 fm (915 m), 16 en más de 1000 fm (1830 m) y dos a profundidades mayores a 2000 fm (3660 m). La profundidad máxima alcanzada fue de más de 4200 m. Las muestras obtenidas revelaron la presencia de una fauna muy particular alrededor de las islas británicas y frente a la península ibérica, en un área de muestreo de aproximadamente 1500 mn de largo por 100-150 mn de ancho (Thomson 1873, Gage y Tyler 1992). Estos hallazgos motivaron, en parte, la planeación del viaje alrededor de la Tierra del buque de investigación H.M.S “Challenger” (Lám. 1 A) de 1872 24 Biodiversidad y comunidades del talud continental del Pacífico mexicano a 1876, que efectuó muestreos en profundidades de hasta 5500 m (Soule 1970). El H.M.S. “Challenger”, construido en Inglaterra y botado el 13 de febrero de 1858, era un barco de tipo “corvette” de 61 m de eslora. En aquella época, y a pesar de parecerse a los galeones españoles del siglo XVIII, los barcos grandes contaban con velas y motores de vapor que les daban seguridad y versatilidad; era el caso del “Challenger”. Curiosamente, este buque participó, en 1862, en las operaciones bélicas de los EE.UU. en contra de México e, incluso, participó en la ocupación de Veracruz. Asignado a la primera campaña de exploración marítima alrededor del mundo, la renombrada “Challenger Expedition”, el H.M.S. “Challenger” fue modificado de lo que era un barco de guerra a un buque de investigación, instalando laboratorios y plataformas de muestreo, así como almacenes para conservar los especímenes capturados. Ya se contaba con termómetros de precisión así como botellas para muestrear agua y dragas para conseguir sedimentos. Entre sus equipos más importantes, tenía a disposición unos 291000 m de cable y redes para muestrear en aguas profundas. Los resultados de esta campaña, consignados en 34 volúmenes, representan todavía hasta hoy la base de muchos estudios taxonómicos de la fauna de aguas profundas a nivel mundial. Lám. 1. A, “Challenger”; B. “Albatross”; C. “Velero IV”; D, batiscafo “FNRS 2”. A B C D O peraciones oceanográficas en aguas profundas 25 Posteriormente, una amplia serie de campañas de vasto alcance fueron organizadas por países europeos o por los EE.UU. con el fin de ampliar los conocimientos acerca de esta fauna profunda. Fueron los cruceros de los barcos “Travailleur” y “Talisman” (Francia), “Hirondelle” y “Princesse Alice” (Mónaco), “Ingolf” (Dinamarca), “Michael Sars” (Noruega), “Valdivia” (Alemania), “Blake” y “Albatross” (EE.UU.), “Albatross” (Suecia; bautizado en honor al ”Albatross” británico) y “Galathea” (Dinamarca). El “Galathea” consiguió las muestras más profundas de la fauna marina recolectadas a la fecha, con la exploración de la fosa de las Filipinas, a 10190 m de profundidad. Esta serie de expediciones, que llevaron los nombres de los barcos en los cuales se realizaron, se iniciaron a mediados del siglo XIX y terminaron en 1952 (Soule 1970, Groueff 1973, Gage y Tyler 1992). De particular interés para México fueron las campañas del “Albatross” de los EE.UU. (Lám. 1 B). A cargo de la Comisión de Pesca del gobierno de los EE.UU. el “Albatross” fue construido en 1882, específicamente para realizar investigaciones acerca de los peces y de las condiciones hidrológicas en las áreas visitadas. A cambio del pago de los costos de operaciones del buque, Alexander Agassiz realizó tres expediciones en el Pacífico, de las cuales la primera (1891) fue la más significativa para la fauna profunda del Pacífico de México (véase infra). Sin embargo, es de señalar que durante la segunda expedición (1899-1900) en el Pacífico sur y centro, se alcanzó la mayor profundidad de arrastre para esta época: 4137 fm (7570 m) en una cuenca del Pacífico central (Soule 1970, Groueff 1973). Otro personaje histórico, Allan Hancock, nacido el 26 de julio 1875 en San Francisco, mostraría durante su vida un particular interés para la investigación científica en el mar. Después de obtener su certificado para operar embarcaciones mayores (lo que le valió el título de “Capitán”), A. Hancock financió expediciones a bordo de la serie de barcos que mando construir, los “Veleros”. Las campañas del “Velero III” fueron probablemente las más importantes para la región que se extiende desde México a Ecuador, pues fueron muchas las campañas de recolecciones de muestras en esta zona y los resultados fueron consignados en una multitud de publicaciones en la serie “Allan Hancock Pacific Expeditions” que son referencias base hasta el día de hoy (e.g., Dawson 1944, Strong y Hertlein 1939, Garth 1958, H.L. Clark 1948). Sin embargo, el “Velero III” estudio esencialmente la flora y la fauna de aguas litorales. A partir de 1948, el “Velero IV” (Lám. 1 C), construido en 1948, efectuó cruceros bajo el mando del propio Capitán Allan Hancock en las costas de California, Baja California y en el golfo de California. Esta embarcación fue una de las pioneras en el uso de fotografía y de equipos específicamente dise26 Biodiversidad y comunidades del talud continental del Pacífico mexicano ñados para operar en aguas profundas, entre otros el “bentoscopo” de O. Barton. La información recabada por el “Velero IV” en aguas profundas ha sido utilizada en una amplia serie de publicaciones (e.g., Parker 1963, Wicksten 1989). La institución “Scripps Institution of Oceanography” de los EE.UU., con base en La Jolla, California, organizó también una serie de cruceros en aguas mexicanas entre 1958 y 1961. Durante estos cruceros, se recolectaron de manera incidental muestras de la fauna de aguas profundas. Solamente dos de estas campañas, la “Baja Slope Expedition” y la “Holt Expedition”, que visitaron las costas de la península de Baja California, tenían como objetivos principales recolectar material biológico, parte del cual fue utilizado por Parker (1963) en su estudio de las comunidades de invertebrados del golfo de California. A partir de los años 50, el polo de interés en la exploración de las aguas profundas se desplazó en la entonces URSS, con la organización de una serie de campañas de investigación de las fosas oceánicas y de diversas partes de los océanos del mundo. Posteriormente, entre 1960 y finales de los años 70, diferentes instituciones de los EE.UU. retomaron cierto liderazgo en la materia, en particular con las expediciones organizadas por la Woods Hole Oceanographic Institution y la Scripps Institution of Oceanography. Con un enfoque más ecológico y adoptando el uso de instrumentos de muestreo más precisos y menos selectivos (i.e., uso de artes permitiendo recolectar un amplio espectro de organismos, desde tallas grandes hasta muy pequeñas), lograron demostrar la existencia de una abundante y muy diversificada fauna en las aguas profundas. De particular interés en esta época fue el uso de redes bien calibradas y de trineos bentónicos confiables, así como de nucleadores de caja que permitían, por primera vez, contar con muestras de sedimentos abundantes y bien estratificadas (Gage y Tyler 1992). En 1946 se fundó el “Institute of Oceanology” de la Academia de Ciencias de la ex-URSS. El primer buque oceanográfico, el “Vitiaz”, un barco re-equipado para la investigación oceánica, tenía una capacidad de 136 tripulantes y una autonomía de 120 días. Comparativamente, el B/O “El Puma de la UNAM tiene una capacidad para 35 personas y cuenta con 30 días de autonomía. El “Vitiaz” contaba con una cadena de anclaje de 11 km para poder inmovilizarse en las aguas más profundas del mundo, una marca jamás igualada desde entonces, y un cabestrante gigantesco, con una capacidad de pesca de hasta 11000 m de profundidad. Además de explorar las aguas territoriales de la URSS, el “Vitiaz” contribuyó al estudio de los océanos Índico (junto con la IIE Expedition), Pacífico y Atlántico entre 1954 y 1979 (Anónimo 2010). Barcos soviéticos participaron O peraciones oceanográficas en aguas profundas 27 muy activamente en los programas IGY-IGC (“International Geophysical Year” y “Year of International Geophysical Cooperation”) en 1957-1959, y en la exploración del océano Índico entre 1960 y 1965, en colaboración con unos 36 barcos de Australia, Francia, Nueva Zelanda, Japón, entre otros (Benson y Rehbock 2002, Britannica 2010). Por otra parte, el uso cada vez más común de métodos de observación directa revolucionó nuestra percepción de los procesos biológicos y ecológicos en ambientes profundos. El sueño del científico −poder observar con sus propios ojos los ambientes profundos y las especies animales que allí viven− estaba por concretarse. En 1930, William Beebe, zoólogo y naturalista nacido en Brooklyn, y Otis Barton, miembro de una familia bien establecida, diseñaron la primera “batisfera”, que consistía en una esfera de grueso metal, sostenida con un largo cable, con escotilla de acceso y ventanilla de observación en cristal reforzado. Esta batisfera se usó para realizar las primeras exploraciones in situ de las grandes profundidades marinas. En 1930, la primera inmersión alcanzó 183 m, en las aguas cercanas a las islas Galápagos. En 1934, la batisfera llegó a unos 908 m de profundidad. En 1948, O. Barton bajó en solitario en una versión modificada de la batisfera, el “bentoscopo”, hasta 1370 m (Wikipedia 2010). El primer “submarino” de exploración en aguas profundas fue concebido en 1937-1939 por Auguste Piccard, de nacionalidad suiza, con el apoyo del “Fonds National pour la Recherche Scientifique” (FNRS) de Bélgica. Profesor en la Université Libre de Bruxelles, Piccard tuvo que interrumpir sus trabajos durante la segunda guerra mundial y retomó su proyecto en 1946. Es hasta 1948 que se realizaron los primeros ensayos de su sumergible frente a la costa de Senegal, con el apoyo de barcos de la marina francesa. El “submarino” utilizado, mejor conocido como “batiscafo” y bautizado FNRS 2 (Lám. 1 D), logró su primera inmersión sin equipaje hasta 1380 m de profundidad. El proyecto fue retomado por la marina francesa y, después de haber recibido varias modificaciones que aumentaban la seguridad del batiscafo, se terminó la construcción del FNRS 3. Sin embargo, debido a múltiples problemas administrativos y de política interna, Piccard fue alejado del proyecto y de la toma de decisiones por lo que buscó otros apoyos. Posteriormente, logró conseguir la ayuda incondicional del gobierno italiano al inicio de los años 50’s y, después de establecerse en la ciudad porteña de Trieste, en el norte del mar Adriático, Piccard puso en marcha la construcción del batiscafo “Trieste” (Lám. 2 A.). Durante los primeros ensayos realizados en Italia, el “Trieste” alcanzó una profundidad de 3150 m. El resto es historia... Con el apoyo de la marina de los EE.UU., 28 Biodiversidad y comunidades del talud continental del Pacífico mexicano el “Trieste” realizó su hundimiento controlado en el “Challenger Deep”, en la fosa de Las Marianas. Alcanzó la profundidad de 10916 m el 26 de enero 1960 (Soule 1970, Groueff 1973). La epopeya del “Trieste” motivó a diversas empresas y centros de investigación a construir sumergibles capaces de explorar las aguas profundas. Los formatos, tamaños y capacidad de inmersión variaron mucho, desde equipos Lam. 2. A, batiscafo “Trieste”; B, submarino de investigación “Denise”; C, “Calypso”; D, submarino de investigación “Alvin”. A B C D O peraciones oceanográficas en aguas profundas 29 limitados a inmersiones hasta un máximo de 100-200 m (e.g., “Star”, “Shelf Diver”, “Benthos”), hasta sumergibles de mayor capacidad (e.g., “Dive Saucer”, “Deep Star”, “Alvin”, “Aluminaut”, “Deep Quest”). Sin lugar a dudas, el campeón absoluto fue el “Archimède”, sin límites teóricos de profundidad (hasta 12000 m, más allá de la profundidad máxima conocida en los océanos: 11524 m, en la fosa de Mindanao). Todos estos sumergibles eran basados en los mismos principios: una esfera o habitáculo de acero o de titanio (con la excepción del “Johnson Sea-Link”, equipado con una esfera de plexiglás de 10 cm de espesor), con pequeñas ventanas de plexiglás muy gruesas, una escotilla de acceso y equipos de control y de comunicación que forman el habitáculo donde pueden permanecer 2-4 personas. La estructura externa consiste de un sistema de flotación no compresible, equipos de filmación y de muestreo, y lastre para el hundimiento. Son pocos los submarinos con tripulación capaces de alcanzar profundidades superiores a los 1000 m con toda seguridad. El “Cyana” francés llegaba hasta 3000 m y el “Alvin” (EE.UU.) hasta 4500 m. Otros fueron diseñados para llegar hasta 6000 m: el “Nautile” (Francia), el “Sea Cliff” de la marina de los EE.UU., los sumergibles rusos “Mir I” y “Mir II” y el japonés “Shinkai 6500” (Soule 1970, Gage y Tyler 1992). Entre estos sumergibles, los más celebres por la enorme publicidad que recibieron en los medios son, sin lugar a dudas, el “Denise”, el “Alvin”, el “Cyana” y el “Ben Franklin”. El “Denise” (Lám. 2 B) era un pequeño submarino con un habitáculo de 2 m de diámetro, con capacidad para dos investigadores que alcanzaba inicialmente hasta 350 m de profundidad. Recorrió el mundo a bordo de la célebre “Calypso” (Lám. 2 C) del Comandante Jacques-Yves Cousteau. En operaciones desde 1959, fue el primer equipo de manejo simple y rutinario que permitió explorar las aguas profundas. Fue sustituido, en 1965, con equipos con un alcance de hasta 500 m (Soule 1970). El “Alvin” (Lám. 2 D.), construido en 1964, fue concebido para alcanzar hasta 4500 m de profundidad con 2-3 miembros de equipaje. Propiedad de la marina de los EE.UU. y operado por la Woods Hole Oceanographic Institution (WHOI), fue utilizado por primera vez en 1965 frente a las Bahamas. Su nombre es derivado de uno de los ingenieros pioneros de WHOI, Allyn Vine, y ha realizado más de 4400 sumersiones en todos los lugares del globo, incluyendo México. Quizás sus acciones más famosas fueron la localización en 1966 de una bomba de hidrógeno perdida frente a España, el descubrimiento de las ventilas hidrotermales en 1970 30 Biodiversidad y comunidades del talud continental del Pacífico mexicano Lam. 3. A, submarino de investigación “Cyana”; B, submarino de investigación “Ben Franklin”; C, sistema de observación submarina por control remoto (ROV, Remote Operated Vehicle); D, “Glomar Challenger”. (A, cortesía del Ifremer/Genavir/A. Bonfiglio). B A C D y su participación en la exploración del naufragio del “Titanic” en 1986 (WHOI 2007). El “Cyana” (Lám. 3 A), propiedad del IFREMER (Francia), ha realizado más de 1300 inmersiones. Con un alcance de hasta 3000 m de profundidad fue puesto en operaciones en 1969 y podía llevar tres miembros de equipaje. Inicialmente conocido como el proyecto SP 3000, del cual el Comandante Jacques-Yves Cousteau fue uno de los instigadores, “Cyana” participó en 1974, junto con el “Archimède” y el “Alvin”, en la expedición “Famous” (“French American Mid Ocean Undersea Survey”) en la dorsal medio-Atlántica, con el fin de explorar y estudiar esta zona de extensión de los fondos del océano. “Cyana” fue retirado de servicio en 2003 (Laubier 2010, IFREMER 2010). El “Ben Franklin” (Lám. 3 B) fue construido en Suiza, en colaboración con la compañía Grumman Aerospace, la misma compañía que construyó el módulo lunar del “Apolo XI” que efectuó el histórico “aterrizaje” en la Luna con Neil Armstrong y Buzz Aldrin en 1969. Con un alcance práctico de unos 700 m de profundidad, O peraciones oceanográficas en aguas profundas 31 su recorrido más famoso lo realizó dentro de la corriente del “Gulf Stream”, donde permaneció a la deriva a una profundidad de unos 200 m (o más), desde Florida hasta el Atlántico NO, viajando un total de 1444 mn (unos 2650 km) en 30 días (Soule 1970). Actualmente existe una amplia serie de submarinos con equipaje capaces de bajar hasta profundidades intermedias y que han sido utilizados para diversas tareas, desde la exploración científica, el manejo industrial y las actividades recreativas. El “Dual Deep Worker 3” (DDW 3), por ejemplo, es un pequeño submarino con capacidad para alcanzar 2000 pies (unos 700 m) de profundidad. Ha sido utilizado ocasionalmente por el equipaje del “Arctic Sunrise” de Greenpeace para monitorear problemas ambientales y por el realizador de películas James Cameron (Farlex 2010). Otro elemento fundamental vendría a agregarse a la exploración de los fondos en aguas profundas: los sistemas de filmación y recolección de material por control remoto. Desde 1966, cuando se perdieron cuatro bombas termonucleares cerca de Palomares, Almería, en las aguas del sudeste de España, se utilizó un aparato robotizado equipado con sonares, cámaras y brazos articulados controlado con un cable desde la superficie para recuperar una de estas. Este equipo, el “CURV” (“Cable Controlled Research Vehicule”) permitía bajar en la columna de agua y hasta el fondo con un sistema de controles a distancia, sin personas a bordo. El CURV fue el precursor de los conocidos “” (“Remote Operated Vehicle”) (Lám. 3 C), de uso ya muy común en las operaciones de exploraciones oceanográficas. Son “ROV” similares que sirvieron para explorar los restos del “Titanic” y para reparar y colmatar la fuga de petróleo de la plataforma Deepwater Horizon, administrada por la British Petroleum en el golfo de México, en abril de 2010. En el Pacífico mexicano, se han utilizado este tipo de equipos para explorar las chimeneas hidrotermales. Además de sus cámaras de registro de imágenes, están equipadas de brazos, redes, trampas, palas, tubos de aspiración y recipientes para recolectar muestras del fondo y especímenes vivos. Equipos más modernos llamados “ROPOS” (“Remotely Operated Platform for Ocean Science”) son usados para recolectar y almacenar informaciones oceanográficas, al igual que las unidades REMUS (“Remote Environmental Monitoring Units”). Todas están conectadas al barco nodriza mediante un cable de control y de transmisión instantánea de datos e imágenes. La tecnología moderna está actualmente orientada hacia el uso de equipos autónomos, o “AUV” (“Autonomous Underwater Vehicle”), que pueden permanecer en el agua por meses, moverse de manera autónoma usando sus com32 Biodiversidad y comunidades del talud continental del Pacífico mexicano putadoras, almacenar datos y detectar modificaciones en el medio ambiente. Una vez concluido su ciclo de actividades, algunos son capaces de regresar a su base de manera autónoma (Alden 2010). Más adelante en el siglo XX, se organizaron varios programas de muestreo internacionales y multidisciplinarios, cuyos objetivos principales eran elucidar las relaciones entre el medio ambiente y las comunidades existentes, así como analizar la dinámica entre estas comunidades y los flujos de materia orgánica desde la superficie, fuentes esenciales para el balance energético de las especies que no pueden contar con los procesos de producción primaria (i.e., fotosíntesis de las micro y macro algas) en aguas desprovistas de luz. Algunos de estos programas siguen vigentes. El “Glomar Challenger” (Lám. 3 D) emprendió entre 1968 y 1983 el mayor reconocimiento de los fondos a nivel del globo, realizando perforaciones en el sedimento de más de un kilómetro en profundidades mayores a 6700 m (Soule 1970). La colección de núcleos de sedimentos obtenidos por este barco es inigualada hasta la fecha. Las campañas MUSORSTOM fueron organizadas por Francia en el océano Pacifico oeste, y dieron lugar a la publicación de una muy extensa serie de trabajos taxonómicos sobre virtualmente todos los grupos de invertebrados y peces de aguas profundas de esta región (e.g., Forest 1985, Hanley y Burke 1991, Vadon 1991, Chan y Crosnier 1997, Galil 1997, McCosker 1999). Las aguas profundas del Pacífico de México A pesar de todas estas actividades a nivel internacional, hasta recientemente no se contaba con un programa global y bien organizado de exploración sistemática de las aguas profundas del Pacífico mexicano. Este hecho era evidentemente contradictorio con la importancia económica y ecológica de los múltiples ambientes en la región. El Pacífico de México se extiende desde la frontera oeste de México con los Estados Unidos de América del Norte (EE.UU.) hasta la frontera con Guatemala (incluyendo el golfo de California). Incluyendo ambas costas del continente americano , México cuenta con una plataforma continental que cubre unos 393000 km2 (Wri-Org 2010). Según Hendrickx (1993), considerando la totalidad de la superficie de la Zona Económica Exclusiva (ZEE) del Pacífico de México (un total de 2364200 km2), el área que corresponde a la plataforma continental (profundidades inferiores a 200 m), donde se realizan la enorme mayoría de las actividades pesqueras y de exploración, cubre solamente 154300 km2, o sea un 6.5 % de O peraciones oceanográficas en aguas profundas 33 esta superficie total. Las zonas con profundidades de 200 a 2000 m cubren unos 288900 km2 (12.2 %), pero aquellas con profundidades >2000 m totalizan unos 1921000 km2 (81.3 %), lo cual permite evaluar la importancia de los ecosistemas de aguas profundas en el país. A pesar de ello, la exploración de las aguas profundas del Pacífico mexicano (en particular las áreas tropicales y subtropicales) por parte de grupos de investigación nacionales, hasta fechas recientes, ha sido incidental o inexistente; así lo refleja la literatura disponible. Las razones son esencialmente: 1) la dificultad de muestrear en alta mar, y más aún en aguas oceánicas, donde las profundidades rebasan rápidamente los 1000 m y las operaciones de muestreo son difíciles y largas; 2) el alto costo para mover una embarcación de mediano o gran calado en aguas oceánicas, en particular considerando las distancias que se deben recorrer para cubrir la ZEE. Sin embargo, resulta incongruente que ni siquiera la parte más accesible de estas aguas profundas, en la porción central y sur del golfo de California, haya sido explorada adecuadamente. La accesibilidad a los ambientes naturales en aguas profundas es extremadamente complicada y el nivel de complejidad aumenta drásticamente con el aumento de la profundidad. La presión del agua aumenta de una atmósfera cada 10 m. Si bien las estructuras metálicas tales como las dragas o los trineos equipados de redes no sufren daños debido a la presión, otros equipos son muy sensibles a estas variaciones (e.g., los medidores de parámetros dentro del agua). Por otro lado, el bajar un equipo de muestreo de 100 kg de peso a 2000 m de profundidad con un cable de acero de 2 km de longitud y 1 cm de espesor equivale a bajar un peso de un kilogramo desde lo alto de un edificio de 100 m de altura amarrándolo a un hilo de medio milímetro de grosor. Por si fuera poco, las corrientes superficiales y de fondo, la deriva de la embarcación por el viento y los movimientos del oleaje hacen todavía más difíciles las operaciones de muestreo. Por los mismos motivos, las operaciones en alta mar deben realizarse en embarcaciones grandes, capaces de resistir los posibles embates de las marejadas o de las tormentas que pueden desatarse repentinamente. Para evitar percances y pérdidas de tiempo en alta mar por averías, los equipos que se usan para muestrear en aguas profundas deben ser muy resistentes, robustos, relativamente pesados y de despliegue rápido. Evitar problemas técnicos o descomposturas es primordial, pues el tiempo de uso de los buques grandes está limitado por el alto costo de operación de los mismos: desde 5000 hasta 50000 USD o más por día de operación. Para aprovechar mejor el tiempo de barco, es muy recomendable adoptar una estrategia multidisciplinaria, 34 Biodiversidad y comunidades del talud continental del Pacífico mexicano formar grupos de trabajo con objetivos comunes o, dentro de lo posible, compartir las campañas con colegas con interés distintos en la misma área de muestreo. Si bien el H.M.S. “Challenger”, pionero de la investigación en aguas profundas, visitó parte de la costa del Pacífico americano, no llegó hasta México. Su zona de muestreo en el Pacífico Este se limitó a las aguas de Chile, entre el continente y las islas Juan Fernández (Wicksten 1989). En 1875, el barco de investigaciones U.S. “Narragansett” visitó la costa oeste de México, pero no muestreó en aguas profundas. Para tener las primeras informaciones acerca de la fauna profunda del Pacífico de México, se tendría que esperar la llegada del “Albatross” de los EE.UU. que visitó la costa del Pacífico Este desde las islas Galápagos hasta Guaymas, en la parte central del golfo de California, en 1891 (véase Faxon 1895). El material recolectado por esta expedición dio como resultado la publicación de una serie de monografías editadas por el “Museum of Comparative Zoology” de Harvard, EE.UU. Estos trabajos representan todavía hoy en día la base para el estudio de la fauna de aguas profundas del Pacífico este tropical, particularmente debido a los muestreos realizados entre las islas Galápagos y Ecuador, frente a Panamá-Colombia, alrededor Fig. 1. Derrotero del “Albatross” entre Ecuador y México en 1891 (adaptado de Faxon, 1895). O peraciones oceanográficas en aguas profundas 35 de las islas Malpelo y del Coco (“Cocos Island”), y en el golfo de California surcentro (Fig. 1). Entre estos trabajos destacan el estudio de los moluscos (e.g., Dall 1908), de los cangrejos, de los camarones y de otros macrocrustáceos (e.g., Faxon 1893, 1895), de las estrellas de mar (e.g., Ludwig 1905), de las ofiuras (e.g., Lutken y Mortensen 1899), de los erizos (e.g., A. Agassiz 1898), de los pepinos de mar (e.g., Ludwig 1894) y de los peces (e.g., Garman 1899). Las embarcaciones de la serie “Velero”, que operaron en aguas mexicanas bajo el mando del Capitán Allan Hancock, trabajaron sobre todo la zona litoral. El “Velero IV” exploró aguas tanto someras como profundas y efectuó algunas incursiones en la costa de Baja California y en el golfo de California a partir de 1948, año en el cual inició sus operaciones. La información relacionada con estos muestreos está muy dispersa en la literatura. De manera incidental, otros barcos de instituciones norteamericanas (e.g., “T. Washington”, “Orca”, “E. W. Scripps”, “Horizon”, “Agassiz”, “S.F. Baird”, “Calafia”) también han visitado las costas de Baja California y del golfo de California entre 1957 y 1972. Han operado diversos tipos de artes en aguas profundas (dragas de penetración y de arrastre, nucleadores, trampas de control remoto) con la subsecuente captura de especímenes hasta profundidades de más de 3000 m (véase Parker 1963, Luke 1977, SIO 1992, Hendrickx 2012a, 2012b). Otras embarcaciones que operaron de manera más sostenida en aguas profundas del Pacífico mexicano entre 1958 y 1960 participaron en dos expediciones: la “Baja Slope Expedition” y la “Holt Expedition”, ambas patrocinadas por la Scripps Institution of Oceanography (Parker 1963). Algunos intentos de muestreo exploratorio en la parte norte-central del golfo de California se realizaron en 1971-1972, esencialmente para determinar la presencia de bancos de peces [e.g., Merluccius sp. (Mathews et al. 1974)]. Por otro lado, algunos investigadores nacionales y extranjeros han mantenido una fuerte actividad exploratoria en las zonas de chimeneas hidrotermales (i.e., en la cuenca de Guaymas, golfo de California; en la zona del 21º N, “dorsal o cresta del Pacífico este”) a bordo de submarinos de los EE.UU. (véase Blake 1985, Escobar-Briones y Soto 1993). Por su parte, Solís-Weiss y Hilbig (1992) y Solís-Weiss y HernándezAlcántara (1994), por ejemplo, estudiaron poliquetos recolectados entre 2000 y 2020 m de profundidad por el “Alvin” en 1988 en la cuenca de Guaymas, en el golfo de California. El reciente interés por descubrir nuevos recursos pesqueros más allá del límite de la plataforma continental ha motivado la organización de proyectos de pesca 36 Biodiversidad y comunidades del talud continental del Pacífico mexicano exploratoria en diversos países, por ejemplo en Perú (Vélez et al. 1992, Kameya et al. 1997), en Chile (e.g., Retamal 1993, Arana y Vega 2000, Arana 2003), en El Salvador (Hendrickx y López 2006) y en Costa Rica (e.g., Wehrtmann y Nielsen-Muñoz 2009, Wehrtmann et al. 2010).). Sin embargo, en el Pacífico de México, hasta la fecha, las investigaciones han sido limitadas y muy desproporcionadas comparadas con la extensión de aguas profundas que encontramos frente a las costas del país. La zona del mínimo de oxígeno (ZMO) Una parte significativa de las aguas profundas del mundo se caracterizan por las muy bajas concentraciones de oxígeno disponible. Estas áreas, conocidas como las zonas del mínimo de oxígeno (ZMO) son extremadamente hostiles para la vida marina y, donde interceptan la pendiente del talud continental, dan origen a franjas bentónicas anóxicas o severamente hipóxicas donde la riqueza en especies baja drásticamente. En aguas todavía más profundas, el oxígeno se recupera y las comunidades que allí se encuentran son ricas en especies, a veces muy abundantes. En estos ecosistemas, las concentraciones de oxígeno y la profundidad son factores muy importantes para la supervivencia de las especies, las cuales deben contar con adaptaciones muy especificas para poder sobrevivir y reproducirse (Díaz y Rosenberg 1995, Levin y Gage 1998, Rogers 2000, Levin et al. 2001, McClain y Rex 2001, McClain 2004). La ZMO del Pacífico este es la más importante a nivel mundial y ha sido detectada desde el golfo de California central hasta la porción central de Chile, y hasta los 50° N en el Pacífico NE (Díaz y Rosenberg 1995). Representa un ambiente muy adverso para la fauna, tanto en el ámbito pelágico como en el bentónico. A nivel del fondo, la ZMO actúa como barrera fisiológica entre la fauna de la plataforma y del talud. En México, abarca la totalidad del País, salvo el extremo norte del golfo de California (véase Díaz y Rosenberg 1995, Helly y Levin 2004). Es extremadamente amplia en el sur del golfo de California y a lo largo de la costa SO de México (Hendrickx 2001, Hendrickx y Serrano 2007, Serrano 2012). Este fenómeno tiene consecuencias muy negativas sobre el ambiente marino, ya que limita la anchura y (sobre todo) la profundidad de la franja costera en la cual los organismos no adaptados a concentraciones de oxígeno reducidas o extremadamente bajas logran vivir (véase Childress y Seibel 1998, Rogers 2000, Levin 2003, Hendrickx y Serrano 2010). A pesar de su importancia sobre la disponibilidad de los recursos marinos en el Pacífico mexicano, O peraciones oceanográficas en aguas profundas 37 poco se sabe acerca de la ubicación exacta (latitudinal y batimétrica), la amplitud y las variaciones anuales o interanuales de la ZMO en el área. Entre las pocas evaluaciones de la extensión de este fenómeno en aguas mexicanas, existe un trabajo recapitulativo publicado a principios de los 60 por Parker (1963), quien utilizó toda la información disponible en esta época para proponer un mapa de distribución de la ZMO en el golfo de California. La información publicada posteriormente se refiere esencialmente a datos aislados (e.g., hidrocalas, algunos perfiles o transectos, mediciones en algunas localidades) (e.g., Gaxiola-Castro et al. 2002, Sánchez-Velasco et al. 2004) y no hay un estudio integral o sostenido en la columna de agua a lo largo de la ZEE en México, en áreas suficientemente amplias para ser consideradas representativas del nivel de influencia de este fenómeno sobre la biología y la distribución de las especies en la región (Hendrickx y Serrano 2010). Aún la información recabada durante el proyecto TALUD, por mas amplia que sea, es insuficiente para poder entender con claridad este fenómeno en las aguas mexicanas (véase Serrano 2012). El proyecto TALUD Los recursos pesqueros de la plataforma continental y de las aguas oceánicas superficiales del Pacífico mexicano son relativamente bien conocidos. En el golfo de California, a pesar de caracterizarse por una diversidad faunística muy alta que alcanza más de 6,000 especies macro-bentónicas y macro-pelágicas (Brusca et al. 2005), las pesquerías se limitan esencialmente a las aguas más someras (e.g., pesca de camarones Penaeidae, recolección de moluscos, pesca de escama). En el Pacífico SO se vive una situación parecida, aunque localmente las condiciones batimétricas (i.e., plataforma muy angosta, fondos rocosos en aguas someras) dificultan hasta cierto punto la explotación de los recursos. A pesar de eso, ciertas áreas del SO de México están pobladas por comunidades ricas y abundantes que ofrecen cierto potencial pesquero para las comunidades locales (Hendrickx et al. 1997, Godínez-Domínguez y González-Sansón 1998, 1999, Arciniega-Flores et al. 1998, 2008). En el noroeste del Pacífico mexicano, en el ámbito pelágico, además de la pesca deportiva existe una flota atunera muy activa que cuenta con aproximadamente 137 barcos (CIAT 2010). La pesquería de los atunes en México es una de las mejor establecidas, con una captura anual en 2009 de 131621 toneladas, casi exclusivamente en el Pacífico (SAGARPA 2010). Los progresos técnicos realizados recientemente en el manejo de redes de pesca en aguas profundas (i.e., por debajo de los 400 m de profundidad), progresos 38 Biodiversidad y comunidades del talud continental del Pacífico mexicano motivados en gran parte por la escasez o la insuficiencia de recursos para la pesca tradicional en aguas someras (i.e., hasta los 100-150 m), han dado como resultado la explotación de recursos antes considerados de difícil acceso. En el Mediterráneo, por ejemplo, después de haber realizado las investigaciones pertinentes, la pesca de Aristeus antennatus (Risso, 1816) y de Nephrops norvegicus (Linnaeus, 1758) a más de 400 m se ha vuelto rutinaria (Sardà y Demestre 1987, Cartes y Sardà 1989, Sardà y Valladares 1990). Investigaciones más recientes indican que muchas otras especies comparten este hábitat (Cartes et al. 1994); muchos géneros registrados (e.g., Gennadas, Solenocera, Acanthephyra, Pasiphaea, Nematocarcinus, Plesionika) tienen uno o varios representantes en aguas del golfo de California y del Pacífico mexicano tropical (Hendrickx y Estrada-Navarrete 1996, Wicksten y Hendrickx 2003). Frente a Perú, los arrastres exploratorios han dado excelentes resultados, con el descubrimiento de áreas propicias para la pesca de camarones de aguas profundas tales como Heterocarpus vicarius Faxon, 1893, H. hostilis Faxon, 1893, Nematocarcinus agassizii Faxon, 1893, Haliporoides diomedeae (Faxon, 1893), Plesionika trispinus Squires y Barragán, 1976, Psathyrocaris fragilis Wood-Mason, 1893 y Pasiphaea magna Faxon, 1893 (Kameya et al. 1997). Nuevamente, varias de estas especies se encuentran distribuidas hasta el golfo de California sur o central (Wicksten y Hendrickx 1992, Hendrickx 1995a, 2001, 2003a, 2004, Hendrickx y Estrada Navarrete 1996). Existe, en consecuencia, una verdadera necesidad de explorar estos fondos para obtener, en conjunto con información básica acerca de la composición y de la dinámica de las comunidades naturales, información acerca de un posible potencial pesquero. Por otro lado, varias instituciones dedicadas a la investigación en el mar y en los océanos han desarrollado instrumentos de medición o de muestreo adaptados a las condiciones adversas encontradas en aguas profundas. En particular, se desarrollaron diferentes modelos de trineos bentónicos, aptos a recoger la macrofauna que vive apoyada sobre el sustrato, redes de media agua con capacidad hasta 400-500 m de profundidad o más (King y Iversen 1962, Guennegan y Martin 1985), así como instrumentos que penetran en el sedimento y logran traer hasta la superficie trozos del fondo marino poco o nada perturbados: los nucleadores de caja (Sorbe 1983, Gage y Tyler 1992). En la columna de agua, sistemas equipados de sensores colocados en una estructura de titanio (CTD, “Conductivity-Temperature-Density”), a veces equipados con sensores para medir las concentraciones de oxígeno disuelto o el pH, están diseñados para realizar mediciones continuas hasta varios miles de metros de profundidad (Idronaut 2010). O peraciones oceanográficas en aguas profundas 39 Los objetivos del proyecto TALUD, el cual inició en 1989, fueron: 1) determinar la composición faunística a nivel específico de los invertebrados y peces batiales en profundidades mayores a 200 m, con especial énfasis en la fauna localizada por debajo de la ZMO y en el interfase “zona anóxica-zona hipóxica”; 2) determinar la composición de los sedimentos en la franja batial explorada y medir las características hidrologicas básicas (i.e., oxígeno disuelto, temperatura y salinidad) en la columna de agua, con énfasis en las condiciones predominantes en el ambiente epibentónico; 3) poner especial énfasis en las mediciones de oxigenación con el fin de establecer un modelo de distribución de este parámetro en la zona de muestreo y extrapolar los resultados a la porción tropical-subtropical del Pacífico mexicano; 4) determinar la composición faunística a nivel de grupos de la meiofauna presente en las muestras de sedimentos y estudiar detalladamente los copépodos Harpacticoidea; 5) analizar la presencia de copépodos parásitos en las muestras de peces recolectados de zonas profundas y proceder a su análisis taxonómico; 6) estudiar la fauna epi y endobentónica de invertebrados (principalmente los poliquetos, moluscos, crustáceos y equinodermos) asociados con las aguas profundas y analizar la composición de las comunidades acorde con los parámetros ambientales; 7) estudiar las comunidades de crustáceos pelágicos presentes entre la superficie y hasta 1500 m de profundidad; 8) estudiar la fauna de peces asociados con las aguas profundas y analizar la composición de las comunidades acorde con los parámetros ambientales. Como parte de los resultados de los cruceros TALUD, se obtuvieron indicadores claros de que, tanto en el golfo de California como a lo largo de la costa del SO de México (Jalisco hasta Chiapas), la franja costera somera donde existen especies con potencial pesquero es extremadamente reducida debido a la influencia de la ZMO. La magnitud de este fenómeno es tan fuerte que, en algunas zonas, la ZMO ha sido detectada en aguas someras, a partir de 50 m de profundidad (Hendrickx y Serrano 2007). Por lo anterior, el proyecto se enfocó muy rápidamente al estudio de las comunidades que se ubican por debajo de esta ZMO, en particular en el umbral donde se inicia la recuperación del oxígeno disuelto. Materiales y métodos Con una eslora de 50 m, un desplazamiento de 1058 tm y una velocidad de crucero de 12.5 nudos, el B/O “El Puma” fue el primer buque mexicano dedicado 100 % a la investigación oceanográfica. Puede llevar, además de la tripulación de 15 personas, un total de 20 científicos acomodados en confortables cabinas. 40 Biodiversidad y comunidades del talud continental del Pacífico mexicano Cuenta con una serie de cabestrantes para realizar diversas operaciones (e.g., pesca, hidrocalas, núcleos, dragados, sondeos, arrastres pequeños), tres plataformas hidráulicas de abatimiento ajustables, una grúa hidráulica de 12 ton de capacidad y un estructura de tipo “unigan” de ocho ton de capacidad en la popa. En total cuenta con cuatro laboratorios, una biblioteca, una sala de juntas, dos congeladores de gran capacidad y dos cuartos de preparación de equipos y recuperación de muestras (Figs. 2, 3). Fig. 2. Vista lateral del B/O “El Puma”. (http://www.buques.unam.mx/planos_ PUMA.htm) Fig. 3. Plano de las cubiertas de botes y de abrigo donde se encuentran todas las plataformas, los puestos de trabajo y los laboratorios para las operaciones oceanográficas. O peraciones oceanográficas en aguas profundas 41 Desde 1981, después de la llegada a México del primer buque oceanográfico de la UNAM, el B/O “El Puma” (Lám. 4 A), el Laboratorio de Invertebrados Bentónicos (LIB) inició una amplia serie de cruceros oceanográficos orientados hacia el estudio de las comunidades bentónicas y pelágicas en la plataforma continental del Pacífico mexicano tropical y subtropical. Estas campañas fueron enfocadas, en primer termino, al estudio de la plataforma de Sinaloa, en el SE del golfo de California, con los cruceros SIPCO I, II y III (1981-82). Posteriormente, se organizaron tres campañas que cubrieron la totalidad del golfo de California (CORTES 1, 2 y 3; 1982 y 1985) con el apoyo parcial del CONACYT (proyecto PCCBBNA-021996) (Hendrickx y Salgado-Barragán 1992). En estas campañas se contó con la participación de académicos del Instituto de Ciencias del Mar y Limnología y de la Facultad de Ciencias de la UNAM. Además, el grupo de académicos del LIB participó de manera ocasional o activamente en otras campañas dirigidas por otros científicos o bien estudiaron material recolectado por otros proyectos: (e.g., campañas BIOCAPESS frente a Sinaloa, en 1991-92; campañas GUAYTEC en el golfo de California central, en 1987; campañas CEEMEX frente a las costas de Sinaloa, en 1990-1991, y en el golfo de Tehuantepec, en 19911992 (véase Hendrickx 1990, Hendrickx y Salgado-Barragán 1994, Hendrickx et al. 1997, Hendrickx y Vázquez-Cureño 1998). Sin ninguna excepción, el material biológico y los datos ambientales recolectados u obtenidos a lo largo de estas campañas oceanográficas, dieron como resultado, por lo menos, una publicación por campaña por parte del propio LIB (i.e., SIPCO, 6 publicaciones; CORTES, 21 publicaciones; GUAYTEC, 2 publicaciones; CEEMEX, 3 publicaciones; BIOCAPESS, 1 publicación). Además, 14 artículos científicos y cuatro libros resultaron de la síntesis de información de varias de estas campañas. La participación de grupos de investigación asociados en las campañas organizadas por el mismo LIB o el acceso, por parte de otros grupos, a material recolectado durante estas campañas, ha generado una lista impresionante de contribuciones que todavía sigue creciendo (e.g., Caso 1986, HernándezAlcántara y Solís-Weiss 1991, 1993a, 1993b, 1994, 1998a, 1998b, Licea et al. 1995, Moreno et al. 1996). Esta amplia experiencia en la organización de actividades en alta mar y en el manejo de operaciones a bordo de “El Puma” fue aprovechada para la planeación, la organización y la realización de las campañas TALUD, entre 1989 y 2009. Todas las operaciones realizadas durante el proyecto TALUD fueron efectuadas a bordo del B/O “El Puma”, de la Universidad Nacional Autónoma de México 42 Biodiversidad y comunidades del talud continental del Pacífico mexicano Lam. 4. A, B/O “El Puma”; B, mismo, vista de popa a proa indicándose el cabrestante geológico de uso múltiple (w) y el sistema de tambores para redes (tr); C, mismo, vista hacia la popa. Las flechas indican el winche del CTD (w), la plataforma hidrológica con su pescante (ph), los tambores para redes (tr) y el unigan de pesca de popa (u); D, mismo, unigan de pesca de popa. A C B D (UNAM). Hasta la fecha se han efectuado en total 13 campañas. De éstas, los TALUD I-III se realizaron cuando B/O “El Puma” estaba todavía bajo la responsabilidad del Instituto de Ciencias del Mar y Limnología (ICMyL). En las campañas TALUD IV-XIII, el buque ya había pasado bajo la responsabilidad de la Coordinación de la Investigación Científica de la misma UNAM. El uso del B/O “El Puma” como plataforma de trabajo resultó indispensable para el éxito del proyecto TALUD. En efecto, los muestreos en aguas profundas reO peraciones oceanográficas en aguas profundas 43 quieren de una embarcación grande, con equipos de muestreo que permitan contar con cables resistentes y suficientemente largos para alcanzar el nivel del fondo y resistir las fuertes tracciones ocasionadas por los equipos durante los muestreos, ya sea a nivel del fondo o dentro de la columna de agua. Durante las campañas TALUD se necesitó, por ejemplo, soltar cerca de 5000 m de cable para poder efectuar muestreos de la fauna bentónica a profundidades del orden de 2350 m. Este tipo de equipamiento no se encuentra en embarcaciones pequeñas. Campañas del proyecto TALUD En el contexto del proyecto TALUD, se realizaron tres campañas preliminares en la franja del talud continental del golfo de California por parte del LIB entre 1989 y 1991. Posteriormente, con el apoyo de CONACYT (proyecto 31805-N), se realizaron las campañas oceanográficas TALUD IV-VII (2000-2001). Las campañas TALUD VIII-XII (2005-2008) recibieron apoyo parcial del PAPIIT (PAPIIT, DGAPA, UNAM, proyecto IN 217306-3). Estas actividades permitieron obtener la serie de Fig. 4. Posición de las estaciones de muestreo visitadas durante: A. las campañas TALUD I-III; B. la campaña TALUD IV; C. la campaña TALUD V; D. la campaña TALUD VI. 44 A B C D Biodiversidad y comunidades del talud continental del Pacífico mexicano O peraciones oceanográficas en aguas profundas 45 1 2 3 4 5 6 7 8 9 1A 1B 1C 6 7 8 Est. TALUD I TALUD I TALUD I TALUD I TALUD I TALUD I Crucero TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II Est. Crucero 23-mar-90 24-mar-90 24-mar-90 24-mar-90 23-mar-90 24-mar-90 24-mar-90 24-mar-90 25-mar-90 13-dic-89 13-dic-89 13-dic-89 12-dic-89 12-dic-89 12-dic-89 Fecha Fecha 23º03'36" 22º59'36" 22º58'12" 22º56'24" 22º55'54" 22º55'12" 22º54'18" 22º52'42" 22º22'48" 22º54'00" 22º53'00" 22º52'12" 23º15'54" 23º30'42" 23º33'00" Lat. N Lat. N 106º25'48" 106º26'18" 106º30'12" 106º31'24" 106º32'30" 106º34'00" 106º35'12" 106º35'36" 106º07'12" 106º30'00" 106º31'00" 106º31'54" 107º31'12" 107º09'01" 107º14'00" Long. O Long. O No No No No No No No No No No No No No No No Karling Karling Tipo de arrastre Red 80' Red 80' Red 80' Zoopl Red 80' Red 80' Tipo de arrastre Red 80' Red 80' Red 80' Red 80' Agassiz Agassiz Agassiz Agassiz Red 80' Prof. de arrastre (m) 170 220-230 310 1550 220 130 Prof. de arrastre (m) 61 90 133 162 285 380 495 565 55 Prof. núcleos (m) No No No No No No Prof. núcleos (m) No No No No No No No No No 2.0 0.35 0.30 0.25 0.16 0.44 0.23 0.23 ND ND ND ND ND ND ND Oxígeno (ml l-1) Oxígeno (ml l-1) Cuadro 1. Estaciones de muestreo visitadas durante las campañas TALUD I-XIII entre 1989 y 2009. Se incluye la fecha, la posición y las actividades principales desarrolladas en cada estación. ND, no hay datos; K, draga Karling; IK, red de media agua Isaac Kids; H, hidrocala; MN, micronecton; No, no se realizó la operación. Cuadro 1. Continúa. Est. 10 11 12 13 14 14 15 17 19 20A 20B 21 22 24 26 27A 27B 28 29 30 Crucero TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II 46 Biodiversidad y comunidades del talud continental del Pacífico mexicano 25-mar-90 25-mar-90 25-mar-90 25-mar-90 24-mar-90 24-mar-90 25-mar-90 26-mar-90 26-mar-90 26-mar-90 26-mar-90 26-mar-90 25-mar-90 25-mar-90 27-mar-90 27-mar-90 27-mar-90 27-mar-90 27-mar-90 27-mar-90 Fecha 22º33'30" 22º22'36" 22º25'30" 22º24'24" 22º25'36" 22º25'36" 22º23'48" 21º40'48" 21º37'42" 21º47'07" 21º47'07" 21º48'42" 21º48'24" 21º50'36" 23º21'54" 24º05'06" 24º11'42" 24º00'00" 23º59'12" 23º59'54" Lat. N 106º13'00" 106º17'30" 106º18'30" 106º19'54" 106º21'12" 106º21'12" 106º22'30" 105º57'54" 106º12'00" 106º17'36" 106º17'36" 106º19'24" 106º22'00" 106º23'42" 107º10'00" 107º40'36" 107º35'24" 107º40'04" 107º44'12" 107º46'06" Long. O No No No No No No No No No No No No No No No No No No No No Karling Tipo de arrastre Red 80' Red 80' Red 80' Agassiz Agassiz IK Agassiz No No Red 80' Agassiz Agassiz Agassiz IK No Red 80' Red 80' Red 80' Red 80' Agassiz Prof. de arrastre (m) 82 130 188 282 370 370 465 ND ND 167 167 280 416 500 ND 64 39 90 130 225 Prof. núcleos (m) No No No No No No No H H No No No No No H No No No No No 0.90 0.34 0.10 ND 0.35 0.35 0.18 ND ND 0.25 0.25 0.13 0.23 0.14 ND 2.50 4.20 0.70 0.41 0.34 Oxígeno (ml l-1) O peraciones oceanográficas en aguas profundas 47 Est. 31 32 33 34 35 36 38A 38B 39 40A 40B 41 42 43 44 46 47 48 Crucero TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II TALUD II Cuadro 1. Continúa. 27-mar-90 28-mar-90 28-mar-90 28-mar-90 28-mar-90 30-mar-90 28-mar-90 28-mar-90 28-mar-90 28-mar-90 28-mar-90 29-mar-90 29-mar-90 29-mar-90 29-mar-90 29-mar-90 30-mar-90 30-mar-90 Fecha 23º59'18" 23º58'42" 23º58'06" 23º59'12" 23º54'12" 24º21'54" 24º55'54" 24º57'24" 24º54'42" 24º54'54" 24º54'00" 24º53'36" 24º51'00" 24º48'06" 24º46'00" 24º38'24" 23º41'42" 23º04'54" Lat. N 107º49'30" 107º52'36" 107º54'36" 107º55'36" 107º55'48" 108º13'00" 108º36'30" 108º33'42" 108º39'30" 108º39'42" 108º40'09" 108º42'00" 108º43'30" 108º43'30" 108º42'30" 108º27'00" 107º33'06" 107º28'36" Long. O No No No No No No No No No No No No No No No No No No Karling Tipo de arrastre Agassiz Agassiz No No No No Agassiz Agassiz Red 80' Red 80' No Red 80' Agassiz Agassiz Agassiz No No No Prof. de arrastre (m) 272 380 ND ND ND ND 64 40 94 131 ND 215 300 371 550 ND ND ND Prof. núcleos (m) No No H H H H No No No No H No No No No H H H 0.20 0.20 ND ND ND ND 2.80 3.00 2.50 ND ND 0.30 0.18 0.18 0.14 ND ND ND Oxígeno (ml l-1) Cuadro 1. Continúa. Est. 2 3A 3B 5 6 9 10A 10B 10 13 14 14A 14B 15 16A 16B 17 19 19B 20A Crucero TALUD III TALUD III TALUD III TALUD III TALUD III TALUD III TALUD III TALUD III TALUD III TALUD III TALUD III TALUD III TALUD III TALUD III TALUD III TALUD III TALUD III TALUD III TALUD III TALUD III 48 Biodiversidad y comunidades del talud continental del Pacífico mexicano 17-ago-91 17-ago-91 17-ago-91 24-ago-91 18-ago-91 18-ago-91 18-ago-91 18-ago-91 18-ago-91 18-ago-91 19-ago-91 19-ago-91 19-ago-91 20-ago-91 20-ago-91 20-ago-91 19-ago-91 20-ago-91 20-ago-91 24-ago-91 Fecha 22º44'36" 22º35'48" 22º36'36" 23º22'00" 23º17'54" 23º41'00" 23º44'18" 23º43'24" 23º41'54" 23º38'30" 24º44'48" 24º38'48" 24º39'12" 24º45'06" 24º52'00" 24º47'00" 24º33'00" 25º12'00" 25º18'24" 25º12'36" Lat. N 106º32'00" 106º37'42" 106º35'54" 107º10'00" 107º30'18" 107º23'36" 107º38'36" 107º39'06" 107º31'48" 107º58'48" 108º22'48" 108º26'54" 108º37'48" 108º19'12" 108º35'06" 108º38'30" 108º50'54" 109º07'00" 109º18'36" 109º06'03" Long. O No No No No No No No No No No No No No No No No No No No No Karling Tipo de arrastre Agassiz Agassiz IK IK Agassiz Agassiz Agassiz IK Agassiz IK Agassiz Agassiz Agassiz Agassiz Agassiz Agassiz IK IK IK Agassiz Prof. de arrastre (m) 520-535 980-1000 275 365 (698) 996-1148 491-535 968 630 823 0-720 720-732 1016-1020 1188-1208 444-496 216-224 579-608 770 410 (920) 600 (1890) 966 Prof. núcleos (m) No No No No No No No No No No No No No No No No No No No No ND ND ND ND ND <0.1 ND ND ND ND <0.1 0.15 ND <0.1 ND ND ND ND ND ND Oxígeno (ml l-1) O peraciones oceanográficas en aguas profundas 49 Est. 24 24A 25 A1 25 A1 25 A2 25B 26 27 28 Est. 1 3 4 5 6 7 12 13 Crucero TALUD III TALUD III TALUD III TALUD III TALUD III TALUD III TALUD III TALUD III TALUD III Crucero TALUD IV TALUD IV TALUD IV TALUD IV TALUD IV TALUD IV TALUD IV TALUD IV Cuadro 1. Continúa. 23-ago-00 23-ago-00 23-ago-00 23-ago-00 23-ago-00 23-ago-00 24-ago-00 24-ago-00 21-ago-91 24-ago-91 21-ago-91 21-ago-91 21-ago-91 23-ago-91 23-ago-91 23-ago-91 23-ago-91 Fecha Fecha 21º59'36" 21º58'28" 21º59'00" 22º03'18" 22º00'30" 22º00'22" 23º14'48" 23º17'30" 25º33'36" 25º45'12" 25º51'00" 25º51'00" 25º50'54" 25º03'18" 26º10'18" 26º32'48" 26º28'54" Lat. N Lat. N 106º23'42" 106º28'53" 106º35'00" 106º41'18" 106º47'48" 106º49'18" 107º28'24" 107º29'51" 109º42'01" 109º46'48" 109º57'00" 109º57'00" 109º56'54" 109º47'24" 109º58'24" 109º51'00" 109º45'42" Long. O Long. O No K K K No No No No No No No No No No No No No Karling Karling Tipo de arrastre Agassiz Agassiz Agassiz IK IK IK Agassiz Agassiz Agassiz Tipo de arrastre No Trineo Trineo Trineo No MN No Trineo Prof. de arrastre (m) 1043 1027-1060 1280-1360 200 (2000) 230 (ND) 150 (ND) 712-730 395-415 221-244 Prof. de arrastre (m) 460-490 785-800 1200-1290 1340-1360 ND 500 (1970) ND 860 Prof. núcleos (m) No No No No No No No No No Prof. núcleos (m) H 800 1228 1540 1970 No 1045 1550 0.33 0.28 0.84 1.60 ND ND ND 0.34 ND ND ND ND ND ND ND ND ND Oxígeno (ml l-1) Oxígeno (ml l-1) 14 24-ago-00 14 bis 24-ago-00 15 17 18 19 20 21 22 24 25 26 27 28 29 31 TALUD IV TALUD IV TALUD IV TALUD IV TALUD IV TALUD IV TALUD IV TALUD IV TALUD IV TALUD IV TALUD IV TALUD IV TALUD IV TALUD IV TALUD IV TALUD IV Biodiversidad y comunidades del talud continental del Pacífico mexicano 27-ago-00 25-ago-00 25-ago-00 25-ago-00 28-ago-00 26-ago-00 26-ago-00 26-ago-00 26-ago-00 26-ago-00 28-ago-00 27-ago-00 25-ago-00 24-ago-00 Est. Crucero Fecha Cuadro 1. Continúa. 50 25º46'02" 24º15'12" 24º15'18" 24º27'24" 24º29'30" 24º17'20" 24º58'00" 24º53'12" 24º56'24" 24º59'0" 24º52'30" 24º57'48" 24º14'42" 23º23'30" 23º13'24" 23º13'24" Lat. N 109º39'59" 108º17'10" 108º24'06" 108º35'16" 108º55'48" 108º50'30" 108º55'18" 108º59'24" 109º05'36" 109º12'06" 109º37'00" 109º37'00" 108º10'59" 107º47'48" 107º41'48" 107º41'48" Long. O No K K K No No No K K No K No No No No No Karling No Trineo Trineo Trineo No MN No Trineo Trineo Trineo Trineo MN No MN Tipo de arrastre Trineo No 908-944 1196-1200 1500-1540 ND 1325 (ND) ND 778-800 1200-1274 1605-1620 2024-2040 1280 (2080) ND 1500 (2350) Prof. de arrastre (m) 2220-2250 ND H (635) H (605620) 960 1240 1510 1850 No 549 848 1235 1565 2120 No Prof. núcleos (m) 2120 H (20502100) No ND 0.43 0.70 ND ND ND 0.18 0.25 0.76 1.35 2.1 ND ND 2.23 2.45 ND Oxígeno (ml l-1) O peraciones oceanográficas en aguas profundas 51 37 Est. TALUD IV Crucero 2 3 4 5 6 11 12 14 15 32 33 34 35 36 TALUD IV TALUD IV TALUD IV TALUD IV TALUD IV TALUD V TALUD V TALUD V TALUD V TALUD V TALUD V TALUD V TALUD V TALUD V Est. Crucero Cuadro 1. Continúa. 13-dic-00 13-dic-00 13-dic-00 13-dic-00 13-dic-00 17-dic-00 14-dic-00 14-dic-00 14-dic-00 Fecha 28-ago-00 27-ago-00 27-ago-00 27-ago-00 27-ago-00 27-ago-00 Fecha 21º59'54" 21º59'14" 22º04'39" 22º00'57" 22º00'00" 23º14'00" 23º18'00" 23º16'20" 23º23'24" Lat. N 24º53'49" 25º46'00" 25º45'54" 25º40'42" 25º53'59" 25º51'59" Lat. N 106º23'53" 106º28'30" 106º34'42" 106º40'00" 106º48'05" 107º00'00" 107º26'59" 107º40'45" 107º47'54" Long. O 109º36'48" 109º42'06" 109º48'06" 109º54'24" 110º11'17" 110º11'00" Long. O No K No K K K K K No Karling No No K K K No Karling Tipo de arrastre No Trineo No Trineo Trineo Trineo Trineo Trineo Trineo MN Tipo de arrastre No Trineo Trineo Trineo MN Prof. de arrastre (m) ND 1060-1080 1240 2000-2100 1000 (2100) (20402080) Prof. de arrastre (m) ND 730 ND 1515-1620 1950-2010 850-870 1160-1170 2080-2140 2340 Prof. núcleos (m) 460 730 1190 1495 1925 825 1120 2020 No No Prof. núcleos (m) H (860) 1040 1244 2020 No 0.22 0.13 0.56 1.15 1.88 0.07 0.54 1.98 ND Oxígeno (ml l-1) ND 0.25 0.55 0.79 1.68 ND Oxígeno (ml l-1) 52 Biodiversidad y comunidades del talud continental del Pacífico mexicano Est. 15 17 18 19 20 24 25 26 27 29 31 32 34 35 36 Crucero TALUD V TALUD V TALUD V TALUD V TALUD V TALUD V TALUD V TALUD V TALUD V TALUD V TALUD V TALUD V TALUD V TALUD V TALUD V Cuadro 1. Continúa. 17-dic-00 17-dic-00 17-dic-00 17-dic-00 17-dic-00 15-dic-00 15-dic-00 15-dic-00 15-dic-00 16-dic-00 16-dic-00 16-dic-00 16-dic-00 17-dic-00 14-dic-00 Fecha 25º45'53" 25º46'01" 25º40'41" 25º54'30" 25º54'30" 24º14'39" 24º15'12" 24º16'24" 24º14'44" 24º57'50" 24º51'46" 24º56'18" 24º58'22" 25º14'36" 23º21'30" Lat. N 109º39'49" 109º42'39" 109º54'24" 110º11'23" 110º11'24" 108º11'01" 108º17'09" 108º24'18" 108º35'18" 108º55'14" 108º57'59" 109º11'48" 109º12'08" 109º24'15" 107º48'12" Long. O No No No No No No K K K No K K No No No Karling No Trineo No No MN No Trineo Trineo Trineo No Trineo Trineo No MN Tipo de arrastre MN Prof. de arrastre (m) 1350 (2384) ND 940-990 1180-1200 1470-1525 ND 800-860 1280-1310 ND 1290 (2040) ND 850-900 ND ND 1340 (1990) 645 810 1225 1970 No 590 900 1160 1515 525 815 1220 1580 No Prof. núcleos (m) No 0.06 0.10 0.61 1.3 ND 0.07 0.15 0.38 1.20 ND 0.13 0.53 1.23 ND ND Oxígeno (ml l-1) O peraciones oceanográficas en aguas profundas 53 11 12 13 15 TALUD VI TALUD VI TALUD VI TALUD VI 18 19 20 22 25 26 27 29 29 3 4 5 7 TALUD VI TALUD VI TALUD VI TALUD VI TALUD VI TALUD VI TALUD VI TALUD VI TALUD VI TALUD VI TALUD VI TALUD VI TALUD VI Est. Crucero Cuadro 1. Continúa. 15-mar-01 15-mar-01 15-mar-01 15-mar-01 16-mar-01 16-mar-01 16-mar-01 16-mar-01 16-mar-01 14-mar-01 14-mar-01 14-mar-01 14-mar-01 13-mar-01 13-mar-01 13-mar-01 14-mar-01 Fecha 24º14'56" 24º16'24" 24º14'48" 24º17'26" 24º51'41" 24º56'18" 25º01'12" 25º16'24" 25º16'24" 23º14'59" 23º18'36" 23º14'59" 23º14'42" 22º00'01" 22º00'51" 22º00'42" 22º21'39" Lat. N 108º16'17" 108º24'18" 108º35'11" 108º50'05" 108º57'54" 109º06'42" 109º11'36" 109º24'54" 109º24'54" 106º59'00" 107º26'56" 107º30'03" 107º30'00" 106º28'06" 106º34'42" 106º39'55" 107º01'42" Long. O K K K No K K K K No K K No No K K K No Karling Trineo Trineo Trineo No Trineo Trineo Trineo Trineo MN Trineo Trineo No MN Tipo de arrastre Trineo Trineo Trineo MN Prof. de arrastre (m) 770-780 1190-1250 1470-1530 1305 (2100) 825-855 1050-1160 ND 1230 (2390) 850-890 1160-1200 1250-1440 ND 830-850 1190-1270 1580-1600 2080 1440 (2080) 820 1200 1530 No 850 1270 1585 No No 760 1110 1345 No Prof. núcleos (m) 780 1300 1535 No 0.29 0.73 1.27 ND 0.22 1.40 1.44 ND ND 0.17 0.50 0.50 ND 0.21 ND 1.74 ND Oxígeno (ml l-1) Cuadro 1. Continúa. Est. 32 33 34 36 Est. 3 4 5 11 12 13B 18 18 bis 19 20 25 26 Crucero TALUD VI TALUD VI TALUD VI TALUD VI Crucero TALUD VII TALUD VII TALUD VII TALUD VII TALUD VII TALUD VII TALUD VII TALUD VII TALUD VII TALUD VII TALUD VII TALUD VII 54 Biodiversidad y comunidades del talud continental del Pacífico mexicano 05-jun-01 05-jun-01 05-jun-01 06-jun-01 06-jun-01 06-jun-01 07-jun-01 10-jun-01 07-jun-01 07-jun-01 08-jun-01 08-jun-01 Fecha 17-mar-01 17-mar-01 17-mar-01 17-mar-01 Fecha 22º13'18" 22º03'18" 22º00'24" 23º16'54" 23º18'18" 23º30'18" 24º14'30" 24º52'46" 24º16'12" 24º14'48" 24º52'48" 24º25'24" Lat. N 25º46'15" 25º45'49" 25º43'50" 25º53'15" Lat. N 106º31'36" 106º34'42" 106º39'54" 106º59'48" 107º26'48" 107º44'00" 108º16'24" 108º58'05" 108º23'42" 108º35'12" 108º58'00" 109º05'21" Long. O 109º42'08" 109º48'40" 109º53'59" 110º10'08" Long. O K K No K K K K No K No K K Karling No K K No Karling Tipo de arrastre Trineo Trineo Trineo Trineo Trineo Trineo Trineo No Trineo Trineo Trineo Trineo Tipo de arrastre No Trineo Trineo No Prof. de arrastre (m) ND 1120-1160 1240-1270 1360 (2000) Prof. de arrastre (m) 740-750 1200-1230 1490-1520 780-790 1040-1120 1400-1450 950-1010 ND 1160-1180 1480-1520 780-850 1180-1220 Prof. núcleos (m) 750 1190 1520 760 1130 1470 830 865 1200 1530 905 1260 Prof. núcleos (m) 780 1105 1295 No 0.77 0.82 1.81 0.15 0.18 1.04 ND ND 0.08 0.79 0.04 0.35 Oxígeno (ml l-1) 0.28 0.57 0.86 ND Oxígeno (ml l-1) O peraciones oceanográficas en aguas profundas 55 Est. 27 32B 33B 34B Est. 2 3 6 7 9 10 11 12 13 15 16 Crucero TALUD VII TALUD VII TALUD VII TALUD VII Crucero TALUD VIII TALUD VIII TALUD VIII TALUD VIII TALUD VIII TALUD VIII TALUD VIII TALUD VIII TALUD VIII TALUD VIII TALUD VIII Cuadro 1. Continúa. 18-abr-05 18-abr-05 18-abr-05 16-abr-05 16-abr-05 17-abr-05 17-abr-05 17-abr-05 17-abr-05 17-abr-05 17-abr-05 08-jun-01 09-jun-01 09-jun-01 09-jun-01 Fecha Fecha 25º23'06" 25º24'48" 25º21'00" 24º37'06" 24º32'36" 24º51'18" 25º04'00" 25º01'48" 24º58'12" 24º54'24" 25º02'00" 25º01'30" 26º03'00" 26º06'30" 26º05'30" Lat. N Lat. N 110º18'24" 110º34'48" 110º17'00" 109º25'6" 109º30'30" 109º48'24" 110º04'00" 110º06'54" 110º16'06" 110º25'36" 110º27'00" 109º12'00" 109º55'24" 110º06'42" 110º10'30" Long. O Long. O K No No K K No No K K K No K K K K Karling Karling Trineo Trineo MN Tipo de arrastre Trineo Trineo Trineo Trineo Tipo de arrastre No Trineo MN MN Trineo Trineo Trineo MN Prof. de arrastre (m) 1580-1600 850-880 1260-1300 1500-1520 Prof. de arrastre (m) 2150 1600 580 (1700) 510 (1600) 1650-1680 1500 895-920 1150 (1280) 1625 (2100) 2100 (?) 1030 2100 1030 No Prof. núcleos (m) 1600 780 1295 1500 Prof. núcleos (m) 2200 2180?? No No 1650 1550 920 No 1.02 0.20 ND ND 0.39 (?) ND ND 0.84 ND 0.20 ND 1.39 0.15 0.60 0.78 Oxígeno (ml l-1) Oxígeno (ml l-1) Cuadro 1. Continúa. Est. 17B 18 20 21 22 Est. 1 2 3 4 5 6 7A 7B 8 10 11 14 Crucero TALUD VIII TALUD VIII TALUD VIII TALUD VIII TALUD VIII Crucero TALUD IX TALUD IX TALUD IX TALUD IX TALUD IX TALUD IX TALUD IX TALUD IX TALUD IX TALUD IX TALUD IX TALUD IX 56 Biodiversidad y comunidades del talud continental del Pacífico mexicano 10-nov-05 11-nov-05 11-nov-05 11-nov-05 11-nov-05 12-nov-05 15-nov-05 15-nov-05 12-nov-05 12-nov-05 12-nov-05 15-nov-05 18-abr-05 19-abr-05 19-abr-05 19-abr-05 19-abr-05 Fecha Fecha 24º37'54" 24º38'42" 24º32'52" 24º28'42" 24º33'20" 24º49'38" 25°11'30" 25°11'19" 25º07'28" 24º56'24" 24º53'33" 25º31'30" 24º24'24" 25º50'00" 25º56'56" 26º02'18" 26º03'42" Lat. N Lat. N 109º22'36" 109º17'36" 109º29'04" 109º34'06" 109º49'30" 109º47'58" 109°41'06" 109°42'45" 109º49'48" 110º16'42" 110º25'38" 110º07'24" 110º50'06" 110º34'00" 110º43'00" 110º37'06" 110º23'54" Long. O Long. O No K K K No MN No No K No No No K No K K K Karling Karling Tipo de arrastre Trineo MN Trineo Trineo Trineo Tipo de arrastre MN Trineo Trineo Trineo No No MN MN Trineo Trineo No Trineo Prof. de arrastre (m) 700-750 690 (1300) 1140-1150 1380 2200 Prof. de arrastre (m) 1050 (2285) 2195-2300 1454-1620 1901-2120 ND 920 (1650) 1510 (2377) 850 (2392) 1657 969-1225 ND 2010-2244 Prof. núcleos (m) 700 No 1150 1380 2200 Prof. núcleos (m) No 2250 1625 1395 470 No No No 1595 1254 875 2000 ND 2.37 1.19 0.85 ND ND ND ND 1.16 0.11 (?) ND 1.86 0.13 ND 0.30 0.51 1.27 Oxígeno (ml l-1) Oxígeno (ml l-1) O peraciones oceanográficas en aguas profundas 57 Est. 12 15 15 B 16 16B 17 18 19 20B 21B 22 23 Est. 3 4 5 6 7 Crucero TALUD IX TALUD IX TALUD IX TALUD IX TALUD IX TALUD IX TALUD IX TALUD IX TALUD IX TALUD IX TALUD IX TALUD IX Crucero TALUD X TALUD X TALUD X TALUD X TALUD X Cuadro 1. Continúa. 09-feb-07 09-feb-07 09-feb-07 09-feb-07 10-feb-07 12-nov-05 13-nov-05 15-nov-05 13-nov-05 13-nov-05 13-nov-05 13-nov-05 14-nov-05 14-nov-05 14-nov-05 14-nov-05 15-nov-05 Fecha Fecha 28º16'38" 28º16'06" 28º14'50" 28º01'34" 27º53'09" 25°30’24” 25º21'27" 25º21'32" 24º23'48" 25º26'54" 25º19'54" 25º49'38" 25º53'09" 25º58'07" 26º04'42" 26º03'42" 26º03'59" Lat. N Lat. N 112º35'12" 112º32'50" 112º24'53" 112º17'25" 112º16'42" 110°26’30” 110º18'18" 110º18'32" 110º36'42" 110º38'01" 110º47'42" 110º34'45" 110º41'54" 110º40'04" 110º34'48" 110º20'36" 110º24'45" Long. O Long. O No K K K No No No No No No No No No No No No No Karling Karling Tipo de arrastre MN Trineo No Trineo No Trineo MN Trineo Trineo Trineo Trineo MN Tipo de arrastre Trineo Trineo Trineo Trineo MN Prof. de arrastre (m) 1150 (1595) 1985-2290 ND 997-1021 ND 626-846 ND 1078-1084 1229-1343 1349-1369 2214-2309 1450 (2318) Prof. de arrastre (m) 377-379 587-633 820-837 1037-1043 900 (1191) Prof. núcleos (m) No 2018 1983 1015 610 800 No 1050 1192 1330 2220 No Prof. núcleos (m) 379 625 No No No 0.96 0.39 0.11 0.30 ND ND 1.78 ND 0.15 ND 0.03 ND 0.24 0.57 0.57 1.63 ND Oxígeno (ml l-1) Oxígeno (ml l-1) Cuadro 1. Continúa. Est. 8 9 10 11 12 13 14 15 17 18 19 20 21 22 23 23 25 26 Crucero TALUD X TALUD X TALUD X TALUD X TALUD X TALUD X TALUD X TALUD X TALUD X TALUD X TALUD X TALUD X TALUD X TALUD X TALUD X TALUD X TALUD X TALUD X 58 Biodiversidad y comunidades del talud continental del Pacífico mexicano 14-feb-07 14-feb-07 13-feb-07 13-feb-07 14-feb-07 14-feb-07 10-feb-07 10-feb-07 10-feb-07 11-feb-07 11-feb-07 11-feb-07 11-feb-07 11-feb-07 12-feb-07 12-feb-07 12-feb-07 13-feb-07 Fecha 26º39'04" 26º35'16" 27º14'31" 27º02'46" 27º00'58" 27º00'30" 28º05'56" 27º52'51" 27º48'30" 27º34'16" 28º01'31" 27º49'46" 27º44'53" 27º40'24" 27º08'00" 27º09'06" 27º13'30" 27º14'41" Lat. N 111º18'20" 111º05'59" 111º14'39" 110º52'57" 110º12'23" 111º12'00" 112º26'50" 112º15'53" 112º17'12" 111º40'30" 111º51'10" 111º43'18" 111º36'58" 111º39'54" 111º44'10" 111º46'54" 111º36'08" 111º36'15" Long. O K K K K No No K K K No K K K K K K K No Karling Trineo Trineo Trineo Trineo No MN Tipo de arrastre Trineo Trineo Trineo MN Trineo Trineo Trineo Trineo Trineo Trineo Trineo MN Prof. de arrastre (m) 975-1007 1205-1215 1396-1422 1220 (1800) 465-486 668-704 905-943 1528-1530 1289-1326 1526 1786-1816 Aprox. 1250 (1785) 1864-1865 1575-1586 ND aprox. 1250 (1770) 837-840 1292-1301 830 1310 1900 1640 1754 1750 Prof. núcleos (m) No 1216 1426 No 460 680 901 1560 1330 1440 1930 No 0.17 0.36 0.65 0.88 0.48 ND 0.26 0.31 0.44 ND 0.14 0.12 0.20 0.51 0.46 0.51 0.58 ND Oxígeno (ml l-1) O peraciones oceanográficas en aguas profundas 59 Est. 27 29 30 Est. 1 2 3 5 6 6A 8 9 11 12 13 14 15 Crucero TALUD X TALUD X TALUD X Crucero TALUD XI TALUD XI TALUD XI TALUD XI TALUD XI TALUD XI TALUD XI TALUD XI TALUD XI TALUD XI TALUD XI TALUD XI TALUD XI Cuadro 1. Continúa. 08-jun-07 08-jun-07 08-jun-07 08-jun-07 08-jun-07 09-jun-07 09-jun-07 07-jun-07 07-jun-07 07-jun-07 07-jun-07 07-jun-07 07-jun-07 15-feb-07 15-feb-07 15-feb-07 Fecha Fecha 17º06'30" 17º10'23" 17º23'28" 17º49'24" 17º45'00" 17º32'24" 17º23'00" 16º52'00" 16º47'42" 16º50'18" 16º52'35" 16º16'57" 16º58'00" 26º40'18" 26º35'36" 26º36'50" Lat. N Lat. N 101º40'16" 101º37'21" 101º25'06" 101º59'30" 102º00'06" 102º02'28" 102º04'00" 100º22'00" 100º28'12" 100º42'09" 100º47'34" 100º54'30" 100º57'00" 110º37'00" 110º35'44" 110º21'10" Long. O Long. O No No No No No No No K No No No No No No No No Karling Karling No No No No No No No Tipo de arrastre No Trineo Trineo Tipo de arrastre Trineo No No No No MN Prof. de arrastre (m) ND 1383-1439 1203-1213 Prof. de arrastre (m) 740-790 ND ND ND ND 1400 (1960) ND ND ND ND ND ND ND 1778 1404 695 580 1195 1539 2140 Prof. núcleos (m) 1474 1395 1249 Prof. núcleos (m) 880 990 1380 1550 2000 No 1.48 0.20 0.17 ND 0.65 ND ND 0.14 0.24 0.92 1.25 1.73 ND 0.77 0.55 ND Oxígeno (ml l-1) Oxígeno (ml l-1) Cuadro 1. Continúa. Est. 15B 16 17 18 19 19B 20 23 24 25 26 27 28 29 30 30B Crucero TALUD XI TALUD XI TALUD XI TALUD XI TALUD XI TALUD XI TALUD XI TALUD XI TALUD XI TALUD XI TALUD XI TALUD XI TALUD XI TALUD XI TALUD XI TALUD XI 60 Biodiversidad y comunidades del talud continental del Pacífico mexicano 10-jun-07 10-jun-07 10-jun-07 10-jun-07 10-jun-07 11-jun-07 11-jun-07 11-jun-07 11-jun-07 11-jun-07 09-jun-07 09-jun-07 09-jun-07 09-jun-07 09-jun-07 09-jun-07 Fecha 17º53'18" 18º30'25" 18º29'21" 18º23'22" 18º33'12" 18º41'00" 18º48'05" 19º18'00" 19º26'23" 19º28'10" 17º48'00" 18º07'30" 18º05'02" 17º58'12" 17º56'01" 17º56'00" Lat. N 103º11'30" 103º54'28" 104º01'18" 104º13'16" 104º32'12" 104º33'07" 104º32'36" 105º26'19" 105º19'00" 105º17'30" 102º30'42" 103º02'32" 103º04'07" 103º05'21" 103º08'24" 103º10'00" Long. O No No No No No No No No No No No No No No No No Karling No No No No No No No No No No Tipo de arrastre No No No No No MN Prof. de arrastre (m) ND ND ND ND ND 1490 (1750) ND ND ND ND ND ND ND ND ND ND 2677 994 1502 1800 1918 1305 1050-1120 1590-1595 1225 860 Prof. núcleos (m) CTD (285) 250 560 1270 1843 No 1.94 0.09 0.27 0.80 0.89 0.03 0.54 1.13 0.60 0.12 ND 0.12 ND 0.68 1.68 ND Oxígeno (ml l-1) O peraciones oceanográficas en aguas profundas 61 Est. 2 3 4 5 8 9 10 13 14 15 15C 16 17 18 19 20 22 Crucero TALUD XII TALUD XII TALUD XII TALUD XII TALUD XII TALUD XII TALUD XII TALUD XII TALUD XII TALUD XII TALUD XII TALUD XII TALUD XII TALUD XII TALUD XII TALUD XII TALUD XII Cuadro 1. Continúa. 31-mar-08 31-mar-08 27-mar-08 27-mar-08 31-mar-08 31-mar-08 28-mar-08 29-mar-08 29-mar-08 28-mar-08 30-mar-08 30-mar-08 30-mar-08 31-mar-08 28-mar-08 28-mar-08 28-mar-08 Fecha 18º08'03" 18º05'08" 17º58'08" 17º56'30" 17º54'20" 18º11'27" 16º58'28" 17º04'16" 17º10'26" 17º11'03" 17º45'16" 17º36'20" 17º25'33" 17º27'51" 16º49'18" 16º54'35" 16º59'39" Lat. N 103º02'24" 103º03'33" 103º05'30" 103º09'57" 103º10'45" 103º52'29" 100º55'20" 101º39'28" 101º37'37" 101º28'05" 102º00'29" 102º01'59" 102º07'20" 102º10'43" 100º30'52" 100º44'10" 100º58'07" Long. O No No No No No No K K K K K K K No K K No Karling No No No No No MN Trineo Trineo Trineo Trineo Trineo Trineo Trineo MN Tipo de arrastre Trineo Trineo MN Prof. de arrastre (m) 990-1088 1380-1456 1200 (1995) 1977-1925 1880-1940 1392-1420 1180-1299 1199-1100 1415-1476 2080-2125 1530 (1880) ND ND ND ND ND 1340 (2200) 296 740 1283 1625 1992 No 1849 1928 1520 1290 1200 1245 2018 No Prof. núcleos (m) 980 1386 No ND ND 0.64 1.59 ND ND 1.48 1.49 0.75 0.51 0.36 0.80 1.61 ND 0.20 0.80 ND Oxígeno (ml l-1) Cuadro 1. Continúa. Est. 23 24 25 26 27 28 28B 29 30 30B Est. B B 34 36 37 Crucero TALUD XII TALUD XII TALUD XII TALUD XII TALUD XII TALUD XII TALUD XII TALUD XII TALUD XII TALUD XII Crucero TALUD XIII TALUD XIII TALUD XIII TALUD XIII TALUD XIII 62 Biodiversidad y comunidades del talud continental del Pacífico mexicano 13-ene-09 13-ene-09 14-ene-09 15-ene-09 15-ene-09 02-abr-08 26-mar-08 26-mar-08 Fecha 01-abr-08 01-abr-08 01-abr-08 01-abr-08 02-abr-08 02-abr-08 02-abr-08 Fecha 26º19'54" 26º17'04" 26º33'36" 26º07'12" 26º03'54" 19º19'37" 19º22'05" 19º30'37" Lat. N 18º33'43" 18º28'00" 18º26'45" 18º33'27" 18º40'28" 18º50'19" 18º56'00" Lat. N 110º29'12" 110º27'53" 111º02'00" 110º30'53" 110º20'06" 105º26'20" 105º16'18" 105º19'16" Long. O 103º57'45" 104º14'10" 104º16'10" 104º28'21" 104º35'51" 104º34'14" 104º59'57" Long. O No No No No No K K K Karling K K K K K K No Karling Trineo Trineo Trineo Tipo de arrastre Agassiz No Agassiz Agassiz Agassiz Tipo de arrastre Trineo Trineo Trineo Trineo Trineo Trineo MN Prof. de arrastre (m) 1058-1088 1535-1542 1858-1879 2125-2165 1040-1095 1101-1106 1450 (1425) 1609-1643 1350-1380 865-1045 Prof. de arrastre (m) 1295-1330 ND 1485-1530 2360-2300 2056-2195 No 1350 825 Prof. núcleos (m) 1780 1785 1484 2326 1860 Prof. núcleos (m) 1150 1507 1627 1956 1158 1080 No 0.87 0.87 0.95 1.83 1.68 1.03 0.71 0.11 Oxígeno (ml l-1) 0.22 0.95 1.39 1.97 0.26 0.27 ND Oxígeno (ml l-1) muestras de la endo y epifauna más grande jamás vista para las aguas profundas del Pacífico mexicano. Las campañas TALUD se dividieron en cinco etapas: Etapa 1 (TALUD I-III; 1989-1990). Frente a Sinaloa, SE del golfo de California (Fig. 4 A; Cuadro 1). Esta etapa puede ser considerada como la etapa preliminar o de exploración. La primera campaña del proyecto TALUD se efectuó en 1989 (TALUD I). El conocimiento local acerca de la ubicación y de la extensión de la ZMO era muy somero, razón por la cual se optó por realizar transectos desde la plataforma continental externa (aproximadamente entre 110 y 180 m de profundidad) hacia aguas más profundas. Rápidamente se encontraron problemas técnicos muy serios, pues parte de los fondos de la zona intermedia entre la plataforma y el talud superior resultó cubierta de ramas y troncos de árboles hundidos. Los daños ocasionados a las redes utilizadas fueron considerables y se optó por limitar el número de estaciones visitadas. Además, se pudo constatar que esta zona relativamente somera, al límite entre la plataforma y el talud continentales, se carecterizaba por tener concentraciones de oxígeno disuelto muy bajas. Sin lugar a dudas, los mejores resultados de esta primer etapa en lo que se refiere a la fauna profunda correspondieron al TALUD III, ya que se efectuaron 22 arrastres, de los cuales 15 fueron por debajo de los 700 m, y se recolectaron muchas especies de la fauna típicamente asociada con la franja bentónica ubicada al limite inferior o por debajo de la ZMO. Etapa 2 (TALUD IV-VII; 2000-2001). Frente a Sinaloa, SE del golfo de California (Figs. 4 B-D, 5 A; Cuadro 1). Basándose en los resultados de los tres cruceros anteriores, se modificaron algunos aspectos esenciales del estudio (e.g., intervalo de profundidad de los muestreos, equipos utilizados, académicos involucrados) y se efectuaron cuatro cruceros, intentando visitar las mismas localidades cuatro veces. En algunas ocasiones eso no fue posible; sin embargo, la serie de datos y de información obtenida fue muy amplia y resultó ser un avance muy significativo para el estudio de las aguas profundas en México. En esta segunda etapa se visitaron 90 estaciones y se realizaron 57 arrastres profundos. Etapa 3 (TALUD VIII y IX; 2005). Baja California Sur, SO del golfo de California (Fig. 5 B, C; Cuadro 1). Estos dos cruceros fueron organizados frente a la porción SE de la península de Baja California, entre las islas Cerralvo y Carmen. Las dificultades encontradas durante estos cruceros fueron esencialmente por causa de la proximidad de la costa rocosa y la irregularidad de los fondos. De hecho, en una de las estaciones, los equipos de arrastre pegaron una zona rocosa, se perdió la draga O peraciones oceanográficas en aguas profundas 63 Fig. 5. Posición de las estaciones de muestreo visitadas durante: A. la campaña TALUD VII; B. la campaña TALUD VIII; C. la campaña TALUD IX; D. las campañas TALUD X-XIII. A B C D Karling (ruptura del cable de arrastre) y se doblaron los patines del trineo bentónico. A pesar de las dificultades encontradas en una zona caracterizada por la abundancia de salientes rocosos profundos, se efectuaron 37 estaciones de muestreo y 23 arrastres bentónicos en esta etapa. Etapa 4 (TALUD X y XIII; 2007 y 2009). Golfo de California central (Fig. 5 D; Cuadro 1). Con el fin de completar el estudio del talud superior del golfo de California y realizar el análisis de la composición de las comunidades presentes por debajo de la ZOM, se efectuaron dos campañas en el área comprendida entre el norte de isla Carmen y el sur de isla Tiburón. En el crucero TALUD X se realizaron unas de las estaciones de muestreo más someras del proyecto posteriores a 1990, entre 379 y 633 m. Eso se debió a la variación del nivel de la ZOM comparativamente con los cruceros anteriores en el SE y SO del golfo de California. En total se visitaron 25 estaciones en esta campaña, incluyendo 18 arrastres con el trineo bentónico. La campaña TALUD XIII fue muy corta (cuatro arrastres) y consistió en la recuperación de algunas estaciones que no se pudieron realizar al final de la campaña TALUD X por mal tiempo. Estas estaciones fueron todas ubicadas en la cuenca del Carmen y proximidades. 64 Biodiversidad y comunidades del talud continental del Pacífico mexicano Etapa 5 (TALUD XI y XII; 2007 y 2008). Costa SO de México, Jalisco-Guerrero (Fig. 5 D; Cuadro 1). Debido a un error de maniobra del buque en la estación 2 del TALUD XI, el cable de arrastre del trineo y de la draga Karling fue cortado causando el hundimiento de los equipos. Como consecuencia, no se pudo realizar la gran mayoría de los arrastres programados en esta campaña (de 18 solo se realizó uno) y, considerando que al momento de la pérdida de los equipos el buque se encontraba justo al norte de Acapulco, se optó por realizar las hidrocalas y los muestreos de sedimentos con el nucleador de caja camino de regreso al puerto de embarque. Al año siguiente, se consiguió tiempo de uso del buque para recorrer de nuevo esta zona y, esta vez, se visitaron 27 estaciones y se realizaron 18 arrastres completos con éxito. A lo largo del proyecto TALUD, en total se realizaron 666 operaciones de muestreo. Estos muestreos comprenden: 142 hidrocalas; 125 muestreos con el nucleador múltiple; 99 muestreos de sedimentos con la draga Karling y 80 con el nucleador de caja; 179 muestras de la fauna bentónica con la red camaronera de 80’ (21 muestras), con las dragas tipo Agassiz (41) y con el trineo bentónico (117); y 41 muestreos de la fauna pelágica, la mayoría (28) con una red para micronecton y con la red Isaacs Kidd (12) (Cuadro 1). Una lista de los equipos e instalaciones utilizadas durante las campañas del proyecto TALUD aparece en el Cuadro 2. En la totalidad de las operaciones de muestreo se utilizaron los localizadores vía satélite para determinar la posición exacta de los muestreos y las ecosondas para revisar el perfil del fondo y determinar las profundidades de cada estación al momento de efectuar un muestreo o una hidrocala. En los TALUD I-IX se utilizó una ecosonda analógica Edo Western con impresión directa en papel, mientras que a partir del TALUD X se contó con una ecosonda digital SIMRAD con sistema de conservación de datos en soporte digital. Considerando el amplío abanico de grupos faunísticos estudiados y la necesidad de efectuar algunos análisis directamente a bordo, se usaron todos los laboratorios de la cubierta de trabajo así como los “winches” (malacates o cabrestantes) hidráulicos, de uso múltiple y del CTD (Lám. 4 B-D), y los congeladores de popa. Hidrocalas En cada estación de muestreo, se procuró realizar una hidrocala entre la superficie y el fondo para obtener información acerca de los perfiles de temperatura, de salinidad y de oxígeno disuelto. Este último parámetro es particularmente importante O peraciones oceanográficas en aguas profundas 65 Hidrocalas y soporte para CTD Winkler Hidrocala (salinidad, temperatura, concentración oxigeno) Hidrocala (salinidad, temperatura, concentración oxigeno) Zooplancton Micronecton Estructura Rosette Titulador de oxígeno CTD O2 Sea-bird 19 Red pelágica Isaacs-Kidd Red Bongo CTD SBE-911-plus Monitoreo de la profundidad Ecosondas (analógica y digital) Marco de popa y cabestrante de uso múltiple Cabestrante del CTD Tambor de redes CTD, rosette y nucleador múltiple Maniobras de recuperación de redes y trineo Redes de arrastre y nucleador de caja Hidrocalas y soporte para CTD ICML, UNAM B/O El Puma B/O El Puma B/O El Puma B/O El Puma Adquisición CONACyT TALUD II y III TALUD I TALUD XII y XIII Analógica: TALUD I a IX. Digital: TALUD X-XIII TALUD IV-XIII Todas B/O El Puma B/O El Puma Todas Todas Todas B/O El Puma B/O El Puma B/O El Puma C: Según la longitud de cable disponible D: 2 redes de 500 m, boca de 0.3 m2 C: Hasta 500 m de profundidad D: ca 3 m2 C: Aprox. 5500 m de cable sintético C: Aprox. 3000 m de cable conductor C: Analógica, impresión directa. Digital, conservación de datos en formato electrónico C: 12 botellas; 10500 m C: Indefinida C: 3500 m; sensor O2 hasta 2000 m. C: 6500 m C: Aprox. 3000 m de cable de acero C: Redes grandes y cable sintético Características Malacate hidrológico Campañas donde se uso Uso Equipo Biodiversidad y comunidades del talud continental del Pacífico mexicano Origen Cuadro 2. Equipos e instalaciones utilizadas durante las campañas del proyecto TALUD. C, capacidad; D, dimensiones. 66 O peraciones oceanográficas en aguas profundas 67 Sedimentos Sedimentos-meiofauna Sedimentos-meiofauna Draga Smith McIntyre Nucleador de caja Nucleador múltiple Macrofauna superficial Macrofauna (grandes muestras de crustáceos y peces) Todos los grupos Draga Agassiz (grande) Congeladores Laboratorios (varios) Macrofauna superficial Draga Agassiz (chica) Trineo bentónico Sedimentos hasta 7 cm de profundidad Macrofauna superficial Micronecton Red para micronecton Draga Karling Uso Equipo Cuadro 2. Continúa. B/O El Puma Construcción local (CONACYT) Construcción local (CONACyT-PAPIIT) Construcción local (CONACyT) Construcción local (ICML) B/O El Puma B/O El Puma B/O El Puma B/O El Puma Construcción local Origen Todas TALUD II y III TALUD IIII a XII TALUD XIII TALUD IV-XII TALUD IV-XII TALUD IV-IX TALUD I-III TALUD VIII-XIII TALUD IV-XIII Campañas donde se uso C: Espacio de trabajo para 12 personas simultáneamente C: Hasta 1500 m de profundidad; boca de 0.81 m2 C: Hasta 600-700 m; 20 l C: Hasta 2500 m de profundidad; 80 l C: Hasta 2020 m de profundidad; 6 núcleos C: Hasta 80 l de capacidad. D: 1.10 m por 0.40 m D: Anchura, 2.35 m; altura, 0.90 m D: Anchura, 1.80 m; altura, 0.80 m D: Anchura, 2.50 m; altura, 1.0 m C: hasta -20 °C; 30 m3 Características en el contexto del proyecto TALUD, pues, como se vio anteriormente la ZMO representa un factor limitativo en los patrones de distribución batimétrica y geográfica de las especies pelágicas y bentónicas en casi la totalidad de la costa del Pacífico mexicano (Hendrickx y Serrano 2010). Para bajar los equipos en la columna de agua, se utilizó el cabestrante especialmente diseñado para el uso de la estructura Rosette y de los CTD de control directo (i.e., con una conexión directa a través del cable de descenso-ascenso). Desafortunadamente, en la mayoría de los cruceros TALUD los equipos de medición de salinidad, temperatura y oxígeno in situ (CTD) del B/O El Puma estaban descompuestos o no confiables por no haber recibido su mantenimiento rutinario y sus calibraciones en la fábrica. En la mayoría de los cruceros se midió la concentración de oxígeno por medio del método Winkler, utilizando muestras de agua recolectadas en botellas instaladas en una estructura tipo Rossette (Lám. 5 A), con sistema de cierre controlado vía el cable del cabestrante hidrológico. Estas mediciones se hicieron con un sistema de titulación por micropipeta instalado a bordo. En el TALUD III se contó con algunos datos de oxígeno disuelto obtenido con un CTD O2 (i.e., un CTD autónomo equipado de un sensor para medición del oxígeno disuelto en el agua) propiedad de la University of South Carolina, EE.UU. Los perfiles correspondientes (véase un ejemplo en la Fig. 6) sirvieron para reconocer la amplitud del fenómeno en la zona y convencieron a los participantes del proyecto TALUD de la necesidad de poder contar con un equipo similar. A partir de la campaña TALUD VII se consiguió un CTD O2 Seabird 19 (Lám. 5 A-B) con capacidad de medición hasta 3500 m (2000 en el caso del sensor de oxígeno) que permitió realizar de manera rutinaria los perfiles en las siguientes campañas; además, se siguió con el análisis de muestras de agua por el método Winkler en tres niveles (subsuperficie, media agua, cerca del fondo). Durante el crucero TALUD XI y a solicitud del Instituto Tecnológico del Mar, Mazatlán, se conservaron muestras de agua recolectadas a nivel del fondo para análisis químicos. Sedimentos De manera muy ocasional se utilizó una draga Smith-McIntyre (Lám. 5 C) en los cruceros TALUD I y II. Desafortunadamente, las muestras de sedimentos de estas campañas no fueron analizadas. Debido a las altas profundidades, en el crucero TALUD III no se intentó conseguir muestras de sedimentos ya que no se contaba con un equipo adecuado para realizar este tipo de muestreo. En los cruceros TALUD IV-VII se utilizaron dos tipos de recolectores de sedimentos, cada uno con un pro68 Biodiversidad y comunidades del talud continental del Pacífico mexicano Fig. 6. Perfiles de oxígeno disuelto medidos a través de la columna de agua durante la campaña TALUD III. pósito distinto. Para la meiofauna se diseñó un nucleador múltiple con una capacidad de seis pequeños núcleos de aproximadamente 25 cm de longitud (Lám. 5 D). Construido en un taller local, este equipo, de un peso total de solamente 40 kg, resultó extraordinariamente económico y eficiente y permitió obtener muestras en profundidades de más de 2000 m. Colocado por debajo de la estructura Rosette, se operó con el malacate hidrológico hasta que tocara fondo; esta operación de ambos equipos de manera simultánea permitió ahorrar una considerable cantidad de horas de operación en alta mar. Para la macrofauna que vive en el sedimento, se utilizó una pequeña draga de arrastre cuyo diseño fue inspirado de las dragas tipo Karling. Construida en un taller local, la draga Karling fue diseñada para recolectar el sedimento superficial hasta una profundidad de 7 cm, con una capacidad máxima de almacenaje de 80 litros de lodo (Lám. 6 A, B). Este equipo se colocó en “tren” por detrás del trineo bentónico (o red de patines) utilizado a partir del TALUD IV (véase infra) y permitió realizar un muestreo del sedimento superficial de manera simultánea con el muestreo de la fauna apoyada sobre el fondo (epiO peraciones oceanográficas en aguas profundas 69 Lam. 5. A, estructura Rosette equipada con botellas y el CTD del proyecto; B, estructura Rosette en maniobra de inmersión; C, draga geológica Smith McIntyre; D, nucleador múltiple para meiofauna siendo recuperado en la plataforma hidrológica. B A C D fauna), ahorrándose de esta manera varias horas de muestreo. La draga Karling fue utilizada de manera rutinaria en los cruceros TALUD IV-XII, con algunas excepciones debido a fallas técnicas (Cuadro 1). Se conservaron fracciones de estas mues70 Biodiversidad y comunidades del talud continental del Pacífico mexicano tras para los análisis granulométricos y del contenido de materia orgánica (por calcinación). El resto fue filtrado en una estructura de filtración diseñada para este propósito y utilizada con anterioridad en las campañas SIPCO y CORTES (Lám. 7 A). Esta estructura contó con tamices apilados con aperturas de malla de ca. 5 mm (tamiz superior) hasta 500 μm (tamiz inferior). En los últimos cruceros TALUD se sustituyó la malla más fina por una de 300 μm. A partir del TALUD X se contó con un nucleador de caja tipo Reineck (Lám. 6 C, D), de aproximadamente 80 litros de capacidad máxima. Este tipo de equipo tiene la ventaja de proporcionar muestras muy homogéneas del sedimento del fondo, las cuales permiten obtener, a su vez, submuestras directamente en el puente, casi como si se estaría muestreando in situ. De hecho, al usar este equipo, se consiguieron muestras muy precisas del sedimento superficial para el análisis de la meiofauna y de algunos parámetros ambientales del sedimento (e.g., composición del sedimento, concentración de materia orgánica por titulación, temperatura del sedimento, pH). Además, durante el crucero TALUD XII y a solicitud del Instituto Tecnológico del Mar, Mazatlán, se conservaron muestras de sedimentos para análisis químicos. Arrastres pelágicos En el TALUD I solo se realizó un muestreo de zooplancton utilizando una estructura tipo Bongo parecida a la que se usó en los cruceros SIPCO (1981-1982) y CORTES (1982-1985), con una luz de malla de 500 μm (Lám. 7 B). En el TALUD II, por primera vez se utilizó una red pelágica de tipo Isaacs-Kidd (Lám. 8 A) prestada por el grupo del Dr. V. Arenas (ICML, UNAM). Este mismo equipo se utilizó de manera más intensiva en el TALUD III y se lograron recolectar 10 muestras, las cuales fueron analizadas parcialmente en el contexto de un estudio de los camarones pelágicos del Pacífico mexicano (Hendrickx y Estrada-Navarrete 1996). En los cruceros TALUD IV a XII se usaron dos estructuras con redes de media agua para micronecton construidas en talleres locales, de aproximadamente 0.8 m2 de superficie de boca y con una malla de 1 mm de apertura (Lám. 7 C). Los muestreos se efectuaron oblicuamente, hasta una profundidad de 500 m y 1500 m, sin parada en el nivel más profundo para evitar un sobre-muestreo del estrato más profundo. En el transcurso del estudio, se le dio prioridad a las muestras a 1500 m de profundidad, pues en este caso se lograba muestrear la fauna pelágica ubicada por debajo de la ZMO. En la mayoría de los muestreos de zooplancton y de micronecton de las campañas TALUD se uso un medidor continuo de “tiempo y profundidad” Benthos O peraciones oceanográficas en aguas profundas 71 Lam. 6. A, draga de arrastre endobentónica tipo Karling en proceso de recuperación; B, misma, vista frontal; C, nucleador de caja en proceso de inmersión; D, mismo en cubierta de operaciones. A B C D precalibrado (Lám. 7 D) que registro in situ la profundidad alcanzada por el equipo y la duración del muestreo. Arrastres bentónicos En los años 80, la plataforma de operaciones del B/O “El Puma” recibió de la Universidad Autónoma de Baja California Sur (UABCS) una draga tipo Agassiz que esta institución había utilizado en algunos de sus cruceros. Este equipo, con una anchura de 2.5 m y una altura de 1.0 m, tenía la ventaja de ser simétrico, por lo que no importa de que lado toca el fondo cuando se está bajando. La draga Agassiz fue el primer equipo de arrastre profundo que se utilizó en el proyecto y se desplegó en las campañas TALUD II y III, en conjunto con una red tipo camaronera de 80’ disponible en el barco. Ambos equipos fueron usados según las circunstancias: la red de 80’ en aguas someras y la draga Agassiz en aguas más profundas (Cuadro 1). El TALUD II 72 Biodiversidad y comunidades del talud continental del Pacífico mexicano Lám. 7. A, estructura de filtración de sedimentos con tamices de diferentes aperturas; B, estructura bongo para muestreo de zooplancton; C, red para micronecton en proceso de recuperación; D, misma, al termino del muestreo; se aprecia a la izquierda el medidor de tiempo y profundidad Benthos. A C B D presentó la particularidad de ser una combinación entre muestreos para el proyecto CEEMEX en aguas de la plataforma continental (véase Hendrickx et al. 1998) y muestreos propios del proyecto TALUD en aguas más profundas, por lo que se utilizó O peraciones oceanográficas en aguas profundas 73 en varias ocasiones la red de tipo camaronera de 80’. A partir del crucero TALUD IV se descartó el uso de redes y se construyó en el astillero Constructora y Reparadora de Buques, S.A. de C.V., en Mazatlán, Sinaloa, un trineo bentónico inspirado de varios diseños disponibles en la literatura (Sorbe 1983, Guennegan y Martin 1985) (Lám. 8 B). Este trineo bentónico, con una boca de 0.9 m de altura por 2.95 m de anchura, fue equipado de una malla tipo camaronera de 5.5 cm de apertura equipada de una red interna con malla más fina (ca 2.5 cm) para asegurar la captura de organismos pequeños. Este equipo se utilizó con mucho éxito en las campañas IV-XII. En la campaña TALUD XIII, se construyó en las instalaciones del ICMyL en Mazatlán una draga tipo Agassiz de tamaño menor a aquella utilizada en los cruceros TALUD II-III (Lám. 8 C). Este equipo se uso en la cuenca del Carmen, donde las condiciones de muestreo son muy difíciles debido a la poca anchura de los pisos batimétricos. Este equipo fue bajado en posición vertical, con el buque parado, iniciándose el arrastre una vez que tocaba fondo; esta estrategia aumentó la precisión del muestreo y permitió evitar el largo recorrido necesario para el despliegue del trineo bentónico usado en las campañas IV a XII. Instituciones participantes El proyecto TALUD es fundamentalmente un proyecto del Instituto de Ciencias del Mar y Limnología de la UNAM. Sin embargo, durante su desarrollo, se incorporaron académicos de otras instituciones con diferentes especialidades. Consecuentemente, el grupo TALUD está actualmente conformado por nueve científicos (de los cuales siete son investigadores principales) perteneciendo a cuatro instituciones; además, algunos investigadores participaron de manera incidental u ocasionalmente en las campañas o bien recibieron muestras específicamente recolectadas para algún tipo de estudio (Cuadro 3). La tripulación del B/O “El Puma” tubo un papel destacado en todos los cruceros que se llevaron a cabo entre 1989 y 2009. En total, fueron 30 los tripulantes quienes se embarcaron durante estas campañas (Cuadro 4) y el equipo científico siempre pudo contar con su ayuda y con una actitud positiva y constructiva. Durante las campañas, se fomentó la participación de estudiantes, tanto a nivel de licenciatura como de posgrado, y se pudo contar con el apoyo de algunos ayudantes voluntarios con experiencia demostrada. En total, fueron 70 los miembros del personal científico perteneciendo a estas dos categorías (Cuadro 5). 74 Biodiversidad y comunidades del talud continental del Pacífico mexicano Lam. 8. A, red de media agua tipo Isaacs Kidd en proceso de recuperación; B, trineo bentónico en proceso de recuperación; C, draga tipo Agassiz. A B C Apoyos recibidos Una parte significativa de los gastos ocasionados por la realización de las campañas (con excepción de los gastos de uso del buque) fueron cubiertos por los presupuestos institucionales de los investigadores participantes. Todos los cruceros organizados recibieron el apoyo de la UNAM, ya sea a través del ICMyL o del CTIC, en lo que se refiere al gasto operativo del buque (i.e., combustibles, avituallamiento, etc .). Un apoyo recibido de Conacyt (proyecto 31805-N; véase infra) sirvió para compensar parte de los gastos ocasionados por la operación de el B/O “El Puma” durante las campañas TALUD IV-VII. En total se usaron unos 90 días de navegación de los cuales unos 22 fueron utilizados para llegar a las áreas de trabajo. O peraciones oceanográficas en aguas profundas 75 Cuadro 3. Lista del personal académico que participó en las campañas oceanográficas TALUD I-XIII entre 1989 y 2009. Se indican los temas principales de interés y de actividades. NP, no participó en las campañas. Nombre Institución Temas de interés principales Campañas TALUD Michel E. Hendrickx (1) ICML-UNAM I, III y IV-XIII Nuria Méndez Ubach Samuel Goméz Noguera David Serrano ICML-UNAM ICML-UNAM Moluscos-CrustáceosEquinodermos Poliquetos Meiofauna IV-XIII IV, VI-X, XIII Hidrología IX-XIII Peces VIII, X-XIII Peces VIII-X, XII Hugo Aguirre Juan Madrid Vera José Salgado Barragan Marcelo García Guerrero(2) Pablo Zamorano de Haro(3) Jorge Ruelas(5) A. Carolina Ruiz Fernandéz María Ana Tovar Hernández Albert van der Heiden Facultad de Ciencias del mar UAS Instituto Nacional de la Pesca Instituto Nacional de la Pesca ICML-UNAM ICML-UNAM (postdoc) RARE Conservation, BCS Instituto Tecnológico del Mar ICML-UNAM Plancton-Bentos-Winkler IV-XIII Crustáceos VII, VIII Moluscos-Registros X Agua-Sedimentos IV CTD-Winkler; peces VI ICML-UNAM (postdoc) ICML-UNAM Poliquetos XII, XIII Peces I-III CSIC Barcelona CSIC Barcelona CSIC Barcelona CSIC Barcelona CSIC Barcelona Secretaría de Marina ICML-UNAM ICML-UNAM (DF) Peces Peces Hidrología Crustáceos Cefalópodos Hidrología Esponjas Sedimentos II II II II II II IV, V III (4) Pedro Rubies Beatriz Morales Nin Mario Manriquez Francisco Sarda Pilar Sanchez Eladio García Patrón José Luis Carballo Adolfo Molina Cruz 76 Biodiversidad y comunidades del talud continental del Pacífico mexicano Cuadro 3. Continúa. Nombre Institución Temas de interés principales R. Thunell University of South Sedimentos Carolina ICML-UNAM Fitoplancton Roberto Cortés Altamirano(5) Arturo Toledano Granados(6) ICML-UNAM Arturo Nuñez Pasten(6) ICML-UNAM Moluscos-BentosPlancton Plancton-Bentos Sergio Rendón Rodríguez(6) Alfredo Galaviz Solís(6) Alberto Castro del Río(6) Humberto Bojórquez Leyva(6) ICML-UNAM ICML-UNAM ICML-UNAM ICML-UNAM Plancton-Bentos Registros Registros-Sedimentos CTD-Winkler Campañas TALUD III NP IV-VII III-VII y IX-XII XII, XIII IV-IX, XI, XIII II-V, X, XI V Responsable del proyecto y de las campañas Actualmente CIDIR Oaxaca (3) Actualmente INE México D.F. (4) Actualmente CIAD Mazatlán. Responsable del TALUD II. (5) Análisis de muestras de los cruceros (6) Apoyo técnico (1) (2) Cuadro 4. Lista de los miembros de la tripulación del B/O “El Puma” que participaron en las campañas TALUD I-XIII entre 1989 y 2009. Cap. Alt., Capitán de altura; Ing., Ingeniero; Oper. esp., Operador especialista; Ctm., Contramaestre; Tim., Timonel; Coc., Cocinero; A. coc., Ayudante de cocinero, Cam., Camarista. Cap. Alt. Marcelino González Durán (QEPD) Cap. Alt. Pascual Barajas Flores Cap. Alt. Héctor Ulises Gutiérrez Granja Cap. Alt. Ezequiel Benjamín Vázquez Ibarra Cap. Alt. Adrián Cantú Alvarado Cap. Alt. Manuel A. Martínez Díaz Cap. Alt. José Osuna Astorga Ing. Jesús Octavio Chávez Osuna Ing. Julio Alfonso Ruiz Ramírez Oper. esp. Miguel Angel Aguilar Guzmán Oper. esp. Arturo Trujillo Meza Oper. esp. Rodolfo Fernández Rodríguez Ctm. Juan Toto Fiscal Ctm. Federico Zamora Lizárraga Tim. Hernán Tirado Galindo Tim. Abel Francisco Mendia González Tim. Laurencio Triana Valles Tim. Adrián Rodríguez Castillo O peraciones oceanográficas en aguas profundas 77 Cuadro 4. Continúa. Ing. Aurelio Ovalle Martínez Ing. Luís Castañeda Segura Ing. René Abel García Torres Ing. Fernando Lavín Zatarain Ing. Manuel Suárez Sierra Ing. José de Jesús Hernández Flores Coc. Baudelio Muñóz Mota Coc. Ramón Flores Murguía Coc. Ernesto Soriano Padilla A. coc. Mauricio Rodríguez Aragón Cam. Manuel Rodríguez Aragón Cam. José Félix Flores Sandoval Cuadro 5. Lista de estudiantes y ayudantes que participaron en las campañas oceanográficas TALUD I-XIII entre 1989 y 2009. Nombre TALUD Nombre TALUD Luis Del Cerro Héctor Plascencia González Luis Miguel Valadez Manzano Sandra Guido Sánchez Saúl Rogelio Guerrero Galván Jaime Rodríguez García Felipe Silva Martínez Gustavo Colado Uribe Israel Castro Leal Yolanda Ortíz Gaona Claudia M. Agraz Hernández Manuel Alvarez Mendoza Laura Vázquez Cureño II II,III II II II II II II II II II II III VI VI VII VII VII VIII VIII-X VIII VIII VIII, X, XII, XIII VIII VIII VIII, X Dora Patricia Sánchez Vargas A. De M. Mexia Hernández G. Rodríguez Ochoa III III III Flor Delia Estrada Navarrete Sergio Mussot Pérez J.C. Milan A. José Salgado Barragán Jesús Alonso Esparza Haro Efraín Barranco Ramírez III III III III III IV, V Verónica Maldonado Sánchez Manuel Peñuelas Román Ismael Diego Núñez Riboni Pilar Pérez Pérez Enrique Ávila Torres Oscar Ricardo Guzón Zatarain Pablo Zamorano De Haro Julio César Herrera Arriaga Febe Elizabeth Vargas Arriaga Francisco Neptalí Morales Serna Ramón Vázquez Núñez Manuel Ayón Parente Manuel Leonardo Camacho Cruz José Soledad Ibarra Rivera Alondra Martínez Hernández Kissy Fabiola Rodríguez Soberanes Daniel de Jesús Moreno Flores Eva Visauta Girbau Betel Martínez Guerrero Georgina Quevedo Pacheco Juan Luis Sánchez Tellez Alba Lucia Castellanos Cendales 78 Biodiversidad y comunidades del talud continental del Pacífico mexicano IX-XII IX IX IX-XI IX, X X, XI X, XI XI XI Cuadro 5. Continúa. Nombre TALUD Nombre TALUD Alejandro Nava Aybar César P. Sánchez Cañedo Iyari Myotzi Bustos Hernández María del Carmen Espinosa Pérez Cristina Vega Juárez Alfredo Flores Reyes Benjamín Yáñez Chávez Héctor Nava Bravo Miguel Angel Díaz Flores Laura Leonor Navarro Zazueta Ernesto Sánchez Rojas Zaira L. Hernández Inda IV IV IV-VII IV-VI Lilia Catherinne Soler Jiménez Luis Sauma Castillo Dilian N. Anguas Cabrera Carlo Magno Zárate Montes XI XI XI-XIII XII, XIII IV V-VII V-VII V-VII V, VI V, VI V VI, VII XII, XIII XII XII, XIII XIII XIII XIII XIII XIII José Antonio Cruz Barraza VI Ana Cristina Quankiu Rascón Carolina Salas Singh Agustina Ferrando Ostoni Jesús Ramón Rendón Martínez Osiris Chávez Vargas Francisco Vásquez Melchor Omar Hernández Tovalin María de los Ángeles Barrón García Ricardo Colima Palacios XIII En el periodo correspondiente a 2000-2002 se consiguió el apoyo de Conacyt a través del proyecto 31805-N ($ 525,000.00 MN en tres años). En 2006, se logró conseguir un apoyo de $ 403, 600.00 MN por parte de la UNAM, a través del proyecto PAPIIT IN217306-3 (Dirección General de Apoyo al Personal Académico, UNAM), también por una duración de tres años. De manera conservadora, se estima que la participación presupuestal adicional por parte del personal académico participante (i.e., gastos menores, tanto para actividades de laboratorio como en adquisición de material y de reactivos usados en el buque; mantenimiento de equipos) fue del orden de $ 120,000.00 MN. Resultados La gran cantidad de información obtenida y de material biológico recolectado durante el proyecto TALUD han permitido alcanzar una producción científica destacada que consiste en 21 comunicaciones en reuniones científicas (Cuadro 6) y 46 publicaciones (sin contar los 13 capítulos de este libro) en diversos medios de difusión del conocimiento, tanto a nivel nacional como internacional. Estas publicaciones incluyen 38 basadas integralmente en resultados de las campañas TALUD, así como otras O peraciones oceanográficas en aguas profundas 79 Cuadro 6. Contribuciones presentadas en foros nacionales e internacionales basadas en resultados y avances obtenidos a partir de los datos oceanográficos y biológicos obtenidos durante las campañas TALUD. Lugar y tipo de evento Título de la presentación y formato Première Conférence Européenne sur les Crustacés. Paris, Francia. 31 de agosto al 5 de septiembre de 1992. 2nd European Crustacean Conference. Lieja, Bélgica. 1-6 de septiembre de 1996. Crustacés de profondeur du golfe de Californie, Mexique. M.E. Hendrickx. Oral. Seventh Benelux Congress of Zoology. Bruselas, Bélgica. 24-25 de noviembre de 2000. EDFAM Conference “Life History, Assessment and Management of Crustacean Fisheries”. La Coruña, España. 8-12 de octubre de 2001. Eighth Benelux Congress of Zoology. Nijmegen, Netherlands. 23-24 de noviembre de 2001. Eighth Benelux Congress of Zoology. Nijmegen, Netherlands. 23-24 de noviembre de 2001. The Crustacean Society Meeting. Glasgow, Escocia. 18-23 de julio de 2005. XVIII Congreso Nacional de Zoología. Monterrey, N.L., México. 4-7 de octubre de 2005. X Congreso Asociación de Investigadores del Mar de Cortés, A.C. Mazatlán, Sin., México. 25-28 de octubre de 2005. XIV Congreso Nacional de Oceanografía. Manzanillo, Colima, México. 15-19 de mayo de 2006. 80 Distribution, biology and biochemical composition of Heterocarpus vicarius Faxon (Crustacea: Decapoda: Caridea: Pandalidae) from the SE Gulf of California, Mexico. M.E. Hendrickx, F. Páez-Osuna y H.M. Zazueta-Padilla. Cartel. Exploring the SE Gulf of California continental slope: 100 years after the “Albatross”. M.E. Hendrickx. Oral. Deep water shrimps on the continental slope of the SE Gulf of California, Mexico. M.E. Hendrickx. Cartel. Estimation of body size and weight of four species of deep water shrimps in the SE Gulf of California, Mexico. M.E. Hendrickx. Cartel. Fishery potential of the Pacific lobsterette Nephropsis occidentalis in the SE Gulf of California, Mexico. M.E. Hendrickx. Cartel. Diversity and ecology of deep water decapod crustaceans in the southern Gulf of California, Mexico. M.E. Hendrickx. Cartel. El papel de la profundidad y la concentración de oxígeno sobre la comunidad de moluscos del talud continental del sureste del golfo de California. P. Zamorano, M.E. Hendrickx y A. Toledano Granados. Oral. Análisis latitudinal de la comunidad de moluscos en el mar profundo del Mar de Cortés. P. Zamorano y M.E. Hendrickx. Oral. Análisis de sensibilidad aplicado a los índices de diversidad: un ejemplo de la comunidad de moluscos de aguas profundas en el sur del golfo de California. P. Zamorano y M.E. Hendrickx. Oral. Biodiversidad y comunidades del talud continental del Pacífico mexicano Cuadro 6. Continúa. Lugar y tipo de evento Título de la presentación y formato X Reunión Nacional de Malacología y Conquiliología. Guadalajara, Jal. México. 18-22 de marzo de 2007. The Crustacean Society Meeting. Coquimbo, Chile. 14-17 de octubre de 2007. Biocenosis y distribución de los moluscos de aguas profundas en el Pacífico mexicano: una evaluación de los avances. P. Zamorano y M.E. Hendrickx. Oral. Description of the male genital structures of Hymenopenaeus doris (Faxon. 1893) based on a single specimen captured in the Gulf of California, Mexico. M.E. Hendrickx. Cartel. Efecto de la zona de mínimo de oxígeno sobre los corredores pesqueros a lo largo del Pacífico mexicano. M.E. Hendrickx y D.E. Serrano-Hernández. Oral. Pelagic shrimps collected during the TALUD IIIVII cruises, R/V “El Puma” , SE Gulf of California, Mexico. M.E. Hendrickx. Cartel. Proyecto TALUD: El mar profundo en el Pacífico mexicano. P. Zamorano, M. E. Hendrickx, N. Méndez, S. Gómez, H. Aguirre, J. Madrid y D. Serrano. Oral. Caracterización de las comunidades de anélidos poliquetos del talud continental del sureste del golfo de California. N. Méndez. Cartel. Distribución de Melinnampharete eoa (Polychaeta: Ampharetidae) en el golfo de California, México. N. Méndez. Cartel. Collection and quantification of deep-sea macrobenthos: a case study of the polychaete Melinnampharete gracilis. N. Méndez. Oral. Genetic paternity test of Cephalurus cephalus, lollipop catshark (Scyliorhinidae), by random amplified polymorphic DNA techniques. H. AguirreVillaseñor, S.A. García Gasca, J. Madrid-Vera y C. Salas-Singh. Cartel. New depth record of Coryphaenoides capito (Garman, 1899) bighead grenadier (Gadiformes: Macrouridae) from the Gulf of California, Mexico. H. Aguirre Villaseñor, D.J. Moreno-Flores, J. MadridVera y C. Salas-Singh. Cartel. Dibranchus spinosus (Garman, 1899) distribution in the Mexican central Pacific. C. Salas-Singh y H. Aguirre Villaseñor. Cartel. Congreso Nacional de Ciencia y Tecnología del Mar. Nuevo Vallarta, Nay., México. 29-31 de octubre de 2007. The Crustacean Society Meeting. Galveston, Texas, USA. 9-13 de junio de 2008. Seminario de Divulgación INE-ICML “La Frontera Final: El Océano Profundo”. México, D. F. 1 de diciembre de 2009. XIV Congreso Nacional de Ciencia y Tecnología del Mar, Nuevo Vallarta, Nay., México. 29-31 de octubre de 2007 II Simposio Latinoamericano de Polychaeta. Mar del Plata, Argentina. 9-11 de marzo de 2009. 12th Deep-Sea Biology Symposium. Reykjavík, Islandia. 7-11 de junio de 2010. 12th Deep-Sea Biology Symposium. Reykjavík, Islandia. 7-11 de junio de 2010. 12th Deep-Sea Biology Symposium. Reykjavík, Islandia. 7-11 de junio de 2010. 12th Deep-Sea Biology Symposium. Reykjavík, Islandia. 7-11 de junio de 2010. O peraciones oceanográficas en aguas profundas 81 ocho en las cuales se utilizó parte de estos resultados (Cuadro 7). Además existen, en varios rubros, trabajos concluidos que se encuentran en revisión en diversos medios de comunicación para su posible publicación. Se produjeron también seis tesis de licenciatura y de maestría (Monzalvo Santos 2003, Ibarra Rivera 2006, Zamorano de Haro 2006, Rodríguez Soberanes 2007, Moreno Flores 2009, Zarate Montes 2011) y hay una de doctorado en proceso y prácticamente concluida (Castillo Velázquez, datos no publicados). Los problemas para la clasificación de las especies, muchas de las cuales son mal conocidas y han sido citadas escasamente en la literatura, han retrasado la publicación de algunos resultados en revistas formales. Eso se complica más todavía por el hecho que, hoy en día, es prácticamente imposible conseguir el préstamo de material comparativo depositado en museos en el extranjero. Las trabas existentes a nivel de las aduanas locales y las restricciones internacionales para el manejo y el envío de muestras conservadas en líquido son obstáculos que imposibilitan, muy a menudo, completar los estudios de tipo taxonómico. Un rápido análisis de los resultados alcanzados durante el proyecto TALUD permite concluir que nunca se había conseguido tal cantidad de información acerca de la fauna de aguas profundas y de su hábitat en el Pacífico de México. De hecho, desde el punto de vista faunístico, la información obtenida por los académicos que participaron en las campañas TALUD es la mayor obtenida desde que se publicaron los resultados de las exploraciones realizadas por el buque “Albatross”. A partir de los resultados obtenidos durante las campañas TALUD y basándose en datos comparativos disponibles en la literatura, se puede resumir la situación de la siguiente manera. El número de especies en profundidades mayores a los 200 m respecto al número total de especies conocidas en el golfo de California es muy variable según el grupo faunístico: 245 de 680 para los anélidos Polychaeta, 33 de 1446 para los moluscos Gastropoda, 51 de 560 para los moluscos Bivalvia, 23 de 286 para los cangrejos Brachyura, 42 de 165 para el conjunto de los camarones y 20 de 144 para el grupo de langostas (y afines) (Hendrickx 1992a, 1996a, Hendrickx et al. 2005). En el caso de los poliquetos, tres familias contienen 44 especies casi exclusivas de fondos profundos (Ampharetidae 99 %; Maldanidae, 100 %; Sternaspidae, 66 %) (Méndez 2006, 2007). En el caso de los camarones, salvo contadas excepciones, solamente los Caridea (e.g., Pandalidae, Glyphocrangonidae, Crangonidae) tienen representantes en aguas profundas del golfo de California (Hendrickx 2003a). Los moluscos de aguas profundas alrededor de la península de Baja California y en el golfo de California ofrecen también un patrón particular de distribución (Zamorano y Hendrickx 2009, 2011). En el 82 Biodiversidad y comunidades del talud continental del Pacífico mexicano Cuadro 7. Contribuciones publicadas, aceptadas, en prensa o remitidas basadas en su totalidad o en parte en datos obtenidos durante las campañas TALUD Citas (año de publicación) Temática Información completa del proyecto TALUD (publicados) Hendrickx, M.E. (1996) Decápodos; distribución Hendrickx, M.E., F. Osuna-Páez, y H.M. Morfología; composición química Zazueta-Padilla (1998) Hendrickx, M.E. (2001) Comunidades de decápodos Hendrickx, M.E. (2002a) Camarones; nueva especie Hendrickx, M.E. (2002b) Camarones; distribución Hendrickx, M.E. (2003a) Camarones; pesquerías; abundancia Hendrickx, M.E. (2003b) Langostinos; distribución, y abundancia Hendrickx, M.E. (2003c) Langostas; distribución, y abundancia Hendrickx, M.E. (2004) Camarones; pesquerías; abundancia Hendrickx, M.E. y J. López (2006) Caracoles; distribución; abundancia Zamorano, P., M.E. Hendrickx y A. Toledano Moluscos; distribución Granados (2006) Méndez, N. (2006) Poliquetos; distribución; abundancia Méndez, N. (2007) Poliquetos; ecología Zamorano, P., M.E. Hendrickx y A. Toledano Moluscos; distribución Granados (2007) Hendrickx, M.E. y D. Serrano (2007) Hidrología; oxígeno mínimo Zamorano, P. y M.E. Hendrickx (2007) Moluscos; ecología Hendrickx, M.E. y M. García-Guerrero (2007) Camarones; larva Hendrickx, M.E. (2008b) Decápodos; distribución Ibarra Rivera, J.S. y M.E. Hendrickx (2008) Lofogastridos; taxonomía; claves; distribución Hendrickx, M.E. (2008c) Isópodos; distribución Méndez, N. (2009) Poliquetos; distribución; abundancia Zamorano, P. y M.E. Hendrickx (2009) Moluscos; distribución; zoogeografía Hendrickx, M.E. (2010) Camarones; nueva especie Hendrickx, M.E. y D. Serrano (2010) Hidrología; oxígeno mínimo; pesquerías Massin, C. y M.E. Hendrickx (2010) Holoturias; nueva especie Gómez, S., G.B. Deets, J.E. Kalman y F.N. Copépodos; parásitos; nueva especie Morales-Serna (2010) Hendrickx, M.E. y M. Ayón-Parente (2011) Langostinos; taxonomía O peraciones oceanográficas en aguas profundas 83 Cuadro 7. Continúa. Citas (año de publicación) Temática Hendrickx, M.E. y M. Ayón-Parente (2011) Langostinos; taxonomía Aguirre-Villaseñor, H. y R. Castillo-Velázquez Peces; distribución (2011) Zamorano, P. y M.E. Hendrickx (2010) Moluscos; comunidades; zoogeografía Hendrickx, M.E., C. Mah y C.M. Zárate-Montes Estrellas de mar; taxonomía; distribución (2010) Massin, C. y M.E. Hendrickx (2011) Holoturias; taxonomía; nuevas especies Serrano, D. y M.E. Hendrickx (2011) Hidrología; oxígeno mínimo; pesquerías Hendrickx, M.E., M. Ayón-Parente y D. Serrano Langostinos; distribución; zoogeografía (2011) Trabajos aceptados (A), en prensa (P) o remitidos (R) Zamorano, P., M.E. Hendrickx, N. Méndez, S. Aguas profundas de México Goméz, D. Serrano, H. Aguirre, J. Madrid y N. Morales-Serna 2012 (A) Zamorano, P. y M.E. Hendrickx (A) Moluscos; distribución; ecología Winfield, I., M. Ortiz y M.E. Hendrickx (R) Amphipoda; nueva especie Aguirre-Villaseñor, H., C. Salas-Singh, J. Martínez Peces; distribución y J. Madrid-Vera (R) Zamorano, P. y M.E. Moluscos; distribución Hendrickx (P) Información parcial del proyecto TALUD (publicados) Hendrickx, M.E. (1996b) Camarones; taxonomía; claves; distribución Hendrickx, M.E. y F.D. Estrada-Navarrete (1996) Camarones; taxonomía; claves; distribución Schiff, H. y M.E.Hendrickx (1997) Crustáceos; visión Hendrickx, M.E. (2003) Malacostraca; colección Ayón-Parente, M. y M.E. Hendrickx (2007) Isópodos, y decápodos; distribución Hendrickx, M.E. ( 2008a) Decápodos; distribución Hendrickx, M.E. y M.K. Wicksten (2011) Caridea; distribución Zamorano, P. y M.E. Hendrickx. (Aceptado) Lucinoma caso de los equinodermos, un fenómeno similar se presenta y solamente se conoce un número reducido de especies en grandes profundidades, aunque los estudios derivados de los cruceros TALUD han permitido describir ya tres especies nuevas (Massin y Hendrickx 2010, 2011). Desafortunadamente, no se cuenta con fuentes de información tan amplias para la zona SO del Pacífico mexicano y, en consecuencia, sería prematuro efectuar alguna comparación. En el caso de los peces, se 84 Biodiversidad y comunidades del talud continental del Pacífico mexicano pudo revisar el estatus taxonómico y la distribución de los Macrouridae y de algunas otras especies (Moreno Flores 2009, Aguirre-Villaseñor y Castillo-Velázquez datos no publicados, H. Aguirre-Villaseñor y J. Madrid-Vera, com. pers.). A partir de los 100-150 m, junto con una notable disminución de la temperatura del agua y de la concentración de oxígeno disuelto, se observa una franca disminución de la biodiversidad. En condiciones de oxigenación deficiente (< 1.0 ml l-1 O2), el número de especies de macro-crustáceos encontradas varía entre dos y seis. Ciertas especies (e.g., Heterocarpus vicarius, Pleuroncodes planipes Stimpson, 1860, Cancer johngarthi Carvacho, 1989, Solenocera mutator Burkenroad, 1938, Squilla biformis Bigelow, 1891) adaptadas a estas condiciones deficientes se vuelven dominantes o incluso exclusivas (Hendrickx 1985 1992b, 1995a, 1995b, 1995c, 1996a, 1996b, Hendrickx y Wicksten 1989, Hendrickx y Salgado Barragán 1991). Conforme aumenta la profundidad, las condiciones a nivel del fondo (valores epibentónicos) se mantienen muy adversas, casi anóxicas (i.e., ca 0.0 ml l-1 O2), y la macrofauna desaparece en su totalidad. En la columna de agua un fenómeno similar puede ser detectado a partir de los 100 m de profundidad (Parker 1963, Mathews et al. 1974, Alvarez-Borrego y Lara-Lara 1991, Hendrickx 1995a, 1996a, Hendrickx y Serrano 2007, 2010, Serrano y Hendrickx, 2011). Las condiciones favorables a la presencia de especies con metabolismo aeróbico reaparecen a partir de los 500-750 m (según la latitud y las condiciones oceánicas) y una fauna totalmente distinta aparece (Hendrickx 1992b, 1996a). Estas condiciones fueron comprobadas durante los cruceros TALUD (Hendrickx 2001, Zamorano et al. 2007, Ibarra-Rivera y Hendrickx 2008, Massin y Hendrickx 2011, Hendrickx et al. 2011). Los estudios realizados indican que esta fauna presenta características particulares en cuanto a su abundancia, su morfología y su fisiología (Hendrickx 1996a, 2001, 2003b, Schiff y Hendrickx 1997). Dada su gran abundancia, los poliquetos encontrados (e.g., la especie tubícola Amage scutata Moore, 1923) constituyen un componente importante de la infauna (Méndez 2006). Su importancia radica en el hecho que, en este hábitat con diversidad moderada, constituyen una parte significativa dentro de las cadenas tróficas, modifican la textura del sustrato y podrían servir de indicadores de “salud” (por su carácter sedentario) del medio ambiente profundo (Méndez 2007). Además, las campañas TALUD permitieron detectar la presencia de especies potencialmente importante para la pesca, tales como Heterocarpus affinis y Nephropsis occidentalis (Hendrickx 2003a, 2003c, 2004) y de taxones nuevos dentro del grupo de los crustáceos (Hendrickx 2002a, 2010, Roman-Contreras y Boyko 2007). El ámbito pelágico sigue un patrón semejante. En la franja de la plataforma exterO peraciones oceanográficas en aguas profundas 85 na o talud superior son pocas las especies epipelágicas (e.g., Lucifer typus H. Milne Edwards, 1837) (Hendrickx y Estrada-Navarrete 1994) y raras las especies mesopelágicas que toleran la disminución de la concentración de oxígeno disuelto (e.g., Gennadas sordidus Kemp, 1910, con una superficie branquial mayor que le permite tolerar ambientes hipóxicos) (Hendrickx y Estrada Navarrete 1989, 1996). En cambio, la fauna de Lophogastrida (géneros Gnathophausia y Neognathophausia) es abundante y se detectó la presencia de cinco especies en los muestreos realizados durante las campañas TALUD; este material, en particular, es el más abundante jamás registrado en aguas del golfo de California y del SO de México para este grupo de crustáceos (Ibarra-Rivera y Hendrickx 2008). En el caso específico de la meiofauna, se puede afirmar que las investigaciones previas en estas aguas son inexistentes; de hecho, el único grupo de especies de la meiofauna estudiado de manera sistemática en el Pacífico mexicano es aquel de los copépodos harpacticoideos (Gómez-Noguera y Hendrickx 1997). Aún en este caso, toda la información disponible se refiere a especies de aguas someras o costeras y existe un desconocimiento total de la biodiversidad, de la abundancia y de la importancia trófica de estas comunidades en fondos oceánicos del Pacífico este tropical (Gómez-Noguera y Morales-Serna 2012). El estudio paralelo de los copépodos parásitos de peces recolectados durante las campañas TALUD ha conducido a la descripción de una especie nueva (Gómez et al. 2010). En cuanto a los peces, se hicieron algunos descubrimientos importantes relacionados con la distribución de las especies y de su biología. Al igual que en el caso de los invertebrados, la fauna encontrada es totalmente distinta a la fauna de la plataforma continental y bien adaptada a los ambientes profundos (H. AguirreVillaseñor y J. Madrid-Vera, com. pers., Aguirre-Villaseñor y Moreno-Flores 2012, Aguirre-Villaseñor et al. 2012). Sobre la base del material recolectado durante los cruceros TALUD, se pudo encontrar y describir en total siete especies nuevas: una de copépodos, tres de crustáceos decápodos y tres de holoturias (Cuadro 7). Se estima, además, que quedan por describirse por lo menos otra especie de crustáceos decápodos, una estrella de mar, dos moluscos y un número indefinido (pero seguramente superior a 30) de copépodos harpacticoideos. Conclusiones El año 2010 fue considerado como el año internacional de la biodiversidad. Muchos 86 Biodiversidad y comunidades del talud continental del Pacífico mexicano países, instituciones e individuos celebraron este evento, incluyendo la UNAM, pero el dramático rezago a nivel mundial para conocer la flora y la fauna que nos rodea no ha sido entendido por las autoridades. Tal como lo señalan Feldmann y Manning (1992) y Salazar-Vallejo et al. (2007), seguimos padeciendo de una extraordinaria y muy preocupante falta de recursos en infraestructura humana, en equipos y en materiales para lograr alcanzar un ritmo adecuado en el estudio de las especies que viven en los ambientes naturales. Como lo subrayaron algunos especialistas, las especies desaparecen a un ritmo mucho mas rápido que el ritmo que hemos decidido adoptar para estudiarlas. La fauna que vive en los ambientes profundos del planeta es privilegiada, pues los efectos más peligrosos de las actividades humanas no han impactado de manera decisiva en su hábitat (Groombridge y Jenkins 2000). Pero las cosas están cambiando. Son cada vez mayores las cantidades de productos tóxicos y de desperdicios que la “civilización” está arrojando mar adentro; son cada vez más frecuentes los accidentes en las plataformas de perforaciones petroleras y la profundidad de operación de estas es cada vez mayor; aún si no llegan directamente a los grandes fondos marinos, los desechos que se dispersan en los mares y en las zonas costeras van llegando cada vez con mayor frecuencia a las grandes extensiones oceánicas, donde se degradan poco a poco, afectando los procesos biológicos y ecológicos de la zona eufótica (donde la luz sirve de motor a la vida), perturbando así las cadenas tróficas y modificando la dinámica en la transferencia de “energía” (vía las partículas que se hunden) hacia las aguas profundas. Los estudios de la biodiversidad están esencialmente basados en los estudios taxonómicos; a su vez, la taxonomía requiere de la exploración sostenida y prudente de los ambientes naturales, con el fin de poder completar la recolección o la observación de las plantas y de los animales que allí viven (Feldmann y Manning 1992). Las colecciones biológicas que resultaron de las grandes expediciones realizadas desde el siglo XIX todavía están disponibles para estudios adicionales y para propósitos comparativos. En el caso de los mares de México, la mayoría del material recolectado durante esta época se encuentra depositado en la Smithsonian Institution en Washington, en el Harvard Museum de la Universidad de Harvard, en el Museum of Natural History del condado de Los Angeles, California y en la colección de invertebrados marinos de la Scripps Institution of Oceanography, en La Jolla, California. Es a final de los años 30’s que se inició en México la formación de colecciones oficiales, acorde con el decreto presidencial que otorgó al Instituto de Biología de la UNAM la responsabilidad de organizar y cuidar grandes colecciones nacionales de la flora y la fauna O peraciones oceanográficas en aguas profundas 87 del País. Otras colecciones, hoy en día de gran importancia para el conocimiento biológico de México, fueron establecidas posteriormente (Llorente-Bousquets et al. 1999). En lo que se refiere a las especies marinas, las más importantes son probablemente aquellas resguardadas en la UNAM, tanto en su sede principal como en las sedes foráneas, en la Universidad de Nuevo León, en Monterrey y en el CIQRO, hoy Ecosur, en Chetumal. Otras colecciones interesantes están surgiendo en otros estados, y eso a pesar de la escasez de apoyos para este tipo de actividades. Las muestras recolectadas durante las campañas TALUD representaron, en muchos casos, un incremento significativo del número de especímenes conocidos para varias especies de aguas profundas del Pacifico este. El material recolectado y consignado en colecciones reconocidas a nivel nacional e internacional representa, sin lugar a dudas, el mayor conjunto de especímenes y de especies de estos ambientes disponibles en México. Considerando la extensión de las zonas oceánicas en la ZEE de México, donde la profundidad excede los 500 m, resulta apabullante el constatar que las campañas TALUD recorrieron menos del 10 % de esta superficie y que la red de estaciones fue muy poco densa. Por ejemplo, durante el proyecto TALUD se realizó en promedio un arrastre bentónico por aproximadamente 12000 km2 de extensión oceánica. Queda por explorarse la totalidad de la zona bajo la influencia de la corriente de California, todo el área del golfo de Tehuantepec, y toda la franja oceánica con profundidades superiores a 2000-2300 m en de la ZEE. El reto es enorme y seguramente rebasa la capacidad actual del País en materia de infraestructura y de apoyos financieros para la operación de embarcaciones en alta mar. Basándose en la experiencia adquirida durante el proyecto TALUD y considerando que la parte inexplorada más amplia tiene profundidades más allá de los 3000 m, un programa de exploración muy superficial de estas áreas requeriría de, por lo menos, cuatro años de muestreo continuo (unos 1200 días de actividades en alta mar) y 12 años de trabajo en los laboratorios para clasificar, estudiar y analizar las muestras y los datos obtenidos. El reto que representa el estudio de una de las “Últimas Fronteras” de México en materia de conocimiento faunístico básico está a la altura del carácter misterioso y atractivo que ofrecen las comunidades naturales que hicieron de las aguas profundas su dominio. Pero los retos pertenecen a unos de los aspectos más atractivos de la investigación científica. Sin ellos, no tendríamos tanta motivación ni tantos deseos de superarnos. El solo hecho que esta fauna está allí, intacta, esperándonos, es suficiente para convencernos que hay que salir a buscarla. 88 Biodiversidad y comunidades del talud continental del Pacífico mexicano Agradecimientos Se agradece al personal científico que participó en el proyecto TALUD y cuyo apoyo y dedicación permitieron llevar a cabo las actividades de muestreo durante las campañas. Se reconoce aquí la labor del personal técnico que participó de manera ocasional o regular en estas campañas: José Salgado Barragán, Arturo Núñez Pasten, Arturo Toledano Granados, Alfredo Galaviz Solís, Alberto Castro del Río, Sergio Rendón Rodríguez, Humberto Bojórquez. De igual manera, se agradece la participación de estudiantes y de ayudantes voluntarios de diversas instituciones durante las campañas. Se reconoce el apoyo recibido por parte de la Universidad Nacional Autónoma de México para la realización de los cruceros TALUD a bordo del B/O “El Puma”, ya sea vía el Instituto de Ciencias del Mar y Limnología (ICMyL), vía la Coordinación Técnica de la Investigación Científica (CTIC) o vía la Dirección General de Apoyo al Personal Académico (DGAPA; proyecto PAPIIT IN 217306-3). Asimismo, se agradece el apoyo recibido por parte del Conacyt, México (proyecto 31805-N). Agradecemos al Ifremer y a Danièle Lemercier por la autorización de reproducir la fotografía de la lámina 3, A. Se agradece a Mercedes Cordero Ruiz por la edición final de este manuscrito así como la preparación de las figuras y láminas. Finalmente, se agradece al Instituto Nacional de Ecología (INE), México, por invitar a los miembros del proyecto TALUD a elaborar esta obra, en particular a los editores Pablo Zamorano y Margarita Caso Chávez. Referencias Agassiz, A. 1898. Reports on the dredging operations off the west coast of Central America, Mexico and the Gulf of California by the U.S.S. “Albatross” during 1891. Preliminary report on the Echini. Bulletin of the Museum of Comparative Zoology at Harvard University 32(5): 71-86. Aguirre-Villaseñor, H. y R. Castillo-Velázquez. 2011. New depth record of Cherublemma emmelas, black brotula (Ophidiiformes: Ophidiidae) from the Gulf of California, Mexico. Revista Mexicana de Biodiversidad 82: 713-715. Aguirre-Villaseñor, H. y C. Salas-Singh. 2012. New records of the lollipop catshark Cephalurus cephalus (Scyliorhinidae) from the Gulf of California, Mexico. Revista Mexicana de Biodiversidad 83. Aguirre-Villaseñor, H., C. Salas-Singh y J. Madrid-Vera. 2012. Nuevo registro de la quimera prieta del Pacífico este, Hydrolagus melanophasma, James, Ebert, Long y Didier, O peraciones oceanográficas en aguas profundas 89 2009, en el Pacífico central mexicano. Pp. 395-404. En: P. Zamorano, M.E. Hendrickx y M. Caso (eds.). Biodiversidad y comunidades del talud continental del Pacífico mexicano. Instituto Nacional de Ecología, Secretaría del Medio Ambiente y Recursos Naturales (Semarnat). Aguirre-Villaseñor, H., C. Salas-Singh, J. Martínez y J. Madrid-Vera. New Eastern Pacific records of Hydrolagus melanophasma James, Ebert, Long and Didier with annotations of a juvenile female. Journal of Fish Biology (Aceptado). Alden, A. 2010. http://geology.about.com/od/regional_geology/a/deepseaobservs_3.htm. Alvarez-Borrego, S., y J.R. Lara-Lara. 1991. The physical environment and primary productivity of the Gulf of California. Pp. 555-567. En: J.P. Dauphin y B. Simoneit (eds.). The Gulf and Peninsular province of the Californias. American Association of Petroleum Geologist Memoir 47: 555-567. Anónimo, 2010. http://world-ocean.ru/en/history/vitiaz/past.php. Arana E.P. 2003. Experiencia chilena en faenas de pesca en aguas profundas y distantes: Evolución y Perspectivas. Pp. 57-79. En: R.E. Yánez (ed.). Actividad Pesquera y de acuicultura en Chile. 1. Aspectos globales. Ediciones Universitarias de Valparaíso. Chile. Arana, P.M. y R. Vega. 2000. Pesca exploratoria con espineles en aguas profundas en torno a la isla Robinson Crusoe (Archipiélago de Juan Fernández), Chile. Investigaciones Marinas 28: 219-230. Arciniega-Flores, J., V. Landa-Jaime y G. González-Sanson. 1998. Distribution and abundance of soft-bottom stomatopod crustaceans off the coast of Jalisco and Colima, Mexico. Ciencias Marinas 24: 169-181. Arciniega-Flores, J., J.E. Michel-Morfín y V. Landa-Jaime. 2008. Estructura poblacional de Portunus xantusii affinis (Brachyura: Portunidae) en fondos blandos de las costas de Jalisco y Colima, México. Pp. 1-16. En: M.E. Hendrickx (ed.) Contributions to the Study of East Pacific Crustaceans 5 (1). [Contribuciones al Estudio de los Crustáceos del Pacífico Este 5 (1)]. Instituto de Ciencias del Mar y Limnología, UNAM. México, D.F. Ayón-Parente, M. y M.E. Hendrickx. 2007. New records of crustaceans (Isopoda and Decapoda) along the Pacific coast of Mexico. Pp. 59-61 En: M.E. Hendrickx (ed.) Contributions to the Study of East Pacific Crustaceans 4(2) [Contribuciones al Estudio de los Crustáceos del Pacífico Este 4(2). Instituto de Ciencias del Mar y Limnología, UNAM. México, D.F. Benson, K.R. y P.F. Rehbock. 2002. Oceanographic History. The Pacific and Beyond. University of Washington Press, Seattle and London. 556 pp. Blake, J.A. 1985. Polychaeta from the vicinity of deep-sea geothermal vents in the eastern Pacific I: Euphrosinidae, Phyllodocidae, Hesionidae, Nereididae, Glyceridae, Dorvilleidae, 90 Biodiversidad y comunidades del talud continental del Pacífico mexicano Orbiniidae, and Maldanidae. Bulletin of the Biological Society of Washington 6: 67-101. Britannica. 2010. http://www.britannica.com/facts/5/925385/International-IndianOcean-Expedition-as-discussed-in-Indian-Ocean. Brusca, R.C., L.T. Findley, P.A. Hastings, M.E. Hendrickx, J. Torre Cosio y A.M. van der Heiden. 2005. Macrofaunal diversity in the Gulf of California. Pp. 179-203. En: Cartron, J.-L. E., G. Caballos y R.S. Felger (eds.). Biodiversity, Ecosystems and Conservation in Northern Mexico. Oxford University Press. Cartes, J.E. y F. Sardà. 1989. Feeding ecology of the deep-water aristeid crustacean Aristeus antennatus. Marine Ecology Progress Series 54: 229-238. Cartes, J.E., J.C. Sorbe y F. Sardà. 1994. Spatial distribution of deep-sea decapods and euphausiids near the bottom in the northwestern Mediterranean. Journal of Experimental Marine Biology and Ecology 179: 131-144. Caso, M.E. 1986. Los equinodermos del golfo de California colectados en las campañas SIPCO I-II-III a bordo del B/O “El Puma”. Anales del Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México 13 (1): 91-184. Castillo Velázquez, R. 2012. Alimentación, reproducción y edad y crecimiento de Cherublemma emmelas en el Golfo de California. Tesis de Doctorado. Universidad Nacional Autónoma de México, Posgrado en Ciencias del Mar y Limnología. Chan, T.-Y. y A. Crosnier. 1997. Crustacea Decapoda: Deep-sea shrimps of the genus Plesionika Bate, 1888 (Pandalidae) from French Polynesia, with descriptions of five new species. Pp. 187-234. En: A. Crosnier (ed.), Résultats des Campagnes Musorstom. Vol. 18. Mémoires du Museum National d’Histoire Naturelle, Paris 176. Childress J.J. y B.A. Seibel. 1998. Life at stable low oxygen levels: adaptations of animals to oceanic oxygen minimum layers. Journal of Experimental Biology 201: 1223-1232. CIAT. 2010. Comisión Interamericana del Atún Tropical. http://www.iattc.org/VesselListsSPN.htm. Clark, H.L. 1948. A report on the Echini of the warmer Eastern Pacific, based on the collections of the “Velero” III. Allan Hancock Pacific Expedition 8 (5): 225-351. Dall, W.H. 1908. Reports on the dredging operations off the west coast of Central America to the Galapagos, to the west coast of Mexico, and in the Gulf of California, in charge of Alexander Agassiz, carried on by the U.S. Fish Commission steamer “Albatross”, during 1891, Lieut. Commander Z.L. Tanner, U.S.N., commanding. XXXVII. Reports on the scientific results of the expedition to the eastern tropical Pacific, in charge of Alexander Agassiz, by the U.S. Fish Commission steamer “Albatross”, from October, 1904, to March, 1905, Lieut. Commander L.M. Garret, U.S.N., commanding. XIV. The Mollusca and Brachiopoda. Bulletin of the Museum of Comparative Zoology at O peraciones oceanográficas en aguas profundas 91 Harvard University 43 (6): 205-487. Dawson, E.Y. 1944. The marine algae of the Gulf of California. Allan Hancock Pacific Expeditions 3 (10): 189-454. Diaz, R.J. y R. Rosenberg. 1995. Marine benthic hypoxia: A review of its ecological effects and the behavioural responses of benthic macrofauna. Oceanography and Marine Biology Annual Review 33: 245-303. Escobar Briones, E. y L. A. Soto. 1993. Bentos del mar profundo en México. Pp. 106116. En: S.I. Salazar-Vallejo y N.E. González (eds.). Biodiversidad Marina y Costera de México. Comisión Nacional de Biodiversidad y CIQRO, México. Facebook. 2011. http://www.facebook.com/pages/Oceans/112038895488787. Farlex 2010. http://encyclopedia.farlex.com/Deep-sea+exploration. Faxon, W. 1893. Reports on the dredgings operations off the west coast of Central America to the Galapagos, to the west coast of Mexico, and in the Gulf of California, in charge of Alexander Agassiz, carried on by the U.S. Fish Commision Steamer “Albatross”, during 1891, Lieut. Commander Z.L. Tanner, U.S.N., commanding. VI. Preliminary descriptions of new species of Crustacea. Bulletin of the Museum of Comparative Zoology at Harvard University 24 (7): 149-220. Faxon, W. 1895. Reports on an exploration off the west coast of Mexico, Central and South America, and off the Galapagos Islands, in charge of Alexander Agassiz, carried on by the U.S. Fish Commission steamer “Albatross”, during 1891, Lieut. Commander Z.L. Tanner, U.S.N., commanding. XV. The stalk-eyed Crustacea. Memoirs of the Museum of Comparative Zoology at Harvard College, 18: 1-292. Feldmann, R.M. y R.B. Manning. 1992. Crisis in systematic biology in the “Age of Biodiversity”. Journal of Paleontology 66 (1): 157-158. Forest, J. 1985. La campagne MUSORSTOM II (1980). Compte rendu et liste des stations / The MUSORSTOM II Expedition (1980). Report and list of stations. Pp. 7-30. En: Résultats des campagnes MUSORSTOM Vol 2. Mémoires du Muséum national d’Histoire naturelle, Paris. Zoologie 133. Gage, J.D. y P.A. Tyler. 1992. Deep-sea biology: a natural history of organisms at the deep-sea floor. Cambridge University Press. Gran Bretaña. 504 pp. Galil, B. 1997. Crustacea Decapoda: A revision of the Indo-Pacific species of the genus Calappa Weber, 1795 (Calappidae). Pp. 271-335. En: A. Crosnier (ed.). Résultats des Campagnes MUSORSTOM, Vol. 18. Mémoires du Muséum national d’Histoire Naturelle, Paris. Sér. A, Zoologie 176. Garman, S. 1899. Reports on an exploration off the west coasts of Mexico, Central and South America, and off the Galapagos Islands, in charge of Alexander Agassiz, by the 92 Biodiversidad y comunidades del talud continental del Pacífico mexicano U.S. Fish Commission Steamer “Albatross,” during 1891, Lieut. Commander Z.L. Tanner, U.S.N., Commanding. XXVI. The Fishes. Memoirs of the Museum of Comparative Zoology at Harvard College 24: 1-431. Garth, J.S. 1958. Brachyura of the Pacific coast of America; Oxyrhyncha. Allan Hancock Pacific Expeditions 21 (1): 1-459. Gaxiola-Castro G., S. Álvarez-Borrego, S. Nájera-Martínez, A.R. Zirino. 2002. Efecto de las ondas internas en el fitoplancton del Golfo de California. Ciencias Marinas 28: 297-309. Godinez-Domínguez, E. y G. González-Sanson. 1998. Variation of the bathymetric distribution patterns of the macrobenthic fauna on the continental shelf off Jalisco and Colima, Mexico. Ciencias Marinas 24: 337-351. Godinez-Domínguez, E. y G. González-Sanson. 1999. Diversity of soft-bottom macroinvertebrates from the continental shelf off Jalisco and Colima, Mexico. Ciencias Marinas 25: 609-627. Gómez-Noguera, S. E. y M.E. Hendrickx. 1997. Distribution and abundance of meiofauna in a subtropical coastal lagoon in the southeastern Gulf of California Mexico. Marine Pollution Bulletin 34 (7): 582-587. Gómez, S. y F.N. Morales-Serna. 2012. Meiofauna de mar profundo del golfo de California: algunos aspectos acerca de la distribución y abundancia de Copepoda. Pp. 123-144. En: P. Zamorano, M.E. Hendrickx y M. Caso (eds.). Biodiversidad y comunidades del talud continental del Pacífico mexicano. Instituto Nacional de Ecología, Secretaría del Medio Ambiente y Recursos Naturales (Semarnat). Gómez, S., G.B. Deets, J.E. Kalman y F.N. Morales-Serna. 2010. Lophoura brevicollum n. sp. (Copepoda: Siphonostomatoida: Sphyriidae), a parasite of the smooth grenadier Nezumia liolepis (Gilbert, 1890) (Pisces: Macrouridae) from the Eastern Pacific, and a new record and new host of Lophoura unilobulata Castro R. and Gonzalez. Journal of Crustacean Biology 30 (1): 129-140. Grassle, J.F. 1989. Species diversity in deep-sea communities. Trends in Ecology and Evolution 4 (1): 12-15. Groombridge, B. y M.D. Jenkins. 2000. Global biodiversity. Earth’s living resources in the 21st century. UNEP – World Conservation Monitoring Centre. 352 pp. Groueff, S. 1973. L’homme et la mer. Larousse. Paris, Francia. 395 pp. Guennegan, Y. y V. Martin. 1985. Annexe 3. Techniques de prélevement. Pp. 571-602. En: L. Laubier y Cl. Monniot (eds.). Benthos. Peuplements profonds du golfe de Gascogne. IFREMER, Francia. . Hanley, J.R. y M. Burke. 1991. Polychaeta Polynoidae: Scaleworms of the Chesterfield Islands and Fairway Reefs, Coral Sea. Pp. 9-82. En: A. Crosnier (ed.). Résultats des O peraciones oceanográficas en aguas profundas 93 Campagnes MUSORSTOM, Vol. 8. Mémoires du Muséum national d’Histoire Naturelle Paris. Sér. A, Zoologie 151. Helly, J.J. y L.A. Levin. 2004. Global distribution of naturally occurring marine hypoxia on continental margins. Deep-Sea Research I 51: 1159-1168. Hendrickx, M.E. 1985. Diversidad de los macroinvertebrados bentónicos acompañantes del camarón en el área del Golfo de California y su importancia como recurso potencial. Cap. 3. Pp. 95-148. En: A. Yañez-Arancibia (ed.). Recursos potenciales pesqueros de México: La pesca del camarón. Programa Universitario de Alimentos, Instituto de Ciencias del Mar y Limnología, Instituto Nacional de la Pesca, Universidad Nacional Autónoma de México. México, D.F. Hendrickx, M.E. 1990. The stomatopod and decapod crustaceans collected during the GUAYTEC II Cruise in the Central Gulf of California, Mexico, with the description of a new species of Plesionika Bate (Caridea: Pandalidae). Revista de Biología Tropical 38 (1): 35-53. Hendrickx, M.E. 1992a. Crustacés de profondeur du golfe de Californie, Mexique. P. 64. En: Proceedings of the First European Crustacean Conference. Aug. 31-Sept. 5, 1992. Paris, Francia. Hendrickx, M.E. 1992b. Distribution and zoogeographic affinities of decapod crustaceans of the Gulf of California, Mexico. Proceedings of the San Diego Society of Natural History 20: 1-11. Hendrickx, M.E. 1993. Crustáceos decápodos del Pacífico Mexicano. Pp. 271-318. En: S.I. Salazar-Vallejo y N.E. González (eds.). Biodiversidad Marina y Costera de México. Comisión Nacional de Biodiversidad y CIQRO, México. Hendrickx, M.E. 1995a. CAMARONES. Pp. 417-537 En: W. Fischer, F. Krupp, W. Schneider, C. Sommer, K.E. Carpenter y V.H. Niem (eds.). Guía FAO para la identificación de especies para los fines de la pesca. Pacífico centro-oriental. Vol. I. Plantas e Invertebrados. FAO, Roma, Italia. . Hendrickx, M.E. 1995b. INTRODUCCIÓN. Pp. 1-7. En: W. Fischer, F. Krupp, W. Schneider, C. Sommer, K.E. Carpenter y V.H. Niem (eds.). Guía FAO para la identificación de especies para los fines de la pesca. Pacífico centro-oriental. Vol. I. Plantas e Invertebrados. FAO, Roma, Italia. Hendrickx, M.E. 1995c. CANGREJOS. Pp. 565-636. En: W. Fischer, F. Krupp, W. Schneider, C. Sommer, K.E. Carpenter y V.H. Niem (eds.). Guía FAO para la identificación de especies para los fines de la pesca. Pacífico centro-oriental. Vol. I. Plantas e Invertebrados. FAO, Roma, Italia. Hendrickx, M.E. 1996a. Habitats and biodiversity of decapod crustaceans in the SE Gulf of California, Mexico. Revista de Biología Tropical 44 (2A): 603-617. 94 Biodiversidad y comunidades del talud continental del Pacífico mexicano Hendrickx, M.E. 1996b. Los camarones Penaeoidea bentónicos (Crustacea: Decapoda: Dendrobranchiata) del Pacífico mexicano. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad e Instituto de Ciencias del Mar y Limnolología, Universidad Nacional Autónoma de México. México, D.F. 148 pp. Hendrickx, M.E. 1996c. New records of deep-water decapod crustaceans in the southeastern Gulf of California, Mexico. Revista de Biología Tropical 44 (2B): 945-947. Hendrickx, M.E. 2001. Occurrence of a continental slope decapod crustacean community along the edge of the minimum oxygen zone in the southeastern Gulf of California, Mexico. Belgian Journal of Zoology 131 (Suppl. 2): 95-109. Hendrickx, M.E. 2002a. A new deep water species of Odontozona Holthuis (Decapoda, Stenopodidae) from the Southern Gulf of California, Mexico. Crustaceana 75 (3-4): 405-412. Hendrickx, M.E. 2002b. New bathymetric record for the genus Sicyonia in the eastern Pacific. Oceanides 17 (2): 129. Hendrickx, M.E. 2003a. Size and abundance of deep water shrimps on the continental slope of the SE Gulf of California, Mexico. Pp. 227-234. En: M.E. Hendrickx (ed.) Contributions to the Study of East Pacific Crustaceans 2. [Contribuciones al Estudio de los Crustáceos del Pacífico Este 2]. Instituto de Ciencias del Mar y Limnología, UNAM. México, D.F. Hendrickx, M.E. 2003b. Geographic and bathymetric distributions of species of Munidopsis (Crustacea: Decapoda: Galathaeidae) in the SE Gulf of California, Mexico. Pp. 21-30. En: M.E. Hendrickx (ed.) Contributions to the Study of East Pacific Crustaceans 2. [Contribuciones al Estudio de los Crustáceos del Pacífico Este 2]. Instituto de Ciencias del Mar y Limnología, UNAM. México, D.F. Hendrickx, M.E. 2003c. Distribution and size of the deep water Pacific lobsterette, Nephropsis occidentalis Faxon, 1893 (Crustacea: Decapoda: Astacidea: Nephropidae) in the SE Gulf of California, Mexico. Crustaceana 76 (2): 207-215. Hendrickx, M.E. 2004. Distribution and estimation of body size and weight of four species of deep water shrimps in the SE Gulf of California, Mexico. Crustaceana 76 (9): 1025-1036. Hendrickx, M.E. 2008a. New records and notes on decapod crustaceans in the east Pacific. Crustaceana 81 (8): 999-1262. Hendrickx, M.E. 2008b. Second capture of Odontozona foresti Hendrickx, 2002 (Stenopodidea) and Munida bapensis Hendrickx, 2000 (Galatheoidea) in western Mexico. Pp. 47-50. En: M.E. Hendrickx (ed.) Contributions to the Study of East Pacific Crustaceans 5 (1). [Contribuciones al Estudio de los Crustáceos del O peraciones oceanográficas en aguas profundas 95 Pacífico Este 5 (1)]. Instituto de Ciencias del Mar y Limnología, UNAM. México, D.F. Hendrickx, M.E. 2008c. Rediscovery of Rocinela murilloi Brusca and Iverson, 1985 (Crustacea: Isopoda: Aegidae) in the Gulf of California, Mexico, and ecological data associated with its capture. Crustaceana 81(10): 1259-1262. Hendrickx, M.E. 2011. A new species of Glyphocrangon (Decapoda: Caridea: Glyphocrangonidae) from off the coast of western Mexico. Zootaxa 2372: 358-366. Hendrickx, M. E., M. Ayón Parente and D. Serrano. 2011. Additional record of Janetogalathea californiensis (Anomura: Galatheidae) from the central Gulf of California, Mexico, with notes on its distribution. Hidrbiologica 21(1) 89-94. Hendrickx, M.E. 2012a. Los Glyphocrangonidae y Crangonidae (Crustacea: Decapoda: Caridea) recolectados durante los cruceros TALUD en el Pacífico mexicano. Pp. 319356. En: P. Zamorano, M.E. Hendrickx y M. Caso (eds.). Biodiversidad y comunidades del talud continental del Pacífico mexicano. Instituto Nacional de Ecología, Secretaría del Medio Ambiente y Recursos Naturales (Semarnat). Hendrickx, M.E. 2012b. Distribución de Stereomastis pacifica (Faxon, 1893) (Crustacea; Decapoda; Palinura; Erynoidea; Polychelidae) en el Pacífico mexicano. Pp. 357-371. En: P. Zamorano, M.E. Hendrickx y M. Caso (eds.). Biodiversidad y comunidades del talud continental del Pacífico mexicano. Instituto Nacional de Ecología, Secretaría del Medio Ambiente y Recursos Naturales (Semarnat). Hendrickx, M.E. y M. Ayón-Parente. 2011. Rediscovery of Uroptychus pubescens Faxon, 1893 (Decapoda: Anomura: Chirostylidae) in the southern Gulf of California, Pacific coast of Mexico. Crustaceana 84 (12-23): 1411-1420. Hendrickx, M.E. y F.D. Estrada Navarrete. 1989. Processa pippinae Wicksten and Méndez, 1985: a pelagic processid shrimp from the Gulf of California. Revista de Biología Tropical 37 (1): 109-112. Hendrickx, M.E. y F.D. Estrada-Navarrete. 1994. Temperature related distribution of Lucifer typus (Crustacea: Decapoda) in the Gulf of California. Revista de Biología Tropical 42 (3): 581-586. Hendrickx, M.E. y F.D. Estrada-Navarrete. 1996. Los camarones Pelágicos (Crustacea: Dendrobranchiata y Caridea) del Pacífico mexicano. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad e Instituto de Ciencias del Mar y Limnolología, Universidad Nacional Autónoma de México. México, D.F. 157 pp. Hendrickx, M.E. y M. García-Guerrero. 2007. Description of the first zoea of Acanthephyra brevicarinata Hanamura, 1983 (Caridea: Oplophoridae) from deep water of the Gulf of California, Mexico. Pp. 31-37. En: M.E. Hendrickx (ed.) Contributions to the Study 96 Biodiversidad y comunidades del talud continental del Pacífico mexicano of East Pacific Crustaceans 4(2). [Contribuciones al Estudio de los Crustáceos del Pacífico Este 4(2). Instituto de Ciencias del Mar y Limnología, UNAM. México, D.F. Hendrickx, M.E. y J. López. 2006. Geographic and depth distribution of Bathybembix bairdii (Dall, 1889) (Mollusca, Gastropoda, Trochidae) in the East Pacific. Oceanides 21 (1-2): 93-99. Hendrickx, M.E. y J. Salgado Barragán. 1991. Los estomatópodos (Crustacea: Hoplocarida) del Pacífico mexicano. Publ. esp. del Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México 10: 1-200. Hendrickx, M.E. y J. Salgado Barragán. 1992. New records of two species of brachyuran crabs (Decapoda: Brachyura) from tropical coastal lagoons, Pacific coast of Mexico. Revista de Biología Tropical 40 (1): 149-150. Hendrickx, M.E. y J. Salgado Barragán. 1994. Stomatopods (Crustacea: Hoplocarida) collected off the coasts of Sinaloa, Mexico, during the BIOCAPESS Cruises IV, V and VI (August 1991, March and June 1992). Anales del Instituto de Biología, Universidad Nacional Autónoma de México 65 (2): 217-231. Hendrickx, M.E. y D. Serrano. 2007. Efecto de la zona de mínimo de oxígeno sobre los corredores pesqueros a lo largo del pacifico mexicano. Pp. 1-2. En: XIV Congreso Nacional de Ciencia y Tecnología del Mar. Memorias. Conferencia. Octubre 29 al 31 de 2007, Nuevo Vallarta, Nay. Hendrickx, M.E. y D. Serrano. 2010. Impacto de la zona de mínimo de oxígeno sobre los corredores pesqueros en el Pacífico mexicano. Interciencia 35 (1): 12-18. Hendrickx, M.E. y L.A. Vázquez-Cureño. 1998. Composition and zoogeographical affinities of the stomatopod and decapod crustaceans collected during the CEEMEX P4 cruise in the Gulf of Tehuantepec, Mexico. Bulletin de l’Institut royal des Sciences naturelles de Belgique 68: 135-144. Hendrickx, M.E. y M.K. Wicksten. 1989. Los Pandalidae (Crustacea: Caridea) del Pacífico mexicano, con una clave para su identificación. Caldasia 16 (76): 71-86. Hendrickx, M.E. y M.K. Wicksten. 2011. New distribution ranges and records of caridean shrimps (Crustacea: Decapoda: Caridea) from the west coast of Mexico. Hidrobiologica 21 (1): 26-33. Hendrickx, M.E., F. Osuna-Páez y H.M. Zazueta-Padilla. 1998. Biology and biochemical composition of the deep water shrimp Heterocarpus vicarius (Crustacea: Decapoda: Caridea: Pandalidae) from the southeastern Gulf of California, Mexico. Bulletin of Marine Science 63 (2): 265-275. Hendrickx, M.E., R.C. Brusca y L.T. Findley. 2005. A Distributional Checklist of the Macrofauna of the Gulf of California, Mexico. Part I. Invertebrates. [Listado y Distribución de la MacroO peraciones oceanográficas en aguas profundas 97 fauna del Golfo de California, México, Parte I. Invertebrados]. M.E. Hendrickx, R.C. Brusca y L.T. Findley (eds.). Arizona-Sonora Desert Museum. Tucson, Arizona. 429 pp. Hendrickx, M.E., C. Mah y C.M. Zárate Montes. 2011. Deep-water Asteroidea (Echinodermata) collected during the TALUD cruises in the Gulf of California, Mexico. Revista Mexicana de Biodiversidad 82: 798-824. Hernández-Alcántara, P. y V. Solís-Weiss. 1991. New records of errantiate polychaetous annelids from the Gulf of California. Bulletin of Marine Science 48 (2): 251-260. Hendrickx, M.E., M. Demestre, A. Esparza-Haro y J. Salgado-Barragán. 1997. Stomatopod and decapod crustaceans collected during the CEEMEX P5 and P7 cruises to the Gulf of Tehuantepec, Mexico. Oceanides 11: 1-28. Hernández-Alcántara, P. y V. Solís-Weiss. 1993a. Distribución latitudinal y batimétrica de los anélidos poliquetos del orden Terebellomorpha de la plataforma continental del Golfo de California, México. Cuadernos Mexicanos de Zoología 1 (2): 65-72. Hernández-Alcántara, P. y V. Solís-Weiss. 1993b. New records of sedentariate Polychaetous annelids from the continental shelf of the Gulf of California. Bulletin of Marine Science 53 (3): 1027-1041. Hernández-Alcántara, P. y V. Solís-Weiss. 1994. Los espiónidos (Polychaeta: Spionidae) del golfo de California y Golfo de Tehuantepec, México. Revista de Biología Tropical 42 (3): 567-577. Hernández-Alcántara, P. y V. Solís-Weiss. 1998a. Capitellids (Polychaeta: Capitellidae) from the continental shelf of the Gulf of California, Mexico, with the description of a new species, Notomastus angelicae. Proceedings of the Biological Society of Washington 111 (3): 708-719. Hernández-Alcántara, P. y V. Solís-Weiss. 1998b. Parasitism among polychaetes. A rare case illustratred by a new species: Labrorostratus zaragozensis, n. sp. (Oenonidae) found in the Gulf of California, Mexico. Journal of Parasitology 84 (5): 978-982. Herring, P.J. y M.R. Clarke. 1971. Deep Oceans. Praeger Publishers, New York. 320 pp. Ibarra Rivera, J.S. 2006. Los géneros Gnathophausia y Neognathophausia (Malacostraca: Lophogastrida) en el Pacífico mexicano. Tesis profesional. Instituto Tecnológico de Los Mochis, Sinaloa. 70 pp. Ibarra-Rivera, J.S. y M.E. Hendrickx. 2008. Los géneros Gnathophausia y Neognathophausia (Malacostraca: Lophogastrida) en el Pacífico mexicano. Pp. 55-93. En: M.E. Hendrickx (ed.) Contributions to the Study of East Pacific Crustaceans 5 (1). [Contribuciones al Estudio de los Crustáceos del Pacífico Este 5 (1)]. Instituto de Ciencias del Mar y Limnología, UNAM. México, D.F. Idronaut. 2010. http://www/gnc-inc.com/320a4/pdf. 98 Biodiversidad y comunidades del talud continental del Pacífico mexicano IFREMER, 2010. http://www.ifremer.fr/exploration/engin/cyana.htm. Kameya, A., R. Castillo, L. Escudero, E. Tello, V. Blaskovic, J. Córdova, Y. Hooker, M. Gutiérrez y S. Mayor. 1997. Localización, distribución y concentración de langostinos rojos de profundidad. Crucero BIC Humboldt 9607-08 18 de julio a 6 de agosto de 1996. Publicación Especial, Instituto del Mar de Perú. 47 pp. King, J.E. y R.T.B. Iversen. 1962. Midwater trawling for forage organisms in the Central Pacific 1951-1956. Fishery Bulletin 62 (210): 271-321. Laubier, L. 2010. http://www.universalis.fr/encyclopedie/cyana. Levin, L.A. 2003. Oxygen minimum zone benthos: adaptation and community response to hypoxia. Oceanography and Marine Biology: An Annual Review 41: 1-45. Levin, L.A. y J.D. Gage. 1998. Relationships between oxygen, organic matter and the diversity of bathyal macrofauna. Deep-Sea Research II 45: 129-163. Levin, L.A., R.J. Etter, M.A. Rex, A.J. Gooday, C.R. Smith, J. Pineda, C.T. Stuart, R.R. Hessler y D. Pawson. 2001. Environmental influences on regional deep-sea species diversity. Annual Review Ecology System 32: 51-93. Licea, S., J.L. Moreno, H. Santoyo y G. Figueroa. 1995. Dinoflagelados del golfo de California. Universidad Autónoma de Baja California Sur, SEP-FOMES, PROMARCO. México. 163 pp. Llorente-Bousquets, J., P. Koleff Osorio, H. Benítez Díaz y L. Lara Morales. 1999. Síntesis del estado de las colecciones biológicas mexicanas. Resultados de la encuesta “Inventario y diagnóstico de la actividad taxonómica en México” 1996-1998. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, México. México. 143 pp. Ludwig, H. 1894. Reports of an Exploration off the West Coast of Mexico, Central and South America, and off the Galapagos Islands, in Charge of Alexander Agassiz, by the U.S. Fish Commission Steamer “Albatross”, during 1891, Lieutenant Commander Z.L. Tanner, U.S.N., Commanding, XII: The Holothurioidea. Memoirs of the Museum of Comparative Zoology at Harvard College 17 (3): 1-183. Ludwig, H. 1905. Asteroidea. En: Reports on an Exploration off the West Coasts of Mexico, Central and South America, and off the Galapagos Islands, in Charge of Alexander Agassiz, by the U.S. Fish Commission Steamer “Albatross”, during 1891, Lieut. Commander ZL. Tanner, U.S.N., Commanding and Reports on the Scientific Results of the Expedition to the Tropical Pacific, in Charge of Alexander Agassiz, on the U.S. Fish Commission Steamer “Albatross”, from August, 1899, to March, 1900, Commander Jefferson F. Moser, U.S.N. Commanding. Memoirs of the Museum of Comparative Zoology at Harvard College 32:1-292. Luke, S.R. 1977. Catalog of the benthic invertebrate collections. I. Decapod Crustacea O peraciones oceanográficas en aguas profundas 99 and Stomatopoda. University of California. SIO Ref. Ser. 77-9. 72 p. Lütken, Chr. F. y Th. Mortensen. 1899. Reports on an exploration off the west coasts of Mexico, Central and South America, and off the Galapagos Islands, in charge of A. Agassiz, during 1891 on the “Albatross”. XXV. The Ophiuridae. Memoirs of the Museum of Comparative Zoology at Harvard College 23 (2): 93-208. Massin, C. y M.E. Hendrickx. 2010. A new species of deep-water Holothuroidea (Echinodermata) of the genus Synallactes from off western Mexico. Scientia Marina 74 (3): 599-603. Massin, C. y Hendrickx, M.E. 2011. Deep-water Holothuroidea (Echinodermata) collected during the TALUD cruises off the Pacific coast of Mexico, with the description of two new species. Revista Mexicana de Biodiversidad 82: 413-443. Mathews, C.P., J.L. Granados y J. Arvizu-Martinez. 1974. Results of the exploratory cruises of the Alejandro de Humboldt in the Gulf of California. CalCOFI Technical Report 17: 101-111. McClain, C.R. 2004. Connecting species richness, abundance and body size in deep-sea gastropods. Global Ecology and Biogeography 13 (4): 327-334. McClain, C.R. y M.A. Rex. 2001. The relationship between dissolved oxygen concentration and maximum size in deep-sea turrid gastropods: an application of quantile regression. Marine Biology 139: 681-685. McCosker, J.E. 1999. Pisces anguilliformes: Deepwater snake eels (Ophichthidae) from the New Caledonian region, Southwest Pacific Ocean. Pp. 180: 571-588. En: A. Crosnier (ed.). Résultats des Campagnes MUSORSTOM, Vol. 20. Mémoires du Muséum national d’Histoire Naturelle Paris. Sér. A, Zoologie. Méndez, N. 2006. Deep-water polychaetes (Annelida) from the southeastern Gulf of California, Mexico. Revista de Biología Tropical 54 (3): 773-785. Méndez, N. 2007. Relationships between deep-water polychaete fauna and environmental factors in the southeastern Gulf of California, Mexico. Scientia Marina 71 (3): 605-622. Méndez, N. 2009. Distribution and extraordinary abundance of the deep-sea Melinnampharete gracilis Hartman, 1969 (Polychaeta: Ampharetidae) in the Gulf of California, Mexico. Cahiers de Biologie Marine 50: 273-276. Monzalvo Santos, I.K. 2003. Determinación de mercurio en dos especies de peces bentónicos (Cherublemma emmelas y Zalietus elater) de la zona del talud del Golfo de California. Tesis de Licenciatura, Escuela de Biología, UAS. 82 pp. Moreno Flores, D.J. 2009. Distribución de la familia Macrouridae Gilbert y Hubbs 1916, en el sur del golfo de California. Tesis de Maestría, Ciencias del Mar y Limnología, UNAM. 74 pp. 100 Biodiversidad y comunidades del talud continental del Pacífico mexicano Moreno, J.L., S. Licea y H. Santoyo. 1996. Diatomeas del golfo de California. UABCS, SEP-FOMES, PROMARCO. 165 pp. Parker, R.H. 1963. Zoogeography and ecology of some macroinvertebrates, particularly mollusks, in the Gulf of California and the continental slope off Mexico. Videnskabelige Meddelelser fra Dansk Naturhistorisk Forening 126:1-178. Retamal R., M.A. 1993. Crustáceos decápodos abisales de la zona Iquique-Arica. Estudios Oceanológicos 12: 1-8. Rodríguez Soberanes, K. 2007. Variación espacial de la densidad de copépodos bentónicos y su relación con la profundidad, concentración de oxígeno disuelto, concentración de carbono orgánico y granulometría del sedimentos en el talud continental de Sinaloa. Tesis profesional. Instituto Tecnológico de Mazatlán. Mazatlán, Sinaloa. 107 pp. Rogers, A.D. 2000. The role of the oceanic oxygen minima in generating biodiversity in the deep sea. Deep-Sea Research II 47: 119-148. Román-Contreras, R. y C. Boyko. 2007. A new genus and species of Bopyrid isopod infesting the crab Munidopsis depressa (Anomura: Galatheidae) from the Gulf of California, with notes on its ecology. Journal of Crustacean Biology 27 (2): 370-379. SAGARPA. 2010. Boletín enero 2010. Salazar-Vallejo, S.I., E. Escobar-Briones, N.E. González, E. Suárez-Morales, F. Alvarez, J.A. de León-González y M.E. Hendrickx. 2007. Iniciativa Mexicana en Taxonomía: Biota Marina y Costera. Ciencia y Mar 11 (32): 3-11. Sánchez-Velasco, L., V.M. Godínez-Sandoval, B. Shirasago y M. Peguero-Icaza. 2004. Datos hidrográficos del golfo de California: Campaña oceanográfica GOLCA0308 (del 1 al 7 de agosto del 2003). Informe Técnico 16560. Comunicaciones Académicas. Serie Oceanografía Física. CICESE. 44 pp. Sardà, F. y M. Demestre. 1987. Estudio biológico de la gamba Aristeus antennatus (Risso, 1816) en el mar catalán (NE de España). Investigaciones Pesqueras 51 (Suppl. 1): 213-232. Sardà, F. y F.J. Valladares. 1990. Gastric evacuation of different foods by Nephrops norvegicus (Crustacea Decapoda) and estimation of soft tissue ingested, maximum food intake and cannibalism in captivity. Marine Biology 104: 25-30. Schiff, H. y M.E. Hendrickx. 1997. An introductory survey of ecology and sensory receptors of tropical eastern Pacific crustaceans. Italian Journal of Zoology 64: 13-30. Scripps Institution of Oceanography. 1992. The marine shrimp of the Scripps Institution of Oceanography Benthic Invertebrate collection catalogue. Mimeographed report. 106 pp. Serrano, D. 2012. La zona del mínimo oxígeno en el Pacífico mexicano. Pp. 105-119. O peraciones oceanográficas en aguas profundas 101 En: P. Zamorano, M.E. Hendrickx y M. Caso (eds.). Biodiversidad y comunidades del talud continental del Pacífico mexicano. Instituto Nacional de Ecología, Secretaría del Medio Ambiente y Recursos Naturales (Semarnat). Serrano, D. y M.E. Hendrickx. 2011. Debajo de la zona del mínimo de oxígeno. Investigación y Ciencia 417: 13-15. Smith, R.H. 1992. Planet Earth. Time-Life, EE.UU. 113 pp. Smith, C.R., L.A. Levin y S. Mullineaux. 1998. Deep-sea biodiversity: a tribute to Robert R. Hessler. Deep-Sea Research II 45: 1-11. Solís-Weiss, V. y P. Hernández-Alcántara. 1994. Amphisamytha fauchaldi: A new species of Ampharetid (Annelida: Polychaeta) from the hydrothermal vents at Guaymas Basin, Mexico. Bulletin of the Southern California Academy of Sciences 93 (3): 127-134. Solís-Weiss, V. y B. Hilbig, 1992. Redescription of Ophryotrocha platykephale Blake (Polychaeta, Dorvilleidae) from the Guaymas Basin hydrothermal vents. Bulletin of the Southern California Academy of Sciences 91 (2): 92-96. Sorbe, J.P. 1983. Description d’un traîneau destiné à l’échantillonnage quantitatif étagé de la faune suprabenthique néritique. Annales de l’Institut Océanographique, París. 59 (2): 117-126. Soule, G. 1970. The greatest depths: Probing the seas to 20000 feet (6100 m) and below. Macrae Smith. Philadelphia, EE.UU. 194 pp. Strong, A.M. y L.G. Hertlein. 1939. Marine mollusks from Panama collected by the Allan Hancock Expedition to the Galapagos Islands, 1931-1932. Allan Hancock Pacific Expeditions 2 (12): 177-245. Thomson, C.W. 1873. The Depths of the Sea. Octavio. MacMillan and Co. London. 527 pp. Vadon, C. 1991. Echinodermata: Ophiuridae profonds de Nouvelle-Calédonie. Formes paedomorphes. Pp. 335-356. En: A. Crosnier (ed.). Résultats des Campagnes MUSORSTOM, Vol. 8. Mémoires du Muséum national d’Histoire naturelle, Paris. Sér. A, Zoologie 151. Vélez, J., A. Kameya, C. Yamashiro, N. Lostaunau y O. Valiente. 1992. Investigación del recurso potencial langostino rojo de profundidad a bordo del BIC “Fridtjof Nansen” (25 de abril - 25 de mayo, 1990). Informe. Instituto del Mar del Perú, C.E.E. 104: 3-24. Wehrtmann, I.S. y V. Nielsen-Muñoz. 2009. The deepwater fishery along the Pacific Costa Rica, Central America. En: P. Arana, J.A.A. Pérez y P.R. Pezzutto (eds.). Deep-sea fisheries off Latin America. Latin American Journal of Aquatic Research 37 (3): 543-554. Wehrtmann, I.S., J. Herrera-Correal, R. Vargas y P. Hernáez. 2010. Squat lobsters (Decapoda: Anomura: Galatheidae) from deepwater Pacific Costa Rica: species diversity, 102 Biodiversidad y comunidades del talud continental del Pacífico mexicano spatial and bathymetric distribution. Nauplius 18 (1): 69-77. WHOI. 2007. http://www.whoi.edu/page.do?pid=8422. Wicksten, M.K. 1989. Ranges of offshore decapod crustaceans in the eastern Pacific Ocean. Transactions of the San Diego Society of Natural History 21 (19): 291-316. Wicksten, M.K. y M.E. Hendrickx. 1992. Checklist of Penaeoid and Caridean shrimps (Decapoda: Penaeoidea, Caridea) from the eastern tropical Pacific. Proceedings of the San Diego Society of Natural History 9: 1-11. Wicksten, M.K. y M.E. Hendrickx. 2003. An updated checklist of benthic marine and brackish water shrimps (Decapoda: Penaoidea, Stenopodidea, Caridea) from the Eastern Tropical Pacific. Pp. 49-76. En: M.E. Hendrickx (ed.) Contributions to the Study of East Pacific Crustaceans 2. [Contribuciones al Estudio de los Crustáceos del Pacífico Este 2]. Instituto de Ciencias del Mar y Limnología, UNAM. México, D.F. Wikipedia. 2010. http://es.wikipedia.org/wiki/Batisfera. Wri-Org. 2010. http://earthtrends.wri.org/text/coastal-marine/variable-62.html. Zamorano de Haro, P. 2006. Moluscos del Talud Continental del Pacífico Mexicano (Taxonomía, Biodiversidad, Ecología, Biogeografía). Tesis de Maestría. Universidad Nacional Autónoma de México, Posgrado en Ciencias del Mar y Limnología. 124 pp. Zamorano, P. y M.E. Hendrickx. 2007. Biocenosis y distribución de los moluscos de aguas profundas en el Pacífico mexicano: una evaluación de los avances. Pp. 48-49. En: E. Ríos-Jara, M.C. Esqueda-González y C.M. Galván-Villa (eds.). Estudios sobre la malacología y conquiliología en México. Universidad de Guadalajara, México. Zamorano, P. y M.E. Hendrickx. 2009. Análisis latitudinal y batimétrico de la comunidad de moluscos de mar profundo en el golfo de California, México. Brenesia 71-72: 41-54. Zamorano, P. y M.E. Hendrickx. 2011. A comparative analysis of deep-water mollusks from both sides of the Baja California Peninsula, Mexico. Cahiers de Biologie Marine 52: 13-22. Zamorano, P. y M.E. Hendrickx. (Aceptado). Distribution of Lucinoma heroica (Dall, 1901) in the minimum oxygen zone in the Gulf of California, Mexico. Marine Biodiversity Records (JMBA). Zamorano, P., M.E. Hendrickx y A. Toledano Granados. 2006. Distribution and ecology of deep water mollusks from the continental slope, southeastern Gulf of California, Mexico. Marine Biology 150 (5): 883-892. Zamorano, P., M.E. Hendrickx y A. Toledano Granados. 2007. New geographic and depth records for deep-water mollusks in the Gulf of California, Mexico. Nuevos registros geográficos y batimétricos para moluscos de mar profundo en el Golfo de California, México. Revista Mexicana de Biodiversidad 78 (2): 311-318. O peraciones oceanográficas en aguas profundas 103 Zamorano, P., M.E. Hendrickx, N. Méndez, S. Goméz, D. Serrano, H. Aguirre, J. Madrid y N. Morales-Serna. 2012. Explorando las aguas profundas del Pacífico mexicano: el proyecto TALUD. En: INE (ed.). La frontera final: el océano profundo. Instituto Nacional de Ecología, Secretaría del Medio Ambiente y Recursos Naturales (Semarnat). México, D.F. Zarate Montes, C. 2011. Taxonomía y ecología de equinodermos de aguas profundas del Pacífico mexicano. Tesis de Maestría. Universidad Nacional Autónoma de México, Posgrado en Ciencias del Mar y Limnología. 106 pp. 104 Biodiversidad y comunidades del talud continental del Pacífico mexicano